Изобретение относится к таксации поваленных модельных деревьев и может быть использовано в природообустройстве и инженерной экологии, экологическом мониторинге и охране окружающей среды путем анализа результатов измерений и испытаний модельных деревьев, причем преимущественно ели и других видов с геометрической формой кроны, в экологической оценке лесных, сельских и городских ландшафтов. Изобретение может также найти применение в лесном хозяйстве при оценке и мониторинге качества структуры кроны у модельных деревьев древостоев.
Известен способ анализа древесного ствола (см., например, учебник: Н.П.Анучин. Лесная таксация. Изд-е 5-е, доп. - М.: Лесная промышленность, 1982. - С.424-426), включающий выбор пробной площади, выбор модельного дерева на пробной площади, описание значений свойств выбранного модельного дерева и места его произрастания, нанесение на ствол отметок о северной и южной сторонах, установление местоположения шейки корня, спиливание дерева около корневой шейки ствола, измерение расстояний на стволе до первого мертвого и первого живого сучков, до начала кроны и других таксационных показателей, разметку ствола на отдельные секции кратной длины (0.5, 1 или 2 м) или некратной длины в зависимости от цели анализа ствола и требуемой точности исследования после первой секции длиной 2,6 м, разметку середин секций, выпиливание кружков начиная от основания ствола к его вершине по серединам секций, причем первый рез делают на месте отметки около корневой шейки, а второй - отступя от него к вершине на толщину кружка, каждому отпиленному кружку придают отметки о номерах пробной площади, модельного дерева и отпиленного кружка, причем обозначения делают на несколько сглаженной верхней части кружков, а нижние части кружков в камеральных условиях используют для подсчета годичных колец и измерения диаметров.
Недостатком этого способа является отсутствие измерений кроны поваленного дерева по структуре ветвей и сучьев (до валки дерева в его кроне были и сучья как омертвевшие ветви). При этом ветви и сучья обрезаются бензомоторными сучкорезами или обрубаются топором заподлицо с поверхностью ствола, что не позволяет проводить измерения в зоне оснований ветвей и сучьев на стволе модельного дерева.
Известен также способ анализа древесного ствола (Пат.2201593 Российская Федерация, МПК7 G01N 33/46, А01G 23/00, А01G 23/02. Способ анализа древесного ствола. / Верхунов П.М., Мазуркин П.М. (РФ); заявитель и патентообладатель Марийск. гос.тех. ун-т.- №2001116223/13; заявл. 13.06.01; опубл. 27.03.2003, Бюл. №5), включающий описание значений свойств выбранного дерева и места его произрастания, нанесение на ствол отметок о северной и южной сторонах, установление местоположения шейки корня, измерение расстояний на стволе от шейки корня до первого мертвого сучка и первой живой ветви до начала кроны, разметку ствола выполняют на секции некратной длины в зависимости от расположения оснований ветвей и сучьев, причем секции некратной длины размечают по крайней мере дважды - вначале по местам наименьших диаметров между основаниями сучьев сваленного модельного дерева, а затем по вершинам этих неровностей с измерением соответствующих расстояний от корневой шейки с оценкой нескольких образующих на неровной поверхности ствола.
Недостатком является то, что прототип не позволяет комплексно измерять, а затем и проводить анализ в камеральных условиях данных измерений, ствол совместно с основаниями ветвей и сучьев модельного дерева. При этом каждый вид дерева имеет уникальную структуру кроны, причем наиболее геометрически и математически правильной является крона ели. Причем ельники с березняками с содержанием ели наиболее распространены в России, поэтому метод модельных деревьев еще долго будет существовать в лесной таксации.
Технический результат - повышение комплексности анализа древесных стволов с основаниями сучьев и ветвей по модельным деревьям.
Этот технический результат достигается тем, что способ анализа ствола модельного дерева, включающий описание значений свойств выбранного дерева и места его произрастания, нанесение на ствол отметок о северной и южной сторонах, установление местоположения шейки корня, измерение расстояний на стволе от шейки корня до первого мертвого сучка и первой живой ветви до начала кроны, разметка ствола на секции некратной длины в зависимости от расположения оснований ветвей и сучьев, причем секции некратной длины размечают по крайней мере дважды - вначале по местам наименьших диаметров между основаниями сучьев сваленного модельного дерева, а затем по вершинам этих неровностей с измерением соответствующих расстояний от корневой шейки с оценкой нескольких образующих на неровной поверхности ствола, отличающийся тем, что до разметки ствола в зоне кроны на секции некратной длины ствол у модельного дерева частично очищают от сучьев с оставлением остатков с основаниями сучьев, после разметки по основаниям остатков сучьев с измерением соответствующих расстояний от корневой шейки подсчитывают количество сучьев на одной высоте ствола, а затем измеряют угол примыкания остатка у каждого сучка и относительно северной стороны определяют азимут расположения каждой бывшей ветви и сучка у росшего до валки модельного дерева, а до разделки ствола на кружки и секции остатки всех сучков удаляют обрубкой топором или обрезкой бензомоторной сучкорезкой, причем данные измерений подвергают в камеральных условиях статистической обработке по высоте ствола дополнительно с оценкой нескольких образующих на неровной поверхности ствола модельного дерева.
Остатки с основаниями сучьев в ходе очистки ствола от них оставляют длиной 5 - 15 см от поверхности ствола, угол примыкания остатка сучка и азимут его расположения на стволе от северного направления измеряют транспортиром с точностью до пяти градусов, а остатки сучков принимаются располагающимися в одной мутовке при разнице их высот от корневой шейки по стволу не более пяти сантиметров.
По количеству остатков сучков в мутовках модельного дерева ели статистическим моделированием выявляют по формуле:
nвм=n1+n2+n3,
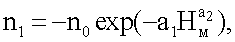
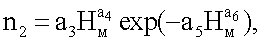
n3=Acos(πHм/p),
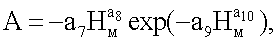
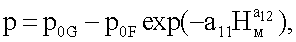
где nвм - количество живых ветвей в каждой мутовке ели, шт.;
n1 - первая составляющая статистической закономерности, показывающая динамику численности мертвых сучьев в растущей кроне, шт.;
n2 - вторая составляющая статистической закономерности, показывающая динамику численности живых ветвей в растущей кроне, шт.;
n3 - третья составляющая статистической закономерности, показывающая волновую динамику колебательной адаптации численности живых ветвей в мутовке растущей кроны к условиям окружающей среды, шт.;
Нм - высота мутовки от корневой шейки до середины мутовки, м;
n0 - ретроспективная численность омертвевших ветвей всей мутовки на уровне корневой шейки у дерева ели, шт.;
А - половина амплитуды колебательного возмущения ели как кризисный отклик (отрицательный знак) в ходе роста на влияние условий (фенотипических факторов окружающей среды) места произрастания с ростом дерева в высоту по мутовкам от корневой шейки, шт.;
р - половина периода колебательного возмущения мутовок по высоте на стволе растущего дерева ели, м;
p0G - половина генетически возможной в семени ели протяженности ствола, по продолжительности жизни живых ветвей, от уровня корневой шейки до первой мутовки, м;
p0F - половина фенотипически переменной (снижающейся с ходом роста дерева в высоту по мутовкам) протяженности ствола, в процессе жизнедеятельности дерева ели по формированию мутовок, от уровня корневой шейки до последней мутовки перед терминальным побегом, м;
a1…a12 - параметры статистической модели, зависящие от хода роста ствола ели по мутовкам кроны по распределениям живых ветвей по отдельным мутовкам, вычисляемые по измеренным данным высоты мутовки по стволу от корневой шейки и численности живых ветвей в мутовках.
Сущность технического решения заключается в том, что ветви вырастают в мутовках не случайным, а вполне определенным образом. Поэтому количество живых ветвей в мутовке вырастает исходя из взаимодействия двух сил: с одной стороны, организм существует и ель живет по генетически заданной и подкрепленной прошлым ростом и развитием кроны, а с другой - противодействие этому напору жизни дерева в ходе его биологического времени со стороны фенотипических факторов среды нарастает и постепенно прекращает дальнейшую генерацию мутовок.
Положительный эффект заключается в том, что этот взаимный процесс, генетически стартовавший от корневой шейки, весьма просто и наглядно проявляется через генерацию мутовок по численности живых ветвей в них. Высота мутовки от корневой шейки является неким условным временем (с помощью спилов можно и узнать возраст появления каждой мутовки). При этом на модельных деревьях это выполнить можно с высокой точностью (измерение высоты с точностью 0,01 м или ±5 мм). На учетных деревьях подсчитать число ветвей в мутовках нелегко, но можно, однако из-за скрытности для геодезических приборов трудно пока реализуемо измерение высоты мутовок по стволу. На практике положительный эффект достигается также и тем, что двухстадийная очистка ствола от сучьев известна и поэтому предлагаемый способ вполне реализуем при массовых измерениях большого числа деревьев ели в процессе проведения лесозаготовительных работ.
Существенной новизной является измерение поверхности ствола по меньшей мере трижды:
во-первых, по снижениям диаметров поперечных сечений ствола для технической оценки стволов модельных деревьев и ельника по объему и приросту объемов древесины, то есть в технической лесной таксации;
во-вторых, по возвышениям диаметров сечений стволов для инженерно-экологической оценки ельника и всей территории, на которой произрастают исследуемые одиночные или групповые деревья ели в парках и культурных ландшафтах;
в-третьих, по возвышениям на мутовках ели с определением углов расположения между ветвями в одной мутовке, а также количества и углов примыкания ветвей во всех мутовках кроны ели.
Существенной новизной становится также и то, что появляется практическая возможность измерения деревьев ели без соотнесения их к какому-то классу роста, что облегчает инженерно-экологические изыскания в ходе заготовки кругляка на лесных участках лесного фонда.
В связи с этим предлагаемое техническое решение обладает существенными признаками новизны, простотой реализации в производственных условиях, положительным эффектом как в лесном хозяйстве и лесозаготовках, так и в инженерной экологии, а также перспективой расширения областей практического применения методологии ландшафтной экологии с растущими на территории этих лесных, агроэкологических, сельских и городских территорий деревьями ели без их разрушения в растущем состоянии. При этом повторные измерения учетных деревьев ели позволят наладить долгосрочный мониторинг окружающей эти учетные деревья ели среды. Хотя при измерениях модельных деревьев образуются только однократные во времени данные, но они дополняют существующую методологию лесной таксации на модельных деревьях ели новым способом измерения кроны по распределениям ветвей в мутовках.
Из научно-технической и патентной литературы материалов, порочащих новизну предлагаемого способа, не обнаружено.
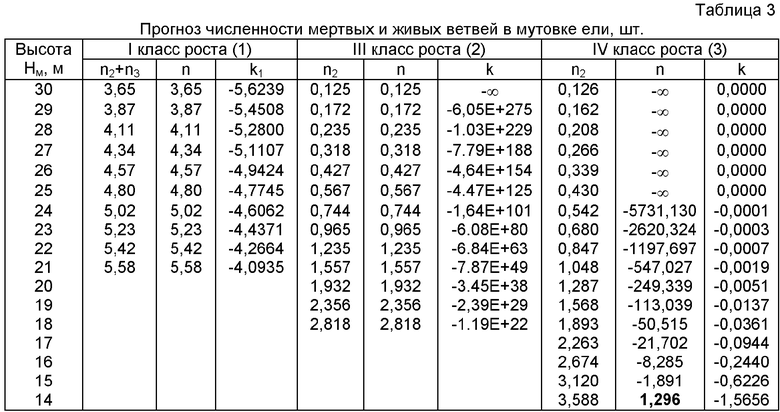
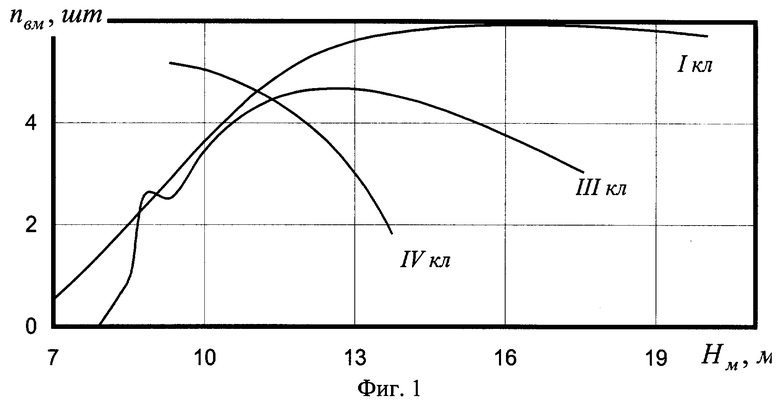
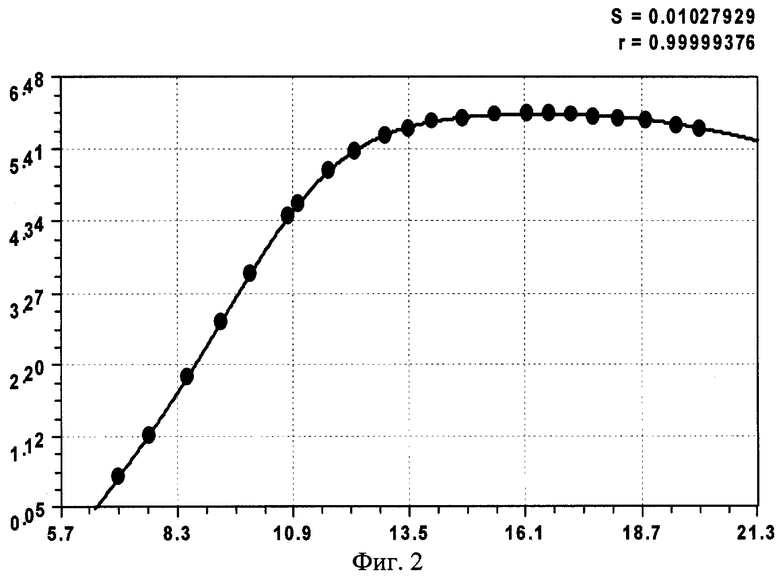
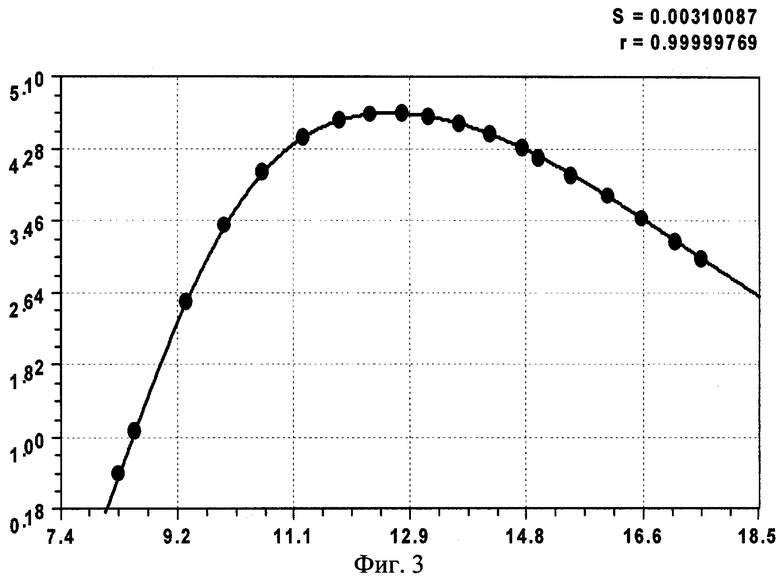
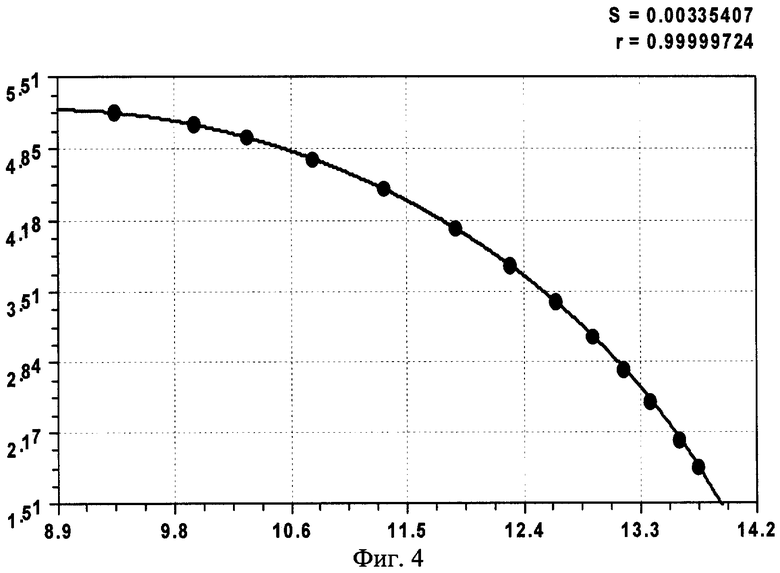
На фиг.1 приведены три графика распределений численности ветвей в мутовках ели различного класса роста, причем заметно, что с ухудшением условия произрастания численность живых ветвей быстрее убывает с высотой ствола; на фиг.2 показан график с коэффициентом корреляции 0,999994 и абсолютной погрешностью менее 0,01 м для деревьев ели первого класса роста; на фиг.3 - то же на фиг.2 для деревьев ели третьего класса роста; на фиг.4 - то же на фиг.2 для множества модельных деревьев ели четвертого класса роста.
Способ анализа ствола модельного дерева, например модельного дерева ели, включает такие действия.
Наиболее простым в реализации предлагаемого способа является измерение модельного дерева ели, срубленного и лежащего на земле. В этом случае используются общеизвестные способы и инструменты таксации модельного дерева мерной лентой. Измерение учетного дерева требует использования новых способов с применением геодезических приборов, но при этом ветви в мутовках затрудняют визирование точек, расположенных на основаниях ветвей.
До разметки ствола в зоне кроны на секции некратной длины ствол у модельного дерева частично очищают (грубая очистка ствола от сучьев). При этом оставляют остатки с основаниями бывших ветвей, то есть сучьев.
Мерной лентой на сваленном модельном дереве с остатками сучьев измеряют высоту дерева от корневой шейки до верхушки терминального побега. Затем в кроновой зоне лежащего ствола прикладывают мерную ленту, начиная от корневой шейки ствола.
После разметки по основаниям остатков сучьев с измерением соответствующих расстояний от корневой шейки подсчитывают количество сучьев на одной высоте ствола. Для многих видов деревьев в мутовке оказывается только по одной ветви.
Затем измеряют угол примыкания остатка у каждого сучка и относительно северной стороны определяют азимут расположения каждой бывшей ветви и сучка у росшего до валки модельного дерева, а затем до разделки ствола на кружки и секции остатки всех сучков удаляют обрубкой топором или обрезкой бензомоторной сучкорезкой.
Данные измерений подвергают в камеральных условиях статистической обработке по высоте ствола дополнительно с оценкой нескольких образующих на неровной поверхности ствола модельного дерева.
Остатки с основаниями сучьев в ходе предварительной очистки ствола от них оставляют длиной 5-15 см от поверхности ствола, угол примыкания остатка сучка и азимут его расположения на стволе от северного направления измеряют транспортиром с точностью до пяти градусов, а остатки сучков принимаются располагающимися в одной мутовке при разнице их высот от корневой шейки по стволу не более пяти сантиметров.
По количеству остатков сучков в мутовках модельного дерева ели статистическим моделированием выявляют по формуле:
nвм=n1+n2+n3,
n3=Acos(πHм/p),
где nвм - количество живых ветвей в каждой мутовке ели, шт.;
n1 - первая составляющая статистической закономерности, показывающая динамику численности мертвых сучьев в растущей кроне, шт.;
n2 - вторая составляющая статистической закономерности, показывающая динамику численности живых ветвей в растущей кроне, шт.;
n3 - третья составляющая статистической закономерности, показывающая волновую динамику колебательной адаптации численности живых ветвей в мутовке растущей кроны к условиям окружающей среды, шт.;
Нм - высота мутовки от корневой шейки до середины мутовки, м;
n0 - ретроспективная численность омертвевших ветвей всей мутовки на уровне корневой шейки у дерева ели, шт.;
А - половина амплитуды колебательного возмущения ели как кризисный отклик (отрицательный знак) в ходе роста на влияние условий (фенотипических факторов окружающей среды) места произрастания с ростом дерева в высоту по мутовкам от корневой шейки, шт.;
р - половина периода колебательного возмущения мутовок по высоте на стволе растущего дерева ели, м;
p0G - половина генетически возможной в семени ели протяженности ствола, по продолжительности жизни живых ветвей, от уровня корневой шейки до первой мутовки, м;
p0F - половина фенотипически переменной (снижающейся с ходом роста дерева в высоту по мутовкам) протяженности ствола, в процессе жизнедеятельности дерева ели по формированию мутовок, от уровня корневой шейки до последней мутовки перед терминальным побегом, м;
a1…a12 - параметры статистической модели, зависящие от хода роста ствола ели по мутовкам кроны по распределениям живых ветвей по отдельным мутовкам, вычисляемые по измеренным данным высоты мутовки по стволу от корневой шейки и численности живых ветвей в мутовках.
Способ анализа ствола модельного дерева реализуется, например, при обработке лесного модельного дерева ели в ельнике, следующим образом.
Вначале выбирают пробную площадку ельника, а затем отбирают модельные деревья ели по их внешним признакам. Выбор модельного дерева и описание его таксационных признаков на пробной площади выполняют по намеченной очередности валки, наносят на ствол отметки о северной и южной сторонах, устанавливают местоположение шейки корня. Измеряют также таксационные показатели кроны.
Спиливание дерева выполняют около корневой шейки ствола. На срубленном стволе измеряют расстояния до первого мертвого и первого живого сучков, до начала кроны и другие таксационные показатели.
Мерной лентой на сваленном модельном дереве с остатками сучьев измеряют высоту дерева от корневой шейки до верхушки терминального побега. Затем в кроновой зоне лежащего ствола прикладывают мерную ленту, начиная от корневой шейки ствола.
После измерения поваленного дерева ели выполняют предварительную обрубку или очистку ствола от сучьев с сохранением его оснований мутовок и терминального побега, а затем размечают ствол на отдельные секции кратной (0.5, 1 или 2 м), кратно-некратной длины (в зависимости от качества формы ствола по отдельным зонам по длине от комля к вершине) или все секции некратной длины (обычно стволы низкокачественных лесных деревьев и одиноко растущих деревьев).
Все измерения записывают в журнал, причем для мутовок показывают высоту их расположения в метрах от корневой шейки ствола с точностью ±0,005 м.
После разметки по основаниям остатков сучьев с измерением соответствующих расстояний от корневой шейки подсчитывают количество сучьев на одной высоте ствола. Для многих видов деревьев в мутовке оказывается только по одной ветви, но при этом значимо различаются параметры каждой ветви по сравнению с деревом ели.
Затем измеряют угол примыкания остатка у каждого сучка и относительно северной стороны определяют азимут расположения каждой бывшей ветви и сучка у росшего до валки модельного дерева, а затем до разделки ствола на кружки и секции остатки всех сучков удаляют обрубкой топором или обрезкой бензомоторной сучкорезкой.
Данные измерений подвергают в камеральных условиях статистической обработке по высоте ствола дополнительно с оценкой нескольких образующих на неровной поверхности ствола модельного дерева.
Остатки с основаниями сучьев в ходе предварительной очистки ствола от них оставляют длиной 5 - 15 см от поверхности ствола, угол примыкания остатка сучка и азимут его расположения на стволе от северного направления измеряют транспортиром с точностью до пяти градусов, а остатки сучков принимаются располагающимися в одной мутовке при разнице их высот от корневой шейки по стволу не более пяти сантиметров.
По количеству остатков сучков в мутовках модельного дерева ели статистическим моделированием выявляют по формуле:
nвм=n1+n2+n3,
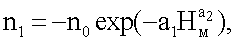
n3=Acos(πHм/p),
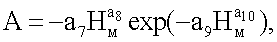
где nвм - количество живых ветвей в каждой мутовке ели, шт.;
n1 - первая составляющая статистической закономерности, показывающая динамику численности мертвых сучьев в растущей кроне, шт.;
n2 - вторая составляющая статистической закономерности, показывающая динамику численности живых ветвей в растущей кроне, шт.;
n3 - третья составляющая статистической закономерности, показывающая волновую динамику колебательной адаптации численности живых ветвей в мутовке растущей кроны к условиям окружающей среды, шт.;
Нм - высота мутовки от корневой шейки до середины мутовки, м;
n0 - ретроспективная численность омертвевших ветвей всей мутовки на
уровне корневой шейки у дерева ели, шт.;
А - половина амплитуды колебательного возмущения ели как кризисный отклик (отрицательный знак) в ходе роста на влияние условий (фенотипических факторов окружающей среды) места произрастания с ростом дерева в высоту по мутовкам от корневой шейки, шт.;
р - половина периода колебательного возмущения мутовок по высоте на стволе растущего дерева ели, м;
p0G - половина генетически возможной в семени ели протяженности ствола, по продолжительности жизни живых ветвей, от уровня корневой шейки до первой мутовки, м;
p0F - половина фенотипически переменной (снижающейся с ходом роста дерева в высоту по мутовкам) протяженности ствола, в процессе жизнедеятельности дерева ели по формированию мутовок, от уровня корневой шейки до последней мутовки перед терминальным побегом, м;
а1...а12 - параметры статистической модели, зависящие от хода роста ствола ели по мутовкам кроны по распределениям живых ветвей по отдельным мутовкам, вычисляемые по измеренным данным высоты мутовки по стволу от корневой шейки и численности живых ветвей в мутовках.
Примеры. В статье (Старостин В.А. О распределении ветвей по толщине в кроне ели в лесных культурах. // Межвуз. сб. научн. тр. «Лесоводство, лесные культуры и почвоведение». - Л.: ЛТА, 1991. - С.105-109) приведены статистические табличные данные по средним значениям параметров живой кроны у деревьев ели разных классов роста.
Экспериментальные данные В.А. Старостина имеют комплексный характер и поэтому позволяют определить закономерности по численности nвм живых ветвей в мутовке некой среднестатистической ели, характеризующей качество роста всего ельника.
За переменную можно принять высоту расположения мутовки ели.
Для мутовок ели получены модели (табл.1 и фиг.1):
- первого класса роста (фиг.2)
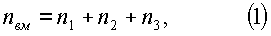
n1=-5,89280exp(-0,012765Hм 1,48765),
n2=0,26007Hм 2,17251exp(-0,19249Hм 0,93010),
n3=Acos(πHм/p),
A=-8,17169·10-12Hм 19,09708exp(-1,53583Hм 1,06652),
p=7,98040-7,81950exp(-0,024989 Hм 1,08556),
- второго класса роста (фиг.3)
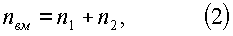
n1=-5,14022exp(-1,2070·10-5Hм 5,28132),
n2=9,81576·10-5Hм 7,03393exp(-0,55931Hм 0,99994);
- третьего класса роста (фиг.4)
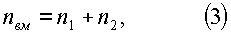
n1=-4,00638·10-5exp(0,78245Hм),
n2=0,10674Hм 3,26919exp(-0,36519Hм).
Сопоставление конструкций выявленных закономерностей показывает, что наиболее сложной и доводимой до полноты структуры является уравнение (1), а остальные две модели являются по конструкции только частными случаями. При этом точность математического описания столь высока (см. коэффициенты корреляции на фиг.2-4 в правом верхнем углу), что абсолютная погрешность: во-первых, не превышает 0,01 м; во-вторых, даже такие малые возмущения имеют волновой характер. Этот факт указывает на то, что распределение численности ветвей в мутовках по высоте дерева ели (на других видах деревьев пока эксперименты не проводились) вполне может стать экологическим критерием, весьма чувствительным к малейшим изменениям в условиях места произрастания, в том числе лесных, сельских и даже городских условиях.
Сравнение трех классов роста у популяций деревьев ели показывает, что максимальная относительная погрешность вышеприведенных формул не превышает 1%. При этом чем хуже условия роста и развития дерева, тем меньше образуется ветвей в мутовках на вершине кроны. При этом с ухудшением условий произрастания повышается высота от корневой шейки до первой живой мутовки ели.
В табл.2 приведены данные по отдельным составляющим полученных статистических закономерностей и приведены значения коэффициента приспособляемости k (k1 и k2 для первого класса роста) и динамичности k∂ поведения мутовки к условиям произрастания на материнском стволе.
Изменение приспособляемости и динамичности живых ветвей в мутовке ели
У дерева ели первого класса возраста наблюдается наиболее сложная динамика роста дерева и развития структуры кроны по мутовкам.
Смертность по численности ветвей в мутовках с высотой убывает по закону гибели (спада), а у деревьев ели четвертого класса возраста, наоборот, смертность мутовок только нарастает в ходе жизнедеятельности ели. Однако быстрое убывание смертности мутовок, как это видно из данных по деревьям ели третьего класса роста, не является благом.
При этом оказывается, что живым ветвям нет противодействия по смертности ветвей в мутовках, поэтому коэффициент приспособляемости по второй составляющей, то есть приспособляемости живых ветвей в мутовках, критически резко нарастает и быстро достигает бесконечности.
Такого биоэнергетического напряжения ветви не выдерживают, и деревья ели третьего класса возраста начинают отставать в росте и развитии с максимума 4,690 шт. ветвей в одной мутовке на высоте 12,30 м.
Деревья ели первого класса роста чувствуют себя спокойно, так как отношение
k1=n2/n1 у них растет медленно. Даже на последней мутовке из живых ветвей численностью 7,677 шт. не появились только 1,958 шт. мертвых в зародышах почек ветвей, фактически образуя мутовку численностью в 5,73 шт. с произрастающими на высоте 20,02 м ствола ветвями. При этом адаптация среднего дерева происходила по колебательному возмущению с двумя максимумами. Поэтому целью дальнейших исследований является изучение проявлений кризисных колебаний в организме дерева, которые должны, по-видимому, совпасть с колебаниями свойств окружающей растущее дерево среды.
А у деревьев ели четвертого класса роста максимальная напряженность уже была на высоте 9,32 м, когда в момент измерений там находилась первая мутовка с живыми ветвями в начале кроны.
Еще яснее картина динамики проявляется с прогнозами в росте измеренных деревьев ели (табл.3) по полученным закономерностям.
Нижние мутовки ели имеют живые и отмершие ветви. Самоочищением дерево сбрасывает неживые ветви, образуя отпад. При этом плотность расположения ветвей в одной мутовке зависит от двух противоположно направленных процессов. Первый из них по естественной составляющей закономерности является биотехническим возбуждением дерева на рост новых живых мутовок. Вторая составляющая для деревьев высоких классов роста является законом гибели мертвых сучьев.
Для деревьев первого класса роста появилась и третья, хотя и малая по биоэнергетике, но решающая в достижении долголетия. Поэтому из табл.3 видно, что ель могла бы достичь высоты от 20,02 м относительно легко, сохраняя и на дальнейшее способность генерирования живых ветвей в последующих мутовках. Хотя напряжение по генерированию мутовок по табл. 3 нарастает до коэффициента приспособляемости 5,6239, однако это гораздо меньше напряжения 7,42 у третьей снизу мутовки для деревьев ели второго класса роста.
Как видно из формулы (3), для слабо растущих деревьев ели мертвые сучья увеличивают свою популяцию в мутовке по закону экспоненциального роста. Тем самым «влияние смерти» сказывается на всей высоте ослабленного дерева, а на вновь появляющихся живых ветвях лежит «печать смерти» у тех материнских деревьев, которые значительно отстали в росте. Поэтому по табл.3 дается прогноз, что деревья ели четвертого класса роста не поднимутся выше 14 м высоты.
Предлагаемый способ применим не только к популяциям деревьев, но и к отдельным деревьям.
Для них всех общая закономерность будет иметь вид уравнения
nвм=n1+n2+n3,
n3=Acos(πHм/p),
где nвм - количество живых ветвей в каждой мутовке ели, шт.;
n1 - первая составляющая статистической закономерности, показывающая динамику численности мертвых сучьев в растущей кроне, шт.;
n2 - вторая составляющая статистической закономерности, показывающая динамику численности живых ветвей в растущей кроне, шт.;
n3 - третья составляющая статистической закономерности, показывающая волновую динамику колебательной адаптации численности живых ветвей в мутовке растущей кроны к условиям окружающей среды, шт.;
Нм - высота мутовки от корневой шейки до середины мутовки, м;
n0 - ретроспективная численность омертвевших ветвей всей мутовки на уровне корневой шейки у дерева ели, шт.;
А - половина амплитуды колебательного возмущения ели как кризисный отклик (отрицательный знак) в ходе роста на влияние условий (фенотипических факторов окружающей среды) места произрастания с ростом дерева в высоту по мутовкам от корневой шейки, шт.;
p - половина периода колебательного возмущения мутовок по высоте на стволе растущего дерева ели, м;
p0G - половина генетически возможной в семени ели протяженности ствола, по продолжительности жизни живых ветвей, от уровня корневой шейки до первой мутовки, м;
p0F - половина фенотипически переменной (снижающейся с ходом роста дерева в высоту по мутовкам) протяженности ствола, в процессе жизнедеятельности дерева ели по формированию мутовок, от уровня корневой шейки до последней мутовки перед терминальным побегом, м;
а1…а12 - параметры статистической модели, зависящие от хода роста ствола ели по мутовкам кроны по распределениям живых ветвей по отдельным мутовкам, вычисляемые по измеренным данным высоты мутовки по стволу от корневой шейки и численности живых ветвей в мутовках.
Эффективность нового способа проявляется в том, что он позволяет анализировать модельные деревья комплексно по стволам и мутовкам кроны. По вырубленным ельникам можно будет судить о прошлом поведении деревьев, прежде всего по деревьям ели из сложного древостоя.
Наибольший эффект будет достигнут многократными измерениями мутовок ели на учетных молодых деревьях, которые будут доступны измерениям мутовок мерной лентой и расположены в различных условиях произрастания в естественных и искусственных (плантации) условиях.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ АНАЛИЗА СТВОЛА И КРОНЫ ДЕРЕВА ЕЛИ | 2008 |
|
RU2376749C1 |
| СПОСОБ АНАЛИЗА ВЕТВЕЙ КРОНЫ ДЕРЕВА ЕЛИ | 2011 |
|
RU2495417C2 |
| СПОСОБ АНАЛИЗА ДРЕВЕСНОГО СТВОЛА | 2001 |
|
RU2201593C2 |
| СПОСОБ АНАЛИЗА СПИЛОВ ДРЕВЕСНОГО СТВОЛА | 2003 |
|
RU2265841C2 |
| СПОСОБ АНАЛИЗА КРОНЫ УЧЕТНОЙ ЕЛИ ПО ИСПЫТАНИЯМ ХВОИНОК ГОДИЧНЫХ ВЕТОЧЕК | 2011 |
|
RU2489717C2 |
| СПОСОБ АНАЛИЗА КРОНЫ И МЕСТА ПРОИЗРАСТАНИЯ БЕРЕЗЫ НА СКЛОНЕ ОВРАГА | 2012 |
|
RU2525262C2 |
| СПОСОБ ФОРМИРОВАНИЯ РЕЗОНАНСНОЙ ДРЕВЕСИНЫ ЕЛИ ЕВРОПЕЙСКОЙ | 2016 |
|
RU2644585C2 |
| Способ сбора почек в хвойных молодняках | 1983 |
|
SU1335180A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ХАРАКТЕРА ИЗМЕНЕНИЯ ШИРИНЫ ВОДОПРОВОДЯЩЕГО СЛОЯ НА СПИЛЕ ДЕРЕВА В ЗАВИСИМОСТИ ОТ ПРЕОБЛАДАЮЩИХ ВЕТРОВ | 2006 |
|
RU2331877C2 |
| СПОСОБ ОТБОРА ОБРАЗЦОВ ДРЕВЕСИНЫ ДЛЯ ИЗУЧЕНИЯ СЕЗОННОГО РОСТА КУСТАРНИКОВ В ВЫСОКОГОРЬЯХ | 2022 |
|
RU2797019C1 |
Способ включает описание значений свойств выбранного дерева и места его произрастания. На ствол наносят отметки о северной и южной сторонах, устанавливают местоположение шейки корня, измеряют расстояния на стволе от шейки корня до первого мертвого сучка и первой живой ветви до начала кроны. В зоне кроны ствол у модельного дерева частично очищают от сучьев с оставлением остатков с основаниями сучьев. Затем осуществляют разметку ствола на секции некратной длины в зависимости от расположения оснований ветвей и сучьев. Секции некратной длины размечают по крайней мере дважды - вначале по местам наименьших диаметров между основаниями сучьев сваленного модельного дерева, а затем по вершинам этих неровностей с измерением соответствующих расстояний от корневой шейки с оценкой нескольких образующих на неровной поверхности ствола. После разметки по основаниям остатков сучьев с измерением соответствующих расстояний от корневой шейки подсчитывают количество сучьев на одной высоте ствола. Затем измеряют угол примыкания остатка у каждого сучка и относительно северной стороны определяют азимут расположения каждой бывшей ветви и сучка у росшего до валки модельного дерева. До разделки ствола на кружки и секции остатки всех сучков удаляют обрубкой топором или обрезкой бензомоторной сучкорезкой. Данные измерений подвергают в камеральных условиях статистической обработке по высоте ствола дополнительно с оценкой нескольких образующих на неровной поверхности ствола модельного дерева. Способ обеспечивает повышение комплексности анализа древесных стволов с основаниями сучьев и ветвей по модельным деревьям. 2 з.п. ф-лы, 4 ил.
1. Способ анализа ствола модельного дерева, включающий описание значений свойств выбранного дерева и места его произрастания, нанесение на ствол отметок о северной и южной сторонах, установление местоположения шейки корня, измерение расстояний на стволе от шейки корня до первого мертвого сучка и первой живой ветви до начала кроны, разметка ствола на секции некратной длины в зависимости от расположения оснований ветвей и сучьев, причем секции некратной длины размечают по крайней мере дважды - вначале по местам наименьших диаметров между основаниями сучьев сваленного модельного дерева, а затем по вершинам этих неровностей с измерением соответствующих расстояний от корневой шейки с оценкой нескольких образующих на неровной поверхности ствола, отличающийся тем, что до разметки ствола в зоне кроны на секции некратной длины ствол у модельного дерева частично очищают от сучьев с оставлением остатков с основаниями сучьев, после разметки по основаниям остатков сучьев с измерением соответствующих расстояний от корневой шейки подсчитывают количество сучьев на одной высоте ствола, а затем измеряют угол примыкания остатка у каждого сучка и относительно северной стороны определяют азимут расположения каждой бывшей ветви и сучка у росшего до валки модельного дерева, а до разделки ствола на кружки и секции остатки всех сучков удаляют обрубкой топором или обрезкой бензомоторной сучкорезкой, причем данные измерений подвергают в камеральных условиях статистической обработке по высоте ствола дополнительно с оценкой нескольких образующих на неровной поверхности ствола модельного дерева.
2. Способ анализа ствола модельного дерева по п.1, отличающийся тем, что остатки с основаниями сучьев в ходе очистки ствола от них оставляют длиной 5-15 см от поверхности ствола, угол примыкания остатка сучка и азимут его расположения на стволе от северного направления измеряют транспортиром с точностью до 5°, а остатки сучков принимаются располагающимися в одной мутовке при разнице их высот от корневой шейки по стволу не более 5 см.
3. Способ анализа ствола модельного дерева по п.1, отличающийся тем, что по количеству остатков сучков в мутовках модельного дерева ели статистическим моделированием выявляют по формуле:
nвм=n1+n2+n3
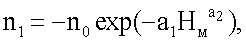
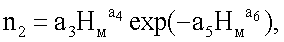
n3=Acos(πHм/p),
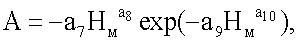
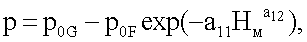
где nвм - количество живых ветвей в каждой мутовке ели, шт.;
n1 - первая составляющая статистической закономерности, показывающая динамику численности мертвых сучьев в растущей кроне, шт.;
n2 - вторая составляющая статистической закономерности, показывающая динамику численности живых ветвей в растущей кроне, шт.;
n3 - третья составляющая статистической закономерности, показывающая волновую динамику колебательной адаптации численности живых ветвей в мутовке растущей кроны к условиям окружающей среды, шт.;
Нм - высота мутовки от корневой шейки до середины мутовки, м;
n0 - ретроспективная численность омертвевших ветвей всей мутовки на уровне корневой шейки у дерева ели, шт.;
А - половина амплитуды колебательного возмущения ели как кризисный отклик (отрицательный знак) в ходе роста на влияние условий (фенотипических факторов окружающей среды) места произрастания, с ростом дерева в высоту по мутовкам от корневой шейки, шт.;
р - половина периода колебательного возмущения мутовок по высоте на стволе растущего дерева ели, м;
р0G - половина генетически возможной в семени ели протяженности ствола, по продолжительности жизни живых ветвей, от уровня корневой шейки до первой мутовки, м;
p0F - половина фенотипически переменной (снижающейся с ходом роста дерева в высоту по мутовкам) протяженности ствола, в процессе жизнедеятельности дерева ели по формированию мутовок, от уровня корневой шейки до последней мутовки перед терминальным побегом, м;
a1…a12 - параметры статистической модели, зависящие от хода роста ствола ели по мутовкам кроны по распределениям живых ветвей по отдельным мутовкам, вычисляемые по измеренным данным высоты мутовки по стволу от корневой шейки и численности живых ветвей в мутовках.
| СПОСОБ АНАЛИЗА ДРЕВЕСНОГО СТВОЛА | 2001 |
|
RU2201593C2 |
| СПОСОБ ИЗМЕРЕНИЯ УЧЕТНОГО ДЕРЕВА | 2002 |
|
RU2224418C1 |
| АНУЧИН Н.П | |||
| Лесная таксация | |||
| - М.: Гослесбумиздат, 1960, с.380-390 | |||
| ТЕСЛЮК Н.К | |||
| Методы измерительной таксации | |||
| - М.: Лесная промышленность, 1978, с.101-110. | |||

