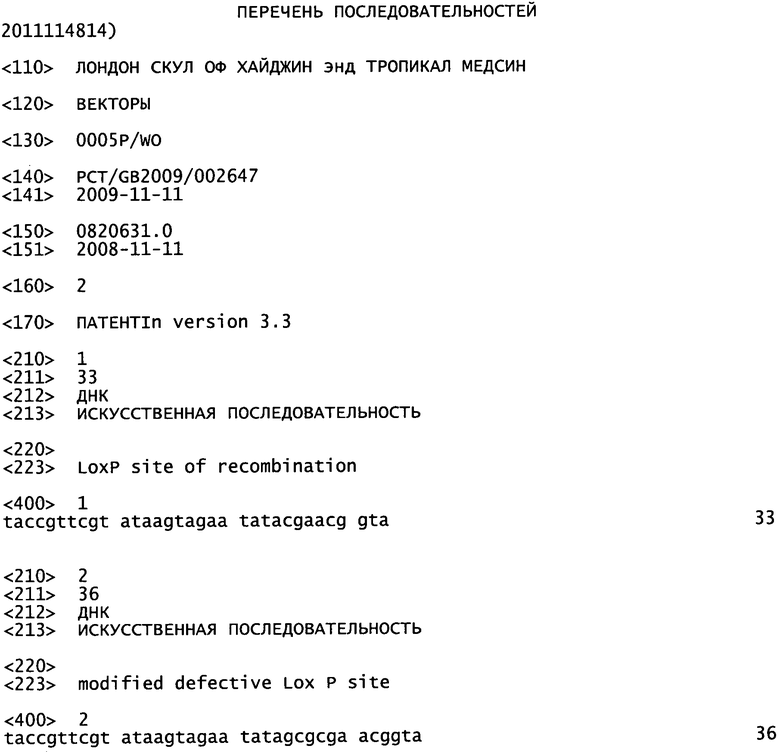
Настоящее изобретение относится к вектору переноса, применяемому для встраивания гена в генетический локус последовательности бакуловируса. Также настоящее изобретение относится к способам применения вектора переноса и получаемым бакмидам и бакуловирусам.
Систему экспрессии на основе бакуловируса применяли для экспрессии тысяч белков в клетках эукариот в целях структурных и биохимических исследований (14). Помимо того, что система на основе бакуловируса позволяет осуществлять экспрессию отдельных рекомбинантных белков, ее также применяли для совместной экспрессии нескольких белов, которые образуют комплексы (15-22). Это важно, поскольку скрининг всего генома для поиска взаимодействующих молекул показывает, что многие жизненно-важные функции живых клеток обусловлены комплексами белков из нескольких субъединиц (23). Для достижения совместной экспрессии белков с помощью системы на основе бакуловируса применяли две основные стратегии. Наиболее распространенный подход - это совместное инфицирование клетки двумя или более вирусами, каждый из которых экспрессирует один рекомбинантный белок (19, 24, 25). Однако, часто выход комплексов белков при гаком подходе варьирует в значительной степени, а совместное инфицирование одних и тех же клеток двумя разными бакуловирусами не подчиняется закону распределения Пуассона (26). Таким образом, хотя способ совместного инициирования технически прост, практически он ограничен получением только относительно простых комплексов для приложений, которые не требуют большого количества очищенного белка.
Для приложений, таких как структурные исследования и крупные испытания вакцин, для которых требуются большие количества белков, и для более сложных комплексов. образуемых множеством субъединиц, применяли альтернативный подход совместной экспрессии белков с нескольких одинаковых кассет экспрессии, которые встраивают в локус полиэдрина или Р10 (15-18, 22, 27). Этот подход имеет то преимущество, что каждая клетка в культуре, инфицированная рекомбинантным вирусом, воспроизводимо экспрессирует все белки, необходимые для образования белкового комплекса. При сравнении подходов совместного инфицирования и применения только одного бакуловируса, второй подход продемонстрировал значительно больший выход рекомбинантного комплекса (28). Однако экспрессия нескольких белков с применением одного рекомбинантного вируса не лишена недостатков. Хотя палочковидный бакуловирус, является довольно устойчивым к вставкам в его геном, гены трансфецируют в вирус посредством гомологичной рекомбинации, и вектор переноса должен иметь такой размер, чтобы с ним было легко работать в клетках Е.coli. Соответственно, существует ограничение по числу генов, которые можно встраивать в вектор переноса. На практике это означает, что редко можно экспрессировать более четырех белков из одного локуса. Кроме того, бакуловирус содержит последовательности и экспрессирует белки, которые запускают гомологичную рекомбинацию (29-32). Соответственно, вирусы, которые содержат большое количество повторов, склонны к перестройке и рекомбинации (21, 33, 34).
Подход совместной экспрессии с применением одного бакуловируса модифицировали посредством встраивания сайта loxP в локус chiA/cathepsin бакмиды, который уже содержит мишень Tn7 в локусе полиэдрина (20). Это позволяет осуществить встраивание множества экспрессионных кассет в каждый из этих локусов посредством рекомбинации в Е.coli. Как можно было ожидать исходя из того, что указанная система основана на повторяющейся дупликации кассеты экспрессии, которую модифицировали в целях экспрессии разных генов, существуют данные, что вставки в этой системе в определенной степени генетически нестабильны (21).
Недавно, для изучения бакуловируса появились новые возможности благодаря применению системы рекомбинации ЕТ, которая позволила производить селективное выключение вирусных генов (35-49). Однако потенциал данного метода для конструирования и облегчения экспрессии белков в генетических локусах помимо локуса р10 и полиэдрина не изучали.
Настоящее изобретение относится к способу эффективной экспрессии множества рекомбинантных белков (т.е., белков; не происходящих от бакуловируса) с генома одного бакуловируса. Данный способ позволяет эффективно вводить кассеты экспрессии отдельных белков в разные локусы в геноме вируса при помощи системы рекомбинации RT. Кроме того, данный способ дает возможность экспрессии комплексов с множеством субъединиц.
Экспрессия отдельных белков с применением бакуловируса в качестве системы экспрессии сейчас общепринята. Основная методология данной системы экспрессии очень мало изменилась с тех пор, как она была получена впервые. Поскольку геном бакуловируса состоит из 133 кб двухцепочечной ДНК, он слишком велик для того, чтобы работать с ним напрямую. Соответственно, гены, кодирующие чужеродные белки, с использованием клеток Е.coli клонируют в векторы, содержащие кассету экспрессии из вируса насекомых AcMNPV, которые затем вводят с вирусной ДНК в клетки насекомых, где белки вируса и клетки опосредуют гомологичную рекомбинацию, которая приводит к продукции инфекционного (применительно к клеткам насекомых) вируса, экспрессирующего чужеродный белок. Были получены различные варианты указанной системы, включая геномы вирусов, которые способны реплицироваться только в случае происхождения рекомбинации (1, 2), и проведение рекомбинации в Е.coli при помощи бакмиды, основанной на применении транспозона Tn7 (3) для ускорения отбора рекомбинантного вируса. Экспрессию комплекса из нескольких белков в клетках насекомых также осуществляли при помощи экспрессирующей системы бакуловируса, которая кратко описана выше. Изначально для совместного инфицирования восприимчивых клеток применяли несколько вирусов, каждый из которых экспрессировал отдельный белок, и комплекс белков очищали после экспрессии. Однако этот способ в лучшем случае неэффективен и не достаточно воспроизводим для промышленного масштаба. В качестве альтернативы создали векторы, которые включают несколько единиц экспрессии, но способны к рекомбинации с единственным генетическим локусом вируса (4-6). Эти векторы имели то преимущество, что они давали возможность получения всех субъединиц рекомбинантных белков в каждой инфицированной клетке и давали значительно более высокий выход комплекса рекомбимантных белков. Кроме того, поскольку все белки экспрессирует только один вирус, воспроизводимость результата инфекций была гораздо выше. Такие векторы имеют два недостатка. Во-первых, поскольку векторы содержат повторы, и бакуловирус экспрессирует белки, которые запускают гомологичную рекомбинацию, экспрессия, особенно с конструкций, содержащих три или четыре единицы экспрессии, не всегда достаточно генетически стабильна при большом количестве пересевов вирусов (которые необходимы при увеличении объема для промышленной экспрессии белков с применением бакуловирусов). Во-вторых, есть практическое ограничение по размеру вставки, которую легко можно сохранять и с которой можно работать в Е.coli, что ограничивает количество белков, которые можно экспрессировать с таких векторов. Другой подход, который был предложен для производства нескольких белков, состоит во встраивании чужеродного белка (белков) в один генетический локус в бакуловирусе, последующей селекции вируса, экспрессирующего этот белок, и применение генома этого вируса для рекомбинации во втором генетическом локусе для экспрессии других белков (7). Однако из-за необходимости высокой степени технической подготовки и большого времени, которых требует этот подход (16 дней для первого локуса и 25 дней для каждого последующего локуса), этот подход применяли очень редко. Последние разработки включали модификацию ВАС на основе системы транспозона Tn7 посредством встраивания сайта LoxP во второй генетический локус в бакуловирусе, чтобы также сделать возможным встраивание дополнительных генов в этот локус в Е.coli (8). Однако этот способ все еще не лишен проблем, связанных с тем, что наличие повторов вследствие дупликации промоторов в бактериальных векторах означает, что конструкция будет генетически нестабильна при множественных пересевах вируса, и векторы переноса будут быстро достигать размера, максимально допустимого для манипуляции в Е.coli.
Способ, предложенный в настоящем изобретении, позволяет преодолеть по меньшей мере некоторые из проблем, присущих ранее существующим способам, такие как генетическая нестабильность и потребность в множественных циклах рекомбинации.
Согласно первому аспекту настоящего изобретения предложен вектор переноса для встраивания гена в генетический локус последовательности бакуловируса, содержащий:
кассету экспрессии, содержащую эукариотический промотор, функционально связанный с геном; кассету двойной селекции, содержащую:
(i) экспрессируемую последовательность, кодирующую первый селектируемый маркер; и
(ii) экспрессируемую последовательность, кодирующую второй селектируемый маркер; и
последовательности, фланкирующие указанные кассету экспрессии и кассету двойной селекции, причем указанные последовательности в существенной степени соответствуют последовательностям генетического локуса в последовательности бакуловируса.
Вектор переноса, предложенный в настоящем изобретении, можно применял, для эффективного встраивания гена в генетический локус последовательности бакуловируса и для отбора последовательностей бакуловируса, которые содержат указанный ген. В частности, было показано, что применение кассеты двойной селекции значительно увеличивает число идентифицированных положительных клонов (т.е., последовательностей бакуловируса, содержащих встроенный ген) по сравнению с отбором при помощи одиночного селектируемого маркера. Это дает значительные преимущества по сравнению с применением одиночного селектируемого маркера. Трансформируется только небольшое количество бакуловирусов, и, следовательно, маркер экспрессируется только на очень низком уровне. Например, когда маркером является устойчивость к антибиотику, можно применять только очень низкую концентрацию антибиотика. Это приводит к селекции вариантов бактерий, которые являются устойчивыми к используемому антибиотику. Следовательно, некоторые бактерии, растущие на чашках, могут не содержать модификацию. Применение двух способов положительной селекции позволяет преодолеть вероятность таких ложно-положительных результатов отбора.
Вектором переноса, предложенным в соответствии с настоящим изобретении, может быть любой подходящий вектор, при условии, что он способен к рекомбинации с последовательностью бакуловируса путем гомологичной рекомбинации и обеспечивать встраивание гена в последовательность бакуловируса. Подходящие векторы переноса хорошо известны специалистам в данной области техники. Основные свойства вектора переноса обсуждаются ниже.
Ген, который нужно вставить в последовательность бакуловируса, может представлять собой любой гетерологичный ген (т.е., ген, не происходящий от бакуловируса). Предпочтительно, указанный гетерологичный ген кодирует субъединицу белкового комплекса. Указанный гетерологичный ген может кодировать белок вируса, компонент рецептора или комплекса шаперонов. Особенно предпочтительно, чтобы гетерологичный ген кодировал белок вируса, который вместе с другими белками вируса может образовывать вирусоподобную частицу (ВПЧ).
Генетическим локусом, в который встраивают ген в последовательности бакуловируса, может быть любой подходящий локус, который дает возможность экспрессировать такой встроенный ген, а также не нарушает способность последовательности бакуловируса к репликации. Также применяемый локус не должен влиять на способность бакуловируса к инфицированию клеток, т.е., к репликации или распространению из клетки в клетку. Такие локусы включают локусы полиэдрина и р10. Другие дополнительные подходящие локусы, обнаруженные авторами настоящего изобретения, включают ctx, egt, 39k, orf51, gp37, iap2 и odv-e56.
Последовательностью бакуловируса может быть любая подходящая последовательность бакуловируса, которая может реплицироваться в клетке насекомого и прокариотической клетке, такой как Е.coli. В частности, можно применять любой геном бакуловируса, который содержит репликой ВАС. Подходящие последовательности бакуловируса включают бакмиду AcMNPV.
Термин «кассета экспрессии» относится к комбинации элементов, необходимых для экспрессии гена. Соответственно, кассета экспрессии включает подходящий промотор, который делает возможной экспрессию кодируемого гена в клетках эукариот (например, клетках насекомых), и подходящую последовательность сигнала полиаденилирования, фланкирующую указанную последовательность гена. Кассеты экспрессии для экспрессии генов в клетках эукариот хорошо известны специалистам в данной области техники.
Промоторы, которые особо предпочтительны, включают вирусные поздние промоторы бакуловируса (например, р35) или вирусные очень поздние промоторы (например, промоторы полиэдрина и р10). Подходящие последовательности сигнала полиаденилирования включают последовательность полиаденилирования триптофангидроксилазы (Тпг) или последовательности полиаденилирования из генов бакуловируса.
Кассета экспрессии согласно настоящему изобретению может содержать более одного гена (например, 2, 3 или 4 гена), что приводит к экспрессии более чем одного гена. Однако обычно предпочтительно, чтобы кассета экспрессии содержала единственный ген.
Было показано, что кассета двойной селекции улучшает идентификацию и выделение клонов, которые успешно включили ген. Кассета двойной селекции содержит 2 различных селектируемых маркера. Также указанная кассета содержит любые элементы, необходимые для экспрессии маркеров в клетке бактерии, такие как один или более бактериальных промоторов. Этап отбора последовательностей, которые содержат ген, включает отбор последовательностей, содержащих оба селектируемых маркера. Данный этап отбора проводят в клетках бактерий, например, Е.coli. Селектируемыми маркерами могут быть любые маркеры, которые позволяют отбирать клетки, содержащие кассету. Например, маркеры могут быть визуальными, такие как маркер, вызывающий изменение внешнего вида или окраски колонии клеток. В качестве альтернативы, маркер может придавать устойчивость, например, к антибиотику. Два указанных селектируемых маркера могут быть маркерами одного типа, например, представлять собой устойчивость к двум разным антибиотикам, или маркерами разного типа, например, представлять собой устойчивость к антибиотику и визуальный маркер. Первый селектируемый маркер предпочтительно представляет собой визуальный маркер, такой как фрагмент LacZalpha, придающий синюю окраску клеткам, экспрессирующим фрагмент LacZalpha, в присутствии IPTG (изопропилтиогалактозида) и X-gal. Второй селектируемый маркер предпочтительно представляет собой любой маркер, придающий устойчивость к антибиотику, такую как устойчивость к хлорамфениколу, тетрациклину, нуромицину, ампициллину, пенициллину, апрамицину, канамицину или флеомицину. Особенно предпочтительно, чтобы второй маркер придавал устойчивость к флеомицину.
Согласно предпочтительному варианту реализации указанная кассета двойной селекции фланкирована последовательностями рекомбинации LoxP. Последовательности рекомбинации LoxP соединяются вместе в присутствии рекомбиназы Cre, что приводит к делеции последовательности, которая находится между ними. Соответственно, кассету двойной селекции можно удалить, что позволяет применять одни и те же селектируемые маркеры при введении любых дополнительных генов в последовательность бакуловируса. Предпочтительно модифицировать сайты LoxP так, чтобы между сайтами мог происходить только один цикл рекомбинации. Подходящие модифицированные сайты LoxP хороню известны специалистам в данной области техники, например, модифицированные сайты Lox66 и Lox71.
Последовательности, фланкирующие кассету экспрессии и кассету двойной селекции согласно настоящему изобретению, в существенной степени соответствуют последовательностям указанного генетического локуса, что дает возможность осуществления гомологичной рекомбинации между вектором переноса и последовательностью бакуловируса. Каждая фланкирующая последовательность предпочтительно должна иметь длину по меньшей мере 20 пар оснований (по), более предпочтительно длину по меньшей мере 30 по, и еще более предпочтительно, длину по меньшей мере 50 по, и должна содержать последовательность, позволяющую осуществлять специфичную рекомбинацию с последовательностью указанного генетического локуса.
Согласно второму аспекту настоящего изобретения предложен способ получения рекомбинантной бакмиды, включающий:
объединение бакмиды и вектора переноса, соответствующего первому аспекту настоящего изобретения, для обеспечения возможности гомологичной рекомбинации; и
селекцию рекомбинантной бакмиды, которая содержит кассету двойной селекции.
Отбор рекомбинантной бакмиды можно проводить при помощи стандартных методик в зависимости от кассеты двойной селекции. Способ получения рекомбинантной бакмиды может дополнительно включать детектирование наличия гена в бакмиде или продукта его экспрессии. Можно применять подходящие методики скрининга в зависимости от конкретного гена.
Кроме того, когда кассета селекции фланкирована последовательностями рекомбинации LoxP, способ предпочтительно дополнительно включает также индукцию рекомбинации между сайтами LoxP, например, посредством воздействия на бакмиду рекомбиназой Cre, для удаления кассеты двойной селекции. Предпочтительно, чтобы рекомбиназа Cre была под контролем индуцируемого промотора, такого как промотор арабинозы. Преимущество удаления кассеты двойной селекции состоит в том, что с бакмидой можно рекомбинировать дополнительный вектор переноса, причем указанный дополнительный вектор переноса может содержать ту же кассету двойной селекции. Отбор дополнительно рекомбинированной бакмиды можно проводить при помощи тех же методик, которые применяют после первого цикла рекомбинации.
Обычно способ получения рекомбинантной бакмиды реализуют в прокариотической клетке, поскольку системой на основе клетки прокариот можно легче и быстрее манипулировать. Предпочтительно, чтобы система на основе клетки прокариот обеспечивала возможность гомологичной рекомбинации. Такие системы включают систему рекомбинации лямбда ред, которая описана в заявке на Европейский патент ЕР-А-1291420. Предпочтительно, чтобы способ получения рекомбинантной бакмиды был реализован в E.coli. Особенно предпочтительно, чтобы способ был реализован с применением клеток E.coli линии EL350, которые содержат и интегрированный профаг лямбда, экспрессирующий exo, bet и gam под контролем регулируемого температурой промотора lambdaPL, и рекомбиназу Cre под контролем промотора, индуцируемого арабинозы.
Способ получения рекомбинантной бакмиды можно повторять несколько раз так, чтобы бакмида могла содержать несколько гетерологичных генов, т.е., 1, 2, 3, 4, 5, 6, 7, 8 или более гетерологичных генов. При попытках применения разных промоторов для экспрессии каждого встроенного гена, и если каждый ген встроен в отличный генетический локус, можно сократить или избежать повторов в бакмиде и тем самым гарантировать хорошую генетическую стабильность бакмиды. Также предпочтительно, чтобы между встроенными гетерологичными генами был расположен по меньшей мере один незаменимый ген. Такое расположение гарантирует, что если между любыми из встроенных последовательностей произойдет гомологичная рекомбинация, незаменимый ген будет удален, и образуется нежизнеспособная бакмида.
Согласно третьему аспекту настоящего изобретения предложена рекомбинантная бакмида, полученная посредством способа, соответствующего второму аспекту настоящего изобретения. Указанная бакмида содержит встроенный ген, а также может содержать остатки последовательностей LoxP. Эти остатки можно применять в качестве маркеров для идентификации бакмид. Предпочтительно, чтобы бакмида содержала по меньшей мере 5, 6, 7, 8 или 9 гетерологичных генов.
Согласно четвертому аспекту настоящего изобретения также предложен способ получения рекомбинантного бакуловируса, включающий культивирование эукариотических клеток, содержащих рекомбинантную бакмиду, полученную посредством способа, соответствующего второму аспекту настоящего изобретения при условиях, в которых образуется бакуловирус.
Предпочтительно, эукариотические клетки представляют собой клетки насекомых, более предпочтительно, их получают из насекомых Spodoptera frugiperda или Trichoplusia ni, например, клетки насекомых Sf21, Sf9 или TN 5B-1-4. Бакуловирус можно выделить при помощи стандартных методик и можно протестировать экспрессию встроенных гетерологичных генов.
Согласно пятому аспекту настоящего изобретения также предложен рекомбинантный бакуловирус, полученный посредством способа, соответствующего четвертому аспекту настоящего изобретения.
Согласно шестому аспекту настоящего изобретения также предложена рекомбинантная бакмида или рекомбинантный бакуловирус, которые экспрессируют совокупность гетерологичных белков, отличающиеся тем, что каждый белок экспрессируется с отдельного генетического локуса указанных бакмиды или бакуловируса. Также предпочтительно, чтобы между каждым генетическим локусом, экспрессирующим гетерологичный белок, был расположен по меньшей мере один незаменимый ген. Такое расположение гарантирует, что если между генетическими локусами, экспрессирующими гетерологичный ген, произойдет гомологичная рекомбинация, указанный незаменимый ген будет удален, и образуется нежизнеспособная бакмида или бакуловирус. Предпочтительно, чтобы указанные бакмида или бакуловирус экпрессировали по меньшей мере 3, более предпочтительно по меньшей мере 5, а наиболее предпочтительно по меньшей мере 8 белков. Указанные белки являются гетерологичными, т.е, их не кодируют существующие в природе бакуловирусы. Также предпочтительно, чтобы кодируемые гетерологичные белки были субъединицами белкового комплекса, такого как ВПЧ, комплекс рецептора или комплекс шаперонов.
Также предпочтительно, чтобы отдельные генетические локусы рекомбинантной бакмиды или бакуловируса были выбраны из: ctx, egt, 39k, orf51, gp37, iap2, odv-e56 и p10. Положение таких локусов специально описано применительно к бакуловирусу AcMNPV в Таблице 1. Специалисты в данной области техники могут легко определить положение локусов у других штаммов бакуловирусов и бакмид на основании этой информации.
Согласно седьмому аспекту настоящего изобретения также предложены рекомбинантная бакмида или рекомбинантный бакуловирус, в которых кассету экспрессии, кодирующая гетерологичный белок, встраивают в один или более из следующих генетических локусов: ctx, egt, 39k, orf51, gp37, iap2 и odv-e56.
Было показано, что указанные генетические локусы специфичным образом способствуют высокому уровню экспрессии гетерологичного белка, не нарушая ключевых функций бакмиды или бакуловируса.
Предпочтительно, чтобы рекомбинантная бакмида или рекомбинантный бакуловирус кодировали совокупность белков, и каждая кассета экспрессии была встроена в отдельный генетический локус. Также предпочтительно, чтобы рекомбинантная бакмида или рекомбинантный бакуловирус кодировали по меньшей мере 3, более предпочтительно но меньшей мере 5, а наиболее предпочтительно по меньшей мере 8 белков. Также предпочтительно, чтобы кодируемые гетерологичные белки были субъединицами белкового комплекса, такого как ВПЧ, комплекс рецептора или комплекс шаперонов.
Согласно восьмому аспекту настоящего изобретения также предложен вектор переноса для встраивания гена в генетический локус последовательности бакуловируса, содержащий:
кассету экспрессии, содержащую эукариотический промотор, функционально связанный с геном; и
последовательности, фланкирующие кассету экспрессии, причем указные последовательности в значительной степени соответствуют последовательностям генетического локуса в последовательности бакуловируса, при этом указанный генетический локус выбирают из числа: ctx, egt, 39k, orf51, gp37, iap2 и odv-e56.
Как указано выше, было показано, что при вставке гена в перечисленные генетические локусы можно получить высокий уровень экспрессии гена без нарушения ключевых функций последовательности бакуловируса.
Согласно девятому аспекту настоящего изобретения также предложен способ получения рекомбинантной бакмиды, включающий:
объединение бакмиды и вектора переноса, соответствующего восьмому аспекту настоящего изобретения, для обеспечения прохождения гомологичной рекомбинации; и
селекцию рекомбинантной бакмиды, содержащей кассету экспрессии.
Способы получения такой рекомбинантной бакмиды и отбора такой бакмиды хорошо известны специалистам в данной области техники. Согласно настоящему изобретению также предложена рекомбинантная бакмида, полученная указанным способом.
Согласно десятому аспекту настоящего изобретения также предложен способ получения рекомбинантного бакуловируса, включающий получение рекомбинантной бакмиды посредством способа, предложенного в настоящем изобретении, и культивирование эукариотической клетки, содержащей указанную бакмиду с получением бакуловируса.
Способы получения такого рекомбинантного бакуловируса хороню известны специалистам в данной области техники. Согласно настоящему изобретению также предложена рекомбинантный бакуловирус, полученный указанным способом.
Согласно настоящему изобретению также предложена клетка, содержащая вектор переноса, бакмиду или бакуловирус, соответствующие любому из аспектов настоящего изобретения. Согласно настоящему изобретению также предложен способ получения одного или более белков, включающий культивирование рекомбинантной бакмиды или бакуловируса, соответствующих любому из аспектов настоящего изобретения, при подходящих условиях. Указанные один или более белков кодируются одним или более генами и могут объединяться, образуя белковый комплекс, такой как ВПЧ, комплекс рецептора или комплекс шаперонов. Один или более белок можно изолировать при помощи стандартных методик, хорошо известных специалистам в данной области техники.
Ниже настоящее изобретение в качестве примера описано применительно к следующим фигурам.
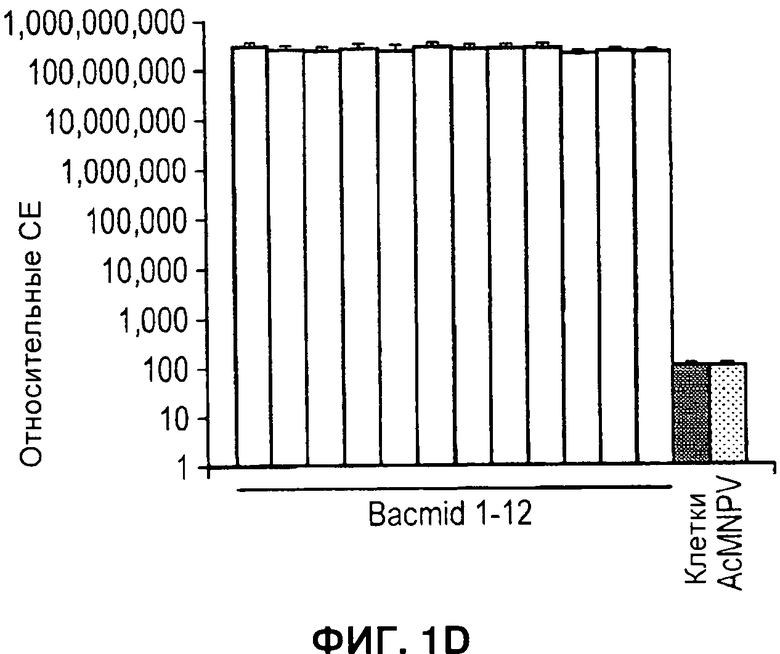
На Фигуре 1 показан улучшенный отбор рекомбинантов, полученных при клонировании ЕТ. А) Схема ДНК, применяемой для рекомбинации гена cat (хлорамфениколацетилтрансферазы) и двойных рекомбинаций. Обе конструкции содержат одинаковые фланкирующие последовательности бакуловируса (AcMNPV), кассету экспрессии люциферазы Renilla (Pp35-Rluc-Tph) и сайты loxP (LoxP). Двойной селектируемый маркер также содержит ген устойчивости к зеоцину (ZeoR) и фрагмент LacZa. В конструкции, устойчивой к хлорамфениколу, его заменяют на ген cat. В) Количество положительных колоний бактерий после рекомбинации ЕТ. ДНК, применяемую для рекомбинации, получали либо посредством ПЦР (полимеразной цепной реакции), либо путем расщепления рестрикционной эндонуклеазы, как указано. С) Правильность встраивания кассеты экспрессии Rluc подтверждали посредством ПЦР с применением одного праймера в Т-ДНК и одного фланкирования целевого локуса в ДНК бакуловируса. Надлежащий продукт ПЦР (показан стрелкой) можно получить только после надлежащей интеграции. Показаны результаты 12 независимых рекомбинаций (1-12), отсутствие образца контроля ПЦР (13), образец немодифицированной бакмиды (14) и образец Т-ДНК (15). Дорожка М - это маркер ДНК. D) Активность люциферазы Renilla (относительные световые единицы) через 48 часов после инфицирования в лизате клеток, инфицированных пересевом 2 рекомбинантных бакмид 1-12 с панели С. Фоновая активность эквивалентна активности лизатов неинфицированных клеток, и количество клеток, инфицированных немодифицированной бакмидой (AcMNPV) было более чем в 106 раз меньше.
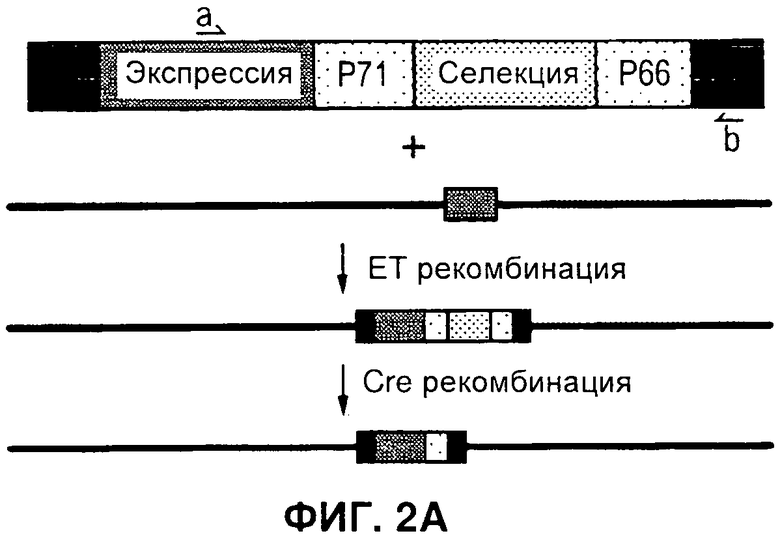
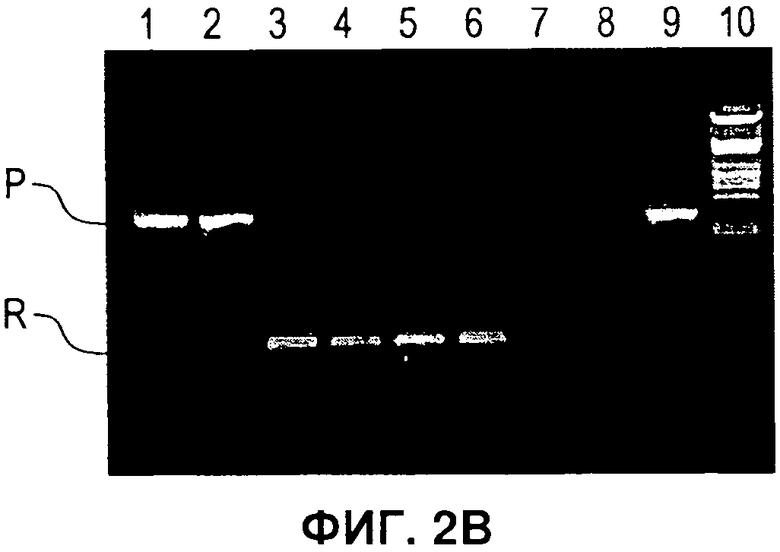
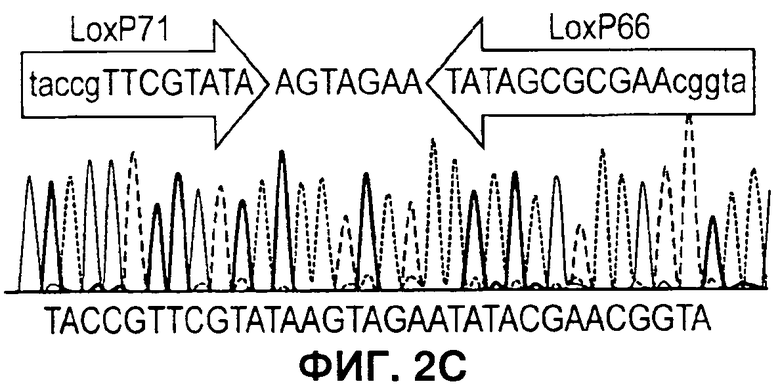
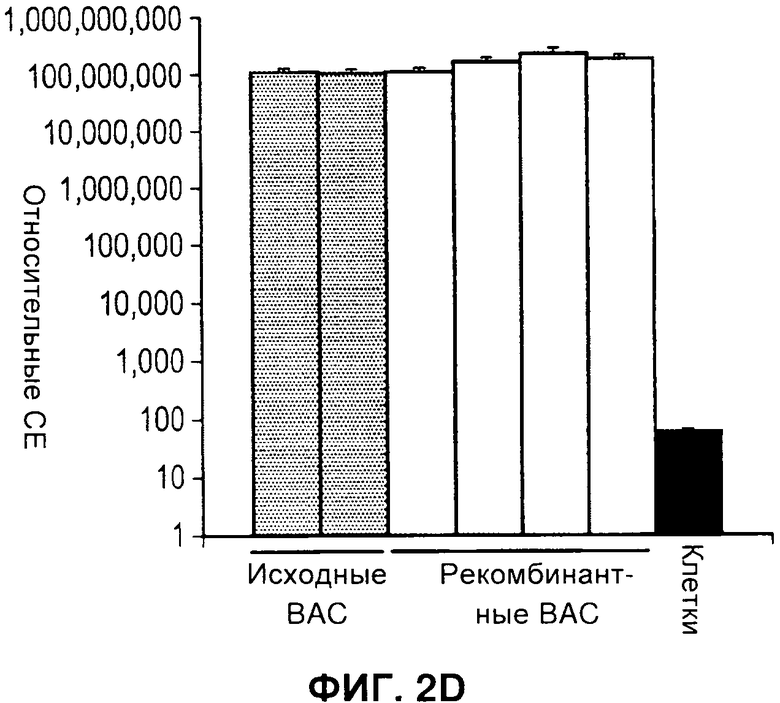
На Фигуре 2 показано селективное удаление маркерных генов. А) Схема, иллюстрирующая стратегию ЕТ рекомбинации кассеты экспрессии (экспрессии) в ДНК бакмиды AcMNPV и селективное удаление только маркерных генов (отбор) посредством опосредуемой Cre рекомбинации. В) ПЦР с применением праймеров а и b в А, дорожки 1-2: два независимых рекомбинанта бакмиды после ЕТ рекомбинации, дорожки 3-6: четыре независимых рекомбинанта после опосредуемой Cre рекомбинации в целях удаления селектируемых маркеров бактерий, дорожка 7: нет образца, дорожка 8: образец ДНК немодифицированной бакмиды, дорожка 9: образец ДНК плазмиды, содержащий кассету селектируемого маркера. дорожка 10: маркер ДНК. Продукты ПЦР, соответствующие размерам, прогнозируемым для исходных и рекомбинантных продуктов опосредуемой Cre рекомбинации, помечены как Р и R, соответственно. С) Файл трассировки последовательностей репрезентативного рекомбинанта из анализа ПЦР на панели В, подтверждающий наличие дефективного loxP, включающего плечи loxP71 и 1охР66, которые отображают рекомбинант, неспособный подвергнуться дальнейшей опосредуемой Cre рекомбинации. D) Активность люциферазы Renilla для исходных и рекомбинантных бакмид, показанных на панели В, при их трансфекции в клетки насекомых. Активность люциферазы Renilla оценивали через 48 часов после инфицирования после пересева рекомбинантого вируса. Фоновая активность неинфицированных клеток отмечена обозначением «Клетки».
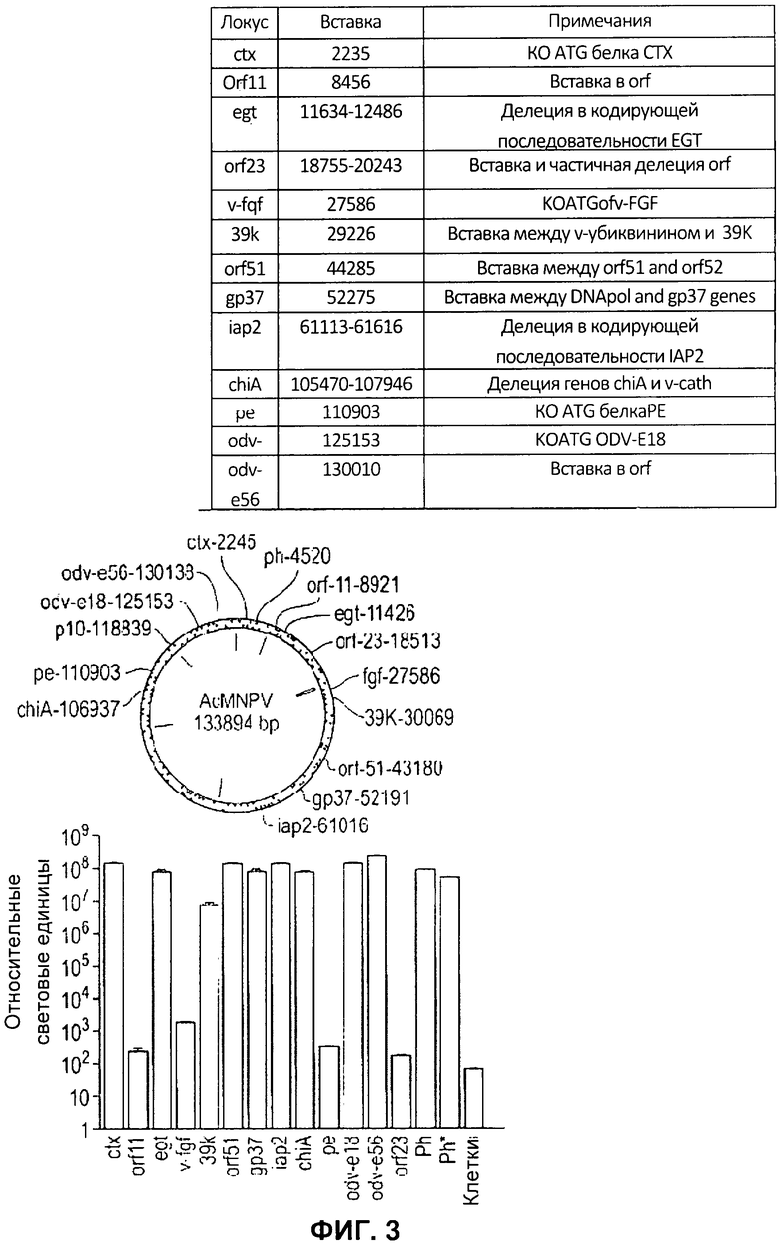
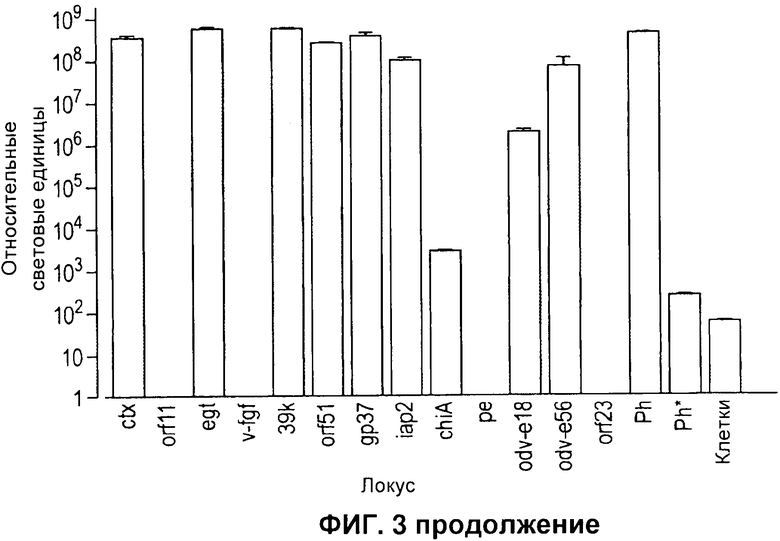
На Фигуре 3 показана идентификация дополнительных сайтов экспрессии рекомбинантных белков в геноме AcMNPV. А) Схема AcMNPV, иллюстрирующая относительные положения локусов, применяемых для экспрессии белков. В) Таблица, в которой указаны локусы, применяемые для встраивания, и любые дополнительные изменения, произведенные в локусе. С) Относительная активность люциферазы Renilla через 48 часов после инфицироваиия вирусом, модифицированным таким образом, чтобы он содержал дополнительную кассету экспрессии люциферазы светлячка в каждом указанном локусе. Все вирусы содержат одинаковую кассету экспрессии люциферазы Renilla в локусе р10 под контролем промотора р35. Планки погрешностей отражают стандартное отклонение для пяти повторов каждого локуса. D) Нормированная активность люциферазы светлячка, отражающая относительную экспрессию кассеты из промотора полиэдрина - терминатора полиэдрина люциферазы светлячка, встроенной в каждый локус, как указано. Планки погрешностей отражают стандартное отклонение для 5 повторов для каждого локуса. Активность люциферазы светлячка нормировали на контроль люциферазы Renilla, экспрессируемой с того же генома. Вставки люциферазы светлячка в локусы orfl 1, v-fgf, ре и orf23 были исключены, поскольку уровень активности люциферазы Renilla был более чем на 2 логарифмических единицы ниже, чем у контрольного вируса. Вирус Ph содержит ген люциферазы светлячка в локусе полиэдрина, а ген люциферазы Renilla в локусе р10. Вирус Ph* содержит такую же вставку гена люциферазы Renilla в локусе р10, но не содержит вставки гена люциферазы светлячка.
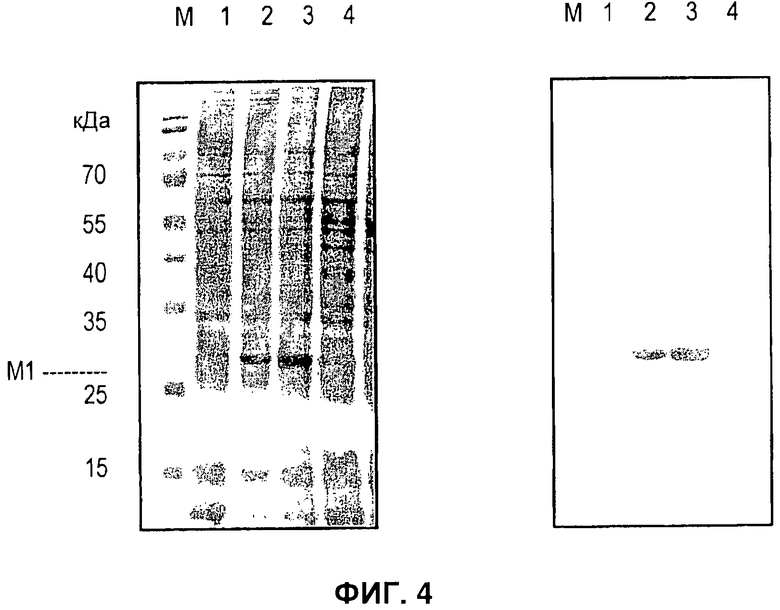
На Фигуре 4 показана экспрессия белка оболочки вируса гриппа M1 с локусов egt и полиэдрина. На левой панели показан окрашенный Кумасси полиакриламидный гель с додецилсульфатом натрия (SDS PAGE) всего белка из клеток, инфицированных контрольным бакуловирусом (дорожки 1 и 4), egt-M1 (дорожка 2) и YM1-M1 (дорожка 3). Размеры белков-маркеров (дорожка М) указаны слева от геля. На панели справа показан результат вестерн-блоттинга двойного геля, анализируемого при помощи антисыворотки анти-Н7М7.
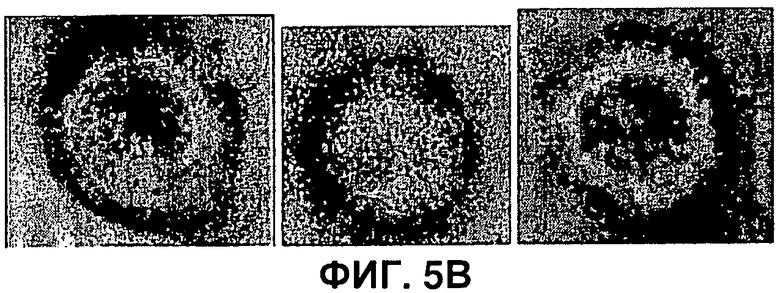
На Фигуре 5 показана совместная экспрессия белков оболочки вируса гриппа M1 и НА. а также образование ВПЧ. А) На панели слева показан окрашенный Кумасси гель всех белков; на панели справа показан результат вестерн-блоттинга двойного геля, анализируемого при помощи антисыворотки против вируса гриппа H7N7. Клетки инфицировали двойственным бакуловирусом, экспрессирующим и НА, и M1 (дорожка 1), бакуловирусом, экспрессирующим только НА (дорожка 2), клетки контроля (дорожка 3), бакуловирусом, экспрессирующим только M1 (дорожка 4). В) Электронно-микроскопические снимки ВПЧ вируса гриппа, с отрицательной окраской. С) ВПЧ вируса гриппа после иммунного окрашивания золотом (НА); частицы золота показаны стрелками.
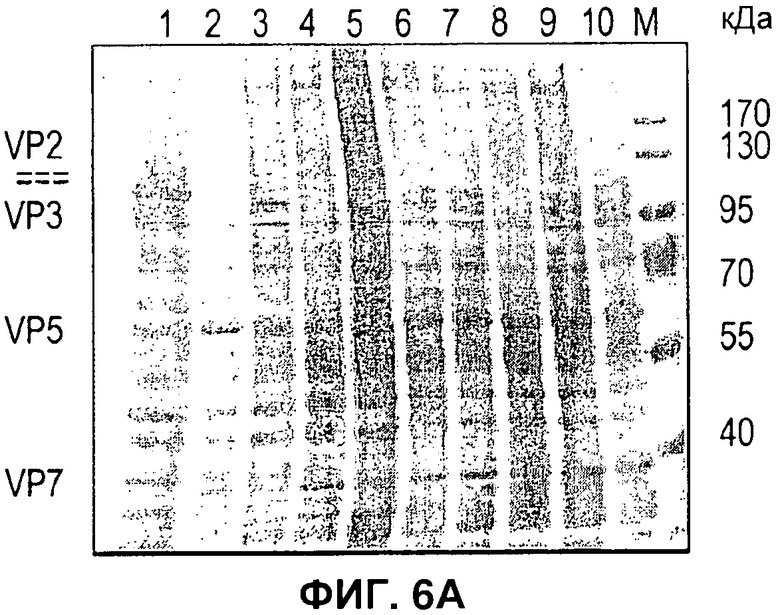
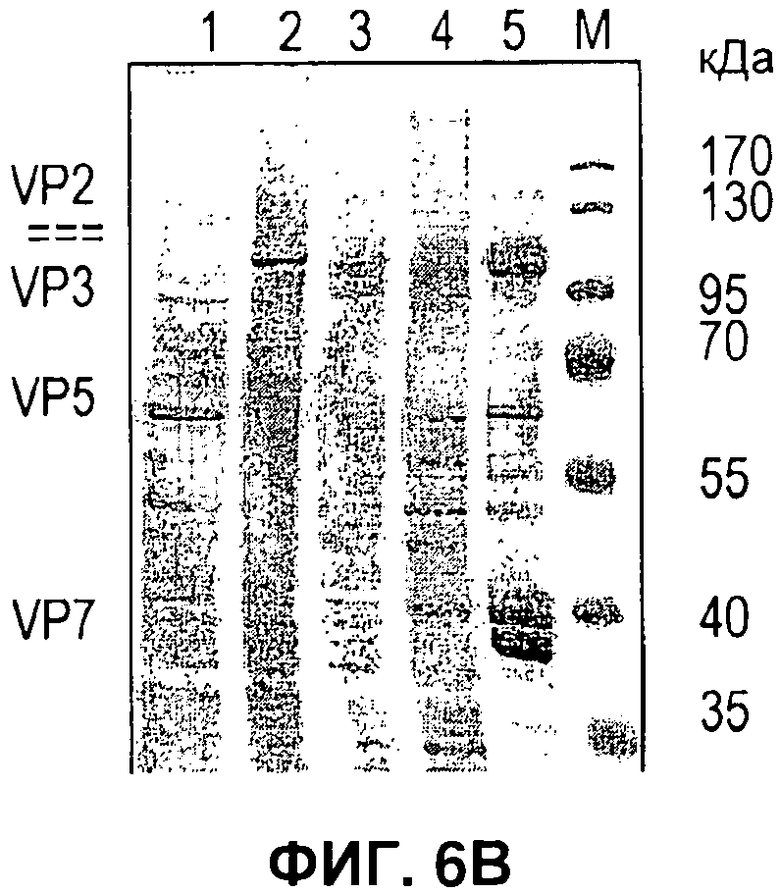
На Фигуре 6 показано получение бакуловируса, одновременно экспрессирующего 4 белка. A) Лизат клеток, инфицированных бакуловирусом, экспрессирующим VP2 (дорожка 1); VP5 (дорожка 2), VP3 (дорожка 3) и VP7 (дорожка 4), неинфицированных клеток (дорожка 5) или всех 4 белков (дорожки 6-10), положение белков-маркеров (М) и размер в кДа, как указано. B) Лизат клеток, экспрессирующих VP5, VP2, VP3, VP7 (дорожки 1-4) и частично очищенные ВПЧ (дорожка 5). С) В таблице указан локус встраивания и промотор вируса, применяемый для экспрессии каждого белка ВПЧ. D) Электронно-микроскопические снимки отрицательной окраски при иммунном окрашивании золотом (для VP5), показывающие очищенные ВПЧ BTV.
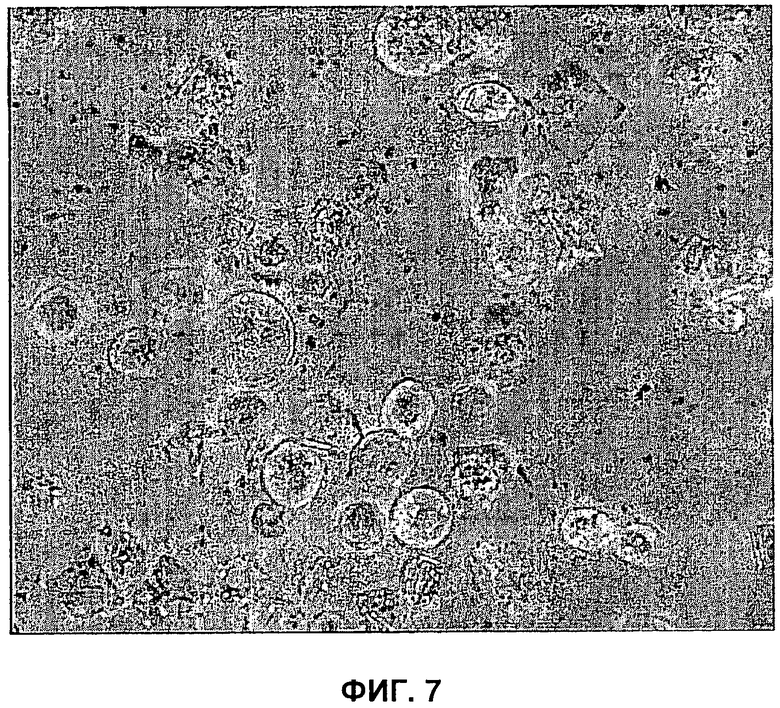
На Фигуре 7 показаны кристаллические пластинки, образованные при гиперэкспрессии ССТ5. Четырехугольные агрегаты белков в питательной среде очень позднего инфицирования бакуловирусом, экспрессирующим ССТ5.
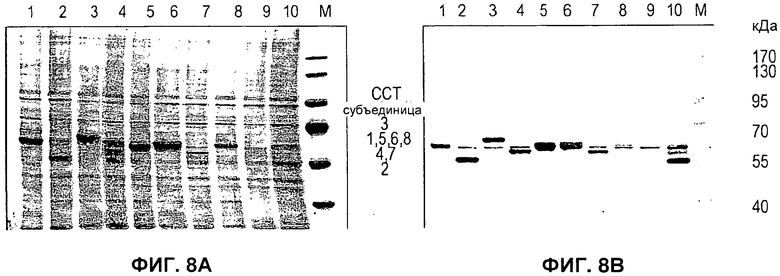
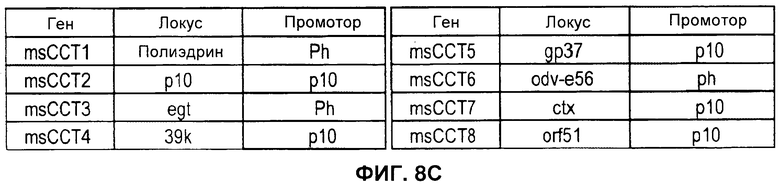
На Фигуре 8 показана экспрессия ССТ в клетках насекомых. А) Окрашенный Кумасси лизат клеток, инфицированных бакуловирусом, экспрессирующим ССТ1-ССТ8 (дорожки 1-8, соответственно), только клетки (дорожка 9) или только бакуловирус, одновременно экспрессирующий 7 субъединиц ССТ (дорожка 10). В) Результат вестерн-блоттинга двойного геля с применением антисыворотки из поликлональных анти-ССТ антител мыши. C) В таблицах указаны разные генетические локусы и промоторы, применяемые для экспрессии каждой из субъединиц ССТ.
ПРИМЕРЫ
Система экспрессии на основе бакуловируса обладает твердо установленным потенциалом для получения больших количество правильно скрученных белков эукариот для исследований ферментов и структурных исследований.
Однако становится все очевиднее, что многие, если не большинство, белки активны в клетках в составе комплексов, образованных продуктами нескольких различных генов. При возрастающем темпе открытий в области структуры белков существует реальная потребность в системах для быстрого и надежного получения и очистки комплексов белков. В частности, это справедливо для крупных комплексов, которые требуют одновременной экспрессии, а также скручивания и процессинга нескольких субъединиц белков в клетке эукариот. Ранее изобретатели разработали векторы переноса на основе бакуловируса, способные к экспрессии 2, 3, 4 и 5 белков с генома одного бакуловируса. В предыдущих исследованиях главная цель изобретателей состояла в получении системы, способной синтезировать большие количества комплексов белков вируса, содержащих не эквимолярные количества кодируемых вирусом белков для исследований ферментов и структурных исследований. Одно из наблюдений, сделанных при работе с указанными системами, состоит в том, что отдельный бакуловирус, экспрессирующий несколько генов, образует желаемые комплексы белков гораздо эффективнее, чем клетки насекомых, инфицированные совместно несколькими бакуловирусами, экспрессирующими отдельные гены. В настоящем исследовании изобретатели применили новые технологии для разработки этого наблюдения для получения рекомбинантных бакуловирусов, экспрессирующих несколько белков для создания биологически значимых комплексов белков млекопитающих.
В частности, изобретатели адаптировали и усовершенствовали новые технологии конструирования хромосом бактерий для быстрого и эффективного получения рекомбинантных бакуловирусов, одновременно экспрессирующих несколько белков. Новые разработанные системы в 30 раз более эффективны, чем традиционные процессы, и они позволяют производить стандартную вставку генов в любой локус в геноме бакуловируса. Эти исследования позволили идентифицировать 7 новых генетических локусов в геноме бакуловируса, которые позволяют проводить экспрессию рекомбинантных белков на высоком уровне. Кроме того, изобретатели продемонстрировали, что можно проводить многократно повторяющиеся циклы рекомбинации, что позволяет генерировать геномы вирусов, экспрессирующие несколько белков в комплексе в результате вставок отдельных генов. Например, изобретатели экпрессировали комплексы белков ВПЧ для вируса гриппа A (подтип Н7) и вируса катаральной лихорадки овец (серотип 1), а также 8 субъединиц комплекса шаперонов млекопитающих ССТ (TCP), который находится в центре внимания в исследованиях рака.
МАТЕРИАЛЫ И МЕТОДЫ
Встраивание отдельного гена в несколько локусов у E.coli получают посредством рекомбинации лямбда ред (Lambda red).
Изобретение включает применение локусов бакуловируса, которые ранее не применяли для экспрессии нескольких белков, и включение селектируемых маркеров, которые можно удалять из генома бакуловируса и в дальнейшем применять повторно. В отличие от других систем гены рекомбинантных белков не фланкируют повторами, чтобы повысить их генетическую стабильность.
Векторы переноса содержат область AcMNPV для управления гомологичной рекомбинацией, кассету экспрессии (промотор AcMNPV, полилинкер, сигнал полиаденилирования) и кассету селекции бактерий. Применяемый нромотор AcMNPV получают из позднего (например, р35) или очень позднего (полиэдрин, р10) гена вирусов. Кассета селекции бактерий включает мутантный сайт LoxP - селектируемый маркер бактерий - мутантный сайт LoxP. Мутантные сайты LoxP являются вариантами LoxP66 и LoxP71 (9). Сайты LoxP для каждого селектируемого маркера конструируют так, чтобы при рекомбинации маркер удалялся путем разрушения сайта LoxP, который остается. Каждый селектируемый маркер содержит разные мутантные плечи LoxP, чтобы не происходила рекомбинация между генами разных селектируемых маркеров, введенными в один и тот же бакуловирус.
Выбор системы, применяемой для экспрессии рекомбинантных белков с нескольких локусов
План оригинального исследования включал применение гомологичной рекомбинации в клетках насекомых или альтернативный способ ЕТ рекомбинации в E.coli, который даст возможность эффективного встраивания кассет экспрессии для рекомбинантных белков в разных локусах в геноме бакуловируса. Ранее полученные нами данные указывают на то, что линеаризация вектора переноса приводит к значительному увеличению частоты рекомбинации (до ~30%) в локусе (р10), который не демонстрировал положительного отбора, помимо производимого рекомбинантного белка. В начале проекта исследования были рассмотрены две системы (ЕТ рекомбинация и рекомбинация в клетках насекомых) в связи с их воспроизводимостью и временем, которое занимает вся вставка рекомбинантного гена в конкретный локус и проверка экспрессии получаемого белка. Что касается времени. которое занимает каждый цикл экспрессии, система ЕТ рекомбинации происходила быстрее. в основном вследствие сокращения времени, требуемого для подготовки генома бакуловируса к вставке генов во второй генетический локус бакуловируса. Кроме того, было возможно создать подход, позволяющий подтверждать генетические вставки независимо от экспрессии трансгена, и, следовательно, потребность в выращивании вируса в клетках насекомых для проверки экспрессии становилась менее критичной, чем для системы, полностью основанной на клетках насекомых. Во-вторых, хотя в систему бакуловируса можно вводить маркер и вставляемые гены совместно, число таких маркеров ограничено, и каждый из них можно применять только один раз. Поскольку одной из задач проекта была экспрессия комплекса ССТ мыши (ТСР1) (8 субъединиц), на ранней стадии в качестве приоритетного направления исследования был выбран подход ЕТ рекомбинации, поскольку эта система была быстрее, и она в большей степени способна приспособиться к множественным генетическим вставкам.
ПРИМЕР 1
Разработка реагентов, позволяющих проводить эффективный отбор ЕТ рекомбинантов.
Первые эксперименты с применением подхода ЕТ были неудачными. Были проведены эксперименты, в которых ген-репортер (люцифераза) вводили в искусственную хромосому бактерии, содержащую полный геном AcMNPV (бакмиду) и проводили отбор по гену устойчивости к хлорамфениколу, который был интегрирован с геном-репортером люциферазы. Хотя были обнаружены колонии бактерий, устойчивых к хлорамфениколу, последующий анализ показал, что они не содержали должным образом модифицированной бакмиды с геном-репортером люциферазы. Аналогичные результаты были получены, когда ген-репортер, предназначенный для экспрессии в клетках насекомых только в присутствии реплицирующегося AcMNPV, был заменен на GFP (зеленый флюоресцентный белок). Однако когда конструкцию с GFP применили для рекомбинации популяции E.coli, без отбора с хлорамфениколом, и приготовили ДНК бакмиды для всей трансформированной популяции E.coli, то указанная ДНК при трансфекции ее в клетки насекомых привела к образованию нескольких очагов флюоресценции (Sf21). Эти данные показали, что проблема заключалась не в рекомбинации самой по себе, а в следующем за рекомбинацией отборе содержащих вставки бактерий. Чтобы преодолеть эту проблему, изобретатели разработали новую кассету селекции, которую включали в систему двойных маркеров на основе фрагмента LacZa и гена устойчивости к Зеоцину, фланкируемых модифицированными сайтами рекомбинации LoxP (Фигура 1, А). При применении данной системы в чашки для отбора добавляли достаточное количество Зеоцина, чтобы сократить, но не уничтожить фоновый рост колоний, и отбирали рекомбинанты на основании синего фенотипа колоний в присутствии IPTG и X-gal. Чтобы оценить относительную эффективность выделения рекомбинантов при применении систем хлорамфеникола (cat) и двойной селекции, изобретатели провели эксперимент, в котором компетентные для рекомбинации Е.coli, содержащие немодифицированную бакмиду, подвергали электропорации с введением 30 нг (~12,5 фмоль) либо кассеты на основе хлорамфеникола, либо кассеты двойной селекции (Фигура 1, В). Поскольку идеальная система для введения последовательностей, кодирующих чужеродные белки, не может включать многократную ПЦР встроенного гена, изобретатели также сравнили эффективность рекомбинации между ДНК, амплифицированной посредством ПЦР, которая является стандартом для ЕТ рекомбинации. и ДНК, выделенной при помощи рестрикционных эндонуклеаз, очищенной в геле. Для ПЦР разработали такие праймеры, чтобы концы продуктов ПЦР соответствовали концам фрагментов, выделенных при помощи рестрикционных эндонуклеаз.
Двойной отбор привел к 20-кратному увеличению количества положительных колоний по сравнению с отбором только на основе хлорамфеникола, когда рекомбинирующие фрагменты ДНК генерировали посредством Г1ЦР, и к 30-кратному увеличению, когда фрагменты ДНК выделяли путем расщепления ДНК плазмиды рестрикционными эндонуклеазами (Фигура 1, В). Эти различия были значимыми (критерий Стьюдента, р=0,03). Для отбора на основе хлорамфеникола между количеством колоний, полученных с ДНК из ПЦР и с ДНК, сгенерированной рестрикционными эндонуклеазами, различий не было. Однако для двойного отбора среднее количество колоний было в четыре раза больше для ДНК, полученной при помощи рестрикционных эндонуклеаз, чем для ДНК, амплифицированной путем ПЦР (критерий Стьюдента, р=0,05). Чтобы подтвердить, что рекомбинанты, полученные при новом двойном отборе, были истинными, представляющими модифицированную бакмиду, содержащую встроенную конструкцию экспрессии, проводили ПЦР на ДНК бакмиды, выделенной из положительных колоний. Один праймер разработали для последовательности внутри маркера устойчивости к Зеоцину, а другой праймер был направлен на последовательность, присутствующую только в ДНК бакмиды, фланкирующей правильный сайт встраивания, но не в Т-ДНК.
Таким образом, продукт ПЦР можно получить только тогда, когда между линейной ДНК, применяемой для рекомбинации, и бакмидой произошла рекомбинация. Посредством данного способа тестировали образцы ДНК 12 отдельных бакмид (Фигура 1C, дорожки 1-12), все они были положительны в отношении продукта, указывающего на правильно направленную рекомбинацию. Напротив, ни ДНК одной только бакмиды, ни ДНК плазмиды. содержащей фрагмент ДНК, применяемый для рекомбинации, не могли служить матрицей для получения продукта ПЦР (Фигура 1C, дорожки 14 и 15, соответственно). Чтобы получить дальнейшее подтверждение того, что рекомбинация привела к образованию инфекционного рекомбинантного бакуловируса, клетки Sf21 трансфецировали теми же 12 клонами ПЦР-положительных бакмид, и пересеяли дважды, затем через 48 часов после инфицирования оценивали активность люциферазы Renilla в клетках, инфицированных каждым из рекомбинантов. Клетки, инфицированные каждым из 12 рекомбинантных вирусов, обладали активностью люциферазы Renilla, которая в 106 раз превышала фоновую активность (Фигура 1, D). На основании указанных данных в дальнейших исследованиях изобретатели перешли к применению двойного отбора.
ПРИМЕР 2
Опосредуемое Cre удаление селектируемого маркера позволяет проводить несколько циклов рекомбинации с одним и тем же двойным отбором.
Для экспрессии комплексов белков из нескольких разных субъединиц с применением вставок в один локус, разработали такую систему двойного маркера, чтобы кассета селекции была фланкирована модифицированными сайтами LoxP. Указанные сайты объединяют в себе как мутации lox66 и lox71, которые ограничивают опосредуемую Cre рекомбинацию до одного цикла (1), так и мутацию в спенсере, снижающую гомологию с сайтами loxP дикого типа. Таким образом, инкубация модифицированной бакмиды с рекомбиназой Cre приводит к удалению двойного селектируемого маркера и к инактивации сайта рекомбинации lox, но оставляет на месте кассету экспрессии бакуловируса (Фигура 2А). Чтобы подтвердить, что эту стратегию можно успешно применять для конструирования множественных вставок в ДНК бакмиды, Cre-рекомбинацию применяли для удаления двойного маркера из бакмиды, в которую был встроен ген-репортер люциферазы Renilla. Рекомбинацию проводили в E.coli с применением линии клеток EL350 (2), которая содержит и интегрированный профаг лямбда, экспрессирующий exo, be, и gam под контролем регулируемого температурой промотора, и рекомбиназу Cre под контролем индуцируемого промотора арабинозы. Клетки EL350, содержащие три бакмиды, модифицированные в целях включения вставки люциферазы Renilla для двойной селекции (Фигура 1), индуцировали арабинозой, затем наносили на чашки, содержащие канамицин для отбора бакмид и X-gal для отбора колоний, которые потеряли селектируемый маркер при опосредуемой Cre рекомбинации. ДНК бакмиды очищали от четырех предполагаемых рекомбинантов, а опосредованную Cre рекомбинацию подтверждали ПЦР с применением праймеров, фланкирующих селектируемые маркеры (Фигура 2В). Все четыре рекомбинанта содержали продукты ПЦР, соответствующие но размеру продуктам, ожидаемым при успешной Cre-рекомбинации. В дальнейшем это подтверждали секвенированием по модифицированному сайту loxP 4 рекомбинантов (Фигура 2С). Рекомбинанты содержали поврежденный сайт loxP s, который включал мутации и loxP71, и loxP66, и не был компетентен в отношении последующих циклов рекомбинации. Чтобы получить дальнейшее подтверждение того, что рекомбинанты бакмид, полученные при опосредуемой Cre рекомбинации, жизнеспособны в клетках насекомых, ДНК бакмид трансфецировали в клетки насекомых и оценивали активность люциферазы после двух пересевов, как прежде. Все рекомбинанты демонстрировали активность люциферазы Renilla, которая была эквивалентна активности для исходных бакмид до Cre-рекомбинации (Фигура 2D).
ПРИМЕР 3
Идентификация генетического локуса в геноме бакуловируса, подходящего для интенсивной экспрессии гетерологичных белков.
Несмотря на работу по экстенсивной экспрессии белков, которую проводили на системе экспрессии на основе бакуловируса, в основном экспрессия была сосредоточена на замещении генов полиэдрина или р10 в целях экспрессии рекомбинантных белков. В относительно небольшом числе литературных источников описывается применение альтернативных локусов для экспрессии рекомбинантных белков. Чтобы проверить, можно ли эффективно применять отбор в разных генетических локусах бакуловируса, на одной из бакмид, уже содержащих ген люциферазы Renilla, проводили второй цикл рекомбинации. В этих экспериментах тот же промотор полиэдрина - люцифераза светлячка - терминатор полиэдрина вставляли независимо всего в 13 разных генетических локусов (ctx, 25 orf11, egt, orf23, v-fgf, 39k, orf51, gp37, iap2, chiA, pe, odv-el8 и odv-e56), генерируя бакуловирус с двойственной экспрессией для белков люциферазы Renilla и люциферазы светлячка (Фигура 3А). Локусы выбирали в качестве сайтов встраивания в этих экспериментах путем определения, какие гены бакуловируса не являются ключевыми для роста вируса в культуре ткани, и какое расположение генов бакуловируса в конкретных локусах благоприятствует вставке дополнительной кассеты экспрессии. Тем не менее, в некоторые локусы вносили некоторые дополнительные изменения, в частности изменения, которые привели к выключению конкретных генов бакуловируса (Фигура 3В). Рекомбинантные вирусы пересевали дважды в клетки насекомых 5/21, и на третьем пересеве клетки собирали через 48 часов после инфицирования, лизировали и оценивали в них активность люциферазы светлячка и Renilla. Активность люциферазы Renilla применяли в качестве маркера репликации вирусов и экспрессии белка, поскольку все рекомбинанты содержали одинаковую запускаемую промотором р35 кассету люциферазы Renilla в локусе р10. Экспрессию люциферазы светлячка в каждом новом локусе сравнивали с экспрессией у вируса, содержащего ген люциферазы светлячка в локусе полиэдрина и такого же эталонного гена люциферазы Renilla. Из 13 исследуемых локусов, 9 демонстрировали активность люциферазы Renilla, превышающей фоновую активность по меньшей мере в 105 раз, и из них 8 (ctx, egt, orf51, gp37, iap2, chiA, odv-el8 и odv-e56) демонстрировали активность люциферазы Renilla, которая была равна или превышала активность, измеренную только для исходной люциферазы Renilla, и для контролей люциферазы светлячка в локусе полиэдрина (Фигура 3С). Четыре локуса (prf11, v-fgf, ре и orf23), которые привели к образованию вируса, демонстрирующего активность люциферазы Renilla, превышавшую фоновую активность не более 10 раз, исключили из дальнейшего анализа. Для остальных вирусов активность люциферазы Renilla применяли в качестве эталонного значения для нормировки активности люциферазы светлячка и вычисления меры относительной экспрессии люциферазы светлячка с каждого локуса (Фигура 3D). 7 локусов (ctx, egt, 39k, orf51, gp37, iap2 и odv-e56) демонстрировали активность люциферазы светлячка, превышавшую по меньшей мере в 10 раз фоновую активность, и активность, аналогичную фоновой, когда тот же ген экспрессировался с локуса полиэдрина (Фигура 3D). Два вируса (chiA и odv-elS) демонстрировали высокий уровень активности люциферазы Renilla, но относительно низкую экспрессию люциферазы светлячка.
Данное исследование показало, что возможна интенсивная экспрессия чужеродных белков с нескольких генетических локусов в геноме бакуловируса, и позволило идентифицировать семь локусов (ctx, egt, 39k, orf51, gp37, iap2 и odv-e56) помимо полиэдрина и р10, которые обеспечивают хорошую экспрессию. Из этих сайтов 39k, orf51 и gp37 обеспечивают встраивание в ДНК, фланкирующую кодирующую область гена, что непосредственно не нарушает экспрессию белков. Напротив, можно ожидать, что встраивание в ctx, egt, iap2 и odv-e56 будет препятствовать экспрессии соответствующих белков с данных генов. Первые три из указанных генов ранее описывали как не ключевые гены для роста вируса в культуре клеток (3-6). Также сообщали об укорочении белка ODV-E56 (7).
Среди локусов, которые не давали хорошей экспрессии гена-репортера люциферазы светлячка, четыре (orf11, v-fgf, ре и orf23) также приводили к сниженной экспрессии белка-маркера люциферазы Renilla, который присутствовал во всех рекомбинантах. Поскольку исследование было сосредоточено на идентификации сайтов, подходящих для встраивания экспрессирующих конструкций с очень поздним промотором, точную причину такой сниженной экспрессии не выясняли. Возможные объяснения включают локус-специфичные эффекты на репликацию или транскрипцию вируса и нарушение целостности элементов ключевых промоторов или энхансеров для фланкирующих генов. Низкий уровень экспрессии люциферазы светлячка в вирусах с вставками в локусы chiA и odv-e18 был непредвиденным по разным причинам. В других публикациях есть сведения о том, что регистрировали вставку кассет экспрессии рекомбинантных белков в локус chiA (8, 9).
Возможно, что сниженный уровень экспрессии, наблюдаемый в отношении гена люциферазы светлячка в этом локусе в наших экспериментах, связан с эффектами на гены, фланкирующие вставку. Для odv-el8 в последних публикациях, где применяли тот же мутантный вирус, выдвигают предположение, что данный белок является ключевым для наработки отпочкованных вирусов и перемещения от клетки к клетке (10, 11). Указанные исследования были основаны на мутантах, в которых имела место делеция в кодирующей последовательности локуса odv-el8 и в вышележащем фланкирующем гене. В экспериментах, где локус odv-el8 инактивировали посредством мутации ATG кодирующей последовательности на GAT и впоследствии вставляли кассету люциферазы светлячка в этой точке в гене, удавалось выделить инфекционный вирус. Экспрессия люциферазы Renilla при третьем пересеве эквивалентна экспрессии в исходном вирусе, что указывает на то, что нарушения способности этого мутантного вируса к репликации не происходило. Однако, если принять во внимание снижение уровня люциферазы светлячка ~ на 2 лог. единицы по сравнению с уровнем у вируса без указанной мутации, нельзя исключать возможность того, что маленькая популяция вирусов, в которой мутация подверглась репарации, дополняла вторую популяцию, экспрессирующую ген-репортер.
ПРИМЕР 4
Экспрессия комплексов белков с применением мультилокусной экспрессии у бакуловируса
Пример 3 посвящен применению генов-репортеров для количественной оценки потенциала разных локусов бакуловируса к экспрессии рекомбинантного белка. Чтобы выяснить, можно ли экспрессировать и выделить комплексы рекомбинантных белков, были выбраны в качестве объекта три специфических комплекса с разным числом субъединиц белков. Вирусоподобные частицы для вируса гриппа (А/изоляция/масса/1/80) подтипа Н7 получали путем совместной экспрессии белков вируса M1 и НА (комплекс из 2 белков), а для вируса катаральной лихорадки овец (BTV) серотипа 1 - путем совместной экспрессии VP2, VP3, VP5 и VP7 (комплекс из 4 белков). Кроме того, изобретатели выбрали в качестве объекта изучения экспрессию всех 8 субъединиц комплекса щаперонов мыши ССТ, чтобы применять его для дальнейших функциональных и структурных исследований.
ВПЧ вируса гриппа А
Для этих экспериментов все гены вируса гриппа брали у адаптированного для линии мышей штамма H7N7 (А/изоляция/масса/1/80), полученного от исследователей в Марбургском университете имени Филиппа, Германия.
Чтобы проверить, способна ли мультилокусная система экспрессии производить комплексы из двух белков, сначала кодирующую последовательность белка M1 вируса гриппа A вставляли в локус egt под контролем промотора полиэдрина. Для сравнения тот же ген вставляли в традиционный вектор экспрессии бакуловируса (pAc-YM1), направляемый локусом полиэдрина. После рекомбинации оба получаемых бакуловируса экспрессировали белок M1. Уровень экспрессии белка с локусов egt и полиэдрина был значительно выше, чем уровень экспрессии любого другого белка вируса или клетки в клетках, инфицированных бакуловирусом, при оценке методом SDS-PAGE с окраской Кумасси (Фигура 4).
Чтобы сгенерировать двойственный вирус, экспрессирующий и M1, и HA, кассету селекции бактерий удаляли из экпрессирующей M1 бакмиды посредством Cre-рекомбинации, а ген НА из того же штамма вируса гриппа (А/изоляция/масса/1/80) вставляли в локус р10 посредством второго цикла рекомбинации ЕТ. Совместную экспрессию M1 и НА из полученного вируса подтверждали посредством SDS-PAGE и вестерн-блоттинга, как указано выше (Фигура 5А). Кроме того, ВПЧ вируса гриппа изолировали из среды культивирования инфицированных клеток, очищали посредством ультрацентрифугирования в градиенте плотности и визуализировали посредством ЭМ анализа с отрицательным окрашиванием (Фигура 5В). Чтобы гарантировать, что визуализированные ВПЧ были действительно вирусами гриппа, частицы иммунологически метили золотом с применением антител, специфичных для НА вируса гриппа (Фигура 5С).
В совокупности указанные данные служили надежным доказательством того, что мультилокусный подход можно успешно применять для экспрессии рекомбинантных белков, и что на практике он может служить для получения такого рекомбинантного вируса посредством двух циклов встраивания системы двойного селектриуемого маркера. Другие исследовали активно изучали ВПЧ вируса гриппа и показали, что он обладает иммуногенностью у мышей и хорьков. Этот способ конструирования ВПЧ имеет то преимущество, что он позволяет получить геном бакуловируса с M1, уже заранее интегрированным и готовым к экспрессии. Таким образом, для получения ВПЧ необходимо провести только один цикл рекомбинации для добавления гена НА из любого активно формирующегося подтипа вируса гриппа, который требуется для производства вакцины. Для вакцин против гриппа, основанных на ВПЧ вируса гриппа, способ может упростить необходимое клонирование и потенциально увеличить скорость, с которой можно получить новые типы ВПЧ.
ВПЛ BTV1
В отличие от вируса гриппа, у которого ВПЧ можно сформировать путем экспрессии всего двух белков, ВПЧ BTV требует координированной экспрессии четырех структурных белков (VP2, VP3, VP5 и VP7). Чтобы лучше продемонстрировать применимость новой системы для рекомбинации, получали вирусы, которые экспрессировали каждый белок отдельно и комбинацию всех четырех структурных белков BTV (Фигура 6). Одно из преимуществ новой системы состоит в том, что вирусы, экспрессирующие отдельные белки и несколько белков, можно получить из одного и того же набора векторов переноса. Это облегчает получение контрольных вирусов, экспрессирующих отдельные белки, которые служат маркерами положения белков, экспрессируемых одновременно в полном комплексе. Для ВПЧ BTV применяли набор локусов, отличный от набора локусов, применяемых в примере с вирусом гриппа (Фигура 6С). Кроме того, новую систему сочетали с бакмидой с выключенным orfl629, как описано у Zhao el al., (12), чтобы можно было применять традиционный вектор переноса в локусе полиэдрина для экспрессии одной из субъединиц комплекса. Образование ВПЧ подтверждали по морфологии частиц при ЭМ с отрицательной окраской в сочетании с иммунным окрашиванием золотом одного из белков наружного капсида (VP5) (Фигура 6D).
ССТ мыши
Применение настоящего изобретения не ограничивается образованием вирусоподобных частиц. На самом деле, поскольку многие белки присутствуют в клетках в форме комплексов из более чем одной субъединицы, эффективное формирование указанных комплексов может найти применение в целом ряде областей. Чтобы продемонстрировать применимость указанной системы в других исследованиях, выбрали комплекс шаперонов ССТ. Данный комплекс вовлечен в исследования рака и вследствие того, что он состоит из 8 субъединиц, представляет собой значительную проблему в плане экспрессии белков. Действительно, не было возможности экспрессировать данный комплекс в системе па основе бакуловируса в отсутствии векторов и способов, предложенных в настоящем изобретении. Были сконструированы восемь векторов переноса, каждый из которых экспрессирует одну из субъединиц ССТ мыши и нацелен на отличный локус, идентифицированный в Примере 3 выше. Их применяли для генерирования рекомбинантного вируса, экспрессирующего каждую субъединицу в отдельности и сочетания субъединиц. Для одной из субъединиц (ССТ5) отметили нетипичный фенотип, когда в клетке была гиперэкспрессия указанного белка. В клетке отмечали кристаллические иглы и, в конце инфицирования, когда клетки разрывались, в среде культивирования отмечали четырехугольные пластинки кристаллического вещества (Фигура 7). Для данного белка создали схему очистки, и ее сопровождали условиями для предварительной кристаллизации и веществом инфицированных клеток для сотрудника, реализующего часть проекта, связанного с ССТ.
Хотя экспрессия некоторых других субъединиц ССТ приводит к образованию видимых агрегатов в клетках в конце инфицирования, экспрессия ни одного из белков не приводила к образованию агрегатов с таким регулярным видом. Все 8 субъединиц ССТ проявляли высокий уровень экспрессии (Фигура 8А) и давали перекрестную реакцию с поликлональной антисывороткой, полученной на ССТ мышей. Имела место некоторая перекрестная реакция между одной из эндогенных для клетки насекомых субъединиц ССТ и поликлональным антителом (Фигура 8В, дорожка 9). Однако данный эндогенный сигнал был связан только с 3 субъединицами (ССТ1, ССТ5 и ССТ6), которые накапливались в инфицированных клетках в большом количестве и, следовательно, его было можно четко определить. Все остальные субъединицы мигрировали как белки с незначительно различающейся молекулярной массой, и, таким образом, можно было отделить от белков клеток насекомых на основании миграции.
ВЫВОДЫ
Для всех трех комплексов белков имели место явные различия в аккумуляции одного и того же рекомбинантного белка при экспрессии его одного или в присутствии других белков. Почти во всех случаях имело место снижение уровня экспрессии белка, когда с того же вируса экспрессировали несколько белков. В какой-то степени этого можно было ожидать. Принимая во внимание конкуренцию за белки, необходимые для транскрипции, процессинга РНК, трансляции и скручивания белка, можно предсказать, что два гена бакуловируса с высоким уровнем экспрессии могут демонстрировать более низкую экспрессию при работе вместе, чем при работе по отдельности. Однако, что было неожиданно как для примера с ВПЧ BTV, так и для примера с ССТ, так это то, что снижение экспрессии белка варьировало для разных генетических локусов. Например, в случае BTV, VP2, VP5, VP3 и VP7 вес приводили к значительной и сходной аккумуляции белка, когда они экспрессировались со своего собственного локуса. Однако при их сочетании в одном бакуловирусс, экспрессирующем все четыре белка, VP2 и VP3 накапливались в меньшей степени, чем VP5 и VP7 (Фигура 6А). Данный эффект нельзя объяснить эффектом положения при геномной интеграции, поскольку и в одиночных, и в «учетверенных» вирусах гены экспрессировались с одинаковых генетических локусов. Одним из возможных объяснений может быть то, что данный эффект был связан с разными промоторами, которые применяли для разных генов. И VP5, и VP7 экспрессировались под контролем промотора р10, а VP2 и VP3 - под контролем промотора полиэдрина. Это может согласовываться с одним сообщением в литературе, согласно которому делеция гена р10 приводила к повышению экспрессии с локуса полиэдрина 13. Таким образом, можно было бы предсказать, что дупликация промотора полдиэдрина в присутствии двух промоторов р10 будет снижать экспрессию с промотора полиэдрина запускаемых генов. Однако это не может в полной мере объяснять эффект, наблюдаемый в случае комплекса ССТ. И ССТ5, и ССТ2 экспрессировались с промотора р10 и давали высокий стационарный уровень рекомбинантного белка, когда экспрессировались по одному (Фигура 8А и В, дорожки 2 и 5). Однако при сочетании их в одном вирусе экспрессия ССТ5 значительно снижалась по сравнению с ССТ2 (Фигура 8А и В, дорожка 10). На основании этих данных можно предположить, что другая цис-действующая последовательность, присутствующая в локусе р10, может вносить вклад в повышение относительной экспрессии ССТ2 и BTV VP5, которые оба были встроены в данный локус. Относительно низкая кажущаяся экспрессия НА вируса гриппа при встраивании его гена в тот же сайт может быть в большей степени связана с круговоротом данного белка в ЭПР, чем с транскрипционными эффектами. И ССТ2, и VP5 накапливаются в цитоплазме.
Изобретатели повысили эффективность системы ЕТ рекомбинации применительно к бакуловирусу до такой степени, что ее можно применять для стандартного встраивания кассет экспрессии для рекомбинантных белков. Кроме того, они идентифицировали семь генетических локусов (ctx, egt, 39k, orf51, gp37, iap2 и odv-e56), которые можно применять для интенсивной экспрессии белков и продемонстрировали, что комплексы из нескольких белков можно собирать при помощи данной системы, приведя три примера (ВПЧ вируса гриппа А и BTV, и комплекс ССТ).
Все документы, цитируемые выше, включены в настоящую заявку посредством ссылок.
Цитируемая литература
1. Albert, H., Dale, E. С, Lee, E. & Ow, D. W. Plant J 7, 649-59. (1995).
2. Lee, E. C et al. Genomics 73, 56-65 (2001).
3. Eldridge, R., Li, Y. & Miller, L. K. J Virol 66, 6563-71 (1992).
4. Flipsen, J. Т., Mans, R. M., Kleefsman, A. W., Knebel-Morsdorf, D. & Vlak, J. M. J Virol 69, 4529-32(1995).
5. O'Reilly, D. R. & Miller, L. K. Science 245, 1110-2 (1989).
6. Griffiths, C. M. et al. J Gen Virol 80 (Pt 4), 1055-66 (1999).
7. Braunagel, S. C, Elton, D. M., Ma, H. & Summers, M. D. Virology 217, 97-110 (1996).
8. Berger, I., Fitzgerald, D. J. & Richmond, T. J. Nat Biotechnol 22, 1583-7 (2004).
9. Fitzgerald, D. J. et al. Nat Methods 3, 1021-32 (2006).
10. McCarthy, С. В., Dai, X., Donly, С & Theilmann, D. A. Virology 372, 325-39 (2008).
11. McCarthy, С. В. & Theilmann, D. A. Virology 375, 277-91 (2008).
12. Zhao, Y., Chapman, D. A. & Jones, I. M. Nucleic Acids Res 31, E6-6. (2003).
13. Chaabihi, H. et al. J Virol 67, 2664-71 (1993).
14. Summers, M. D. Adv Virus Res 68, 3-73 (2006).
15. Emery, V. С & Bishop, D. H. Protein Eng 1, 359-66 (1987).
16. French, T. J., Marshall, J. J. & Roy, P. J Virol 64, 5695-700 (1990).
17. Latham, T. & Galarza, J. M. J Virol 75, 6154-65. (2001).
18. Pushko, P. et al. Vaccine 19, 142-53. (2000).
19. Ye, L. et al. Virology 351, 260-70 (2006).
20. Berger, I., Fitzgerald, D. J. & Richmond, T. J. Nat Biotechnol 22, 1583-7 (2004).
21. Fitzgerald, D. J. et al. Nat Methods 3, 1021-32 (2006).
22. French, T. J. & Roy, P. J Virol 64, 1530-6 (1990).
23. Devos, D. & Russell, R. B. Curr Opin Struct Biol 17, 370-7 (2007).
24. O'Neal, C M., Clements, J. D., Estes, M. K. & Conner, M. E. J Virol 72, 3390-3 (1998).
25. Palomares, L. A., Lopez, S. & Ramirez, О. Т. Biotechnol Bioeng 78, 635-44. (2002).
26. Mena, J. A., Ramirez, O. T. & Palomares, L. A. BMC Biotechnol 7, 39 (2007).
27. Weyer, U. & Possee, R. D. J Gen Virol 72 (Pt 12), 2967-74 (1991).
28. Bertolotti-Ciarlet, A., Ciarlet, M., Crawford, S. E., Conner, M. E. & Estes, M. K. Vaccine 21, 3885-900(2003).
29. Kamita, S. G., Maeda, S. & Hammock, B. D. J Virol 77, 13053-61 (2003).
30. Crouch, E. A. & Passarelli, A. L. J Virol 76, 9323-34 (2002).
31. Mikhailov, V. S., Okano, K. & Rohrmarm, G. F. J Virol 77, 2436-44 (2003).
32. Pijlman, G. P. et al. J Virol 76, 5605-11 (2002).
33. Pijlman, G. P., van den Born, E., Martens, D. E. & Vlak, J. M. Virology 283, 132-8 (2001).
34. Pijlman, G. P., van Schijndel, J. E. & Vlak, J. M. J Gen Virol 84, 2669-78 (2003).
35. Vanarsdall, A. L., Pearson, M. N. & Rohrmarm, G. F. Virology 367, 187-95 (2007).
36. Vanarsdall, A. L., Mikhailov, V. S. & Rohrmann, G. F. Virology 364, 475-85 (2007).
37. Okano, K., Vanarsdall, A. L. & Rohrmann, G. F. Virology 359, 46-54 (2007).
38. Vanarsdall, A. L., Okano, K. & Rohrmann, G. F. J Virol 80, 1724-33 (2006).
39. Okano, K., Vanarsdall, A. L. & Rohrmann, G. F. J Virol 78, 10650-6 (2004).
40. Vanarsdall, A. L., Okano, K. & Rohrmann, G. F. Virology 326, 191-201 (2004).
41. Fang, M., Dai, X. & Theilmann, D. A. J Virol 81, 9859-69 (2007).
42. Wang, Y. et al. Virology 367, 71-81 (2007).
43. Yamagishi, J., Burnett, E. D., Harwood, S. H. & Blissard, G. W. Virology 365, 34-47 (2007).
44. Xi, Q., Wang, J., Deng, R. & Wang, X. Virus Genes 34, 223-32 (2007).
45. Li, Y. et al. Virus Genes 31, 275-84 (2005).
46. Stewart, T. M., Huijskens, I., Willis, L. G. & Theilmann, D. A. J Virol 79. 4619-29 (2005).
47. Kamita, S. G. et al. Proc Natl Acad Sci USA 102, 2584-9 (2005).
48. Milks, M. L., Washburn, J. O., Willis, L. G., Volkman, L. E. & Theilmann, D. A. Virology 310, 224-34(2003).
49. Zhao, Y., Chapman, D. A. & Jones, I. M. Nucleic Acids Res 31, E6-6 (2003).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУСТАБИЛЬНОЕ ПОЛУЧЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2011 |
|
RU2577982C2 |
| НУКЛЕАЗА PaCas9 | 2018 |
|
RU2706298C1 |
| СТАБИЛЬНОЕ ПОЛУЧЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2011 |
|
RU2577979C2 |
| Лентивирусная плазмида (варианты), способ ее получения (варианты), набор праймеров для получения лентивирусного плазмидного вектора (варианты) | 2018 |
|
RU2680537C1 |
| НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ С ДЕФИЦИТОМ ДПНРНК | 2014 |
|
RU2721855C2 |
| АНТИГЕННЫЕ ПЕПТИДЫ NEISSERIA | 2000 |
|
RU2281956C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИВАЛЕНТНОЙ ВАКЦИНЫ ОТ ГРИППА | 2018 |
|
RU2701953C1 |
| ПОЛИВАЛЕНТНАЯ ВАКЦИНА ПРОТИВ ГРИППА | 2018 |
|
RU2706191C1 |
| Кассета, предназначенная для получения плазмидных векторов, используемых для создания клеток-продуцентов вирусоподобных частиц (ВПЧ) вируса гриппа | 2018 |
|
RU2680703C1 |
| РЕКОМБИНАНТНЫЙ ГЕРПЕСВИРУС КОИ (KHV) И ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЯ, ВЫЗЫВАЕМОГО KHV | 2012 |
|
RU2662768C2 |




Изобретение относится к области генетической инженерии. Предложен вектор переноса для встраивания гена в генетический локус последовательности бакуловируса, содержащий кассету экспрессии, содержащую эукариотический промотор, функционально связанный с указанным геном, удаляемую кассету двойной селекции, которая фланкирована последовательностями рекомбинации LoxP, причем указанные последовательности LoxP модифицированы, чтобы обеспечить прохождение только одного цикла рекомбинации, содержащую экспрессируемую последовательность, кодирующую первый селектируемый маркер, экспрессируемую последовательность, кодирующую второй селектируемый маркер, последовательности, фланкирующие указанные кассету экспрессии и кассету двойной селекции, причем указанные последовательности в существенной степени соответствуют последовательностям указанного генетического локуса в последовательности бакуловируса, что обеспечивает гомологическую рекомбинацию между указанным вектором переноса и указанным бакуловирусом. Также предложены способ получения рекомбинантной бакмиды, способ получения рекомбинантного бакуловируса, рекомбинантная бакмида, рекомбинантный бакуловирус, клетка и способ получения белка. Изобретение позволяет увеличить эффективность экспрессии рекомбинантных белков. 7 н. и 12 з.п. ф-лы, 8 ил., 1 табл., 4 пр.
1. Вектор переноса для встраивания гена в генетический локус последовательности бакуловируса, содержащий:
кассету экспрессии, содержащую эукариотический промотор, функционально связанный с указанным геном;
удаляемую кассету двойной селекции, которая фланкирована последовательностями рекомбинации LoxP, причем указанные последовательности LoxP модифицированы, чтобы обеспечить прохождение только одного цикла рекомбинации, содержащую:
(i) экспрессируемую последовательность, кодирующую первый селектируемый маркер; и
(ii) экспрессируемую последовательность, кодирующую второй селектируемый маркер; и
последовательности, фланкирующие указанные кассету экспрессии и кассету двойной селекции, причем указанные последовательности в существенной степени соответствуют последовательностям указанного генетического локуса в последовательности бакуловируса, что обеспечивает гомологическую рекомбинацию между указанным вектором переноса и указанным бакуловирусом.
2. Вектор переноса по п. 1, отличающийся тем, что указанный первый селектируемый маркер представляет собой визуальный маркер.
3. Вектор переноса по п. 2, отличающийся тем, что указанный первый маркер представляет собой фрагмент LacZalpha.
4. Вектор переноса по п. 1, отличающийся тем, что указанный второй селектируемый маркер придает устойчивость к антибиотику.
5. Вектор переноса по п. 4, отличающийся тем, что указанный ген устойчивости к антибиотику придает устойчивость к флеомицину.
6. Вектор переноса по любому из пп. 1-5, отличающийся тем, что указанные последовательности, фланкирующие кассеты экспрессии и двойной селекции в существенной степени соответствуют одному из следующих генетических локусов: ctx, egt, 39k, orf51, gp37, iap2, odv-e56 и p10.
7. Способ получения рекомбинантной бакмиды, включающий:
а) объединение бакмиды и вектора переноса, соответствующего любому из пп. 1-6, для обеспечения прохождения гомологичной рекомбинации; и
б) селекцию рекомбинантной бакмиды, которая содержит указанные кассету экспрессии и кассету двойной селекции указанного вектора переноса по п. 1, и необязательно, повторение стадий а) и б) некоторое число раз с получением бакмиды, которая содержит соответствующее число генов.
8. Способ по п. 7, отличающийся тем, что указанный вектор переноса представляет собой вектор переноса по п. 1, дополнительно включающий осуществление рекомбинации между последовательностями LoxP с удалением кассеты селекции из бакмиды.
9. Способ получения рекомбинантного бакуловируса, включающий получение рекомбинантной бакмиды способом по п. 7 или 8 и культивирование эукариотической клетки, содержащей указанную бакмиду с получением бакуловируса.
10. Рекомбинантная бакмида для экспрессии множества гетерологичных белков, которые взаимодействуют с образованием комплекса белков, полученная способом по п. 7, которая содержит кассету экспрессии и кассету двойной селекции указанного вектора переноса по п. 1.
11. Рекомбинантная бакмида по п. 10, отличающаяся тем, что указанный комплекс белков представляет собой вирусоподобную частицу или комплекс шаперонов.
12. Рекомбинантная бакмида по п. 10, которая экспрессирует совокупность белков, характеризующаяся тем, что каждый белок экспрессируется с отдельного генетического локуса указанной бакмиды.
13. Рекомбинантная бакмида по п. 12, отличающаяся тем, что отдельные генетические локусы выбраны из следующих: ctx, egt, 39k, orf51, gp37, iap2 и odv-e56.
14. Рекомбинантный бакуловирус для экспрессии множества гетерологичных белков, которые взаимодействуют с образованием комплекса белков, полученный способом по п. 9, при получении рекомбинантной бакмиды способом по п. 7, содержащий кассету экспрессии и кассету двойной селекции указанного вектора переноса по п. 1.
15. Рекомбинантный бакуловирус по п. 14, отличающийся тем, что указанный комплекс белков представляет собой вирусоподобную частицу или комплекс шаперонов.
16. Рекомбинантный бакуловирус по п. 14, который экспрессирует совокупность белков, характеризующийся тем, что каждый белок экспрессируется с отдельного генетического локуса указанного бакуловируса.
17. Рекомбинантный бакуловирус по п. 16, отличающийся тем, что отдельные генетические локусы выбраны из следующих: ctx, egt, 39k, orf51, gp37, iap2 и odv-e56.
18. Клетка для получения по меньшей мере одного белка, содержащая вектор переноса по любому из пп. 1-6, бакмиду для получения по меньшей мере одного белка по любому из пп. 10, 11, 12 или 13, или бакуловирус для получения по меньшей мере одного белка по любому из пп. 14, 15, 16 или 17.
19. Способ получения одного или более белков, включающий культивирование эукариотической клетки, которая трансфецирована рекомбинантной бакмидой по любому из пп. 10, 11, 12 или 13, или бакуловирусом по любому из пп. 14, 15, 16 или 17 при условиях, которые делают возможной продукцию указанных белков.
| WO 2007054250 A1, 18.05.2007 | |||
| WO03074714 A1, 12.09.2003 | |||
| US 0006461863 B1, 08.10.2002 | |||
| ПОЛИПЕПТИД, СПОСОБ ПОЛУЧЕНИЯ (ВАРИАНТЫ), ДНК (ВАРИАНТЫ), ВЕКТОР (ВАРИАНТЫ), КЛЕТКА, ПРИМЕНЕНИЕ ПОЛИПЕПТИДА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) | 1993 |
|
RU2177480C2 |
| ОПОСРЕДОВАННАЯ РЕЦЕПТОРАМИ ДОСТАВКА ГЕНОВ С ИСПОЛЬЗОВАНИЕМ ВЕКТОРОВ НА ОСНОВЕ БАКТЕРИОФАГОВ | 1998 |
|
RU2209088C2 |

