Данная заявка заявляет эффект приоритета заявки США 09/081385, поданной 14 мая 1998, находящейся в процессе одновременного рассмотрения. Для целей ведения дела в США приоритет указанной заявки включен тем самым здесь в качестве полной ссылки.
Данное изобретение относится в общем к области трансдукции сигналов (передачи сигналов) между клетками, через цитокины и их рецепторы. Более конкретно, оно относится к ферментативной активности, которая расщепляет и высвобождает рецептор для TNF, обнаруженный на клеточной поверхности, и последующим биологическим эффектам. Определенными вариантами осуществления данного изобретения являются композиции, которые влияют на подобную ферментативную активность и могут быть включены в лекарственные средства для лечения заболеваний.
Цитокины играют центральную роль в коммуникации между клетками. Секреция цитокина из одной клетки в ответ на стимул может стимулировать соседнюю клетку к подходящему биологическому ответу - такому как стимуляция, дифференцировка или апоптоз. Высказывается гипотеза, что на важные биологические события может влиять не только высвобождение цитокинов из первой клетки, но также связывание с рецепторами на второй клетке, которое опосредует последующий ответ. Данное изобретение, описанное в этой патентной заявке, обеспечивает новые соединения для воздействия на трансдукцию сигнала от фактора некроза опухолей.
Цитокин, известный как фактор некроза опухолей (TNF или TNF-α), является структурно родственным лимфотоксину (LT или TNF-β). Они имеют около 40 процентов гомологии аминокислотной последовательности (Old, Nature 330:602-603, 1987). Эти цитокины высвобождаются макрофагами, моноцитами и природными клетками-киллерами и играют роль в воспалительных и иммунологических событиях. Эти два цитокина вызывают широкий спектр эффектов как in vitro, так и in vivo, в том числе (i) васкулярный тромбоз и некроз опухолей; (ii) воспаление; (iii) активацию макрофагов и нейтрофилов; (iv) лейкоцитоз; (v) апоптоз и (vi) шок. TNF связывали с разнообразными патологическими состояниями, в том числе с различными формами рака, артритом, псориазом, эндотоксиновым бактериально-токсическим шоком, сепсисом, аутоиммунными заболеваниями, инфекциями, ожирением и кахексией. TNF, по-видимому, играет роль в трех факторах, способствующих регуляции веса тела: потреблении, расходовании и накоплении энергии (Rothwell, Int. J. Obesity 17:S98-S101, 1993). В случае септицемии увеличенные концентрации эндотоксина, по-видимому, повышают уровни TNF (Beutler et al., Science 229:869-871, 1985).
Делались попытки изменения хода заболевания лечением пациента ингибиторами TNF, с варьирующимися степенями успеха. Например, ингибитор TNF дексанабинол обеспечивал защиту против опосредованных TNF действий после травматического повреждения головного мозга (Shohami et al., J. Neuroimmun. 72:169-77, 1997). Некоторое улучшение при болезни Крона было получено лечением антителами против TNF (Neurath et al., Eur. J. Immun. 27:1743-50, 1997).
TNF и LT человека опосредуют их биологические активности связыванием специфически с двумя различными гликопротеиновыми рецепторами плазматической мембраны (с размерами 55 кДа и 75 кДа, известными как TNF-R р55 и TNF-R р75 соответственно). Эти два рецептора имеют 28%-ную гомологию аминокислотной последовательности в их внеклеточных доменах, которые состоят из четырех повторяющихся обогащенных цистеинами районов (Tartaglia and Goeddel, Immun. Today 13:151-153, 1992). Однако эти рецепторы не имеют значимой гомологии последовательности в их внутриклеточных доменах и опосредуют различные внутриклеточные ответы на активацию рецепторов. В соответствии с различными активностями TNF и LT большая часть клеток человека экспрессирует низкие уровни рецепторов TNF: приблизительно 2000-10000 рецепторов на клетку (Brockhaus et al., Proc. Natl. Acad. Sci. USA 87:3127-3131, 1990).
На экспрессию рецепторов TNF как на лимфоидных, так и на нелимфоидных клетках можно подействовать экспериментально многими различными агентами, такими как бактериальный липополисахарид (ЛПС), форболмиристатацетат (диэфир форбола, ФМА; активатор протеинкиназы С), интерлейкин-1 (IL-1), интерферон-гамма (IFN-γ) и IL-2 (Gatanaga et al., Cell Immunol. 138:1-10, 1991; Yui et al., Placenta 15:819-835, 1994). Было показано, что комплексы TNF человека, связанные с его рецептором, интернализуются из клеточной мембраны, и затем рецептор подвергается либо деградации, либо рециркуляции (Armitage, Curr. Opin. Immunol. 6:407-413, 1994). Было сделано предположение, что активность рецептора TNF может модулироваться при помощи пептидов, которые связываются внутриклеточно с этим рецептором или которые связываются с сайтом связывания лиганда, или которые действуют на выделение рецептора из клетки. См., например, патентные публикации WO 95/31544, WO 95/33051, WO 96/06142 и ЕР 568 925.
TNF-связывающие белки (TNF-BP) были идентифицированы при повышенных уровнях в сыворотке и моче лихорадящих пациентов, пациентов с почечной недостаточностью и раковых пациентов и даже некоторых здоровых индивидуумов. Опухоли головного мозга и яичников человека продуцировали высокие уровни TNF-BP в сыворотке. Эти молекулы были очищены, охарактеризованы и клонированы (Gatanaga et al., Lymphokine Res. 9:225-229, 1990a; Gatanaga et al., Proc. Natl. Acad. Sci. USA 87:8781-8784, 1990b). TNF-BP человека состоит из белков 30 кДа и 40 кДа, которые идентичны N-концевым внеклеточным доменам TNF-рецепторов р55 и р75 соответственно (Патент США 5395760; ЕР 418-014). Такие белки были предложены для применения в лечении эндотоксинового бактериально-токсического шока. Mohler et al., J. Immunol. 151:1548-1561, 1993.
Имеется несколько механизмов, возможных для продуцирования секретируемых белков, сходных с мембраносвязанными рецепторами. Один из них включает в себя трансляцию из альтернативно сплайсированных мРНК, не имеющих трансмембранного и цитоплазматического районов. Другой включает в себя протеолитическое расщепление интактных мембранных рецепторов с последующим выделением расщепленного рецептора из клетки. Растворимая форма TNF-R р55 и р75, по-видимому, не образуется из сплайсинга мРНК, так как in vitro в клетках человека может быть детектирована только полноразмерная мРНК рецептора (Gatanaga et al., 1991). Карбокси-концевое секвенирование и исследования мутаций на TNF-R р55 человека показывает, что между остатками Asn 172 и Val 173 может существовать сайт расщепления (Gullberg et al., Eur. J. Cell. Biol. 58:307-312, 1992).
Имеются сообщения, что специфический ингибитор металлопротеазы, ингибитор протеазы TNF-α (TAPI), блокирует выделение растворимых TNF-R р75 и р55 (Crowe et а1., J. Exp. Med. 181:1205-1210, 1995; Mullberg et al., J. Immunol. 155:5198-5205, 1995). Процессинг про-TNF на клеточной мембране с высвобождением лиганда TNF, по-видимому, зависит от фермента, подобного металлопротеазе матрикса (Gearing et al., Nature 370:555-557, 1994). Именно семейство структурно родственных деградирующих матрикс ферментов играет главную роль в ремоделировании и репарации ткани, связанных с развитием и воспалением (Birkedal-Hansen et al., Crit. Rev. Oral Biol. Med. 4:197-250, 1993). Эти ферменты имеют Zn2+ в их каталитических доменах и Са2+ стабилизирует значительно их третичную структуру.
В Европейской патентной заявке ЕР 657536А1 Wallach et al. предполагают, что можно было бы получить фермент, который расщепляет рецептор TNF 55000 кДа, путем нахождения мутированной формы этого рецептора, которая не расщепляется этим ферментом, но все еще связывается с ним. Единственным предлагаемым источником для этого фермента является детергентный экстракт мембран клеток, который, по-видимому, имеет протеазную активность. Если бы было возможно получить фермент согласно этой схеме, то этот фермент предположительно содержал бы простирающийся через мембрану район. Эта патентная заявка не описывает какую-либо протеазу, которая была действительно получена.
В более ранней патентной заявке в данной серии (Международной патентной публикации WO 98/20140) описаны способы получения выделенного фермента, который отщепляет как TNF-R р55, так и TNF-R р75 от клеточных поверхностей. Подходящим источником является культуральная среда клеток, которые были стимулированы форболмиристатацетатом (ФМА). Эта ферментативная активность была названа TRRE (фермент высвобождения рецептора TNF). В других исследованиях TRRE высвобождался непосредственно при ФМА-стимуляции, что указывает на то, что он предварительно синтезируется в неактивной форме, чтобы быстро превращаться в активную форму при стимуляции. Доказательством прямого отщепления TNF-R является то, что выделение его из клетки начинается очень быстро (˜5 минут) с максимальным выделением в пределах 30 минут. TRRE является специфическим для TNF-R и не расщепляет рецепторы IL-1, CD30, ICAM-1 или CD11b. Активность TRRE усиливается добавлением Са++ или Zn++ и ингибируется ЭДТА и фенантролином.
При условии, что TNF участвует в разнообразных патологических состояниях, желательно было бы получить различные факторы, которые позволили бы модулировать выделение рецептора для регуляции тем самым трансдукции (передачи) сигнала от TNF в сайт заболевания.
Это описание обеспечивает новые соединения, которые стимулируют ферментативное расщепление и высвобождение рецепторов TNF из клеточной поверхности. Девять новых ДНК-клонов были отобраны после повторяемого скрининга в анализе, который тестирует способность усиления высвобождения рецептора. Полинуклеотидные последовательности данного изобретения и белки, кодируемые ими, являются потенциально диагностическими инструментами и терапевтическими соединениями, которые могут быть использованы для коррекции трансдукции сигнала TNF выгодным образом.
Одним вариантом осуществления данного изобретения является выделенный полинуклеотид, содержащий нуклеотидную последовательность со следующими свойствами: а) эта последовательность экспрессируется на уровне мРНК в Т-клетках Jurkat; b) когда клетки COS-1, экспрессирующие TNF-рецептор, генетически трансформируют для экспрессии этой последовательности, эти клетки имеют увеличенную ферментативную активность расщепления и высвобождения этого рецептора. Если полинуклеотидная последовательность экспрессируется в клетках Jurkat, то она может быть обнаружена в экспрессионной библиотеке клеток Jurkat, депонированной АТСС (Номер доступа TIB-152). Известно, что этот полинуклеотид может быть получен из других клеточных линий или получен рекомбинантными способами.
В изобретение включены полинуклеотиды, в которых эта нуклеотидная последовательность содержится в виде любой из последовательностей SEQ ID NO:1-10. Вариантами изобретения являются также полинуклеотиды, содержащие по меньшей мере 30 и предпочтительно больше последовательных нуклеотидов в указанной нуклеотидной последовательности или по меньшей мере 50 последовательных нуклеотидов, которые гомологичны указанной последовательности, при значительном уровне, предпочтительно при уровне 90% или более. В изобретение включены также антисмысловые и рибозимные полинуклеотиды, которые ингибируют экспрессию модулятора TRRE.
Другим вариантом данного изобретения являются выделенные полипептиды, содержащие аминокислотную последовательность, кодируемую полинуклеотидом данного изобретения. Неограничительными примерами являются последовательности, показанные в SEQ ID NO:147-SEQ ID NO:158. В данное изобретение включены фрагменты и слитые (химерные) белки и они предпочтительно содержат по меньшей мере 10 последовательных остатков, кодируемых полинуклеотидом данного изобретения, или по меньшей мере 15 последовательных аминокислот, которые гомологичны при значительном уровне, предпочтительно при уровне по меньшей мере 80%. Предпочтительные полипептиды стимулируют расщепление и высвобождение рецепторов TNF из клеточной поверхности, в частности клеток COS-1, генетически трансформированных для экспрессии рецептора TNF. Эти полипептиды могут иметь простирающийся через мембрану домен или могут не иметь простирающегося через мембрану домена и могут необязательно продуцироваться посредством процесса, который включает в себя секрецию из клетки. В изобретение включены видовые гомологи с желательной последовательностью и искусственные мутанты с дополнительными выгодными свойствами.
Другим вариантом данного изобретения является антитело, специфическое для полипептида данного изобретения. Предпочтительными являются антитела, которые связывают TRRE-модуляторный белок, но не другие вещества, обнаруживаемые в сравнимых количествах в пробах тканей человека.
Другим вариантом данного изобретения является тест-способ определения измененной TRRE-активности в пробе клеток или ткани, в котором используют полинуклеотид или антитело данного изобретения для обнаружения присутствия или отсутствия соответствующего TRRE-модулятора. Этот тест-способ может быть в случае необходимости использован для диагностики или оценки клинического состояния в связи с атипичными уровнями TNF или трансдукции сигнала TNF.
Другим вариантом данного изобретения является способ увеличения или уменьшения трансдукции сигнала от цитокина в клетку (в том числе сигнала TNF, но не только), предусматривающий контактирование этой клетки с полинуклеотидом, полипептидом или антителом данного изобретения.
Следующим вариантом данного изобретения является способ скрининга полинуклеотидов на способность модулировать TRRE-активность. Этот способ включает в себя обеспечение клеток, которые экспрессируют как TRRE, так и TNF-рецептор; генетическое изменение этих клеток подлежащими скринингу полинуклеотидами; клонирование этих клеток и идентификацию клонов с желательной активностью.
Еще одним вариантом данного изобретения является способ скрининга веществ на способность влиять на TRRE-активность. Обычно он предусматривает инкубирование клеток, экспрессирующих рецептор TNF, с TRRE-модулятором данного изобретения в присутствии или в отсутствие испытуемого вещества и измерение действия на выделение этого рецептора TNF.
Продукты данного изобретения могут быть использованы в приготовлении лекарственного средства для лечения организма человека или животного. Это лекарственное средство содержит клинически эффективное количество для лечения заболевания, такого как сердечная недостаточность, кахексия, воспаление, эндотоксиновый бактериально-токсический шок, артрит, множественный склероз, сепсис и рак. Эти композиции могут быть использованы для введения субъекту, у которого предполагают наличие или риск возникновения такого заболевания, необязательно в комбинации с другими формами лечения, пригодными для состояния субъекта.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 является схематическим представлением плазмиды pCDTR2. Эта плазмида экспрессирует TNF-R р75, ˜75 кДа-форму рецептора TNF. PCMV обозначает здесь цитомегаловирус; BGHpA обозначает сигнал полиаденилирования бычьего гормона роста.
Фигура 2 является линией, изображающей уровни TNF-R р75, обнаруженные на клетках COS-1, генетически измененных для экспрессии рецептора. Результаты, полученные для трансформированных клеток, обозначенных C75R ( , устремленная вверх линия), сравниваются с результатами их исходных клеток COS-1 (
, устремленная вверх линия), сравниваются с результатами их исходных клеток COS-1 ( , базовая линия). Число рецепторов рассчитывали при помощи анализа Скетчарда (вставка).
, базовая линия). Число рецепторов рассчитывали при помощи анализа Скетчарда (вставка).
Фигура 3 является графиком выживания, показывающим, что TRRE снижает смертность в мышах, которым вводили липополисахарид (ЛПС) для индукции септического перитонита. (◆) только ЛПС; () ЛПС плюс контрольный буфер; () ЛПС плюс TRRE (2000 Е); ( ) ЛПС плюс TRRE (4000 Е).
) ЛПС плюс TRRE (4000 Е).
Фигура 4 является растровой репродукцией диаграммы в виде столбцов, показывающей действие 9 новых клонов на TRRE-активность на клетках C75R (клетках COS-1, трансфицированных для экспрессии TNF-рецептора). Каждый из этих клонов увеличивает TRRE-активность более чем в 2 раза.
Фигура 5 является графиком выживания, показывающим способность 4 новых клонов сохранять жизнь мышей, которым вводили ЛПС. (◆) солевой раствор; () БСА; () Меу-3 (100 мкг); (Х) Меу-3 (10 мкг); (*) Меу-5 (10 мкг); () Меу-8 (10 мкг).
Было обнаружено, что некоторые клетки, участвующие в пути трансдукции TNF, экспрессируют ферментативную активность, которая заставляет рецепторы TNF отделяться от клеточной поверхности. Ферментативная активность отщепления и высвобождения рецепторов TNF была обозначена как TRRE. Форболмиристатацетат индуцирует высвобождение TRRE из клеток в культуральную среду. TRRE-белок был очищен в качестве примера из супернатанта клеток TNF-1 (пример 2). Эта протеаза несет некоторые отличительные признаки семейства металлопротеаз и быстро высвобождается из клеток при активации.
Для выяснения природы этого белка проводили функциональное клонирование. Клетки Jurkat были выбраны как хороший источник TRRE. кДНК из библиотеки Jurkat экспрессировали и клеточный супернатант тестировали на способность высвобождать рецепторы TNF из клеточных поверхностей. Клонирование и тестирование продукта экспрессии проводили посредством нескольких циклов и были получены девять клонов, которые более чем удваивали TRRE-активность в этом тесте (фигура 4). На уровне ДНК все 9 клонов имели различные последовательности.
Продукты экспрессии белка из этих клонов тестировали в липополисахаридной модели животного для сепсиса. Белок из трех различных клонов успешно предохранял животных от летальной дозы ЛПС (фигура 5). Это указывает на важную роль этих молекул в способности справляться с патологическими состояниями, опосредованными TNF.
Количество новых стимулирующих TRRE клонов, полученных из экспрессионной библиотеки, было удивительным. Субстратная специфичность TRRE, выделенного в примере 2, отличает рецепторы TNF 75 кДа и 55 кДа от рецепторов других цитокинов и белков поверхности клеток. Было мало оснований для предположения заранее, что клетки могут иметь девять различных протеаз для рецептора TNF. Возможно, что один из этих клонов кодирует TRRE, выделенный в примере 2, или родственный белок. Возможно, что некоторые из других клонов имеют протеолитическую активность для расщепления рецепторов TNF в том же самом сайте или в другом сайте, что вызывает высвобождение растворимой формы из клетки. Гипотезой данного изобретения является то, что некоторые из этих клонов могут сами не иметь протеолитической активности, но играют роль в стимулировании TRRE-активности вторичным образом.
Эта возможность согласуется со сделанными наблюдениями, поскольку существует эндогенный уровень TRRE-активности в клетках, использованных в этом тесте. Тест расщепления включает в себя мониторинг высвобождения рецептора TNF из клеток С75, которые являются клетками COS-1, генетически измененными для экспрессии TNF-R р75. Стандартный тест проводят путем контактирования этих трансформированных клеток с жидкостью, которая, как считают, содержит TRRE. Уровень эндогенной активности TRRE выявляется из скорости самопроизвольного высвобождения рецептора, даже в отсутствие добавления экзогенного TRRE (приблизительно 200 единиц). Таким образом, дополнительные белки, которые стимулируют TRRE-активность, увеличивали бы активность, измеряемую в этом тесте. Возможны многочисленные механизмы стимуляции, включающие в себя белки, которые активируют зимогенную форму TRRE, белки, которые освобождают TRRE от других компонентов клеточной поверхности, или белки, которые стимулируют секрецию TRRE из внутреннего компартмента клетки. Не требуется обязательное понимание этого механизма для применения продуктов данного изобретения в большинстве описанных вариантов.
Ожидается, что некоторые из этих клонов будут иметь активность не только в отношении стимулирования расщепления рецептора TNF, но будут также оказывать действие на другие белки клеточной поверхности. Вплоть до того, что последовательности расщепления или вспомогательные белки являются общими между различными рецепторами, некоторые клоны могли бы стимулировать фенотипические изменения (такие как высвобождение рецептора) для семейства родственных субстратов.
Данное изобретение обеспечивает полипептиды, которые стимулируют TRRE-активность, полинуклеотиды, которые кодируют такие полипептиды, и антитела, которые связывают такие пептиды. Связывание TNF с его рецептором опосредует ряд биологических эффектов. Расщепление TNF-рецептора посредством TRRE уменьшает трансдукцию сигнала посредством TRRE. Потенциаторы TRRE-активности обладают таким же действием. Таким образом, продукты данного изобретения могут быть использованы для модуляции трансдукции сигналов цитокинами, что имеет важное значение в способности справляться с патологическими состояниями, на которые действуют цитокины. Продукты данного изобретения могут быть также использованы в диагностических способах для определения, когда атипичная TRRE-активность действует на трансдукцию сигнала неподходящим образом. Тест-системы, описанные в этом описании, обеспечивают способ для скрининга дополнительных соединений, которые могут влиять на TRRE-активность и, следовательно, на трандукцию сигналов от TNF.
На основании краткого изложения данного изобретения и руководствуясь иллюстрациями в разделе примеров, специалист с квалификацией в данной области легко поймет, какие способы следует использовать в применении на практике данного изобретения. Нижеследующее подробное описание дается в интересах читателя для лучшего понимания изобретения.
Определения и основные способы
В применении в данном описании, "TRRE-активность" обозначает способность композиции расщеплять и высвобождать рецепторы TNF из поверхности экспрессирующих их клеток. Предпочтительным тестом является расщепление из трансфицированных клеток COS-1, описанное в примере 1. Однако TRRE-активность может быть измерена на любых клетках, которые несут рецепторы TNF с размером 55 кДа или 75 кДа. Другие свойства фермента TRRE, полученного ФМА-индукцией клеток ТНР-1 (приведенные в примере 2), не должны быть обязательно свойством TRRE-активности, измеряемой в этом тесте.
Унифицированная активность TRRE определяется как 1 рг растворимого TNF-R р75, высвобождаемого из клеточной поверхности в стандартном тесте, после коррекции на самопроизвольное высвобождение. Измерение TRRE-активности объясняется дополнительно в примере 1.
"TRRE-модулятор" представляет собой соединение, которое обладает способностью либо увеличения, либо уменьшения TRRE-активности для процессинга TNF на поверхности клеток. Те соединения, которые увеличивают TRRE-активность, могут быть названы TRRE-стимуляторами, а те, которые уменьшают TRRE-активность, могут быть названы TRRE-ингибиторами. TRRE-стимуляторы включают в себя соединения, которые обладают протеолитической активностью в отношении TNF-R, и соединения, которые увеличивают активность протеаз TNF-R. Примерами TRRE-стимуляторов являются девять полинуклеотидных клонов, описанных в примере 5, и их белковые продукты. Ингибиторы TRRE-активности могут быть получены с использованием тестов скрининга, описанных ниже.
Термин "полинуклеотид" относится к полимерной форме нуклеотидов любой длины, либо дезоксирибонуклеотидов, либо рибонуклеотидов, или их аналогов. Полинуклеотиды могут иметь трехмерную структуру и могут выполнять любую функцию, известную или неизвестную. Далее приведены неограничительные примеры полинуклеотидов: ген или фрагмент гена, экзоны, интроны, (мРНК), рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, зонды нуклеиновых кислот и праймеры. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и аналоги нуклеотидов. В случае их присутствия, модификации нуклеотидной структуры могут быть произведены до или после сборки полимера. Термин полинуклеотид обозначает взаимозаменяемо двух- и одноцепочечные молекулы. Если нет иных указаний или требований, любой вариант данного изобретения, описанный здесь, который является полинуклеотидом, включает как двухцепочечную форму, так и каждую из двух комплементарных одноцепочечных форм, известных или предполагаемых для образования данной двухцепочечной формы.
"Гибридизация" обозначает реакцию, в которой один или несколько полинуклеотидов взаимодействуют с образованием комплекса, который стабилизируется через образование водородных связей между основаниями нуклеотидных остатков. Реакции гибридизации могут проводиться в условиях различной "строгости". Рассматриваемые при этом условия включают в себя температуру, ионную силу и присутствие дополнительных растворенных веществ в реакционной смеси, таких как формамид. Условиями увеличивающейся строгости являются: 30°С, в 10Х SSC (0,15 М NaCl, 15 мМ цитратный буфер); 40°С, в 6Х SSC или при приблизительно 40°С, в 0,5Х SSC или при приблизительно 30°С, в 6Х SSC, причем SSC содержит 50% формамид. ДСН и источник фрагментированной ДНК (такой как сперма лосося) обычно также присутствуют во время гибридизации. Более высокие условия строгости требуют более высокого минимума комплементарности между гибридизующимися элементами для образования стабильного гибридизационного комплекса. См. "Molecular Cloning: A Laboratory Manual", Second Edition (Sambrook, Fritsch & Maniatis, 1989).
Понятно, что пуриновые и пиримидиновые азотистые основания со сходными структурами могут быть функционально равноценными в смысле спаривания оснований по Уотсону-Крику; и взаимозаменяемость сходных азотистых оснований, в частности урацила и тимина, или модификация азотистых оснований, такая как метилирование, не является существенной заменой.
Процент идентичности последовательности для полинуклеотиов или полипептидов рассчитывают сопоставлением сравниваемых последовательностей и затем подсчетом числа общих остатков в каждом сопоставляемом положении. Никакого "пенальти" не налагается из-за присутствия вставок или делеций, но они допускаются только в том случае, когда требуется согласовать явно увеличенное число аминокислотных остатков в одной из сопоставляемых последовательностей. В том случае, когда одна из сравниваемых последовательностей указана как являющаяся "последовательной", "гэпы" (пробелы) в этой последовательности не допускаются во время сравнения. Процентная идентичность выражается как количество (в %) остатков в тестируемой последовательности, которые являются идентичными остаткам в сравнительной или ссылочной последовательности.
В применении здесь, "экспрессия" полинуклеотида обозначает образование РНК-транскрипта. Последующая трансляция в белок или другие эффекторные соединения может также иметь место, но не требуются, если нет иных указаний.
"Генетическое изменение" обозначает процесс, в котором генетический элемент вводят в клетку другим способом, чем посредством митоза или мейоза. Этот элемент может быть гетерологичным для данной клетки или он может быть дополнительной копией или улучшенной версией элемента, уже присутствующего в данной клетке. Генетическое изменение может быть выполнено, например, трансдукцией клетки рекомбинантной плазмидой или другим полинуклеотидом с использованием любого способа, известного в данной области, такого как электропорация, осаждение фосфатом кальция или контактирование с комплексом полинуклеотид-липосома. Генетическое изменение может быть также выполнено, например, трансдукцией или инфекцией ДНК- или РНК-вирусом или вирусным вектором. Предпочтительно, чтобы генетическое изменение наследовалось потомством данной клетки, но это не является общим требованием, если нет иных указаний.
Термины "полипептид", "пептид" и "протеин" используются здесь взаимозаменяемо для обозначения полимеров аминокислот любой длины. Этот полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты и может прерываться неаминокислотами. Эти термины охватывают также аминокислотный полимер, который был модифицирован; например, образованием дисульфидной связи, гликозилированием, липидированием, ацетилированием, фосфорилированием или любой другой манипуляцией, такой как конъюгация с меченым компонентом.
"Слитый полипептид" представляет собой полипептид, содержащий районы в отличающемся положении в последовательности, в сравнении с тем, что имеет место в природе. Эти районы могут нормально существовать в отдельных белках и быть соединены вместе в слитом полипептиде; они могут нормально существовать в том же самом белке, но быть помещены в новом расположении в слитом полипептиде; или они могут быть синтетически перегруппированы. "Функционально эквивалентный фрагмент" полипептида отличается от нативной последовательности добавлением, делецией или заменой аминокислотных остатков или любой их комбинацией при сохранении функционального свойства этого фрагмента, относящегося к тому контексту, в котором он используется. Слитые белки и функционально эквивалентные фрагменты включены в определение полипептидов, используемое в этом описании.
Понятно, что укладка и биологическая функция белков может использовать аккомодацию инсерций, делеций и замен в аминокислотной последовательности. Некоторые аминокислотные замены являются более легко переносимыми. Например, замена аминокислоты с гидрофобными боковыми цепями, ароматическими боковыми цепями, полярными боковыми цепями, боковыми цепями с положительным или отрицательным зарядом или боковыми цепями, содержащими два или менее атомов углерода, другой аминокислотой с боковой цепью со сходными свойствами может происходить без нарушения по существу идентичности этих двух последовательностей. Способы определения гомологичных районов и оценки в баллах степени гомологии описаны в Altschul et al. Bull. Math. Bio. 48:603-616, 1986 и Heniroff et al. Proc. Natl. Acad. Sci. USA 89:10915-10919, 1992. Замены, которые сохраняют функциональность полипептида или придают новое и выгодное свойство (такое как повышенная активность, стабильность или пониженная иммуногенность), являются особенно предпочтительными.
Термин "антитело" (взаимозаменяемо используемый во множественной форме) обозначает молекулу иммуноглобулина, способную к специфическому связыванию с мишенью, такой как полипептид, посредством по меньшей мере одного сайта узнавания антигена, локализованного в вариабельной области молекулы иммуноглобулина. В применении здесь этот термин включает в себя не только интактные антитела, но также эквиваленты антител, которые включают в себя по меньшей мере один антигенсвязывающий сайт желательной специфичности. Они включают в себя, но не ограничиваются ими, ферментативно или рекомбинантно полученные фрагменты антител, слитые белки, гуманизированные антитела, одноцепочечные вариабельные области, двойные антитела и цепи антител, которые подвергаются антигениндуцируемой сборке.
Термин "выделенный" ("выделенное") полипептид, полинуклеотид, белок, антитело или другое вещество относится к получению этого вещества, освобожденного по меньшей мере от некоторых других компонентов, которые могут также присутствовать там, где природно встречается это вещество или подобное вещество или откуда оно первоначально получено. Так, например, выделенное вещество может быть получено с использованием способа очистки для обогащения его из исходной смеси. Обогащение может быть измерено на абсолютной основе, такой как вес на объем раствора, или оно может быть измерено относительно второго, потенциально примесного вещества, присутствующего в исходной смеси. Чем больше обогащение вариантов данного изобретения, тем более предпочтительными они являются. Так, например, 2-кратное обогащение является предпочтительным, 10-кратное обогащение является более предпочтительным, 100-кратное обогащение является еще более предпочтительным, 1000-кратное обогащение является даже еще более предпочтительным. Вещество может быть обеспечено также в выделенном состоянии при помощи способа искусственной сборки, такого как химический синтез или рекомбинантная экспрессия.
Термин "клетка-хозяин" обозначает клетку, которая была генетически изменена или которая способна к трансформации, путем введения экзогенного полинуклеотида.
Термин "клиническая проба" включает в себя множество типов проб, полученных из субъекта и применимых в процедуре in vitro, такой как диагностический тест. Это определение включает в себя образцы твердых тканей, полученные хирургическим удалением, образец патологии или образец биопсии, клетки, полученные из клинического субъекта, или их потомство, полученное из культуры, жидкие пробы, такие как кровь, сыворотка, плазма, цереброспинальная жидкость и моча, и любые фракции или экстракты таких проб, которые содержат потенциальное указание на это заболевание.
Если нет иных указаний, практика данного изобретения будет использовать общепринятые способы молекулярной биологии, микробиологии, рекомбинантных ДНК и иммунологии, в пределах квалификации в данной области. Такие способы объясняются в стандартной литературе, такой как "Molecular Cloning: A Laboratory Manual", Second Edition (Sambrook, Fritsch & Maniatis, 1989), "Oligonucleotide Synthesis" (M.J. Gait, ed., 1984), "Animal Cell Culture" (R.I.Freshney, ed., 1987); ряд "Methods in Enzymology" (Academic Press, Inc.); "Handbook of Experimental Immunology" (D.M.Weir & C.C.Blackwell, Eds.), "Gene Transfer Vectors for Mammalian Cells" (J.M.Miller & M.P.Calos, eds., 1987), и "Current Protocols in Molecular Biology (F.M.Ausubel et al., eds., 1987 и "Current Protocols in Immunology" (J.E.Coligan et al., eds., 1991). Читатель может также выбрать ссылку на более раннюю патентную заявку, относящуюся к TRRE, Международную заявку WO 98/020140.
Для целей ведения дела в США и в других юрисдикциях, где это позволено, все патенты, патентные заявки, статьи и публикации, указанные в любом месте в этом описании, включены тем самым в качестве ссылки во всей их полноте.
Полинуклеотиды
Полинуклеотиды данного изобретения могут быть получены любым подходящим способом в данной области. С использованием данных, обеспеченных в данном описании, последовательности менее ˜50 п.н. легко получают химическим синтезом, либо через коммерческую службу, либо при помощи известного способа синтеза, такого как триэфирный способ или фосфитный способ. Предпочтительным способом является тверодофазный синтез с использованием мононуклеозидфосфорамидитных связывающих единиц (Hirose et al., Tetra, Lett. 19:2449-2452, 1978; US Patent No. 4415732).
Для использования в антисмысловой терапии полинуклеотиды могут быть получены при помощи химического способа, который дает более стабильные фармацевтические препараты. Неограничительные примеры включают в себя тиол-дериватизованные нуклеозиды (патент США 5578718) и олигонуклеотиды с модифицированными скелетами (патенты США с номерами 5541307 и 5378825).
Полинуклеотиды данного изобретения могут быть также получены ПЦР-амплификацией матрицы с желательной последовательностью. Олигонуклеотидные праймеры, охватывающие желательную последовательность, отжигают с матрицей, удлиняют их ДНК-полимеразой и затем расплавляют при более высокой температуре таким образом, чтобы матрица и удлиненные олигонуклеотиды разъединились. Этот цикл повторяют до тех пор, пока не будет получено желательное количество амплифицируемого полинуклеотида (патенты США с номерами 4683195 и 4683202). Подходящие матрицы включают в себя библиотеку Т-клеток Jurkat и другие экспрессионные библиотеки человека и животных, которые содержат последовательности, кодирующие TRRE-модулятор. Библиотека Т-клеток Jurkat доступна из Американской коллекции типовых культур, 10801 University BLVD., Manassas VA 20110, U.S.A. (ATCC # TIB-152). Мутации и другие адаптации могут быть выполнены во время амплификации конструированием подходящих праймеров или могут быть включены после генетического сплайсинга.
Количества промышленного масштаба больших полинуклеотидов наиболее удобно получать встраиванием желательной последовательности в подходящий клонирующий вектор и репродуцированием клона. Способы клонирования нуклеотидов даны в Sambrook, Fritsch & Maniatis (supra) и в патенте США с номером 5552524. Примеры клонирования и способы экспрессии иллюстрированы в примере 6.
Предпочтительные полинуклеотидные последовательности являются на 50%, 70%, 80%, 90% или 100% идентичными одной из последовательностей, приведенных в качестве примеров в этом описании; в порядке увеличения предпочтительности. Длина последовательных остатков в идентичной или гомологичной последовательности, сравнимой с примерной последовательностью, может быть приблизительно 15, 30, 50, 75, 100, 200 или 500 остатков в порядке увеличения предпочтительности вплоть до длины целого клона. Нуклеотидные замены, которые обусловливают консервативную замену или сохраняют функцию кодируемого полипептида (в смысле гибридизационных свойств или кодируемого продукта) являются особенно предпочтительными заменами.
Полинуклеотиды данного изобретения могут быть использованы для измерения измененной активности TRRE в пробе клеток или ткани. Это предусматривает контактирование пробы с полинуклеотидом в условиях, позволяющих полинуклеотиду гибридизоваться специфически с нуклеиновой кислотой, которая кодирует модулятор TRRE-активности, если она присутствует в этой пробе, и определение полинуклеотида, который гибридизовался, как результата стадии а). Специфичность этого теста может быть обеспечена несколькими путями. Один способ предусматривает использование специфического зонда - полинуклеотида данного изобретения с последовательностью, достаточно длинной и достаточно идентичной детектируемой последовательности, так что он связывает эту мишень, но не другую нуклеиновую кислоту, которая может присутствовать в этой пробе. Этот зонд обычно является меченым (либо непосредственно, либо посредством вторичного реагента), так что он может быть затем детектирован. Подходящие метки включают в себя 32P и 33Р, хемилюминесцентные и флуоресцентные реагенты. После реакции гибридизации непрореагировавший зонд отмывают, так чтобы можно было определить количество гибридизованного зонда. Сигнал может быть усилен с использованием разветвленных зондов (патент США с номером 5124246). В другом способе полинуклеотид является праймером для реакции ПЦР. Специфичность обеспечивается способностью спаренных зондов амплифицировать представляющую интерес последовательность. После подходящего числа ПЦР-циклов количество присутствующего амплифицированного продукта коррелирует с количеством последовательности-мишени, исходно присутствующей в этой пробе.
Такие тесты применимы как в исследованиях, так и в диагностике или оценке патологического состояния. Например, TNF-активность играет роль в элиминации опухолевых клеток (пример 4) и рак может избежать этого процесса элиминации активацией TRRE-активности в затронутой заболеванием ткани. Таким образом, при некоторых условиях высокая экспрессия модуляторов TRRE может коррелировать с прогрессированием рака. Диагностические тесты применимы также в мониторинге терапии, например, когда генную терапию применяют для увеличения TRRE-активности.
Полинуклеотиды данного изобретения могут быть также использованы для получения полипептидов и приготовления лекарственных средств, как поясняется ниже.
Полипептиды
Короткие полипептиды данного изобретения могут быть получены твердофазным химическим синтезом. Принципы твердофазного химического синтеза можно найти в Duglas & Penney, Bioorganic Chemistry, Springer-Verlag NY pp 54-92 (1981) и в патенте США №4493795. Автоматизированный твердофазный пептидный синтез может проводиться с использованием устройства, такого как пептидный синтезатор PE-Applied Biosystems, Foster City CA).
Более длинные полипептиды удобно получать экспрессионным клонированием. Полинуклеотид, кодирующий желаемый полипептид, оперативно (функционально) связывают с регуляторными элементами для транскрипции и трансляции и затем трансфицируют в подходящую клетку-хозяин. Экспрессия может выполняться в прокариотах, таких как Е. coli (ATCC Accession (Номер доступа АТСС) No 31446 или 27325), эукариотных микроорганизмах, таких как дрожжи Saccharomyces cerevisiae, или высших эукариотах, таких как клетки насекомых или млекопитающих. Ряд экспрессионных систем описан в патенте США №5552524. Экспрессионное клонирование доступно в таких коммерческих службах, как Lark Technologies, Houston TX. Получение белка из 4 примеров клонов данного изобретения в клетках насекомых иллюстрируется в примере 6. Белок очищают из продуцирующей клетки-хозяина стандартными способами химии белков, такими как аффинная хроматография и ВЖХ. Продукты экспрессии в случае необходимости для облегчения аффинной очистки получают с последовательностью-меткой (тэгом), которая может быть затем удалена.
Предпочтительные последовательности являются на 40%, 60%, 80%, 90% или 100% идентичными одной из последовательностей, приведенных в качестве примеров в этом описании; в порядке увеличения предпочтительности. Длина этой идентичной или гомологичной последовательности, сравниваемой с нативным полинуклеотидом человека, может быть приблизительно 7, 10, 15, 20, 30, 50 или 100 остатков в порядке увеличения предпочтительности, вплоть до длины всего кодирующего района.
Полипептиды могут быть тестированы на способность модулировать TRRE в тесте расщепления TNF-R. Полипептид контактируют с рецептором (предпочтительно экспрессируемым на поверхности клетки, такой как клетка С75), и определяют способность этого полипептида увеличивать или уменьшать расщепление и высвобождение рецептора. Расщепление TNF-R приведенными в качестве примеров полипептидами данного изобретения иллюстрируется в примере 7.
Полипептиды данного изобретения могут быть использованы в качестве иммуногенов для выработки антител. Большие белки будут индуцировать образование смеси антител, тогда как короткие пептидные фрагменты будут индуцировать образование антител против небольшого района интактного белка. Клоны антител могут быть картированы в отношении сайта связывания белка путем получения коротких перекрывающихся пептидов длиной приблизительно 10 аминокислот. Перекрывающиеся пептиды могут быть получены на найлоновой мембране-подложке стандартным химическим F-Moc-способом с использованием набора SPOTS™ из Genosis в соответствии с инструкциями изготовителя.
Полипептиды данного изобретения могут быть также использованы для действия на трансдукцию сигнала TNF, как объясняется ниже.
Антитела
Поликлональные антитела могут быть получены инъецированием позвоночного животного полипептидом данного изобретения в иммуногенной форме. Иммуногенность полипептида может быть усилена связыванием с носителем, таким как KLH, или объединением с адъювантом, таким как адъювант Фрейнда. Обычно примирующая (праймирующая) инъекция сопровождается бустер-инъекцией спустя приблизительно 4 недели и антисыворотку собирают спустя одну неделю. Нежелательная активность, перекрестно-реагирующая с другими антигенами, если она присутствует, может быть удалена, например, пропусканием препарата через адсорбенты, приготовленные из этих антигенов, прикрепленных к твердой фазе, и сбором несвязавшейся фракции. Если желательно, активность специфических антител может быть дополнительно очищена комбинацией способов, которые могут включать в себя протеин А-хроматографию, осаждение сульфатом аммония, ионообменную хроматографию, ВЖХ и иммуноаффинную хроматографию с использованием иммунизирующего полипептида, связанного с твердым носителем. Фрагменты антител и другие производные могут быть получены стандартными иммунохимическими способами, такими как подвергание антитела расщеплению ферментами, такими как папаин или пепсин.
Получение моноклональных антител описано в таких стандартных ссылках, как Harrow & Lane (1988), патенты США с номерами 4491632, 4472500 и 4444887 и Methods in Enzymology 73B:3 (1981). Вкратце, млекопитающее иммунизируют и собирают антитела-продуцирующие клетки (обычно спленоциты). Клетки иммортализуют слиянием с непродуцирующей миеломой, трансфекцией вирусом Эпстейна-Барр или трансформацией онкогенной ДНК. Эти обработанные клетки клонируют и культивируют и отбирают клоны, которые продуцируют антитело желательной специфичности.
Другие способы получения молекул специфических антител (оптимально в форме одноцепочечных вариабельных областей) включают в себя контактирование библиотеки иммунокомпетентных клеток или вирусных частиц с целевым антигеном и выращивание положительно отобранных клонов. Иммунокомпетентный фаг может быть сконструирован для экспрессии сегментов вариабельных областей иммуноглобулина на поверхности фага. См. Marks et al., New Eng. J. Med. 335:730, 1996, Международные патентные заявки WO 9413804, WO 9201047, WO 9002809 и McGuiness et al., Nature Biotechnol. 14:1449, 1996.
Антитела данного изобретения могут быть использованы в иммуноанализах на модуляторы TRRE. Общие способы иммуноанализа могут быть найдены в "The Immunoassay Handbook", Stockton Press NY, 1994 и "Methods of Immunological Analysis", Weinheim: VCH Verlags gesellschaft mbH, 1993). Антитело объединяют с тест-пробой в условиях, в которых это антитело будет связываться специфически с любым модулятором, который может присутствовать, но не с какими-либо другими белками, возможно, присутствующими в пробе. Образовавшийся комплекс может быть измерен in situ (патенты США с номерами 4208479 и 4708929) или физическим отделением его от непрореагировавших реагентов (патент США 3646346). Тесты отделения обычно включают в себя меченый TRRE-реагент (конкурентный анализ) или меченое антитело (сэндвич-анализ) для облегчения детектирования и количественного определения этого комплекса. Подходящими метками являются радиоактивные изотопы, такие как 126I, ферменты, такие как β-галактозидаза, и флуоресцентные метки, такие как флуоресцеин. Антитела данного изобретения могут быть также использованы для обнаружения TRRE-модуляторов в фиксированных срезах посредством иммуногистологии. Антитело контактируют с тканью, непрореагировавшее антитело отмывают и затем детектируют связанное антитело - обычно с использованием меченого антииммуноглобулинового реагента. Иммуногистология покажет не только, присутствует ли модулятор, но и где он локализован в этой ткани.
Обнаружение TRRE-модуляторов представляет интерес для исследовательских целей и для клинического применения. Как указано ранее, высокая экспрессия TRRE-модуляторов может коррелировать с прогрессированием рака. Диагностические тесты применимы также в мониторинге TRRE-модуляторов, которые вводят в ходе терапии.
Антитела данного изобретения могут быть также использованы для приготовления лекарственных средств. Антитела с терапевтическим потенциалом включают в себя антитела, которые действуют на активность TRRE - либо стимуляцией клиренса TRRE-модулятора, либо блокированием его физиологического действия. Антитела могут быть подвергнуты скринингу на целевую активность в соответствии с тестами, описанными в следующем разделе.
Скрининг-тесты
Данное изобретение обеспечивает ряд способов скрининга для отбора и разработки получения продуктов, которые модулируют TRRE и, следовательно, влияют на трансдукцию сигнала TNF.
Один способ скрининга является способом скрининга на полинуклеотиды, которые способны модулировать TRRE-активность. Для проведения этого скрининга получают клетки, которые экспрессируют как TRRE, так и рецептор TNF. Подходящие клеточные линии могут быть сконструированы из любой клетки, которая экспрессирует уровень функциональной TRRE-активности. Эти клетки можно идентифицировать тестированием культурального супернатанта на способность высвобождать мембраносвязанный TNF-R. Уровень экспрессии TRRE должен быть умеренным, так чтобы можно было детектировать увеличение активности. Затем клетки могут быть генетически изменены для экспрессии либо TNF-R р55, либо TNF-R р75, что иллюстрируется в примере 1. Примером является линия C75R: клетки COS-1, генетически измененные для экспрессии формы 75 кДа TNF-R. Высвобождение TNF-R из этой клетки измеряют либо тестированием остаточного связывания меченого TNF-лиганда с этой клеткой, либо при помощи иммуноанализа супернатанта на высвобожденный рецептор (пример 1).
Этот тест-скрининг проводят контактированием клеток, экспрессирующих TRRE и TNF-R, с полинуклеотидами, которые должны быть подвергнуты скринингу. Определяют действие полинуклеотида на ферментативное высвобождение TNF-R из клетки и отбирают полинуклеотиды с желательной активностью (либо стимулирующей, либо ингибирующей TRRE активностью). В варианте этого способа клетки, экспрессирующие TRRE-активность, но не TNF-R (такие как нетрансфицированные клетки COS-1), контактируют с тестируемым полинуклеотидом. Затем собирают культуральную среду и используют для определения TRRE-активности с использованием второй клетки, экспрессирующей TNF-R (такой как клетка С75).
Этот тип теста-скрининга применим для отбора полинуклеотидов из экспрессионной библиотеки, в которой предполагают наличие кодирующих последовательностей для модуляторов TRRE. Экспрессионная библиотека клеток Jurkat (АТСС Accession No. TIB-152) является примером такой библиотеки. Другими клетками, из которых могут быть сконструированы подходящие библиотеки, являются клетки, о которых известно, что они экспрессируют высокие уровни TRRE, в частности, после ФМА-стимуляции, такие как клетки ТНР-1, U-937, HL-60, ME-180, MRC-5, Raji, К-562 и моноциты здорового человека. Скрининг включает в себя экспрессию ДНК из этой библиотеки в выбранной клеточной линии, используемой для скрининга. Отбирают лунки с желательной активностью и ДНК выделяют, необязательно после репликации или клонирования этих клеток. Повторяющиеся циклы функционального скрининга и отбора могут привести к идентификации новых полинуклеотидых клонов, которые стимулируют или ингибируют TRRE-активность. Это иллюстрируется ниже в примере 5. Могут быть проведены дополнительные эксперименты на отобранных полинуклеотидах для определения, модулирует ли полинуклеотид TRRE-активность внутри клетки или через действие его белкового продукта. Длинная открытая рамка считывания предполагает роль белкового продукта, и исследование аминокислотной последовательности в отношении сигнального пептида и пересекающего мембрану района может помочь определить, секретируется этот белок из клетки или экспрессируется в поверхностной мембране.
Этот тип скрининга применим также для дополнительного развития получения полинуклеотидов данного изобретения. Например, могут быть получены экспрессионные конструкции, которые кодируют функциональные пептидные фрагменты, слитые белки и другие варианты. Минимальный размер полинуклеотидной последовательности, которая все еще кодирует TRRE-модулирующую активность, может быть определен путем удаления части этой последовательности и затем использования этого теста-скрининга для определения, присутствует ли еще эта активность. Мутированные и удлиненные последовательности могут быть тестированы таким же образом.
Этот тип теста-скрининга применим также для разработки соединений, которые влияют на TRRE-активность путем блокирования мРНК, кодирующей модулятор TRRE. Особый интерес представляют рибозимы и антисмысловые олигонуклеотиды. Рибозимы представляют собой эндорибонуклеазы, которые катализируют расщепление РНК в специфическом сайте. Они содержат полинуклеотидную последовательность, которая комплементарна сайту расщепления на мишени, и дополнительную последовательность, которая обеспечивает третичную структуру для осуществления расщепления. Конструкция рибозимов описана в патентах США с номерами 4987071 и 5591610. Антисмысловые олигонуклеотиды, которые связывают мРНК, содержат короткую последовательность, комплементарную этой мРНК (обычно длиной 8-25 оснований). Предпочтительный химический способ для конструирования антисмысловых олигонуклеотидов описан в одном из разделов выше. Специфичность обеспечивается как комплементарной последовательностью, так и свойствами химической структуры. Антисмысловые молекулы, которые ингибируют экспрессию рецепторов поверхности клетки, описаны в патентах США с номерами 5135917 и 5789573. Скрининг включает в себя контактирование клетки, экспрессирующей TRRE-активность и TNF-R, с соединением и определение действия на высвобождение рецептора. Рибозимы и антисмысловые молекулы, эффективные в изменении экспрессии стимулятора TRRE, уменьшали бы высвобождение TNF-R. Рибозимы и антисмысловые молекулы, эффективные в изменении экспрессии ингибитора TRRE, увеличивали бы высвобождение TNF-R.
Другой способ скрининга, описанный в данном описании, предназначен для тестирования способности полипептидов модулировать TRRE-активность (пример 7). Клетки, экспрессирующие как TNF-R, так и умеренный уровень TRRE-активности, контактируют с тестируемыми полипептидами и сравнивают скорость высвобождения рецептора со скоростью самопроизвольного высвобождения. Увеличенная скорость высвобождения показывает, что этот полипептид является стимулятором TRRE, тогда как уменьшенная скорость высвобождения показывает, что этот полипептид является ингибитором TRRE. Этот анализ может быть использован для тестирования активности новых полипептидов и разработки вариантов полипептидов, уже известных в качестве модуляторов TRRE. Минимальный размер полипептидной последовательности, которая все еще кодирует модулирующую TRRE активность, может быть определен приготовлением меньшего фрагмента полипептида и затем использования этого анализа-скрининга для определения, присутствует ли еще эта активность. Мутированные и удлиненные последовательности могут тестироваться таким же образом.
Другой способ скрининга данного изобретения является способом для скрининга веществ, которые препятствуют действию модулятора TRRE на уровне белка. Этот способ включает в себя инкубирование клеток, экспрессирующих рецептор TNF (таких как клетки C75R), с полипептидом данного изобретения, имеющим стимулирующую TNF активность. Имеется две возможности для подачи модулятора TRRE в этом тесте. В одном случае полипептид добавляют к среде этих клеток в качестве реагента вместе с тестируемым веществом. В другом случае клетки генетически изменяют для экспрессии модулятора TRRE на высоком уровне, и этот тест требует только, чтобы тестируемое вещество контактировало с этими клетками. Этот вариант делает возможным скрининг с высокой пропускной способностью большого числа тестируемых соединений.
В каждом случае скорость высвобождения рецептора сравнивают в присутствии и в отсутствие тестируемого вещества для идентификации соединений, которые увеличивают или уменьшают TRRE-активность. Должны проводиться параллельные эксперименты, в которых активность вещества в выделении рецептора из клетки испытывают в отсутствие добавленного полипептида (с использованием клеток, которые не экспрессируют этот полипептид). Это будет определять, проявляется ли активность тестируемого вещества через его действие на добавляемый стимулятор TRRE или через некоторый другой механизм.
Этот тип скрининга применим для идентификации антител, которые действуют на активность модулятора TRRE. Индуцируют образование антител против модулятора TRRE, как описано в предыдущем разделе. Если антитело уменьшает активность TRRE в этом тесте скрининга, то оно имеет терапевтический потенциал снижения TRRE-активности in vivo. Скрининг моноклональных антител с использованием этого теста может также способствовать идентификации сайтов связывания или каталитических сайтов в этом полипептиде.
Этот тип теста скрининга применим также для скрининга с высокой пропускной способностью соединений с небольшой молекулой, которые способны действовать на уровень рецепторов TNF на клетке, посредством их влияния на модулятор TRRE. Низкомолекулярные соединения, имеющие эту желательную активность, часто являются предпочтительными для фармацевтических композиций, так как они часто являются более стабильными и менее дорогими для получения.
Лекарственные средства и их применение
Как описано ранее, применимость некоторых продуктов, являющихся вариантами данного изобретения, заключается в действии на трансдукцию сигнала от цитокинов (в частности, от TNF). Продукты, которые стимулируют TRRE-активность, обладают способностью уменьшать количество рецепторов TNF на поверхности клеток, что будет уменьшать трансдукцию сигнала от TNF. Напротив, продукты, которые ингибируют TRRE-активность, предотвращают расщепление рецепторов TNF, усиливая трансдукцию сигнала.
Способность воздействовать на трансдукцию сигнала TNF представляет существенный интерес в управлении клиническими состояниями, в которых передача сигнала TNF способствует патологии этого состояния. Такие состояния включают в себя:
- Сердечную недостаточность. Считается, что IL-1β и TNF являются центральными медиаторами для превращения воспалительного процесса в хронический, рекрутинга и активации воспалительных клеток. Воспаление ослабляет сердечную функцию в случае застойной сердечной недостаточности, при отторжении трансплантата, миокардите, сепсисе и ожоговом шоке.
- Кахексию. Общую потерю веса и истощение, встречающиеся в ходе хронических заболеваний, таких как рак. Считают, что TNF влияет на аппетит, расходование энергии и скорость обмена веществ.
- Болезнь Крона. Воспалительный процесс, опосредованный TNF, приводит к утолщению стенки кишечника, происходящему из лимфедемы и инфильтрации лимфоцитов.
- Эндотоксиновый бактериально-токсический шок. Шок, индуцированный высвобождением эндотоксинов из грамотрицательных бактерий, таких как Е. coli, включает в себя TNF-медиируемое воспаление.
- Артрит. TNF стимулирует экспрессию синтетазы оксида азота, которая, как считают, участвует в патогенезе заболевания.
Другими представляющими интерес состояниями являются множественный склероз, сепсис, воспаление, вызываемое микробной инфекцией, и заболевания, имеющие аутоиммунную этиологию, такие как диабет типа I.
Полипептиды данного изобретения, которые стимулируют TRRE-активность, могут вводиться с целью уменьшения или нормализации трансдукции сигнала TNF. Например, в случае застойной сердечной недостаточности или болезни Крона полипептид предоставляют с регулярными интервалами для ослабления воспалительных последствий. Это лечение проводят в случае необходимости с другими агентами, которые влияют на трансдукцию сигнала TNF (такими как антитела к TNF или антагонисты рецептора) или которые ослабляют степень воспаления другими путями.
Полинуклеотиды данного изобретения могут быть также использованы для стимуляции TRRE-активности при помощи генной терапии. Кодирующую последовательность функционально (оперативно) связывают с регуляторными элементами для транскрипции и трансляции в клетках человека. Затем ее обеспечивают в форме, которая будет стимулировать вхождение и экспрессию этой кодирующей последовательности в клетки в месте заболевания. Формы, пригодные для локальной инъекции, включают в себя "оголенную" (депротеинизированную) ДНК, полинуклеотиды, упакованные с катионными липидами, и полинуклеотиды в форме вирусных векторов (таких как аденовирусные и AAV-конструкции). Способы генной терапии, известные практикующему врачу с квалификацией в данной области, будут включать в себя способы, описанные в патентах США с номерами 5399346, 5827703 и 5866696.
Способность воздействовать на трансдукцию сигнала TNF представляет также интерес в том случае, когда считают, что TNF играет благоприятную роль в устранении заболевания. В частности, TNF играет роль в индуцировании некроза твердых опухолей. Таким образом, продукты данного изобретения могут вводиться раковым пациентам для ингибирования TRRE-активности, с увеличением в результате трансдукции сигнала TNF и улучшения благоприятного действия.
Варианты данного изобретения, которые ингибируют TRRE-активность, включают в себя антисмысловые полинуклеотиды.
Способом придания длительно сохраняющейся ингибиторной активности является терапия с введением антисмысловых генов. Конструируют генетическую конструкцию, которая будет экспрессировать РНК в клетке, которая, в свою очередь, будет снижать транскрипцию гена-мишени (патент США №5759829). У людей, более частой формой антисмысловой терапии является введение антисмысловой молекулы непосредственно, в форме короткого стабильного полинуклеотидного фрагмента, который комплементарен сегменту целевой мРНК (патенты США с номерами 5135917 и 5789573) - в этом случае, транскрипта, который кодирует TRRE-модулятор. Другим вариантом данного изобретения, который ингибирует TRRE, являются рибозимы, сконструированные, как описано в приведенном ранее разделе. Функция рибозимов в ингибировании трансляции мРНК описана в патентах США с номерами 4987071 и 5591610.
После того как обнаружено, что продукт данного изобретения обладает подходящей TRRE-модулирующей активностью в тестах in vitro, описанных в данном описании, предпочтительно его эффективность тестируют также в животной модели TNF-медиируемого процесса заболевания. В примере 3 описана ЛПС-модель для сепсиса, которая может быть использована для испытания стимуляторов TRRE-активности. В примере 4 описана модель некроза опухолей, в которой TRRE-ингибиторы могли бы тестироваться на способность усиливать вызывающую некроз активность. Специалистам в данной области будут известны другие модели животных, пригодные для испытания действий на трансдукцию сигнала TNF или воспаление. Другими иллюстрациями являются реперфузионные модели сердечной ишемии Weyrich et al. (J. Clin. Invest. 91:2620, 1993) и Garcia-Criado et al. (J. Am. Coll. Surg. 181:327, 1995); реперфузионная модель легочной ишемии Steinberg et al. (J. Heart Lung Transplant. 13:306, 1994), модель воспаления легких Международной патентной заявки WO 9635418; модель бактериального перитонита Sharar et al. (J. Immunol. 151:4982, 1993), модель колита Meenan et al. (Scand. J. Gastroenterol. 31:786, 1996) и модель диабета von Herrath et al. (J. Clin. Invest. 98:1324, 1996). Модели септического шока описаны в Mack et al. J. Surg. Res. 69:399, 1997 и Seljelid et al. Scand. J. Immunol. 45:683-7.
Для применения в качестве активного ингредиента в фармацевтической композиции полипептид, полинуклеотид или антитело данного изобретения обычно очищают от других реакционноспособных или потенциально иммуногенных компонентов, присутствующих в смеси, в которой они получены. Обычно каждый активный ингредиент обеспечивают в по меньшей мере 90% гомогенности и более предпочтительно 95% или 99% гомогенности, как определено функциональным анализом, хроматографией или электрофорезом в ДСН-полиакриламидном геле. Затем активный ингредиент компаундируют в лекарственное средство в соответствии с общепринятыми процедурами для приготовления фармацевтических препаратов, такими как описанные в Remington's Pharmaceutical Sciences, 18th Ed. (1990), E.W.Martin ed., Mack Publishing Co., PA. Стадии в компаундировании лекарственного средства зависят отчасти от предполагаемого использования и способа введения и могут включать в себя стерилизацию, смешивание с подходящими нетоксичными и немешающими наполнителями и носителями, разделение на дозированные единицы и заключение в устройство для доставки. Лекарственное средство обычно упаковывают с приложением информации о предполагаемом применении.
Режим введения будет зависеть от характера состояния, подлежащего лечению. Для состояний, которые, как ожидается, требуют умеренного дозирования и которые находятся при хорошо перфузируемых участках (например, сердечной недостаточности), приемлемым является системное введение. Например, лекарственное средство может быть приготовлено для внутривенного введения, внутримышечной инъекции или подъязычного или интраназального всасывания. Если возможно вводить активный ингредиент локально, этот способ обычно является предпочтительным. Локальное введение будет как увеличивать концентрацию активного ингредиента в месте заболевания, так и минимизировать действие на рецепторы TNF на других тканях, не вовлеченных в процесс заболевания. Состояния, которые пригодны для введения непосредственно в месте заболевания, включают в себя рак и ревматоидный артрит. Инъекцию непосредственно в твердые опухоли можно выполнять, когда они расположены близко от кожи или когда их можно достичь при помощи эндоскопической процедуры. Активные ингредиенты могут быть также введены в участок опухоли во время хирургической резекции посредством имплантации в гелеобразном матриксе или в подходящей мембране, такой как Gliadel® (Guilford Sciences). В том случае, когда прямое введение невозможно, введение может осуществляться через артериолы, ведущие к месту заболевания. Альтернативно, фармацевтическая композиция может быть приготовлена таким образом, что она усиливает накопление активного ингредиента в месте заболевания. Например, активный ингредиент может быть инкапсулирован в липосоме или в матриксе другой структуры, который имеет на его поверхности антитело или лиганд, способные связывать белок клеточной поверхности на клетке-мишени. Подходящие нацеливающие агенты включают в себя антитела против раковых антигенов, лиганды для тканеспецифических рецепторов (например, серотонин для легочного нацеливания). Для композиций, которые снижают трансдукцию сигнала TNF, подходящей нацеливающей молекулой может быть лиганд TNF, так как ткань-мишень может, по-видимому, обнаруживать на поверхности высокую плотность рецептора TNF.
Эффективные количества композиций данного изобретения являются количествами, которые изменяют TRRE-активность по меньшей мере на приблизительно 10%, обычно по меньшей мере на приблизительно 25%, более предпочтительно по меньшей мере на приблизительно 50% или 75%. Если желательно почти полное устранение TRRE-активности, предпочтительные композиции уменьшают TRRE-активность по меньшей мере на 90%. Если желательно увеличение TRRE-активности, предпочтительные композиции увеличивают TRRE-активность по меньшей мере в 2 раза. Минимальное эффективное количество активного соединения будет зависеть от подлежащего лечению состояния, от конкретного выбранного для применения модулятора TRRE и от того, будет ли введение системным или местным (локальным). Для системного введения эффективным количеством будет обычно количество модулятора TRRE, которое может вызвать изменение в активности фермента на 100-50000 единиц - в типичном случае на приблизительно 10000 единиц. Весовое количество белка, нуклеиновой кислоты или антитела выбирают соответственно на основе удельной активности активного соединения в единицах на грамм.
Нижеследующие примеры дают дополнительное руководство для специалиста, применяющего на практике данное изобретение, и не предназначены для ограничения каким бы то ни было образом данного изобретения.
ПРИМЕРЫ
Пример 1: Тест-система для определения TRRE-активности
Этот пример иллюстрирует тест-систему, при помощи которой измеряют TRRE-активность в отношении TNF-R человека в его нативной конформации в мембране клеточной поверхности.
Мембраносвязанный TNF-R был выбран в качестве субстрата как имеющий микроокружение, сходное с микроокружением этого субстрата для TRRE in vivo. Мембраносвязанный TNF-R требует также более специфическую активность, что могло бы дифференцировать менее специфические протеазы. Клетки, экспрессирующие повышенный уровень формы р75 TNF-R, конструировали трансфекцией кДНК в клетки обезьяны COS-1, которые экспрессируют мало TNF-R размером как 75 кДа, так и 55 кДа.
Процедура конструирования этих клеток была следующей: кДНК TNF-R р 75 человека клонировали из библиотеки кДНК λgt10, полученной из моноцитарных клеток U-937 (Clontech Laboratories, Palo Alto, CA). Первые 300 п.н. как на 5'-, так и на 3'-концах клонированного фрагмента секвенировали и сравнивали с сообщенной последовательностью кДНК TNF-R р75 человека. Клонированная последовательность была фрагментом 2,3 т.п.н., охватывающим положения 58-2380 сообщенной последовательности TNF-R р75, которая включает в себя полную длину последовательности, кодирующей TNF-R р75, от положения 90 до положения 1475. Затем кДНК 2,3 т.п.н. TNF-R р75 субклонировали в сайт множественного клонирования эукариотического экспрессионного вектора pCDNA3. Ориентацию кДНК TNF-R р75 подтверждали рестикционным картированием.
Фигура 1 иллюстрирует конечную конструкцию 7,7 т.п.н. pCDNA3. Она несет ген устойчивости к неомицину для отбора трансфицированных клеток в G418, и экспрессия TNF-R р75 запускается промотором цитомегаловируса. Затем pCDNA3 трансфицируют в клетки почек обезьяны COS-1 (ATCC CRL 1650) с использованием способа осаждения ДНК фосфатом кальция. Выбранный клон в среде G418 идентифицировали и субклонировали. Этот клон был назван C75R.
Для определения уровня экспрессии TNF-R р75 на клетках C75R 2×106 клеток на лунку высевали в 24-луночный культуральный планшет и инкубировали в течение 12-16 часов в 5% СО2 при 37°С. Затем их инкубировали с 2-30 нг рекомбинантным 125I-TNF человека (радиоактивно меченым по способу с хлорамином Т) в присутствии или в отсутствие 100-кратного избытка немеченого TNF человека при 4°С в течение 2 часов. После трех промывок охлажденным на льду ЗФР клетки лизировали 0,1 н. NaOH и связанную радиоактивность определяли в счетчике Pharmacia Clinigamma (Uppsala, Sweden).
Фигура 2 показывает полученные результаты. C75R имел очень высокий уровень специфического связывания радиоактивно меченого 125I-TNF, тогда как исходные клетки COS-1 не имели такого уровня. При помощи анализа Скетчарда было определено, что число TNF-R, экспрессируемых на C75R, равно 60000-70000 рецепторов на клетку (фигура 2, вставка). Рассчитанная величина Kd была 5,5×10-10 М. Эта величина Kd хорошо согласуется с величинами, сообщенными ранее для нативного TNF-R р75.
TRRE получали ФМА-стимуляцией клеток ТНР-1 (WO 9802140). Клетки ТНР-1 (АТСС 45503), растущие в логарифмической фазе, собирали и ресуспендировали до 1×106 клеток/мл в RPMI-1640, дополненной 1% ФТС, и инкубировали с 10-6 М ФМА в течение 30 минут в 5% CO2 при 37°С. Клетки собирали и промывали один раз бессывороточной средой для удаления ФМА и ресуспендировали в таком же объеме среды RPMI-1640 с 1% ФТС. После 2 часов инкубации в 5% CO2 при 37°С клеточную суспензию собирали, центрифугировали и бесклеточный супернатант собирали в качестве источника TRRE.
Для измерения действия TRRE на мембраносвязанный TNF-R в конструкциях клеток COS-1 проводили следующий эксперимент. Клетки C75R высевали при плотности 2×105 клеток на лунку в 24-луночный культуральный планшет и инкубировали в течение 12-16 часов в 5% СО2 при 37°С. Среду в этих лунках отсасывали, заменяли только свежей средой или средой с TRRE и инкубировали в течение 30 минут при 37°С. Затем среду заменяли свежей средой, содержащей 30 нг/мл 125I-меченого TNF. После 2 часов при 4°С клетки лизировали 0,1 н. NaOH и измеряли уровень связанной радиоактивности. Уровень специфического связывания C75R 125I-TNF значимо снижался после инкубации с TRRE. Радиоактивность была 1393 имп/мин на клетках, инкубированных с TRRE, в сравнении с 10567 имп/мин на клетках, не обработанных TRRE, что означало 87% потерю связывающей способности.
Для определения размера TNF-R р75, выделяемого из C75R посредством TRRE, проводили следующий эксперимент. 15×106 клеток C75R высевали в планшет 150 мм для культуры клеток и инкубировали при 5% СО2 при 37°С в течение 12-16 часов. Среду с TRRE инкубировали с клетками C75R в планшете 150 мм в течение 30 минут и полученный супернатант собирали и центрифугировали. Концентрированную пробу наносили на 10% акриламид-ДСН для электрофореза и электрофоретически переносили на поливинилидендифторидную мембрану (Immobilon). Иммуноокрашивание давало единственную полосу 40 кДа, сходную с размером, обнаруживаемым в биологических жидкостях. Таким образом, трансфицированные клетки COS-1 экспрессировали высокие уровни TNF-R р75 человека в форме, сходной с нативным TNF-R.
Следующий способ определения был использован для рутиннного измерения TRRE-активности. Клетки C75R и клетки COS-1 высевали в 24-луночные культуральные планшеты при плотности 2,5×106 клеток/мл/лунка и инкубировали в течение ночи (в течение 12-16 часов) в 5% CO2 при 37°С. После отсасывания среды в лунке 300 мкл TRRE-среды инкубировали в каждой лунке планшетов C75R и COS-1 в течение 30 минут в 5% СО2 при 37°С (соответствующих А и С, упоминаемых ниже, соответственно). Одновременно клетки C75R в 24-луночных планшетах инкубировали также с 300 мкл свежей среды или буфера. Супернатанты собирали, центрифугировали и затем анализировали на концентрацию растворимого TNF-R р75 при помощи ELISA.
Анализ ELISA для выделившегося TNF-R (WO 9802140) выполняли следующим образом. Поликлональные антитела к TNF-R р75 человека генерировали иммунизацией самок Ново-Зеландских белых кроликов (Yamamoto et al. Cell. Immunol. 38:403-416, 1978). IgG-фракцию сыворотки иммунизированных кроликов очищали с использованием аффинной колонки с белком G (Pharmacia Fine Chemicals, Uppsala, Sweden) (Ey et al. (1978) Immunochemistry 15:429-436, 1978). Затем IgG-фракцию метили пероксидазой хрена (Sigma Chemical Co., St. Louis, МО) (Tijssen and Kurstok, Anal. Biochem. 136:451-457, 1984). В первой стадии этого анализа 5 мкг немеченого IgG в 100 мкл 0,05 М карбонатного буфера (рН 9,6) связывали с 96-луночным микропланшетом для ELISA (Corning, Corning, NY) инкубированием в течение ночи при 4°С. Индивидуальные лунки промывали три раза 300 мкл 0,2% Твина-20 в забуференном фосфатом солевом растворе (ЗФР). В каждую лунку добавляли 100 мкл проб и стандарты рекомбинантного рецептора и инкубировали при 37°С в течение 1-2 часов. Затем лунки промывали таким же образом, добавляли 100 мкл меченого пероксидазой хрена IgG и инкубировали в течение 1 часа при 37°С. Лунки промывали еще раз и окраску проявляли в течение 20 минут при комнатной температуре с субстратами ABTS (Pierce, Rockford, IL) и 30% Н2О2 (Fisher Scientific, Fair Lawn, NJ). Развитие окраски измеряли при 405 нм.
При инкубировании клеток C75R с TRRE-средой растворимый TNF-R р75 высвобождался в супернатант, который можно было измерить при помощи ELISA. Количество высвобождаемых рецепторов соответствовало количеству добавленного TRRE. Имелся также некоторый уровень самопроизвольного высвобождения в клетках C75R, инкубированных только со средой. Высказывается гипотеза, что это было обусловлено эндогенным источником протеолитического фермента, происходящего из обезьяны, гомолога TRRE человека.
Были проведены следующие расчеты. А = (количество растворимого TNF-R р75 в С75R-планшете, обработанном содержащей TRRE пробой); т.е. общее количество растворимого sTNF-R в С75R-планшете. В = (количество растворимого TNF-R р75, самопроизвольно высвобождаемого в С75R-планшете, обработанном только средой или буфером, содержащим тот же самый реагент, что и соответствующие пробы, но без экзогенного TRRE); т.е. самопроизвольное высвобождение sTNF-R из клеток C75R. С = (количество растворимого TNF-R р75 в COS-1-планшете, обработанном TRRE-пробой, или уровень фона растворимого TNF-R р75, высвобождаемого ТНР-1); т.е. деградированая величина перенесенных (предсуществующих) sTNF-R в TRRE-пробе во время 30-минутной инкубации в COS-1-планшете. Это соответствует уровню фона sTNF-R, деградированного в С75R-планшете. Нетто-высвобождение растворимого TNF-R р75, продуцируемого только TRRE-активностью, существующей в исходной пробе, рассчитывают следующим образом: (Нетто-высвобождение растворимого TNF-R р75 только под действием TRRE) = А-В-С.
Унифицированную активность (активность, принимаемую за единицу активности) TRRE определяли следующим образом: 1 пг нетто-высвобождения растворимого TNF-R р75 (А-В-С) в ходе этого анализа является одной единицей (Е) TRRE-активности.
С использованием этого анализа временной ход выделения рецептора посредством TRRE измеряли в следующем эксперименте. TRRE-среду инкубировали с клетками C75R и клетками COS-1 в течение варьируемых периодов времени. Затем супернатанты собирали и анализировали на уровень растворимого TNF-R р75 при помощи ELISA и рассчитывали нетто-активность TRRE. Детектируемые уровни растворимого рецептора выделялись TRRE в пределах 5 минут и увеличивались вплоть до 30 минут. Более продолжительные периоды инкубирования показали, что уровень TRRE оставался относительно константным после 30 минут, предположительно из-за истощения субстратов. Таким образом, было определено, что оптимальным временем инкубирования является период в 30 минут.
Распределения индукции TRRE и известных ММР при ФМА-стимуляции являются совершенно различными. Для индукции ММР, моноцитарных клеток U-937, клетки фибросаркомы НТ-1080 или макрофаги перитонеального экссудата (РЕМ) обычно должны стимулироваться в течение одного-двух дней с использованием ЛПС или ФМА. С другой стороны, в сравнении с этой пролонгированной индукцией, TRRE высвобождается очень быстро в культуральном супернатанте после 30 минут ФМА-стимуляции. Гипотеза, что TRRE и sTNF-R образуют комплекс in vitro, была подтверждена экспериментом, в котором 25% TRRE-активность извлекали из колонки с растворимым TNF-R р75. Это означает, что свободный TRRE способен связываться с его каталитическим продуктом, sTNF-R. Остальные 75%, которые не соединялись с аффинной колонкой, могут быть уже связанными с sTNF-R или могут не иметь достаточной аффинности для связывания с sTNF-R даже, когда он находится в свободной форме.
Пример 2: Характеристика TRRE, полученного из клеток ТНР-1
TRRE, полученный ФМА-стимуляцией клеток ТНР-1, частично очищали из культуральной среды (WO 9802140). Сначала белок из этой среды концентрировали осаждением 100%-насыщенным сульфатом аммония при 4°С. Осадок осаждали центрифугированием при 10000×g в течение 30 минут и ресуспендировали в ЗФР в приблизительно двойном объеме относительно осадка. Затем этот раствор диализовали при 4°С против 10 мМ Трис-HCl, 60 мМ NaCl, рН 7,0. Эту пробу наносили на анионообменную колонку для хроматографии, колонку А-25 диэтиламиноэтил (ДЭАЭ)-Сефадекса (Pharmacia Biotech) (2,5×10 см), предварительно уравновешенную 50 мМ Трис-HCl, 60 мМ NaCl, рН 8,0. Затем TRRE элюировали линейным градиентом ионной силы 60-250 мМ NaCl, 50 мМ Трис-HCl, рН 8,0. Каждую фракцию измеряли на поглощение при 280 нм и анализировали на активность TRRE. ДЭАЭ-фракции с наивысшей удельной активностью (наивысшей величиной единиц TRRE/A280) объединяли и использовали в характеристиках TRRE, описанных в этом примере.
В следующем эксперименте выясняли субстратную специфичность этого фермента с использованием иммуногистохимических способов. Использовали флуоресцеинизотиоцианат(ФИТЦ)-конъюгированные анти-С054, ФИТЦ-конъюгированные козьи антикроличьи и антимышиные антитела, мышиные моноклональные антитела анти-СD30, анти-CD11b и анти-IL-1R (Serotec, Washington D.C.) Кроличьи поликлональные антитела анти-TNF-R р55 и анти-TNF-R р75 получали в соответствии с Yamamoto et al. (1978) Cell Immunol. 38:403-416. Клетки ТНР-1 обрабатывали в течение 30 минут 1000 и/или 5000 Е/мл TRRE, элюированного из ДЭАЭ-Сефадекс-колонки, и затем переносили в полистирольные пробирки 12×75 мм (Fisher Scientific, Pittsburgh, PA) при 1×105 клеток на 100 мкл на пробирку. Затем эти клетки осаждали центрифугированием при 350×g в течение 5 минут при 4°С и окрашивали непосредственно 10 мкл ФИТЦ-конъюгированных антител анти-СD54 (разведенных в холодной смеси ЗФР/0,5% азид натрия), опосредованно ФИТЦ-конъюгированным антимышиным антителом после обработки мышиными моноклональными антителами анти-CD11b, анти-IL-1R и анти-СD30 и также опосредованно ФИТЦ-конъюгированным антикроличьим антителом после обработки кроличьими поликлональными антителами анти-TNF-R р55 и анти-TNF-R р75.
Клетки ТНР-1, окрашенные каждым из этих антител без обработки TRRE, использовали в качестве отрицательных контролей. Пробирки инкубировали в течение 45 минут при 4°С, встряхивали каждые 15 минут, промывали дважды ЗФР/2% ФТС, повторно осаждали и затем ресуспендировали в 200 мкл 1% параформальдегида. Эти меченые клетки ТНР-1 анализировали с использованием клеточного сортера с возбуждением флуоресценции (FACS) (Becton Dickenson, San Jose, CA) с аргоновым лазером 15 мВ с возбуждением 488 нм. Флуоресцентные сигналы дискриминировали на основе рассеяния света вперед или под прямым углом для исключения мертвых клеток и агрегатов из анализа. Дискриминированные флуоресцентные сигналы (104) детектировали при фильтре 585 ВР и анализировали с использованием программного обеспечения Lysis II. Величины выражали в виде процента положительных клеток, который рассчитывали делением интенсивности флуоресценции средних каналов (MFI) окрашенных клеток ТНР-1, обработанных TRRE, на MFI клеток без обработки TRRE (клеток отрицательного контроля).
Для испытания TNF-цитолитического действия in vitro при обработке TRRE цитолитический тест на клетках L929 выполняли согласно способу, описанному Gatanaga et al. (1990b). Вкратце, клетки L929, прикрепленную клеточную линию мышиных фибробластов, высевали (70000 клеток/0,1 мл на лунку в 96-луночном планшете) и инкубировали в течение ночи. Монослойные клетки L929 предобрабатывали в течение 30 минут 100, 500 или 2500 Е/мл частично очищенного TRRE и затем экспонировали с серийными разведениями рекомбинантного TNF человека в течение 1 часа. После промывания планшета RPMI-1640 с 10% ФТС для удаления TRRE и TNF клетки инкубировали в течение 18 часов в RPMI-1640 с 10% ФТС, содержащей 1 мкг/мл актиномицина В, при 37°С в 5% СО2. Затем культуральные супернатанты отсасывали и в каждую лунку добавляли 50 мкл 1% раствора фиолетового кристаллического красителя. Планшеты инкубировали в течение 15 минут при комнатной температуре. После промывания планшетов водопроводной водой и сушки на воздухе клетки, окрашенные фиолетовым кристаллическим, лизировали добавлением 100 мкл на лунку 100 мМ HCl в метаноле. Поглощение при 550 нм измеряли с использованием планшет-ридера EAR 400 AT (SLT-Labinstruments, Salzburg, Austria).
Для выяснения того, укорачивает ли TRRE также размер ˜55 кДа TNF-R частично очищенный TRRE применяли к клеткам ТНР-1, которые экспрессируют низкие уровни как TNF-R р55, так и TNF-R р75 (приблизительно 1500 рецепторов на клетку согласно анализу Скетчарда). Элюат TRRE из колонки ДЭАЭ-Сефадекса добавляли к клеткам ТНР-1 (5×106 клеток/мл) при конечной концентрации TRRE 1000 Е/мл и инкубировали в течение 30 минут. Концентрацию растворимых TNF-R р55 и TNF-R р75 в этом супернатанте измеряли при помощи анализа (ELISA) растворимых TNF-R р55 и TNF-R р75. Было обнаружено, что TRRE укорачивает как TNF-R р55, так и TNF-R р75 человека на клетках ТНР-1 и высвобождает 2382 и 1682 пг/мл растворимых TNF-R р55 и TNF-R р75 соответственно.
Таким образом, TRRE, полученный ФМА-стимуляцией клеток ТНР-1, способен ферментативно расщеплять и высвобождать TNF-R р75 человека на клетках C75R и как TNF-R р55, так и TNF-R р75 человека на клетках ТНР-1.
Частичное ингибирование TRRE-активности получали под действием хелатообразователей, таких как 1,10-фенантролин, ЭДТА и ЭГТА (%% остающейся TRRE-активности были 41%, 67% и 73% соответственно при концентрации 2 мМ). С другой стороны, ингибиторы сериновых протеаз, такие как ФМСФ, AEBSF и 3,4-DCl, и ингибиторы сериновых и цистеиновых протеаз, такие как TLCK и ТРСК, не оказывали влияния на ингибирование TRRE. TRRE слегка активировался в присутствии Mn2+, Са2+, Mg2+ и Со2+ (%% остающейся TRRE-активности были 157%, 151%, 127% и 123% соответственно), тогда как в присутствии Zn2+ и Cu2+ происходило частичное ингибирование (%% остающейся TRRE-активности были 23% и 47% соответственно) (WO 9802140).
TRRE-фракции из наиболее активной ДЭАЭ-фракции (60 мМ - 250 мМ NaCl) можно было очистить дополнительно. В одном способе (WO 0802140) эти фракции концентрировали до 500 мкл с использованием фильтра Centriprep-10 (мембрана с отсечением MW 10000) (Amicon). Эту концентрированную пробу подвергали электрофорезу в 6% ПААГ при неденатурирующих нативных условиях. Гель нарезали горизонтально на полоски по 5 мм и каждую из них элюировали в 1 мл ЗФР. Затем элюаты анализировали согласно анализу примера 1 на TRRE-активность.
Пример 3: TRRE-активность ослабляет септический шок
Нижеследующий протокол использовали для определения действий TRRE в предотвращении смертности в модели септического шока. Мышей инъецировали летальными или сублетальными уровнями ЛПС и затем контрольным буфером или TRRE. Затем собирали пробы периферической крови с интервалами для определения, ингибировал ли TRRE индуцированное TNF продуцирование других цитокинов в кровотоке. Животных оценивали на способность TRRE ингибировать клинические эффекты шока и затем эвтанизировали и ткани исследовали гистопатологическими способами.
Детали были следующими: взрослых мышей Balb/c помещали в приспособление для фиксирования тела и инъецировали внутривенно через хвостовую вену 0,1 мл раствора, содержащего 10 нг - 10 мг ЛПС в забуференном фосфатом солевом растворе (ЗФР). Эти уровни ЛПС индуцируют уровни от слабого до летального шока в этом штамме мышей. Шок происходит из изменений в проницаемости сосудов, потери жидкости и дегидратации и часто сопровождается симптомами, включающими в себя летаргию, сгорбленное, неподвижное положение, мятую шерсть, прекращение приема пищи, цианоз и, в серьезных случаях, смерть в пределах 12-24 часов. Контрольные мыши получали инъекцию ЗФР. Различные количества (2000 или 4000 Е) очищенного TRRE человека инъецировали в/в в объеме 0,1 мл в пределах часа до или после инъекции ЛПС. Сыворотку (0,1 мл) собирали иглой 27 G и шприцом IV на 1 мл из хвостовой вены при 30, 60 и 90 минутах после инъекции ЛПС. Сыворотку гепаринизировали и хранили в замороженном виде при -20°С. Пробы из многочисленных экспериментов анализировали при помощи ELISA на присутствие sTNF-R, TNF, IL-8 и IL-6. Животных подвергали мониторингу на протяжении следующих 12 часов на клинические эффекты шока. Выбранных животных эвтанизировали в периоды от 3 до 12 часов после обработки, проводили аутопсию и различные органы и ткани фиксировали в формалине, заливали в парафин, делали срезы и окрашивали гематоксилином-эозином (Г и Э). Срезы тканей подвергали гистопатологическому и иммунопатологическому исследованию.
Фигура 3 показывает полученные результаты. (◆) только ЛПС; () ЛПС плюс контрольный буфер; ( ) ЛПС плюс TRRE (2000 Е); (
) ЛПС плюс TRRE (2000 Е); ( ) ЛПС плюс TRRE (4000 Е).
) ЛПС плюс TRRE (4000 Е).
Мыши, инъецированные только ЛПС или ЛПС и контрольным буфером, умирали вскоре после инъекции. 50% испытуемых животных умирали спустя 8 часов (ЛПС) или 9 часов (ЛПС плюс контрольный буфер) и 100% животных умирали через 15 часов. В противоположность этому животные, получавшие TRRE, полученный, как описано в примере 1, дали гораздо лучшие результаты. Когда инъекции ЛПС сопровождались инъекциями 2000 Е TRRE, смерть задерживалась, и коэффициенты смертности были более низкими. Только 40% этих животных были мертвы через 24 часа. Когда 4000 Е TRRE инъецировали вместе с ЛПС, все животные были живыми спустя 24 часа. Таким образом, TRRE способен противодействовать смертности испытуемых животных, индуцированной ЛПС.
Пример 4: TRRE-активность уменьшает активность некротизации опухолей
Следующий протокол использовали для исследования действий TRRE на некроз опухолей у испытуемых животных, в которых были индуцированы опухоли и в которые затем инъецировали TNF.
В день 0 кожные опухоли Meth А индуцировали на абдоминальной стенке пятнадцати мышей BALB/c чрескожной инъекцией 2×205 клеток опухоли Meth А. В день 7 мышей делили на три группы из пяти мышей каждая и обрабатывали следующим образом:
- Группа 1: инъецированные внутривенно TNF (1 мкг/мышь).
- Группа 2: инъецированные внутривенно TNF (1 мкг/мышь) и инъецированные внутриопухолевой инъекцией TRRE, полученного, как описано в примере 1 (400 единиц/мышь, 6, 12 часов после инъекции TNF).
- Группа 3: инъецированные внутривенно TNF (1 мкг/мышь) и инъецированные внутриопухолевой инъекцией контрольной среды (6, 12 часов после инъекции TNF).
В день 8 некроз опухолей измеряли с получением следующих результатов: Группа 1: 100% некроз (5/5); Группа 2: 20% (1/5); Группа 3: 80% (4/5). Инъекции TRRE значительно снизили способность TNF индуцировать некроз в опухолях Meth А в мышах BALB/c.
Поскольку добавление TRRE-активности устраняет благоприятную некротизирующую активность TNF, ингибирование эндогенной TRRE-активности могло бы стимулировать выгодные эффекты TNF.
Пример 5: Девять новых полинуклеотидных клонов, которые действуют на TRRE-активность
Было обнаружено, что ряд клеток экспрессируют высокие уровни TRRE-активности, особенно после ФМА-стимуляции. Этими клетками являются клеточные линии, обозначаемые ТНР-1, U-937, HL-60, ME-180, MRC-5, Raji, K-562. Клетки Jurkat имеют высокую TRRE-активность (850 TRRE Е/мл при 10-2 ФМА). В этом эксперименте получали экспрессионную библиотеку Т-клеток Jurkat (АТСС # TIB-152) и использовали ее для получения 9 полинуклеотидных клонов, которые увеличивают TRRE-активность.
Отбор экспрессионных последовательностей в этой библиотеке производили посредством повторяющихся циклов трансфекции в клетки COS-1 с последующим анализом супернатанта, как в примере 1, на расщепление и высвобождение рецептора TNF. Использовали стандартные способы генетического манипулирования. Вкратце, ДНК 106 клеток Jurkat экстрагировали с использованием набора InVitrogen для экстракции плазмид в соответствии с инструкциями изготовителя. кДНК встраивали в вектор ZAP Express™/EcoRI (номер по каталогу 938201, Stratagene, La Jolla CA). Библиотеку делили на 48 групп ДНК и трансформировали в клетки COS-1 при помощи способа трансфекции с CaCl2. Как только эти клетки вырастали, проводили определение TRRE, и были отобраны пять положительных групп. Получали ДНК из каждой из этих пяти групп и трансфицировали в Е. coli, с 15 чашками на группу. ДНК получали из этих клеток и затем ее трансфицировали в клетки COS-1 еще раз. Клетки выращивали и опять определяли TRRE-активность. Были отобраны две положительные группы, и их трансфицировали в Е. coli с получением 98 колоний. Из 96 из этих колоний получали ДНК и трансфицировали ее в клетки COS-1. Опять проводили определение TRRE-активности и обнаружили, что девять клонов существенно увеличивали TRRE-активность в этом анализе. Эти клоны были обозначены 2-8, 2-9, 2-14, 2-15, Р2-2, Р2-10, Р2-13, Р2-14 и Р2-15.
Фигура 4 является диаграммой в виде столбцов, показывающей TRRE-активность, наблюдаемую при испытании этих 9 клонов с клетками С75 в стандартном анализе (пример 1).
Затем эти девять клонов секвенировали согласно следующей процедуре:
1. Плазмидную ДНК получали при помощи модифицированной процедуры щелочного лизиса.
2. Секвенирование ДНК проводили с использованием реакций Краситель-Дезокситерминации цепи (ABI). В качестве меток использовали специфические в отношении оснований флуоресцентные красители.
3. Реакции секвенирования анализировали на 5,75% гелях Long RangerТМ при помощи автоматизированного секвенатора ABI 373A-S или на 5,0% гелях Long RangerТМ при помощи автоматизированного секвенатора ABI 377.
4. Последующий анализ данных выполняли с использованием программного обеспечения SequencherТМ 3.0.
В реакциях секвенирования использовали стандартные праймеры Т7Х, Т3Х, -40, -48 Reverse (обратные) и ВК Reverse (обратный) (BKR). Для каждого клона синтезировали несколько дополнительных внутренних секвенирующих праймеров (перечисленных ниже).
Анализ последовательностей NCBI BLAST (Basic Local Alignment Search Tool) проводили для определения, были ли другие последовательности значимо сходными с этими последовательностями. Как ДНК-последовательности этих клонов, так и соответствующие открытые рамки считывания (ORF) (если они имелись) сравнивали с последовательностями, доступными в базах данных.
Были получены и секвенированы следующие клоны:
Клон 2-9 (AIM2): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:11-SEQ ID NO:38. Последовательность AIM2 представлена в SEQ ID NO:1. Комплементарная цепь последовательности AIM2 представляет собой SEQ ID NO:147. Самая длинная открытая рамка считывания (ORF) в последовательности AIM2 имеет длину 474 АА (аминокислоты) и представлена в SEQ ID NO:148.
Клон 2-8 (AIM3): Две частичные последовательности с длиной 739 и 233 были получены и обозначены AIM3T3 и AIM3T7. Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:39-SEQ ID NO:46. Последовательности AIM3T3 и AIM3T7 представлены в SEQ ID NO:2 и SEQ ID NO:3 соответственно. BLAST-поиск выявил, что последовательность AIM3T3 может быть гомологичной рибосомной РНК 28S мыши (М. musculus) (Hassouna et al. Nucleic Acids Res. 12:3563-3583, 1984) и генам пре-рРНК 45S М. musculus (номер доступа Х82564). Комплементарная последовательность последовательности AIM3T3 обнаружила 99% сходство на протяжении 408 п.н., начиная с нуклеотида 221 SEQ ID NO:2, с первой и 97% сходство на том же самом протяжении с последней.
Клон 2-14 (AIM4): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:14-SEQ ID NO:65. Последовательность AIM4 представлена в SEQ ID NO:4. Комплементарная цепь последовательности AIM4 представляет собой SEQ ID NO:149. Самая длинная открытая рамка считывания (ORF) в последовательности AIM4 имеет длину 236 АА (аминокислот) и представлена в SEQ ID NO:150. AIM4 имеет значимые совпадения при сопоставлении с последовательностями арфаптина 2 человека, мРНК ADE2H1, обнаруживая гомологии с SAICAR-синтазой, мРНК полипиримидиновый тракт-связывающего белка (гетерогенного ядерного рибонуклеопротеина I), несколькими генами РТВ для полимиридиновых тракт-связывающих белков, мРНК для белка porl. Арфаптин 2 человека является предполагаемым белком-мишенью фактора АДФ-рибозилирования, который взаимодействует с RAC1 посредством прямого связывания с ним. RAC1 участвует в активных перемещениях (волнах возбуждения) в мембране клетки. Арфаптин 2 имеет возможные трансмембранные сегменты, потенциальные сайты фосфорилирования СК2, сайт фосфорилирования РКС и последовательность присоединения к клетке RGD.
Клон 2-15 (AIM5): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:66-SEQ ID NO:80. Последовательность AIM5 представлена в SEQ ID NO:5. BLAST-поиск выявил, что последовательность AIM5 проявляет некоторое сходство с фактором инициации 5А (elF-5A) человека Koettnitz et al. (1995) Gene 159:283-284, 1995 и фактором инициации 4D человека (elF 4D) Smith-McBride et al. (1989) J. Biol. Chem. 264:1578-1583, 1989.
Клон Р2-2 (AIM6): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:81-SEQ ID NO:93. Последовательность AIM6 представлена в SEQ ID NO:6. Самая длинная открытая рамка считывания (ORF) в последовательности AIM6 имеет длину 1038 АА (аминокислот) и представлена в SEQ ID NO:151.
Клон Р2-10 (AIM7): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:94-SEQ ID NO:106. Последовательность AIM7 представлена в SEQ ID NO:7. Самая длинная открытая рамка считывания (ORF) в последовательности AIM7 имеет длину 849 АА (аминокислот) и представлена в SEQ ID NO:152. BLAST-поиск выявил, что этот клон может быть родственным мРНК рецептора инсулиноподобного фактора роста II человека (Morgan et al. Nature 329:301-307, 1987 или мРНК катион-независимого рецептора маннозо-6-фосфата человека (Oshima et al. J. Biol. Chem. 263:2553-2562, 1988). Последовательность AIM7 показала примерно 99% идентичность обеим этим последовательностям на протяжении 2520 нуклеотидов, начиная с нуклеотида 12 SEQ ID NO:7, и 99% сходство с последней на том же самом протяжении.
Клон Р2-13 (AIM8): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:107-SEQ ID NO:118. Последовательность AIM8 представлена в SEQ ID NO:8. Самая длинная открытая рамка считывания (ORF) в последовательности AIM8 имеет длину 852 АА (аминокислоты) и представлена в SEQ ID NO:153.
Клон Р2-14 (AIM9): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:119-SEQ ID NO:124. Последовательность AIM9 представлена в SEQ ID NO:9. Самая длинная открытая рамка считывания (ORF) имеет длину 149 АА (аминокислот).
Клон Р2-15 (AIM10): Внутренние праймеры, использованные для секвенирования, показаны в SEQ ID NO:125-SEQ ID NO:146. Последовательность AIM10 представлена в SEQ ID NO:10. Самая длинная открытая рамка считывания (ORF) в последовательности AIM10 имеет длину 693 АА (аминокислоты) и представлена в SEQ ID NO:154. Последовательность 10 при BLAST-поиске в неизбыточных базах данных при NCBI имеет совпадения при сопоставлении с мРНК Е1b-55кДа-связанного белка человека, локуса HSA7509 (номер доступа AJ007509, NID g3319955).
Клональные ДНК могут быть непосредственно инъецированы в тест-животных для определения способности этих нуклеиновых кислот индуцировать TRRE-активность, противодействовать септическому шоку и/или влиять на некроз опухолей, как описано подробно в примерах 3 и 4. Альтернативно, из клональных ДНК могут быть генерированы белки или РНК для подобного тестирования.
Пример 6: Экспрессия новых полученных клонов
В примере 5 описаны 9 новых клонов, которые усиливают TRRE-активность в системе анализа клеточной поверхности. Эти клоны были получены в фагмидном векторе рВК-СМВ.
Следующее исследование было выполнено по контракту через коммерческую лабораторию Lark Technologies, Houston, TX. Клоны удаляли из челночных векторов и встраивали в экспрессионные векторы следующим образом. Рекомбинантную плазмиду (рВК-CMV, содержащую вставку) расщепляли подходящими рестрикционными ферментами (рестриктазами), такими как Spel, Xbal, EcoRI или др., как было необходимо. Бакуловирусный транспортный вектор (pAcGHLT-A Baculovirus Transfer Vector, PharMingen, San Diego, Cat. No. 21460P) также разрезали подходящей рестриктазой (рестриктазами) в пределах или вблизи от сайта множественного клонирования для получения вставки, выделенной из челночного вектора.
Субклонируемый представляющий интерес фрагмент выделяли из продуктов расщепления при помощи электрофореза в низкоплавкой агарозе и очищали из геля с использованием набора для экстракции гелей Qiaquick согласно Lark SOP MB 020602. В случае необходимости вектор-реципиент обрабатывали щелочной фосфатазой в соответствии с Lark SOP MB 090201. Этот фрагмент лигировали в выбранный сайт вектора pAcGHLT-A. Эту рекомбинантную плазмиду трансформировали в клетки Е. coli XL1 Blue MRF' и трансформированные бактериальные клетки отбирали на чашках с LB-агаром, содержащих ампициллин (100 мкг/мл). Устойчивые к ампициллину колонии выскребали и выращивали на LB-бульоне, содержащем ампициллин, для получения плазмид.
Плазмидную ДНК получали с использованием процедуры щелочного минилизата (Lark SOP MB 010802) и расщепляли подходящей рестриктазой (рестриктазами). Подтверждали, что отобранные клоны имели правильный размер. Субклоны расщепляли другой подходящей рестриктазой (рестриктазами) для определения правильной ориентации вставки для подтверждения присутствия фрагментов правильного размера (размеров). Субклон выращивали в 100 мл LB-бульона, содержащего ампициллин (100 мкг/мл), и плазмидную ДНК получали с использованием набора для получения плазмид Qiagen Midi Plasmid Preparation (Lark SOP MB 011001). Концентрацию ДНК определяли измерением поглощения при 260 нм и расщеплением рестриктазами подтверждали, что эта проба ДНК происходит из правильного субклона.
Таким образом получали экспрессионные конструкции для Mey3, Mey5, Mey6, Mey8, имеющие теперь представляющую интерес кодирующую последовательность, слитую с геном GST с полигистидиновой меткой (тэгом), сайтом протеинкиназы А и сайтом расщепления тромбином. Ген GST и теперь данный слитый белок находятся под контролем промотора полиэдрина. PharMingen (San Diego, CA) встраивал этот вектор со вставкой в функциональные частицы бакуловируса путем ко-инсерции транспортного вектора (pAcGHLT-A) в восприимчивую клеточную линию S насекомых вместе с линеаризованной вирусной ДНК (PharMingen, San Diego, CA, BaculoGold viral DNA, Cat. No. 21100D). Эти функциональные вирусные частицы выращивали опять на клетках насекомых для получения исходного материала с высоким титром. Затем производили получение белка путем инфицирования крупномасштабной культуры клеток в Tini cell. Клетки собирали, когда выход белка достигал максимума и до того, как вирус убивал эти клетки. Слитые белки собирали на глутатион-агарозной колонке, промывали и высвобождали глутатионом.
Белки, собранные из этой аффинной колонки, определяли количественно измерением OD280 и анализировали на гелях с использованием электрофореза в ДСН-ПААГ и Вестерн-блоттинга с меченым антителом анти-GST (PharMingen, San Diego, CA, mAbGST Cat. No. 21441A) для подтверждения того, что все присутствующие полосы содержали GST-часть.
Четыре из десяти последовательностей клонировали, экспрессировали в инфицированных бакуловирусом клетках насекомых и затем очищали.
во вставке
(мг/мл)
Гели показали присутствие белка GST, кроме более крупных белков, которые были также положительными с антителом анти-GST в Вестерн-анализах. Mey3 воспроизводимо обнаруживал присутствие белков около 32 кДа, 56 кДа, полосы около 60-70 кДа и другие полосы, большие чем 70 кДа. Меу5 постоянно имел белки, мигрирующие как приблизительно 34 кДа, 38 кДа, 58 кДа, около 60-70 кДа и другие, большие чем 70 кДа. Mey6 имел белковые полосы около 34 кДа, 56 кДа, 58 кДа и полосы около 60-70 кДа. Меу8 имел белковые полосы около 36 кДа, 58 кДа и полосы около 60-70 кДа. Все указанные полосы были положительными в отношении GST. Эти полосы могут представлять целевой слитый белок или продукт деградации/расщепления, образующийся во время выращивания и очистки.
Пример 7: Анализ продуктов экспрессии на действие на активность расщепления TNF-R
Следующий способ использовали для измерения TRRE-активности Mey 3, 5, 6 и 8. Клетки C75R и клетки COS-1 высевали в 24-луночные культуральные планшеты при плотности 2,5×105 клеток/мл на лунку и инкубировали в течение ночи (в течение 12-16 часов) в 5% СО2 при 37°С. После отсасывания среды в лунке, 300 мкл 1 мкг Меу 3, 5 и 8 инкубировали в каждой лунке обоих планшетов C75R и COS-1 в течение 30 минут в 5% СО2 при 37°С (соответствующих А и С, упоминаемым ниже, соответственно). Одновременно клетки C75R в 24-луночных планшетах инкубировали также с 300 мкл свежей среды или буфера (что соответствует В, упоминаемому ниже). Супернатанты собирали, центрифугировали и затем анализировали на концентрацию растворимого TNF-R р75 при помощи ELISA, как описано в примере 1. Были получены следующие результаты:
Пример 8: Эффективность продуктов экспрессии в лечении септического шока
Протокол, описанный в примере 3, использовали для испытания действий продуктов экспрессии из новых клонов в предупреждении смертности в модели септического шока.
Различные количества рекомбинантных клонов Mey 3, 5 и 8 (10-100 мкг/мышь) инъецировали в/в в объеме 0,05 мл в пределах часа до или после инъекции летальной дозы ЛПС. Сыворотку (0,1 мл) собирали с использованием иглы 27 G и шприца на 1 мл из хвостовой вены при 30, 60 и 90 минутах после инъекции ЛПС. Эту сыворотку гепаринизировали и хранили в замороженном виде при -20°С. Пробы из многочисленных экспериментов анализировали при помощи ELISA на присутствие растворимого TNF-R, TNR-лиганда, IL-8 и IL-6. Животных подвергали мониторингу на протяжении следующих 12 часов на клинические эффекты шока. Выбранных животных эвтанизировали в периоды от 3 до 12 часов после обработки, проводили аутопсию и различные органы и ткани фиксировали в формалине, заливали в парафин, делали срезы и окрашивали гематоксилином-эозином (Г и Э). Срезы тканей подвергали гистопатологическому и иммунопатологическому исследованию.
Фигура 5 показывает полученные результаты. (◆) солевой раствор; ( ) БСА; () Mey-3 (100 мкг); (X) Mey-3 (10 мкг); (*) Mey-5 (10 мкг); () Mey-8 (10 мкг).
) БСА; () Mey-3 (100 мкг); (X) Mey-3 (10 мкг); (*) Mey-5 (10 мкг); () Mey-8 (10 мкг).
Мыши, инъецированные только ЛПС или ЛПС, контрольным буфером или контрольным белком (БСА), умирали быстро. Все животные в этой группе были мертвы при 24 часах. В противоположность этому, когда инъекции ЛПС сопровождались инъекциями 10-100 мкг Меу 3, 5 и 8, смерть задерживалась и коэффициенты смертности были более низкими. Ни одно из животных не было мертвым при 24 часах, когда они получали Mey 3 и Mey 5. Только 66% животных были мертвыми при 24 часах, когда они получали Mey 8. Таким образом, Mey 3, 5 и 8 были способны противодействовать смертности испытуемых животных, индуцируемой посредством ЛПС.
Список последовательностей приведен в конце описания.





































































| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА D И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799044C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2708032C2 |
| ЭРИТРОФЕРРОН И ERFE-ПОЛИПЕПТИДЫ И СПОСОБЫ РЕГУЛЯЦИИ МЕТАБОЛИЗМА ЖЕЛЕЗА | 2013 |
|
RU2684216C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ, ОБУСЛОВЛЕННОЙ РАКОМ КОСТИ, ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ | 2005 |
|
RU2389509C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ, ВЫЗВАННОЙ ОСТЕОАРТРИТОМ, ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ И СОДЕРЖАЩИХ ЕГО КОМПОЗИЦИЙ | 2006 |
|
RU2429013C2 |
| ВАРИАНТЫ ГЕМАГГЛЮТИНИНА И НЕЙРАМИНИДАЗЫ ГРИППА | 2010 |
|
RU2535970C2 |
| ХИМЕРНЫЕ МОЛЕКУЛЫ ФАКТОРА VII | 2010 |
|
RU2563231C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ В-КЛЕТОК | 2013 |
|
RU2650805C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ГУМАНИЗИРОВАННОГО АНТИ-CD19 ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА | 2014 |
|
RU2711975C2 |
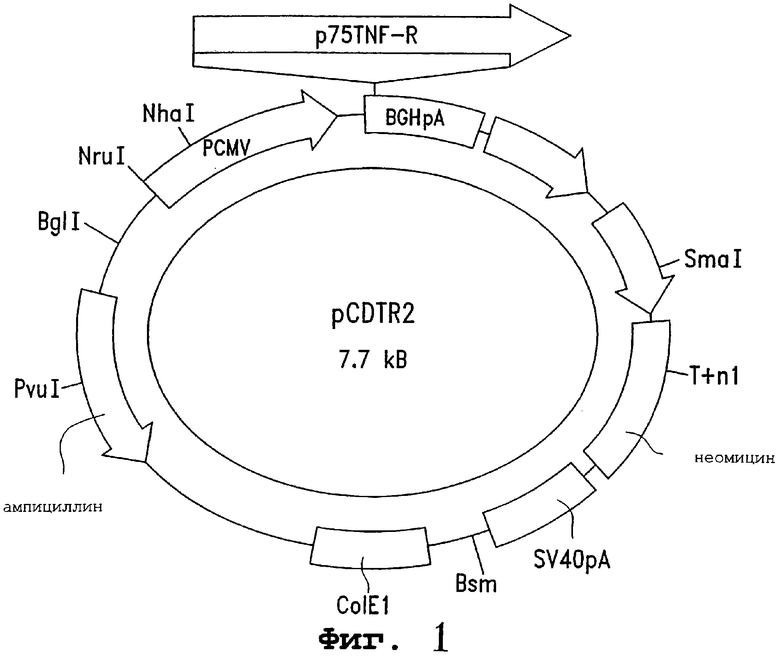
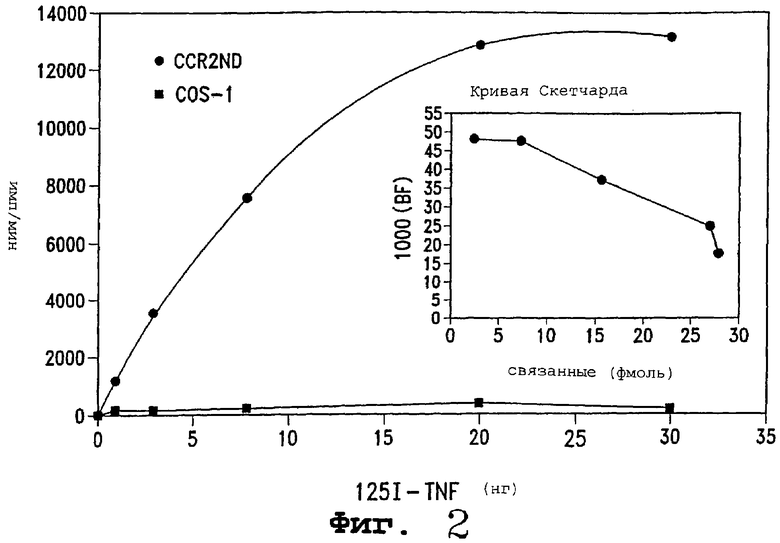
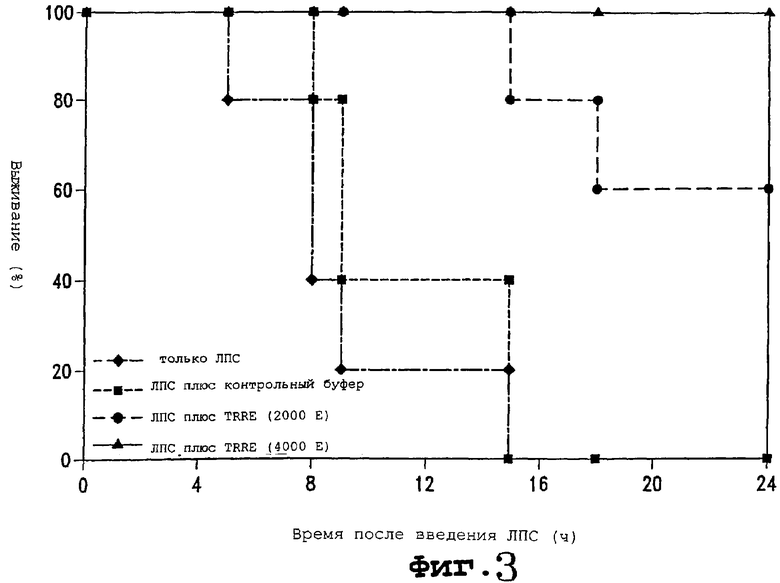
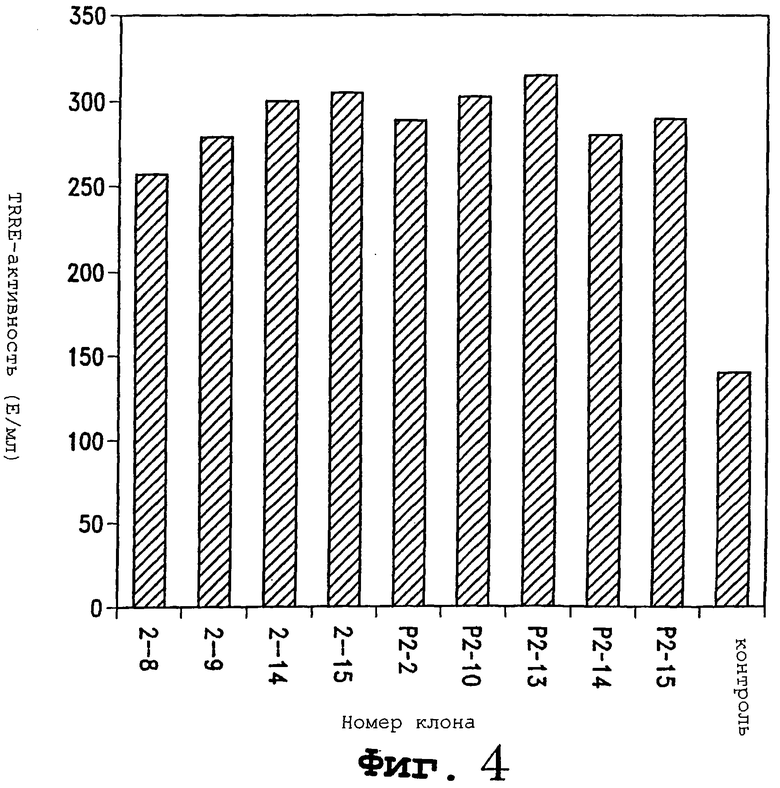
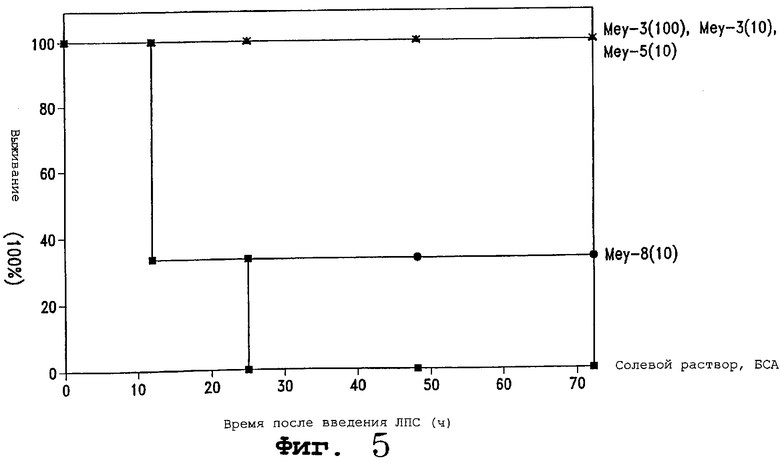
Изобретение относится к области биотехнологии. Предложены белки и полинуклеотиды, которые стимулируют ферментативное расщепление и высвобождение рецепторов TNF. Также предложены способы идентификации дополнительных соединений, которые влияют на выделение рецептора TNF из клеток. В качестве активного ингредиента в фармацевтической композиции продукты данного изобретения увеличивают или уменьшают трансдукцию сигнала TNF, ослабляя тем самым патологию заболевания. 16 с. и 11 з.п. ф-лы, 5 ил., 3 табл.
| GONZALEZ I | |||
| et al | |||
| ПРИБОР ДЛЯ ОПРЕДЕЛЕНИЯ ОСАДКИ ВАЛОВ ПАРОВЫХ ТУРБИН | 1917 |
|
SU283A1 |
| Proc | |||
| Nath | |||
| akad | |||
| sci | |||
| USA, 1985, v.82, р.7666-7670 | |||
| OSHIMA A | |||
| et al | |||
| The human cation-independent mannoze-E-phosphate receptor | |||
| J | |||
| biol | |||
| chem | |||
| Механическая топочная решетка с наклонными частью подвижными, частью неподвижными колосниковыми элементами | 1917 |
|
SU1988A1 |
| KATSURA K | |||
| et al | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Bioch | |||
| bioph | |||
| res | |||
| сом | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |

