Изобретение относится к сельскому хозяйству, в частности к способам селекции.
Известен способ закрепления гетерозисного эффекта при получении вегетативно размножающихся многолетних растений риса (см., например, F.Hu, P.Xu, J.Li, J.Zhou, Е.Sacks, Z.Li, D.Tao Rice heterosis fixing by means of vegetative propagation from Oryza longistaminata. // Abstracts 4th International Symposium on Hybrid rice, 2002, p.95), заключающийся в переносе генов, ответственных за формирование корневища у диких родственников риса (например, от вида Orisa longistominata, у которого эти гены расположены в третьей и четвертой хромосомах и ассоциированы с локусами OSR 16, RM 119 соответственно).
Известен способ закрепления гетерозисного эффекта (см., например, Bennett J., Xuezhi Bi, Kathiresan, Chaudhury A., Ivahova A., Payne Т., Dennis L., and Khush G. Molecular tools for achieving synthetic apomixes for hybrid rice. // Abstracts 4 International Symposium on Hybrid rice, 2002, p. 12), заключающийся в переводе гибридов на апомиктичное размножение, методом переноса генов, ответственных за апомиктичное размножение из других видов, включая и родственные рису: кукуруза, сорго, пшеница.
Известен способ закрепления гетерозисного эффекта (см., например, V.Krishnasamy, 2002), заключающийся в получении синтетических семян гибридов. Под синтетическими семенами в данном случае подразумевается использование соматических эмбрионов, в качестве эндосперма которых служит смесь регуляторов роста, элементов питания и антибиотиков.
Недостатком указанных выше способов является то, что для закрепления гетерозисного эффекта они предлагают полное сохранение комплекса генов исходного гибрида, включая летальные и полулетальные гены, приводящие к расщеплению в последующих поколениях при половом размножении. Их основная задача заключается в переводе гибридов на вегетативное или апомиктичное размножение, кроме того, ни один из приведенных способов не реализован.
Известен способ закрепления гетерозиса гибридов в последующих поколениях, принятый нами в качестве прототипа, описанный в патенте №2453253, М.Кл. A01K 67/00, A01K 67/04, опубликованный 27.07.2000, Бюл. №21 (ч.2), включающий получение гомозиготных особей, осуществление возвратных скрещиваний с абсолютно гомозиготными особями (для растений дигаплоидами, полученными через культуру пыльников) для удаления летальных и полулетальных генов из генотипа гетерозисного гибрида и восстановления комплекса генов исходного гибрида.
Недостатком способа является его трудоемкость, выражающаяся в проведении 5-6 возвратных скрещиваний между дигаплоидом и исходной особью, то есть растением гетерозисного гибрида, пыльцу которого использовали для получения популяции дигаплоидных растений. Для проведения возвратного скрещивания необходимо поддерживать жизнеспособность гибрида в течение нескольких его обычных жизненных циклов, например для риса период его вегетации 3-5 месяцев, после чего растения погибают. Для удаления летальных и полулетальных генов, а также сохранения комплекса генов, определяющего гетерозисный эффект, необходимо получить гибрид между дигаплоидом и исходным растением гетерозисного гибрида, от которого получали пыльцу для создания дигаплоидов. Получение дигаплоидов требует продолжительного времени от 7 месяцев, цикл вегетации исходного растения к этому времени уже закончен и оно очень ослаблено. Гибридизация дигаплоидов на ослабленные материнские растения приводит к очень низкой завязываемости и жизнеспособности гибридных семян, эта процедура повторяется 5-6 раз.
Задачей заявленного изобретения является повышение эффективности процесса восстановления комплекса генов исходного гибрида и удаления из его генотипа полулетальных, а также неэффективно действующих генов, приводящих к снижению продуктивности в последующих поколениях.
Технический результат - создание на основе гетерозисных гибридов высокопродуктивных сортов с аналогичной урожайностью.
Указанный технический результат при реализации способа достигается тем, что в способе закрепления гетерозиса гибридов в последующих поколениях, включающем получение гомозиготных особей через культуру пыльников для удаления летальных и полулетальных генов из генотипа гетерозисного гибрида, согласно предлагаемому изобретению для восстановления комплекса генов исходного гибрида проводят гибридизацию генотипически контрастных дигаплоидных линий, при этом в гибридизацию включают только наиболее продуктивные дигаплоидные линии, а контрастность дигаплоидных линий оценивают по комплексу признаков морфологических, физиологических, биохимических, молекулярных маркеров (SSR, SNP, и.т.д.), по вкладам генетических систем в продуктивность образца, и по совокупности всех предложенных методов.
В результате гибридизации различных дигаплоидных линий между собой, а не с исходным гибридом, происходит объединение лучших генов, обуславливающих высокую продуктивность исходного гибрида и, следовательно, восстановление комплекса генов, определяющих гетерозисный эффект, которые были распределены при кроссинговере и получении дигаплоидов по различным образцам. Это дает возможность отказаться от продолжительного и очень сложного поддержания жизнеспособности растения (в течение нескольких обычных его жизненных циклов) исходного гибрида и избежать проблем, связанных с получением от него потомства.
Включение в гибридизацию только наиболее продуктивных дигаплоидных линий позволяет отбраковать образцы, сохранившие или получившие во время культивирования на питательной среде летальные, полулетальные и сублетальные гены, и, тем самым, обеспечить более эффективное очищение генотипа исходного гибрида.
Выделение контрастных дигаплоидных линий позволяет выявить образцы, несущие различные гены, определяющие гетерозисный эффект, и в результате обеспечивает ускорение процесса их накопления в создаваемом генотипе.
Гибридизация контрастных высокопродуктивных образцов, полученных при скрещивании различных дигаплоидных линий, еще более сокращает процесс накопления исходного комплекса генов. Проведение 2 скрещиваний контрастных дигаплоидных линий почти в 2 раза сокращает процесс накопления исходного комплекса генов гетерозисного гибрида, что в итоге позволяет получать высокопродуктивные сорта уже на первом этапе.
Проведенный анализ уровня техники, включающий поиск по патентным и научно-техническим источникам информации, и выявление источников, содержащих сведения об аналогах заявленного изобретения, позволил установить, что не обнаружен аналог, характеризующийся признаками, тождественными всем существенным признакам заявленного изобретения. Определение из перечня выявленных аналогов прототипа, как наиболее близкого по совокупности существенных признаков аналога, дозволил выявить совокупность существенных по отношению к усматриваемому заявителем техническому результату отличительных признаков в заявленном способе, изложенных в формуле изобретения.
Следовательно, заявляемое изобретение соответствует условию «новизна».
Для проверки соответствия заявленного изобретения требованию «изобретательского уровня» заявитель провел дополнительный поиск известных способов и технических решений с целью выявления признаков, совпадающих с отличительными от ближайшего аналога признаками завяленного изобретения. Результаты проверки показывают, что заявленное изобретение не следует для специалиста явным образом из известного уровня технологии и техники, в частности заявленным изобретением не предусматриваются известные преобразования.
Следовательно, заявленное изобретение соответствует требованию «изобретательский уровень» по действующему законодательству.
Сущность предлагаемого способа заключается в следующем.
Для закрепления гетерозисного эффекта по предлагаемому нами способу получали популяцию дигаплоидных линий из пыльцы гетерозисного гибрида (рисунок 1). Оценивали их продуктивность и жизнеспособность, выделяли для дальнейшей работы наиболее продуктивные дигаплоидные линии. Для выявления линий, несущих различные гены исходного комплекса генов, оценивали контрастность генотипов, выделенных высокопродуктивных дигаплоидных линий различными методами по комплексу: морфологических, физиологических, биохимических признаков, молекулярных маркеров, по вкладам генетических систем в продуктивность образца, и по совокупности всех предложенных методов. Для восстановления комплекса генов, определяющих гетерозисный эффект, проводили гибридизацию контрастных дигаплоидных линий, так как при получении дигаплоидов гены, определяющие высокую продуктивность гетерозисного гибрида, распределяются между различными особями. Получали дигаплоиды из пыльцы полученного гибрида через культуру пыльников. Оценивали их продуктивность и жизнеспособность, выделяли для дальнейшей работы наиболее продуктивные дигаплоидные линии, оценивали их продуктивность. Осуществление такой последовательности дает возможность получения высокопродуктивного сорта на первом этапе. Если же на первом этапе не удалось объединить в одном генотипе достаточного числа генов исходного комплекса генов, и, следовательно, полученные сорта не достигают урожайности исходного гибрида, переходили к следующему этапу, гибридизации линий, полученных при скрещивании различных контрастных дигаплоидных образцов. Из пыльцы полученного гибрида еще раз получали дигаплоидные линии. Выделяли наиболее продуктивные из них, размножали и оценивали продуктивность полученных сортов. Лучший сорт передается на госсортоиспытание.
Возможность селективной элиминации генов в культуре пыльников показана нами на организменном и молекулярном уровнях. Материалом для исследования послужили гибриды первого, второго поколения, родительские формы 13 гибридных комбинаций, а также дигаплоидные линии, полученные с использованием культуры пыльников из пыльцы гибридов первого поколения. Для закрепления гетерозисного эффекта в последующих поколениях использовали возвратные скрещивания исходного гибрида с дигаплоидами, полученными из его пыльцы. Получение дигаплоидов проводили для элиминации полулетальных аллелей, за которые в данном случае принимали любые неэффективно работающие локусы, понижающие жизнеспособность образца. Попадание летального гена в гаплоидный пронуклеус приводит зародыш к гибели на одной из стадий развития, выживают только особи, в генотипе которых минимальное количество летальных генов и максимальное благоприятных. При получении растений через культуру пыльников частота выживших особей не превышает 0,5%, следовательно, большая часть генотипов элиминируется. Вероятность выживания выше у генотипов с минимальным количеством летальных генов, что позволяет очистить генотип исходного гибрида от летальных и полулетальных генов, сохранив при этом комплекс благоприятных генов.
Возвратные скрещивания особей, лишенных вредных генов в гомозиготном и гетерозиготном состояниях, приводят к возникновению потомства, в генотипе которого также не может быть полулеталей в гомозиготном состоянии. Это и наличие большого числа генов модификаторов позволяет не только сохранить, но даже и усилить гетерозис по всех последующих беккроссных поколениях.
Следовательно, проблему закрепления гетерозиса можно решить посредством получения гибридов без леталей и полулеталей. При этом основной генотип исходного гибрида должен быть не только сохранен, но и улучшен. Высокая жизнеспособность беккроссных поколений, начиная с первого, представляет большой интерес, так как эти поколения по существу происходят в результате самооплодотворения, обычно сильно снижающего жизнеспособность. Высокая жизнеспособность объясняется тем, что беккроссы не приводят, как простое бисексуальное размножение, к появлению в генотипе потомства вредных генов в гомозиготном состоянии. Беккроссирование в данном случае проводили для возвращения в генотип гибрида части генов исходного комплекса, повышающих жизнеспособность. Беккроссное поколение получено в 13 гибридных комбинациях риса. Анализ продуктивности, полученный при беккроссировании растений, проводили при сравнении: с исходным гибридом; дигаплоидами, полученными из пыльцы этого гибрида; вторым поколением гибрида; родительскими формами. Так как сорта многолинейны (гены, определяющие морфотип, селекционер контролирует, но по устойчивости эффективности физиологических процессов полиморфизм внутри сорта сохраняется), то и гибриды первого поколения одной и той же комбинации, но полученные при гибридизации генетически различных линий одного сорта также генетически также различны. Нами было заложено два варианта опытов. В первом получали беккроссное поколение при скрещивании с растением из той же гибридной комбинации, посеянным позднее после получения дигаплоидных линий. Во втором - с исходным лучшим растением гибридной комбинации, отобранным для получения дигаплоидов (жизнеспособность его поддерживалась в течение всего получения дигаплоидов и выращивания их до фазы цветения).
В первом варианте опыта наблюдалось расщепление, что подтверждает многолинейность исходных сортов и необходимость получения беккроссного поколения при скрещивании только с растением - донором пыльцы. Во втором варианте опыта показано отсутствие расщепления в беккроссном поколении и, следовательно, возможность очищения генотипа гетерозисного гибрида от неэффективно действующих генов в культуре пыльников (таблица 1-2).
В гибридных комбинациях, по которым было получено большое количество дигаплоидных линий, а следовательно, была возможность отбора высокопродуктивной дигаплоидной линии для проведения возвратного скрещивания, продуктивность полученных растений была выше, чем у исходного гибрида (таблица 3).
При скрещивании гибрида с низкопродуктивными дигаплоидами продуктивность растений, полученных при возвратном скрещивании, ниже, чем у исходного гибрида, однако дисперсионный анализ достоверных различий по продуктивности исходного гибрида и возвратного скрещивания не показал. Следовательно, при получении дигаплоидов для очищения от неблагоприятно действующих генов необходимо создавать большее количество их линий, так как часто оказываются жизнеспособными низкопродуктивные и стерильные линии. Продолжительное культивирование растительных тканей на среде вызывает появление большого числа мутаций как ядерных, так и соматических, кроме того, часть линий, несущих летали, также выживает. Однако эти линии легко браковать по продуктивности, так как все летали находятся в гомозиготном состоянии. Следовательно, необходимо проводить отбор по продуктивности среди дигаплоидных линий. Полученные нами данные показывают, что при получении дигаплоидов происходит очищение исходного гибрида от леталей и полулеталей, принесенных родительскими формами. Для подтверждения селективной элиминации генов в культуре пыльников проводили молекулярное маркирование популяции дигаплоидов подвида японика (полученной в результате гибридизации сортов BR 29 и IR 69428), состоящей из 90 линий с использованием 200 маркеров (таблицы 4-5). 100 из них равномерно распределены по геному с расстоянием между ними 20 сМ, остальные 100, связанные с признаками, определяющими устойчивость к стрессам (засухе, засолению, высоким и низким температурам), были отобраны по литературным или интернет данным, или предоставлены группами, работающими в IRRI (международного института риса) с маркированием локусов, определяющих адаптивность.
Изучение частоты встречаемости различных аллелей полиморфных маркеров показало селективную элиминацию некоторых аллелей, что подтверждает возможность использования культуры пыльников для удаления генов, снижающих жизнеспособность.
После удаления полулетальных генов при получении дигаплоидов проводили гибридизацию контрастных дигаплоидных линий одной гибридной комбинации. При получении дигаплоидных линий на основе гетерозисного гибрида комплекс генов, определяющих гетерозис, распределяется между дигаплоидными линиями, для его объединения в одном генотипе проводили гибридизацию контрастных высокопродуктивных линий. Изучение величины гетерозиса по признакам, обуславливающим высокую продуктивность в 226 гибридных комбинациях, показало, что только 16,6% комбинаций были гетерозисными, между дигаплоидными линиями одной гибридной комбинации процент гетерозисных был 52,92%, это является подтверждением того, что использование дигаплоидных линий в гибридизации значительно увеличивает вероятность получения гетерозисных комбинаций.
После удаления полулетальных генов при получении дигаплоидов проводили гибридизацию контрастных дигаплоидных линий одной гибридной комбинации. При получении дигаплоидных линий на основе гетерозисного гибрида комплекс генов, определяющих гетерозис, распределяется между дигаплоидными линиями, для его объединения в одном генотипе проводили гибридизацию контрастных высокопродуктивных линий. Изучение величины гетерозиса по признакам, обуславливающим высокую продуктивность в 226 гибридных комбинациях, показало, что только 16,6% комбинаций были гетерозисными, между дигаплоидными линиями одной гибридной комбинации процент гетерозисных был 52,92%, это является подтверждением того, что использование дигаплоидных линий в гибридизации значительно увеличивает вероятность получения гетерозисных комбинаций. Для еще большей эффективности гибридизации в нее включают контрастные линии, что увеличивает процент гетерозисных комбинаций до 82,3%, а следовательно, позволяет собрать воедино комплекс генов исходного гибрида. С использованием данной технологии в 2010 году переданы на Государственное сортоиспытание 2 сорта риса Ивушка и Крепыш.
Изобретение относится к биохимии растений. Получают гомозиготные особи через культуру пыльников с последующим восстановлением комплекса генов исходного гибрида, путем гибридизации генотипически контрастных дигаплоидных линий. При этом в гибридизацию включают только наиболее продуктивные дигаплоидные линии, а контрастность дигаплоидных линий оценивают по комплексу признаков морфологических, физиологических, биохимических, молекулярных маркеров (SSR, SNP, и.т.д.), по вкладам генетических систем в продуктивность образца, и по совокупности всех предложенных методов. Предложенный авторами способ позволяет более эффективно, чем с использованием предложенных ранее методов, очистить генотип исходного гибрида, ускорить процесс накопления генов, определяющих гетерозисный эффект, отказаться от продолжительного и очень сложного поддержания жизнеспособности растения (в течение нескольких обычных его жизненных циклов) исходного гибрида и избежать проблем, связанных с получением от него потомства. Сокращает процесс накопления исходного комплекса генов, определяющих гетерозисный эффект и удаления генов, снижающих продуктивность и жизнеспособнось гибрида практически вдвое. 1 ил., 5 табл.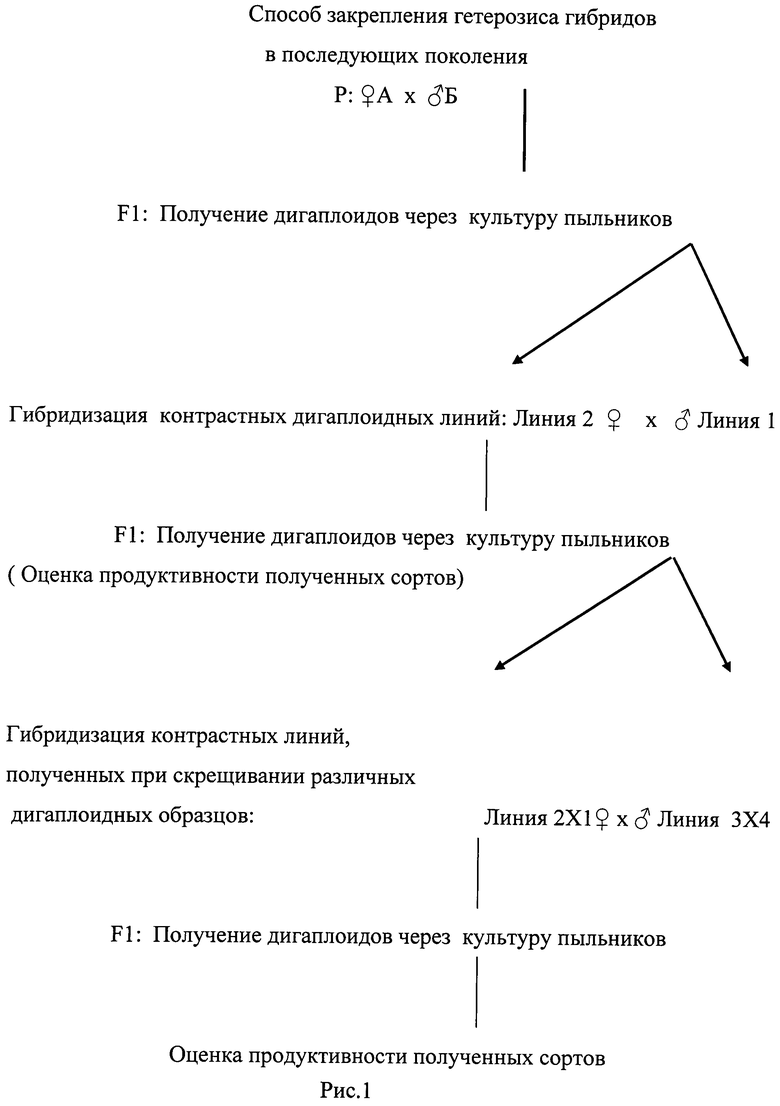
Способ закрепления гетерозиса гибридов сельскохозяйственных растений в последующих поколениях, включающий получение гомозиготных особей через культуру пыльников с последующим восстановлением комплекса генов, определяющих гетерозисный эффект исходного гибрида, отличающийся тем, что восстановление комплекса генов проводят путем гибридизации генотипически контрастных дигаплоидных линий, а контрастность дигаплоидных линий оценивают по комплексу признаков морфологических, физиологических, биохимических, молекулярных маркеров (SSR, SNP и т.д.), по вкладам генетических систем в продуктивность образца и по совокупности всех предложенных методов.
| СПОСОБ ЗАКРЕПЛЕНИЯ ГЕТЕРОЗИСА ГИБРИДОВ В ПОСЛЕДУЮЩИХ ПОКОЛЕНИЯХ | 1999 |
|
RU2153253C1 |
| КОСТЫЛЕВ П.И | |||
| Биотехнологии и оценочный этап селекции риса | |||
| - Зерновое хозяйство России, 2009, №1, найдено online [21.02.2012]. | |||

