Изобретение относится к коньюгатам иммуноглобулина, способу их получения и использованию.
Общая концепция целенаправленной доставки антинеопластических веществ к опухоли, используя моноклональные антитела (Моав), известна, и в настоящее время определяется ее терапевтическое значение. Обычно такой подход включает получение коньюгатов антитела с токсическим веществом, способным селективно локализоваться и убивать клетки опухоли. Основное внимание уделялось конструированию иммунотоксинов из A-цепей токсинов растений, бактерий и антител таких, которые связывают антиген и, проникая внутрь, ведут к гибели клеток. В действительности многие Моав, которые считались специфичными к опухолям, оказались также реакционноспособными по отношению к субпопуляциям нормальных клеток, и, следовательно, может оказаться невозможным использовать такие сильные токсины из-за того, что потенциально они могут повредить нормальные ткани. Безопасной альтернативой растительным токсинам является соединение антител с обычными противоопухолевыми препаратами, такими как доксорубицин, виндезин, хлорамбуцил, мелфалан и метотрексат. Из-за неспецифичной токсичности обычно используемых антинеопластических агентов предпринимались попытки повысить их терапевтический индекс путем соединения их с Моав и с ассоциированными с опухолью антигенами.
Попытки подавить отторжение трансплантата, обусловленное вырабатываемыми тимусом T-клетками, часто сосредоточивались на снижении активности T-клеток путем использования антитимоцитглобулина. Недавно с развитием знаний о моноклональных антителах стало возможно определить подклассы T-клеток в соответствии с их функциями in vitro и в зависимости от присутствия специфичных поверхностных антигенов, определяемых Моав. Это стимулировало поиск механизма контроля T-клетками отторжения трансплантата, сосредоточиваясь на основном разделении на подклассы хелпер/промотор и цитотоксин/супрессор, определяемые L3T4- и Ly-2-антигенами мыши. Хотя ОКТЗ Моав, анти-пан-T-клеточный реагент, показал потенцию in vivo, в недавнем исследовании мы обнаружили, что 20 Моав были индифферентны в опытах in vivo на мышах к 20 разным антигенам лимфоцитов мыши и потому они бесполезны при исследованиях in vivo. Поэтому следует опробовать средства, с помощью которых эти высоко специфичные Моав можно сделать более эффективными и такими, чтобы они полностью удаляли клетки-мишени. Использование цитотоксичных препаратов в соединении с Моав является одним из таких подходов.
Клинические испытания конъюгатов препарат-Моав включают иммунохимиотерапию рака и облегчение различных иммунорегулируемых заболеваний и отторжений аллотрансплантата. Множество исследований показали специфичную цитотоксичность токсинов и препаратов по отношению к опухолевым клеткам в случае соединения этих агентов с Моав, соединенными с ассоциированными с опухолью антигенами.
Однако мало уделялось внимания in vivo испытаниям способности конъюгатов препарат-Моав разрушать T-клетки и изучению T-клеточной иммунорегуляции при отторжении трансплантата, хотя конъюгаты токсин-антитело широко использовались in vitro для разрушения T-клеток перед пересадкой костной ткани.
Антракциклины представляют собой важную группу антинеопластических агентов, используемых при химиотерапии рака, причем доксорубицин и даунорубицин продемонстрировали свою эффективность против твердых опухолей. Однако соединение даунорубицина и доксорубицина с антителами привело к существенному снижению эффективности препаратов, если это соединение осуществлялось посредством аминогруппы. Недавно даунорубицин соединили с Моав посредством C-14, используя бромодаунорубицин. Эти конъюгаты показали активность in vitro. Однако сообщения об исследованиях in vivo отсутствуют (Gallego et al., Inl. J. Cancer, 1984, 33, 737 - 744). Кроме того, было показано, что конъюгаты даунорубицина с Моав обладают неспецифичной токсичностью в концентрациях > 10 мкг/мл.
Мы обнаружили, что идарубицин (4) деметоксидаунорубицин, Ida можно соединить с Моав, и эти конъюгаты обладают селективной и сильной противоопухолевой активностью in vitro и in vivo. Неспецифичная токсичность конъюгатов Ida-Моав in vitro не наблюдалась при концентрациях < 8 мг/кг.
Конъюгаты Ida-Моав обладают более высокой эффективностью in vitro и in vivo, чем конъюгаты даунорубицина с Моав.
Кроме того, мы оценили способность конъюгатов Ida-Моав специфично уничтожать популяции клеток путем присоединения Ida к Моав, которые, как известно, реагируют с различными субпопуляциями лимфоцитов (L3T4+, Ly-2+ и Thy-16). Мы обнаружили, что это удивительно эффективный способ уничтожения клеток. Например, анти-Ly-2.1 Моав, который обладает сильным воздействием in vivo на Ly-2.1+-клетки, можно превратить в эффективный цитотоксичный агент путем соединения с цитотоксичным агентом Ida.
используя Моав к Ly-2- и L3T4-антигенам, мы показали, что конъюгаты Ida-Моав способны элиминировать клетки-мишени in vitro и in vivo. Можно получить конъюгаты Ida-Моав, которые обладают более высокой способностью специфичной уничтожать подклассы T-клеток, ответственных за отторжение трансплантата, по сравнению с антилимфоцит-глобулинами или анти-пан-T-клеточными реагентами такими, как ОКТЗ Моав.
Изобретение раскрывает конъюгаты иммуноглобулина, включающие Ida, соединенные с моноклональными антителами или их фрагментами, включающими по меньшей мере один из участков связывания антигена данного антитела.
Ida описан в патенте США N 4077988. Предпочтительно 2 - 8 молекул Ida ковалентно связаны с каждой молекулой антитела или фрагмента антитела, более предпочтительно 2 - 6. Обычно, но не обязательно, молекулы Ida присоединены в положении 14 к моноклональному антителу или фрагменту антитела. Предпочтительно они связаны непосредственно, хотя возможна связь через инертный носитель или сшивающий агент. Ida может быть связан с моноклональным антителом или фрагментом антитела посредством аминогруппы. Это можно осуществить, используя разлагающийся цептидный соединитель, декстрановый носитель, кислотночувствительный соединитель или полиглутаминовую кислоту.
Обычно каждое антитело или фрагмент антитела специфичны к антигену на поверхности клеток, против которых необходимо воздействовать Ida. Например, антитело или фрагмент антитела могут быть специфичны к соответствующей ткани-мишени такой, как неоплазм у человека. Примерами неоплазмов у человека, на которые может быть желательно направить действие Ida, являются опухоли груди, ободочной кишки, легких, простаты, яичников, тимуса и другие раки, саркомы и лейкемии. Антитело и фрагмент антитела может быть специфичен к неоплазмам у животных. Можно использовать антитело или фрагмент антитела специфичный к рецептору трансферина /TFR/ человека, который присутствует на делящихся клетках, эритроидных клетках-предшественниках и клетках различных опухолей. Подходящее анти-TFR моноклональное антитело можно использовать против рецептора трансферина на клетках LiCR LON-UHy-2 (HMy-2).
Если конъюгаты иммуноглобулина предполагается использовать для уничтожения специфичных популяций T-лимфоцитов, то антитело или фрагмент антитела специфичен к клеточному поверхностному антигену, который сам по себе специфичен к этим T-лимфоцитам. Поэтому антитело или фрагмент антитела может быть специфичен к популяции хелперов, супрессоров или цитотоксичных T-лимфоцитов.
Предпочтительно моноклональное антитело или фрагмент антитела того же класса, что и то, против которого предполагается использовать конъюгат иммуноглобулина. Моноклональное антитело или фрагмент антитела человека или мыши обычно поэтому используют, когда предполагается вводить соединение человеку. Предпочтительно, чтобы антитело или фрагмент антитела был класса 1C. Фрагмент антитела может быть Fab, Fab' или F(ab)2-фрагмент.
Можно также использовать мономер IgM, который можно получить из IgM-антитела путем протеолитического ферментативного переваривания.
Конъюгаты иммуноглобулина получают по настоящему изобретению способом, который включает соединение Ida с моноклональным антителом или фрагментом антитела. Предпочтительно 14-гало-Ida взаимодействует с моноклональным антителом или фрагментом антитела. 14-заместитель может быть фтором, хлором, бромом или иодом, но предпочтительно бромом. 14-бромо-Ida описан в патенте США N 4125697. Коньюгацию, таким образом, можно осуществить способом, который включает
а) смешение моноклонального антитела или фрагмента антитела с молярным избытком 14-гало-Ida,
b) проведение реакции при температуре 18-37oC,
с) удаление осадка,
d) удаление непрореагировавших исходных материалов путем гельфильтрации и
е) удаление препарата (Ide) путем абсорбционной хроматографии или ионообменной хроматографии.
Стадию (а) обычно проводят в смешивающемся с водой органическом растворителе таком, как N, N-диметилформамид. Предпочтительно молярный избыток 14-гало-Ida на стадии (а) от 0 до 50 раз. На стадии (b) реакцию предпочтительно проводят в течение 1-8 ч. Обычно реакцию проводят при комнатной температуре.
Можно использовать и другие способы получения коньюгата. Если необходимо, чтобы коньюгат имел инертный носитель или сшивающий агент, расположенный между Ida и моноклональным антителом или фрагментом антитела, то носитель или сшивающий агент обычно сначала присоединяют к атому углерода C-14 Ida и затем присоединяют к антителу или фрагменту антитела. Как указано выше, антитело или фрагмент антитела может быть связан с Ida посредством аминогруппы путем использования расщепляющегося пептидного соединения-спейсера, декстранового носителя, кислотночувствительного спейсера или полиглутаминовой кислоты.
Коньюгаты иммуноглобулина по настоящему изобретению можно использовать для лечения рака у людей и млекопитающих. Можно применять терапевтически эффективное количество конъюгата. Рак может представлять собой твердую опухоль, асцитную опухоль или лейкемию. Неоплазмы у человека, которые можно лечить настоящими коньюгатами, указаны выше, можно вводить два и более коньюгат, в которых моноклональное антитело или фрагмент антитела в каждом коньюгате имеет различную специфичность.
Коньюгат можно вводить в виде инъекции. Можно его применять парентерально, например внутривенно. Его можно вводить локально или непосредственно в опухоль. Количество коньюгата, вводимое больному, зависит от различных факторов, таких как вид подлежащей лечению опухоли, состояние больного. Обычная доза составляет 10-200 кг коньюгата на 1 м2 тела пациента. Настоящие коньюгаты можно применять совместно с другими химиотерапевтическими агентами или с агентами, усиливающими активность настоящих коньюгатов, например вазоактивными агентами или фактором омертвления опухоли.
Коньюгаты иммуноглобулина можно также использовать для специфичного удаления подкласса Т-лимфоцитов из популяции клеток. Человеку или животному можно ввести цитотоксично эффективное количество коньюгата, включающего моноклональное антитело или фрагмент антитела, направленный против клеточного поверхностного антигена, присутствующего на подлежащих удалению лимфоцитах. При альтернативном варианте популяцию клеток можно инкубировать in vitro с таким коньюгатом, Можно использовать комбинацию коньюгатов против двух или более клеточных поверхностных антигенов. Подлежащие удалению лимфоциты могут представлять собой хелперы, супрессоры или цитотоксичные Т-клетки.
Этот аспект настоящего изобретения можно использовать для предотвращения отторжения трансплантированной ткани у реципиента. Настоящие коньюгаты можно использовать как иммуносупрессивные агенты. Реципиенту вводят такое количество коньюгата, которое эффективно предотвращает отторжение трансплантата. В данном случае предпочтительно удаляют цитотоксичные Т-клетки.
Рак у животных и человека можно лечить путем удаления Т-клеток-супрессоров введением соответствующего коньюгата. Аутоимунные заболевания можно лечить путем введения коньюгата для удаления T-клеток-хелперов. В каждом случае применяются указанные выше способ введения коньюгата и доза.
Иммуноглобулиновые коньюгаты приготавливают в виде фармацевтических композиций с фармацевтически приемлемым носителем или разбавителем. Можно использовать любой подходящий носитель или разбавитель. Подходящие носители или разбавители включают физиологический раствор и раствор декстрозы Рингера.
Следующие примеры иллюстрируют настоящее изобретение.
На приложенных иллюстрациях фиг. 1-12 относятся к примеру 1, а фиг. 13-17 - к примеру 2.
На фиг. 1 представлена структурная формула производных антрациклина.
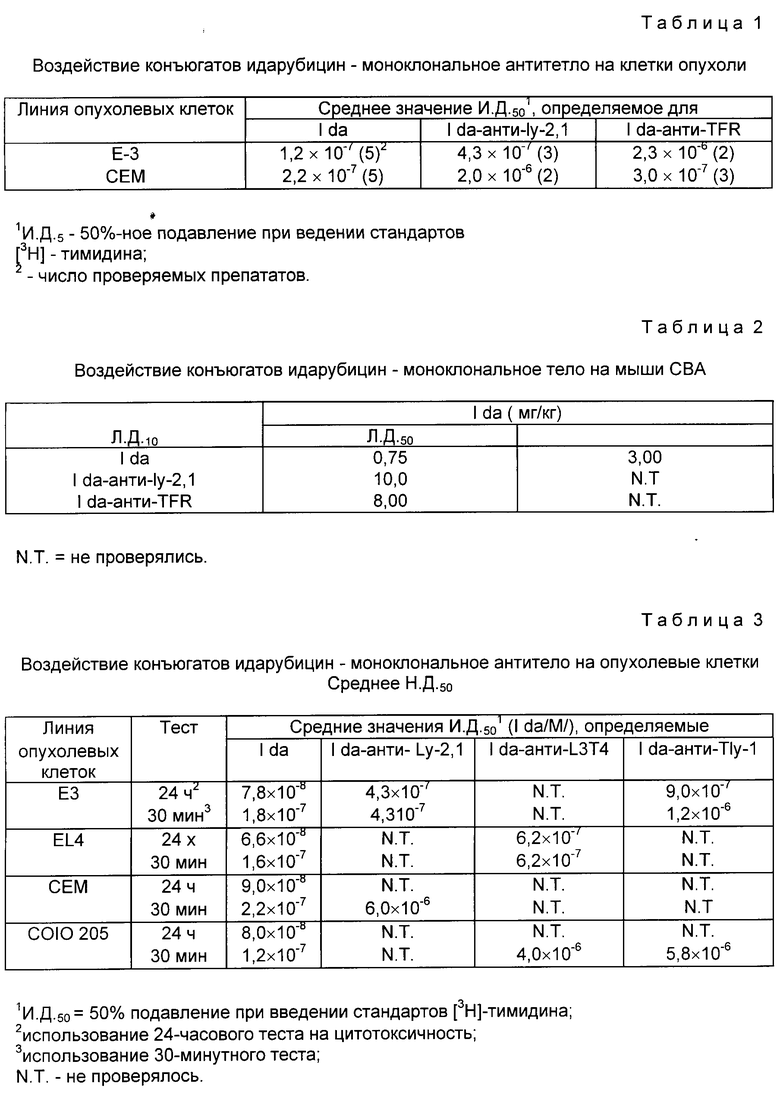
На фиг. 2 представлено соединение идарубицина /Ida/ с анти-Ly-2.1 (0,5 мг). Количество молей Ida, внедренных на моль анти-Ly-2.1 (1)- левая ордината, выделение протеина; (2) правая ордината, показано как функция количества нмолей Ida в реакционной смеси (абсцисса).
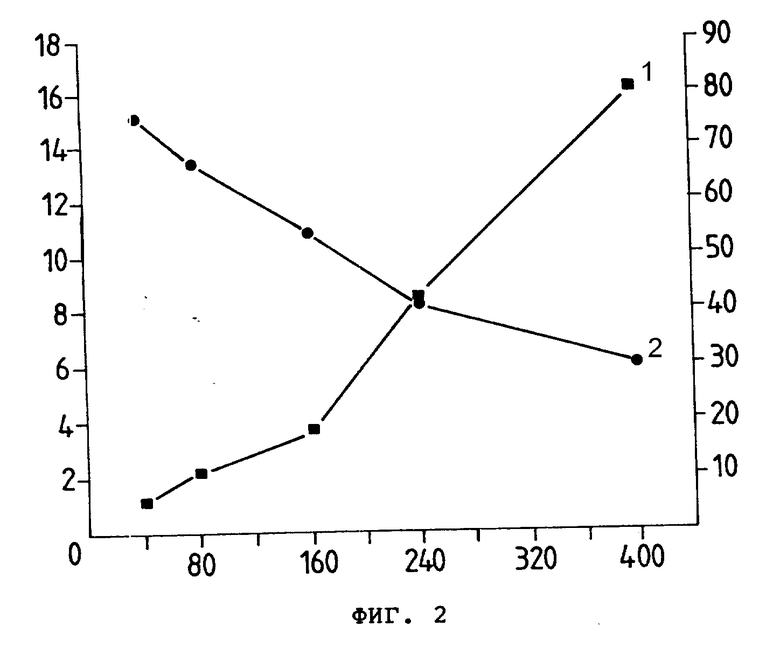
На фиг. 3 представлен титр антитела как % разеткообразующих клеток (ордината) в зависимости от разбавления (•10-1) (абсцисса) коньюгатов анти-Ly-2.1 на клетках-мишенях ITT (I) 75 NS E3. Последовательные разбавления раствора 0,5 мг/мл анти-Ly-2.1 (3) или анти-Ly-2.1 проводили с помощью раствора 2 (4) или 8 (5) моль Ida моль антитела.
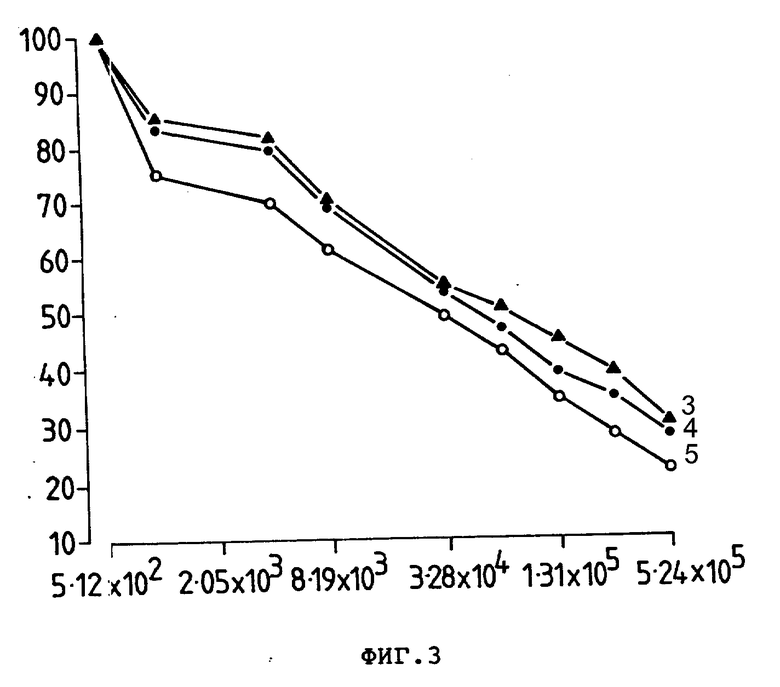
На фиг. 4 представлено ингибирующее действие Ida (6) или Ida-анти-Ly-2.1, 5 моль Lda/моль антителе (7), на E3-клетках при 24-часовом тесте, где приведен % ингибирования внедрения [3 H]-тимидина (ордината) в зависимости от концентрации Ida(М) (абсцисса).
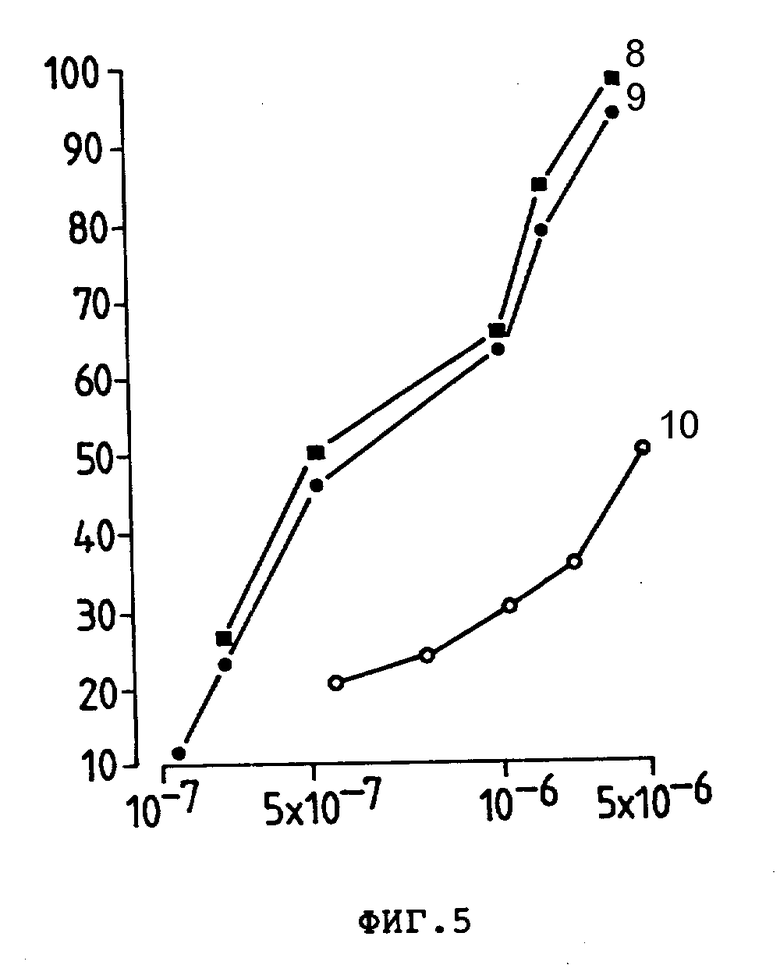
На фиг. 5 представлено ингибирующее действие Ida (8), Ida-анти -Ly-2.1, 5 моль Ida/моль антитела (9) или Ida анти-TFR, 5 моль антитела (10) на /Ly-2+/ E3-клетки при 30-минутном тесте, где приведен % ингибирования внедрения [3H] -тимидина (ордината) в зависимости от концентрации Ida (M) (абсцисса).
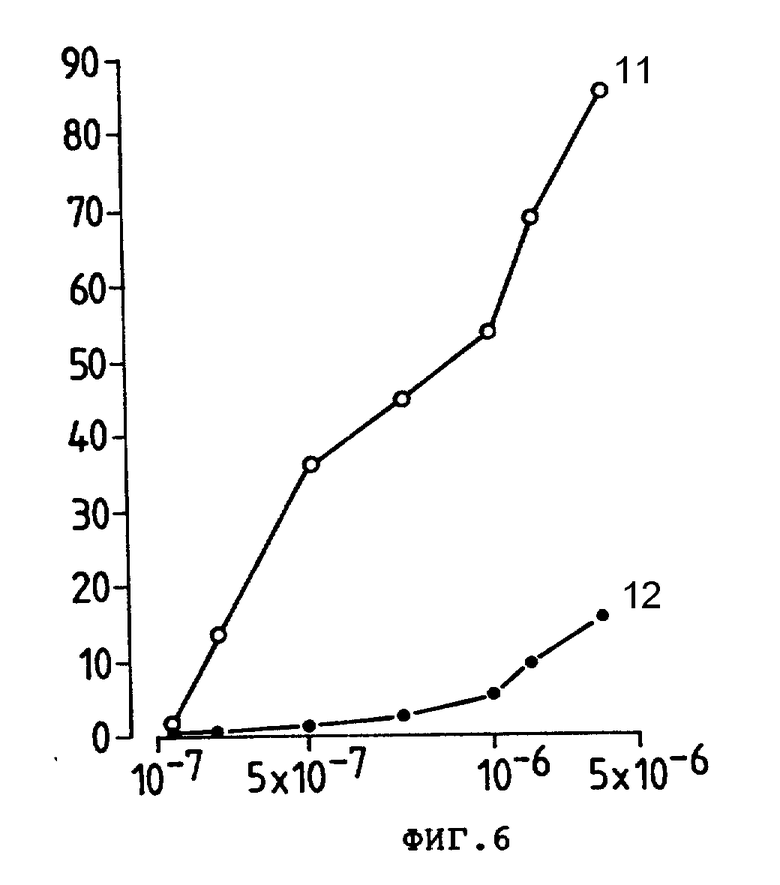
На фиг. 6 представлено ингибирующее действие Ida-анти-Ly-2.1, 5 моль Ida моль антитела (11) и коньюгата с анти-Ly-2.1 (12) на E3-клетки-мишени в 30-минутном тесте на специфичность, где приведен % ингибирования внедрения [3H] тимидина (ордината) в зависимости от концентрации Ida- (N) (абсцисса).
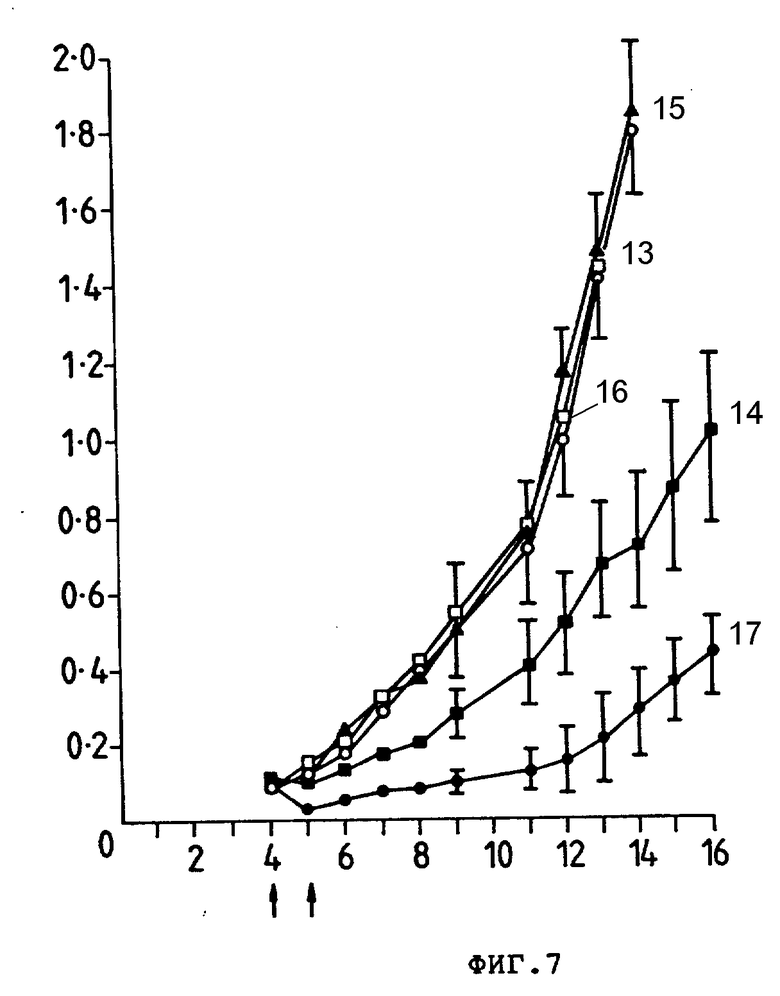
На фиг. 7 представлен рост тимомы E3 у мышей CBF1, которым подкожно ввели 2•106 клетках. Группам по 10 мышей внутривенно ввели препараты, обозначенные стрелкой; PBS (13), Ida (14), анти-Ly-2.1 (15), Ida-анти-TFR (16) или Ida-анти-Ly-2.1(17), Средний размер опухоли (см2) (ордината) показан в зависимости от количества дней, прошедших после введения опухолевых клеток (абсцисса). Гистограмма ошибок показывает ± стандартную ошибку среднего значения.
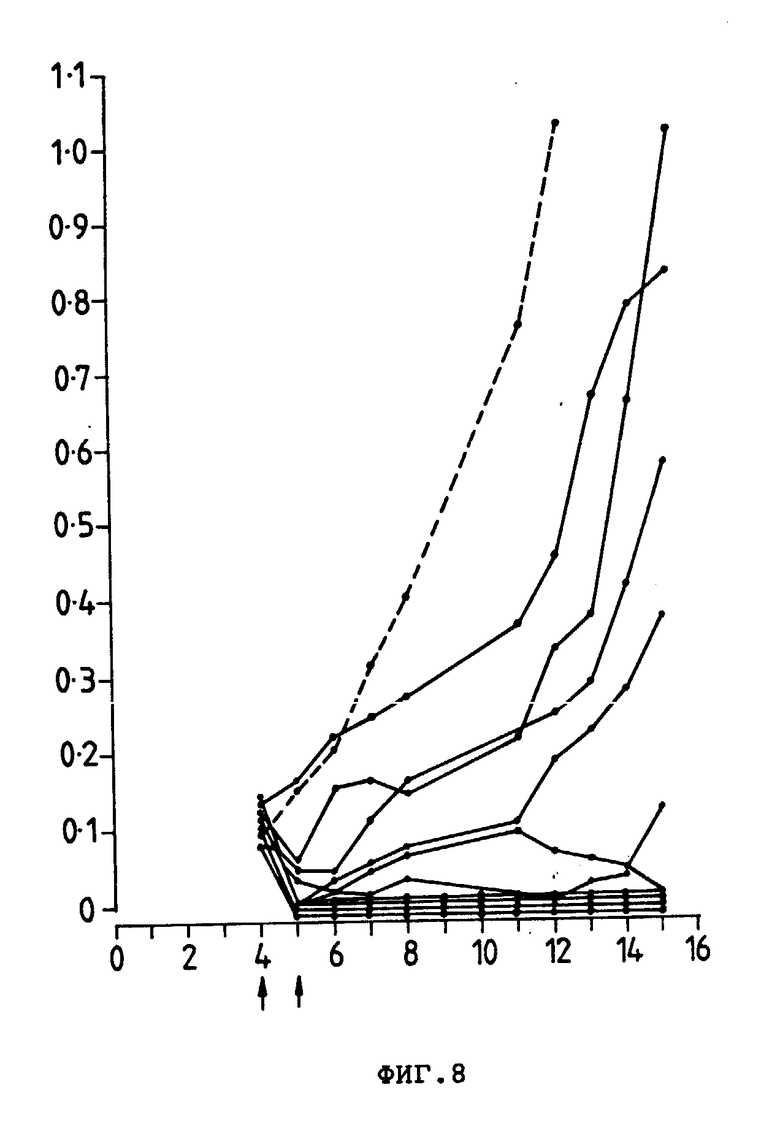
На фиг. 8 представлены индивидуальные кривые роста опухоли у мышей CBF1, которым подкожно ввели 2,0•106 клеток E3 опухоли и которым не 4- и 5-й день внутривенно ввели Ida-анти-Ly-2.1-коньюгат. Размер опухоли (см2) (ордината) показан в зависимости от количества дней, прошедших после введения опухолевых клеток (абсциссах).
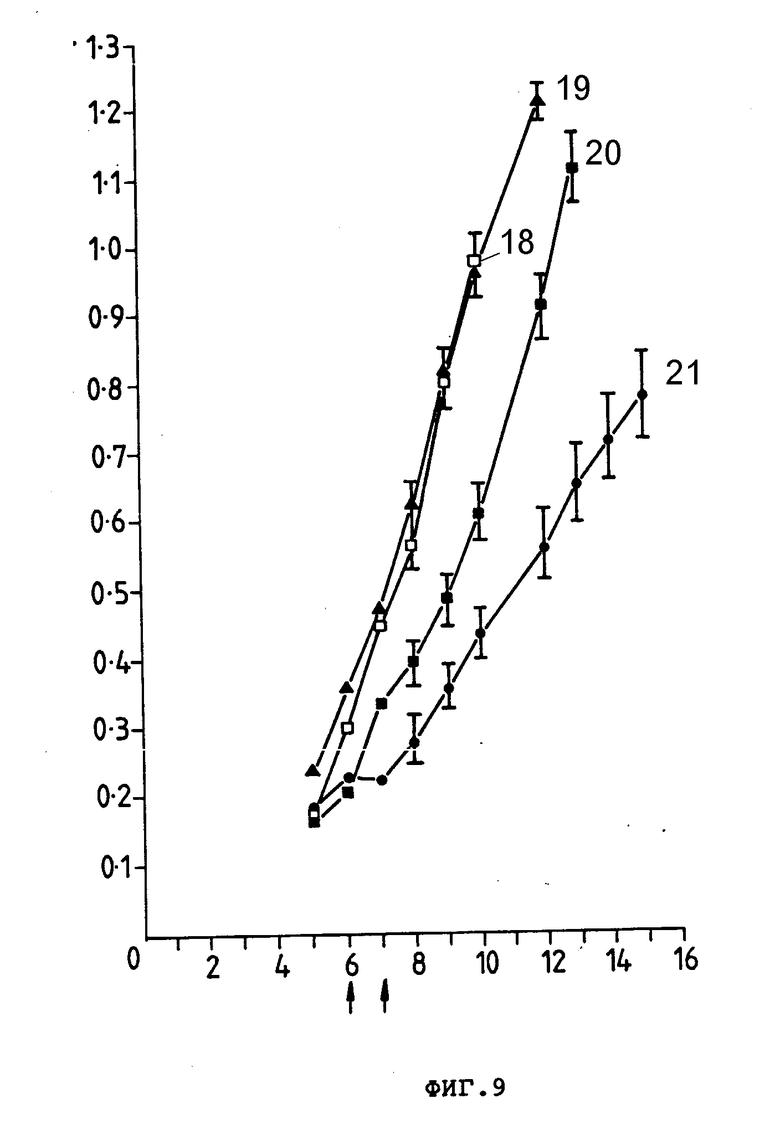
На фиг. 9 представлен рост тимоны E3 у CBF1-мышей, которым подкожно ввели 3,0•106клеток. Группам по 10 мышей внутривенно ввели препараты, показанные стрелкой; PBS (18), анти-Ly-2.1 (19), Ida (20) или Ida-анти-Ly-2.1 коньюгат (21). Средние значения размера опухоли (см2) (ордината) представлены в зависимости от количества дней, прошедших после введения опухолевых клеток ( абсцисса). Гистограмма ошибок показывает ± стандартная ошибка от среднего значения.
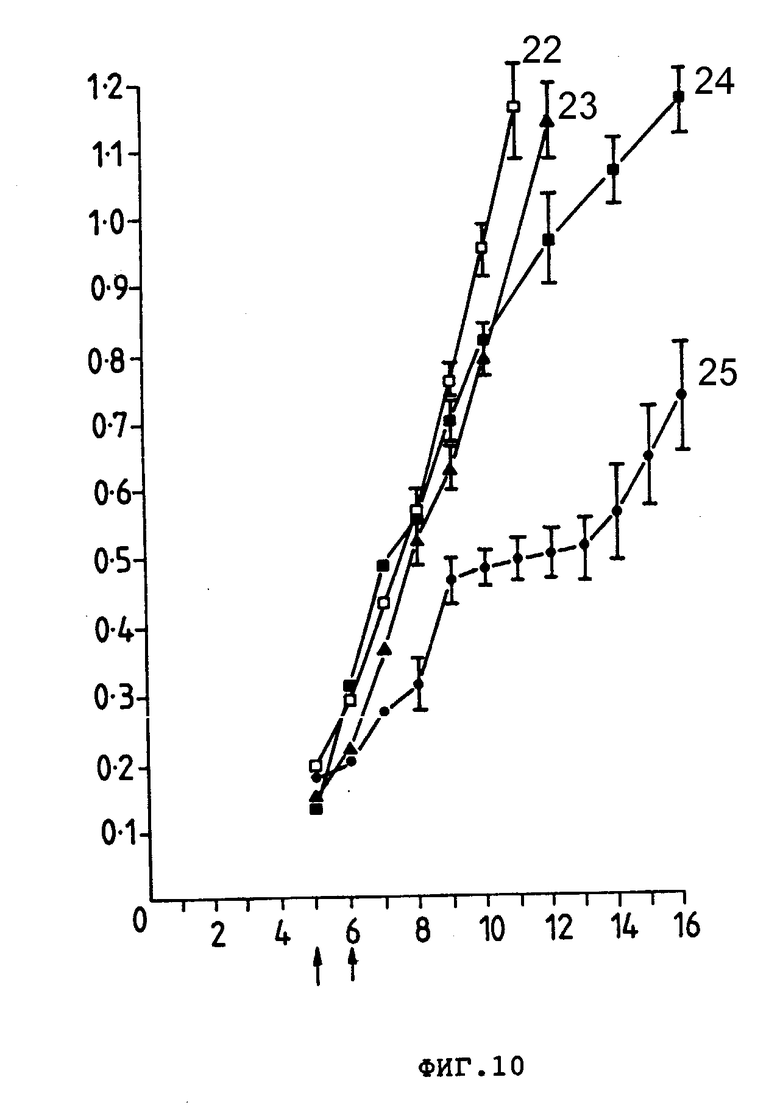
На фиг. 10 представлен рост тимоны E3 у мышей CBF, которым подкожно ввели 3,0•106 клеток. Группам по 10 мышей вводили внутривенно препараты, показанные стрелкой: PBS (22), анти-Ly-2.1 (23), Ida (24) или Ida-анти-Ly-2.1 коньюгат (25). Средние размеры опухоли (см2) (ордината) представлены в зависимости от количества дне после введения опухолевых клеток (абсцисса). Гистограмма ошибок показывает ± стандартная ошибка среднего значения.
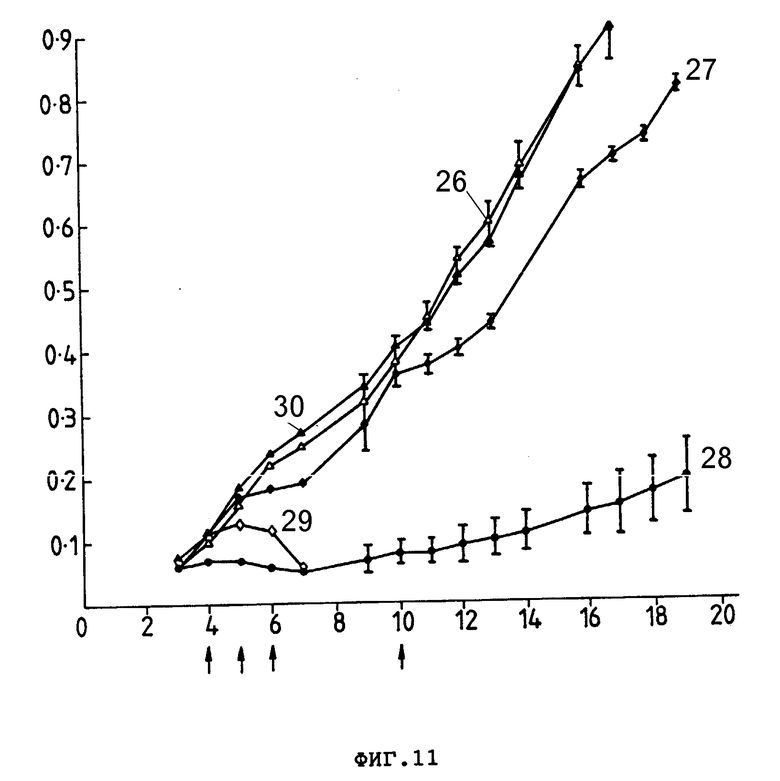
На фиг. 11 представлен рост ксенотрансплантата опухоли COLO 205 человека у безволосых мышей, которым подкожно ввели 2•106 клеток. Группам по 10 мышей внутривенно вводили препараты, показанные стрелками: PBS (26), свободный Ida (27), Ida-250-30,6-коньюгат (28), смесь Ida и 250-30,6 (29) и 250-30,5 (30). Средний размер опухоли (см2) (ордината) представлен в зависимости от количества дней, прошедших после введения клеток опухоли (абсцисса). Гистограмма представляет ± стандартная ошибка от среднего значения размера опухоли.
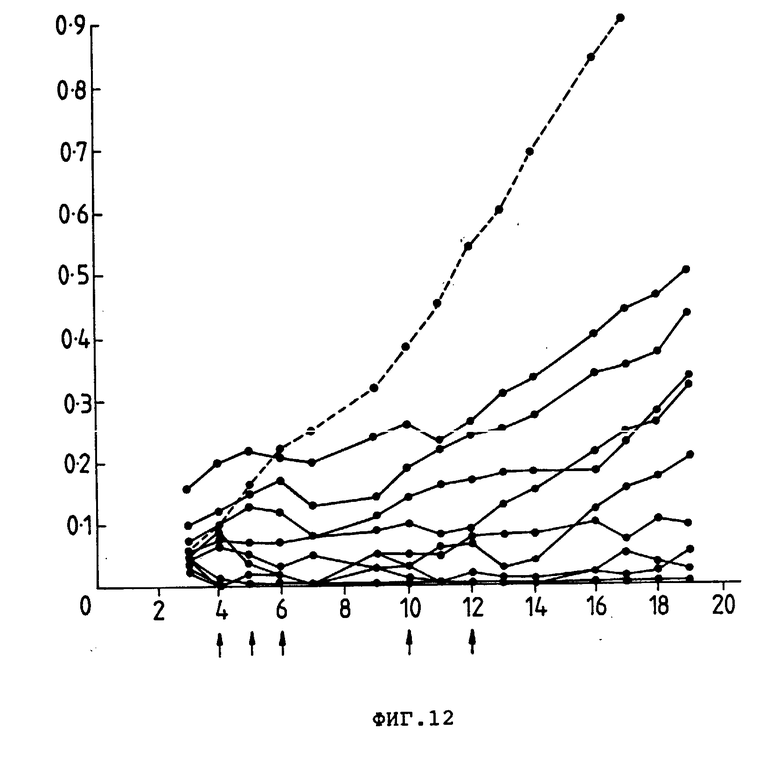
На фиг. 12 представлены индивидуальные кривые роста опухоли, ксенотрансплантированной безволосым мышам, которых обработали внутривенно (стрелка) Ida-250-30,6-коньюгатом. Пунктирная линия показывает среднее значение размера опухоли у мышей, обработанных PBS. Размер опухоли (см2) (ордината) представлен в зависимости от количества дней, прошедших после введения опухолевых клеток (абсцисса).
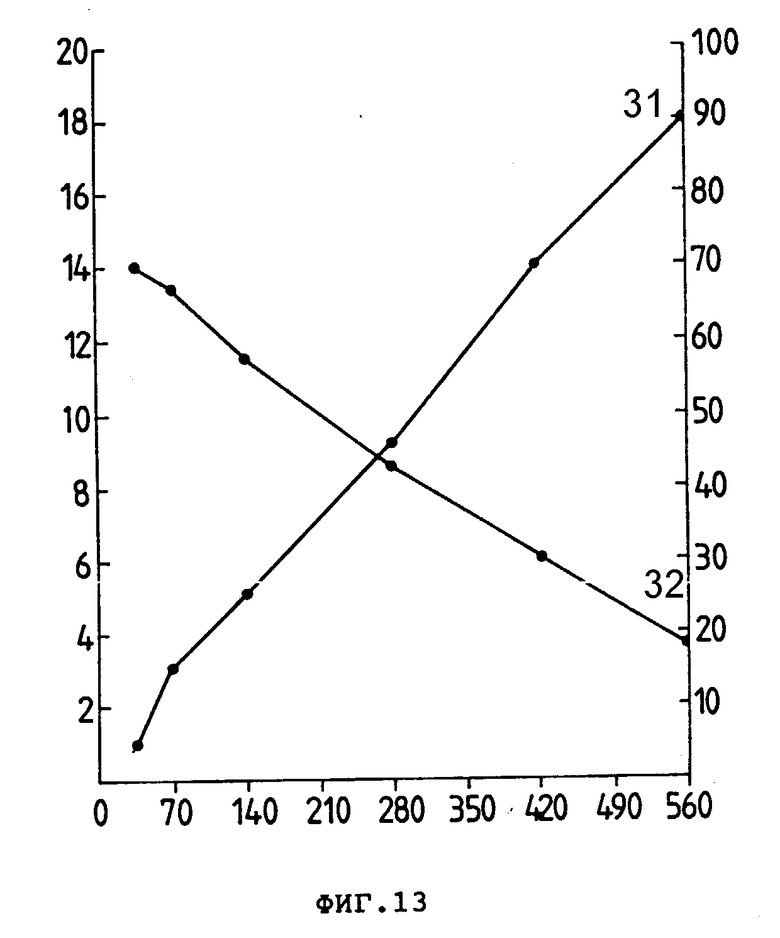
На фиг. 13 показано соединение идарубицина (Ida) с анти-L3T4 (0,5 мг). Количество молей Ida,внедренных на моль анти-L3T4 (31) (левая ордината), и выделение протеина (32) (правая ордината) показаны как функция количества н-молей Ida в реакционной смеси (абсцисса).
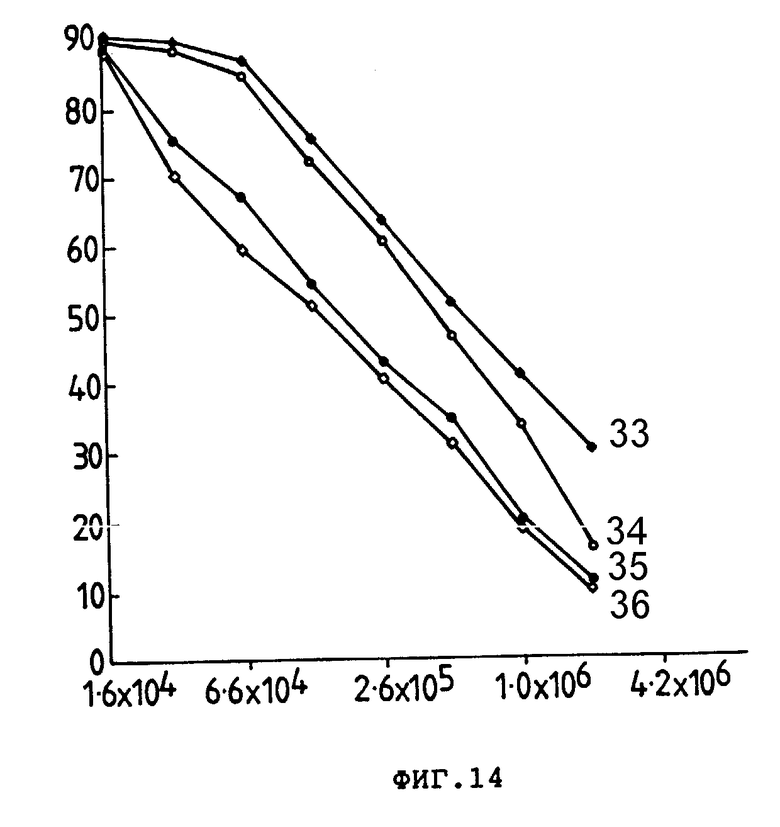
На фиг. 14 представлен титр антитела как % розеткообразующих клеток (ордината) в зависимости от разведения антитела (•10-1) (абсцисса) анти-TLy-I-конъюгатов ITT (1) 75 NS на E3-клетках-мишенях. Серию разведений провели на растворе 1 мг/мл анти-TLy-I (33) или конъюгата раствором 1 (34), 4 (35) или 7 (36) моль Ida/моль анти-TLy-I.
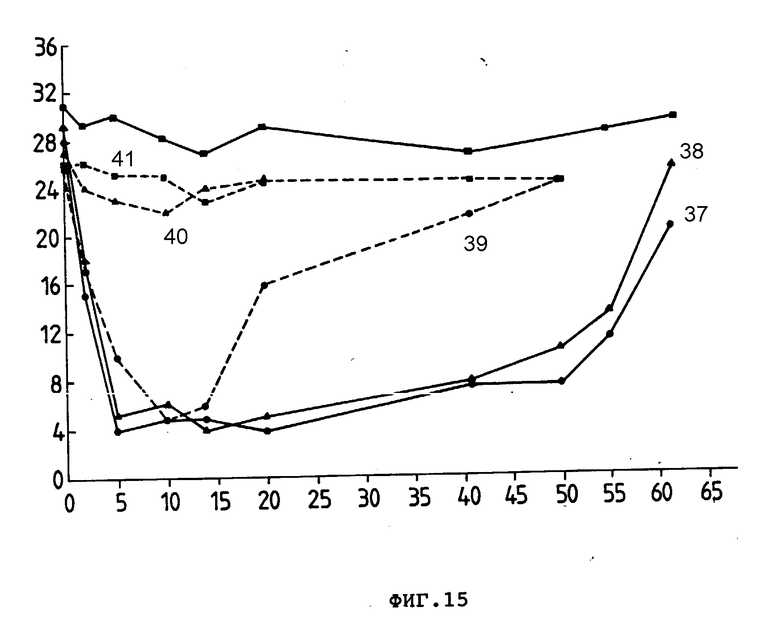
На фиг. 15 представлено воздействие Ida-Моав и Моав на количество L3T4+ - и Ly-2+-клеток в селезенке у мышей, обработанных Ida-анти-L3T4 (37), анти-L3T4 (38), Ida-анти-Ly-2.1 (39), анти-Ly-2.1 (40) или необработанных (41), где % розеткообразующих клеток (ордината) представлен в зависимости от времени в днях (абсцисса).
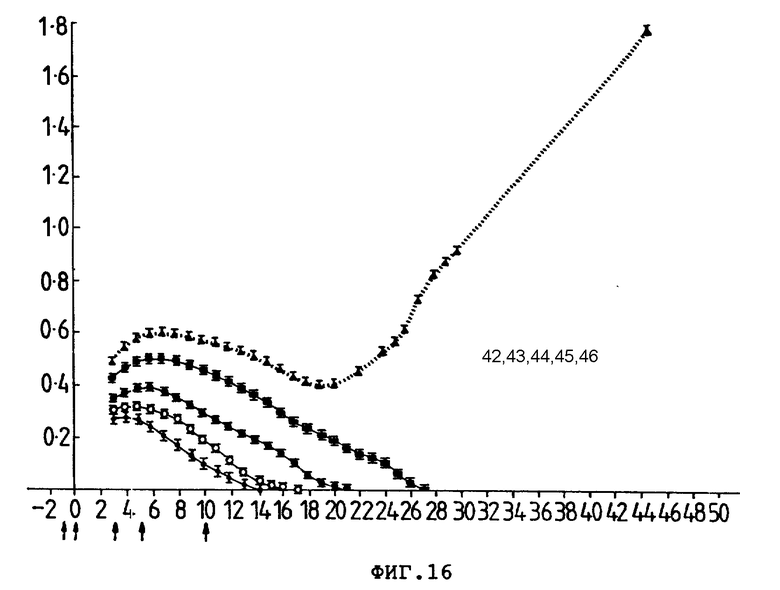
На фиг. 16 предварительно воздействие совместной обработки Ida-Моав-конъюгата на выживание трансплантата P388D1 опухоли (по разностям H-2 и не-H-2) у CBA-мышей. Группам из 10-15 мышей подкожно ввели 8•106 клеток P388D 1 опухоли и внутривенно (стрелки) один из препаратов: (I) PBS (42). (II) анти-L3T4 и анти-Ly-2.1 (43). (III) Ida-анти-L3T4 (44). (IV) Ida-анти-Ly-2.1 (45) и (V) Ida-анти-L3T4 и Ida-анти-Ly-2.1 (46). Средний размер (см2) опухоли (ордината) представлен в зависимости от количества дней после введения клеток опухоли (абсцисса). Гистограмма ошибок представляет ± стандартная ошибка среднего значения.
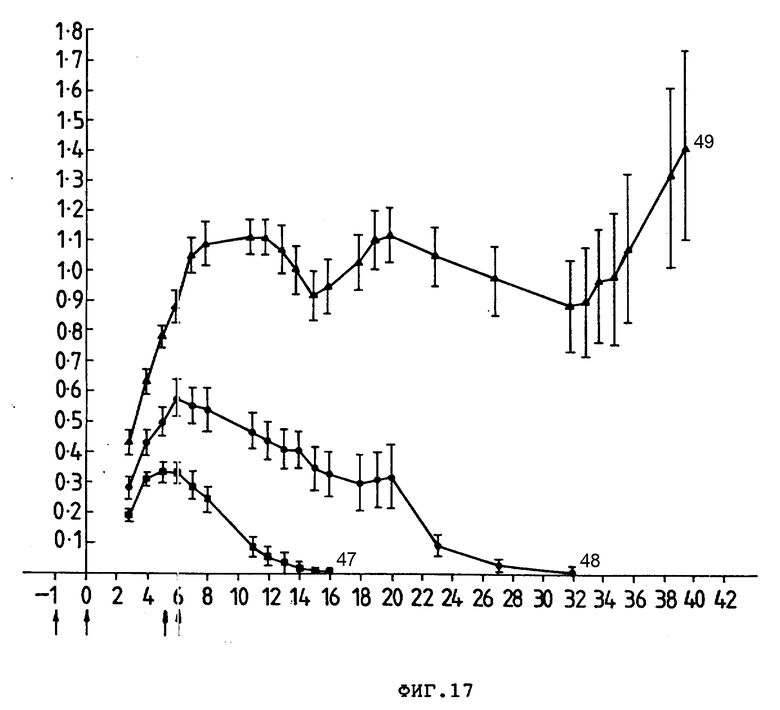
На фиг. 17 представлено воздействие Ida-анти-ILy-1-конъюгата на выживание трансплантата опухоли P388D1 у мышей CBA. Группам по 10 мышей подкожно ввели 1•106 клеток опухоли P383D1 и внутривенно (стрелки) один из препаратов: (I) PBS (47). (II) анти-TLy-1 (48) и (III) Ida-анти-TLy-1- (49). Средние размеры (см2) опухоли представлены (ордината) в зависимости от количества дней после введения клеток опухоли (абсцисса). Гистограмма ошибок представляет ± стандартная ошибка среднего значения.
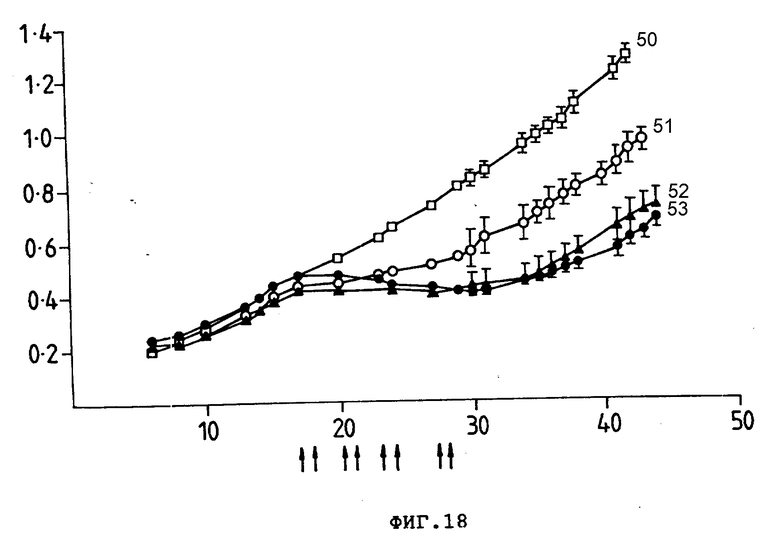
На фиг. 18 представлен рост ксенотрансплантата COLO205 (30,6+, 17,1+) у безволосных мышей, которым ввели 8•106 клетки каждой мыши. Группам по 10 мышей внутрибрюшинно ввели (показано стрелками) PBS (50), 17,1-Ida (51), 30,6-Ida (52) или смесь 30,6-Ida и 17,1-Ida (53). Среднее значение размера опухоли (см2) (ордината) представлено в зависимости от количества дней после введения клеток опухоли (абсцисса). Гистограмма ошибок показывает ± стандартная ошибка среднего значения. Общая доза Ida составила 200 мкг.
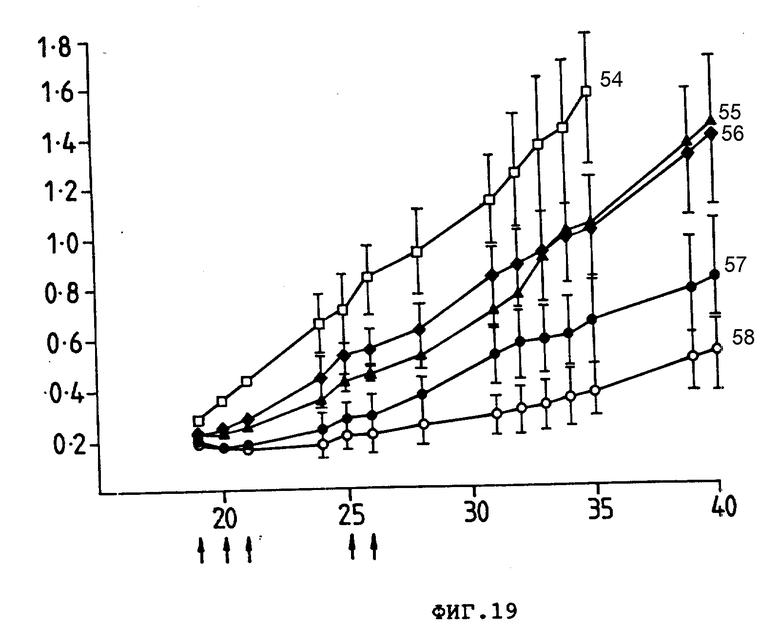
На фиг. 19 показан рост ксенотрансплантата опухоли обводочной кишки человека у безволосных мышей, который трансплантировали в виде фрагмента опухоли (1-5 мг). Группам мышей по 10 штук внутривенно ввели (обозначены стрелкой) PBS (54), 17,1-Ida (55). JGT-13-Ida (56), 27,1-Ida (57), 30,6-Ida (58). Средний размер опухоли (см2) (ордината) представлен в зависимости от количества дней после трансплантации опухоли (абсцисса). Гистограмма ошибок представляет ± стандартная ошибка среднего значения. Общая доза Ida составила 80 мкг.
Пример 1. Материалы и способы. Опухолевые клетки.
Линии клеток, исследуемые в данном примере включали вариант [E3] III(1)NSE3(Ly-2+) мышиной тимомы (Snuth ctal. , S.Natl Cancel 1985, 76, 503-510) ELy(Ly-2-, AR-), АИНФОМЫ (norowtz et al., Science 1968, 160, 533-535), CEM-линию (TFR+) человеческих клеток (Foley et al., Cancer 1955, 18, 522-529) и линию (-250-30,6+) человеческих клеток COLO205. Клетки сохранялись in vitro в среде Игла, модифицированной Дульбекко (DME), или в среде RPMI 1640 ( Flow laboratories, Sydney, Avstralia), в которую добавляли 10%-ную нагретую сыворотку новорожденного теленка (Flaw), 2 мМ глютамина (Commoniwealth Screen Laboratories, Sydney), 100 мкг/мл стремтомицина (Glaco, Melbauro, Australia) и 100 И.Е./мл пенициллина (Commoniwealth Screen Laboratoris). Опухоль E3 поддерживалась in vivo серийным пассажем в (C57BL(6•BALB/c/F1 мышь/мышь CBF1. Клетки из асцитической жидкости промыли и центрифугировали (400g•5 мин) два раза в фосфатно-буферном солевом растворе (PBS), pH 7,3, повторно суспендировали в PBS и подкожно ввели (S,c) в абдоминальную стенку мыши: эти операции привели к прощупываемым опухоли до лечения. Мышь подвергали серии внутривенных (i.v) или внутриопухолевых (i.t) обработок и ежесуточно измеряли последующий размер опухолей при помощи калибра-квадрата, производя измерения по перпендикулярным осям опухолей. Данные записывались как средний размер опухоли (произведение диаметров ± средняя квадратическая ошибка).
Мыши. CBA, (C57BL)6•BALB/c/CBP1мыши) и голые мыши (nu/nu) /были получены в Departament Pothology, University of Melbone. Экспериментальные группы из 8-10 мышей одного пола и возраста использовали в каждом эксперименте.
Моноклональные антитела. Использовали следующие моноклональные антитела: (I) анти-Ly-2.1(IgC1), реагирующее с мышиной Ly-2.1 специфичностью (Uogarth et al Immunology, 1982, 46, 135-144), (ii)A3C6 (анти-TFR) (IgC1), реагирующее с транфериновым рецептором человека (TFR) (Panaccio et al., Immunology and Cell Biology) и (iii) антитело, обозначенное 250-30,6, реагирующее с антигеном, присутствующим на клетках карциномы ободочной кишки человека.
Moab выделили из асцитической жидкости осаждением при помощи 40%-ного сульфата аммония, растворением в PBS и диализом тем же буферным раствором. Эти неочищенные препараты либо абсорбировали на протеин-A-сефарозе (Pharmacia Inc Piscataway, New Lersey), интенсивно промывали PBS (pH 7,3) и разбавляли 0,2 М глицин/HCl (pH 2,8), либо пропускали через синюю колонну Afligel (Bio-Rad Zaboratories Ptz Ltd Sydney). После нейтрализации Moab подвергли диализу по отношению к PBS, взяли аликвотную пробу и хранили при -70oC. A3C6 получили внутрибрюшинной иммунизацией мыши CBA с недельными интервалами в течение трех недель клетками LiCR-LON-HMy-2(HMy-2) (ОКТ9+Vl) в количестве 2•106, удаляя селезенки через трое суток после последней инъекции и сливая с клетками P3-NS1-AC4-1(NS-1).
Получение и количественная оценка конъюгатов.
Интактные анти-Ly-2.1, анти-IFR Moab или 250 - 30,6 (1 - 2 мг/мл в борном буферном растворе, pH 8) смешали с молярным избытком (1 - 50) 14-бром-4-деметоксидаунорубицина (Br-Ida), растворенного в N,N-диметилформамиде (ДМФ) при 10 мг/мл. Реакцию поддерживали при комнатной температуре 4 ч перед центрифугированием (400 g•5 мин) для удаления любого осадка. Свободный Br-Ida и другие исходные вещества удалили гель-фильтрационной хроматографией, используя колонну Sephadex C-25 (PD-10, Pharmacia), и конъюгаты затем пропустили через колонну Porapax Q (Millipore) для удаления любого поглощенного лекарства (Miederevieser et al. J Chromatog, 1971, 54, 215 - 223). Количество Ida, включенного в конъюгаты лекарство - Moab, определялось абсорбционной спектрометрией при 483 мм /E498 = 3,4•103 М-1 см-1 и протеиновой оценкой (Bredford, Anol Biochem 1976, 72, 248 - 253).
Активность антитела.
Тест на розеткообразование, использующий овечий антимышиный иммуноглобулин (SANG), применили для определения активности антитела Ida-Moab-конъюгатов по сравнению со свободными моноклональными антителами (Moab), которые подвергались тем ж процедурам, которые используются в методе сцепления (Parish and Mc. Kenzie, S. Iumunol Method, 1978, 20, 173 - 183).
Активность лекарства.
(a) 24-часовой тест на подавление: 100 мкл клеток (2 - 5•106 мл) внесли в микротетровальную чашку с плоским дном и выращивали 1 ч при 37oC. Свободный идарубицин (Ida), растворенный в PBS, и конъюгаты Ida-Moab асептически отфильтровали и в стерильном PBS выполнили разбавление 50 мкл свободного Ida или конъюганта, добавили в клетки, используя двойные лунки для каждой пробы; контрольные лунки получали 50 мкл PBC, и клетки выращивали при 37oC, 7% CO2 в течение 24 ч.
(b) 30-минутный тест на подавление: 200 мкл клеток (2 - 5•106 мл) собрали в стерильные пробирки Эппендорфа, повторно суспендировали в стерильном лекарстве или конъюгате и смешивали 30 мин при 37oC. Затем клетки центрифугировали (400 g•5 мин), повторно суспендировали в среде выращивания и 100 мкл аликвоты высеяли в микротитровальную чашку, используя двойные лунки для каждой пробы до инкубационного периода 16 - 24 ч. После инкубационного периода в обеих этих пробах добавили 50 мкг питательной среды, содержащей I и Ci[3H] -тимидина (удельная активность 5Ci/ммоль); Amersham/ и чашки выращивали 2 - 4 ч. Клетки затем собрали, обезводили, отдельные пробы разделили и посчитали на бетасцинтилляционном счетчике. Введение [3H]-тимидина выражалось как процент подавления до введения контрольных проб. Стандартная ошибка для любой точки определялась двойными определениями и не превышала 5% для любой заданной экспериментальной точки.
Токсичность. Группам из 10 - 20 CBA-мышей давали одну внутривенную инъекцию различных доз Ida или Ida-анти-Ly-2.1, и наблюдалась выживаемость мышей по отношению к дозе лекарства, выраженной в мг/кг. Органы этих мышей удалили и взвешивали до фиксации в формалине и окрашивании гемотоксилином и эозином.
Результаты. Получение и определение характеристик конюгатов.
Br-ida (фиг. 1) ковалентно соединили с несколькими Moab: против человеческого TER, против антигена, присутствующего на раковых клетках ободочной кишки человека (антитело 250 - 30,6), и против Ly-2-аллоантигена мыши. Условия реакции установили для конъюгации, изменяя молярный избыток Br-Ida, добавленного в Moab, и идя на компромисс между более высоким введением Ida и более низкими протеиновыми выделениями. Ida-анти-Ly-2.1 (фиг. 2), Ida-анти-TER и Ida-250-30,6 (данные не показаны) включали 3 - 5 молекул Ida при протеиновых выделениях больше 50%. Реакция Br-Ida с моноклональными антителами (Moab) может обеспечить два типа связей (фиг. 1C и D). Для установления того, что было представлено, конъюгаты подвергли воздействию pH 4,5 или 9 в течение 48 ч, освобожденное свободное лекарство поглощали в Porapab Q и пробы повторно количественно определяли спектрометрией. 50% связанного лекарства было освобождено при воздействии основания (pH 9), в то время как при pH 4,5 потерь не наблюдалось. Это позволяет предположить, что по меньшей мере 50% лекарства имеют сложноэфирную связь (фиг. 1D), так как сложноэфирная связь чувствительна к основным условиям, пока аминная связь является устойчивой.
Активность антитела.
Титры антитела до и после конъюгации измеряли методом розеткообразования и они определялись как разбавление, при котором 50% клеток-мишеней демонтрировали розетки. Ida-анти-Ly-2.1-конъюгаты, содержащие 2 и 8 молекул Ida, имели титры антитела по отношению к клеткам B3 1:56000 и 1:33000 соответственно, в то время как титр неконъюгатированных антител был равен 1:8000 (фиг. 3). Ida-250-30,6 - конъюгаты, содержащие 2 и 6 молекул Ida, имели титры антител против клеток COLO 205 1:16000 и 1:11000 соответственно, в то время как титр неконъюгатированных антител был 1:33000. Таким образом, наблюдается некоторая потеря активности антител из-за процедуры конъюгации; конъюгаты с менее 6 молекулами Ida-молекулой Moab использовали для исследований in vitro и in vivo. Было отмечено, растворимость и активность антител Ida-анти-Ly-2.1-конъюгатов значительно снижаются ниже этих уровней внесения (данные не приводятся). Максимальное число молекул Ida, которые можно полезно вносить, сильно меняется в зависимости от индивидуального антитела.
In vitro-активность и конъюгаты индарубицина и индарубицина-Моав.
In vitro-цитоксичность Ida и двух Ida-Moab-конюгатов на линии клеток мыши ITT (1) 75 NSE 3 (Ly2+ TFR-) и линии клеток CEM человека (Ly - 2-TFR+) измеряли в 24-часовом тесте подавления и определяли И.Д.50 (50% подавления при внесении стандартов [3H]-тимидина). Результаты показаны на фиг. 4 и в табл. 1. И. Д.50 для Ida находилась в диапазоне 1,0 - 2,5•10-7 М для обеих проверяемых линий клеток (фиг. 4, табл. 1). И.Д.50 для Ida-анти-Ly-2.1 на E3 была в 4 раза больше (фиг. 4) и для Ida-анти-TER на CEM значения И.Д.50 были в 1 - 2 раза больше, чем для свободного Ida (табл. 1). Следовательно, свободный Ida был более цитотоксичным по отношению к E3 и CEM, чем Ida-анти-Ly-2.1 и Ida-анти-TFR соответственно. Однако эти конъюгаты Ida-Moab проявляют 10-кратную меньшую цитотоксичность к нереагирующим линиям клеток (табл. 1), отмечая таким образом, что их цитотоксичное воздействие было специфичным и являлось результатом задержки активности антител (фиг. 3).
1 И.Д.50 - 50%-ное подавление при введении стандартов [3H]-тимидина;
2 число проверяемых препаратов.
Конъюгаты Ida-250-30,6 были слегка менее активными, чем свободный Ida. Свободный Ida имел И. Д.50 6 • 10-8М, в то время как конъюгат имел И.Д.50 3,55 • 10-7М на линии клеток-мишеней COLO 205.
Для изучения, проявляют ли конъюгаты избирательность в их цитотоксичном действии по отношению к клеткам-мишеням Ida-анти-LY-2.1 и Ida-анит-TFR выращивали 30 мин с E3 (Ly-2+)-клетками до выливания несвязанного конъюгата и измерения цитотоксичности. Ida-анти-Ly-2.1-конъюгат имел И.Д.50 6,2 • 10-7М по сравнению с И.Д.50 5.2 • 10-7М для свободного Ida (фиг. 5). В противоположность этому нереагирующий Ida-анти-TFR-конъюгат имел И.Д.50 5 • 10-6М, т.е. в 10 раз больше, чем для свободного Ida, показывая, что активность связывания антител Ida-анти-Ly-2.1-конъюгата приводит к его избирательной цитотоксичности. Аналогично Ida-250-30,6-конъюгат и свободное лекарство выращивали 30 мин с COLO 205 (250-30,6+Ve) и E3 (250-30,6-Ve) линиями клеток пере промывкой и проведением испытания на цитотоксичность. Обе линии клеток проявили аналогичную реакцию на дозу свободного лекарства, т.е. 9,2 • 10-7М для COLO 205 и 9,8 • 10-7М для E3. Однако, Ida-250-30,6-конъюгат был в 4 раза более токсичным по отношению в COLO 205, чем к нереактивной линии клеток E3 антитела. Аналогичные результаты получили, используя линию клеток CEM и Ida-анти-Ly-2.1 как нереактивный стандарт (данные не показаны). Для дополнительной убежденности, что цитотоксичность конъюгатов Ida-Moab для клеток-мишеней была специфической и имела место в сайте связывания антитела, мы провели исследования по ингибированию цитотоксичности конъюгата, используя свободные Moab. При концентрации Ida 4 • 10-6М (2 мкг анти-Ly-2.1) цитотоксичности конъюгата анти-Ly-2.1 на E3-клетках была снижена на 70% после добавления 50 мкг (250 мкг/мл) анти-Ly-2.1. (фиг. 6), указывая, что цитотоксичность конъюгата Ida-анти-Ly-2.1 непосредственно связана с ее характеристикой связывания антител. Аналогичные контрольные результаты получили с 260-30,6. Следует отметить, что во всех тестах свободные анти-Ly-2.1, анти-TFR и 250 - 30,6 были нецитотоксичными (данные не показаны).
In vitro-лечение клеток мышиной тимомы ITT(1) 75 NS T3.
Для оценки подавления роста твердых опухолей группы мышей CBF1 (10 особей в группе), привитых 2 • 106R3-клетками подкожно в абдоминальной области, лечили внутривенными инъекциями одного из следующих препаратов: (I) PBS); (U) анти-Ly-2/1; (iii) (Ida; (iv) Ida-анти-TFR, или (v) Ida-анти-Ly-2/1. Мыши получали 20 мкг Ida и/или 1200 мкг анти-Ly-2.1 соответственно на 4- и 5-е сутки (размер опухоли 0,1 см2) после инокуляции.
В пределах 24 ч первой обработки обработанные Ida анти-Ly-2.1 мыши имели средний размер опухоли, равный 20% опухоли леченных PBS мышей (т.е. 80% уменьшение массы опухоли (фиг. 7)). Очевидно, что один препарат анти-Ly-2.1 и Ida, ковалентно связанный с неспецифическим анти-TFR и Moab, не влияли на рост опухоли E3. Опухоли мышей, получающих только Ida, уменьшались до 50%; однако 3 из этих мышей погибли, а другие имели 25%-ное снижение массы тела. Индивидуальные кривые роста опухолей мышей, принимающих Ida-анти-Ly-2.1, показали снижение 9 из 10 опухолей во время курса лечения (фиг. 8); 5 из 10 опухолей полностью регрессировали и не появились повторно (200 сут) и те опухоли, которые продолжали расти после завершения лечения (5 из 10), росли меньшими скоростями, чем опухоли мышей, которые лечили PBS и Ida-анти-TFR. Был проведен дополнительный эксперимент для оценки внутривенного лечения больших опухолей, используя Ida-анти-Ly-2.1. Группам CBF1-мышей (10 особей в группе) инокулировали 3 • 106 E3-клеток, и мыши затем получили 15 мкг и 900 мкг Ida и инти-Ly-2.1 соответственно на 6 (размер опухоли 0,2 см2) и 7-е сутки после инокуляции опухоли (фиг. 9). Мыши, обработанные Ida-анти-Ly-2, имели средний размер опухоли на 50% меньше, чем мыши, обработанные PBS, и 66% опухоли мышей, обработанных Ida на седьмые сутки. Эта тенденция продолжалась до окончания исследования (18-е сут). Индивидуальные кривые роста опухолей 10 мышей CBF1, принимавших Ida-анти-Ly-2.1, показывают, что было 4 регрессии и одно полное исчезновение массы опухоли ( >200 сут. данные не приводятся). Следовательно, Ida-анти-Ly-2.1 был эффективен против больших опухолей, и в обоих экспериментах антиопухолевая активность Ida была значительно улучшена при связывании с анти-Ly-2.1 Moab.
In vitro - лечение опухоли ободочной кишки человека COLO 205.
Воздействие Ida-анти-250-30,6-конъюгата оценивалось на голых мышах (пи/пи), порождающих ксенотранcплантаты COLO 205.
Инъекции подкожно в абдоминальную стенку 2 • 106 клеток привели к пальпируемой опухоли через 4 сут (примерно 0,1 см2). Группы из 10 мышей затем лечили внутривенными инъекциями одного из следующих препаратов: (I) PBS; (ii) 250 - 306; (iii) Ida плюс 250 - 30,6 (неконъюгированные); (vI) Ida; (v) конъюгат Ida-250-30,6. Всего ввели 275 мкг Ida сериями внутривенных инъекций (5) на 4-, 5-, 6-, 10-, и 12-у сутки после инокуляции опухоли.
В группах мышей, которые лечили PBS или неконъюганированным 250-30,6, не наблюдался терапевтический эффект. При применении только Ida 2 из 10 мышей выжили, в то время как все мыши из группы, получавший неконъюганированный Ida плюс 250-30,6, характеризовались резким уменьшением размера опухоли. Эти результаты показаны на фиг. 11. В противоположном этому кривые роста опухолей для отдельных мышей (фиг. 12) показывают, что 5 из 10 мышей имели опухоли, которые регрессировали на 7-е сутки, эти опухоли затем продолжали расти, хотя 2 из 10 мышей остались без опухолей. Эти мыши не проявляли симптомов токсичности.
Внутриопухолевое лечение.
Внутриопухолевая терапия оказалась полезным способом для иммунотерапии опухолей человека и животных. Следовательно, провели исследования для определения характеристик противоопухолевой активности конъюгатов Ida-анти-Ly-2.1 при введении их непосредственно в твердую E3-опухоль. У групп из 10 мышей CBF1, которым подкожно имплантировали 3 • 106 E3-клеток, развились опухоли (0,1-0,2 см2) на 5-е сутки после инокуляции опухоли. Лечение состояло из 2 инъекций на 5- и 5-е сутки после имплантации опухоли, причем мыши получали одно из следующих лечений: (1) PBS; (2) Ida; (3) анти-Ly-2.1 или (4) Ida-анти-Ly-2.1 (общее количество Ida 30 мкг). Ida-анти-Ly-2.1 проявил наибольшую антиопухолевую активность, свободный Ida и только анти-Ly-2.1 не проявили влияние на рост опухоли при введении прямо в опухоль. Мыши, которые получали Ida-анти-Ly-2.1, имели средний размер опухоли, равный 60% опухоли мышей, которые получали PBS на 8-е сутки, и 30% опухоли мышей, которые получали PBS на 13-е сутки (фиг. 10). Индивидуальные кривые роста опухолей (данные одну полную регрессию, в то время как остальные мыши проявили задержанное снижение роста опухоли на 3-и сутки после завершения лечения.
Токсичность.
Для экспериментов по острой токсичности группам из 10 CBA (Ly-2.1+)-мышей давали одну инъекцию различных доз Ida, Ida-анти-Ly-2.1 или Ida-анти-TFR. Все мыши, которым вводили Ida, показали начальную потерю веса до 25% от исходного веса, однако у мышей, обрабатываемых любым конъюгатом Ida-Moab, не наблюдалась потеря веса. В табл. 2 показана токсичность Ida и Ida-Moab-конъюгатов, выраженная в значениях Л.Д.50 и Л.Д.10. Как показано, Л. Д. 10 Ida-анти-Ly-2.1 была 10 мг/кг Ida по сравнению с только 0,75 мг/кг для свободного Ida. В дополнение конъюгат Ida-анти-TFR имел Л.Д.10 8 мг/кг. Конъюганты Ida-Moab не проверялись на Л.Д.50 - дозу. Эти результаты показывают самый большой терапевтический индекс конъюгатов Ida-Моав по сравнению со свободным Ida.
Гистопатологические результаты.
Острый эффект: внутривенное введение свободного Ida (1 мг/кг) приводит в атрофии белой пульпы в селезенке на 15-е сутки после лечения и некоторой гипертрофии сердечных мышечных волоком (данные не приводятся). В противоположность этому одна доза Ida-анти-Ly-2.1 (2,4 мг/кг) не вызывала неспецифической тканевой токсичности через 15 и 30 сутки, хотя некоторое набухание гепатоцидов наблюдалось на 15-е сутки.
Пример 2. Вещества и способы.
Мыши. На факультете патологии Мельбурнского университета получили мышей вида DBA/2 x BALB/c/F1 и 6BA. В каждом эксперименте использовали группы из 10-20 мышей одного пола и возраста.
Опузолевые клетки.
Линии клеток, изучаемые в данном исследовании, включают вариант E3/Ly-2+ мышиной тимомы ITT(1) 75 NS (E3), EL4-лимфомы (L3T4+) (Horowitz et al, Seience, 1968, 160, 533-535), COL0 карциномы ободочной кишки человека (Semple etal, Cancer Ru 1976, 38, 1345-355) и CEM легкомии T-клеток человека (Ly-2-. L3T4-) (Foley et al., Cancer, 1965, 18, 522-529). Клетки сохранялись in vitro, как описано в примере 1. Линии макрофагов P338D1 сохранялась in vivo серийным пассажем в мышь (DBA/2 • BA/B/c/F1. Клетки из асцитной жидкости промывали и центрифугировали (400g • 5 мин) два раза в фосфатбуферном солевом растворе (PBS, pH 7,3), повторно суспендировали в PES и подкожно вводили в абдоминальную стенку мыши, где они развивались в пальпируемые опухолевые трансплантаты. Мыши затем подвергались сериям внутривенных лечений, размер опухолей ежесуточно измеряли при помощи калибра-квадрата, измеряя опухоли по перпендикулярным осям. Данные записывались как средний размер опухоли (произведения двух диаметров ±стандартная ошибка.
Моноклональные антитела.
Моноклональные антитела к мышиным клеточным поверхностным антигенам L3T4, Ly-2 и ThY-1, которые характеризуют отдельные субпопуляции лимфоцитов, использовались как модель.
Использовались следующие Моав: (i) анти-Ly-2.1 (мышиный IgC2a), реактивное со специфичность мышиного Ly-2.1 (Horgarth et al, Immunology 1982, 46, 135-144), (ii) H129.19 (анти-L3T4) (IgC2 крысы), реактивное со специфичностью мышиного L3T4 (Pierreset al., J.Immunology 1984, 132, 2775-2782) и (iii) анти-TLy-1 (IgG2 крысы), реактивное с мышиными T-клетками (Marshak-Rothstech et al. , J. Immunology 1979, 122, 2491-2497). Антитела выделили, очистили и хранили, как в примере 1. Активность антител определяли тестом на розеткообразование с овечьим антимышиным иммуноглобулином (CAMC), как описано в примере 1.
Получение конъюгатов идарубицин-моноклональное антитело.
Moa b (1-2 мг/мл) мешали с 5-20 молярным избытком 14-бром-4-деметиоксидаунорубицина (Br-Ida), растворенного в N, N-диметилформамиде (10 мг/мл), 4 ч при pH 8 (0,05 M боратный буферный раствор) и комнатной температуре. Провели реакцию и очистку, и количество Ida в результирующем конъюгате определили, как описано в примере 1.
Активность лекарства.
Два теста (24 ч и 30 мин) провели для оценки активности лекарства аналогично примеру 1, используя клетки при концентрации 1-5•106 мг/мл.
Серология.
Для определения титра антитела и числа клеток L3T4+ и Ly-2+ в этих экспериментах использовали розеткообразования. Этот метод включает связывания SAMG и он также определяет иммуноглобулин крыс (Ig), связанный с овечьими эритроцитами (SRC) с целью обнаружения антител на поверхности лимфоидных клеток. Ig+ селезеночные клетки имели поверхностный Ig, удаляемый при помощи блокировки с SAMG (25 мкл и 2 мл клеток при 107 мкл), клетки затем сохраняли на льду в среде, содержащей 0,01% азида натрия для предотвращения ресинтеза иммуноглобулина.
Результаты.
Исследование проводилось в раздельных фазах:
(a) in vitro - определение характеристики трех различных Ida-Моав-конъюгатов;
(b) последующая демонстрация их использования в активном удалении подкласса T-клеток до и во время отторжения аллотрансплантата опухолевых клеток.
Приготовление и определение характеристик конъюгатов.
Br-Ida ковалентно связали с Moab против мышиных L3T4, Ly-2 и Thy-1 антигенов. Условия реакции устанавливались для конъюгации, изменяя молярный избыток Br-Ida, добавляемого к Moab и соблюдая компромисс между более высоким введением Ida и более низкими выделениями протеина. Ida-анти-L3T4 (фиг. 13), Ida-анти-Ly-2.1 и Ida-анти-Thy-1 (данные не приведены) включали 3-6 молекул Ida при выделениях протеина более 50%.
Активность антител.
Титры антител до и после конъюгации измерялись методом розеткообразования и определялись как разбавление, при котором 50% клеток-мишеней E3 образуют розетки. Ida-анти-Thy-1-конъюганты, содержащие 1,4 и 7 молекул Ida, имели титры антител 1:425000, 1:170000 и 1:130000, соответственно, в то время как титр немодифицированного антитела был равен 1:550000 (фиг. 14). Таким образом, после конъюгации с Ida наблюдается некоторая потеря активности антител. Однако конъюгаты с менее чем 4 молекулами Ida-молекулы c Moab использовали для in vito и in vito - исследований. Аналогично, активность антител обоих конъюгатов Ida-анти-Ly-2.1 и Ida-анти-L3T4 (данные не приводятся) резко уменьшается при введении более 6 молекул Ida-молекулы с Moab.
Активность лекарства in vitro.
Цитотоксичность конъюгатов Ida-Моав проверяли на различных реактивных клетках-мишенях, используя 24-часовой тест, и сравнивали со свободным Ida. Активность свободного Ida в 4-10 раз выше, чем Ida-Moab-конъюгатов при И.Д. 50 (50%-ное ингибирование во введении стандартов [3H]-тимидина) свободного Ida, имеющей место при 6,6-9,0•10-8N, против проверяемых линий опухолевых клеток. Было видно, что Ida-анти-Ly-2.1-конъюгат является наиболее цитотоксичным конъюгатом (И.Д.50-4,3•10-7М) при проверке против линии клеток ITT(I) 75NS E3, которая является вариантом нативной линии клеток, улучшенной для антигена Ly-2. Для изучения избирательности воздействия конъюгата на клетки-мишени Ida-Moab-конъюгаты выращивали 3 мин с клетками-мишенями, затем вымыли несвязанный конъюгат и измеряли цитотоксичность. Используя этот 30-минутный тест, выявили, что нереактивные Ida-Moab-конъюгаты имели И.Д.50 в 10-50 раз больше, чем И.Д.50 свободного Ida, показали этим, что связывание антитела является существенным для цитотоксичности Ida-Moab-коньюгата. Следует отметить, что ни одно Moab, использованное в данном исследовании, не оказывало цитотоксического воздействия на клетки-мишени in vitro в отсутствии комплемента.
Результаты сведены в табл. 3.
In vivo - потенция конъюгатов идарубицин-моноклональное антитело.
In vivo-иммунодепрессивная потенция конъюгатов Ida-Moab сравнивалась с этой характеристикой одного Моав при помощи проверки способности этих конъюгатов избирательно убирать из селезенки клетки L3T4+ или Ly-2+. Мыши CBA получали 4 внутривенных инъекции анти-Ly-2.1 или анти-L3T4-конъюгата (30 мкг Ida/1,5 мг Моав) в 0-, 2-, 5- и 10-й день, и число Ly-2+- или L3T4+- клеток в селезенке контролировали при помощи теста на розеткообразование. Для каждого лечения клетки селезенки двух мышей, которые получали препарат, исследовались каждый день и результаты усреднялись (фиг. 15). Наблюдалось резкое снижение числа L3T4+-клеток от примерно 30% от общего числа селезеночных клеток у нормальных мышей на 0 день до примерно 4% у мышей, которые получали Ida-анти-L3T4, на 20-й день. Эти L3T4+-клетки оставались истощенными в течение более чем 60 сут до того, как стало наблюдаться постепенное увеличение. Истощение после in vivo-лечения одним анти-L3T4 также привело к резкому снижению числа клеток L3T4+ (от 30 к 5%).
Конъюгат Ida-анти-Ly-2.1 снижал число клеток селезенки Ly-2+ от 25 до 5% на 10-й день. Однако число клеток Ly-2+ начинало увеличиваться на 15-й день и возвращалось к норме на 40-50-й день. Было интересно отмечать, что одно анти-Ly-2.1 Моав было неспособно истощить Ly-2+-клетки в значительной степени. Колебания в числе клеток Ly-2+ и L374+ у нелеченых контрольных мышей, как предполагается, связаны с естественными колебаниями между особями. Таким образом, стало ясно, что оба Ida-анти-L374 и Ida-анти-Ly-2.1 могут удалять L3T4+- или Ly-2+-клетки соответственно из селезенок обработанных мышей более эффективно, чем одно Moab. Влияние Ida-анти-Ly-2.1-конъюгата важно, так как анти-Ly-2.1 Моав не оказывает никакого влияния при использовании только его одного, но может превратиться в потенциальный иммунодепрессивный агент при связи с Ida. Истощение, поддерживаемое конъюгатами лекарство - Моав, было, следовательно, пригодным для изучения роли клеток Ly-2+ и L3T4+ in vivo в реакции отторжения трансплантата.
Воздействие Ida-анти-L374, Ida-анти-Ly-2.1 и Ida-анти-TLy-1 на выживание опухолевого трансплантата.
В этих экспериментах Ida-анти-L374-, Ida-анти-Ly-2.1- и Ida-анти-TLy-1-конъюгаты использовались для увеличения времени выживания опухолевых трансплантатов P388D1 у CBA-мышей. Аллотрансплантаты осуществлялись по H-2 (класс 1 и 11) и не-H-2 районам.
(a). Комбинированное лечение конъюгатами.
Группам из 10-15 мышей CBA подкожно вводили опухолевые клетки P388D1 в количестве 8•106 и внутривенно на -1-, 0 (день инокуляции опухоли)-, 3-, 5- и 10-е сутки один из следующих препаратов: (I) PBS, (ii) анти-L3T4 и анти-Ly-2.1, (iii) Ida-анти-L3T4, (iv) Ida-анти-Ly-2.1 и (v) Ida-анти-L3T4 и Ida-анти-Ly-2.1. Суммарное количество введенных Ida и Моав было 125 мкг и 5,75 мк соответственно.
Опухолевые трансплантаты мышей, которые получали PBS и Моав, выживших в течение 14-17 сут с максимальным средним размером опухоли 0,31 см2, показали, что оба неконъюгированные моноклональные антитела вместе неспособны эффективно истощать L3T4+- и Ly-2+-клетки так, чтобы опухолевый трансплантат мог выжить (фиг. 16). В дополнение Ida-анти-Ly-2.1 или Ida-анти-L3T4 по отдельности были способны только продлить выживание трансплантата (20-28 сут) при максимальных средних размерах опухолей 0,40-0,50 см2. По-видимому, один Ida-анти-Ly-2.1 или Ida-анти-L3T4 не может удалить все T-клетки, и сильное антигенное заражение (как видно при всех различиях MHC) способно вызвать пролиферацию оставшихся Ly-2+- и L3T4+-клеток и реакцию отторжения. В противоположность этому комбинация Ida-анти-Ly-2.1- и Ida-анти-L3T4-конъюгатов позволила 14/15 опухолевым транспластатам P388D1 избежать отторжения и увеличиться в размере, пока мышей не умертвили (50-й день - 2 см2). Было отмечено, что несколько из этих обработанных конъюгатами опухолей P388D1 демонстрировали определенную регрессию в размере на 8-16-е суки; однако постоянное истощение L3T4+- и Ly-2+-клетки (10-й день) позволило опухолевым трансплантатам P388D1 установить стойкий рост опухоли.
(b) Ida-анти-TLy-1-конъюгат.
Группам из 10 мышей CBA подкожно вводили опухолевые клетки P388D1 в количестве 107 и внутривенно на -1-, 0-, 5- и 6-й день один из следующих препаратов: (I) PBS, (ii) анти-TLy-1 и (iii) Ida-анти-TLy-1. Суммарное количество введенного Ida и анти-TLy-1 было 130 мкг и 5,4 мк соответственно, и опухолевые трансплантаты обработанных PBS мышей жили в течение 15 сут. Одно анти-TLy-1 Моав поддерживало выживание опухолевого трансплантата в течение 28-32 сут при максимальном среднем размере опухоли 0,53 см2 (6-е сутки) (фиг. 17). У мышей, которые получали Ida-анти-Thy-1 30% опухолевых трансплантатов полностью были отторгнуты на 40-е сутки, в то время как оставшиеся 70% продолжали расти, что, в конечном счете (32-е сутки), позволило увеличить средний размер опухоли группы. Следовательно, Ida-анти-Thy-1 аналогично комбинации Ida-анти-Ly-2.1 и Ida-анти-L3T4 был способен истощить Ly-2+- и L3T4+-клетки так, что большинство опухолевых трансплантатов P388D1 выживало у мышей CBA при отсутствии нормальной реакции отторжения.
Пример 3. Приготовление конъюгаты в соответствии с процедурой, описанной в примере 1, между моноклональным антителом 17,1 и Ida и между Ida и моноклональным антителом 30,6. Клетки COLO 205 карциномы ободочной кишки человека (30,6+, 17,1+) подкожно инокулировали безволосым мышам в количестве 8•106 клеток/мышь, как описано в примере 2. Затем инокулированные мыши подвергались серии внутрибрюшинных инъекций. Размер опухоли измеряли калибром-квадратом, измеряя ее по перпендикулярным осям опухолей. Данные записывались как средний размер опухоли (произведение двух квадратов ± стандартная ошибка). Результаты показаны на фиг. 18.
Пример 4. Приготовили конъюгаты в соответствии с процедурой, описанной в примере 1, между Ida и моноклональным антителом 17,1, между Ida и моноклональным антителом ICT-13, между Ida и моноклональным антителом 27,1 и между Ida и моноклональным антителом 30,6. Безволосым мышам имплантировали ксенотрансплантат опухоли: ободочной кишки человека LIM2210 (1-5). Затем имплантированным мышам сделали серию внутривенных инъекций. Размер опухоли измерили калибром-квадратом, измеряя ее по перпендикулярным осям опухолей. Данные записывали как средний размер опухоли (произведение двух диаметров ± стандартная ошибка). Результаты показаны на фиг. 19.
Изобретение относится к области медицины и касается коньюгатов иммуноглобулина с производным 14-бромдаунорубицина, обладающих противоопухолевой активностью. Сущность изобретения заключается в разработке коньюгатов моноклонального антетела с 14-бром- 4-деметоксидаунорубицина, при этом коньюгат содержит от 2 до 8 молекул лекарства, ковалентно связанного с антителом или фрагментом, а также фармацевтической композиции, содержащей данный коньюгат, а также приемлемый разбавитель или носитель. Технический результат заключается в повышении специфичности доставки лекарства к опухоли. 2 с.п.ф-лы, 3 табл., 19 ил.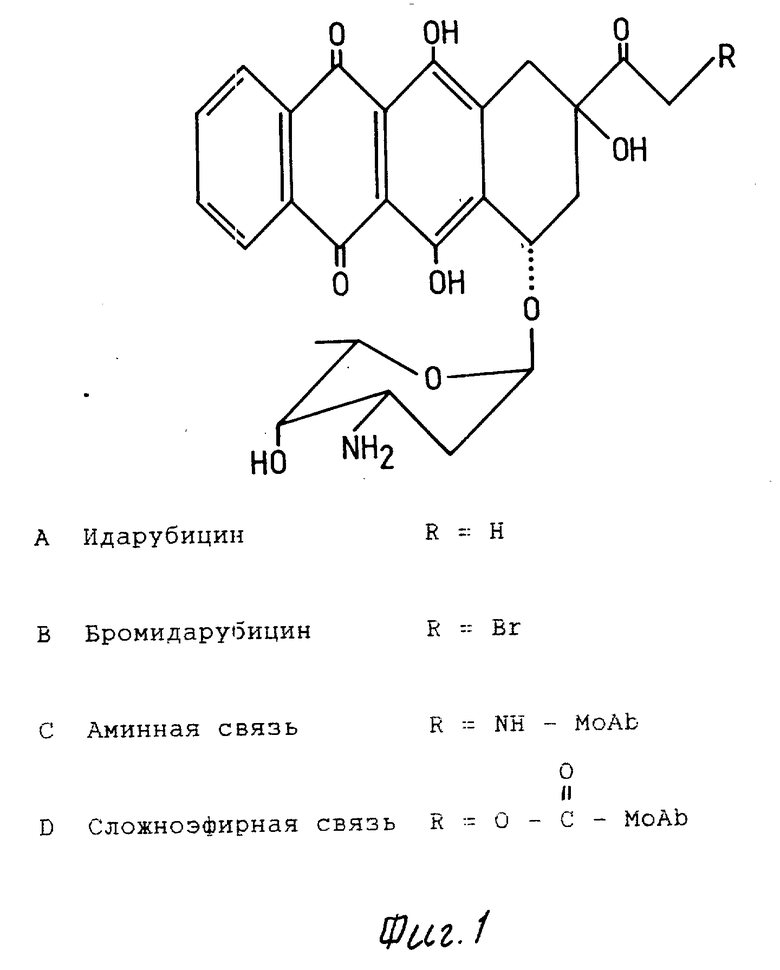
| Gallego et al., Int | |||
| J.Cancer, v.33, 737-744, 1984. |
