Настоящее изобретение заявляет преимущество временной заявки США, серийный № 61/247518, поданной 30 сентября 2009 года, которая включена в настоящую заявку посредством ссылки.
Область изобретения
Настоящее изобретение относится к области конъюгирования бактериальных капсульных сахаридов, в частности, капсульных полисахаридов Staphylococcus aureus типа 5 или типа 8, с носителями для образования гликоконъюгатов. Гликоконъюгаты являются полезными для иммунизации.
Предпосылки изобретения
Капсульные сахариды бактерий используют в течение многих лет в вакцинах против капсулированных бактерий. Поскольку сахариды являются T-независимыми антигенами, они, однако, являются низко иммуногенными. Конъюгация с носителем может преобразовать T-независимые антигены в T-зависимые антигены, усиливая, таким образом, вторичные иммунные ответы и обеспечивая возможность развития защитного иммунитета. Наиболее эффективные сахаридные вакцины, поэтому, основаны на гликоконъюгатах, и прототип конъюгат-вакцины был против Haemophilus influenzae типа b ('Hib') [например, см. главу 14 ссылочного документа 97].
Другая бактерия, для которой были описаны конъюгат-вакцины, представляет собой Staphylococcus aureus (S.aureus). Различные полисахариды были выделены из S.aureus для использования в гликоконъюгатах. Два полисахарида, которые представляют особый интерес, это капсульные полисахариды типа 5 и типа 8. Приблизительно 60% человеческих штаммов S.aureus представляют собой тип 8, и приблизительно 30% представляют собой тип 5. Большая работа с конъюгатами типа 5 и типа 8 была выполнена Fattom et al., и это описано в документах, таких как ссылочные документы 1-9. Способ Fattom для конъюгации типа 5 и типа 8 полисахаридов типично включает тиолирование очищенного полисахарида с использованием цистамина. Реакция основана на присутствии карбоксилатных групп в капсульном полисахариде. Эти группы взаимодействуют с цистамином в присутствии карбодиимида, например, EDAC. Дериватизированный полисахарид затем конъюгируют с белком-носителем, таким как Pseudomononas aeruginosa эндотоксин A (ETA), типично через линкер [2]. Другие исследователи осуществляли конъюгацию очищенных капсульных полисахаридов типа 5 и типа 8 путем восстановительного аминирования [10 и 11]; глутаральдегидного связывания [10]; или взаимодействия гидроксильных групп полисахаридов с агентами цианилирования, такими как CDAP [12] или трихлорид циануровой кислоты [13].
Хотя было показано, что конъюгат-вакцины, полученные способом Fattom, являются безопасными и иммуногенными для человека [5], все еще остается необходимость в других и более лучших способах получения конъюгатов капсульных полисахаридов S.aureus типа 5 или типа 8.
Раскрытие изобретения
Изобретение основано на способе конъюгации, который можно использовать вместо способов конъюгации, раскрытых в предшествующем уровне техники. В отличие от этих способов, способ по настоящему изобретению не включает конъюгацию через гидроксильную или карбоксилатную группы в полисахариде. Способ поэтому оставляет эти группы в форме, которая ближе, чем известные из уровня техники, к форме, наблюдаемой в природном полисахариде. Вместо использования этих групп, способ включает образование реакционноспособных альдегидных групп в полисахариде для использования в конъюгации. Полученные конъюгаты могут иметь другие, предпочтительно улучшенные, иммунологические свойства по сравнению с конъюгатами предшествующего уровня техники.
Изобретение поэтому обеспечивает альтернативные или улучшенные способы конъюгирования капсульного полисахарида S.aureus типа 5 или типа 8 с белком-носителем и конъюгаты, полученные такими способами. Изобретение также обеспечивает промежуточные соединения, которые являются полезными в способах по настоящему изобретению, и способы получения этих промежуточных соединений.
В первом аспекте изобретение обеспечивает способ получения конъюгата капсульного полисахарида S.aureus типа 5 или типа 8 и молекулы носителя, включающий следующие стадии: (a) деполимеризация капсульного полисахарида с получением полисахаридного фрагмента; (b) окисление фрагмента для введения альдегидной группы в, по меньшей мере, один сахаридный остаток во фрагменте с получением окисленного сахаридного остатка; и (c) связывание окисленного сахаридного остатка с молекулой носителя через альдегидную группу с получением, таким образом, конъюгата. Связывание на стадии (c) может быть прямым, или оно может осуществляться через линкерную молекулу. Изобретение также обеспечивает конъюгат, полученный или который можно получить этим способом.
Капсульный полисахарид
Изобретение основано на капсульных полисахаридах S.aureus типа 5 и типа 8. Структуры капсульных полисахаридов типа 5 и типа 8 были описаны в ссылочных документах 14 и 15 следующим образом:
Тип 5
→4)-β-D-ManNAcA(3OAc)-(1→4)-α-L-FucNAc(1→3)-β-D-FucNAc-(1→
Тип 8
→3)-β-D-ManNAcA(4OAc)-(1→3)-α-L-FucNAc(1→3)-β-D-FucNAc-(1→
Полученные в последнее время данные спектроскопии ЯМР [16] привели к пересмотру этих структур:
Тип 5
→4)-β-D-ManNAcA-(1→4)-α-L-FucNAc(3OAc)-(1→3)-β-D-FucNAc-(1→
Тип 8
→3)-β-D-ManNAcA(4OAc)-(1→3)-α-L-FucNAc(1→3)-α-D-FucNAc(1→
Полисахарид может быть химически модифицирован по сравнению с капсульным полисахаридом, присутствующим в природе.
Например, полисахарид может быть де-O-ацетилирован (частично или полностью), де-N-ацетилирован (частично или полностью), N-пропионирован (частично или полностью) и т.п. Деацетилирование может происходить до, в процессе или после конъюгации, но типично происходит до конъюгации. В зависимости от конкретного полисахарида, деацетилирование может влиять или не влиять на иммуногенность, например, для NeisVac-C™ вакцины используют де-O-ацетилированный полисахарид, тогда как Menjugate™ является ацетилированной, но обе вакцины являются эффективными. Эффект деацетилирования и т.п. можно оценить при помощи рутинных анализов. Например, влияние O-ацетилирования на капсульные полисахариды S.aureus типа 5 или типа 8 обсуждается в ссылочном документе 6. Природные полисахариды, как указано в этом документе, имеют 75% O-ацетилирования. Эти полисахариды индуцировали антитела как к полисахаридному остову молекулы, так и к O-ацетильным группам. Полисахариды с 0% O-ацетилирования тем не менее индуцировали антитела к полисахаридному остову молекулы. Оба типа антител были опсоническими против штаммов S.aureus, которые различались по содержанию O-ацетильных групп. Соответственно, капсульные полисахариды типа 5 или типа 8, используемые в настоящем изобретении, могут иметь O-ацетилирование от 0 до 100%. Например, степень O-ацетилирования капсульного полисахарида типа 5 может быть 10-100%, 10-100%, 20-100%, 30-100%, 40-100%, 50-100%, 60-100%, 70-100%, 80-100%, 90- 100%, 50-90%, 60-90%, 70-90% или 80-90%. Альтернативно, можно использовать капсульный полисахарид типа 5 с 0% O-ацетилирования. Подобным образом, степень O-ацетилирования капсульного полисахарида типа 8 может быть 10-100%, 10-100%, 20-100%, 30-100%, 40-100%, 50-100%, 60-100%, 70-100%, 80-100%, 90-100%, 50-90%, 60-90%, 70-90% или 80-90%. Альтернативно, можно использовать капсульный полисахарид типа 8 с 0% O-ацетилирования. В одном варианте воплощения степень O-ацетилирования капсульных полисахаридов типа 5 и типа 8 может быть 10-100%, 20-100%, 30-100%, 40-100%, 50-100%, 60-100%, 70-100%, 80-100%, 90-100%, 50-90%, 60-90%, 70-90% или 80-90%. В других вариантах воплощения используют капсульные полисахариды типа 5 и типа 8 с 0% O-ацетилирования. Степень N-ацетилирования капсульного полисахарида типа 5, используемого в настоящем изобретении, может быть 0-100%, 50-100%, 75-100%, 80-100%, 90-100% или 95-100%. Типично, степень N-ацетилирования капсульного полисахарида типа 5 составляет 100%. Подобным образом, степень N-ацетилирования капсульного полисахарида типа 8, используемого в настоящем изобретении, может быть 0-100%, 50-100%, 75-100%, 80-100%, 90-100% или 95-100%. Типично, степень N-ацетилирования капсульного полисахарида типа 8 составляет 100%. В одном варианте воплощения степень N-ацетилирования капсульных полисахаридов типа 5 и типа 8 может быть 0-100%, 50-100%, 75-100%, 80-100%, 90-100% или 95-100%. Типично, степень N-ацетилирования капсульного полисахарида типа 5 и типа 8 составляет 100%.
Степень O-ацетилирования полисахарида можно определить любым способом, известным из уровня техники, например, способом протонного ЯМР (например, как описано в ссылочных документах 17, 18, 19 или 20). Еще один способ описан в ссылочном документе 21. Подобные способы можно использовать для определения степени N-ацетилирования полисахарида. O-ацетильные группы можно удалить путем гидролиза, например, путем обработки основанием, таким как безводный гидразин [22] или NaOH [6]. Подобные способы можно использовать для удаления N-ацетильных групп. Для поддержания высоких уровней O-ацетилирования в капсульных полисахаридах типа 5 и/или 8 обработки, которые приводят к гидролизу O-ацетильных групп, сводятся к минимуму, например, обработки при предельных значениях pH.
Капсульные полисахариды можно очистить с использованием известных процедур, как описано в ссылочных документах, указанных в настоящей заявке. Типичный способ включает фенол-этанольную инактивацию клеток S.aureus, центрифугирование, обработку лизостафином, обработку РНКазой/ДНКазой, центрифугирование, диализ, обработку протеазой, дополнительный диализ, фильтрование, осаждение этанолом/CaCl2, диализ, лиофилизацию, анионообменную хроматографию, диализ, лиофилизацию, вытеснительную хроматографию с исключением по размеру, диализ и лиофилизацию [1]. Альтернативный способ включает обработку в автоклаве клеток S.aureus, ультрафильтрацию полисахаридсодержащего супернатанта, концентрирование, лиофилизацию, обработку метаперйодатом натрия для удаления тейхоевой кислоты, дополнительную ультрафильтрацию, диафильтрацию, высокоэффективную вытеснительную жидкостную хроматографию с исключением по размеру, диализ и лиофилизацию [23], предпочтительно используют способ очистки, описанный в ссылочном документе 24.
Изобретение, однако, не ограничивается полисахаридами, очищенными из природных источников, и полисахариды можно получить другими способами, такими как общий или частичный синтез.
Молекула носителя
Изобретение включает использование молекул носителей, которые типично представляют собой белки. Как правило, ковалентная конъюгация сахаридов с носителями повышает иммуногенность сахаридов, поскольку превращает их из T-независимых антигенов в T-зависимые антигены, обеспечивая, таким образом, возможность примирования для иммунологической памяти. Конъюгация особенно полезна для педиатрических вакцин [например, ссылочный документ 25] и является широко известным приемом [например, см. обзор в ссылочных документах 26-34].
Предпочтительными белками-носителями являются бактериальные токсины, так как дифтерийный или столбнячный токсины или токсоиды или их мутанты. Авторами настоящего изобретения было обнаружено, что мутантный CRM197 дифтерийный токсин [35] является подходящим. Pseudomonas aeruginosa экзотоксин А (ETA) и его нетоксичный мутантный рекомбинантный экзопротеин (rEPA) используют в качестве белков-носителей для капсульных полисахаридов S.aureus типа 5 или типа 8 ([1] и [2]). Также используют S.aureus α-гемолизин (α-токсин) ([10] и [36]), овальбумин [13] и человеческий сывороточный альбумин [11]. Эти носители можно использовать в настоящем изобретении.
Другие подходящие белки-носители включают белковый комплекс внешней мембраны N.meningitidis [37], синтетические пептиды [38, 39], белки теплового шока [40, 41], коклюшные белки [42, 43], цитокины [44], лимфокины [44], гормоны [44], факторы роста [44], человеческий сывороточный альбумин (типично рекомбинантный), искусственные белки, включающие множественные человеческие CD4+ T-клеточные эпитопы из различных происходящих из патогенов антигенов [45], такие как N19 [46], белок D из H.influenzae [47-49], пневмококковый поверхностный белок PspA [50], пневмолизин [51] или его нетоксичные производные [52], железосодержащие белки [53], токсин А или B из C.difflcile [54], GBS белок [55], GAS белок [56] и т.п.
Другие подходящие белки-носители включают S.aureus белковые антигены, например S.aureus белковые антигены, описанные ниже.
Можно использовать более одного белка-носителя, например, для снижения риска супрессии носителя. Таким образом, различные белки-носители можно использовать для капсульных полисахаридов типа 5 и типа 8, например, полисахарид типа 5 может быть конъюгирован с CRM197, тогда как полисахарид типа 8 может быть конъюгирован с rEPA. Также можно использовать более чем один белок-носитель для конкретного полисахаридного антигена, например, полисахарид типа 5 может быть в двух группах, где одна группа конъюгирована с CRM197, а другая конъюгирована с rEPA. Однако такой же белок-носитель типично используют для всех полисахаридов.
Один белок-носитель может нести более чем один полисахаридный антиген [57, 58]. Например, один белок-носитель может иметь конъюгированные с ним капсульные полисахариды типа 5 и типа 8. Для достижения этой цели можно смешать разные полисахариды до осуществления способа конъюгации. Типично, однако, когда имеются отдельные конъюгаты для каждого полисахарида, при этом различные полисахариды смешивают после конъюгации. Эти отдельные конъюгаты могут быть основаны на одном и том же носителе.
Деполимеризация
На стадии (a) способа по настоящему изобретению капсульный полисахарид подвергают деполимеризации с получением полисахаридного фрагмента. Сообщалось о деполимеризации капсульного полисахарида типа 8 путем обработки ультразвуком перед осуществлением конъюгации [3]. Авторами было сделано заключение, что низкомолекулярный тип 8 не был иммуногенным. Не смотря на то, что эти авторы, поэтому, отдавали предпочтение высокомолекулярным полисахаридам, в настоящем изобретении, к удивлению, используют полисахаридные фрагменты с более низкой молекулярной массой по сравнению с природными капсульными полисахаридами.
Полноразмерные полисахариды могут быть деполимеризованы с получением более коротких фрагментов для использования в изобретении различными способами. Авторами настоящего изобретения было обнаружено, что способы, которые приводят к расщеплению (1→3) гликозидных связей между α-L-FucNAc(3OAc) и β-D-FucNAc остатками в капсульном полисахариде типа 5, являются особенно подходящими. Когда эти способы применяют к капсульному полисахариду типа 5, они приводят к полисахаридному фрагменту, содержащему β-D-FucNAc-(1→часть на его нередуцирующем конце. Эта часть включает две вицинальные гидроксильные группы. Подобным образом, когда эти способы применяют к капсульному полисахариду типа 8, считается, что они приводят к полисахаридному фрагменту, содержащему α-D-FucNAc-(1→часть на его нередуцирующем конце, при этом указанная часть также включает две вицинальные гидроксильные группы. Вицинальные гидроксильные группы в полисахаридном фрагменте типа 5 или типа 8 обеспечивают осуществление последующей конъюгации фрагмента с молекулой носителя, как описано ниже.
Соответственно, в следующем аспекте изобретение обеспечивает способ обработки капсульного полисахарида S.aureus типа 5, включающий стадию деполимеризации капсульного полисахарида, с получением полисахаридного фрагмента, содержащего β-D-FucNAc-(1→часть на его нередуцирующем конце. В родственном аспекте изобретение обеспечивает способ обработки капсульного полисахарида S.aureus типа 8, включающий стадию деполимеризации капсульного полисахарида с получением полисахаридного фрагмента, содержащего α-D-FucNAc-(1→часть на его нередуцирующем конце. Капсульный полисахарид может представлять собой капсульный полисахарид S.aureus типа 5 или типа 8, как описано выше в разделе "Капсульный полисахарид". Изобретение также обеспечивает полисахаридный фрагмент, полученный или который можно получить любым из этих способов.
Авторами настоящего изобретения было обнаружено, что деполимеризацию можно осуществить при помощи кислотного гидролиза. Для кислотного гидролиза предпочтительно использовать мягкую кислоту, например уксусную кислоту, во избежание побочных реакций по другим группам в полисахариде. Специалист в данной области сможет определить подходящие кислоты и условия (например, концентрацию, температуру и/или время) для гидролиза. Например, авторами настоящего изобретения было обнаружено, что обработка полисахарида при 2 мг/мл 2% уксусной кислотой (об./об.) при 90°C в течение 3 часов является подходящей. Авторами настоящего изобретения также было обнаружено, что обработка, при 2 мг/мл 5% уксусной кислотой при 90°C в течение 30 минут, 5 или 6 часов является подходящей. Обработка другими кислотами, например, трифторуксусной или другими органическими кислотами, также может быть подходящей. В частности, авторами настоящего изобретения было обнаружено, что эффективность деполимеризации можно повысить, в частности, для капсульного полисахарида типа 8, с использованием хлористоводородной кислоты. Например, авторами настоящего изобретения было обнаружено, что обработка полисахарида 2M раствором хлористоводородной кислоты при 100°C в течение 30 минут является подходящей. Авторами настоящего изобретения также было обнаружено, что обработка 2M раствором хлористоводородной кислоты при 100°C в течение 1, 1,5, 2 или 2,5 часов является подходящей. Такая обработка хлористоводородной кислотой может привести к де-O-ацетилированию полисахарида, например, как описано ниже.
Другие способы деполимеризации полисахарида могут быть подходящими. Эти способы включают нагревание, микрофлюидизацию [59], обработку ультразвуком [3], окисление-восстановление [60] или озонолиз [61].
Полисахаридные фрагменты можно идентифицировать при помощи хроматографии, например, вытеснительной хроматографии с исключением по размеру. Специфические молекулярные массы можно измерить методом гель-фильтрации по сравнению с пуллулановыми стандартами, такими как поставляемые компанией Polymer Standard Service [62]. Типично, фрагмент по настоящему изобретению представляет собой смесь фрагментов с массами, находящимися в определенных пределах. Для деполимеризованного капсульного полисахарида типа 5 молекулярная масса фрагмента типично варьирует в пределах 1-500 кДа, например в пределах от 5 до 100 кДа, в частности от 10 до 50 кДа, и более конкретно от 20 до 30 кДа. Подобным образом, для деполимеризованного капсульного полисахарида типа 8 молекулярная масса фрагмента может варьировать в пределах 1-500 кДа, например в пределах от 5 до 100 кДа, в частности от 10 до 50 кДа, и более конкретно от 20 до 30 кДа. В некоторых вариантах воплощения низкомолекулярные типа 5 и/или типа 8 полисахаридные фрагменты выбраны для использования в изобретении. Например, фракции, полученные гель-фильтрацией, соответствующие низкомолекулярным фрагментам, могут быть выбраны и объединены. Низкомолекулярные полисахаридные фрагменты типично имеют молекулярную массу, которая варьирует в пределах от 5 до 20 кДа.
Деполимеризация может привести к изменению степени O-ацетилирования капсульного полисахарида. В качестве примера, авторами настоящего изобретения было обнаружено, что кислотный гидролиз может привести к снижению степени O-ацетилирования. В некоторых вариантах воплощения степень O-ацетилирования фрагмента может быть 10-90%, 20-70%, 30-50%, в частности, 35-45%. В других вариантах воплощения степень O-ацетилирования фрагмента может быть 0-10%, 0-5%, 0-2%, в частности 0%.
Введение альдегидной группы
На стадии (b) способа фрагмент подвергают окислению для введения альдегидной группы в, по меньшей мере, один сахаридный остаток во фрагменте. Эта стадия может включать введение более чем одной альдегидной группы в сахаридный остаток. В частности, можно ввести две альдегидные группы. Например, когда деполимеризация на стадии (a) приводит к полисахаридному фрагменту типа 5, содержащему β-D-FucNAc-(1→часть на его нередуцирующем конце, две вицинальные гидроксильные группы в этой части могут быть окислены для введения двух альдегидных групп в эту часть. Таким образом, β-D-FucNAc-(1→часть может представлять собой сахаридный остаток стадии (b). Подобным образом, когда деполимеризация приводит к полисахаридному фрагменту типа 8, содержащему α-D-FucNAc-(1→часть на его нередуцирующем конце, две вицинальные гидроксильные группы в этой части могут быть окислены для введения двух альдегидных групп. Таким образом, α-D-FucNAc-(1→часть может представлять собой сахаридный остаток стадии (b).
Соответственно, в следующем аспекте изобретение представляет способ обеспечения производного капсульного полисахарида S.aureus типа 5, включающий стадию окисления капсульного полисахарида S. aureus типа 5, содержащего β-D-FucNAc-(1→часть на его нередуцирующем конце, для преобразования двух вицинальных гидроксильных групп в β-D-FucNAc-O→части в две альдегидные группы. В родственном аспекте изобретение представляет способ обеспечения производного капсульного полисахарида S. aureus типа 8, включающий стадию окисления капсульного полисахарида S.aureus типа 8, содержащего α-D-FucNAc-(1→часть на его нередуцирующем конце, для преобразования двух вицинальных гидроксильных групп в α-D-Fuc Ac-(1→части в две альдегидные группы. Капсульный полисахарид может представлять собой полисахаридный фрагмент, описанный выше в разделе "Деполимеризация". Изобретение также обеспечивает производное капсульного полисахарида S.aureus, полученное или которое можно получить любым из этих способов.
Типичные реакции для получения альдегидов включают использование перйодатных солей и, в частности, мета-перйодатов (например, мета-перйодат натрия или калия, например, NaIO4), для окисления вицинальных гидроксильных групп [63]. Специалист в данной области сможет определить подходящие условия для окисления. Например, авторами настоящего изобретения было обнаружено, что обработка полисахарида при 2 мг/мл при помощи NaIO4 при 1:1 отношении (масс./масс.) при комнатной температуре в течение 1-2 часов в темноте является подходящей. Авторами настоящего изобретения также было обнаружено, что обработка полисахарида при 2 мг/мл при помощи 93 мМ NaIO4 при комнатной температуре в течение 8 часов в темноте является подходящей. Можно использовать другие условия окисления, например, с использованием тетроксида осмия и т.п.
Связывание с молекулой носителя
Связывание окисленного сахаридного остатка с молекулой носителя на стадии (c) способа можно осуществлять непосредственно или через линкер. Можно использовать любую подходящую реакцию конъюгации с любым подходящим линкером, если это желательно.
Когда окисление на стадии (b) приводит к полисахаридному фрагменту типа 5, содержащему β-D-FucNAc-(1→часть на его нередуцирующем конце, в котором две альдегидные группы были введены в эту часть, связывание на стадии (c) может осуществляться через одну из этих альдегидных групп. Таким образом, окисленная β-D-FucNAc-(1→часть может представлять собой окисленный сахаридный остаток стадии (c). Подобным образом, когда окисление приводит к полисахаридному фрагменту типа 8, содержащему α-D-FucNAc-(1→часть на его нередуцирующем конце, в котором две альдегидные группы были введены в эту часть, связывание на стадии (c) может осуществляться через одну из этих альдегидных групп, таким образом, окисленная α-D-FucNAc-(1→часть может представлять собой окисленный сахаридный остаток стадии (c).
Соответственно, в следующем аспекте изобретение представляет способ обеспечения конъюгированного капсульного полисахарида S.aureus типа 5, включающий стадию связывания с молекулой носителя капсульного полисахарида S.aureus типа 5, содержащего β-D-FucNAc-(1→часть на его нередуцирующем конце, который был окислен для преобразования двух вицинальных гидроксильных групп в две альдегидные группы, где связывание осуществляется через одну из этих альдегидных групп. В родственном аспекте, изобретение представляет способ обеспечения конъюгированного капсульного полисахарида S.aureus типа 8, включающий стадию связывания с молекулой носителя капсульного полисахарида S.aureus типа 8, содержащего α-D-FucNAc-(1→часть на его нередуцирующем конце, который был окислен для преобразования двух вицинальных гидроксильных групп в две альдегидные группы, где связывание осуществляется через одну из этих альдегидных групп. Капсульный полисахарид может представлять собой капсульный полисахарид, описанный выше в разделе "Введение альдегидной группы". Молекула носителя может представлять собой носитель, описанный выше в разделе "Молекула носителя". Изобретение также обеспечивает конъюгированный капсульный полисахарид, полученный или который можно получить любым из этих способов.
Присоединение окисленного сахаридного остатка или линкера к носителю типично происходит через группу амина (-NH2), например, в боковой цепи лизина или остатка в белке-носителе, или аргининового остатка. Присоединение к носителю также может осуществляться через сульфгидрильную (-SH) группу, например, в боковой цепи цистеинового остатка. Авторами настоящего изобретения было обнаружено, что прямое связывание можно удобным образом осуществить путем взаимодействия альдегидной группы в окисленном сахаридном остатке с группой амина в носителе путем восстановительного аминирования. Поэтому прямое связывание такого характера является предпочтительным в настоящем изобретении. В отличие от этого, ссылочный документ 2 предполагает, что линкеры могут быть предпочтительны в конъюгатах S.aureus типа 5 и 8. Если это желательно, связывание через линкер можно использовать в настоящем изобретении, например, путем взаимодействия альдегидной группы в окисленном сахаридном остатке с группой амина в линкере путем восстановительного аминирования, или путем преобразования альдегидной группы в группу амина путем восстановительного аминирования с получением группы амина для присоединения линкера.
Восстановительное аминирование представляет собой стандартную процедуру в органической химии, и его широко используют в получении конъюгатов капсульных полисахаридов для использования в вакцинах, включая капсульные полисахариды S.aureus [10]. В одном варианте воплощения альдегидная группа в окисленном сахаридном остатке взаимодействует с группой амина в носителе или линкере. Это можно удобным образом осуществить путем сочетания полисахарида с носителем или линкером в присутствии подходящего восстановителя (например, цианоборогидридов, таких как цианоборогидрид натрия NaBH3CN; боран-пиридина; триацетоксиборогидрида натрия; борогидридной обменной смолы; и т.п.). В другом варианте воплощения альдегидную группу преобразовывают в группу амина путем восстановительного аминирования с получением группы амина для присоединения линкера. Восстановительное аминирование включает либо аммиак, либо первичный амин (NH2R). Это можно удобным образом осуществить с использованием аммониевой соли (например, хлорида аммония) в сочетании с подходящим восстановителем (например, указанным выше). Специалист в данной области сможет определить подходящие условия для восстановительного аминирования. Например, авторами настоящего изобретения было обнаружено, что обработка полисахарида при 10 мг/мл белком-носителем при отношении 4:1 полисахарид:белок (масс./масс.) и NaBH3CN при отношении 2:1 полисахарид:NaBH3CN является подходящей.
Связывание через линкерную группу можно осуществить с использованием любой известной процедуры, например процедур восстановительного аминирования, описанных выше. В одном варианте воплощения можно использовать бифункциональный линкер с получением первой группы для связывания с альдегидной группой в окисленном сахаридном остатке, и второй группы для связывания с носителем. Например, можно использовать бифункциональный линкер формулы X1-L-X2, где X1 может взаимодействовать с альдегидом; X2 может взаимодействовать с носителем; и L представляет собой связывающую группу в линкере. Типичная X1 группа представляет собой группу амина. Типичные L группы представляют собой алкильные группы с прямой цепью, содержащие от 1 до 10 атомов углерода (например, C1, C2, C3, C4, C5, C6, C7, C8, C9, С10), например -(CH2)4- или -(CH2)3-. В другом варианте воплощения можно использовать бифункциональный линкер с получением первой группы для связывания с группой амина, образованной из альдегидной группы, в окисленном сахаридном остатке (например, путем восстановительного аминирования, как описано выше), и второй группы для связывания с носителем (типично для связывания с амином в носителе). Например, можно использовать гомобифункциональный линкер формулы X-L-X, где две X группы являются одинаковыми и могут взаимодействовать с аминами; и где L представляет собой связывающую группу в линкере. Типичная X группа представляет собой N-оксисукцинимид. L типично имеет формулу -L'-L2-L'-, где L' представляет собой карбонил. Типичные L2 группы представляют собой алкильные группы с прямой цепью, содержащие от 1 до 10 атомов углерода (например, C1, C2, C3, C4, Cs, C6, C7, C8, C9, C10), например -(CH2)4-. Типичный линкер, таким образом, представляет собой N-гидроксисукцинимидный диэфир адипиновой кислоты (SIDEA):

Другие X группы представляют собой такие, которые образуют сложные эфиры при сочетании с HO-L-OH, такие как норборан, п-нитробензойная кислота и сульфо-N-гидроксисукцинимид. Следующие бифункциональные линкеры, которые являются реакционно-способными с аминами, для использования с настоящим изобретением включают акрилоилгалогениды (например, хлорид) [65], галогенацилгалогениды [66], дисукцинимидилглутарат, дисукцинимидилсуберат, этиленгликоль бис[сукцинимидилсукцинат] и т.п.
Линкер, как правило, добавляют в молярном избытке к полисахариду. Взаимодействие линкер/полисахарид, как правило, осуществляют в апротонном растворителе (например, DMSO, этилацетат и т.п.), поскольку линкеры обычно нерастворимы в воде. Однако когда используют водорастворимые линкеры, тогда подходящими являются большое количество различных растворителей, включая протонные растворители, такие как вода. Подходящие линкеры включают сульфонированные формы, такие как сульфонированные SIDEA:
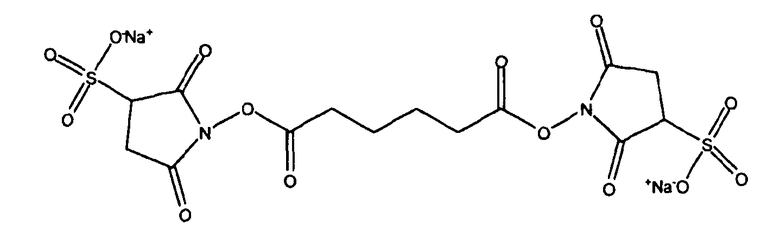
Когда используют линкер, конъюгат будет включать линкерную группу. Эта группа не образуется ни в полисахариде, ни в носителе, но представляет собой третью молекулу, используемую в процессе получения конъюгата, и ее легко отличить как от полисахарида, так и белка-носителя в конечном конъюгированном продукте. Линкерная группа может включать атомы, такие как углерод, водород, кислород и/или азот. Линкеры, которые включают углерод и водород, являются типичными, и также типично используют линкеры, которые дополнительно включают кислород и/или азот. Линкеры, которые включают атомы азота, могут включать атом углерода, связанный с атомом азота, который, в свою очередь, связан со вторым атомом углерода (-C-N-C-). Линкеры, которые включают атом кислорода, типично включают его как часть карбонильной группы. Линкерные группы с молекулярной массой в пределах 30-500 Да являются типичными. Линкеры, содержащие две карбонильные группы, также являются типичными.
Особенно полезная линкерная группа представляет собой -NH-C(О)-(CH2)n-C(О)-, где n имеет значение 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. Типично значение n равно 4. Концевая группа -NH- в этом линкере обычно связана с атомом углерода из полисахаридной части. Концевая группа -C(O)- обычно связана с атомом азота в аминокислотной боковой цепи в носителе. Предпочтительную линкерную группу удобным образом можно ввести способом, включающим: восстановительное аминирование альдегида в окисленном сахаридном остатке; взаимодействие полученной -NH2 группы с бифункциональным линкером, который представляет собой диэфир (например, дисукцинимидиловый эфир) диовой кислоты (например, адипиновой кислоты, HOOC-(CH2)4-COOH); и восстановительное аминирование продукта (см. Фиг. 6 [64]).
Другие химические приемы, которые можно использовать для присоединения линкера к -NH2 группе в полисахариде, включают:
- акрилоилирование (например, путем взаимодействия с акрилоилхлоридом), с последующим присоединением по методу Михаэля либо к ε-ΝΗ2 аминокислотной боковой цепи, либо к -SH цистеиновой боковой цепи [65]. Полученный линкер представляет собой -NH-C(О)-(CH2)2- (пропионамидо).
- взаимодействие с галогенацилгалогенидом, с последующим взаимодействием с ε-ΝΗ2 аминокислотной боковой цепи или с -SH цистеиновой боковой цепи [66]. Линкер представляет собой -NH-C(О)-CH2-.
Конъюгаты с отношением полисахарид:белок (масс./масс.) в пределах от 1:20 (т.е. избыток белка) до 20:1 (т.е. избыток полисахарида) типично получают способом по настоящему изобретению. Отношения в пределах от 1:10 до 1:1 являются предпочтительными, в частности отношения в пределах от 1:5 до 1:2, и наиболее предпочтительно около 1:3. В отличие от этого, конъюгаты капсульного полисахарида типа 5 и типа 8, полученные способами предшествующего уровня техники, имеют тенденцию к более высоким отношениям, например в пределах от 0,73:1 до 1,08:1 в ссылочных документах 1, 2 и 3. В конкретных вариантах воплощения настоящего изобретения отношение полисахарид:белок (масс./масс.) для капсульного полисахаридного конъюгата типа 5 находится в пределах от 1:10 до 1:2; и/или отношение полисахарид:белок (масс./масс.) для капсульного полисахаридного конъюгата типа 8 находится в пределах от 1:5 до 7:10.
Композиции могут включать небольшое количество свободного носителя [67]. Когда данный белок-носитель присутствует как в свободной, так и конъюгированной форме в композиции по настоящему изобретению, неконъюгированная форма предпочтительно составляет не более чем 5% от общего количества белка-носителя в композиции в целом, и более предпочтительно присутствует в количестве менее чем 2% масс.
После конъюгации свободные и конъюгированные полисахариды можно разделить. Существует множество подходящих способов, включая гидрофобную хроматографию, тангенциальную ультрафильтрацию, диафильтрацию и т.п. [см. также ссылочные документы 68 и 69 и т.п.].
Сочетания конъюгатов и других антигенов
Помимо обеспечения индивидуальных конъюгатов, описанных выше, изобретение обеспечивает композицию, включающую конъюгат по настоящему изобретению и один или более дополнительных антигенов. Композиция типично представляет собой иммуногенную композицию.
Дополнительный антиген (антигены) может включать дополнительные конъюгаты по настоящему изобретению, и, таким образом, изобретение обеспечивает композицию, включающую более чем один конъюгат по настоящему изобретению. В частности, настоящее изобретение обеспечивает композицию, включающую конъюгат капсульного полисахарида типа 5 по настоящему изобретению и конъюгат капсульного полисахарида типа 8 по настоящему изобретению. Альтернативно, дополнительный антиген (антигены) может представлять собой конъюгаты капсульного полисахарида типа 5 или типа 8, полученные способами, отличными от способов, раскрытых в настоящем изобретении, например, способами, раскрытыми в ссылочных документах 1-13 выше. Дополнительный антиген (антигены) может также представлять собой конъюгаты капсульного полисахарида типа 5 или типа 8, полученные способами, раскрытыми в ссылочных документах 59, 70, 71, 72, 73, и 74, и, в частности, способами, проиллюстрированными в этих документах. Соответственно, изобретение обеспечивает композицию, включающую конъюгат капсульного полисахарида типа 5 и конъюгат капсульного полисахарида типа 8, где один из конъюгатов (конъюгат типа 5 или конъюгат типа 8) представляет собой конъюгат по настоящему изобретению, а другой конъюгат не является конъюгатом по настоящему изобретению.
Дополнительный антиген (антигены) может включать другие антигены S.aureus, включая белковый и сахаридный антигены, описанные ниже.
Дополнительный антиген (антигены) может включать антигены из не относящихся к S.aureus патогенов. Таким образом, композиции по настоящему изобретению дополнительно могут включать один или более отличных от S.aureus антигенов, включая дополнительные бактериальные, вирусные или паразитические антигены. Они могут быть выбраны из следующих:
- белковый антиген из N.meningitidis серологической группы B, такой как антигены, описанные в ссылочных документах 75-81, при этом белок '287' (см. ниже) и производные (например, 'ΔG287') являются особенно полезными;
- везикулярный препарат внешних мембран (OMV) из N.meningitidis серологической группы B, такой как раскрытые в ссылочных документах 82, 83, 84, 85 и т.п.;
- сахаридный антиген из N.meningitidis серологической группы A, C, W135 и/или Y, такой как олигосахарид, раскрытый в ссылочном документе 86, из серологической группы C или олигосахариды ссылочного документа 87;
- сахаридный антиген из Streptococcus pneumoniae [например, ссылочные документы 88-90; главы 22 и 23 ссылочного документа 97];
- антиген из вируса гепатита А, такой как инактивированный вирус [например, 91, 92; глава 15 ссылочного документа 97];
- антиген из вируса гепатита B, такой как поверхностные и/или сердцевинные антигены [например, 92, 93; глава 16 ссылочного документа 97];
- антиген из вируса гепатита C [например, 94];
- антиген из Bordetella pertussis, такой как коклюшный голотоксин (PT) и волокнистый гемагглютинин (FHA) из B.pertussis, необязательно также в сочетании с пертактином и/или агглютиногенами 2 и 3 [например, ссылочные документы 95 и 96; глава 21 ссылочного документа 97];
- дифтерийный антиген, такой как дифтерийный токсоид [например, глава 13 ссылочного документа 97];
- столбнячный антиген, такой как столбнячный токсоид [например, глава 27 ссылочного документа 97];
- сахаридный антиген из Haemophilus influenzae B [например, глава 14 ссылочного документа 97];
- антиген из N.gonorrhoeae [например, 75, 76, 77];
- антиген из Chlamydia pneumoniae [например, 98, 99, 100, 101, 102, 103, 104];
- антиген из Chlamydia trachomatis [например, 105];
- антиген из Porphyromonas gingivalis [например, 106];
- полиомиелитный антиген(антигены) [например, 107, 108; глава 24 ссылочного документа 97], такой как IPV;
- антиген (антигены) бешенства [например, 109], такой как лиофилизированный инактивированный вирус [например, 110, RabAvert™];
- антигены кори, свинки и/или краснухи [например, главы 19, 20 и 26 ссылочного документа 97];
- антиген (антигены) гриппа [например, главы 17 и 18 ссылочного документа 97], такие как поверхностные белки гемагглютинина и/или нейраминидазы;
- антиген из Moraxella catarrhalis [например, 111];
- антиген из Streptococcus pyogenes (группа А streptococcus) [например, 112, 113, 114];
- антиген из Streptococcus agalactiae (группа B streptococcus) [например, 56, 115-117];
- антиген из S.epidermidis [например, капсульный полисахарид типа I, II и/или III, получаемый из штаммов ATCC-31432, SE-360 и SE-10, как описано в ссылочных документах 118, 119 и 120.
Когда используют сахаридный или углеводный антиген, его типично конъюгируют с носителем для усиления иммуногенности. Конъюгация H.influenzae B, менингококковых и пневмококковых сахаридных антигенов хорошо известна.
Токсичные белковые антигены могут быть детоксифицированы, если это необходимо (например, детоксификация коклюшного токсина химическими и/или генетическими способами [96]).
Когда дифтерийный антиген включен в композицию, она типично также включает столбнячный антиген и коклюшные антигены. Подобным образом, когда включен столбнячный антиген, композиция типично также включает дифтерийный и коклюшный антигены. Подобным образом, когда включен коклюшный антиген, композиция также типично включает дифтерийный и столбнячный антигены.
Антигены могут быть адсорбированы на алюминиевой соли.
Один тип предпочтительной композиции включает дополнительные антигены, которые поражают субъектов с ослабленным иммунитетом, и, таким образом, конъюгаты S.aureus по настоящему изобретению можно сочетать с одним или более антигенами из следующих отличных от S.aureus патогенов: Steptococcus agalactiae, Staphylococcus epidermis, вирус гриппа, Enterococcus faecalis, Pseudomonas aeruginosa, Legionella pneumophila, Listeria monocytogenes, Neisseria meningitidis и вирус парагриппа.
Другой тип предпочтительной композиции включает дополнительные антигены из бактерий, ассоциированных с нозокомиальными инфекциями, и поэтому конъюгаты S.aureus по настоящему изобретению можно сочетать с одним или более антигенами из следующих отличных от S.aureus патогенов: Clostridium difficile; Pseudomonas aeruginosa; Candida albicans; и внекишечная патогенная Escherichia coli.
Антигены в композиции типично могут присутствовать в концентрации, по меньшей мере, 1 мкг/мл каждый. Как правило, концентрация любого присутствующего антигена должна быть достаточной, чтобы вызвать иммунный ответ против этого антигена.
В качестве альтернативы использования антигенов, которые представляют собой белки, в композиции по настоящему изобретению, можно использовать нуклеиновую кислоту, кодирующую такой антиген [например, ссылочные документы 121-129]. Белковые компоненты композиций по настоящему изобретению, таким образом, можно заменить нуклеиновой кислотой (обычно ДНК, например, в форме плазмиды), которая кодирует белок.
В практических условиях может существовать верхний предел количества антигенов, включенных в композиции по настоящему изобретению. Количество антигенов (включая антигены S.aureus) в композиции по настоящему изобретению может быть меньше чем 20, меньше чем 19, меньше чем 18, меньше чем 17, меньше чем 16, меньше чем 15, меньше чем 14, меньше чем 13, меньше чем 12, меньше чем 11, меньше чем 10, меньше чем 9, меньше чем 8, меньше чем 7, меньше чем 6, меньше чем 5, меньше чем 4 или меньше чем 3. Количество S.aureus антигенов в композиции по настоящему изобретению может быть меньше чем 6, меньше чем 5 или меньше чем 4.
Фармацевтические композиции и способы
Изобретение обеспечивает фармацевтическую композицию, включающую (a) конъюгат по настоящему изобретению и (b) фармацевтически приемлемый носитель. Типично 'фармацевтически приемлемые носители' включают любой носитель, который сам не индуцирует продукцию антител, вредных для субъекта, принимающего композицию. Подходящие носители типично представляют собой крупные, медленно метаболизируемые макромолекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, аминокислотные сополимеры, сахарозу [130], трегалозу [131], лактозу и липидные агрегаты (такие как масляные капли или липосомы). Такие носители хорошо известны специалистам в данной области со средней квалификацией. Вакцины также могут содержать разбавители, такие как вода, физиологический раствор, глицерин и т.п. Кроме того, могут присутствовать вспомогательные вещества, такие как смачивающие вещества или эмульгаторы, регулирующие pH буферные вещества и подобные. Стерильный апирогенный фосфатно-буферный физиологический раствор является типичным носителем. Подробное обсуждение фармацевтически приемлемых эксципиентов можно найти в ссылочном документе 132.
Композиции по настоящему изобретению могут быть в водной форме (т.е. растворы или суспензии) или в сухой форме (например, лиофилизированные). Если используют сухую вакцину, тогда она должна быть восстановлена в жидкой среде непосредственно перед инъекцией. Лиофилизация конъюгатных вакцин известна из уровня техники, например, Menjugate™ продукт представлен в лиофилизированной форме, тогда как NeisVac-C™ и Meningitec™ представлены в водной форме. Для стабилизации конъюгатов в процессе лиофилизации может быть типичным включение в композицию сахарного спирта (например, маннита) или дисахарида (например, сахарозы или трегалозы), например, в концентрации в пределах от 1 мг/мл до 30 мг/мл (например, около 25 мг/мл).
Композиции могут быть представлены во флаконах, или они могут быть представлены в готовых заполненных шприцах. Шприцы могут поставляться с иглами или без них. Шприц включает одну дозу композиции, тогда как флакон может включать одну дозу или несколько доз.
Водные композиции по настоящему изобретению также являются подходящими для восстановления других вакцин из лиофилизированной формы. Когда композиция по настоящему изобретению предназначена для такого восстановления для немедленного использования, изобретение обеспечивает набор, который может включать два флакона или может включать один уже заполненный шприц и один флакон, при этом содержимое шприца используют для реактивации содержимого флакона непосредственно перед инъекцией.
Композиции по настоящему изобретению могут быть упакованы в виде форм, включающих стандартную дозу или многократные дозы. Для форм, включающих многократные дозы, флаконы являются более предпочтительными, чем предварительно заполненные шприцы. Эффективные объемы доз можно определить рутинным путем, но типично для человека доза композиции имеет объем 0,5 мл, например, для внутримышечной инъекции.
pH композиции типично находится в пределах от 6 до 8, например около 7. Стабильный pH можно поддерживать с использованием буфера. Если композиция включает соль гидроксида алюминия, типично используют гистидиновый буфер [133]. Композиция может быть стерильной и/или апирогенной. Композиции по настоящему изобретению могут быть изотоничным по отношению к человеку.
Композиции по настоящему изобретению являются иммуногенными и более предпочтительно представляют собой композиции вакцин. Вакцины в соответствии с изобретением могут быть либо профилактическими (т.е. для предотвращения инфекции), либо терапевтическими (т.е. для лечения инфекции), но типично они являются профилактическими. Иммуногенные композиции, используемые в качестве вакцин, включают иммунологически эффективное количество антигена (антигенов), а также любые другие компоненты, по мере необходимости. Под 'иммунологически эффективном количеством' подразумевается, что введение такого количества субъекту, либо в виде разовой дозы, либо в виде части серии доз, является эффективным для лечения или профилактики. Это количество варьирует в зависимости от здоровья и физического состояния субъекта, подлежащего лечению, возраста, таксономической группы субъекта, подлежащего лечению (например, не относящиеся к человеку приматы, приматы и т.п.), способности иммунной системы субъекта синтезировать антитела, желаемой степени защиты, композиции вакцины, оценки лечащим врачом медицинской ситуации и других соответствующих факторов. Как ожидают, такое количество находится в относительно широком диапазоне, который можно определить путем рутинных испытаний.
В каждой дозе количество каждого отдельного сахаридного антигена, как правило, находится в пределах 1-50 мкг (измерено как масса сахарида), например около 1 мкг, около 2,5 мкг, около 4 мкг, около 5 мкг или около 10 мкг.
S.aureus поражает различные области организма, и поэтому композиции по настоящему изобретению можно получить в различных формах. Например, композиции можно получить в виде препаратов для инъекций, в виде жидких растворов или суспензий. Композиции можно получить для внутрилегочного введения, например, в виде препарата для ингаляций с использованием тонкодисперсного порошка или спрея. Композицию можно получить в виде суппозитория или пессария. Композицию можно получить для назального, внутриушного или внутриглазного введения, например, в виде спрея, капель, геля или порошка [например, ссылочные документы 134 и 135]. Сообщалось об успехе, полученном при назальном введении пневмококковых сахаридов [136, 137], Hib сахаридов [138], MenC сахаридов [139] и смесей Hib и MenC сахаридных конъюгатов [140].
Композиции по настоящему изобретению могут включать антимикробное средство, в частности, когда они упакованы в многодозовом формате.
Композиции по настоящему изобретению могут включать детергент, например Tween (полисорбат), такой как Tween 80. Детергенты, как правило, присутствуют на низких уровнях, например <0,01%.
Композиции по настоящему изобретению могут включать натриевые соли (например, хлорид натрия) для сообщения тоничности. Концентрация 10±2 мг/мл NaCl является типичной.
Композиции по настоящему изобретению, как правило, включают буфер. Фосфатный буфер является типичным.
Композиции по настоящему изобретению, как правило, можно вводить в сочетании с другими иммунорегуляторными средствами. В частности, композиции обычно включают один или более адъювантов. Такие адъюванты включают, но не ограничиваются этим:
A. Минералсодержащие композиции
Минералсодержащие композиции, подходящие для использования в качестве адъювантов в настоящем изобретении, включают минеральные соли, такие как алюминиевые соли и кальциевые соли. Изобретение включает минеральные соли, такие как гидроксиды (например, оксигидроксиды), фосфаты (например, гидроксифосфаты, ортофосфаты), сульфаты и т.п. [например, см. главы 8 и 9 ссылочного документа 141], или смеси различных минеральных соединений (например, смесь фосфатного и гидроксидного адъюванта, необязательно с избытком фосфата), при этом соединения принимают любую подходящую форму (например, гелевую, кристаллическую, аморфную и т.п.), и адсорбция солью (солями) является типичной. Минералсодержащие композиции также могут быть сформулированы в виде частицы металлической соли [142].
Алюминиевые соли могут быть включены в вакцины по настоящему изобретению таким образом, чтобы доза Al3+ находилась в пределах от 0,2 до 1,0 мг на дозу.
Типичный алюминийфосфатный адъювант представляет собой аморфный гидроксифосфат алюминия с PO4/Al молярным отношением в пределах от 0,84 до 0,92, включен при 0,6 мг Al3+/мл. Можно использовать адсорбцию с низкой дозой фосфата алюминия, например в пределах от 50 до 100 г Al3+ на конъюгат в расчете на дозу. Когда используют фосфат алюминия и когда желательно не адсорбировать антиген на адъюванте, предпочтительно включение свободных фосфатных ионов в раствор (например, путем использования фосфатного буфера).
B. Масляные эмульсии
Композиции масляных эмульсий, подходящие для использования в качестве адъювантов в настоящем изобретении, включают сквален-водные эмульсии, такие как MF59 (5% Сквалена, 0,5% Tween 80 и 0,5% Span 85, сформулированные в виде субмикронных частиц с использованием микрофлюидизатора) [Глава 10 ссылочного документа 141; см. также ссылочные документы 143-145]. MF59 используют в качестве адъюванта в FLUAD™ вакцине, включающей трехвалентную субъединицу вируса гриппа.
Особенно полезными адъювантами для использования в композициях являются субмикронные эмульсии масло-в-воде. Предпочтительные субмикронные эмульсии масло-в-воде для использования в настоящем изобретении представляют собой сквален/водные эмульсии, необязательно содержащие различные количества MTP-PE, такие как субмикронная эмульсия масло-в-воде, содержащая 4-5% масс./об. сквалена, 0,25-1,0% масс./об. Tween 80 (полиоксиэтиленсорбитан моноолеат) и/или 0,25-1,0% Span 85 (сорбитан триолеат) и, необязательно, N-ацетилмурамил-L-аланил-D-изоглуатминил-L-аланин-2-(1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфосфорилокси)этиламин (MTP-PE). Субмикронные эмульсии масло-в-воде, способы их получения и иммуностимулирующие средства, такие как мурамиловые пептиды, для использования в композициях подробно описаны в ссылочных документах 143 и 146-147.
Полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (1FA) также можно использовать в качестве адъювантов в настоящем изобретении.
C. Композиции сапонинов [глава 22 ссылочного документа 141]
Композиции сапонинов также можно использовать в качестве адъювантов в настоящем изобретении. Сапонины представляют собой гетерологичную группу стеролгликозидов и тритерпеноидных гликозидов, которые присутствуют в коре, листьях, стеблях, корнях и даже цветках широкого ряда видов растений. Сапонины, выделенные из коры дерева Quillaia saponaria Molina, были тщательно изучены в качестве адъювантов. Сапонин также можно получать коммерческим путем из Smilax ornata (сарсапорель), Gypsophilla paniculata (гипсофила) и Saponaria officianalis (мыльный корень). Композиции адъювантов на основе сапонинов включают очищенные композиции, такие как QS21, а также липидные композиции, такие как ISCOM.
Композиции сапонинов очищают с использованием ВЭЖХ и ОФ-ВЭЖХ. Были идентифицированы специфические фракции, очищенные с использованием этих процедур, включая QS7, QS17, QS18, QS21, QH-A, QH-B и QH-C. Предпочтительно, сапонин представляет собой QS21. Способ получения QS21 раскрыт в ссылочном документе 148. Композиции сапонинов также могут включать стерол, такой как холестерин [149].
Сочетания сапонинов и холестеринов можно использовать для образования уникальных частиц, которые называют иммуностимулирующими комплексами (ISCOM) [глава 23 ссылочного документа 141]. ISCOM типично также включают фосфолипид, такой как фосфатидилэтаноламин или фосфатидилхолин. Любой известный сапонин можно использовать в ISCOM. Предпочтительно, ISCOM включает один или более из QuilA, QHA и QHC. ISCOM более подробно описаны в ссылочных документах 149-151. Необязательно, ISCOM могут быть лишены дополнительного детергента (детергентов) [152].
Обзор, касающийся разработки адъювантов на основе сапонинов, можно найти в ссылочных документах 153 и 154.
D. Виросомы и вирусоподобные частицы
Виросомы и вирусоподобные частицы (VLP) также можно использовать в качестве адъювантов в настоящем изобретении. Эти структуры, как правило, содержат один или более белков из вируса, необязательно объединенных или сформулированных с фосфолипидом. Они, как правило, являются непатогенными, нереплицирующими и, как правило, не содержат никакого природного вирусного генома. Вирусные белки могут быть рекомбинантно получены или выделены из полных вирусов. Эти вирусные белки, подходящие для использования в виросомах или VLP, включают белки, выделенные из вируса гриппа (такие как HA или NA), вируса гепатита B (такие как сердцевинные или капсидные белки), вируса гепатита E, вируса кори, вируса Sindbis, ротавируса, вируса ящура, ретровируса, вируса Norwalk, вируса папилломы человека, ВИЧ, РНК-фагов, Qβ-фага (такие как белки оболочки), GA-фага, fr-фага, AP205 фага и Ty (такие как ретротранспозон Ty белок pl). VLP более подробно обсуждаются в ссылочных документах 155-160. Виросомы более подробно обсуждаются, например, в ссылочном документе 161.
E. Бактериальные или микробные производные
Адъюванты, подходящие для использования в настоящем изобретении, включают бактериальные или микробные производные, такие как нетоксичные производные энтеробактериального липополисахарида (LPS), производные липида А, иммуностимулирующие олигонуклеотиды и ADP-рибозилирующие токсины и их детоксифицированные производные.
Нетоксичные производные LPS включают монофосфориллипид (MPL) и 3-O-деацилированный PL (3dMPL). 3dMPL представляет собой смесь 3-де-O-ацилированного монофосфориллипида А с 4, 5 или 6 ацилированными цепями. Предпочтительная форма "мелких частиц" де-O-ацилированного монофосфориллипида А раскрыта в ссылочном документе 162. Такие "мелкие частицы" 3dMPL являются достаточно мелкими для стерильного фильтрования через 0,22-мкм мембрану [162]. Другие нетоксичные LPS производные включают миметики монофосфориллипида А, такие как аминоалкилглюкозаминидфосфатные производные, например RC-529 [163, 164].
Производные липида А включают производные липида А из Escherichia coli, такие как OM-174. OM-174 описан, например, в ссылочных документах 165 и 166.
Иммуностимулирующие олигонуклеотиды, подходящие для использования в качестве адъювантов в настоящем изобретении, включают нуклеотидные последовательности, содержащие CpG мотив (динуклеотидная последовательность, содержащая неметилированный цитозин, связанный фосфатной связью с гуанозином). Было показано, что двухцепочечные РНК и олигонуклеотиды, содержащие палиндромные или поли(dG) последовательности, также являются иммуностимулирующими.
CpG могут включать нуклеотидные модификации/аналоги, такие как фосфоротиоатные модификации, и могут быть двухцепочечными или одноцепочечными. Ссылочные документы 167, 168 и 169 раскрывают возможные аналоговые замены, например, замену гуанозина 2'-дезокси-7-деазагуанозином. Адъювантный эффект CpG олигонуклеотидов также обсуждается в ссылочных документах 170-175.
CpG последовательность может быть направлена на TLR9, например, мотив GTCGTT или TTCGTT [176]. CpG последовательность может быть специфической для индукции Th1 иммунного ответа, например, CpG-A ODN, или она может быть более специфической для индукции B-клеточного ответа, например, CpG-B ODN. CpG-A и CpG-B ODN обсуждаются в ссылочных документах 177-179. Предпочтительно, CpG представляет собой CpG-A ODN.
Предпочтительно, CpG олигонуклеотид сконструирован таким образом, что 5'-конец является доступным для распознавания рецептором. Необязательно, две CpG олигонуклеотидные последовательности могут быть связаны по их 3'-концам с образованием "иммуномеров". См., например, ссылочные документы 176 и 180-182.
Бактериальные ADP-рибозилирующие токсины и их детоксифицированные производные можно использовать в качестве адъювантов в настоящем изобретении. Предпочтительно, белок происходит из E.coli (E.coli термолабильный энтеротоксин "LT"), холеры ("CT") или коклюша ("PT"). Использование детоксифицированных ADP-рибозилирующих токсинов в качестве адъювантов для введения через слизистую оболочку описано в ссылочном документе 183, и в качестве парентеральных адъювантов - в ссылочном документе 184. Токсин или токсоид предпочтительны в форме голотоксина, включающего как А, так и B субъединицы. Предпочтительно, А субъединица содержит детоксифицирующую мутацию; предпочтительно, B субъединица не является мутированной. Предпочтительно, адъювант представляет собой детоксифицированный LT мутант, такой как LT-K63, LT-R72 и LT-G192. Использование ADP-рибозилирующих токсинов и их детоксифицированных производных, в частности, LT-K63 и LT-R72, в качестве адъювантов можно найти в ссылочных документах 185-192. Различные ссылки на аминокислотные замены предпочтительно основаны на расположениях А и B субъединиц ADP-рибозилирующих токсинов, описанных в ссылочном документе 193, который специально включен в настоящую заявку посредством ссылки во всей его полноте.
F. Человеческие иммуномодуляторы
Человеческие иммуномодуляторы, подходящие для использования в качестве адъювантов в настоящем изобретении, включают цитокины, такие как интерлейкины (например, IL-1, IL-2, IL-4, IL-5, IL-6, IL-7, IL-12 [194] и т.п.) [195], интерфероны (например, интерферон-γ), макрофагальный колониестимулирующий фактор и фактор некроза опухоли.
G. Биоадгезивы и мукоадгезивы
Биоадгезивы и мукоадгезивы также можно использовать в качестве адъювантов в настоящем изобретении. Подходящие биоадгезивы включают этерифицированную гиалуроновую кислоту в виде микросфер [196] или мукоадгезивы, такие как сшитые производные поли(акриловой кислоты), поливиниловый спирт, поливинилпирролидон, полисахариды и карбоксиметилцеллюлозу. Хитозан и его производные также можно использовать в качестве адъювантов в настоящем изобретении [197].
H. Микрочастицы
Микрочастицы также можно использовать в качестве адъювантов в настоящем изобретении. Микрочастицы (т.е. частицы диаметром от ~100 нм до ~150 мкм, более предпочтительно диаметром от ~200 нм до ~30 мкм и наиболее предпочтительно диаметром от ~500 нм до ~10 мкм), образованные из веществ, которые являются биоразлагаемыми и нетоксичными (например, поли(α-оксикислота), полиоксимасляная кислота, полиортоэфир, полиангидрид, поликапролактон и т.п.), с поли(лактид-ко-гликолид)ом, являются предпочтительным, и их необязательно обрабатывают, чтобы они имели отрицательно-заряженную поверхность (например, при помощи SDS) или положительно-заряженную поверхность (например, катионным детергентом, таким как CTAB).
I. Липосомы (главы 13 и 14 ссылочного документа 141)
Примеры липосомных композиций, подходящих для использования в качестве адъювантов, описаны в ссылочных документах 198-200.
J. Композиции полиоксиэтиленового простого эфира и полиоксиэтиленового сложного эфира
Адъюванты, подходящие для использования в настоящем изобретении, включают полиоксиэтиленовые простые эфиры и полиоксиэтиленовые сложные эфиры [201]. Такие композиции дополнительно включают поверхностно-активные вещества на основе сложного эфира полиоксиэтилена и сорбитана в сочетании с октоксинолом [202], а также поверхностно-активные вещества на основе полиоксиэтиленалкиловых простых эфиров или сложного эфира в сочетании с, по меньшей мере, одним дополнительным неионным поверхностно-активным веществом, таким как октоксинол [203]. Предпочтительные полиоксиэтиленовые простые эфиры выбраны из следующей группы: полиоксиэтилен-9-лауриловый эфир (лаурет 9), полиоксиэтилен-9-стеориловый эфир, полиоксиэтилен-8-стеориловый эфир, полиоксиэтилен-4-лауриловый эфир, полиоксиэтилен-35-лауриловый эфир и полиоксиэтилен-23-лауриловый эфир.
K. Полифосфазен (PCPP)
PCPP композиции описаны, например, в ссылочных документах 204 и 205.
L. Мурамиловые пептиды
Примеры мурамиловых пептидов, подходящих для использования в качестве адъювантов в настоящем изобретении, включают N-ацетил-мурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетил-нормурамил-L-аланил-D-изоглутамин (нор-MDP) и N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-sw-глицеро-3-гидроксифосфорилокси)этиламин MTP-PE).
M. Имидазохинолоновые соединения.
Примеры имидазохинолоновых соединений, подходящих для использования в качестве адъювантов в настоящем изобретении, включают Imiquamod и его гомологи (например, "Resiquimod 3M"), описанные более подробно в ссылочных документах 206 и 207.
N. Тиосемикарбазоновые соединения.
Примеры тиосемикарбазоновых соединений, а также способы формулирования, получения и определения соединений, которые все являются подходящими для использования в качестве адъювантов в настоящем изобретении, включают такие, которые описаны в ссылочном документе 208. Тиосемикарбазоны являются особенно эффективными в стимуляции мононуклеарных клеток человеческой периферийной крови для продукции цитокинов, таких как TNF-α.
O. Триптантриновые соединения.
Примеры триптантриновых соединений, а также способы формулирования, получения и определения соединений, которые все являются подходящими для использования в качестве адъювантов в настоящем изобретении, включают такие, которые описаны в ссылочном документе 209. Триптантриновые соединения являются особенно эффективными в стимуляции мононуклеарных клеток человеческой периферийной крови для продукции цитокинов, таких как TNF-α.
Изобретение также может включать сочетания аспектов одного или нескольких адъювантов, описанных выше. Например, следующие сочетания можно использовать в качестве композиций адъювантов в настоящем изобретении: (1) сапонин и эмульсия масло-в-воде [210]; (2) сапонин (например, QS21)+нетоксичное LPS производное (например, 3dMPL) [211]; (3) сапонин (например, QS21)+нетоксичное LPS производное (например, 3dMPL)+холестерин; (4) сапонин (например, QS21)+3dMPL+IL-12 (необязательно+стерол) [212]; (5) сочетания 3dMPL, например, с QS21 и/или эмульсией масло-в-воде [213]; (6) SAF, содержащий 10% сквален, 0,4% Tween 80™, 5% плюроник-блок-полимер L121 и thr-MDP, либо микрофлюидизированный в субмикронной эмульсии, либо обработанный путем перемешивания с встряхиванием для образования эмульсии с более крупным размером частиц; (7) система адъюванта Ribi™ (RAS), (Ribi Immunochem), содержащая 2% сквалена, 0,2% Tween 80 и один или более бактериальных клеточнооболочных компонентов из группы, включающей монофосфорилипид (MPL), трегалоза димиколят (TDM) и скелет клеточной оболочки (CWS), предпочтительно MPL+CWS (Detox™); и (8) одна или более минеральных солей (такие как алюминиевая соль)+нетоксичное производное LPS (такое как 3dMPL).
Другие вещества, которые действуют как иммуностимулирующие средства, раскрыты в главе 7 ссылочного документа 141.
Использование адъювантов на основе алюминиевой соли является особенно полезным, и антигены, как правило, адсорбируют на такие соли. Для конъюгатов Menjugate™ и NeisVac™ используют гидроксидный адъювант, тогда как для Meningitec™ используют фосфатный адъювант. Композиции по настоящему изобретению могут включать некоторые антигены, адсорбированные на гидроксиде алюминия, но также включать другие антигены в ассоциации с фосфатом алюминия. Однако обычно используют только одну соль, например гидроксид или фосфат, но не обе сразу. Не все конъюгаты должны быть адсорбированы, т.е. некоторые из них или все могут быть свободными в растворе.
Способы лечения
Изобретение также обеспечивает способ усиления иммунного ответа у млекопитающего, включающий введение млекопитающему фармацевтической композиции по настоящему изобретению. Иммунный ответ предпочтительно является защитным и предпочтительно включает антитела. Способ может вызывать бустерный ответ.
Млекопитающим предпочтительно является человек. Когда вакцина предназначена для профилактического применения, человек предпочтительно представляет собой ребенка (например, ребенок, начинающий ходить, или младенец) или подростка; когда вакцина предназначена для терапевтического применения, человек предпочтительно представляет собой взрослого человека. Вакцину, предназначенную для детей, также можно вводить взрослым, например, для определения безопасности, дозы, иммуногенности и т.п. Предпочтительным классом людей для лечения являются пациенты, подверженные риску развития нозокомиальной инфекции, в частности, пациенты с поздней стадией почечного заболевания и/или находящиеся на гемодиализе. Другие пациенты, подверженные риску развития нозокомиальной инфекции, также являются предпочтительными, например, пациенты с иммунодефицитом или перенесшие хирургическую операцию, особенно операцию на сердце, или травму. Другим предпочтительным классом людей для лечения являются пациенты, подверженные риску развития бактериемии. Еще одним предпочтительным классом являются пациенты, страдающие от, или которые перед этим подвергались воздействию, вируса гриппа, поскольку S.aureus связан с пост-инфекционной пневмонией у этих пациентов.
Изобретение также обеспечивает композицию по настоящему изобретению для использования в качестве лекарственного средства. Лекарственное средство предпочтительно способно повышать иммунный ответ у млекопитающего (т.е. это иммуногенная композиция), и более предпочтительно представляет собой вакцину.
Изобретение также обеспечивает использование конъюгата по настоящему изобретению для получения лекарственного средства для повышения иммунного ответа у млекопитающего.
Эти применения и способы предпочтительно предназначены для профилактики и/или лечения заболевания, вызванного S.aureus, например кожных инфекций, таких как импетиго, фурункулы, фолликулярный целлюлит, ячмени, фурункулы, карбункулы, токсический эпидермальный некролиз и абсцессы, септический артрит, пневмония, мастит, флебит, менингит, инфекции мочевых путей, остеомиелит, эндокардит, синдром токсического шока (TSS), септицемия и нозокомиальные инфекции.
Один способ проверки эффективности терапевтического лечения включает мониторинг S.aureus инфекции после введения композиции по настоящему изобретению. Один способ проверки эффективности профилактического лечения включает мониторинг иммунных ответов против S.aureus антигенов после введения композиции.
Предпочтительные композиции по настоящему изобретению могут обеспечивать титр антител у пациента, который выше критерия для серопротекции для каждого антигенного компонента для приемлемого процентного количества субъектов-людей. Антигены с соответствующим титром антител, выше которого считается, что у хозяина имеет место сероконверсия против антигена, хорошо известны, и такие титры опубликованы организациями, такими как ВОЗ. Предпочтительно сероконверсия наблюдается у более чем 80% статистически значимой выборки субъектов, более предпочтительно у более чем 90%, еще более предпочтительно у более чем 93%, и наиболее предпочтительно у 96-100%.
Композиции по настоящему изобретению, как правило, можно вводить непосредственно пациенту. Прямую доставку можно осуществить путем парентеральной инъекции (например, подкожно, интраперитонеально, внутривенно, внутримышечно или в интерстициальное пространство ткани) или путем ректального, перорального, вагинального, местного, чрескожного, интраназального, внутриглазного, внутриушного, легочного или другого пути введения через слизистую оболочку. Внутримышечное введение в область бедра или плеча является предпочтительным. Инъекцию можно вводить через иглу (например, гиподермическую иглу), но альтернативно можно использовать инъекцию без иглы. Типичная внутримышечная доза составляет 0,5 мл.
Изобретение можно использовать, чтобы вызвать системный и/или мукозальный иммунитет.
Дозы для лечения могут включать схему введения разовой дозы схему введения нескольких доз. Несколько доз можно использовать в схеме первичной иммунизации и/или в схеме бустерной иммунизации. После схемы введения первичной дозы можно использовать схему введения бустерной дозы. Подходящий интервал времени между первичными дозами (например, в пределах 4-16 недель) и между первичной и ревакцинацией можно определить рутинным способом.
Антигены S.aureus
Как указано выше, в композиции по настоящему изобретению можно включить один или более дополнительных антигенов S.aureus. Антигены могут представлять собой белковые или сахаридные антигены. Белковые антигены S.aureus можно использовать в качестве белков-носителей для конъюгатов по настоящему изобретению, белков-носителей для других конъюгатов или в качестве неконъюгированных белковых антигенов. Сахаридные антигены S.aureus можно использовать в качестве сахаридов для другого конъюгата или в качестве неконъюгированных сахаридных антигенов.
Подходящие сахаридные антигены S.aureus включают экзополисахарид S.aureus, который представляет собой поли-N-ацетилглюкозамин (PNAG). Этот полисахарид присутствует как в S.aureus, так и в S.epidermidis, и может быть выделен из любого такого источника [214, 215]. Например, PNAG может быть выделен из S.aureus штамма MN8m [216]. Сахаридный антиген может представлять собой полисахарид, имеющий размер, который возникает в процессе очистки экзополисахарида из бактерий, или это может быть полисахарид, полученный путем фрагментации такого полисахарида, например, размер может варьировать от более 400 кДа до 75-400 кДа, или от 10 до 75 кДа, или вплоть до 30 повторяющихся звеньев. Сахаридный антиген может иметь разные степени N-ацетилирования, и, как описано в ссылочном документе 217, PNAG может быть менее чем 40% N-ацетилирован (например, менее чем 35, 30, 20, 15, 10 или 5% N-ацетилирован; деацетилированный PNAG также известен как dPNAG). Деацетилированные эпитопы PNAG могут вызвать образование антител, которые способны опосредовать опсоническое убивание. Получение dPNAG описано в ссылочном документе 218. PNAG может быть или не быть O-сукцинилированным, например, он может быть O-сукцинилирован по менее чем 25, 20, 15, 10, 5, 2, 1 или 0,1% остатков. PNAG может быть конъюгирован с молекулой носителя, как описано выше, или, альтернативно, может быть неконъюгированным.
Другим подходящим сахаридным антигеном S.aureus является антиген типа 336, который представляет собой β-связанный гексозамин с отсутствием O-ацетилирования [219, 220]. Антиген типа 336 является перекрестно реагирующим с антителами, направленными против 336 штамма (ATCC 55804). Антиген типа 336 может быть конъюгирован с молекулой носителя, как описано выше, или, альтернативно, может быть неконъюгированным.
Подходящие белковые антигены S.aureus включают следующие антигены S.aureus (или антигены, включающие его иммуногенный фрагмент (фрагменты)) [например, см. ссылочные документы 221-228]: AhpC, AhpF, аутолизинамидаза, аутолизинглюкозаминидаза, коллаген-связывающий белок CAN, EbhB, GehD липаза, гепарин-связывающий белок HBP (17 кДа), ламининовый рецептор, MAP, MntC (также известен как SitC), MRP1I, Npaза, ORF0594, ORF0657n, ORF0826, PBP4, RAP (РНК III активирующий белок), Sai-1, SasK, SBI, SdrG, SdrH, SSP-1, SSP-2 и витронектин-связывающий белок.
Дополнительные подходящие белковые антигены S.aureus включают антиген clfA; антиген clfB; антиген sdrE2; антиген sdrC; антиген sasF, антиген emp; антиген sdrD; антиген spa; антиген esaC; антиген esxA; антиген esxB; антиген sta006; антиген isdC; антиген Hla; антиген sta011; антиген isdA; антиген isdB; и антиген sta073, описанные ниже. Один или более (т.е. 1, 2, 3, 4, 5, 6 или больше) из этих антигенов могут присутствовать в композиции по настоящему изобретению. Из этих антигенов, специально предусмотрено использование одного или более (т.е. 1, 2, 3, 4, 5, 6 или больше) из антигена esxA; антигена esxB; антигена sta006; антигена Hla; антигена sta011; и/или антигена sta073.
Например, композиция по настоящему изобретению дополнительно может включать одно из следующих сочетаний белковых антигенов S.aureus:
(1) Антиген esxA, антиген esxB, антиген sta006 и антиген Hla. Антигены esxA и esxB могут быть успешно объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB с антигеном esxB ниже от антигена esxA. Антиген Hla может представлять собой детоксифицированный мутант, например, включающий H35L мутацию.
(2) Антиген esxA, антиген esxB, антиген sta006 и антиген sta011. Антигены esxA и esxB могут быть объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB.
(3) Антиген esxA, антиген esxB и антиген sta011. Антигены esxA и esxB могут быть успешно объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB.
(4) Антиген esxA, антиген esxB, антиген Hla, антиген sta006 и антиген sta011. Антигены esxA и esxB могут быть объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB. Антиген Hla может представлять собой детоксифицированный мутант, например, включающий H35L мутацию.
(5) Антиген esxA, антиген esxB и антиген Hla. Антигены esxA и esxB могут быть успешно объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB. Антиген Hla может представлять собой детоксифицированный мутант, например, включающий H35L мутацию.
(6) Антиген Hla, антиген sta006 и антиген sta011. Антиген Hla может представлять собой детоксифицированный мутант, например, включающий H35L мутацию.
(7) Антиген esxA и антиген esxB. Антигены esxA и esxB могут быть успешно объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB.
(8) Антиген esxA, антиген esxB и антиген sta006. Антигены esxA и esxB могут быть успешно объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB.
(9) Антиген esxA, антиген esxB, антиген sta011 и антиген sta073. Антигены esxA и esxB могут быть объединены в виде гибридного полипептида, как обсуждается ниже, например, гибрида EsxAB.
(10) Антиген sta006 и антиген sta011.
Другие антигены Staphylococcus aureus раскрыты в ссылочном документе 229.
clfA
Антиген 'clfA' аннотируется как 'фактор слипания A'. В NCTC 8325 штамме clfA представляет собой SAOUHSC_00812 и имеет аминокислотную последовательность SEQ ID NO:1 (GI:88194572). В Newman штамме он представляет собой nwmn_0756 (GI:151220968).
Полезные антигены clfA могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:1, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:1; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:1, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти clfA белки включают варианты SEQ ID NO:1. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:1. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:1, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:1. Может быть полезным исключение конечных 368 C-концевых аминокислот SEQ ID NO:1. Может быть полезным исключение первых 39 N-концевых аминокислот SEQ ID NO:1. В других фрагментах исключен один или более белковых доменов.
SEQ ID NO:2 является полезным фрагментом SEQ ID NO:1 ('ClfA40-559'). В этих фрагментах исключена длинная повторяющаяся область по направлению к C-концу SEQ ID NO:1.
clfB
Антиген 'clfB' аннотируется как 'фактор слипания B'. В NCTC 8325 штамме clfB представляет собой SAOUHSC_02963 и имеет аминокислотную последовательность SEQ ID NO:3 (Gl:88196585). В Newman штамме он представляет собой nwmn_2529 (GI:151222741).
Полезные антигены clfB могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:3, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:3; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:3, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти clfB белки включают варианты SEQ ID NO:3. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:3. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:3, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:3. Может быть полезным исключение конечных 40 C-концевых аминокислот SEQ ID NO:3. Может быть полезным исключение первых 44 N-концевых аминокислот SEQ ID NO:3. В других фрагментах исключен один или более белковых доменов. ClfB в его природном виде представляет собой длинный белок, и поэтому использование фрагментов является полезным, например, для очистки, манипулирования, слияния, экспрессии и т.п.
SEQ ID NO:4 является полезным фрагментом SEQ ID NO:3 ('ClfB45-552'). Этот фрагмент включает наиболее доступный домен ClfB, и его легче использовать в промышленном масштабе. Он также уменьшает схожесть антигена с человеческими белками. Другие полезные фрагменты, основанные на 3-доменной модели ClfB, включают: ClfB45-360 (также известный как CLfB-N12; SEQ ID NO:5); ClfB212-542 (также известный как CLfB-N23; SEQ ID NO:6), и ClfB360-542 (также известный как CLfB-N3; SEQ ID NO:7).
sdrE2
Антигены 'sdrE2' аннотируется как 'Ser-Asp-обогащенный фибриноген/костный сиалопротеин-связывающий белок SdrE'. В Newman штамме sdrE2 представляет собой NWNN_0525 и имеет аминокислотную последовательность SEQ ID NO:8 (GI:151220737).
Полезные sdrE2 антигены могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:8, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:8; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:8, где n имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sdrE2 белки включают варианты SEQ ID NO:8. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:8. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:8, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:8. Может быть полезным исключение конечных 38 C-концевых аминокислот SEQ ID NO:8. Может быть полезным исключение первых 52 N-концевых аминокислот SEQ ID NO:8. В других фрагментах исключен один или более белковых доменов. SdrE2 в его природном виде представляет собой длинный белок, и поэтому использование фрагментов является чрезвычайно полезным, например, для очистки, манипулирования, слияния, экспрессии и т.п.
SEQ ID NO:9 является полезным фрагментом SEQ ID NO:8 ('SdrE53-632'). Этот фрагмент включает наиболее доступный домен SdrE2, и его легче использовать в промышленном масштабе. Он также уменьшает схожесть антигена с человеческими белками.
sdrC
'Антиген sdrC' означает 'sdrC белок'. В NCTC 8325 штамме sdrC представляет собой SAOUHSC_00544 и имеет аминокислотную последовательность SEQ ID NO:10 (GI:88194324).
Полезные антигены sdrC могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:10, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:10; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:10, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sdrC белки включают варианты SEQ ID NO:10. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:10. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:10, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:10. Может быть полезным исключение конечных 38 C-концевых аминокислот SEQ ID NO:10. Может быть полезным исключение первых 50 N-концевых аминокислот SEQ ID NO:10. В других фрагментах исключен один или более белковых доменов. SdrC в его природном виде представляет собой длинный белок, и поэтому использование фрагментов является полезным, например, для очистки, манипулирования, слияния, экспрессии и т.п.
SEQ ID NO:11 является полезным фрагментом SEQ ID NO:10 ('SdrC51-518'). Этот фрагмент включает наиболее доступный домен SdrC, и его легче использовать в промышленном масштабе. Он также уменьшает схожесть антигена с человеческими белками.
sasF
'Антиген sasF' аннотируется как 'sasF белок'. В NCTC 8325 штамме sasF представляет собой SAOUHSC_02982 и имеет аминокислотную последовательность SEQ ID NO:12 (GI:88196601).
Полезные антигены sasF могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:12, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:12; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:12, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sasF белки включают варианты SEQ ID NO:12. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:12. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:12, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:12. Может быть полезным исключение конечных 39 C-концевых аминокислот SEQ ID NO:12. Может быть полезным исключение первых 37 N-концевых аминокислот SEQ ID NO:12. В других фрагментах исключен один или более белковых доменов.
emp
Антиген 'emp' аннотируется как 'связывающий белок внеклеточного матрикса и плазмы'. В NCTC 8325 штамме emp представляет собой SAOUHSC_00816 и имеет аминокислотную последовательность SEQ ID NO:13 (Gl:88194575). В Newman штамме он представляет собой nwmn_0758 (GI:151220970).
Полезные антигены emp могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:13, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:13; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:13, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти emp белки включают варианты SEQ ID NO:13. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:13. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:13, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:13. Может быть полезным исключение первых 26 N-концевых аминокислот SEQ ID NO:13. В других фрагментах исключен один или более белковых доменов.
SEQ ID NO:14, 15, 16 и 17 являются полезными фрагментами SEQ ID NO:13 ('Emp35-340', 'Emp27-334', 'Emp35-334' и 'Emp27-147', соответственно).
sdrD
Антиген 'sdrD' аннотируется как 'sdrD белок'. В NCTC 8325 штамм sdrD представляет собой SAOUHSC_00545 и имеет аминокислотную последовательность SEQ ID NO:18 (GI:88194325).
Полезные антигены sdrD могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:18, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше(например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:18; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:18, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sdrD белки включают варианты SEQ ID NO:18. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:18. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:18, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:18. Может быть полезным исключение конечных 38 C-концевых аминокислот SEQ ID NO:18. Может быть полезным исключение первых 52 N-концевых аминокислот SEQ ID NO:18. В других фрагментах исключен один или более белковых доменов. SdrD в его природном виде представляет собой длинный белок, и поэтому использование фрагментов является чрезвычайно полезным, например, для очистки, манипулирования, слияния, экспрессии и т.п.
SEQ ID NO:19 является полезным фрагментом SEQ ID NO:18 ('SdrD53-592'). Этот фрагмент включает наиболее доступный домен SdrD, и его легче использовать в промышленном масштабе. Он также уменьшает схожесть антигена с человеческими белками. Другим полезным фрагментом, с таким же C-концевым остатком, является SdrD394-592 (также известный, как SdrD-N3; SEQ ID NO:20).
spa
Антиген 'spa' аннотируется как 'белок A' или 'SpA'. В NCTC 8325 штамме spa представляет собой SAOUHSC_00069 и имеет аминокислотную последовательность SEQ ID NO:21 (GI:88193885). В Newman штамме он представляет собой nwmn_0055 (GI:151220267). Все S.aureus штаммы экспрессируют структурный ген для spa, хорошо охарактеризованный фактор вирулентности, закрепленный на клеточной оболочке поверхностный белковый продукт которого содержит пять высоко гомологичных иммуноглобулин-связывающих доменов, обозначенных как E, D, A, B и C [230]. Эти домены демонстрируют ~80% идентичность на аминокислотном уровне, включают 56-61 остатков в длину и организованы в виде тандемных повторов [231]. SpA синтезируется в виде белка-предшественника с N-концевым сигнальным пептидом и C-концевым сортинг-сигналом [232, 233]. Закрепленный на клеточной оболочке spa в большом количестве присутствует на поверхности стафилококков [234, 235]. Каждый из его иммуноглобулин-связывающих доменов состоит из антипараллельных α-спиралей, которые собраны в три пучка спиралей и могут связываться с Fc-доменом иммуноглобулина G (IgG) [236, 237], VH3 тяжелой цепью (Fab) IgM (т.е. B-клеточным рецептором) [238], фактором фон Виллебранда на его A1 домене [239] и/или TNF-α рецептором 1 (TNFR1) [240], который расположен на поверхностях эпителия дыхательных путей.
Полезные антигены spa могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:21, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:21; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:21, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти spa белки включают варианты SEQ ID NO:21. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:21. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:21, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:21. Может быть полезным исключение конечных 35 C-концевых аминокислот SEQ ID NO:21. Может быть полезным исключение первых 36 N-концевых аминокислот SEQ ID NO:21. В других фрагментах исключен один или более белковых доменов. Ссылочный документ 241 предполагает, что индивидуальные IgG-связывающие домены могут быть полезными иммуногенами, отдельно или в сочетании.
SEQ ID NO:22 является полезным фрагментом SEQ ID NO:21 ('Spa37-325'). Этот фрагмент содержит все пять SpA Ig-связывающих доменов и включает наиболее доступный домен SpA. Он также уменьшает схожесть антигена с человеческими белками. Другие полезные фрагменты могут не включать 1, 2, 3 или 4 из природных A, B, C, D и/или E доменов. Как сообщается в ссылочном документе 241, другие полезные фрагменты могут включать только 1, 2, 3 или 4 из природных A, B, C, D и/или E доменов, например, включают только SpA(A) домен, но не B-E, или включают только SpA(D) домен, но не A, B, C или E, и т.п. Таким образом, spa антиген, полезный в настоящем изобретении, может включать 1, 2, 3, 4 или 5 IgG-связывающих доменов, но в идеальном случае содержит 4 или меньше. Когда антиген включает только один тип spa домена (например, только Spa(A) или SpA(D) домен), он может включать более чем одну копию этого домена, например, несколько SpA(D) доменов в одной полипептидной цепи. Индивидуальный домен в антигене может быть мутирован по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислотам относительно SEQ ID NO:21 (например, см. ссылочный документ 241, раскрывающий мутации по остаткам 3 и/или 24 домена D, по остатку 46 и/или 53 домена A и т.п.). Такие мутанты не должны устранять способность антигена вызвать образование антитела, которое распознает SEQ ID NO:21, но могут устранять связывание антигена с IgG. В некоторых аспектах spa антиген включает такое замещение, как (a) одна или более аминокислотных замен в IgG Fc-связывающем суб-домене SpA домена A, B, C, D и/или E, которое разрушает или уменьшает связывание с IgG Fc, и (b) одна или более аминокислотных замен в VH3-связывающем суб-домене SpA домена A, B, C, D, и/или E, которое разрушает или уменьшает связывание с VH3. В некоторых вариантах воплощения вариант SpA включает по меньшей мере или не более чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или больше пептидов домена D варианта SpA.
esaC
'esaC' антиген аннотируется как 'esaC'. В NCTC 8325 штамме esaC представляет собой SAOUHSC_00264 и имеет аминокислотную последовательность SEQ ID NO:23 (GI:88194069).
Полезные антигены esaC могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:23, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:23; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:23, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100 или больше). Эти esaC белки включают варианты SEQ ID NO:23. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:23. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:23, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:23. В других фрагментах исключен один или более белковых доменов.
esxA
'esxA' антиген аннотируется как 'белок'. В NCTC 8325 штамме esxA представляет собой SAOUHSC_00257 и имеет аминокислотную последовательность SEQ ID NO:24 (GI:88194063).
Полезные антигены esxA могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:24, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:24; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:24, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90 или больше). Эти esxA белки включают варианты SEQ ID NO:24. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:24. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:24, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:24. В других фрагментах исключен один или более белковых доменов.
esxB
'esxB' антиген аннотируется как 'esxB'. В NCTC 8325 штамме esxB представляет собой SAOUHSC_00265 и имеет аминокислотную последовательность SEQ ID NO:25 (GI:88194070).
Полезные антигены esxB могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:25, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:25; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:25, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100 или больше). Эти esxB белки включают варианты SEQ ID NO:25. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:25. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:25, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:25. В других фрагментах исключен один или более белковых доменов.
sta006
'sta006' антиген аннотируется как 'феррихром-связывающий белок', и он также был указан как 'FhuD2' в литературе [242]. В NCTC 8325 штамме sta006 представляет собой SAOUHSC_02554 и имеет аминокислотную последовательность SEQ ID NO:26 (GI:88196199). В Newman штамме он представляет собой nwmn_2185 (GI:151222397).
Полезные антигены sta006 могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:26, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:26; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:26, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sta006 белки включают варианты SEQ ID NO:26. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:26. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:26, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:26. Может быть полезным исключение первых 17 N-концевых аминокислот SEQ ID NO:26. В других фрагментах исключен один или более белковых доменов. Мутантные формы sta006 описаны в ссылочном документе 243. Антиген sta006 может быть липидированным, например, с ацилированным N-концевым цистеином.
isdC
'isdC' антиген аннотируется как 'белок'. В NCTC 8325 штамме isdC представляет собой SAOUHSC_01082 и имеет аминокислотную последовательность SEQ ID NO:27 (GI:88194830).
Полезные антигены isdC могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:27, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:27; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:27, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200 или больше). Эти isdC белки включают варианты SEQ ID NO:27. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:27. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:27, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:27. Может быть полезным исключение конечных 39 C-концевых аминокислот SEQ ID NO:27. Может быть полезным исключение первых 28 N-концевых аминокислот SEQ ID NO:27. В других фрагментах исключен один или более белковых доменов. Полезные фрагменты IsdB раскрыты в ссылочном документе 249.
Ссылочный документ 244 раскрывает антигены, которые включают полезные эпитопы как из IsdB, так и из IsdH.
Hla
Антиген 'Hla' представляет собой 'альфа-гемолизиновый предшественник', также известный как 'альфа токсин' или просто 'гемолизин'. В NCTC 8325 штамме Hla представляет собой SAOUHSC_01121 и имеет аминокислотную последовательность SEQ ID NO:28 (GI:88194865). В Newman штамме он представляет собой nwmn_1073 (GI:151221285). Hla представляет собой важную детерминанту вирулентности, продуцируемую большинством штаммов S.aureus, обладающую порообразующей и гемолитической активностью. Анти-Hla антитела могут нейтрализовать вредные эффекты токсина в животных моделях, и Hla является особенно полезным для зашиты от пневмонии.
Полезные антигены Hla могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:28, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:28; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:28, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти Hla белки включают варианты SEQ ID NO:28. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:28. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:28, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:28. Может быть полезным исключение первых 26 N-концевых аминокислот SEQ ID NO:28. Усечение по C-концу также можно использовать, например, оставляя только 50 аминокислот (остатки 27-76 SEQ ID NO:28) [245]. В других фрагментах исключен один или более белковых доменов.
Hla токсичность может быть исключена в композициях по настоящему изобретению путем химической инактивации (например, с использованием формальдегида, глутаральдегида или других образующих поперечные связи реагентов). Вместо этого, однако, предпочтительно использовать мутантные формы Hla, которые устраняют его токсическую активность, сохраняя при этом его иммуногенность. Такие детоксифицированные мутанты уже известны из уровня техники. Один полезный антиген Hla содержит мутацию по остатку 61 SEQ ID NO:28, который представляет собой остаток 35 зрелого антигена (т.е. после исключения первых 26 N-концевых аминокислот). Таким образом, остаток 61 может не быть гистидином, а вместо этого может представлять собой, например, Ile, Val или предпочтительно Leu. His-Arg мутацию в этом положении также можно использовать. Например, SEQ ID NO:29 представляет собой последовательность зрелого мутанта Hla-H35L, и полезный антиген Hla включает SEQ ID NO:29. Другая полезная мутация заменяет длинную петлю короткой последовательностью, например, для замены 39мера по остаткам 136-174 SEQ ID NO:28 тетрамером, таким как PSGS (SEQ ID NO:30), как в SEQ ID NO:31 (которая также включает H35L мутацию) и SEQ ID NO:32 (которая не включает H35L мутацию).
Дополнительные полезные антигены Hla раскрыты в ссылочных документах 246 и 247.
SEQ ID NO:33, 34 и 35 представляют собой три полезных фрагмента SEQ ID NO:28 ('Hla27-76', 'Hla27-89' и 'Hla27-79', соответственно). SEQ ID NO:36, 37 и 38 представляют собой соответствующие фрагменты из SEQ ID NO:29.
sta011
Антиген 'sta011' аннотируется как 'липопротеин'. В NCTC 8325 штамме sta011 представляет собой SAOUHSC_00052 и имеет аминокислотную последовательность SEQ ID NO:39 (GI:88193872).
Полезные антигены sta011 могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:39, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:39; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:39, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sta011 белки включают варианты SEQ ID NO:39. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:39. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:39, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:39. Может быть полезным исключение первых 23 N-концевых аминокислот SEQ ID NO:39. В других фрагментах исключен один или более белковых доменов. Антиген sta006 может быть липидированным, например, с ацилированным N-концевым цистеином.
Вариантные формы SEQ ID NO:39, которые можно использовать для получения антигенов sta011, включают, но не ограничиваются этим, SEQ ID NO:40, 41 и 42 с различными Ile/Val/Leu заменами.
isdA
Антиген 'isdA' аннотируется как 'IsdA белок'. В CTC 8325 штамме isdA представляет собой SAOUHSC_01081 и имеет аминокислотную последовательность SEQ ID NO:43 (GI:88194829). В Newman штамме он представляет собой nwmn_1041 (GI:151221253).
Полезные антигены isdA могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:43, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:43; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:43, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти isdA белки включают варианты SEQ ID NO:43. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:43. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:43, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:43. Может быть полезным исключение конечных 38 C-концевых аминокислот SEQ ID NO:43. Может быть полезным исключение первых 46 N-концевых аминокислот SEQ ID NO:43. Усечение для исключения C-концевого 38мера SEQ ID NO:43 (начиная с LPKTG мотива) также является полезным. В других фрагментах исключен один или более белковых доменов.
SEQ ID NO:44 является полезным фрагментом SEQ ID NO:43 (аминокислоты 40-184 SEQ ID NO:43; 'IsdA40-184'), который включает гемм-связывающий сайт природного белка и включает наиболее доступный домен антигена. Он также уменьшает схожесть антигена с человеческими белками. Другие полезные фрагменты раскрыты в ссылочных документах 248 и 249.
IsdA не обладает хорошей адсорбцией на алюминийгидроксидных адъювантах, поэтому IsdA, присутствующий в композиции, может быть неадсорбированным или может быть адсорбирован на альтернативном адъюванте, например фосфате алюминия.
isdB
Антиген 'isdB' аннотируется как 'нейрофиламентный белок isdB'. В NCTC 8325 штамме isdB представляет собой SAOUHSC_01079 и имеет аминокислотную последовательность SEQ ID NO:45 (GI:88194828). IsdB был предложен для использования в качестве вакцинного антигена как таковой [250], но это может не предотвращать пневмонию.
Полезные антигены isdB могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:45, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:45; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:45, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти isdB белки включают варианты SEQ ID NO:45. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:45. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:45, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:45. Может быть полезным исключение конечных 36 C-концевых аминокислот SEQ ID NO:45. Может быть полезным исключение первых 40 N-концевых аминокислот SEQ ID NO:45. В других фрагментах исключен один или более белковых доменов. Полезные фрагменты IsdB раскрыты в ссылочных документах 249 и 251, например, не содержащие 37 аминокислот внутренней области SEQ ID NO:45.
В некоторых вариантах воплощения композиции по настоящему изобретению не включают isdB антиген.
sta073
Антиген 'sta073' аннотируется как 'бифункциональный аутолизиновый предшественник'. В NCTC 8325 штамме sta073 представляет собой SAOUHSC_00994 и имеет аминокислотную последовательность SEQ ID NO:46 (GI:88194750). В Newman штамме он представляет собой nwmn_0922 (GI:151221134). С использованием протеомного анализа было обнаружено, что этот белок секретируется или является поверхностно-экспонированным.
Полезные антигены sta073 могут вызвать образование антитела (например, при введении человеку), которое распознает SEQ ID NO:46, и/или могут включать аминокислотную последовательность: (a) имеющую идентичность 50% или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или больше) с SEQ ID NO:46; и/или (b) включающую фрагмент из, по меньшей мере, 'n' последовательных аминокислот из SEQ ID NO:46, где 'n' имеет значение 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или больше). Эти sta073 белки включают варианты SEQ ID NO:46. Предпочтительные фрагменты (b) включают эпитоп из SEQ ID NO:46. Другие предпочтительные фрагменты не включают одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из C-конца и/или одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или больше) из N-конца SEQ ID NO:46, с сохранением при этом по меньшей мере одного эпитопа SEQ ID NO:46. Может быть полезным исключение первых 24 N-концевых аминокислот SEQ ID NO:46. В других фрагментах исключен один или более белковых доменов.
Sta073 не обладает хорошей адсорбцией на алюминийгидроксидных адъювантах, поэтому Sta073, присутствующий в композиции, может быть неадсорбированным или может быть адсорбирован на альтернативном адъюванте, например, фосфате алюминия.
Гибридные полипептиды
Белковые антигены S.aureus, используемые в настоящем изобретении, могут присутствовать в композиции в качестве индивидуальных отдельных полипептидов. Однако когда используют более чем один антиген, они не обязательно могут присутствовать в качестве отдельных полипептидов. Вместо этого, по меньшей мере, два (например, 2, 3, 4, 5 или больше) антигена могут экспрессироваться в виде одной полипептидной цепи ('гибридный' полипептид). Гибридные полипептиды обладают двумя основными преимуществами: во-первых, полипептиду, который может быть нестабильным или плохо экспрессируется как таковой, можно помочь путем добавления подходящего гибридного партнера, который разрешает эту проблему; во-вторых, коммерческое производство упрощается, поскольку необходимо использовать только одну экспрессию и очистку для получения двух полипептидов, которые оба являются антигенно полезными.
Гибридный полипептид может включать две или более полипептидных последовательностей из каждого из антигенов, перечисленных выше, или два или более вариантов одного и того же антигена в случаях, когда последовательность имеет частичную вариабельность в различных штаммах.
Гибриды, состоящие из аминокислотных последовательностей из двух, трех, четырех, пяти, шести, семи, восьми, девяти или десяти антигенов, являются полезными. В частности, гибриды, состоящие из аминокислотных последовательностей из двух, трех, четырех или пяти антигенов, являются предпочтительными, такими как два или три антигена.
Различные гибридные полипептиды можно смешать вместе в одну композицию. Гибриды можно объединить с негибридными антигенами, выбранными из первой, второй или третьей групп антигенов. В таких сочетаниях антиген может присутствовать в более чем одном гибридном полипептиде и/или в виде негибридного полипептида. Однако предпочтительно, когда антиген присутствует либо в виде гибрида, либо в виде негибрида, но не в том и в другом виде.
Гибридные полипептиды могут быть представлены формулой NH2-A-{-X-L-}n-B-COOH, где: X представляет собой аминокислотную последовательность антигена S.aureus, описанную выше; L представляет собой необязательную линкерную аминокислотную последовательность; А представляет собой необязательную N-концевую аминокислотную последовательность; B представляет собой необязательную C-концевую аминокислотную последовательность; n представляет собой целое число, имеющее значение 2 или больше (например, 2, 3, 4, 5, 6 и т.п.). Обычно n имеет значение 2 или 3.
Когда -X- группа содержит лидерную пептидную последовательность в форме ее дикого типа, она может быть включена или не включена в гибридный белок. В некоторых вариантах воплощения лидерные пептиды должны быть исключены, но только не пептиды -X- группы, расположенной по N-концу гибридного белка, т.е. лидерный пептид X1 должен быть сохранен, но лидерную пептиды X2... Xn исключаются. Это является эквивалентным исключению всех лидерных пептидов и использованию лидерного пептида X1 как группы -A-.
Для каждых n случаев {-X-L-}, линкерная аминокислотная последовательность -L- может присутствовать или отсутствовать. Например, когда n=2, гибрид может представлять собой NH2-X1-L1-X2-L2-COOH, NH2-X1-X2-COOH, NH2-X1-L1-X2-COOH, NH2-X1-X2-L2-COOH и т.п. Линкерная аминокислотная последовательность (последовательности) -L- типично является короткой (например, 20 или меньше аминокислот, т.е. 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2, 1). Примеры включают короткие пептидные последовательности, которые облегчают клонирование, полиглициновые линкеры (т.е. включающие Glyn, где n=2, 3, 4, 5, 6, 7, 8, 9, 10 или больше) и гистидиновые метки (т.е. Hisn, где n=3, 4, 5, 6, 7, 8, 9, 10 или больше). Другие подходящие линкерные аминокислотные последовательности должны быть очевидными для специалистов в данной области. Полезный линкер представляет собой GSGGGG (SEQ ID NO:47) или GSGSGGGG (SEQ ID NO:48), где Gly-Ser дипептид образован из BamHl сайта рестрикции, способствуя, таким образом, клонированию и манипуляции, и (Gly)4 тетрапептид является типичным поли-глициновым линкером. Другие подходящие линкеры, в частности, для использования в качестве конечных Ln, представляют собой ASGGGS (SEQ ID NO:49, например, кодируемый SEQ ID NO:50) или Leu-Glu дипептид.
-A- представляет собой необязательную N-концевую аминокислотную последовательность. Она типично является короткой (например, 40 или меньше аминокислот, т.е. 40, 39, 38, 37, 36, 35, 34, 33, 32, 31, 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2, 1). Примеры включают лидерные последовательности для направления траффика белка, или короткие пептидные последовательности, которые способствуют клонированию или очистке (например, гистидиновые метки, т.е. Hisn, где n=3, 4, 5, 6, 7, 8, 9, 10 или больше). Другие подходящие N-концевые аминокислотные последовательности должны быть очевидными для специалистов в данной области. Когда X1 не содержит свой собственный N-концевой метионин, -A- предпочтительно представляет собой олигопептид (например, с 1, 2, 3, 4, 5, 6, 7 или 8 аминокислотами), который обеспечивает N-концевой метионин, например, Met-Ala-Ser, или просто Met остаток.
-B- представляет собой необязательную C-концевую аминокислотную последовательность. Она типично является короткой (например, 40 или меньше аминокислот, т.е. 39, 38, 37, 36, 35, 34, 33, 32, 31, 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2, 1). Примеры включают последовательности для направления траффика белка, короткие пептидные последовательности, которые способствуют клонированию или очистке (например, включающие гистидиновые метки, т.е. Hisn, где n=3, 4, 5, 6, 7, 8, 9, 10 или больше, такие как SEQ ID NO:51), или последовательности, которые повышают стабильность белка. Другие подходящие C-концевые аминокислотные последовательности должны быть очевидными для специалистов в данной области.
Один гибридный полипептид по настоящему изобретению может включать антигены как EsxA, так и esxB. Они могут быть расположены в любом порядке, от N- к C-концу. Последовательности SEQ ID NO:52 ('EsxAB'; кодируемый SEQ ID NO:53) и 54 ('EsxBA') представляют собой примеры таких гибридов, и обе содержат гексапептидные линкеры ASGGGS (SEQ ID NO:49).
Общая информация
При осуществлении настоящего изобретения используют, если не указано иное, традиционные способы химии, биохимии, молекулярной биологии, иммунологии и фармакологии, известные специалистам в данной области. Такие процедуры подробно описаны в литературе. См., например, ссылочные документы 252-259 и т.п.
"GI" нумерация использована выше. GI номер, или "GenInfo Идентификатор", представляет собой ряд цифр, присваиваемых последовательно каждой записываемой последовательности, обработанной NCBI, когда последовательности добавляют в его базы данных. GI номер не имеет никакого сходства с номером доступа зарегистрированной последовательности. Когда последовательность обновляется (например, для коррекции или для добавления дополнительной характеристики или информации), тогда она получает новый GI номер. Таким образом, последовательность, ассоциированная с определенным GI номером, никогда не меняется.
При указании процента идентичности последовательностей между двумя аминокислотными последовательностями подразумевается, что при их выравнивании указанный процент аминокислот одинаковый при сравнении этих двух последовательностей. Это выравнивание и процент гомологии или идентичность последовательностей можно определить с использованием программ, известных из уровня техники, например, которые описаны в разделе 7.7.18 ссылочного документа 260. Предпочтительное выравнивание определяют при помощи алгоритма поиска гомологии Смита-Ватермана с использованием поиска аффинных гэпов, со штрафом за раскрытие гэпа 12 и штрафом за удлинение гэпа 2, BLOSUM матрицы 62. Алгоритм поиска гомологии Смита-Ватермана раскрыт в ссылочном документе 261.
Когда изобретение касается "эпитопа", этот эпитоп может быть B-клеточным эпитопом и/или T-клеточным эпитопом. Такие эпитопы можно идентифицировать эмпирическим путем (например, с использованием PEPSCAN [262, 263] или подобных способов), или они могут быть предвиденными (например, с использованием антигенного индекса Джеймсона-Вольфа [264], подходов на основе матрицы [265], MAPITOPE [266], TEPITOPE [267, 268], нейросетей [269], OptiMer & EpiMer [270, 271], ADEPT [272], Tsites [273], гидрофильности [274], антигенного индекса [275] или способов, раскрытых в ссылочных документах 276-280 и т.п.). Эпитопы представляют собой части антигена, которые распознаются и связываются с антиген-связывающими сайтами антител или T-клеточных рецепторов, и они также могут быть указаны как "антигенные детерминанты".
Когда исключается "домен" антигена, это может включать исключение сигнального пептида, цитоплазматического домена, трансмембранного домена, внеклеточного домена и т.п.
Термин "включающий" охватывает как "включающий", так и "состоящий из", например композиция, "включающая" X, может состоять исключительно из X или может включать что-либо дополнительно, например, X+Y.
Термин "около", используемый вместе с числовым значением x, означает, например, x+10%.
Термин "по существу" не исключает "полностью", например композиция, которая "по существу не содержащая" Y, может быть полностью свободна от Y. Где это необходимо, термин "по существу" могут быть опущен из определения изобретения.
Когда изобретение обеспечивает способ, включающий несколько последовательных стадий, изобретение также может обеспечивать способ, включающий меньше чем общее количество стадий. Например, изобретение обеспечивает способ, включающий следующие стадии: (a) деполимеризация капсульного полисахарида S.aureus типа 5 или типа 8 с получением полисахаридного фрагмента; и (b) окисление фрагмента для введения альдегидной группы в, по меньшей мере, один сахаридный остаток во фрагменте с получением окисленного сахаридного остатка. Дополнительную стадию (c) осуществлять необязательно, чтобы способ попадал в объем настоящего изобретения, поскольку продукт стадий (a) и (b) имеет применение в качестве промежуточного соединения в получении конъюгата, и его можно использовать, хранить, экспортировать и т.п. для отдельного и последующего использования. Такое последующее использование может включать осуществление стадии (c). Альтернативно, продукт стадий (a) и (b) можно объединить с молекулой носителя различными способами, например, через гидроксильную или карбоксильную группу, которая сохраняется в окисленном сахаридном остатке.
Подобным образом, когда исходное полисахаридное вещество уже частично обработано, тогда изобретение охватывает способы, включающие только последние стадии способа. Например, изобретение охватывает способ, включающий стадию связывания окисленного сахаридного остатка в капсульном полисахариде типа 5 или типа 8 с молекулой носителя через альдегидную группу, где исходное вещество для такого способа представляет собой капсульный полисахарид типа 5 или типа 8, который был предварительно деполимеризован с получением полисахаридного фрагмента и затем окислен для введения альдегидной группы в сахаридный остаток.
Эти различные стадии могут быть осуществлены в очень разное время разными людьми в разных местах (например, в разных странах).
Должно быть понятно, что кольца сахаров могут существовать в открытой и закрытой форме, и что, хотя в структурных формулах, представленных в настоящей заявке, показаны закрытые формы, открытые формы также охватываются настоящим изобретением. Подобным образом, должно быть понятно, что сахара могут существовать в пиранозной и фуранозной формах, и что, хотя в структурных формулах, представленных в настоящей заявке, показаны пиранозные формы, охватываются также фуранозные формы. Также охватываются различные аномерные формы сахаров.
Первичный амин может быть представлен формулой NH2R. R группа типично является электрон-донорной и включает C1-8углеводород, в частности, C1-8алкил, особенно метил. R часто представляет собой -CH3, -C2H5 или -C3H7. Углеводородная группа может быть замещена одной или несколькими группами, такими как: галоген (например, Cl, Br, F, I), тригалогенметил, -NO2, -CN, -N+(C1-6алкил)2О-, -SO3H, -SOC1-6алкил, -SO2C1-6алкил, -SO3C1-6алкил, -OC(=О)OCl-6алкил, -C(=О)H, -C(=О)C1-6алкил, -OC(=О)C1-6алкил, -N(C1-6алкил)2, C1-6алкил, -N(C1-6алкил)2, C(=О)N(C1-6алкил)2, -N(C1-6алкил)C(=О)О(C1-6алкил), -N(C1-6алкил)C(=О)N(C1-6алкил)2, -CO2H, -OC(=О)N(C1-6алкил)2, -N(C1-6алкил)C(=О)C1-6алкил, -N(C1-6алкил)C(=S)C1-6алкил, -N(C1-6алкил)SO2N(C1-6алкил)2, -CO2C1-6алкил, -SO2N(C1-6алкил)2, -C(=О)NH2, -C(=S)N(C1-6алкил)2, -N(C1-6алкил)SO2C1-6алкил, -N(C1-6алкил)C(=S)N(C1-6алкил)2, -NH-C1-6алкил, -S-C1-6алкил или -О-C1-6алкил. Термин 'углеводород' включает линейные, разветвленные или циклические одновалентные группы, состоящие из углерода и водорода. Углеводородные группы, таким образом, включают алкильную, алкенильную и алкинильную группы, циклоалкильную (включая полициклоалкильную), циклоалкенильную и арильную группы, а также их сочетания, например, алкилциклоалкильную, алкилполициклоалкильную, алкиларильную, алкениларильную, циклоалкиларильную, циклоалкениларильную, циклоалкилалкильную, полициклоалкилалкильную, арилалкильную, арилалкенильную, арилциклоалкильную и арилциклоалкенильную группы. Типичные углеводородные группы представляют собой C1-14 углеводородную группу, более конкретно C1-8 углеводородную группу.
Краткое описание фигур
Фиг. 1 представляет схему получения конъюгата капсульный полисахарид S.aureus типа 5-CRM197 с использованием линкера, представляющего собой дигидразин адипиновой кислоты, и химии с использованием карбодиимида.
Фиг. 2a представляет SDS-PAGE анализ конъюгата капсульный полисахарид S.aureus типа 5-CRM197, полученного с использованием линкера, представляющего собой дигидразин адипиновой кислоты, и химии с использованием карбодиимида. Фиг. 2b представляет S300 Sephacryl™ хроматограмму конъюгата капсульный полисахарид S.aureus типа 5-CRM197, полученного с использованием линкера, представляющего собой дигидразин адипиновой кислоты, и химии с использованием карбодиимида.
Фиг. 3 представляет S300 Sephacryl™ хроматограмму деполимеризованного капсульного полисахарида типа 5.
Фиг. 4A сравнивает ID 1Н сигналы в 1Н аномерной области для деполимеризованного и природного капсульного полисахарида типа 5. Отмечены некоторые заметные отличия. Фиг. 4B и 4C сравнивает аномерную и метил-фукозную области, соответственно, 2D (1Н, 1Н) скалярных спектров связывания для этих полисахаридов. Отмечены некоторые заметные отличия.
Фиг. 5a представляет SDS-PAGE анализ конъюгата капсульный полисахарид S.aureus типа 5-CRM197, полученного с использованием способа по настоящему изобретению. Фиг. 5b представляет S300 Sephacryl™ хроматограмму конъюгата капсульный полисахарид S.aureus типа 5-CRM197, полученного с использованием способа по настоящему изобретению.
Фиг. 6 представляет S300 Sephacryl™ хроматограмму капсульных полисахаридов S.aureus типа 5, деполимеризованных в разных условиях.
Фиг. 7 представляет SDS-PAGE анализы конъюгатов капсульный полисахарид S.aureus типа 5-CRM197, полученных с использованием способов по настоящему изобретению.
Фиг. 8 представляет IgG ответ на различные антигены в модели абсцесса почки мыши, вызванного S.aureus инфекцией.
Фиг. 9 сравнивает IgG и IgM ответы на различные антигены в модели абсцесса почки мыши.
Фиг. 10 представляет защитные ответы на различные антигены в модели абсцесса почки мыши.
Фиг. 11 сравнивает IgG ответ на различные конъюгаты в модели абсцесса почки мыши.
Фиг. 12 сравнивает защитные ответы на различные конъюгаты в модели абсцесса почки мыши.
Фиг. 13A и 13B сравнивают защитные ответы на другие конъюгаты в модели абсцесса почки мыши.
Фиг. 14 сравнивает IgG и IgM ответы на другие конъюгаты в модели абсцесса почки мыши.
Фиг. 15 сравнивает защитные ответы на еще один конъюгат в сочетании с различными средствами в модели абсцесса почки мыши.
Фиг. 16 сравнивает ответы на различные антигены в мышиной летальной модели инфекции S.aureus.
Фиг. 17 представляет SEC-ВЭЖХ хроматограмму капсульного полисахарида S.aureus типа 8, деполимеризованного при помощи 2M хлористоводородной кислоты.
Фиг. 18 представляет ЯМР-спектр капсульного полисахарида S.aureus типа 8, деполимеризованного при помощи 2M хлористоводородной кислоты.
Способы осуществления изобретения
Получение и характеристика конъюгата
Очищенный капсульный полисахарид S.aureus типа 5 конъюгировали с CRM 197 с использованием химии на основе карбодиимида и линкера, представляющего собой дигидразин адипиновой кислоты, аналогично способу, используемому в ссылочном документе 2 (см. ниже). В этом способе капсульный полисахарид конъюгируют с дериватизированным CRM197 с использованием EDC (Фиг. 1). Реакция включает карбоксильные группы капсульного полисахарида. Карбодиимид (EDC) активирует карбоксильные группы для связывания с -NH2 группой из дериватизированного белка-носителя (CRMadh) с образованием амидной связи. Дериватизированный CRMadh получали с использованием такой же химии на основе карбодиимида.
Получение CRMadh:
К раствору CRM197 добавляли 100 мМ MES pH6,0 буфера для достижения конечной концентрации 10-12 мг/мл. Затем добавляли 3,5 мг/мл ADH (дигидразида адипиновой кислоты) и 0,15 (EDC/CRM, масс./масс.) и реакцию осуществляли при слабом перемешивании в течение 1 часа при комнатной температуре. Смесь затем диализовали против первого 200 мМ NaCl, 10 мМ MES pH7,3 буфера и затем против 5 мМ MES pH7,0 буфера с использованием 6-8 кДа мембраны (SpectraPor). Продукт был охарактеризован при помощи MicroBCA, SDS-Page (3-8%), ВЭЖХ и MS. Было обнаружено, что CRMadh был дериватизирован 6-8 линкером ADH.
Реакция конъюгации:
Реакцию конъюгации осуществляли при концентрации капсульного полисахарида 2 мг/мл в 50 мМ MES буфере pH6,04. Дериватизированный белок-носитель, CRMadh, добавляли к раствору капсульного полисахарида до конечной концентрации 4,0 мг/мл. Раствор выдерживали при комнатной температуре в течение 3 часов. Отношение полисахарид:белок в реакционной смеси составляло 1:2 (масс./масс.), отношение полисахарид:EDC составляло 1:6,66 (эквивалент/эквивалент), и отношение полисахарид:сульфоNHS составляло 1:0,53 (эквивалент/эквивалент).
Через 3 часа образование конъюгата проверяли при помощи SDS-PAGE с использованием NuPAGE® 3-8% трис-ацетатного геля (Invitrogen) (Фиг. 2a). После конъюгации конъюгат очищали при помощи гель-фильтрационной хроматографии (осуществляли на Akta™ системе (G&E Healthcare) с использованием S300 Sephacryl™ смолы (G&E Healthcare), с 10 мМ NaPi, 10 мМ NaCl, pH7,2 буфер подвижной фазы). Детекцию конъюгата осуществляли при 215 нм, 254 нм и 280 нм (Фиг. 2b). Раствор конъюгата хранили при -20°C до последующего использования. Общее содержание сахарида в конъюгате определяли при помощи HPAEC-PAD анализа, а содержание белка при помощи микроВСА анализа, как описано в ссылочном документе 281 (таблица 1).

Очищенные капсульные полисахариды S.aureus типа 5 и типа 8 отдельно конъюгировали с CRM197 с использованием способа по настоящему изобретению (см. ниже).
Деполимеризация
Очищенный капсульный полисахарид растворяли в дистиллированной воде при 2 мг/мл. Добавляли уксусную кислоту до конечной концентрации 2% (об./об.) и реакционную смесь выдерживали при 90°C в течение 3 часов (или в течение ночи в случае партии B). Раствор затем нейтрализовали при помощи 1M NaOH, и деполимеризованный полисахарид очищали на колонке для гель-фильтрации (осуществляли на Akta™ системе (G&E Healthcare) с использованием S300 Sephacryl™ смолы (G&E Healthcare), с 10 мМ NaPi, 10 мМ NaCl, pH 7,2 буфером подвижной фазы). Детекцию сахарида осуществляли при 215 нм (Фиг. 3). Объединенные фракции диализовали против дистиллированной воды с использованием 1 кДа мембраны (SpectraPor) и лиофилизировали.
Сайт расщепления, как было обнаружено с использованием 1Н ЯМР, находится в положении (1→3) гликозидных связей в полисахариде типа 5. Вкратце, образцы природного и деполимеризованного капсульного полисахарида типа 5 лиофилизировали для удаления протонированного водного растворителя и растворяли в оксиде дейтерия (99,9% дейтерия, Sigma-Aldrich). Все спектры ЯМР записывали при 50°C на спектрометре Bruker Avance III 400 МГц с использованием 5-мм широкополостного зонда и пакета программ TopSpin 2.1 (Bruker) для сбора и обработки данных. 1D 1Н спектры собирали с использованием стандартного одноимпульсного эксперимента при ширине спектра 4000 Гц и со сбором 32k точек данных. Трансмиттер был установлен на остаточную HDO частоту (4,79 м.д.). Спектры получали количественно с использованием общего время рецикла для обеспечения полного восстановления каждого сигнала (5x Время Продольной Релаксации T1). Спектры обрабатывали, осуществляя преобразование Фурье, после применения 0,2 Гц функции расширения линии. 2D (1Н, 1Н) скалярные корреляционные спектры регистрировали при помощи DQF-COSY последовательности импульсов. Собирали 4096 точек данных в F2 домене и 256 в F1 домене.
ID 1Н сигналы для природного полисахарида сравнивали с опубликованными значениями, и было обнаружено, что они согласуются (таблица 2).
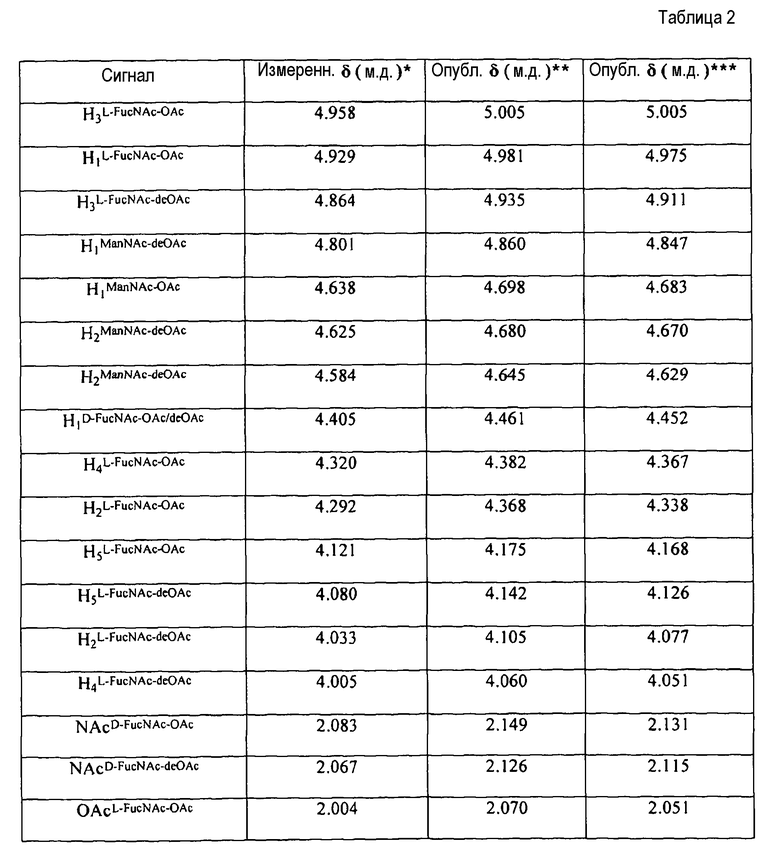

** Jones C. Carbohydr. Res. 2005, 340(6), 1097-1106 - HDO сигнал при 4,484 м.д.
*** Jones C. Carbohydr. Res. 2005, 340(6), 1097-1106 - H1L-FucNAc = 5,005 м.д., поэтому HDO сигнал при 4,532 м.д. вместо 4,484 м.д.
Фиг. 4A сравнивает ID 1Н сигналы в 1Н аномерной области для деполимеризованного и природного полисахаридов. Фиг. 4B и 4C сравнивают аномерную и метил-фукозную области, соответственно, 2D (1Н, 1Н) скалярных спектров связывания для этих полисахаридов. Данные показывают, что обработка уксусной кислотой приводила к расщеплению (1→3) гликозидных связей между α-L-FucNAc(3OAc) и β-D-FucNAc остатками в капсульном полисахариде типа 5.
Окисление
Деполимеризованный капсульный полисахарид растворяли в дистиллированной воде при 2 мг/мл. Добавляли NaIO4 при отношении полисахарид:NaIO4 1:1 (масс./масс.) и реакционную смесь выдерживали при комнатной температуре в течение 1-2 часов в темноте. Раствор затем диализовали против дистиллированной воды с использованием 1кДа мембраны (SpectraPor) и лиофилизировали снова.
Конъюгация
Окисленный капсульный полисахарид растворяли в 200 мМ NaPi, 1M NaCl, pH7,2 буфере при концентрации 10 мг/мл. К раствору добавляли CRM197 при отношении полисахарид:белок 4:1 (масс./масс.) и добавляли NaBH3CN (Aldrich) при отношении сахарид:NaBCNH3 2:1 (масс./масс.). Раствор выдерживали при 37°C в течение 2 дней. SDS-PAGE использовали для подтверждения образования конъюгата (см. Фиг. 5a для типа 5 конъюгата). После конъюгации конъюгат очищали гель-фильтрационной хроматографией (осуществляли на Akta™ системе (G&E Healthcare) с использованием S300 Sephacryl™ смолы (G&E Healthcare), с 10 мМ NaPi, 10 мМ NaCl, pH7,2 буфером подвижной фазы). Детекцию конъюгата осуществляли при 215 нм, 254 нм и 280 нм (см. Фиг. 5b для типа 5 конъюгата). Раствор конъюгата хранили при -20°C до последующего использования. Общее содержание сахарида в конъюгате определяли при помощи HPAEC-PAD анализа, а содержание белка при помощи микроВСА анализа (см. таблицу 3a для типа 5 конъюгата и таблицу 3b для типа 8 конъюгата).
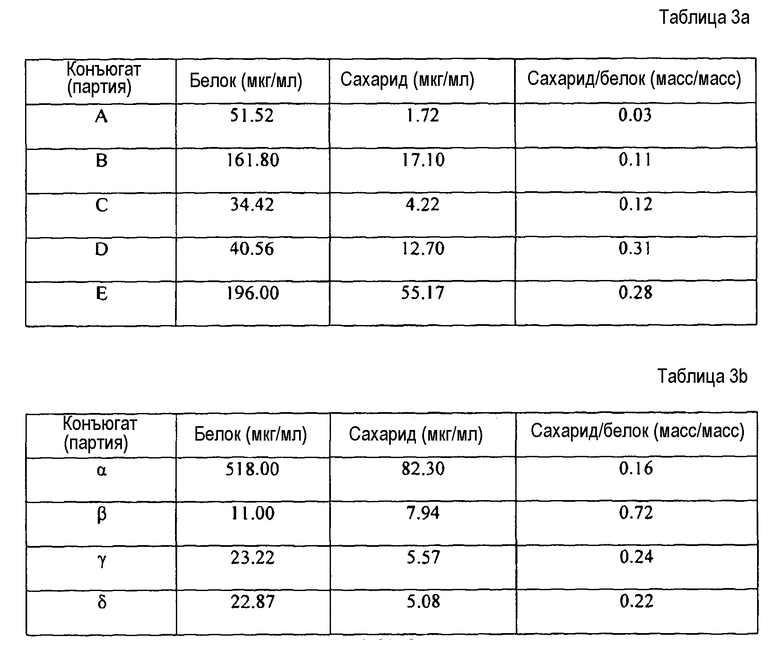
Очищенный S.aureus типа 5 конъюгировали с CRM197 с использованием другого способа по настоящему изобретению. В этом способе стадии деполимеризации, окисления и конъюгации осуществляли, как описано выше, за исключением того, что стадию конъюгации осуществляли с дериватизированным белком-носителем, описанным выше (CRMadh), вместо CRM197. Общее содержание сахарида в конъюгате определяли при помощи HPAEC-PAD анализа, а содержание белка при помощи МикроВСА анализа (таблица 4).

Альтернативные способы деполимеризации
В других исследованиях испытывали различные условия для деполимеризации очищенного капсульного полисахарида. Полисахарид растворяли в дистиллированной воде при 2 мг/мл. Добавляли уксусную кислоту до конечной концентрации 2% или 5% (об./об.) и реакционную смесь выдерживали при 90°C в течение 30 минут, 3 часов, 5 часов или 6 часов. Раствор затем нейтрализовали и очищали на колонке для гель-фильтрации, как описано выше. Детекцию сахарида осуществляли при 215 нм и фракции объединяли (Фиг. 6).
Объединенные фракции затем окисляли и диализовали против воды, как описано выше. Фракции конъюгировали с CRM197 или CRMadh, как описано выше, и полученные конъюгаты очищали при помощи гель-фильтрационной хроматографии, как описано выше (Фиг. 7).
В другом исследовании использовали хлористоводородную кислоту при 0,5M вместо уксусной кислоты для капсульного полисахарида типа 8 и реакционную смесь выдерживали при 90°C в течение 2,5 часов, с осуществлением отбора проб каждые 30 минут. Образцы анализировали при помощи ЯМР и SEC-ВЭЖХ. Не наблюдали никакой деполимеризации, и уровень O-ацетилирования оставался почти без изменения. В отличие от этого, когда использовали хлористоводородную кислоту при 2M и реакционную смесь выдерживали при 100°C, деполимеризацию наблюдали уже через 30 минут. Уровень O-ацетилирования постепенно снижался в течение 2,5 часов (Фиг. 17 и 18 (ацетильные пики представлены кружками)).
Исследование методом иммунизации - модель абсцесса (1)
Общий протокол анализа: Мышей подвергали иммунизации в соответствии со схемой, описанной ниже, и вводили внутривенную инъекцию бактериальной суспензии S.aureus. Культуру S.aureus центрифугировали, промывали два раза и разводили в PBS перед введением. Дополнительные разведения были нужны для желаемого прививочного материала, который был экспериментально выверен методом посева на агаре и колониеобразования. Для сбора органов мышей умерщвляли и их почки удаляли и гомогенизировали в 1% Triton X-100. Аликвоты затем разбавляли и высевали на агаровой среде для определения КОЭ в трех повторах. Для гистологии ткань почки инкубировали при комнатной температуре в 10% формалине в течение 24 часов. Ткани заключали в парафин, тонко секционировали, окрашивали гематоксилином/эозином и исследовали под микроскопом.
CD1 мышей в возрасте 3 недели подвергали иммунизации в дни 0 и 11 путем интраперитонеальной инъекции с использованием 5 мкг дозы антигена в объеме инъекции 200 мкл. У мышей брали кровь в дни 0 и 20 и осуществляли введение S.aureus в день 21. Органы собирали в день 25. Иммунизации осуществляли в группах из восьми мышей в соответствии со следующей схемой:
Группа 1 - только квасцы
Группа 2 - только капсульный полисахарид типа 5
Группа 3 - капсульный полисахарид типа 5 плюс квасцы
Группа 4 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 1)
Группа 5 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 1) плюс квасцы
Конъюгат индуцировал специфический IgG ответ против полисахарида типа 5. Композиция квасцов давала улучшенный ответ (Фиг. 8). Конъюгат также индуцировал специфический IgM ответ против полисахарида типа 5 (Фиг. 9). Композиция конъюгата с квасцами также давала лучшую защиту от почечной инфекции (Фиг. 10).
Исследование методом иммунизации - модель абсцесса (2)
CD1 мышей в возрасте 3 недели подвергали иммунизации в дни 1, 14 и 28 путем интраперитонеальной инъекции с использованием 5 мкг дозы антигена в объеме инъекции 200 мкл. У мышей брали кровь в дни 0, 27 и 37 и осуществляли введение S.aureus в день 38. Органы собирали в день 42. Иммунизации осуществляли в группах из восьми мышей в соответствии со следующей схемой:
Группа 1 - только квасцы
Группа 2 - капсульный полисахарид типа 5 плюс квасцы
Группа 3 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 2) плюс квасцы
Группа 4 - конъюгат капсульный полисахарид типа 5-CRMadh (партия A') плюс квасцы
Конъюгаты индуцировали специфический IgG ответ против полисахарида типа 5. Конъюгаты по настоящему изобретению (представлены партией A') давали особенно высокий титр (Фиг. 11). Конъюгаты по настоящему изобретению давали лучшую защиту от почечной инфекции (Фиг. 12).
Исследование методом иммунизации - модель абсцесса (3)
CD1 мышей в возрасте 3 недели подвергали иммунизации в дни 1, 14 и 28 путем интраперитонеальной инъекции с использованием 5 мкг дозы (или 0,5 мкг дозы в случае партии A) антигена в объеме инъекции 200 мкл. У мышей брали кровь в дни 0, 27 и 37 и осуществляли введение S.aureus (выращен в жидкой или твердой среде) в день 38. Органы собирали в день 42. Иммунизации осуществляли в группах из восьми мышей в соответствии со следующей схемой:
Группа 1 - только квасцы
Группа 2 - капсульный полисахарид типа 5 плюс квасцы
Группа 3 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 2) плюс квасцы
Группа 4 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 3) плюс квасцы
Группа 5 - конъюгат капсульный полисахарид типа 5-CRMadh (партия A') плюс квасцы
Группа 6 - капсульный полисахарид типа 5-CRM конъюгат (партия A) плюс квасцы
Группа 7 - конъюгат капсульный полисахарид типа 5-CRM (партия B) плюс квасцы
Конъюгаты по настоящему изобретению (представлены партиями A', А и B) давали защиту от почечной инфекции (Фиг. 13А и 13B). Конъюгаты по настоящему изобретению давали высокие титры специфических IgG антител с низкими титрами IgM антител (Фиг. 14).
Исследование методом иммунизации - модель абсцесса (4)
CD1 мышей в возрасте 3 недели подвергали иммунизации в дни 1 и 14 путем интраперитонеальной инъекции с использованием дозы 1 мкг антигена в объеме инъекции 200 мкл. У мышей брали кровь в дни 0, 13 и 27 и осуществляли введение S.aureus в день 28. Органы собирали в день 32. Иммунизации осуществляли в группах из восьми или девяти мышей в соответствии со следующей схемой:
Группа 1 - конъюгат капсульный полисахарид типа 8-CRM (партия α) плюс квасцы
Группа 2 - конъюгат капсульный полисахарид типа 8-CRM (партия α) плюс MF59
Группа 3 - только квасцы
Группа 4 - только MF59
Конъюгаты по настоящему изобретению давали защиту от почечной инфекции (Фиг. 15). Композиция квасцов давала лучшую защиту по сравнению с MF59 композицией.
Исследование методом иммунизации - летальная модель (1)
Общий протокол анализа: Мышей подвергали иммунизации в соответствии со схемой, описанной ниже, и вводили интраперитонеальную инъекцию бактериальной суспензии S.aureus. Культуры S.aureus центрифугировали, промывали два раза и разводили в PBS перед введением. Дополнительные разведения были нужны для желаемого прививочного материала, который был экспериментально выверен методом посева на агаре и колониеобразования. Животных отслеживали в течение 14 дней и летальное заболевание регистрировали.
CD1 мышей подвергали иммунизации путем интраперитонеальной инъекции с использованием 5 мкг дозы антигена в объеме инъекции 200 мкл. Иммунизации осуществляли в группах из двенадцати мышей в соответствии со следующей схемой, перед введением 5×108 КОЭ S.aureus типа 5:
Группа 1 - PBS плюс квасцы
Группа 2 - конъюгат капсульный полисахарид типа 5-CRM (партия C) плюс квасцы
Группа 3 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 3) плюс квасцы
Конъюгаты по настоящему изобретению (представлены партией C) обеспечивали более высокое выживание (Фиг. 16).
Исследование методом иммунизации - летальная модель (2)
CD1 мышей подвергали иммунизации путем интраперитонеальной инъекции с использованием 2 мкг (сахарид) и 10 мк (белок, если присутствует) доз антигена в объеме инъекции 200 мкл. Иммунизации осуществляли в группах из двенадцати мышей в соответствии со следующей схемой, перед введением 5×108 КОЭ S.aureus типа 5:
Группа 1 - PBS плюс квасцы
Группа 2 - конъюгат капсульный полисахарид типа 5-CRM (партия D) плюс квасцы
Группа 3 - конъюгат капсульный полисахарид типа 5-CRMadh (партия 4) плюс квасцы
Группа 4 - конъюгат капсульный полисахарид типа 5-CRM (партия D) плюс белки EsxAB, Sta006 и Sta011 и квасцы
Группа 5 - конъюгат капсульный полисахарид типа 5-CRM (партия D) плюс белки HlaH35L, Sta006 и Sta011 и квасцы
Данные выживания представлены в таблице 5.

Конъюгаты по настоящему изобретению (представлены партией D) обеспечивали более высокое выживание. Выживание повышалось при добавлении белковых антигенов S.aureus.
Исследование методом иммунизации - летальная модель (3)
CD1 мышей подвергали иммунизации путем интраперитонеальной инъекции с использованием доз антигена 2 мкг (полисахарид типа 5, если присутствует) и 1 мкг (полисахарид типа 8, если присутствует) и 10 мк (белок, если присутствует) в объеме инъекции 200 мкл. Иммунизации осуществляли в группах из двенадцати мышей в соответствии со следующей схемой, перед введением 5×l08 КОЭ S.aureus типа 5:
Группа 1 - PBS плюс квасцы
Группа 2 - конъюгат капсульный полисахарид типа 5-CRM (партия E) плюс белки EsxAB, Sta006 и Sta0l1 и квасцы
Группа 3 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия β) плюс белки EsxAB, Sta006 и Sta011 и квасцы
Группа 4 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия β) плюс белки EsxAB, Sta011 и Sta073 и квасцы
Данные выживания представлены в таблице 7.

Исследование методом иммунизации - летальная модель (4)
CD1 мышей подвергали иммунизации путем интраперитонеальной инъекции с использованием доз антигена 2 мкг (капсульный полисахарид типа 5, если присутствует) и 1 мкг (капсульный полисахарид типа 8, если присутствует) и 10 мк (белок, если присутствуют) доз антигена в объеме инъекции 200 мкл. Иммунизации осуществляли в группах из двенадцати мышей в соответствии со следующей схемой, перед введением 5×108 КОЭ S.aureus типа 5:
Группа 1 - PBS плюс квасцы
Группа 2 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия γ первая доза, партия δ вторая доза) плюс белки EsxAB, Sta006 и Sta011 и квасцы
Группа 3 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия γ первая доза, партия δ вторая доза) плюс квасцы
Группа 4 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия γ первая доза, партия δ вторая доза) плюс белок EsxAB и квасцы
Группа 5 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия γ первая доза, партия δ вторая доза) плюс белок Sta006 и квасцы
Группа 6 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия γ первая доза, партия δ вторая доза) плюс белок Sta011 и квасцы
Группа 7 - конъюгат капсульный полисахарид типа 5-CRM (партия E) и конъюгат капсульный полисахарид типа 8-CRM (партия γ первая доза, партия δ вторая доза) плюс белки Sta006 и Sta011 и квасцы
Группа 8 - конъюгат капсульный полисахарид типа 5-CRM (партия E) плюс белки HlaH35L, Sta006 и Sta011 и квасцы
Группа 9 - конъюгат капсульный полисахарид типа 5-CRM (партия E) плюс белок HlaH35L и квасцы
Данные выживания представлены в таблице 8.

Должно быть понятно, что изобретение было описано только в качестве примера, и можно осуществить модификации, не выходя при этом за рамки объема и сути настоящего изобретения.
ССЫЛОЧНЫЕ ДОКУМЕНТЫ
[I] Fattom et al. (1990) Infect Immun. 58(7):2367-74.
[2] Fattom et al. (1992) Infect Immun. 60(2):584-9.
[3] Fattom et al. (1993) Infect Immun. 61(3):1023-32.
[4] Fattom et al. (1996) Infect Immun. 64(5):1659-65.
[5] Welch et al. (1996) J Am Soc Nephrol. 7(2):247-53.
[6] Fattom et al. (1998) Infect Immun. 66(10):4588-92.
[7] Fattom et al. (1993) Vaccine 17(2):126-33.
[8] Fattom et al. (2002) N Engl J Med. 346(7):491-6.
[9] Robbins et al. (2005) Ann N Y Acad Sci. 754:68-82.
[10] Reynaud-Rondier et al. (1991) FEMS Microbiology Immunology 76:193-200.
[11] Tollersrud et al. (2001) Vaccine. 19(28-29):3896-903.
[12] WO03/061558.
[13] Gilbert et al. (1994) Vaccine. 12(4):369-74.
[14] Moreau et al. (1990) Carbohydrate Res. 339(5):285-91.
[15] Fournier et al (1984) Infect. Immun. 45(1):87-93.
[16] Jones (2005) Carbohydrate Res. 340(6):1097-106.
[17] Lemercinier and Jones (1996) Carbohydrate Res. 296:83-96.
[18] Jones and Lemercinier (2002) J Pharm Biomed Anal. 30(4):1233-47.
[19] WO05/033148.
[20] WO 00/56357.
[21] Hestrin (1949) J. Biol. Chem. 180:249-261.
[22] Konadu et al. (1994) Infect. Immun. 62:5048-5054.
[23] Gilbert et al. (1994) J. Microb. Meth. 20:39-46.
[24] Патентная заявка США № 61/256905 'PURIFICATION OF STAPHYLOCOCCUS AUREUS TYPE 5 AND TYPE 8 CAPSULAR SACCHARIDES' (NOVARTIS AG). Assignee reference no. 53615-US-PSP.
[25] Ramsay et al. (2001) Lancet 357(9251):195-196.
[26] Lindberg (1999) Vaccine 17 Suppl 2:S28-36.
[27] Buttery & Moxon (2000) J R Coll Physicians Lond 34:163- 168.
[28] Ahmad & Chapnick (1999) Infect Dis Clin North Am 13:113-33, vii.
[29] Goldblatt (1998) J. Med. Microbiol. 47:563-567.
[30] Европейский патент № 0477508.
[31] Патент США № 5306492.
[32] WO98/42721.
[33] Dick et al. В Conjugate Vaccines (eds. Cruse et al.) Karger, Basel, 1989, 10:48-114.
[34] Hermanson Bioconjugate Techniques, Academic Press, San Diego (1996) ISBN:0123423368.
[35] Research Disclosure, 453077 (Jan 2002).
[36] Herbelin et al. (1997) J Dairy Sci. 80(9):2025-34.
[37] EP-A-0372501.
[38] EP-A-0378881.
[39] EP-A-0427347.
[40] WO93/17712.
[41] WO94/03208.
[42] WO98/58668.
[43] EP-A-0471177.
[44] WO91/01146.
[45] Falugi et al. (2001) Eur J Immunol 31:3816-3824.
[46] Baraldo et al. (2004) Infect Immun 72(8):4884-7.
[47] EP-A-0594610.
[48] Ruan et al. (1990) J Immunol 145:3379-3384.
[49] WO00/56360.
[50] WO02/091998.
[51] Kuo et al. (1995) Infect Immun 63:2706-13.
[52] Michon et al. (1998) Vaccine. 16:1732-41.
[53] WO01/72337.
[54] WO00/61761.
[55] WO2004/041157.
[56] WO02/34771.
[57] WO99/42130.
[58] WO2004/011027.
[59] WO2007/113222.
[60] Патент США № 6045805
[61] Патенты США №№ 6027733 & 6274144.
[62] www.polymer.de
[63] Патенты США №№ 4356170 и 4663160.
[64] Патент США № 4711779.
[65] WO00/10599.
[66] Патент США № 4057685.
[67] WO96/40242.
[68] Lei et al. (2000) Dev Biol (Basel) 103:259-264.
[69] WO00/38711; Патент США № 6146902.
[70] WO2004/080490.
[71] WO2006/032475.
[72] WO2006/032500.
[73] WO2006/065553.
[74] WO2007/118979.
[75] WO99/24578.
[76] WO99/36544.
[77] WO99/57280.
[78] WO00/22430.
[79] Tettelin et al. (2000) Science 287:1809-1815.
[80] WO96/29412.
[81] Pizza et al. (2000) Science 287:1816-1820.
[82] WO01/52885.
[83] Bjune et al. (1991) Lancet 338(8775):1093-1096.
[84] Fukasawa et al. (1999) Vaccine 17:2951-2958.
[85] Rosenqvist et al. (1998) Dev. Biol. Stand. 92:323-333.
[86] Costantino et al. (1992) Vaccine 10:691-698.
[87] WO03/007985.
[88] Watson (2000) Pediatr Infect Dis J 19:331-332.
[89] Rubin (2000) Pediatr Clin North Am 47:269-285, v.
[90] Jedrzejas (2001) Microbiol Mol Biol Rev 65:187-207.
[91] Bell (2000) Pediatr Infect Dis J 19:1187-1188.
[92] Iwarson (1995) APMIS 103:321-326.
[93] Gerlich et al. (1990) Vaccine 8 Suppl:S63-68 & 79-80.
[94] Hsu et al. (1999) Clin Liver Dis 3:901-915.
[95] Gustafsson et al. (1996) N. Engl. J. Med. 334:349-355.
[96] Rappuoli et al. (1991) TIBTECH 9:232-238.
[97] Vaccines (2004) eds. Plotkin & Orenstein. ISBN 0-7216-9688-0.
[98] WO02/02606.
[99] Kalman et al. (1999) Nature Genetics 21:385-389.
[100] Read et al. (2000) Nucleic Acids Res 28:1397-406.
[101 Shirai et al. (2000) J. Infect. Dis. 181 (Suppl 3):S524-S527.
[102] WO99/27105.
[103] WO00/27994.
[104] WO00/37494.
[105] WO99/28475.
[106] Ross et al. (2001) Vaccine 19:4135-4142.
[107] Sutter et al. (2000) Pediatr Clin North Am 47:287-308.
[108] Zimmerman & Spann (1999) Am Fam Physician 59:113-118, 125-126.
[109] Dreesen (1997) Vaccine 15 Suppl:S2-6.
[110] MMWR Morb Mortal Wkly Rep 1998 Jan 16; 47(1):12, 19.
[111] McMichael (2000) Vaccine 19 Suppl 1:S101-107.
[112] WO02/34771.
[113] Dale (1999) Infect Dis Clin North Am 13:227-43, viii.
[114] Ferretti et al. (2001) PNAS USA 98:4658-4663.
[115] WO03/093306.
[116] WO2004/018646.
[117] WO2004/041157.
[118] Ichiman and Yoshida (1981) J. Appl. Bacteriol. 51:229.
[119] US4197290.
[120] lchiman et al. (1991) J. Appl. Bacteriol. 71:176.
[121] Robinson & Torres (1997) Seminars in Immunology 9:271-283.
[122] Donnelly et al. (1997) Annu Rev Immunol 15:617-648.
[123] Scott-Taylor & Dalgleish (2000) Expert Opin Investig Drugs 9:471-480.
[124] Apostolopoulos & Plebanski (2000) Curr Opin Mol Ther 2:441-447.
[125] Ilan (1999) Curr Opin Mol Ther 1:116-120.
[126] Dubensky et al. (2000) Mol Med 6:723-732.
[127] Robinson & Pertmer (2000) Adv Virus Res 55:1-74.
[128] Donnelly et al. (2000) Am J Respir Crit Care Med 162(4 Pt 2):S190-193.
[129] Davis (1999) Mt. Sinai J. Med. 66:84-90.
[130] Paoletti et al. (2001) Vaccine 19:21 18-2126.
[131] WO00/56365.
[132] Gennaro (2000) Remington: The Science and Practice of Pharmacy. 20th edition, ISBN:0683306472.
[133] WO03/009869.
[134] Almeida & Alpar (1996) J. Drug Targeting 3:455-467.
[135] Agarwal & Mishra (1999) Indian J Exp Biol 37:6-16.
[136] WO00/53221.
[137] Jakobsen et al. (2002) Infect Immun 70:1443- 1452.
[138] Bergquist et al. (1998) APMIS 106:800-806.
[139] Baudner et al. (2002) Infect Immun 70:4785-4790.
[140] Ugozzoli et al. (2002) J Infect Dis 186:1358-1361.
[141] Vaccine Design. (1995) eds. Powell & Newman. ISBN:030644867X. Plenum.
[142] WO00/23105.
[143] WO90/14837.
[144] Podda (2001) Vaccine 19:2673-80.
[145] Frey et al. (2003) Vaccine 21:4234-7.
[146] Патент США № 6299884.
[147] Патент США № 6451325.
[148] Патент США № 5057540.
[149] WO96/33739.
[150] EP-A-0109942.
[151] WO96/11711.
[152] WO00/07621.
[153] Barr et al. (1998) Advanced Drug Delivary Reviews 32:247-271.
[154] Sjolanderet et al. (1998) Advanced Drug Delivary Reviews 32:321-338.
[155] Niikura et al. (2002) Virology 293:273-280.
[156] Lenz et al. (2001) J Immunol 166:5346-5355.
[157] Pinto et al. (2003) J Infect Dis 188:327-338.
[158] Gerber et al. (2001) Virol 75:4752-4760.
[159] WO03/024480.
[160] WO03/024481.
[161] Gluck et al. (2002) Vaccine 20:B10-B16.
[162] EP-A-0689454.
[163] Johnson et al. (1999) Bioorg Med Chem Lett 9:2273-2278.
[164] Evans et al. (2003) Expert Rev Vaccines 2:219-229.
[165] Meraldi et al. (2003) Vaccine 21:2485-2491.
[166] Pajak et al. (2003) Vaccine 21:836-842.
[167] Kandimalla et al. (2003) Nucleic Acids Research 31:2393-2400.
[168] WO02/26757.
[169] WO99/62923.
[170] Krieg (2003) Nature Medicine 9:831-835.
[171] McCluskie et al. (2002) FEMS Immunology and Medical Microbiology 32:179-185.
[172] WO98/40100.
[173] Патент США № 6207646.
[174] Патент США № 6239116.
[175] Патент США № 6429199.
[176] Kandimalla et al. (2003) Biochemicval Society Transactions 31 (part 3):654-658.
[177] Blackwell et al. (2003) J Immunol 170:4061-4068.
[178] Krieg (2002) Trends Immunol 23:64-65.
[179] WO01/95935.
[180] Kandimalla et al. (2003) BBRC 306:948-953.
[181] Bhagat et al. (2003) BBRC 300:853-861.
[182] WO03/035836.
[183] WO95/17211.
[184] WO98/42375.
[185] Beignon et al. (2002) Infect Immun 70:3012-3019.
[186] Pizza et al. (2001) Vaccine 19:2534-2541.
[187] Pizza et al. (2000) Int J Med Microbiol 290:455-461.
[188] Scharton-Kersten et al. (2000) Infect Immun 68:5306-5313.
[189] Ryan et al. (1999) Infect Immun 67:6270-6280.
[190] Partidos et al. (1999) Immunol Lett 67:209-216.
[191] Peppoloni et al. (2003) Expert Rev Vaccines 2:285-293.
[192] Pine et al. (2002) J Control Release 85:263-270.
[193] Domenighini et al. (1995) Mol Microbiol 15:1165-1167.
[194] WO99/40936.
[195] WO99/44636.
[196] Singh et al. (2001) J Cont Release 70:267-276.
[197] WO99/27960.
[198] Патент США № 6090406.
[199] Патент США № 5916588.
[200] EP-A-0626169.
[201] WO99/52549.
[202] WO01/21207.
[203] WO01/21152.
[204] Andrianov et al. (1998) Biomaterials 19:109-115.
[205] Payne et al. (1998 Adv Drug Delivary Review 31:185-196.
[206] Stanley (2002) Clin Exp Dermatol 27:571-577.
[207] Jones (2003) Curr Opin Investig Drugs 4:214-218.
[208] WO04/60308.
[209] WO04/64759.
[210] WO99/11241.
[211] WO94/00153.
[212] WO98/57659.
[213] Европейские патентные заявки №№ 0835318, 0735898 и 0761231.
[214] Joyce et al. (2003) Carbohydrate Research 338:903.
[215] Maira-Litran et al. (2002) Infect. Immun. 70:4433.
[216] WO2004/043407.
[217] WO2007/113224.
[218] WO2004/043405.
[219] WO98/10788.
[220] WO2007/053176.
[221] WO2007/113222.
[222] WO2005/009379.
[223] WO2009/029132.
[224] WO2008/079315.
[225] WO2005/086663.
[226] WO2005/115113.
[227] WO2006/033918.
[228] WO2006/078680.
[229] Kuroda et al. (2001) Lancet 357(9264):1225-1240; см. также стр. 1218-1219.
[230] Sjodahl (1977) J. Biochem. 73:343-351.
[231] Uhlen et al. (1984) J Biol. Chem. 259:1695-1702 & 13628 (Corr.).
[232] Schneewind et al. (1992) Cell 70:267-281.
[233] DeDent et al. (2008) EMBO J. 27:2656-2668.
[234] Sjoquist et al. (1972) Eur. J. Biochem. 30:190-194.
[235] DeDent et al. (2007) J. Bacteriol. 189:4473-4484.
[236] Deisenhofer et al., (1978) Hoppe-Seyh Zeitsch. Physiol. Chem. 359:975-985.
[237] Deisenhofer (1981) Biochemistry 20:2361-2370.
[238] Graille et al. (2000) Proc. Nat. Acad. Sci. USA 97:5399-5404.
[239] O'Seaghdha et al. (2006) FEBS J. 273:4831-41.
[240] Gomez et al. (2006) J. Biol. Chem. 281:20190-20196.
[241] WO2007/071692.
[242] Sebulsky & Heinrichs (2001) J Bacteriol 183:4994-5000.
[243] Sebulsky et al. (2003) J Biol Chem 278:49890-900.
[244] WO2005/009378.
[245] Rable & Wardenburg (2009) Infect Immun 77:2712-8.
[246] WO2007/145689.
[247] WO2009/029831.
[248] WO2005/079315.
[249] WO2008/152447.
[250] Kuklin et al. (2006) Infect Immun. 74(4):2215-23.
[251] WO2005/009379.
[252] Gennaro (2000) Remington: The Science and Practice of Pharmacy. 20th edition, ISBN:0683306472.
[253] Methods in Enzymology (S. Colowick and N. Kaplan, eds., Academic Press, Inc.)
[254] Handbook of Experimental Immunology, Vols. I-IV (D.M. Weir and C.C. Blackwell, eds, 1986, Blackwell Scientific Publications)
[255] Sambrook et al. (2001) Molecular Cloning: A Laboratory Manual, 3rd edition (Cold Spring Harbor Laboratory Press).
[256] Handbook of Surface and Colloidal Chemistry (Birdi, K.S. ed., CRC Press, 1997).
[257] Ausubel et al. (eds) (2002) Short protocols in molecular biology, 5th edition (Current Protocols).
[258] Molecular Biology Techniques: An Intensive Laboratory Course, (Ream et al., eds., 1998, Academic Press).
[259] PCR (Introduction to Biotechniques Series), 2nd ed. (Newton & Graham eds., 1997, Springer Verlag).
[260] Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987) Supplement 30.
[261] Smith & Waterman (1981) Adv. Appl. Math. 2:482-489.
[262] Geysen et al. (1984) PNAS USA 81:3998-4002.
[263] Carter (1994) Methods Mol Biol 36:207-23.
[264] Jameson, BA et al. 1988, CABIOS 4(1):181-186.
[265] Raddrizzani & Hammer (2000) Brief Bioinform 1(2):179-89.
[266] Bublil et al. (2007) Proteins 68(1):294-304.
[267] De Lalla et al. (1999) J. Immunol. 163:1725-29.
[268] Kwok et al. (2001) Trends Immunol 22:583-88.
[269] Brusic et al. (1998) Bioinformatics 14(2):121-30.
[270] Meister et al. (1995) Vaccine 13(6):581-91.
[271] Roberts et al., (1996) AIDS Res Hum Retroviruses 12(7):593-610.
[272] Maksyutov & Zagrebelnaya (1993) Comput Appl Biosci 9(3):291-7.
[273] Feller & de la Cruz (1991) Nature 349(6311):720-1.
[274] Hopp (1993) Peptide Research 6:183-190.
[275] Welling et al. (1985) FEBS Lett. 188:215-218.
[276] Davenport et al. (1995) Immunogenetics 42:392-297.
[277] Tsurui & Takahashi (2007) J Pharmacol Sci. 105(4):299-316.
[278] Tong et al. (2007) Brief Bioinform. 8(2):96-108.
[279] Schirle et al. (2001) J Immunol Methods. 257(1-2):1-16.
[280] Chen et al. (2007) Amino Acids 33(3):423-8.
[281] Bardotti et al. (2008) Vaccine. 26(18):2284-96.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОЧИСТКА КАПСУЛЬНЫХ САХАРИДОВ STAPHYLOCOCCUS AUREUS ТИПА 5 И ТИПА 8 | 2010 |
|
RU2579900C2 |
| КОМПОЗИЦИИ ДЛЯ ИММУНИЗАЦИИ ПРОТИВ STAPHYLOCOCCUS AUREUS | 2010 |
|
RU2508126C2 |
| МОЛЕКУЛА, СОДЕРЖАЩАЯ SPR0096 и SPR2021 | 2012 |
|
RU2636350C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ АНТИГЕНОВ STAPHYLOCOCCUS AUREUS | 2010 |
|
RU2536981C9 |
| ЖИДКИЕ ВАКЦИНЫ ДЛЯ МНОЖЕСТВЕННЫХ СЕРОГРУПП МЕНИНГОКОККОВ | 2004 |
|
RU2378010C2 |
| ЖИДКИЕ ВАКЦИНЫ ДЛЯ МНОЖЕСТВЕННЫХ СЕРОГРУПП МЕНИНГОКОККОВ | 2009 |
|
RU2595845C2 |
| ГИПО- И ГИПЕРАЦЕТИЛИРОВАННЫЕ МЕНИНГОКОККОВЫЕ КАПСУЛЬНЫЕ САХАРИДЫ | 2004 |
|
RU2362784C2 |
| МНОЖЕСТВЕННЫЕ ВАРИАНТЫ МЕНИНГОКОККОВОГО БЕЛКА NMB 1870 | 2003 |
|
RU2336091C2 |
| ПОЛИПЕПТИДНЫЕ ВАКЦИНЫ ДЛЯ ШИРОКОЙ ЗАЩИТЫ ПРОТИВ РЯДОВ ПОКОЛЕНИЙ МЕНИНГОКОККОВ С ПОВЫШЕННОЙ ВИРУЛЕНТНОСТЬЮ | 2003 |
|
RU2333007C2 |
| АДЪЮВАНТНЫЙ МЕНИНГОКОККОВЫЙ БЕЛОК, СВЯЗЫВАЮЩИЙ ФАКТОР Н | 2010 |
|
RU2557315C2 |





























































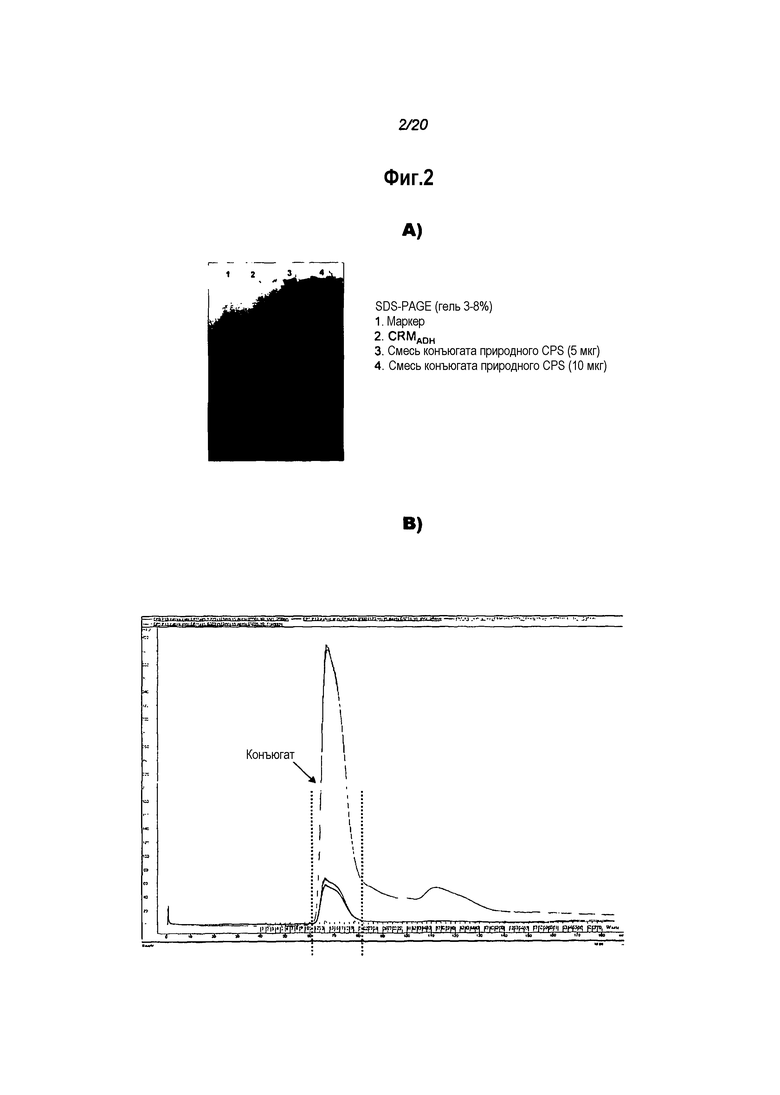

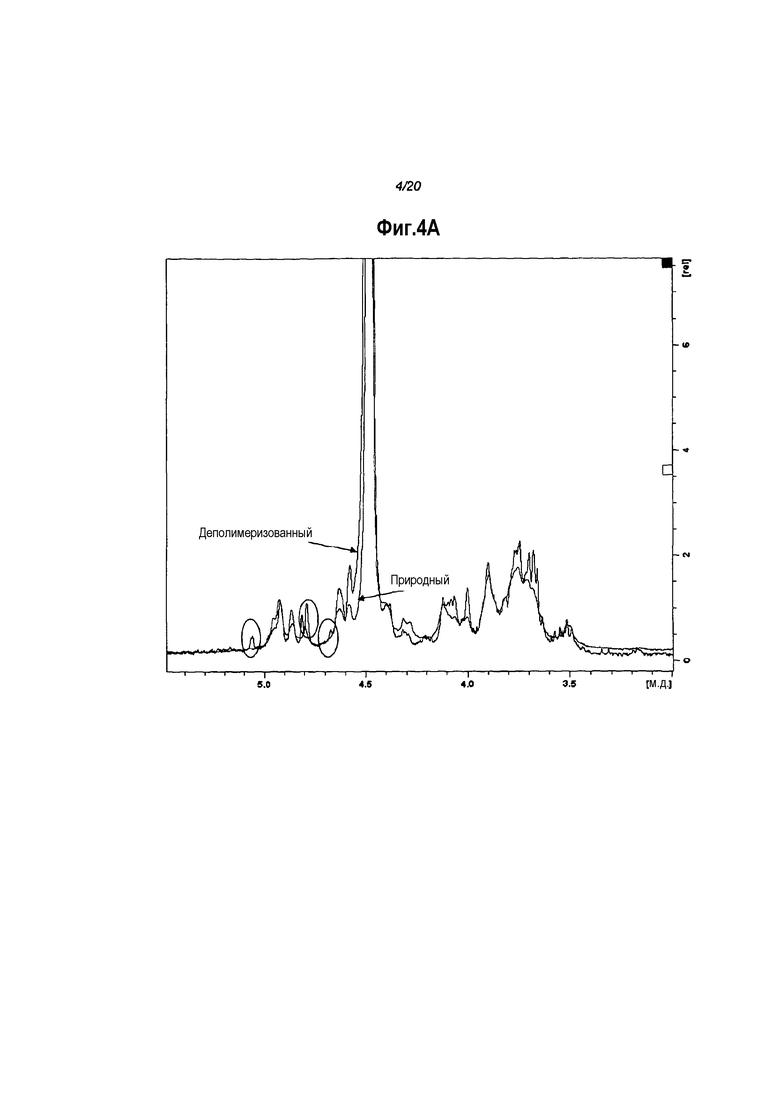



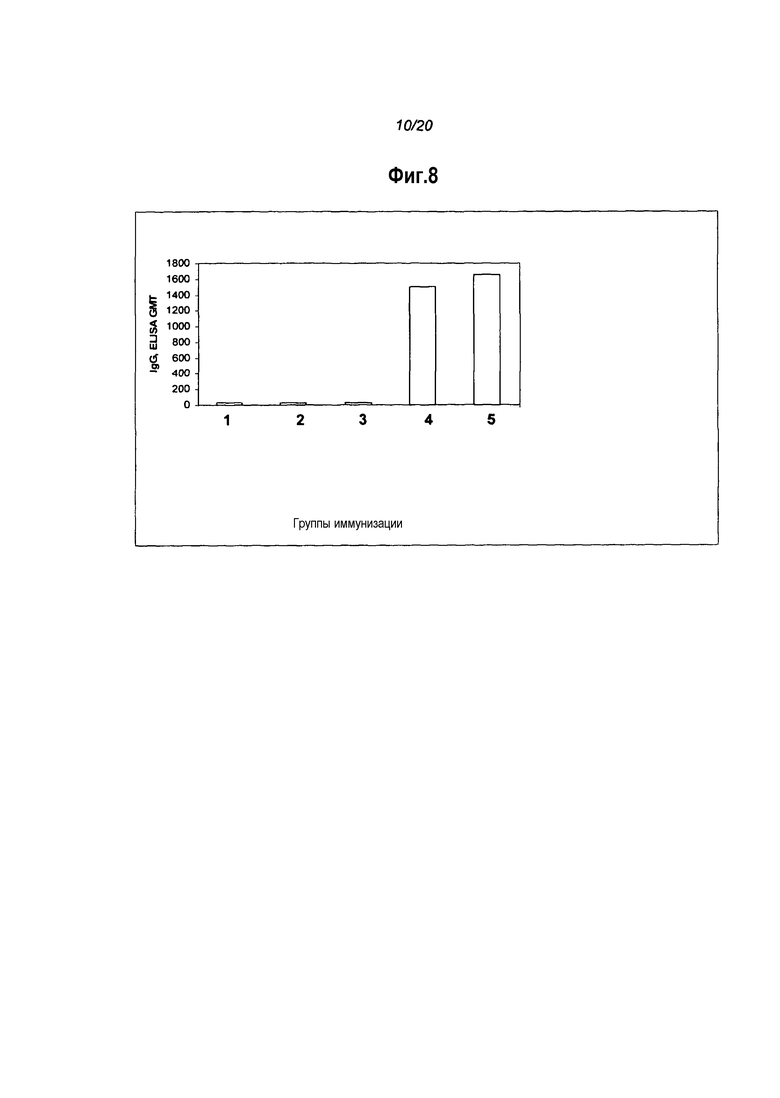

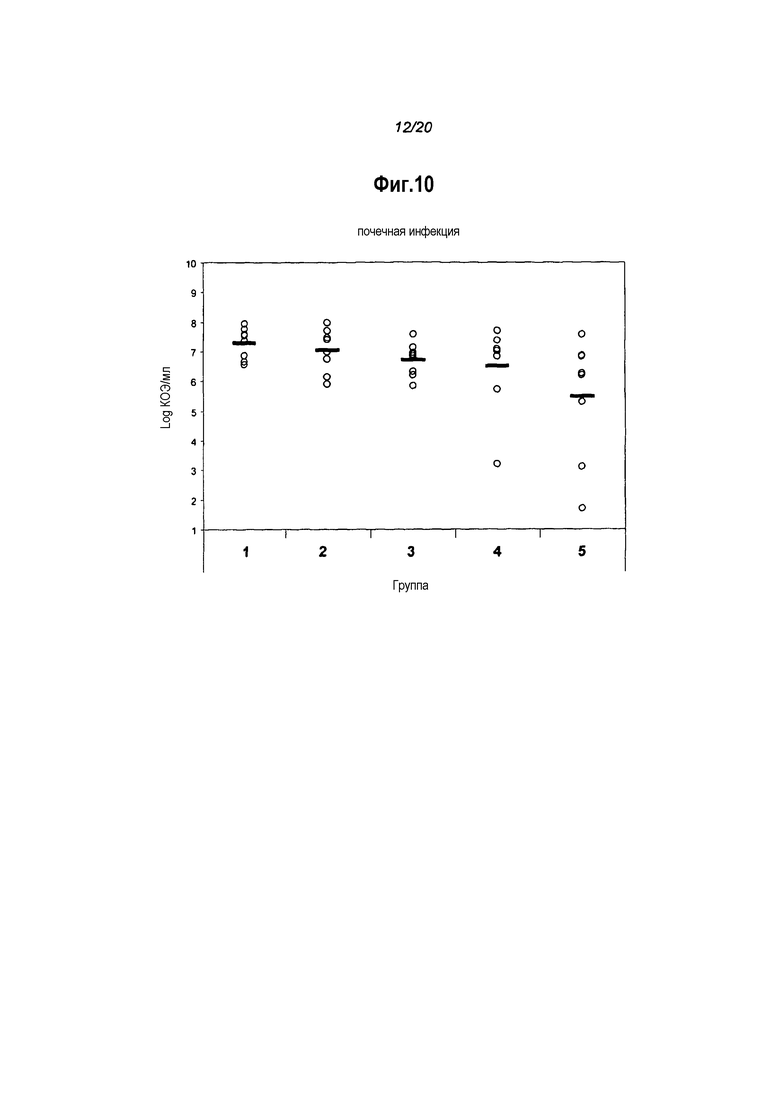

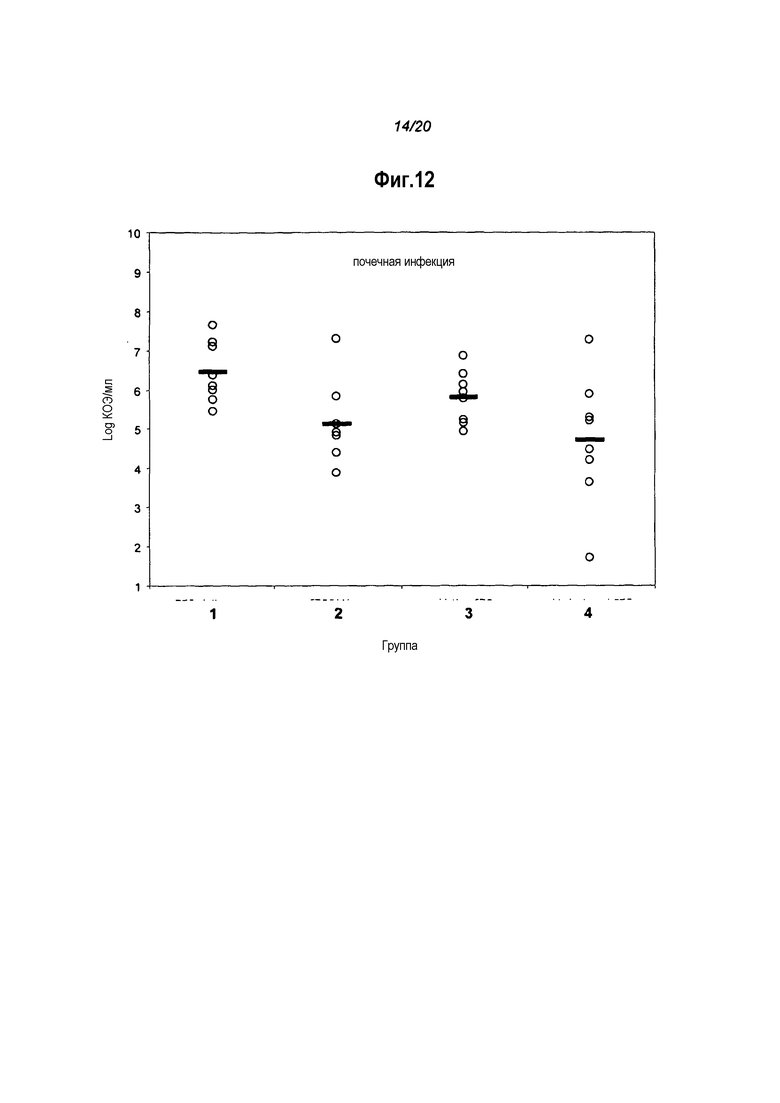



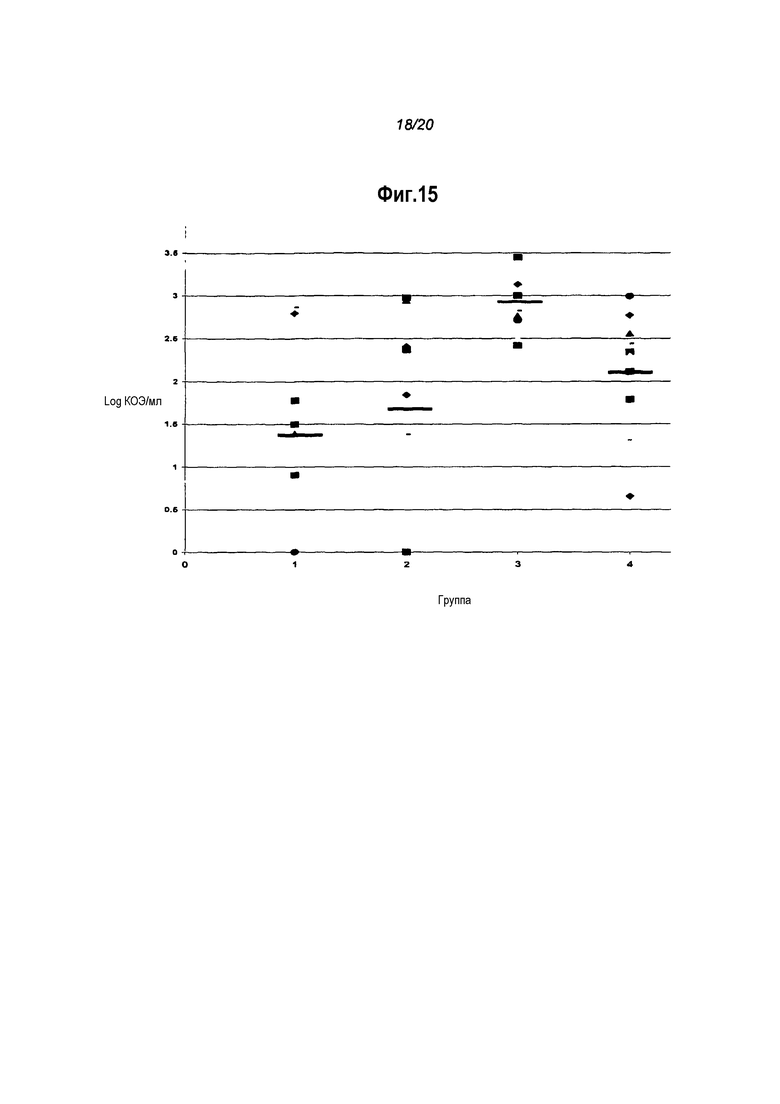


Настоящее изобретение относится к биохимии, в частности к способу получения конъюгата капсульного полисахарида S.aureus типа 5 или типа 8 и белка-носителя. Способ включает стадию деполимеризации капсульного полисахарида путем кислотного гидролиза с получением полисахаридного фрагмента, стадию окисления фрагмента для введения альдегидной группы в по меньшей мере один сахаридный остаток во фрагменте с получением окисленного сахаридного остатка и стадию связывания окисленного сахаридного остатка с белком-носителем через альдегидную группу. На стадии окисления две вицинальные гидроксильные группы полисахарида преобразуются в альдегидные. Настоящее изобретение обеспечивает конъюгат капсульного полисахарида S.aureus типа 5 или типа 8 и белка-носителя, полученный указанным способом, и иммуногенную композицию, включающую указанный конъюгат. Настоящее изобретение позволяет улучшить иммунологические свойства получаемого конъюгата. 3 н. и 10 з.п. ф-лы, 18 ил., 8 табл., 1 пр.
1. Способ получения конъюгата капсульного полисахарида S. aureus типа 5 или типа 8 и белка-носителя, включающий следующие стадии: (а) деполимеризация капсульного полисахарида путем кислотного гидролиза с получением полисахаридного фрагмента; (b) окисление фрагмента для введения альдегидной группы в, по меньшей мере, один сахаридный остаток во фрагменте с получением окисленного сахаридного остатка; и (с) связывание окисленного сахаридного остатка с белком-носителем через альдегидную группу с получением, таким образом, конъюгата;
где стадия (b) включает:
(i) стадию окисления капсульного полисахарида S.aureus типа 5, содержащего β-D-FucNAc-(1→часть на его нередуцирующем конце, для преобразования двух вицинальных гидроксильных групп в β-D-FucNAc-(1→части в две альдегидные группы; или
(ii) стадию окисления капсульного полисахарида S. aureus типа 8, содержащего α-D-FucNAc-(1→часть на его нередуцирующем конце, для преобразования двух вицинальных гидроксильных групп в α-D-FucNAc-(1→части в две альдегидные группы.
2. Способ по п. 1, где деполимеризацию осуществляют путем кислотного гидролиза с использованием уксусной кислоты.
3. Способ по п. 1, где средняя молекулярная масса фрагмента находится в пределах от 5 до 100 кДа.
4. Способ по п. 1, где степень О-ацетилирования фрагмента составляет 10-90%.
5. Способ по п. 1, где связывание представляет собой прямое связывание путем взаимодействия альдегидной группы с группой амина в носителе путем восстановительного аминирования.
6. Способ по п. 1, где связывание осуществляют через линкер путем взаимодействия альдегидной группы с группой амина в линкере путем восстановительного аминирования.
7. Способ по п. 6, где линкер присоединен к белку-носителю.
8. Способ по п. 1, где связывание приводит к отношению полисахарид : белок (масс./масс.) в пределах от 1:5 до 1:2.
9. Конъюгат капсульного полисахарида S. aureus типа 5 или типа 8 и белка-носителя для получения лекарственного средства для повышения иммунного ответа у млекопитающего, где указанные капсульный полисахарид S. aureus типа 5 или типа 8 и белок-носитель соединены через альдегидную связь, образованную в результате окисления вицинальной группы полисахарида, где указанный конъюгат получен способом по любому из пп. 1-8.
10. Иммуногенная композиция, включающая эффективное количество конъюгата по п. 9.
11. Композиция по п. 10, дополнительно включающая один или более белковых антигенов S. aureus, выбранных из группы, включающей антиген clfA; антиген clfB; антиген sdrE2; антиген sdrC; антиген sasF; антиген emp; антиген sdrD; антиген spa; антиген esaC; антиген esxA; антиген esxB; антиген sta006; антиген isdC; антиген Hla, антиген sta011; антиген isdA; антиген isdB; и антиген sta073.
12. Композиция по п. 11, где один или более белковых антигенов S. aureus выбраны из группы, состоящей из антигена esxA; антигена esxB; антигена sta006; антигена Hla; антигена sta011; и антигена sta073.
13. Композиция по п. 12, где композиция включает белковые антигены S. aureus в соответствии с одним из сочетаний (1)-(10), представленных ниже:
(1) антиген esxA, антиген esxB, антиген sta006 и антиген Hla;
(2) антиген esxA, антиген esxB, антиген sta006 и антиген sta011;
(3) антиген esxA, антиген esxB и антиген sta011;
(4) антиген esxA, антиген esxB, антиген Hla, антиген sta006 и антиген sta011;
(5) антиген esxA, антиген esxB и антиген Hla;
(6) антиген Hla, антиген sta006 и антиген sta011;
(7) антиген esxA и антиген esxB;
(8) антиген esxA, антиген esxB и антиген sta006;
(9) антиген esxA, антиген esxB, антиген sta011 и антиген sta073; и
(10) антиген sta006 и антиген sta011.
| US 5306492 А, 26.04.1994 | |||
| LEE JEAN C | |||
| et al., Genetic analysis of type 5 capsular polysaccharide expression by Staphylococcus aureus, Journal of bacteriology, August 1994, Vol.176 | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| US20070110762 A1, 17.05.2007 | |||
| US 6027733 A, 22.02.2000 | |||
| FATTOM A., et al | |||
| Effect of conjugation methodology, carrier protein, and adjuvants of the | |||

