Изобретение относится к способам, используемым в сельском хозяйстве.
Общеизвестен естественный способ сохранения гетерозиса в ряду последующих поколений посредством вегетативного размножения у растений, дополнительно обладающих половым размножением. Многочисленные исследования в этой области на других растениях, не способных к вегетативному размножению, и животных не увенчались полным успехом (1), потому что природа гетерозиса все еще оставалась большой загадкой генетики (2). В литературе не было высказано даже сколько-нибудь реальных теоретических подходов к кардинальному решению этой важной проблемы.
У некоторых животных гетерозис можно закрепить клонированием. Однако этим способом пока получают единицы идентичных матери потомков. У тутового шелкопряда клонирование разработано более успешно, но для практического использования в плане сохранения гетерозиса оно не приемлемо по двум причинам: из-за большой трудоемкости массового получения партеногенетического потомства и меньшей, по сравнению с самцами, продуктивности женского пола, из которого состоят клоны (3).
Перспективные результаты получены авторами после того, как они разработали метод мейотического партеногенеза и получили от партеногенетических клонов абсолютно гомозиготных самцов тутового шелкопряда (4). Их возвратное скрещивание с партеногенетическим клоном гибридного происхождения позволило закреплять гетерозис в беккроссных поколениях (5). Но это было открытием лишь принципиальной возможности закрепления гетерозиса. Практического же значения этот метод не имел и, следовательно, не мог быть запатентован в качестве способа. Это объяснялось тем, что гомозиготных самцов могли получать только от высокожизнеспособных женских партеноклонов с высокой склонностью к партеногенезу. У коммерческих пород и гибридов абсолютных гомозиготов практически получать не удавалось, поэтому мейотический партеногенез был использован только для рекогносцировочных опытов, направленных на выяснение возможности решения проблемы. Изобретение способа закрепления гетерозиса у тутового шелкопряда, пригодного для производства, стало возможным после открытия авторами односпермичного андрогенеза (1998, неопубликовано).
Сущность изобретения. Гетерозис проявляется только в первом поколении гибрида. В последующих поколениях, начиная со второго, он резко затухает. Поэтому, чтобы вырастить гетерозисный гибрид приходится каждый раз вновь повторять межсортовую или межпородную гибридизацию. Этот процесс технически сложный и весьма трудоемкий, а применительно ко многим растительным культурам он просто невыполним, хотя их гибриды, если бы они были получены, дали бы удивительно высокие урожаи по сравнению с родительскими формами. Примером этого служат многие сельскохозяйственные растения.
Эти проблемы были бы кардинально решены, если бы удалось разработать эффективный способ закрепления гетерозиса в последующих поколениях. Такой способ одновременно открыл бы совершенно новый подход к созданию еще более выдающихся по гетерозису гибридов. Известно, что любой промышленный гибрид получают посредством скрещивания огромной массы индивидуумов двух родительских форм. А эти индивидуумы весьма дифференцированы по комбинационной способности. Поэтому производство довольствуется средним гетерозисом по всем вместе взятым индивидуальным гибридам, каждый из которых происходит от половых клеток двух родителей. В то время как редкие индивидуальные гибриды обладают поистине фантастическим гетерозисом, в следующем поколении он безвозвратно теряется. Предлагаемый способ позволит закреплять этот мощный гетерозис в последующих поколениях гибрида и размножать его в неограниченных количествах.
Одной из причин гетерозиса считали благоприятное действие на развитие и жизнедеятельность организма гетерозиготности вообще всех генов, независимо от их специфичности (гипотеза "сверхдоминирования"). Авторы на тутовом шелкопряде экспериментально доказали, что гетерозис возникает в результате двух главных причин. Первая - интеграция в генотипе гибридов большого количества скоординированных в своем действии благоприятных генов, контролирующих жизнеспособность. Вторая - переход в гетерозиготное состояние не всех генов генотипа, а только рецессивных деталей, полулеталей и субвиталей (4). На фиг. 1 приведены доказательства этого.
Следовательно, снижение гетерозиса в последующих поколениях гибридов в основном объясняется неизбежным при скрещивании гибрида в его пределах переходом части рецессивных деталей и полулеталей в гомозиготное состояние и нарушением в процессе мейоза комплекса благоприятных генов, повышающих жизнеспособность. Поэтому авторы пришли к выводу, что закрепить гетерозис в последующих поколениях можно, если в генотипе гибрида полностью сохранить или даже улучшить комплекс всех благоприятных генов и практически полностью удалить из генотипа рецессивные летали и полулетали. Эта задача решена авторами следующим способом.
В качестве исходного материала выбирают генетически отдаленные две породы, от скрещивания которых возникают наиболее высокогетерозисные гибриды. От этих двух пород получают серию индивидуальных гибридов, каждый из которых происходит только от двух родителей. Путем сравнительных испытаний выбирают 10 лучших по гетерозису индивидуальных гибридов. От каждого гибрида получают абсолютно гомозиготных потомков методом односпермического андрогенеза, выполнение которого доступно селекционерам. Для этого неосемененных самок любой породы облучают γ- лучами в дозе 80 кр. Затем самки спариваются с самцами индивидуальных гибридов. Отложенные яйца в возрасте 60-80 минут после откладки при температуре 25oC прогревают 210 минут в воде, нагретой до 38oC.
Абсолютные гомозиготы в подавляющем большинстве погибают на разных стадиях развития из-за того, что в гаплоидном генотипе, доставшимся им от отца, содержится много летальных, полулетальных и субвитальных генов. При диплоидизации ядра пронуклеуса они переходят в гомозиготное состояние, чаще всего несовместимое с нормальным развитием организма. Выживают только те гомозиготы, которым в ходе мейоза не досталось или досталось, но очень мало, вредных генов, в основном слабого действия (5).
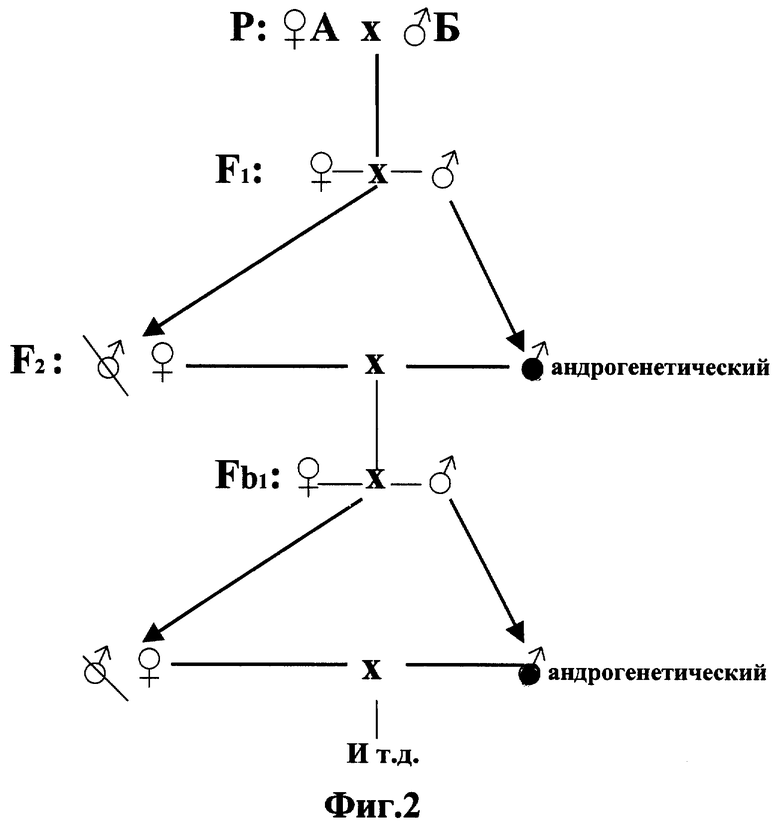
Выращенных абсолютно гомозиготных особей возвратно (беккросс) скрещивают с исходным гибридом, получая, таким образом, первое беккроссное поколение (фиг. 2). Созревание исходного гибрида и абсолютных гомозиготов должно быть синхронизировано путем задержки начала выращивания первого на время, равное продолжительности цикла развития взятого объекта.
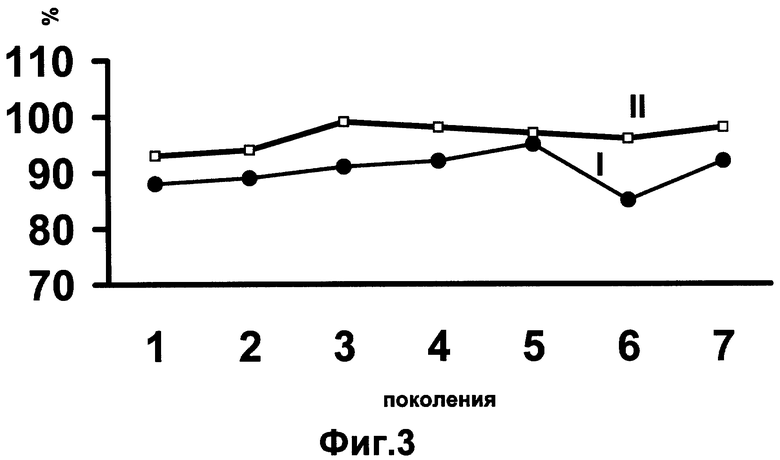
Простые расчеты показывают, что в беккроссном потомстве новые гомозиготы сильных по вредности генов не могут появиться, а гомозиготы субвитальных генов, если они не были элиминированны у выживших гомозиготных андрогенов, подавляются комплексом благоприятных генов, доставшихся от исходного гибрида. Вот поэтому гетерозис сохраняется во всех беккроссных поколениях (фиг. 3).
С первыми и последующими беккроссными поколениями поступают точно так же, как и с исходным гибридом (фиг. 2). Дальнейшие беккроссы приводят, во-первых, к почти полному удалению деталей и полулеталей из генотипа гибрида и, во-вторых, к сохранению той численно преобладающей части генов, которые обеспечили гетерозис в исходном гибриде.
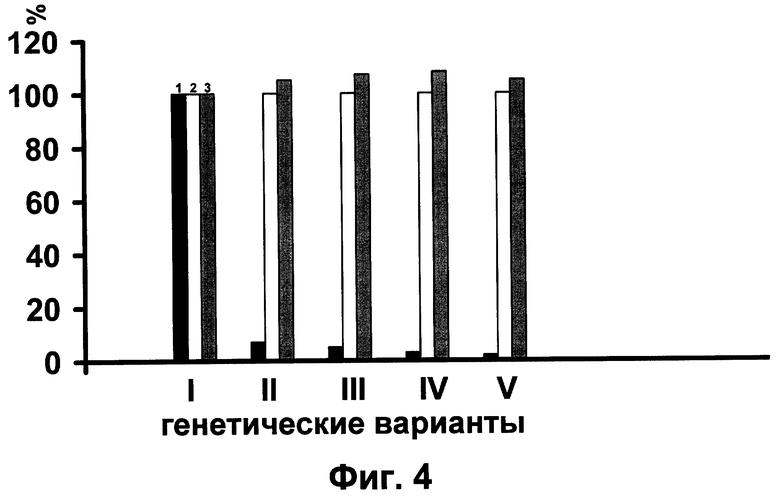
После 5 или 6 беккроссов очищенный от вредных генов гибрид массово размножают путем внутригибридного скрещивания. У потомства, полученного в результате такого размножения, гетерозис не только сохраняется на уровне исходного гибрида, но и даже несколько усиливается (фиг. 4), что свидетельствует о полном решении проблемы закрепления гетерозиса у тутового шелкопряда.
Полная общность генетических основ гетерозиса и его затухания у животных и растений позволяет данное изобретение рекомендовать для закрепления гетерозиса у сельскохозяйственных растений, у которых возможно получение от гибридов абсолютно гомозиготных особей андрогенного происхождения. Их получают путем стимулирования эмбрионального развития гаплоидной пыльцы с последующим превращением ее зародышевых клеток в диплоидные, развивающиеся в жизнеспособные фертильные растения. Методика варьирует в зависимости от биологических особенностей культуры.
Графические материалы.
Фиг. 1 А. Показана прямая зависимость между урожаем коконов тутового шелкопряда - основной показатель гетерозиса (1) и уровнями гетерозиготности (2) генетических вариантов гибрида неочищенных от леталей и полулеталей. Показатели урожая и гетерозиготности исходного гибрида первого варианта (1) приняты за 100%.
Б. Показано полное отсутствие зависимости между урожаем коконов (1) и уровнями гетерозиготности (2) у генетических вариантов, очищенных от леталей и полулеталей. Это доказывает несостоятельность гипотезы гетерозиса "сверхдоминирования" и возможность сохранения гетерозиса в беккроссных поколениях.
Фиг. 2. Схема очищения гибридов тутового шелкопряда от рецессивных леталей и полулеталей посредством возвратных скрещиваний гибридов с полученными от них абсолютно гомозиготными самцами А и Б породы. F1, F2 - гибрид первого и второго поколения. Fb1, Fb2 - первое и второе беккроссное поколение.
Фиг. 3. Жизнеспособность исходного гибрида (1) и беккросных поколений (II), полученных по схеме, представленной на фиг. 2.
Фиг. 4. Демонстрирует показатели частоты вредных генов в гетерозиготном состоянии (1), массы кокона (2), жизнеспособности (3) у исходного гибрида (I) и трансформируемого гибрида после четырех последовательных возвратных скрещиваний с гомозиготными самцами (II), а также у трех последовательных инбредных поколений (III-V). Выкормка каждого генетического варианта проводилась одновременно с контрольным партеногенетическим гибридом, показатели которого принимались за 100%. Во всех генетических вариантах гетерозис выше, чем у исходного гибрида, что свидетельствует о кардинальном решении проблемы закрепления гетерозиса.
Стабильное сохранение гетерозиса во всех беккроссных поколениях уже свидетельствовало о принципиальной эффективности разрабатываемого способа. Но беккроссные поколения не применимы в практике из-за сложности их получения. Поэтому в заключительном эксперименте на тутовом шелкопряде изучали возможность закрепления гетерозиса уже не в беккроссных, а в нормальных поколениях.
В этом заключительном опыте исходный гибрид вначале подвергался четырем беккроссам с гомозигонтыми самцами. В результате частота гетерозигот по леталям и полулеталям снизилась до 6,2% со 100% в исходном материале. Далее беккроссные поколения размножали инбридингом. Каждое инбредное поколение получали посредствам скрещивания брата с сестрой в пределах каждой отдельно взятой семьи. В результате частота вредных генов, погашенных нормальными аллелями, снизилась в первом инбредном поколении до 4,7, а во втором и третьем - до 3,5 и 2,6% соответственно. Инбредное размножение исключительно пагубно влияет на все хозяйственные показатели нормального инбредного потомства. Но в нашем опыте оно не только не оказало угнетающего действия на инбредное потомство, а, наоборот, привело к повышению у него средней массы одного кокона и жизнеспособности по сравнению с исходным, контрольным гибридом (фиг. 4). Следовательно, проблема закрепления гетерозиса у гибридов последующих поколений кардинально решена.
БИБЛИОГРАФИЧЕСКИЕ ДАННЫЕ
1. Инге-Вечтомов С. И. 1989. Генетика с основами селекции. М. "Высшая школа", на стр. 557.
2. Хатт Ф. 1969. Генетика животных. Пер. с англ. под ред. д-ра биол. наук Я.Л. Глембоцкого. М., "Колос", на стр. 322.
3. Струнников В. А. 1998. Клонирование животных: теория и практика. - Природа, N 7, с.3 -9.
4. Струнников В.А. 1987. Генетические методы селекции и регуляции пола тутового шелкопряда. М. ВО "Агропромиздат", на стр. 35.
5. Струнников В.А. 1994. Природа гетерозиса и новые методы его повышения. - М. Наука, 108 с.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ВЫСОКОПРОДУКТИВНЫХ РАСТЕНИЙ- САМООПЫЛИТЕЛЕЙ НА ОСНОВЕ ЭФФЕКТА ЗАКРЕПЛЕННОГО ГЕТЕРОЗИСА | 2003 |
|
RU2254709C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИЗОГЕННОЙ ЛИНИИ ПО РЕЦЕССИВНОМУ ПРИЗНАКУ СЕМЕНИ У РАСТЕНИЙ | 1999 |
|
RU2151495C1 |
| СПОСОБ ЗАКРЕПЛЕНИЯ ГЕТЕРОЗИСА ГИБРИДОВ В ПОСЛЕДУЮЩИХ ПОКОЛЕНИЯХ | 2011 |
|
RU2465771C1 |
| СПОСОБ ЗАКРЕПЛЕНИЯ ГЕТЕРОЗИСА ГИБРИДОВ | 2019 |
|
RU2759222C2 |
| Способ создания гомозиготных гомостильных линий у гречихи | 1990 |
|
SU1738167A1 |
| Способ создания мужски-стерильных F1-гибридов лука репчатого, устойчивых к заболеваниям | 2023 |
|
RU2834769C1 |
| СПОСОБ СОЗДАНИЯ ИЗОГЕННОЙ ЛИНИИ ПО ДОМИНАНТНОМУ ПРИЗНАКУ СЕМЕНИ У РАСТЕНИЙ | 2005 |
|
RU2290785C1 |
| СПОСОБ СЕЛЕКЦИИ ГАПЛОИДОВ И ДВОЙНЫХ ГАПЛОИДОВ РАСТЕНИЙ | 1994 |
|
RU2158502C2 |
| КУКУРУЗНЫЕ ПРОДУКТЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2012 |
|
RU2650764C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИЗОГЕННОЙ ЛИНИИ ПО ДОМИНАНТНОМУ ПРИЗНАКУ СЕМЕНИ У РАСТЕНИЙ | 1999 |
|
RU2151494C1 |
Способ предназначен для использования в сельском хозяйстве. Цель - значительное повышение эффективности гибридизации сельскохозяйственных культур. Гетерозис гибридов проявляется только в первом поколении. Установлено, что затухание гетерозиса в последующих поколениях гибрида в основном происходит из-за перехода рецессивных леталей, полулеталей и субвиталей в гомозиготное состояние и нарушения комплекса благоприятных скоординировано действующих генов. Устранение этих явлений приводит к закреплению гетерозиса в последующих поколениях. Оно выполняется посредством возвратных скрещиваний гибрида с искусственно полученными от него абсолютно гомозиготными андрогенетическими сыновьями, после чего генетически трансформируемый гибрид практически полностью очищается от вредных генов и одновременно сохраняет в целости комплекс благоприятных генов, определяющих гетерозис. Это позволяет полностью сохранить гетерозис в последующих уже промышленных поколениях, полученных в результате несложных внутригибридных скрещиваний, что доказано экспериментами на тутовом шелкопряде. Способ также предназначен для сельскохозяйственных растений, у которых возможно получение андрогенетических абсолютно гомозиготных особей. 3 з.п.ф-лы, 4 ил.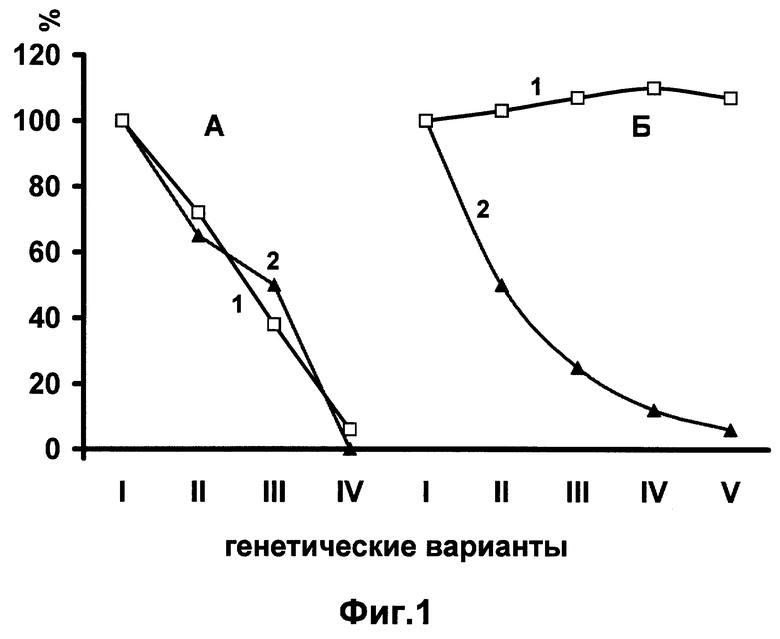
| Струнников В.А | |||
| Природа гетерозиса и новые методы его повышения | |||
| - М.: Наука, 1994, с.41 | |||
| Струнников В.А | |||
| Генетические методы селекции и регуляции пола тутового шелкопряда | |||
| - М.: ВО "Агропромиздат", 1987, с.35. |

