[Область техники]
В рамках настоящего изобретения предложены композиция для индукции созревания дендритных клеток и способ созревания дендритных клеток. В частности, в рамках настоящего изобретения предложены композиция для индукции созревания дендритных клеток, содержащая гибридный белок ESAT-6/Rv2299c (рекомбинантный белок, состоящий из ESAT-6 и Rv2299c), полученный из М. tuberculosis, и способ дифференцировки незрелых дендритных клеток в зрелые дендритные клетки с применением указанной композиции.
[Уровень техники]
Mycobacterium tuberculosis (Mtb) является одним из наиболее широко распространенных патогенов человека, и треть населения мира инфицирована М. tuberculosis (Kaufmann, 2001). Поскольку коровья бацилла Кальмета-Герена (БЦЖ), которая является единственной вакциной, используемой в настоящее время, не обеспечивает значительный защитный эффект в отношении туберкулеза (ТВ) (Brandt et al, 2002), существует потребность в более эффективной вакцине, которая может заменить БЦЖ или усилить действие БЦЖ. Для разработки перспективных вакцин-кандидатов против туберкулеза (ТБ) крайне важно идентифицировать и охарактеризовать антигены микобактерий, ассоциированные с профилактической индукцией иммунного ответа. В то же время, многие антигены уже использованы для создания γпротивотуберкулезных вакцин, находящихся в настоящее время на различных этапах клинических испытаний (Ahsan, 2015).
Иммунный ответ с участием Тh1 необходим для контроля инфекции Mtb, о чем свидетельствует повышенная восприимчивость к микобактериальным инфекциям у мышей или людей с нарушением или мутацией генов, связанных с путями IFN-γ и IL-12 (van Crevel et al., 2002). Соответственно, многие исследования вакцин против туберкулеза ранее были сосредоточены на антигенах, которые являются сильными стимуляторами Т-клеток, таких как комплекс антигена 85 (Ag85) и ESAT-6 (Yuk and Jo, 2014). Поскольку ответы Т-клеток, необходимые для борьбы с инфекцией, редко приводят к элиминации Mtb из организма инфицированных людей или животных, (Feldman and Baggenstoss, 1938, Rhoades et al., 1997, Grace and Ernst, 2016), указанные антигены способны вызывать сильный профилактический иммунный ответ, но не приводят к индукции полноценного иммунного ответа, приводящего к элиминации патогена (Weinrich Olsen et al., 2001, Aagaard et al., 2011).
Наиболее эффективными антигенпрезентирующими клетками иммунной системы являются дендритные клетки (ДК), которые представляют собой критически важные клетки, которые представляют собой связующий элемент между врожденным иммунитетом и приобретенным иммунитетом.
Mtb способна модулировать функцию ДК, для того, чтобы задержать начало ответа с участием Т-клеток, что позволяет выиграть время для пролиферации данного микроорганизма (Gallegos et al., 2008, Hanekom et al., 2003, Wolf et al., 2007, Cooper, 2009). Это говорит о том, что указанные микроорганизмы могут выживать в неактивном состоянии. Соответственно, ранняя активация ДК и миграция ДК в лимфатические узлы, а также стимуляция Т-клеток являются ключевыми факторами для индукции эффективного предотвращения инфекции Mtb. Данные результаты позволяют предположить, что антигены микобактерий, которые индуцируют эффективную иммунную профилактику в организме хозяина посредством активации ДК, являются многообещающими мишенями для разработки противотуберкулезных вакцин. Действительно, ДК, инфицированные БЦЖ или подвергшиеся воздействию антигена Mtb, демонстрировали значительный профилактический эффект в отношении инфекции нормальными и большими количествами вирулентной Mtb в модели на мышах (Demangel et al., 1999, Rubakova et al., 2007). Тем не менее, хотя было выявлено несколько белков микобактерий, которые активируют ДК с последующей стимуляцией иммунного ответа Th1 (Byun et al., 2012b), о механизме их специфического противотуберкулезного иммунного ответа и защитном действии белка в качестве вакцины известно мало.
Если в качестве противотуберкулезной вакцины использовать гибридный белок, состоящий из активирующего ДК антигена и стимулирующего Т-клетки антигена, этот белок может обеспечивать два положительных ответа; индукция реакции поляризации Th1 самим ДК-активирующим белком и профилактическое усиление иммунитета, вызванное стимулирующим Т-клетки антигеном, посредством активации ДК. Поэтому предполагается, что профилактическое действие этого гибридного белка может быть более эффективным, чем эффект вакцины, включающей только Т-клеточный антиген. Сообщалось, что белок теплового шока микобактерий (HSP), такой как HSP65, вызывает сильный профилактический иммунитет против туберкулеза или эффективен в качестве адъюванта (Silva, 1999). В рамках настоящего изобретения выявлен белок Rv2299c, семейства HSP90, эффективно индуцировавший созревание ДК, и исследован механизм противотуберкулезного иммунного ответа посредством активации ДК, приводящей к устойчивому ответу Th1-типа. Затем было исследовано профилактическое действие на клинические изоляты Mtb HN878 белка Rv2299c или гибридного белка RV2299c/ESAT6 в качестве вакцины. Указанные результаты свидетельствуют о том, что ДК, созревшие в результате воздействия Rv2299c, индуцируют реакции Th1-клеток, направленные на обеспечение противотуберкулезной иммунной активности, и гибридный белок, включающий Rv2299c/ESAT-6, в качестве вакцины нового типа на основе белка, активирующего ДК, является многообещающей вакциной-кандидатом для усиления (бустинга) действия БЦЖ.
[Раскрытие сущности изобретения]
[Техническая задача]
Поскольку дендритные клетки играют крайне важную роль в усилении иммунной функции самого организма, как описано выше, разработка нетоксичного иммуномодулятора для индукции мощного иммунного ответа путем стимулирования дифференцировки дендритных клеток, чтобы вызвать их созревание, и четкое понимание их механизма действия является важным вопросом в области клеточной иммунотерапии с использованием дендритных клеток.
Соответственно, целью настоящего изобретения является обеспечение композиции, стимулирующей созревание дендритных клеток.
Другой целью настоящего изобретения является обеспечение способа индукции созревания дендритных клеток.
Еще одной целью настоящего изобретения является обеспечение терапевтического агента против Mycobacterium tuberculosis, содержащего дендритные клетки, созревание которых обеспечено способом, описанным выше.
[Техническое решение]
Для решения этих проблем в соответствии с настоящим изобретением предложена композиция для активации дендритных клеток, содержащая в качестве активного ингредиента гибридный белок Rv2299c, основанный на белке Rv2299c/ESAT-6.
Кроме того, в соответствии с настоящим изобретением предложена композиция для индукции созревания дендритных клеток, отличающаяся тем, что гибридный белок Rv2299c/ESAT-6 представляет собой белок, полученный из М. tuberculosis.
Гибридный белок Rv2299c/ESAT-6 предпочтительно содержится в композиции в количестве от 1 мкг/мл до 20 мкг/мл.
Кроме того, предложен способ индукции созревания незрелых дендритных клеток путем обработки незрелых дендритных клеток гибридным белком Rv2299c/ESAT-6.
Предпочтительным является способ, при котором после обработки незрелых дендритных клеток гибридным белком Rv2299c/ESAT-6 клетки культивируют в течение от 12 часов до 36 часов, и гибридный белок Rv2299c/ESAT-6 содержится в композиции в количестве от 1 мкг/мл до 20 мкг/мл.
Предложен способ индукции созревания незрелых дендритных клеток, при котором гибридный белок Rv2299c/ESAT-6 увеличивает выработку по меньшей мере одного из белков, выбранного из группы, состоящей TNF-α, IL-12р70, IL-6 и IL-1β во время созревании незрелых дендритных клеток.
Кроме того, в настоящем изобретении созревание незрелых дендритных клеток обусловлено стимуляцией TLR (toll-подобного рецептора)-4 с помощью Rv2299c или стимуляцией активируемой митогеном протеинкиназой (МАРK) и NF-κB.
Дендритные клетки, полученные способом, описанным выше, характеризуются положительными иммунологическими свойствами в отношении кластера дифференцировки (CD) 80, CD86, главного комплекса гистосовместимости (МНС) класса I и II.
Кроме того, в настоящем изобретении предложена композиция для усиления иммунитета, которая включает композицию для индукции созревания дендритных клеток.
[Преимущества настоящего изобретения]
Преимущество настоящего изобретения заключается в том, что клетки с помощью способа обеспечения созревания дендритных клеток согласно настоящему изобретению можно успешно дифференцировать незрелые клетки в зрелые дендритные клетки, с целью эффективной активации иммунного ответа организма.
[Описание чертежей]
Фиг. 1А представляет собой изображение ДНС-ПААГ и Вестерн-блот-анализа для оценки чистоты Rv2299c.
Фиг. 1В представляет собой изображение, иллюстрирующее снижение активности фагоцитоза и усиление функционального созревания за счет белков в ДК, обработанных Rv2299c или LPS.
Фиг. 1С представляет собой изображение, иллюстрирующее, что экспрессия молекул класса I и II и костимулирующих молекул, таких как CD80 и CD86 в ДК, обработанных Rv2299c в течение 24 часов, значимо увеличивается дозозависимым образом.
Фиг. 1D и 1Е представляет собой график и изображение, соответственно, иллюстрирующие, что доля IL-12р70-положительных клеток значимо увеличена в ДК, обработанных Rv2299c.
Фиг. 2 представляет собой график, иллюстрирующий, что активность созревания дендритных клеток, вызванного Rv2299c, может быть обусловлена загрязнением эндотоксином.
Фиг. 3А и 3В представляют собой графики, иллюстрирующие, что Rv2299c активирует ДК посредством пути TLR4, индуцируя экспрессию поверхностных молекул и провоспалительных цитокинов.
Фиг. 3С представляет собой график, иллюстрирующий, что выработка цитокинов (TNF-α, IL-6 и IL-1β) в дендритных клетках, обработанных Rv2299c, зависит от TRIF и MyD88.
Фиг. 3D, 3Е, и 3F представляют собой изображения, иллюстрирующие результаты конфокальной микроскопии (3D), график анализа FACS (3Е) и связывание с TLR4, в отличие от TLR2, показывающие взаимодействие дендритных клеток с Rv2299c.
Фиг. 4А представляет собой изображение, иллюстрирующее фосфорилирование Р38 и ERK 1/2 и фосфорилирование и деградацию IκВ-α в ДК, обусловленные Rv2299c.
Фиг. 4В представляет собой изображение, указывающее на индукцию важного изменения сайта р65 из цитоплазмы в ядро.
Фиг. 4С и 4D. представляют собой графики, иллюстрирующие, что индуцируемая Rv2299c экспрессия (4С) и выработка провоспалительных цитокинов в значительной степени устраняются костимулирующими молекулами на поверхности ДК всеми фармакологическими ингибиторами, за исключением ингибитора JNK.
Фиг. 5А и 5В представляют собой графики анализа MLR (реакция смешанных лимфоцитов), выполненные для точного описания свойств активности Rv2299c в отношении взаимодействия между ДК и Т-клеткой, где Фиг. 5А показывает, что ДК, обработанная Rv2299c, индуцирует пролиферацию Т-клеток, а Фиг. 5 В показывает, что пролиферация наивных Т-клеток индуцируется в фенотипе Th1.
Фиг. 6А и 6В представляют собой графики, полученные с помощью проточной цитометрии, выполненной с целью определить, может ли стимулированная Rv2299c ДК стимулировать CD4+.
Фиг. 7. представляет собой график, иллюстрирующий, что Rv2299c ингибирует рост Mtb и индуцирует активность Т-клеток, обладающих бактерицидной активностью, посредством стимуляции созревания ДК.
Фиг. 8А представляет собой изображение, иллюстрирующее цитотоксичность рекомбинантного гибридного белка Rv2299c/ESAT-6.
Фиг. 8В и 8С представляют собой графики, иллюстрирующие экспрессию молекул МНС и экспрессию провоспалительных цитокинов, таких как TNF-α, IL-12 и IL-1β, в рекомбинантном гибридном белке Rv2299c/ESAT-6.
Фиг. 9А представляет собой график, иллюстрирующий, что рост Mtb был в значительной степени ингибирован добавлением Т-клеток, активированных ESAT-6-стимулированными ДК.
Фиг. 9В представляет собой график, иллюстрирующий, что высокая выработка провоспалительных цитокинов, таких как IFN-γ и TNF-α, достигается, когда Mtb-инфицированные МКМ культивируют совместно с Т-клетками, активированными ДК, стимулированными гибридным белком.
Фиг. 10А представляет собой схему, иллюстрирующую эффект прайм-бустинга БЦЖ на высоковирулентном штамме HN878 под действием гибридного белка Rv2299c/ESAT-6.
Фиг. 10В представляет собой график, иллюстрирующий, что БЦЖ отдельно, гибридный белок ESAT-6 и гибридный белок Rv2299c/ESAT - все снизили бактериальную нагрузку в легких.
Фиг. 10С и 10D представляют собой графики, иллюстрирующие профилактический эффект гибридного белка Rv2299c/ESAT-6 против заражения штаммом HN878 с точки зрения долгосрочного сокращения числа бактерий.
Фиг. 11А, 11В, 12А и 12В представляют собой графики, иллюстрирующие, что изменения в фенотипе Т-клеток, вызванные профилактической инокуляцией гибридным белком Rv2299c/ESAT-6, были оценены по окрашиванию цитокинов, и графики, иллюстрирующие ко-экспрессию IFN-γ, TNF-α и IL-2 и т.п.
[Принцип изобретения]
До настоящего времени был выполнен ряд исследований белков, полученных из М. tuberculosis, при иммунном ответе иммуноцитов, но исследования Rv2299c не известны.
Незрелые дендритные клетки созревают, образуя зрелые дендритные клетки. Зрелые дендритные клетки теряют способность усиливать экспрессию молекул клеточной поверхности и различных цитокинов, которые захватывают и одновременно стимулируют антиген. В частности, зрелые дендритные клетки экспрессируют антигены МНС типа I и типа II в более высоких количествах по сравнению с незрелыми дендритными клетками и регулируют кластер дифференцировки (CD) 80+, CD83+, CD86+ и CD14-. Повышенная экспрессия МНС вызывает увеличение антигенной плотности дендритных клеток, тогда как активация костимулирующих молекул CD80 и CD86 приводит к улучшению сигнала активации Т-клеток через костимулирующую молекулу-партнера, например, CD28, на Т-клетках.
Зрелые дендритные клетки по настоящему изобретению могут быть получены приведением в контакт с Rv2299c или гибридным белком Rv2299c/ESAT-6, полученным рекомбинантным способом. Настоящее изобретение может индуцировать созревание дендритных клеток посредством композиции, содержащей Rv2299c или гибридный белок Rv2299c/ESAT-6. Rv2299c или гибридный белок Rv2299c/ESAT-6 эффективно получают, например, но не ограничиваясь указанным, рекомбинантным способом, причем Rv2299c может содержаться в композиции в концентрации от 1 мкг/мл до 20 мкг/мл, предпочтительно от 5 мкг/мл до 10 мкг/мл.
Созревание дендритных клеток можно отслеживать способами, известными в данной области техники. Маркеры клеточной поверхности могут быть обнаружены с помощью анализов, известных в данной области, таких как проточная цитометрия и иммуногистохимия. Клетки также можно отслеживать по выработке цитокинов (например, с помощью ИФА, FACS и других иммуноанализов).
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Следующие примеры представлены для иллюстрации настоящего изобретения в более понятной форме, и настоящее изобретение не ограничено данными примерами.
<Пример 1>
Материалы и методы
1.1 Получение бактериального штамма и рода Mycobacterium
Mtb H37Rv (АТСС 27294) и H37Ra (АТСС 25177) были приобретены в Американском банке штаммов (Американская коллекция типовых культур, АТСС, Манассас, Вирджиния), a Mtb НМ878 был получен из Международного научно-исследовательского института туберкулеза (ITRC, Чангвон, Кёнсан-Намдо, Южная Корея). Микобактерия коровьей БЦЖ (штамм Пастера 1173Р2) была предоставлен доктором Брошем из Института Пастера (Париж, Франция). Все микобактерии, использованные в этих исследованиях, были приготовлены, как описано выше (Cha et al., 2015b).
1.2 Животные, вакцинация и инфицирование аэрозольным путем
Не содержащие специфических патогенов самки мышей C57BL/6 возрастом 5-6 недель, а также трансформантные мыши с Т-клеточным рецептором OT-I и ОТ-II (TCR) (фон C57BL/6), C57BL/6 (Н-2Kb и I-Ab), Мыши, нокаутированные по C57BL/6J TLR2 (TLR2-/-; B6.129-Tlr2tm1Kir/J) и мыши, нокаутированные по CLBL/10 TLR4 (TLR4-/-; C57BL/10ScNJ) были приобретены у The Jackson Laboratory (Бар-Харбор, штат Мэн), США). Мышей содержали при постоянной температуре (24±1°С) и влажности (50±5%) в изолированных условиях в помещениях для биологически опасных животных BL-3 Медицинского исследовательского центра Университета Йонсей. Животным обеспечивали стерильное коммерческое питание для мышей, а также произвольный доступ к воде при стандартизированных условиях контроля освещенности (цикл с 12-часовым освещением и 12-часовой темнотой). Мышей контролировали ежедневно, и никаких клинических признаков или заболеваний во время этих экспериментов ни одна из мышей не показала.
Для вакцинации мышей сначала вакцинировали БЦЖ Пастера 1173Р2 путем подкожной инъекции (2,0×105 КОЕ/мышь). Через шесть недель после иммунизации БЦЖ вводили субъединичные вакцины три раза каждые три недели, а также DDA-липосому (DDA - диметилдиоктадециламмоний) (5 мкг/инъекцию), включающую монофосфориллипид-А (MPL, 5 мкг/инъекцию). Через четыре недели после последней иммунной вакцинации собирали клетки селезенки и легких и использовали для исследования иммуногенности (уровней секреции IFN-γ и циклов Т-клеток, вырабатывающих JEN-γ, и Ag-специфических Т-клеток). Чтобы изучить профилактический эффект при введении только субъединичной вакцины, иммунизированную только БЦЖ группу и группу, в которой осуществляли прайминг БЦЖ и бустинг субъединичной вакциной, инфицировали аэрозолями со штаммами Mtb H37Rv (АТСС 27294) или Mtb HN878, как описано выше (Cha et al., 2015b, Lee et al., 2009). В кратком изложении, мышей подвергали воздействию инфицированного воздуха в ингаляционной камере (Glas-Col, Терре-Хот, Индиана, США), в течение 60 минут с заданной дозой H37Rv или HN878 для воздействия Mtb с количеством жизнеспособных частиц около 200 КОЕ. Через 8 или 16 недель после заражения в соответствующих группах собирали клетки селезенки и легких, и использовали проточную цитометрию для оценки цикла многофункциональных Т-клеток и подтипов Т-клеток.
1.3 Антитело и реагент
Рекомбинантный колониестимулирующий фактор макрофагов мыши (M-CSF), гранулоцитарно-колониальный фактор макрофагов мыши (GM-CSF) и интерлейкин-4 (IL-4) были приобретены у Creagene (Gyeonggi, Южная Корея). Набор флуоресцеин изотиоцианат (FITС)-аннексин V/пропидий йодид был приобретен у Research & Development System (Миннеаполис, Миннесота, США). Декстран-FITC (молекулярный вес 40000 Да) был получен от Sigma (Сент-Луис, Миссури, США). Липополисахарид из E.coli O111:В4 был приобретен у InvivoGen (Сан-Диего, Калифорния, США). Эндотоксиновый фильтр (END-X) и смола для удаления эндотоксина (END-X В15) были получены от АСС (Ист Фолмут, Массачусетс, США). OT-I пептид (OVA257-264) и ОТ-II пептид (OVA323-339) были синтезированы производителем Peptron (Тэджон, Южная Корея). Моноклональное антитело к фосфорилированному ERK1/2, моноклональное aнти-ERK1/2 антитело, моноклональное антитело к фосфорилированному р38, моноклональное антитело к р38, поликлональное анти-NF-κB антитело (р65), моноклональное поликлональное антитело к фосфорилированному IκВ-α, моноклональное анти-IκВ-α антитело, поликлональное антитело против ламина В и поликлональное антитело против β-актина были получены от CST (Данверс, Массачусетс, США). HRP-конъюгированное антитело против IgG мыши и HRP-rhddor антитело против антител кролика были получены от Calbiochem (Сан-Диего, Калифорния, США), a mAb против β-актина (АС-15) было приобретено у Sigma. CD11c, р65, IFN-g и СD62L-нацеленные FITC-конъюгированные mAb, IL-12p70, IL-10 и CD3-нацеленные АРС-конъюгированные mAb, CD4 и СD8-нацеленные РеrСР-Су5.5-конъюгированные mAb, CD8+- нацеленные АРС-Су7-конъюгированные mAb, CD80, CD86, МНС класса I, МНС класса II, IFN-γ и СD44-нацеленные фикоэритрин (РЕ)-конъюгированные mAb, CD11c и IL-2-нацеленные РЕ-Су7-конъюгированные mAb, и СD3е-нацеленные на eFluor 450-конъюгированные mAb были приобретены у eBioscience (Сан-Диего, Калифорния, США). Фикоэритрин (РЕ)-конъюгированные анти-IgG1 мыши, aнти-IgG2a мыши и aнти-IgG2b мыши, АРС-конъюгированные aнти-IgG2a мыши и анти-IgG1 мыши, FITC-конъюгированные aнти-IgG2b мыши и РЕ-Су7-конъюгированные анти-IgG1 мыши и aнти-IgG2b мыши были получены от eBioscience (Сан-Диего, Калифорния, США. Наборы ИФА для определения TNF-α, IL-1β, IFN-γ, IL-2, IL-4, IL-6, IL-10 и IL-12p70 были получены от eBioscience.
1.4 Экспрессия и очистка рекомбинантного белка
Для получения рекомбинантного белка Rv2299c соответствующие гены амплифицировали полимеразной цепной реакцией (ПЦР) с использованием геномной ДНК М. tuberculosis H37Rv АТСС27294 в качестве матрицы и следующих праймеров: Rv2299c прямой праймер, 5'-CATATGAACGCCCATGTCGAGCAGTTG-3', и обратный, 5'-
GAATTCGGCAAGGTACGCGCGAGACGTTC-3' ESAT-6 прямой, 5'-AAGCTTATGACAGAGCAGCAGTGGAAT-3', и обратный, 5'- CTCGAGTGCGAACATCCCAGTGACGTT-3'. ПЦР-продукт Rv2299c расщепляли с помощью NdeI и EcoRI, a ESAT-6 расщепляли с помощью HindIII и XhoI. Эти продукты были встроены в вектор pET22b(+) (Novagen, Мэдисон, Висконсин, США), и результат был секвенирован. Рекомбинантную плазмиду трансформировали в клетки Е. coli BL21 тепловым шоком в течение 1 минуты при 42°С. Сверхэкспрессированные белки были получены с небольшими модификациями способа, как описано выше (Cho et al., 2006). В кратком изложении, рекомбинантные плазмиды, содержащие E.coli, инкубировали при 37°С до достижения ими оптической плотности (OD) 0,4-0,5 на 600 нм, а затем их индуцировали с помощью 1 мМ изопропил-D-тиогалактопиранозида (IPTG; ELPIS-Biotech), Тэджон, Южная Корея). Затем бактериальные клетки собирали центрифугированием и суспендировали в смеси 20 мМ Трис-HCl (рН 8,0), 0,5 М хлорида натрия, 5 мМ имидазола и 1 мМ фенилметилсульфонилфторида (Sigma); и разрушали ультразвуком. Рекомбинантные белки очищали с помощью хроматографического анализа на никель-нитрилоацетатной агарозе (Ni-NTA) в соответствии с инструкциями производителя (Qiagen, Чатсворт, Калифорния, США). Соответствующие результаты стадий очистки анализировали с помощью 13,5% полиакриламидного геля в присутствии додецилсульфата натрия (ДНС-ПААГ) с окрашиванием красителем кумасси бриллиантовым синим и иммуноблотом с использованием антитела против His (Santa Cruz). Очищенные белки собирали, концентрировали и диализовали с помощью забуференного фосфатом солевого раствора (PBS, рН 7,4). Чтобы устранить загрязнение эндотоксином, диализированные рекомбинантные белки инкубировали с полимиксин-В-агарозой (PmB, Sigma) при 4°С в течение 6 часов. Наконец, рекомбинантные белки, очищенные от эндотоксина, фильтровали и стерилизовали и замораживали при -70°С. Концентрации белка измеряли с помощью набора для анализа белка ВСА (Pierce, Рокфорд, Иллинойс), используя бычий сывороточный альбумин в качестве стандарта. Остаточный LPS (липополисахарид) рекомбинантных белков определяли с использованием биологического теста с использованием эндотоксина (LAL) (Lonza, Базель, Швейцария) в соответствии с инструкциями производителя. Чистоту рекомбинантных белков оценивали окрашиванием кумасси бриллиантовым синим (СВ) и вестерн-блоттингом с использованием антигистидиновых антител.
1.5 Культура клеток
Как недавно было описано, из костного мозга мышей были получены культивированные и очищенные ДК (Byun et al., 2012b). Макрофаги, происходящие из костного мозга (МКМ), получали с использованием рекомбинантного M-CSF, как описано выше. В кратком изложении, клетки костного мозга, выделенные из мышей C57BL/6, отделяли от эритроцитов (RBC) с помощью растворяющего буфера (4,15 г/500 мл хлорида аммония и буфер 0,01 М трис-HCl (рН 7,5±2)) и промывали средой RPMI 1640. Затем их высевали на 6-луночный культуральный планшет (106 клеток/мл, 3 мл/лунку) и культивировали с использованием среды RPMI 1640 с добавкой 100 Ед./мл пенициллина/стрептомицина (Lonza), 10% фетальной бычьей сыворотки (Lonza), 50 мМ меркаптоэтанола (Lonza), 0,1 мМ заменимой аминокислоты (Lonza), 1 мМ пирувата натрия (Sigma), 20 нг/мл GM-CSF и 10 нг/мл IL-4 (ДККМ) или 20 нг/мл M-CSF (МКМ) при 37°С в присутствии 5% углекислого газа.
1.6 Анализ цитотоксичности
Анализы цитотоксичности выполняли с использованием набора для окрашивания аннексии V/пропидий йодид (PI) в соответствии с инструкциями производителя (BD Biosciences). Клетки окрашивали FITC-конъюгированным аннексином V и PI. Анализ окрашенных клеток проводили с использованием цитометра FACSCanto II с FACSDiva, а результаты анализировали с использованием программного обеспечения FlowJo (Tree Star, Ашленд, Орегон, США).
1.7 Анализ экспрессии поверхностной молекулы с помощью проточного клеточного анализа
На 6-й день ДККМ собирали, промывали PBS и ресуспендировали в промывочном буфере для сортировки флуоресцентно-активированных клеток (2% FBS и 0,1% азида натрия в PBS). Клетки предварительно культивировали с 0,5% BSA в PBS в течение 30 минут и промывали PBS. Клетки окрашивали FITC-конъюгированным анти-CD11c с РЕ-конъюгированным анти-Н-2Kb (МНС класса I), анти-I-антителом (МНС класса II), анти-СD80 и анти-СD86 при 4°С в течение 45 минут. Клетки трижды промывали PBS и ресуспендировали с 500 мкл PBS. Флуоресценцию измеряли с помощью проточной цитометрии, а данные анализировали с использованием программного обеспечения для анализа данных CellQuest.
1.8 Способность ДККМ поглощать антитела по данным измерения Rv2299c
ДККМ (2×105 клеток) поддерживали в равновесии при 37°С или 4°С в течение 45 минут и сенсибилизировали в концентрации 1 мг/мл флуоресцеин-конъюгированным декстраном. Для остановки реакции добавляли холодный буфер для окрашивания. Клетки трижды промывали, окрашивали РЕ-конъюгированным анти-CD11c-антителом и затем анализировали с помощью цитометра FACSCanto. Неспецифическое связывание декстрана с ДК определяли путем культивирования ДК с FITC-конъюгированным декстраном при 4°С, и полученные фоновые значения вычитали из значений специфического связывания.
1.9 Установление удаления LPS-загрязнения для Rv2299c
Чтобы определить, что созревание ДК, индуцированных Rv2299c, происходило не из-за загрязнения эндотоксином или LPS в белковых препаратах, проводили предварительную обработку полимиксином В (РmВ) (Sigma), термическую денатурацию и деградацию протеиназой K (Sigma) и выполняли анализ. ДК предварительно инкубировали с 50 мкг/мл РmВ в течение 1 часа при комнатной температуре перед обработкой 100 нг/мл LPS и 10 мкг/мл Rv2299c. Для термической денатурации LPS или Rv2299c инкубировали при 100°С в течение 1 часа. Для деградации протеиназы K, LPS или Rv2299c расщепляли растворимой протеазой K в концентрации 10 мкг/мл при 37°С в течение 1 часа. Для дезактивации ферментов, их нагревали при 100°С в течение 15 минут и затем добавляли в культуральный раствор ДККМ. Через 24 часа уровни TNF-α и IL-6 в супернатантах ДККМ анализировали с помощью ИФА.
1.10 Конфокальный сканирующий лазерный микроскоп
ДК наносили на ночь на покровное стекло, покрытое поли-L-лизином. После обработки Rv2299c клетки фиксировали 4% параформальдегидом, пермеабилизировали 0,1% раствором тритона Х-100. Перед инкубацией клеток с 2% BSA в антителе против Rv2299c, содержащим PBS/T при комнатной температуре в течение 2 часов, клетки блокировали 2% бычьим сывороточным альбумином (BSA) в PBS, содержащим 0,1% Tween-20 (PBS/T), в течение 2 часов. После промывания PBS/T клетки повторно культивировали с Су-3-конъюгированным вторичным антителом в темной комнате в течение 1 часа и затем окрашивали раствором 1 мкг/мл DAPI при комнатной температуре в течение 10 минут. Форму клеток и чувствительность к флуоресценции наблюдали с использованием конфокального лазерного микроскопа (Zeiss LSM 510 Meta; Carl Zeiss Ltd, Велвин Гарден Сити, Великобритания). Их изображения получали с использованием программного обеспечения LSM510 Meta и обрабатывали с помощью измерителя изображений LSM.
1.11 Иммунопреципитация
ДК (1×107) культивировали с 10 мг/мл Rv2299c в течение 6 часов, и клеточный осадок растворяли в лизирующем буфере (10 мМ Трис-HCl (рН 7,4), 1% NP-40, 0,25% дезоксихолата натрия, 150 мМ НСl, 1 мМ ЭДТА, 1 мМ PMSF, 1 мкг/мл апротинина, 1 мкг/мл лейпептина, 1 мкг/мл пепстатина, 1 мМ Na3VO4 и 1 мМ NaF). Чтобы предотвратить неспецифическое связывание, к 1 мг клеточного лизата добавляли 50 мкл нормальной сыворотки (Санта-Крус) и 100 мкл 50% суспензии гранул сорбента белок А или G-сефарозы (Invitrogen, Карлсбад, Калифорния) добавляли к 1 мг клеточного лизата, чтобы заранее определить стабильность клеточного лизата. После инкубации при 4°С в течение 2 часов смесь гранул и клеточного лизата центрифугировали при 10000 g при 4°С в течение 5 минут и собирали супернатант для дальнейшего эксперимента. Rv2299c (His), TLR2 и ТLR4-родственные белки инкубировали с IgG против антител мыши в качестве контрольных антител против анти-ТLR2 и TLR4 при 4°С в течение 1 часа и с IgG против антител мыши в качестве контрольного антитела против анти-Rv2299 с (His), а затем иммунопреципитировали путем инкубации с сорбентом белок А или G-сефарозой при 4°С в течение 24 часов. Гранулы собирали, промывали и кипятили в 5Х буфере для образцов в течение 5 минут. Белки разделяли на 10% ДНС-ПААГ и переносили на мембрану из дифторида винилиденхлорида (Millipore). Затем мембраны исследовали с помощью антител против TLR2, TLR4 и His, как описано.
1.12 Анализ методом иммуноблоттинга
После стимуляции воздействием 10 мкг/мл Rv2299c, ДК суспендировали в 100 мкл лизирующего буфера, содержащего 50 мМ Трис-НСl (рН 7,5), 150 мМ НСl, 1% Тритон-Х100, 1 мМ ЭДТА, 50 мМ NaF, 30 мМ Na4PO7, 1 мМ фенилметансульфонилфторида, 1 мкг/мл апротинина и 1 мМ перванадата. Образцы цельноклеточного лизата разделяли в SDS-полиакриламидных гелях и переносили на нитроцеллюлозные мембраны. Мембраны блокировали 5% обезжиренным молоком, инкубировали с антителом в течение 2 часов, а затем инкубировали с вторичными антителами, конъюгированными с HRP, в течение 1 часа при комнатной температуре. Эпитопы на белках-мишени, включая МАРK и NF-κВ, которые специфически распознаются антителами, визуализировали с использованием набора для ECL-определения (GE Healthcare, Литтл Чалфонт, Великобритания).
1.13 Получение экстракта ядер
Экстракты ядер, полученные из клеток, готовили следующим образом. ДК обрабатывали 100 мл буфера для лизиса (10 мМ HEPES (рН 7,9), 10 мМ КСl, 0,1 мМ ЭДТА, 0,5% нонидет Р-40, 1 мМ дитиотреитола (DTT) и 0,5 мМ ПМСФ) на льду в течение 10 минут. После центрифугирования при 4000 об/мин в течение 5 минут осадок ресуспендировали в 100 мкл экстракционного буфера (20 мМ HEPES (рН 7,9), 40 мМ хлорида натрия, 1 мМ ЭДТА, 1 мМ DTT и 1 мМ PMSF), клетки культивировали на льду в течение 30 минут. После центрифугирования при 12000 об/мин в течение 10 минут супернатант, содержащий экстракты ядер, собирали и хранили при -80°С до тех пор, когда он потребуется.
1.14 Обработка ДК фармакологическим ингибитором для анализа сигнального пути
Все фармакологические ингибиторы были приобретены у Calbiochem. В качестве контроля растворителя в культуру добавляли диметилсульфоксид (Sigma) при 0,1% (об./об.). ДК промывали PBS и предварительно обрабатывали ингибиторами в среде RPMI 1640, содержащей глутамин, в течение 1 часа перед обработкой Rv2299c в течение 24 часов. Ингибиторы использовали в следующих концентрациях: U0126 (10 мкМ), SB203580 (20 мкМ), SP600125 (10 мкМ) и Bay11-7082 (20 мкМ). Во всех экспериментах, в которых использовались ингибиторы, тестовые концентрации определяли после тщательных экспериментов по титрованию для оценки жизнеспособности ДК с использованием анализов МТТ.
1.15 Анализ пролиферации Т-клеток in vitro
Реактивные Т-клетки, участвующие в ответе наивных Т-клеток, выделяли из общей массы мононуклеарных клеток, полученных от мышей BALB/c, с помощью колонки MACS (Miltenyi Biotec). Как OVA-специфичные CD8+, так и CD4+реактивные Т-клетки, соответственно, были получены из спленоцитов мышей ОТ-1 и ОТ-2. Эти Т-клетки окрашивали с помощью 1 мкМ CFSE (Invitrogen), как описано выше (Jeong et al., 2009). ДК (2×105 клеток на лунку), обработанные пептидом OVA в течение 24 часов в присутствии 10 мкг/мл Rv2299c, совместно культивировали с окрашенными CFSE CD8+и CD4+Т-клетками (2×106) при соотношении ДК к Т-клеткам 1:10. На 3-й или 4-й день после совместного культивирования соответствующие Т-клетки окрашивали РеrСР-су5.5-конъюгированным CD4+mAb, РЕ-су5-конъюгированным анти-CD4+mAb, РЕ-су5-конъюгированным анти-СD8+mAb, Аlеха647- конъюгированным анти-ССR3 mAb или РЕ-конъюгированным анти-CXCR3 mAb и анализировали с помощью проточного цитометра. Супернатанты собирали, и измеряли выработку IFN-γ, IL-2 и IL-4 с помощью ИФА.
1.16 Анализ активации эффекторов/Т-клеток памяти
Как описано выше, реактивные Т-клетки, участвующие в ответе аллогенных Т-клеток, выделяли из общей массы мононуклеарных клеток, полученных от мышей BALB/c, зараженных микобактериями, с помощью колонки MACS (Miltenyi Biotec). Окрашивание АРС-конъюгированным mAb против CD3 (BD Biosciences) показало, что препарат состоял в основном из CD3+клеток (>95%). Произведенные в крупном масштабе ДК (2×105 клеток на лунку), полученные от мышей дикого типа (WT), TLR2-/- и TLR4-/- C57BL/6 после промывки обрабатывали Rv2299c в течение 24 часов. Клетки совместно культивировали с 2×106 реактивными аллогенными Т-клетками при соотношении Т-клеток к ДК 1:10. На 4-й день совместного культивирования клетки окрашивали РеrСР-су5.5-конъюгированным анти-СD4+ mAb, PerCP-су5.5-конъюгированным анти-СD8+ mAb, FITC-конъюгированным aнти-CD62L mAb и РЕ-конъюгированным анти-СD44 mAb и анализировали проточным цитометром.
1.17 Измерение цитокинов
Для обнаружения IL-6, IL-1β, TNF-α, IFN-γ, IL-4, IL-2, IL-12p70 и IL-10 в супернатанте культуры использовали сэндвич-иммуноферментный анализ (ИФА) как описано выше (Byun et al., 2012b). Отдельные клетки, полученные из легких вакцинированных или инфицированных мышей, стимулировали PPD (2 мкг/мл) или антиген-специфическими пептидами CD4 или CD8 Т-клеток (2 мкг/мл) при 37°С в течение 24 часов. Уровни IFN-γ-цитокинов в культуральных супернатантах измеряли с использованием коммерческого набора ИФА (eBioscience) в соответствии с протоколом производителя.
1.18 Анализ внутриклеточного цитокина
Клетки сначала блокировали 10% (об./об.) нормальной козьей сывороткой при 4°С в течение 15 минут и окрашивали FITC-конъюгированными антителами к CD11c+ при 4°С в течение 30 минут. В качестве отрицательного контроля использовали клетки, окрашенные подходящим изотипически сходным иммуноглобулином (Ig). Клетки фиксировали и пермеабилизировали с помощью набора Cytofix/Cytoperm (BD Biosciences), в соответствии с инструкциями производителя. IL-12p70, IL-10, IL-2, IL-4 и IFN-γ в клетках определяли в пермеабилизирующем буфере с помощью антител, конъюгированных с флуоресцеином (BD Biosciences). Чистые белковые производные (PPD) были предоставлены доктором Бреннаном из Aeras (Роквилл, Мэриленд, США). Для внутриклеточного окрашивания цитокинов одиночно-клеточные суспензии вакцинированных животных (2×106 клеток) стимулировали PPD (2 мкг/мл) и антигенспецифическими пептидами CD4 или CD8 Т-клеток (2 мкг/мл) в присутствии GolgiStop при 37***°С в течение 12 часов (BD Biosciences). После заражения HN878, PPD использовали отдельно в качестве стимулятора для внутриклеточного окрашивания цитокинов. Клетки сначала блокировали блоком Fc (анти-CD16/32) при 4°С в течение 15 минут и окрашивали BV421-конъюгированными анти-СD3, РеrСр-Су5,5-конъюгированными анти-СD4, АРС-Су7-конъюгированными анти-CD8 и FITC-конъюгированными анти-CD62L антителами. Клетки фиксировали и пермеабилизировали с помощью набора Cytofix/Cytoperm (BD Biosciences), в соответствии с инструкциями производителя. TNF-α, IL-2 и IFN-γ в клетках детектировали в буфере для пермеабилизации с использованием АРС-конъюгированных анти-TNF-α, РЕ-Су7- конъюгированных анти-IL-2 и РЕ-конъюгированных анти-IFN-α антител. Все антитела были приобретены у eBioscience (Сан-Диего, Калифорния), если не указано иное. Клетки анализировали с помощью проточного цитометра FACSverse с использованием коммерчески доступной программы FlowJo (Treestar, Inc., Сан-Карлос, Калифорния, США).
1.19 Подсчет количества бактерий и гистопатологический анализ После последней вакцинации, а затем через 16 недель заражения HN878 от 6 до 7 мышей в каждой группе подвергали эвтаназии с помощью диоксида углерода, и их легкие и селезенку гомогенизировали. Количество жизнеспособных микроорганизмов определяли путем посева на агар Middlehrook 7H11 (Difco Laboratories, Детройт, Мичиган, США) путем серийного разбавления гомогената органа (половина левого легкого и половина селезенки) и добавляли 10% OADC (Difco Laboratories), амфотерицин В (Sigma-Aldrich, Сент-Луис, Миссури, США) и 2 мкг/мл гидразида 2-тиофенкарбоновой кислоты (Sigma-Aldrich). После инкубации при 37°С в течение 4 недель подсчитывали число колоний. Для гистопатологического анализа верхнюю долю правого легкого окрашивали гематоксилином и эозином и оценивали на предмет тяжести воспаления. Как описано выше (Cha et al., 2015а), уровни воспаления в легких оценивали с использованием программы ImageJ (National Institutes of Health, Бетесда, Мэриленд). Кроме того, воспалительный ответ оценивали по размеру поражения и составу иммунных клеток. Данные по КОЕ и легочному воспалению были представлены в виде медианы log10 КОЕ ± межквартильный размах (IQR).
1.20 Статистический анализ
Все эксперименты были повторены не менее трех раз, и были получены согласованные результаты. Уровень значимости для сравнения между образцами определяли по распределению множественных сравнительных тестов Тьюки с использованием статистического программного обеспечения (GraphPad Prism Software, версия 4.03; GraphPad Software, Сан-Диего, Калифорния). Данные на графике выражены в виде среднего значения ±SEM. Статистически значимым считается каждое значение *р<0,05, **р<0,01 или ***р<0,001.
<Пример 2>
Рекомбинантный белок Rv2299c индуцирует созревание и активацию дендритных клеток (ДК).
2.1 Экспериментальный метод
(A) (a) Rv2299c был успешно индуцирован в E.coli BL21 при добавлении 1 мМ изопропил-β-D-тиогалактозида (IPTG) в течение 6 часов. Белки были в основном локализованы в тельцах включения. Молекулярная масса N-концевой His-метки Rv2299c была близка к 72 кДа по данным ДНС-ПААГ (М: маркер молекулярной массы; дорожка 1: отсутствие индукции, дорожка 2: индукция). Рекомбинантный Rv2299c был получен в клетках BL21 и очищен смолой NTA. Очищенные белки подвергали (b) ДНС-ПААГ и (с) Вестерн-блот-анализу с использованием антител мыши против His 1:1000. (В) ДККМ обрабатывали Rv2299c в концентрации 1, 5 или 10 мкг/мл в течение 30 минут, а затем культивировали в отсутствие или в присутствии LPS в течение 24 часов. Клетки культивировали с декстраном (FITC) при 37°С в течение 30 минут и затем при 4°С в течение 30 минут и оценивали с помощью FAC-анализа поглощения декстрана (FITC). Показана доля на наличие декстран (FIТС)-положительных CD11c+ (РЕ)-положительных клеток. Результаты представляют четыре экспериментальных результата. Гистограмма показывает среднее значение ± SEM для доли декстран-FIТС-позитивных CD11c+ клеток для трех независимых экспериментов, а также показана статистическая значимость (***р<0,001) для обработанных клеток по сравнению с контролем. (С) ДККМ культивировали в присутствии 1, 5 или 10 мкг/мл Rv2299c в течение 24 часов и анализировали с помощью двухцветной проточной цитометрии. Клетки были отсортированы по сигналу для исключения клеток CD11c. Среда: необработанный контроль; LPS: положительный контроль (100 мкг/мл LPS). ДК окрашивали анти-СD80, анти-СD86, анти-МНС класса I или анти-МНС класса II. Результаты представляют три экспериментальных результата.
Среднюю интенсивность флуоресценции рассчитывали из приведенного выше графика. Каждый столбец представляет средний уровень флуоресценции образца. Данные представлены в виде среднего значения ± SEM (n=5). *р<0,05, **р<0,01 и ***р<0,001 по сравнению с необработанной контрольной группой. (D) ДККМ стимулировали воздействием 1, 5 или 10 мкг/мл Rv2299c и 100 нг/мл LPS в течение 24 часов. Анализ количества выработанных TNF-α, IL-6, IL-1β, IL-12p70 и IL-10 выполняли с помощью ИФА. Результаты получены из репрезентативного эксперимента в одном из трех проведенных экспериментов. Данные представлены в виде среднего значения ± SEM (n=5). Значимость *р<0,05, **р<0,01 и ***р<0,001 указана по сравнению с необработанной контрольной группой. (Е) Анализ экспрессии IL-12p70 и IL-10 в CD11с+ДК путем внутриклеточного окрашивания цитокинов.
2.2 Анализ экспериментальных результатов
Имеется мало информации об иммунологической роли белка Rv2299c в семействе микобактериальных HSP. Рекомбинантные белки Rv2299c в E.coli BL21 очищали, и исследовали их иммунный ответ. Чистоту Rv2299c оценивали с помощью ДНС-ПААГ и Вестерн-блоттинга (Фиг. 1А). Содержание эндотоксина в полученном Rv2299c составляло менее 15 пг/мл (<0,1 ед./мл) с помощью LAL-анализа. ДК и макрофаги играют важную роль в инициации и активации профилактических иммунных реакций против микобактерий. Таким образом, было изучено влияние Rv2299c на активацию макрофагов или созревание ДК. Rv2299c не вызывал дополнительной активации макрофагов. Активность фагоцитоза в качестве маркера созревания ДК определяли по воздействию декстран-FITC. Количество дважды положительных клеток (CDllc+ и декстран-FIТС-положительных) было снижено в ДК, обработанных Rv2299c или LPS, используемых в качестве положительного контроля, что указывает на пониженную активность белка и повышенное функциональное созревание белка (см. Фиг. 1В). Далее, в ДК, обработанных Rv2299c в течение 24 часов, дозозависимым образом значимо усиливалась экспрессия молекул МНС класса I и II и костимулирующих молекул, таких как CD80 и CD86 (см. Фиг. 1С). Эти результаты показывают, что Rv2299c эффективно индуцирует созревание ДК.
Затем было проанализировано, связано ли Rv2299c-oпоcpeдованное созревание ДК с секрецией провоспалительных или противовоспалительных цитокинов. Как показано на Фиг. 1D, Rv2299c значимо стимулировал ДК секретировать высокие уровни TNF-α, IL-6 и IL-1β, тогда как необработанные ДК секретировали пренебрежимо малое количество цитокинов. Затем определяли выработку IL-12p70 и IL-10 со значимыми эффектами в отношении развития опосредованного Т-клетками иммунного ответа. В отличие от LPS, Rv2299c значимо индуцировал секрецию IL-12p70, в отличие от IL-10 (см. Фиг. 1D). Анализ FACS показал, что в IL-10-положительных клетках никаких изменений не было, но доля IL-12р70-положительных клеток была увеличена в ДК, обработанных Rv2299c, по сравнению с результатами, полученными для необработанных ДК (см. Фиг. 1Е). Хотя содержание эндотоксина определяло общее время выработки рекомбинантных белков, было подтверждено, что способность Rv2299c индуцировать созревание ДК не была обусловлена загрязнением LPS в результате обработки протеиназой К или термической денатурации (см. Фиг. 2). Кроме того, обработка полимиксином В не влияла на активность Rv2299c, но активность LPS была значимо ингибирована активностью LPS. С этой точки зрения полученные результаты свидетельствуют о том, что Rv2299c индуцирует секрецию провоспалительных цитокинов в ДК, и что такие ДК, созревшие под действием Rv2299c, могут стимулировать иммунный ответ Th1-типа.
<Пример 3>
Rv2299c индуцирует созревание дендритных клеток (ДК) посредством TLR4-пути.
3.1 Экспериментальный метод
(А) Гистограммы показывают экспрессию CD86 или МНСII в Rv2299c-обработанных CD11с+-отсортированных ДК, полученных от мышей WT, TLR2/ и TLR4/. ДК, полученные от мышей WT, TLR2/ и TLR4/, обрабатывали Rv2299c (10 мкг/мл) в течение 24 часов. На каждой панели показана доля положительных клеток. Гистограмма показывает среднее значение ± SEM соотношения для каждой поверхностной молекулы по отношению к клеткам CD11c+ в трех независимых экспериментах. Статистическая значимость (***р<0,001) выражается в виде числа Rv2299c-oбpaбoтанных TLR2/к Rv2299c-обработанным WT ДК. Все данные выражены как среднее значение ± SD (n=3), и статистическая значимость (***р<0,001) указана для обработки по сравнению с Rv2299c-oбpaбoтанными WT ДК. (В) ДК, полученные от мышей WT, TLR2/ и TLR4/, обрабатывали Rv2299c или LPS в течение 24 часов. Количество выработанных TNF-α, IL-6, или IL-1β в ДК, обработанных Rv2299c или LPS, полученных от мышей WT, TLR2/ и TLR4/, измеряли методом ИФА. Все данные представлены в виде среднего значения ± стандартное отклонение (n=3), и для обработанных клеток показана статистическая значимость (***р<0,001) по сравнению с WT ДК, обработанным Rv2299c. (С) ДК, полученные от мышей WT, MyD88/ и TRIF/, обрабатывали Rv2299c (10 мкг/мл) и LPS (100 нг/мл) в течение 24 часов. Количество выработанных TNF-α, IL-6, или IL-1β в Rv2299c- или LPS-обработанных ДК, полученных от мышей WT, MyD88/ и TRIF/, измеряли методом ИФА. Все данные представлены в виде среднего значения ± стандартное отклонение и статистическая значимость (***р<0,001) показана для обработанных клеток по сравнению с Rv2299c-обработанными WT ДК. (D) Интенсивность флуоресценции aнти-Rv2299c в сочетании с ДК, обработанными Rv2299c. ДК, полученные от мышей WT, TLR2/ и TLR4/, обрабатывали Rv2299c (10 мкг/мл) в течение 1 часа, фиксировали и окрашивали DAPI и Су3-конъюгированными анти-His-антителами (ссылка: 10 мкМ). (Е) ДККМ, полученные от мышей WT, TLR2/ и TLR4/, обрабатывали Rv2299c (10 мкг/мл) в течение 1 часа и окрашивали Аlеха488-конъюгированным анти-His mAb. Доля положительных клеток показана на каждой панели. Гистограмма показывает среднее ± SEM доли Rv2299c-Alexa488 в клетках CD11c+, полученных в трех независимых экспериментах. Статистическая значимость (***р<0,001) выражается в виде числа Rv2299c-oбpaбoтанныx TLR4/, по сравнению с Rv2299c-oбpaбoтанными WT ДК. (F) Иммунопреципитация (IP) с анти-His, aнти-TLR2 или aнти-TLR4 антителами и иммуноблоттинг с анти-His, aнти-TLR2 или aнти-TLR4 антителами. ДК обрабатывали Rv2299c (10 мкг/мл) в течение 6 часов. Клетки собирали, и клеточные лизаты иммунопреципитировали с помощью антитела против IgG мыши, антитела против IgG мыши, анти-His, aнти-TLR2 или анти-TLR4. Затем белки визуализировали иммуноблоттингом с анти-His, aнти-TLR2 или aнти-TLR4 антителами. Сумма обозначает среднее для общего клеточного лизата.
3.2 Анализ экспериментальных результатов
Было выявлено несколько компонентов Mtb, которые активируют ДК посредством пути Toll-подобного рецептора (TLR) (Harding and Boom, 2010). Поэтому был исследован вопрос, могут ли TLR распознавать Rv2299c в ДК и активировать их. Экспрессия поверхностных молекул (см. Фиг. 3А) и провоспалительных цитокинов (см. Фиг. 3В) была значимо ингибирована в ДК, полученных от мышей TLR4-/-, по сравнению с ДК, полученными от мышей дикого типа (WT) или TLR2-/- мышей. Это означает, что Rv2299c может быть агентом активации для TLR4. MyD88 является общей адаптерной молекулой для всех TLR, но молекула TRIF является необходимой для TLR4-опосредованной активации МуD88-независимого сигнального пути (Takeda et al., 2003). Определение выработки провоспалительных цитокинов в ДК, полученных от WT, MyD88-/- и TRIF-дефицитных мышей, показывает, что MyD88- и TRIF-зависимые пути участвуют в Rv2299c-индуцированной выработке TNF-α, IL-6 и IL-1β. (см. Фиг. 3С). Затем было исследовано, взаимодействует ли Rv2299c с молекулами TLR4 в ДК. Конфокальная микроскопия с Су3-конъюгированными поликлональными aнти-Rv2299c антителами показала, что Rv2299c преимущественно связывается с поверхностью ДК, полученных из WT и TLR2-/-, в отличие от TLR4-/- ДК (см. Фиг. 3D). Кроме того, связывание между молекулами Rv2299c и TLR4 было подтверждено анализом FACS (см. Фиг. 3Е). Чтобы подтвердить взаимодействие между Rv2299c и TLR4, были проведены анализы иммунопреципитации для aнти-TLR2 или aнти-TLR4 антител и анти-His антител. Было обнаружено, что Rv2299c связывается с TLR4, в отличие от TLR2 (см. Фиг. 3F). Эти результаты показывают, что Rv2299c индуцирует созревание ДК зависимым от TLR4 образом, что приводит к повышенной экспрессии молекул клеточной поверхности и провоспалительных цитокинов.
<Пример 4>
Пути МАРK и NF-κВ ассоциированы с созреванием Rv2299c-опосредованной дендритной клетки (ДК).
4.1 Экспериментальный метод.
(А) ДК обрабатывали 10 мкг/мл белков Rv2299c и наносили на график экспрессию белка в зависимости от времени. Клеточные лизаты помещали на ДНС-ПААГ и проводили иммуноблот-анализ с использованием соответствующих специфических антител к фосфо-р38 (р-р38), р38, фосфо-ERK1/2 (p-ERK1/2), фосфо-1κВ-а и р65 NF-κВ. (В) Влияние Rv2299c на клеточную локализацию субъединицы р65 NF-κВ в ДК. ДК помещали на предметное стекло закрытой стеклянной камеры и обрабатывали Rv2299c в течение 1 часа. После стимуляции определяли иммунореактивность субъединицы р65 внутриклеточного NF-***кВ по иммунофлуоресценции, как описано в разделе «Материалы и методы». (С и D) ДК обрабатывали р38 (SB203580, 20 мкМ), ERK1/2 (U0126, 10 мкМ), фармакологические ингибиторы JNK (SP600125, 20 мкМ), Bay11-7082 (20 мкМ) или DMSO (контроль-носитель) в течение 1 часа, после чего их обрабатывали 10 мкг/мл белков Rv2299c в течение 24 часов ч. Экспрессию CD80 и CD86 анализировали проточной цитометрией. Гистограмма показывает отношение соответствующих клеток CD11c+ к поверхностным молекулам (среднее значение ±SEM для трех отдельных экспериментов). Количества TNF-α, IL-6 и IL-1β в культуральной среде измеряли с помощью ИФА. Показано среднее ± SEM для трех независимых экспериментов, и статистическая значимость (*р<0,05, **р<0,01
или ***р<0,001) указана для обработки по сравнению с Rv2299c-обработанным контролем.
4.2 Анализ экспериментальных результатов
МАРK и NF-κВ являются критически важными сигнальными молекулами для регуляции созревания ДК и секреции провоспалительных цитокинов (Bansal et al., 2010b, Pathak et al., 2007). Поэтому было исследовано, были ли активированы МАРK и NF-κВ в ответ на Rv2299c. Как и ожидалось, Rv2299c вызывал фосфорилирование р38 и ERK1/2 и фосфорилирование и деградацию IκВ-α в ДК (см. Фиг. 4А) и индуцировал значимое изменение сайта р65 от цитозоля к ядру (см. Фиг. 4В). Чтобы подтвердить роль МАРK и NF-κB в Rv2299c-индуцированной выработке провоспалительных цитокинов и экспрессии костимулирующей молекулы, ДК предварительно обрабатывали ингибитором р38 (SB203580), ингибитором ERK1/2 (U0126), ингибитором JNK (SP600125) или ингибитором NF-κВ (Bay 11-0782) в течение 1 часа, прежде чем подвергнуть воздействию Rv2299c. Все фармакологические ингибиторы, за исключением ингибитора JNK, в значимой степени устраняли индуцированную Rv2299c экспрессию костимулирующих молекул (см. Фиг. 4С) и выработку провоспалительных цитокинов (см. Фиг. 4D) на поверхности ДК. Эти результаты показывают, что сигнальные пути МАРK и NF-κВ являются необходимыми для выработки провоспалительных цитокинов и экспрессии маркеров созревания ДК, индуцированной Rv2299c.
<Пример 5>
Созревшие под действием Rv2299c дендритные клетки (ДК) индуцируют пролиферацию наивных Т-клеток.
5.1 Экспериментальный метод
(А) Трансформированные OVA-специфические CD8+ Т-клетки и трансформированные OVA-специфические CD4+ Т-клетки выделяли и окрашивали CFSE. Их культивировали одновременно с ДК, обработанными Rv2299c (10 мкг/мл) или LPS (100 нг/мл), в течение 96 часов. OVA-специфические CD8+ Т-клетки и OVA-специфические CD4+ Т-клетки, соответственно, сенсибилизировали воздействием OVA257-264 (1 мкг/мл) и OVA323-339 (1 мкг/мл). Т-клетки по отдельности и Т-клетки, культивированные одновременно с необработанными ДК, служили контролем. Пролиферацию OT-I+ и OT-II+ Т-клеток оценивали с помощью проточной цитометрии. (В) Культивированные супернатанты и IFN-γ, IL-2 и IL-4, собранные через 24 часа после этого, измеряли с помощью ИФА. Показано среднее значение ± SEM для трех независимых экспериментов, а статистическая значимость (*р<0,05) указана для обработанных клеток по сравнению с соответствующим контролем (ДК-клетки/OVА257-264-сенсибилизированные ДК или Т-клетки/OVА323-339-сенсибилизированные ДК). "n.s" обозначает отсутствие значимого эффекта обработки.
5.2 Анализ экспериментальных результатов
Чтобы достоверно проиллюстрировать свойство Rv2299c активно влиять на взаимодействие между ДК и Т-клетками, проводили анализ изогенной MLR с использованием OT-I-TCR-трансформированных CD8+ Т-клеток и трансформированных ОТ-II TCR CD4+Т-клеток. ДК, сенсибилизированные OVA257-264 иди OVA323-339, культивировали совместно с трансформированными CFSE-мечеными OVA-специфичными CD4+ или CD8+ Т-клетками в течение 72 часов. ДК, обработанные Rv2299c или LPS, индуцировали пролиферацию Т-клеток в значимо большей степени по сравнению с необработанными ДК (см. Фиг. 5А). Кроме того, наивные CD4+ и CD8+ Т-клетки, сенсибилизированные Rv2299с-обработанными ДК, вырабатывались при значительно более высоких уровнях IFN-γ и IL-2 по сравнению с необработанными ДК (р<0,05-0,01), тогда как секреция IL-4 не увеличивалась независимо от Rv2299 с-стимуляции (см. Фиг. 5В). Эти результаты предполагают, что обработанные Rv2299c ДК индуцируют пролиферацию наивных Т-клеток в направлении фенотипа Th1.
<Пример 6>
Созревшие под действием Rv2299c дендритные клетки (ДК) индуцировали расширение популяций эффекторных Т-клеток/Т-клеток памяти.
6.1 Экспериментальный метод
(А) Т-клетки, полученные от мышей BALB/c, инфицированных Mycobacterium tuberculosis. ДК культивировали с 10 мкг/мл белков Rv2299c в течение 24 часов. Созревшие под действием ДК промывали и культивировали совместно с аллогенными Т-клетками при соотношении ДК к Т-клеткам 1:10 в течение 3 дней. Спленоциты окрашивали aнти-CD4, анти-CD8, aнти-CD62L и анти-СD44 mAb. На гистограмме показаны клетки CD62LlowCD44highT в селезенке. На гистограмме приведено соотношение (среднее ± SEM) для CD4+/CD8+CD44high и CD4+/CD8+CD62Llow Т-клеток в трех независимых экспериментах. (В) Экспрессию IFN-γ, IL-2 или IL-4 в клетках CD3+/CD4+и CD3+/CD8+анализировали в Т-клетках кишечных лимфатических узлов, совместно культивированных с ДК-сенсибилизированными ДК или LPS-сенсибилизированными ДК с помощью внутриклеточного окрашивания IFN-γ, IL-2 или IL-4. Доля дважды положительных клеток в Т-клетках показана в верхнем правом углу, и результаты отражают три независимых эксперимента. Статистическая значимость (**р<0,01 или ***р<0,001) указана для сравнения с необработанным ДК, a "n.s" означает отсутствие значимого эффекта.
6.2 Анализ экспериментальных результатов
Чтобы оценить, способны ли Rv2299с-стимулированные ДК в особенности стимулировать CD4+, полученные от мышей, инфицированных Mtb, использовали проточную цитометрию для анализа изменения экспрессии CD62L и CD44 в CD4+селезеночных Т-клетках, индуцированных Rv2299c-обработанными ДК. ДК получали из костного мозга мышей WT, TLR2-/- или TLR4-/- и стимулировали их созревание с помощью Rv2299c или LPS. Как созревшие под воздействием Rv2299c ДК, полученные от инфицированных мышей, так и сингенные CD4+Т-клетки совместно культивировали в течение 72 часов. Затем созревшие под воздействием Rv2299c ДК, полученные от мышей WT или TLR2-/-, в отличие от мышей TLR4-/-, показали пониженную экспрессию CD62L и повышенную экспрессию CD44 в CD4+ Т-клетках по сравнению с контрольными ДК или обработанными LPS ДК (см. Фиг. 6А). Доля CD4-IFN-γ или СD4-IL-2-положительных клеток была значимо выше в совместной культуре с созревшими под воздействием Rv2299c ДК, полученными от мышей WT или TLR2-/-, в отличие от мышей TLR4-/-, по сравнению с контрольными ДК или обработанными LPS ДК (см. Фиг. 6В). Кроме того, число СD4-IL-4-положительных клеток не увеличивалось из-за совместного культивирования с антиген-стимулированными ДК. Эти данные свидетельствуют о том, что созревшие под воздействием Rv2299c ДК индуцируют экспансию популяций эффекторных Т-клеток/Т-клеток памяти и ТLR4-зависимым образом управляют иммунным ответом типа Th1.
<Пример 7>
Т-клетки, активированные созревшими под воздействием Rv2299c дендритными клетками (ДК), ингибируют рост Mtb в клетках.
7.1 Экспериментальный метод
МКМ инфицировали H37Rv (MOI=1) в течение 4 часов. Затем инфицированные МКМ обрабатывали амикацином (200 мкг/мл) в течение 2 часов и дважды промывали PBS. Затем предварительно приготовленную смесь добавляли в каждую лунку планшета и культивировали в течение 3 дней. Полученная смесь представляла собой активированные антигеном ДК, которые совместно культивировали с CD4+ Т-клетками при соотношении ДК к Т-клеткам 1:10 в течение 3 дней. Активирующие ДК антигены представляли собой LPS (100 нг/мл) и Rv2299c (10 мкг/мл). (А) Количество Mtb в зараженных макрофагах. (В) Супернатант анализировали с помощью ИФА. Показанные данные представляют собой среднее значение ± SD (n=3); *р<0,05, **р<0,01 или ***р<0,001 относится к значимости для обработанных клеток по сравнению с необработанными клетками, "n.s" обозначает отсутствие значимого эффекта данной обработки.
7.2 Анализ экспериментальных результатов
Чтобы подтвердить, что созревшие под воздействием Rv2299c ДК действительно играют роль в контроле Mtb, было исследовано, могут ли Т-клетки, активированные созревшими под воздействием Rv2299c ДК, усиливать бактерицидное действие в макрофагах. Наивные Т-клетки селезенки от неинфицированных мышей активировали путем совместного культивирования с созревшими под воздействием Rv2299c ДК в течение 72 часов и затем добавляли к Mtb-инфицированным МКМ. Как показано на Фиг. 7, простое добавление инактивированных Т-клеток значимо ингибировало рост Mtb в клетках. Интересно, что рост Mtb был значимо ингибирован обработкой Т-клеток, активированных Rv2229 с-стимулированными ДК, по сравнению с инактивированными Т-клетками или Т-клетками, активированными LPS-стимулированными ДК. Уровни провоспалительных цитокинов, IFN-γ и IL-17, ассоциированных с противотуберкулезной активностью, были значимо повышены при добавлении к ним Т-клеток, активированных Rv2299c-стимулированными ДК, по сравнению с инактивированными Т-клетками или Т-клетками, активированными LPS-стимулированными ДК. Эти результаты показывают, что ингибирование роста Mtb ассоциировано с этой выработкой цитокинов (см. Фиг. 7). Полученные результаты показывают, что Rv2299c индуцирует активацию Т-клеток с бактерицидной активностью посредством созревания ДК.
<Пример 8>
Гибридный белок Rv2299c/ESAT-6 усиливает иммунный ответ ESAT-6.
8.1 Экспериментальные методы для подтверждения того, что гибридные белки индуцирует созревание дендритных клеток (ДК)
(А) ДККМ культивировали в течение 24 часов в присутствии 1, 2, 5, 10 или 20 мкг/мл ESAT-6 или гибридных белков и анализировали проточной цитометрией. Положительным контролем был стауроспорин. ДК окрашивали анти-CD11c, аннексином V и PI. Показана доля положительных клеток (окрашенных аннексином V и PI клеток) в каждом квадранте. Результаты представляют три эксперимента. (В) ДККМ культивировали в присутствии 1, 5 или 10 мкг/мл гибридных белков, 2 мкг/мл ESAT-6 или 10 мкг/мл Rv2299c в течение 24 часов и анализировали с помощью проточной цитометрии с двухцветной маркировкой. Клетки были отсортированы для исключения клеток CD11c. ДК окрашивали анти-СD40, анти-СD80, анти-СD86 или анти-МНС класса II. Полученные данные представляют три экспериментальных результата. Средняя интенсивность флуоресценции рассчитывали из приведенного выше графика. Каждый столбик представляет средний уровень флуоресценции образца. Данные представлены как среднее ± SEM (n=5). *р<0,05, **р<0,01 и ***р<0,001 по сравнению с необработанным контролем, ESAT-6 или Rv2299c. (С) Супернатант измеряли с помощью ИФА. Результаты получены из одного репрезентативного эксперимента из трех проведенных экспериментов. Данные представлены как среднее ± SEM (n=5). *р<0,05, **р<0,01 и ***р<0,001 по сравнению с необработанным контролем, ESAT-6 или Rv2299c.
8.2 Экспериментальный метод для подтверждения ингибирования роста Mtb в макрофагах гибридными белками
МКМ инфицировали H37Rv (MOI=1) в течение 4 часов. Затем инфицированные МКМ обрабатывали амикацином (200 мкг/мл) в течение 2 часов и дважды промывали PBS. Затем предварительно приготовленную смесь добавляли в каждую лунку планшета и культивировали в течение 3 дней. Смесь представляла собой активированные антигеном ДК, которые совместно культивировали с CD4+ Т-клетками при соотношении ДК к Т-клеткам 1:10 в течение 3 дней. Антигенами, активировавшими ДК, были ESAT-6 (2 мкг/мл), Rv2299c и гибридный белок (10 мкг/мл). (А) Количество Mtb в зараженных макрофагах. (В) Супернатант анализировали с помощью ИФА. Показанные данные представляют собой среднее значение ± SD (n=3); *р<0,05, **р<0,01 или ***р<0,001 относится к значимости для обработанных клеток по сравнению с необработанными клетками, "n.s" обозначает отсутствие значимого эффекта данной обработки.
8.3 Анализ экспериментальных результатов
Предполагалось, что ДК-активированные белки могут усиливать профилактический иммунитет стимулирующих Т-клетки антигенов. Чтобы проверить это предположение, ESAT-6, основной кандидат на Т-клеточную вакцину, экспрессировали и очищали в белках, слитых с Rv2299c в E.coli, и исследовали его иммуногенность. Во-первых, определяли цитотоксичность рекомбинантного слитого с Rv2299c ESAT-6 в МКМ окрашиванием аннексином V и йодидом пропидия (PI). ESAT-6 показал цитотоксичность выше 5 мкг/мл, но гибридные белки не показали токсичности при 10 мкг/мл (см. Фиг. 8А). Поэтому в последующих экспериментах ESAT-6 использовали в концентрации 2 мкг/мл. Костимуляция, экспрессия молекул МНС и выработка провоспалительных цитокинов, таких как TNF-α, IL-1β и IL-12, значимо повышались в ДК, стимулированных гибридными белками, по сравнению с одиночным белком (см. Фиг. 8 В). Затем определяли бактерицидную активность Т-клеток, стимулированных созревшими под действием гибридных белков ДК. Как показано на Фиг. 9А, внутриклеточный рост Mtb был значимо ингибирован добавлением Т-клеток, активированных ESAT-6-стимулированными ДК, по сравнению с МКМ-клетками без каких-либо Т-клеток или ДК, но Т-клетки, активированные Rv2299c-cтимyлиpoванными ДК или стимулированными гибридным белком ДК, значимо ингибировали внутриклеточный рост Mtb по сравнению с другими условиями. Была выявлена высокая выработка провоспалительных цитокинов, таких как IFN-γ и TNF-α, когда Mtb-инфицированные МКМ культивировали совместно с Т-клетками, активированными стимулированными гибридным белком ДК (см. Фиг. 9В).
<Пример 9>
Гибридные белки Rv2299c/ESAT-6 вызывают значимый "прайм-бустерный" эффект БЦЖ в отношении высоковирулентного штамма Mtb HN878.
9.1 Экспериментальный метод
(А) График вакцинации и план эксперимента, (В) Различие в степени бактериального роста в одном легком мыши показано в легком мышей, вакцинированных только БЦЖ, только белком, гибридным белком БЦЖ-Rv2299c/ESAT-6 и контролем с адъювантом (только MPL-DDA) через 16 недель после заражения Mtb HN878. (С) Срезы легких каждой мыши, вакцинированной через 16 недель после заражения Mtb HN878 (n=6 животных/группа), окрашивали гематоксилином и эозином (D) Процент участков воспаленной ткани в срезах ткани легких. G1: инфицированный контроль, G2: контроль инокуляции MPL/DDA, G3: только БЦЖ, G4: вакцинация гибридным белком БЦЖ-Rv2299c/ESAT-6/MPL-DDA, G5: группа инокуляции ESAT-6/MPL-DDA. *Р<0,05, **р<0,01 и ***р<0,001 по сравнению с группой, получавшей только MPL-DDA.
9.2 Анализ экспериментальных результатов
В конечном итоге, было оценено, является ли гибридный белок Rv2299c/ESAT-6 потенциальной вакциной против туберкулеза. Во-первых, для оценки на модели на мышах вакцинного эффекта белка Rv2299c отдельно использовали штамм Mtb Erdman. Однако с точки зрения снижения бактериальной нагрузки в легких и селезенке, инокуляция Rv2299c/MPL-DDA не показала значимого защитного действия в отношении штамма Erdman.
Затем было оценено, является ли гибридный белок Rv2299c/ESAT-6 потенциально БЦЖ-прайм-бустером в модели заражения Mtb HN878, высокопатогенного штамма М. tuberculosis. Предыдущие исследования показали, что вакцинация мышей с использованием только БЦЖ обеспечивает меньшую защиту от изолятов W-Beijing по сравнению с Mtb H37Rv через 10 недель после заражения (Lopez et al., 2003). Поэтому, как описано выше, вакцинную эффективность белка в модели на мышах проверяли с использованием клинически высоковирулентного штамма HN878, одного из семейств W-Beijing (Cha et al., 2015b). Через четыре недели после последней вакцинации мышей заражали штаммом Mtb HN878, и определяли бактериальную нагрузку в их легких через 16 недель (см. Фиг. 10A). Как показано на Фиг. 10В, только БЦЖ, ESAT-6 и гибридный белок Rv2299c/ESAT-6 все значимо снижали бактериальную нагрузку в легких по сравнению с контрольной группой, получавшей адъювант. Только БЦЖ и гибридный белок показали сходные профилактические эффекты, а вакцинация ESAT-6 показала более высокую бактериальную нагрузку в легких. Однако три эти вакцины не показали большой разницы между собой. Также была исследована способность гибридного белка быть бустером вакцины БЦЖ. Через 16 недель после заражения у мышей, получавших бустинг гибридным белком, было обнаружено значимо меньшее количество бактерий, чем у мышей, получавших вакцину БЦЖ. Также оценивали воспаление легких после вакцинации. Через 16 недель после заражения в группе, вакцинированной гибридным белком Rv2299c/ESAT-6, наблюдали значимое снижение воспаления легких по сравнению с контролем БЦЖ (р<0,01). В целом, гибридный белок Rv2299c/ESAT-6 продемонстрировал профилактический эффект против заражения штаммом HN878 с точки зрения долгосрочного снижения количества бактерий. Кроме того, через 16 недель после заражения группа, вакцинированная гибридным белком Rv2299c/ESAT-6, показала значимое снижение процента воспаления легких по сравнению с инфицированным контролем (р<0,01) (см. Фиг. 10С и 10D).
<Пример 10>
Иммуногенность антиген-специфических Тh1 иммунных ответов у мышей, инфицированных Mtb HN878
10.1 Экспериментальный метод подтверждения индукции антиген-специфических многофункциональных Т-клеток в легком мыши с иммунитетом
Каждую группу мышей вакцинировали и умерщвляли, как описано в разделе «Материалы и методы». Через 4 недели после последней вакцинации каждую группу мышей (n=6) подвергали эвтаназии, и их легочные клетки стимулировали меченым антигеном в присутствии GolgiStop при 37°С в течение 12 часов. Долю антиген-специфических CD4+ Т-клеток, вырабатывающих IFN-γ, TNF-α и/или IL-2, в клетках, выделенных из легких вакцинированных мышей, анализировали с использованием многоцветной проточной цитометрии отсортировывая лимфоциты CD4+. На круговой диаграмме представлено среднее количество клеток, совместно экспрессирующих IFN-γ, TNF-α и/или IL-2. Данные выражали в виде среднего значения ± SD, полученного от 5 мышей в каждой группе, а для определения значимости использовали t-критерий для независимой выборки. *Р<0,05, **р<0,01 и ***р<0,001 по сравнению с группой, в которой использовали только MPL-DDA (t-критерий для независимой выборки).
10.2 Экспериментальный метод подтверждения сохранения многофункциональных Т-клеток в селезенке после заражения штаммом Mycobacterium tuberculosis HN878.
После 16 недель заражения каждую группу мышей (n=6) подвергали эвтаназии, и их клетки селезенки (2,0×106 клеток) стимулировали каждым антигеном (5 мкг/мл) в присутствии GolgiStop при 37°С в течение 12 часов. Долю антиген-специфических CD4+ Т-клеток, вырабатывающих IFN-γ, TNF-α и/или IL-2, в клетках, выделенных из легких каждой группы мышей, анализировали с использованием многоцветной проточной цитометрии отсортировывая лимфоциты CD4+. На круговой диаграмме представлено среднее количество клеток, совместно экспрессирующих IFN-γ, TNF-α и/или IL-2. Данные выражали в виде среднего значения ± SD, полученного от 5 мышей в каждой группе, а для определения значимости использовали t-критерий для независимой выборки. *Р<0,05, **р<0,01 и ***р<0,001 по сравнению с группой, в которой использовали только MPL-DDA (t-критерий для независимой выборки).
10.3 Анализ вышеприведенных экспериментальных результатов
Th1-опосредованные иммунные ответы и многофункциональные Т-клетки играют важную роль в профилактическом иммунитете против Mtb. Поэтому затем были оценены изменения в фенотипе Т-клеток, вызванные вакцинацией гибридным белком Rv2299c/ESAT-6. Через 16 недель после заражения клетки легкого (см. Фиг. 11А и 11 В) и клетки селезенки (см. Фиг. 12А и 12В) выделяли и стимулировали каждым иммунизирующим антигеном, а фенотип реагирующих CD4+ Т-клеток оценивали многоцветным внутриклеточным окрашивание цитокинов. По сравнению с другими группами уровни трижды положительных CD4+ Т-клеток (ко-экспрессирующие IFN-γ, TNF-α и IL-2) в легких были увеличены и поддерживались только в группе, вакцинированной гибридным белком Rv2299c/ESAT-6 (см. Фиг. 11А, 11В, 12А и 12В). Кроме того, была показана повышенная частота дважды положительных многофункциональных CD4+Т-клеток (TNF-α+IL-2+CD4+ Т-клеток) после заражения в группе, вакцинированной гибридным белком Rv2299c/ESAT-6 (см. Фиг. 11А, 11В, 12А и 12В).
<Пример 11>
Обсуждение
Было обнаружено, что субъединичные белковые вакцины потенциально полезны для замены бустинга БЦЖ или индуцированных БЦЖ иммунных реакций (Dalmia and Ramsay, 2012). Предлагается новая стратегия разработки вакцин для улучшения профилактического эффекта кандидатов на вакцины против туберкулеза. Наиболее важным этапом в разработке вакцины является выбор оптимального антигена для включения в мультибелковую вакцину. ДК регулируют последующее развитие типов Th1 или Th2 при поляризации наивных Т-клеток и дренировании лимфатических узлов. Таким образом, вакцины на основе белка, активированного ДК, могут улучшить профилактический иммунитет других вырабатываемых антигенов. В этом исследовании было обнаружено, что белок Rv2299c, способный активировать ДК, является значимой мишенью для разработки противотуберкулезной вакцины. Это показывает, что гибридный белок Rv2299c/ESAT-6 достоверно снижает бактериальную нагрузку в легких после заражения высоковирулентными клиническими штаммами Mtb HN878.
Основной принцип разработки вакцины против туберкулеза, предназначенной для индукции Т-клеточного иммунитета, основан на предположении, что сильный Th1-ответ специфически против антигенов туберкулеза, является важным механизмом противотуберкулезного иммунитета. Антигены, которые не индуцируют Th1-ответ, не обладают вакцинной эффективностью против Mtb, но не все белки, которые индуцируют сильный ответ Th1, вызывают значимый защитный эффект (Orr et al., 2014). Вакцина MVA85A, которая вызывает надежный Th1-ответ против Ag85A, не проявляет выраженного бустинг-эффекта БЦЖ у людей (Tameris et al., 2013). Важно, чтобы вакцина вызывала ответ Т-клеток памяти и давала долгосрочный иммунный ответ. ДК играют критическую роль в эффективной активации Т-клеток и индукцией ответов Т-клеток памяти. Известно, что мыши, вакцинированные Mtb-инфицированными ДК (Tascon et al., 2000) или ДК, обработанными экстрактом антигена (Rubakova et al., 2007) защищены от заражения Mtb. Эти результаты подтверждают стратегию разработки противотуберкулезной вакцины, которая нацелена на ДК in vivo (Sinha et al., 2007). В этом исследовании, для проверки гипотезы о том, что ДК-активированные белки являются потенциальными кандидатами на вакцину, важным вопросом исследования является выбор надежного ДК-активированного белка. Во-первых, микобактериальные HSP выбираются потому, что микобактериальные HSP ассоциированы со способностью активировать ДК иммунных клеток, такие как макрофаги и ДК (Wang et al., 2001, Wang et al., 2002, Franco et al., 2008) и адъювантную активность (Srivastava et al., 1998). Это исследование показывает, что Rv2299c (HSP90) среди микобактериальных HSP индуцирует созревание ДК посредством передачи сигналов TLR4 и активации МАРK и NF-κВ. Эта активация приводит к повышению экспрессии костимулирующих молекул, стимулирующих иммунные ответы Th1-типа и секрецию провоспалительных цитокинов. Недавно сообщалось, что ДК, выделенные из клеток селезенки мышей, обработанных Rv2299c, приводят к повышенной экспрессии молекул МНС и костимулирующих молекул, а нагруженные ДК опухолевые антигены с обработкой Rv2299c вызывают усиленный эффект противоопухолевого иммунитета (Vo et al., 2015). Эти результаты и результаты нашего исследования показывают, что Rv2299c обладает адъювантной активностью благодаря активации ДК in vivo и ex vivo.
Сообщалось, что некоторые микобактериальные белки активируют ДК, заставляя их индуцировать ответ инфекции Thl посредством пути TLR2 (Bansal et al., 2010а, Byun et al., 2012a) или путь TLR4 (Choi et al., 2015, Kim et al., 2013), или индуцировать ответ инфекции Th2 (Bansal et al., 2010b) посредством пути TLR2 (Bansal et al., 2010b). Это свидетельствует, что каждый антиген обладает различной активностью в дендритных клетках. Рекомбинантный микобактериальный HSP70 передает сигнал через TLR4 и TLR2, тогда как микобактериальный HSP65 передает сигнал только через TLR4 вызывая активацию иммунных клеток (Bulut et al., 2005). HSP90 человека вызывает биологическую активность посредством пути TLR4 (Thuringer et al., 2011, Hutchinson et al., 2009). Это исследование демонстрирует, что Rv2299c семейства HSP90 активирует ДК зависимым от TLR4, MyD88 и TRIF образом.
Это продемонстрировано с помощью анализа с соосаждением и экспериментов с использованием мышей, нокаутных по TRL4 или TRL2.
Предполагается, что для надежной вакцины на основе ДК важно активировать Т-клетки с антимикобактериальной активностью. Однако о фактическом противомикобактериальном эффекте, вызываемом активируемым ДК белком, который индуцирует иммунный ответ Th1, известно мало. Это исследование демонстрирует, что Т-клетки, активированные созревшими под воздействием Rv2299c ДК, значимо ингибируют рост Mtb в макрофагах и индуцируют большой объем выработки цитокинов по сравнению с инактивированными Т-клетками. Т-клетки, неспецифически активированные созревшими под воздействием LPS ДК, не проявляли бактерицидную активность. Эти результаты демонстрируют, что Rv2299c индуцирует специфическую активацию наивных CD+ Т-клеток с профилактической активностью.
Сообщалось, что субъединичная вакцина, состоящая из двух-четырех антигенов гибридных белков, вызывает сильный профилактический ответ и эффект, эквивалентный профилактике, индуцируемой одной только БЦЖ. Большинство людей вакцинированы БЦЖ в младенчестве, но эффективность этой вакцины со временем уменьшается (Bertholet et al., 2010). Поэтому бустинг БЦЖ-индуцированного профилактического иммунитета может быть наиболее рациональной стратегией. В связи с этим были исследованы профилактический иммунный ответ и эффекты гибридных белков, состоящих из Rv2299c/ESAT-6. ESAT-6 выбран в качестве партнера для гибрида, поскольку ESAT-6 является одним из основных антигенов, включаемых в вакцину в современных клинических испытаниях. В этом исследовании мыши, инокулированные гибридным белком, показали такую же эффективность вакцины, что и мыши, инокулированные только БЦЖ. Было высказано предположение, что вторичные иммунные ответы Т-клеток памяти не зависят от ДК, и для инициирования вторичных ответов может быть достаточно других не специализированных антигенпрезентирующих клеток (Bertram et al., 2004, Dawicki and Watts, 2004), в то время как другие исследования показали, что активация Т-клеток памяти в основном зависит от ДК в ответ на системную или локальную инфекцию (Zammit et al., 2005; Wakim et al., 2008). В этом исследовании было обнаружено, что созревшие под действием Rv2299c ДК индуцируют экспансию эффекторных Т-клеток/Т-клеток памяти CD4+, полученных от Mtb-инфицированных мышей. Это свидетельствует, что Rv2299c может индуцировать вторичный иммунный ответ Т-клеток памяти у субъектов, вакцинированных БЦЖ. Бустинг БЦЖ с помощью белка ESAT-6, слитого с Rv2299c, уменьшает патологию и обеспечивает долговременную защиту от Mtb по сравнению с одной БЦЖ. В частности, поскольку вакцинация БЦЖ у мышей обеспечивает меньшую защиту от штамма W-Beijing по сравнению с Mtb H37Rv, было определено воздействие гибридного белка на Mtb HN878, вирулентный клинический штамм, полученный в семействе W-Beijing (Lopez et al., 2003). Этот штамм вызывает смерть и обширную патологию легких у инфицированных мышей C57BL/6.
Профилактический эффект вакцинации гибридным белком Rv2299c/ESAT-6 исследовали в отношении штамма Mtb HN878, высоковирулентного штамма W-Beijing, на модели на мышах. Мыши, иммунизированные гибридным белком Rv2299c/ESAT-6, были в существенной степени профилактически защищены после заражения штаммом Mtb HN878 судя по оценке легочной патологии и роста бактерий (см. Фиг. 10). Вакцинация гибридным белком Rv2299c/ESAT-6 показала значимо сниженную бактериальную нагрузку в легких, а также заметно уменьшала патологию легких, тем самым продемонстрировав замечательную профилактическую эффективность против Mtb HN878.
В целом, вакцинно-индуцированная профилактика против Mtb-инфекции конкретно ассоциирована с Ag-специфическими многофункциональными IFN-γ+TNFα+IL-2+ и TNF-α+IL-2+CD4+ Т-клетками в легких. Важно отметить, что тестирование на Ag-иммунизированньгх мышах показывает, что иммунизация гибридным белком Rv2299c/ESAT-6 заметно индуцирует выработку многофункциональных CD4+ Т-клеток в легких до и после заражения (см. Фиг. 11 и 12). Исходя из этой тенденции, Rv2299c может служить главным элементом успешной вакцины против Mtb-инфекции. Иммунизация гибридным белком Rv2299c/ESAT-6, в отличие от иммунизации одной БЦЖ или одним белком, индуцировала специфичный для белка Rv2299c/ESAT-6 ответ многофункциональных CD4+ Т-клеток в легком. Способность этой вакцины-кандидата индуцировать специфичные для гибридного белка Rv2299c/ESAT-6 ответы многофункциональных Т-клеток в легком после внутримышечной инъекции согласуется с наблюдаемым профилактическим эффектом. Кроме того, этот ответ, вызванный иммунизацией гибридным белком Rv2299c/ESAT-6, сохранялся до 16 недель после заражения. Это свидетельствует, что их уровень значимо увеличивается после распознавания инфекции Mtb первичными многофункциональными Т-клетками памяти CD4+. Эти результаты могут указывать на важность соответствующего обеспечения способности вакцин-кандидатов индуцировать иммунные реакции в легких.
В заключение, полученные результаты позволяют предположить, что Rv2299c является отличным кандидатом для рационального дизайна мультиантигенеых эффективных противотуберкулезных вакцин. Поэтому необходимы более подробные исследования для изучения специфической эффективности вакцины и эффективных профилактических механизмов гибридного белка Rv2299c/ESAT-6 на моделях на мышах или морских свинках.


--->
Список последовательностей
<110> Quratis Inc.
<120> КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ
ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6
<130> QURATIS1-6P
<150> KR 2016-0070517
<151> 2016-06-07
<160> 5
<170> KoPatentIn 3.0
<210> 1
<211> 647
<212> PRT
<213> Искусственная последовательность
<220>
<223> Rv2299c
<400> 1
Met Asn Ala His Val Glu Gln Leu Glu Phe Gln Ala Glu Ala Arg Gln
1 5 10 15
Leu Leu Asp Leu Met Val His Ser Val Tyr Ser Asn Lys Asp Ala Phe
20 25 30
Leu Arg Glu Leu Ile Ser Asn Ala Ser Asp Ala Leu Asp Lys Leu Arg
35 40 45
Ile Glu Ala Leu Arg Asn Lys Asp Leu Glu Val Asp Thr Ser Asp Leu
50 55 60
His Ile Glu Ile Asp Ala Asp Lys Ala Ala Arg Thr Leu Thr Val Arg
65 70 75 80
Asp Asn Gly Ile Gly Met Ala Arg Glu Glu Val Val Asp Leu Ile Gly
85 90 95
Thr Leu Ala Lys Ser Gly Thr Ala Glu Leu Arg Ala Gln Leu Arg Glu
100 105 110
Ala Lys Asn Ala Ala Ala Ser Glu Glu Leu Ile Gly Gln Phe Gly Ile
115 120 125
Gly Phe Tyr Ser Ser Phe Met Val Ala Asp Lys Val Gln Leu Leu Thr
130 135 140
Arg Lys Ala Gly Glu Ser Ala Ala Thr Arg Trp Glu Ser Ser Gly Glu
145 150 155 160
Gly Thr Tyr Thr Ile Glu Ser Val Glu Asp Ala Pro Gln Gly Thr Ser
165 170 175
Val Thr Leu His Leu Lys Pro Glu Asp Ala Glu Asp Asp Leu His Asp
180 185 190
Tyr Thr Ser Glu Trp Lys Ile Arg Asn Leu Val Lys Lys Tyr Ser Asp
195 200 205
Phe Ile Ala Trp Pro Ile Arg Met Asp Val Glu Arg Arg Thr Pro Ala
210 215 220
Ser Gln Glu Glu Gly Gly Glu Gly Gly Glu Glu Thr Val Thr Ile Glu
225 230 235 240
Thr Glu Thr Leu Asn Ser Met Lys Ala Leu Trp Ala Arg Pro Lys Glu
245 250 255
Glu Val Ser Glu Gln Glu Tyr Lys Glu Phe Tyr Lys His Val Ala His
260 265 270
Ala Trp Asp Asp Pro Leu Glu Ile Ile Ala Met Lys Ala Glu Gly Thr
275 280 285
Phe Glu Tyr Gln Ala Leu Leu Phe Ile Pro Ser His Ala Pro Phe Asp
290 295 300
Leu Phe Asp Arg Asp Ala His Val Gly Ile Gln Leu Tyr Val Lys Arg
305 310 315 320
Val Phe Ile Met Gly Asp Cys Asp Gln Leu Met Pro Glu Tyr Leu Arg
325 330 335
Phe Val Lys Gly Val Val Asp Ala Gln Asp Met Ser Leu Asn Val Ser
340 345 350
Arg Glu Ile Leu Gln Gln Asp Arg Gln Ile Lys Ala Ile Arg Arg Arg
355 360 365
Leu Thr Lys Lys Val Leu Ser Thr Ile Lys Asp Val Gln Ser Ser Arg
370 375 380
Pro Glu Asp Tyr Arg Thr Phe Trp Thr Gln Phe Gly Arg Val Leu Lys
385 390 395 400
Glu Gly Leu Leu Ser Asp Ile Asp Asn Arg Glu Thr Leu Leu Gly Ile
405 410 415
Ser Ser Phe Val Ser Thr Tyr Ser Glu Glu Glu Pro Thr Thr Leu Ala
420 425 430
Glu Tyr Val Glu Arg Met Lys Asp Gly Gln Gln Gln Ile Phe Tyr Ala
435 440 445
Thr Gly Glu Thr Arg Gln Gln Leu Leu Lys Ser Pro His Leu Glu Ala
450 455 460
Phe Lys Ala Lys Gly Tyr Glu Val Leu Leu Leu Thr Asp Pro Val Asp
465 470 475 480
Glu Val Trp Val Gly Met Val Pro Glu Phe Asp Gly Lys Pro Leu Gln
485 490 495
Ser Val Ala Lys Gly Glu Val Asp Leu Ser Ser Glu Glu Asp Thr Ser
500 505 510
Glu Ala Glu Arg Glu Glu Arg Gln Lys Glu Phe Ala Asp Leu Leu Thr
515 520 525
Trp Leu Gln Glu Thr Leu Ser Asp His Val Lys Glu Val Arg Leu Ser
530 535 540
Thr Arg Leu Thr Glu Ser Pro Ala Cys Leu Ile Thr Asp Ala Phe Gly
545 550 555 560
Met Thr Pro Ala Leu Ala Arg Ile Tyr Arg Ala Ser Gly Gln Glu Val
565 570 575
Pro Val Gly Lys Arg Ile Leu Glu Leu Asn Pro Ser His Pro Leu Val
580 585 590
Thr Gly Leu Arg Gln Ala His Gln Asp Arg Ala Asp Asp Ala Glu Lys
595 600 605
Ser Leu Ala Glu Thr Ala Glu Leu Leu Tyr Gly Thr Ala Leu Leu Ala
610 615 620
Glu Gly Gly Ala Leu Glu Asp Pro Ala Arg Phe Ala Glu Leu Leu Ala
625 630 635 640
Glu Arg Leu Ala Arg Thr Leu
645
<210> 2
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Rv2299c, прямой праймер
<400> 2
catatgaacg cccatgtcga gcagttg 27
<210> 3
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Rv2299c, обратный праймер
<400> 3
gaattcggca aggtacgcgc gagacgttc 29
<210> 4
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ESAT-6, прямой праймер
<400> 4
aagcttatga cagagcagca gtggaat 27
<210> 5
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ESAT-6, обратный праймер
<400> 5
ctcgagtgcg aacatcccag tgacgtt 27
<---
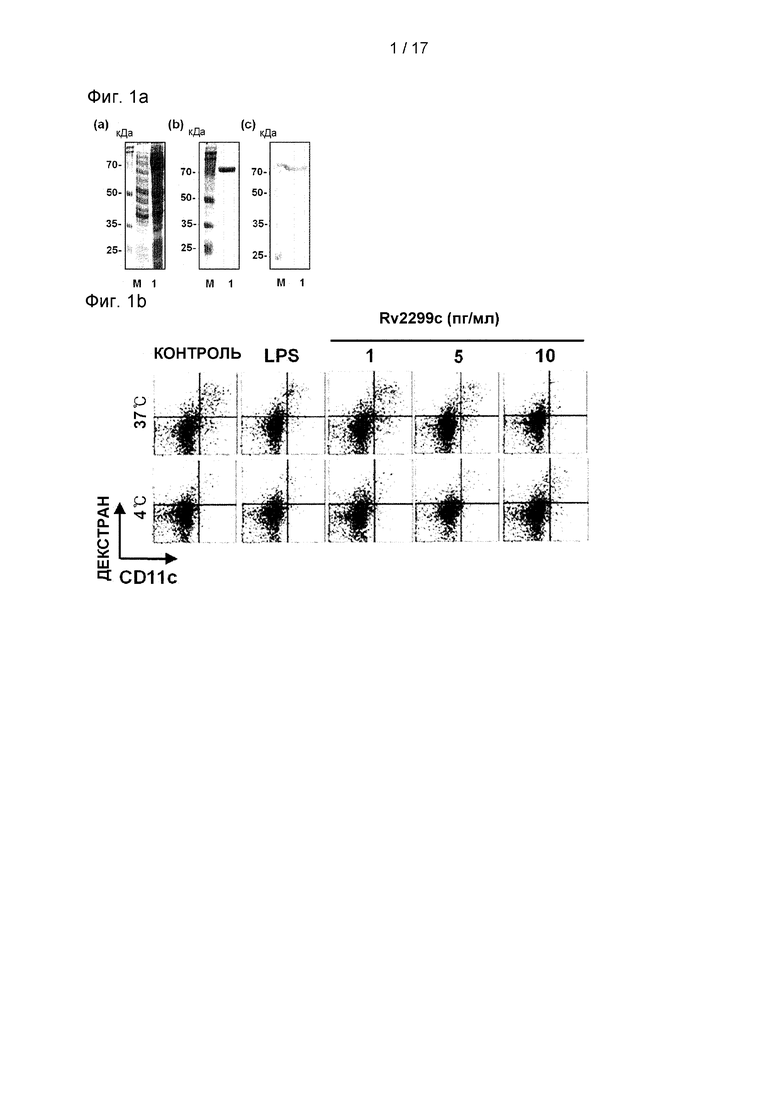
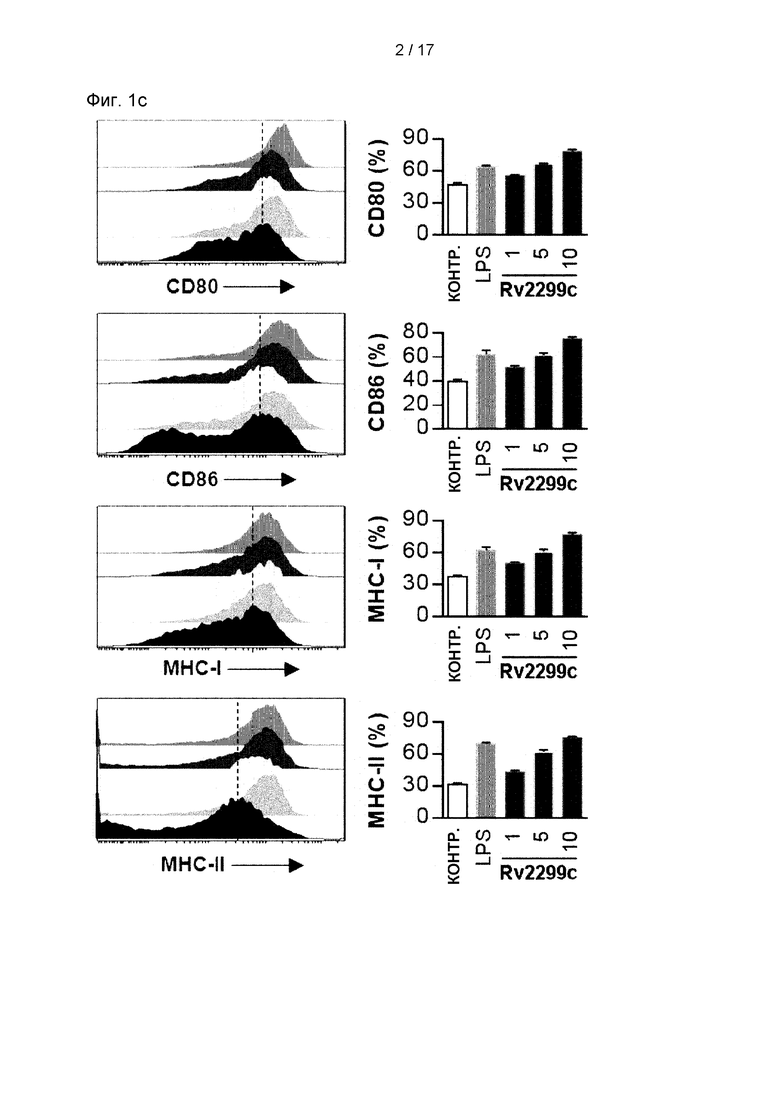
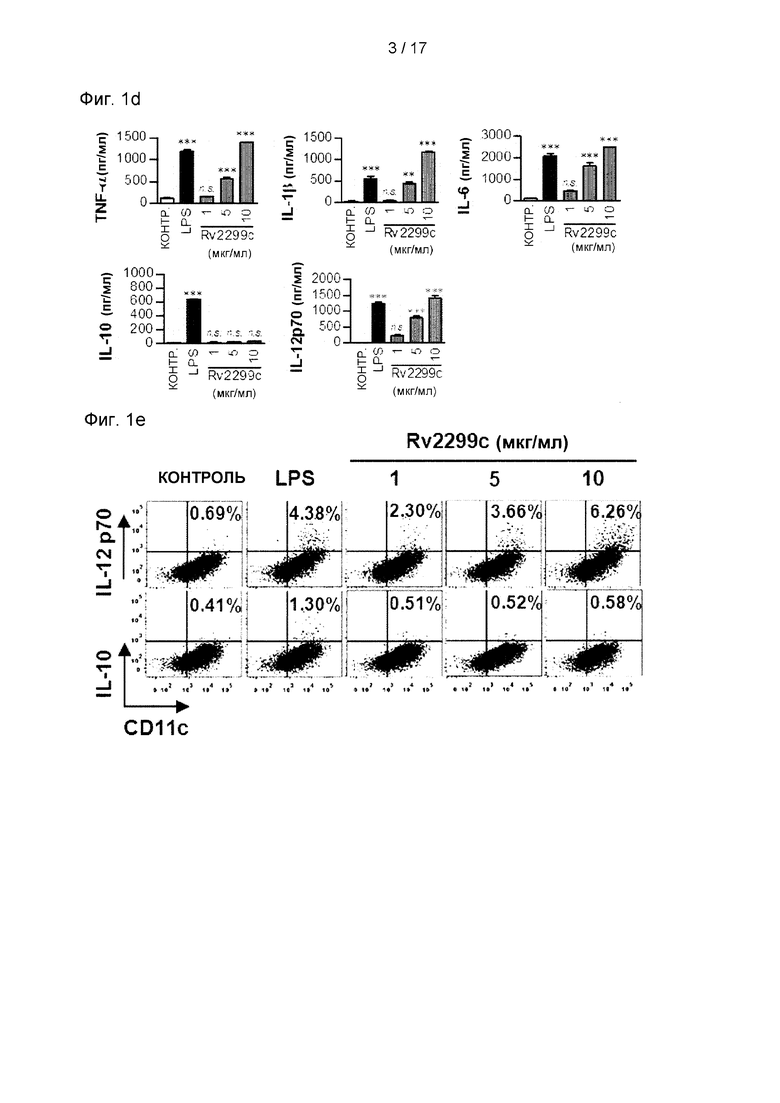
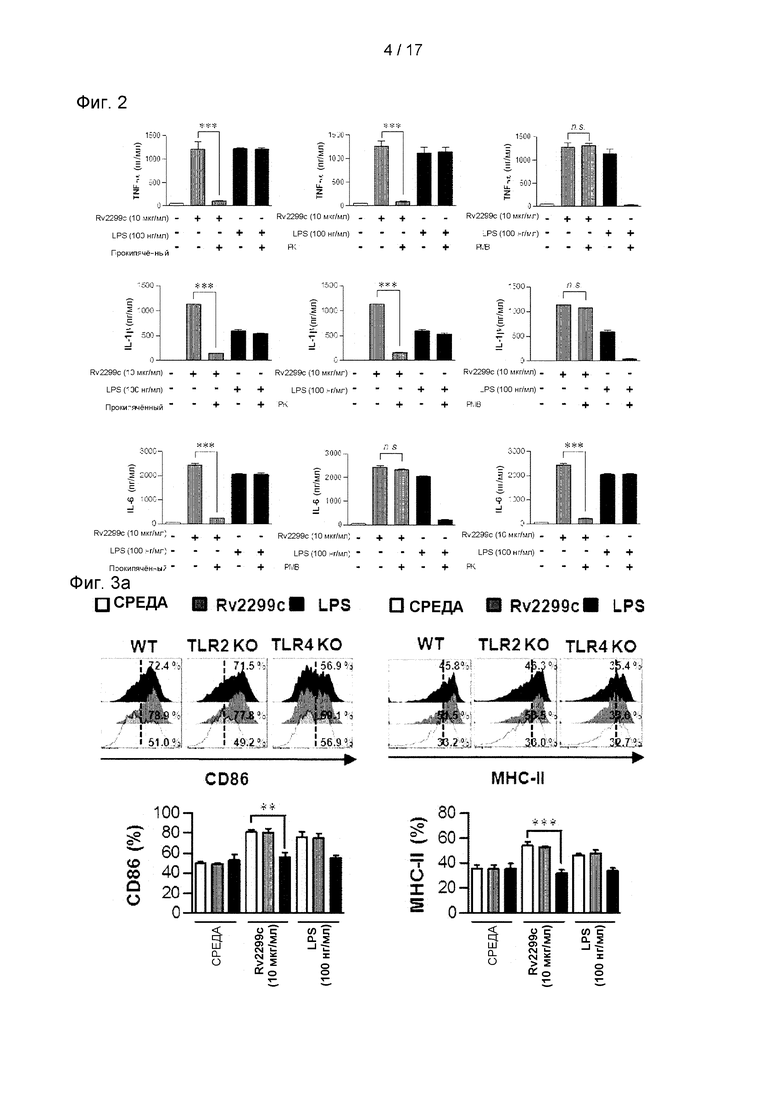
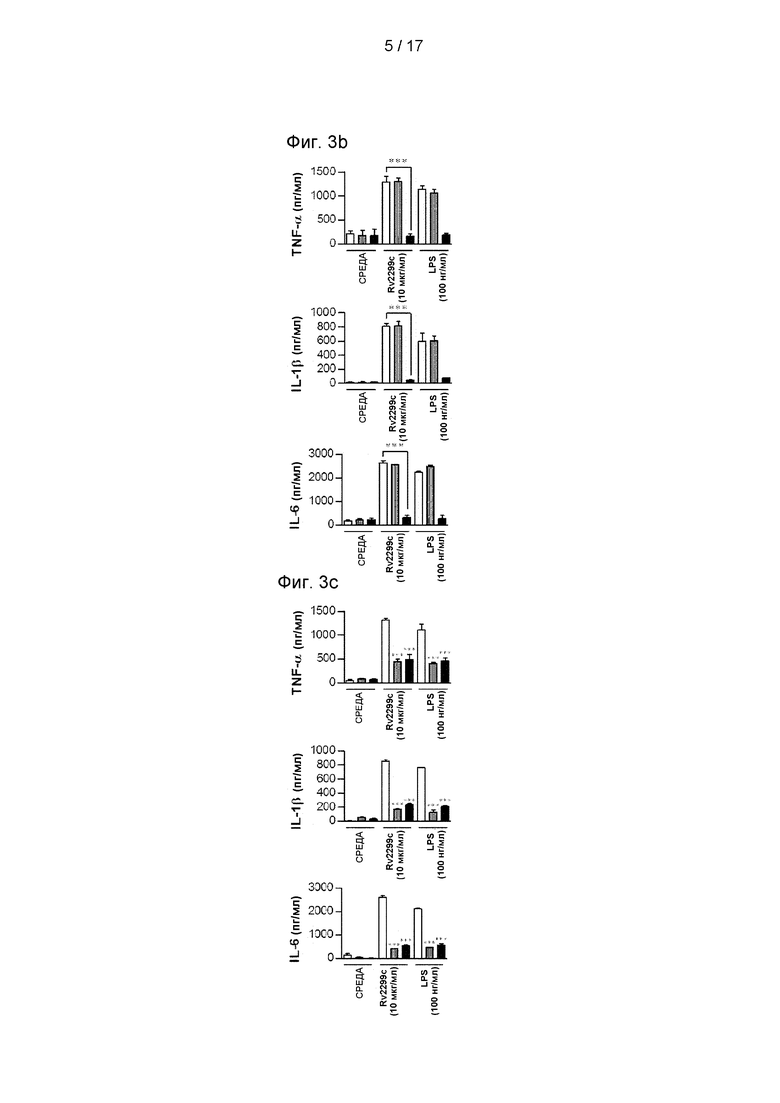
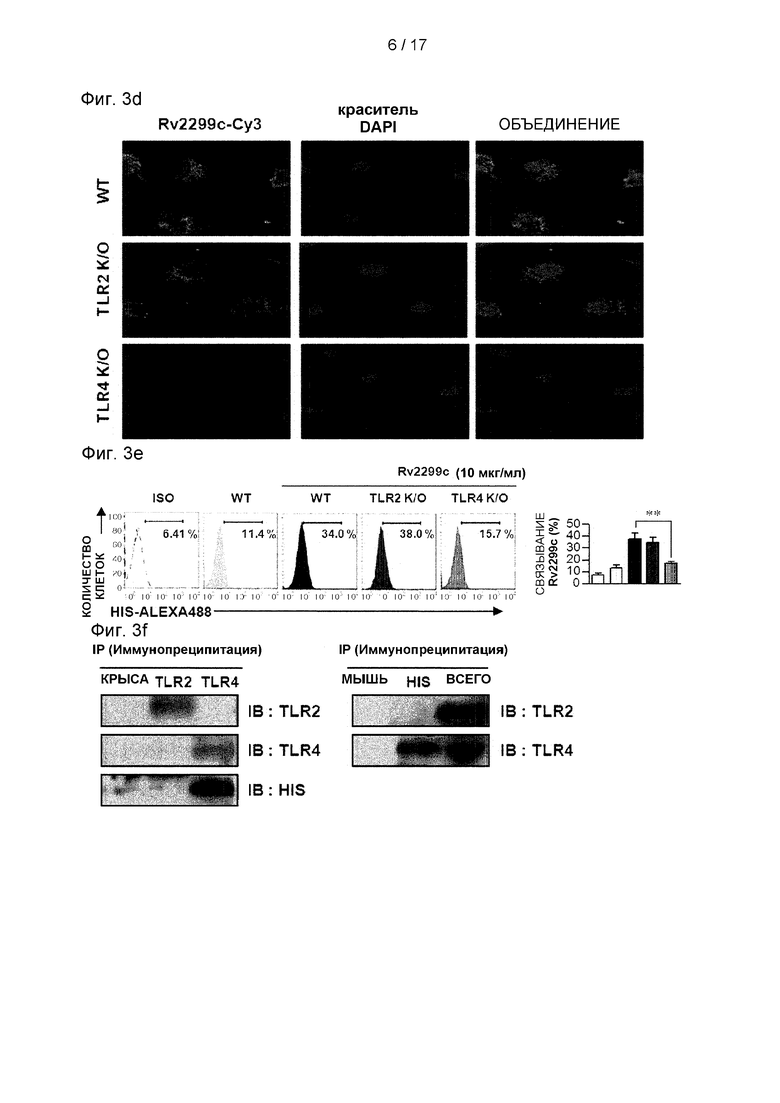
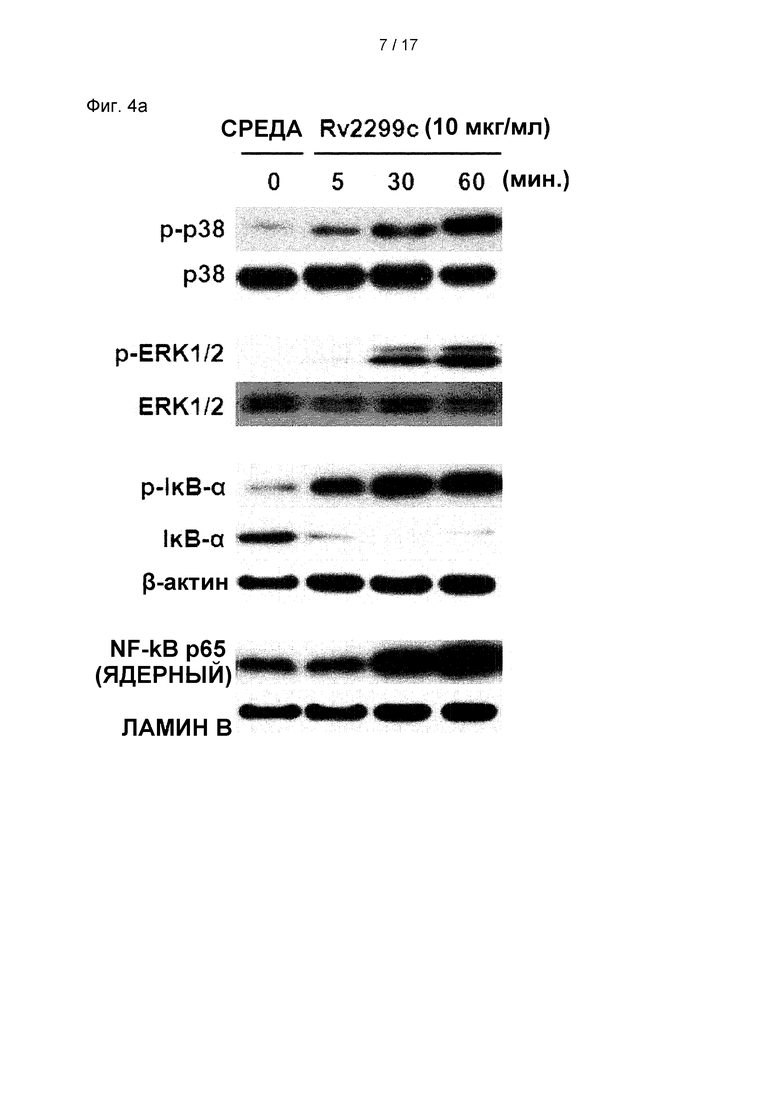
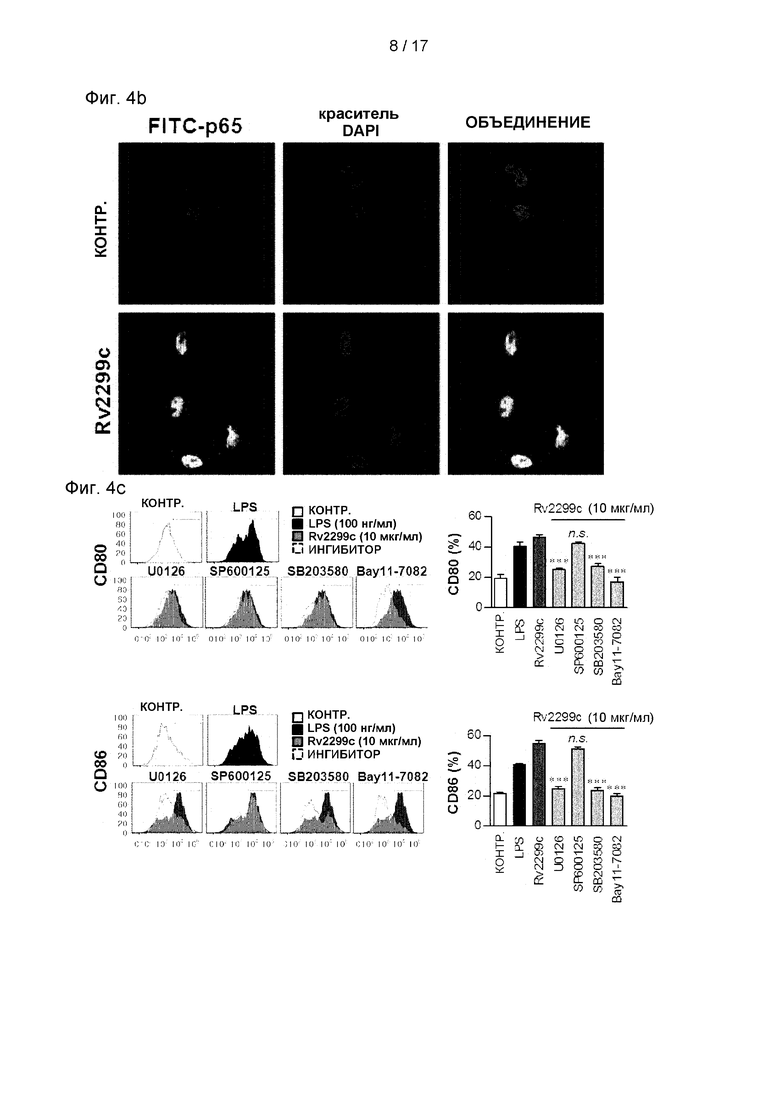
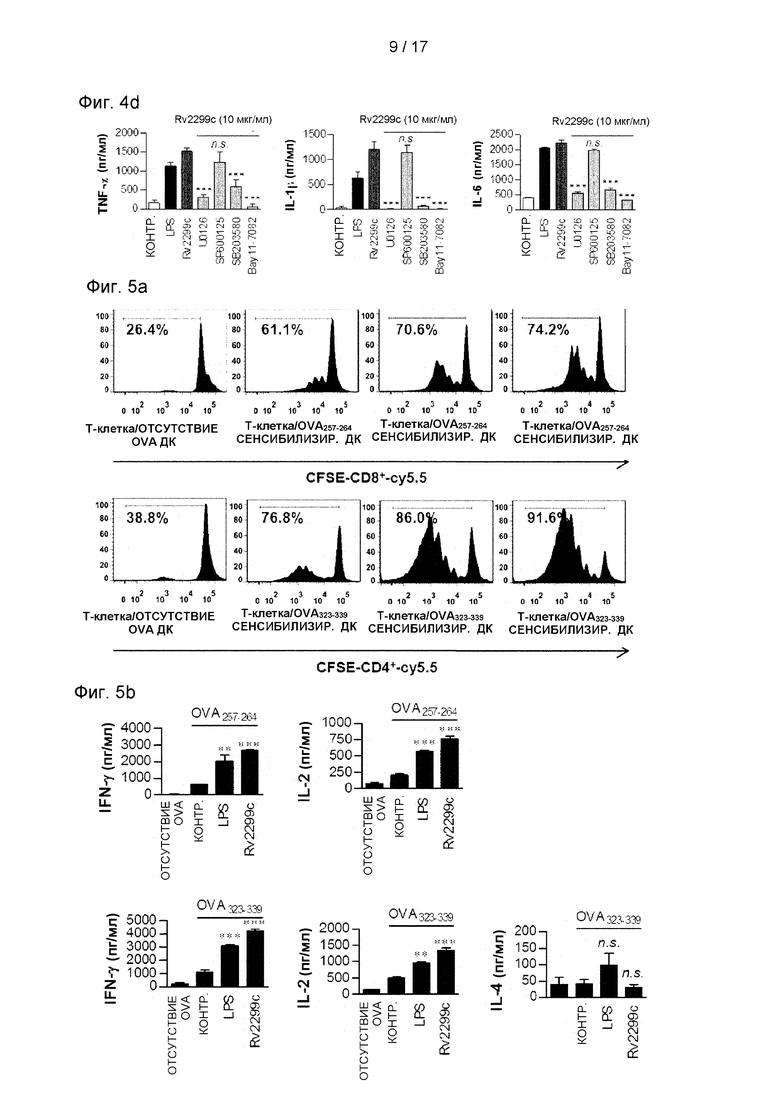
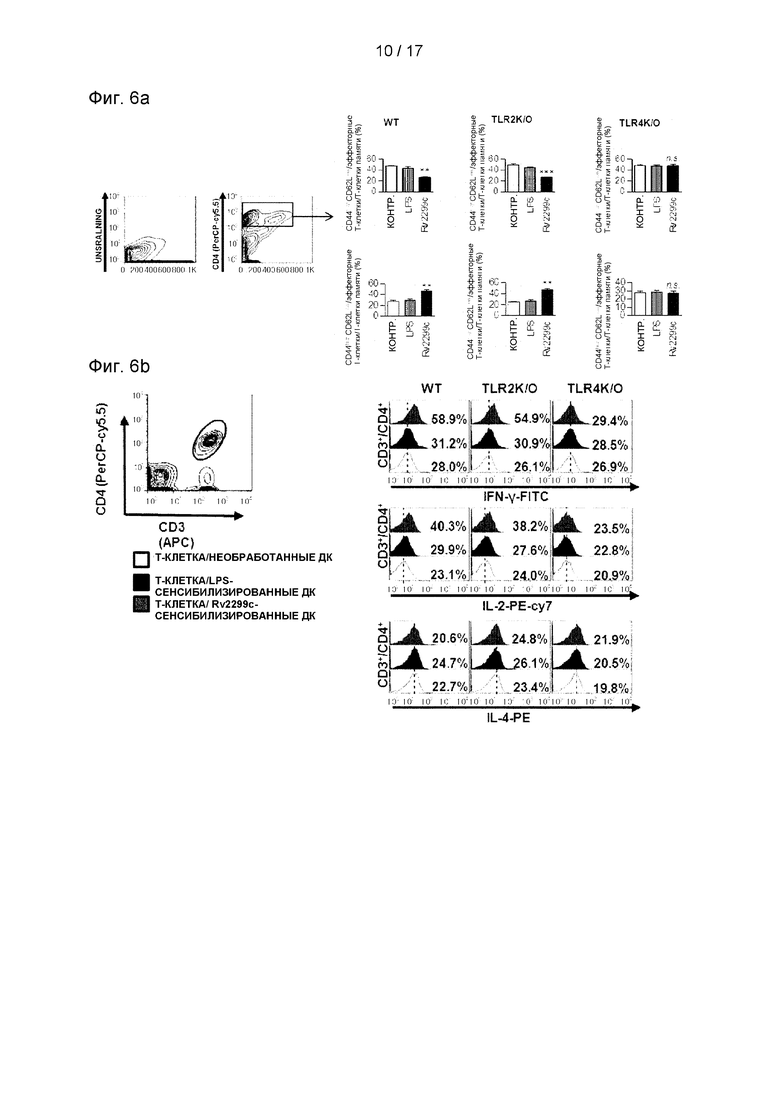
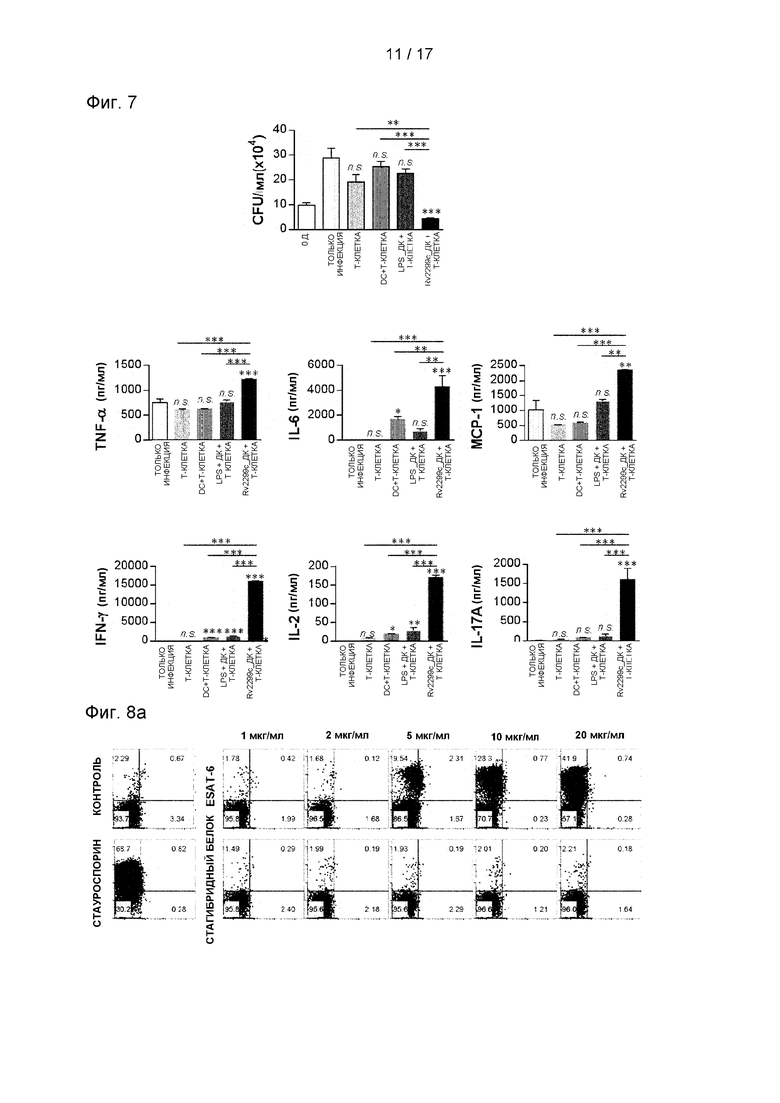
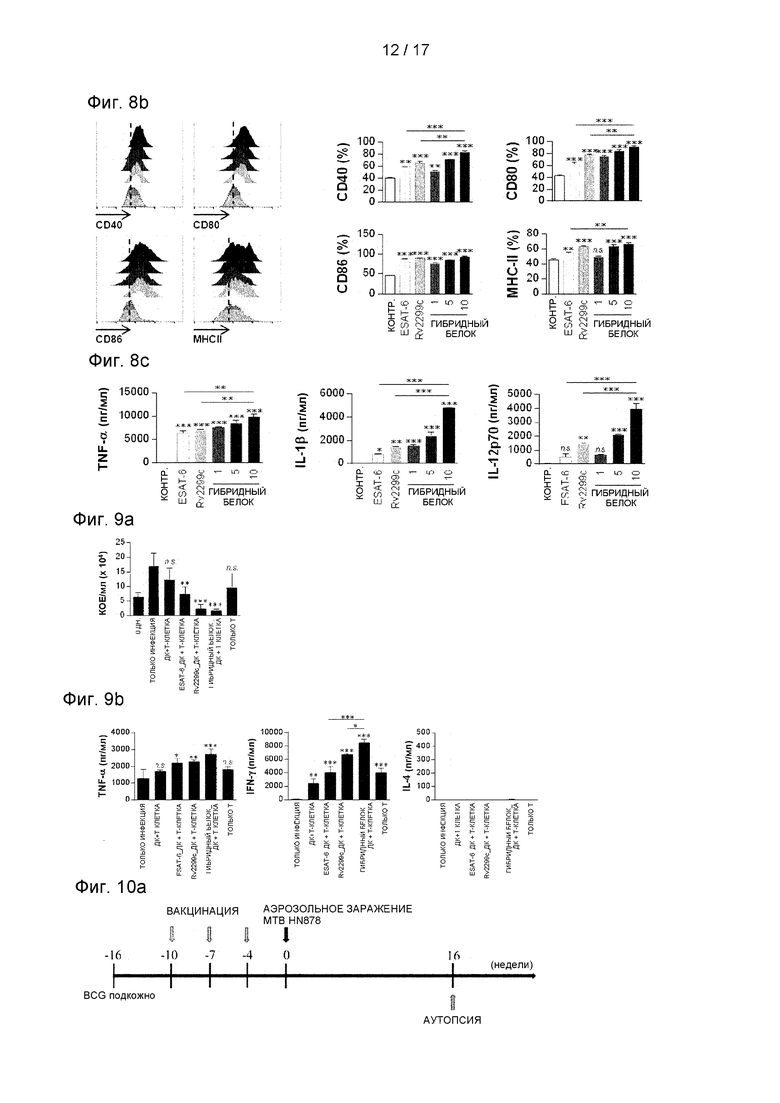
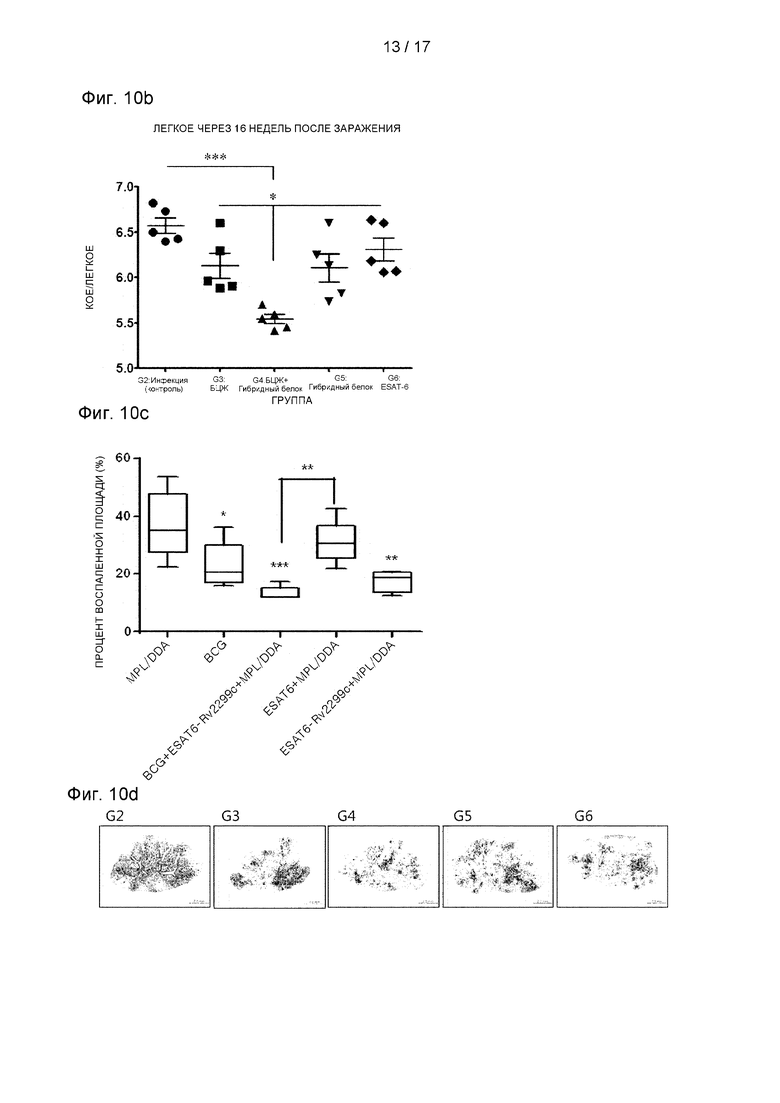
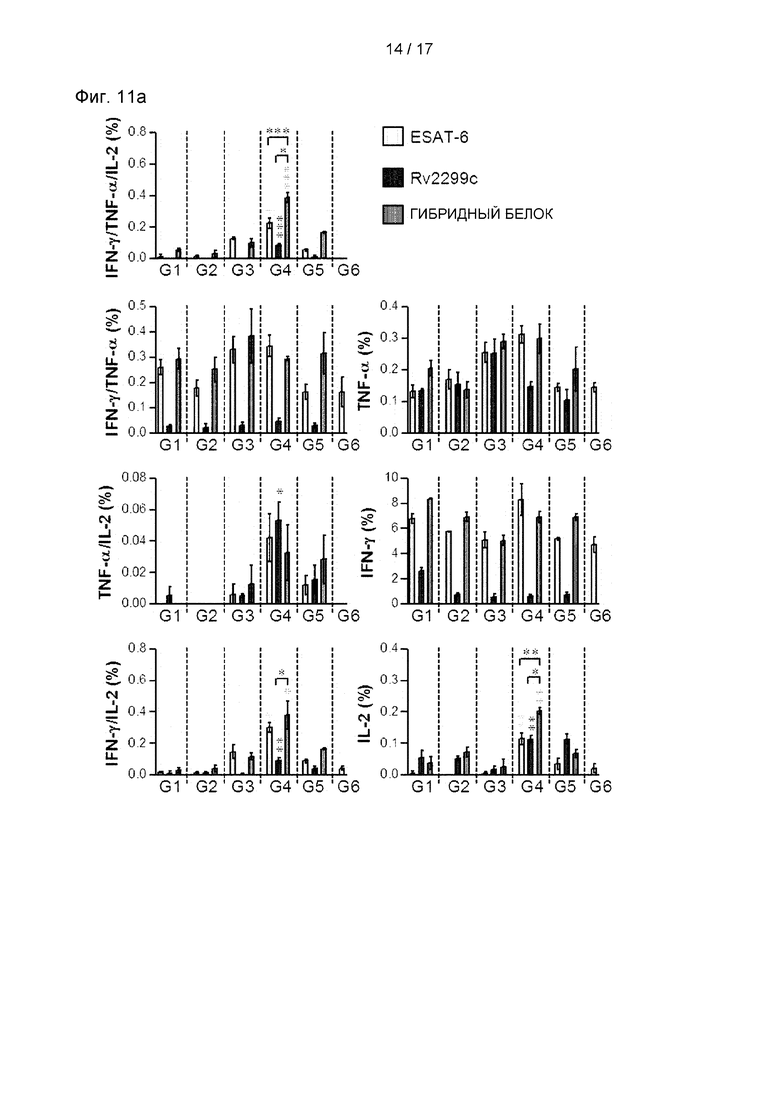
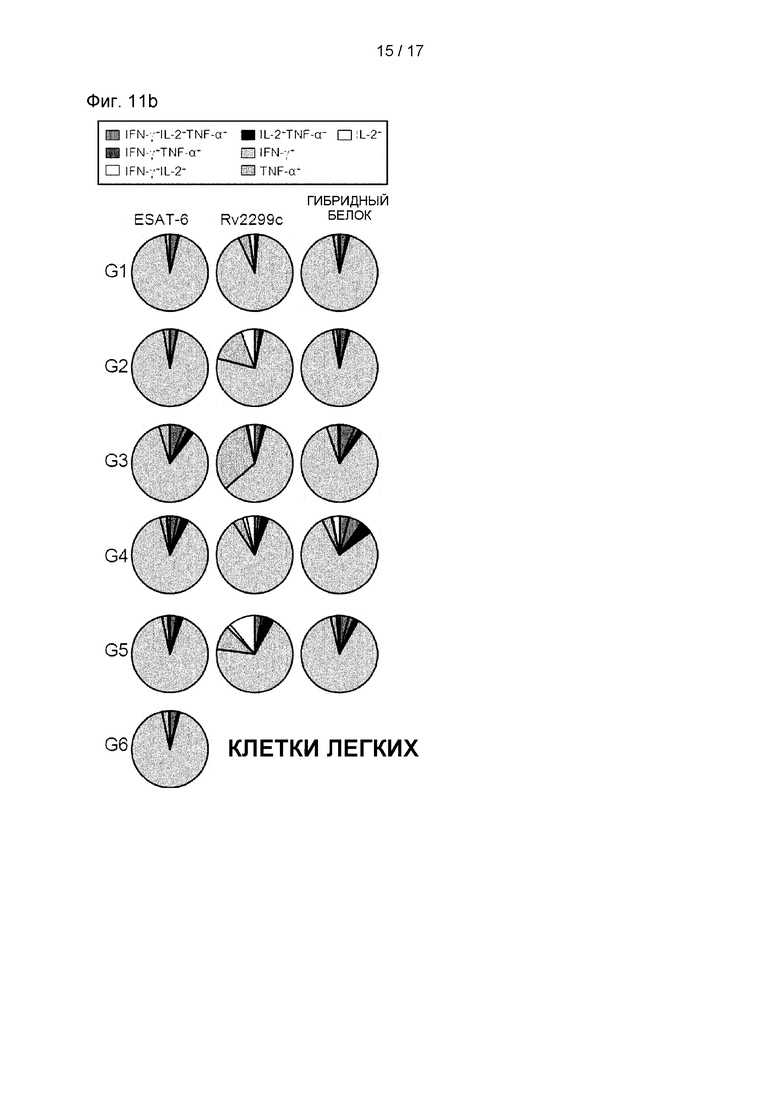
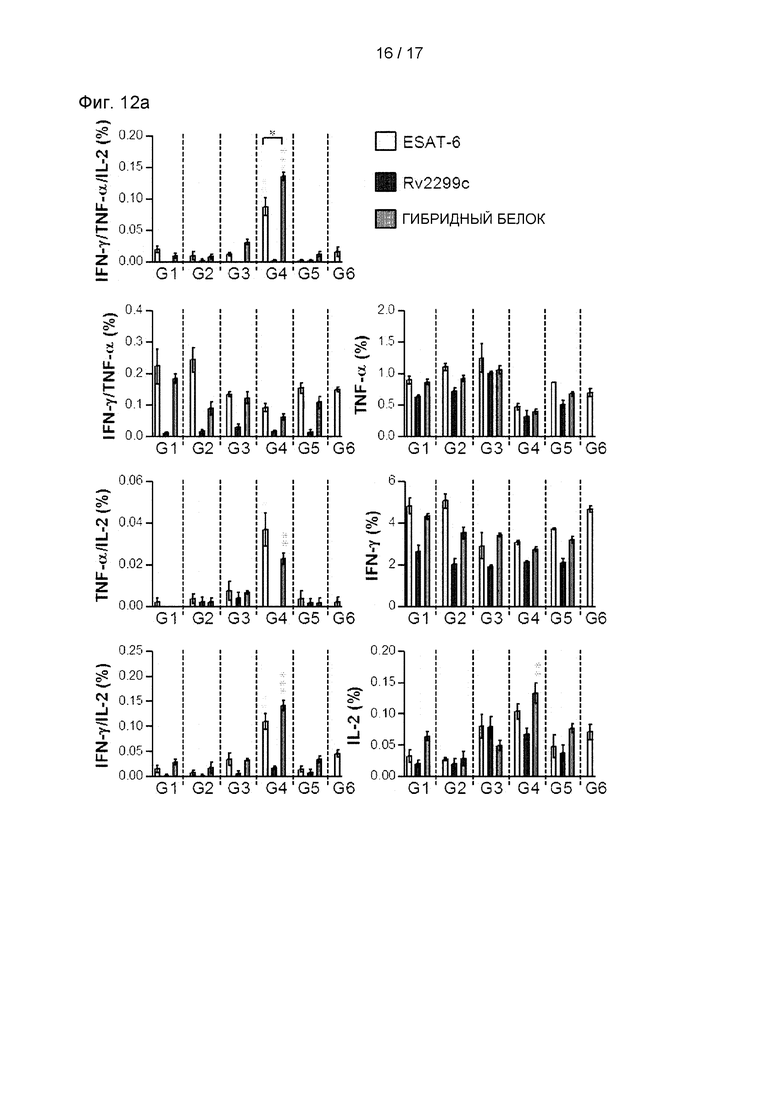
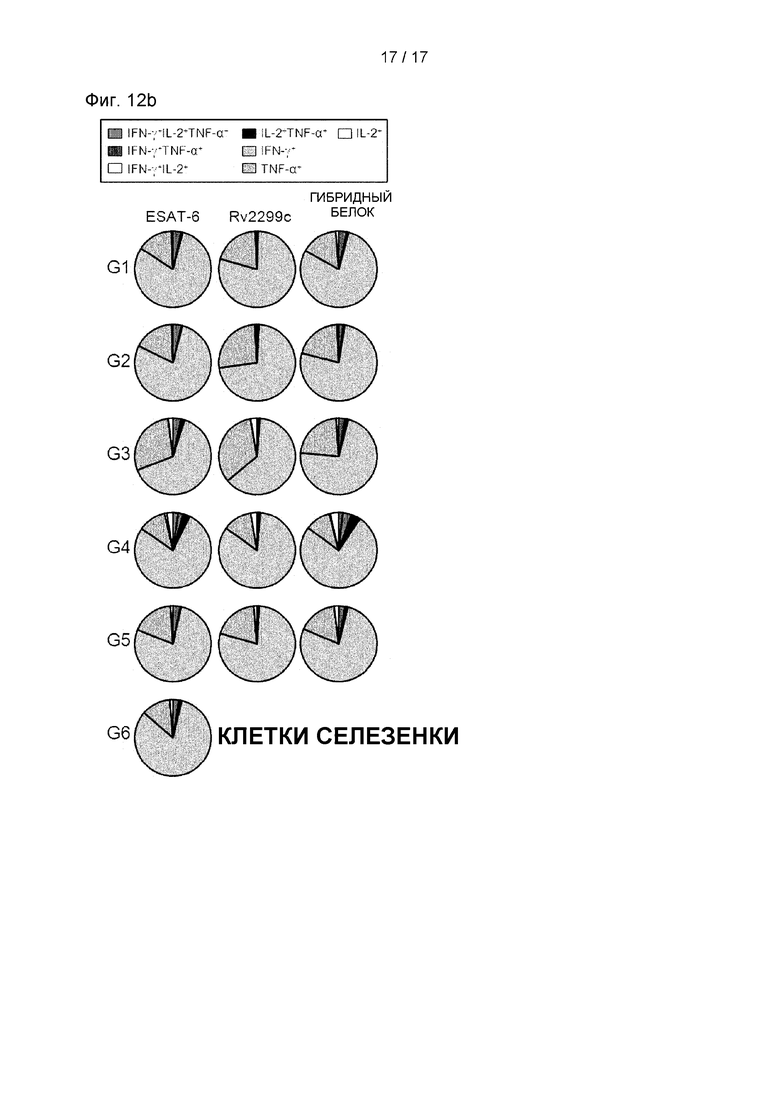
Настоящее изобретение относится к области биотехнологии, конкретно к гибридным иммуномодуляторным белкам, и может быть использовано для индукции созревания дендритных клеток (ДК) и усиления иммунитета. Гибридный белок Rv2299c/ESAT-6 состоит из белка ESAT-6, слитого с Rv2299c, и способен к индукции созревания ДК. Способ индукции созревания незрелых ДК включает дифференцировку мононуклеарных клеток, выделенных из крови, в незрелые ДК; и обработку указанных незрелых ДК гибридным белком Rv2299c/ESAT-6 для обеспечения созревания указанных незрелых ДК в зрелые ДК. При этом указанный гибридный белок Rv2299c/ESAT-6 применяют в концентрации от 1 мкг/мл до 20 мкг/мл. Изобретение обеспечивает получение нетоксичного иммуномодулятора для индукции иммунного ответа путем стимулирования дифференцировки ДК. 3 н. и 6 з.п. ф-лы, 12 ил., 11 пр.
1. Применение гибридного белка Rv2299c/ESAT-6 в качестве активного ингредиента для индукции созревания дендритных клеток, при этом белок Rv2299c имеет аминокислотную последовательность SEQ ID NO: 1.
2. Применение по п.1, отличающееся тем, что указанный гибридный белок Rv2299c/ESAT-6 представляет собой белок, происходящий из M. tuberculosis.
3. Применение по п.1, отличающееся тем, что указанный гибридный белок Rv2299c/ESAT-6 применяют в концентрации от 1 мкг/мл до 20 мкг/мл.
4. Способ индукции созревания незрелых дендритных клеток, включающий:
(a) осуществление дифференцировки мононуклеарных клеток, выделенных из крови, в незрелые дендритные клетки; и
(b) осуществление обработки указанных незрелых дендритных клеток гибридным белком Rv2299c/ESAT-6 для обеспечения созревания указанных незрелых дендритных клеток в зрелые дендритные клетки, при этом указанный гибридный белок Rv2299c/ESAT-6 применяют в концентрации от 1 мкг/мл до 20 мкг/мл.
5. Способ по п.4, отличающийся тем, что после обработки указанных незрелых дендритных клеток гибридным белком Rv2299c/ESAT-6 указанные клетки культивируют от 12 часов до 36 часов.
6. Способ по п.4, отличающийся тем, что указанный гибридный белок Rv2299c/ESAT-6 увеличивает продукцию по меньшей мере одного белка, выбранного из группы, состоящей из TNF-α, IL-12p70, IL-6 и IL-lβ, во время созревания незрелых дендритных клеток.
7. Способ по п.4, отличающийся тем, что созревание указанных незрелых дендритных клеток вызвано стимуляцией TLR (toll-подобного рецептора)-4 гибридным белком Rv2299c/ESAT-6.
8. Способ по п.4, отличающийся тем, что созревание незрелых дендритных клеток вызвано стимуляцией митоген-активируемой протеинкиназы (МАРК) и NF-κB.
9. Применение гибридного белка Rv2299c/ESAT-6 в качестве активного ингредиента для усиления врожденного и/или приобретённого иммунитета, при этом белок Rv2299c имеет аминокислотную последовательность SEQ ID NO: 1.
| KR 20130039061 А, 19.04.2013 | |||
| WANG X | |||
| ET AL., Early Secreted Antigenic Target of 6-kDa Protein of Mycobacterium tuberculosis Primes Dendritic Cells To Stimulate Th17 and Inhibit Th1 Immune Responses, THE JOURNAL OF IMMUNOLOGY, 2012, v | |||
| Питательный кран для вагонных резервуаров воздушных тормозов | 1921 |
|
SU189A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| ВВЕДЕНИЕ ДЕНДРИТНЫХ КЛЕТОК, ПОДВЕРГНУТЫХ ЧАСТИЧНОМУ СОЗРЕВАНИЮ IN VITRO, ДЛЯ ЛЕЧЕНИЯ ОПУХОЛЕЙ | 2003 |
|
RU2348418C2 |

