Изобретение относится к области биотехнологии, а именно к получению аурелина - антимикробного пептида сцифоидной медузы Aurelia aurita, который может найти применение в качестве лекарственного средства в медицинской и ветеринарной практике.
Эндогенные антимикробные пептиды - эволюционно древние факторы врожденного иммунитета, играющие ключевую роль в защите многоклеточных организмов от инфекции [Bulet P. et al. (2004) Immunol Rev. 198: 169-84]. Сходные по строению и функции пептиды были выделены из тканей позвоночных и беспозвоночных животных, а также растений. У беспозвоночных, лишенных лимфоцитарного иммунитета, выработка защитных молекул пептидной природы составляет один из главных механизмов противодействия инфекции. Природные антимикробные пептиды могут стать прототипами новых антибиотиков широкого спектра действия, способных решить проблему резистентности к существующим антимикробным средствам.
Аурелин - катионный антимикробный пептид массой 4,3 кДа, выделенный из мезоглеи (соединительной ткани) сцифоидной медузы Aurelia aurita и состоящий из 40 аминокислотных остатков (SEQ ID No. 1) [Ovchinnikova T.V. et al. (2006) Biochem Biophys Res Commun. 348(2): 514-23]. Шесть остатков цистеина в составе молекулы образуют три внутримолекулярные дисульфидные связи. Аурелин проявляет антимикробную активность в отношении грамположительных и грамотрицательных бактерий. Наличие шести остатков цистеина в полипептидной цепи аурелина стало основанием для предварительного отнесения нового пептида к семейству дефенсинов - наиболее распространенному и консервативному семейству антимикробных пептидов. К дефенсин-подобным пептидам относятся дрозомицин, гелиомицин, термицины, скорпин, бутинин, ASABF (антибактериальные пептиды нематод), митилины и митицины. Кроме того, по 6 остатков цистеина содержат не обладающие сходством с дефенсинами молекулы тахистатинов, пенеидинов и микроплюзина. В то же время, уникальный порядок расположения остатков цистеина в аурелине отличает его от всех перечисленных пептидов. Его ближайшими гомологами в настоящее время можно считать группу токсинов из яда морских анемон (класс Anthozoa, тип Coelenterata), блокирующих калий-селективные каналы нервных клеток: BgK, ShK, HmK, AeK, AsKS.
Известен наиболее близкий к заявленному способ получения аурелина, заключающийся в измельчении мезоглеи медузы Aurelia aurita, экстракции 5% уксусной кислотой, последовательной ультрафильтрации через фильтры с размером пор 300 и 10 кДа, упаривании фильтрата, разделении компонентов с помощью препаративного гель-электрофореза и очистки активной фракции с помощью обращенно-фазовой ВЭЖХ [Ovchinnikova T.V. et al. (2006) Biochem Biophys Res Commun. 348(2): 514-23]. Недостатками данного способа являются сложная схема очистки целевого пептида и использование труднодоступного природного сырья.
Изобретение решает задачу расширения ассортимента биологически активных пептидов и получения антимикробного пептида аурелина сцифоидной медузы Aurelia aurita
Поставленная задача решается за счет конструирования экспрессирующего плазмидного вектора pE-Trx-Aur, состоящего из двух фрагментов ДНК:
- BglII/XhoI-фрагмента с нуклеотидной последовательностью SEQ ID No. 2, который содержит промотор транскрипции Т7 РНК-полимеразы, lac-оператор, участок связывания рибосомы и участок, кодирующий аффинную метку, белок-носитель тиоредоксин и антимикробный пептид аурелин;
- BglII/XhoI-фрагмента плазмиды pET31b(+), который содержит терминатор транскрипции Т7 РНК-полимеразы, сайт инициации репликации, ген β-лактамазы и ген lac-репрессора (lacI);
за счет штамма-продуцента гибридного белка Trx-Aur путем трансформации штамма Escherichia coli BL21(DE3) указанным вектором pE-Trx-Aur;
а также за счет способа получения антимикробного пептида аурелина, включающего культивирование штамма-продуцента Escherichia coli BL21(DE3)/pE-Trx-Aur, разрушение клеток, аффинную очистку гибридного белка Trx-Aur на металлохелатном носителе, расщепление гибридного белка Trx-Aur бромцианом по остатку метионина, введенному между последовательностями аурелина и тиоредоксина, и очистку целевого пептида методом обращенно-фазовой ВЭЖХ.
С целью предотвращения преждевременной гибели микроорганизма-продуцента во время экспрессии в его клетках антимикробных пептидов и связанной с этим потери продуктивности, требуется временно нейтрализовать токсичность синтезируемого продукта. Заявленный плазмидный вектор pE-Trx-Aur обеспечивает высокий уровень экспрессии аурелина в E.coli за счет включения его аминокислотной последовательности в состав гибридного белка Trx-Aur (SEQ ID No. 3), несущего последовательность тиоредоксина А. Тиоредоксин способен накапливаться в высокой концентрации (до 40%) в цитоплазме E.coli в растворимой форме и применяется для сверхэкспрессии биологически активных полипептидов [LaVallie E.R. et al. (1993) Nature BioTechnology. 11: 187-193]. Необходимость использования белка-носителя обусловлена не только токсичностью зрелого аурелина для бактериальной клетки, но и возможностью его деградации в гетерологичной системе. Для высвобождения целевого пептида из молекулы гибридного белка Trx-Aur между последовательностями аурелина и тиоредоксина вводят остаток метионина, позволяющий проводить избирательное расщепление гибридного полипептида бромцианом.
При расщеплении гибридного белка Trx-Aur бромцианом целесообразным является использование в качестве партнера для гетерологичной экспрессии модифицированного тиоредоксина A E.coli, содержащего замену внутреннего остатка метионина на остаток другой аминокислоты, например, на лейцин (M37L). Известно, что подобное незначительное изменение первичной структуры не оказывает влияния на пространственную структуру и свойства тиоредоксина [Rudresh et al. (2002) Protein Eng. 15(8): 627-33]. Отсутствие внутренних остатков метионина в последовательности белка-носителя уменьшает число индивидуальных полипептидных фрагментов, образующихся в результате реакции с бромцианом. В предпочтительном случае расщепление гибридного белка дает всего один побочный продукт, значительно отличающийся по физико-химическим свойствам от целевого пептида, что облегчает процесс очистки последнего.
Очистку целевого пептида упрощают за счет включения в состав гибридного белка Trx-Aur аффинной метки - аминокислотной последовательности, позволяющей проводить очистку гибридного полипептида методом аффинной хроматографии на сорбентах с иммобилизованными (хелатированными) ионами металлов, т.е. металлохелатную очистку. В качестве такой последовательности обычно используют последовательность из 4-10 остатков гистидина, чаще всего - последовательность из шести остатков гистидина, которая вводится в N-концевую или C-концевую область гибридного белка, либо в область, разделяющую белок-носитель и целевой белок [Terpe K. (2003) Appl Microbiol Biotechnol. 60(5): 523-33]. В качестве аффинной метки для металлохелатной очистки также используют фрагменты HAT и 6xHN [U.S. Pat. No. 7,176,298].
В состав заявленного вектора pE-Trx-Aur включают регуляторные элементы, контролирующие экспрессию гибридного белка: промотор и терминатор для РНК-полимеразы бактериофага Т7, lac-оператор, консенсусный сайт связывания бактериальной рибосомы, стартовый и стоп-кодоны. Для подавления базальной экспрессии гена гибридного белка в состав вектора pE-Trx-Aur включают ген lac-репрессора (lacI). В состав заявленного вектора входят сайт инициации репликации (Ori) плазмиды pBR322 и маркерный ген β-лактамазы, детерминирующий устойчивость трансформированных плазмидой pE-Trx-Aur клеток Escherichia coli к ампициллину и позволяющий проводить отбор клеток, содержащих векторную ДНК, путем выращивания на соответствующей селективной питательной среде.
Конструирование плазмидного вектора pE-Trx-Aur, экспрессирующей гибридный белок Trx-Aur, содержащий последовательность аурелина, может быть осуществлено путем лигирования BglII/XhoI-фрагмента плазмиды pET-31b(+) (Novagen), содержащего область инициации репликации, Т7 терминатор, гены β-лактамазы и lacI, со вставкой, кодирующей ген гибридного белка. Искусственный ген, который кодирует гибридный белок, может быть получен химическим синтезом набора олигонуклеотидных фрагментов с последующей сборкой и амплификацией промежуточных и конечного продуктов при помощи полимеразной цепной реакции (ПЦР). Выбор структуры олигонуклеотидных праймеров для синтеза каждого из структурных элементов (промотора, оператора, сайта связывания рибосомы, тиоредоксина, аффинной последовательности, аурелина) основывают на данных по их нуклеотидным последовательностям, доступных из открытых источников (банки данных GenBank, EMBL-Bank, DDBJ). Матрицей для амплификации последовательности тиоредоксина служит бактериальный геном, либо плазмида рЕТ-32а(+) (Novagen). Замену метионинового ко дона в составе тиоредоксина (M37L) при необходимости осуществляют на стадии сборки методом направленного мутагенеза при помощи ПЦР. Перед лигированием для получения липких концов очищенный ампликон и плазмидный вектор обрабатывают рестриктазами. Продуктами лигазной реакции трансформируют компетентные клетки Е.coli штамма с выключенной системой рекомбинации и рестрикции ДНК, например, DH5α, DH10B или XL1-Blue. Отбор клонов, содержащих плазмидный вектор со вставкой, проводят при помощи ПЦР и рестрикционного анализа выделенной векторной ДНК. Правильность сборки конструкции определяют секвенированием векторной ДНК.
Штамм-продуцент гибридного белка Trx-Aur, содержащего последовательность целевого пептида аурелина, получают путем трансформации препаратом векторной ДНК pE-Trx-Aur компетентных клеток Е.coli и отбора клонов, обладающих способностью экспрессировать рекомбинантный белок. Наличие и уровень экспрессии рекомбинантного белка контролируют с помощью ПААГ-электрофореза в денатурирующих условиях. В качестве родительского штамма для создания штамма-продуцента используют Е.coli BL21(DE3). Преимущество использования данного штамма в качестве основы для создания штамма-продуцента заключается в том, что BL21 (DE3) обладает фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого гибридного белка Trx-Aur и загрязнения препарата наиболее активными протеазами Е.coli. В хромосомную ДНК BL21 (DE3) интегрируют ген Т7-РНК полимеразы, который совместно с Т7 промотором и Т7 терминатором в плазмиде pE-Trx-Aw обеспечивает продукцию гибридного белка Trx-Aur клетками Е.coli при индукции изопропилтио-β-D-галактозидом или лактозой. Базальная экспрессия (до момента добавления индуктора) Т7 РНК-полимеразы и гибридного белка Trx-Aur поддерживается на минимальном уровне благодаря наличию системы контроля на основе lac-операторов и генов lac-репрессора, присутствующих как в плазмиде pE-Trx-Aur, так и в хромосоме штамма-продуцента.
Клетки штамма-продуцента сохраняют культурально-морфологические и физиолого-биохимические признаки родительского штамма Е.coli. Клетки мелкие, палочковидной формы, грамотрицательные, 1×3,5 мкм, подвижные, хорошо растут на обычных питательных средах (МПА, МПБ, LB-бульон, LB-агар, минимальная среда с глюкозой). Рост в жидких средах характеризуется ровным помутнением, осадок легко седиментирует. Клетки растут при температуре от 4°С до 42°С при оптимуме рН от 6,8 до 7,5; в качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевого экстракта. В качестве источника углерода при росте на минимальной среде используют аминокислоты, глицерин, углеводы. Клетки проявляют устойчивость к ампициллину (до 500 мкг/мл), обусловленную наличием в плазмиде pE-Trx-Aur гена β-лактамазы.
Штамм-продуцент хранят на чашках и косяках при температуре 4°С, пересевая на свежие среды один раз в месяц, а также при температуре минус 70°С в среде LB с добавлением 10-30% глицерина.
Биосинтез продукта (экспрессию) проводят следующим образом: клетки штамма-продуцента выращивают в питательной среде (например, LB, MBL или М9) с добавлением необходимого селектирующего агента (100 мкг/мл ампициллина) при температуре 20-37°С до достижения культурой средней или поздней логарифмической фазы роста, ступенчато увеличивая объем культуральной жидкости путем последовательных пересевов материала, после чего индуцируют синтез гибридного белка добавлением изопропилтио-β-D-галактозида или лактозы и инкубируют дополнительно в течение 2-24 часов при температуре 20-37°С. Увеличения выхода гибридного белка достигают с помощью принудительной аэрации культуральной жидкости и культивирования штамма-продуцента на обогащенных питательных средах (например, с добавлением глюкозы, глицерина, дикарбоновых кислот, аминокислот, неорганических солей, в т.ч. содержащих микроэлементы).
Очистка антимикробного пептида аурелина включает следующие обязательные стадии: разрушение клеток, металлохелатную очистку гибридного белка Trx-Aur, обработку гибридного белка бромцианом, разделение продуктов реакции и очистку целевого пептида методом обращенно-фазовой ВЭЖХ.
Для очистки клеточного материала от примесей, содержащихся в культуральной жидкости, особенно при выделении гибридного белка из клеточной культуры с низкой плотностью, желательным является предварительное концентрирование клеток с помощью центрифугирования или фильтрации. Разрушение (лизис) клеток осуществляют физическим или химическим способом или комбинацией способов, например, с помощью ультразвукового дезинтегратора, Френч-пресса, дезинтегратора Гаулина, с помощью осмотического шока, детергентов, хаотропных агентов, гидролитических ферментов (лизоцим, ДНКаза). С целью снижения нагрузки на аффинный сорбент нерастворимые примеси из клеточного лизата могут быть удалены центрифугированием или фильтрацией. Для металлохелатной очистки может быть использован сорбент, содержащий такие хелатирующие группы? как иминодиацетат (IDA), нитрилотриацетат (NTA) или карбоксиметиласпартат (СМА, TALON) в комплексе с катионами Ni2+, Со2+, или Cu2+ [Chaga G.S. (2001) J. Biochem Biophys Methods. 49(1-3): 313-34]. Элюирование гибридного белка проводят, уменьшая рН буфера или увеличивая концентрацию имидазола, либо добавляя в буфер ЭДТА. Реакцию с бромцианом проводят в стандартных условиях: растворяют белок в 60-90% трифторуксусной, уксусной или муравьиной кислоте или в 6М хлориде гуанидина с добавлением 0,1М соляной кислоты, добавляют бромциан в 10-200-кратном стехиометрическом избытке по отношению к числу остатков метионина в образце, инкубируют в темноте в течение 16-20 ч. При необходимости проводят очистку продуктов реакции расщепления гибридного белка с помощью повторной металлохелатной хроматографии, отделяя таким образом белок-носитель и нерасщепленный гибридный белок. Очистку аурелина проводят методом обращенно-фазовой ВЭЖХ в системе ацетонитрил-вода с добавлением 0,1% ТФУ, используя градиент концентрации ацетонитрила. Степень чистоты пептида может быть повышена путем повторной очистки методом обращенно-фазовой ВЭЖХ.
Идентичность рекомбинантного и природного аурелина устанавливают методами ПААГ-электрофореза в денатурирующих услових, МАЛДИ-времяпролетной масс-спектрометрии, секвенированием N-концевой аминокислотной последовательности по методу Эдмана, тестированием антимикробной активности методом серийных разведении в жидкой питательной среде. Степень очистки определяют методами ПААГ-электрофореза в денатурирующих условиях и МАЛДИ-времяпролетной масс-спектрометрии, а также с помощью повторной обращенно-фазовой ВЭЖХ.
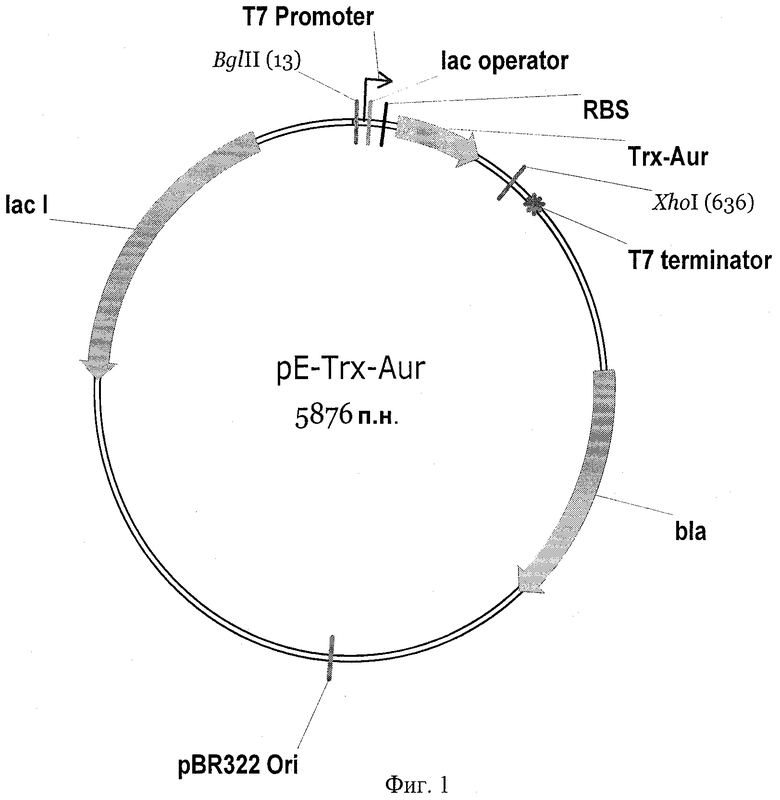
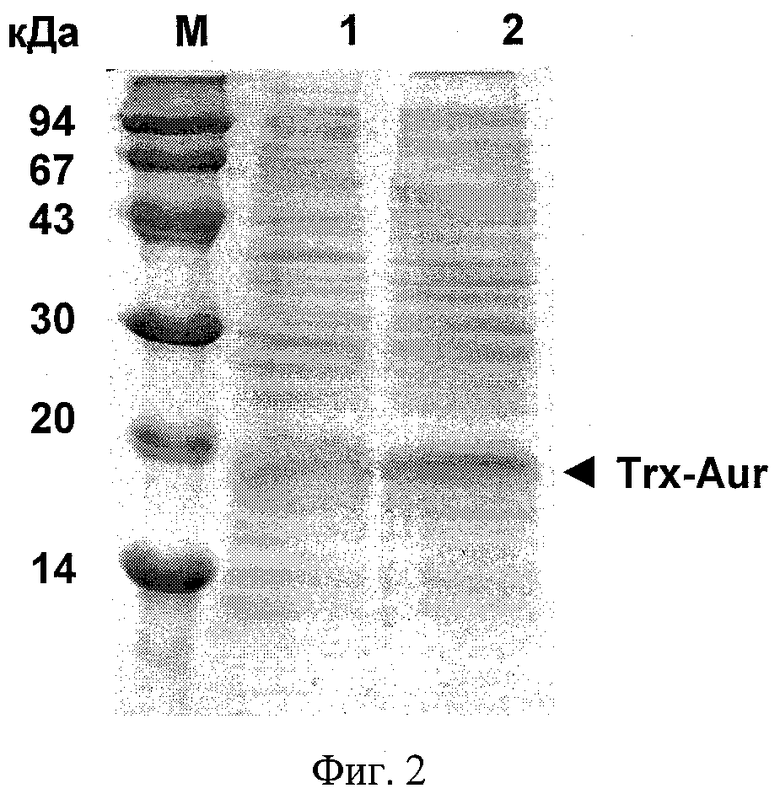
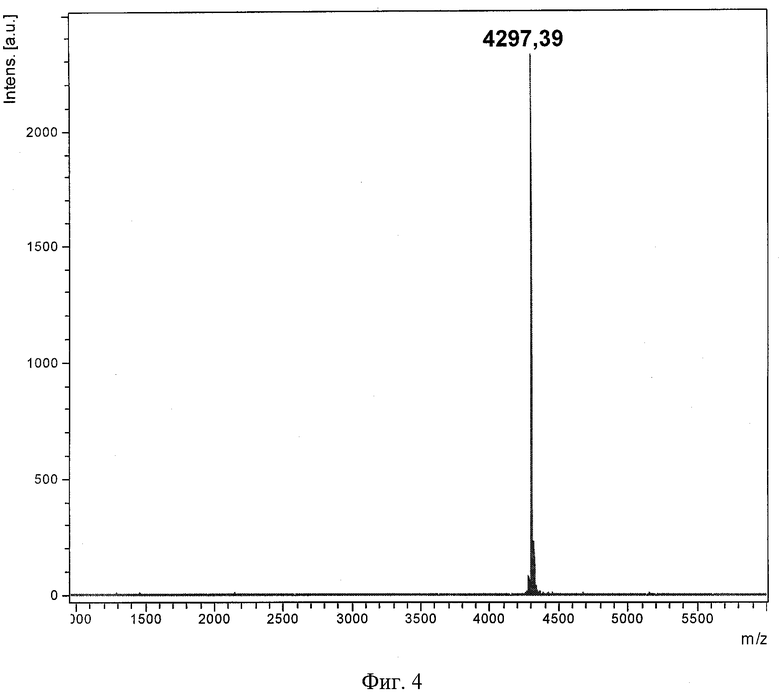
На фиг.1 представлена физическая карта плазмидного вектора для экспрессии антимикробного пептида аурелина: BglII, XhoI - сайты рестрикции; pBR322 Ori - участок инициации репликации плазмиды; bla - ген устойчивости к β-лактамным антибиотикам; lacI - ген lac-репрессора; Т7 promoter - промотор транскрипции; Т7 terminator - терминатор транскрипции; lac operator - сайт связывания lac-репрессора; RBS - сайт связывания рибосомы; Trx-Aur - последовательность, кодирующая гибридный белок, содержащий аурелин. На фиг.2 показана электрофореграмма клеточного лизата, содержащего гибридный белок Trx-Aur: M - стандарт молекулярной массы; 1 и 2 - суммарный клеточный лизат двух клонов после экспрессии. На фиг.3 представлена хроматограмма очистки рекомбинантного аурелина методом обращенно-фазовой ВЭЖХ. На фиг.4 приведен MALDI масс-спектр аурелина, полученного генно-инженерным способом.
Изобретение иллюстрируют примеры.
Пример 1
Конструирование плазмидного вектора pE-Trx-Aur
Нуклеотидную последовательность SEQ ID No. 2, содержащую промотор транскрипции Т7 РНК-полимеразы, lac-оператор, участок связывания рибосомы и участок, кодирующий гибридный полипептид (последовательно связанные гистидиновый октамер, тиоредоксин с заменой M37L, сайт расщепления бромцианом и аурелин), получают химико-ферментативным синтезом с помощью ПНР. Олигонуклеотиды, используемые в ПЦР, синтезируют твердофазным фосфоамидитным методом с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов - 5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O-(β-цианэтилдиизопропиламино)-фосфитов, активированных тетразолом.
Фрагмент, кодирующий белок-носитель тиоредоксин (M37L), получают методом ПЦР-амплификации и направленного мутагенеза с помощью ген-специфических праймеров, используя в качестве исходной матрицы плазмиду рЕТ32а(+), содержащую ген тиоредоксина. Остальные участки последовательности pE-Trx-Aur получают путем последовательного отжига и элонгации взаимно перекрывающихся олигонуклеотидов, а также отжига, элонгации и амплификации промежуточных продуктов синтеза. На завершающей стадии синтеза последовательность амплифицируют с помощью праймеров, несущих на 5'-концах сайты узнавания рестриктаз BglII и XhoI. Продукт амплификации гидролизуют указанными рестриктазами, очищают электрофорезом в 1,5% агарозном геле, полосу ДНК величиной 629 п.н. выделяют из геля методом электроэлюции в 15% раствор ПЭГ-6000 и лигируют с фрагментом ДНК размером 5,3 тыс.п.н., полученным в результате обработки плазмиды pET31b(+) рестриктазами BglII и XhoI. В результате лигазной реакции получают кольцевую ковалентно замкнутую ДНК размером 5876 п.н. (фиг.1). Продуктами лигазной реакции трансформируют компетентные клетки E.coli DH10B, приготовленные с помощью 0,1 М хлорида кальция. После трансформации суспензию бактерий смешивают с питательной средой LB, растят 1 ч при 37°С и высевают на чашки Петри с LB-агаром, содержащим 50 мкг/мл ампициллина.
Первичный отбор клонов, содержащих нужную плазмиду, осуществляют методом «ПЦР с клонов» с использованием праймеров на плазмидный остов и вставку. Отобранные клоны подращивают в жидкой питательной среде и выделяют плазмидную ДНК, которую анализируют на наличие вставки с помощью рестрикционного анализа. Окончательное строение плазмид, содержащих требуемый фрагмент, подтверждают определением нуклеотидной последовательности секвенированием по Сэнгеру. По данным секвенирования отбирают плазмиду со вставкой, нуклеотидная последовательность которой полностью соответствует запланированной (SEQ ID No. 2).
Пример 2
Получение штамма-продуцента Escherichia coli BL21(DE3)/pE-Trx-Aur, вырабатывающего гибридный белок Trx-Aur, и определение его продуктивности.
Проводят трансформацию компетентных клеток Е.coli BL21(DE3), приготовленных с помощью 0,1М хлорида кальция, плазмидой pE-Trx-Aur, описанной в примере 1. После трансформации суспензию бактерий смешивают с питательной средой LB, растят 1 ч при 37°С и высевают на чашки Петри с LB-агаром, содержащим 50 мкг/мл ампициллина и 0,5% глюкозы. Чашки инкубируют при 37°С в течение 18 ч.
Бактериологической петлей переносят выросшие колонии в 10 мл жидкой среды LB, содержащей 150 мкг/мл ампициллина, подращивают до оптической плотности OD600 0,7 на термостатируемой качалке со скоростью вращения 200 мин-1 при температуре 37°С, отбирают 0,3 мл культуры для последующего электрофоретического анализа, добавляют индуктор - изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают инкубацию при температуре 37°С в течение 5 ч, отбирая каждый час пробы по 0,3 мл для определения оптической плотности OD600 и последующего электрофоретического анализа. Равные аликвоты суспензии клеток, отобранных до внесения индуктора и после завершения роста, центрифугируют, отделяют супернатант и анализируют осадок клеток ПААГ-электрофорезом в денатурирующих условиях. Для этого образцы лизируют буфером, содержащим 2% додецилсульфат натрия (SDS), на водяной бане в течение 5 мин. Электрофорез проводят в 15% ПААГ в присутствии SDS. Гель прокрашивают 0,1% кумасси G-250 в присутствии 25% изопропилового спирта. Появление отчетливой полосы в районе 18 кДа в образцах, отобранных после индукции, свидетельствует о способности штамма синтезировать гибридный полипептид Trx-Aur, имеющий молекулярную массу 17,8 кДа (фиг.2). Относительное содержание гибридного белка, составляющее около 15%, определяют путем сканирования и денситометрического анализа окрашенных гелей.
Пример 3
Получение рекомбинантного антимикробного пептида аурелина
Клетки штамма-продуцента Escherichia coli BL21(DE3)/pE-Trx-Aur, полученного согласно примеру 2, выращивают в жидкой среде LB с добавлением 100 мкг/мл ампициллина и 1% глюкозы до достижения культурой оптической плотности OD600 0,7 при температуре 37°С. Индукцию синтеза белка проводят с помощью изопропил-β-D-галактозида в концентрации 1,0 мМ, после чего инкубируют 4 часа при температуре 37°С. Уровень экспрессии гибридного белка, а также содержание гибридного белка, белка-носителя и аурелина в препарате на последующих стадиях очистки определяют с помощью ПААГ-электрофореза в трис-трициновой буферной системе в денатурирующих условиях.
По окончании ферментации клеточную биомассу отделяют центрифугированием при 4000 g в течение 10 мин, ресуспендируют в буфере А (50 мМ трис-HCl, 0,5 М NaCl, 20 мМ имидазол, рН 7,8, 1 мМ PMSF) и разрушают клетки с помощью ультразвукового гомогенизатора. Полученный лизат центрифугируют (осветляют) при 25000 g в течение 20 мин. Все работы по получению осветленного лизата проводят при температуре 4°С. Очистку гибридного белка Trx-Aur, содержащего в качестве аффинной метки октагистидиновую последовательность, осуществляют с помощью металлохелатной хроматографии на препаративной колонке с Ni-NTA агарозой, уравновешенной буфером А. Для этого осветленный лизат наносят на колонку и элюируют связавшийся с носителем гибридный белок буфером В (50 мМ трис-HCl, 0,5 М NaCl, 0,5 М имидазол, рН 7,8). Детектирование ведут при длине волны 280 нм. Элюат, содержащий гибридный белок, диализируют относительно воды в течение 16 ч через мембрану с размером пор 12 кДа и лиофильно высушивают.
Очищенный гибридный белок Trx-Aur растворяют в 80% трифторуксусной кислоте в концентрации 10 мг/мл, добавляют равную массу бромциана (1 г бромциана на 1 г белка) и выдерживают при температуре 25°С в защищенном от света месте в течение 16 ч. Реакцию останавливают добавлением пятикратного объема деионизированной воды, после чего образец лиофильно высушивают. Процедуру добавления воды и упаривания повторяют трижды.
Продукты реакции растворяют в 80% трифторуксусной кислоте в концентрации 5%. Нерастворимый осадок отделяют центрифугированием при 10000 g в течение 10 мин и отбрасывают. Разделение продуктов реакции расщепления и очистку аурелина проводят с помощью обращенно-фазовой ВЭЖХ на препаративных колонках в системе ацетонитрил-вода с добавлением 0,1% ТФУ (фиг.3). Детектирование ведут при длине волны 214 нм. Фракцию, содержащую аурелин, собирают и лиофильно высушивают. Выход пептида в пересчете на 1 л клеточной культуры составляет 10 мг.
Для определения N-концевой аминокислотной последовательности аурелина применяют автоматическое микросеквенирование на приборе Precise 491 cLC Protein Sequencing System (PE Applied Biosystems). Идентификацию фенилтиогидантоин-производных аминокислот проводят на анализаторе 120А РТН Analyzer (PE Applied Biosystems). В основе методики автоматического определения аминокислотной последовательности пептидов и белков лежит метод химической деградации полипептидной цепи по методу Эдмана, позволяющий последовательно отщеплять N-концевые аминокислотные остатки в виде фенилтиогидантоинов и идентифицировать отщепленные производные аминокислот методом обращенно-фазовой ВЭЖХ. В результате автоматического микросеквенирования устанавливают идентичность аминокислотных последовательностей генно-инженерного и природного аурелина.
Для определения молекулярной массы очищенного белка используют МАЛДИ-времяпролетный масс-спектрометрический анализ на приборе Reflex III (Bruker Daltonics), оснащенном УФ-лазером с длиной волны 336 нм. В качестве матрицы используют 2,5-дигидроксибензойную кислоту в смеси, содержащей 20% ацетонитрил и 0,1% трифторуксусную кислоту. Полученный пик с m/z 4297,39 (фиг.4) соответствует молекулярному иону природного аурелина.
Пример 4
Тестирование антимикробной активности аурелина.
Антимикробную активность очищенного рекомбинантного и природного пептида определяют методом радиальной диффузии пептида в агарозном геле в отношении тест-культур бактерий: Bacillus megaterium VKM41, Staphylococcus aureus 209p, Micrococcus luteus В 1314. Тест-культуры выращивают на LB-arape (2LB-arape с 1% сахарозы для М. luteus) в течение 16 ч при 37°С. Выросшие колонии переносят в 3 мл жидкой среды Ѕ LB и инкубируют на роторной качалке при 37°С до достижения культурой плотности OD620 1,0-1,5. Аликвоту клеточной суспензии добавляют к 12 мл расплавленной и охлажденной до 40°С обедненной среды (10 мМ фосфатный буфер, рН 7,4, 0,03% TSB, 1% агароза) и полученной смесью заливают чашки диаметром 90 мм. Конечная концентрация бактерий составляет 105 КОЕ/ мл. В слое агарозы высверливают лунки диаметром 2 мм и вносят по 5 мкл раствора образца в 5% ацетонитриле с добавлением 0,1% трифторуксусной кислоты. В качестве контроля используют 5% ацетонитрил с 0,1% трифторуксусной кислоты. Чашки инкубируют, не переворачивая, в течение 3 ч при 37°С, после чего на первый слой агарозы наслаивают 12 мл обогащенной среды (LB-агар или 2LB-arap с 1% сахарозы для M.luteus). Чашки переворачивают и инкубируют в течение 24 ч при 37°С. Антимикробное действие пептида определяют по диаметру зон ингибирования роста тест-культуры. Результаты тестирования активности природного аурелина и пептида, полученного заявленным способом, представлены в таблице.
Показано, что очищенный аурелин в количестве 5 мкг/лунку проявляет антимикробную активность и ингибирует рост всех тестируемых бактерий. Наиболее чувствительной к пептиду культурой является В. megaterium. Активность рекомбинантного аурелина совпадает в пределах погрешности измерения с активностью природного пептида.


Изобретение относится к области биотехнологии, а именно к получению антимикробного пептида аурелина сцифоидной медузы Aurelia aurita. Конструируют плазмидный вектор pE-Trx-Aur для экспрессии аурелина в клетках Escherichia coli в составе гибридного белка Trx-Aur, состоящий из двух фрагментов ДНК, нуклеотидная последовательность которых приведена в описании. Трансформируют данным вектором родительский штамм Escherichia coli BL21(DE3), получая штамм-продуцент гибридного белка Trx-Aur. Для получения пептида аурелина проводят культивирование клеток полученного штамма-продуцента, затем осуществляют лизис клеток, аффинную очистку гибридного белка Trx-Aur на металлохелатном носителе, расщепление гибридного белка Trx-Aur бромцианом по остатку метионина, введенному между последовательностями аурелина и тиоредоксина, и очистку целевого пептида методом обращенно-фазовой ВЭЖХ. Изобретение позволяет получить биологически активный аурелин по упрощенной технологии и без использования труднодоступного природного сырья. 3 н.п. ф-лы, 4 ил., 1 табл.
1. Плазмидный вектор рЕ-Trx-Aur для экспрессии в клетках Escherichia coli антимикробного пептида аурелина в составе гибридного белка Trx-Aur, состоящий из двух фрагментов ДНК:
BglII/XhoI-фрагмента с нуклеотидной последовательностью SEQ ID No. 2, содержащего промотор транскрипции Т7 РНК-полимеразы, lac-оператор, участок связывания рибосомы и участок, кодирующий аффинную метку, белок тиоредоксин и аурелин;
BglII/XhoI-фрагмента плазмиды рЕТ31b(+), содержащего терминатор транскрипции Т7 РНК-полимеразы, сайт инициации репликации, ген β-лактамазы и ген lac-penpeccopa (lacI).
2. Штамм бактерии Escherichia coli BL21(DE3)/pE-Trx-Aur - продуцент гибридного белка Trx-Aur, получаемый путем трансформации клеток родительского штамма BL21(DE3) плазмидным вектором pE-Trx-Aur.
3. Способ получения антимикробного пептида аурелина, включающий экспрессию гибридного белка Trx-Aur в штамме-продуценте Escherichia coli BL21(DE3)/pE-Trx-Aur, лизис клеток, аффинную очистку гибридного белка Trx-Aur на металлохелатном носителе, расщепление гибридного белка Trx-Aur бромцианом по остатку метионина, введенному между последовательностями аурелина и тиоредоксина, и очистку целевого пептида методом обращенно-фазовой ВЭЖХ.
| МАССОВЫЙ РАСХОДОМЕР | 0 |
|
SU231144A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| СПОСОБ ПРОИЗВОДСТВА ПЕКТИНА И ПИЩЕВЫХ ВОЛОКОН ИЗ САХАРНОЙ СВЕКЛЫ | 2004 |
|
RU2261868C1 |
| JP 2008074886 A, 03.04.2008 | |||
| JP 2003321497 A, 11.11.2003. | |||

