Изобретение относится к области сельского хозяйства, в частности селекции озимой ржи, и может быть использовано в селекции этой и других перекрестноопыляющихся злаковых культур на продуктивность и адаптивность с признаками пониженной пыльцевой продуктивности (экономичного расходования пластических веществ) и элементов продуктивности компенсаторного действия.
Известен способ повышения продуктивности гибридов первого поколения путем сбалансированного метаболизма формирования гетерозисного эффекта, контролируемого ядерными и цитоплазматическими детерминантами и их межгеномным взаимодействием [17] . Недостаток способа в том, что не конкретизируются методы подбора пар при гибридизации, а также критерии отбора растений из гибридной популяции.
Известен способ накопления сухой массы зерна колосом злаковых растений в экстремальных по засухе условиях не только за счет вклада ассимилятов текущего фотосинтеза, но и использованием запасных, а также реутилизацией структурных безазотистых веществ [3]. Однако критические климатические условия во время вегетации растений способствуют преждевременному усыханию растений, их частичной гибели и, как результат, резкому падению продуктивности. Поэтому необходима селекция генотипов, устойчивых к засухе, способных постоянно депонировать ассимиляты в различных органах растения и экономно реутилизировать их на налив зерна.
Известен способ повышения продуктивности колоса активизацией оттока пластических веществ из запасающих органов растений посредством увеличения числа "аттрагирующих" центров [13]. При этом ускоренный отток пластических веществ к репродуктивным органам вызывает быстрое старение и отмирание органов растения [16] в очередности лист, стебель, корень, чешуи колоса [4]. К тому же, увеличение числа "аттрагирующих" центров не всегда сопровождается эффективным использованием пластических веществ и ростом продуктивности колоса.
Известен способ повышения продуктивности растения использованием компенсационных корреляций, проявляющихся в отрицательной связи между размером и числом одноименных органов [8]. Климатические условия, уровень минерального питания по разному сказываются на росте и развитии растения, особенно в отдельные фазы, приводя часто к нарушению взаимосвязи, например, между числом зерен в колосе и их крупностью и снижению урожайности. В этой связи необходим поиск эффективных гибридных комбинаций, отдельных элементов растения, увеличение которых не способствует значительному снижению других.
Известен способ отбора в поколениях гетерозисных гибридов пшеницы трансгрессивных растений, гомозиготных по доминантным признакам продуктивности суммирующего действия наследуемых от разных родителей ([12] - прототип).
Недостаток прототипа в том, что при подборе родительских форм для гибридизации не учитывается плазменный контроль адаптивности у материнской формы. Не задействованы источники пониженной пыльцевой продуктивности, хотя для пшеницы их роль значительно меньше, чем для ржи. Отбор трансгрессивных растений проводился в межсортовых гибридах без анализа величины гетерозиса первого поколения и элементов продуктивности F2 компенсаторного действия.
Известно, что пыльники двух нижних цветков средних колосков, выполняя ингибирующую роль, являются аттрагирующими центрами первого порядка, их удаление до цветения способствует повышению озерненности колоска пшеницы и массы зерна с колоса в целом [13]. Способ трудоемкий, не обеспечивает широкого спектра формообразования, проблематичен отпор ядерно-плазменных трансгрессий.
Ветроопыляемые культуры, к которым относится и рожь, формируют огромное количество пыльцы, затрачивая на это гораздо больше, чем самоопылители, пластических веществ (Cruden, 1977), например один гектар посевов райграса, если нет скашивания, за сезон цветения дает 210 кг пыльцы [6]. Тогда как оптимальное количество пыльцы, необходимое для оплодотворения одной яйцеклетки у культурных растений, составляет около 200 пыльцевых зерен. Приспособленность ветроопыляемых растений к опылению выражена редукцией частей цветка, увеличением продуктивности пыльцы и величины рылец, поэтому оптимизация соотношения этих элементов должна способствовать повышению эффективности расходования пластических веществ.
Известен способ повышения урожая кукурузы путем уменьшения расхода питательных веществ на формирование пыльцы и структурных элементов метелки, а именно за счет недоразвития метелок с меньшим числом веточек, наблюдаемое у инбредных гомозиготных по lg1 и lg2 линий [15]. Способ не обеспечивает генетическое разнообразие, но факт уменьшения пыльцевой продуктивности линий без снижения их урожая заслуживает внимания. Кукуруза - раздельнополый злак, частичная редукция метелки может быть не связана с повышением продуктивности початка.
Аттрагирующим центром первого порядка у дикой многолетней ржи является узел кущения в отличие от культурной, у которой аттрагирующие центры первого порядка с момента выхода растений в трубку перемещаются от узла кущения в колос [7]. Очевидно, эволюционное становление онтогенеза культурной однолетней ржи связано со сменой аттрагирующих центров от узла кущения к колосу и предпосылкой этому может быть генетическая гомозиготная детерминация формирования крупных пыльников с большим количеством пыльцы. Вероятно, именно этот признак ржи возможно ослабить в пользу улучшения использования пластических веществ и повышения коэффициента хозяйственной годности создаваемых сортов.
В создании самофертильных линий для селекции гибридной ржи широко используются в качестве одной из родительских форм дикие виды с мелкими пыльниками и склонностью к самоопылению, в частности S. vavilovii (Grossh), S. iranicum (Kobyl. ). Высокая самофертильность получаемых гибридов сопровождается достаточно хорошей пыльцевой продуктивностью, чтобы обеспечить перекрестное опыление [14].
Оптимальное сочетание перекрестного опыления и самоопыления обеспечивает гарантированное оплодотворение яйцеклетки меньшим числом пыльцевых зерен при одновременном снижении расхода пластических веществ на формирование пыльника и пыльцы.
Клейстогамия относится к одному из компенсаторных механизмов экономичного расходования ассимилятов до образования зерновки [2]. Дикий клейстогамный вид S. silvеstre Host. имеет самые маленькие размеры пыльников, отличается уникальными признаками и свойствами, но в силу генетического барьера нескрещиваемости с культурной рожью, низкой фертильностью гибридов первого поколения в селекции используется ограничено [1].
Нами на основе цитогеномов дикой лесной клейстогамной ржи и гексаплоидного тритикале созданы вторичные источники аллоплазматической короткостебельной ржи с пониженной пыльцевой продуктивностью, а следовательно, экономичным расходованием пластических веществ в период споро- и гаметогенеза - МГР-1, Фиолтри.
При выполнении НИР было обнаружено новое свойство, а именно целенаправленный подбор пар при гибридизации, когда в качестве материнской формы используются образцы с плазмонным контролем адаптивности и интенсивности роста, а в качестве отцовской - источинки пониженной пыльцевой продуктивности, обеспечивает синтез гибридов с конкурсным гетерозисом и выделение из них трансгрессивных растений по элементам продуктивности компенсаторного действия.
Предложенный способ позволяет иметь технический результат, заключающийся в получении адаптивной, продуктивной популяции озимой ржи на основе выделенных трансгрессивных растений с пониженной пыльцевой продуктивностью и элементами продуктивности компенсаторного действия из гибридной комбинации, созданной целенаправленным подбором родительских форм.
Сопоставительный анализ заявляемого технического решения с прототипом показал, что заявляемый способ отличается от прототипа тем, что получение трансгрессивных растений с элементами продуктивности компенсаторного действия проводят в гибридных комбинациях с конкурсным гетерозисом, у которых в качестве материнского компонента используются формы с плазмонным контролем адаптивности и интенсивности роста, а в качестве отцовского - источник пониженной пыльцевой продуктивности.
Изучение патентной и научно-технической литературы свидетельствует о том, что получение трансгрессивных растений предложенным способом ново и в селекции с рожью не применялось.
Предложенный способ предусматривает использование известных сведений: детерминация гетерозиса путем ядерно-плазменного баланса генотипа [17]; закрепление гетерозиса в трансгрессивных линиях пшеницы [12] и гороха [11]; снижение потерь урожая зерна при сохранении главного побега растений от разрушительного действия реутилизации структурных веществ при засухе до или в ранней фазе налива зерновки [3]; использование клейстогамии или иных источников пониженной пыльцевой продуктивности в качестве механизма экономичного расхода пластических веществ до формирования зерновки [2]; частичная детерминация морозостойкости цитоплазматическими генами [9].
Цитогенетическая детерминация интенсивности весеннего отрастания побегов после действия низкотемпературного фактора у Популяции 50 морозостойкой (П-50М) обусловливает реципрокный эффект и ядерно-плазменный гетерозис в комбинации с высокопродуктивным сортом Саратовская 5 (табл.1).
Между оценкой интенсивности весеннего отрастания (показатель морозо- и зимостойкости популяции), с одной стороны, и продуктивной кустистостью, числом зерен в колосе, озерненностью, продуктивностью растения, с другой, установлена корреляционная зависимость. При выращивании на стеллажах коэффициенты корреляции были 0,85±0,12, 0,94±0,08, 0,91±0,10, 0,70±0,17; в полевых условиях - 0,58±0,19, 0,74±0,16, 0,76±0,15, 0,54±0,20 соответственно. Иначе, сопряженность показателей элементов структуры урожая растения и интенсивности отрастания позволяет опосредовано судить о ядерно-плазменной детерминации гетерозиса по элементам структуры урожая и о возможности создания продуктивной, адаптивной, с интенсивным poстом популяции с высоким наследованием данных элементов.
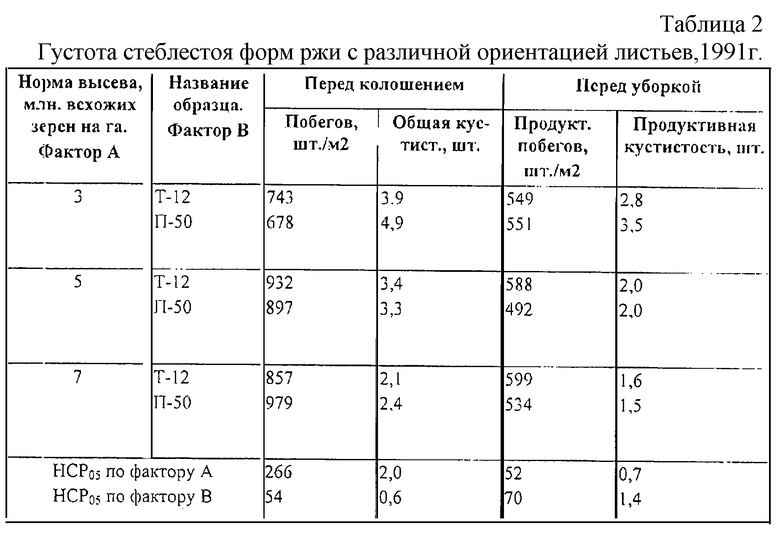
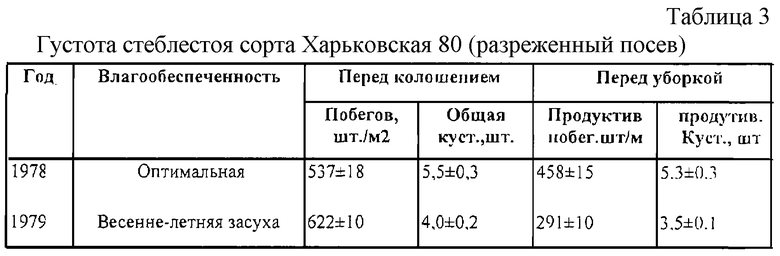
Компенсация изреженности стеблестоя за счет продуктивной кустистости растений ржи является необходимым селектируемым признаком из-за частичной гибели или повреждения растений в периоды зимовки и весенне-летней вегетации. В онтогенезе реализация общей кустистости растений ржи заметно снижается. Как правило, резкое изреживание стеблестоя идет за счет усыхания подгонов или гибели ослабленных растений в период споро- и гаметогенеза и особенно в засушливые годы (табл. 2 и 3).
Короткостебельная безлигульная Популяция 50 как источник вертикальной ориентации листьев, повышенной плотности стеблестоя и лучшей сохранности растений к уборке при загущении агроценоза не может реализовать потенциал густоты стеблестоя (табл. 2). Причиной тому могут быть недостаточная засухоустойчивость, слабая корневая система, более высокое, чем у Таловской 12, поражение болезнями.
Высокостебельный (относительно сортов с геном Н1) сорт озимой ржи Харьковская 60, отличающиеся хорошей продуктивностью, высокой зимостойкостью и средней засухоустойчивостью, также изреживает стеблестой как в оптимальный по увлажнению год, так и в засушливый [10]. Причем неблагоприятные климатические условия, начиная с ранних этапов развития растения, оказывают существенное влияние на процесс стеблеобразования, поэтому важным моментом в селекции продуктивных сортов является отбор генотипов, способных поддерживать высокую густоту продуктивного стеблестоя, независимо от условий произрастания.
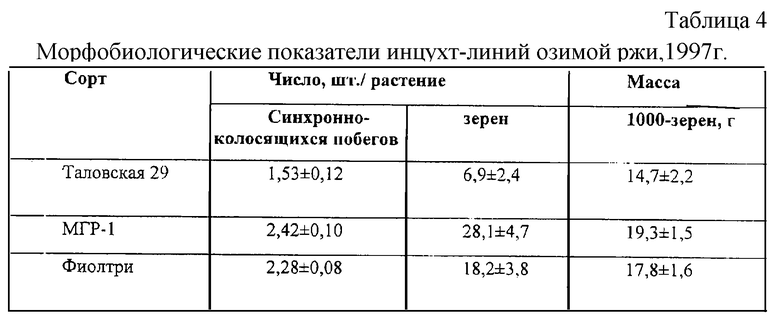
Популяции озимой ржи МГР-1, созданной на основе цитогенома дикой лесной клейстогамной ржи, и Фиолтри, созданной на основе цитогенома гексаплоидного тритикале служат источниками пониженной пыльцевой продуктивности. Инцухт-линии, выделенные из них, отличаются от инцухт-линий районированного сорта Таловская 29 синхронностью образования колосоносных побегов, склонностью к самоопылению, меньшей инцухт-депрессией при формировании зерна (табл. 4). Вероятно, имеется определенная связь пониженной пыльцевой продуктивности и аллелей автофертильности, что обеспечивает высокую завязываемость зерна в различных условиях опыления. Отсюда, высокая озерненность колоса озимой ржи может быть достигнута меньшим числом пыльцевых зерен.
За время цветения цветка ржи (в среднем 32 мин) на рыльце попадает в среднем 67 пыльцевых зерен, обеспечивая тем самым нормальное образование семян [5] . Р.Б. Нокс (1985) считает, что для оплодотворения одной яйцеклетки культурных растений необходимо около 200 пыльцевых зерен, формируется же пыльцы гораздо больше. У ржи на каждую формирующуюся завязь приходится 36300 пыльцевых зерен, тогда как у пшеницы 5600 [5]. Естественно, значительная часть пыльцы ржи обеспечивает активную физиологическую среду при оплодотворении, но и довольно много ее просто теряется, т.е. растением неэффективно расходуются синтезируемые пластические вещества.
Воздушно-сухая масса главного побега озимой ржи Искра в оптимальном по климатическим условиям 1988 году в период цветение - полная спелость в сплошном посеве увеличилась в 2,9 раза, с 28 до 80 г при среднем значении Кхоз= 0,37. Очевидно, этот период является наиболее плодотворным по ассимиляционной и аттракционной деятельности сформированных органов растений.
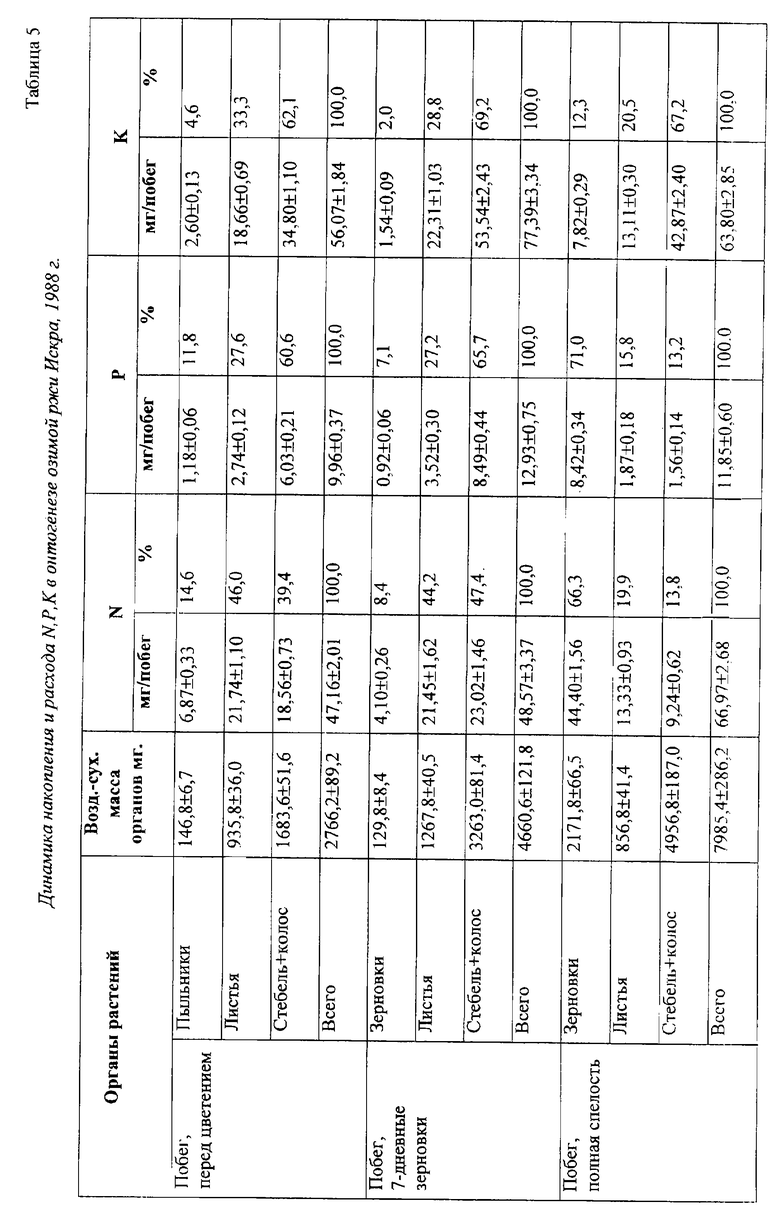
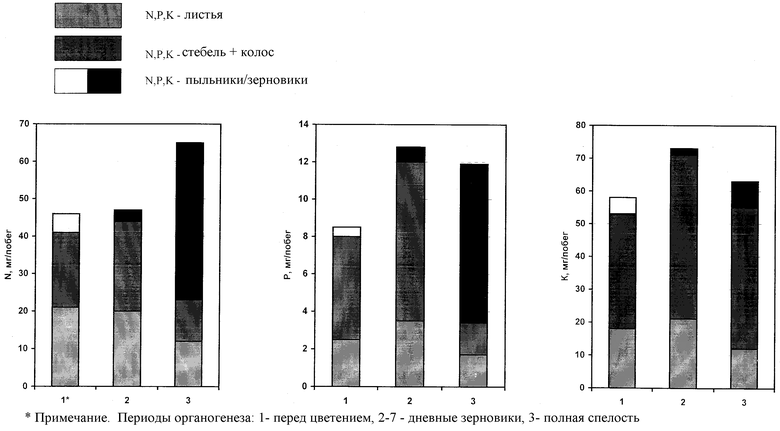
Динамика накопления и расхода N, Р, К растениями ржи в данный период показывает существенный вынос пыльниками макроэлементов при цветении, превышающим накопленное в недельных зерновках количество азота, фосфора и калия (см. чертеж, табл. 5). Потеря определенной части пыльцы при цветении сопровождается бесследной утратой для растения накопленных макроэлементов. Отсюда, крайне важна проблема эффективного распределения запасных и структурных веществ в реализации потенциальной продуктивности растения, улучшения качества продукции.
К фазе полной спелости половина (50,8%) массы азота зерновок формируется за счет аттракции из листьев и стеблей главного побега, другая часть - за счет поступлений из почвы. Во время налива зерна в зерновки утилизируется большая часть N, Р, К вегетативной массы главного побега, чему предшествует отток питательных веществ боковых побегов. Накопление белка в зерне обусловлено в основном степенью реутилизации азота отмирающих вегетативных органов [3].
Снижение пыльцевой продуктивности опосредовано, через более эффективное использование азота, фосфора и калия, способствует улучшению развития вегетативных органов растения и повышению продуктивности.
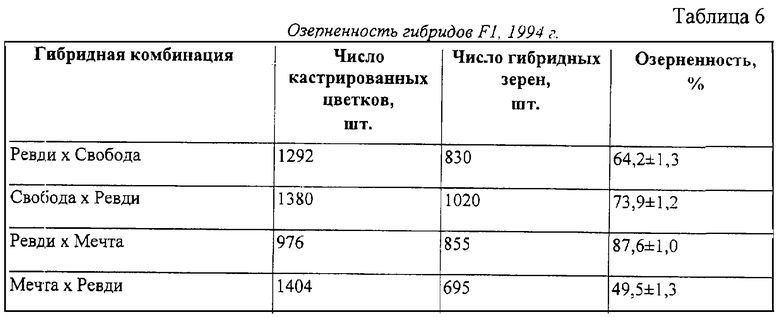
Согласно предлагаемому способу на основе гибрида Таловская 15 х МГР-1 создан вторичный источник пониженной пыльцевой продуктивности Мечта, отличающийся еще и тем, что имеет более высокую конкурентоспособность микроспор и конкурсный гетерозис по продуктивности (табл.6). Урожаи зерна в конкурсном сортоиспытании 1996 года у сорта-стандарта Таловская 15 составил 5,07 т/га, у сортообразцов Мечта - 5,25 и МГР-1 - 5,18 т/га (HCР05=0,20 т/га).
Высокая жизнеспособность пыльцы источников пониженной пыльцевой продуктивности наследуемый признак, указывает на сбалансированность физиолого-биохимических процессов во время роста и развития растений.
Полученные в ходе выполнения НИР результаты позволяют заключить, что возможно создание высокопродуктивной, адаптивной популяции озимой ржи, эффективно расходующей пластические вещества путем гибридизации, когда в качестве материнской формы используются образцы с плазмонным контролем адаптивности и интенсивности роста, а в качестве отцовской - источники пониженной пыльцевой продуктивности, обеспечивающие возникновение конкурсного ядерно-плазменного гетерозиса гибридов и трансгрессивных по признакам компенсаторного действия, подпадающих обычно под отбор. Таковыми являются синхронное колошение, высокая озерненность колоса, выполненность и форма зерновки.
На основе анализа известных решений в науке и технике делаем вывод, что наше техническое решение является новым и неочевидным. Сущность предлагаемого изобретения может быть понята из следующего описания.
Подбирают или создают необходимые для гибридизации компоненты с плазмонным контролем адаптивности и интенсивности роста.
Подбирают или создают источники пониженной пыльцевой продуктивности, отличающиеся экономичным расходованием пластических веществ в период споро- и гаметогенеза. В данном способе исследователи имели в наличии подобные источники.
Получают гибриды между указанными формами, причем в качестве отцовской используют источники пониженной пыльцевой продуктивности.
Отбирают из высеянных гибридов F1 гибриды с конкурсным гетерозисом.
Высевают отобранные гибриды следующего поколения и отбирают растения, трансгрессивные по синхронному колошению побегов, озерненности колоса, крупности, выполненности и выровненности зерна компенсаторного действия. Формируют популяцию озимой ржи на основе отобранных растений.
Литература
1. Кобылянский В. Д. Рожь. Генетические основы селекции. - М.: Колос, 1982. - 241 с.
2. Коновалов Ю. Б. Формирование продуктивности колоса яровой пшеницы и ячменя. - М.: Колос, 1981. - 176 с.
3. Кумаков В.А. Физиология формирования урожая яровой пшеницы и проблемы селекции// С.-х. биология. - 1995. - 5. -С. 3-19.
4. Маренков А.Я., Маренков Н.Я. Влияние минеральных удобрений и орошения на реутилизацию углеводов в процессе формирования зерна пшеницы// Вестник с. -х. Науки Казахстана. - 1975. 3. - С. 15-19.
5. Молчан И.М. О физиологическом значении процесса оплодотворения у растений// Известия ТСХА. - 1966. - Вып.3. -С. 17-29.
6. Нокс Р.Б. Биология пыльцы /Пер. с англ. и предисл. С.А.Резниковой. - М.: Агромпромиздат, 1985. - 83 с.
7. Скрипчинский В.В. Некоторые биологические свойства многолетних и однолетних форм кустовых злаков и проблема эволюции рода Secale //Вопросы эволюции, биогеографии, генетики и селекции/ АН СССР. - М.-Л., 1960. - С. 242-249.
8. Синнот Э.В. Морфогенез растений /Пер. с англ. - М.: ИЛ, 1963. - 603 с.
9. Титаренко А. В. Генетические основы создания нового исходного материала и селекции озимой ржи в Центрально-Черноземной зоне // Автореф. дис. . .. доктора с.-х. наук. - Санкт-Петербург, 1994. - 43 с.
10. Титаренко Л.П. Селекционная ценность исходного материала тетраплоидной озимой ржи для условий Центрально-Черноземной зоны // Автореф. дис. ... канд. с.-х. наук. - Харьков, 1988. - 18 с.
11. Фомин В. С. Пути повышения эффективности селекции гороха в ЦЧЗ // Диссертация доктора с.-х. наук в форме научного доклада. - Немчиновка, 1995. - 45 с.
12. Хупацария Т.И. Методика выделения трансгрессивных форм из межсортовых гибридов яровой пшеницы, их изучение и взаимосвязь элементов продуктивности растений // Автореф. Дис. ... канд.биол.наук. - М., 1974. - 19 с.
13. Evans L.T., Bingham J., Roskams M.A. The pattern of grain set within ears of wheat// Austr. J.Biol.Sci. - 1972. - V.25, N1. - P.1-8.
14. Friedt W., Linol V., Walter H. et al. The value of inbred lines from Secale cerealexSecale vavilovii via classical inbreeding and androgenetic haploids// Z.Pflanzenzuch. - 1983. - Bd.91, N2. - S.89-103.
15. Johnson R. , Lambert R. Tassel trivia: little - known fasts about corn// Illinois Res. - 1975. - V.17, N4. -P.5.
16. Leopold A.C., Niedergans - Kamien E., Janick J. Experimental modification of plant senescense// Plant Physiol. - 1959. - V.34, N5. - P. 570-573.
17. Srivastava H. K. , Sarkissian I.V. Heterosis, complementation and homeostasis in mitochondria of wheat //Genetics. - 1969.- V.G1; suppl.2, part 2. - P. 57-63.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СЕЛЕКЦИИ ОЗИМОЙ РЖИ | 1992 |
|
RU2080054C1 |
| СПОСОБ СЕЛЕКЦИИ РАСТЕНИЙ ЗЛАКОВЫХ КУЛЬТУР | 1997 |
|
RU2125365C1 |
| СПОСОБ ОТБОРА МОРОЗОСТОЙКИХ ФОРМ АЛЛОПЛАЗМАТИЧЕСКОЙ РЖИ | 2000 |
|
RU2192124C2 |
| Способ получения аналогов тетраплоидной ржи | 1991 |
|
SU1807842A3 |
| СПОСОБ ЗАРАЖЕНИЯ ПРОСА ГОЛОВНЕЙ | 1993 |
|
RU2090054C1 |
| Способ гибридизации полиплоидов зерновых культур | 1989 |
|
SU1655389A1 |
| СПОСОБ ОТБОРА ХОЗЯЙСТВЕННО-ЦЕННЫХ ФОРМ ЗЕРНОВЫХ КУЛЬТУР | 1992 |
|
RU2068635C1 |
| СПОСОБ ОТБОРА ВЫСОКОПРОДУКТИВНЫХ ФОРМ ОЗИМОЙ РЖИ ПРИ СЕЛЕКЦИИ НА ЗАСУХОУСТОЙЧИВОСТЬ | 2008 |
|
RU2368132C1 |
| Способ получения форм ржи | 1986 |
|
SU1632402A1 |
| Способ повышения плодовитости аутотетраплоидной ржи | 1986 |
|
SU1431716A1 |

Изобретение предназначено для использования в области сельского хозяйства в селекции. Способ включает подбор родительских форм. В качестве материнской используют форму с плазмонным контролем адаптивности и интенсивности роста. В качестве отцовской - источник пониженной пыльцевой продуктивности. Проводят гибридизацию, размножают полученные гибриды, и в гибридах с конкурсным гетерозисом отбирают растения по элементам продуктивности компенсаторного действия. При этом в качестве элементов продуктивности компенсаторного действия используют синхронное колошение, озерненность колоса, крупность, выполненность и выравненность зерна. Изобретение позволяет получить адаптивную продуктивную популяцию озимой ржи. 1 з.п.ф-лы, 1 ил., 6 табл.
| ХУПАЦАРИЯ Т.И | |||
| Методика выделения трансгрессивных форм из межсортовых гибридов яровой пшеницы, их изучение и взаимосвязь элементов продуктивности растений | |||
| Автореферат дисс | |||
| канд | |||
| биол | |||
| наук | |||
| - М., 1974, с.3-17 | |||
| СИННОТ Э.В | |||
| Морфогенез растений | |||
| - М.: ИЛ, 1963, с.393-425 | |||
| ТИТАРЕНКО А.В | |||
| Генетические основы создания нового исходного материала и селекции озимой ржи в Центрально-Черноземной зоне | |||
| Автореферат дисс | |||
| доктора с.-х | |||
| наук | |||
| - С.-П., 1994, с.10-30. |
