Изобретение относится к области биотехнологии, а именно к получению буфорина-2 - антимикробного пептида жабы Bufo bufo gargarisans, который может найти применение в медицинской и ветеринарной практике в качестве антибиотика широкого спектра действия.
Буфорин-2 (TRSSRAGLQFPVGRVHRLLRK) - фрагмент N-концевой части гистона Н2А жабы дальневосточной Bufo bufo gargarisans [U.S. Pat. No.5,936,063]. Буфорин-2 обладает широким спектром бактерицидного и фунгицидного действия, механизм которого состоит в образовании нестабильных тороидальных пор, быстрой транслокации антибиотика через мембрану и последующем связывании с нуклеиновыми кислотами. Пространственная структура буфорина-2 включает короткий неупорядоченный N-концевой участок (TRSS) и два спиральных участка (RAGLQF и VGRVHRLLRK), разделенных остатком пролина. Эксперименты с укороченными синтетическими производными буфорина-2 показали, что удаление неупорядоченного участка не приводит к снижению антимикробной активности пептида, а минимальным фрагментом, обладающим таковой, является α-спиральный участок VGRVHRLLRK. Среди производных буфорина-2 были найдены пептиды, сохраняющие, в отличие от природного буфорина-2, активность в растворах с высокой концентрацией солей. Запатентован ряд искусственных аналогов буфорина-2 на основе его α-спирального фрагмента, в том числе аналоги, включающие повторяющиеся тетрапептидные участки RLLR [U.S. Pat. No.6,531,446]. Кроме того, заявлен дополнительный набор аналогов [WO 03080652].
Активность в отношении метициллин-резистентных штаммов Staphylococcus epidermidis создает предпосылки для использования препаратов на основе буфорина-2 в хирургии и трансплантологии. Эксперименты на животных моделях показали, что буфорин-2 способствует снижению концентрации бактериального эндотоксина и фактора некроза опухолей TNF-α в плазме крови, препятствуя, таким образом, развитию септического шока. При совместном применении буфорина-2 с традиционными антибиотиками наблюдается явление синергизма.
Известен способ получения модифицированного буфорина-2, состоящий в экспрессии в Е.coli гибридного белка, содержащего короткий отрицательно заряженный участок, нейтрализующий активность буфорина-2, и катионный участок, соответствующий зрелому пептиду [U.S. Pat. No. 6,183,992, C07K 14/435]. Отрицательно заряженный участок содержит 2 остатка цистеина, участвующих в образовании дисульфидных связей и, таким образом, стабилизирующих структуру телец включения, в которых накапливается белок. Конструируют ген, кодирующий мультимер гибридного белка, в составе которого чередуются указанные выше катионные и анионные участки. Буфорин-2 получают путем расщепления гибридного белка бромцианом по остаткам метионина, ограничивающим с двух сторон участки, соответствующие зрелому пептиду. В качестве метода очистки целевого пептида от основных примесей, в первую очередь от анионного фрагмента, используют ионообменную хроматографию.
Известен наиболее близкий к заявленному способ получения модифицированного буфорина-2, являющийся усовершенствованной разновидностью вышеописанного способа [Lee J.H., Kim M.S., Cho J.H., Kim S.C. Enhanced expression of tandem multimers of the antimicrobial peptide buforin II in Escherichia coli by the DEAD-box protein and trxB mutant // Appl. Microbiol. Biotechnol. - 2002. - Vol.58 (6)]. Используют штаммы E.coli, мутантные по тиоредоксин-редуктазе (TrxB), промотор фага Т7 заменяют промотором tac, а также коэкспрессируют мРНК-стабилизирующий белок.
К недостатку обоих способов можно отнести то, что получаемый пептид не является идентичным природному буфорину-2: после расщепления гибридного белка бромцианом в составе целевого пептида остается дополнительный (22-й) С-концевой остаток гомосерина.
Изобретение решает задачу получения антимикробного пептида буфорина-2, идентичного натуральному.
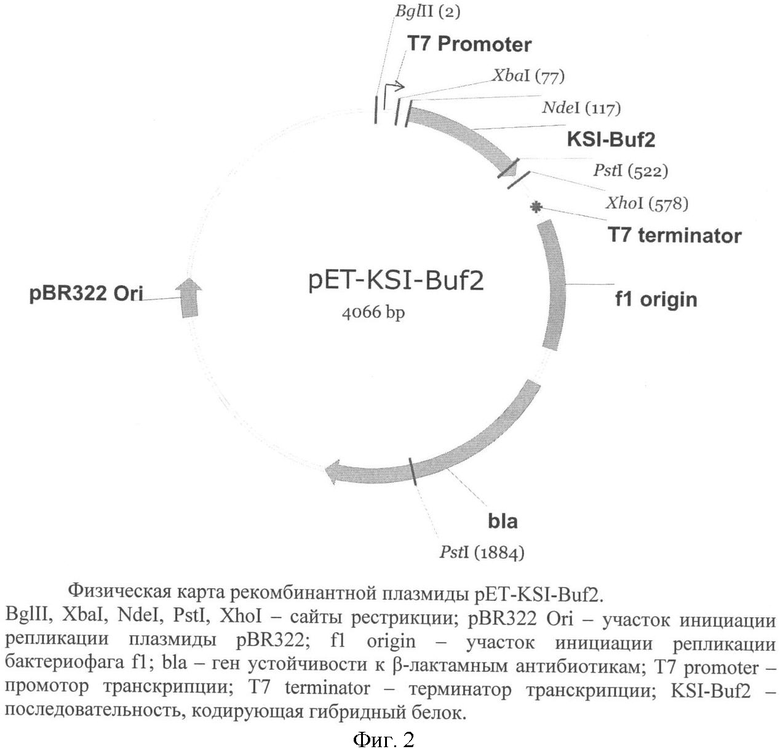
Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pET-KSI-Buf2 размером 4066 п.о., кодирующей гибридный белок KSI-Buf2, содержащий антимикробный пептид буфорин-2, состоящей из:
- Bg1II/XhoI-фрагмента ДНК, приведенного на фиг.1, содержащего промотор транскрипции Т7 РНК-полимеразы, участок связывания рибосомы и последовательность, кодирующую модифицированную кетостероид-изомеразу Pseudomonas testosteroni и буфорин-2, разделенные остатком метионина;
- Bg1II/XhoI-фрагмента ДНК плазмиды рЕТ20b(+), содержащего терминатор транскрипции Т7-РНК-полимеразы, сайт инициации репликации и ген β-лактамазы, детерминирующий устойчивость трансформированных плазмидой pET-KSI-Buf2 клеток Escherichia coli к ампициллину, в качестве генетического маркера;
за счет штамма Escherichia coli BL21(DE3)/pET-KSI-Buf2 - продуцента гибридного белка, содержащего антимикробный пептид буфорин-2;
а также за счет способа получения антимикробного пептида буфорина-2, включающего культивирование штамма-продуцента Escherichia coli BL21(DE3)/pET-KSI-Buf2, разрушение клеток, выделение и отмывку телец включения, содержащих гибридный белок KSI-Buf2, солюбилизацию гибридного белка и расщепление его бромцианом в кислой среде по остатку метионина, растворение целевого пептида и его очистку методом обращенно-фазовой ВЭЖХ.
Преимущество предлагаемого изобретения заключается, во-первых, в том, что в результате осуществления способа получается пептид, полностью идентичный природному буфорину-2. На фиг.1 приведена аминокислотная последовательность гибридного белка KSI-Buf2, содержащего буфорин-2, и соответствующая ей нуклеотидная последовательность гена. Введение остатка метионина непосредственно перед N-концевым остатком буфорина-2 обеспечивает возможность избирательного химического расщепления полипептидной цепи бромцианом с высвобождением целевого пептида, не содержащего каких-либо дополнительных аминокислотных остатков по сравнению с природным буфорином-2. Использование бактериальной кетостероид-изомеразы в качестве партнера для рекомбинантной экспрессии позволяет нейтрализовать токсичность антимикробного пептида для штамма-продуцента и благодаря этому достичь высоких выходов гибридного белка (более 50% от суммарного клеточного белка). Достоинством способа является простота технологии очистки буфорина-2. Отсутствие внутренних остатков метионина в последовательности белка-носителя сводит к минимуму число индивидуальных полипептидных фрагментов, образующихся в результате реакции с бромцианом, и, таким образом, облегчает процесс очистки целевого пептида. Высокий уровень экспрессии KSI-Buf2 и его крайне низкая растворимость в воде позволяют опустить стадию хроматографической очистки гибридного белка и ограничиться отмывкой нерастворимой фракции суммарного клеточного белка. Для биосинтеза рекомбинантного белка применяются оптимальные регуляторные элементы, контролирующие его экспрессию: промотор и терминатор для высокопроцессивной РНК-полимеразы бактериофага Т7, консенсусный сайт связывания бактериальной рибосомы, стартовый и стоп-кодоны.
Конструирование новой плазмидной ДНК pET-KSI-Buf2, экспрессирующей гибридный белок KSI-Buf2, осуществляют на основе большого фрагмента Bg1II-XhoI плазмиды рЕТ20b(+). Искусственный ген, кодирующий модифицированную кетостероид-измеразу (KSI) и буфорин-2, фланкированный сайтами рестриктаз Bg1II и XhoI, получают химическим синтезом набора олигонуклеотидных фрагментов с последующей их сборкой и амплификацией при помощи полимеразной цепной реакции (ПЦР). Матрицей для амплификации последовательности KSI служит плазмида рЕТ-31b(+). Перед лигированием для получения липких концов очищенный ампликон и векторную плазмиду обрабатывают рестриктазами Bg1II и XhoI. Продуктами лигазной реакции трансформируют компетентные клетки Е.coli DH-10B или аналогичного штамма с выключенной системой рекомбинации и рестрикции ДНК. Отбор положительных клонов проводят при помощи ПЦР с использованием специфических праймеров и последующим рестриктным анализом выделенной плазмидной ДНК. Структуру гена, кодирующего гибридный белок, содержащий буфорин-2, определяют секвенированием по методу Сэнгера.
Рекомбинантная плазмидная ДНК pET-KSI-Buf2, кодирующая гибридный белок KSI-Buf2, содержащий антимикробный пептид буфорин-2, характеризуется следующими признаками:
- имеет размер молекулы 4066 п.о.;
- кодирует гибридный белок KSI-Buf2, содержащий модифицированную последовательность бактериальной кетостероид-изомеразы (KSI) Pseudomonas testosteroni и буфорин-2;
- состоит из: Bg1II/XhoI-фрагмента ДНК, содержащего промотор транскрипции Т7-РНК-полимеразы, участок связывания рибосомы и последовательность гибридного полипептида KSI-Buf2, содержащего кетостероид-изомеразу и буфорин-2, и Bg1II/XhoI-фрагмента ДНК плазмиды рЕТ20b(+), содержащего терминатор транскрипции Т7-РНК-полимеразы, сайт инициации репликации и ген β-лактамазы;
- имеет уникальную совокупность признаков: промотор и терминатор транскрипции РНК-полимеразы бактериофага Т7; сайт связывания рибосомы; искусственный ген, кодирующий гибридный белок KSI-Buf2, содержащий бактериальную кетостероид-изомеразу и буфорин-2; ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pET-KSI-Buf2 клеток к ампициллину.
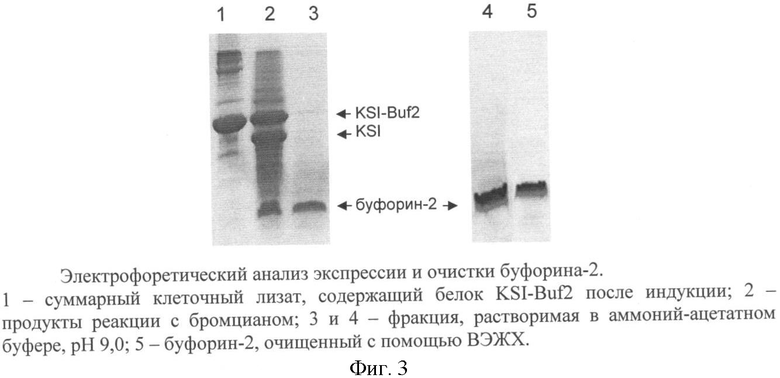
Для получения штамма-продуцента гибридного белка KSI-Buf2 препаратом плазмидной ДНК pET-KSI-Buf2 трансформируют компетентные клетки Escherichia coli BL21(DE3) и проводят отбор клонов, обладающих способностью экспрессировать рекомбинантный белок. Наличие и уровень экспрессии рекомбинантных белков контролируют с помощью денатурирующего ПААГ-электрофореза.
Клетки растут при температуре от 4°С до 40°С при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы. Клетки проявляют устойчивость к ампициллину (до 500 мкг/мл), обусловленную наличием в плазмиде pET-KSI-Buf2 гена β-лактамазы (bla).
Преимущества штамма Е.coli BL21(DE3) в качестве основы для создания штамма-продуцента заключаются в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого рекомбинантного белка и загрязнения препарата наиболее активными протеазами Е.coli. В хромосомную ДНК BL21 (DE3) интегрирован ген Т7-РНК полимеразы, что, совместно с использованием Т7 промотора и Т7 терминатора в плазмиде pET-KSI-Buf2, обеспечивает быструю и эффективную продукцию белка клетками Е.coli при индукции изопропилтио-β-D-галактозидом или лактозой.
Клетки Е.coli BL21(DE3)/pE-KSI-Buf2 являются суперпродуцентом. При индукции изопропилтио-β-D-галактозидом происходит эффективный биосинтез гибридного белка KSI-Buf2, содержащего буфорин-2, который накапливается в клетках в количестве 50-70% от суммарного белка бактерии.
Биосинтез продукта проводят следующим образом: клетки штамма Е.coli BL21(DE3)/pET-KSI-Buf2 выращивают в богатой среде (например, LB) с добавлением 100-200 мкг/мл ампициллина до достижения культурой оптической плотности OD600 0,5-1,0 при температуре 37°С, ступенчато увеличивая объем культуральной жидкости путем последовательных пересевов материала, после чего индуцируют синтез белка изопропил-β-D-галактозидом в концентрации 0,1-1,0 мМ или лактозой и растят еще 6-12 часов при температуре 25-37°С. Увеличение выхода рекомбинантного белка может быть достигнуто с помощью принудительной аэрации культуральной жидкости и использования обогащенных питательных сред (например, с добавлением глюкозы).
Получение антимикробного пептида буфорина-2 из клеток продуцента включает следующие стадии: отделение бактерий от культуральной среды с помощью центрифугирования, их разрушение одним из обычно применяемых способов (ультразвуковой дезинтеграцией, химическим лизисом с использованием детергентов и хаотропных агентов), отмывку буферными растворами телец включения от водорастворимых компонентов клетки, солюбилизацию телец включения, обработку полученной белковой массы бромцианом, разделение продуктов реакции, очистку целевого пептида методом обращенно-фазовой ВЭЖХ.
Тельца включения, полученные после дезинтеграции биомассы, обрабатывают по следующей схеме:
1. Тельца включения подвергают двукратной отмывке 25-100 мМ фосфатным или трис-HCl буфером (рН 6,5-8,5) с добавлением 1% неионогенного детергента Triton Х-100 и 4М мочевины.
2. Отмытые тельца включения растворяют в 80% трифторуксусной кислоте, либо в 70% муравьиной кислоте, либо в 6М гуанидине-HCl с добавлением 0,1М HCl и добавляют равную массу бромциана. Реакционную смесь инкубируют в темноте 16-20 ч при температуре 20-25°С. Реакцию останавливают добавлением трех-пятикратного объема воды, после чего упаривают образец под вакуумом при температуре 37°С. Процедуру добавления воды и упаривания повторяют несколько раз.
3. Продукты реакции растворяют в 0,2-0,5 М аммоний-ацетатном буфере (рН 9-10) или другом буфере, обеспечивающем высокую растворимость буфорина-2 и плохо растворяющем гибридный белок KSI-Buf2 и белок-носитель KSI. Нерастворимый осадок отделяют центрифугированием. Раствор диализуют против 1% уксусной кислоты через мембрану с порогом задержки 1000 Да, после чего лиофильно высушивают.
4. Навеску препарата растворяют в 5% ацетонитриле с добавлением 0,1% трифторуксусной кислоты и подвергают очистке методом обращенно-фазовой ВЭЖХ в градиенте концентрации ацетонитрила. Фракцию, содержащую рекомбинантный буфорин-2, лиофильно высушивают.
5. При необходимости может быть проведена дополнительная очистка целевого пептида методом ВЭЖХ.
6. Идентичность полученного буфорина-2 природному пептиду устанавливают методами ПААГ-электрофореза в трис-трициновой системе, MALDI-времяпролетной масс-спектрометрии, секвенированием N-концевой аминокислотной последовательности по методу Эдмана, а также тестированием антимикробной активности методом радиальной диффузии в агаризованной питательной среде либо методом серийных разведении в жидкой питательной среде. Степень очистки буфорина-2 определяют методами ПААГ-электрофореза в трис-трициновой системе и MALDI-времяпролетной масс-спектрометрии. Выход буфорина-2, полученного заявленным способом, с чистотой не менее 95% составляет не менее 5 мг/л культуры.
На фиг.1 представлена нуклеотидная последовательность Bg1II/XhoI-фрагмента ДНК, содержащего промотор транскрипции Т7-РНК-полимеразы, участок связывания рибосомы и участок, кодирующий гибридный полипептид KSI-Buf2; приведена соответствующая аминокислотная последовательность KSI-Buf2. На фиг.2 представлена физическая карта плазмиды pET-KSI-Buf2. На фиг.3 показаны картины ПААГ-электрофореза гибридного белка и буфорина-2 на разных стадиях очистки. На фиг.4 представлена хроматограмма очистки буфорина-2 методом обращенно-фазовой ВЭЖХ. На фиг.5 приведен масс-спектр очищенного буфорина-2.
Изобретение иллюстрируют следующие примеры.
Пример 1.
Конструирование рекомбинантной плазмидной ДНК pET-KSI-Buf2.
Нуклеотидную последовательность, содержащую промотор транскрипции Т7-РНК-полимеразы, участок связывания рибосомы и участок, кодирующий гибридный полипептид KSI-Buf2, получают химико-ферментативным синтезом. Химический синтез олигонуклеотидов, используемых для сборки нуклеотидной последовательности, выполняют твердофазным фосфоамидитным методом с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов - 5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O-(β-цианэтилдиизопропиламино)-фосфитов, активированных тетразолом. Фрагмент, кодирующий белок-носитель KSI, получают методом ПЦР-амплификации с помощью ген-специфических праймеров, используя в качестве исходной матрицы ген KSI в плазмиде рЕТ-31b(+). Остальные участки последовательности получают путем последовательного отжига и элонгации взаимно перекрывающихся олигонуклеотидов. На завершающей стадии синтеза последовательность амплифицируют с помощью праймеров, несущих на 5'-концах сайты узнавания рестриктаз Bg1II и XhoI. Продукт амплификации гидролизуют указанными рестриктазами, очищают электрофорезом в 1,5% агарозном геле, полосу ДНК величиной 576 п.о. выделяют из геля методом электроэлюции в 15% раствор ПЭГ-6000 и лигируют с фрагментом ДНК размером 3,5 тыс. п.о., полученным в результате обработки плазмиды рЕТ20b(+) рестриктазами Bg1II и XhoI. В результате лигазной реакции образуется кольцевая ковалентно замкнутая ДНК размером 4066 п.о. Продуктами лигазной реакции трансформируют компетентные клетки E.coli DH-10B, приготовленные по стандартному протоколу с помощью 0,1 М хлорида кальция. После трансформации суспензию бактерий смешивают с питательной средой LB, растят 1 ч при 37°С и высевают на чашки Петри с LB-агаром, содержащим 50 мкг/мл ампициллина.
Первичный отбор клонов, содержащих нужную плазмиду, осуществляют методом «ПЦР с клонов». Отобранные клоны используют для подращивания в жидкой среде и выделения плазмидной ДНК, которую анализируют на наличие вставки с помощью рестриктного анализа. Окончательное строение плазмид, содержащих требуемый фрагмент, подтверждают определением нуклеотидной последовательности секвенированием по Сэнгеру. По данным секвенирования отбирают ту плазмиду, у которой нуклеотидная последовательность лигированной вставки полностью идентична первоначально запланированной (фиг.1).
Пример 2.
Получение штамма-продуцента E.coli BL21(DE3)/pET-KSI-Buf2, вырабатывающего рекомбинантный белок KSI-Buf2, и определение его продуктивности.
Проводят трансформацию клеток Е.coli BL21(DE3) плазмидой pET-KSI-Buf2, как описано в примере 1. Петлей переносят выросшие колонии в 10 мл жидкой среды LB, содержащей 150 мкг/мл ампициллина, подращивают до оптической плотности OD600 0,7 на термостатируемой качалке со скоростью вращения 200 мин-1 при температуре 37°С, отбирают аликвоту культуры для последующего анализа, добавляют индуктор - изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают инкубацию при температуре 37°С в течение 5 ч, отбирая каждый час пробы по 0,3 мл для определения оптической плотности OD600 и последующего электрофоретического анализа. Равные аликвоты суспензии клеток, отобранных до внесения индуктора и после завершения роста, центрифугируют, отделяют супернатант и анализируют осадок клеток ПААГ-электрофорезом в денатурирующих условиях. Для этого образцы лизируют буфером, содержащим 1% SDS, 1% β-меркаптоэтанола, 8М мочевину на водяной бане в течение 5 мин. Электрофорез проводят в 15% SDS-ПААГ. Гель прокрашивают 0,025% кумасси G-250 по стандартной методике. Появление отчетливой полосы в районе 16 кДа в образцах, отобранных после индукции, свидетельствует о способности штамма синтезировать KSI-Buf2. Относительное содержание рекомбинантного белка определяют путем сканирования и денситометрического анализа окрашенных гелей.
Пример 3.
Получение рекомбинантного антимикробного пептида буфорина-2.
Культивирование клеток штамма-продуцента Escherichia coli BL21(DE3)/pET-KSI-Buf2 проводят следующим образом.
Клетки штамма BL21(DE3)/pET-KSI-Buf2 выращивают в жидкой среде LB с добавлением 150 мкг/мл ампициллина и 1% глюкозы до достижения культурой оптической плотности OD600 1,0 при температуре 37°С. Индукцию синтеза белка проводят с помощью изопропил-β-D-галактозида в концентрации 1,0 мМ, после чего инкубируют 10 часов при температуре 37°С. Уровень экспрессии гибридного белка, а также содержание гибридного белка и целевого пептида в препарате на разных стадиях очистки определяют с помощью SDS-ПААГ в трис-трициновой буферной системе (фиг.3).
По окончании ферментации клетки продуцента гибридного белка (биомассу) отделяют центрифугированием при 4000 g в течение 10 мин, ресуспендируют в лизирующем буфере (100 мМ Na2HPO4/NaH2PO4 (рН 7,8), 0,5М NaCl, 4M мочевина, 1% Triton X-100) и разрушают с помощью ультразвукового гомогенизатора. Все работы по выделению гибридного белка проводят при температуре 4°С. Нерастворимую фракцию клеточного белка (тельца включения), содержащую 70% гибридного белка, отделяют центрифугированием. Тельца включения дважды ресуспендируют в 50 мМ фосфатном буфере (рН 7,8) и центрифугируют, после чего лиофильно высушивают.
Очищенный гибридный белок растворяют в 80% трифторуксусной кислоте в концентрации 2%, добавляют равную массу бромциана (1 г бромциана на 1 г белка) и выдерживают при температуре 25°С в защищенном от света месте при постоянном встряхивании в течение 20 ч. Реакцию останавливают добавлением пятикратного объема деионизированной воды, после чего упаривают образец при 37°С. Процедуру добавления воды и упаривания повторяют трижды.
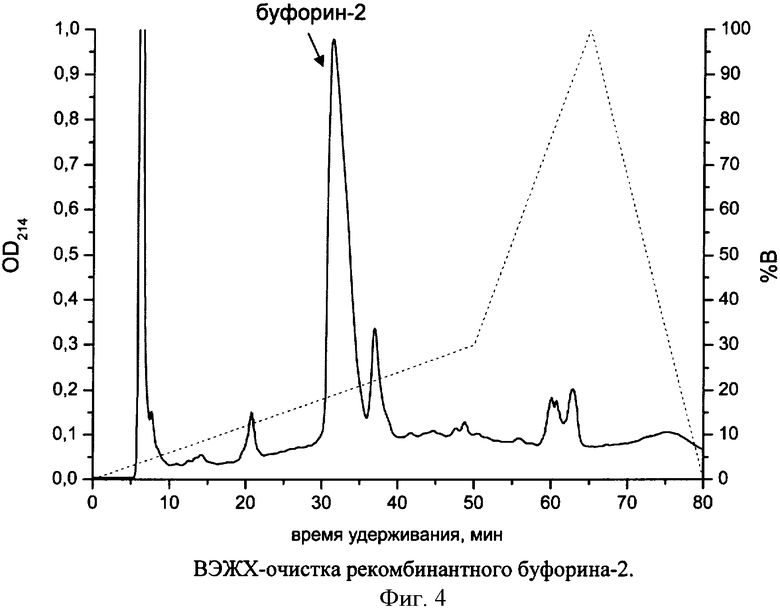
Продукты реакции растворяют в 200 мМ аммоний-ацетатном буфере (рН 9,5). Нерастворимый осадок отделяют центрифугированием. Раствор диализуют против 1% уксусной кислоты через мембрану с порогом задержки 1 кДа, после чего лиофильно высушивают и перерастворяют в концентрации 10 мг/мл в 5% ацетонитриле с добавлением 0,1% трифторуксусной кислоты. Нерастворимый осадок отделяют центрифугированием и отбрасывают. Очистку буфорина-2 проводят с помощью обращенно-фазовой ВЭЖХ на препаративных колонках в системе, состоящей из буфера А (5% ацетонитрил, 0,1% ТФУ) и буфера В (80% ацетонитрил, 0,1% ТФУ) (фиг.4). Буфорин-2 элюируют линейным градиентом буфера В от 0 до 30% в течение 50 мин. Детектирование ведут при длине волны 214 нм. Фракцию элюата, содержащую буфорин-2, собирают и лиофильно высушивают.
Для определения N-концевой аминокислотной последовательности буфорина-2 применяют автоматическое микросеквенирование на приборе Precise 491 cLC Protein Sequencing System (PE Applied Biosystems, США). Идентификацию фенилтиогидантоин-производных аминокислот проводят на анализаторе 120А РТН Analyzer (Applied Biosystems, США). В основе методики автоматического определения аминокислотной последовательности пептидов и белков лежит метод химической деградации полипептидной цепи по методу Эдмана, позволяющий последовательно отщеплять N-концевые аминокислотные остатки в виде фенилтиогидантоинов и идентифицировать отщепленные производные аминокислот методом обращенно-фазовой ВЭЖХ. В результате автоматического микросеквенирования установлена полная идентичность аминокислотных последовательностей генно-инженерного и природного буфорина-2 (TRSSRAGLQFPVGRVHRLLRK).
Для определения молекулярной массы буфорина-2 используют MALDI-времяпролетный масс-спектрометрический анализ на приборе Reflex III (Bruker Daltonics). В качестве матрицы используют 0,15М 2,5-дигидроксибензойную кислоту в смеси, содержащей 25% метанол и 0,1% трифторуксусную кислоту. Образец облучают УФ-лазером с длиной волны 337 нм. Полученный пик с m/z 2433,00 (фиг.5) соответствует молекулярному иону природного буфорина-2 (расчетная молекулярная масса составляет 2434,79 Да). Выход буфорина-2 составляет 10 мг/л клеточной культуры.
Пример 4.
Тестирование антимикробной активности буфорина-2.
Антимикробную активность буфорина-2 определяют методом радиальной диффузии пептидов в агарозном геле с тест-культурами микроорганизмов: Escherichia coli, штамм С600 (грамотрицательная бактерия); Bacillus subtilis, штамм L1 (грамположительная бактерия); Staphylococcus aureus, штамм 209Р (грамположительная бактерия). Тест-культуры пересевают из консерва на чашку Петри с LB-агаром и выращивают в течение ночи. Выросшие колонии переносят в 3 мл 1/2 LB и инкубируют на роторной качалке при 37°С до достижения культурой оптической плотности OD600 1,0-1,5. Аликвоту клеточной суспензии (1-4·106 КОЕ) добавляют к 12 мл расплавленной и охлажденной до 40°С обедненной среды (9 мМ фосфатный буфер, рН 6,5, 1 мМ цитрат натрия, 0,03% TSB, 1% агароза). Конечная концентрация бактерий составляет 2·105 КОЕ/мл. Полученной смесью заливают чашки диаметром 90 мм. В слое агарозы высверливают лунки диаметром 2 мм и вносят по 5 мкл раствора образца (буфорина-2 или стрептомицина, используемого в качестве контрольного антибиотика) в 10% ацетонитриле с добавлением 0,1% трифторуксусной кислоты. Чашки инкубируют, не переворачивая, в течение 3 часов при 37°С. На первый слой агарозы наслаивают 12 мл обогащенной среды (2LB с 1% агарозы). Чашки переворачивают и инкубируют в течение 18 ч при 37°С. Антимикробное действие пептида определяют по диаметру зон ингибирования роста тест-культуры. Результаты тестирования активности буфорина-2 представлены в таблице.
Антимикробная активность буфорина-2


Изобретение относится к области генной инженерии, конкретно к микробиальному продуцированию рекомбинантных белков, и может быть использовано в биотехнологии, медицине и ветеринарии. Конструируют рекомбинантную плазмидную ДНК pET-KSI-Buf2 размером 4066 п.о., кодирующую гибридный белок KSI-Buf2, содержащий антимикробный пептид буфорин-2. Путем трансформации клеток родительского штамма бактерий Escherichia coli BL21(DE3) рекомбинантной плазмидной ДНК pET-KSI-Buf2 получают штамм Escherichia coli BL21(DE3)/pET-KSI-Buf2 - продуцент гибридного белка, содержащего антимикробный пептид буфорин-2. Затем проводят культивирование штамма Escherichia coli BL21(DE3)/pET-KSI-Buf2 с последующим разрушением клеток, выделением и отмывкой телец включения, содержащих гибридный белок KSI-Buf2, солюбилизацией гибридного белка и расщеплением его бромцианом в кислой среде по остатку метионина, растворением целевого пептида и его очисткой методом обращенно-фазовой ВЭЖХ. Изобретение позволяет получить идентичный природному буфорин-2 с высоким выходом и по упрощенной технологии. 3 н.п. ф-лы, 5 ил., 1 табл.
BglII/XhoI-фрагмента ДНК, приведенного на рис.1, содержащего промотор транскрипции Т7 РНК-полимеразы, участок связывания рибосомы и последовательность, кодирующую модифицированную кетостероид-изомеразу Pseudomonas testosteroni и буфорин-2, разделенные остатком метионина;
BglII/XhoI-фрагмента ДНК плазмиды рЕТ20b(+), содержащего терминатор транскрипции Т7-РНК-полимеразы, сайт инициации репликации и ген β-лактамазы, детерминирующий устойчивость трансформированных плазмидой pET-KSI-Buf2 клеток Escherichia coli к ампициллину, в качестве генетического маркера.
| LEE J.H | |||
| et al., Enhanced expression of tandem multimers of the antimicrobial peptide buforin II in Escherichia coli by the DEAD-box protein and trxB mutant | |||
| Appl | |||
| Microbiol | |||
| Biotechnol., 2002, v.58, n.6, p.790-796 | |||
| RAO A | |||
| et al., Application of the "codon-shuffling" method | |||
| Synthesis and selection of de novo proteins as antibacterials, J | |||
| Biol. |

