ОПИСАНИЕ
Область техники
Настоящее изобретение относится к вакцине на основе В-клеток, нагруженной лигандом Т-клеток-природных киллеров и антигеном для профилактики и лечения инфекционных заболеваний и злокачественных заболеваний, более конкретно к иммунотерапевтической и профилактической вакцине, действие которой опосредовано В-клетками, нагруженными α-галактозилцерамидом, разновидностью гликолипида, который вместе с антигеном может стимулировать Т-клетки-природные киллеры.
Предшествующий уровень техники
В основном опухолевые антигены не могут быть эффективно представлены антиген-представляющими клетками, то есть иммунный ответ не может быть эффективно индуцирован. Противоопухолевая вакцина представляет собой новую терапевтическую вакцину, которая характеризуется активацией опухолеспецифической иммунной системы (например, введение опухолевых антигенов в антиген-представляющие клетки) для индукции мощного иммунного ответа для разрушения злокачественных клеток.
Из доступных подходов к вакцинации известно, что клеточные вакцины с использованием антиген-представляющих клеток (АПК), таких как дендритные клетки (ДК), являются надежными для выработки эффективного Т-клеточного иммунитета (Rosenberg, S. A. et al., Nat. Med., 10, 909-915, 2004). Поскольку было показано, в частности, что вакцины на основе ДК эффективно индуцируют Aг-специфические эффекторные Т-клетки и Т-клетки памяти, они были предложены для противоопухолевой иммунотерапии в ряде клинических испытаний (Rosenberg, S. A. et al., Nat. Med., 10, 909-915, 2004). Дендритные клетки являются идеальными антиген-представляющими клетками (АПК) для иммунотерапии, поскольку они могут захватывать Аг, а затем мигрировать в лимфоидные органы, где они представляют этот Аг соответствующим Т-клеткам. Более важно то, что они обеспечивают мощную ко-стимуляцию Т-клеток (Figdor, C. G. et al., Nat. Med., 10, 475-480, 2004; и Banchereau, J. et al., Cell, 106, 271-274, 2001). Идея ДК-вакцины является традиционной как для экспериментальных, так и для клинических исследований. Однако ДК сравнительно редки в крови и лимфоидных тканях и увеличить их численность ex vivo из моноцитов крови затруднительно, и то и другое являются основными препятствиями для их широкого применения в вакцинах (Schultze, J. L. et al., Trends Immunol., 25, 659-664, 2004).
В-клетки представляются перспективным альтернативным источником для клеточных вакцин в силу их изобилия в лимфоидных тканях и крови и легкого способа размножения ex vivo (Schultze, J. L. et al., Trends Immunol., 25, 659-664, 2004; von Bergwelt-Baildon, M.S. et al., Blood, 99, 3319-3325, 2002; и Schultze, J. L. et al., J. Clin. Invest., 100, 2757-2757, 1997), а также в силу того, что они собираются в лимфоидных органах после их паретнерального введения.
Несмотря на эти преимущества, В-клетки игнорируются в качестве источника клеточных вакцин, поскольку они слабо иммуногенны. Действительно, накопленные доказательства показывают, что они непосредственно индуцируют иммунологическую толерантность и у CD4 и у CD8 Т-клеток, вероятно, вследствие отсутствия ко-стимуляции (Bennett, S. R. et al., J. Exp. Med., 188, 1977-1983, 1998; и Eynon, E. E. et al., J. Exp. Med., 175, 131-138, 1992). Однако «активированные» В-клетки могут примировать как CD4, так и CD8 Т-клетки (von Bergwelt-Baildon, M. S. et al., Blood, 99, 3319-3325, 2002; Schultze, J. L. et al., J. Clin. Invest., 100, 2757-2765, 1997; Lapointe, R. et al., Cancer Res., 63, 2836-2843, 2003; и Heit, A. et al., J. Immunol., 172, 1501-1507, 2004), давая возможность предположить, что при активизации соответствующими стимулами В-клетки могут действовать как иммуногенные АПК, способные индуцировать Аг-специфический Т-клеточный иммунитет.
Точно установлено, что iNKТ-клетки играют ключевую роль в разнообразии иммунных ответов и в иммунопатологии в целом. Хотя они представляют менее 1% лимфоцитов у мышей, iNKТ-клетки направляют ответ на собственные и экзогенные Аг и определяют индукцию толерантности или иммунитета (Kronenberg, M., Annu. Rev. Immunol., 23, 877-900, 2005; и Park, S. H. & Bendelac, A., Nature, 406, 788-792, 2000). Они действуют как суппрессоры иммунитета при опухолях, диабете и в иммунологически привилегированном участке (Sonoda, K. H., et al., J. Exp. Med., 190, 1215-1226, 1999).
Напротив, опосредованная лигандом активация iNKТ-клеток приводит к активации T-, B- и NK-клеток, а также ДК. Инъекция αGalCer, лиганда iNKT генерирует противоопухолевый иммунитет посредством NK- и Т-клеток (Moodycliffe, A. M., et al., Nat. Immunol., 1, 521-525, 2000).
Альфа-галактозилцерамид (αGalCer) представляет собой разновидность гликолипида, экстрагированного из морской губки, который является лигандом Т-клеток-природных киллеров, имеющих Т-клеточный рецептор (TCR) Vα14+, и представленный как CD1d на антиген-представляющей клетке (АПК) (Kawano et al., Science, 278: 1626, 1997). Активация Т-клеток-природных киллеров приводит к масс-продукции IFN-γ и IL-4, которые могут контролировать иммунные ответы либо на инфекционное заболевание либо на злокачественное заболевание (Chen et al., J. Immunol., 159: 2240, 1997; Wilson et al., Proc. Natl. Acad. Sci. U.S.A., 100: 10913, 2003).
У мышей, которым совместно вводили белковый Аг и αGalCer, развивается гуморальный и клеточно-опосредованный иммунитет, в том числе цитотоксические Т-клеточные ответы (Hermans, I. F., et al., J. Immunol, 171, 5140-5147, 2003; и Stober, D. et al., J. Immunol., 170, 2540-2548, 2003). Более того, недавнее исследование показало, что ДК, нагруженные αGalCer, генерируют более продолжительные iNKТ-клеточные ответы, чем свободная форма αGalCer, что позволяет предположить, что адъювантность iNKT-лигандов могла бы быть усилена, если ее направить на специальные АПК.
Авторы настоящего изобретения подтвердили, что представление iNKT-лиганда на В-клетках могло бы перевести их из толерогенных в иммуногенные, тем самым генерируя мощный иммунитет против Аг, представленного на молекулах MHC В-клеток. Для верификации этого подтверждения и таким образом для осуществления этого изобретения авторы настоящего изобретения дополнительно подтвердили эффективность αGalCer-нагруженных, сенсибилизированных пептидом В-клеток и В-клеток, нагруженных αGalCer, трансдуцированных аденовирусом, в выработке антиген-специфического иммунитета и противоопухолевой активности.
Техническая задача
Задачей настоящего изобретения является предоставление иммунотерапевтической/профилактической вакцины и противоопухолевой вакцины, способной индуцировать опосредованные антиген-представляющими В-клетками антиген-специфические иммунные реакции. Т-клетки-природные киллеры, стимулированные αGalCer на молекуле CD1d нагруженных антигеном В-клеток, переводят толерогенные В-клетки в иммуногенные антиген-представляющие клетки.
Техническое решение
Настоящее изобретение относится к иммунотерапевтической/профилактической вакцине и противоопухолевой вакцине, действие которой опосредовано В-клетками, нагруженными лигандом Т-клеток-природных киллеров и антигеном.
Настоящее изобретение относится также к противоопухолевой вакцине, действие которой опосредовано В-клетками, нагруженными лигандом Т-клеток-природных киллеров и экспрессирующими опухолевый антиген.
Настоящее изобретение дополнительно относится к активатору Т-клеток-природных киллеров, опосредованному В-клетками, нагруженными αGalCer.
Кроме того, настоящее изобретение относится к индуктору цитотоксического ответа, опосредуемого В-клетками, экспрессирующими опухолевый антиген.
Далее настоящее изобретение описывается подробно.
Настоящее изобретение относится к иммунотерапевтической/профилактической вакцине и противоопухолевой вакцине, содержащей фармацевтически эффективное количество В-клеток, выбранных из группы, состоящей из a) CD19+ В-клеток, полученных нагрузкой их лигандом Т-клеток-природных киллеров, b) CD19+ В-клеток, полученных путем их нагрузки лигандом и сенсибилизации антигеном, и c) CD19+ В-клеток, полученных путем их нагрузки лигандом и переносом гена, кодирующего пептидный антиген, посредством чего CD19+ В-клетки экспрессируют антиген, и фармацевтически приемлемый носитель.
Лиганд Т-клеток-природных киллеров включает в себя альфа-галактуронозилцерамид и альфа-глюкуронозилцерамид, происходящие из Sphingomonas spp. (Mattner, J. et al. Nature 434:525, 2005, Kinjo, Y. et al. Nature 434:520, 2005), фосфатидилинозитолтетраманнозид, происходящий из M. tuberculosis (Fischer, K. et al. PNAS 101:10685, 2004), аутоантигены изоглоботригексозилцерамид (Zhou, D. et al. Science 306:1786, 2004) и ганглиозид GD3 (Wu, D. Y. et al. J. Exp. Med. 198:173, 2003), фосфатидилхолин (J. Immunol. 175:977, 2005), бета-галактозилцерамид (β GalCer, Ortaldo JR et al. J. Immunol. 172:943), липофосфогликан и гликоинозитол фосфолипиды гликозидной связи на поверхности лейшманий (J. Exp. Med. 200:895, 2004), αGalCer производные бета-аномерного GalCer и альфа-аномерного GalCer (J. Immunol. 173:3693, 2004), варианты αGalCer (J. Am. Chem. Soc. 126:13602, 2004) и бактериальный липидный антиген, такой как мономиколят глюкозы, происходящий из Nocardia falcinica (Moody, D. B. et al. J. Exp. Med. 192:965, 2000).
Поскольку уже точно установлено, что ДК, нагруженные αGalCer, активируют iNKТ-клетки (van der Vliet HJ, et al., J Immunol Methods., 247(1-2): 61-72, 2001), авторы настоящего изобретения исследовали вопрос о том, будут ли В-клетки, нагруженные αGalCer, оказывать подобное воздействие.
ДК и CD19+ В-клетки (Фиг. 1) выделяли у мыши. Каждую клетку сенсибилизировали различными концентрациями αGalCer, затем совместно культивировали с NKT гибридомой. Затем измеряли уровень IL-2 в культуральном супернатанте. В результате и В-клетки, нагруженные αGalCer (B/αGalCer), и ДК, нагруженные αGalCer (ДК/αGalCer), стимулировали продукцию IL-2 NKT гибридомой (см. Фиг. 30). Следовательно, это подтверждает, что и B/αGalCer, и ДК/αGalCer могут активировать iNKТ-клетки ex vivo.
Для изучения того, могут ли B/αGalCer и ДК/αGalCer, введенные in vivo, активировать иммунные клетки или нет, авторы настоящего изобретения вводили B/αGalCer или ДК/αGalCer от мышей C57BL/6 внутривенно сингенным мышам и измеряли уровень клеток, продуцирующих IL-4 и IFN-γ, c помощью ELISPOT. В частности, мышам C57BL/6 дикого типа или Jα281-/- внутривенно вводили носитель, αGalCer, В-клетки, нагруженные αGalCer, или ДК, нагруженные αGalCer. Неделю спустя клетки селезенки отделяли для получения отдельных клеток, которые помещали на планшет ELISPOT. Носитель или αGalCer добавляли к планшету, с последующим культивированием в течение 6 часов для стимуляции. Исследование ELISPOT проводили для определения клеток, секретирующих IL-4 и IFN-γ, в соответствии с инструкциями производителя (набор IL-4 ELISPOT, набор IFN-γ ELISPOT; R&D system).
В результате инъекция и B/αGalCer и ДК/αGalCer индуцировала высокие количества клеток, продуцирующих IL-4 и IFN-γ (Фиг. 2). Однако следует отметить, что значительное число этих цитокин-продуцирующих клеток выявлялось даже при отсутствии повторной стимуляции αGalCer. Тем не менее, авторы настоящего изобретения полагают, что индукция клеток, продуцирующих IL-4 и IFN-γ, зависит от iNKТ-клеток, поскольку этого не происходило у мышей Jα281-/-, котороые лишены популяции iNKT (см. Фиг. 2).
Авторы настоящего изобретения внутривенно вводили B/αGalCer, меченные CFSE (сложным эфиром сукцинимидила и карбоксифлуоресцеина) нативным мышам, а через 24 или 48 часов после введения анализировали молекулы ко-стимуляции на клетках CFSE+ методом проточной цитометрии для определения того, какие изменения, если они есть, были индуцированы в В-клетках, нагруженных αGalCer, после введения. В результате высокие уровни экспрессии CD86, а не CD80 были индуцированы в течение 24 часов (Фиг. 3). CD40 и MHC II также слегка активировались. В заключение in vivo введение B/αGalCer индуцировало активацию В-клеток в течение 24 или 48 часов.
Далее, авторы настоящего изобретения изучили то, может ли совместная сенсибилизация αGalCer и MHC I-рестриктированного пептида на В-клетках примировать пептид-специфические CD8+ Т-клетки. С этой целью авторы изобретения впервые осуществили адоптивный перенос CFSE-меченых OVA-специфических CD8+ Т-клеток мышам, затем ввели сенсибилизированные наполнителем В-клетки (одни B), αGalCer-сенсибилизированные В-клетки (B/αGalCer), наполнитель + В-клетки, сенсибилизированные пептидом (B/pep), или αGalCer + В-клетки, сенсибилизированные пептидом (B/αGalCer/pep). Затем были получены лимфоциты из селезенки и лимфатических узлов, с последующим измерением CD8+ Т-клеточного ответа у мышей, с перенесенными В-клетками. В результате небольшое деление OVA-специфических CD8+ Т-клеток было индуцировано у мышей, получавших только В; однако слабое деление OVA-специфичных CD8+ Т-клеток, возможно, Аг-неспецифического происхождения, было отмечено в группе B/αGalCer. Введение B/pep индуцировало значительное деление OVA-специфических CD8+ Т-клеток. А у мышей, получивших B/αGalCer/pep, было показано усиленное деление CD8+ Т-клеток, с более чем 40% полученных в результате клеток, продуцирующих IL-2, и наиболее неожиданно более чем 90% полученных в результате клеток, продуцирующих IFN-γ на значительно более высоких уровнях, чем группа B/pep (см. Фиг. 4). Эти результаты дают возможность предположить, что значительно большая скорость CD8+ Т-клеточной активации могла быть достигнута посредством нагрузки αGalCer на B/pep.
Авторы настоящего изобретения исследовали вопрос о том, мог ли подход к вакцинам на основе В-клеток индуцировать цитотоксический иммунитет. В частности, авторы изобретения вводили внутривенно группам мышей C57BL/6 одни B, B/αGalCer, B/pep или B/αGalCer/pep, а затем определяли in vivo активность CTL. В результате только B/αGalCer/pep полностью лизировали сенсибилизированные пептидом мишени (Фиг. 5). А в группе, обработанной B/αGalCer/pep, было показано значительное увеличение числа IFN-γ-продуцирующих CD8+ Т-клеток в отношении этого пептида (Фиг. 6, слева). При проведении последующей иммунизации с использованием OVA-покрытых сингенных спленоцитов для определения реактивности CTL мыши, которым вводили одни B или B/αGalCer, реагировали нормально на прайминг и генерировали значительное количество пептид-специфических IFN-γ-продуцирующих CD8+ Т-клеток (Фиг. 6, справа). Однако когда спленоциты от мышей, иммунизированных B/pep, повторно стимулировали родственным пептидом in vitro, число IFN-γ-продуцирующих CD8+ Т-клеток снижалось по сравнению с мышами, которым вводили одни В, что дает возможность предположить, что те мыши были толерантны к этому пептиду. Напротив, у мышей, вакцинированных B/αGalCer/pep, обнаруживалось гораздо большее число пептид-реактивных CD8+ Т-клеток, чем обнаруживалось либо в группе, получающей B/αGalCer или одни B, что предполагает, что это вторичный иммунный ответ.
Авторы изобретения сравнили эффективность стратегии вакцин на основе В-клеток в генерировании цитотоксичности с ДК-вакцинами. С этой целью определяли минимальное число клеток, необходимое для достижения полного лизиса клеток-мишеней in vivo. Принимая во внимание то, что площадь поверхности ДК гораздо больше, чем площадь поверхности В-клеток, можно сделать заключение, что B/αGalCer/pep были столь же эффективны, как и ДК/αGalCer/pep, в генерировании цитотоксичности. В то же время профиль in vivo цитотоксичности группы, обработанной ДК/pep, был очень похож на таковой группы, обработанной ДК/αGalCer/pep, что указывает на то, что нагрузка αGalCer на ДК дополнительно не усиливала вакцинную эффективность ДК/pep.
Авторы изобретения далее изучали, какие типы иммунных клеток вовлечены в выработку CTL-ответа. С этой целью мышам вводили элиминирующие антитела за 4 дня до или 4 дня после их вакцинации B/αGalCer/pep, а затем проводили исследование CTL in vivo. В результате выработки активности CTL не препятствовали ни элиминация CD4+, ни элиминация NK1.1+ клеток, независимо от сроков проведения элиминации. С другой стороны, элиминация CD8 полностью блокировала уничножение клеток-мишеней (Фиг. 9).
Можно было бы утверждать, что сенсибилизированные пептидом В-клетки действуют не как АПК, а как резервуары пептида, из которого ДК хозяина извлекают пептиды для индукции CTL-ответов. Для исследования этой возможности использовали мышей bm-1. Клетки этих мышей могут нести OVA-пептид на их молекулах MHC класса I, но полученный в результате комплекс не распознается когнитивными CD8+ Т-клетками вследствие мутации в области H-2K. В-клетки этой линии экспрессировали CD1d нормально и стимулировали активацию iNKT в ответ на αGalCer, что указывает на то, что взаимодействие между B и iNKT было интактным. Еще раз, когда B/αGalCer/pep получали с использованием В-клеток мышей дикого типа, а затем их вводили мышам недикого типа, генерировалась полная in vivo OVA-специфическая цитотоксичность. Однако когда B/αGalCer/pep, полученный из В-клеток мышей bm-1, вводили мышам недикого типа, генерировать OVA-специфическую цитолитическую активность не удавалось, что дает возможность предположить, что ДК или другие специфические АПК у мышей реципиентов не отвечают за выработку CTL (Фиг. 11).
Авторы изобретения далее изучали возможность генерирования CTL в тех случаях, когда αGalCer и пептид были сенсибилизированы по отдельности, а затем введены вместе. В результате у мышей, вакцинированных с использованием 'B/αGalCer плюс B/pep', не вырабатывалась in vivo цитотоксичность (Фиг. 12). Этот результат демонстрирует, что пептид и αGalCer должны быть представлены на той же В-клетке для генерирования OVA-специфической цитотоксичности.
Авторы изобретения исследовали также то, будет ли вакцинация с использованием B/αGalCer/pep генерировать противоопухолевый иммунитет. Для тестирования профилактической противоопухолевой активности группы мышей вакцинировали однократно, используя одних только B, B/αGalCer, B/pep, B/αGalCer/pep, ДК/pep или ДК/αGalCer/pep до подкожной трансплантации им меланомы, трансфицированной OVA. В результате наблюдался слегка замедленный профиль опухолевого роста у мышей, вакцинированных B/αGalCer, хотя в конце концов у всех мышей развивались опухоли. Напротив, у мышей, получавших B/αGalCer/pep, ДК/pep или ДК/αGalCer/pep, опухоли не развивались (см. Фиг. 13). Для изучения того, установилась ли у этих мышей длительная противоопухолевая активность, повторно заражали выживших мышей без опухоли теми же опухолевыми клетками через 70 дней после первого заражения опухолью. Авторы изобретения не наблюдали опухолевый рост у тех мышей, демонстрируя, что вакцинация с использованием B/αGalCer/pep создавала противоопухолевый иммунитет памяти (см. Фиг. 14).
Далее, авторы изобретения изучали вопрос о том, будет ли вакцинация с использованием B/αGalCer/pep уничтожать уже существующие опухоли. В результате опухолевый рост полностью подавлялся у мышей, вакцинированных с использованием ДК/pep, ДК/αGalCer/pep или B/αGalCer/pep (см. Фиг. 15 и Фиг. 16).
Для определения того, может ли эта схема введения вакцины на основе В-клеток применяться для конкретного опухолевого Аг, авторы изобретения выбрали модель Her-2/neu. Они наблюдали значительный уровень цитотоксичности, специфичной в отношении Her-2/neu in vivo у мышей, которым вводили нагруженные αGalCer, Her-2/neu 63-71-сенсибилизированные В-клетки (см. Фиг. 17). Для проверки терапевтической противоопухолевой активности в этой модели авторы изобретения вводили опухолевые клетки, экспрессирующие Her-2/neu, мышам до вакцинации их αGalCer-нагруженных, Her-2/neu 63-71-сенсибилизированных В-клеток. После инокуляции опухоли выживаемость была несколько лучше у тех мышей, которых вакцинировали B/αGalCer или B/pep, чем у мышей, вакцинированных только B (см. Фиг. 18). Напротив, все мыши, вакцинированные с использованием B/αGalCer/pep выжили на протяжении всего эксперимента. В модели солидной опухоли мышей с трансплантированными опухолями вакцинировали вакциной на основе В-клеток или вакциной на основе ДК. В результате рост опухолевых клеток ингибировался у мышей, вакцинированных αGalCer-нагруженными, Her-2/neu 63-71-сенсибилизированными В-клетками, также как и у мышей, вакцинированных ДК-вакциной (см. Фиг. 19).
Следовательно, доказано, что вакцина на основе В-клеток согласно изобретению является такой же эффективной, как и вакцины на основе ДК в выработке как профилактического, так и терапевтического противоопухолевого иммунитета.
Настоящее изобретение относится также к иммунотерапевтической/профилактической вакцине и противоопухолевой вакцине, действие которой опосредовано В-клетками, нагруженными лигандом Т-клеток-природных киллеров, и экспрессирующих антиген.
В отличие сенсибилизированных пептидом клеточных вакцин клеточная вакцина, трансдуцированная вирусным вектором для экспрессии целого антигена, может применяться ко всем, без ограничения, в гаплотипах главного комплекса гистосовместимости и может идуцировать различные эпитоп-специфические иммунные ответы, в частности как гуморальный иммунный ответ, так и опосредованный клетками иммунный ответ, одновременно В-клетки трансдуцировали аденовирусом (AdHM), имеющим ген, кодирующий внеклеточный и трансмембранный домен Her-2/neu антигена, ассоциированного с опухолью. В результате на αGalCer презентацию В-клеток не влияла трансдукция AdHM (см. Фиг. 21). Через неделю после иммунизации лизис клеток-мишеней не наблюдался у мышей, иммунизированных В-клетками и B/αGalCer. Напротив, мощный цитотоксический Т-клеточный иммунный ответ наблюдался у мышей, иммунизированных B/AdHM и B/AdHM/αGalCer (см. Фиг. 22 и Фиг. 24).
Цитотоксический T-лимфоцитарный ответ, опосредованный нагруженными αGalCer, трансдуцированными аденовирусом В-клетками (B/AdHM/αGalCer), продолжался выше и дольше, чем ответ, опосредованный В-клетками, трансдуцированными аденовирусом (B/AdHM), хотя различие не было значительным. Эти результаты указывают на то, что нагрузка αGalCer В-клеток в отличие от пептидной сенсибилизации не влияет на стимуляцию цитотоксического Т-клеточного ответа. Однако когда В-клеточную вакцину, нагруженную αGalCer, вводили мышам, NKТ-клетки стимулировались, и тем самым NK-клетки активировались по меньшей мере in vivo, в отличие от мышей, которым вводили В-клеточную вакцину, трансдуцированную аденовирусом (см. Фиг. 25).
Для определения эффективности стратегии вакцин на основе В-клеток в выработке цитотоксического Т-лимфоцитарного ответа авторы изобретения исследовали лизис клеток-мишеней путем вакцинации мышей субоптимальной дозой, которая была недостаточной для индукции полных цитотоксических Т-лимфоцитарных реакций. В результате В-клетки, трансдуцированные аденовирусом для экспрессии антигена, могли индуцировать антиген-специфический Т-лимфоцитарный ответ эффективно, даже несмотря на то, что требовалось большое количество В-клеток по сравнению с сенсибилизированной пептидом В-клеточной вакциной. Поскольку приблизительно 20% всех В-клеток, трансдуцированных аденовирусом, фактически экспрессировали Her-2/neu, эффективность В-клеток, трансдуцированных аденовирусом, была принудительно занижена. В случае переноса небольшого количества В-клеток NKТ-клеточная активация, индуцированная В-клетками, нагруженными αGalCer, была полезна для лизиса клеток-мишеней, индуцированного В-клетками, трансдуцированными аденовирусом.
Вирус, который инфицирует В-клетки для экспрессии опухолевого антигена, может быть проиллюстрирован аденовирусом, ретровирусом, вирусом коровьей оспы, вирусом оспы и вирусом синдбис, но не всегда ими ограничен. Помимо использования вирусного вектора для переноса гена, кодирующего антиген, могут быть использованы следующие способы: 1) связывание ДНК с липосомой для защиты ДНК от разрушения ферментом или абсорбции в эндосому; 2) связывание ДНК с молекулярным конъюгатом или синтетическим лигандом для повышения эффективности трансфекции (например, асиалогликопротеином, трансферрином и полимерным IgA); 3) конструирование новой системы трансдукции ДНК с использованием PTD (домен белковой трансдукции) для повышения эффективности трансдукции гена, кодирующего антиген (например: Mph-1); и 4) введение пептида или белка в В-клетки для презентации антигена.
Настоящее изобретение дополнительно относится к активатору Т-клеток-природных киллеров, опосредованному В-клетками, нагруженными αGalCer. Как объяснялось выше, В-клетки согласно изобретению, нагруженные αGalCer, как дендритные клетки, нагруженные αGalCer, стимулируют гибридому Т-клеток-природных киллеров in vitro и таким образом индуцируют секрецию IL-2 (см. Фиг. 30) и эффективно активируют iNKТ-клетки, которые могли индуцировать активацию различных иммунных клеток in vivo (см. Фиг. 2). Следовательно, В-клетки согласно изобретению, нагруженные αGalCer, могут быть использованы в качестве активатора Т-клеток-природных киллеров как дендритные клетки, нагруженные αGalCer.
Настоящее изобретение также относится к индуктору цитотоксического ответа, опосредованного В-клетками, экспрессирующими опухолевый антиген.
В отличие от В-клеточной вакцины, сенсибилизированной пептидом для индукции опосредованного клетками иммунного ответа, В-клеточная вакцина, трансдуцированная аденовирусом, может индуцировать как опосредованный клетками иммунный ответ, так и гуморальный иммунный ответ одновременно. Для подтверждения, могла ли В-клеточная вакцина, трансдуцированная AdHM индуцировать антитело, специфичное к Her-2/neu, мышей иммунизировали B/AdHM и B/AdHM/αGalCer соответственно, а затем измеряли титр анти-Her-2/neu антител в сыворотке. В результате титр анти-Her-2/neu антитела удерживался на высоком уровне в течение длительного времени в обеих группах, иммунизированных B/AdHM и B/AdHM/αGalCer (см. Фиг. 27). В отличие от групп, иммунизированных только В-клетками (B) и B/αGalCer, гуморальный иммунный ответ генерировался в обеих группах, иммунизированных B/AdHM и B/AdHM/αGalCer (см. Фиг. 28).
Через одну неделю после иммунизации вакциной на основе В-клеток авторы настоящего изобретения выполнили внутривенную инъекцию мышиных/крысиных опухолевых клеток, экспрессирующих Her-2/neu (TAUF), с последующим изучением выживаемости. В результате мыши, которым вводили либо В-клетки, нагруженные αGalCer (B/αGalCer), либо В-клетки, трансдуцированные AdHM (B/AdHM), выживали дольше, чем те, которым вводили только В-клетки. Более того, противоопухолевый эффект у мышей, которым вводили В-клетки, трансдуцированные аденовирусом, экспрессируют Her-2/neu и нагруженные αGalCer был более значительным, чем у мышей, иммунизированных B/AdHM (см. Фиг. 29). Следовательно, было подтверждено, что В-клеточная вакцина, нагруженная αGalCer и трансдуцированная аденовирусом, может эффективно индуцировать иммунные реакции для профилактики опухолей.
Для подтверждения терапевтического эффекта В-клеточной вакцины мышей BALB/c иммунизировали B, B/αGalCer, B/AdHM и B/AdHM/αGalCer соответственно, с последующим внутривенным введением TAUF в хвостовую вену для приживления рака легких. В результате самое короткое время выживания наблюдалось в группе мышей, обработанных только В-клетками, а результат в группе, обработанной B/αGalCer вакциной, был сходно коротким. В то же время период выживания был более или менее пролонгированным в группе мышей, обработанной вакциной B/AdHM, по сравнению с группами, обработанными В или B/αGalCer, но образование опухоли не было полностью подавлено. Но период выживания был гораздо длиннее в группе, обработанной B/AdHM/αGalCer, по сравнению со всеми остальными группами, и, следовательно, противоопухолевый эффект в группе B/AdHM/αGalCer был больше, чем в любых других иммунизированных группах (см. Фиг. 29).
Вакцина согласно изобретению может дополнительно к лиганду Т-клеток-природных киллеров и В-клеткам содержать один или несколько эффективных ингредиентов, обладающих тем же или сходным эффектом. Вакцина также может включать в себя в дополнение к упомянутым выше эффективным ингредиентам один или несколько фармацевтически приемлемых носителей для введения. Фармацевтически приемлемый носитель может быть выбран или получен перемешиванием нескольких ингредиентов, выбранных из группы, состоящей из солевого раствора, стерильной воды, раствора Рингера, забуференного солевого раствора, раствора декстрозы, раствора мальтодекстрозы, глицерина и этанола. Можно добавлять другие общие добавки, такие как антиокислитель, буферный раствор, бактериостатический агент и т.д. для получения инъекционных растворов, таких как водный раствор, суспензия и эмульсия, разбавители, диспергирующие агенты, поверхностно-активные вещества, связующие и смазывающие вещества можно дополнительно добавлять. Вакцина согласно изобретению в дальнейшем может быть получена в подходящих формах для каждого заболевания или в соответствии с ингредиентами по способу, представленному у Remington's Pharmaceutical Science (в последней редакции), Mack Publishing Company, Easton PA.
Вакцину согласно изобретению можно вводить парентерально и такое парентеральное введение включает в себя подкожную инъекцию, внутривенную инъекцию, внутримышечную инъекцию и внутригрудную инъекцию. Для получения вакцины в виде состава для парентерального введения В-клетки, нагруженные лигандом Т-клеток-природных киллеров, В-клетки, нагруженные лигандом Т-клеток-природных киллеров и пептидом, или В-клетки, трансдуцированные вирусом, экспрессирующим опухолевый антиген, смешивают со стабилизатором или буферным агентом для получения раствора или суспензии, которые потом предоставляли в виде ампул или флаконов (см. стандарт для получения вирусного вектора, используемого для трансдукции дендритных клеток).
Вакцину можно вводить эффективной дозой для индукции иммунного ответа у пациента. Например, вакцину можно вводить людям однократно или несколько раз в день дозировкой 1·103~1·109 клеток/кг, и более предпочтительно 1·104 клеток/кг ~ 1·108 клеток/кг. Для получения В-клеточной вакцины, нагруженной αGalCer, среда должна быть дополнена αGalCer в концентрации 1-2 мкг/мл на 1·106-1·107 В-клеток/мл. Для получения В-клеточной вакцины, сенсибилизированной совместно αGalCer и пептидом, среда должна быть дополнена αGalCer в концентрации 1-2 мкг/мл на 1·106~1·107 В-клеток/мл и пептидом в концентрации 1-10 мкг/мл на 1·106~1·107 В-клеток/мл.
По-видимому, αGalCer не индуцирует токсичность у грызунов и обезьян (Nakata et al., Cancer Res., 58: 1202-1207, 1998). При введении мышам 2200 мкг/кг побочных эффектов не отмечалось (Giaccone et al., Clin. Cancer Res., 8: 3702, 2002). В результате клинического испытания отмечалась слабая головная боль как побочный эффект в соответствии с системным введением αGalCer (Mie Nieda et al., Blood, 103: 383-389, 2004, Giaccone et al., Clin. Cancer Res., 8: 3702, 2002), которую можно предотвратить приемом парацетамола. Это небольшая, если вообще есть, возможность показать небольшой системный побочный эффект (Giaccone et al., Clin. Cancer Res., 8: 3702, 2002). В настоящем изобретении αGalCer не вызывал дозо-зависимой токсичности (50-4800 мкг/м2) и продемонстрировал резистентность на протяжении всего исследования с повышением дозы, что указывает на то, что αGalCer является весьма безопасным веществом.
Настоящее изобретение включает в себя антигенные пептиды вирусного, бактериального, грибкового, паразитарного и опухолевого происхождения и введенный антиген в В-клетки, опосредованный вирусным вектором, включает в себя антиген вирусного, бактериального, грибкового, паразитарного и опухолевого происхождения.
«Антиген» означает любое вещество (например, белок, пептид, опухолевые клетки, гликопротеин, гликолипид, живой вирус, убитый вирус, ДНК и т.д.), которое способно индуцировать иммунный ответ при распознавании его иммунной системой, когда оно проникает в хозяина. Антиген может быть получен в очищенном или неочищенном виде, но очищенная форма является предпочтительной. Антиген согласно изобретению включает в себя патогенный белок, рекомбинантный белок, пептид, полисахарид, гликопротеин, липополисахарид и молекулу ДНК (полинуклеотид), опухолевые клетки, живые вирусы и убитые вирусы.
Антигены, представленные в приведенном ниже перечне антигенов, могут быть предоставлены в качестве инструмента для индукции иммунного ответа вместо ассоциированного с опухолью антигена в Примерах изобретения, но не всегда ограничены этим перечнем. Перечень включает в себя антиген вируса гриппа (гемагглютинин и нейраминидазный антигены), антиген Bordetella pertussis (коклюшевый токсин, нитевидный гемагглютинин и пертактин), антиген вируса папилломы человека (HPV) (гликопротеин), антиген Helicobacterpylori (капсульные полисахариды серогруппы A, B, C, Y и W-135), столбнячный токсоид, дифтерийный антиген (дифтерийный токсоид), пневмококковый антиген (капсульный полисахарид Streptococcus pnemoniae 3 типа, туберкулезный антиген, антиген вируса иммунодефицита человека (HIV) (GP-120, GP-160, p18, Tat, Gag, Pol и Env), холерный антиген (В субъединица холерного токсина), антиген стафилококка (стафилококковый энтеротоксин B), антиген шигеллы (полисахариды шигеллы), антиген вируса везикулярного стоматита (гликопротеин вируса везикулярного стоматита), антиген цитомегаловируса (CMV), антиген гепатита (антигены гепатита A(HAV), B(HBV), C(HCV), D(HDV) и G(HGV)) (коровый антиген и поверхностные антигены), антиген респираторно-синцитиального вируса (RSV), антигены простого герпеса или их сочетания (например, дифтерии, коклюша и столбняка, DPT), антиген Borrelia sp. (например, антигены OspA, OspB и OspC), антиген Candida albicans (например, MP65) и антиген плазмодия (например, белок CS).
Опухолевый антиген образуется путем соматической мутации гена дикого типа. Злокачественный антиген включает в себя антиген, специфичный к опухоли, придающий генетическую нестабильность опухолевой клетке, и ассоциированный с опухолью антиген, эндогенный аутоантиген, который экспрессируется больше в опухолевых клетках и временно экспрессируется на стадии развития опухоли или экспрессируется в специфической ткани ограниченно. Примером специфичного к опухоли антигена может служить HPV E6/E7, который представляет собой антиген злокачественного вируса, а ассоциированный с опухолью антиген может быть проиллюстрирован gp100, тирозиназой, TRP-1 (родственный тирозиназе белок-1), TRP-2 (родственный тирозиназе белок-2), MAGE, MUC-1, CEA (карнцероэмбриональный антиген), p53, альфа-фетопротеин и Her-2/neu, и т.д. (Rosenberg SA., Nature, 17; 411 (6835): 380-4, 2001).
Описание чертежей
Применение предпочтительных вариантов осуществления изобретения лучше всего понятно со ссылкой на сопровождающие чертежи, на которых:
на Фиг. 1 - Фиг. 3 представлены графики, иллюстрирующие реципрокную активацию между iNKТ-клетками и В-клетками, нагруженными αGalCer;
на Фиг. 1 представлена серия графиков, иллюстрирующих, что В-клетки очищали из селезенки после истощения CD11c+ клеток с последующим измерением экспрессии CD19, CD11c и CD1d с помощью проточной цитометрии;
на Фиг. 2 представлена серия графиков, иллюстрирующих, что мышам C57BL/6 дикого типа или мышам Jα281-/- вводили только наполнитель, свободную форму αGalCer, или В-клетки, нагруженные αGalCer (B/αGalCer) или ДК (ДК/αGalCer), а затем получали спленоциты от мышей-реципиентов и анализировали клетки, продуцирующие IL-4 и IFN-γ, анализировали с помощью анализа ELISPOT;
на Фиг. 3 представлена серия графиков, иллюстрирующих, что В-клетки, нагруженные наполнителем или αGalCer, окрашивали CFSE и вводили мышам C57BL/6. Анализировали CFSE+ клетки из спленоцитов;
на Фиг. 4 - Фиг. 6 представлены графики, демонстрирующие, что В-клетки, сенсибилизированные совместно αGalCer и пептидом (OVA257-264), активируют пептид-специфические CD8+ Т-клетки;
на Фиг. 4 представлена серия графиков, иллюстрирующих, что CD8+ Т-клетки, специфичные к OVA257-264, меченные CFSE (OT-I Т-клетки), переносили мышам C57BL/6, которым вводили B/наполнитель, B/αGalCer, B/pep или B/αGalCer/pep соответственно. Затем исследовали пролиферацию OT-1 Т-клеток и секрецию цитокинов с помощью проточной цитометрии;
на Фиг. 5 представлена серия графиков, иллюстрирующих, что мышей C57BL/6 вакцинировали указанными формами В-клеток (только B, B/αGalCer, B/pep или B/αGalCer/pep). Одну, три или пять недель спустя проводили in vivo исследование CTL путем введения сингенных мишеней, меченных CFSE;
CFSEhigh: сенсибилизированная пептидом мишень;
CFSElow: несенсибилизированный пептидом контроль;
на Фиг. 6 представлена серия графиков, иллюстрирующих, что ответ CD8+ Т-клеток, продуцирующих IFN-γ, на повторную стимуляцию OVA257-264 in vitro рассчитывали 7 дней спустя после введения вакцин на основе В-клеток (слева) или 7 дней после дополнительного прайминга CTL с использованием сингенных спленоцитов, нагруженных OVA257-264 (справа);
на Фиг. 7 и Фиг. 8 представлены графики сравнения эффективности выработки цитотоксических Т-лимфоцитов между клеточной вакциной на основе В-клеток и на основе ДК;
на Фиг. 7 представлена серия графиков, иллюстрирующих, что B/αGalCer/pep и ДК/αGalCer/pep, сенсибилизированные пептидом OVA257-264, серийно разводили, затем вводили мышам, а затем измеряли цитотоксичность in vivo в отношении OVA257-264;
на Фиг. 8 представлена серия графиков, иллюстрирующих, что B/наполнитель/pep и ДК/наполнитель/pep, сенсибилизированные OVA257-264, вводили аналогичным образом, как описано на Фиг. 7, а затем измеряли цитотоксичность in vivo в отношении OVA257-264;
на Фиг. 9 и Фиг. 10 представлены графики, показывающие типы клеток, вовлеченных в CTL-ответ, индуцированный вакциной, на основе В-клеток;
на Фиг. 9 представлена серия графиков, иллюстрирующих, что В-клетки, совместно сенсибилизированные αGalCer и OVA257-264, вводили мышам. Мыши-реципиенты получали моноклональные антитела, элиминирующие иммунные клетки, затем их тестировали на in vivo цитотоксичность в отношении OVA257-264;
на Фиг. 10 представлен график, иллюстрирующий, что В-клетки, сенсибилизированные совместно αGalCer и пептидом OVA257-264, вводили мышам дикого типа, Jα281-/- или MHC класс II-/-, с последующим стандартным анализом высвобождения 51Cr, для исследования цитотоксичности in vitro в отношении OVA257-264;
на Фиг. 11 и Фиг. 12 представлены графики, демонстрирующие, что В-клетки, сенсибилизированные совместно с использованием αGalCer и пептида OVA257-264, действуют как непосредственные антиген-представляющие клетки для CD8+ Т-клеток;
на Фиг. 11 представлена серия графиков, иллюстрирующих, что В-клетки мышей дикого типа или мышей bm-1 были сенсибилизированы совместно αGalCer и OVA257-264 до инъекции их мышам дикого типа, затем измеряли цитотоксичность in vivo в отношении OVA257-264;
на Фиг. 12 представлена серия графиков, иллюстрирующих, что мышей C57BL/6 вакцинировали В-клетками, сенсибилизированными совместно αGalCer и OVA257-264, или сочетанием В-клеток, сенсибилизированных OVA257-264, и В-клеток, сенсибилизированных αGalCer', затем измеряли in vivo цитотоксичность в отношении OVA257-264;
на Фиг. 13 - Фиг. 16 представлены графики, иллюстрирующие, что вакцина на основе В-клеток может обеспечить как профилактический, так и терапевтический противоопухолевый иммунитет против меланомы B16, трансфицтированной OVA;
на Фиг. 13 представлен график, иллюстрирующий, что мышей C57BL/6 вакцинировали указанной клеточной вакциной (только B, B/αGalCer, B/pep, B/αGalCer/pep, ДК/pep или ДК/αGalCer/pep). Семь дней спустя опухолевые клетки MO-5 вводили подкожно мышам, а затем измеряли массу опухоли;
на Фиг. 14 представлен график, иллюстрирующий, что рост повторно введенных опухолевых клеток подавлялся у мышей без опухолей, представленных на Фиг. 13. Через семьдесят дней после первой инокуляции опухоли мышей без опухолей повторно заражали подкожно клетками MO-5 (2·l05) и измеряли массу опухоли;
на Фиг. 15 и Фиг. 16 представлены графики, иллюстрирующие, что мышей C57BL/6 вакцинировали указанной клеточной вакциной (только B, B/αGalCer, B/pep, B/αGalCer/pep, ДК/pep или ДК/αGalCer/pep). Один день или девять дней спустя опухолевые клетки MO-5 подкожно вводили мышам, а затем измеряли массу опухоли;
* p<0,05 по сравнению с контрольной группой «только В» (Фиг. 13, Фиг. 15 и Фиг. 16) или с интактными мышами (Фиг. 14);
на Фиг. 17 - Фиг. 19 представлены графики, иллюстрирующие, что вакцина на основе В-клеток запускает терапевтический противоопухолевый иммунитет против опухолей, экспрессирующих Her-2/neu;
на Фиг. 17 представлена серия графиков, иллюстрирующих, что мышей BALB/c вакцинировали В-клетками или ДК после совместного культивирования с αGalCer или наполнителем плюс Her-2/neu 63-71 (hP63), как указано, а затем проводили исследования CTL in vivo;
на Фиг. 18 и Фиг. 19 представлены графики, иллюстрирующие, что мышей BALB/c заражали внутривенно (Фиг. 18) или подкожно (Фиг. 19) CT26-Her2/neu, способом, аналогичным описанному выше, с последующей иммунизацией мышей В-клетками или ДК и определяли выживаемость этих мышей (Фиг. 18) или рост опухоли (Фиг. 19);
на Фиг. 20 и Фиг. 21 представлены графики, иллюстрирующие испытание для проверки того, можно ли способ трансдукции В-клеток целым антигеном, с использованием вирусного вектора применять для В-клеточной вакцины или нет;
на Фиг. 20 представлена серия графиков, иллюстрирующих, что В-клетки заражали аденовирусом (AdHM), экспрессирующим внеклеточный домен и трансмембранный домен Her-2/neu, ассоциированного с опухолью антигена, а затем измеряли экспрессию Her-2/neu на поверхности В-клетки;
на Фиг. 21 представлен график, иллюстрирующий, что В-клетки, трансдуцированные AdHM и нагруженные αGalCer, могли активировать клетки DN32.D3. В частности, клетки DN32.D3 культивировали соответственно только с В-клетками, В-клетками, нагруженными αGalCer, В-клетками, трансдуцированными AdHM, и В-клетками, трансдуцированными AdHM и нагруженными αGalCer, а затем измеряли уровень IL-2 в каждом супернатанте с помощью ELISA;
на Фиг. 22 - Фиг. 24 представлены графики, иллюстрирующие, что В-клетки, трансдуцированные ассоциированным с опухолью антигеном Her-2/neu, эффективно индуцируют цитотоксический иммунный ответ;
на Фиг. 22 представлен график, иллюстрирующий, что мышей иммунизировали соответственно В-клетками, выделенными из селезенки мышей BALB/c, а затем трансдуцированными аденовирусом (B/AdHM), В-клетками, дополнительно нагруженными αGalCer (B/AdHM/αGalCer), только В-клетками, и В-клетками, нагруженными αGalCer (B/αGalCer). Каждой мыши переносили клетки-мишени, нагруженные цитотоксическим Т-клеточным эпитопом для исследования антигенспецифического цитотоксического ответа Т-лимфоцитов in vivo;
на Фиг. 23 представлен график сравнения продолжительности цитотоксического ответа Т-клеток в каждой группе мышей, иммунизированных B/AdHM и B/AdHM/αGalCer;
на Фиг. 24 представлен график сравнения уровней IFN-γ секретируемого CD8+ Т-клетками до и после стимуляции каждой мыши, иммунизированной B/AdHM и B/AdHM/αGalCer, с цитотоксическим Т-клеточным пептидом in vitro;
интактные: группа мышей, не обработанных вакциной;
на Фиг. 25 и Фиг. 26 представлены графики, иллюстрирующие, что В-клеточные вакцины, трансдуцированные аденовирусом (B/AdHM/αGalCer), могли эффективно активировать клетки-природные-киллеры и индуцировать цитотоксический Т-клеточный ответ;
на Фиг. 25 представлен график, на котором показана активация клеток-природных киллеров;
на Фиг. 26 представлен график, на котором показан лизис клеток-мишеней в зависимости от дозы клеточной вакцины, который исследовали для сравнения эффективности в индукции цитотоксических Т-клеток;
на Фиг. 27 и Фиг. 28 представлены графики, иллюстрирующие, что В-клеточная вакцина, трансдуцированная AdHM (B/AdHM и B/AdHM/αGalCer), могла индуцировать Her-2/neu-специфическое антитело;
на Фиг. 27 представлен график, иллюстрирующий, что мышей BALB/c иммунизировали B/AdHM и B/AdHM/αGalCer соответственно, а затем измеряли титр анти-Her-2/neu-антител для исследования связывания анти-Her-2/neu-антитела в сыворотке с мышиными/крысиными опухолевыми клетками, экспрессирующими Her-2/neu на его поверхности (TAUF);
на Фиг. 28 представлен график, демонстрирующий гуморальный иммунный ответ в каждой группе;
на Фиг. 29 представлена серия графиков, показывающих противоопухолевую активность В-клеточной вакцины, трансдуцированной аденовирусом. В частности, рак легких индуцировали у мышей инокуляцией TAUF, с последующей иммунизацией В-клеточной вакциной;
на Фиг. 30 представлена серия графиков, иллюстрирующих секрецию IL-2, которая является индексом для активации Т-клеток-природных киллеров, путем активации Т-клеток-природных киллеров, индуцированных под действием В-клеток, нагруженных αGalCer, в соответствии с концентрацией αGalCer, и активации Т-клеток-природных киллеров, индуцируемых под действием В-клеток или ДК, нагруженных αGalCer, в соответствии с клеточной концентрацией;
на Фиг. 31 - Фиг. 35 представлены графики, иллюстрирующие, что специфические иммунные клетки могли элиминироваться введением антитела, способного истощать CD4+, CD8+ или NK1.1-клетки in vivo;
на Фиг. 36 представлен график, показывающий противоопухолевую иммунную активность в отношении тимомы (EG-7), индуцированную вакциной на основе В-клеток;
*p < 0,001: статистическая значимость по сравнению с группой, обработанной только В-клетками.
Способ осуществления изобретения
Практические и в настоящее время предпочтительные варианты осуществления настоящего изобретения являются иллюстрирующими, как показано в следующих Примерах.
Однако должно быть понятно, что специалисты в данной области, приняв во внимание это описание, могут производить модификации и усовершенствования в пределах сущности и объема настоящего изобретения.
Пример 1
Двунаправленная активация В-клеток и iNKТ-клеток, нагруженных αGalCer
1-1. Сравнение iNKТ-клеточной активации, индуцированной В-клетками под действием ДК
Поскольку уже четко установлено, что ДК, нагруженные αGalCer, активируют iNKТ-клетки (Kim, S., et al., Synthesis, 847, 2004), авторы настоящего изобретения исследовали, оказывают ли В-клетки, нагруженные αGalCer, эффект, сходный с ДК, нагруженными αGalCer, или нет.
Использовали самок мышей C57BL/6 и BALB/c в возрасте 6-10 недель. Мышей OT-I и C57BL/6bm1 (bm1) покупали в Jackson Laboratory, а мыши Jα281-/- и мыши MHC II-/- были любезно предоставлены Dr. Chung (Seoul National University, Korea) (Kim JH et al., Am J Pathol., 167(5):1231-41, 2005) и Dr. Park (Korea University, Korea) (Park, S. H., et al., J. Exp. Med. 193:893, 2001) соответственно. Всех мышей держали в специальных беспатогенных условиях в Центре животных для фармацевтических исследований Сеульского национального университета Кореи (Animal Center for Pharmaceutical research at Seoul National University, Korea). Получали антитела из гибридом, а именно GK1.5 (анти-CD4: ATCC номер: TIB-207), 2.43 (анти-CD8; ATCC номер: TIB-210), и PK136 (анти-NK1.1; ATCC номер: HB-191) и вводили их внутрибрюшинно (150 (мкг/мышь) для истощения соответствующих подгрупп лимфоцитов in vivo.
В частности, для отделения В-клеток из каждой мыши у мыши выделяли селезенку с последующей гомогенизацией. После истощения CD11c+ клеток из селезенки с помощью анти-CD11c микробус (Miltenyibiotec) авторы настоящего изобретения выделяли чистые В-клетки, используя анти-B220 микробусы (Miltenyibiotec). Более 99% этих клеток были CD19-позитивными (см. Фиг. 1).
Для отделения дендритных клеток из селезенки полученную от мыши селезенку помещали в 7 мл среды, содержащей коллагеназу D (1 мг/мл, Roche) и ДНазу I (50 мкг/мл, Sigma-Aldrich), с последующим взаимодействием при 37°C в течение 30 минут. В среду добавляли EDTA (конечная концентрация 10 мM, pH 7,2), с последующим проведением дополнительной реакции в течение еще 5 минут. Дендритные клетки отделяли от спленоцитов с помощью 15,5% градиента плотности Accudenz (Accurate Chemical & Scientific).
Очищенные клетки (В-клетки или ДК) совместно культивировали с αGalCer (1 мкг/мл) или с наполнителем (0,5% полисорбат) в течение 14 часов в CО2 инкубаторе. Затем клетки совместно культивировали с гибридомой Т-клеток-природных киллеров, DN32.D3 (Claire Forestier et al., The Journal of Immunology, 171: 4096, 2003); затем измеряли IL-2 как индикатор активации NKT с помощью сэндвич ELISA.
В результате ДК, нагруженные αGalCer (ДК/αGalCer), стимулировали DN32.D3 к продукции IL-2. Подобным образом В-клетки, нагруженные αGalCer (B/αGalCer), эффективно стимулировали DN32.D3 клетки к продукции IL-2, уравнивая скорость продукции IL-2 группой ДК при более высоких соотношениях с гибридомой, но ухудшая при более низких соотношениях (см. Фиг. 30).
1-2. Активация Т-клеток-природных киллеров в зависимости от иммунного ответа
Для исследования, могли ли B/αGalCer и ДК/αGalCer, введенные in vivo, активировать иммунные клетки или нет, авторы настоящего изобретения вводили B/αGalCer или ДК/αGalCer от мышей C57BL/6 внутривенно сингенным мышам и измеряли уровень клеток, продуцирующих IL-4 и IFNγ, с помощью ELISPOT. В частности, мышам C57BL/6 дикого типа или мышам Jα281-/- внутривенно вводили наполнитель, αGalCer, В-клетки, нагруженные αGalCer, или ДК, нагруженные αGalCer. Одну неделю спустя селезенку отделяли для получения отдельных клеток, которые помещали на планшет ELISPOT. В планшет добавляли наполнитель или αGalCer с последующим культивированием в течение 6 часов для стимуляции. ELISPOT проводили для определения количества клеток, секретирующих IL-4 и IFN-γ, в соответствии с инструкциями производителя (наборы IL-4 ELISPOT, IFN-γ ELISPOT; R&D system).
В результате инъекция как B/αGalCer, так и ДК/αGalCer индуцировала высокие количества клеток, продуцирующих IL-4 и IFN-γ (Фиг. 2). Однако следует отметить, что заметное количество этих цитокин-продуцирующих клеток выявлялось даже в отсутствие повторной стимуляции с использованием αGalCer. Тем не менее, авторы настоящего изобретения полагают, что индукция клеток, продуцирующих IL-4 и IFN-γ, зависит от iNKТ-клеток, поскольку этого не происходит у мышей Jα281-/-, лишенных популяции iNKT (см. Фиг. 2).
1-3. В-клеточная активация под действием активации Т-клеток-природных киллеров
Для определения какие, если есть, изменения были индуцированы в В-клетках после инъекции, авторы настоящего изобретения внутривенно вводили B/αGalCer, меченные CFSE (карбоксифлуоресциен сукцинимидиловым эфиром), а затем анализировали молекулы костимуляции на CFSE+ клетках. В частности, лимфоциты получали из селезенки и лимфатического узла мышей C57BL/6 и клетки CD11c+ удаляли с использованием анти-CD11c микробус (Miltenyibiotec). В-клетки разделяли с использованием анти-B220 микробус (Miltenyibiotec). Разделенные В-клетки были помечены 10 мкM CFSE, которые вводили в хвостовую вену сингенной мыши. Для мечения CFSE CFSE (Molecular Probe) разводили в среде RPMI в соответствующей концентрации, с последующим проведением реакции при 37°C в течение 15 минут для мечения. Клетки промывали средой RPMI три раза для удаления оставшегося CFSE. Двадцать четыре и сорок восемь часов после мечения молекулы костимуляции на клетках CFSE+, отделенные от клеток селезенки и лимфатического узла мыши, исследовали с помощью проточной цитометрии.
В результате высокие уровни экспрессии CD86, но не CD80 индуцировались в течение 24 часов. CD40 и MHC II также слега активировались (Фиг. 3). Даже через 48 часов после инъекции не наблюдалось активации CD80 и все другие результаты оставались главным образом в соответствии с 24-часовом уровнем. То есть В-клетки активировались Т-клетками-природными киллерами, активированными под действием B/αGalCer в пределах 24 часов.
Принимая во внимание результаты Примеров <1-1>, <1-2> и <1-3>, было подтверждено, что как ДК/αGalCer, так и B/αGalCer способны к полной активации iNKТ-клеток как in vitro, так и in vivo, и эта активация in vivo является двунаправленной (поскольку были активированы как iNKT, так и В-клетки).
Пример 2
B/αGalCer, сенсибибилизированные пептидом, способствуют активации пептид-специфических CD8+ Т-клеток
Авторы настоящего изобретения изучали, могла ли ко-сенсибилизация αGalCer и MHC I-рестриктированного пептида на В-клетках примировать пептид-специфические CD8+ Т-клетки. С этой целью авторы изобретения сначала адаптивно переносили CFSE-меченые OVA-специфичные CD8+ Т-клетки (OT-I) мышам C57BL/6, затем вводили В-клетки, сенсибилизированные носителем (только В-клетки), B/αGalCer, наполнитель и В-клетки, сенсибилизированные пептидом OVA257-264 (B/pep), или В-клетки, сенсибилизированные αGalCer и OVA257-264 (B/αGalCer/pep). Сорок восемь часов спустя из селезенки получали отдельные клетки, а затем наблюдали пролиферацию клеток OT-I. В то же время клетки OT-I, секретирующие IL-2 и IFN-γ, культивировали с 1 мкМ OVA257-264 пептида и 1 мкл/мл GolgiPlug в течение четырех часов, а затем проводили внутриклеточное окрашивание цитокинов с использованием набора Cytoperm/Cytofix (BD Pharmingen).
В результате небольшое деление OVA-специфичных CD8+ Т-клеток (OT-I клеток) было индуцировано у мышей, получающих только В; однако слабое деление OVA-специфичных CD8 Т-клеток Аг-неспецифичным образом отмечался в группе B/αGalCer (Фиг. 7, 1,2±0,5% по сравнению с 5,4±1,8%). Инъекция B/pep индуцировала значительное деление OT-I, но очень немногие из этих клеток продуцировали IL-2 (<4%) и относительно немногие продуцировали IFN-γ (38%) после повторной стимуляции in vitro. Напротив, у мышей, которым вводили B/αGalCer/pep, наблюдалось усиленное деление OT-I, с более 40% полученных в результате клеток, продуцирующих IL-2, и наиболее неожиданно более 90% продуцирующих IFN-γ на значительно более высоких уровнях, чем в группе B/pep (Фиг. 4).
Эти результаты дают возможность предположить, что гораздо более высокая скорость CD8+ Т-клеточной активации могла быть достигнута нагрузкой αGalCer на B/pep.
Пример 3
Длительный цитотоксический Т-клеточный ответ и преодоление иммунологической толерантности под действием B/αGalCer/pep
3-1. Длительный цитотоксический Т-клеточный ответ, индуцированный действием B/αGalCer/pep
Авторы настоящего изобретения исследовали, могла ли методика вакцины на основе В-клеток индуцировать цитотоксический иммунитет. С этой целью авторы изобретения внутривенно вводили группам мышей C57BL/6 только B, B/αGalCer, B/pep или B/αGalCer/pep, а затем определяли in vivo активность CTL. Кроме того, авторы изобретения исследовали, если цитотоксический иммунный ответ, индуцированный вакциной на основе В-клеток, мог быть продолжительным. В частности, мышей C57BL/6 иммунизировали только В, B/αGalCer, B/pep и B/αGalCer/pep соответственно. На первой, третьей и пятой неделе иммунизации проводили in vivo исследование CTL. Сингенные лимфоциты либо нагружали OVA257-264 пептидом либо оставляли нетронутыми до мечения 20 мкМ (CFSEhigh) или 2 мкM (CFSElow) CFSE. Равные количества двух популяций смешивали и вводили мышам внутривенно. На следующий день получали отдельные клетки и CFSEhigh и CFSElow и измеряли с помощью проточной цитометрии. Чем ниже число CFSEhigh, тем выше цитотоксический иммунный ответ.
В результате только B/αGalCer/pep полностью лизировал сенсибилизированные пептидом мишени, а также поддерживал полную цитотоксичность даже 5 недель после однократной инъекции. В то же время ни B/pep, ни B/αGalCer не индуцировали каких-либо пептид-специфических CTL-ответов у мышей. Только в группе, обработанной B/αGalCer/pep, было показано существенно повышение числа IFN-γ-продуцирующих CD8+ Т-клеток в отношении OVA257-264 (Фиг. 6, слева).
3-2. Подтверждение преодоления иммунологической толерантности под действием B/αGalCer/pep
В основном, введение одного только антигена индуцирует иммунологическую толерантность. Следовательно, авторы настоящего изобретения исследовали, можно ли преодолеть иммунологическую толерантность, индуцированную антигеном, или нет, когда В-клетки нагружены αGalCer вместе с пептидом. В частности, мышей C57BL/6 иммунизировали аналогичным образом, описанным в Примере <3-1>. Неделю спустя получали спленоциты, с последующей стимуляцией сингенными спленоцитами, покрытыми OVA, в течение одной недели. CD8+ Т-клетки, секретирующие IFN-γ, в стимулированных спленоцитов определяли с помощью внутриклеточного цитокинового окрашивания.
В результате мыши, которым ввели только В или B/αGalCer, отвечали нормально на прайминг и генерировали значительное количество пептид-специфических IFN-γ-продуцирующих CD8+ Т-клеток (Фиг. 6, справа). Однако у мышей, которым вводили B/pep, не было показано увеличения количества IFN-γ-продуцирующих CD8+ Т-клеток, что дает возможность предположить, что эти мыши были толерантными к этому пептиду.
Напротив, мыши, вакцинированные B/αGalCer/pep, приобретали гораздо большее число пептид-реактивных CD8+ Т-клеток, чем мыши либо в группе, получавшей только В, либо B/αGalCer. В это время мышам, вакцинированным только В или B/αGalCer, не вводили никакого антигена, поэтому антиген-специфичный иммунный ответ не индуцировался у этих мышей и in vitro стимуляция спленоцитами, покрытыми OVA, была первой и единственной стимуляцией, вызывающей иммунный ответ. Однако мышам, вакцинированным B/αGalCer/pep, уже ввели антиген, поэтому in vitro стимуляция спленоцитами, покрытыми OVA, представляла собой вторичный иммунный ответ. В заключение B/αGalCer/pep преодолевает иммунологическую толерантность, демонстрируя иммуногенность и мощный вторичный иммунный ответ.
Пример 4
Вакцина на основе В-клеток является таким же эффективным генератором CTL, как и вакцина на основе ДК
Авторы настоящего изобретения сравнили эффективность вакцины на основе В-клеток в выработке цитотоксичности с действием ДК-вакцины.
С этой целью авторы изобретения определи минимальное количество клеток, необходимое для достижения полного лизиса мишени in vivo. В частности, В-клетки или ДК культивировали совместно с αGalCer или наполнителем в течение 16-18 часов, а затем сенисибилизировали 1 мкг/мл OVA257-264 в течение часа. Серийно разведенные клетки вводили внутривенно сингенным мышам и проводили анализ CTL in vivo. Как объяснялось в приведенном выше Примере, сенсибилизированные пептидом мишени были помечены как CFSEhigh, а контрольные клетки, не сенсибилизированные пептидом, были помечены как CFSElow. Равные количества двух популяций перемешивали и вводили мышам внутривенно. Несколько позже спленоциты мышей исследовали с помощью проточной цитометрии для подсчета соотношения CFSElow: CFSEhigh для количественного определения антиген-специфичного лизиса клеток-мишеней.
В результате у мышей, которым вводили ДК/αGalCer/pep, наблюдался полный лизис клеток-мишеней только с 16000 клеток. Интересно то, что однократной вакцинации 80000 B/αGalCer/pep клеток было достаточно для установления полного пептид-специфического лизиса, тогда как вакцинация 16000 клеток генерировала умеренную цитотоксичность (Фиг. 7). Однако с учетом того, что площадь поверхности ДК значительно больше, чем площадь поверхности В-клеток, можно сказать, что B/αGalCer/pep является столь же эффективным, что и ДК/αGalCer/pep в выработке цитотоксичности. ДК/pep эффективно генерировал OVA-специфическую цитотоксичность, в то время как обработка B/pep, несмотря на количество тестируемых клеток, нет (Фиг. 8). Следует отметить, что профиль цитотоксичности in vivo в группе, обработанной ДК/pep, был очень похож на профиль в группе, обработанной ДК/αGalCer/pep, указывая на то, что нагрузка αGalCer на ДК не усиливала вакцинную эффективность ДК/pep.
Пример 5
Необходимое условие выработки CTL под действием B/αGalCer/pep
Авторы изобретения исследовали вопрос о том, какого типа иммунные клетки участвуют в развитии CTL-ответа. С этой целью мышам вводили элиминирующие антитела (элиминирующее антитело анти-CD4: GK1.5, элиминирующее антитело анти-CD8; 2.43, элиминирующее антитело анти-NK1.1; PK136). За четыре дня до или четыре для после их вакцинировали B/αGalCer/pep, а затем проводили анализ CTL in vivo.
В результате сроки элиминации не отличались, поскольку выработка CTL активности не препятствовала элиминация ни CD4+, ни NK1.1+ клеток (Фиг. 9). С другой стороны, элиминация CD8 полностью блокировала уничтожение клеток-мишеней. В соответствии с этими результатами у мышей MHC II-/- (лишенных CD4+ Т-клеток) развивались нормальные CTL-ответы, тогда как у мышей Jα281-/- (лишенных iNKТ-клеток) не удалось сделать то же самое (Фиг. 10).
Пример 6
Презентация антигена и стимуляция цитотоксических Т лимфоцитов В-клетками
Можно было бы утверждать, что сенсибилизированные пептидом В-клетки действуют не как АПК, а как резервуары пептида, из которых ДК хозяина извлекают пептиды для индукции CTL-ответов. Для изучения этой возможности авторы настоящего изобретения использовали мышей bm-1 (Norbury, C. C. et al., Science 304, 1318-1321, 2004). Клетки этих мышей могут нагружать OVA пептид на их MHC I, но полученный в результате комплекс не распознается когнитивными CD8+ Т-клетками вследствие мутации в области H-2K. В-клетки этой линии нормально экспрессировали CD1d и стимулировали активацию iNKT в ответ на αGalCer, указывая на то, что взаимодействие между В и iNKT было интактным.
Еще раз когда B/αGalCer/pep получали с использованием В-клеток мышей дикого типа, а затем вводили мышам дикого типа, генерировалась полная in vivo OVA-специфичная цитотоксичность. Однако когда B/αGalCer/pep, полученный из В-клеток мышей bm-1, вводили мышам дикого типа, не удавалось генерировать OVA-специфичную цитолитическую активность, что дает возможность предположить, что ДК или другие профессиональные АПК у мышей-реципиентов не отвечали за выработку CTL (Фиг. 11).
Авторы настоящего изобретения исследовали, возможно ли генерировать CTL, когда αGalCer и пептид были сенсибилизированы по отдельности, а затем введены вместе. С этой целью мышам C57BL/6 внутривенно вводили 'B/αGalCer плюс B/pep' или только B/αGalCer/pep и проводили анализ in vivo CTL. В результате у мышей, вакцинированных 'B/αGalCer плюс B/pep', не удалось генерировать in vivo цитотоксичность (Фиг. 12). Этот результат показывает, что пептид и αGalCer должны быть представлены на одной и той же В-клетке для выработки цитотоксичности.
Пример 7
Противоопухолевое действие B/αGalCer/pep
7-1. In vivo тест с использованием OVA моделей
Авторы настоящего изобретения исследовали, могла ли вакцинация с использованием B/αGalCer/pep генерировать противоопухолевый иммунитет. Для проверки профилактической противоопухолевой активности группы мышей вакцинировали однократно только B, B/αGalCer, B/pep, B/αGalCer/pep, ДК/pep, или ДК/αGalCer/pep перед подкожной трансплантации им OVA-трансфектированной меланомы B16 (MO-5) (Dr. Kenneth Rock, University of Massachusetts: Falo, L. D., et al., Nat. Med. 1: 649, 1995). В результате наблюдалась небольшая задержка в профиле опухолевого роста у мышей, вакцинированных B/αGalCer, хотя у всех мышей в конечном итоге развились опухоли (Фиг. 13). Напротив, у мышей, получавших B/αGalCer/pep, ДК/pep или ДК/αGalCer/pep, рост опухоли не развивался.
Для изучения того, устанавливается ли у этих мышей длительная противоопухолевая активность, авторы изобретения повторно заражали выживших мышей опухолями MO-5 через 70 дней после первой инокуляции опухоли. В результате авторы изобретения не наблюдали опухолевого роста у этих мышей (Фиг. 14), продемонстрировав таким образом, что вакцинация B/αGalCer/pep устанавливала иммунитет памяти в отношении этой опухоли. Когда авторы изобретения провели аналогичные эксперименты, используя опухоль другого происхождения, OVA-трансфектированную тимому (EG-7), наблюдались очень похожие и согласующиеся результаты (Фиг. 36).
Авторы настоящего изобретения исследовали вопрос о том, могла ли вакцинация B/αGalCer/pep устранять уже существующую опухоль. С этой целью авторы изобретения установили две терапевтические модели; мышей вакцинировали (i) один день или (ii) девять дней после подкожной трансплантации, когда опухоли стали пальпируемыми.
В однодневной модели вакцинация либо ДК/pep либо ДК/αGalCer/pep почти полностью подавляла рост опухоли. Интересно, что опухолевый рост также полностью был сокращен у мышей, вакцинированных B/αGalCer/pep (Фиг. 15). В девятидневной модели ни одна из этих вакцинаций полностью не разрушала растущую опухоль вследствие очень агрессивной природы меланомы B16. Однако у мышей, вакцинированных B/αGalCer/pep, рост опухоли был менее выражен, чем в группе мышей «только В», и напоминал то, что наблюдалось в группах, вакцинированных ДК/pep или ДК/αGalCer/pep (Фиг. 16).
7-2. In vivo тест с использованием моделей Her-2/neu
Для исследования того, можно ли применять вакцину на основе В-клеток для конкретного опухолевого Аг, эксперименты проводили с использованием TAUF (Penichet ML et al., Lab Anim Sci., 49: 179-88, 1999), опухолевой клеточной линии, экспрессирующей Her-2/neu, образом, аналогичным описанному в приведенном выше Примере <7-1>. Этот опухолевый антиген хорошо охарактеризован и известен его CTL эпитоп. Еще раз авторы наблюдали значительный уровень Her-2/neu-спепцифичной цитотоксичности in vivo у мышей, которым вводили В-клетки, нагруженные αGalCer, сенсибилизированные Her-2/neu63-71 (Фиг. 17).
Для проверки противоопухолевой активности в этой модели авторы настоящего изобретения внутривенно или подкожно вводили опухолевые клетки, экспрессирующие Her-2/neu (TAUF) мышам BALB/c до вакцинации их В-клетками, сенсибилизированными Her-2/neu63-71, нагруженными αGalCer. После внутривенной инокуляции опухоли выживаемость была несколько лучше у мышей, вакцинированных B/αGalCer или B/pep, чем у мышей, вакцинированных только В (Фиг. 18). Напротив, все мыши, вакцинированные B/αGalCer/pep, выживали на протяжении всего эксперимента. Эти выжившие мыши также были резистентны повторному заражению TAUF. Подобные результаты наблюдались в модели подкожного роста опухоли (Фиг. 19).
Совместно было доказано, что схема введения вакцины на основе В-клеток является такой же эффективной, что и вакцин на основе ДК в выработке как профилактического, так и терапевтического противоопухолевого иммунитета.
Пример 8
Введение целого антигена с использованием антиген-экспрессирующего вирусного вектора
Сенсибилизированная пептидом клеточная вакцина в большинстве случаев не может быть использована, поскольку применение пептида ограничено гаплотипами главного комплекса гистосовместимости (MHC) и может только представлять единичный эпитоп. С другой стороны, опосредованный вирусом целый антиген может быть широко использован без ограничения в гаплотипах MHC и может индуцировать как гуморальный иммунный ответ, так и иммунный ответ, опосредованный клетками, что указывает на то, что он может индуцировать различные эпитоп-специфичные иммунные ответы. Основываясь на этом, авторы изобретения исследовали, могли ли В-клетки, трансдуцированные целым антигеном посредством вирусного вектора, эффективно индуцировать иммунный ответ. Сначала авторы настоящего изобретения получили аденовирус (AdHM), доставляющий ген, кодирующий внеклеточный домен и трансмембранный домен, ассоциированный с опухолью антигена Her-2/neu (HM), а затем инъекционно вводили в мышиную модель.
В частности, В-клетки отделяли от спленоцитов мышей BALB/c, а затем культивировали для трансдукции с 100 MOI аденовируса, который вводят HM (AdHM, Viromed) в бессывороточных условиях при 37°C в течение 90 минут. Затем туда добавлялась сыворотка с последующим дополнительным культивированием в течение 8 часов, в течение 24 часов и в течение 48 часов соответственно. Клетки окрашивали PE меченным анти-Her-2/neu антителом (BD biosciences #340552) для проточной цитометрии. Измеряли уровень Her-2/neu, экспрессированного на поверхности клетки (исследовали эффективность аденовирусной трансдукции для В-клеток и процентное содержание PE+ клеток во всех В-клетках).
Через 8 часов, 24 часа и 48 часов культивирования В-клеток с аденовирусом было подтверждено, что эффективность трансдукции была более 20% во всех условиях культивирования (Фиг. 20). В частности, когда В-клетки культивировали совместно с AdHM в течение более 8 часов, В-клетки были полностью трансдуцированы AdHM, и, следовательно Her-2/neu экспрессировался на поверхностях В-клеток. На экспрессию Her-2/neu на поверхности В-клеток не влияло культивирование с αGalCer. Кроме того, на способность В-клетки к презентации αGalCer также не влияло (Фиг. 21). Как объяснялось выше, представленные выше результаты указывают на то, что когда αGalCer представлен клеткам DN32.D3 молекулой CD1d на В-клетке, активированные DN32.D3 клетки секретируют IL-2. Следовательно, измерение уровня IL-2 в культуральном супернатанте может подтвердить способность В-клеток к презентации αGalCer, и в настоящем изобретении была подтверждена секреция IL-2 в культуральном супернатанте, подтверждая, что В-клетки могут презентировать αGalCer.
Пример 9
Цитотоксический Т-клеточный ответ, индуцированный В-клетками, трансдуцированными аденовирусом
В-клетки выделяли из спленоцитов мышей BALB/c, которые трансдуцировали 100 M.O.I. аденовируса, экспрессирующего внеклеточный домен Her-2/neu и трансмембранный домен, в бессывороточных условиях в инкубаторе при 37°C в течение 60 минут. Затем туда добавляли сыворотку с последующим дополнительным культивированием в течение 24 часов для получения B/AdHM клеток. AdHM-трансдуцированные В-клетки культивировали в сыворотке, содержащей среду, дополненную 1-2 мкг/мл αGalCer, в течение 23 часов, с получением в результате В-клеток, нагруженных αGalCer (B/AdHM/αGalCer). Полученные В-клетки промывали RPMI более трех раз для удаления оставшегося аденовируса и αGalCer.
Авторы настоящего изобретения вводили В-клеточную вакцину внутривенно мышам для исследования, могли ли В-клетки, трансдуцированные AdHM, индуцировать цитотоксический иммунный ответ. Мыши, которым вводили одни только В-клетки (B) и В-клетки, нагруженные αGalCer (B/αGalCer), использовались в качестве отрицательного контроля. Измеряли Her-2/neu специфичные Т-клеточные ответы, индуцированные В-клетками, трансдуцированными AdHM (B/AdHM) и В-клетками, сенсибилизированными совместно αGalCer и AdHM (B/AdHM/αGalCer). После иммунизации вводили клетки-мишени, нагруженные пептидом эпитопа цитотоксической Т-клетки (Her-2/neu63-71, Anygene), с последующим исследованием in vivo активности CTL.
В результате через одну неделю после иммунизации лизис клеток-мишеней не наблюдался в группе, иммунизированной только В и B/αGalCer, тогда как цитотоксический Т-клеточный ответ наблюдался в группе мышей, иммунизированных B/AdHM и B/AdHM/αGalCer (Фиг. 22). Трансдуцированная аденовирусом В-клеточная активация может эффективно индуцировать in vivo цитотоксический Т-клеточный ответ без помощи Т-клеток-природных киллеров, указывая на то, что они могут быть использованы в качестве антиген-представляющих клеток. Цитотоксический Т-клеточный ответ, индуцированный B/AdHM и B/AdHM/αGalCer, постепенно снижался, но вплоть до 7 недель после иммунизации, этот ответ длился на значительном уровне (Фиг. 23). Через одну неделю после иммунизации измеряли секрецию IFN-γ у CD8+ Т-клеток вместе с цитотоксическим Т-клеточным ответом. В результате количество активированных CD8+ Т-клеток было увеличено в группе мышей, обработанных B/AdHM и B/AdHM/αGalCer, по сравнению с группой, обработанной только В-клетками, и более конкретно число CD8+ Т-клеток в группе, обработанной B/AdHM/αGalCer, было немного выше, чем в группе, обработанной B/AdHM (Фиг. 24).
Пример 10
Активация клеток-природных киллеров В-клетками, нагруженными αGalCer, и улучшение вакцинного эффекта В-клеточной вакцины, трансдуцированной аденовирусом
10-1. Активация клеток-природных киллеров В-клетками, нагруженными αGalCer
Активированные клетки-природные киллеры подсчитывали исходя из секретирующих IFN-γ. В частности, В-клетки выделяли из спленоцитов мышей, а затем культивировали в среде, содержащей 1 мкг/мл GolgiPlug, в течение 5 часов. Клетки окрашивали антимышиным IFN-γ:АПК, CD3:PE и CD49b:FITC антителом (все Biolegend). Клетки-природные киллеры состояли из тех, которые экспрессировали CD49b, маркера клеток-природных киллеров, но не CD3.
Цитотоксический Т-клеточный ответ в группе, обработанной В-клетками, сенсибилизированными совместно аденовирусом и αGalCer (B/AdHM/αGalCer), был выше и длился дольше, чем в группе, обработанной В-клетками, трансдуцированными аденовирусом (B/AdHM), но различие не было значимым. Эти результаты указывают на то, что нагрузка αGalCer не усиливает цитотоксический Т-клеточный ответ, в отличие от сенсибилизированной пептидом вакцины на основе В-клеток. Но введение вакцины на основе В-клеток, нагруженных αGalCer, стимулировало Т-клетки-природные киллеры in vivo, в отличие от вакцины на основе В-клеток, трансдуцированных аденовирусом, и тем самым активировало клетки-природные киллеры (Фиг. 25). Можно ожидать противоопухолевый эффект путем активации клеток-природных киллеров, и тем самым индуцируя цитотоксический Т-клеточный ответ.
10-2. Улучшение вакцинного эффекта вакцины на основе В-клеток, нагруженных совместно αGalCer и аденовирусом
Для исследования эффективности индукции цитотоксического Т-клеточного ответа В-клеточной вакцины авторы настоящего изобретения вводили клеточную вакцину в субоптимальной дозе, которой недостаточно для индукции полного цитотоксического Т-клеточного ответа, а затем определяли лизис клеток-мишеней. Группу положительного контроля обрабатывали В-клетками, нагруженными αGalCer и пептидом эпитопа цитотоксической Т-клетки (B/αGalCer/pep). Спустя одну неделю после иммунизации каждой В-клеточной вакциной эффективность индукции цитотоксического Т-клеточного ответа сравнивали между группами, обработанными соответственно B/AdHM и B/AdHM/αGalCer (Фиг. 26). В группе, которой вводили B/αGalCer/pep, считалось что большинство В-клеток представляют антиген, но только 20% всех В-клеток, трансдуцированных аденовирусом, могли работать как антиген-представляющие клетки. Таким образом, считается, что В-клеточная вакцина, трансдуцированная аденовирусом для введения целого антигена, может индуцировать антиген-специфичный цитотоксический Т-клеточный ответ эффективно, даже несмотря то, что доза выше, чем доза сенсибилизированной пептидом В-клеточной вакцины. Кроме того, когда вводили небольшое количество В-клеток (неполная эффективная доза), оно также могло индуцировать лизис клеток-мишеней с помощью Т-клеток-природных киллеров, активированных нагруженным αGalCer.
Пример 11
В-клеточная вакцина, трансдуцированная аденовирусом, индуцирующая антиген-специфический антительный ответ
Вакцина на основе В-клеток, трансдуцированных аденовирусом, обладает преимуществом одновременно индуцировать гуморальный иммунный ответ и иммунный ответ, опосредованный клетками, в отличие от сенсибилизированной пептидом В-клеточной вакцины, индуцирующей только клеточно-опосредованный иммунный ответ. Для подтверждения, могла ли В-клеточная вакцина на основе AdHM индуцировать антитело, специфичное к Her-2/neu, мышей BALB/c иммунизировали B/AdHM и B/AdHM/αGalCer соответственно. Для измерения титра анти-Her-2/neu антитела в сыворотке, связывание антитела с TAUF клетками, мышиные/крысиные опухолевые клетки, экспрессирующие Her-2/neu на поверхности, исследовали с помощью проточной цитометрии.
Для определения антительного ответа образцы крови получали на 1-, 3-, 5- и 7-й неделе от иммунизации путем кровопускания из глаза. Образцы крови мышей стояли при комнатной температуре в течение 2 часов, с последующим центрифугированием при 8000 в течение 10 минут для отделения сыворотки. Для измерения титра анти-Her-2/neu антитела в сыворотке сыворотку серийно разводили в два раза от 1:50 (с использованием PBS, содержащего 1% FBS, 0,09% азида), с последующим культивированием с TAUF, опухолевыми клетками, экспрессирующими Her-2/neu, при 4°C в течение 60 минут. TAUF клетки, инкубированные с мышиной сывороткой, промывали буфером, с последующим окрашиванием мышиного антитела, связывающегося с TAUF клетками, используя меченное FITC козье антитело против мышиного IgG в качестве вторичного антитела. Затем количество мышиных антител, связавшихся с TAUF, измеряли с помощью проточного цитометра (FACSCaliber, BD Biosciences), и титр антител рассчитывали путем стандартизации сыворотки интактной мыши, и принимая среднюю интенсивность флуоресценции, повышенную более чем в 1,8 раз за положительный ответ.
В результате согласуясь с результатом индукции цитотоксических Т-клеток, титры анти-Her-2/neu антител в группах, иммунизированных B/AdHM и B/AdHM/αGalCer, удерживались высокими (Фиг. 27), и гуморальный ответ также был индуцирован в вышеуказанных двух группах, в отличие от групп, обработанных В-клетками и B/αGalCer (Фиг. 28).
Пример 12
Профилактическое и терапевтическое действие на злокачественные опухоли путем иммунизации вакциной на основе В-клеток, трансдуцированных аденовирусом, нагруженных αGalCer
2·105 клеток TAUF вводили в хвостовую вену мышей BALB/c, опухолевой модели Her-2/neu для индукции опухоли. Затем мышей, несущих опухоль, иммунизировали В-клеточными вакцинами с последующим исследованием выживаемости.
Авторы изобретения проверили, могла ли В-клеточная вакцина, трансдуцированная аденовирусом, индуцировать опосредованный клетками иммунный ответ и гуморальный иммунный ответ вместе, и посредством которого обладает противоопухолевой активностью. TAUF клетки внутривенно вводили мышам для индукции рака легких, при котором определяли профилактический и терапевтический эффект В-клеточной вакцины. В-клеточную вакцину вводили для иммунизации в концентрации 2·106, а 7 дней спустя 2·105 опухолевых клеток внутривенно вводили мышам, с последующим изучением выживаемости. Период выживания был длительным в группах, обработанных В-клетками, нагруженными αGalCer (B/αGalCer), и В-клетками, трансдуцированными аденовирусом, экспрессирующими опухолевый антиген (B/AdHM), по сравнению с выживаемостью в группе, обработанной только В-клетками (B). Когда мышей иммунизировали В-клеточной вакциной, нагруженной αGalCer, трансдуцированной аденовирусом, экспрессирующей опухолевый антиген (B/AdHM/αGalCer), мыши выживали на протяжении всего эксперимента. Эти результаты указывают на то, что B/AdHM/αGalCer вакцина обладает лучшим противоопухолевым эффектом, чем B/αGalCer или B/AdHM вакцина (Фиг. 29, вверху). И было решено, что иммунный ответ, индуцированный В-клеточной вакциной, может эффективно предотвращать рост опухоли.
Далее, для подтверждения терапевтичского эффекта В-клеточной вакцины 5·104 опухолевых клеток (TAUF) вводили инъекционно в хвостовую вену для индукции рака легких. Три дня спустя группы мышей иммунизировали 1·106 В-клеток, B/αGalCer, B/AdHM и B/AdHM/αGalCer соответственно. Самое короткое время наблюдалось в группе, обработанной только В-клетками, а время выживания группы B/αGalCer было сходным. В то же время время выживания в группе, иммунизированной B/AdHM вакциной, было несколько удлинено по сравнению с животными групп, обработанных только В-клетками и B/αGalCer. Напротив, период выживания был существенно удлинен в группе, иммунизированной B/AdHM/αGalCer, что согласовалось с результатом профилактических моделей, указывая на то, что вакцина B/AdHM/αGalCer обладает лучшим противоопухолевым эффектом, чем В-клетки, B/αGalCer или B/AdHM вакцина (Фиг. 29, внизу).
Пример составления лекарственного средства. Получение инъекционного раствора
Инъекционный раствор противоопухолевой вакцины согласно изобретению готовили следующим образом:
1-2 мкг/мл αGalCer, 5·106 клеток/мл В-клеток, 1-2 мкг/мл пептида, 1 г 5'-хлор-3,2'-дигидроксихалкона или 5'-хлор-2,3'-дигидроксихалкон гидрохлорида, 0,6 г NaCl и 0,1 г аскорбиновой кислоты растворяли в дистиллированной воде, получая в результате 100 мл раствора для инъекций. Раствор разливали в бутылку, который стерилизовали при 120°C в течение 30 минут.
Промышленная применимость
Как объяснялось выше, В-клетки согласно изобретению, которые нагружены лигандом Т-клеток-природных киллеров, в частности αGalCer, и антигеном, не только индуцируют цитотоксический Т лимфоцитарный ответ на уровне, аналогичном уровню, индуцируемым общепринятой ДК вакциной, но также оказывают профилактическое и терапевтическое действие на солидную опухоль и метастазирующую опухоль. В-клетки, трансдуцированные аденовирусом, кодирующим опухолевый антиген, могут индуцировать антиген-специфичные иммунные ответы таким образом, что вакцина, опосредованная В-клетками, может быть использована в качестве профилактического и терапевтического средства для лечения опухолей. Кроме того, вакцина по изобретению может индуцировать иммунный ответ даже без помощи CD4+ Т-клеток. Следовательно, ее можно эффективно вводить пациентам, инфицированным ВИЧ, имеющим иммунный дефицит, вызванный недостатком CD4+ Т-клеток.
Специалистам в данной области будет понятно, что концепции и конкретные варианты осуществления, раскрытые в приведенном выше описании, могут быть использованы в качестве основы для модификации или создания других вариантов осуществления для выполнения тех же задач согласно изобретению. Специалистам в данной области также будет понятно, что каждые эквивалентные варианты осуществления не отклоняются от сущности и объема изобретения, как изложено в прилагаемой формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННЫЙ ПРЕПАРАТ ДЛЯ УВЕЛИЧЕНИЯ ИММУНОГЕННОСТИ ВАКЦИНЫ (ВАРИАНТЫ) | 2008 |
|
RU2491090C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ УСИЛЕНИЯ ИМУННОГО ОТВЕТА НА АНТИГЕНЫ | 2008 |
|
RU2537188C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6 | 2017 |
|
RU2733886C2 |
| ПОЛИЭПИТОПНЫЕ ИММУНОГЕННЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2598265C2 |
| ВНУТРИКЛЕТОЧНЫЙ ДОМЕН БЕЛКА HER-2/NEU ДЛЯ ПРЕДУПРЕЖДЕНИЯ ИЛИ ЛЕЧЕНИЯ МАЛИГНИЗАЦИЙ | 1996 |
|
RU2236461C2 |
| CDH3-ПЕПТИД И ВКЛЮЧАЮЩЕЕ ЕГО ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2008 |
|
RU2483078C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК, ОБЛАДАЮЩИХ АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2596920C2 |
| Полиэпитопная противоопухолевая вакцинная конструкция, содержащая эпитопы опухоль-ассоциированных антигенов, фармацевтическая композиция и ее применение для стимуляции специфического противоопухолевого иммунного ответа | 2016 |
|
RU2684235C2 |
| Способ активации цитотоксических Т-лимфоцитов в популяции мононуклеарных клеток крови при помощи мембранных везикул, полученных из генетически модифицированных опухолевых клеток меланомы М14 со сверхэкспрессией интерлейкина 2 | 2022 |
|
RU2790867C1 |
| СПОСОБ ИММУНОТЕРАПИИ ХРОНИЧЕСКОГО ВИРУСНОГО ГЕПАТИТА С | 2016 |
|
RU2637631C2 |

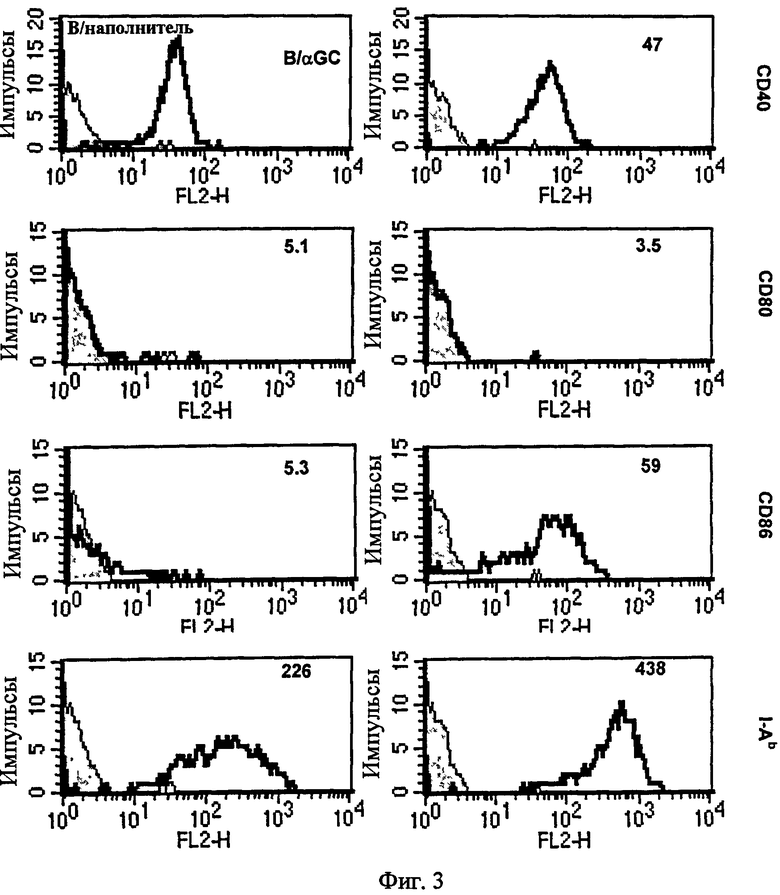
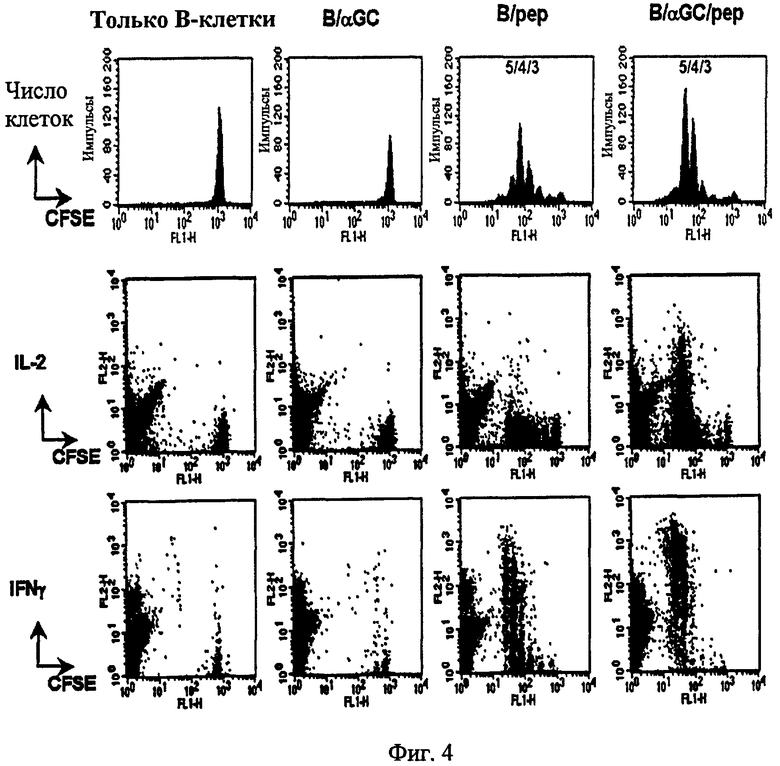
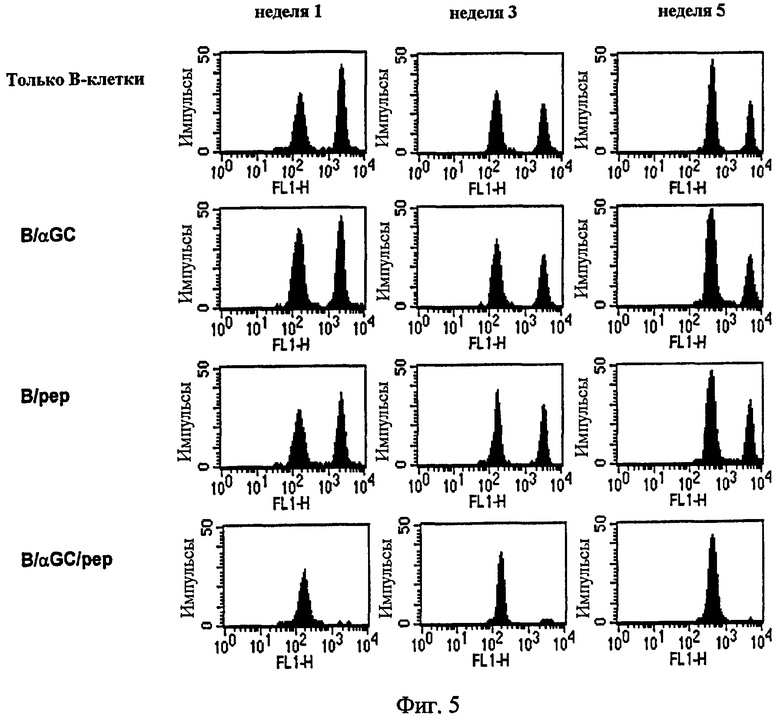
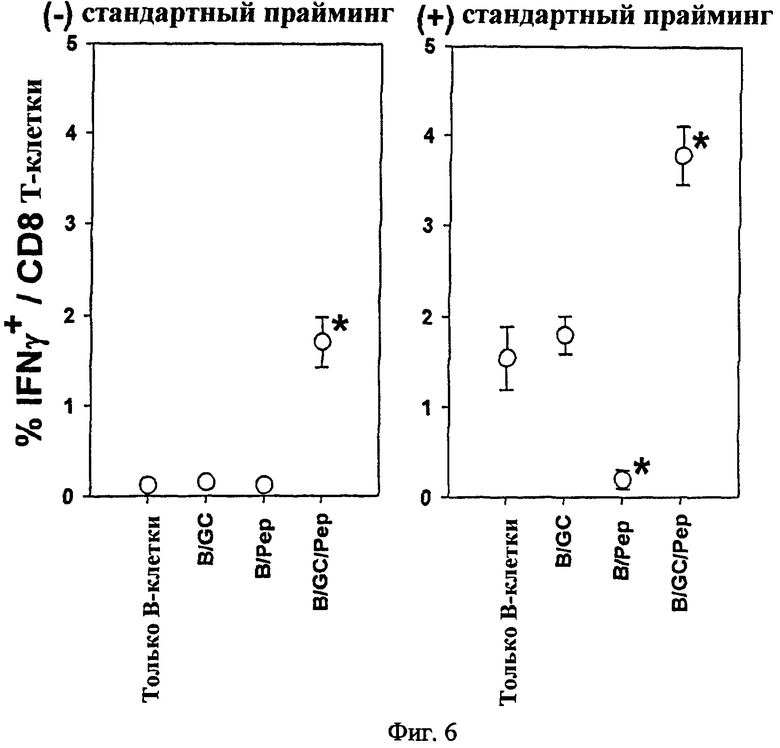
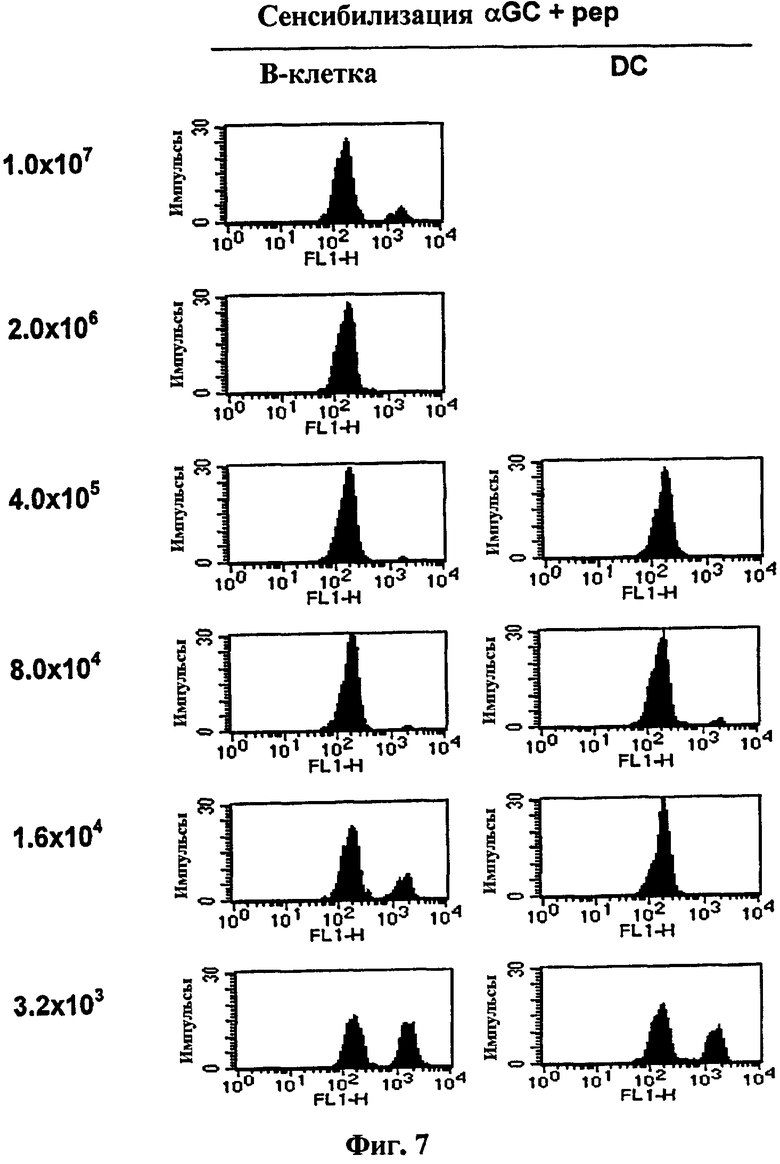
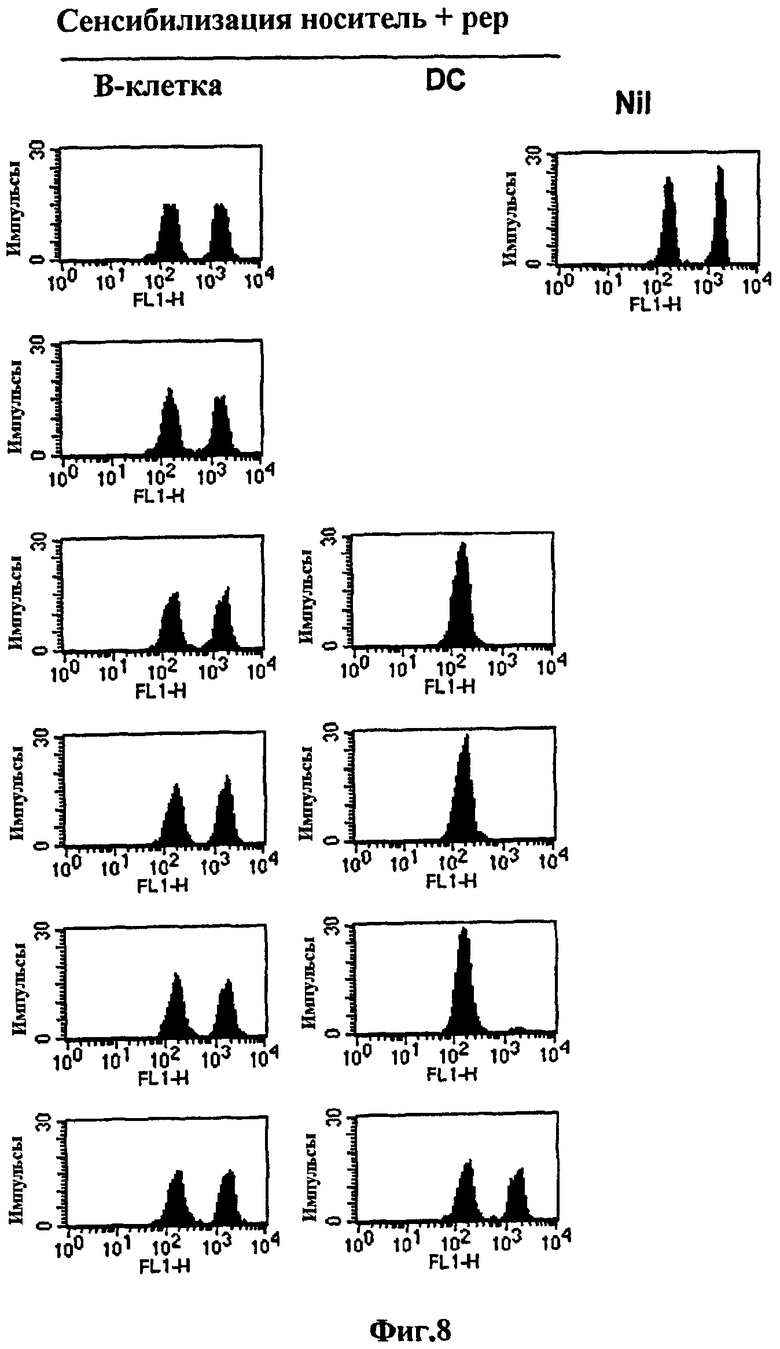
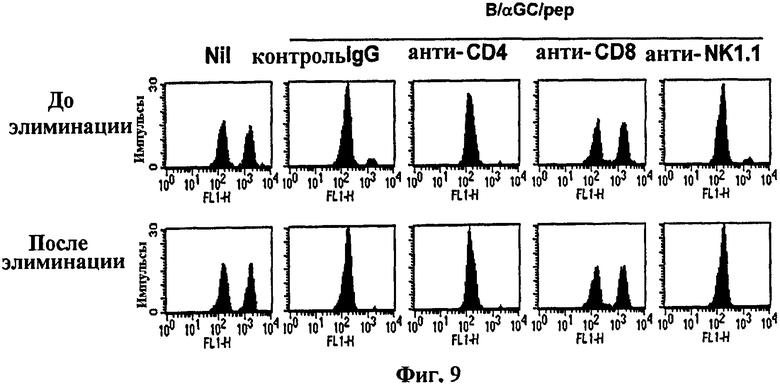
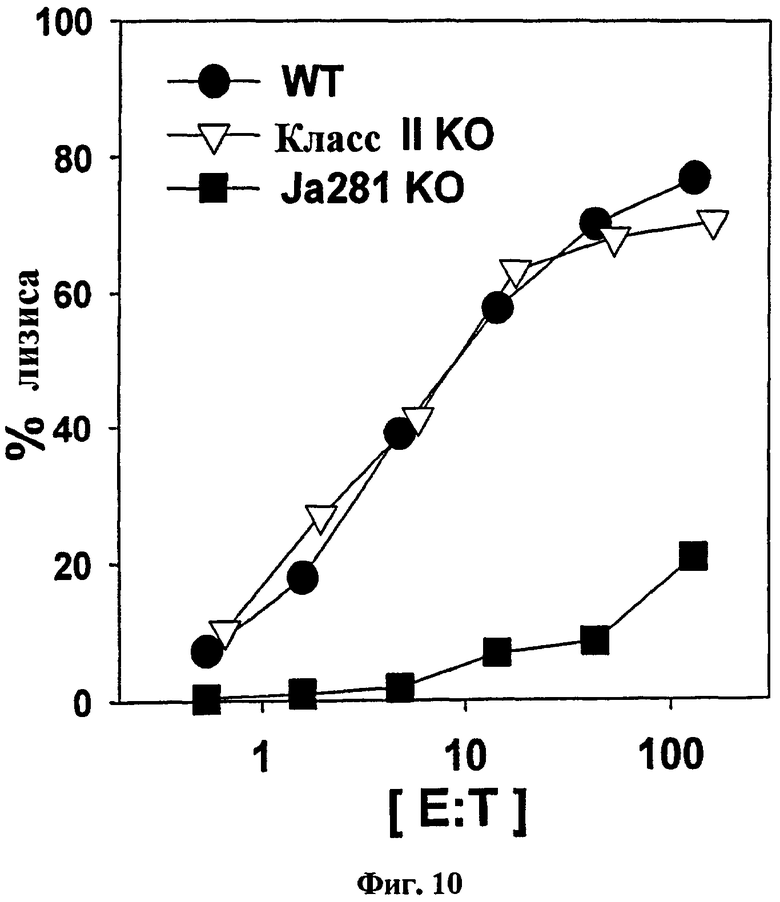
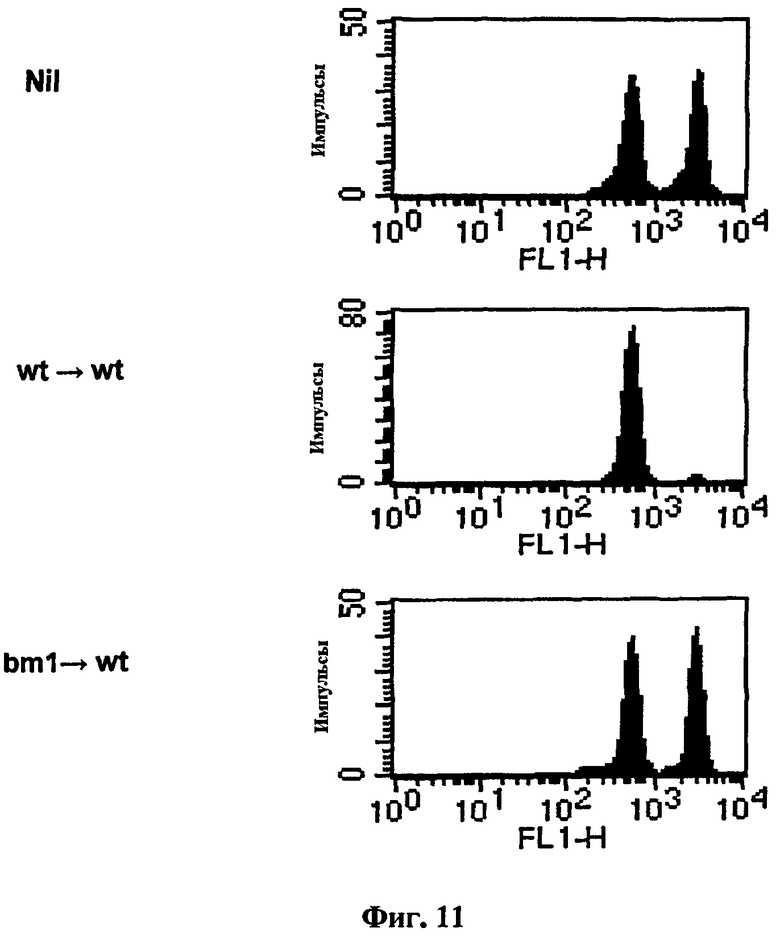
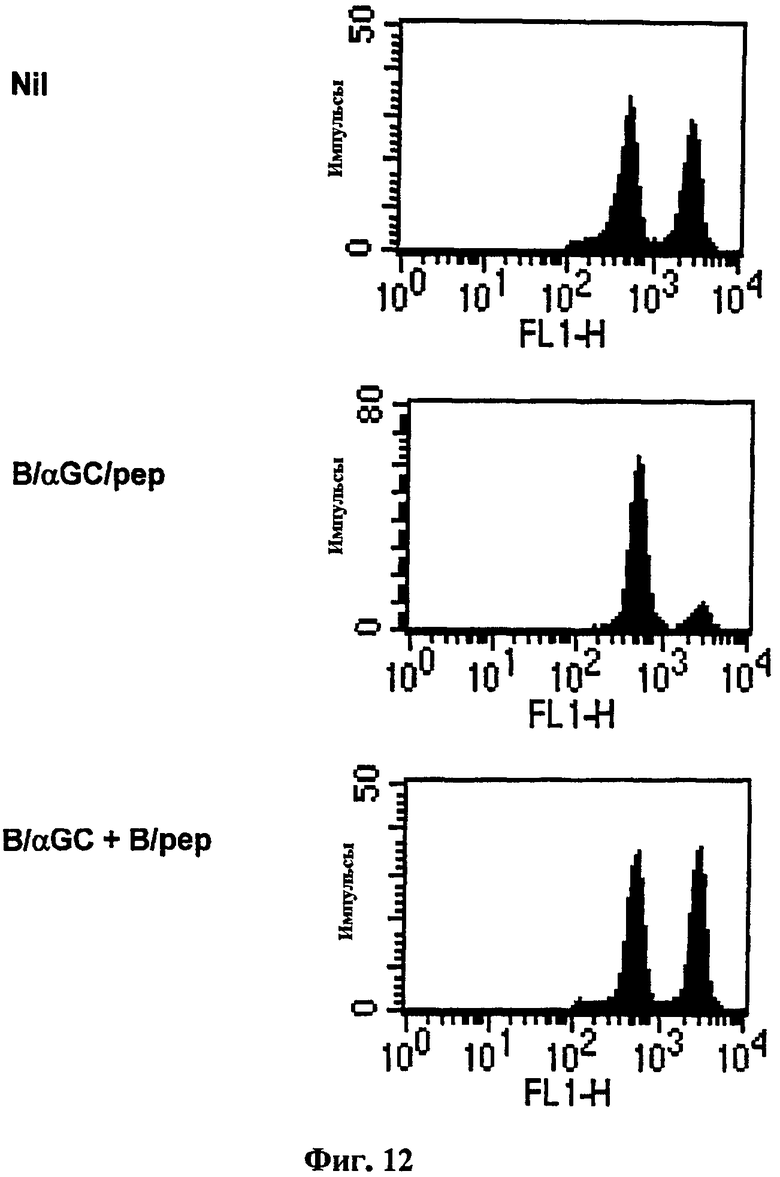
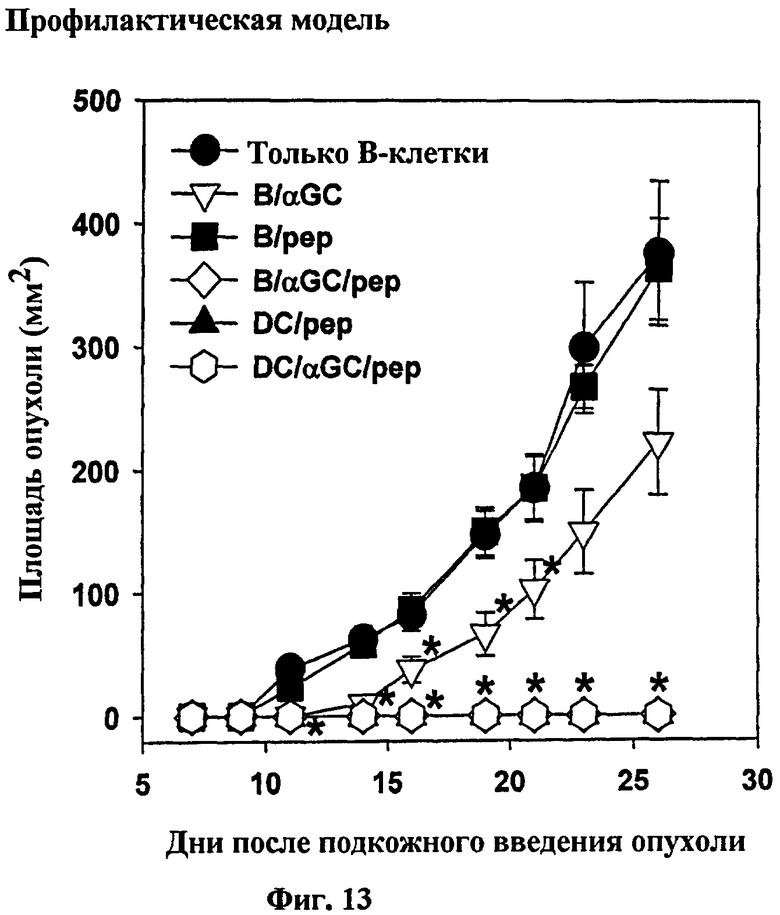
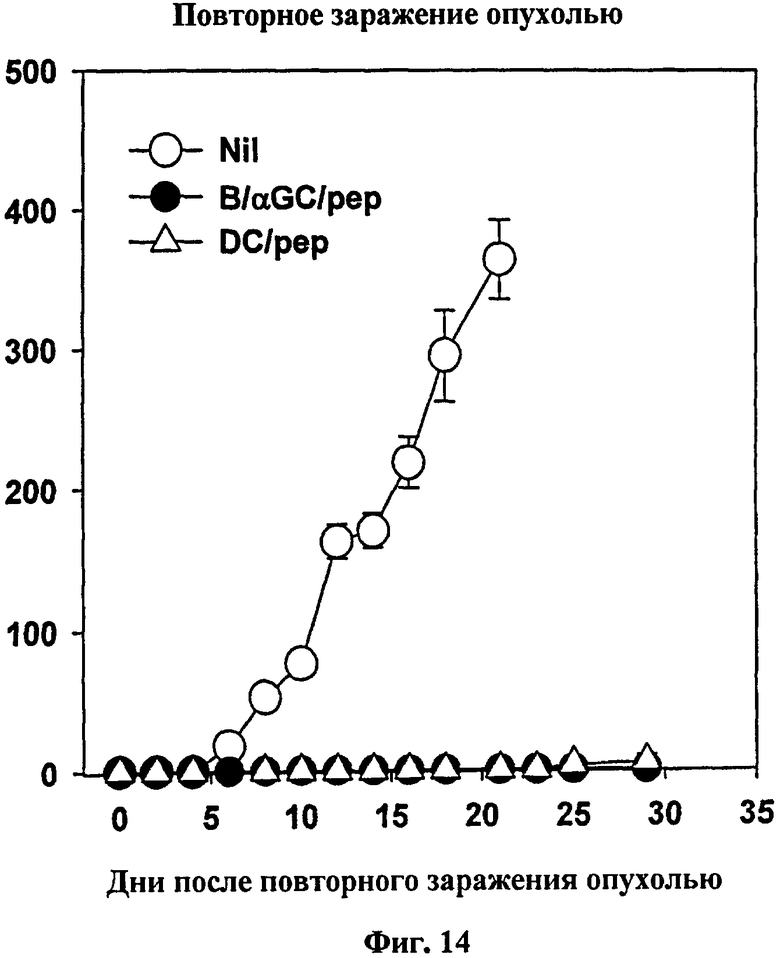
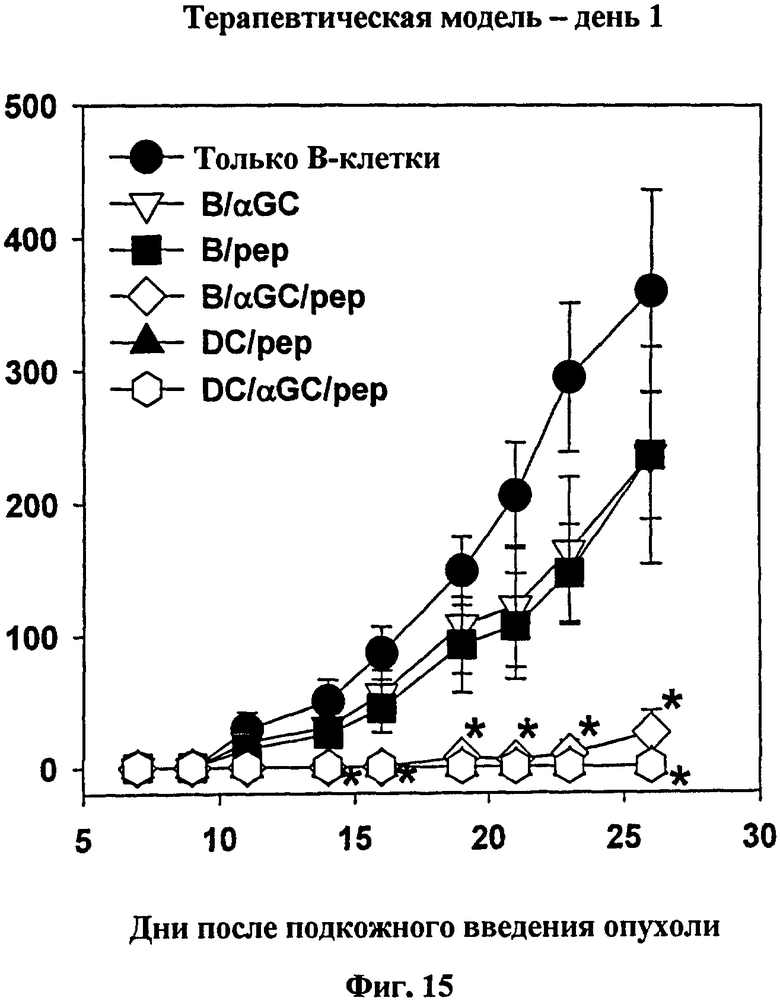

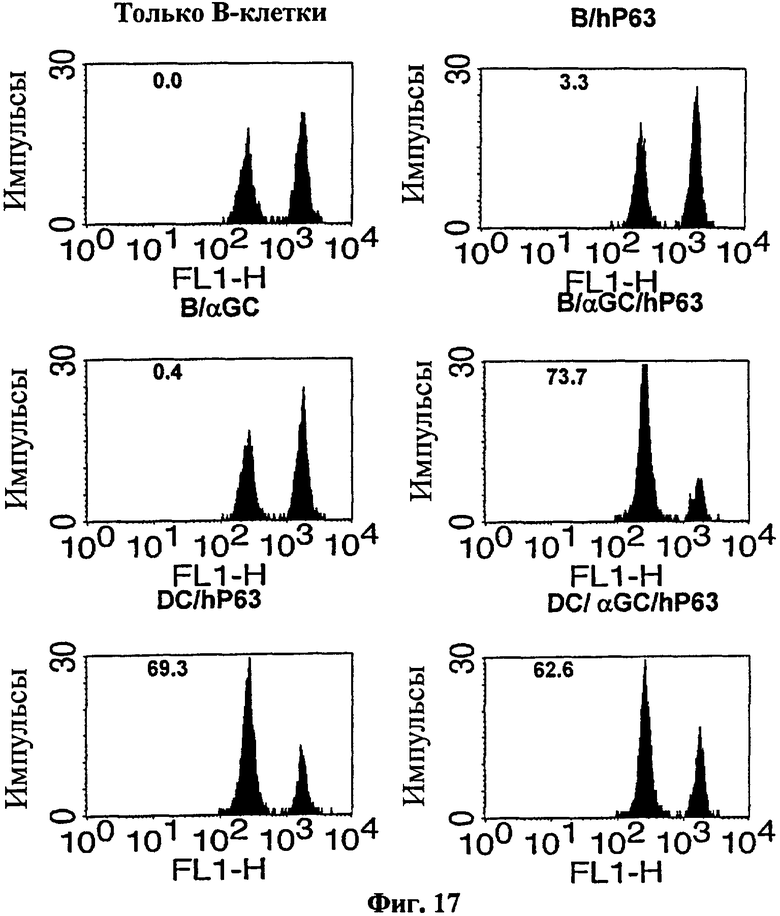
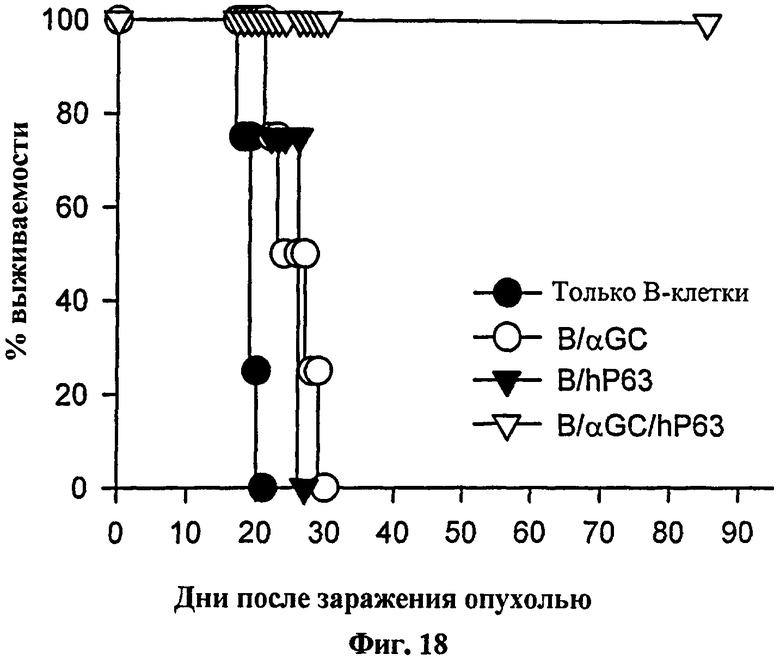
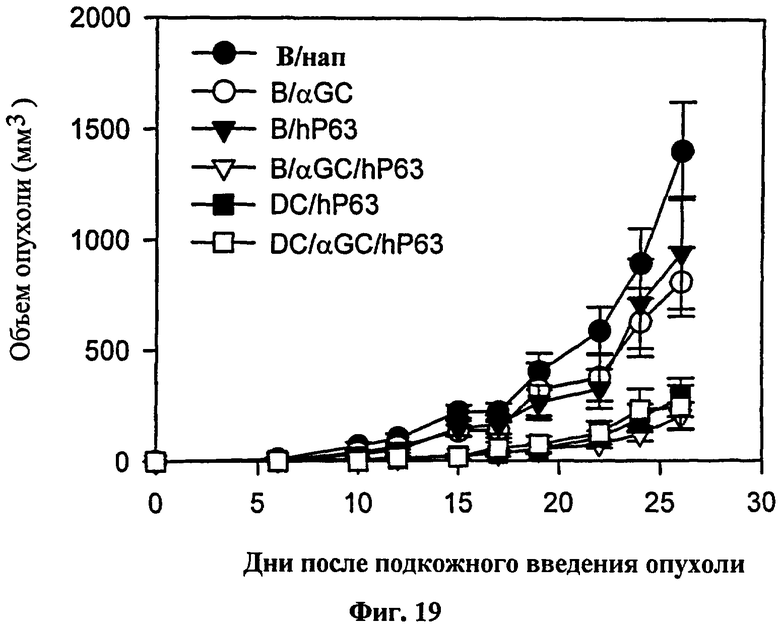

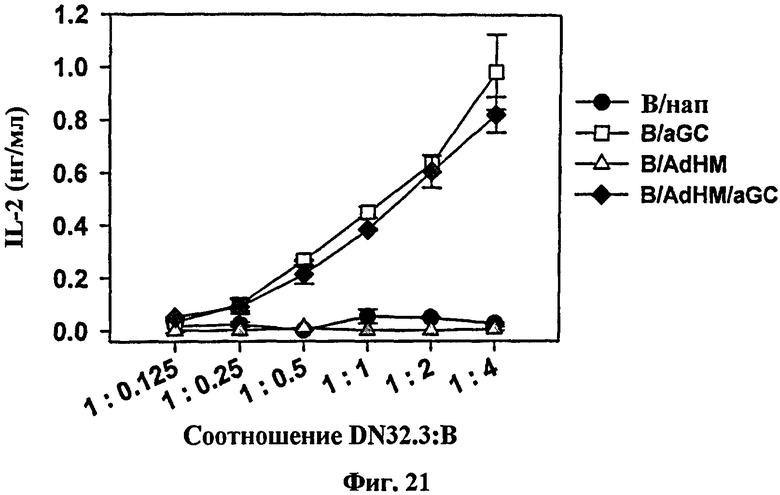


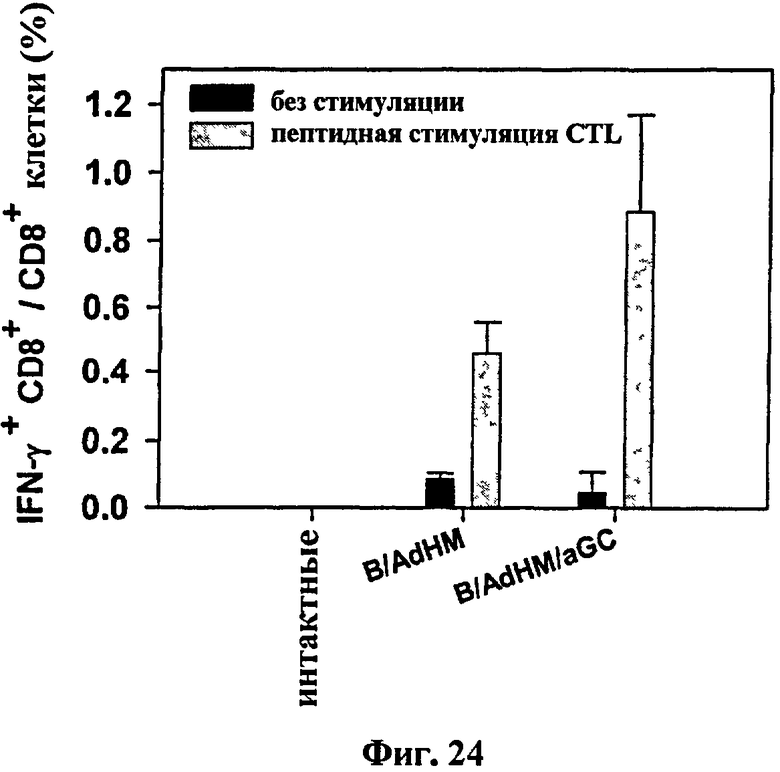

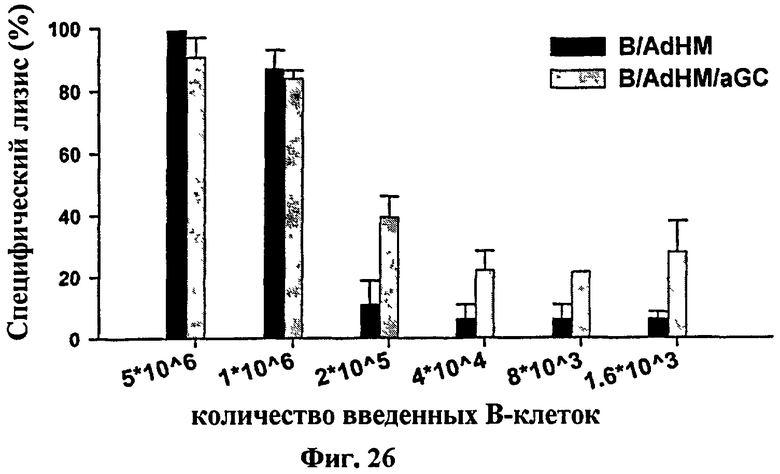
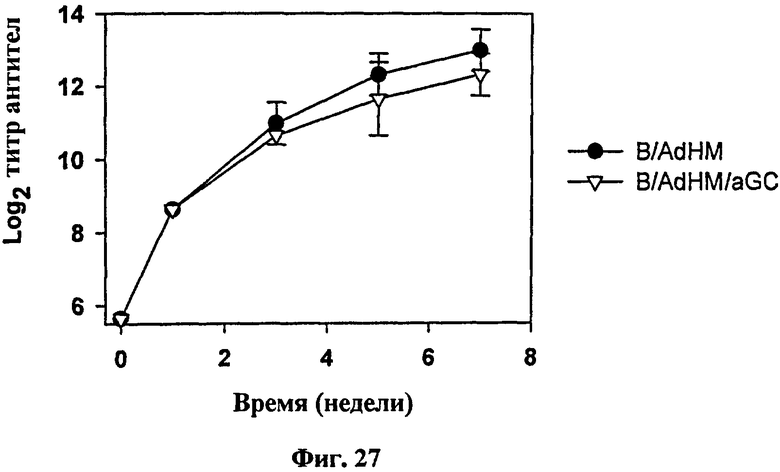
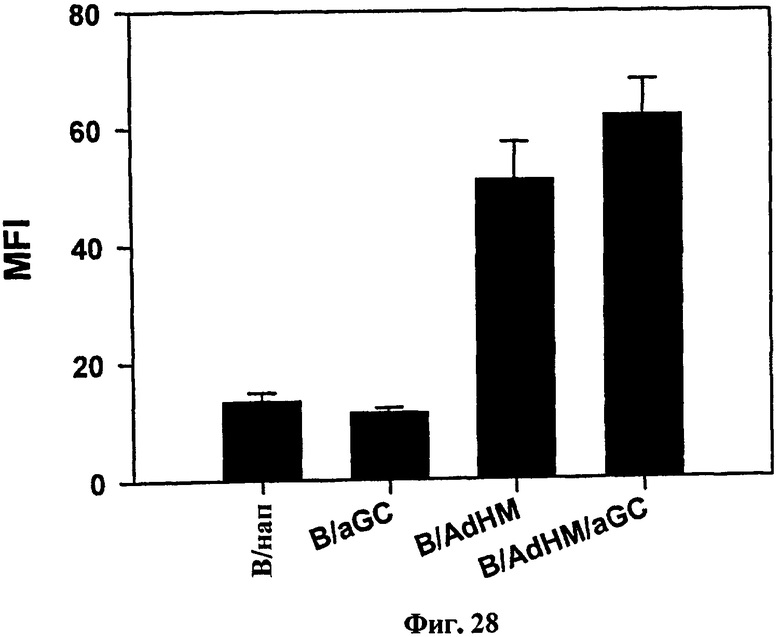
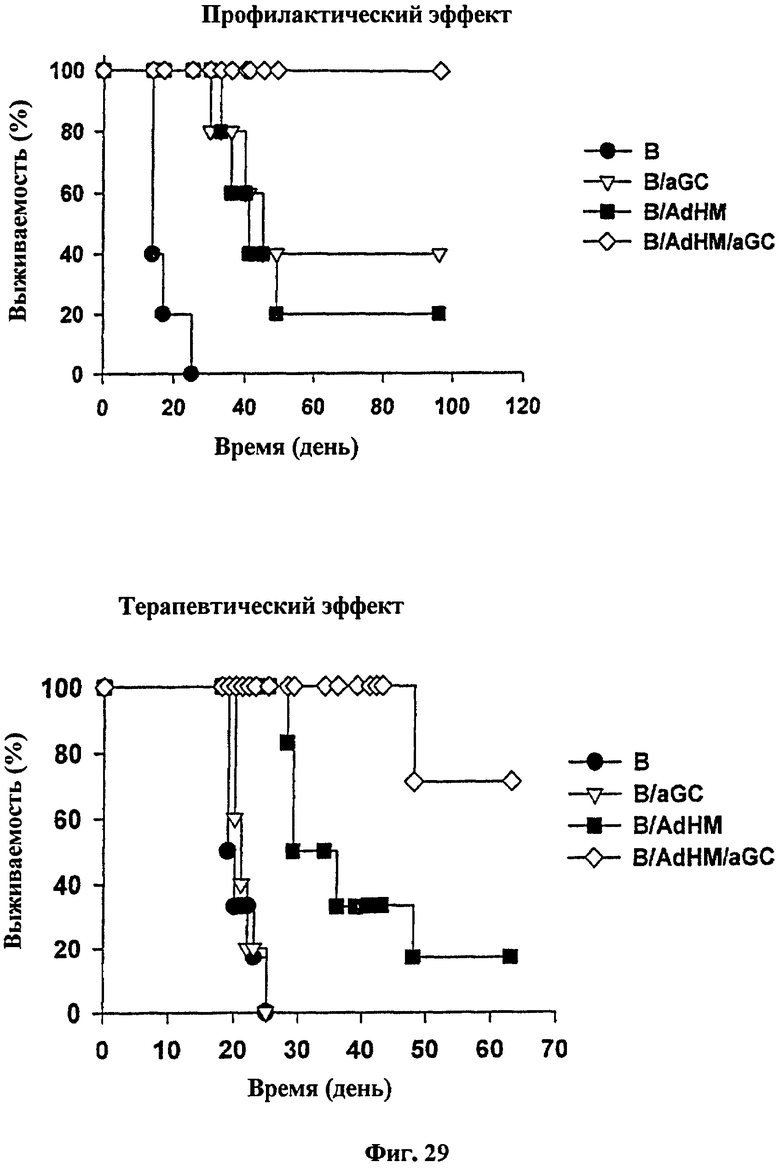
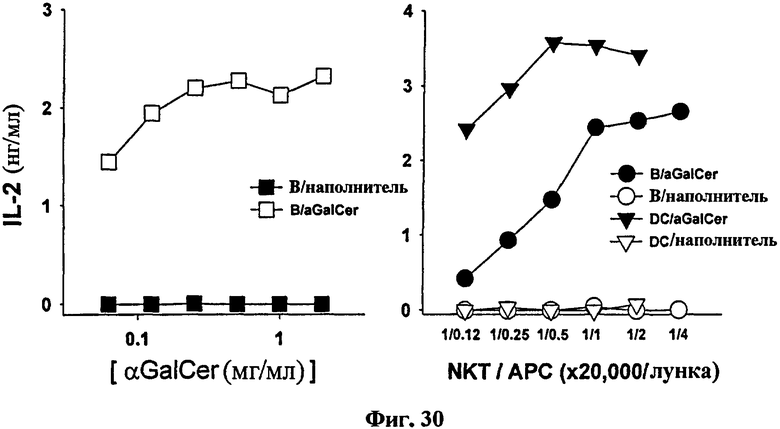

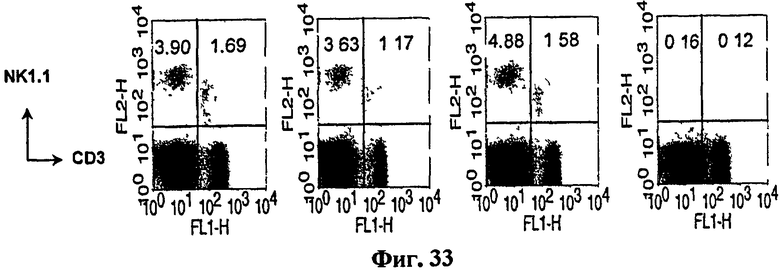
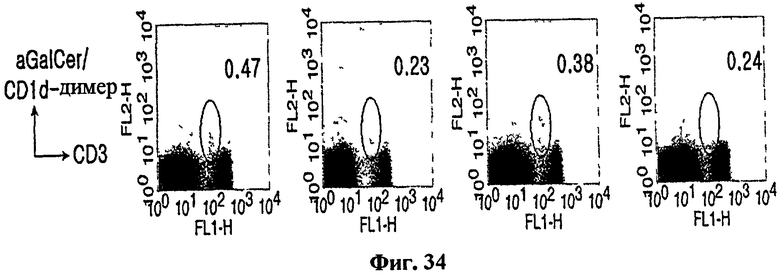
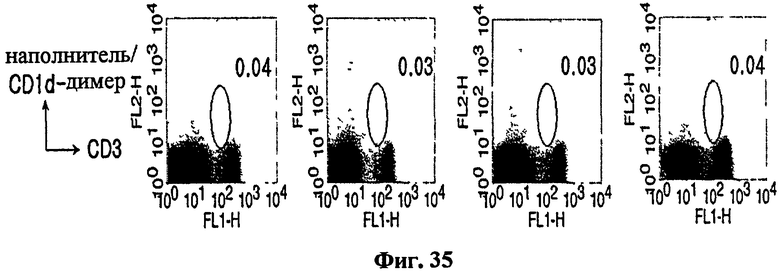

Изобретение относится к биотехнологии. Описана иммунотерапевтическая или профилактическая вакцина, применяемая при/против вирусного инфекционного заболевания, бактериального инфекционного заболевания или рака, содержащая фармацевтически эффективное количество В-клеток, выбранных из группы, состоящей из В-клеток, полученных путем нагрузки их лигандом Т-клеток-природных киллеров и сенсибилизирующим антигеном, и В-клеток, полученных путем нагрузки их лигандом Т-клеток-природных киллеров и переноса гена, кодирующего пептидный антиген или белковый антиген, посредством которого В-клетки экспрессируют антиген, и фармацевтически приемлемый носитель. Предложен способ индукции цитотоксического ответа у нуждающегося в лечении пациента, предусматривающий введение этому пациенту описанной вакцины. Изобретение расширяет арсенал средств иммунотерапии или профилактики вирусного инфекционного заболевания, бактериального инфекционного заболевания или рака. 2 н. и 9 з.п. ф-лы, 36 ил.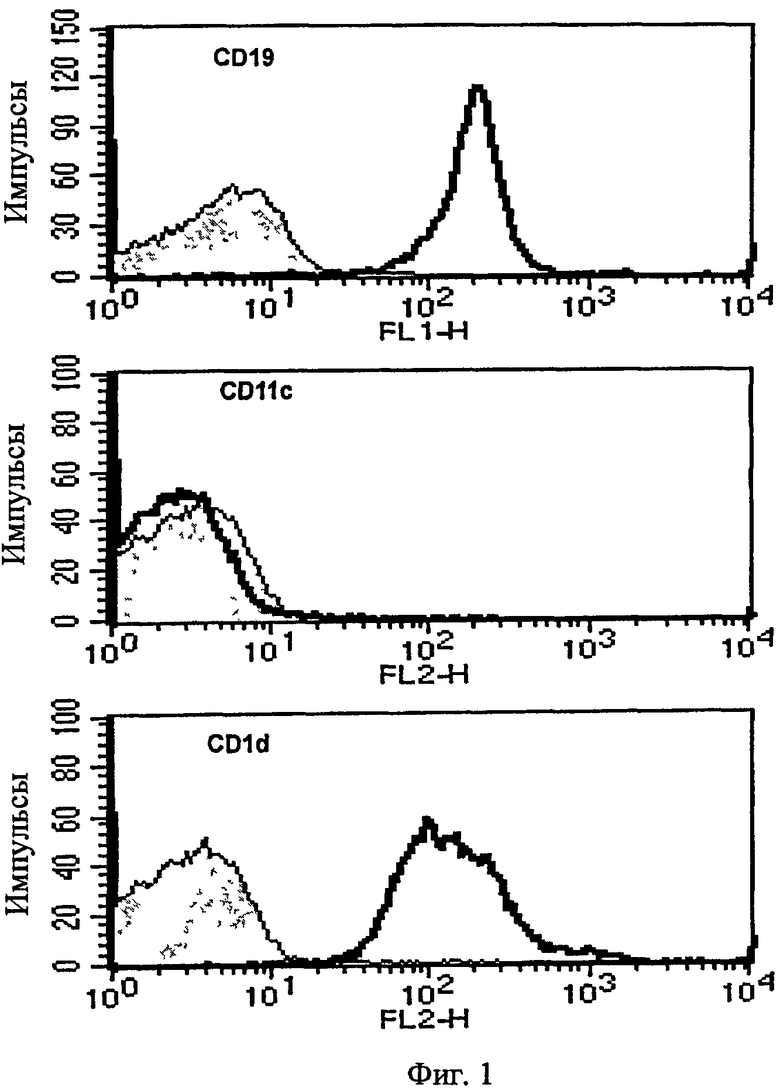
1. Иммунотерапевтическая или профилактическая вакцина, применяемая при/против вирусного инфекционного заболевания, бактериального инфекционного заболевания или рака, содержащая фармацевтически эффективное количество В-клеток, выбранных из группы, состоящей из
a) В-клеток, полученных путем нагрузки их лигандом Т-клеток-природных киллеров и сенсибилизирующим антигеном, и
b) В-клеток, полученных путем нагрузки их лигандом Т-клеток-природных киллеров и переноса гена, кодирующего пептидный антиген или белковый антиген, посредством которого В-клетки экспрессируют антиген, и фармацевтически приемлемый носитель.
2. Иммунотерапевтическая или профилактическая вакцина по п.1, в которой лиганд выбран из группы, состоящей из α-галактозилцерамида, α-галактуронозилцерамида, α-глюкуронозилцерамида, фосфатидилинозитолтетраманнозида, изоглоботригексозилцерамида, ганглиозида GD3, фосфатидилхолина, β-галактозилцерамида, липофосфогликана, гликоинозитолфосфолипида, производных aGalCer, р-аномерного галактозилцерамида и α-аномерного галактозилцерамида, бактериального липидного антигена и вариантов αGalCer.
3. Иммунотерапевтическая или профилактическая вакцина по п.1, в которой антиген происходит из любых патогенов, включая патогенные бактерии, вирусы и паразиты или опухолевый антиген.
4. Иммунотерапевтическая или профилактическая вакцина по п.3, в которой антиген, происходящий из патогенной бактерии, выбран из группы, состоящей из антигена Bordetella pertussis (коклюшевый токсин, нитчатый гемагглютинин и пертактин), столбнячного токсоида, дифтерийного антигена (дифтерийный токсоид), антигена Helicobacter pylori (капсульные полисахариды серогруппы А, В, С, Y и W-135), антигена пневмококка (капсульный полисахарид Streptococcus pneumoniae 3 типа), туберкулезного антигена, холерного антигена (В-субъединица холерного токсина), антигена стафилококка (стафилококковый энтеротоксин В), антигена шигеллы (полисахариды шигеллы), антигена Borrelia sp., антигена Candida albicans и антигена плазмодия.
5. Иммунотерапевтическая или профилактическая вакцина по п.3, в которой антиген вирусного происхождения выбран из группы, состоящей из антигена вируса гриппа (гемагглютинин и нейраминидазный антиген), антиген вируса папилломы человека (HPV) (гликопротеин), антиген вируса везикулярного стоматита (гликопротеин вируса везикулярного стоматита), антиген цитомегаловируса (CMV), антиген вируса гепатита (антиген гепатита A(HAV), B(HBV), C(HCV), D(HDV) и G(HGV)), (коровый антиген и поверхностный антиген), антиген респираторно-синцитиального вируса (RSV), антиген вируса простого герпеса (HSV), антиген вируса иммунодефицита человека (HIV) (GP-120, GP-160, p18, Tat, Gag, Pol и Env) и их сочетания.
6. Иммунотерапевтическая или профилактическая вакцина по п.3, в которой опухолевый антиген выбран из группы, состоящей из HPV E6/E7, gp100, тирозиназы, TRP-1 (родственного тирозиназе белка-1), TRP-2 (родственного тирозиназе белка-2), MAGE, MUC-1, СЕА (раково-эмбрионального антигена), р53, альфа-фетопротеина и Her-2/neu.
7. Иммунотерапевтическая или профилактическая вакцина по п.1, в которой антиген представляет собой пептид, липополисахарид, полисахарид, гликопротеин или полинуклеотид.
8. Иммунотерапевтическая или профилактическая вакцина по п,1, в которой пептидный или белковый антиген экспрессируется посредством трансдукции рекомбинантным вирусом.
9. Иммунотерапевтическая или профилактическая вакцина по п.8, в которой рекомбинантный вирус выбран из группы, состоящей из аденовируса, адено-ассоциированного вируса, ретровируса, вируса коровьей оспы, вируса оспы и вируса синдбис, имеющих ген, кодирующий антиген.
10. Иммунотерапевтическая или профилактическая вакцина по п.2, в которой В-клетки получены путем нагрузки их альфа-галактозилцерамидом и введения антигена, индуцирующая цитотоксический ответ.
11. Способ индукции цитотоксического ответа у нуждающегося в лечении пациента, предусматривающий введение этому пациенту вакцины по любому из пп.1-10.
| FAIS P | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J | |||
| Cancer | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| RU 94036709 A1, 27.08.1996. | |||

