ОБЛАСТЬ ТЕХНИКИ
Настоящая группа изобретений относится к медицине и медицинской технике, в частности, к способам и устройствам для мониторинга уровня глюкозы в крови путем измерения с помощью калориметрического метода теплового эффекта и скорости метаболизма локального участка живой ткани. Также настоящее изобретение относится к способам получения биополимерной молекулы с заданными биологическими свойствами и заданной пространственной структурой. Использование группы изобретений позволит осуществлять неинвазивный мониторинг уровня сахара в крови пациентов, страдающих диабетом, в том числе для ранней диагностики сахарного диабета.
УРОВЕНЬ ТЕХНИКИ
Одним из широко распространенных и опасных заболеваний современности является сахарный диабет, наиболее распространенная форма которого, составляющая более 90-95% от общего числа диабетиков, это - сахарный диабет 2-го типа (СД2), который является одной из самых актуальных проблем современной медицины. Социальная значимость этого хронического заболевания определяется несколькими факторами, важнейшим из которых является высокая распространенность заболевания во всем мире недостаточно высокая эффективность методов лечения заболевания, способы излечения которого на сегодняшний день все еще находятся в стадии разработки. Согласно прогнозу Международной федерации диабета (IDF), число пациентов, страдающих диабетом, к 2035 увеличится мире до 592 млн. человек с 422 млн. в 2018.
Федеральная статистика по здоровью США показала, что 12,3% американцев возрастом 20 лет и выше живет с диабетом. Причем около 3% об этом даже не подозревают, тем самым усугубляя свое положение. Другие 37% живут с пре-диабетом (состояние, характеризующееся повышенным уровнем сахар в крови). Для сравнения, 10 лет назад этот показатель составлял всего 27% вместо 37. Исследователи из Центров по Контролю Заболеваний США утверждают, если человек с пре-диабетом не меняет свой образ жизни в лучшую сторону, то в 30% случаев у него развивается диабет 2 типа в течение 5 лет. Согласно прогнозу Международной федерации диабета (IDF), расходы мирового здравоохранения на лечение диабета к 2035 вырастут до 936 млрд. долларов с 586 млрд. в 2012.
Диабет часто приводит к развитию осложнений, таких как слепота, почечные нарушения, нервные заболевания и сердечно-сосудистые заболевания. Диабет является ведущим заболеванием, приводящим к слепоте в возрасте от 20 до 74 лет. Примерно от 12000 до 24000 человек в год теряют зрение по причине диабета. Диабет является ведущей причиной заболеваний почек, примерно в 40% новых случаев. Примерно 40-60% больных диабетом имеют предрасположенность к различным формам нервных заболеваний, которые могут привести к ампутации конечностей. Больные диабетом примерно в 2-4 раза более предрасположены к сердечным заболеваниям, в частности, инфаркту миокарда.
Диабет представляет собой заболевание, связанное с недостаточной продукцией или неэффективным использованием инсулина клетками организма. Несмотря на то, что причины заболевания поняты не полностью, некоторые факторы, такие как генетические, окружающей среды, вирусные, идентифицированы.
Существует две основных разновидности диабета: 1 типа и 2 типа.
Диабет 1 типа (известный как инсулин - зависимый диабет) является аутоиммунным заболеванием, при котором выработка инсулина полностью прекращается, и чаще всего развивается в детстве и юности. Больные диабетом 1 типа нуждаются в ежедневных инъекциях инсулина.
Диабет 2 типа является метаболическим заболеванием, вызванным тем, что организм не в состоянии производить достаточное количество инсулина или неэффективно использует его. Больные диабетом 2 типа составляют примерно 90-95% от общего числа диабетиков. В США заболеваемость диабетом 2 типа приближается к эпидемиологическому порогу, в основном благодаря увеличению числа американцев пожилого возраста и преобладанию малоподвижного образа жизни, ведущего к ожирению.
Инсулин способствует проникновению глюкозы в клетку с последующим ее расщеплением для получения энергии для всех метаболических процессов. У диабетиков глюкоза не может проникнуть в клетку, накапливаясь в крови, а клетки испытывают энергетический голод.
Больные диабетом 1 типа самостоятельно вводят инсулин с помощью специального шприца и картриджа. Также возможна непрерывная подкожная инъекция инсулина посредством имплантированной помпы. Инсулин обычно получают из поджелудочной железы свиньи или синтезируют химически.
Общепринятые медицинские методы лечения диабета предписывают пациентам, принимающим инсулин, осуществлять самостоятельный мониторинг содержания сахара в крови. Опираясь на знание уровня сахара в крови, пациенты могут регулировать дозу инсулина при очередной инъекции. Регулировка необходима, поскольку уровень сахара в крови меняется в течение дня и день ото дня в силу различных причин. Несмотря на важность такого мониторинга, несколько проведенных исследований показали, что доля пациентов, которые осуществляют такой мониторинг хотя бы раз в день, падает с возрастом. Это падение происходит в основном из-за того, что метод мониторинга, который сегодня используется, связан с инвазивным отбором пробы крови из пальца. Многие пациенты считают взятие пробы крови из пальца более болезненной процедурой, чем инъекция инсулина.
Научно-исследовательские и опытно-конструкторские разработки (НИОКР), направленные на решение проблемы сахарного диабета, активно ведутся в следующих технологических направлениях:
1) Создание неинвазивного (бескровного) метода непрерывного мониторинга сахара крови пациентов;
2) Создание инвазивных методов измерения сахара крови на основе биосенсоров нового поколения;
3) Разработка носимых электронных устройств превентивной медицины для ранней диагностики сахарного диабета на основе биосенсоров и биотехнологий нового поколения;
4) Создание и внедрении в клиническую практику новых эффективных и безопасных противодиабетических лекарственных средств.
Таким образом, создание неинвазивного (бескровного) метода мониторинга сахара крови пациентов, страдающих диабетом, и создание на его основе медицинского прибора, способного заменить промышленные инвазивные глюкометры, является одной из актуальных нерешенных проблем современной медицины и здравоохранения. Задача разработки новых методов и устройств превентивной медицины для ранней диагностики сахарного диабета, основанных на неинвазивной непрерывной регистрации сахарных кривых, тесно связано с разработкой носимых электронных устройств для неинвазивного непрерывного мониторинга сахара крови на основе биосенсоров и биотехнологий нового поколения.
Коммерческий глюкометр для измерения уровня глюкозы в крови инвазивным способом, представляет из себя биосенсор, в котором используется фермент глюкозоксидаза для расщепления содержащейся в крови глюкозы. В процессе расщепления фермент сначала окисляет глюкозу и использует два электрона для восстановления ФАД (компонент фермента) в ФАД-Н2, который, в свою очередь, окисляется в несколько ступеней электродом. Результирующий ток пропорционален концентрации глюкозы. В этом случае электрод является преобразователем, а фермент-биоселективным элементом.
Известны способы и устройства для определения уровня сахара крови неинвазивным способом: [1-10].
Метод калориметрии, как известно, широко используется в биологии для изучения тепловых процессов на молекулярном и клеточном уровнях [1-10]. Метод микрокалориметрии также успешно используется для исследования тепловых процессов (выделения и поглощения тепла) в отдельных органах, в частности, в активных мышцах и нервных волокнах.
Возросшая за последнее десятилетие активность исследований, направленных на создание метода микрокалориметрии для изучения в физиологических условиях in vivo, тепловых процессов, сопряженных с метаболизмом локального участка живой ткани человека, в значительной степени обусловлена работами по созданию неинвазивного метода мониторинга сахара крови (неинвазивного глюкометра) основанного на мониторинге теплопродукции локального участка ткани человека.
Известными методами физиологической калориметрии являются методы: прямой калориметрии и непрямой калориметрии. Метод прямой калориметрии предусматривает непосредственное определение суммарного количества выделяемого тепла с помощью калориметрической камеры для живых объектов.
Метод непрямой калориметрии позволяет определять количество выделяемого тепла косвенным путем, на основе учета динамики дыхательного газообмена с помощью респираторных камер и различных систем. Различают две возможные модификации метода непрямой калориметрии: метод полного газового анализа (учет поглощенного O2 и выделенного CO2) и метод неполного газового анализа (учет поглощенного O2).
Известен способ измерения скорости базального метаболизма человеческого организма с помощью калориметра всего тела (прямая калориметрия), описанный в US 4386604 А, опубл. 07.06.1983. По измерению температуры воздуха и суммарного количества воды, испаряющейся с поверхности всего тела, определяют суммарную теплоотдачу всего тела и вычисляют скорость базального метаболизма. Основным недостатком х способа является то, что для его реализации требуются громоздкие, стационарные и дорогостоящие калориметрические камеры всего тела. Кроме того, метод прямой калориметрии характеризуется низкой точностью.
Известен способ и устройство для микрокалориметрического измерения скорости локального метаболизма ткани, содержания воды межклеточной ткани, концентрации биохимических компонентов крови и давления в сердечно-сосудистой системе, описанный в [11], в котором по измерению теплового эффекта и скорости метаболизма локального участка ткани определяют содержание сахара в крови (RU 2396897). Величина скорости метаболизма определяется путем измерения суммарного количества воды, испаряющейся в процессе неощутимой перспирации с поверхности локального участка кожи, и измерения температуры и влажности окружающего воздуха. Упомянутый способ измерения позволяет реализовать высокую точность в лабораторных условиях с контролируемыми параметрами микроклимата (температура и влажность воздуха) помещения, в котором проводится измерение, при неизменных значениях климатических параметров внешней среды (температура и влажность окружающей среды, атмосферное давление). Основным недостатком упомянутого способа, ограничивающим его практическое применение, является то, что результаты измерений зависят от физико-климатических факторов окружающей среды; в каждом случае при изменении климатических факторов, при неизменных значениях параметров микроклимата помещения, в котором проводятся измерения, требуется новая калибровка измерительного канала.
Известны спектральные методы для неинвазивного измерения концентрации глюкозы в крови [1]. Основным недостатком известных неинвазивных устройств для измерения концентрации глюкозы в крови, основанных на методах спектроскопии диффузного отражения и абсорбционной спектроскопии, является высокая погрешность измерений, обусловленная низким отношением регистрируемого сигнала к шуму, которое ограничивает точность при спектральных измерениях на характеристических частотах молекул глюкозы крови в кровеносных капиллярах, расположенных на глубине 0,5-1 мм от поверхности эпидермиса.
Известен метод оптико-калориметрической спектроскопии, в основе которого лежит поглощение света с возбуждением уровней энергии молекул вещества биологического объекта, последующая безызлучательная релаксация уровней и нагрев объекта. В отличие от метода изотермической калориметрической спектроскопии, описанной выше, в методе оптико-калориметрической спектроскопии температура вещества является переменной величиной; информационным параметром в этом методе является изменение температуры исследуемого образца. Степень нагрева определяется величиной поглощающей способности вещества, интенсивностью света и эффективностью конкурирующих процессов (флуоресценции, фотохимического и фотоэлектрического эффектов). Главное достоинство оптико-калориметрических методов состоит в возможности регистрации спектров поглощения сильно рассеивающих сред, что очень важно в биологии и медицине. Поскольку измеряемым параметром является изменение температуры, то в качестве детекторов применяются неселективные приемники излучения, т.е. отсутствует ограничение по длинам волн со стороны приемника.
Однако, ни одна из существующих неинвазивных непрерывных методик мониторинга сахара в крови не позволяет в реальном времени точно отслеживать содержание и временную динамику биохимических компонентов живой ткани, которые являются основным фактором, используемым для ранней диагностики и терапии сахарного диабета.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Техническая задача и технический результат настоящего изобретения состоят в разработке принципиально нового метода спектроскопии живой ткани в условиях in vivo - абсорбционной изотермической калориметрической спектроскопии, позволяющей определять в реальном времени содержание биохимических компонентов живой ткани, и устройства, позволяющего осуществить этот метод.
Разработанные способ и устройство для неинвазивного непрерывного мониторинга сахара в крови применяются для ранней диагностики сахарного диабета и мониторинга сахара в крови в процессе сахаропонижающей терапии и основаны на использовании биосенсоров нового поколения.
Дополнительной технической задачей и техническим результатом настоящего изобретения является создание нового класса биосенсоров и биологических катализаторов для применения в самых разных областях человеческой деятельности, помимо ранней диагностики диабета: в сферах разработки лекарственных препаратов нового поколения, биомедицины и белковой и генной инженерии, пищевой промышленности, безопасности и мониторинга качества окружающей среды.
Одним из перспективных направлений применения группы изобретений является создание и внедрение в клиническую практику новых эффективных и безопасных лекарственных средств, в частности, одной из перспективных мишеней для создания новых противодиабетических средств является глюкокиназа. Дополнительной технической задачей и техническим результатом настоящего изобретения является разработка способа получения биополимерной молекулы с заданными биологическими свойствами и заданной пространственной структурой. Предлагаемый способ получения биополимерной молекулы позволяет определить первичную структуру биополимерной молекулы, которая при заданном биохимическом составе растворителя, при определенных условиях (температура, давление, кислотность), может свернуться в биополимерную молекулу с гетерофазной структурой, обладающую функциями биосенсора и/или биологического катализатора, с биоселективностью к заданной молекуле S растворителя, катализирующего биохимическую реакцию с участием заданной молекулы S. Полимерная цепь биополимерной молекулы с заданными свойствами, структура которой определяется предлагаемым способом, на следующем этапе технологической цепочки синтезируется методами белковой и/или генной инженерии. На основе биополимерных молекул с гетерофазной структурой, полученных предлагаемым способом, могут быть созданы новое поколение биологических катализаторов и эффективных и безопасных лекарственных средств для терапии, в частности, сахарного диабета и других хронических социально значимых заболеваний. Кроме того, предлагаемый способ включает этап определения трехмерной пространственной структуры и биологических свойств биополимерной молекулы. Предлагаемый метод определения трехмерной пространственной структуры и биологических свойств биополимерной молекулы на основе известной первичной структуры полимерной цепи имеет важное практическое применение для разработки новых лекарственных препаратов и новых методов лечения. Проблема предсказания пространственной структуры биополимерной молекулы и ее связь с биологической функцией является одной из крупнейших нерешенных проблем современной науки, решение которой сделало возможным создание принципиально новых биотехнологий, которые могут применяться в белковой и генной инженерии.
Предлагаемый способ и устройство для его осуществления, ориентированные на неинвазивный непрерывный мониторинг сахара крови, позволяют определять содержание сахара в крови путем измерения с помощью принципиально нового метода спектроскопии живой ткани в условиях in vivo - абсорбционной изотермической калориметрической спектроскопии, позволяющая определять в реальном времени содержание биохимических компонентов живой ткани. Величина мощности электромагнитного излучения на характеристической частоте исследуемого биохимического компонента, поглощенной в локальном объеме вещества живой ткани, измеряется не по изменению температуры ткани, а по изменению величины осмотического давления (или количества воды в межклеточном пространстве), характеризующего объем межклеточного вещества, находящегося в гетерофазном конденсированном состоянии, при температуре фазового перехода (соответствующей термодинамическому равновесию кристаллической и жидкой фаз межклеточного вещества). При этом величина поглощенной мощности электромагнитного излучения, падающей на поверхность исследуемого локального участка эпидермиса живой ткани, определяется по количеству межклеточного вещества, изменившего фазовое состояние (плавление глобулярной фазы гиалуроновой кислоты), методом измерения в реальном времени осмотического давления межклеточного вещества.
Предлагаемый способ ранней диагностики и терапии сахарного диабета может быть реализован с помощью принципиально нового метода спектроскопии живой ткани в условиях т vivo - абсорбционной изотермической калориметрической спектроскопии, позволяющая определять в реальном времени содержание биохимических компонентов живой ткани.
Указанный технический результат достигается путем осуществления способа изотермической калориметрической спектроскопии биохимических компонентов живой ткани пациента, включающего следующие этапы:
- накладывают на поверхность кожи пациента с дозированным давлением по меньшей мере один тепло- и водонепроницаемый аппликатор, образующий закрытую систему в локальной области ткани под аппликатором;
- оказывают локальное воздействие на участок ткани под аппликатором электромагнитным излучением на одной или нескольких длинах волн, соответствующих характеристическим частотам поглощения биохимических компонентов межклеточного и/или внутриклеточного вещества;
- измеряют величину физиологического параметра, характеризующего термодинамическое фазовое состояние межклеточного вещества под аппликатором и ее временную динамику, в зависимости от мощности падающего электромагнитного излучения;
- определяют концентрацию биохимического компонента межклеточного вещества и/или межклеточной жидкости и/или крови и ее временную динамику на основе временной динамики измеренного физиологического параметра.
В частных вариантах воплощения изобретения интенсивность электромагнитного излучения может быть постоянной или переменной, изменяющейся с постоянной скоростью и/или модулированной частотой и/или амплитудой.
В частных вариантах воплощения изобретения физиологическим параметром, характеризующим термодинамическое фазовое состояние межклеточного вещества, является осмотическое давление межклеточного вещества и/или количество воды в межклеточном пространстве ткани и/или эластическое давление живой ткани под аппликатором.
В частных вариантах воплощения изобретения биохимические компоненты межклеточного и/или внутриклеточного вещества выбирают из группы, включающей в себя воду, гиалуроновую кислоту, глюкозу, триглицериды и другие биохимические компоненты межклеточного вещества, клетки и крови.
В частных вариантах воплощения изобретения длину волны воздействующего на участок ткани электромагнитного излучения выбирают из диапазона электромагнитного излучения, который определяют по характеристическим частотам поглощения биохимических компонентов живой ткани в оптическом и/или ближнем инфракрасном и/или среднем и дальнем инфракрасном и/или терагерцовом и/или микроволновом диапазоне.
В частных вариантах воплощения изобретения концентрацию биохимического компонента межклеточного вещества и/или межклеточной жидкости и/или крови дополнительно измеряют на основе метода спектроскопии диффузного отражения.
В частных вариантах воплощения изобретения концентрацию биохимического компонента межклеточного вещества и/или межклеточной жидкости и/или крови дополнительно измеряют на основе метода спектроскопии комбинационного рассеяния света.
В частных вариантах воплощения изобретения биохимическим компонентом является глюкоза в крови, концентрацию которой определяют по концентрации глюкозы, связанной с мономерами полимерной цепи межклеточного вещества.
В частных вариантах воплощения изобретения осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют на основе спектральных измерений на длинах волн, соответствующих характеристическим частотам поглощения биохимических компонентов межклеточного и/или внутриклеточного вещества.
В частных вариантах воплощения изобретения спектральные измерения основаны на методе двухчастотной спектроскопии, в которой длина волны электромагнитного излучения выбирается по характеристической частоте тетрамера гиалуроновой кислоты межклеточного вещества в терагерцовой области, а длина волны спектрального датчика осмотического давления межклеточного вещества выбирается по характеристическим частотам поглощения воды в роговом слое эпидермиса в инфракрасной области.
В частных вариантах воплощения изобретения осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса на основе измерения физических характеристик рогового слоя эпидермиса, которые выбирают из группы, включающей в себя электрофизические характеристики, спектральные и оптико-акустические характеристики, теплофизические характеристики.
В частных вариантах воплощения изобретения осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса на основе измерений спектральных характеристик рогового слоя эпидермиса на длинах волн, соответствующих характеристическим частотам воды в роговом слое, с помощью спектрального метода, который выбирают из группы, включающей: ИК спектроскопию, Раман - спектроскопию, оптико-акустическая спектроскопию, разностную двухлучевую спектроскопию.
В частных вариантах воплощения изобретения осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса путем измерения электрических характеристик рогового слоя эпидермиса, которые выбирают из группы, включающей в себя поперечную электропроводность рогового слоя на постоянном и/или переменном токе и диэлектрическую проницаемость.
В частных вариантах воплощения изобретения концентрацию глюкозы в крови определяют по на основе временной динамики физиологического параметра, причем физиологическим параметром является осмотического давления межклеточного вещества области ткани под аппликатором, причем указанное межклеточное вещество под аппликатором является природным биосенсором с гетерофазной структурой с селективностью к молекуле глюкозы и чувствительностью к внешним тепловым потокам, при этом индивидуальную калибровку локальной области ткани пациента определяют по содержанию гиалуроновой кислоты в межклеточном веществе и количества воды в объеме ткани под аппликатором, измеряемых спектральным методом.
В частных вариантах воплощения изобретения способ применим для ранней диагностики сахарного диабета на основе временной динамики концентрации биохимического компонента.
В частных вариантах воплощения изобретения способ применим для ранней диагностики сахарного диабета на основе временной динамики осмотического давления межклеточного вещества.
Указанный технический результат также достигается за счет разработки и создания устройства для изотермической калориметрической спектроскопии биохимических компонентов живой ткани, содержащего тепло и водонепроницаемый аппликатор, выполненный с возможностью наложения на кожу пациента с дозированным давлением, датчик температуры, один или несколько датчиков физиологических параметров, характеризующих термодинамическое фазовое состояние межклеточного вещества под аппликатором, один или несколько источников электромагнитного излучения, устройство для создания калибровочного воздействия, при этом датчики физиологических параметров расположены под аппликатором, при этом сигналы с вышеуказанных датчиков поступают последовательно на входы блока усилителей и/или синхронного детектора и/или аналогово-цифрового преобразователя, установленных на верхней поверхности аппликатора, в блок обработки информации и блок отображения информации.
В частных вариантах воплощения изобретения источник электромагнитного излучения выполнен с возможностью излучения с постоянной и/или модулированной интенсивностью и/или частотной модуляцией.
В частных вариантах воплощения изобретения датчики физиологических параметров, характеризующие термодинамическое состояние межклеточного вещества под аппликатором, выполнены в виде датчиков осмотического давления межклеточного вещества или датчиков количества воды в межклеточном пространстве ткани в локальном объеме под аппликатором и/или датчика эластического давления живой ткани под аппликатором.
В частных вариантах воплощения изобретения устройство для создания калибровочного воздействия представляет собой источник тепловой мощности, выполненный в виде резистора и/или элемента Пельтье и/или источник электрического тока и/или напряжения; устройство для создания дозированного давления на поверхность аппликатора.
В частных вариантах воплощения изобретения диапазон излучения источников электромагнитного излучения выбирают из группы, включающей в себя оптический и/или ближний инфракрасный; средний инфракрасный; дальний инфракрасный и/или терагерцовый; микроволновый диапазон излучения.
В частных вариантах воплощения изобретения датчиком осмотического давления межклеточного вещества или датчиком количества воды в межклеточном пространстве ткани в локальном объеме под аппликатором представляет собой электрометрический датчик, основанный на измерении электрофизических характеристик рогового слоя эпидермиса и ткани под аппликатором, принцип измерения которого выбирается из группы, включающей измерение поперечной электропроводности рогового слоя эпидермиса на постоянном и/или переменном токе; измерение диэлектрической проницаемости рогового слоя эпидермиса; измерение электропроводности и/или диэлектрической проницаемости ткани под аппликатором.
В частных вариантах воплощения изобретения датчиком осмотического давления межклеточного вещества или датчиком количества воды в межклеточном пространстве ткани в локальном объеме под аппликатором представляет собой электрометрический датчик, основанный на измерении электрофизических характеристик рогового слоя эпидермиса и ткани под аппликатором, принцип измерения которого выбирается из группы, включающей измерение поперечной электропроводности рогового слоя эпидермиса на постоянном и/или переменном токе; измерение диэлектрической проницаемости рогового слоя эпидермиса; измерение электропроводности и/или диэлектрической проницаемости ткани под аппликатором.
В частных вариантах воплощения изобретения длину волны электромагнитного излучения источника спектрального датчика определяют по характеристическим частотам поглощения биохимических компонентов межклеточного вещества и/или внутриклеточного вещества, которые выбирают из группы, включающей в себя воду, гиалуроновую кислоту, глюкозу и другие биохимические компоненты межклеточного вещества и межклеточной жидкости.
В частных вариантах воплощения изобретения датчиком осмотического давления межклеточного вещества является спектральный датчик, основанный на спектральных измерениях характеристик межклеточного вещества в терагерцовом диапазоне на длине волны, соответствующей энергии поперечной связи между мономерами полимерной цепи гиалуроновой кислоты.
В частных вариантах воплощения изобретения спектральный датчик основан на спектральных измерениях количества воды в роговом слое эпидермиса по характеристическим частотам воды в роговом слое на основе спектрального метода, который выбирают из группы, включающей в себя изотермическую калориметрическую спектроскопию, абсорбционную спектроскопию, спектроскопию диффузионного отражения, спектроскопию комбинационного рассеяния, оптико-акустическую спектроскопию.
В частных вариантах воплощения устройство по изобретению дополнительно содержит спектральное устройство, включающее источник и приемник электромагнитного излучения, для спектральных измерений на основе диффузионной отражательной спектроскопии.
В частных вариантах воплощения изобретения датчик физиологических параметров, характеризующего термодинамическое фазовое состояние межклеточного вещества под аппликатором, выполнен на пьезоэлектрическом способе измерения.
В частных вариантах воплощения изобретения устройство применимо для ранней диагностики сахарного диабета на основе временной динамики концентрации биохимического параметра и/или величины осмотического давления межклеточного вещества.
Указанный технический результат также достигается путем осуществления способа получения биополимерной молекулы с заданными биологическими свойствами и заданной пространственной структурой, состоящей из одной или нескольких субъединиц, способной к самопроизвольной укладке в пространственную конфигурацию с гетерофазной структурой, обладающей специфичностью к одной или нескольким заданным молекулам субстрата S, при определенном составе растворителя, при определенных условиях по температуре и давлению, заключающийся в том, что число субъединиц определяют на основе трехмерной пространственной структуры макромолекулы, при этом первичную структуру каждой субъединицы биополимерной молекулы определяют следующим образом:
- определяют или задают константу равновесия процесса связывания молекулы субстрата S с мономером полимерной цепи биополимерной молекулы посредством образования водородной связи;
- определяют энергию активации и/или энергию водородной связи молекулы субстрата S с мономером полимерной цепи биополимерной молекулы, соответствующую величине константы равновесия;
- по величине энергии активации и/или энергии связи идентифицируют мономеры, которые образуют первичную структуру каждой субъединицы биополимерной макромолекулы, а именно, по меньшей мере, пару мономеров 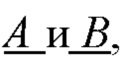 образующих повторяющийся мономер
образующих повторяющийся мономер 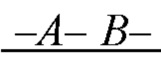 полимерной цепи, в которой А - мономер, содержащая группу, обладающую отрицательным электрическим зарядом, и В - нейтрально незаряженный мономер, с которым молекула субстрата S может образовать водородную связь, энергия которой соответствует заданной константе равновесия;
полимерной цепи, в которой А - мономер, содержащая группу, обладающую отрицательным электрическим зарядом, и В - нейтрально незаряженный мономер, с которым молекула субстрата S может образовать водородную связь, энергия которой соответствует заданной константе равновесия;
- по величине энергии активации и/или энергии связи идентифицируют одновалентные ионы растворителя, соответствующие двум разным соединениям, способным образовать слабую ионную связь с заряженным мономером, при этом один из ионов, ион М, выбирается с константой равновесия близкой к константе равновесия субстрата S;
- определяют кислотность растворителя рН, определяющую отрицательный заряд мономера А, концентрации ионов, температуру и давление;
- определяют число мономеров в цепи;
- получают биополимерную молекулу с заданной первичной структурой с помощью методов генной и/или белковой инженерии;
- получают раствор биополимерной молекулы в растворителе, пространственная структура и характеристики которого соответствуют заданным.
В частных вариантах воплощения изобретения мономер А полимерной цепи обладает положительным зарядом, а ион М растворителя обладает отрицательным зарядом.
В частных вариантах воплощения изобретения ионом M растворителя является одновалентным положительно заряженный ион металла.
В частных вариантах воплощения изобретения первичной структурой биополимерной макромолекулы, состоящей из одной субъединицы, является полисахаридная цепь, повторяющийся мономер которого содержит, по меньшей мере, одну разновидность дисахаридной пары, образованной одним моносахаридом А с отрицательно заряженной группой и одним нейтральным моносахаридом В, не обладающим заряженной группой.
В частных вариантах воплощения изобретения первичной структурой биополимерной макромолекулы, состоящей из одной субъединицы, является полимерная цепь гиалуроновой кислоты, молекулой субстрата является молекула глюкозы, одновалентным ионом металла М является ион натрия, при этом химический состав растворителя близок к химическому составу плазмы крови и межклеточной среды живой системы, при физиологических условиях in vivo.
В частных вариантах воплощения изобретения в состав растворителя добавляют двухвалентный ион и/или вещество, молекула которого обладает способностью образовать двухвалентную ионную связь с отрицательно заряженными мономерами полимерной цепи гиалуроновой кислоты.
В частных вариантах воплощения изобретения биополимерной молекулой, состоящей из одной субъединицы, является полипептидная цепь из аминокислот, с повторяющимися мономерами, содержащими по меньшей мере одну аминокислоту с отрицательно заряженной группой Ra и одну аминокислоту с нейтральной незаряженной группой Rb, одновалентным ионом металла М является ион калия, при этом химический состав растворителя близок к химическому составу внутриклеточной среды живой системы при физиологических условиях in vivo.
В частных вариантах воплощения изобретения в состав растворителя добавляют дополнительные двухвалентные ионы металла или двухвалентные соединения, которые выбирают из группы, включающей ион магния Mg+2, марганца Mn+2, ионы других металлов и соединений.
В частных вариантах воплощения изобретения субстратом является молекула D- глюкозы и/или молекула другого моносахарида.
В частных вариантах воплощения изобретения биополимерная молекула с гетерофазной структурой представляет собой биосенсор с избирательностью к заданной молекуле субстрата S, активный в водной среде, преобразующий сигнал концентрации субстрата S в сигнал, пропорциональный объему макромолекулы и/или внутримолекулярному осмотическому давлению, который измеряется электронным устройством.
В частных вариантах воплощения изобретения биополимерной молекулой является полисахарид гиалуроновой кислоты, обладающий избирательностью к молекуле глюкозы.
В частных вариантах воплощения изобретения биополимерная молекула с заданными биологическими свойствами, состоящая из двух субъединиц, способная к самопроизвольной укладке в пространственную конфигурацию с гетерофазной структурой, обладающая специфичностью к заданной молекуле субстрата S, при определенном составе растворителя, при определенных условиях по температуре и давлению, и способ заключается в том, что для каждой из двух субъединиц определяется первичная структура следующим образом:
- определяют или задают константу равновесия процесса связывания молекулы субстрата S с мономером полимерной цепи биополимерной молекулы посредством образования водородной связи;
- определяют энергию активации и/или энергию водородной связи молекулы субстрата S с мономером полимерной цепи биополимерной молекулы, соответствующую величине константы равновесия;
- по величине энергии связи идентифицируют мономеры, которые образуют первичную структуру первой субъединицы биополимерной макромолекулы, а именно, по меньшей мере, пару мономеров 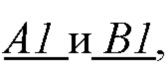 образующих повторяющийся мономер
образующих повторяющийся мономер 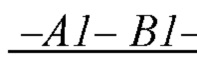 полимерной цепи, в которой А1 - мономер, содержащая группу, обладающую отрицательным электрическим зарядом, и В1 - нейтрально незаряженный мономер, с которым молекула субстрата S может образовать водородную связь, энергия которой соответствует заданной константе равновесия;
полимерной цепи, в которой А1 - мономер, содержащая группу, обладающую отрицательным электрическим зарядом, и В1 - нейтрально незаряженный мономер, с которым молекула субстрата S может образовать водородную связь, энергия которой соответствует заданной константе равновесия;
- идентифицируют мономеры, которые образуют первичную структуру второй субъединицы биополимерной макромолекулы, а именно, по меньшей мере, пару мономеров 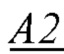 и
и 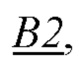 образующих повторяющийся мономер
образующих повторяющийся мономер 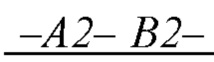 полимерной цепи, в которой А2 - мономер, содержащая группу, обладающую положительным электрическим зарядом, и В2 - нейтрально незаряженный мономер, с которым молекула субстрата S может образовать водородную связь, энергия которой соответствует заданной константе равновесия;
полимерной цепи, в которой А2 - мономер, содержащая группу, обладающую положительным электрическим зарядом, и В2 - нейтрально незаряженный мономер, с которым молекула субстрата S может образовать водородную связь, энергия которой соответствует заданной константе равновесия;
- по величине энергии связи идентифицируют одновалентные ионы растворителя, соответствующие двум разным соединениям или веществам, способным образовать слабую ионную связь с заряженными мономером, при этом один из ионов, ион М, выбирается с константой равновесия близкой к константе равновесия субстрата S;
- определяют кислотность растворителя рН, определяющую отрицательный заряд мономера А, концентрации ионов, температуру и давление;
- определяют число мономеров в цепи;
- получают биополимерную цепь, состоящую из двух субъединиц, с заданной первичной структурой с помощью методов генной и/или белковой инженерии;
- получают раствор биополимерной молекулы в растворителе, характеристики которого соответствуют заданным.
В частных вариантах воплощения изобретения субъединицами являются полипептидные цепи из аминокислот.
В частных вариантах воплощения изобретения для каждой субъединицы определяют расположение в цепи повторяющихся мономеров, характеризующих взаимодействие между субъединицами, образующих уникальную аминокислотную конфигурацию активного центра белковой молекулы, который образуется в процессе пространственной укладки в области пространства между доменами субъединиц.
В частных вариантах воплощения изобретения биополимерная молекула с гетерофазной структурой представляет собой биологический катализатор с заданными характеристиками и избирательностью к заданной молекуле субстрата S, активный в водной среде, катализирующий биохимические реакции с участием молекулы субстрата S.
В частных вариантах воплощения изобретения биополимерная молекула представляет собой фермент с заданными характеристиками на основе биополимерной молекулы с гетерофазной структурой.
В частных вариантах воплощения изобретения первичная структура биополимерной молекулы с заданными биологическими свойствами определяется по характеристикам фермента гексокиназы, константе равновесия и скорости биохимической реакции, которую катализирует фермент.
В частных вариантах воплощения изобретения первичная структура биополимерной молекулы с заданными биологическими свойствами определяется по характеристикам фермента глюкокиназы, константе равновесия и скорости биохимической реакции, которую катализирует фермент.
В частных вариантах воплощения изобретения при заданном составе растворителя и заданной первичной структуре биополимерной молекулы, состоящей из 2 и более субъединиц, определяют четвертичную пространственную структуру макромолекулы следующим образом:
- определяют аминоконцевые группы белковой биополимерной молекулы, которыми отделены полипептидные цепи субъединиц, образующих первичную структуру белковой биополимерной молекулы;
- определяют для каждой субъединицы структуру элементарной цепочки взаимодействия, характеризующей взаимодействие полимерной цепи с растворителем; для этого определяют молекулу субстрата и идентифицируют одновалентный ион в составе растворителя для каждой субъединицы;
- для каждой субъединицы вычисляют константу равновесия, энергию активации и определяют объем трехмерного домена;
- определяют наиболее вероятную трехмерную пространственную конфигурацию доменов, соответствующих субъединицам.
В частных вариантах воплощения изобретения биополимерной молекулой, первичная структура которой состоит из 2 и более субъединиц, является олигомерный белок.
Настоящее изобретение также включает биополимерную молекулу с гетерофазной структурой, представляющую собой биосенсор с селективностью к заданной молекуле субстрата и полученная способом по изобретению, проявляющая активность в водной среде.
Настоящее изобретение также включает биополимерную молекула с гетерофазной структурой, обладающая избирательностью к заданной биологической молекуле субстрата S, представляющую собой биологический катализатор для катализа биохимической реакции с участием молекулы субстрата S, полученную способом по изобретению. Биополимерная молекула с гетерофазной структурой, обладающая избирательностью к заданной молекуле субстрата S, представляющая собой фермент для катализа биохимической реакции с участием молекулы субстрата S, полученная способом по изобретению.
В частных вариантах воплощения изобретения биополимерная молекула характеризуется специфичностью к молекуле глюкозы, а также тем, что:
- биополимерная молекула представляет собой сополимер, которым является полисахарид гиалуроновой кислоты;
- компонентами растворителя с заданной кислотностью одновременно являются ионы натрия и калия, молекула глюкоза.
В частных вариантах воплощения изобретения биополимерная молекула характеризуется специфичностью к молекуле глюкозы, и отличается тем, что биополимерная молекула представляется собой полипептидную цепь, повторяющийся мономер которой содержит аминокислоту с отрицательно заряженной боковой группой, которую выбирают из группы, состоящей из глутаминовой кислоты, аспарагиновой кислоты, глицина, цистеина, тирозина, при этом, аминокислоту с незаряженной боковой группой выбирают из группы, содержащей серин, аспарагин, треонин, глутамин.
В частных вариантах воплощения изобретения аминокислотой с отрицательным зарядом боковой группы является глутаминовая кислота; аминокислотой с незаряженной боковой группой является аспарагин; субстратом биохимической реакции является D-глюкоза.
В частных вариантах воплощения изобретения аминокислотой с отрицательным зарядом боковой группы является аспарагиновая кислота; аминокислотой с незаряженной боковой группой является аминокислота, которую выбирают из группы аспарагин, треонин, цистеин, серин; субстратом биохимической реакции является D-глюкоза.
В частных вариантах воплощения изобретения полипептидная цепь дополнительно содержит аминокислоты с положительно заряженными боковыми группами, которые выбирают из группы лизин, аргинин, гистидин.
В частных вариантах воплощения изобретения растворитель дополнительно содержит двухвалентный ион металла, который выбирают из группы, включающей ион магния Mg+2, марганца Mn+2, ионы других металлов и соединений или другое соединение.
В частных вариантах воплощения изобретения полипептидная цепь первой субъединицы содержит аминокислоту с отрицательно заряженной группой бокового остатка, а полипептидная цепь второй субъединицы содержит аминокислоту с положительно заряженной группой бокового остатка.
В частных вариантах воплощения изобретения аминокислотами активного центра белка являются серин, боковой остаток которого обладает отрицательно заряженной группой, и гистидин, боковой остаток которого обладает положительно заряженной группой.
В частных вариантах воплощения изобретения биополимерная молекула является молекулой для создания биополимерного вещества или фармацевтической композиции на основе гиалуроновой кислоты для лечения ран и воспалений тканей.
В частных вариантах воплощения изобретения биополимерная молекула является молекулой для создания лекарственного препарата или фармацевтической композиции для лечения сахарного диабета.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Прилагаемые чертежи, которые включены в состав настоящего описания и являются его частью, иллюстрируют варианты осуществления группы изобретения и совместно с общим описанием группы изобретения и нижеприведенным подробным описанием вариантов осуществления служат для пояснения принципов настоящего изобретения.
На рисунке 1 показана пространственная (третичная) структура глобулярного белка.
На рис. 2 изображен повторяющийся дисахаридный мономер полимерной цепи гиалуроновой кислоты.
На рис. 3а изображен график зависимости осмотического давления межклеточного вещества эпидермиса человека от мощности падающего инфракрасного теплового излучения (с максимумом спектральной мощности на длине волны λ≈10 мкм).
На рис. 3б изображен график зависимости осмотического давления межклеточного вещества эпидермиса человека от концентрации глюкозы в крови. По оси абсцисс отложена концентрация глюкозы в ммоль/литр; по оси ординат отложена функция Р0 - Росм, где Р0 - константа, значение которой определяется индивидуальной калибровкой. Росм - осмотическое давление межклеточного вещества эпидермиса.
На рис. 4 изображен график зависимости температуры произвольного вещества от количества введенного в систему тепла (heating curve).
На рис. 5 изображена наиболее вероятная конфигурация объединенной системы (слева) полимерная цепь - растворитель, которая реализуется равномерным распределением N/2 адсорбированных ионов Na+ по узлам цепи и однородным распределением N/2 свободных ионов Na+ и N/2 свободных отрицательно заряженных мономеров М- в объеме макромолекулы; справа показана наименее вероятная конфигурация максимальной линейной плотности энтропии (справа).
На рис. 6 показана конфигурации полимерной сетки и сетки водородных связей воды. Сшивки полимерной сетки посредством водородных связей могут реализоваться в разных возможных конфигурациях тетрамера.
На рис. 7 изображена микроскопические формы (G) тетрамера, соответствующие разным числам водородных связей и дискретным значениям энергии связи тетрамера.
На рис. 7а изображена конфигурация (G) тетрамера в равновесных конформациях (R) и (Т), переходящих друг в друга в процессе теплового движения.
На рис. 7б изображена равновесная конфигурация (Na) тетрамера, образованного из 6 звеньев (1-6) не соседних участков цепи (1-5-4 и 2-6-3).
На рис. 8 изображена наиболее вероятная равновесная конфигурация полимерной цепи в растворителе с избытком ионов Na и K, при нулевой концентрации глюкозы: узлы с ионами K, показаны желтым, узлы с ионами Na показаны синим.
На рис. 8а изображена наиболее вероятная конфигурация взаимодействующих элементарных цепочек, которая реализуется в результате спаривания цепочек посредством водородных связей в конфигурации R4  тетрамера;
тетрамера;
На рис. 8б изображена конфигурация квантования внутримолекулярного пространства, возникающая в результате объемных взаимодействий пары элементарных цепочек.
На рис. 8в изображено случайное распределение равновозможных конфигураций тетрамера (Na) в микроскопических ячейках T(R4), D(R2), M(R0), элементарной ячейки: конфигурация R4 в ячейке Т(2).
На рис. 9 изображена функции распределения Θ(α, γ=8) и Θ(α, γ = 139,740210…), полученные из универсальной функции распределения Θ(α, γ), описывающей зависимость энтропии (свободной энергии объемных взаимодействий) полимерной цепи, от состава растворителя, для разных значений константы взаимодействия (параметра порядка) γ; безразмерная концентрация глюкозы α=С/KR выражена в единицах микроскопической константы равновесия KR.
На рис. 10 изображены изомерные конформации (Na) тетрамера образованные посредством водородных связей. Справа схематически показана возможная трехмерная конфигурация изомерной конформации (s), образованной из 6 звеньев полимерной сети, расположенных на плоскости, ориентированной перпендикулярно к прямой, проходящей через 2 молекулы глюкозы.
На рис. 10а изображено распределение мономеров в объеме микроскопической ячейки при концентрациях 2 и 3 мономера/1 микроячейка.
На рис. 11а изображен график относительной энтропии полимерной цепи в наиболее вероятной конфигурации, соответствующей минимуму свободной энергии элементарного объемного взаимодействия, для термодинамических фаз (λ) и (s) от концентрации глюкозы растворителя.
На рис. 11б изображен график свободной энергии объемных взаимодействий в зависимости от концентрации глюкозы.
На рис. 12 изображена диаграмма энергетических уровней тетрамеров (λ, s, g) полимерной сетки природной макромолекулы.
На рис. 13 изображены графики функций распределения (g) процесса: Θ1g (a) - зеленая кривая; Θ2g (α) - синяя кривая; Θ3g (α) - красная кривая.
На рис. 14 изображены графики функций распределения Θ2(α) незаселенных (λ, s, g) тетрамеров: (λ) тетрамер - красная кривая; (s) тетрамер - синяя кривая; (g) тетрамер - фиолетовая кривая.
На рис. 15 изображены графики функции распределения димеров и тетрамеров (λ, g, s): (А,) тетрамер - фиолетовая Θ1λ (α) и красная Θ2λ (α) кривые; (g) тетрамер - голубая Θ1g (α) и синяя Θ2g (α) кривые; (s) тетрамер - темно зеленая Θ1s (α) и зеленая Θ2s (α) кривые.
На рис. 16 изображены графики функции энергия отталкивания идеального газа свободных мономеров в конденсированных фазах (g, λ, s): Ug rep (α) - фаза G (зеленая); Uλ rep (α) - фаза λ (красная); Us rep (α) - фаза s (синяя).
На рис. 17 изображен график функции энергии элементарного объемного взаимодействия (отталкивания) Uλs rep (α) свободных мономеров в фазах (λ, s).
На рис. 18 изображены графики функций энергии парных взаимодействий и внутреней энергия идеального газа свободных мономеров для каждой конденсированной фазы (λ, s, g). На рисунке показаны графики 6 функций: 3 функции энергии притяжения Ug attr (α), Us attr (α); Uλ attr (α); 3 функции энергии отталкивания Ug rep (α), Uλ rep (α), Us rep (α).
На рис. 19 изображены графики энергии элементарного объемного взаимодействия от концентрации глюкозы растворителя: энергия отталкивания точно равна энергии притяжения при любых концентрациях глюкозы растворителя Uλs attr (α) + Uλs rep (α) = 0. На рисунке показаны графики 2 функций, энергии притяжения Uλs attr (α) и энергии отталкивания Uλs rep (α), которые точно равны при любых концентрациях глюкозы растворителя.
На рис. 20 изображен график осмотического давления Posm×V0g, в зависимости от концентрации глюкозы растворителя, при критическом значении микроскопической константы равновесия kλ2 = 5,032…
На рис. 21 изображены графики энергии элементарных объемных взаимодействий Uλs(a) (кривая красного цвета) и обратимая работа сжатия А(α) (синяя кривая), совершаемая за счет свободной химической энергии (водородных связей) мономеров (G) над идеальным газом свободных мономеров, при критическом значении микроскопической константы равновесия kλ2 = 5,032…
На рис. 22 изображен график функции энтальпии макромолекулы от концентрации глюкозы растворителя, при критическом значении константы равновесия kλ2 = 5,032….
На рис. 23 изображен график изотермы энтальпия - концентрация глюкозы в трехфазной области макромолекулы, при критическом значении микроскопической константы равновесия kλ2 = 5,032…, при которой газообразная, жидкая и кристаллические формы могут сосуществовать; газообразная фаза - идеальный газ свободных мономеров полимерной цепи, конденсированная жидкая фаза (λ), конденсированная кристаллическая фаза (s).
На рис. 24 приведена схема изотермического калориметра фазового перехода.
На рис. 25 изображена схема, поясняющая принцип изотермического микрокалориметра живой ткани.
На рис. 26 показана блок схема, поясняющая принцип работы биосенсора глюкозы межклеточного вещества.
На рис. 27 представлена схема спектрального устройства для изотермической калориметрической спектроскопии (ICS) биохимических компонентов межклеточного вещества живой ткани in vivo.
На рис. 28 изображена диссоциация тетрамера, в результате поглощения кванта энергии терагерцового электромагнитного излучения, сопровождается образованием кванта тепловой энергии
На Рис. 29 показана схема изотермической калориметрической спектроскопии биохимических компонентов живой клетки.
На рис. 30 представлена схема абсорбционной изотермической калориметрической спектроскопии живой клетки.
На рис. 31 изображен спектр поглощения воды в широком диапазоне частот (от ультрафиолетового до микроволнового).
На рис. 32 изображены спектры поглощения глюкозы в ИК области и пропускания в ближней ИК области в интервале длин волн от 1,428 мкм (7000 см-1) до 20 мкм (500 см-1).
На рис. 33 изображен спектр поглощения D-глюкозы в ближней ИК области для разных значений концентрации глюкозы.
На рис. 34 схематически показана структура межклеточного вещества, представляющего разветвленную макромолекулу гиалуроновой кислоты, имеющую конфигурацию полимерной сетки, окружающей живую клетку.
На рис. 35 изображены спектральные линии поглощения глюкозы в инфракрасной области спектра, соответствуют характеристическим частотам валентных связей молекулы: С - Н; О - Н; С=O.
На рис. 36 представлены ИК спектры гиалуроновой кислоты, глюкозы, белка и липида: 1 - триглицерид жирной кислоты (бараний жир); 2 - гиалуроновая кислота; 3 - белок (яичный альбумин); 4 - глюкоза.
На рис. 37 изображена схема спектроскопии диффузного отражения для измерения содержания гиалуроновой кислоты и свободных жирных кислот в межклеточном веществе поверхностного слоя эпидермиса живой ткани.
На рис. 38 представлен спектр гиалуроновой кислоты в ближней ИК области.
На рис. 39 представлены спектры поглощения гиалуроновой кислоты в ближней ИК области и спектр пропускания в ИК области.
На рис. 40 представлены теоретические спектры поглощения глюкозы, меланина и воды в красном и ближнем инфракрасном диапазоне.
На рис. 41 представлено схематическое изображение липидного матрикса рогового слоя.
На рис. 42 изображена принципиальная схема спектрального устройства для реализации способа спектроскопии межклеточного вещества локального участка живой ткани.
На рис. 43 представлена схема спектроскопии диффузного отражения для определения осмотического давления межклеточного вещества на основе спектрального измерения временной динамики количества воды в роговом слое эпидермиса на длине волны, соответствующей характеристической частоте воды (1190, 1455, 1945, 945 нм).
На рис. 44 представлены фотографии прототипа и предполагаемый дизайн коммерческого прибора.
На рис. 45 представлена характерная скачкообразная временная динамика сахара крови у пациента Д с сахарным диабетом 1 типа (33 лет, муж.).
На рис. 46 показана скачкообразная динамика сахара крови в форме одиночного импульса, зарегистрированная с помощью изотермического микрокалориметра, и временная динамика уровня инсулина в крови, рассчитанная на основе сахарной кривой.
На рис. 47 представлены характерная сахарная кривая со скачкообразной временной динамикой у пациента с сахарным диабетом 2 типа (66 лет, муж.): характерная динамика сахара крови в форме временной последовательности одиночных импульсов.
На рис. 48 представлена временная динамика осмотического давления межклеточного вещества в процессе изотермического фазового перехода живой ткани (пациента с сахарным диабетом 2 типа) в процессе растворения кристаллической (глобулярной) фазы межклеточного вещества.
На рис. 49 представлены характерные мониторинговые кривые осмотического давления межклеточного вещества здорового пациента и пациента с сахарным диабетом 2 типа.
На рис. 50 (а-д) показаны примеры сахарных кривых со скачкообразной временной динамикой: Рис. 50 а (тест №6 пациент №24), Рис. 50 б (тест №1 пациент №41), Рис. 50 в (тест №5 пациент №41), Рис. 50 г (тест №6 пациент №41). На всех графиках результаты инвазивных измерений показаны кружочками зеленого цвета, а результаты неинвазивного мониторинга фиолетовой кривой.
На рис. 51 представлены результаты тестов на 14 пациентах с сахарным диабетом 1 и 2 типа.
На рис. 52 представлены результаты тестов на 14 пациентов с диабетом 1 и 2 типа.
На рис. 53 - представлены результаты тестов, выполненных на группе из 42 пациентов (252 тестов, 756 инвазивных измерения), полученные путем объединения результатов подгруппы 1 и подгруппы 2.
На рис. 54 показаны структурные формулы аминокислот.
На рис. 55 показана четвертичная структура белков, состоящих из нескольких полипептидных цепей, возникающая в результате ассоциации нескольких субъединиц белка в пространстве.
На рис. 56 схематически показано изменение пространственной структуры белкового фермента гексокиназы в результате связывания молекулы глюкозы.
На рис. 57 схематически показаны основные этапы ферментативной реакции: а) диффузия молекулы АТФ из жидкой фазы в кристаллическую фазу; b) присоединение фосфатной группы (красный) к молекуле D-глюкозы (желтый) с образованием молекулы G6P в результате химической реакции в кристаллической фазе фермента; d) конформационный переход тетрамера со связанной молекулой G6P из кристаллической фазы в жидкую через энергетический барьер, разделяющий жидкую фазу с кристаллической; g) разрушение тетрамера в жидкой фазе с освобождением молекулы продукта G6P.
На рис. 58а представлены кривые насыщения и скорости ферментативной реакции глюкокиназы, в зависимости от концентрации глюкозы.
На рис. 58б представлен график энергетического барьера глюкокиназы.
На рис. 59а представлены кривые насыщения и скорости ферментативной реакции гексокиназы, в зависимости от концентрации глюкозы.
На рис. 59б представлен график энергетического барьера гексокиназы.
На рис. 60 представлены структурные формулы гликозидной связи с аминокислотами.
На рис. 61 представлены структурные формулы гликозидной связи в нуклеозидах.
На рис. 62 схематически показаны ионные связи двухвалентного иона металла в макромолекуле.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Физические основы теплообмена живой ткани с окружающей средой.
Теплообмен представляет собой самопроизвольный и необратимый процесс переноса теплоты, обусловленный градиентом температуры. Различают следующие виды теплообмена: теплопроводность, конвекция, лучистый теплообмен, теплообмен при фазовых превращениях.
Теплоотдача представляет собой теплообмен между поверхностью тела и соприкасающейся с ней средой - теплоносителем (жидкостью, газом).
Испарительное охлаждение представляет собой теплообмен между тканью и окружающей средой, обусловленный испарением воды, поступающей к поверхности эпидермиса из глубинных слоев ткани. Плотность теплового потока, определяется произведением теплоты испарения (теплоты парообразования) на плотность потока воды, испаряющейся с поверхности.
Лучистый теплообмен (радиационный теплообмен, лучистый перенос) представляет собой перенос энергии от одного тела к другому, обусловленный процессами испускания, распространения, рассеяния и поглощения электромагнитного излучения. Каждый из этих процессов подчиняется определенным закономерностям.
Так, в условиях равновесного теплового излучения испускание и поглощение подчиняются закону излучения Планка, закону излучения Стефана-Больцмана, закону излучения Киргоффа.
Существенное отличие лучистого теплообмена от других видов теплообмена (конвекции, теплопроводности) заключается в том, что он может протекать при отсутствии материальной среды, разделяющей поверхности теплообмена, так как электромагнитное излучение распространяется и в вакууме.
Закон излучения Планка устанавливает связь между интенсивностью излучения, спектральным распределением и температурой черного тела. При увеличении температуры, энергия излучения возрастает. Энергия излучения зависит от длины волны. Суммарная энергия, излучаемая черным телом и измеряемая бесконтактным инфракрасным термометром, является суммарной энергией, излучаемой на всех длинах волн. Она пропорциональна интегралу уравнения Планка по длинам волн и описывается в физике законом Стефана-Больцмана.
Лучистый теплообмен между поверхностью ткани и окружающей средой определяется соотношением:
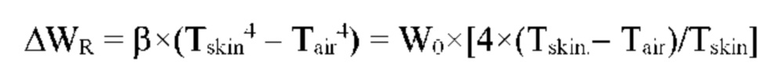
ΔТ << Tskin,
Tskin - температура поверхности кожи,
Tair - температура окружающего воздуха.
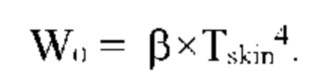
ΔWR - тепловое излучение с поверхности ткани в окружающую среду.
Теплопроводность представляет собой один из видов переноса теплоты от более нагретых частей тела к менее нагретым. Теплопроводность приводит к выравниванию температуры. При теплопроводности перенос энергии осуществляется в результате непосредственной передачи энергии от частиц, обладающих большей энергией, частицам с меньшей энергией. Если относительное изменение температуры Т на расстоянии средней длины свободного пробега частиц мало, то выполняется основной закон теплопроводности (закон Фурье): плотность теплового потока пропорциональна градиенту температуры ΔT:
WΔT = -λ×ΔT,
где λ - коэффициент теплопроводности или теплопроводность, не зависящий от ΔT. Коэффициент λ зависит от агрегатного состояния вещества, его молекулярного строения, температуры, давления, состава и т.д.
Конвекция представляет собой перенос теплоты в жидкостях и газах потоками вещества. Конвекция приводит к выравниванию температуры вещества. При стационарном подводе теплоты к веществу в нем возникают стационарные конвекционные потоки. Интенсивность конвекции зависит от разности температур между слоями, теплопроводности и вязкости среды.
Испарительное охлаждение, как было отмечено выше, представляет собой теплообмен между тканью и окружающей средой, обусловленный испарением воды, поступающей к поверхности эпидермиса из глубинных слоев ткани посредством транспорта воды по межклеточному пространству. Плотность теплового потока определяется произведением теплоты испарения (теплоты парообразования) на плотность потока воды, испаряющейся с поверхности.
Как известно, в комфортной зоне температур в нормальных условиях транспорт воды посредством потоотделения практически отсутствует и основной вклад в процесс испарительного охлаждения определяется транспортом воды к поверхности тела. В физиологии и медицине этот процесс известен как процесс неощутимой перспирации.
Неощутимая перспирация воды наблюдается в диапазоне температур, в котором у испытуемого не возникает субъективных тепловых ощущений (комфортный температурный диапазон):
Температура окружающей среды: 18-23°С,
Атмосферное давление: 730-770 мм. рт. столба.
Известно, что интенсивность процесса испарительного охлаждения в комфортных условиях в норме составляет 400-700 мл/сутки или 10-8-10-7 г/сек×см2. Это соответствует значениям тепловых потоков 1-10 мВт/см2.
Известно, что физиологический процесс неощутимой перспирации обладает высокой чувствительностью к внешним тепловым потокам. Порог чувствительности процесса к тепловым потокам составляет 0,1 мВт/см2, что эквивалентно чувствительности процесса к перепадам температуры внешней среды, равным 0,05 градуса.
Важным следствием высокой чувствительности физиологического процесса неощутимой перспирации к внешним тепловым потокам, имеющим практическое значение, является зависимость физических характеристик эпидермиса кожи человека от внешних физико-климатических факторов, в частности, от температуры и влажности окружающей среды и атмосферного давления. В частности, результаты измерений физиологического параметра эпидермиса кожи человека - скорости неощутимой перспирации, при неизменных значениях микроклимата помещения, в котором проводятся измерения, оказываются зависящими от физико-климатических факторов окружающей среды (температура и влажность окружающей среды, атмосферное давление). Низкочастотные флуктуации температуры внешнего воздуха в пределах даже одного градуса, при неизменных параметрах микроклимата помещения, в котором проводится измерение, приводят к значимым низкочастотным флуктуациям измеряемого физиологического параметра и, как следствие, увеличению погрешности измерений.
Физиологические и биохимические основы теплопродукции живой ткани
Процесс окисления глюкозы, одного из основных поставщиков энергии в организме, может быть представлен в виде следующего уравнения:
Глюкоза + Кислород ⇒ СО2 + H2O.
Изменение стандартной свободной энергии в этой реакции при физиологических условиях равно:
ΔG = -686 ккал/моль.
Для сравнения, мужчина весом в семьдесят килограмм, поднимаясь по лестнице в течение часа, затрачивает примерно 1000 ккал. Отсюда ясно, что упомянутые выше 686 ккал представляют собой огромное количество энергии. Разумеется, работа, производимая человеком, значительно меньше расходуемой при этом энергии, так как при необратимом процессе не все изменение свободной энергии преобразуется в работу. Реальная эффективность этого преобразования, как будет пояснено ниже, не выше 40%. Более того, пища не "сгорает" сразу и непосредственно в кислороде, высвобождая энергию в форме тепла, а это высвобождение происходит поэтапно и включает ряд довольно сложных химических превращений, каждая из которых дает небольшую "порцию" энергии.
Глюкоза окисляется в организме, образуя диоксид углерода и воду; это один из самых универсальных процессов, который лежит в основе процессов дыхания и пищеварения.
При разрушении каждой молекулы глюкозы, сопровождающемся понижением свободной энергии, высвобождается энергия, достаточная для образования 93 молекул АТФ путем присоединения фосфатных групп к молекулам АДФ. Оказывается, что реально образуются не все 93 молекулы. При этом, весь процесс включает большое число ферментативных реакций. Питательные вещества (углеводы, жирные кислоты и аминокислоты) вступают в серию реакций, образующих цикл Кребса (или цикл трикарбоновых кислот), в процессе которого углеродный остов молекул распадается с образованием СО2, но АТФ здесь не образуется. На следующих этапах реакции происходит перенос электронов с помощью специальных ферментов (дыхательная цепь). На этих этапах синтезируется АТФ, а последний шаг на пути длительного процесса переноса электрона заключается в присоединении его к молекулярному кислороду. Процесс переноса электрона по дыхательной цепи в целом, приводящий к накоплению энергии в молекулах АТФ, называется окислительным фосфорилированием. В результате этого процесса образуется 38 молекул АТФ в расчете на каждую потребленную молекулу глюкозы. Эффективность такого преобразования равна 38/93=40%.
Величину теплопродукции, или тепловой мощности, организма можно количественно оценить исходя из следующих простых соображений.
Энергетическая ценность питания человека составляет около 2400 ккал в сутки. В первом приближении, 2400ккал = 104 Дж, 1 сутки (24 часа) = 86400 секунд = 105 секунд.
Тогда энергия, потребляемая организмом человека в секунду, составит 104/105 = 0,1 кДж×с-1 или 100 Дж×с-1 = 100 Вт; следовательно, тепловая мощность человека приблизительно равна мощности электрической лампочки, имеющей мощность 100 Вт.
При мышечном сокращении АТФ, донор энергии для процесса мышечного сокращения, в ходе реакции с миозином позволяет получать максимум 50 Дж×г-1 энергии. Это означает, что идеальной мышечной системе (т.е. с коэффициентом полезного действия, равным 100%) для подъема груза массой 1 кг на высоту 5 м понадобилось бы израсходовать 2×10-3 моль АТФ. На самом деле, коэффициент полезного действия мышц составляет около 30-40%, остальная же часть высвобождается в виде тепла.
В нормальных условиях жизнедеятельности организма глюкоза является основным энергетическим субстратом. Нормальная концентрация глюкозы в плазме крови человека, в зависимости от условий питания, поддерживается в пределах 50-120 мг %. После еды, в течение фазы всасывания, концентрация глюкозы в системе воротной вены может достигать более 270 мг %. Повышение содержания глюкозы в крови всегда вызывает увеличение секреции инсулина.
В организме человека в состоянии покоя натощак скорость обмена глюкозы составляет в среднем 140 мг/ч на 1 кг массы тела, причем примерно 50% глюкозы потребляется головным мозгом, 20% - мышцами, 20% - эритроцитами и почками, 20% - мышцами и только 10% глюкозы остается на другие ткани.
Скорость утилизации (скорость обмена) глюкозы у здорового человека является линейной функцией концентрации глюкозы в плазме крови. Математическая зависимость утилизации глюкозы от ее концентрации в крови у нормальных людей выражается уравнением:
Ru = 0,02554C + 0,0785,
А у больных некетотическим диабетом:
Ru = 0,004448С + 2,006,
где Ru - скорость утилизации глюкозы, мг/ мин на 1 кг массы тела, а С - концентрация глюкозы в плазме крови, мг %.
Термин "утилизация" глюкозы в физиологическом смысле означает скорость переноса глюкозы из крови в общий фонд глюкозы тканей и выхода из него в процессе метаболизма. С биохимической точки зрения скорость утилизации глюкозы определяется транспортом через цитоплазматическую мембрану и внутриклеточным окислительным фосфорилированием глюкозы. Широко распространенные в литературе термины "скорость оборота", "ассимиляция" и "потребление" глюкозы являются синонимами понятия "утилизация" глюкозы и в любом отношении равнозначны.
Практически во всех тканях в физиологических условиях транспорт глюкозы из межклеточной среды во внутрь клетки представляет собой первичную лимитирующую реакцию в утилизации глюкозы клетками, так как в отсутствие инсулина поток переносимой глюкозы всегда меньше скорости фосфорилирования глюкозы. Равновесие между скоростью транспорта и фосфорилированием глюкозы наступает только при больших концентрациях глюкозы (400-500 мг %). При дальнейшем увеличении концентрации глюкозы лимитирующей реакцией становится фосфорилирование. Другими словами, скорость транспорта глюкозы из межклеточной среды через цитоплазматическую мембрану во внутриклеточную среду является процессом, лимитирующим скорость утилизации глюкозы живой тканью.
Исходя из сделанного выше рассмотрения, представляется логичным и вполне обоснованным заключение о том, что теплопродукция, также, как и скорость утилизации глюкозы, является линейной функцией концентрации глюкозы в крови и, измерение величины локальной теплопродукции позволяет определить уровень глюкозы в крови.
Физика конденсированного состояния биополимерной молекулы.
Объектом настоящего исследования, выполненного в рамках работы над группой изобретений, являются биополимерные молекулы, способные к пространственной самоорганизации и обретению устойчивой молекулярной структуры, обусловленной слабыми внутримолекулярными объемными взаимодействиями не валентной природы.
Примерами биополимерных молекул, способных к пространственной самоорганизации при физиологических условиях, являются полисахариды и белковые молекулы с полипептидной цепью.
Поведение белков всецело определяется исключительной, присущей только им пространственной структурной организацией, которая обусловлена не валентными взаимодействиями и определяется исключительно слабыми объемными взаимодействиями. Лишаясь ее, белки теряют все свои биологические свойства.
Белковые цепи способны самопроизвольно свертываться в строго детерминированные структуры, геометрия и конформационная динамика которых в физиологических (нативных) условиях полностью определяются аминокислотной последовательностью. Трехмерные структуры белков индивидуализированы, очень сложны и имеют строгий порядок, не сводящийся, однако, к периодичности. На Рис.1 схематично показан процесс самоорганизации (фолдинга) биополимерной молекулы, который происходит самопроизвольно, как фазовый переход статистической системы многих взаимодействующих частиц в пространственную конфигурацию с гетерофазной структурой с минимальной свободной энергией. Способность природной полипептидной цепи к пространственной самоорганизации и обретению определенной молекулярной структуры – самая яркая особенность белков, отсутствующая у молекул искусственных полимеров, в том числе у полученных человеком поли-а-аминокислот.
Отсюда следует понимание чрезвычайной сложности задачи экспериментального исследования свойств биополимерных молекул в условиях in vitro. Многие экспериментальные методы исследования структуры вещества, эффективные при исследованиях молекул (структура которых определена ковалентными связями), имеют ограниченную применимость для исследования биополимерных молекул, пространственная структура которых определяется слабыми не валентными взаимодействиями.
Современные методы позволяют определить первичную структура биополимерных молекул, состоящую из последовательности мономеров, связанных прочными ковалентными связями, в то время как, определение пространственной структуры таких молекул продолжает оставаться сложнейшей проблемой. Например, число последовательностей (первичная структура) белковых молекул в базе данных UniParc (на август 2016) составляло более 124 миллионов, в то время как количество структур в базе данных PDB (Protein Data Bank) - лишь чуть больше 121 тысячи, что составляет менее 0,1% от всех известных последовательностей, причем разрыв между двумя этими показателями стремительно нарастает и, вероятно, будет расти и дальше. Такое сильное отставание связано с относительной сложностью современных методов определения структуры.
Другая сторона проблемы заключается в недостаточно глубоком понимании фундаментальной природы слабых не валентных внутримолекулярных взаимодействий. К моменту настоящего исследования, человечество со всеми своими вычислительными мощностями и арсеналом экспериментальных данных до сих пор не научилось строить модели, которые бы описывали процесс белкового фолдинга и предсказывать трехмерную структуру белка на основе его первичной структуры (то есть аминокислотной последовательности), что прямо указывает на отсутствие понимания фундаментальной природы слабых объемных взаимодействий, определяющих биологические свойства биополимерных молекул.
Таким образом, пространственная структура биополимерной молекулы и ее связь с биологической функцией является одной из крупнейших нерешенных проблем современной науки, решение которой возможно при условии достижения прогресса в фундаментальных исследованиях физико-химической природы слабых объемных взаимодействий, определяющих трехмерную структуру биополимерной молекулы.
Следует отметить, что решение проблемы фолдинга прямо связано с научной проблемой биологического катализа - уникального явления, физико-химическая природа которой продолжает оставаться неясной.
Отсюда следует понимание вывода, что решение проблемы фолдинга и биологического синтеза напрямую связано с созданием новых физико-химических методов исследования in vivo биополимерных молекул и возможностью достижения реального прогресса в области физики конденсированного состояния биополимерных структур.
В рамках настоящего исследования данная научная проблема была решена за счет сочетания экспериментального исследования, которое было выполнено с помощью уникального экспериментального метода, основанного на неинвазивных измерениях характеристик межклеточного вещества в условиях in vivo, и теоретического исследования конденсированного состояния биополимерной молекулы, которое было выполнено на основе методов статистической физики с целью обобщения полученных экспериментальных данных.
Биосенсор на основе биополимерной молекулы с гетерофазной структурой. Уникальным модельным объектом для исследований в области физики конденсированного состояния биополимерных молекул, способных к самоорганизации пространственной структуры, как оказалось, является биополимерная молекула полисахарида гиалуроновой кислоты, выбор которой в качестве объекта исследования был обусловлен следующими соображениями:
Высокомолекулярный полисахарид гиалуроновой кислоты является основным компонентом межклеточного вещества, окружающего живую клетку, выполняющего важную роль в самоорганизации физико-химических процессов клеточного метаболизма, важнейшим из которых является процесс физической терморегуляции живой ткани, поддерживающей постоянство температуры. Как известно из опыта, все многообразие млекопитающих организмов, основным межклеточным компонентом которых является биополимерная молекула гиалуроновая кислота, структура которой оставалась неизменной в процессе длительной биологической эволюции, имеют близкую температуру тела (36-38°С), что прямо указывает на гетерофазное состояние межклеточного вещества, обусловленное термодинамическими свойствами биополимерной молекулы гиалуроновой кислоты.
Экспериментальное исследование термодинамических свойств биополимера гиалуроновой кислоты можно проводить в условиях in vivo на основе неинвазивных измерений характеристик межклеточного вещества эпидермиса человека с помощью датчиков, расположенных на поверхности эпидермиса кожи.
биополимерная молекула гиалуроновой кислоты обладает простейшей первичной структурой, показанной на Рис. 2, состоящей из одинаковых повторяющихся мономеров (дисахаридов), состоящих из 2 типов сахарных остатков (звеньев), в отличие от более сложной первичной структуры глобулярных белков (состоящих из 20 типов звеньев), ДНК и РНК (состоящих из 4 типов звеньев), поэтому является уникальным модельным объектом биополимерной молекулы для теоретического исследования. С целью исследования термодинамических свойств межклеточного вещества были проведены экспериментальные исследования in vivo, в которых исследовалась зависимость осмотического давления межклеточного вещества эпидермиса живой ткани человека, характеризующего термодинамическое фазовое состояние межклеточного вещества в локальной области под теплонепроницаемым аппликатором (накладываемым с дозированным давлением на поверхность кожи и образующим замкнутую систему живой ткани в локальной области под аппликатором), от внешнего теплового потока, внешнего давления и концентрации глюкозы в крови.
Экспериментальные исследования зависимости осмотического давления межклеточного от мощности внешнего электромагнитного излучения, падающего на поверхность эпидермиса кожи человека, были проведены в широком диапазоне частот от оптического до микроволнового: оптическое и ближнее инфракрасное излучение 0,4-0,9 мкм; инфракрасное излучение 1-10 мкм; микроволновое излучение 5-10 мм.
На Рис. 3а представлена экспериментальная зависимость осмотического давления межклеточного вещества эпидермиса, которое измерялось неинвазивным методом в реальном времени, от мощности внешнего падающего инфракрасного теплового излучения (с максимумом спектральной мощности на длине волны λ ≈ 10 мкм) при постоянном уровне сахара в крови (во временном интервале эксперимента). В одном эксперименте на одном пациенте (с фиксированным положением датчика на поверхности эпидермиса) было выполнено 5 измерений, соответствующих разным значениям падающей мощности (в единицах мВт/см2): 10; 5,5; 3; 2; 0,5. Пороговая чувствительность межклеточного вещества ограничивается собственными флуктуациями осмотического давления межклеточного вещества и составляет <100 мкВт/см2 для времен наблюдения 102с.; полученное пороговое значение чувствительности эквивалентно перепаду температуры окружающей среды на 0,08° С.
Экспериментально было доказано, что процесс теплообмена между локальным участком эпидермиса живой ткани и окружающей средой является обратимым и изотермическим: обратимые изменения мощности падающего теплового излучения в эксперименте приводят к обратимым изотермическим изменениям осмотического давления в реальном времени.
Как известно, термодинамическая обратимость и изотермический характер теплообмена вещества с внешней средой являются фундаментальными свойствами реального вещества, которые проявляются в тройной точке, в которой вещество находится в гетерофазном состоянии. На рисунке 4 показан график зависимости температуры произвольного вещества от количества введенного в систему тепла (heating curve), наблюдаются участки постоянной температуры вблизи фазовых переходов, в которых вещество находится в гетерофазном состоянии, которые обусловлены тем, что в процессе плавления вещества количество теплоты, равное скрытой теплоте плавления (или испарения) ΔН, поглощается системой без изменения температуры, но с изменением энтропии ΔН/Т = ΔS; эффективная теплоемкость вещества, находящегося в гетерофазном состоянии, равна бесконечности.
Примером системы, в которой реализуется термодинамическое равновесие между тремя фазами вещества, является смесь воды, льда и пара в тройной точке. Как известно, в смеси воды и льда при 273,16 K две фазы находятся в равновесии, и процессы плавления и замерзания протекают обратимо при постоянной энтропии: при нагревании смеси происходит плавление льда, а при охлаждении, наоборот, происходит кристаллизация - образование льда из жидкой фазы, при этом температура воды остается постоянной (свойство изотермии).
Таким образом, результаты экспериментов по измерению порога чувствительности межклеточного вещества к тепловым потокам на основе неинвазивного измерения осмотического давления межклеточного вещества прямо доказывают вывод о гетерофазной структуре межклеточного вещества живой ткани, при физиологической температуре тела 36,6°С. Отсюда следует понимание обобщения, что одинаковая температура теплокровных организмов является следствием гетеро фазного состояния межклеточного вещества теплокровных, которое является фундаментальным свойством межклеточного вещества этих организмов, обусловленным термодинамическими свойствами биополимерной молекулы гиалуроновой кислоты.
Селективность молекулы гиалуроновой кислоты к глюкозе. Особенность биополимерной молекулы гиалуроновой кислоты с гетерофазной структурой заключается в том, что наряду с экстремально высокой чувствительностью к тепловым потокам, молекула обладает избирательностью к молекуле глюкозы, обусловленной связыванием образованием водородных связей между молекулой глюкозы растворителя и мономерами полимерной цепи. С целью исследования избирательности биополимерной молекулы гиалуроновой кислоты к молекуле глюкозы были проведены экспериментальные исследования зависимости осмотического давления межклеточного вещества эпидермиса от концентрации глюкозы в крови.
На Рис. 3б представлена зависимость осмотического давления межклеточного вещества от концентрации глюкозы в крови, полученная экспериментально в клиническом исследовании на группе из 50 пациентов. Как следует из представленного графика, обратная величина осмотического давления межклеточного вещества линейно зависит от концентрации сахара в крови в интервале концентраций 3 ммоль/литр - 25 ммоль/литр. Условия эксперимента описаны подробно в разделе «Практическое применение»
Экспериментально доказано свойство термодинамической обратимости переходов межклеточного вещества из одного состояния в другое - обратимые изменения концентрации глюкозы в крови и, как следствие, скорости метаболизма живой ткани, приводят к обратимым изменениям осмотического давления межклеточного вещества:
обратимые физиологические изменения концентрации глюкозы в крови приводят к обратимым изменениям осмотического давления, аналогичным обратимым изменениям обратимым осмотического давления в процессе теплообмена живой ткани с окружающей средой - уменьшение концентрации глюкозы приводит к увеличение осмотического давления, и наооботот, увеличение концентрации глюкозы в крови приводит к уменьшению осмотического давления - отклику параметра, аналогичному отклику на охлаждение вещества.
Свойство термодинамической обратимости переходов вещества из одного состояния в другое является фундаментальным свойством реального вещества, находящегося в гетерофазном состоянии в тройной точке, поэтому свойство термодинамической обратимости межклеточного вещества живой ткани, обнаруженное экспериментально, является прямым доказательством гетерофазного состояния межклеточного вещества при физиологических условиях.
Межклеточное вещество живой ткани, свойства которой определяются термодинамическими свойствами биополимерной молекулы гиалуроновой кислоты с гетерофазной структурой, таким образом, является природным биосенсором глюкозы, состоящим из двух элементов - распознающего элемента (molecular recogniser) и трансдьюсера (transducer), который преобразует изменения концентрации глюкозы в изменения осмотического давления межклеточного вещества. На рисунке 26 ниже показана блок схема, поясняющая принцип работы биосенсора глюкозы межклеточного вещества.
Таким образом, результаты экспериментальных исследований in vivo доказывают уникальные биосенсорные свойства межклеточного вещества живой ткани, обусловленные гетерофазной структурой биополимерной молекулы гиалуроновой кислоты - межклеточное вещество является уникальным природным биосенсором, обладающим свойством термодинамической обратимости и биоселективностью к молекуле глюкозы и экстремально высокой чувствительностью к внешним тепловым потокам.
Теоретическое исследование было проведено с целью выяснения физико-химического механизма избирательности биополимерной молекулы гиалуроновой кислоты к глюкозе и статистического механизма гетерофазного конденсированного состояния объединенной системы биополимерной молекулы с растворителем вблизи границы фазового перехода на основе обобщения экспериментальных результатов. Описание статистического метода и основные результаты исследования приведены ниже.
Свойство термодинамической обратимости и избирательности роднит полисахаридную молекулу гиалуроновой кислоты с белковыми молекулами, в частности, с ферментами, которые обладают уникальной избирательностью к определенным молекулам субстрата и свойством катализировать биохимические реакции с участием этих молекул. Отсюда следует понимание вывода, что свойство биологической специфичности ферментов, доказанное на основе результатов большого числа биохимических исследований, и избирательность гиалуроновой кислоты к глюкозе имеют единую физико-химическую природу.
В рамках теоретического исследования было выполнено обобщение для ферментных молекул, на основе статистического метода, с помощью которого были получены точные решения для термодинамических фаз вещества в объеме макромолекулы. С этой целью было проведено теоретическое исследование физико-химического механизма ферментов, на примере, ферментов гексокиназы, которые обладают избирательностью к молекуле глюкозы и катализируют биохимическую реакцию с участием глюкозы. Полученные результаты доказали гетерофазный статистический механизм ферментативного катализа.
В данном документе под термином «субстрат S» понимается молекула растворителя, взаимодействующая с мономерами полимерной цепи путем образования слабой не валентной связи. В частности, растворитель по изобретению представляет собой водный раствор ионов натрия, хлора, калия и глюкозы, при этом, разница между концентрациями натрия и хлора равно примерно 40 ммоль/литр (избыток ионов натрия), а содержание глюкозы в интервале гетерофазности может изменяться в широком диапазоне от 3 до 15 ммоль/литр.
В данном документе под термином «мономер» понимается повторяющиеся звенья (структурные единицы) в составе биополимерной молекулы. Примером мономера А является D-глюкуроновая кислота, содержащая отрицательно заряженную группу, примером мономера В является D-Nацетилглюкозамина, нейтрально заряженная, образующие повторяющийся дисахарид полимерной цепи гиалуроновой кислоты. В случае полимерной цепи белковой молекулы, примерами мономеров А являются отрицательно заряженные глутаминовая кислота и аспарагиновая кислота, и положительно заряженные лизин, аргинин, гистидин, примерами мономеров В являются не заряженные серии, треонин, глутамин, аспарагин.
В данном документе под термином - "макромолекула с гетерофазной структурой" понимается… одна - единственная биополимерная молекула в объеме которой при определенных условиях могут сосуществовать несколько термодинамических фаз - кристаллическая фаза (глобула), жидкая (расплавленная глобула), газообразная (клубок).
В данном документе под термином макромолекула с «заданные биологические свойства и заданная пространственная структура» понимается биополимерная молекула, обладающая однозначно определенными биохимическими характеристиками (константа равновесия, скорость биохимической реакции) и однозначно определенной трехмерной пространственной структурой.
В данном документе под термином «растворитель» понимается… вещество, способное растворять другие твердые, жидкие или газообразные вещества, не изменяя их химически. Как правило, при атмосферном давлении и комнатной температуре растворитель является жидким веществом. Примерами являются вода, морская вода, раствор глюкозы.
В данном документе под термином «субъединица» понимается биополимерная (полипептидная) цепь, которая вместе с другими компонентами собирается в мультимерный или олигомерный белковый комплекс.Многие природные ферменты и другие белки состоят из нескольких белковых субъединиц.
В данном документе под термином «биологический катализатор с заданными характеристиками» понимается катализатор с определенными значениями константы скорости и скорости биохимической реакции, которую катализирует.
В данном документе под термином «фермент с заданными характеристиками» понимается белковая макромолекула с однозначно определенной трехмерной пространственной структурой и определенными значениями константы скорости и скорости биохимической реакции, которую катализирует данный фермент.
В данном документе под термином «биосенсор с селективностью к заданной молекуле субстрата» понимается биополимерная молекула, обладающая избирательностью (специфичностью) к определенной молекуле растворителя.
Неравновесная термодинамика конденсированного состояния биополимерной молекулы.
Изотермический микрокалориметр живой ткани.
Как было показано, свойство изотермии теплокровных - относительное постоянство температуры, при обратимых физиологических изменениях концентрации глюкозы в крови (определяющей скорость клеточного метаболизма и интенсивность теплопродукции) и теплового баланса живой ткани с окружающей средой в области физической терморегуляции организма, имеет физическую природу и обусловлено термодинамическими свойствами межклеточного вещества локального микрообъема живой ткани. Межклеточное вещество, в области физиологической нормы, находится в гетерофазном конденсированном состоянии, в котором кристаллическая твердая фаза (глобула) находится в термодинамическом равновесии с метастабильной жидкой фазой (расплавленная глобула) и газообразной фазой (клубок). Теплота, введенная в локальный объем вещества, в результате воздействия на локальный участок поверхности эпидермиса внешнего теплового потока Wпогл, приводит к плавлению твердой фазы вещества - фазовому преобразованию вещества из твердого состояния "глобула" в жидкое состояние "расплавленная глобула"; процесс плавления глобулярной фазы гиалуроновой кислоты протекает, как фазовый переход первого рода с изменением объема вещества (изменившего агрегатное состояние), обусловленным изменением осмотического давления вещества. Изменение осмотического давления межклеточного вещества (количества воды в межклеточном пространстве), в результате плавления глобулярной фазы, приводит к изменению интенсивности межклеточной микроциркуляции - микро гидродинамики (микрофлюидики), направленной к поверхности эпидермиса, обеспечивающей изотермический теплообмен ткани с окружающей средой в процессе испарительного охлаждения с интенсивностью Wисп; тепловой баланс обратимого изотермического процесса, при постоянной энтропии, имеет вид:
ΔWпогл = ΔWисп.
Физическим аналогом системы, в которой реализуется термодинамическое равновесие между тремя фазами вещества, является смесь воды, льда и пара в тройной точке. Как известно [12], в смеси воды и льда при 273,16 K две фазы находятся в равновесии, и процессы плавления и замерзания протекают обратимо при постоянной энтропии: при нагревании смеси происходит плавление льда, а при охлаждении, наоборот, происходит кристаллизация - образование льда из жидкой фазы, при этом температура воды остается постоянной (свойство изотермии).
Процесс плавления вещества, при температуре фазового перехода, происходит изотермически, без изменения температуры; количество теплоты, введенной в систему, можно определить измеряя количество вещества, перешедшего в процессе плавления в жидкое состояние (изотермическая калориметрия - измерение количества теплоты, введенной в объем вещества, при температуре фазового перехода). Принцип изотермической калориметрии на практике используется в известном устройстве - изотермическом калориметре фазового перехода, в котором введенная теплота не изменяет температуры калориметрической системы, а вызывает изменение агрегатного состояния вещества, составляющего часть этой системы (например, таяние льда в ледяном калориметре Бунзена). В процессе плавления вещества количество теплоты, равное скрытой теплоте плавления ΔН, поглощается системой без изменения температуры, но с изменением энтропии ΔН/Т = ΔS. Количество введенной теплоты рассчитывается в этом случае по массе вещества, изменившего агрегатное состояние (например, массе растаявшего льда, которую можно измерить по изменению объема смеси льда и воды), и теплоте фазового перехода. Принцип действия ледяного калориметра заключается в том, что количество введенной в калориметр теплоты измеряется не по изменению температуры, как в калориметрах переменной температуры, а по изменению объема смеси вода-лед, находящейся при температуре плавления льда. Схема изотермического ледяного калориметра показана на Рис. 24. Количество введенной теплоты рассчитывается по массе растаявшего льда, которую можно измерить по изменению объема смеси льда и воды и теплоте фазового перехода.
Принцип действия изотермического калориметра живой ткани аналогичен принципу ледяного калориметра и заключается в том, что количество теплоты, введенной в локальной объем живой ткани или образовавшейся в этом объеме в процессе клеточного метаболизма, измеряется не по изменению температуры живой ткани, как в калориметрах переменной температуры, а по изменению объема межклеточного вещества, определяемого объемом смеси глобула (кристаллическая фаза) - расплавленная глобула (жидкая фаза), находящейся при температуре плавления глобулы гиалуроновой кислоты. Изменения объема межклеточного вещества можно определить путем измерения осмотического давления межклеточного вещества. Схема, поясняющая принцип изотермического микрокалориметра живой ткани, представлена на Рис. 25. Изменение объема межклеточного вещества в процессе плавления кристаллической (глобулярной) фазы гиалуроновой кислоты вещества, вызванного поглощением тепла, определяется методом измерения осмотического давления межклеточного вещества.
Биополимер гиалуроновой кислоты межклеточного вещества живой ткани является природным изотермическим биосенсором с высокой чувствительностью к тепловой энергии, обусловленной маленькой величиной энергии связи ≈ 3,5 kВТ молекулярного комплекса тетрамера: тепловая энергия (количество теплоты), введенная в объем вещества, приводит к изотермическому плавлению кристаллической фазы вещества, протекающему с поглощением энергиин, и, как следствие, к изменению осмотического давления межклеточного вещества, которое можно измерить с помощью датчика, расположенного на поверхности эпидермиса кожи. Как показано выше, конформационные переход Na-тетрамера из конформации (s) кристаллической фазы в конформацию (λ) жидкой фазы может протекать только с поглощением кванта энергии 1kВТ в расчете на один тетрамер. Количество теплоты, которое необходимо сообщить одной единице массы кристаллического вещества в равновесном изобарно-изотермическом процессе, чтобы перевести его из твердого (кристаллического) состояния в жидкое (то же количество теплоты выделяется при кристаллизации вещества) определяется как удельная теплота плавления (энтальпия плавления).
Были проведены экспериментальные исследования зависимости осмотического давления межклеточного от мощности внешнего электромагнитного излучения, падающего на поверхность эпидермиса кожи человека, в широком диапазоне частот от оптического до микроволнового: оптическое и ближнее инфракрасное излучение 0,4-0,9 мкм; инфракрасное излучение 1-10 мкм; микроволновое излучение 5-10 мм. На Рис. 3а показана экспериментальная зависимость осмотического давления межклеточного вещества от мощности падающего на поверхность эпидермиса инфракрасного теплового излучения при постоянной концентрации сахара в крови (в интервале эксперимента 30-60 минут уровень сахара практически не изменяется). Принцип действия изотермического калориметра живой ткани заключается в том, что количество введенной в калориметр теплоты измеряется не по изменению температуры, как в калориметрах переменной температуры, а по изменению объема межклеточного вещества, состоящего из смеси кристаллической (глобула) и жидкой (расплавленная глобула) фаз, находящегося при температуре плавления кристаллической глобулярной фазы.
На Рис. 3а представлена экспериментальная зависимость осмотического давления межклеточного вещества эпидермиса от мощности внешнего падающего инфракрасного теплового излучения (с максимумом спектральной мощности на длине волны λ ≈ 10 мкм) при постоянном уровне сахара в крови (во временном интервале эксперимента). В одном эксперименте на одном пациенте (с фиксированным положением датчика на поверхности эпидермиса) было выполнено 5 измерений, соответствующих разным значениям падающей мощности (в единицах мВт/см2): 10; 5,5; 3; 2; 0,5. Пороговая чувствительность калориметрического спектрометра ограничивается собственными флуктуациями осмотического давления межклеточного вещества и составляет < 100 мкВт/см2 для времен наблюдения 102с.
Скорость потока межклеточной жидкости через эпидермис (скорость массопереноса) определяется градиентом осмотического давления межклеточного вещества. Угол наклона графика на Рис. 3а соответствует коэффициенту массопереноса эпидермиса, значение которого определяется концентрацией глюкозы в крови, увеличение которой приводит к увеличению доли кристаллической фазы вещества в межклеточном пространстве. Концентрацию глюкозы в крови можно определить по углу наклона графика зависимости осмотического давления межклеточного вещества эпидермиса от мощности внешнего падающего теплового излучения; угол наклона графика определяется концентрацией глюкозы в межклеточной жидкости (в крови).
Селективность межклеточного вещества к глюкозе. Особенность изотермического микрокалориметра живой ткани заключается в том, что межклеточное вещество живой ткани, наряду с экстремально высокой чувствительностью к тепловому потоку, обладает селективностью к молекуле глюкозы, концентрация которой определяет концентрацию тетрамеров (сшивок) полимерной цепи гиалуроновой кислоты и, как следствие, определяет долевое соотношение между кристаллической (глобула) и жидкой (расплавленная глобула) термодинамическими фазами вещества. Увеличение концентрации глюкозы в жидкой фазе межклеточного вещества приводит к процессу кристаллизации вещества, которая сопровождается уменьшением объема и осмотического давления; уменьшение концентрации глюкозы приводит к обратному процессу - растворению, которое сопровождается увеличением объема и осмотического давления. Отсюда следует понимание вывода, что увеличение концентрации глюкозы в веществе приводит к отклику вещества, аналогичному отклику на охлаждение.
Межклеточное вещество живой ткани является биосенсором глюкозы, который состоит из двух элементов - распознающего элемента (molecular recogniser) (тетрамер) и трансдьюсера (transducer), который преобразует изменения концентрации глюкозы в изменения осмотического давления межклеточного вещества. На рисунке 26 показана блок схема, поясняющая принцип работы биосенсора глюкозы межклеточного вещества.
Вещество живой ткани представляет природный биосенсор глюкозы, который состоит из двух элементов: распознающего элемента (molecular recogniser), роль которого выполняет кристаллическая фаза макромолекулы, и трансдьюсера (transducer), роль которого выполняет жидкая фаза, который преобразует изменения концентрации глюкозы в изменения осмотического давления межклеточного вещества, которые регистрируются с помощью датчика, расположенного на поверхности эпидермиса.
Калорическое уравнение. Калорическое уравнение состояния, связывающее переменные термодинамического состояния живой ткани в локальном объеме под тепло и водонепроницаемым аппликатором в изотермическом калориметре, определяется из основного уравнения термодинамики, следующим путем.
Основное уравнение термодинамики, описывающее обратимые квазистатические изменения состояния замкнутой системы под аппликатором во временном интервале Δt = τ, равном постоянной времени переходного процесса, можно преобразовать и представить в виде уравнения, описывающего обратимые изменения системы во временном интервале Δt = dt → 0, в котором потоки можно считать постоянными:
В уравнении (44а), связывающем потоки сопряженных процессов переноса энергии и вещества, которые измеряются в реальном эксперименте, приняты следующие обозначения:
WМЕТ - мощность процесса теплопродукции, которая определяется из требования
 при интегрировании в интервале от 0 до τ.
при интегрировании в интервале от 0 до τ.
WΔТ - мощность процесса теплопереноса, поступающего в систему из глубинного слоя, обусловленный разностью температур ΔTskin = Тглуб. - Тпов., которая определяется из требования 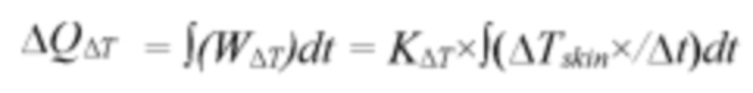 при интегрировании в интервале от 0 до τ.
при интегрировании в интервале от 0 до τ.
KM×(ΔМ/Δt) = J - мощность процесса массопереноса, поступающего в систему из глубинного слоя, обусловленного разностью количества вещества ΔM = Мглуб. - Мпов., которая определяется из требования 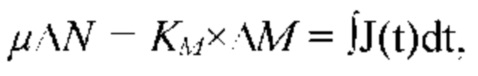 при интегрировании в интервале от 0 до τ, где μ - химический потенциал, N - количество вещества, J(t) - поток вещества.
при интегрировании в интервале от 0 до τ, где μ - химический потенциал, N - количество вещества, J(t) - поток вещества.
Kр×Δр = Wкров. - мощность процесса теплопереноса, который осуществляется капиллярным кровотоком, обусловленным разностью гидравлического давления в системе микроциркуляции Δр = рартер. - рвенул., которая определяется из требования 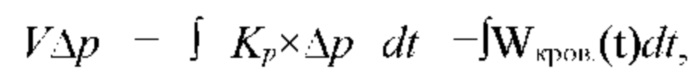 при интегрировании в интервале от 0 до τ.
при интегрировании в интервале от 0 до τ.
Компонента теплопереноса в объеме ткани под аппликатором, обусловленная разностью кровяного давления, в зависимости от знака потока массопереноса, может иметь положительное значение (тепло вводится в систему), отрицательное значение (тепло выводится из системы) или равно нулю. Константа этого процесса определяется выражением Kр = V - V0 > 0, где V0 - объем межклеточного вещества, при нулевом потоке J=0. Объем вещества V > V0, при положительном потоке J > 0; объем вещества V < V0, при отрицательном потоке J < 0.
KΔT, KM, Kр - константы, феноменологические коэффициенты, которые определяются путем калибровки.
Уравнение состояния, связывающее мощности сопряженных процессов переноса энергии и вещества под аппликатором, которые измеряются в реальном эксперименте, вытекает из уравнения (44а) путем деления всех членов на dt и имеет следующий вид:
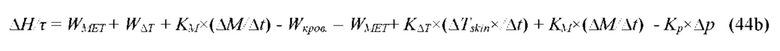
Следует отметить, что основное уравнение термодинамики является интегральной формой уравнения (44b), связывающего потоки процессов переноса энергии и массы. Интегрирование уравнения (44а) в интервале времени от t0 = 0, соответствующего наложению аппликатора с дозированным давлением на поверхность рогового слоя эпидермиса до t = τ приводит к исходному виду основного уравнения термодинамики (3), описывающего обратимые изменения системы в интервале Δt = τ, равном постоянной времени переходного процесса:
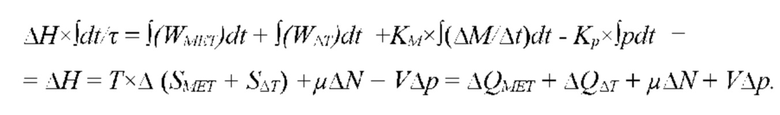
Уравнение (4а) можно записать в виде выражения для расчета величины теплового эффекта метаболизма:
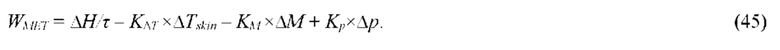
Последнее уравнение (5) можно представить в следующем виде:
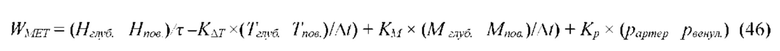
В уравнениях выше приняты следующие обозначения:
Нглуб. - энтальпия глубинного слоя,
Нпов. - энтальпия поверхностного слоя,
Тглуб. - температура глубинного слоя,
Тпов. - температура поверхностного слоя,
Мглуб. - количество воды в глубинном слое,
Мпов. - количество воды в поверхностном слое,
Рартер. - гидростатическое давление на входе капилляра (в системе микроциркуляции),
Рвенул. - гидростатическое давление на выходе капилляра (в системе микроциркуляции),
Параметры KΔT, KM, KP соответствуют коэффициентам пропорциональности и равны константам, которые определяются методом калибровки.
Величина энтальпии определяется количеством воды в единичном объеме, которое при заданных значениях климатических параметров, зависит от содержания гиалуроновой кислоты в межклеточном веществе, которая может изменяться при сахарном диабете и других хронических заболеваниях, поэтому является индивидуальной характеристикой пациента, которую можно определить методом калибровки. Количество воды в единичном объеме ткани, с другой стороны, при заданном содержании гиалуроновой кислоты, определяется климатическими параметрами.
Метод изотермической калориметрической спектроскопии живой ткани.
На основе метода изотермической микрокалориметрии (измерение величины поглощенной живой тканью мощности электромагнитного излучения с помощью изотермического микрокалориметра фазового перехода), описанного выше, может быть создан принципиально новый метод спектроскопии живой ткани в условиях in vivo - абсорбционная изотермическая калориметрическая спектроскопия, позволяющая определять в реальном времени содержание биохимических компонентов живой ткани на основе спектральных измерений на характеристических частотах исследуемых молекул.
Величина мощности электромагнитного излучения на характеристической частоте исследуемого биохимического компонента, поглощенной в локальном объеме вещества живой ткани, измеряется не по изменению температуры ткани, а по изменению величины осмотического давления (или количества воды в межклеточном пространстве), характеризующего объем межклеточного вещества, находящегося при температуре фазового перехода (соответствующей термодинамическому равновесию кристаллической и жидкой фаз межклеточного вещества).
Характеристические частоты биохимических компонентов вещества живой ткани, к которым относятся компоненты межклеточного вещества (гиалуроновая кислота, вода, глюкоза и др.), а также внутриклеточные биохимические молекулы и органоиды, образуют электромагнитный спектр поглощения вещества живой ткани, который в условиях in vivo можно определить методом изотермической калориметрической спектроскопии (ICS - isothermal calorimetric spectroscopy).
Метод изотермической калориметрической спектроскопии живой ткани базируется на определении величины поглощенной мощности электромагнитного излучения, падающей на поверхность исследуемого локального участка эпидермиса живой ткани, по количеству межклеточного вещества, изменившего фазовое состояние (плавление глобулярной фазы гиалуроновой кислоты), методом измерения в реальном времени количества воды или осмотического давления межклеточного вещества. Схема спектрального устройства, основанного на принципе изотермической калориметрической спектроскопии, показана на Рис 27.
Источник электромагнитного излучения с длиной волны, соответствующей характеристической частоте биохимического компонента живой ткани, интегрирован в изотермический микрокалориметр. Введены следующие обозначения: 1 - аппликатор (тепло и водонепроницаемый), выполненный с возможностью наложения на кожу с дозированным давлением, образующий замкнутую термодинамическую систему живой ткани; 2 - источник электромагнитного излучения с перестраиваемой длиной волны; 3 - датчик осмотического давления межклеточного вещества или микро-потока межклеточной гидродинамики.
Зависимость отклика осмотического давления межклеточного вещества от длины волны падающего излучения (при фиксированной мощности) прямо коррелирует со спектром поглощения вещества; отклик (изменения) осмотического давления на мощность падающего излучения возникает пропорционально величине поглощенной мощности, и имеет большую величину на тех длинах волн, энергия которых соответствует энергиям возбуждения колебаний в изучаемых молекулах.
В некоторых вариантах осуществления изобретения излучение модулировано по амплитуде. В спектральной схеме с применением электромагнитного излучения с амплитудной модуляцией в объеме ткани под аппликатором возбуждаются колебания эластического давления \межклеточного вещества, обусловленные колебаниями осмотического давления на частоте модуляции излучения. В этом варианте, величину поглощенной мощности электромагнитного излучения можно определить способом измерения эластического давления ткани под аппликатором на частоте модуляции излучения. Ближайшим аналогом описанного метода является известный метод оптико-акустической (OA) спектроскопии, основанный на эффекте, проявляющемся в виде пульсаций давления газа в замкнутом объеме при поглощении модулированного на звуковой частоте излучения. Возникновение этого эффекта связано с преобразованием части энергии возбужденных излучением молекул в тепловую энергию среды за счет без излучательных переходов. Оптико-акустический эффект возникает за счет преобразования части поглощенной энергии в тепловую, что приводит к образованию акустических колебаний в самом образце либо соприкасающемся с ним газе. Регистрация акустических колебаний непосредственно в веществе осуществляется пьезоэлектрическим датчиком (жидкие и твердые образцы) или микрофоном (газы).
При спектральном измерении многокомпонентной системы в биохимии используют принцип аддитивности, согласно которому поглощение отдельного вещества не зависит от других веществ, обладающих собственным поглощением. Поэтому, при данной длине волны спектральная плотность раствора из смеси компонентов, не взаимодействующих между собой, равна сумме спектральных плотностей отдельных компонентов при той же длине волны. Для оценки степени поглощения исследуемого раствора, содержащего какое-либо соединение, проводится сравнение интенсивности потока излучения, прошедшего через этот раствор, с интенсивностью излучения, прошедшего через раствор сравнения - бланк.
В случае спектроскопии живой ткани для оценки пика поглощения исследуемого соединения, проводится сравнение спектрального сигнала (интенсивности излучения) на длине волны, соответствующей характеристической частоте исследуемого соединения, со спектральным сигналом, обусловленным поглощением чистой воды на данной длине волны, интенсивность которого можно оценить на основе спектрального измерения на другой частоте, соответствующей пику поглощения воды, который не совпадает с пиками поглощения исследуемого вещества.
Возможность практической реализации метода изотермической калориметрической спектроскопии биохимических компонентов живой ткани была подтверждена нами в экспериментах, в которых измерялась зависимость отклика осмотического давления межклеточного вещества эпидермиса от длины волны и мощности падающего электромагнитного излучения; исследовалась зависимость осмотического давления на электромагнитное излучение в оптическом и ближнем инфракрасном (0,4-0,9 мкм), инфракрасном (1 мкм - 10 мкм) и микроволновом диапазонах (5 мм - 10 мм). Зависимость осмотического давления от мощности инфракрасного излучения (с максимумом спектральной мощности на длине волны λ≈10 мкм) показана на Рис. 3.
Метод терагерцовои (THz) калориметрической спектроскопии.
Терагерцовая (THz) спектроскопия живой ткани основана на измерении величины поглощенной мощности терагерцового излучения, падающего на поверхность исследуемого локального участка эпидермиса живой ткани, по количеству межклеточного вещества, изменившего агрегатное состояние, методом измерения в реальном времени количества воды или осмотического давления межклеточного вещества.
В отношении биологических систем терагерцовое излучение дает возможность изучать роль воды в биологических процессах, а также характерные колебательные процессы, которые могут дать характеристику состояния биосистемы. Важным преимуществом терагерцовой спектроскопии с точки зрения медицины является то, что этот метод является бесконтактным и неинвазивным -излучение этого спектрального диапазона не вызывает ионизации (как ультрафиолет) и значительного нагрева объекта (как СВЧ). Обладая высокой чувствительностью к фазовому состоянию воды и вещества живой ткани, терагерцовый диапазон позволяет выяснить уровень связанности молекул воды в биологических системах, уровень сольватации ионов в растворе и уровень гидратации поверхностей на фазовой границе. Как известно, наиболее эффективно спектральные измерения в терагерцовой области проводятся с образцами, представляющими собой плоско - параллельные слои различной толщины (от нескольких миллиметров до десятков микрометров или сотен ангстрем).
На Рис. 12 схематически показана диаграмма энергетических уровней (λ, g, s) тетрамера гиалуроновой кислоты межклеточного вещества, позволяющая рассчитать энергию кванта терагерцового излучения. Энергия перехода между уровнями тетрамера, равная энергии связи ΔЕ≈7kBТ/2=3,5kBТ≈0,115 эВ молекулярного комплекса тетрамера, определяет характеристическую частоту тетрамера, которой соответствует длина волны электромагнитного излучения λ≈152 мкм (65,78 см-1) в терагерцовой области спектра.
Энергия связи тетрамера определяется высотой энергетического барьера между (λ, s) конформациями Na- тетрамера: ΔЕ=Emax-Emin ≈3,5 kBТ, где Emin минимальное значение энергии справа от энергетического барьера.
Длина волны электромагнитного излучения, соответствующая энергии перехода тетрамера в возбужденное состояние ΔЕ=Еmах-Emin ≈3,5 kBТ, определяется только физиологической температурой:
λ=c×h/ΔE=c×h/3,5kBT=532 мкм/3,5≈152 мкм (65,78 см-1),
где с - скорость света, h - постоянная Планка.
Полученное значение длины волны, соответствует терагерцовому (ТГц) диапазону электромагнитного спектра, который расположен в интервале длин волн между 100 мкм (дальнее ПК) и 1 мм (микроволновый).
Воздействие электромагнитного излучения, с длиной волны λ≈152 мкм, соответствующей энергии возбуждения тетрамера 3,5 kBТ, приводит к разрыву водородных связей тетрамера и конформационному переходу Na - тетрамера из конформации (s) кристаллической фазы в конформацию (λ) жидкой фазы, который сопровождается поглощением кванта тепловой энергии, величину которого можно определить с помощью Рис. 16 по высоте энергетического барьера между (λ, s) конформациями Na- тетрамер
ΔЕ=(Emax-Emin)-(Еmах-E0)=3,5 kBТ-2,5 kBТ=1kBТ,
где Е0 - минимальное значение энергии слева от энергетического барьера.
Чувствительность живой ткани к тепловым потокам.
Особенность природного биосенсора живой ткани заключается в том, что тетрамер гиалуроновой кислоты (распознающий элемент, molecular recogniser) обладает высокой чувствительностью не только к электромагнитным квантам hv на характеристической частоте тетрамера (λ≈152 мкм), соответствующей энергии диссоциации тетрамера ΔЕ=3,5 kBТ, но также обладает экстремально высокой чувствительностью к квантам тепловой энергии, которые образуются в веществе в результате поглощения электромагнитного излучения на частотах, которые не совпадают с характеристической частотой тетрамера (λ≈152 мкм). Например, кванты тепловой энергии образуются в межклеточном веществе в результате поглощения электромагнитного излучения на линиях поглощения воды.
Как известно, пространственное распределение биохимических компонентов в микрообъеме живой ткани неоднородно: гиалуроновая кислота, которая является основным компонентом межклеточного вещества, не входит в состав внутриклеточного вещества; концентрация глюкозы в межклеточном пространстве выше, чем ее концентрация внутри клетки, в которой протеакает реакция окисления глюкозы; суммарное количество воды в межклеточном пространстве единичного объема живой ткани намного меньше суммарного количества внутриклеточной воды, поскольку объем клетки много больше объема межклеточного пространства. Отсюда следует понимание вывода, что мощность электромагнитного излучения с длиной волны, соответствующей характеристическим частотам тетрамера гиалуроновой кислоты и других фрагментов этой полимерной макромолекулы, а также частотам молекулы глюкозы, поглощается в основном в объеме межклеточного вещества. С другой стороны, электромагнитное излучение с длиной волны, соответствующей характеристическим частотам воды, в основном поглощается в объеме клетки. Энергия излучения, которая поглощается внутриклеточной водой (и/или другими внутриклеточными молекулами) в изотермическом процессе преобразуется в энергию теплового движения молекул в объеме клетки и межклеточного пространства, окружающего клетку; взаимодействие тетрамера межклеточного вещества с квантом тепловой энергии (тепловое соударение молекулы вещества с тетрамером), который образовался в объеме клетки, приводит изотермическому изменению осмотического давления межклеточного вещества, в результате диссоциации тетрамера. Тепловая энергия, возникающая в объеме межклеточного вещества в результате поглощения энергии внешнего электромагнитного излучения, приводит к изотермическому плавлению кристаллической фазы межклеточного вещества.
Таким образом, живая клетка, окруженная межклеточным веществом, при физиологических условиях, является изотермическим калориметрическим детектором мощности, который преобразует электромагнитную энергию, поглощенную в объеме клетки (или возникающую в клетке в процессе метаболизма), в осмотическое давление межклеточного вещества.
На Рис. 29 показана схема изотермической калориметрической спектроскопии биохимических компонентов живой клетки. Живая биологическая клетка, окруженная межклеточным веществом, является природным изотермическим микрокалориметром мощности электромагнитного излучения, позволяющим определить спектр поглощения вещества живой ткани на основе измерения зависимости осмотического давления межклеточного вещества от спектральной мощности падающего излучения (1, 2, 3, 4, 5): 1-электромагнитное излучение на характеристической частоте гиалуроновой кислоты; 2- электромагнитное излучение на характеристической частоте глюкозы; 3, 4 - электромагнитное излучение на характеристической частоте воды; 5 - электромагнитное излучение на характеристической частоте внутриклеточных биохимических молекул или органелл.
Электромагнитное излучение с длиной волны, соответствующей характеристической частоте воды, поглощается в объеме клетки: энергия такого излучения преобразуется в энергию теплового движения молекул воды и свободных звеньев полимерной цепи, которая приводит к диссоциации тетрамера гиалуроновой кислоты и изотермическому изменению осмотического давления межклеточного вещества. Живая клетка, окруженная межклеточным веществом, является изотермическим калориметром мощности, который преобразует электромагнитную энергию, поглощенную в объеме клетки (или возникающую в клетке в процессе метаболизма), в осмотическое давление межклеточного вещества.
Таким образом, в спектрах поглощения вещества живой ткани (межклеточного и внутриклеточного) наряду с характеристической частотой тетрамера гиалуроновой кислоты в терагерцовой области (λ≈152 мкм), будут проявляются другие частоты, соответствующие различным группам гиалуроновой кислоты, воды, глюкозы и других биохимических молекул. Схема спектрального измерения, по этой причине, должна быть разработана с учетом взаимного влияния на результаты спектральных измерений поглощения мощности близко расположенными линиями поглощения, в частности, терагерцовое излучение вблизи области пика поглощения тетрамера, также будет поглощаться молекулами воды, основная часть которой в живой ткани находится в объеме клетки. По этой причине, отклик осмотического давления межклеточного вещества локального участка живой ткани на терагерцовое излучение с длиной волны λ≈152 мкм будет состоять из двух вкладов: спектрального отклика тетрамера межклеточного вещества и спектрального отклика на поглощение этого излучения молекулами воды. Спектральный вклад, обусловленный поглощением воды, превышает спектральный вклад, обусловленный поглощением электромагнитной энергии тетромером, поскольку, в единичном объеме живой ткани, объем внутриклеточного вещества значительно превышает объем межклеточного вещества. Спектр воды в терагерцовой области интенсивного пика поглощения вблизи характеристической частоты тетрамера, поэтому применение дифференциального метода двухлучевой спектроскопии позволяет выделить сигнал осмотического давления, обусловленный поглощением излучения тетрамером межклеточного вещества, за счет вычитания из общего сигнала вклада, обусловленного поглощением внутриклеточной воды. Для выделения спектрального сигнала тетрамера, необходимо с помошью описанного метода определить интенсивность пика поглощения тетрамера; для повышения точности можно использовать трехлучевую (трехволновую) схему спектроскопии, в которой длины волн соответствуют локальным экстремума пика поглощения (λ≈152 мкм) и двум боковым частотам, соответствующим минимумам пика. При этом вклад в сигнал, обусловленный поглощением внутриклеточной воды, определяют на длине волны, соответсвующей боковым частотам пика поглощения тетрамера гиалуроновой кислоты. Из результирующего спектрального распределения сигнала, измеренного трехлучевым способом, имеющего форму пика, вычитают постоянный сигнал, соответствующий вкладу поглощения молекул воды. В описанной схеме спектрального измерения предполагается, что спектр гиалуроновой кислоты не содержит вблизи линии поглощения тетрамера λ≈152 мкм других пиков поглощения, обусловленных другими фрагментами гиалуроновой кислоты.
THz спектроскопия для определения концентрации глюкозы в крови.
Метод изотермической калориметрической спектроскопии, можно использовать на практике для определения в условиях in vivo содержания биохимических компонентов (межклеточного и внутриклеточного вещества) живой ткани. В частности, метод позволяет определить концентрацию глюкозы в крови на основе измерения концентрации тетрамеров гиалуроновой кислоты межклеточного вещества, пропорциональной концентрации глюкозы, связанной гиалуроновой кислотой. Как было показано, 1/3 от общего числа молекул глюкозы в объеме макромолекулы гиалуроновой кислоты, находится в свободном состоянии, а оставшиеся 2/3 молекул глюкозы связаны с мономерами полимерной цепи гиалуроновой кислоты, из которых 1/3 молекул глюкозы связана непосредственно в молекулярном комплексе тетрамера. Отсюда следует, что концентрацию свободной (не связанной) глюкозы в межклеточной жидкости (и в крови) можно определить путем измерения концентрации глюкозы, связанной в тетрамерах межклеточного вещества (или концентрации глюкозы, связанной с полимерной цепью гиалуроновой кислоты).
Изотермическая THz калориметрическая спектроскопия воды в живой ткани.
Как известно, при спектральном измерении многокомпонентной системы в биохимии используют принцип аддитивности, согласно которому поглощение отдельного вещества не зависит от других веществ, обладающих собственным поглощением. Поэтому, при данной длине волны спектральная плотность раствора из смеси компонентов, не взаимодействующих между собой, равна сумме спектральных плотностей отдельных компонентов при той же длине волны. Для оценки степени поглощения исследуемого раствора, содержащего какое-либо соединение, проводится сравнение интенсивности потока излучения, прошедшего через этот раствор, с интенсивностью излучения, прошедшего через раствор сравнения - бланк.
В случае спектроскопии живой ткани для оценки пика поглощения исследуемого биохимического соединения, проводится сравнение спектрального сигнала (интенсивности излучения) на длине волны, соответствующей характеристической частоте исследуемого соединения, со спектральным сигналом, обусловленным поглощением чистой воды на данной длине волны.
Разработка метода измерения в реальном времени содержания воды в живой ткани, с другой стороны, является актуальной задачей, имеющей прикладное и фундаментальное значение. Метод изотермической калориметрической спектроскопии позволяет в условиях in vivo определять содержание воды (в межклеточном пространстве и объеме клетки) локального участка живой ткани путем измерения электромагнитного спектра поглощения на длинах волн, соответствующих характеристическим частотам воды, которые в настоящее время изучены достаточно подробно для чистой воды. Спектр поглощения воды в широком диапазоне частот (от ультрафиолетового до микроволнового) показан на Рис. 31.
Энергия падающего электромагнитного излучения, длина волны которой соответствует характеристическим частотам воды, поглощается в объеме клетки и преобразуется в энергию теплового движения молекул воды (принцип изотермического калориметра), которое приводит к изотермическому изменению осмотического давления межклеточного вещества в результате плавления кристаллической фазы межклеточного вещества.
Рассмотрим для примера метод спектрального измерения вблизи линии поглощения воды на характеристической частоте 3339 см-1 (2,99 мкм) в интервале 3250 см-1 - 3750 см-1, показанный на Рис. 31.
Распределение спектральной мощности пика поглощения можно определить с помощью схемы трехлучевой (трехволновой) спектроскопии, в которой длины волн соответствуют локальным экстремума пика (длина волны, соответствующая максимуму пика 3339 см-1 (2,99 нм) и две боковые частоты 3250-1 и 2750 см-1, соответствующие минимумам пика. Количество теплоты, поглощенной на заданной длине волны в частотном интервале 2750 см-1 - 3250-1 см-1 определяется по величине изменения осмотического давления межклеточного вещества.
В случае спектральных измерений в терагерагерцовой области (λ≈152 мкм), рассмотренном выше, коэффициент поглощения воды можно принять постоянным, поскольку в спектре поглощения воды вблизи характеристической частоты (λ≈152 мкм) тетрамера межклеточного вещества, как следует из Рис. 31, не наблюдаются локальных пиков поглощения.
Спектральный способ определения концентрации глюкозы по линиям поглощения воды.
Осмотическое давление и количество воды в межклеточном веществе, как показано, определяется концентрацией глюкозы и величиной внешнего теплового потока, падающего на поверхность эпидермиса, определяемой климатическими параметрами. Метод изотермической калориметрической спектроскопии, позволяющий измерить количество воды в локальном микрообъеме живой ткани, может применяться на практике для определения концентрации глюкозы в межклеточном веществе и крови. Как было показано, концентрацию глюкозы в крови можно определить по экспериментальной зависимости осмотического давления от мощности падающего инфракрасного теплового излучения, представленной на Рис. 3а, по углу наклона графика зависимости осмотического давления межклеточного вещества эпидермиса от мощности падающего теплового излучения.
Метод микрокалориметрической спектроскопии, описанный выше, позволяет определить зависимость осмотического давления от тепловой мощности другим способом, с применением модулированного электромагнитного излучения (с амплидной или частотной модуляцией). В схеме измерения с амплитудной модуляциией, применяется излучение на длине волны, соответствующей выбранной характеристической частоте воды, с модуляцией амплитуды (интенсивности). Важная особенность спектрального метода измерения с применением излучения с амплитудной модуляцией, заключается в том, что метод позволяет исключить зависимость сигнала осмотического давления от низкочастотных флуктуации климатических параметров. Метод спектрального измерения с амплитудной модуляцией позволяет выполнить калибровку измерительного устройства неинвазивным способом, путем определения величины энтальпии и других калибровочных параметров с помощью калорического уравнения (45).
В варианте частотной модуляции, спектральные измерения проводятся на нескольких длинах волн, соответствующих характеристическим частотам поглощения воды. Измерение отклика осмотического давления межклеточного вещества на электромагнитное излучение (с одинаковой мощностью) на нескольких разных длинах волн, для которых предварительно определены и известны коэффициенты поглощения, позволяет определить зависимость сигнала осмотического давления межклеточного вещества от мощности электромагнитного излучения, поглощенной в локальной области живой ткани под аппликатором.
Зависимость осмотического давления от поглощенной мощности, полученная описанным спектральным методом с использованием модулированного излучения, позволяет определить концентрацию глюкозы в крови. Важная особенность метода, заключается в том, что спектральный способ измерения на нескольких частотах (по крайней мере на двух), которые можно выполнить в реальном масштабе времени, позволяет исключить вклад в сигнал осмотического давления, обусловленный низкочастотными флуктуациями климатических параметров.
К факторам, ограничивающим точность описанного спектрального метода для измерения концентрации глюкозы, относится близость пиков поглощения воды с пиками поглощения других молекул, например, гиалуроновой кислоты и глюкозы. Как было отмечено выше, в живой биологической ткани спектральные линии поглощения воды, интенсивность пиков которых определяется объемом внутриклеточной воды, расположены вблизи линий поглощения глюкозы. Результирующий сигнал осмотического давления в основном определяется поглощением мощности излучения внутриклеточной водой, вклад которого в сигнал превышает вклад, обусловленный поглощением мощности электромагнитной энергии биохимическими компонентами межклеточного вещества (гиалуроновой кислоты и глюкозы), поскольку объем внутриклеточного вещества, в единичном объеме, значительно превышает объем межклеточного вещества. Схема спектральных измерений на характеристических частотах воды, учитывающая спектр поглощений глюкозы, позволяет увеличить отношение сигнал/шум в измерительной схеме.
Спектры поглощения глюкозы в ИК области и пропускания в ближней ИК области в интервале длин волн от 1,428 мкм (7000 см-1) до 20 мкм (500 см-1), которые в настоящее время исследованы достаточно подробно, показаны на Рис. 32. На Рис. 33 представлены спектры поглощения глюкозы в ближней ИК области в интервале длин волн 1,428 мкм (7000 см-1) - 2,5 мкм (4000 см-1). Как видно из приведенных графиков, колебания концентрации глюкозы в крови приводит к колебаниям интенсивности пиков поглощения глюкозы.
Для определения зависимости осмотического давления межклеточного вещества от мощности падающего электромагнитного излучения, в однолучевой схеме измерений с амплитудной модуляцией излучения, длину волны можно выбрать в одном из интервалов между пиками поглощения глюкозы, например, 7000 см-1 (1,428 мкм), 5000 см-1 (2,0 мкм), 4000 см-1 (2,5 мкм).
В дифференциальной двухлучевой схеме измерений с применением излучения с частотной модуляцией, на двух (или более) длинах волн, можно выбрать частоты, не попадающие в интервалы пиков поглощения глюкозы, боковыми частотами пика поглощения глюкозы, например, 7000 см-1 (1,428 мкм), 5000 см-1(2,0 мкм), 4000 см-1 (2,5 мкм).
В описанной выше схеме спектрального измерения с применением модулированного излучения не учитывается возможный вклад в результирующий сигнал пиков поглощения межклеточной гиалуроновой кислоты, которые расположены вблизи пиков поглощения воды, однако дают в результирующий сигнал вклад, который значительно меньше вклада, обусловленного поглощением на частотах воды, поскольку объем внутриклеточного вещества, в единичном объеме живой ткани, значительно превышает объем межклеточного вещества.
Таким образом, концентрацию глюкозы в крови можно определить на основе изотермической калориметрической спектроскопии двумя разными способами:
метод терагерцовой спектроскопии - измерение концентрации тетрамеров гиалуроновой кислоты межклеточного вещества (пропорциональной концентрации связанной глюкозы в межклеточном веществе) на длине волны терагерцового излучения (λ≈152 мкм), соответствующего характеристической частоте тетрамера;
метод инфракрасной спектроскопии - измерение концентрации тетрамеров гиалуроновой кислоты на основе зависимости осмотического давления межклеточного вещества от поглощенной мощности излучения, которую определяют в реальном времени на основе спектральных измерений с применением излучения с амплитудной модуляцией на одной длине волны (или на нескольких длинах волн), соответствующей характеристической частоте воды в инфракрасной или другой области электромагнитного спектра.
Отношение сигнал/шум в схеме спектрального измерения, описанной выше, можно улучшить за счет дополнительного измерения других биохимических компонентов межклеточного вещества, в частности, содержания гиалуроновой кислоты. Спектроскопия гиалуроновой кислоты и других компонентов межклеточного вещества рассмотрена ниже.
Изотермическая калориметрическая спектроскопия межклеточного вещества.
Спектр поглощения межклеточного вещества (являющегося многокомпонентным биополимером) имеет более сложную структуру спектральных линий поглощения, по сравнению со спектром чистой воды, поскольку наряду с интенсивными пиками поглощения воды (в основном внутриклеточной), содержит пики поглощения, соответствующие характеристическим частотам биохимических компонентов межклеточного вещества, основными компонентами которого являются вода, гиалуроновая кислота и глюкоза, связанная с мономерами полимерной цепи гиалуроновой кислоты, межклеточная (тканевая) жидкость, схожая по составу с плазмой крови, которая состоит из воды, аминокислот, Сахаров, жирных кислот, коферментов. гормонов, нейромедиаторов, солей, а также отходов жизнедеятельности клеток.
Следует также отметить, что характеристические энергии связи молекул различных фармацевтических соединений, а также молекулы соединений и инородных включений в конденсированных средах межклеточного вещества, физиологически не совместимых с метаболизмом живой ткани, также могут оказаться в инфракрасном и терагерцовом диапазоне электромагнитного спектра и проявляться в спектрах поглощения межклеточного вещества.
Для оценки пиков поглощения исследуемой биохимической компоненты межклеточного вещества живой ткани, проводится сравнение спектрального сигнала (интенсивности излучения) на длине волны, соответствующей характеристической частоте исследуемого соединения, со спектральным сигналом, обусловленным поглощением чистой воды на данной длине волны.
На рисунке 34 ниже схематически показана структура межклеточного вещества, представляющего разветвленную макромолекулу гиалуроновой кислоты, имеющую конфигурацию полимерной сетки, окружающей живую клетку. Сшивки полимерной сетки образуются в результате объемных взаимодействий мономеров полимерной цепи с глюкозой растворителя (межклеточной жидкости) в конфигурации тетрамера - молекулярного комплекса, состоящего из двух не соседних дисахаридов полимерной цепи гиалуроновой кислоты и двух молекул D-глюкозы. Наряду с основной характеристической частотой (λ≈152 мкм) в терагерцовой области спектра, соответствующей энергии диссоциации молекулярного комплекса сшивки, в спектре поглощения межклеточного вещества будут проявляться пики поглощения, соответствующие характеристическим частотам дисахаридов гиалуроновой кислоты и молекулы D-глюкозы, связанной мономерами полимерной цепи межклеточного вещества.
Живая клетка, окруженная межклеточным веществом, является природным изотермическим калориметром мощности, позволяющим определить спектр поглощения межклеточного и внутриклеточного вещества живой ткани на основе измерения зависимости осмотического давления межклеточного вещества от спектральной плотности электромагнитного излучения на разных длинах волн (1, 2, 3, 4, 5, 6, 7): 1- терагерцовое излучение на характеристической частоте (λ≈152 мкм) тетрамера гиалуроновой кислоты; 2- излучение на характеристических частотах глюкозы, связанной с мономерами полимерной цепи гиалуроновой кислоты; 3, 4 - излучение на характеристических частотах дисахарида гиалуроновой кислоты; 5, 6 - излучение на характеристических частотах воды: 7 - электромагнитное излучение на характеристической частоте внутриклеточной молекулы.
Поглощение терагерцового излучения. длина волны которой соответствует характеристической частоте тетрамера гиалуроновой кислоты (λ≈152 мкм), приводит к диссоциации молекулярного комплекса тетрамера с преобразованием электромагнитной энергии в энергию теплового движения молекул, в результате которого происходит изотермическое изменение осмотического давления межклеточного вещества. Энергия падающего электромагнитного излучения, длина волны которой отличается от характеристической частоты тетрамера, но соответствует характеристическим частотам биохимических компонентов межклеточного вещества (гиалуроновая кислота, глюкоза, вода и др.), поглощается в объеме вещества и преобразуется в кинетическую энергию теплового движения молекул, которое приводит к изменению осмотического давления межклеточного вещества в результате диссоциации тетрамера гиалуроновой кислоты.
Спектры глюкозы.
На рисунке 32 показан график, поясняющий природу характеристических частот молекулы глюкозы в инфракрасной области.
Спектры гиалуроновой кислоты и свободных жирных кислот.
Инфракрасные спектры исследованных веществ, являющихся компонентами составных частей биологических тканей (глюкоза, гиалуроновая кислота, белок - яичный альбумин, липид -топленый бараний жир), в частотном диапазоне 4000-600 см-1, полученные в этом исследовании на ИК Фурье спектрометре (Tensor 37 фирмы Bruker), представлены на Рис. 36.
В высокочастотной ИК области спектра 4000 см-1 (2,5 мкм) - 3000 см-1 (3,3 мкм), в которой наблюдается интенсивный пик поглощения воды (Рис. 31), в спектрах гиалуроновой кислоты, глюкозы и белка, также наблюдаются интенсивные пики поглощения, обусловленные характеристическими частотами валентной связи О-Н групп. В этой области (4000-3000 см-1) отсутствует пик поглощения жира, который наблюдается рядом в интервале 3040 см-1 (3,29 мкм) -2853 см-1 (3,5 мкм), в которую попадают также линии поглощения всех рассматриваемых компонентов, обусловленные частотами СНn-группировок, присутствующих в структуре этих молекул.
Структуру спектра в более низкочастотной области ИК спектра можно проанализировать на основе выделенного фрагмента спектров всех рассматриваемых компонентов. Очень интенсивная узкая полоса карбоксильных карбонилов 1743 см-1 (5,7 мкм) и менее интенсивная 1553 см-1 (6,4 мкм) присутствует в спектре липида (бараньего топленого жира); эти две полосы, присутствующие также в спектрах всех животных жиров (бараньего топленого жира, свиного, куриного, суркового, рыбьего и растительных масел - оливкового, миндального, льняного, хлопкового, подсолнечного и др.), обусловлены частотами триглицерида жирных (преимущественно ненасыщенных) кислот.
Полоса карбоксильных карбонилов 1743 см-1 (5,7 мкм) присутствует в спектре гиалуроновой кислоты, полимерная цепь которой состоит из большого числа повторяющихся мономеров, каждый из которых состоит из дисахарида, имеющего в структуре группу -СООН. Обе полосы карбонилов 1743 см-1 (5,7 мкм) и 1553 см-1 (6,4 мкм) отсутствуют в спектре глюкозы (фруктозы, сахарозы, целлюлозы, крахмала). Полоса карбоксильных карбонилов 1553 см-1 (6,4 мкм) наблюдается только в спектре белка (яичного альбумина, что позволяет отнести ее к проявлению С=O групп полипептидной связи (1600 см-1-1550 см-1).
Спектральный метод измерения содержания гиалуроновой кислоты.
Измерение содержания гиалуроновой кислоты в живой ткани является актуальной задачей, имеющей самостоятельное практическое значение в косметологии, дерматологии, регенерационной медицине и др.; актуальность задачи обусловлена необходимостью мониторинга гиалуроновой кислоты с целью ранней диагностики диабета, а также в процессе неинвазивного мониторинга сахара крови у пациентов с сахарным диабетом. В процессе клинических исследований на группе пациентов с сахарным диабетом нами было обнаружено влияние динамики содержания гиалуроновой кислоты на временную динамику осмотического давления, при постоянной концентрации глюкозы в крови, во временном интервале эксперимента.
Как видно из сравнения спектров, линии поглощения гиалуроновой кислоты и глюкозы находятся вблизи интенсивного пика поглощения воды в области (4000-3000 см-1), при этом интенсивность пика поглощения воды, определяется поглощением в объеме клетки, поэтому многократно превышает интенсивность пиков глюкозы и гиалуроновой кислоты межклеточного вещества. По этой причине спектроскопия гиалуроновой кислоты и глюкозы, являющихся компонентами межклеточного вещества, сводится к выделению слабых спектральных пиков поглощения компонентов межклеточного вещества на фоне интенсивного спектрального пика воды, обусловленного поглощением воды в объеме клетки, многократно превышающего интенсивность спектральных пиков глюкозы и гиалуроновой кислоты межклеточного вещества; задача спектрального измерения биохимических компонентов межклеточного вещества является более сложной экспериментальной проблемой по сравнению с задачей спектрального измерения количества воды в единице объема живой ткани.
ИК спектроскопия гиалуроновой кислоты. Как известно, распределение гиалуроновой кислоты неравномерно по толщине эпидермиса, ее содержание в роговом слое и верхних слоях эпидермиса выше, чем ее содержание в дерме; авторами исследования предполагается, что транспорт гиалуроновой кислоты в роговой слой осуществляется из кератиноцитов, расположенных непосредственно под роговым слоем эпидермиса; кератиноциты - основные клетки эпидермиса человека (составляют примерно 90% всех клеток эпидермиса), которые содержатся в базальном, шиповатом, зернистом, блестящем, роговом слоях эпидермиса.
Для измерения содержания гиалуроновой кислоты в верхних слоях эпидермиса можно применить метод спектроскопии диффузного отражения, основанный на регистрации электромагнитного излучения, диффузно отраженного от верхних слоев эпидермиса. Схема спектрального измерения содержания гиалуроновой кислоты в эпидермисе, основанная на приеме диффузно отраженного от эпидермиса электромагнитного излучения, показана на Рис. 37.
На рис. 37 изображена схема спектроскопии диффузного отражения для измерения содержания гиалуроновой кислоты и свободных жирных кислот в межклеточном веществе поверхностного слоя эпидермиса живой ткани. Диффузно отраженное от поверхностного слоя эпидермиса излучение собирается под широким углом и передается на детектор.
Для измерения содержания гиалуроновой кислоты в поверхностном слое эпидермиса можно использовать ближнее электромагнитное излучение ближней инфракрасной (или оптической) области с длиной волны, соответствующей характеристической частоте гиалуроновой кислоты. Например, можно выбрать частоту в интервале пика поглощения 7000 см-1 (1,43 мкм), который расположен на спектре рядом с пиком поглощения глюкозы 6000 см-1 (1,67 мкм). Спектр гиалуроновой кислоты в ближней ИК области показан на Рис. 32.
На рис. 38 представлены спектры поглощения гиалуроновой кислоты в ближней ИК области и спектр пропускания в ИК области.
С другой стороны, пики поглощения гиалуроновой кислоты, как было отмечено, наблюдаются вблизи интенсивных пиков поглощения воды. На Рис. 39 представлены спектры поглощения гиалуроновой кислоты в ИК и ближней ИК области спектра, из которых следует, что пики поглощения гиалуроновой кислоты вблизи 35 00 см-1 (2,8 мкм) и 7000 см-1 (1,43 мкм), накладываются на пики поглощения воды (Рис. 31).
Для выделения спектрального сигнала, соответствующего пику поглощения гиалуроновой кислоты, на фоне интенсивного пика поглощения воды, используют однолучевой (одна длина волны) комбинированный спектральный метод, который получается в результате комбинации двух методов - изотермической калориметрической спектроскопии, который позволяет определить содержание воды в эпидермисе (поскольку интенсивность пика поглощения внутриклеточной воды многократно выше интенсивности поглощения межклеточной гиалуроновой кислоты), и спектроскопии диффузного отражения, который позволяет определить суммарное содержание воды и гиалуроновой кислоты в поверхностном слое эпидермиса. Вклад гиалуроновой кислоты выделяют из результирующего спектрального сигнала диффузного отражения путем вычитания из суммарного сигнала временной динамики количества воды в роговом слое эпидермиса под аппликатором, непроницаемом для воды и тепла, приложенном к поверхности с дозированным давлением. Измерение проводится по схеме однолучевой спектроскопии на одной длине волны, соответствующей одной из характеристических частот гиалуроновой кислоты, которую выбирают в частотном интервале спектра, в котором пики поглощения гиалуроновой кислоты накладываются (или близко расположены) на пики поглощения воды (УФ, оптическая, ближняя ИК, ИК, дальняя ИК). В ближней ИК области можно выбрать, например, длину волны 7000 см-1 (1,43 мкм), в ИК области длину волны 3500 см-1 (2,8 мкм). В рассмотренном варианте комбинированной схемы, применение дополнительного спектрального измерительного канала, основанного на регистрации электромагнитного излучения, диффузно отраженного от поверхностного слоя эпидермиса, а также применение измерительного метода синхронного детектирования сигналов, позволяет увеличить отношение сигнал /шум измерительных каналов. Зависимость отклика осмотического давления от мощности падающего электромагнитного излучения с амплитудной модуляцией на длине волны, соответствующей характеристической частоте воды, позволяет определить концентрацию глюкозы на основе метода, описанного выше.
Таким образом, применение комбинации двух спектральных методов (изотермической калориметрической спектроскопии и спектроскопии диффузного отражения) в одном спектральном устройстве позволяет получить лучшее разделение сигналов и определить содержание трех биохимических компонентов живой ткани, глюкозы, гиалуроновой кислоты и воды, с помощью измерений на одной длине волны, соответствующей одной из характеристических частот гиалуроновой кислоты (однолучевая спектроскопия).
Спектральный двухлучевой метод измерения содержания глюкозы.
Комбинированный спектральный метод, описанный выше, основанный на двухлучевой спектроскопии (на двух длинах волн), соответствующих характеристическим частотам глюкозы и гиалуроновой кислоты (вблизи пика поглощения воды), позволяет получить лучшее разделение сигналов и с более высоким отношением сигнал/шум определить концентрацию глюкозы, а также содержание двух других компонентов, гиалуроновой кислоты и воды, низкочастотная временная динамика которых непосредственно влияет на отношение сигнал/шум при измерении концентрации глюкозы. Спектральное измерение на каждой длине волны проводится двумя способами (изотермической калориметрической спектроскопии и спектроскопии диффузного отражения), комбинация которых в одном спектральном устройстве позволяет получить лучшее разделение сигналов. В ближней ИК области можно выбрать, например, длину волны 6000 см-1 (1,67 мкм), соответствующую характеристической частоте глюкозы, и длину волны 7000 см-1 (1,43 мкм), соответствующую характеристической частоте гиалуроновой кислоты.
В другом варианте двухлучевой схемы измерения, концентрацию глюкозы определяют с помощью калорического уравнения (45), в котором величину энтальпии ΔН, которая зависит от содержания гиалуроновой кислоты в межклеточном веществе (и имеет индивидуальные значения для разных пациентов) и количества воды в единичном объеме ткани, которая зависит от климатических параметров, определяют спектральным методом по линиям поглощения гиалуроновой кислоты и воды.
В другом варианте, концентрацию глюкозы определяют по схеме трехлучевой спектроскопии на 3 длинах волн, соответствующих выделенным пикам поглощения глюкозы, воды и гиалуроновой кислоты. Спектральные измерения проводятся в следующих диапазонах спектра: в интервале 3550-3050 см-1 (2,8-3.3 мкм) на одной частоте 3339 см-1 (2,99 нм), соответствующей пику поглощения воды (или трех частотах, соответствующих пикам поглощения воды, гиалуроновой кислоты и глюкозы); на частоте 1637 см-1 (6,08 мкм), соответствующей второму пику поглощения глюкозы; на частоте 1553 см-1 (6,4 мкм), соответствующей третьему пику поглощения глюкозы.
Спектральные линии поглощения глюкозы в оптической и ближней инфракрасной области спектра также находятся вблизи интенсивных пиков поглощения воды (Рис. 31). На Рис. 40 показаны теоретические спектры поглощения глюкозы, меланина и воды в красном и ближнем ИК диапазоне.
Метод измерения концентрации глюкозы в красном и ближнем ИК диапазоне основывается на однолучевой комбинированной схеме, описанной выше. Для выделения спектрального сигнала, соответствующего пику поглощения глюкозы, на фоне интенсивного пика поглощения воды, используют однолучевой комбинированный спектральный метод, который получается в результате комбинации двух методов - изотермической калориметрической спектроскопии, который позволяет определить содержание воды в эпидермисе (поскольку интенсивность пика поглощения внутриклеточной воды многократно выше интенсивности поглощения межклеточной гиалуроновой кислоты), и спектроскопии диффузного отражения, который позволяет определить суммарное содержание воды и глюкозы в поверхностном слое эпидермиса. Комбинация двух спектральных методов (изотермической калориметрической спектроскопии и спектроскопии диффузного отражения) в одном спектральном устройстве позволяет получить лучшее разделение сигналов и определить содержание двух биохимических компонентов живой ткани, глюкозы и воды, с помощью измерений на одной длине волны, соответствующей одной из характеристических частот глюкозы (однолучевая спектроскопия).
Канал измерения на основе изотермической калориметрической спектроскопии позволяет определить содержание воды в объеме эпидермиса, поскольку интенсивность пика поглощения внутриклеточной воды многократно выше интенсивности поглощения глюкозы, концентрация которой в объеме клетки существенно меньше ее концентрации в межклеточном пространстве, в результате процесса окисления глюкозы в объеме клетки; второй канал измерения, на основе спектроскопии диффузного отражения, позволяет определить суммарное содержание воды и глюкозы в поверхностном слое эпидермиса.
В варианте двухлучевой спектроскопии, для получения лучшего разделения, используют две длины волны, например, длина волны в интервале 930-1050 нм, соответствующей ширине пика поглощения воды, и длина волны в интервале 1000-1100 нм, соответствующая пику поглощения глюкозы; дополнительно может использоваться длина волны в интервале 900-970 нм, соответствующая второму пику поглощения воды. В варианте трехлучевой спектроскопии, дополнительно определяют содержание гиалуроновой кислоты, на длине волны, которую выбирают в этом же диапазоне частот, или в другой области ближнего ИК, например, 7000 см-1 (1,43 мкм).
Схема спектральных измерений в оптическом диапазоне должна быть разработана с учетом необходимости индивидуальной калибровки на содержание меланина кожи.
Метод измерения содержания триглицерида жирной кислоты.
Электромагнитное излучение с длиной волны, соответствующей характеристическим частотам свободных жирных кислот, будет интенсивно поглощаться в липидном слое эпидермиса, расположенном под роговым слоем, а также в объеме жирового слоя, расположенного под дермой. Липидный слой эпидермиса, расположенный под роговым слоем, представляет собой 10-25 слоев клеток, ориентированных параллельно поверхности и погруженных в липидный матрикс, составляющий около 10% его объема, имеющий уникальную структуру и химический состав. Кожа содержит, как свободные жирные кислоты, так и жирные кислоты, связанные в триглицериды, фосфолипиды и церамиды. Липиды эпидермиса в основном состоят из: церамидов (50%); холестирина и его эфиров (25%) насыщенных жирных кислот (10%).
С целью определения содержания свободных жирных кислот можно применить спектральный метод, который является комбинацией двух спектральных методов, изотермической калориметрической спектроскопии (для зондирования жирового слоя кожи, расположенного под дермой) и спектроскопии диффузного отражения, основанной на регистрации электромагнитного излучения, диффузно отраженного от межклеточного вещества липидного слоя эпидермиса с высоким содержанием свободных жирных кислот, расположенного под роговым слоем; представляющего собой 10-25 слоев клеток, ориентированных параллельно поверхности и погруженных в липидный матрикс, составляющий около 10% его объема, имеющий уникальную структуру и химический состав. Липиды эпидермиса в основном состоят из: церамидов (50%); холестирина и его эфиров (25%); насыщенных жирных кислот (10%) (рис. 41).
Как видно спектров (Рис. 36), два пика поглощения гиалуроновой кислоты попадают в полосы интенсивных пиков поглощения триглицерида жирных кислот 1743 см-1 (5,7 мкм) и 3040 см-1-2853 см-1 (3,3-3,5 мкм), которые находятся в интервале между двумя пиками поглощения воды 1645 см-1 (6,08 мкм) и 3339 см-1 (3,0 мкм).
Содержание в живой ткани триглицерида жирных кислот и гиалуроновой кислоты можно определить с помощью метода изотермической калориметрической однолучевой спектроскопии на длинах волн, соответствующих характеристическим частотам триглицерида жирной кислоты 1743 см-1 (5,7 мкм) и/или 3040-2853 см-1 (3,3-3,5 мкм).
В другом варианте, спектральный метод для измерения содержания триглицерида жирной кислоты, основан на применении двухлучевой спектральной схемы на длинах волн, соответствующих одной из характеристических частот гиалуроновой кислоты (комбинированный метод, описанной выше), например, 3500 см-1 (2,8 мкм) - 3000 см-1 (3,3 мкм), и одной из характеристических частот триглицерида жирной кислоты, например, 3040 см-1-2853 см-1 (3,3-3,5 мкм). В этом варианте, спектральное измерение в интервале 2,8 мкм - 3,3 мкм проводят на основе комбинированного метода (объединяющего изотермическую калориметрическую спектроскопию и спектроскопию диффузного отражения), а измерение на длине волны триглицерида проводят по схеме спектроскопии диффузного отражения, позволяющей определить содержание триглицерида в поверхностном липидном слое эпидермиса или по схеме изотермической калориметрической спектроскопии, позволяющей определить содержание триглицерида в жировом слое под дермой. Описанная схема измерения позволяет регистрировать сигнал, пропорциональный содержанию триглицерида, или суммарный сигнал, пропорциональный суммарному содержанию глюкозы и липидов, характеризующий интенсивность метаболизма глюкозы и свободных жирных кислот.
Метод трехлучевой спектроскопии глюкозы, гиалуроновой кислоты и триглицерида жирной кислоты.
Концентрацию глюкозы можно определить на основе схемы трех лучевой спектроскопии, в которой применяется метод спектроскопии с помощью модулированного излучения на длине волны, соответствующей характеристическим частотам воды, путем добавления схемы двухлучевой спектроскопии на длинах волн, соответствующих характеристическим частотам гиалуроновой кислоты и триглицерида жирной кислоты. Преимущество этой схемы в том, что интенсивность пиков поглощения практически не меняется за время проведения экспериментов в течении дня (и недели), поэтому измерение сводится к схеме двухлучевой спектроскопии на двух длинах волн, соответствующих характеристическим частотам воды и гиалуроновой кислоты. Однолучевая схема спектрального прибора на длине волны, соответствующей характеристической частоте 1743 см-1 (5,7 мкм) может применяться для мониторинга содержания гиалуроновой кислоты живой ткани.
Метод спектроскопии внутриклеточных биохимических макромолекул и органелл.
Метод микрокалориметрической спектроскопии, описанный выше, в общем случае, позволяет определить концентрацию макромолекул внутриклеточного вещества с применением электромагнитного излучения с частотами (длинами волн), которые совпадает с характеристическими частотами тетрамеров внутриклеточных макромолекул (белки, РНК, ДНК).
Характеристические частоты внутриклеточных белков попадают в дальний инфракрасный диапазон электромагнитного спектра. Например, характерные энергии связи в макромолекуле глобулярного фермента глюкокиназы и других внутриклеточных ферментов имеют величины порядка 7 ккал/моль - 20 ккал/моль или 10 kBТ-30 kBТ, соответствующих интервалу длин волн 20-70 мкм в дальней ИК области спектра.
Концентрацию основных биохимических компонентов межклеточного вещества, в общем случае, можно определять с помощью дифференциального метода по схеме четырех лучевой (четырех волновой) спектроскопии, в котороой используются длины волн, соответствующие характеристическим частотам 3 основных компонентов межклеточного вещества, гиалуроновой кислоты, глюкозы, молекулы воды, а также длина волны, соответствующая характеристической частоте внутриклеточной макромолекулы или комплекса; в схеме измерения, каждая из четырех биохимических соединений, может быть представлена набором из 1 (или более) характеристических частот.Поэтому, число длин волн, характеризующих группу из 2 биохимических компонентов межклеточного вещества, молекулы воды и 1 исследуемой внутриклеточной молекулы, может быть больше 4.
Поглощение внутриклеточными макромолекулами (и биохимическими комплексами) падающего электромагнитного излучения с длинами волн, соответствующих характеристическим частотам внутриклеточных биохимических компонент, будет приводить к изменению теплового баланса в контролируемой области живой ткани под аппликатором (нагреву) и, как следствие, к изменению осмотического давления межклеточного вещества в результате разрыва внутренних межмолекулярных связей тетрамеров гиалуроновой кислоты межклеточного вещества, имеющих характерную энергию связи порядка kBТ.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Предлагаемый способ спектроскопии межклеточного вещества локального участка живой ткани может быть реализован при помощи спектрального устройства, принципиальная схема которого показана на рис. 42. Блок - схема спектрального устройства для изотермической калориметрической спектроскопии биохимических компонентов живой ткани (изотермический калориметрический спектрометр) включает 1- аппликатор, непроницаемый для воды и тепла; 2 -датчик осмотического давления межклеточного вещества; 3 - источник электромагнитного излучения с постоянной и/или модулированной интенсивностью; 4 - датчик физиологического параметра; 5 - блок инструментальных усилителей и/или синхронный детектор и/или аналогово-цифровой преобразователь (АЦП); 6 - блок обработки информации; 7 - блок отображения информации.
Устройство содержит несколько датчиков для измерения физиологических параметров, один из которых предназначен для измерения осмотического давления межклеточного вещества, а также один или несколько источников электромагнитного излучения с постоянной или модулированной интенсивностью.
Устройство включает в себя аппликатор 1 (непроницаемый для тепла и воды, имеющий верхнюю и нижнюю поверхности), выполненный с возможностью наложения на поверхность эпидермиса с дозированным давлением, один или несколько источников 3 электромагнитного излучения, с постоянной или модулированной интенсивностью, с длиной волны, соответствующей характеристической частоте исследуемого биохимического компонента живой ткани, устройство для создания калибровочного воздействия на контролируемый участок ткани под аппликатором, датчик осмотического давления межклеточного вещества (2), датчики температуры и других физиологических параметров (4), расположенные под аппликатором, с внутренней стороны аппликатора на поверхности РСЭ, при этом сигналы с вышеуказанных датчиков поступают последовательно на входы блока 5 усилителей и/или синхронного детектора и/или аналогово-цифрового преобразователя (далее АЦП), установленных на верхней поверхности аппликатора 1; блока 6 обработки информации; блока 7 отображения информации.
Блоком 6 обработки информации является микропроцессор для обработки в реальном времени сигналов от датчиков физиологических параметров локальной области ткани под аппликатором и датчиков климатических параметров окружающей среды.
В одном из вариантов осуществления блок 6 обработки информации расположен на верхней поверхности аппликатора 1, в другом - является удаленным и связан проводной связью с АЦП.
Блоком 7 отображения информации является смартфон или персональный компьютер. При этом связь мультисенсорного устройства с блоком 7 отображения информации может быть в различных вариантах осуществления как беспроводной, так и проводной (посредством USB канала).
В качестве датчиков для измерения динамики физиологических параметров локальной области ткани под аппликатором используют по меньшей мере датчик 2 осмотического давления, с помощью которого определяют количество воды в межклеточном пространстве ткани в локальном объеме под аппликатором на контролируемой глубине, на которой устанавливается локальное термодинамическое равновесие; датчик 4 температуры РСЭ с помощью которого определяют температуру поверхности кожи под аппликатором и датчик эластического давления ткани под аппликатором.
Сигналы датчиков физиологических параметров (датчик осмотического давления 2, датчик температуры 3 и датчик эластического давления 4), расположенных на внутренней поверхности аппликатора 1, поступают на входы инструментальных усилителей и/или аналогово-цифрового преобразователя (блок 10), после которых поступают в блок обработки информации 11, в котором производится обработка сигналов. Обработанная информация передается в устройство отображения 12, на экране которого в реальном времени отображаются текущие значения измеряемых параметров и/или скорости метаболизма параметров.
В случае использования электромагнитного излучения с амплитудной модуляцией сигнала, с целью повышения отношения сигнал /шум применяется метод синхронного (когерентного) детектирования AM сигналов, который позволяет существенно повысить отношение сигнал/шум регистрируемого сигнала и уменьшить его искажения. В этом случае, синхронное детектирование сигнала на выходе датчика осмотического давления осуществляется с помощью инструментального усилителя (со встроенной схемой синхронного детектирования) на частоте AM модуляции электромагнитного излучения.
Рис. 43. Схема спектроскопии диффузного отражения для определения осмотического давления межклеточного вещества на основе спектрального измерения временной динамики количества воды в роговом слое эпидермиса на длине волны, соответствующей характеристической частоте воды (1190, 1455, 1945, 945 нм). Диффузно отраженное от эпидермиса излучение собирается под широким углом и передается на детектор. 1- источник излучения на длине волны, соответствующей характеристической частоте воды в роговом слое эпидермиса; 2 - аппликатор, непроницаемый для воды и тепла; 3- излучение на длине волны, соответствующей характеристической частоте исследуемой биохимической компоненты эпидермиса (живой ткани); 4- приемник электромагнитного излучения.
Для практической реализации мультисенсорного устройства могут использоваться промышленные сертифицированные датчики для измерения климатических параметров, а также сертифицированные сенсоры для измерения температуры рогового слоя и эластического давления эпидермиса. Например, в качестве датчика для измерения температуры рогового слоя могут быть использованы сенсоры фирмы Honeywell.
В качестве датчика для измерения эластического давления могут использовать сенсоры на основе пьезоэлектрического элемента.
Датчик осмотического давления 2, регистрирующий динамику переноса воды в эпидермисе путем регистрации временной динамики содержания воды в роговом слое эпидермиса, может быть основан на разных физическо-химических методах, основанных на разных физических принципах. В частности, для измерения количества воды в роговом слое применимы методы электрометрии, основанные на измерении электрофизических характеристик РСЭ (электропроводность, диэлектрическая проницаемость); спектральные методы, основанные на измерении спектральных характеристик (коэффициенты отражения и поглощения); оптико-акустические методы; теплофизические методы, основанные на измерении теплофизических характеристик (теплопроводность, теплоемкость); электрохимические методы и др. Также датчик количества воды может быть основан на измерении эластического давления. Устройства для измерения количества воды в эпидермисе описаны в патенте [10].
В случае применения электрометрического датчика, количество воды в эпидермисе можно определить путем измерения электрических характеристик (электропроводности и/или диэлектрической проницаемости) рогового слоя на переменном или постоянном токе.
В случае применения спектрального датчика, количество воды в эпидермисе можно определить путем измерения спектральных характеристик (коэффициента отражения и/или коэффициента поглощения) рогового слоя для электромагнитного излучения с длиной волны вблизи линии поглощения, обусловленной содержанием воды в РСЭ. В частности, временную динамику процесса набухания межклеточного вещества можно регистрировать по временной динамике коэффициента отражения рогового слоя эпидермиса. Увеличение количества воды в межклеточном пространстве приводит к изменению спектральной характеристики рогового слоя, обусловленному изменением количества воды в РСЭ.
Спектральный датчик может основываться на способе, в котором осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют путем измерения спектральных характеристик межклеточного вещества в диапазоне спектра электромагнитного излучения, который определяют по характеристическим частотам поглощения межклеточного вещества, и выбирают из группы, включающей в себя характеристические частоты воды в оптическом и/или инфракрасном и/или микроволновом диапазоне; групповые частоты гиалуроновой кислоты межклеточного вещества в инфракрасном и/или терагерцовом и/или микроволновом диапазоне.
В другом варианте, спектральный датчик основывается на способе определения осмотического давления межклеточного вещества методом измерения количества воды в роговом слое эпидермиса путем измерения физических характеристик рогового слоя эпидермиса, которые выбирают из группы, включающей электрофизические характеристики, спектральные и оптико-акустические характеристики, теплофизические характеристики.
В случае спектрального метода, осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса путем измерения спектральных характеристик рогового слоя эпидермиса на характеристических частотах поглощения воды в роговом слое с помощью спектрального метода, который выбирают из группы, включающей, ИК спектроскопию, Раман - спектроскопия, оптико-акустическая спектроскопия, разностная (дифференциальная) двухлучевая спектроскопия.
Дозированное калибровочное воздействие выбрано из группы, включающей в себя внешнее давление, локальную декомпрессию, нагрев, охлаждение, электрический ток или напряжения и магнитное поле.
В качестве устройства 8 для создания калибровочного воздействия можно использовать, например, устройство, генерирующее импульсы электрического напряжения или тока через РСЭ, которые позволяют определить текущее значение и направление микропотоков тканевой жидкости в межклеточном веществе глубинного слоя эпидермиса путем измерения кинетических характеристик массопереноса, обусловленного электрокинетическими явлениями, в частности, электро-осмосом.
В альтернативных вариантах осуществления устройством для создания калибровочного воздействия является источник тепловой мощности, выполненный в виде резистора или элемента Пельтье, или устройство для создания дозированного давления на поверхность аппликатора.
В качестве источников электромагнитного излучения в оптической и инфракрасной областях могут использовать промышленные лазерные диоды (на основе квантово-размерных гетеро-структур и других типов), которые широко распространены и производятся с большим разнообразием характеристик излучения, специализированных для определенных применений. Официальных стандартов по типам и размерам корпусов лазерных диодов не существует, однако иногда крупные производители заключают соглашения об унификации корпусов. Кроме того, существуют услуги по разработке корпуса излучателя по требованиям заказчика.
В качестве приемника электромагнитного излучения в оптическом и инфракрасном диапазоне могут использоваться фотоприемники, которые выпускаются промышленностью. Ведущими компаниями мира выпускается довольно много типов микросхем, в которых приемник излучения (фотоприемник) и усилитель объединены в одном корпусе. Такие системы широко используются в системах дистанционного управления на ИК - лучах (такие модули также называются ИК-ресиверами (IR Receiver). Ведущими компаниями мира выпускается довольно много типов таких микросхем.
Отечественной промышленностью, например, компаниями ООО «ЛЕД Микросенсор НТ» и ООО "АИБИ" (IBSG Co., Ltd.), выпускается широкий спектр оптоэлектронных приборов среднего инфракрасного диапазона - светоизлучающих диодов, светодиодных матриц, фотодиодов для спектрального диапазона от 1,6 мкм до 5,0 мкм, а также электронных устройств для работы с ними (драйверов для светодиодов и усилителей для фотодиодов).
В оптическом и ближнем ИК диапазоне также можно использовать светодиоды отечественного производства: SIM-012ST (950-970 нм); OIS-150-1020 (1020-1060 нм); KM2520F3C03 (930-950 нм); EDEF-1LS3 (740-760 нм); EDEI-1LS3 (830-850 нм). В качестве приемника оптического излучения использован фотодиод, чувствительный к оптическому излучению в диапазоне длин волн от 740 нм до 1060 нм, например, фотодиод типа BPW34.
Источники и приемники терагерцового излучения, миниатюрные по размерам и с характеристиками излучения, приемлемыми для практических применений, пока не выпускаются мировой промышленностью и находятся на стадии НИОКР. В частности, ученые из Принстонского университета (Xue Wu; Kaushik Sengupta, Department of Electrical Engineering, Integrated Microsystems Research Lab, Princeton University, Princeton, NJ, USA) добились радикального уменьшения аппаратуры для работы с электромагнитным излучением терагерцового диапазона. Подобные устройства могут стать важным элементом медицинской диагностики, коммуникаций и разработки лекарств. Две работы ученых на эту тему опубликованы в журнале IEEE Journal of Solid State Circuits. В статье ученые объяснили, как им удалось создать миниатюрную микросхему для генерации терагерцового излучения. Она пока работает в узком диапазоне, однако исследователи поняли, что это препятствие можно преодолеть, создавая сразу несколько волн различных длин на одной микросхеме и комбинируя их, используя точную синхронизацию. Дэниэл Миттлман, вице-председатель Международного общества инфракрасных, миллиметровых и терагерцового излучения, сказал, что работа «очень инновационна и потенциально имеет множество применений», а также отметил, что еще предстоит много работы до внедрения этих разработок в повседневные устройства, они являются перспективными.
Непроницаемый для воды и тепла аппликатор (1) с закрепленными на его нижней поверхности датчиками количества воды (межклеточного осмотического давления 2), температуры 3 и эластического давления эпидермиса на аппликатор 4, фиксируется на поверхности рогового слоя эпидермиса, например, на поверхности кисти руки, с внешним дозированным давлением и прикрепляется в одном из вариантов осуществления с помощью ремешков. Следует, что измерение можно производить на любом участке поверхности тела человека, свободном от волосяного покрова, за исключением участков тела, в которых расположены потовые железы, которые активируются в результате эмоционального воздействия. Примером такого участка являются ладони рук.
Предполагаемые размер, внешний вид и дизайн мультисенсорного устройства показаны на Рис. 44.
Процесс измерения (записи сигналов) начинается непосредственно в момент прижима по меньшей мере одного тепло- и водонепроницаемого аппликатора к поверхности РСЭ или с задержкой через 10-50 секунд с момента прижима.
После наложения аппликатора на поверхность кожи измеряют временную динамику физиологических параметров в локальной замкнутой области ткани под аппликатором в режиме «микрокалориметр», а именно, по меньшей мере тепловой поток через участок эпидермиса под аппликатором или температуру поверхности кожи под аппликатором, эластическое давление ткани под аппликатором, давление, создаваемое аппликатором на поверхность кожи, количество воды в межклеточном пространстве ткани на контролируемой глубине, на которой устанавливается локальное термодинамическое равновесие. Измеряют тепловой поток через участок эпидермиса под аппликатором или температуру на контролируемой глубине Tskin, на которой устанавливается локальное термодинамическое равновесие, при фиксированном давлении аппликатора Psensor.
Для вычисления уровня глюкозы в крови проводят процедуру калибровки и определяют калибровочные параметры с целью определения постоянных коэффициентов, необходимых для вычисления теплового эффекта метаболизма локального участка живой ткани и вычисляют уровень глюкозы в крови, пропорциональный величине теплового метаболизма локального участка живой ткани.
Число датчиков мультисенсорного устройства, описанного выше, может быть увеличено за счет включения в состав устройства дополнительных сенсоров и датчиков физиологических параметров, характеризующих метаболизм локального участка живой ткани, в частности, датчиков биохимических параметров крови (например, содержания лактата в крови), кислотности крови, частоты сердечно сосудистых сокращений. Дополнительные датчики для через-кожных измерений могут быть установлены на внутренней стороне аппликатора и интегрированы с датчиками физиологических и климатических параметров, описанными выше, в единую измерительную схему в составе мультисенсорного устройства.
Методы ранней диагностики сахарного диабета.
Способ диагностики сахарного диабета по динамике сахарной кривой. Современные руководства по медицине определяют нарушение толерантности к глюкозе (НТГ), как концентрации глюкозы в крови в ходе перорального теста толерантности к глюкозе, лежащие в промежутке между нормальными и диабетическими значениями (через 2 часа после приема 75 г глюкозы - от 7,8 до 11,0 ммоль/л); концентрацию глюкозы в процессе теста толерантности измеряют инвазивным методом с помощью глюкометра по нескольким пробам крови, которые берутся в процессе перорального теста. Предполагается, что НТГ является признаком преддиабетического состояния, хотя не у всех лиц с НТГ развивается диабет. В США НТГ имеется у каждого десятого взрослого, причем его частота увеличивается с возрастом, достигая каждого четвертого среди лиц в возрасте 65-74 лет.
Метод изотермической калориметрии живой ткани, описанный выше, открывает новые возможности для создания методов ранней диагностики сахарного диабета, в частности, позволяет определить временную динамику сахара крови в процессе НТГ в режиме непрерывного мониторинга (сахарная кривая). Характерная непрерывная (с частотой 1 измерение в секунду) неинвазивная сахарная кривая здорового пациента в процессе НТГ отличается от сахарных кривых диабетиков и характеризуется наличием участка роста концентрации сахара в крови (продолжительностью примерно 30 минут) и падающего участка (продолжительностью примерно 30 минут).
В процессе клинического исследования, выполненного на группе пациентов с сахарным диабетом, было обнаружено, что сахарные кривые пациентов с сахарным диабетом, регистрируемые непрерывно (с частотой 1 измерение в секунду) с помощью прототипа изотермического микрокалориметра, имеют скачкообразную динамику. Характерные сахарные кривые со скачкообразной временной динамикой, записанные в процессе клинического исследования, показаны на рисунке 45. Уровень сахара крови у пациента Д с сахарным диабетом 1 типа (33 лет, муж.) изменяется самопроизвольно скачком в моменты времени 800 сек и 1400 сек.
Как известно, уровень глюкозы в крови у здоровых пациентов определяется уровнем инсулина, который вырабатывается клетками поджелудочной железы пропорционально уровню глюкозы в крови, динамика которой как правило имеет монотонный характер без скачков. Отклонение механизма поджелудочной железы от нормы может привести к неравномерному скачкообразному поступлению инсулина в кровь и, как следствие, к скачкообразной динамике сахара в крови. На Рис. 46 показана скачкообразная динамика сахара крови в форме одиночного импульса, зарегистрированная с помощью прототипа изотермического микрокалориметра, и временная динамика уровня инсулина в крови, рассчитанная на основе сахарной кривой.
В процессе исследования было обнаружено, что скачкообразная динамика сахара крови может приводить к скачкообразной динамике концентрации сахара в межклеточной жидкости в форме последовательности одиночных импульсов; характерная сахарная кривая показана на рисунке 47. На рисунке 47 представлена характерная сахарная кривая) со скачкообразной временной динамикой у пациента с сахарным диабетом 2 типа (66 лет, муж.): характерная динамика сахара крови в форме временной последовательности одиночных импульсов.
Метод мониторинга сахара крови на основе метода изотермической калориметрии может применяться на практике для ранней диагностики сахарного диабета (1 и 2 типа) по особенностям скачкообразной динамики сахарной кривой.
Способ диагностики сахарного диабета на основе мониторинга энтальпии живой ткани.
Метод изотермической микрокалориметрии живой ткани позволяет определить на ранней стадии сахарный диабет (пред диабет) пациентов по содержанию гиалуроновой кислоты в эпидермисе, которая определяется на основе мониторинга осмотического давления межклеточного вещества (или количества воды в межклеточном пространстве).
В процессе клинического исследования, выполненного с помощью прототипа изотермического микрокалориметра живой ткани, нами было установлено, что величина энтальпии межклеточного вещества у практически здоровых пациентов, значение которого определялось экспериментально (при заданной мощности внешнего теплового потока, обусловленного климатическими параметрами окружающей среды) на основе сравнения неинвазивных показаний прототипа с инвазивными показаниями сертифицированного глюкометра (метод калибровки), не изменяется или изменяется очень медленно со временем, с характерной постоянной времени, превышающей один месяц и более. Результат, полученный экспериментально, находится в хорошем согласии с выводами, полученными на основе теоретического исследования термодинамики конденсированного состояния биополимерной молекулы, описанной выше, которые предсказывают постоянство величины энтальпии в интервале параметров, соответствующем физиологической норме Рис. 22, 23. Межклеточное вещество, в области физиологической нормы, границы которой определяются критическим значением концентрации дисахаридов (мономеров) гиалуроновой кислоты (критическая контурная длина полимерной цепи), при физиологической температуре, находится в гетерофазном состоянии, в котором кристаллическая (глобулярная) фаза вещества находится в термодинамическом равновесии с жидкой (расплавленная глобула) фазой, в которой концентрация мономеров соответствует концентрации насыщенного раствора полимерной цепи гиалуроновой кислоты. Величина энтальпии межклеточного вещества пациента определяется содержанием гиалуроновой кислоты, которое может изменятся при хронических заболеваниях. В частности, при возникновении диабетического состояния, содержание гиалуроновой кислоты может изменяться в результате изменения контурной длины (числа дисахаридов) полимерной цепи; поэтому, содержание гиалуроновой кислоты в межклеточном веществе является биохимическим параметром, мониторинг которого позволяет диагностировать на ранней стадии сахарный диабет.
Увеличение числа звеньев в полимерной цепи, эквивалентное введению в растворитель дополнительного количества дисахаридов гиалуроновой кислоты, при фиксированном объеме вещества, не приводит к увеличению его концентрации в жидкой фазе (пересыщение раствора) - но приводит к увеличению числа мономеров в кристаллической фазе в процессе кристаллизации (процесс выделения из раствора твердой фазы). Увеличение объема вещества за счет увеличения объема растворителя (вода с содержанием глюкозы и ионов натрия) будет приводить к процессу обратному кристаллизации - растворению (переходу вещества из кристаллической фазы в жидкую). Процесс растворения, также, как и процесс плавления, приводит к процессу разрушения кристаллической решетки, который обычно сопровождается охлаждением раствора. Растворение может происходить в изотермических условиях, но в таком случае необходим подвод тепла из вне.
В процессе клинического исследования, выполненного на группе пациентов с сахарным диабетом, было установлено, что величина энтальпии межклеточного вещества, значение которого определялось на основе сравнения неинвазивных показаний прототипа прибора с инвазивными показаниями сертифицированного глюкометра (метод калибровки), может принимать у диабетиков (2 типа) аномальные значения - аномально низкие или аномально высокие; таким образом, энтальпия межклеточного вещества живой ткани пациентов с сахарным диабетом характеризуется 2 значениями энтальпии, соответствующими 2 разным состояниям межклеточного вещества с разными значениям величины энтальпии межклеточного вещества, которые могут у некоторых пациентов могут отличаться более чем на порядок.
Полученные результаты исследования привели к пониманию вывода, что аномально высокое значение энтальпии межклеточного вещества, наблюдаемые у пациентов с сахарным диабетом, связано с повышенным содержанием (избытком) гиалуроновой кислоты в межклеточном веществе эпидермиса исследованных диабетических пациентов. Как известно, при диабете 2 типа содержание гиалуроновой кислоты (НА) в межклеточном веществе повышается, в результате возрастания скорости синтеза НА в условиях гипергликемии (повышенного уровня сахара крови) и, как следствие, приводит к возникновению избыточного количества гиалуроновой кислоты в кристаллической (глобулярной) фазе межклеточного вещества эпидермиса, верхний слой которого характеризуется малым содержанием растворителя в межклеточном пространстве.
Изменение количества растворителя в межклеточном пространстве ткани под аппликатором, в процессе перехода в состояние локального термодинамического равновесия, может привести к фазовому переходу вещества из кристаллического (глобулярного) состояния в жидкое (расплавленная глобула) состояние с увеличением межклеточного объема в результате растворения кристаллической (глобулярной) фазы, находящейся в метастабильном состоянии; процесс растворения, протекает за счет подвода тепла, в объем ткани под аппликатором, потоком межклеточной жидкости. К нарушению термодинамического фазового равновесия межклеточного вещества также могут привести скачкообразное изменения концентрации глюкозы и минеральных ионов в крови.
В процессе клинического исследования у диабетиков 2 типа нами была обнаружена корреляция величины энтальпии межклеточного вещества с инсулинорезистентностью ткани: у диабетиков 2 типа с высокой инсулинорезистентностью наблюдаются аномально низкие значения величины энтальпии ткани, соответствующие преобладанию кристаллической фазы в межклеточном веществе; увеличение доли кристаллической компоненты в веществе приводит к увеличению резистентности межклеточного пространства, обусловленной увеличением коэффициентов переноса вещества для переноса глюкозы и инсулина из капилляра в клетку и увеличению доли связанной глюкозы. В агрегатном состоянии (расплавленная глобула) с аномально высоким уровнем осмотического давления доля свободной глюкозы в локальном объеме межклеточного вещества может увеличивается за счет уменьшения доли связанной глюкозы в кристаллической фазе без увеличения концентрации инсулина и, как следствие, может привести к увеличению концентрации глюкозы в межклеточной жидкости.
Величина энтальпии межклеточного вещества, таким образом, является индивидуальной характеристикой мышечной ткани пациента, имеющей высокую диагностическую ценность. Непрерывное измерение энтальпии межклеточного вещества путем мониторинга содержания гиалуроновой кислоты и/или мониторинга осмотического давления в процессе инсулиновой (или другой сахар понижающей) терапии может быть использовано на практике для ранней диагностики диабета, а также для диагностики в реальном времени инсулинорезистентности живой ткани пациентов с сахарным диабетом (и контроля эффективности сахаро понижающей терапии) на основе мониторинга содержания гиалуроновой кислоты в эпидермисе. На Рис. 48 представлена характерная временная динамика осмотического давления межклеточного вещества (характеризующего количество воды в межклеточном пространстве), которая регистрируется в процессе самопроизвольного перехода межклеточного вещества живой ткани под аппликатором в состояние локального термодинамического равновесия (соответствующего максимуму энтропии); переход живой ткани в новое состояние с высоким значением энтальпии, возникает в результате процесса растворения кристаллической (глобулярной) фазы межклеточного вещества, протекающего за счет подвода тепла потоком межклеточной жидкости, и сопровождается увеличением объема и осмотического давления межклеточного вещества. Осмотическое давление межклеточного вещества, соответствующее равновесному значению энтальпии, которое устанавливается в процессе перехода ткани под аппликатором в состояние локального термодинамического равновесия, превышает исходное значение осмотического давления (соответствующее исходному значению энтальпии) более чем на два порядка (в 100 раз). Причиной фазового перехода межклеточного вещества могут быть скачки уровня сахара в крови и возможно других биохимических и физиологических параметров, и внешних физических факторов (давление, тепловой поток и др.).
Как известно, при инсулинорезистентности чувствительность мышечных клеток к инсулину снижается. В результате нарушается способность глюкозы проникать в клетки мышечной ткани. Глюкоза накапливается в плазме крови. В ответ поджелудочная железа вырабатывает инсулин в еще большем количестве, и его содержание в крови увеличивается выше нормальных значений. В результате глюкоза откладывается в виде жира, ведь жировые клетки хорошо восприимчивы к инсулину, а глюкозе нужно куда-то деваться из крови. Создается своеобразный замкнутый круг.
Аномальное содержание (избыток) гиалуроновой кислоты в межклеточном веществе у диабетиков, обусловленное повышенной скоростью синтеза гиалуроновой кислоты, приводит к кристаллизации межклеточного вещества, которое сопровождается повышенным связыванием глюкозы в кристаллической фазе и увеличением коэффициентов переноса межклеточного вещества для переноса глюкозы и инсулина из капилляра в клетку, поэтому может быть одной из причин, приводящих к развитию сахарного диабета 2 типа и других сердечно-сосудистых заболеваний, связанных с диабетом хронических заболеваний. Фазовый переход межклеточного вещества в агрегатное состояние с аномально высоким значением осмотического давления также приводит к повышению инсулинорезистентности и сопровождается повышением кровяного давления за счет пережатия (схлопывания) капилляров, обусловленного повышением осмотического давления вещества, окружающего капилляр.
Таким образом, метод изотермической калориметрии позволяет по характеру временной динамики осмотического давления межклеточного вещества обнаружить на ранней стадии и предсказать наличие признаков инсулинорезистентности и диабетического состояния у не диагностированных пациентов 2 типа. Метод изотермической калориметрической спектроскопии, описанный выше, также позволяет определить на ранней стадии диабетическое состояние на основе спектральных измерений содержания гиалуроновой кислоты в локальном объеме живой ткани.
Скачки осмотического давления межклеточного вещества, которые наблюдаются у пациентов с сахарным диабетом, могут приводить к скачкам кровяного давления. В частности, скачки осмотического давления внеклеточного вещества в сердце и мозге могут являться возможной причиной инфарктов и инсультов. Скачки осмотического давления в поджелудочной железе могут служить причиной неоднородной концентрации инсулина в крови, в результате скачкообразной динамики скорости образования инсулина в поджелудочной железе. Отсюда следует понимание вывода о природе взаимосвязи между сахарным диабетом и сердечно сосудистыми заболеваниями, которая сегодня доказана клинически. Метод изотермической калориметрии, описанный выше, также позволяет обнаружить на ранней стадии сердечнососудистые заболевания, обусловленные сахарным диабетом.
Воздействие некоторых внешних факторов, физиологических и физических, на ткань может приводить к обратимым переходам живой ткани из одного фазового состояния в другое. К числу таких факторов, относятся, в частности, мышечная нагрузка и температурные воздействия. К внешним воздействиям, которые вызывают обратимые переходы ткани изменения, относятся воздействия, которые приводят к обратимым изменениям фазового состояния межклеточного вещества. К числу таких внешних физических факторов относятся: внешнее давление; локальная декомпрессия; внешняя температура; электромагнитное излучение, вызывающее объемный нагрев ткани; слабый постоянный электрический ток; постоянное магнитное поле; локальная мышечная нагрузка на ткань и др. Метод изотермической микрокалориметрии, дополненный воздействием внешним физическим фактором, позволяет определить величину энтальпии и диагностировать на ранней стадии сахарный диабет, на основе измерения зависимости осмотического давления от внешнего физического фактора.
Нарушение толерантности к глюкозе.
Метод изотермической калориметрии живой ткани, описанный выше, позволяет определить НТГ и чувствительность ткани к инсулину по характеру временной динамики осмотического давления межклеточного вещества (количества воды в межклеточном пространстве), без определения абсолютной величины энтальпии (без калибровки). В процессе клинического исследования у диабетиков 2 типа нами была обнаружена характерная скачкообразная динамика осмотического давления межклеточного вещества, обусловленная скачкообразной динамикой концентрации глюкозы в межклеточном веществе, с инсулинорезистентностью ткани: у диабетиков 2 типа с высокой инсулинорезистентностью наблюдаются аномально низкие значения величины энтальпии ткани, соответствующие преобладанию кристаллической фазы в межклеточном веществе.
Рис. 49. Характерные мониторинговые кривые осмотического давления межклеточного вещества здорового пациента и пациента с сахарным диабетом 2 типа. Временная динамика осмотического давления у пациента с сахарным диабетом имеет скачкообразный характер, обусловленный скачкообразной динамикой концентрации глюкозы в межклеточном веществе и в крови, в отличии от временной динамики здорового пациента, характеризующейся плавными монотонными изменениями, обусловленными плавными монотонными изменениями концентрации глюкозы в межклеточной жидкости и крови.
Следствием скачкообразной динамики осмотического давления межклеточного вещества, имеющим важное значение для практической медицины, является скачкообразный характер кровяного давления. Метод мониторинга осмотического давления применим для ранней диагностики сердечно сосудистых заболеваний, в частности, гипертонии.
Метод изотермической калориметрической спектроскопии, описанный выше, также позволяет определить на ранней стадии диабет и сердечно-сосудистые заболевания, связанные с сахарным диабетом, на основе спектральных измерений содержания гиалуроновой кислоты в локальном объеме живой ткани.
Способ измерения скорости метаболизма и оценка энергетической ценности принимаемой пищи.
Эпидемиологические исследования, проведенные в разных странах, указывают на тесную связь между НТГ и ожирением. Например, в исследовании, проведенном в США, было обнаружено, что средний ИМТ (избыточная масса тела) у лиц, у которых впоследствии развилось НТГ, был достоверно выше, чем у лиц с нормальным ИМТ. В исследовании, проведенном в Израиле, установили, что высокий ИМТ в анамнезе сопровождался повышенной частотой развития НТГ за период 10 лет.
Метод изотермической калориметрической спектроскопии, описанный выше, позволяет определить энергетическую ценность принимаемой пищи на основе спектральных измерений содержания глюкозы и триглицерида свободных жирных кислот в локальном объеме живой ткани. Энергетическая ценность или калорийность - это количество энергии, высвобождаемой в организме человека из продуктов питания в процессе пищеварения, при условии ее полного усвоения. Энергетическая ценность продукта измеряется в килокалориях (ккал) или килоджоулях (кДж) в расчете на 100 г продукта. Килокалория, используемая для измерения энергетической ценности продуктов питания, также носит название «пищевая калория», поэтому, при указании калорийности в килокалориях приставку «кило» часто опускают. Пищевая ценность продукта - это содержание в нем углеводов, жиров и белков из расчета на 100 грамм продукта.
Способ диагностики состояния кожи.
Метод изотермической калориметрии и спектроскопии, описанный выше, позволяет определить физиологическое состояние кожи на основе спектральных измерений содержания гиалуроновой кислоты и триглицерида свободных жирных кислот (и других биохимических компонентов) в локальном объеме живой ткани.
Примеры практического использования. Результаты клинического исследования.
Внешний вид прототипа изотермического калориметра живой ткани, с помощью которого были выполнены клинические исследования с целью подтверждения метода (proof of concept study) показан на рис. 44.
Оценка эффективности метода выполнена двумя разными методами, описанными в протоколе клинического исследования. Группа из 41 пациентов с сахарным диабетом (1 и 2 типа) для выполнения задачи исследования была разбита на две подгруппы из 27 пациентов (подгруппа 1) и 14 пациентов (подгруппа 2), в каждую из которых входят пациенты 1 и 2 типа. Выборка пациентов случайная.
Целью исследования с участием 27 пациентов из первой подгруппы было подтверждение эффективности метода, изучение воспроизводимости результатов тестов и оценка возможности индивидуальной калибровки. С каждым из пациентов было проведено 5-6 тестов продолжительностью 35 минут (с тремя инвазивными измерениями), в среднем 3 теста в течение дня. Всего на 27 пациентах с диабетом 1 и 2 типа было проведено 144 теста с суммарным числом 434 инвазивных измерения.
На Рис. 50 (а-д) показаны примеры сахарных кривых со скачкообразной временной динамикой: Рис. 50а (тест №6 пациент №24), Рис. 50б (тест №1 пациент №41), Рис. 50в (тест №5 пациент №41), Рис. 50г (тест №6 пациент №41). На всех графиках результаты инвазивных измерений показаны кружочками зеленого цвета, а результаты неинвазивного мониторинга фиолетовой кривой.
Показателем эффективности метода в первом методе оценки является корреляция, в интервале погрешности 15%, результатов неинвазивных измерений с результатами инвазивных измерений, при фиксированных значениях индивидуального масштабного коэффициента данного пациента (индивидуальная калибровка).
В рамках исследования было показано, что каждый пациент с сахарным диабетом характеризуется двумя значениями индивидуального масштабного коэффициента К01 и К02, которые имеют постоянное индивидуальное значение для данного пациента; однако, при этом, масштабный коэффициент изменяется в широком диапазоне значений от пациента к пациенту от К01=0,7 (для пациента №1) до К01=480 (для пациента №25).
В соответствии с международным стандартом ISO 15197 (раздел 6.3.3. Minimum system accuracy performance criteria; Data analysis of clinical study according to Clause 6.3.3 of ISO 15197:2013 [p 12, pdf]) приняты следующие критерии оценки точности инвазивных глюкометров:
95% результатов измерений должны попадать в зону А, границы которой определяются из следующих условий:
± 15% при значениях сахара, равных или превышающих 5,55 ммоль/литр;
± 0,83 ммоль/литр при значениях, меньших 5,55 ммоль/литр;
Целью исследования с участием 14 пациентов из второй подгруппы была оценка эффективности метода при измерении слепым методом. С каждым из пациентов было проведено 6 тестов продолжительностью 35 минут (с тремя инвазивными измерениями) в течение двух дней, из которых результаты первых 3 тестов (1, 2, 3) были использованы для калибровки. Для каждого испытуемого проводилась индивидуальная калибровка, в процессе которого по результатам первых 3 тестов (продолжительностью 35 минут) определялись значения индивидуальных масштабных коэффициентов данного пациента К01 и К02.
Следующие 3 теста (4, 5, 6) на следующий день проводились и обрабатывались слепым методом. Всего на 14 пациентах с сахарным диабетом 1 и 2 типа было проведено 83 теста с суммарным количеством инвазивных измерений 249. Результаты тестов представлены на Рис. 51, 52. Результаты тестов подгруппы 2: 14 пациентов с диабетом 1 и 2 типа; 83 теста с суммарным количеством 249 инвазивных измерений. Кривые красного цвета ограничивают зону А. В зону А попадают 246 измерений из 249 или 98% результатов.
Таким образом, результаты сравнительных измерений, полученные на пациентах подгруппы 2 полуслепым и слепым методом, находятся в соответствии с требованиями стандарта ISO 15197 по точности. Результаты тестов, выполненных на группе из 41 пациентов (227 тестов, 683 инвазивных измерения), полученные путем объединения результатов подгруппы 1 и подгруппы 2, представлены на Рис. 53.
Результаты тестов объединенной группы: 42 пациентов с диабетом 1 и 2 типа; 227 тестов с суммарным количеством 683 инвазивных измерений. Кривые ограничивают зону А, зону В (±25%) (не показана на чертежах).
В зону А + В попадают всего 654 измерений из 664 или 98,3% результатов.
Таким образом, результаты клинического исследования, выполненного на группе из 41 пациента с диабетом 1 и 2 типа, находятся в соответствии с требованиями стандарта ISO 15197 по точности.
Проблема фолдинга белка.
Фолдинг белка - процесс пространственной упаковки белковой молекулы, принятия белком строго определенной формы, в которой он выполняет свои функции; процесс самоорганизации белковой молекулы - самопроизвольное сворачивание полимерной цепи при определенных условиях в определенную пространственную структуру, схематически показан на Рис. 1.
Способность природной полипептидной цепи к пространственной самоорганизации и обретению определенной молекулярной структуры, являющейся самой яркой особенностью белков. Процесс самоорганизации белков во многом пока неясен и представляет одну из крупнейших проблем современной науки. Известно, что во многом функции белков можно предсказать по их структуре, хоть и не всегда. Поэтому актуальной научной проблемой является предсказание структуры и, как следствие функции белка, по последовательности аминокислот в полимерной цепи.
Молекулы белков представляют собой линейные полимеры, состоящие из остатков аминокислот (которые являются мономерами), также в состав белков могут входить модифицированные аминокислотные остатки и компоненты не аминокислотной природы, например гликопротеины, характеризующиеся низким содержанием углеводов, углеводный остаток (олигосахарид, имеющий нерегулярную структуру и содержащий глюкозу, маннозу, галактозу и их аминопроизводные, а также N-ацетилнейраминовую кислоту) присоединен к аминокислотам белковой цепи либо N-гликозидной связью - к амидному азоту аспарагина, либо О-гликозидной связью - к гидроксил группе остатков серина, треонина, гидроксилизина. При образовании белка в результате взаимодействия α-карбоксильной группы (-СООН) одной аминокислоты с α-аминогруппой (-NH2) другой аминокислоты образуются пептидные связи. Концы белка называют N- и С-концом, в зависимости от того, какая из групп концевого аминокислотного остатка свободна: -NH2 или -СООН, соответственно.
Как известно, молекула глюкозы может образовать гликозидную валентную связь также с аминокислотами белковых молекул. В гликопротеинах, характеризующихся низким содержанием углеводов, углеводный остаток (олигосахарид, имеющий нерегулярную структуру и содержащий глюкозу, маннозу, галактозу и их аминопроизводные, а также N-ацетилнейраминовую кислоту) присоединен к аминокислотам белковой цепи либо N-гликозидной связью - к амидному азоту аспарагина, либо О-гликозидной связью - к гидроксил группе остатков серина, треонина, гидроксилизина. Структурные формулы гликозидной связи с аминокислотами показаны на Рис. 60.
Как показано, конфигурация тетрамера в биополимерной молекуле гиалуроновой кислоты образуется с участием ионов натрия, концентрация которых в межклеточном пространстве много больше концентрации ионов калия.
С другой стороны, как известно, ионный состав клеток, до тех пор, пока они проявляют свойства живого, отличается от ионного состава окружающей среды. Наиболее существенным отличием является асимметричное распределение одновалентных ионов Na+ и K+: клетки активно накапливают K и выбрасывают в окружающую среду Na. Так создается разность концентраций одновалентных катионов на клеточной мембране.
Отсюда следует понимание вывода, что ионом одновалентного металла М, образующим ионную связь с заряженным звеном полимерной цепи -Rb-M белковых молекул, локализованных в плазматической мембране и цитоплазме клетки, является одновалентный ион K+:
-Rb-M→-Rb-K
Универсальная модель биополимерной молекулы, на основе которой описаны свойства макромолекулы гиалуроновой кислоты, которая как было отмечено, является уникальным модельным объектом для исследования свойств биополимерных молекул, позволяет предсказать на основе точного статистического описания термодинамические свойства произвольной природной макромолекулы, в частности макромолекул белков и ферментов.
В общем виде, структуру биополимерной молекулы белка, способной к пространственной самоорганизации и обретению устойчивой молекулярной гетерофазной структуры, обусловленной слабыми объемными взаимодействиями не валентной природы, можно описать следующим определением: линейная полипептидная цепь из последовательно соединенных амидной (пептидной) связью аминокислот -NH-CH(R)-CO-, повторяющийся мономер которого состоит из двух (или более) типов мономерных звеньев - аминокислот ARa и ARb, с отличающимися боковыми остатками Ra, Rb, один из которых (например, звено ARb) содержит заряженную (отрицательно или положительно) группу Rb, а боковой остаток Ra второго звена (ARa) является незаряженным. В состав растворителя, основным компонентом которого является вода, входят как минимум 3 компоненты: один компонент - молекула (мономерная) субстрата (растворителя) S, например молекула глюкозы, и два (или несколько) низкомолекулярных одновалентных ионов M1 и М2, например ионы натрия и калия, обладающих разным сродством к мономеру, электрический заряд которых противоположен заряду бокового остатка ARb; мономерная молекула растворителя (субстрата) S может образовать две водородные связи с мономерными звеньями полимерной цепи - Ra-S-Rb-; -Ra-S-Rb-; -Ra-S-; -S-Rb; низкомолекулярные одновалентные ионы M1 и М2 могут образовать ионную связь с заряженными звеньями полимерной цепи -Rb-M1. Энергия продольных ковалентных связей Е0 соседних звеньев полимерной цепи много больше характерной тепловой энергии, при физиологических температурах Е0/kBT>>1. Энергия поперечных водородных связей звеньев Ra и Rb с молекулой мономера (субстрата) S растворителя много меньше энергии продольных валентных связей Es<<Е0 полимерной цепи. Низкомолекулярный ион M1 может адсорбироваться мономером полимерной цепи Rb, с образованием слабой ионной связи -Rb-M1.
В общем случае, повторяющийся фрагмент полимерной цепи может состоять из одинаковых пар -А- В- или нескольких типов повторяющихся пар мономерных звеньев -А-В-, -С-В-, …, обладающих способность образовать водородные связи с молекулами субстрата.
Водородные связи, определяющие наиболее вероятную конфигурацию связанной пары элементарных цепочек, соответствующую минимальной свободной энергии объемных взаимодействий, и как следствие наиболее вероятную пространственную конфигурацию тетрамера, образованного парой не соседних мономеров полимерной цепи и парой молекул растворителя с участием ионов, можно определить как сопряженные не валентные связи или сопряженные водородные связи биополимерной молекулы. Элементарная ячейка, соответствующая связанной паре элементарных цепочек (в ее наиболее вероятной конфигурации), образует сопряженную систему - замкнутую систему сопряженных водородных связей с делокализованной электронной плотностью; как известно, общая особенность всех сопряженных систем - растекание электронной плотности водородных связей по всей сопряженной системе.
Структурные формулы аминокислот, образующих полипептидные цепи белковых молекул, показаны в таблице на рисунке 54.
Как известно, магний функционирует в качестве кофактора в более чем 300 известных ферментативных реакций, включенных в широкий спектр метаболической активности. Выработка энергии, метаболизм глюкозы, окисление жирных кислот и активизация аминокислот - везде требуется магний. Магний - один из важнейших минералов, являясь основным внутриклеточным элементом, активизирует ферменты, регулирующие углеводный обмен, стимулирует образование белков, регулирует хранение и высвобождение энергии в АТФ, снижает возбуждение в нервных клетках, расслабляет сердечную мышцу.
Как было показано, тетрамер в объеме макромолекулы, полимерная цепь которой содержит отрицательно заряженные группы, может образоваться только с участием одновалентного иона (Na, K). Двух валентный ион металла (Mg2+, Mn2+) не может участвовать в образовании тетрамера - сшивка из 2 не соседних мономеров полимерной цепи, 2 молекул субстрата и 2 одновалентных катионов растворителя, однако может образовать сшивку между двумя не соседними мономерами полимерной цепи без участия молекул растворителя. На рисунке 62 схематично показана сшивка, образованная 2 валентным ионом магния между не соседними мономерами полимерной цепи, посредством ионной связи.
Общепринятая классификация. В соответствии с общепринятой классификацией, выделяют 4 уровня структурной организации белков: первичную, вторичную, третичную и четвертичную структуры белка. Первичная структура (последовательность аминокислотных остатков) полипептида определяется структурой его гена или генетическим кодом, а структуры более высоких порядков формируются в процессе сворачивания белка. Хотя пространственная структура белка в целом определяется его аминокислотной последовательностью, она является довольно лабильной и может зависеть от внешних условий, поэтому более правильно говорить о предпочтительной или наиболее энергетически выгодной конформации белка.
Первичная структура - последовательность аминокислотных остатков в полипептидной цепи, которую, как правило, описывают, используя однобуквенные или трехбуквенные обозначения для аминокислотных остатков. Важными особенностями первичной структуры являются консервативные мотивы - устойчивые сочетания аминокислотных остатков, выполняющие определенную функцию и встречающиеся во многих белках. Консервативные мотивы сохраняются в процессе эволюции видов, по ним часто удается предсказать функцию неизвестного белка.
Вторичная структура - локальное упорядочивание фрагмента полипептидной цепи, стабилизированное водородными связями. Ниже приведены некоторые самые распространенные типы вторичной структуры белков:
α - спирали - плотные витки вокруг длинной оси молекулы. Один виток составляет 3,6 аминокислотных остатка, шаг спирали равен 0,54 нм (на один аминокислотный остаток приходится 0,15 нм). Спираль стабилизирована водородными связями между Н и О пептидных групп, отстоящих друг от друга на 4 звена;
β - листы (складчатые слои) - несколько зигзагообразных полипептидных цепей, в которых водородные связи образуются между относительно удаленными друг от друга (0,34 нм на аминокислотный остаток) аминокислотами в первичной структуре или разными цепями белка (а не близко расположенными, как имеет место быть в α - спирали).
Третичная структура - пространственное строение полипептидной цепи, образуется при свертывании полипептидной в компактную трехмерную систему (в случае ферментов это, как правило, сферическая глобула). Структурно состоит из элементов вторичной структуры, стабилизированных различными типами взаимодействий, в которых гидрофобные взаимодействия играют важнейшую роль.
Как показано выше, объемные взаимодействия простой полимерной цепи с растворителем можно описать, как парное объемное взаимодействие элементарных цепочек, образующих связанную пару, наиболее вероятная конфигурация которой показана на Рис. 5а. Наиболее вероятная конфигурация взаимодействующих элементарных цепочек реализуется в результате спаривания цепочек посредством водородных связей в конфигурации 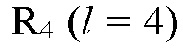 тетрамера; каждая элементарная цепочка находится в своей наиболее вероятной линейной конфигурации с максимальной энтропией, соответствующей минимуму свободной энергии взаимодействия цепочки с растворителем.
тетрамера; каждая элементарная цепочка находится в своей наиболее вероятной линейной конфигурации с максимальной энтропией, соответствующей минимуму свободной энергии взаимодействия цепочки с растворителем.
Исследования принципов укладки белков показали, что между уровнем вторичной структуры и атомарной пространственной структурой удобно выделять еще один уровень - мотив укладки (архитектура, структурный мотив). Мотив укладки определяется взаимным расположением элементов вторичной структуры (α-спиралей и β-тяжей) в пределах белкового домена - компактной глобулы, которая может существовать или сама по себе или входить в состав более крупного белка наряду с другими доменами.
Четвертичная структура (или субъединичная, доменная) - взаимное расположение нескольких полипептидных цепей в составе единого белкового комплекса. Белковые молекулы, входящие в состав белка с четвертичной структурой, образуются на рибосомах по отдельности и лишь после окончания синтеза образуют общую надмолекулярную структуру. В состав белка с четвертичной структурой могут входить как идентичные, так и различающиеся полипептидные цепочки. В стабилизации четвертичной структуры принимают участие те же типы взаимодействий, что и в стабилизации третичной. Надмолекулярные белковые комплексы могут состоять из десятков молекул.
Домен белка - элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, фолдинг которой проходит независимо от остальных частей; третичная структура белковой молекулы образуется при свертывании полипептидной в компактную трехмерную систему (в случае ферментов это, как правило, сферическая глобула).
В состав домена обычно входит несколько элементов вторичной структуры. Сходные по структуре домены встречаются не только в родственных белках (например, в гемоглобинах разных животных), но и в совершенно разных белках. Достаточно часто доменам присваивают отдельные названия, так как их присутствие непосредственно влияет на выполняемые белком биологической функции - к примеру, Са2+-связывающий домен кальдомулина, гомеодомен, отвечающий за связывание с ДНК в различных факторах транскрипции и др.
В олигомерных белках, каждая из полипептидных цепей, образующих субъединицу, характеризуется свой вторичной и третичной структурой. Однако у этих белков есть еще один конформационный уровень, называемый четвертичной структурой; под этим термином понимают расположение полипептидных цепей, входящих в состав отдельных полипептидных цепей, относительно друг друга, т.е. способ их совместной укладки и упаковки с образованием нативной конформации олигомерного белка.
Так как домены достаточно «автономны» в формировании своей структуры и выполнении своей функции, с помощью генной инженерии можно «пришить» к одному из белков домен, принадлежащий другому (создав таким образом белок - химеру). Такая химера при удаче будет совмещать функции обоих белков. Так, например, путем слияния ДНК-связывающего домена Cas9 с различными регуляторными доменами удалось получить искусственные факторы транскрипции (crisprTF), избирательно направляемые на нужные участки генома с помощью изготовленных на заказ «РНК-гидов». С помощью Cas9-домена можно также сконструировать искусственные эндонуклеазы рестрикции, репрессоры, ферменты, модифицирующие эпигеном, такие как ДНК-метилазы и деметилазы.
Метод расчета пространственной структуры белка. Наиболее вероятная линейная конфигурация элементарной цепочки отличается от конфигурации первичной структуры полимерной цепи и образуется в результате взаимодействия полимерной цепи с растворителем в равновесной конфигурации макромолекулы, соответствующей минимальной свободной энергии объемных взаимодействий. С другой стороны, объем вещества в конденсированном состоянии, при заданных внешних условиях (давлении, температуре и других характеристиках), определяется исключительно силами объемных взаимодействий, поэтому трехмерную пространственную конфигурацию биополимерной молекулы можно рассчитать на основе конфигурации связанной пары элементарных цепочек, характеризующей равновесную конфигурацию элементарного объемного взаимодействия.
В случае биополимерной молекулы с простой полимерной цепью, не содержащей субъединиц (протомеров), фолдинг белковой молекулы - процесс формирования вторичной структуры, с пространственной укладкой в компактную глобулу, происходит самопроизвольно, как фазовый переход статистической системы многих взаимодействующих частиц в пространственную конфигурацию с гетерофазной структурой с минимальной свободной энергией объемных взаимодействий. Внутримолекулярную гетерофазную структуру и объем домена белковой молекулы с простой цепью с одной субъединицей можно рассчитать, как показано, на основе конфигурации связанной пары элементарных цепочек взаимодействия.
В общем случае, цепь биополимерной молекулы (например, олигомерный белок) может иметь более сложную первичную структуру, состоящую из нескольких соединенных между собой простых цепей (субъединиц, протомеров). Олигомерные белки - белки, содержащие 2 и большее число полипептидных цепей (до нескольких тысяч субъединиц). В олигомерных белках каждая из полипептидных цепей, образующая субъединицу (протомер), характеризуется своей вторичной и третичной пространственной структурой. Примеры биополимерных молекул, состоящих из нескольких субъединиц, показаны на рисунке 55.
Полипептидные цепи в олигомерных белках могут быть либо одинаковыми, либо разными. Число полипептидных цепей в олигомерном белке можно установить по числу аминоконцевых остатков, приходящихся на одну молекулу белка. Олигомерный белок, состоящий из четырех полипептидных цепей, скажем гемоглобин, должен иметь четыре концевых остатка, по одному на каждую цепь. В молекуле инсулина имеется две цепи, связанные друг с другом ковалентными поперечными связями.
Некоторые белки (сложные белки) кроме полипептидной части содержат еще и небелковую - простетическую группу, которая может быть связана с белковой частью как ковалентной, так и слабой нековалентной связью. На рис. 55 представлена четвертичная структура - взаимное расположение субъединиц белка в пространстве, характерна для белков, состоящих из нескольких полипептидных цепей; возникает в результате ассоциации нескольких субъединиц белка в пространстве. В олигомерных белках, каждая из полипептидных цепей, образующих субъединицу, характеризуется свой вторичной и третичной структурой.
Взаимодействие между субъединицами макромолекулы.
Как известно из опыта, трехмерная пространственная структура сложных белков, состоящих из нескольких субъединиц, может изменяться в процессе взаимодействия полимерных цепей, принадлежащих разным субъединицам, с молекулами растворителя. На Рис. 56 показана схема, поясняющая эффект изменения конформации молекулы фермента гексокиназы в процессе связывания молекулы глюкозы.
Объемное взаимодействие между соседними доменами возникает в результате объемных взаимодействий между мономерами (принадлежащими участкам цепей), принадлежащими разным субъединицам: молекула субстрата образует водородную связь с двумя мономерами, которые принадлежат разным субъединицам и имеют разные знаки электрического заряда. Изменение трехмерной пространственной структуры белковой молекулы гексокиназы, состоящей из двух доменов, возникает в результате фазового перехода вещества в локальной области между соседними доменами (соответствующими субъединицам) в гетерофазное конденсированное состояние. Статистический механизм перехода вещества в области между соседними доменами в гетерофазное состояние описан выше в разделе Физика конденсированного состояния полимерной молекулы.
Взаимодействие между биомолекулами.
В общем случае, как известно из опыта, биополимерные молекулы могут взаимодействовать не только с малыми молекулами, но могут образовать комплексы с биомолекулами (чаще всего белком, например клеточным рецептором, но иногда, например, с ДНК) и производить, вследствие такого связывания, те или иные биохимические, физиологические или фармакологические эффекты. Химическое соединение (которое часто, но не всегда, малая молекула), которое образует комплекс с той или иной биомолекулой в биохимии и фармакологии называется лиганд. В случае связывания лиганда с белком лиганд обычно является малой сигнальной молекулой, связывающейся со специфическим участком_связывания на белке-мишени (например, на рецепторе). В случае связывания лиганда с ДНК лиганд обычно также является малой молекулой или ионом, или белком, который связывается с двойной спиралью ДНК. Связывание или ассоциация лиганда с рецептором (так называемый «докинг» лиганда в специфическую «нишу» в рецепторе) обычно обратима и кратковременна. Обратный процесс называется диссоциацией лиганда из связи с рецептором.
Исследования биополимерных комплексов, возникающих в результате ассоциации нескольких макромолекул, связанных не валентными связями, привели к формированию на рубеже 80-90-х годов новой междисциплинарной области знаний, получившая название "супрамолекулярная химия" - это химия межмолекулярных связей, изучающая ассоциацию двух и более химических молекул, а также структуру подобных ассоциатов. Она лежит за пределами классической химии, исследующей структуру, свойства и превращения отдельных молекул. Если последняя имеет дело главным образом с реакциями, в которых происходит разрыв и образование валентных связей, то объектами изучения супрамолекулярной химии служат не валентные взаимодействия - водородная связь, электростатические взаимодействия, гидрофобные силы, структуры "без связи".
Простейший пример супрамолекулярных структур - это комплексы типа «хозяин-гость». Хозяином (рецептором) обычно выступает большая органическая молекула с полостью в центре, а гостем - более простая молекула или ион. Например, циклические полиэфиры различного размера (краун-эфиры) довольно прочно связывают ионы щелочных металлов.
Для супрамолекулярных структур характерны следующие свойства: 1. наличие не одного, а нескольких связывающих центров у хозяина (в краун-эфирах эту роль выполняют атомы кислорода, обладающие не поделенными электронными парами); 2. комплементарность: геометрические структуры и электронные свойства хозяина и гостя взаимно дополняют друг друга (в краун-эфирах это проявляется в том, что диаметр полости должен соответствовать радиусу иона). Комплементарность позволяет хозяину осуществлять селективное связывание гостей строго определенной структуры. В супрамолекулярной химии это явление называют «молекулярным распознаванием» (англ. - molecular recognition). 3. Комплексы с большим числом связей между комплементарными хозяином и гостем обладают высокой структурной организацией.
Примером супрамолекулярной биохимической структуры являются ферменты - молекулы-хозяева. Активный центр каждого фермента устроен таким образом, что в него может попасть только то вещество (субстрат), которое соответствует ему по размерам и энергии; с другими субстратами фермент реагировать не будет. Другим примером супрамолекулярных биохимических структур служат молекулы ДНК, в которых две полинуклеотидные цепи комплементарно связаны друг с другом посредством множества водородных связей. Каждая цепь является одновременно и гостем, и хозяином для другой цепи.
Физика гетерофазного конденсированного состояния биополимерных молекул, описанная выше, позволяет взаимодействие молекул в супрамолекулярных комплексах. Взаимодействие между макромолекулами с образованием супрамолекулярных структур, происходит по механизму, аналогичному механизму взаимодействия между субъединицами олигомерного белка, в результате образования водородных связей между молекулой лиганда и белковой молекулой и ионных связей между ионами растворителя и заряженными участками мономеров полимерной цепи.
Метод определения пространственной структуры сложной макромолекулы.
Трехмерная доменная пространственная структура белка рассчитывается с помощью метода, описанного выше, и состоит из следующих шагов:
- определяют число субъединиц по числу аминоконцевых остатков;
- определяют первичную структуру полипептидной цепи каждой субъединицы (протомера);
- определяют структуру простетической группы;
- определяют структуру элементарной цепочки взаимодействия для каждой субъединицы; для этого определяют ион - молекулярную пару растворителя "молекула субстрата - одновалентный ион" для каждой субъединицы полимерной цепи;
- определяют третичную структуру каждого независимого домена; для этого для каждой субъединицы вычисляют константу равновесия, энергию активации и определяют гетерофазную структуру и объем независимого белкового домена (объем локализации субъединицы полимерной цепи);
- для каждой субъединицы определяют расположение в цепи мономеров, определяющих взаимодействие между субъединицами;
- определяют структуру связанной пары элементарных цепочек, характеризующих взаимодействие между субъединицами.
- трехмерную пространственную конфигурацию (четвертичную структуру) определяют как наиболее вероятную пространственную конфигурацию независимых доменов, соответствующую минимальной свободной энергии объемных взаимодействий в локальной области между доменами.
Гетерофазный-биологический катализ.
Известным эффективным способом ускорения протекания химической реакции неорганических молекул, который широко применяется на практике, является гетерофазный катализ - изменение скорости химической реакции при воздействии катализаторов, образующих самостоятельную фазу и отделенных от реагирующих веществ границей раздела. Например, наиболее распространенный случай, когда твердый катализатор (контакт) ускоряет реакцию между газообразными реагентами или реакцию в растворе. Каталитическая реакция протекает обычно на поверхности твердого катализатора и обусловлена активацией молекул реагентов при взаимодействии с поверхностью. Поэтому для осуществления гетерофазного катализа необходима адсорбция компонентов реакционной смеси из объемной фазы на поверхности катализатора.
Химическими катализаторами, ускоряющими жизненно важные биохимические реакции (около десяти тысяч), протекающие в клетке, являются ферменты - белковые молекулы, полимерная (полипептидная) цепь которых состоит из аминокислот.
Основными признаками ферментативного катализа, отличающими его от лабораторного и производственного катализа, являются строгая избирательность и высокая скорость химической реакции.
До недавнего времени считалось, что обязательным компонентом всех ферментов являются белки. Однако были открыты и стали объектом интенсивных исследований ферменты, построенные из молекул рибонуклеиновых кислот (рибозимы). Интерес к этой группе ферментов резко усилился в связи с разработкой методов молекулярной селекции нуклеиновых кислот, позволившей, в частности, начать направленное конструирование рибозимов с разнообразными типами каталитической активности.
Согласно современным представлениям (ИЮПАК), специфичность и каталитическая активность молекулы фермента определяется активным центром - особым участком молекулы с уникальной комбинацией аминокислотных остатков (и возможно небелковых групп), обеспечивающих непосредственное связывание молекулы фермента с молекулой субстрата и прямое участие в катализе. Процесс связывания молекулы субстрата активным центром фермента сопровождается изменением пространственной структуры ферментной молекулы. Активный центр фермента, как правило, имеет форму узкого углубления или щели, возникающей между соседними доменами. Схема, поясняющая эффект изменения конформации молекулы фермента гексокиназы в процессе связывания молекулы глюкозы показана на Рис. 56.
Наиболее часто в состав активных центров ферментов входят функциональные группы таких аминокислот:
ОН - группы серина, треонина, тирозина;
SH - группы цистеина;
NH - группа гистидина;
СООН - группа глутамата и аспартата;
NH2 - группы аргинина и лизина.
Физическая природа явления биологического катализа в настоящее время неизвестна и относится к числу нерешенных проблем современной науки.
Физика гетерофазного конденсированного состояния биополимерной молекулы, основные положения которой описаны выше, позволяет объяснить и описать на основе точного статистического метода основные свойства биологического катализа - высокую скорость химической реакции и строгую избирательность.
биополимерная молекула с гетерофазной структурой, обладающая избирательностью к молекуле G субстрата, способна катализировать биохимическую реакцию с участием этой молекулы. Ускорение биохимической реакции происходит за счет сближения молекул в кристаллической фазе биополимерной молекулы с гетерофазной структурой. В объеме элементарной ячейки биополимерной молекулы с гетерофазной структурой, как было показано, 1/3 от общего числа молекул субстрата находится в свободном состоянии в растворителе, а оставшиеся 2/3 молекул субстрата находятся в связанном состоянии с мономерами полимерной цепи фермента. Концентрации всех компонентов системы в конденсированной кристаллической (глобулярной) фазе увеличиваются кратно значению параметра порядка, увеличение которого приводит к многократному возрастанию числа эффективных столкновений за счет эффекта сближения.
Скорость гетерофазного биологического катализа.
Рассмотрим физико-химический механизм биологического катализа, на примере фермента гексокиназа и его изомера глюкокиназа, играющих важную роль в биохимической регуляции метаболизма глюкозы в клетке живой ткани.
Ферментативная реакция первой стадии гликолиза, которая катализируется обоими этими ферментами (гексокиназой и глюкокиназой), имеет вид:
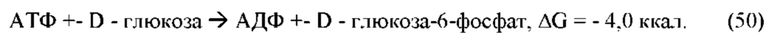
Скорость химической реакции, в общем случае, определяется законом действующих масс (Гульдберг и Вааге) - скорость химической реакции А+В → С, с образованием нового вещества С из двух веществ А иВ, пропорциональна произведению концентраций реагирующих веществ (при прочих одинаковых условиях):

где k - константа скорости, которая не зависит от концентраций А и В.
Кинетическую схему реакции (50), в процессе которой происходит перенос фосфатной группы Р от молекулы АТФ к молекуле глюкозы G (которая не может протекать самопроизвольно без катализатора) можно представить в виде следующей ферментативной реакции:
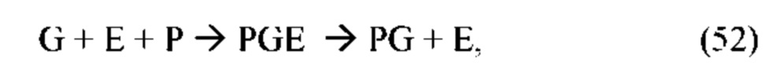
где Е - мономер полимерной цепи, который может образовать слабую ионную связь с одновалентным ионом М растворителя или водородную связь с молекулой субстрата G растворителя.
Ферментативная реакция (51), в рамках рассматриваемого механизма, протекает в несколько этапов в активном центре ферментной молекулы - локальной области внутримолекулярного пространства между доменами, соответствующими субъединицам макромолекулы.
На первом этапе, схематически показанном на Рис. 57а, происходит диффузия молекул АТФ (показана только фосфатная группа АТФ красным) и глюкозы G из растворителя в область активного центра фермента (область газообразной фазы гетерогенной ферментной молекулы с низкой концентрацией субстрата).
На втором этапе (Рис. 57б), происходит абсорбция молекулы АТФ комплексом тетрамера в кристаллической фазе:

Комплекс тетрамера можно определить, как субстрат - ферментный комплекс, который для рассматриваемой реакции можно обозначить символом E2G2, где G обозначает молекулу глюкозы.
Этот этап является обратимым и не сопровождается какими-либо химическими изменениями субстрата.
Переход фосфатной группы Р от молекулы АТФ к молекуле глюкозы G, которая находится в связанном состоянии в комплексе тетрамера E2G2 в кристаллической (глобулярной) фазе (Рис. 44б), происходит в процессе химической реакции, которую схематично можно представить в виде:

На третьем этапе, схематически показанном на Рис. 57 d,g происходит превращение комплекса PGE в продукт GP в результате конформационного перехода тетрамера, который сопровождается переходом молекулы GP через энергетический барьер, разделяющий кристаллическую (глобулярная) фазу, с повышенной концентрацией субстрата, и жидкую (расплавленная глобула) фазу, с пониженной концентрацией субстрата, с последующей десорбцией продукта в результате разрушение комплекса тетрамера (переход из жидкой фазы в газообразную):
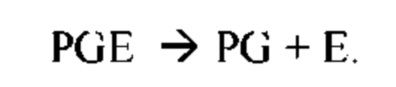
В рамках модели биополимерной молекулы с гетерофазной структурой, описанной выше, в образовании комплекса GE участвуют слабые не ковалентные взаимодействия - ионные и водородные связи, что согласуется с общепринятыми представлениями, в рамках которых предполагается, что в образовании фермент-субстратного комплекса (ФСК, комплекс Михаэлиса) принимают участие не ковалентные взаимодействия. На рис. 57 изображены основные этапы ферментативной реакции: а) диффузия молекулы АТФ из жидкой фазы в кристаллическую фазу; b) присоединение фосфатной группы (красный) к молекуле D- глюкозы (желтый) с образованием молекулы G6P в результате химической реакции в кристаллической фазе фермента; d) конформационный переход тетрамера со связанной молекулой G6P из кристаллической фазы в жидкую через энергетический барьер, разделяющий жидкую фазу с кристаллической; g) разрушение тетрамера в жидкой фазе с освобождением молекулы продукта G6P.
Скорость W ферментативной реакции (51) можно вычислить по формуле (50), если известна константа скорости; константу скорости k можно вычислить, как будет показано ниже, как отношение двух независящих от концентраций констант с помощью выражения: k=kb/К0,
где kb - скорость прохождения продукта PG через энергетический барьер, разделяющий кристаллическую фазу с жидкой; К0 - константа равновесия процесса образования тетрамера из 2 димеров; в этом случае, 1/K0 - константа диссоциации тетрамера с образованием 2 димеров. Здесь учитывается, что в момент достижения вершины энергетического барьера, на которой реализуется конфигурация R2 тетрамера, взаимное притяжение между димерами тетрамера определяется только вкладом внутренней энтропии, величина которой определяет величину константы диссоциации.
Формулу для константы скорости k можно вывести из следующих соображений.
Реагенты G (молекула глюкозы) и Е (мономер полимерной цепи) находятся в равновесии с димерами EG и тетрамерами E2G2. Константа KG диссоциации тетрамера с образованием 2 димеров определяется уравнением:
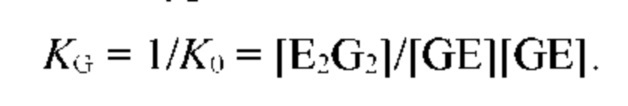
Реагенты GP и Е находятся в равновесии с активированным комплексом EGP (тетрамер-фосфатная группа):

Константа равновесия KGP реакции (53b) определяется уравнением:
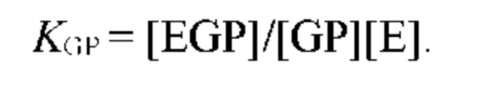
Константа равновесия реакции G+Р → GP, которая не может протекать самопроизвольно, равна константе равновесия ферментативной реакции (53d):

Отсюда следует, что уравнение (50) для скорости реакции можно решить относительно [GP]:
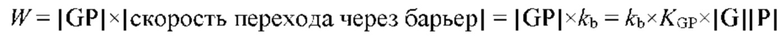 или
или
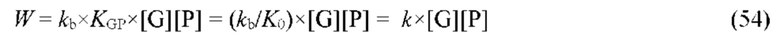
Скорость реакции (50) равна:
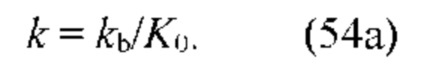
Константа равновесия К0 определяется, как показано, с помощью выражения (8):
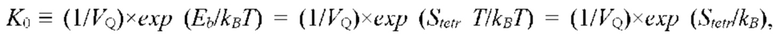 где Eb - энергия связи молекулы глюкозы с тетрамером, которая определяется внутренней энтропией тетрамера Stetr; VQ - квантовый объем молекулы глюкозы, который определяется с помощью выражения [14]:
где Eb - энергия связи молекулы глюкозы с тетрамером, которая определяется внутренней энтропией тетрамера Stetr; VQ - квантовый объем молекулы глюкозы, который определяется с помощью выражения [14]:
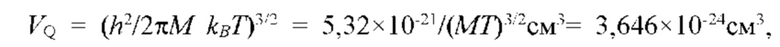 где М - молекулярный вес глюкозы (субстрата); h - постоянная Планка.
где М - молекулярный вес глюкозы (субстрата); h - постоянная Планка.
Скорость kb прохождения молекулы GP через энергетический барьер, разделяющий изомерные конформации тетрамера в жидкой и кристаллических фазах, определяется двумя факторами - высотой энергетического барьера Еа, величина которой определяет вероятность wb прохождения молекулы через барьер, и константой ν, имеющей размерность частоты, соответствующей частоте электромагнитного кванта с энергией kBT:
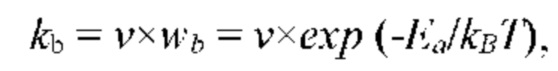 где ν=kBT/h; h - константа Планка.
где ν=kBT/h; h - константа Планка.
Водородная связь, как тип ковалентной валентной связи с делокализованной электронной плотностью, имеет электромагнитную природу. Энергия водородной связи определяется энергией парного взаимодействия молекулы глюкозы с мономером полимерной цепи, которая как показано, квантуется дискретной порцией энергии, кратной кванту тепловой энергии kBT. Частота колебаний ν валентной связи, в общем случае, определяется частотой электромагнитного кванта, которую можно вычислить из требования, что в равновесном состоянии квант электромагнитной энергии валентной связи hν равен кратному числу квантов тепловой энергии n×kBT, по отношению к которому устанавливается равновесие между электронной парой (осциллятором) и термостатом:
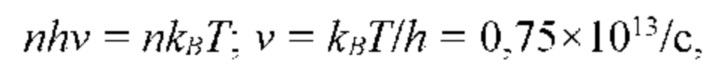 где N=1, 2, 3, … - кратность обменного кванта энергии электромагнитного взаимодействия по отношению к кванту тепловой энергии kBT.
где N=1, 2, 3, … - кратность обменного кванта энергии электромагнитного взаимодействия по отношению к кванту тепловой энергии kBT.
Выражение для константы скорости химической реакции имеет вид:
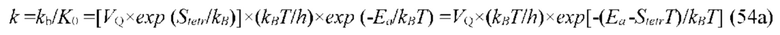
Выражение (54) для скорости реакции (50), таким образом, имеет следующий вид:
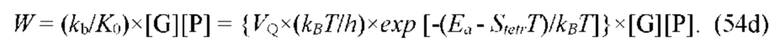
Формула (54d) для скорости реакции реагентов [G] и [Р] является универсальной, применимой для расчета скорости произвольной биохимической реакции, катализатором которой является биополимерная молекула с гетерофазной структурой с избирательностью к молекуле субстрата G.
Следует отметить, что формулу (54 а) можно привести к виду, совпадающему с формулой Аррениуса:
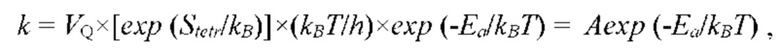
где 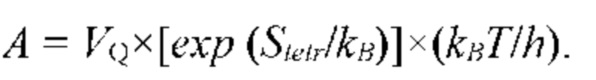
Расчет фермента глюкокиназы.
Термодинамические функции произвольного фермента, в частности, изомеров гексокиназы, можно получить с помощью универсальной функций распределения (14), на основе известных значений константы равновесия Михаэлиса для этих ферментов. Глюкокиназа (КФ 2.7.1.2) - изотип IV фермента гексокиназы (мономерный белок 465 аминокислот и молекулярная масса 50 кДа), в основном присутствующий в гепатоцитах, а также в клетках поджелудочной железы. Катализирует фосфорилирование только D-глюкозы, путем переноса фосфатной группы на глюкозу за счет АТФ
Глюкокиназа характеризуется константой равновесия Km ≈ 10 ммоль/л [31] и вызывает быстрое поглощение глюкозы из крови в печени в абсорбтивный период. Глюкоза при этом поступает в гепатоциты посредством транспортеров типа GLUT2, активность которых не регулируется уровнем инсулина, но сродство которых к глюкозе мало (Km ≈ 10 ммоль/л). Инсулин воздействует на усвоение глюкозы печенью лишь косвенно - он индуцирует синтез глюкокиназы в гепатоцитах. Таким образом, глюкокиназа призвана быстро перевести лишнюю глюкозу крови, возникшую после приема пищи, в гликоген.
Кривые насыщения фермента глюкокиназы, можно получить из универсальной функции распределения (14), в которой квантовое число для вычисления константы взаимодействия γ определяется из условия квантования свободной энергии взаимодействия (31), которое в общем случае можно представить в следующем виде:
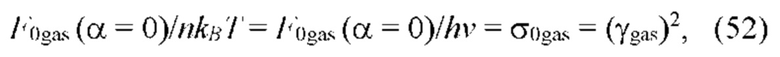
где hν - величина кванта энергии электромагнитного взаимодействия, кратная кванту тепловой энергии kBT; hν=n kB, где n=1, 2, 3, ….
Константе равновесия Km ≈ 10 ммоль/л удовлетворяет значение кванта энергии взаимодействия 3kBT, соответствующего значению квантового числа n=3. Кривые насыщения фермента глюкокиназы, полученные с помощью универсальной функции распределения биополимерной молекулы, показаны на Рис. 58а. Кривые насыщения и скорость ферментативной реакции глюкокиназы, в зависимости от концентрации глюкозы, приведены в единицах константы равновесия k0=5,032 ммоль/л (концентрация глюкозы в крови в норме); значению концентрации глюкозы kmgl=15,096 ммоль/л=3×kms2 на оси абсцисс соответствует значение безразмерной концентрации х=3.
Высоту энергетического барьера Eb=Emax - Emin, разделяющего кристаллическую и жидкую фазы в молекуле глюкокиназы, можно определить непосредственно из графика свободной энергии Uλsattr(α) тетрамера глюкокиназы от концентрации, представленного на Рис. 58б, который можно получить из функций распределения глюкокиназы, представленных на Рис. 58а. Высота энергетического барьера, разделяющего кристаллическую фазу от жидкой фазы, соответствующая энергии одной водородной связи, равна Emax-Emin=3,47×(3kBT)≈10,41 ккал/моль.
Высоту энергетического барьера Еа в единицах ε0=3kBT, можно определить из графика энергетического барьера, представленного в безразмерных единицах на Рис. 58б.
Значения энергии из графика:
Emax=0,1035…
Emin=- 0,188…
Е0=-0,160…
Emax-Emin=0,1035+0,188=0,2915.
Е0-Emin=-0,160+0,188=0,028=kBT.
Разница между энергией связи прямой и обратной реакции равна:
Emin-Е0=0,188-0,160=0,028=3kBT.
Высота энергетического барьера, разделяющего кристаллическую и жидкую фазу:
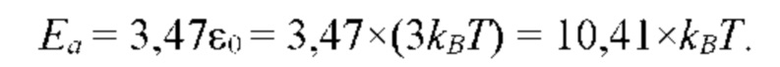
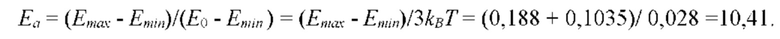
Для вычисления по формуле (54d) скорости ферментативной реакции (50), катализируемой глюкокиназой, при известной высоте энергетического барьера глюкокиназы Еа=3,47×(3kBT), необходимо определить константу диссоциации тетрамера с образованием 2 димеров, которую можно вычислить с помощью выражения:
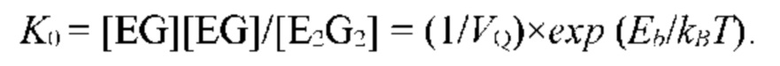
Здесь Eb - энергию связи, которую необходимо затратить для отрыва двух комплексов EG (молекула глюкозы G, связанная одной водородной связью с мономером Е), образующих комплекс тетрамера [E2G2] в конфигурации R2 с 2 водородными связями: EG+EG=E2G2.
Энергия связи Eb определяется внутренней энтропией тетрамера в конфигурации R2 с 2 водородными связями, которую можно вычислить с помощью выражения:
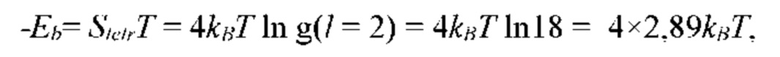
где Stetr=4kB ln 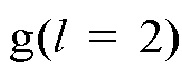 - внутренняя энтропия тетрамера; - число состояний, реализующих микроскопическую форму (конфигурацию) тетрамера R2 с 2 водородными связями. Множитель 4 в формуле соответствует числу водородных связей, возникающих в конфигурации тетрамера R2 в результате делокализации электронной плотности.
- внутренняя энтропия тетрамера; - число состояний, реализующих микроскопическую форму (конфигурацию) тетрамера R2 с 2 водородными связями. Множитель 4 в формуле соответствует числу водородных связей, возникающих в конфигурации тетрамера R2 в результате делокализации электронной плотности.
Откуда следует, что энергия связи, рассчитанная на одну водородную связь, равна:
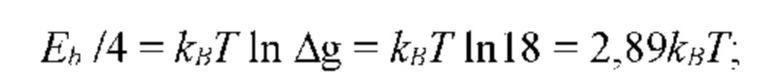
Следует отметить, что термодинамическое равновесие системы устанавливается по отношению к обмену энергией между системой и резервуаром с квантом тепловой энергии kBT, в отличие от электромагнитного равновесия взаимодействующих молекул, которое устанавливается по отношению к кванту электромагнитной энергии ε0=3,47×(3kBT).
Энергия связи Eb равна: 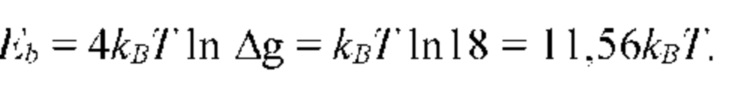
Численное выражение для константы равновесия реакции равно:
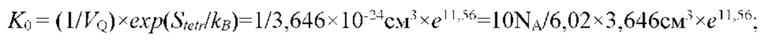
К0=4,385 ммоль/литр,
где NA=6,02×1023 - число Авогадро.
Расчет скорости ферментативной реакции глюкокиназы.
Скорость ферментативной реакции (50), катализируемой глюкокиназой, при известной высоте энергетического барьера глюкокиназы Еа=3,47×(3kBT) и константе диссоциации 1/К0, можно вычислить по формуле (54d).
Высота энергетического барьера Еа в расчете на одну водородную связь, в единицах кванта энергии взаимодействия ε0=3kBT, как показано, равна Еа=Emax-Emin=3,47 ε0.
Энергия активации в расчете на 4 связи с квантом энергии ε0=3kBT равна:
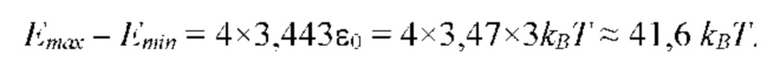
Скорость прохождения через энергетический барьер равна:
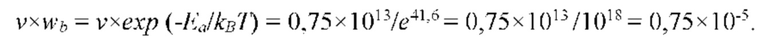
Откуда следует, что численное значение константы скорости равно:
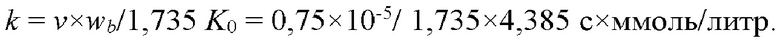
Концентрация глюкозы в элементарной ячейке увеличивается пропорционально увеличению константы взаимодействия (параметра порядка):
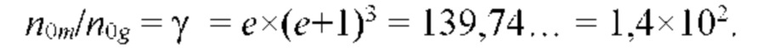
Произведение концентраций реагентов [G] и [Р] равно:
[G][P]=5 ммоль/литр×1×2,08×103 ммоль/литр; где [Р]=1 ммоль/литр - концентрация АТФ, [G]=5 - концентрация глюкозы. Здесь учитывается, что концентрация глюкозы и АТФ, при γ=1, равны соответственно 5 ммоль /литр и 1 ммоль /литр.
Скорость реакции равна:
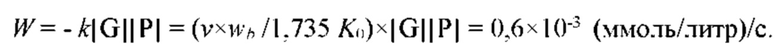
Фактор 1,1735 в расчете скорости реакции, появляется в результате изменения массы молекулы глюкозы после присоединения фосфатной группы. В процессе реакции фосфорилирования D - глюкоза (молекула G) присоединяет фосфатную группу PO3H2 с молекулярной массой 80 и превращается в глюкозо-6- фосфат (молекула GP) с молекулярной массой 260=180+80.
Константы равновесия реакций (53а) и (53b) связаны между собой выражением:
Здесь учитывается, что энергия связи Eb молекулы глюкозы с тетрамером, которая определяется внутренней энтропией тетрамера Stetr, величина которой не зависит от массы молекулы.
Откуда следует, KGP=KG/1,736=1/1,736 К0.
Расчетное значение согласуется с экспериментальными данными [13], при Km=10:
Скорость реакции гексокиназы.
Гексокиназа (АТФ-зависимая D-гексоза-6-фосфотрансфераза) (КФ 2.7.1.1) - цитоплазматический фермент класса трансфераз, подкласса фосфотрансфераз, первый фермент пути гликолиза.
Молекулярный вес 100000-50000, оптимальное значение рН равно 7,6-8,2. Первый изофермент, наиболее характерный для мозга, состоит из одной субъединицы, аминокислотный состав которой установлен. Вопрос о субъединичной структуре остальных изоферментов остается открытым. Для проявления каталитической активности нуждается в ионах Mg2+. Истинным субстратом для этого фермента служит не АТР4, а комплекс MgATP2.
Активность в различных органах (в мкмолях на 1 г влажного веса ткани в 1 мин.) от 23 (мозг) до 3,4 скелетные мышцы.
В активный центр входят остатки серина и гистидина. Важную роль в механизме каталитического действия играют группы SH полипептидной цепи. В организме гексокиназа активируется витаминами, инсулином, а также при связывании с некоторыми биологическими мембранами, например, митохондриальными; ингибируется глюкозо-6-фосфатом и кортикостероидами.
В отличие от глюкокиназы, константа Михаэлиса гексокиназы равна 0,01 ммоль/л, следовательно, гексокиназа, локализованная в клетках большинства тканей организма человека, буквально «вылавливает» глюкозу из плазмы крови, тогда как глюкокиназа катализирует реакцию фосфорилирования глюкозы лишь при высоких ее концентрациях. Соответственно, глюкокиназа и гексокиназа обеспечивают перераспределение потока глюкозы в организме: во время всасывания питательных веществ из кишечника концентрация глюкозы в плазме крови увеличивается, и глюкоза направляется в печень, где подвергается действию глюкокиназы; по окончании пищеварения, на фоне снижения концентрации глюкозы, она направляется в скелетные мышцы, где на нее действует гексокиназа.
Метаболизм глюкозы начинается с необратимой гексокиназной (или глюкокиназной) реакции, в которой катализируется перенос фосфатной группы АТФ на глюкозу с образованием глюкозо-6-фосфата. Гексокиназная реакция служит не только пусковой, но и главной лимитирующей реакцией среди других реакций гликолиза. Субстратная специфичность гексокиназы (КФ 2.7.1.1) относительна - фосфорилированию подвергаются, помимо глюкозы, фруктоза, манноза, галактоза, глюкозамин и некоторые неметаболизирующие сахара типа 2-дезоксиглюкозы. Гексокиназа локализована в плазматической мембране, цитоплазме и частично связана с митохондриями, причем в головном мозге активность этой фракции может достигать 50% суммарной активности фермента. Экспериментально установлено, что в возбуждении гексокиназы принимает участие ион калия, который также участвует в возбуждении других ферментов -пируваткиназы, ускоряющей транспорт фосфорной кислоты от пирувата на аденозиндифосфат, ферментов, участвующих в цикле Кребса, в том числе ферментов, помогающих в синтезе аденозинтрифосфата при окислительном фосфорилировании. Вполне очевидно, что зависимость активности гексокиназы от ионов калия указывает на прямую корреляцию между уровнем калия и уровнем макроэргических фосфатов. Таким образом, важность калия для вышеописанных процессов установлена экспериментально, однако принципы активации калием ферментативных систем пока полностью не выяснены.
Константа равновесия Михаэлиса для реакции гексокиназы равна Khex=0,02 ммоль/литр; отношение констант равновесия равно Kgl/Khex=15/0,02=750.
На Рис. 60а показаны кривые насыщения и скорость ферментативной реакции гексокиназы, в зависимости от безразмерной концентрации глюкозы, масштабированной делением концентрации на 0,5 ммоль/литр.
Высота барьера в расчете на одну водородную связь, как показано, равна
Emax - Emin=3,4436 ε0. Величину кванта энергии взаимодействия во в единицах кванта тепловой энергии kBT можно определить из графика энергетического барьера, представленного на Рис. 60б. Высота энергетического барьера одной валентной связи для прямой реакции E0λhex=2,443×ε0, для обратной реакции E0shex=3,443×ε0; энергия кванта взаимодействия ε0=е kBT.
Величину энергии активации можно определить из графика:
Emax=0,1035…
Emax=-0,1915…
Е0=-0,163…
Emax-Emin=0,1035+0,1915=0,295.
Е0-Emin=-0,161+0,1915=0,0305=kBT.
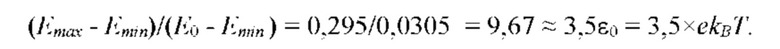
Разница между энергией связи прямой и обратной реакции равна:
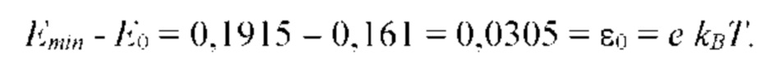
Константа диссоциации тетрамера, как показано, можно вычислить с помощью выражения:
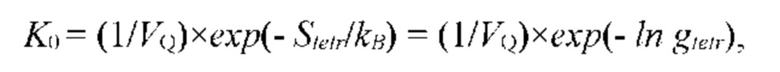
где gtetr - число вырожденных внутренних состояний (обладающих одинаковой энергией), реализующих равновесную конфигурацию тетрамера с 4 валентными связями, при выполнении условия компенсации сил притяжения и отталкивания.
Как следует из принципа квантования свободной энергии (6), описанного выше, при концентрациях глюкозы α≥1, равновесная конфигурация тетрамера, реализующаяся максимальным числом микросостояний gtetr=25, соответствующая минимуму свободной энергии взаимодействия, устанавливается по отношению к кванту энергии взаимодействия ε0, кратного кванту тепловой энергии ε0=n kBT, где n=1, 2, 3, …; ε0≥kBT. Микросостояния, реализующие равновесную конфигурацию тетрамера, при концентрациях глюкозы α≥1 обладают одинаковой энергией и являются вырожденными.
Как показано выше (15), изменение свободной энергии перехода (Na) тетрамера из конформации (λ) в изомерную конформацию (s) равно:
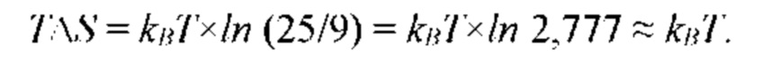
При концентрациях глюкозы α<1, соответствующих значениям константы Михаэлиса KM<5 ммоль/литр, пространственная конфигурация с минимальной свободной энергией объемных взаимодействий не может реализоваться с квантом энергии электромагнитного взаимодействия ε0, кратным кванту тепловой энергии kBT.
Можно показать, что равновесная конфигурация тетрамера, при α<1, может реализоваться при значении кванта энергии взаимодействия ε0=е kBT,
где е - основание натурального логарифма, которое является трансцендентным и иррациональным числом, которое нельзя представить в виде отношения m/n целого числа m к натуральному n, бесконечная непериодическая десятичная дробь, которое приблизительно равна 2,71828….
Число е, как известно, можно представить в виде формулы Эйлера:
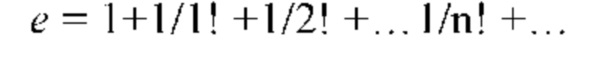
Величина кванта энергии ε0, в этом случае, может принимать квантованные значения εon:, соответствующие разным целым значениям квантового числа n:
ε01=(1+1)×kBT=2kBT;
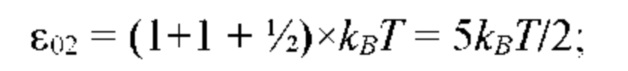
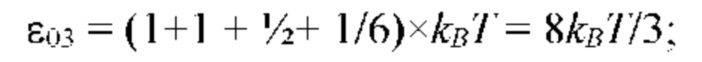
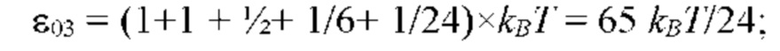
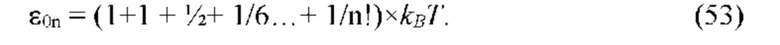
Энергия водородной связи, в этом случае, расщепляются на бесконечное число квантов взаимодействия, энергии которых определяются условием квантования (53), и, как следствие, уровень энергии энергетического барьера расщепляется на бесконечное число близко расположенных энергетических уровней.
В процессе теплового движения могут с равной вероятностью реализоваться конфигурации тетрамера с разными значениями расстояния между мономерами, соответствующие разным значениями энергии воп взаимодействия и энергии водородной связи.
Следует отметить, что разные значения межмолекулярных расстояний в тетрамере гексокиназы, в отличии от глюкокиназы, могут образоваться в том случае, если боковые группы аминокислот имеют большую длину за счет дополнительного мономера и дополнительной ковалентной связи, в результате которой появляется гибкость цепочки боковой группы. Отсюда следует понимание вывода, в случае гексокиназы, аминокислотой с отрицательным зарядом боковой группы является глутаминовая кислота (Глу, Е), боковая группа которой состоит из 3 мономерных звеньев; аминокислотой с незаряженной боковой группой является аспарагин (Асп, N), боковая группа которой состоит из 3 мономерных звеньев; в случае глюкокиназы, аминокислотой с отрицательным зарядом боковой группы является аспарагиновая кислота (Асп, D), боковая группа которой состоит из 2 мономерных звеньев; аминокислотой с незаряженной боковой группой является аминокислота, которую выбирают из группы треонин, цистеин, серии, боковые группы которых состоят из 2 мономерных звеньев.
В активный центр гексокиназы входят остатки серина и гистидина. Важную роль в механизме каталитического действия играют группы SH полипептидной цепи. Гексокиназа активируется витаминами, инсулином, а также при связывании с некоторыми биол. мембранами; ингибируется глюкозо-6-фосфатом и кортикостероидами.
Второе отличие гексокиназы от глюкокиназы заключается в том, что субстратная специфичность гексокиназы (КФ 2.7.1.1) относительна - фосфорилированию подвергаются, помимо глюкозы, фруктоза, манноза, галактоза, глюкозамин и некоторые неметаболизирующие сахара типа 2- дезоксиглюкозы. Можно предположить, что квантование энергии водородной связи (53) позволяет объяснить относительную субстратную специфичность гексокиназы, тем, что перечисленные сахарные молекулы образуют с мономерами полипептидной цепи водородные связи с близкими, но отличающимися значениями энергии, которым соответствуют разные значения расстояния между мономерами тетрамера.
Равновесная конфигурация R2 с 2 водородными связями, соответствующая минимальной свободной энергии при α<1, в соответствии с условием масштабной инвариантности (), может реализоваться при выполнении условия dS/dε≡0, которое может реализоваться единственным способом в результате расщепления на е квантов каждого кванта внутренней энтропии тетрамера, число которых соответствует числу допустимых конфигураций водородных связей в тетрамере. Отсюда следует, что 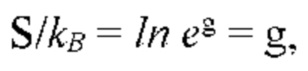 где g - число конфигураций, равное целому числу.
где g - число конфигураций, равное целому числу.
Как показано, при концентрациях глюкозы α≥1, равновесная конфигурация тетрамера, реализующаяся максимальным числом состояний gtetr=25, соответствующая минимуму свободной энергии взаимодействия, устанавливается по отношению к кванту энергии взаимодействия ε0=kBT. Максимум энергии взаимодействия (вершина энергетического барьера), как показано, достигается при концентрации глюкозы, соответствующей особой точке G процесса; тетрамер находится в конформации R2 с 2 водородными связями, которая может с равной вероятностью преобразоваться в конформации R4 или R0.
Число состояний g  реализующих конфигурацию R2 с 2 водородными связями, равно 18, из которых 6 состояний реализуют электронную конфигурацию и 12 состояний реализуют конфигурацию мономеров.
реализующих конфигурацию R2 с 2 водородными связями, равно 18, из которых 6 состояний реализуют электронную конфигурацию и 12 состояний реализуют конфигурацию мономеров.
Число состояний g  реализующих конфигурацию R2 с 2 водородными связями, возникающих при расщеплении энергии водородной связи, при концентрациях глюкозы α<1, равно:
реализующих конфигурацию R2 с 2 водородными связями, возникающих при расщеплении энергии водородной связи, при концентрациях глюкозы α<1, равно:
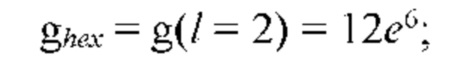
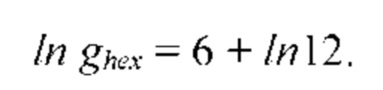
Энергия связи Eb, которую необходимо затратить для отрыва двух комплексов EG (молекула глюкозы G, связанная одной водородной связью с мономером Е), образующих комплекс тетрамера [E2G2] в конфигурации R2 с 2 водородными связями, можно вычислить с помощью выражения:
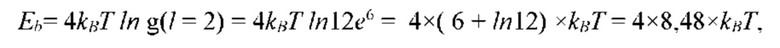
где g - число состояний, реализующих конфигурацию тетрамера R2 с 2 водородными связями.
Множитель 4 в формуле соответствует числу водородных связей в комплексе тетрамера. Откуда следует, что энергия связи, рассчитанная на одну водородную связь, равна:
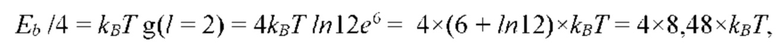
Константа равновесия равна:
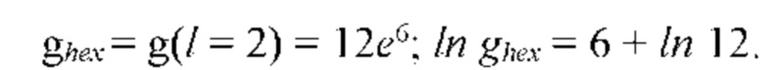
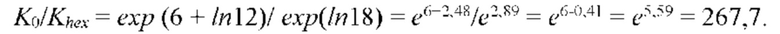
Khtx=К0/261,.7=4,4/267,7=0,016 ммоль/литр.
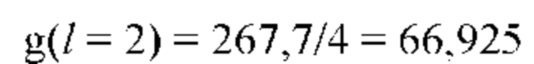
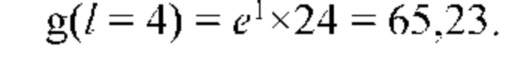
Энергия активации в расчете на 4 связи с квантом энергии ε0=ekBT равна:
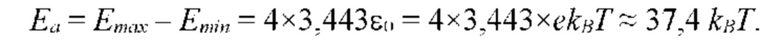
Скорость прохождения через барьер:
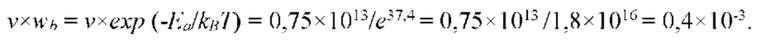
Скорость реакции равна:
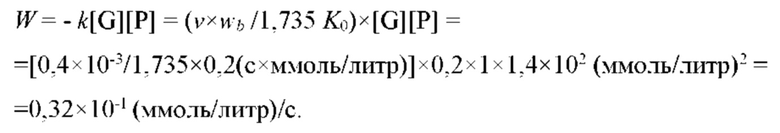
Результат расчета согласуются с экспериментальными данными [13].
Типы химической связи в тетрамерах природных макромолекул
В рамках общепринятой классификации, химическую связь в тетрамере между молекулами субстрата и мономерами полимерной цепи можно определить, как делокализованную водородную связь.
В зависимости от степени обобществления электронной плотности химической связи между несколькими атомами различают локализованную и делокализованную химическую связь. Ковалентная связь считается локализованной, если ее электронная пара находится в поле двух ядер и связывает только два атома. Если электронная плотность химической связи распределена между тремя и более ядрами, то такая связь называется трехцентровой, многоцентровой, а в общем случае - делокализованной. Делокализованная связь - связь, электронная пара которой рассредоточена между несколькими (более 2) ядрами атомов (подобие металлической связи). Характер делокализации электронов химической связи может, в свою очередь, различаться по размерности пространства. Существуют связи, делокализованные в одном измерении, делокализованные в плоскости и делокализованные в трехмерном пространстве. Хорошо известная металлическая связь с позиций изложенной классификации является короткодействующей и дальнодействующей, неполярной, в высшей степени делокализованной (в трех измерениях) связью.
Химическую связь, в зависимости от расстояния, на котором она проявляется, делят на короткодействующую и дальнодействующую. Прочность дальнодействующей связи в десятки раз меньше прочности короткодействующей связи. Дальнодействующую связь также называют слабой или межмолекулярной связью. Короткодействующая связь проявляется на расстояниях, близких к размерам атомов. Она осуществляется между атомами в молекуле, кристалле в пределах от 74 до 400 пм. Энергия разрыва короткодействующей химической связи находится в пределах от 40 до 1000 кДж/моль. Дальнодействующая химическая связь проявляется при переходе вещества из газовой фазы в жидкое или твердое состояние. Она возникает между отдельными атомами, молекулами на расстояниях в несколько раз больших, чем длина обычной короткодействующей связи. Короткодействующие связи образуются в результате такого взаимодействия, что каждый электрон может быть описан самостоятельной волновой функцией - одноэлектронное приближение. Дальнодействующие же связи являются результатом коллективного движения электронов.
Гликозидная связь.
Как было показано, термодинамические свойства биополимерной молекулы можно описать на основе статистической модели, как результат взаимодействия мономеров полимерной цепи с молекулами и ионами растворителя, в результате которого в объеме макромолекулы образуются тетрамеры - молекулярные комплексы, состоящие из 2 не соседних мономеров полимерной цепи и 2 молекул глюкозы растворителя, связанных между собой водородной связью, которую можно рассматривать, как тип слабой делокализованной ковалентной связи между молекулой глюкозы и мономерами полимерной цепи.
Как известно, молекула глюкозы может образовать гликозидную валентную связь также с аминокислотами белковых молекул. В гликопротеинах, характеризующихся низким содержанием углеводов, углеводный остаток (олигосахарид, имеющий нерегулярную структуру и содержащий глюкозу, маннозу, галактозу и их аминопроизводные, а также N-ацетилнейраминовую кислоту) присоединен к аминокислотам белковой цепи либо N-гликозидной связью - к амидному азоту аспарагина, либо О-гликозидной связью - к гидроксил группе остатков серина, треонина, гидроксилизина. Структурные формулы гликозидной связи с аминокислотами показаны на Рис. 61. Связь между аминогруппой или другой группой, содержащей атом азота, с сахаром, часто также называется гликозидной связью, хотя IUPAC этого и не рекомендует. Например, связь между сахаром и азотистым основанием в нуклеозиде называют гликозидной связью. Структурные формулы гликозидной связи в нуклеозидах показаны на Рис. 61.
Фармацевтические композиции на основе гиалуроновой кислоты.
Фармацевтические композиции и препараты на основе гиалуроновой кислоты широко применяются в медицине и косметологии: регенерация клеток кожи в эстетической косметологии; заживление ран и ожогов; защита структур глаза и обеспечения глубины передней камеры в ходе оперативного вмешательства в офтальмо хирургии и при производстве контактных линз; ускоренное размножение клеток костной ткани и быстрое сращивание кости при переломах; доставка лекарств к месту воспаления при патологических очагах и контроль за дозированным высвобождением препарата; ускоренное размножение клеток костной ткани и быстрое сращивание кости при переломах.
Новое понимание термодинамических свойств межклеточного вещества, основанное на новом методе расчета полимерной макромолекулы гиалуроновой кислоты, позволяет разработать принципиально новые фармацевтические композиции на основе гиалуроновой кислоты.
Изобретение также относится с фармацевтическим композициям, которые содержат биополимерную молекулу и один или несколько фармацевтически приемлемых носителей, адъювантов, растворителей и/или наполнителей, таких, которые могут быть введены в организм пациента совместно с биополимерной молекулой, составляющей суть данного изобретения, и которые не разрушают активности биополимерной молекулы, и являются нетоксичными в количествах, достаточных для доставки необходимого количества биополимерной молекулы.
Гиалуроновую кислоту, которая в настоящее время используют на практике, в косметологии и медицине, получают путем биохимического синтеза. Для этих целей используют бактериальные культуры, а конкретно стрептококки, выращенные на растительной основе (пшеничном бульоне). Этот метод базируется на способности некоторых микроорганизмов синтезировать гиалуроновую кислоту. Биохимический метод позволяет добиться большого количества вещества с нужным молекулярным весом и с приемлемой структурой.
Препараты на основе стабилизированной гиалуроновой кислоты, которые применяются в косметологии, долго хранятся в коже, составляют основу препаратов гидрорезерва и запускают регенеративные процессы в дерме. Молекула гиалуроновой кислоты очень чувствительна. Она остро реагирует на химическую модификацию: термическую или механическую; по этой причине препараты следует правильно сохранять в процессе химических реакций. Стабилизированная гиалуроновая кислота получается методом биохимического синтеза, затем следует процесс сшивания, который называется стабилизацией (формирование пересекающихся сшивок между молекулами гиалуроновой кислоты). Молекулы гиалуроновой кислоты подвергаются сшиванию в целях предотвращения их быстрой деградации. Такая гиалуроновая кислота демонстрирует долгие клинические эффекты при ее внедрении в кожу. После сшивания полученные гели проходят очистку, которая представляет собой очень кропотливый процесс и является решающим фактором при ценообразовании препаратов стабилизирующей гиалуроновой кислоты.
В зависимости от уровня стабилизации производятся гели различной вязкости для устранения разнообразных эстетических проблем: мало стабилизированные - для устранения мелких морщин, более стабилизированные и более вязкие - для коррекции носогубных складок и восстановления утраченных объемов.
Стабилизированная гиалуроновая кислота используется в контурной практике и при армировании лица, так как этот вид гиалуроната (натриевая соль гиалуроновой кислоты) хорошо держит объем. То есть, когда надо восполнить утраченные объемы, например, щек, вытолкнуть носогубные складки извне, смоделировать контур лица и заполнить провалы на лице, используется стабилизированная гиалуроновая кислота.
Гиалуроновая кислота должна быть стабилизированной и обладать характеристиками, которые проявляются в нативном состоянии живой ткани, при нормальных физиологических условиях, при которых межклеточное вещество находится в гетерофазном состоянии. Как было показано, нормальные физиологические характеристики живой ткани достигаются, при физиологических значениях температуры и кислотности, при следующих концентрациях биохимических компонентов в межклеточном веществе:

Биополимерные молекулы, описанные в данном изобретении, могут быть использованы для профилактики и/или лечения болезней человека, например, в виде следующих составов (под «биополимерное молекулой» понимается активный ингредиент, представляющий собой биополимерную молекулы по изобретению в гетерофазном состоянию).
Концентрация активного ингредиента в препарате, в частности, эквивалентная концентрации мономеров гиалуроновой кислоты 45 ммоль/литр, равна 15 мг/мл.
Концентрация глюкозы в препарате, эквивалентная концентрации свободной (несвязанной полимером) глюкозы 5 ммоль/литр, в диапазоне 5-15 мг/мл.
Концентрация ионов натрия и калия в водном растворе с рН 7,0-7,5 равна 40 ммоль/литр и 5 ммоль/литр соответственно.



Лечебные композиции на основе ферментов.
Сахарный диабет 2-го типа является актуальной проблемой современного здравоохранения. Несмотря на широкий выбор пероральных гипогликемических препаратов, на сегодняшний день велика потребность в создании и внедрении в клиническую практику новых эффективных и безопасных лекарственных средств. Одной из перспективных мишеней для создания новых противодиабетических средств является глюкокиназа (GK). Она выступает в качестве «сенсора» глюкозы в панкреатических бета-клетках и контролирует скорость синтеза гликогена в печени. Активаторы глюкокиназы представляют собой новый класс противодиабетических препаратов с надежным гипогликемическим эффектом, доказанным в доклинических испытаниях. Очень важно, что активаторы глюкокиназы не вызывают клинически значимой гипогликемии при использовании экспериментальных моделей сахарного диабета и не влияют на уровень липидов, а также не увеличивают массу тела.
В 2003 г. впервые сообщили об успешном применении низкомолекулярного активатора GK (GKA) для снижения уровня глюкозы в животных моделях сахарного диабета типа 2. Скрининг 120000 соединений позволил идентифицировать соединение, обладающее свойствами аллостерического активатора GK. Активный R энантиомер этого соединения, RO82881675, повышал ферментативную активность GK, при пероральном введении снижал уровень глюкозы в крови мышей дикого типа и в моделях диабета, ускорял метаболизм глюкозы в печени и индуцируемую глюкозой секрецию инсулина в изолированных островках поджелудочной железы крысы. Поиск и характеристика других низкомолекулярных GKA стали одним из главных направлений в разработке противодиабетических препаратов в течение следующего десятилетия. К созданию и испытанию таких соединений подключились крупные фармацевтические компании, в том числе "Array Biopharm Inc.", "Astra Zeneca pic, Merck & Co.", "OSI Pharmaceuticals Inc./Eli Lilly", "Pfizer Inc.", "Roche", "DaiichiSankyio", "Takeda", TransTech Pharma Inc./Forest Laboratories Inc./Novo Nordisk A/S", "Advinus", "Hua Medicine". К 2008 году получено уже более 90 патентов на низкомоле8 кулярные аллостерические GKA. Большая часть этих соединений основана на фармакофоре, схема которого представлена на рис. 2. Большинство этих соединений эффективно снижали уровень глюкозы в крови в разных животных моделях диабета. Более 20 потенциальных сахароснижающих препаратов этого класса были допущены до клинических испытаний, однако в большинстве случаев испытания были приостановлены или прекращены на стадии I или II [16]. Главной причиной прекращения испытаний стал повышенный риск развития гипогликемии. В настоящее время продолжаются клинические испытания лишь пяти препаратов. Это GKM8001 индийской фармацевтической компании "Advinus Therapeutics Ltd." (фаза II), DS87309 японской компании "Daiichi8Sankyo" (фаза I), HMS5552 китайской компании "Hua Medicine" (фаза I), PF804937319 компании "Pfizer" (фаза II) и ТТР 399 (GK18399) компаний "TransTech Pharma Inc./Forest Laboratories Inc./Novo Nordisk A/S" (фаза II). Дальнейшие перспективы использования низкомолекулярных аллостерических GKA связывают с возможностью создания гепатоселективных препаратов с низким риском развития гипогликемии.
СПИСОК ЛИТЕРАТУРЫ
1. John L. Smith. The Pursuit of Noninvasive Glucose: "Hunting the Deceitful Turkey". Fourth Edition, 2015.
2. USP # 5,795,305, August 18,1998, Cho et al. Process and device for non-invasive determination of glucose concentration in parts of human body.
3. USP #5,823,966, October 20, 1998, Buchet. Non - invasive continuous blood glucose monitoring.
4. USP # 5,890,489, April 6, 1999, Elden. Method for non - invasive determination of glucose in body fluids.
5. USP # 5,924,996, July 20, 1999, Cho et al. Process and device for detecting the exchange of heat between the human body and the invented device and its correlation to the glucose concentration in human body.
6. USP # 6,522,903, February 18,2003, Berman et al. Glucose measurement assessment utilizing non-invasive assessment methods.
7. USP # 6,517,482, February 11, 2003, Elden et al. Method and apparatus for non- invasive determination of glucose in body fluids.
8. USP # 5,040,541, August 20, 1991, Poppendiek. Whole body calorimeter.
9. USP # 4,386,604, June 7, 1993, Hershey. Determination of the basal metabolic rate of humans with a whole-body calorimeter.
10. Мусин Р.Ф. Патент РФ на изобретение №2629796 "Способ и мультисенсорное устройство для неинвазивного мониторинга уровня глюкозы в крови".
11. Мусин Р.Ф. Патент РФ на изобретение №2396897.
12. Edsall J.T., Gutfreund Н. BIOTHERMODINAMICS. The Study of Biochemical Processes at Equilibrium. JOHN WILLEY & SONS, 1983.00
13. Кендыш И.Н. Регуляция углеводного обмена. Издательство "Медицина", Москва.
14. Peusner Leonardo. Concepts in Bioenergetics. Peusner Biomedical Associates, Prentice - Hall, Inc., Englewood Cliffs, New Jersey, 1974.
15. Lehninger A.L. BIOCHEMISTRY. The Johns Hopkins University, School of Medicine. Worth Publishers, Inc., New York, 1972.
16. Atkins P.W. The second law. Scientific American Library, New York, 1984.
17. Мусин Р.Ф., Морозов B.A., Годик Э.Э., Гуляев Ю.В. (1986). Электрические свойства рогового слоя эпидермиса человека и транспорт воды в нем. - Биофизика, т. 31, вып.3, с. 478-480.
18. Forbath N., Hetenyi С.Glucose dynamics in normal subjects and diabetic patients before and after a glucose load. - Diabetes, 1966, vol. 15, # 11, p.778-789.
19. Hall S. E. H., Saunders J., Sonksen P.H. Glucose and free fatty acid turnover in normal subjects and diabetic patients before and after insulin treatment. - Diabetologia, 1979, vol. 16, # 5, p.297-306.
20. Musin R.F., Godik E.E., Gulyaev Y.V., Morozov V.A. Natural water diffusion through the stratum corneum of the human body epidermis and its electrical properties/ - 3 International conference on water and ions in biological systems. Bucharest, Romania, 1984, p.34.
21. Мусин Р.Ф., Иванова Н.Ю., Мартынов B.A., Морозов В.А., Годик Э.Э., Гуляев Ю.В. (1986). О чувствительности кожи человека к инфракрасным тепловым потокам. -Доклады академии наук СССР, том 289, №3, с. 718-720.
22. Гуляев Ю.В., Годик Э.Э., Мусин Р.Ф., Морозов В.А., Мартынов В.А., Валиев И.В. (1989). Пороги тепловой чувствительности кожи к электромагнитным излучениям. -Сенсорные системы, том 3, №2, с. 209-212.
23. Годик Э.Э., Морозов В.А., Мусин Р.Ф. (1985). О динамике релаксации трибозаряда на поверхности рогового слоя эпидермиса кожи. - Биофизика, том 30, вып. 2, с. 309-312.
24. Musin R.F., Godik Е.Е., Gulyaev Y.V., Morozov V.A., Sudarev A.M. Membrane mechanisms of water transport in epidermis. 4 International conference on water and ions in biological systems. Bucharest, Romania, 1988, p.167-172.
25. Мусин Р.Ф., Морозов B.A., Сударев A.M. (1990). О механизмах транспорта воды в эпидермисе. - Биофизика, том 35, вып. 4, с. 653-656.
26. Yas Kuno. (1959). Human Perspiration. Charles & Thomas Publ., Springfield, Illinois, USA.
27. A.B. Коробков, C.A. Чеснокова. (1986). Атлас по нормальной физиологии. М.: Высшая школа, 351 с.
28. Гомеостаз. Под ред. П.Д. Горизонтова. М., "Медицина", 1976, 464 с., ил.
29. Кальве Э., Прат А. Микрокалориметрия. М.: Изд-во ин. лит. 1963. 477 с.).
30. Ф. Дайсон, Э. Монтролл, М. Кац, М. Фишер. Устойчивость и фазовые переходы. Издательство "Мир", Москва, 1973.
31. Lehninger A., Biochemistry, Worth Publishers, New York, 1972.
32. с. В. Anfmsen. "Molecular Organization and Biological Function (Modern Perspectives in Biology)". New York. Harper & Row, 1966.
33. C.R. Cantor, Paul R. Schimmel. Biophysical Chemistry. Parts 1, 2, 3. W. H. Freeman and Company. San Francisco. 1980. - 536pp.
34. J.M. Guss, D.W.L. Hukins, P.J.C. Smith, W.T. Winter, S. Arnott, R. Moorhouse, DA. Rees Hyaluronic Acid, Molecular Conformations and Interactions in Two Sodium Salts; J. Mol. Biol. v. 95,359 (1975).
35. A.G. Marshall, Biophysical Chemistry, Parts 1,2. John Wiley and Sons, Inc. 1978.
36. L.E. Boltzmann, Lectures on Gas Theory, Berkley, (1964).
37. J.W. Gibbs, Elementary principles in statistical mechanics, developed with especial reference to the rational foundation of thermodynamics, Yale Univ. Press, 1902.
38. J.W. Gibbs, Collected Works, New Haven, (1948).
39. E. Fermi. Thermodynamics. New York, Prentice-Hall, Inc, 1937.
40. E. Fermi, Notes on thermodynamics and statistics, Univ. Chicago Press, 1966.
41. R.P. Feynman. Statistical Mechanics. Set of Lectures. California Institute of Technology. W.A. Benjamin, Inc., Massachusetts (1972).
42. C. Kittel. Thermal Physics. John Willey and Sons, Inc. New York. 1969.
43. C. Bohr, Zbl. Physiologie 17, 682 (1903).
44. J. Monod, J. Wyman, J.P. Changeux, J. Mol., Biol. 12, 88 (1965).
45. J.T. Edsall, H. Gutfreund. Biothermodynamics. John Wiley & Sons, 1983.
46. Paul J. Floiy, The Configuration of Real Polymer Chains, J. Chem. Phys. 17, 303 (1949).
47. Гросберг А.Ю., Хохлов A.P. "Статистическая физика макромолекул" -М.: Наука, 1989.
48. A. Yi! Grosberg, A.R Khokhlov, Statistical Physics of Macromolecules,, 1994.
49. Гросберг А.Ю., Хохлов A.P. Физика цепных молекул. - М.: Знание, 1983.
50. И.М. Лифшиц, А.Ю. Гросберг, А. Р. Хохлов, Объемные взаимодействия в статистической физике полимерной макромолекулы, УФН 127 (3) (1979)
51. Ptitsyn О.В. The molten globule state (Review). In: "Protein Folding" (Т.Е. Creighton, ed.), W.H. Freeman & Co., 1992, 243-300.
52. Hadler N.M. Enhanced Diffusivity of Glucose in a Matrix of Hyaluronic Acid. The Journal of Biological Chemistry. 1980; 255(8): 3532-5.
50. Бродский Я.С. Статистика. Вероятность. Комбинаторика. - М.: Издательство " Мир и Образование", 2008. - 544 с.
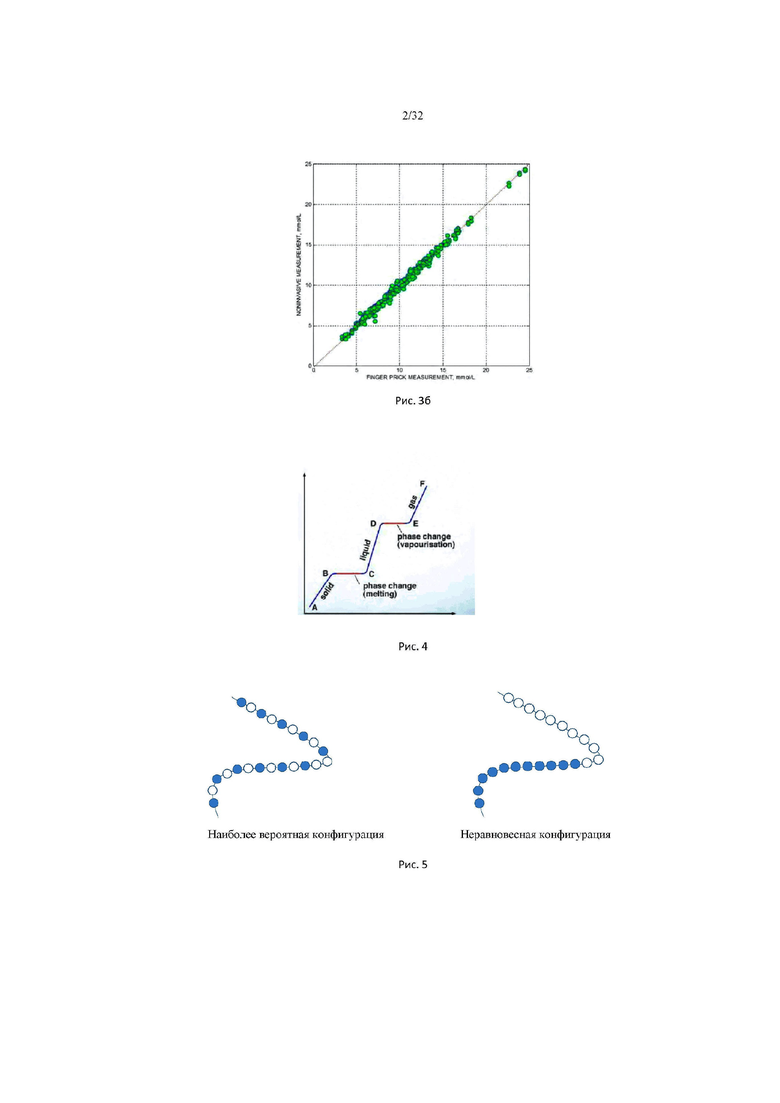
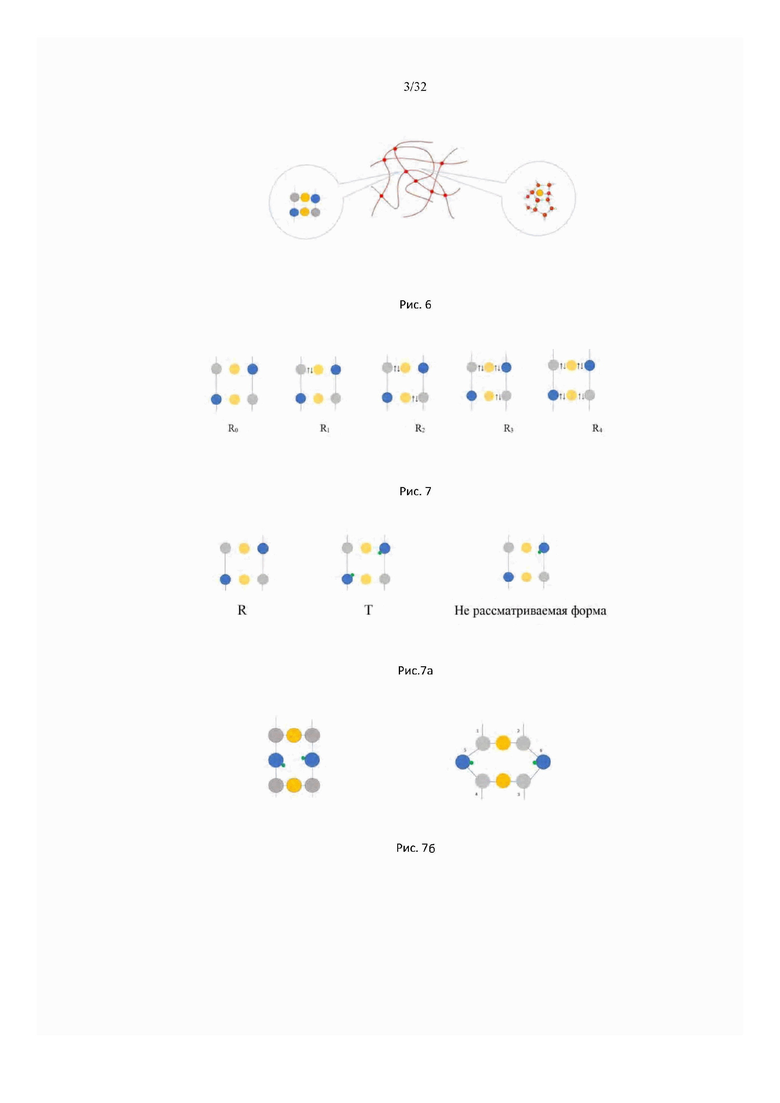
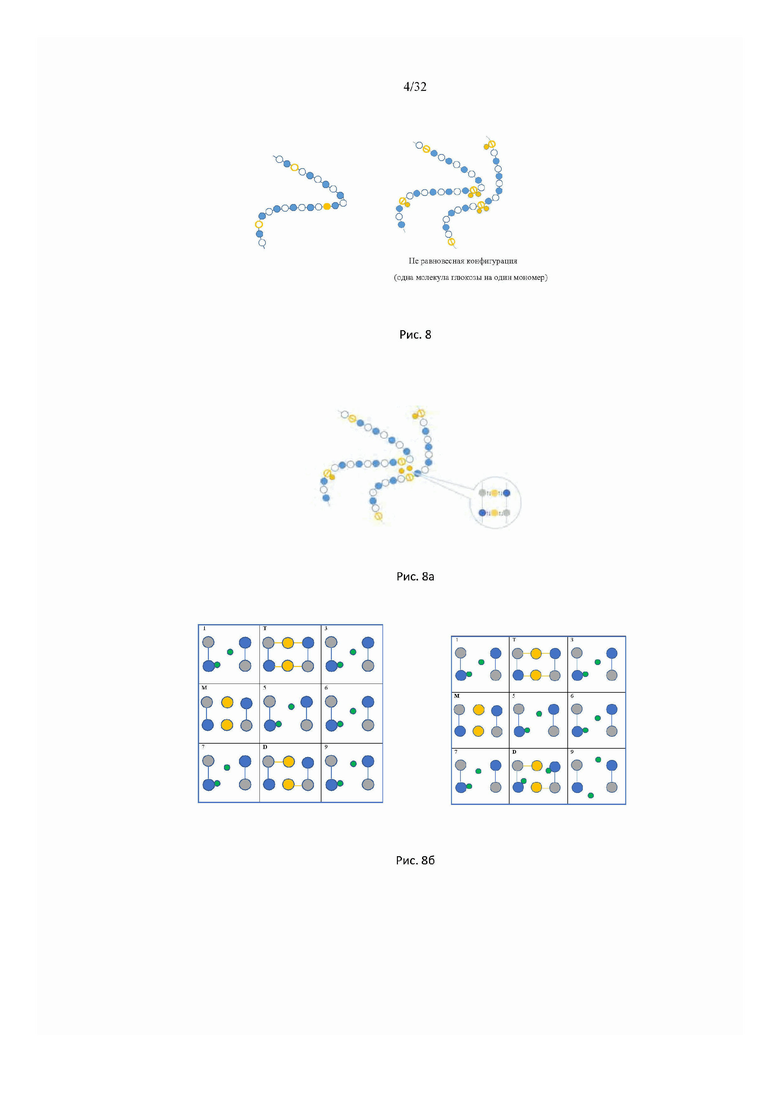
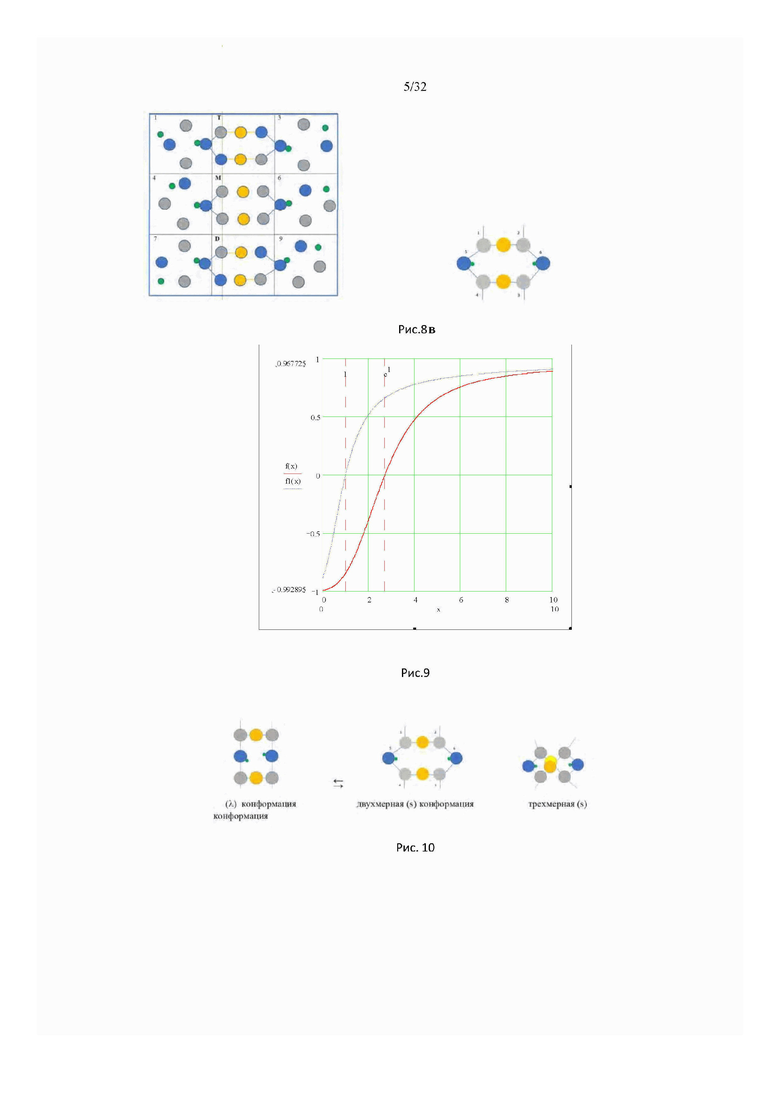
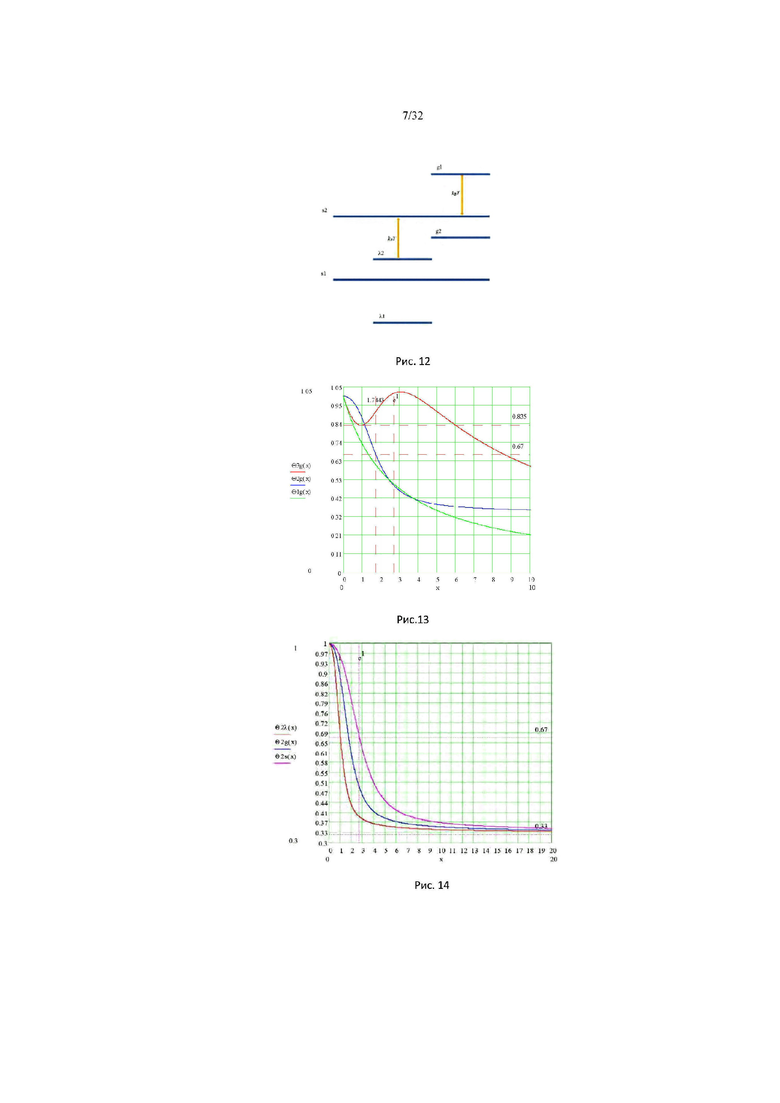
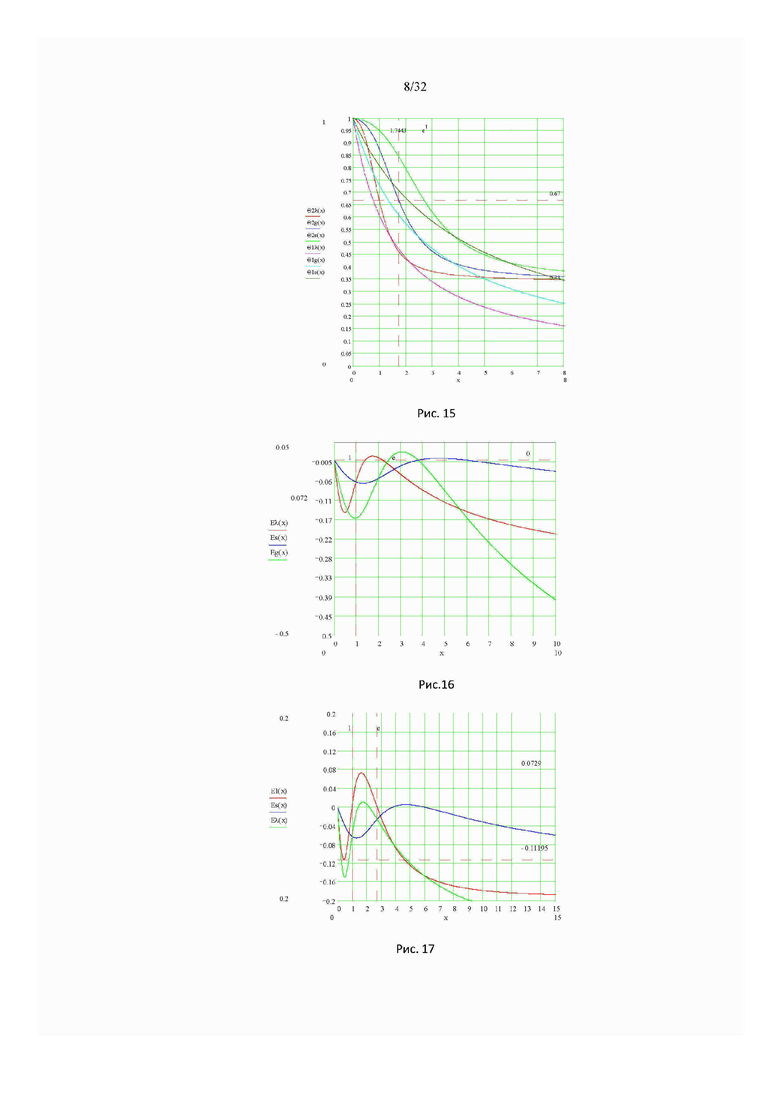
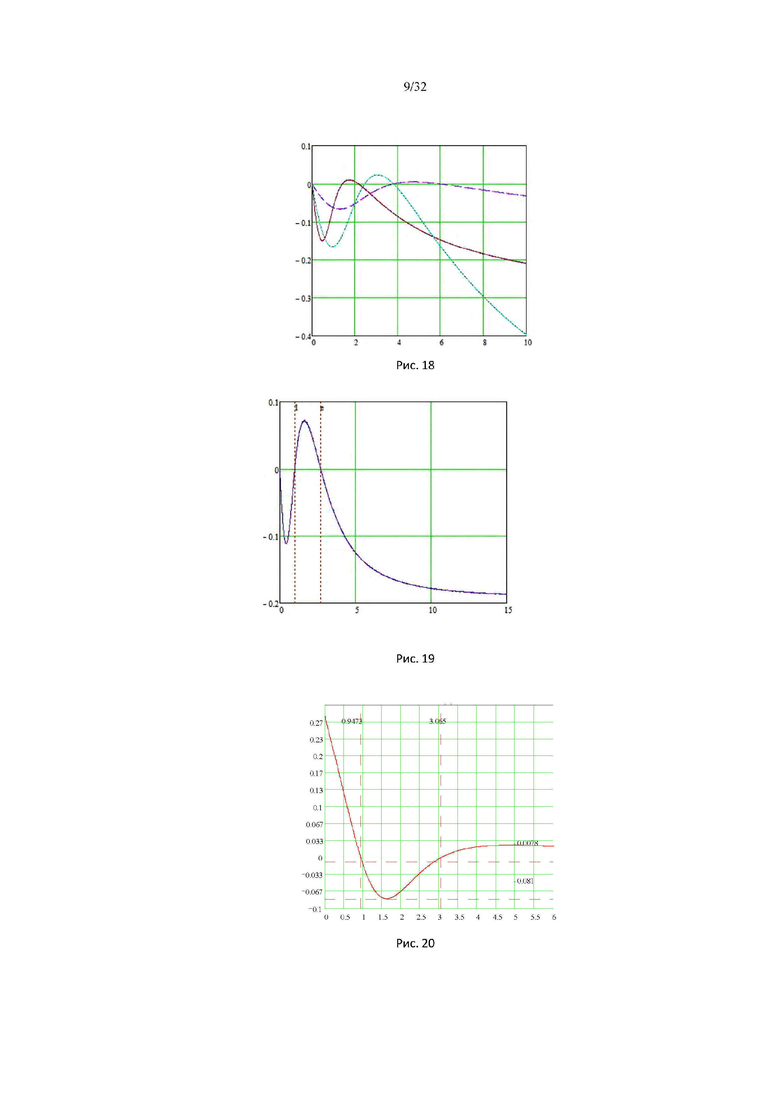
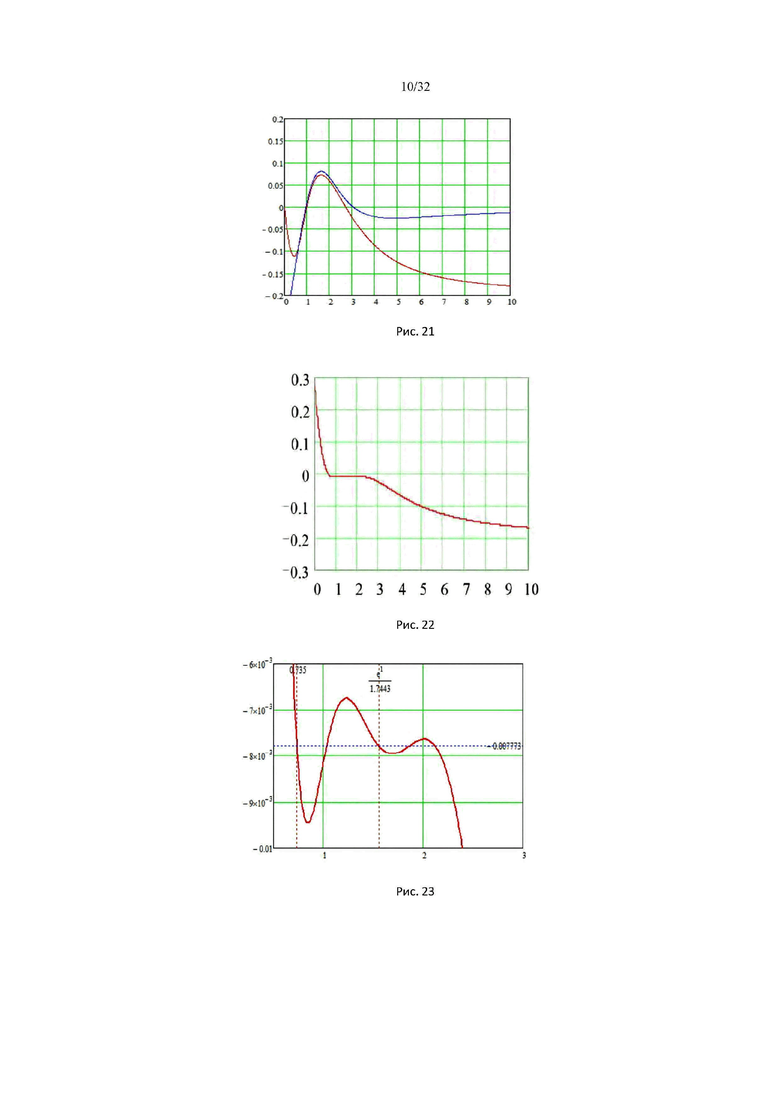
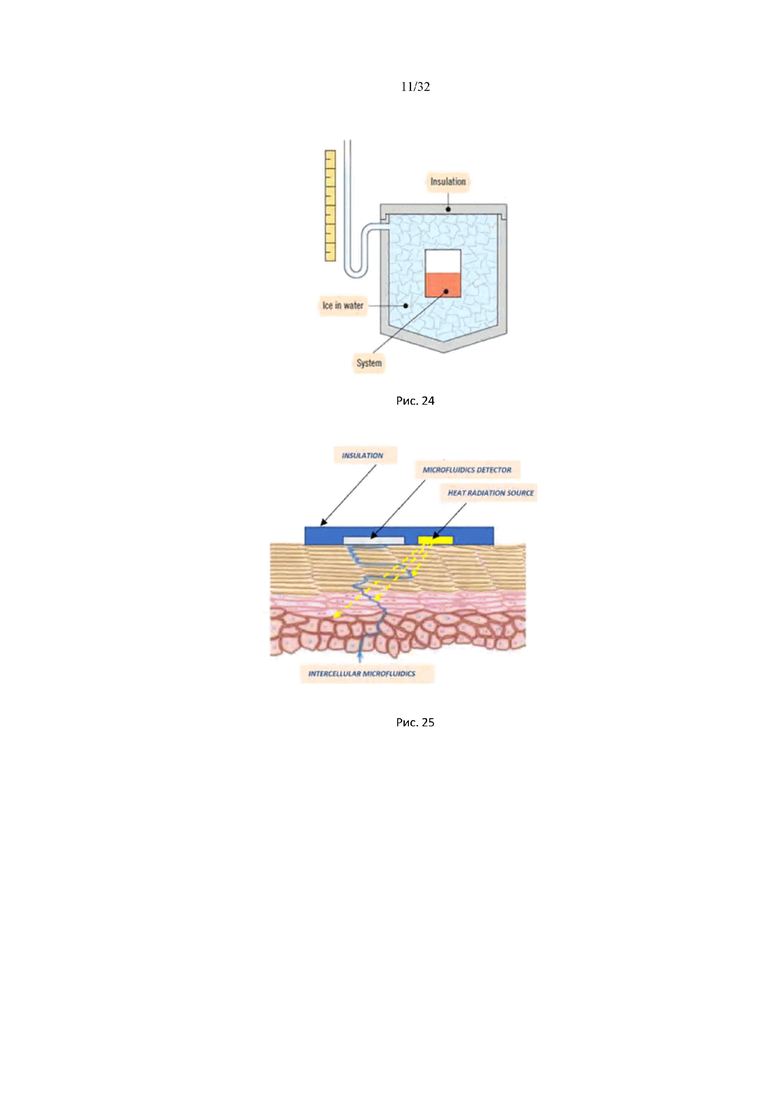
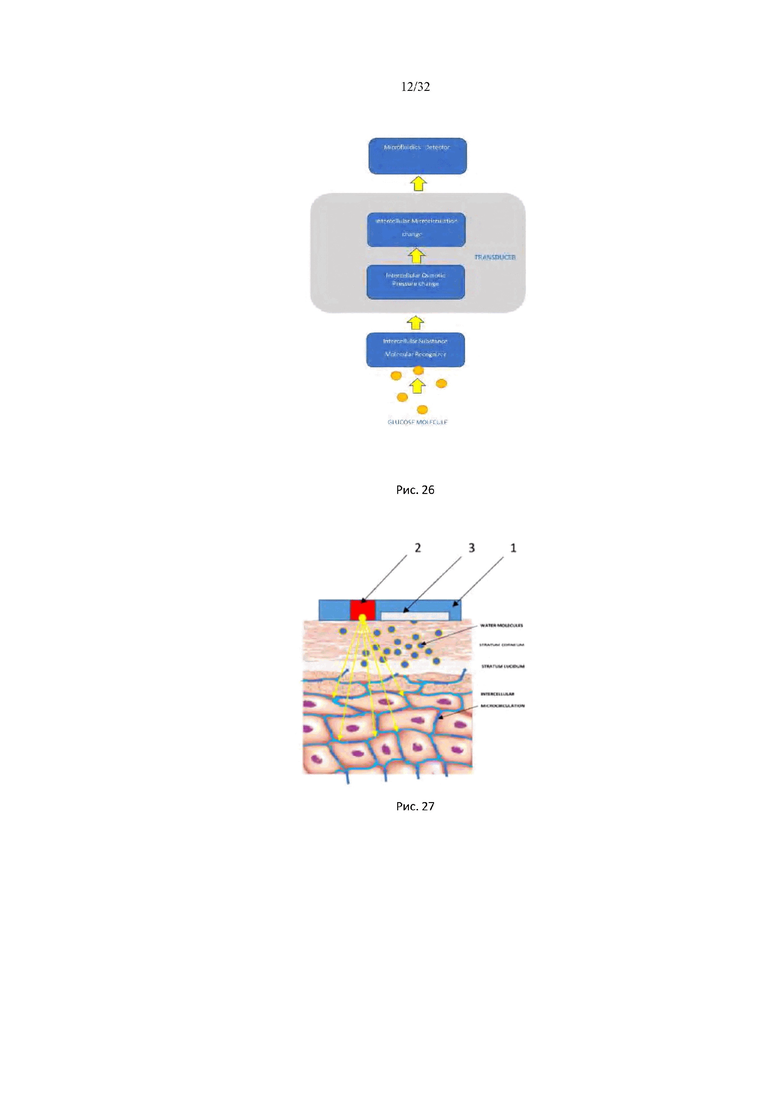
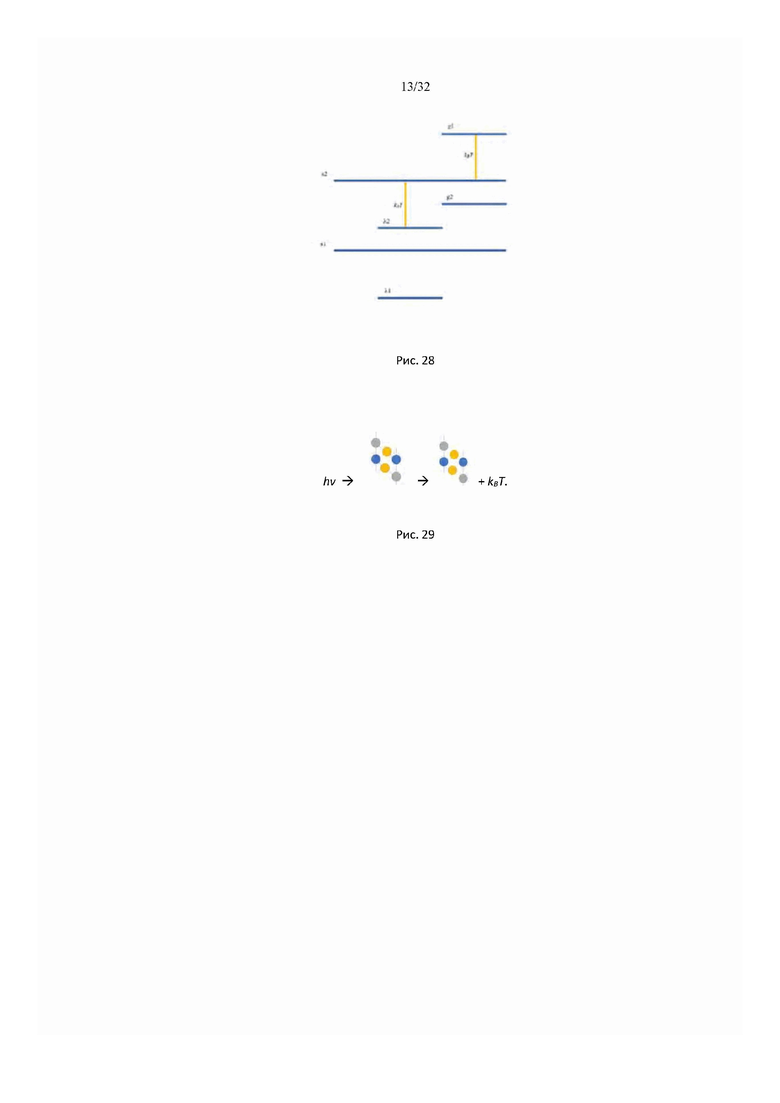
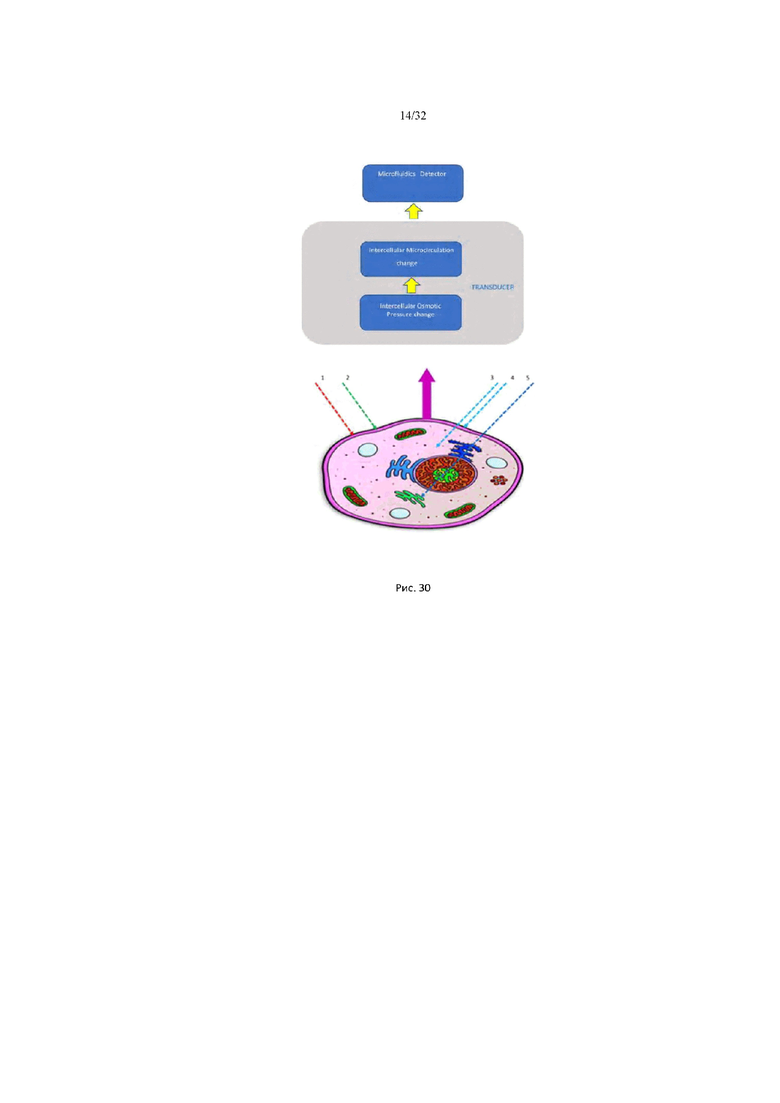
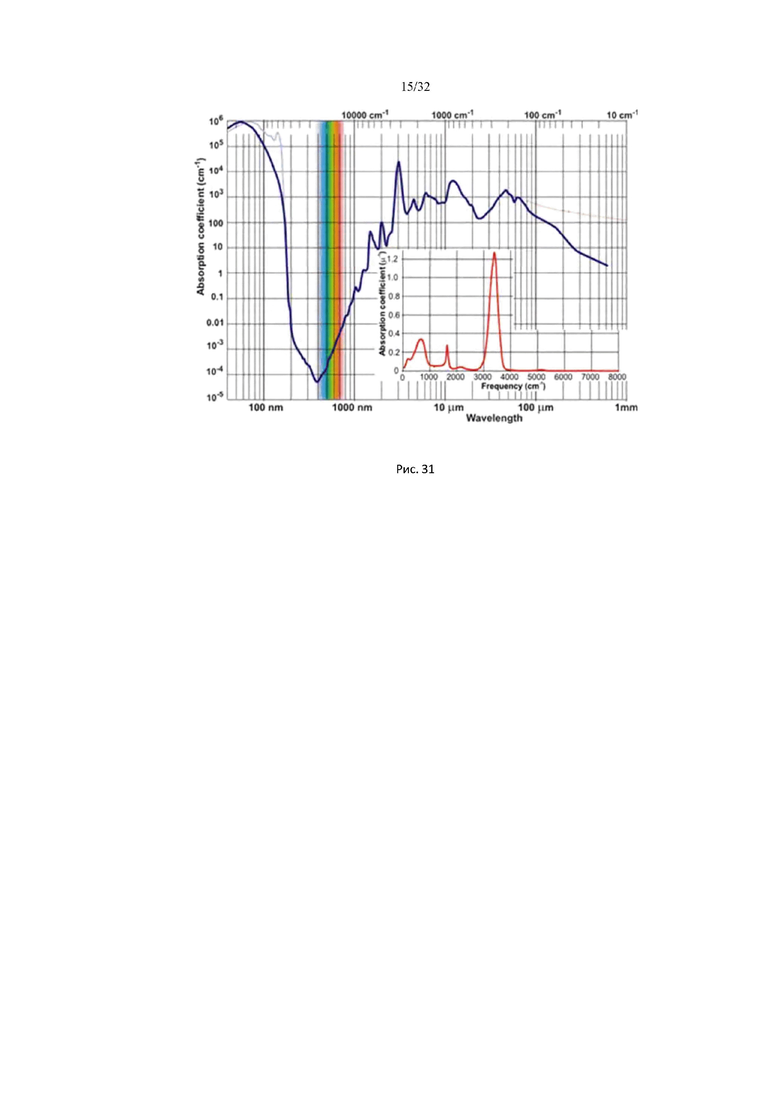
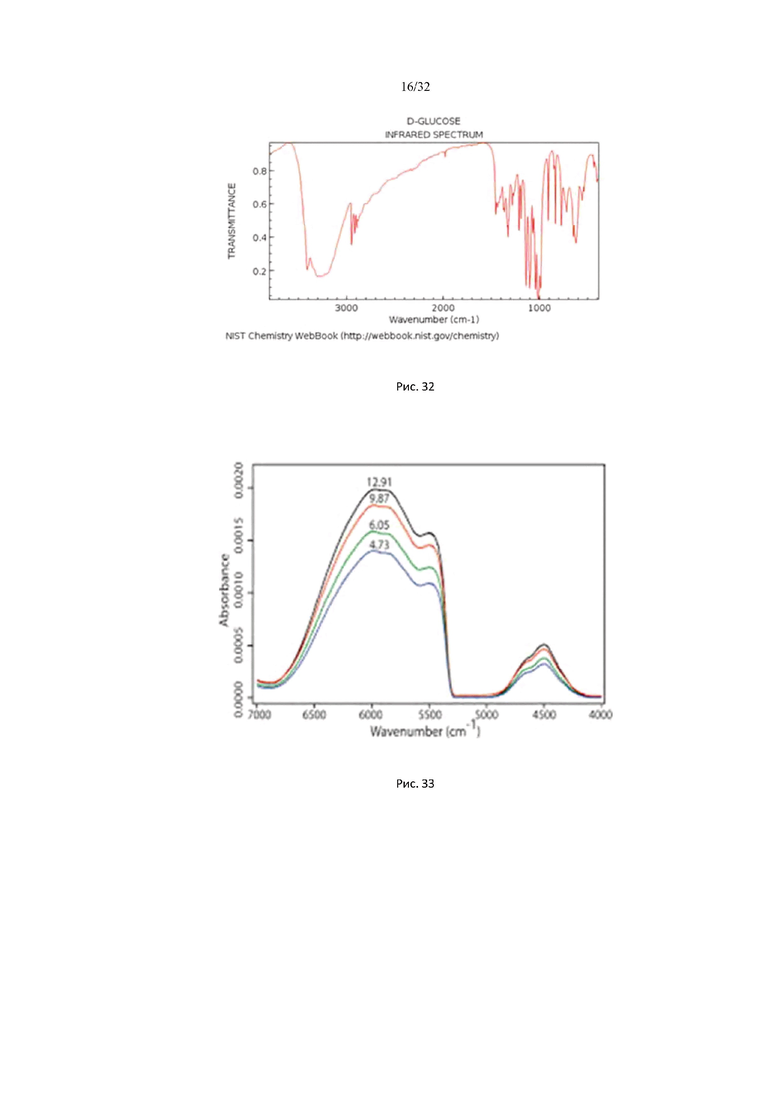

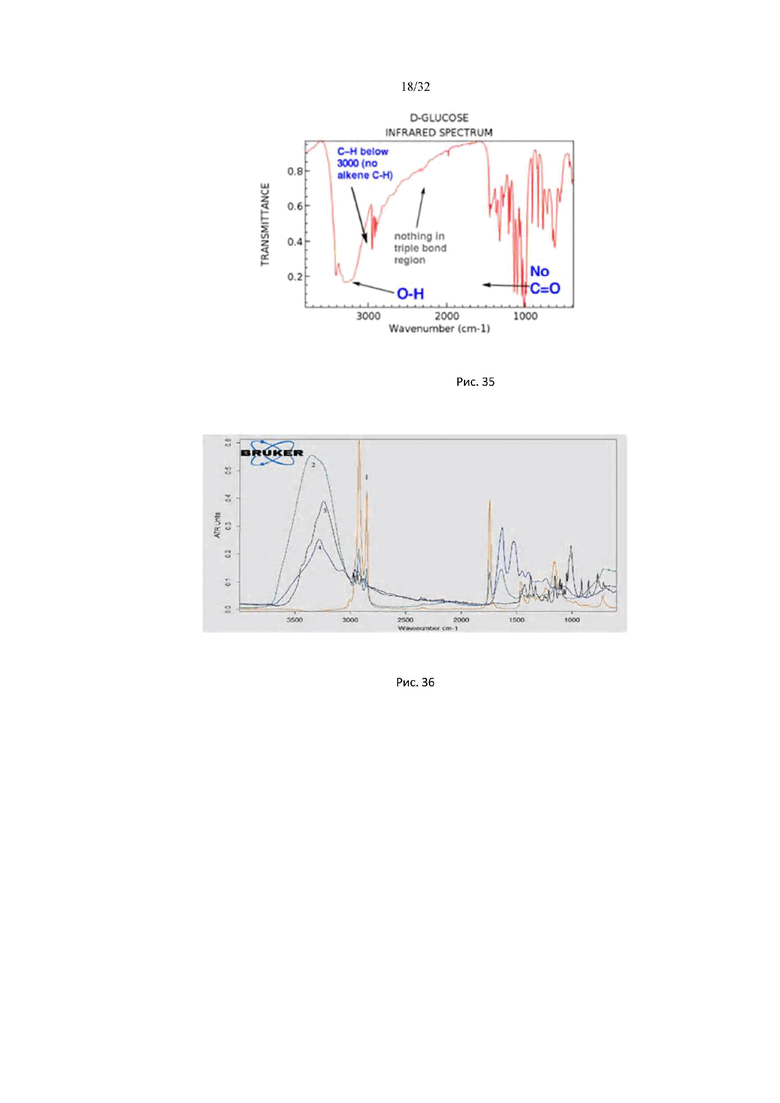
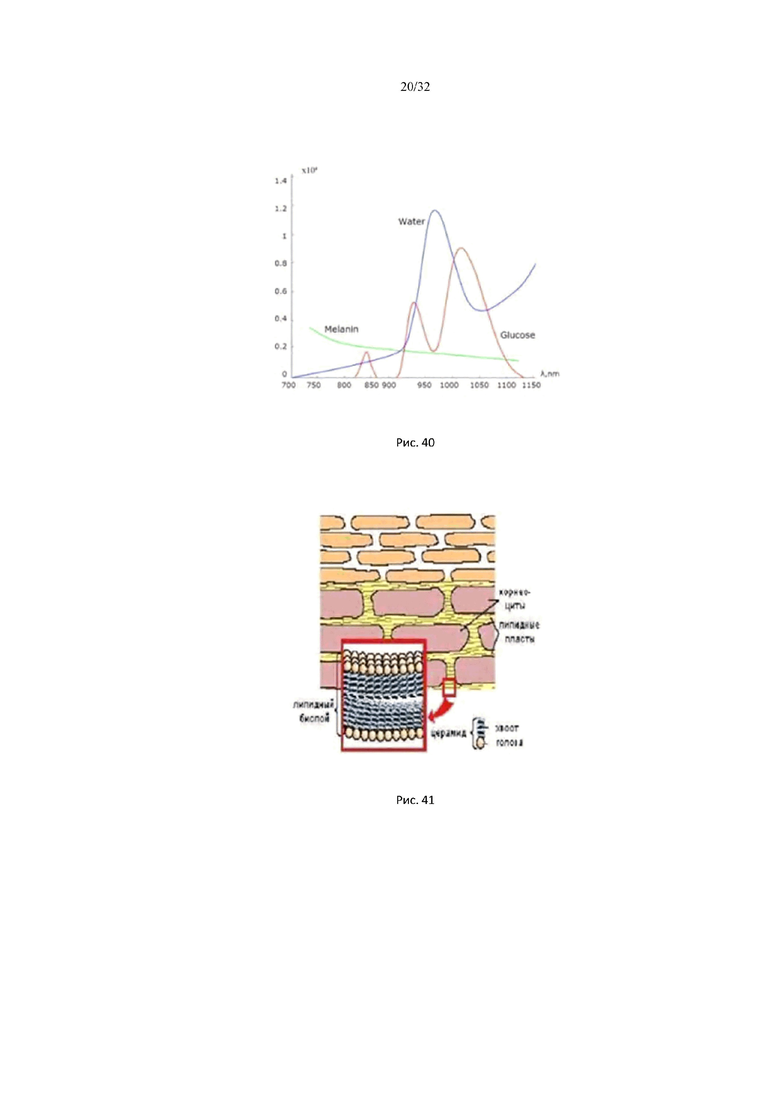
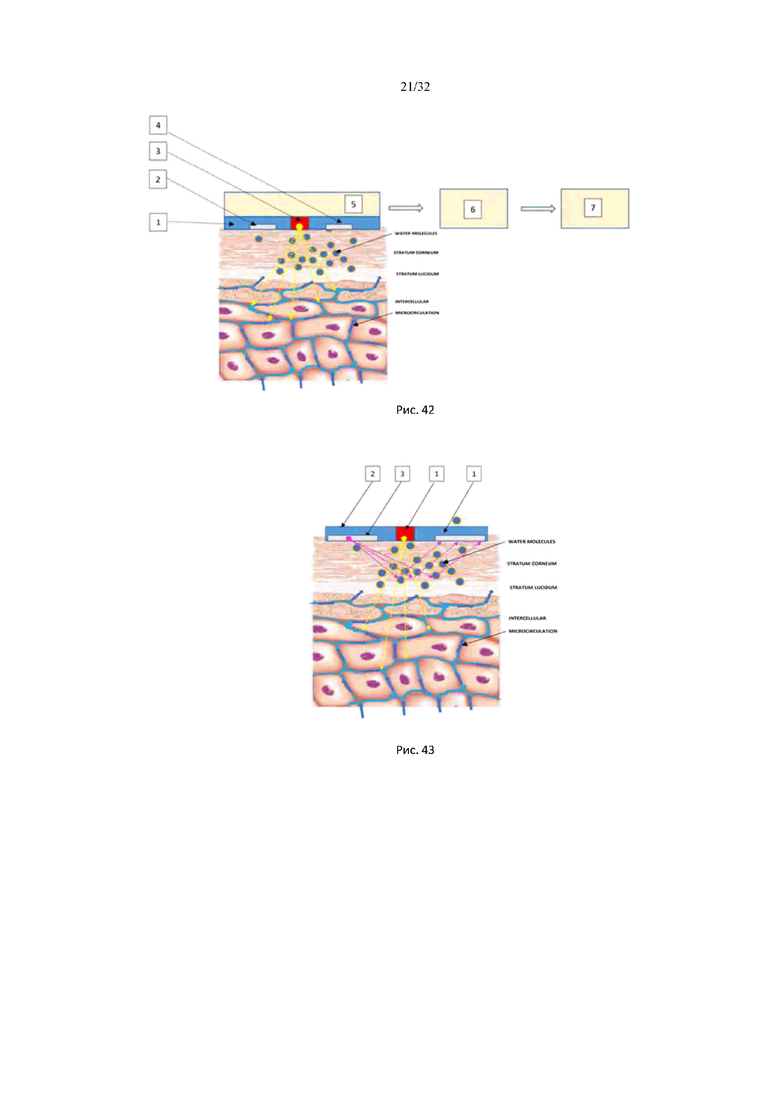
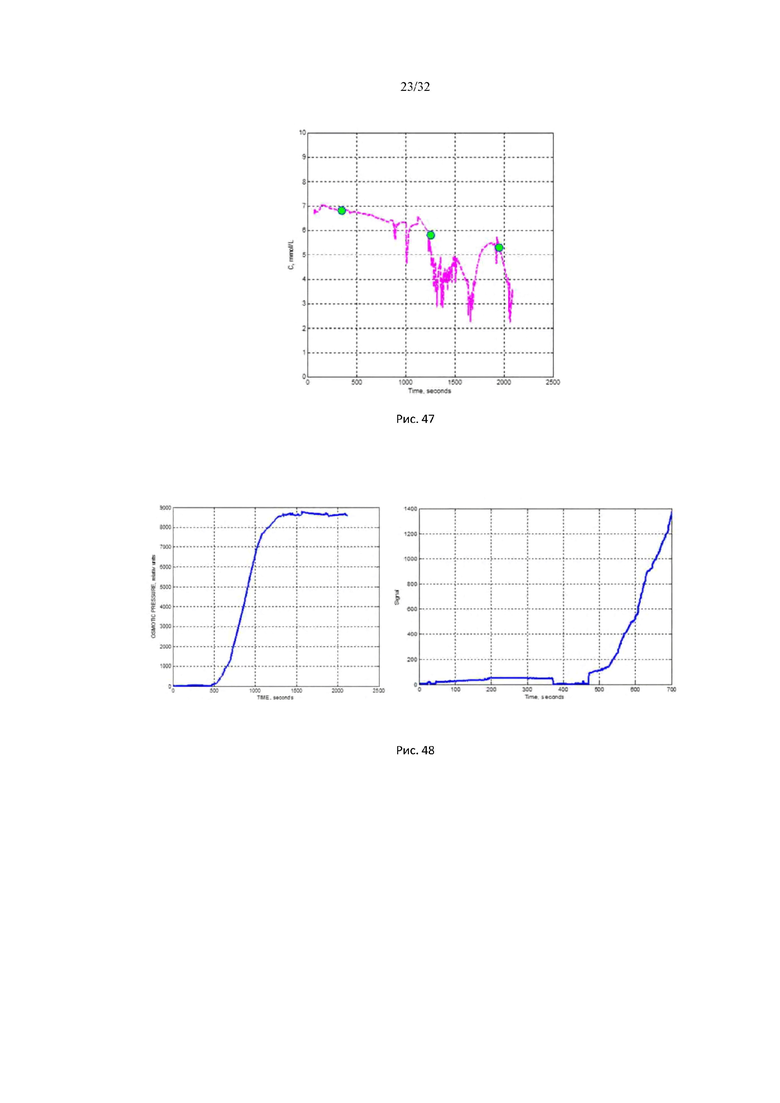
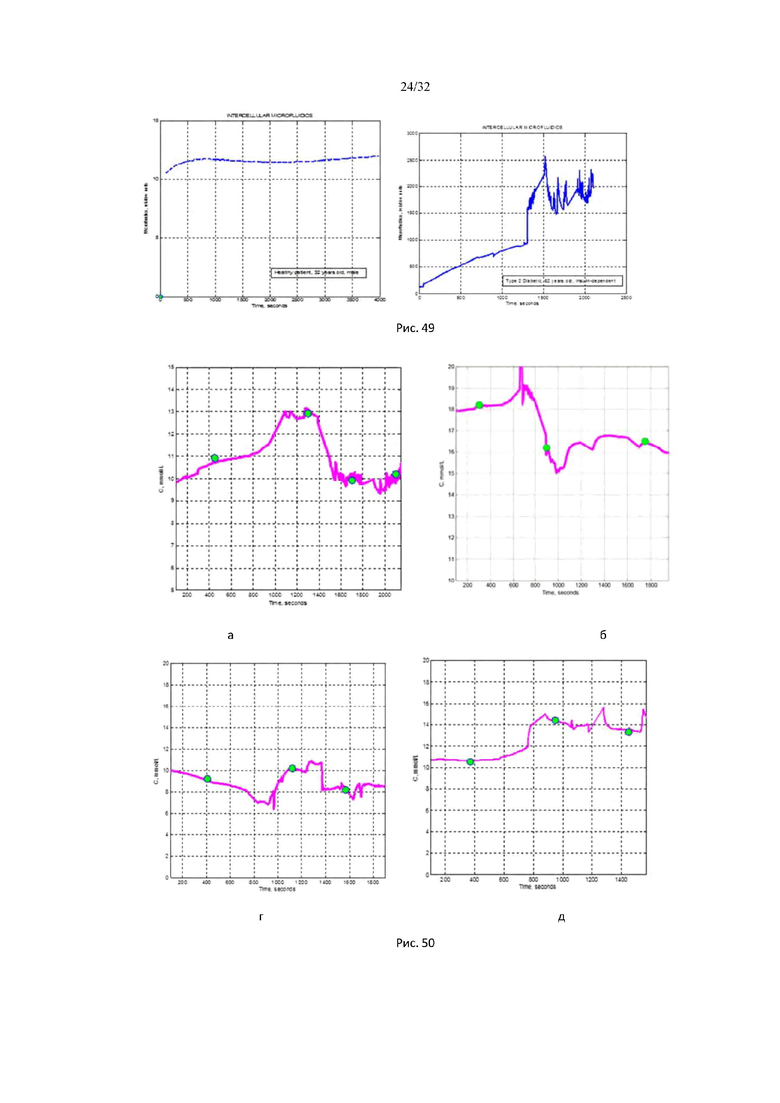
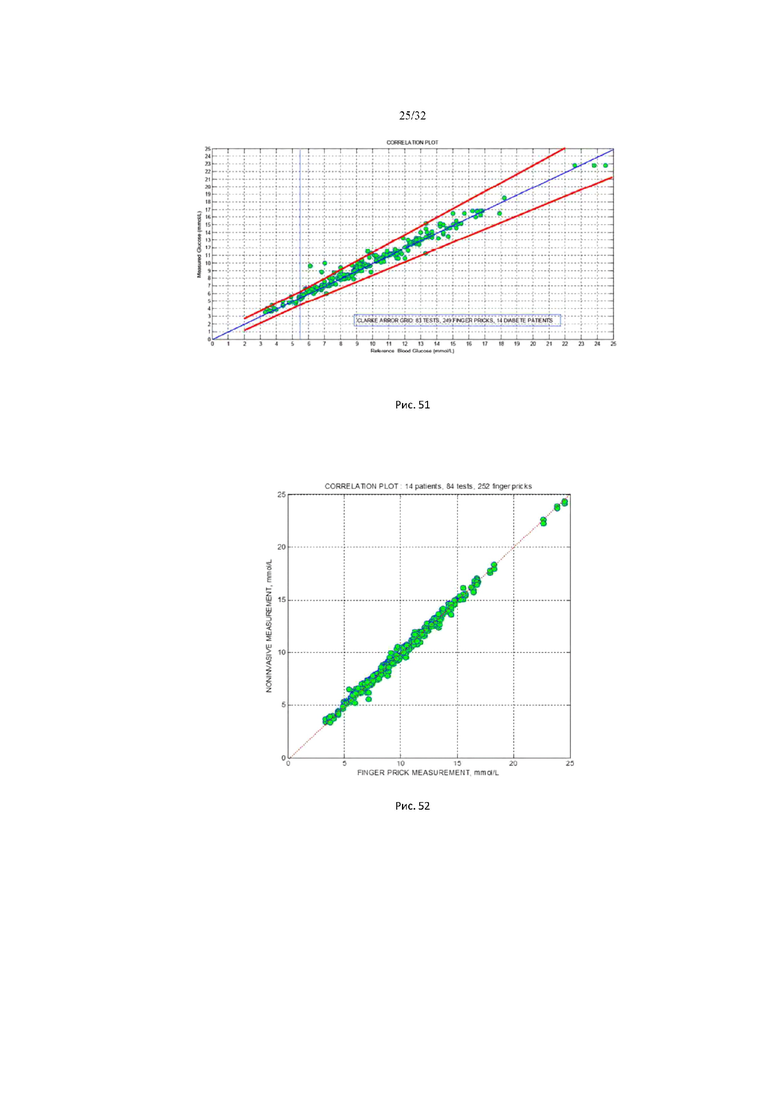
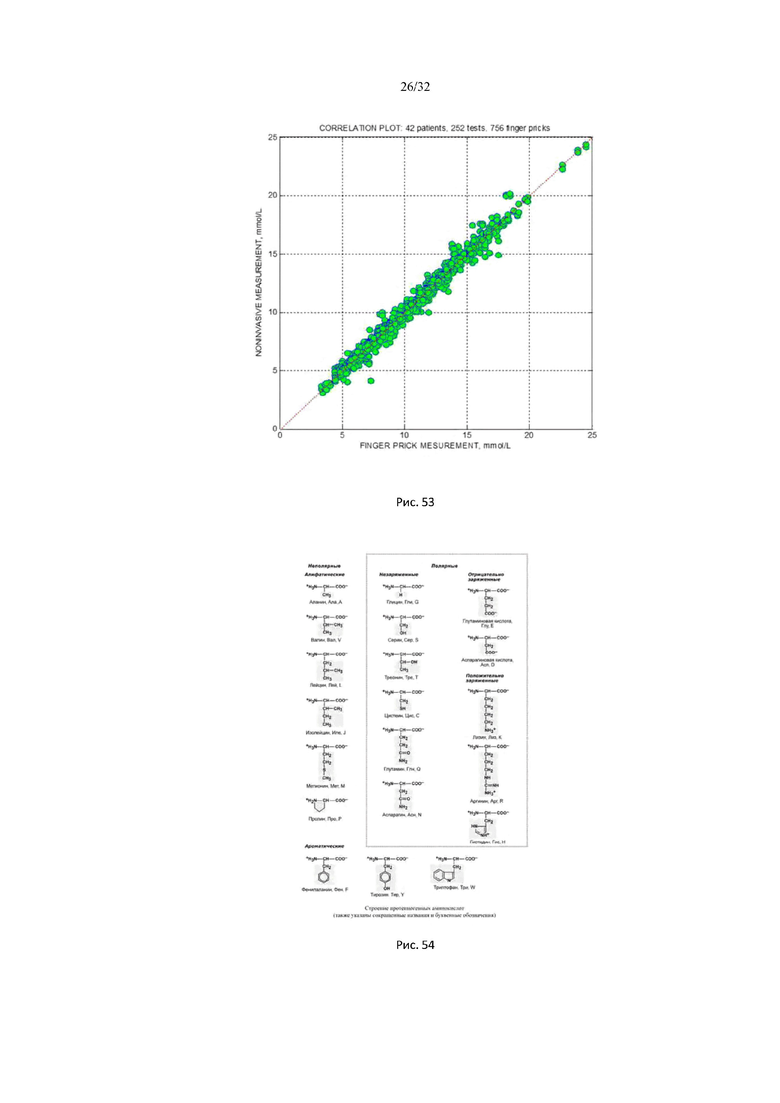

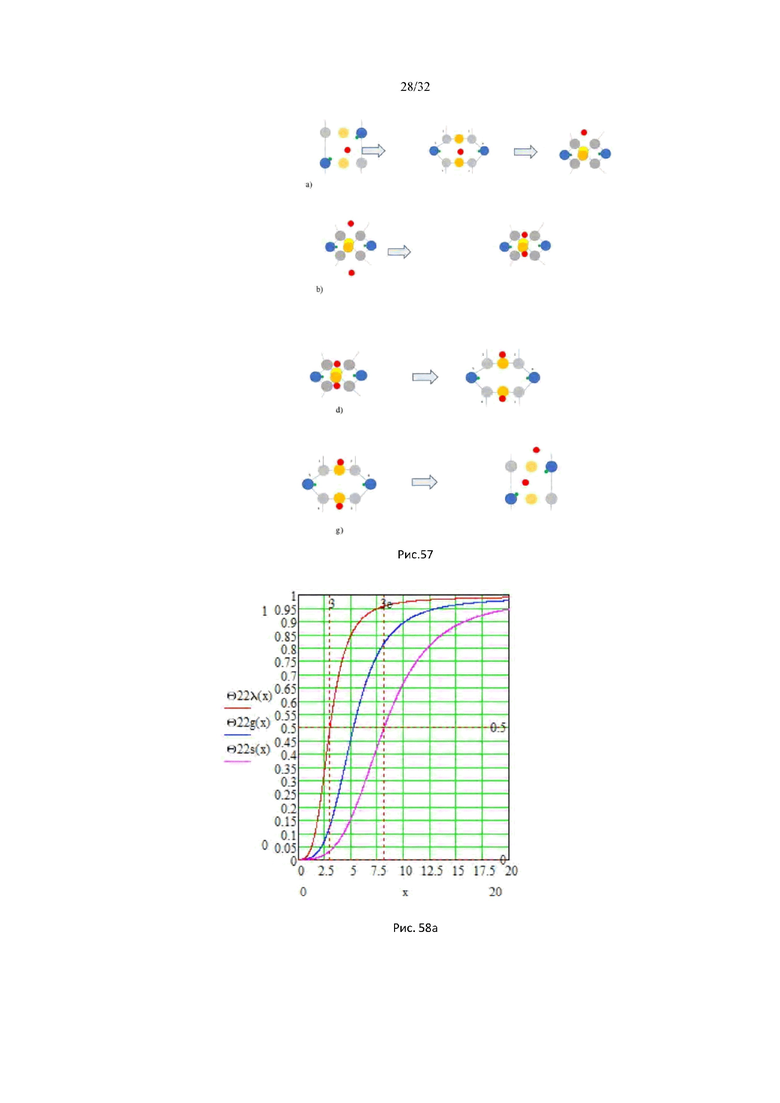
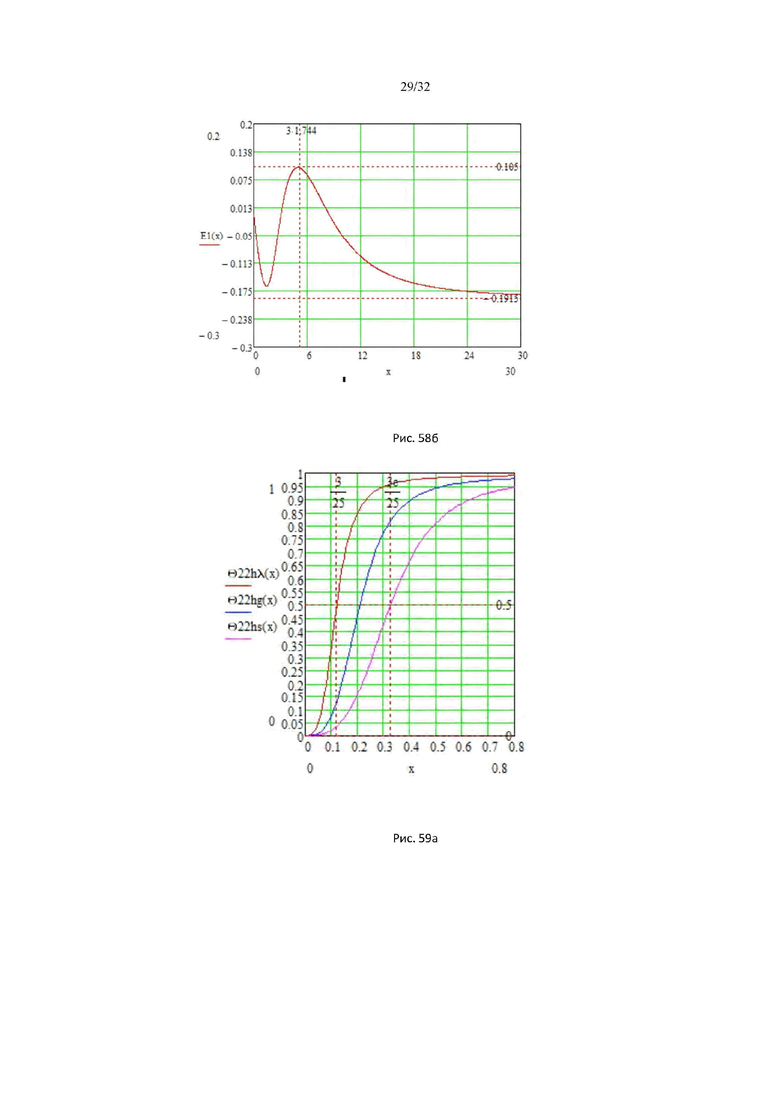
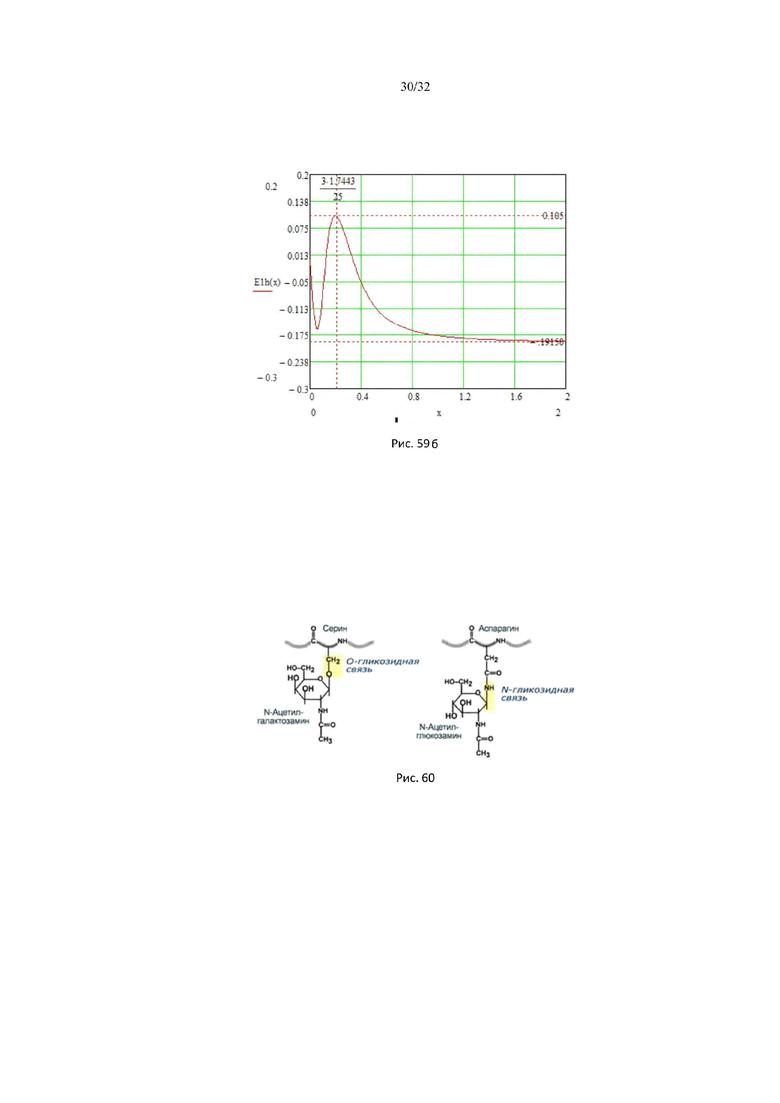
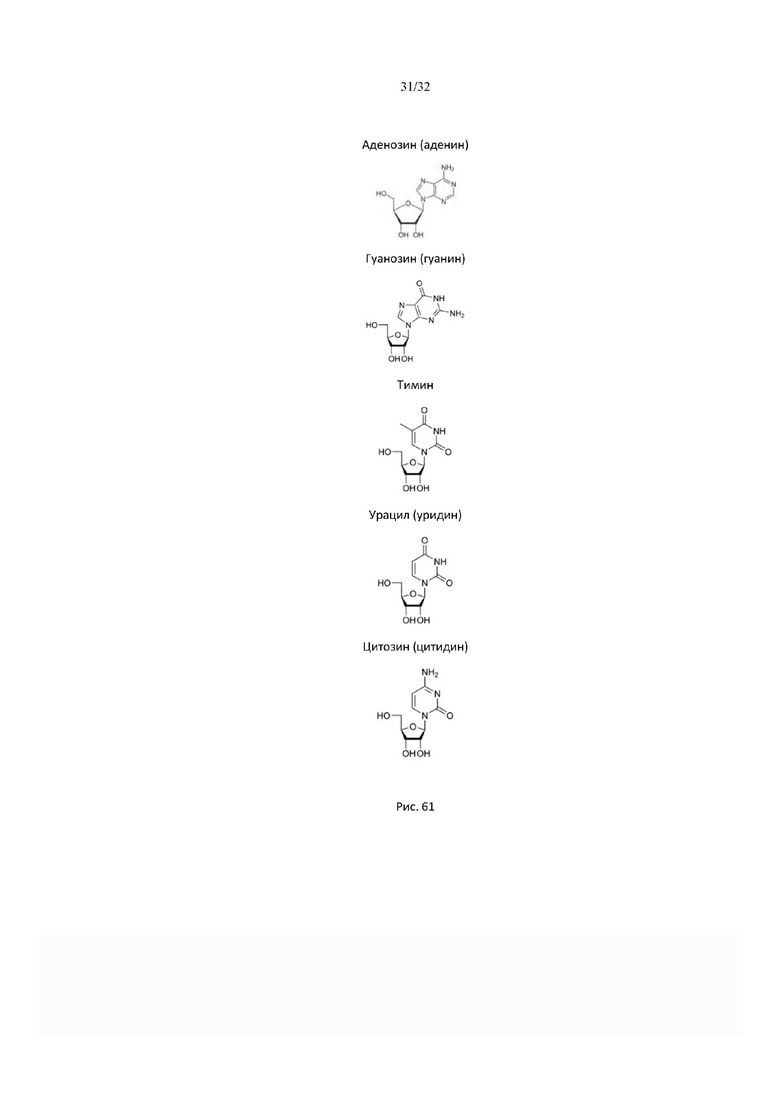
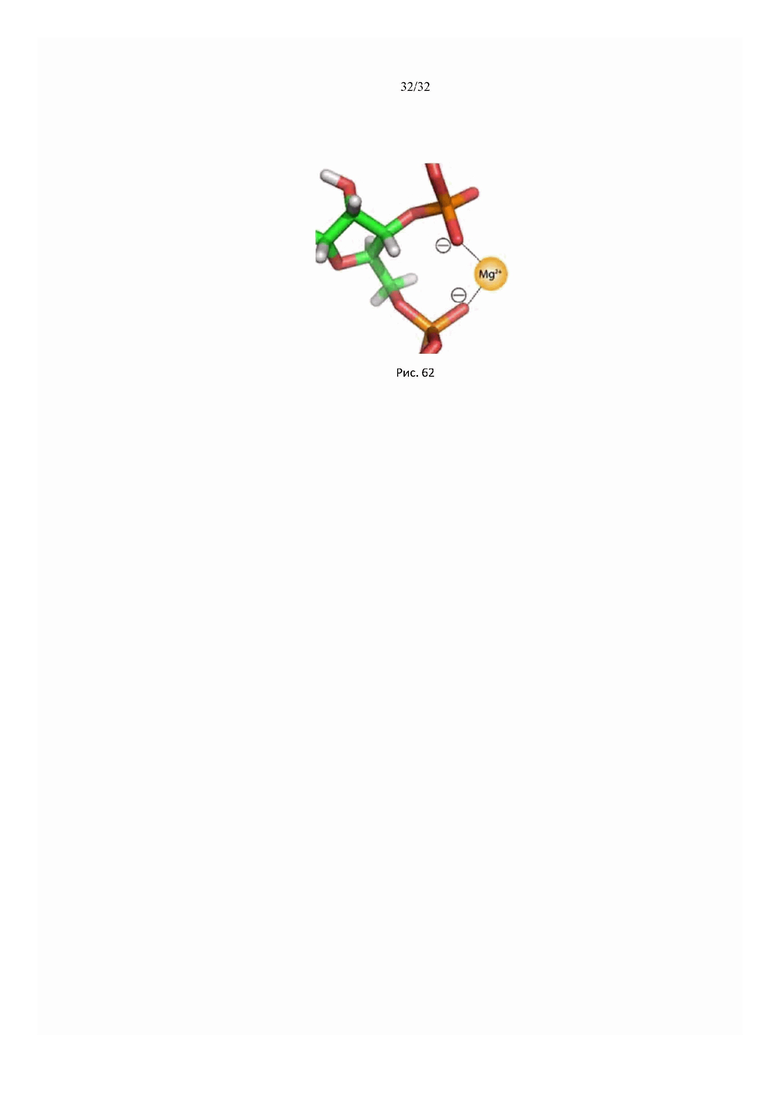
Группа изобретений относится к медицине и медицинской технике, а именно к способу и устройству для изотермической калориметрической спектроскопии биохимических компонентов живой ткани пациента. Способ изотермической калориметрической спектроскопии биохимических компонентов межклеточного и/или внутриклеточного вещества живой ткани пациента, выбранных из: воды, гиалуроновой кислоты, глюкозы, триглицеридов жирных кислот, заключается в том, что накладывают на поверхность кожи пациента с дозированным давлением по меньшей мере один тепло- и водонепроницаемый аппликатор, образующий закрытую систему в локальной области ткани под аппликатором. Оказывают локальное воздействие на участок ткани под аппликатором электромагнитным излучением на одной или нескольких длинах волн, соответствующих характеристическим частотам поглощения биохимических компонентов межклеточного и/или внутриклеточного вещества. Измеряют величину физиологического параметра, характеризующего термодинамическое фазовое состояние межклеточного вещества под аппликатором и его временную динамику, в зависимости от мощности падающего электромагнитного излучения. Определяют концентрацию биохимического компонента межклеточного и/или внутриклеточного вещества и его временную динамику на основе временной динамики измеренного физиологического параметра. Группа изобретений обеспечивает принципиально новый метод спектроскопии живой ткани в условия in vivo. 2 н. и 26 з.п. ф-лы, 62 ил.
1. Способ изотермической калориметрической спектроскопии биохимических компонентов межклеточного и/или внутриклеточного вещества живой ткани пациента, выбранных из: воды, гиалуроновой кислоты, глюкозы, триглицеридов жирных кислот, включающий следующие этапы:
- накладывают на поверхность кожи пациента с дозированным давлением по меньшей мере один тепло- и водонепроницаемый аппликатор, образующий закрытую систему в локальной области ткани под аппликатором;
- оказывают локальное воздействие на участок ткани под аппликатором электромагнитным излучением на одной или нескольких длинах волн, соответствующих характеристическим частотам поглощения биохимических компонентов межклеточного и/или внутриклеточного вещества;
- измеряют величину физиологического параметра, характеризующего термодинамическое фазовое состояние межклеточного вещества под аппликатором и его временную динамику, в зависимости от мощности падающего электромагнитного излучения;
- определяют концентрацию биохимического компонента межклеточного и/или внутриклеточного вещества и его временную динамику на основе временной динамики измеренного физиологического параметра.
2. Способ по п. 1, отличающийся тем, что интенсивность электромагнитного излучения может быть постоянной или переменной, изменяющейся с постоянной скоростью, и/или модулированной частотой, и/или амплитудой.
3. Способ по п. 1, отличающийся тем, что физиологическим параметром, характеризующим термодинамическое фазовое состояние межклеточного вещества, является осмотическое давление межклеточного вещества, и/или количество воды в межклеточном пространстве ткани, и/или эластическое давление живой ткани под аппликатором.
4. Способ по п. 1, отличающийся тем, что длину волны воздействующего на участок ткани электромагнитного излучения выбирают из диапазона электромагнитного излучения, который определяют по характеристическим частотам поглощения биохимических компонентов межклеточного и/или внутриклеточного вещества живой ткани в оптическом, и/или ближнем инфракрасном, и/или среднем и дальнем инфракрасном, и/или терагерцовом, и/или микроволновом диапазоне.
5. Способ по любому из пп. 1-4, отличающийся тем, что концентрацию биохимического компонента межклеточного и/или внутриклеточного вещества живой ткани дополнительно измеряют на основе метода спектроскопии диффузного отражения.
6. Способ по пп. 1-4, отличающийся тем, что концентрацию биохимического компонента межклеточного и/или внутриклеточного вещества живой ткани дополнительно измеряют на основе метода спектроскопии комбинационного рассеяния света.
7. Способ по п. 1, отличающийся тем, что биохимическим компонентом межклеточного и/или внутриклеточного вещества живой ткани является глюкоза в крови, концентрацию которой определяют по концентрации глюкозы, связанной с мономерами полимерной цепи межклеточного вещества.
8. Способ по п. 3, отличающийся тем, что осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют на основе спектральных измерений на длинах волн, соответствующих характеристическим частотам поглощения биохимических компонентов межклеточного и/или внутриклеточного вещества живой ткани.
9. Способ по п. 8, отличающийся тем, что спектральные измерения основаны на методе двухчастотной спектроскопии, в которой длина волны электромагнитного излучения выбирается по характеристической частоте тетрамера гиалуроновой кислоты межклеточного вещества в терагерцовой области, а длина волны спектрального датчика осмотического давления межклеточного вещества выбирается по характеристическим частотам поглощения воды в роговом слое эпидермиса в инфракрасной области.
10. Способ по п. 3, отличающийся тем, что осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса на основе измерения физических характеристик рогового слоя эпидермиса, которые выбирают из группы, включающей в себя электрофизические характеристики, спектральные и оптико-акустические характеристики, теплофизические характеристики.
11. Способ по п. 3, отличающийся тем, что осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса на основе измерений спектральных характеристик рогового слоя эпидермиса на длинах волн, соответствующих характеристическим частотам воды в роговом слое, с помощью спектрального метода, который выбирают из группы, включающей: ИК-спектроскопию, Раман-спектроскопию, оптико-акустическую спектроскопию, разностную двухлучевую спектроскопию.
12. Способ по п. 3, отличающийся тем, что осмотическое давление межклеточного вещества или количество воды в межклеточном пространстве определяют по количеству воды в роговом слое эпидермиса путем измерения электрических характеристик рогового слоя эпидермиса, которые выбирают из группы, включающей в себя поперечную электропроводность рогового слоя на постоянном и/или переменном токе и диэлектрическую проницаемость.
13. Способ по п. 7, отличающийся тем, что концентрацию глюкозы в крови определяют на основе временной динамики физиологического параметра, причем физиологическим параметром является осмотическое давление межклеточного вещества области ткани под аппликатором, причем указанное межклеточное вещество под аппликатором является природным биосенсором с гетерофазной структурой с селективностью к молекуле глюкозы и чувствительностью к внешним тепловым потокам, при этом индивидуальную калибровку локальной области ткани пациента определяют по содержанию гиалуроновой кислоты в межклеточном веществе и количества воды в объеме ткани под аппликатором, измеряемых спектральным методом.
14. Способ по п. 1, отличающийся тем, что способ применим для ранней диагностики сахарного диабета на основе временной динамики концентрации биохимического компонента.
15. Способ по п. 1, отличающийся тем, что способ применим для ранней диагностики сахарного диабета на основе временной динамики осмотического давления межклеточного вещества и/или другого физиологического параметра, характеризующего термодинамическое состояние межклеточного вещества.
16. Устройство для изотермической калориметрической спектроскопии биохимических компонентов межклеточного и/или внутриклеточного вещества живой ткани пациента, выбранных из: воды, гиалуроновой кислоты, глюкозы, триглицеридов жирных кислот, содержащее тепло- и водонепроницаемый аппликатор, выполненный с возможностью наложения на кожу пациента с дозированным давлением, датчик температуры, один или несколько датчиков физиологических параметров, характеризующих термодинамическое фазовое состояние межклеточного вещества под аппликатором, один или несколько источников электромагнитного излучения, устройство для создания калибровочного воздействия, при этом датчики физиологических параметров расположены под аппликатором, при этом сигналы с вышеуказанных датчиков поступают последовательно на входы блока усилителей, и/или синхронного детектора, и/или аналогово-цифрового преобразователя, установленных на верхней поверхности аппликатора, в блок обработки информации и блок отображения информации.
17. Устройство по п. 16, отличающееся тем, что источник электромагнитного излучения выполнен с возможностью излучения с постоянной и/или модулированной интенсивностью и/или частотной модуляцией.
18. Устройство по п. 16, отличающееся тем, что датчики физиологических параметров, характеризующие термодинамическое состояние межклеточного вещества под аппликатором, выполнены в виде датчиков осмотического давления межклеточного вещества или датчиков количества воды в межклеточном пространстве ткани в локальном объеме под аппликатором и/или датчика эластического давления живой ткани под аппликатором.
19. Устройство по п. 16, отличающееся тем, что устройство для создания калибровочного воздействия представляет собой источник тепловой мощности, выполненный в виде резистора и/или элемента Пельтье, и/или источник электрического тока и/или напряжения; устройство для создания дозированного давления на поверхность аппликатора.
20. Устройство по п. 16, отличающееся тем, что диапазон излучения источников электромагнитного излучения выбирают из группы, включающей в себя оптический и/или ближний инфракрасный; средний инфракрасный; дальний инфракрасный и/или терагерцовый; микроволновый диапазон излучения.
21. Устройство по п. 18, в котором датчик осмотического давления межклеточного вещества или датчик количества воды в межклеточном пространстве ткани в локальном объеме под аппликатором представляет собой электрометрический датчик, основанный на измерении электрофизических характеристик рогового слоя эпидермиса и ткани под аппликатором, принцип измерения которого выбирается из группы, включающей измерение поперечной электропроводности рогового слоя эпидермиса на постоянном и/или переменном токе; измерение диэлектрической проницаемости рогового слоя эпидермиса; измерение электропроводности и/или диэлектрической проницаемости ткани под аппликатором.
22. Устройство по п. 18, в котором датчиком осмотического давления межклеточного вещества или датчиком количества воды в межклеточном пространстве ткани в локальном объеме под аппликатором является спектральный датчик, основанный на спектральных измерениях характеристик живой ткани под аппликатором, содержащий источник и приемник электромагнитного излучения.
23. Устройство по п. 22, отличающееся тем, что длину волны электромагнитного излучения источника спектрального датчика определяют по характеристическим частотам поглощения биохимических компонентов межклеточного вещества и/или внутриклеточного вещества.
24. Устройство по п. 18, отличающееся тем, что датчиком осмотического давления межклеточного вещества является спектральный датчик, основанный на спектральных измерениях характеристик межклеточного вещества в терагерцовом диапазоне на длине волны, соответствующей энергии поперечной связи между мономерами полимерной цепи гиалуроновой кислоты.
25. Устройство по п. 22, отличающееся тем, что спектральный датчик основан на спектральных измерениях количества воды в роговом слое эпидермиса по характеристическим частотам воды в роговом слое на основе спектрального метода, который выбирают из группы, включающей в себя изотермическую калориметрическую спектроскопию, абсорбционную спектроскопию, спектроскопию диффузионного отражения, спектроскопию комбинационного рассеяния, оптико-акустическую спектроскопию.
26. Устройство по любому из пп. 16-25, отличающееся тем, что дополнительно содержит спектральное устройство, включающее источник и приемник электромагнитного излучения, для спектральных измерений на основе диффузионной отражательной спектроскопии.
27. Устройство по п. 16, отличающееся тем, что датчик физиологических параметров выполнен на пьезоэлектрическом способе измерения.
28. Устройство по п. 16, отличающееся тем, что устройство применимо для ранней диагностики сахарного диабета на основе временной динамики концентрации биохимического параметра, и/или величины осмотического давления межклеточного вещества, и/или другого физиологического параметра, характеризующего термодинамическое состояние межклеточного вещества.
| СПОСОБ И УСТРОЙСТВО ДЛЯ МИКРОКАЛОРИМЕТРИЧЕСКОГО ИЗМЕРЕНИЯ СКОРОСТИ ЛОКАЛЬНОГО МЕТАБОЛИЗМА ТКАНИ, СОДЕРЖАНИЯ ВОДЫ В МЕЖКЛЕТОЧНОЙ ТКАНИ, КОНЦЕНТРАЦИИ БИОХИМИЧЕСКИХ КОМПОНЕНТОВ КРОВИ И ДАВЛЕНИЯ В СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЕ | 2005 |
|
RU2396897C2 |
| СПОСОБ ОБРАБОТКИ ПОЧВЫ ПОД ДРЕВЕСНЫЕ КУЛЬТУРЫ | 1992 |
|
RU2091996C1 |
| ОЛЬХОВ А.А | |||
| и др | |||
| БИОПОЛИМЕРНЫЕ ВОЛОКНИСТЫЕ МАТЕРИАЛЫ С КОМПЛЕКСАМИ ПОРФИРИНА ДЛЯ МЕДИЦИНЫ / Физика волокнистых материалов: структура, свойства, наукоемкие технологии и материалы (SMARTEX-2017): сб | |||
| материалов ХX Междунар | |||
| науч.-практ | |||
| форума, 22-26 мая 2017 года, Иваново: ИВГПУ, 2017, стр | |||

