Изобретение относится к сельскому хозяйству, в частности к селекции и семеноводству ржи. В настоящее время в производстве используются сорта-популяции, сорта-синтетики и гибридные сорта. Резерв повышения урожайности существующих сортов в настоящее время заложен в использовании явления гетерозиса. В тоже время все современные сорта-популяции и сорта-синтетики имеют существенный общий недостаток - общность генофонда, в связи с этим использование гетерозиса в работе с данными группами сортов малоэффективно. Наиболее полное проявление гетерозиса наблюдается в гибридных сортах (гибридах).
Для создания коммерческих гибридных сортов озимой ржи в качестве исходного материала необходимы гомозиготные линии.
Известен ряд способов получения гомозиготных линий у ржи.
Первый способ - это проведение самоопыления (инбридинга) родоначальных растений в ряде поколений. Имеется два основных недостатка этого метода: наличие у большинства растений ржи генетического механизма самонесовместимости и длительный период создания гомозиготных линий (обычно для этого требуется 6-8, а иногда до 12 и более лет), причем даже при длительном инбридинге не удается добиться полной гомозиготности - та или иная степень гетерозиготности сохраняется. Кроме того, у огромного числа потомств инцухт-линий с каждым циклом инбридинга наблюдается возрастание инцухт-депрессии (вследствие перехода ряда сублетальных, летальных и других рецессивных генов в гомозиготное состояние). Это происходит до тех пор, пока не наступит инбредный минимум. В результате значительная часть первоначального исходного материала теряется в ходе селекции [1].
Известен способ получения гомозиготных линий методом гаплопродьюсера, основанным на использовании скрещивания межвидовых скрещиваний, элиминации хромосом продьюсера и последующего колхицинирования [1].
Недостатками данного способа является трудоемкая технология получения, использования дорогостоящего оборудования и реактивов и крайне низкая эффективность, а также необходимость высокой квалификации сотрудников.
Наиболее близким является способ, основанный на применении культуры пыльников [2, 3], который предусматривает получение гаплоидных растений-регенерантов из пыльников, удвоение их набора хромосом и выращивание фертильных растений, однако данный способ также является трудоемким, требует дорогостоящего оборудования и реактивов, а также работы квалифицированного персонала, при этом также следует отметить низкую его эффективность.
Целью изобретения является повышение выхода гомозиготных линий при упрощении технологии их получения и сокращении сроков создания.
В основу предлагаемого способа положено явление полиэмбрионии. Сущность способа состоит в следующем.
1. Из проращиваемых семян озимой ржи визуально выделяют проростки-близнецы.
2. Сразу после выделения проростки колхицинируют, а затем растения (Со) высаживают в почву (в горшочки для рассады) и помещают в прохладное место на рассеянный свет (до 2 сут). Затем температуру воздуха и освещенность доводят до нормы.
3. В фазе 2-3-х листьев проводят разделение растений близнецов. Растения вновь высаживают в вегетационные сосуды. После того, как растения раскрутятся, проводят клонирование в несколько этапов (по мере того как образуются побеги). По каждому клону (клон - это потомство вегетативно размноженного растения) желательно иметь не менее 10 частей растения.
Клонирование проводят в теплице или поле на специальном участке.
4. Яровизацию клонов проводят или в климокамере, или в естественных условиях (в случае работы с озимыми формами ржи).
5. В фазу выколашивания проводят цитологический анализ материнских клеток пыльцы на стадии мейоза. Анализировать желательно каждое растение каждого клона.
6. Диплоидизированные гаплоиды подвергают самоопылению (JO).
7. В каждом клоне с растений обмолачивают семена (C1J1) отдельно.
Выход растений с полиэмбрионией можно увеличить, используя потомства растений-близнецов.
Настоящий способ позволяет получить исходные линии у различных форм ржи, обладающих как самофертильностью, так и самонесовместимостью.
Существуют несколько приемов преодоления самонесовместимости:
- тепловой шок;
- внутриклоновое переопыление;
- использование пыльцы разного возраста.
Контроль гомозиготности полученных линий проводят цитологическими и биохимическими методами, а также оценкой по потомству.
Сохранность полученных линий обеспечивают микроклонированием, изолированным пересевом и длительным хранением семян.
Экспериментальное подтверждение метода растений-близнецов для получения гомозиготных диплоидизированных гаплоидных линий с целью использования их при создании гетерозисных гибридных сортов озимой ржи.
Основные положения:
1) у озимой ржи существуют растения-близнецы;
2) среди таких растений встречаются гаплоидные генотипы (n набор хромосом);
3) среди сортов озимой ржи имеются различия как по частоте встречаемости растений-близнецов, так и по частоте встречаемости гаплоидов среди растений-близнецов. Подобное утверждение справедливо и в отношении семей любого сорта озимой ржи;
4) из гаплоидов путем колхицинирования возможно создать диплоидизированные гаплоидные растения, а затем и самоопыленные линии.
Из 19 сортообразцов озимой ржи (районированные сорта и селекционные номера) у одного не было найдено близнецов; у девяти сортообразцов было найдено от 1 до 2 растений-близнецов; у двух - от 5 до 6 близнеца; у двух - по 16 близнецов; у двух - по 21 близнецу; у одного - 39 близнеца и у двух сортообразцов - от 49 до 51 растения-близнеца.
Таким образом, у 18 сортообразцов ржи частота встречаемости растений-близнецов варьировала от 0,007 до 3,0%.
У одного гибридной популяции озимой ржи (селекционный номер 1460/86) было обследована 91 семя (по 565 проростков). Выявлено, что в 72 семьях не обнаружено растений близнецов; в 9-ти семьях найдено по 1 близнецу (или 0,18% от числа проростков); в 3-х семьях - по 2 близнеца (0,35%); в 4-х семьях - по 3 близнеца (0,53%); еще в трех семьях - соответственно 4 близнеца (0,71%), 6 близнецов (1,06%) и 12 близнецов (2,12%).
Всего в 1990 г в октябре месяце было просмотрено 15 кг семян ржи (600.000 - 700.000 зерен). При этом найдено 310 зерен, которые дали растения-близнецы (0,05% от числа всех изученных зерен). Таким образом из каждых 10.000 зерен было найдено 1 зерно, обладающее полиэмбрионией.
Растения-близнецы были разделены и высажены в грунт теплицы. После укоренения выжило 436 растений-близнецов разной плоидности (или 70,3% от числа всех генотипов-близнецов). Каждый генотип был расклонирован на 3-6-8-10 или 12 частей, в зависимости от степени кущения. По морфотипу - мелкие строения органов, листьев, побегов - было выделено 186 клонов (а клон - это потомство вегетативно размноженного растения). Это составило 42,7% от числа всех клонов).
Цитологический анализ растений показал, что они имеют гаплоидный набор хромосом. В январе все растения клонов были проколхицинированы и высажены вновь на укоренение в теплицу. Выжило 85 клонов (45,7% от числа обработанных колхицином). Колхицинирование растений ржи проводили в фазу кущения (растения погружались в раствор колхицина целиком, перед этим у них на 2/3 длины обрезали листья).
Режим колхицинирования: 7 ч экспозиции в 0,15%-ном растворе колхицина с добавкой 2%-димитилсульфоксида и 0,3%-ного раствора папаина; 3-5 ч промывки в проточной водопроводной воде; высадка в почву в прохладное (15...18оС) затенение (до 2 сут) место с относительной влажностью воздуха 60-80%.
В феврале растения всех клонов (по 3-12 растений в клоне) были посажены в ящики с почвой и помещены в яровизационную камеру (4оС) на 65 дней. В мае месяце растения были пересажены в поле.
При принудительном самоопылении в полевых условиях растения 41 клона (48,2 от числа прояpовизированных) завязали зерно (J1). Получено 1-159 зерен на клон (0,04-12,0 зерен на каждый колос растения в клоне). Из этого количества часть растений 12 клонов (или 29,3% от числа озерненных клонов) обладали хорошей озерненностью колоса. Надо отметить, что как отдельные растения, так и часть колосьев у отдельных растений оказались не озерненными.
В конце лета семена от 41 клона (линии) были высеяны в поле. Семена 35 линий (или 85,4% от числа высеянных линий) оказались всхожими. Всхожесть составляла 8,7-100% по каждой линии. Перезимовали все 35 линий. Перезимовка варьировала от 42,9 до 100%.
У 8 диплоидизированных линий по таким признакам, как форма куста, высота растений, характер выколашивания и форма колоса, поражаемость болезнями все растения были выравненными. Таким образом, косвенно эти линии можно характеризовать как гомозиготными.
В ходе эксперимента, из 15 кг семян ржи (600-700 тыс.) было найдено 310 зерен, несущих в себе полиэмбрионию и давших проростки-близнецы. Из этих растений было получено 35 диплоидизированных гаплоидных линий (или 11,1% от всех полиэмбриональных семян, или 0,006% от всего количества проанализированных зерен). Следовательно из каждых 100.000 зерен озимой ржи можно найти гаплоидное растение-близнец и получить из него диплоидизированную гомозиготную фертильную линию.
Исследования показали, что у удвоенных гаплоидов озимой ржи можно получить семена. Следовательно, полученные линии можно сохранять и размножать. Однако для сохранения гомозиготности линии необходимо применять инбридинг. Теоретически, если в генотипе у диплоидизированных растений линии есть ген самофертильности, то проблемы с завязываемостью семян не должно быть. Однако если такого гена нет, тогда надо воспользоваться методом внутриклонового переопыления растений, а также повышением температуры воздуха до 30оС в период цветения, и таким образом преодолеть механизм самонесовместимости (т. е. возможно явление псевдосамофертильности). В дальнейшем, диплоидизированные гаплоидные гомозиготные линии необходимо размножать при строгой пространственной изоляции, или же изолировать их под "колпаками" от чужеродной пыльцы. При этом в переопылении внутри линии участвуют все растения этой линии (в этом случае идет как бы "перекрестное" скрещивание и наличие или отсутствие в генотипе гена самофертильности теоретически не должно играть роль). У диплоидизированной гомозиготной линии расщепления по какому-либо признаку было не должно (так как при создании подобной линии в результате колхицинирования гаплоидного организма все аллели - и доминантные и рецессивные - переходят в гомозиготное состояние). Изменение гомозиготности и появление гетерозигот может быть вызвано только мутациями или гибридизацией.
Полученные настоящим способом линии предназначены для получения семян гибридов первого поколения с использованием гаметоцидов, а также при различных селекционно-генетических исследованиях.
Возможность использования в практике селекции полученных линий показана ниже:
Краткий цикл получения коммерческих семян гибридов первого поколения озимой ржи:
- 1-й год - поиск растений близнецов и получение гомозиготных дигаплоидных линий;
- 2 и 3-й годы - испытание и размножение дигаплоидных линий;
- 4 - 8-й годы - получение первых гетерозисных гибридов (с применением гаметоцидов) и их испытание;
- 9-й и последующие годы - получение семян (F1) коммерческих гибридов и их реализация. Организация первого семеноводства родительских компонентов.
Расширенная схема селекции гибридных (коммерческих) сортов озимой ржи.
Схема селекции опирается на применение гомозиготных линий озимой ржи, обладающих высокой общей специфической комбинационной способностью (ОКС и СКС). Линии получают по методу "близнецов". Гибридизация осуществляется при помощи гаметоцидов.
Схематично процесс показан на фиг.1-7.
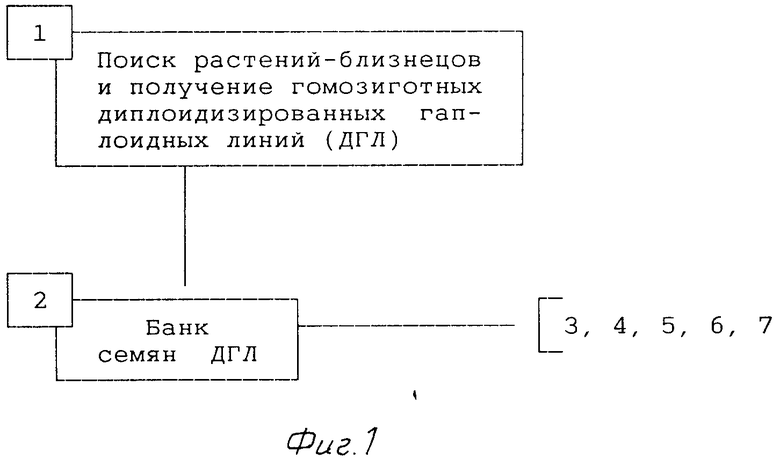
1-й год (фиг.1).
1. При поиске растений-близнецов и получении диплоидизированных гаплоидных линий (ДГЛ) (позиция 1) проводят следующие операции:
- цитологический анализ плоидности растений-близнецов;
- клонирование гаплоидных растений;
- клонирование;
- укоренение;
- яровизация;
- принудительное самоопыление;
- тест на гомозиготность (электрофорез семян J1).
2. В банк хранения (позиция 2) первоначально закладывают 1/4 часть семян линий. В дальнейшем количестве семян, закладывающихся в резерв, может быть увеличено.
Позиции 1 и 2 проводят ежегодно.
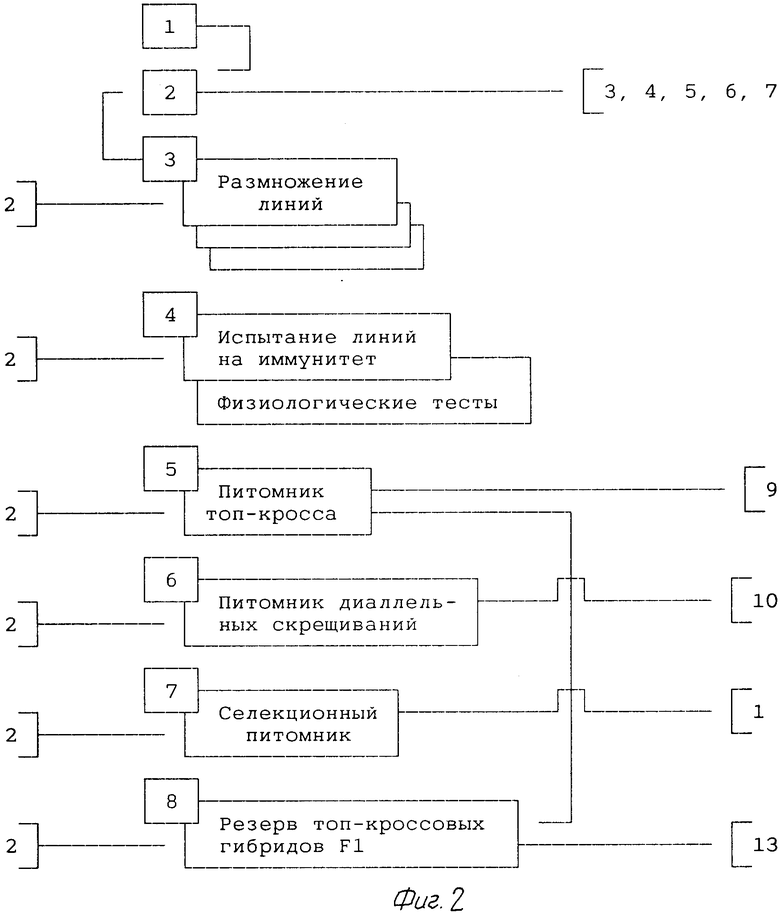
2-й год (фиг.2).
1. При размножении линий (позиция 3) проводят контроль гомозиготности по морфологическим, физиологическим и биохимическим признакам. Электрофорез проводят на 1-2 зернах с каждого растения.
Размножение проводят самоопылением растений под индивидуальными изоляторами (при наличии системы самофертильности), переопылением нескольких растений ДГЛ под общим изолятором или выращиванием ДГЛ на пространственно-изолированных участках.
2. Испытание ДГЛ на иммунитет (позиция 4) проводят в лабораторных и полевых условиях на искусственно инфицированных фонах. Для теста по каждой болезни высевают 5 зерен ДГЛ.
3. Для определение комбинационной способности закладывают питомник топ-кросса (позиция 5) или питомник диаллельных скрещиваний (позиция 6).
В питомнике топ-кросса (позиция 5) предусматривают 2 тестера: лучшая по продуктивности ДГЛ и районированный сорт или смесь семян всех испытываемых ДГЛ.
Часть семян топ-кроссовых гибридов оставляют в резерве (позиция 8). Остальная часть семян идет на следующий год в питомник испытания топ-кроссовых гибридов (позиция 9).
Из питомника диаллельных скрещиваний (позиция 6) семена диаллельных гибридов F1 поступают на следующий год в питомник испытаний (позиция 10).
4. Для оценки продуктивности ДГЛ закладывают селекционный питомник (позиция 7). Для посева требуется 160 зерен от каждой линии на трехрядковую делянку длиной 1 м. При оценке определяют массу 1000 зерен, продуктивность колоса, выровненность колосьев и другие хозяйственно-ценные признаки, такие как число падения и удельная масса зерна, различные биохимические показатели. Дают иммунологическую характеристику линий на естественном фоне. По результатам этой оценки из семян, находящихся в банке хранения (позиция 2), формируют популяции синтетик и на следующий год закладывают питомник переопыления и первый цикл отбора лучших элит (позиция 11). Общая масса семян переопыленных растений всех линий в селекционном питомнике (позиция 7) может служить новым источником растений-близнецов.
Позиция 3, 4, 5, 6 и 7 проводят в последующем ежегодно. Семена для этих позиций берут из банка семян линий (позиция 2).
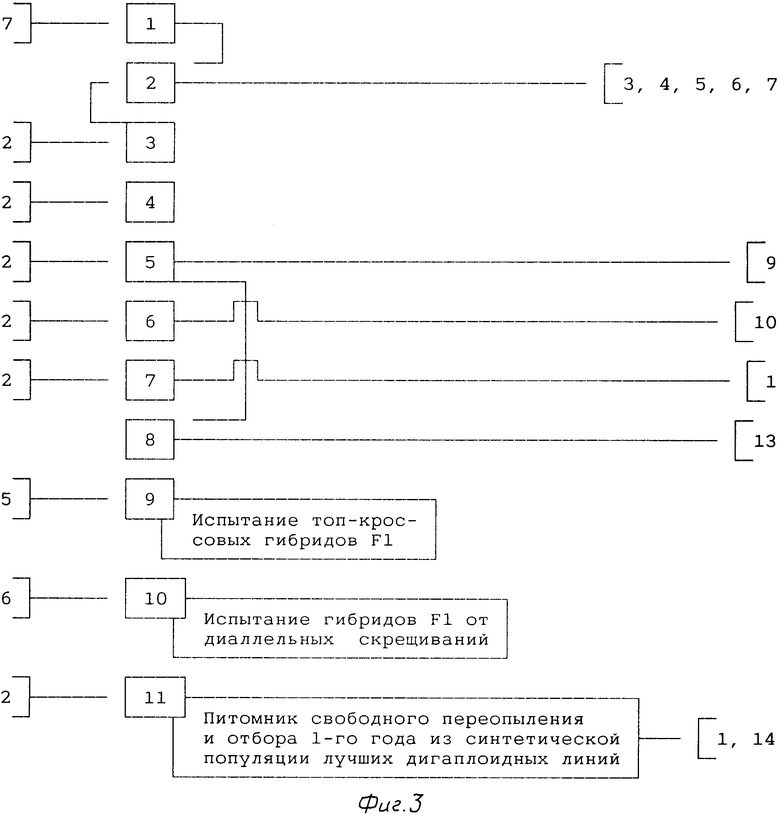
3-й год (фиг.3).
1. Для оценки общей комбинационной способности (ОКС) и специфической комбинационной способности (СКС) закладывают питомник испытания топ-кроссовых гибридов F1 (позиция 9) и питомник испытания гибридов F1 от диаллельных скрещиваний (позиция 10). При этом оба вида питомников желательно закладывать в нескольких местностях (для выявления экологического влияния на урожайность гибридов F1).
2. Синтетическая популяция из лучших по продуктивности ДГЛ (позиция 11) может служить источником новых растений-близнецов. В этом питомнике проводят свободное переопыление и отбор элитных растений. Семена для закладки позиции 11 берут из банка семян линий (позиция 2).
Позиции 9, 10 и 11 проводят ежегодно.
4-й год (фиг.4).
1. По результатам испытаний гибридов F1 от диаллельных скрещиваний и топ-кросса выявляют линии, которые могут служить материнскими или отцовскими компонентами для скрещивания.
Для получения перспективных гибридов F1 закладывают несколько питомников гибридизации (позиция 2). Питомники гибридизации должны быть пространственно изолированы друг от друга и от любых других посевов ржи. Семена для закладки питомников гибридизации берут из банка хранения (позиция 2).
2. На основе результатов испытаний, из лучших топ-кроссовых гибридов F1 формируют популяцию-синтетик и закладывают питомник отбора (позиция 13). Семена для этой цели берут из резерва топ-кроссовых гибридов F1 (позиция 8). Сформированная популяция-синтетик может служить базой для поиска новых растений-близнецов.
3. Из лучших элитных растений синтетической популяции (позиция 11) закладывают питомник свободного переопыления 2-го года (позиция 14). В дальнейшем реккуpентная популяция может служить новым источником растений-близнецов, а также перспективным сортом-синтетиком.
Позиции 12, 13 и 14 проводят ежегодно.
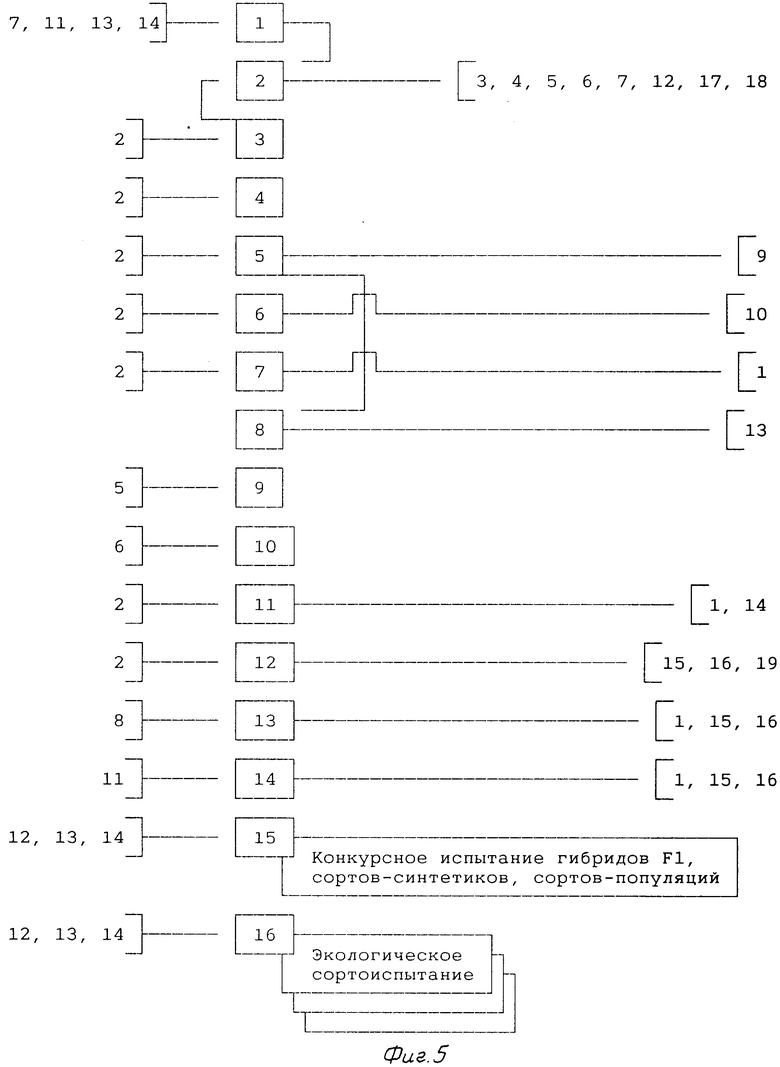
5-й и 6-й годы (фиг.5).
1. Закладывают питомник конкурсного испытания гибридов F1, полученных при помощи гаметоцидов, сортов-синтетиков и сортов-популяций в сравнении со стандартным районированным сортом (позиция 15).
2. Закладывают экологическое сортоиспытание перспективных и районированных гибридных сортов, сортов-синтетиков и сортов-популяций (позиция 16). Экологическое сортоиспытание закладывают в различных почвенно-климатическим условиям местностях.
Семена для закладки конкурсного и экологического сортоиспытаний берут из питомника гибридизации (позиция 12), питомника отбора 1-го года (позиция 13) и питомника свободного переопыления 2-го кода (позиция 14).
3. Родительские компоненты перспективных гибридных сортов должны быть размножены в достаточном количестве на пространственно-изолированных участках (позиция 3).
Позиции 15 и 16 проводят ежегодно.
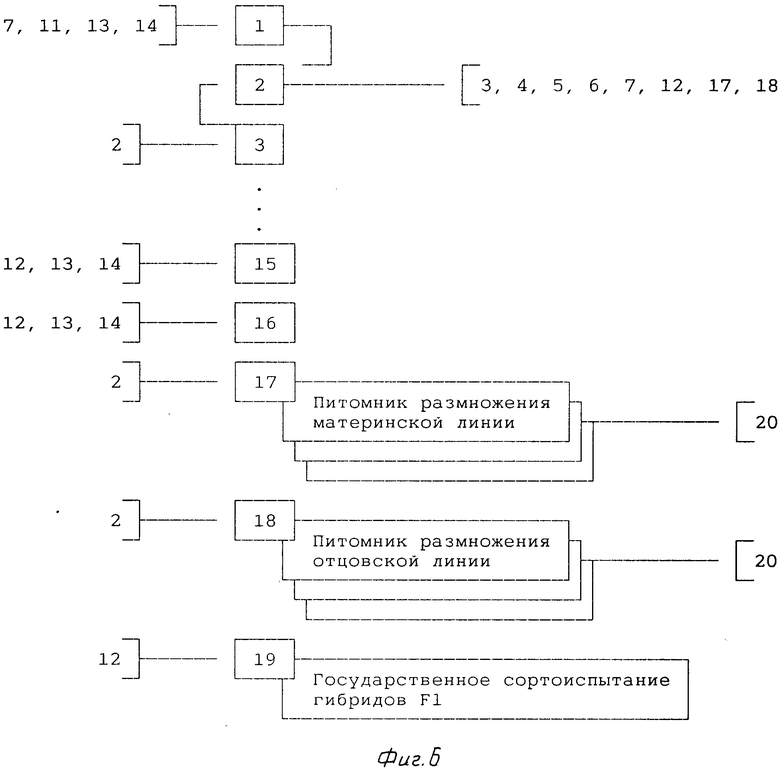
7-й и 8-й годы (фиг.6).
1. По результатам конкурсного и экологического сортоиспытаний перспективные гибриды F1 передают в государственное сортоиспытание (позиция 19).
2. Одновременно с передачей перспективных гибридов F1 в государственное сортоиспытание закладывают несколько питомников размножения pодительских компонентов этих гибридов (позиции 17 и 18). Семена для этого берут в банке семян линий (позиция 2).
Позиции 17, 18 и 19 в дальнейшем проводят по мере необходимости.
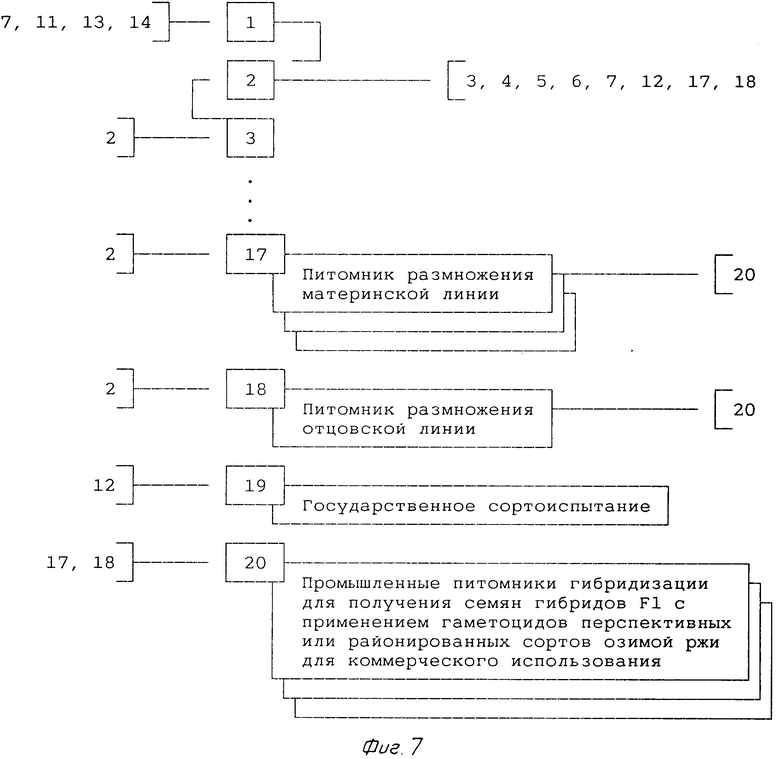
9-й год (фиг.7)
По предварительных результатах государственного сортоиспытания (1-2-х летние данные) закладывают промышленные питомники гибридизации с применением гаметоцидов (позиция 2) для получения семян F1 (перспективного или районированного) гибридного сорта с целью накопления запаса семян F1 для последующей реализации. Семена компонентов берут из питомников размножения этих линий (позиции 17 и 18).
В дальнейшем позиции 17, 18, 19 и 20 проводят ежегодно, пока гибридный сорт находится в районировании и пользуется спросом.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СОЗДАНИЯ ГОМОЗИГОТНЫХ ЛИНИЙ РЖИ | 2013 |
|
RU2551313C2 |
| СПОСОБ СОЗДАНИЯ РАЗДЕЛЬНОПЛОДНЫХ МУЖСКО-СТЕРИЛЬНЫХ ГИБРИДОВ САХАРНОЙ СВЕКЛЫ | 1996 |
|
RU2107430C1 |
| СПОСОБ СОЗДАНИЯ ФОРМ СОИ С ИЗМЕНЕННЫМ ЖИРНО-КИСЛОТНЫМ СОСТАВОМ МАСЛА | 2002 |
|
RU2215406C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГРЕССИВНЫХ РАСТЕНИЙ ОЗИМОЙ РЖИ | 2000 |
|
RU2197815C2 |
| Способ получения аналогов тетраплоидной ржи | 1991 |
|
SU1807842A3 |
| Способ отбора родоначальных растений озимой ржи | 1988 |
|
SU1577727A1 |
| Способ оценки селекционного материала перекрестно опыляющихся многолетних культур | 1984 |
|
SU1259981A1 |
| Способ повышения плодовитости аутотетраплоидной ржи | 1986 |
|
SU1431716A1 |
| СПОСОБ ОТБОРА ВЫСОКОПРОДУКТИВНЫХ ФОРМ ОЗИМОЙ РЖИ ПРИ СЕЛЕКЦИИ НА ЗАСУХОУСТОЙЧИВОСТЬ | 2008 |
|
RU2368132C1 |
| СПОСОБ СОЗДАНИЯ ИСХОДНОГО МАТЕРИАЛА ДЛЯ СЕЛЕКЦИИ ОЗИМЫХ МЯГКИХ ПШЕНИЦ | 1999 |
|
RU2150821C1 |

Использование: биотехнология, сельское хозяйство, селекционно-генетические исследования, семеноводство, растениеводство. Сущность изобретения: гомозиготные линии ржи получают путем выращивания гаплоидных растений, удвоения их набора хромосом, выращивания их до фазы цветения, принудительного самоопыления, получения семян гомозиготных линий и контроля гомозиготности, при этом выращиваемые гаплоидные растения получают из полиэмбриональных семян путем проращивания и разделения близнецовых проростков. 4 з.п. ф-лы, 7 ил.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Wenzel G | |||
| et al.: Theor | |||
| and Appl | |||
| Genet., 1977, v.51, n.2, p.81-86. | |||
