Изобретение относится к области сельского хозяйства, в частности к селекции злаков, например яровой пшеницы, и может быть использовано в селекции этой и других культур на многоузловость корневой системы с интенсивным ростом и развитием растения, засухоустойчивость, скороспелость и другие признаки, контролируемые цитогенами.
Известен способ /6-7/ введения в культуру дикой лесной ржи S.silvestre методом гибридизации искусственно измененных ее и культурной ржи биотипов, обеспечивающих наследственные изменения плазмагенов дикой ржи, действие которых отмечено в ослаблении барьера нескрещиваемости при отдаленной гибридизации и различной выживаемости отдаленных гибридов. Наследственное изменение плазмона дикого вида вызвано мутагенным действием колхицина, которое вкупе с гибридизацией способствовало образованию рекомбинантных зигот, одна из которых дала взрослое полустерильное культурного типа растение. Недостатком способа является применение "митотического яда" колхицина, дающего как положительный, подобно вышеприведенному случаю, так и отрицательный эффект, в виде комплекса цитогенетических нарушений в потомстве, а также его дороговизна и канцерогенность.
Известен способ естественной индукции цитогенов морозостойкости гибридной популяции диплоидной ржи, затрагивающий узел кущения с развитыми вторичными побегами /9/. Способ трудоемкий, длителен во времени, требует специального провокационного фона.
Известен способ спонтанного наследственного изменения плазмона в потомстве генетически несбалансированных анеуплоидных растений аутотетраплоидной ржи, позволяющий выделять продуктивные адаптивные генотипы с новым уровнем ядерно-плазменного гомеостаза /8 /. Однако данный способ лимитирует формобразовательный процесс, требует точного цитологического анализа и оборудования для его выполнения, что не всем и всегда доступно.
Создание перспективных ядерно-плазменных трансгрессий возможно сочетанием эволюционно продвинутого плазмона аутоплоидов с реверсивным диплоидным набором хромосом /10/. Возникновение таких форм - редкое явление как в естественных условиях, так и при гибридизации, для их выделения по фенотипу необходимы определенные навыки.
Известен способ отбора высокожизнеспособных микро- и макрогамет в период гибридизации, обеспечивающий повышение озерненности аутополиплоидов в последующих поколениях, генетической их стабилизации и, соответственно, увеличения продуктивности и адаптивности /5/. Способ не обеспечивает получение измененных по цитоплазмону форм с признаками многоузловости корневой системы, интенсивным ростом и развитием растения.
Известен способ получения изменчивости митохондриона при сомаклональном митозном делении клеток каллусной ткани, индуцированной из незрелых зародышей кукурузы, и использование в селекции измененного таким образом сомаклонального генотипа /13/. Недостаток способа в том, что он не предусматривает получение высокопродуктивных, адаптивных ядерно-плазменных трансгрессий, а затрагивает только придание устойчивости к болезням источникам стерильности определенного типа цитоплазмы.
Известен способ получения отдаленных гибридов с целенаправленным подбором родительских компонентов, использованием культуры зародышей in vitro для преодоления нескрещиваемости и увеличения выхода гибридов с отбором селекционно ценных генотипов в поздних поколениях /Н.В.Цицин. - Теория и практика отдаленной гибридизации. - М.: Наука, 1981. - 160 с. - прототип/.
Недостатком прототипа является то, что первым поколениям гибрида создаются "комфортные" условия с целью максимального их сохранения и размножения, в отбор подпадают лучшие из них. При этом не рассматривается получение цитоплазматической изменчивости и ее использование в конечном итоге при выделении ядерно-плазменных трансгрессий.
В естественных условиях действие стресса по засухе накануне активного деления тотипотентных клеток узла кущения трудно закрепить в потомстве. В большинстве случаев жесткая комплексная весенне-летняя засуха вызывает ускоренное подсыхание верхнего слоя почвы и совпадает по времени с этапом интенсивного кущения яровых и нераскустившихся с осени озимых злаковых растений, обуславливая тем самым плохое или отсутствие вообще образование вторичных узловых побегов. К тому же, повышенная температура воздуха, длинный день и интенсивная солнечная инсоляция, способствующие яровизации яровых культур /4/, до минимума сокращают продолжительность второго этапа и ускоряют переход растений к третьему этапу органогенеза.
Снятие свойственного растениям апикального доминирования, когда точка роста главного побега своим гормональным влиянием сдерживает развитие боковых почек, особенно спящих, возможно при гибели главного побега, что стимулирует пробуждение меристем спящих почек к усилению побегообразования в узлах кущения /1-2/. Однако гибель главного колоса не желательна, так как он служит индикатором селекционной браковки растений.
Известен способ создания стрессовой провокации по дефициту влаги в растении, заключающийся во временном отсутствии защитной функции у пробирочных растений злаков не образующих восковой налет in vitro, что приводит к подсыханию растений при пересадке в грунт /14/.
При разработке НИР нами обнаружено новое свойство известного признака, а именно, подсыхание главного побега у культивируемых и яровизированных in vitro без образования защитного воскового налета растений злаков при пересадке в грунт в фазе трех листьев стимулирует ускоренную и интенсивную регенерацию вторичных узловых побегов с сохранением главного колоса.
Сущность нового технического решения заключается в том, что отбор хозяйственно ценных генотипов из потомств тотипотентных клеток узлов кущения злаковых растений, измененных по контролирующему многоузловость, интенсивность роста и развития растения плазмону, ведут в испытавшим действие стресса на момент начала митозного деления клеток меристем узловых побегов растениях-регенерантах отдаленных гибридов, яровизированных in vitro и высаженных в грунт в фазе трех листьев, с полу- или стерильным главным колосом.
Предложенный способ позволяет получать технический результат, заключающийся в выделении высокопродуктивных адаптивных форм с признаками многоузловости корневой системы и интенсивным ее ростом и развитием в потомстве вторичных побегов полу- или стерильных по главному колосу растений-регенерантов с измененным цитоплазмоном.
Сопоставительный анализ заявляемого технического решения с прототипом показал, что заявляемый способ отличается от известного тем, что создание засухоустойчивого высокопродуктивного сорта с многоузловой, интенсивно растущей и развивающейся корневой системой, контролируемой цитогенами, осуществляют путем отбора генотипов в потомстве вторичных узловых побегов полу- или стерильных по главному колосу пробирочных отдаленных гибридов-регенерантов RF2 яровизированных in vitro и высаженных в грунт в фазе 3 листьев в условиях дефицита влаги.
Изучение патентной и научно-технической литературы свидетельствует о том, что отбор генотипов с многоузловой, интенсивно растущей и развивающейся корневой системой, детерминированных плазмагенами, в потомстве испытавших действие стресса узловых побегов растений-регенерантов RF2 с максимальной разбалансированностью ядерно-плазменного гомеостаза, яровизированных in vitro и пересаженных в грунт в фазе трех листьев, дающих в дальнейшем полу- или стерильный главный колос, является новым и в селекционной работе со злаками не применялся.
Предложенный способ предусматривает использование известных сведений. Генетическая изменчивость плазмона на фоне максимальной ядерно-плазменной разбалансированности генома возникает под действием естественных стрессовых факторов /8/ или при митозном делении каллусных клеток in vitro /13/. Ускорение роста и развития вторичных побегов достигается стимуляцией к их пробуждению под давлением стресса у яровизированных злаков при переходе от второго к третьему этапам органогенеза /2/.
Межвидовые, межродовые и другие разнохромосомные скрещивания дают высокую разбалансированность генома. Достоинство таких скрещиваний в том, что, используя лучшие по продуктивности и комплексу признаков районированные сорта, можно свести к минимуму формообразование в ядерном геноме и работать с однотипными или нейтральными по фотопериоду генами, но усилить формообразование в стабилизации более совершенного уровня ядерно-плазменного гомеостаза растений.
На примере межвидовых гибридов от скрещивания районированных сортов яровой твердой пшеницы Светлана и озимой мягкой пшеницы Тарасовская 29 отмечена низкая озерненность колоса в F1 и падение ее по причине возрастающей несбалансированности генома во втором поколении /табл. 1/.
Данный факт объясняется повышением концентрации генотипов с ядерно-плазменной разбалансированностью генома соматических клеток растений F2), поскольку в соматических клетках первого поколения свойственная исходному цитогеному материнская часть ядерного генома остается неизменной и уровень снижения озерненности обусловлен только цитогенетической разбалансированностью зиготы и тканей эндосперма. Выявленная ядерно-плазменная разбалансированность соматических клеток растений F2 простых скрещиваний линейных сортов аналогична F1 тройных скрещиваний и именно при митозе в таких клетках имеется предпосылка к активизации формообразования в стабилизации нового уровня ядерно-плазменного гомеостаза растений.
Культура незрелых зародышей гибридов in vitro /3/ позволяет сохранять рекомбинанты и расширить формообразование плазмагенов за счет выживания продуктивных вторичных узловых побегов.
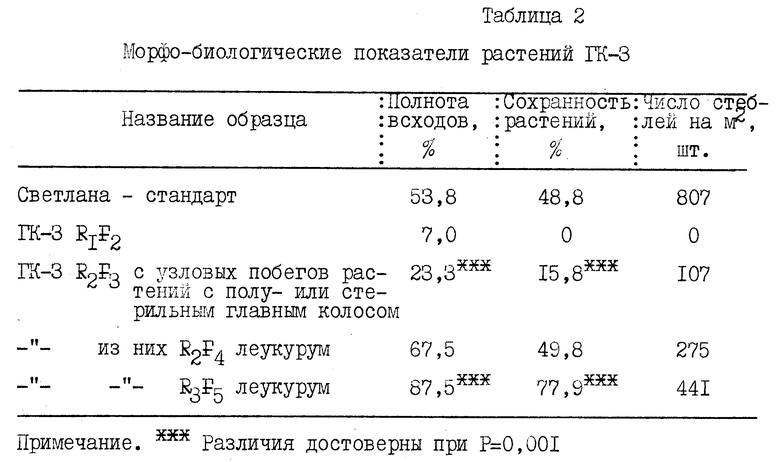
В засушливых условиях при посеве зерновок R1F2 ГК-3 /Светлана х Тарасовская 29/ в почву получена низкая полнота всходов и гибель растений к уборке /табл. 2/. При высадке яровизированных in vitro растений R1F1 в фазе трех листьев из 100 гибридных зерен получено 28 растений со средней продуктивной кустистостью 3,78 побега и общим сбором зерен 690 штук, из них в R2F2 получено 158 растений со средней продуктивной кустистостью 4,6 побега и сбором 1317 зерен.
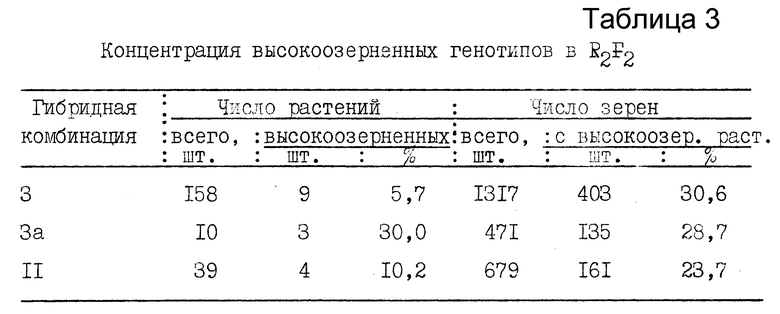
В данном случае на фоне максимальной ядерно-плазменной разбалансированности соматических клеток рекомбинанты F2 отдаленных гибридов имеют высокую стерильность главного колоса. При этом растения с нормально озерненным главным колосом, как правило, гено- и фенотипически близки к материнской форме и имеют малую селекционную ценность. Концентрация таких растений в популяциях R2F2 отдаленных гибридов составила 5,7-30,0%, а по числу зерен - 23,7-30,6% /табл. 3/, что в последующих поколениях значительно обременяет популяцию неперспективными генотипами и затрудняет отбор действительно ценных ядерно-плазменных трансгрессий.
В потомстве вторичных побегов регенерантов отдаленных гибридов, отличающихся низкой озерненностью главного колоса, резко повышается концентрация селекционно ценных генотипов. Так, популяция R3F5 ГК-3, сформированная по разновидности материнской формы, скороспелости и крупности зерна, значительно превосходила исходную форму по полноте всходов, сохранности растений к уборке, густоте продуктивного стеблестоя /табл. 2/.
Сорт яровой твердой пшеницы Степь 3, сформированный из смеси 400 генетически стабильных линий R2-5F4-9 ГК-3 - потомств подгонов растений-регенерантов R2F2 с полу- или стерильным главным колосом, характеризовался синхронным колошением с не менее двух продуктивных побегов на растение в разреженном посеве. В загущенном посеве Степь 3 превосходила стандарт - "материнский" сорт Светлану по числу растений с 3-5-ю узлами кущения, сохранности растений к уборке, густоте продуктивного стеблестоя и продуктивности, при равной стандарту крупности зерна /табл. 4/.
Таким образом, стимуляция ускоренной регенерации растений FR2, прояровизированных in vitro и высаженных в грунт накануне кущения, способствует образованию более жизнеспособных, генетически сбалансированных, устойчивых к засухе, скороспелых генотипов с многоузловой корневой системой.
Преимущество сорта Степь 3 по многоузловой, быстро растущей и развивающейся корневой системе сопряжено с такими хозяйственно ценными признаками как эффективное использование осенне-зимних и весенних запасов влаги в почве, опережающий стартовый рост, способность противостоять сорной растительности, скороспелость, засухоустойчивость, высокое качество зерна. Степь 3 достоверно превысила по урожайности: 1/ сорт-стандарт Светлану; 2/линию, выделенную из Светланы методом каллусогенеза незрелых зародышей /12/, имеющую урожайность на уровне стандарта; 3/ отбор R2F4-6 ГК-3 из потомств главного колоса полустерильных растений R2F2
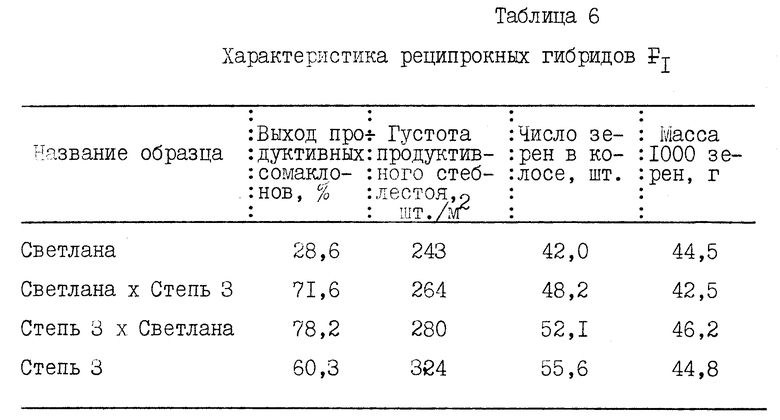
Анализ растений F1 реципрокных гибридов Светлана х Степь 3 показал наличие материнского эффекта по сопряженным с многоузловостью и интенсивностью роста и развития корней показателям: приживаемость каллусных сомаклональных линий в почве, выход продуктивных сомаклональных линий, густота продуктивного стеблестоя, число зерен в колосе, крупность зерна с лучшим проявлением признаков у цитогенома Степь 3 /табл. 6/.
Следовательно, сомаклональная изменчивость сорта как генетически сбалансированной системы затруднена. Генетическая изменчивость, направленная на регулирование в онтогенезе растений ядерно-плазменных отношений с новым, более высоким уровнем гомеостаза не закрепляется в потомствах полустерильного главного колоса растений-регенерантов отдаленных гибридов.
Исходя из чего, генетическая изменчивость регенерирующих соматических клеток узлов кущения, в том числе изменчивость плазмона /табл. 6/, достигается на фоне ядерно-плазменной разбалансированности соматических рекомбинантных клеток под действием стресса при активном митотическом делении узловых тотипотентных клеток и закрепляется в потомстве вторичных узловых побегов злаковых растений с полу- или стерильным главным колосом RF2 отдаленных гибридов, яровизированных in vitro и высаженных в грунт в фазе трех листьев.
На основании анализа известных решений делаем вывод, что наше техническое решение является новым и неочевидным. При этом получен положительный эффект - предложенный способ позволяет создавать сорта с многоузловой корневой системой ускоренного роста и развития.
Сущность предлагаемого изобретения может быть понята из следующего описания:
- подбор родительских компонентов и отдаленная гибридизация злаков;
- культура in vitro незрелых гибридных зародышей по общепринятым методикам;
- яровизация до кущения in vitro яровых злаков при длинном дне и интенсивном освещении, озимых злаков при пониженной температуре;
- пересадка в грунт растений-регенерантов RF2 отдаленных гибридов в фазе трех листьев;
- стимуляция активного кущения растений-регенерантов, не образовавших восковой налет in vitro на листьях главного побега действием стресса по дефициту влаги в конце II и начале III этапов органогенеза;
- выращивание растений-регенерантов с повышенной продуктивной кустистостью в естественных условиях или при имитации среднего уровня дефицита влаги;
- браковка растений-регенерантов с хорошо озерненным главным колосом;
- отбор для пересева и дальнейшей селекционной проработки потомств вторичных узловых побегов рекомбинантных растений-регенерантов с разбалансированным по ядерно-плазменному гомеостазу полу- или стерильным главным колосом.
Пример конкретного исполнения
1. Подбирают родительские компоненты для скрещивания и получают отдаленные гибриды злаков.
2. Культивируют in vitro незрелые гибридные зародыши.
3. Проводят яровизацию in vitro яровых растений на длинном дне и интенсивном освещении, озимых растений при пониженной температуре.
4. Выращивают растения-регенеранты RF2 отдаленных гибридов путем пересадки их в грунт в условиях дефицита влаги в фазе трех листьев до получения продуктивной кустистости и сохранения главного колоса.
5. Анализируют уровень фертильности главного колоса гибридных растений-регенерантов.
6. Отбирают для дальнейшей селекционной проработки гаметы или зерновки со вторичных узловых побегов рекомбинантных растений-регенерантов с полу- или стерильным главным колосом.
Список использованной литературы
1. Велибекова Б. И., Велибеков М.Д., Бондарева Л.Н. Способ определения устойчивости яровых зерновых культур к повреждению шведской мухой //Патент N 2025063 A 01. М 1/00. - 1994. - Бюлл. N 24.
2. Кумаков В.А. Физиология яровой пшеницы. - М.: Колос, 1980. - 200 с.
3. Лукьянюк С. Ф. , Игнатова С.А. Методика культуры тканей и органов в селекции растений. - Одесса, 1983. - 21 с.
4. Разумов В. И. Среда и развитие растений. - Л.: Сельхозизд., 1961. - 367 с.
5. Титаренко А.В., Титаренко Л.П. Способ селекции аутотетраплоидных популяций // А.с. N 1777723 A 01 H 1/04. - 1992. - Бюлл. 44.
6. Титаренко А.В., Титаренко Л.П. Использование Secale silvestre Host. в селекции ржи //VI съезд Белорусского общества генетиков и селекционеров /Тез. докл. - Горки, 1992. - c. 74.
7. Титаренко. А. В. , Титаренко Л.П. Использование Secale silvestre в селекции самоопыленных линий озимой ржи //Изогенные линии и генетические коллекции /Материалы II совещания. - Новосибирск, 1993. - С. 39-40.
8. Титаренко А.В., Титаренко Л.П. Анизоплоидные скрещивания как способ селекции растений //Аграрная наука производству /Тез. докл. - Безенчук, 1993. - С. 59-60.
9. Титаренко А. В., Титаренко Л.П. Индукция морозостойкости цитогенома ржи //Экологические проблемы интродукции растений на современном этапе: вопросы теории и практики / Материалы Междунар. науч. конф.- Краснодар, 1993. - Ч. 2. - С. 667-668.
10. Титаренко А. В. , Титаренко Л.П. Пути активизации формообразования образования в селекции на продуктивность, адаптивность и генетическую стабильность //Защита растений в условиях формирования агропромышленного комплекса: экономика, эффективность, экологичность /Тез. докл. - С.-Петербург, 1995. - С. 255-256.
11. Цицин Н.В. Теория и практика отдаленной гибридизации. - М.: Наука, 1981. - 160 с.
12. Шаяхметов И. Ф., Шитирякова Ф.А., Хабирова М.М. Особенности каллусообразования и регенерации растений в культуре незрелых зародышей яровой твердой пшеницы //С.-х. биология. - 1988. - N 4. - С. 65-67.
13. Kemble R.J., Flavell R.B., Brettell R.Z.S. Mitochondrial DNA analises of fertile and sterile plants derived from tissue culture with the Texas male strile cytoplasm// Theor. and Appl. Genet. - 1982. - V. 62. - N 3. - P. 213-217.
14. Selvapandigan A., Subramani J., Bhatt P.N., Mehta A.R. A simple method for direct transplantation of cultured plants to the field// plant Se. - 1988. V. 56. - N 1. - P. 81-83.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОТБОРА МОРОЗОСТОЙКИХ ФОРМ АЛЛОПЛАЗМАТИЧЕСКОЙ РЖИ | 2000 |
|
RU2192124C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГРЕССИВНЫХ РАСТЕНИЙ ОЗИМОЙ РЖИ | 2000 |
|
RU2197815C2 |
| СПОСОБ СЕЛЕКЦИИ ОЗИМОЙ РЖИ | 1992 |
|
RU2080054C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ УСТОЙЧИВОСТИ ЯРОВЫХ ЗЕРНОВЫХ КУЛЬТУР К ПОВРЕЖДЕНИЮ ШВЕДСКОЙ МУХОЙ | 1992 |
|
RU2025063C1 |
| Способ получения аналогов тетраплоидной ржи | 1991 |
|
SU1807842A3 |
| СПОСОБ ОТБОРА ХОЗЯЙСТВЕННО-ЦЕННЫХ ФОРМ ЗЕРНОВЫХ КУЛЬТУР | 1992 |
|
RU2068635C1 |
| СПОСОБ ЗАРАЖЕНИЯ ПРОСА ГОЛОВНЕЙ | 1993 |
|
RU2090054C1 |
| Способ определения эффективности полиплоидизации злаков | 1990 |
|
SU1708209A1 |
| СПОСОБ ВОЗДЕЛЫВАНИЯ ПОДСОЛНЕЧНИКА В СЕВООБОРОТЕ | 1991 |
|
RU2012180C1 |
| СПОСОБ ВЫРАЩИВАНИЯ ДРЕВЕСНЫХ НАСАЖДЕНИЙ | 1993 |
|
RU2086106C1 |



Изобретение относится к сельскому хозяйству, в частности к селекции растений на продуктивность и адаптивность, может быть использовано в селекции яровой твердой пшеницы и других злаковых культур на многоузловость корневой системы, скорость, засухоустойчивость и другие признаки, контролируемые цитогенами. Селекция продуктивного сорта яровой твердой пшеницы с признаками многоузловости корневой системы с интенсивным ростом и развитием, детерминированного плазмагенами, достигается отбором генотипов в потомствах вторичных побегов полу- или стерильных по главному колосу растений-регенерантов RF2, яровизированных in vitro и высаженных в фазе трех листьев в грунт в условиях дефицита влаги. Данный способ позволяет получить технический результат, заключающийся в выделении высокопродуктивных адаптивных форм с признаками многоузловости корневой системы и интенсивным ее ростом. 6 табл.
Способ селекции растений злаковых культур, включающий подбор родительских пар, отдаленную гибридизацию, культуру зародышей in vitro, регенерацию растений, отбор селекционно ценных генотипов, отличающийся тем, что растения-регенеранты RF2 яровизируют in vitro и в фазе трех листьев высаживают в грунт в условиях дефицита влаги, а в качестве селекционно ценных генотипов отбирают потомства вторичных побегов полу- или стерильных по главному колосу растений.
| Цицин Н.В | |||
| Теория и практика отдаленной гибридизации | |||
| - М.: Наука, 1981, 160 с | |||
| Способ селекции аутотетраплоидных популяций ржи | 1991 |
|
SU1777723A1 |
| Титаренко А.В | |||
| Титаренко Л.П | |||
| Анизоплоидные скрещивания как способ селекции растений | |||
| Аграрная наука производству | |||
| Тез | |||
| докл | |||
| Безенчук, 1993, с.59 - 60. | |||
