Область техники
Настоящее изобретение относится к антителам, в частности к гуманизированным антигенсвязывающим фрагментам (Fab) против бешенства, способу получения указанных антител с использованием дрожжей.
Предшествующий уровень техники
По данным Всемирной организации здравоохранения более 55 тысяч человек в мире ежегодно умирают от бешенства, более половины из них - дети до 15 лет (Всемирная Организация Здравоохранения, информационный бюллетень №99, 2008, декабрь). Переносчиками вируса бешенства являются зараженные животные, главным образом собаки. Только в Москве ежегодно фиксируется более 30 тысяч укусов собаками, при этом в большинстве случаев нельзя исключать инфицирование вирусом бешенства, поскольку досимптомной диагностики бешенства не существует. В случае развития симптомов болезнь становится практически неизлечимой и приводит к быстрой смерти (Hildegund С.J. 2009. Novel Vaccines to Human Rabies // Negl Trop Dis. 3 N9. P.515).
Основным способом борьбы с бешенством на сегодняшний день является иммунизация человека и домашних животных. Всемирная Организация Здравоохранения (ВОЗ) рекомендует поголовную профилактическую вакцинацию домашних животных (кошек и собак) и антирабическую вакцинацию людей в случае их контакта с животным, у которого подозревается бешенство. После контактов 3 категории, к которым относятся укусы, а также для людей с ослабленной иммунной системой ВОЗ рекомендует пассивную иммунизацию, то есть введение антирабического иммуноглобулина (Всемирная Организация Здравоохранения, информационный бюллетень №99, 2008, декабрь). В качестве такого иммуноглобулина применяют лошадиную сыворотку, содержащую антитела против вируса бешенства. Серьезные побочные эффекты вплоть до анафилактического шока, а также быстрый клиренс заметно ограничивают применение сывороток животного происхождения. Энзиматическое расщепление иммуноглобулинов лошадиной сыворотки до Fab'2-фрагментов лишь частично снижает иммунногенность, приводя к существенному удорожанию препарата (Fernandes A, Kaundinya JO, Daftary G, Saxena L, Banerjee S, et al. 2008. Chromatographic purification of equine immunoglobulin G F(ab)2 from plasma // J Chromatogr В Analyt Technol Biomed Life Sci. 876. 109-115). Человеческий иммуноглобулин от вакцинированных добровольцев не может удовлетворить всех потребностей в нем. Кроме того, препараты, полученные с использованием человеческого материала, несут в себе опасность ятрогенного инфицирования.
Источником человеческого иммуноглобулина, предохраняющего от развития симптомов бешенства, могут стать гуманизированные моноклональные антитела, способные нейтрализовать вирус. Ранее авторами настоящего изобретения была получена панель мышиных моноклональных антител к вакцинному штамму вируса бешенства (ВБ) Внуково-32 (Грибенча С.В., Василенко О.В., Фуралев В.А., Клюшник С.Ю., Кузьмицкая Т.М., Свешников П.Г., Баринский И.Ф. 1991. Получение и характеристика гибридом, продуцирующих моноклональные антитела к структурным белкам вируса бешенства, штамм Внуково-32 // Вопросы вирусологии. №4. 318-321). В результате детального изучения свойств антител (Грибенча С.В., Василенко О.В., Кузьмицкая Т.М., Фуралев В.А., Свешников П.Г., Татаров А.Г., Колотвина П.В., Баринский И.Ф. 1991. Взаимодействие моноклональных антител к структурным белкам вакцинного вируса Внуково-32 с другими вирусами группы бешенства // Вопросы вирусологии. №5. 399-402; Грибенча С. В., Василенко О.В., Фуралев В.А., Клюшник С.Ю., Кузьмицкая Т.М., Свешников П.Г., Баринский И.Ф. 1991. Получение и характеристика гибридом, продуцирующих моноклональные антитела к структурным белкам вируса бешенства, штамм Внуково-32 // Вопросы вирусологии. №4. 318-321) было отобрано антитело 1С5, обладающее следующими характеристиками: 1) Взаимодействует с гликопротеидом ВБ; 2) Взаимодействует со всеми исследованными штаммами ВБ. Всего было исследовано 33 штамма, среди которых все основные штаммы ВБ, встречающиеся на территории России, а также штаммы из Центральной Европы, Украины, США и Африки. 3) Обладает вируснейтрализующей активностью порядка 107 LD50 как по отношению к вакцинному штамму Внуково-32, так и по отношению к диким штаммам ВБ, тогда как аналогичный показатель для коммерческого лошадиного иммуноглобулина составил 6,6·102 LD50. 4) Обладает 100% лечебным действием: мыши, зараженные сверхлетальными дозами ВБ (8-10 LD50), выживали без развития симптомов бешенства, если в течение 2-48 часов после заражения им был введен препарат на основе антитела 1С5. Коммерческий лошадиный иммуноглобулин, «Антирабический иммуноглобулин из сыворотки лошади» (АИГ), пр-во ЗАО «БИОЛЕК», Украина, защищал только 16% зараженных животных.
Для снижения неблагоприятных реакций пациентов при использовании антител в терапии указанные антитела или их части должны быть гуманизированы для того, чтобы быть менее иммуногенными, чем их мышиные прототипы. Так, авторы настоящего изобретения на основе моноклональных мышиных антител, обладающих способностью с высокой специфичностью и высоким аффинитетом связываться с вирусом бешенства, и каркасных участков антител человека сконструировали гуманизированные антигенсвязывающие фрагменты (Fab) против вируса бешенства.
Краткое описание изобретения
Целью настоящего изобретения было получение реагента, в частности гуманизированного антигенсвязывающего участка (fragment antigen binding, Fab) против вируса бешенства.
Указанная цель была достигнута путем конструирования гуманизированного антигенсвязывающего фрагмента (Fab) против вируса бешенства на основе ранее изолированного авторами настоящего изобретения моноклонального мышиного антитела против вируса бешенства и демонстрации высокой специфичности указанного антитела в связывании с вирусом бешенства. Также указанная цель была достигнута путем создания способа получения указанных функциональных антител с использованием клеток дрожжей.
Таким образом, настоящее изобретение представляет новое гуманизированное антитело, Гуманизированный антигенсвязывающий фрагмент (Fab) против вируса бешенства, который содержит вариабельный участок тяжелой цепи (VH), соединенный с доменом CH1 человеческого иммуноглобулина IgG1, с последовательностью аминокислот SEQ ID NO: 2 и вариабельный участок легкой цепи (VL), соединенный с доменом Cæ человеческого иммуноглобулина каппа, с последовательностью аминокислот SEQ ID NO: 4.
Также настоящее изобретение представляет изолированный фрагмент ДНК, кодирующий антигенсвязывающий фрагмент (Fab) против вируса бешенства, описанный выше.
Также настоящее изобретение представляет клетку дрожжей, трансформированную фрагментом ДНК, описанным выше, и обладающую способностью к продукции антигенсвязывающего фрагмента (Fab) против вируса бешенства, описанного выше.
Также настоящее изобретение представляет клетку дрожжей, описанную выше, в которой указанными дрожжами являются дрожжи Pichia pastoris.
Также настоящее изобретение представляет способ получения антигенсвязывающего фрагмента (Fab) против вируса бешенства, описанного выше, включающий стадии выращивания клеток дрожжей, описанных выше, в питательной среде и выделения указанного Fab из культуральной жидкости.
Краткое описание чертежей
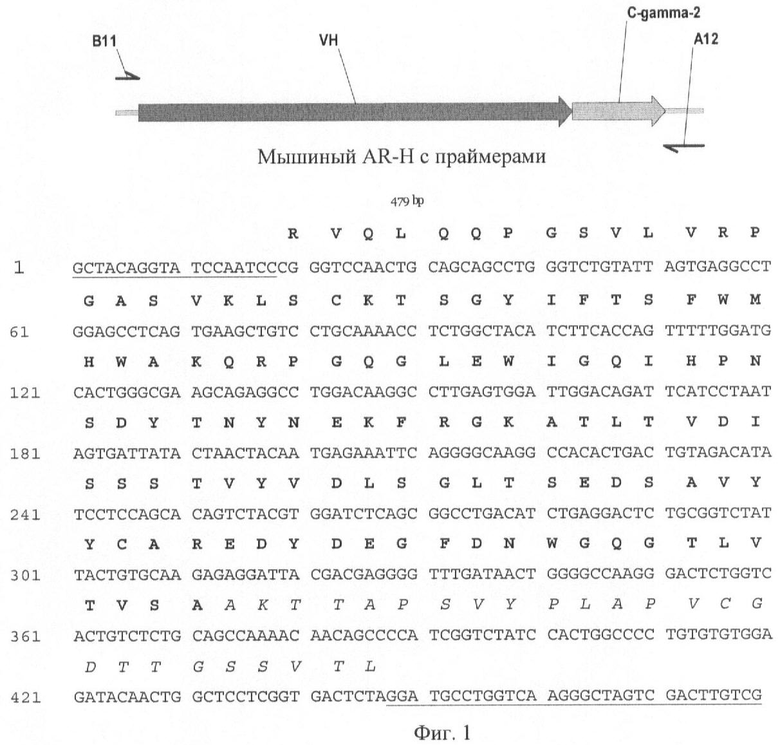
На Фиг.1 показана схема секвенирования, нуклеотидная последовательность клонированного гена тяжелой цепи и аминокислотная последовательность тяжелой цепи Fab-фрагмента мышиного антитела. Нуклеотидные последовательности праймеров, использованные для клонирования гена тяжелой цепи Fab-фрагмента антитела против вируса бешенства, подчеркнуты, аминокислотные последовательности вариабельных доменов тяжелой цепи (IgG2) выделены жирным шрифтом, аминокислотные последовательности константных областей тяжелой цепи (IgG2) мышиных антител выделены курсивом.
На Фиг.2 показана схема секвенирования, нуклеотидная последовательность клонированного гена легкой цепи и аминокислотная последовательность легкой цепи Fab-фрагмента мышиного антитела. Нуклеотидные последовательности праймеров, использованные для клонирования гена легкой цепи Fab-фрагмента антитела против вируса бешенства, подчеркнуты, аминокислотные последовательности вариабельных доменов легкой цепи (IgG2) выделены жирным шрифтом, аминокислотные последовательности константных областей легкой цепи (каппа-тип) мышиных антител выделены курсивом.
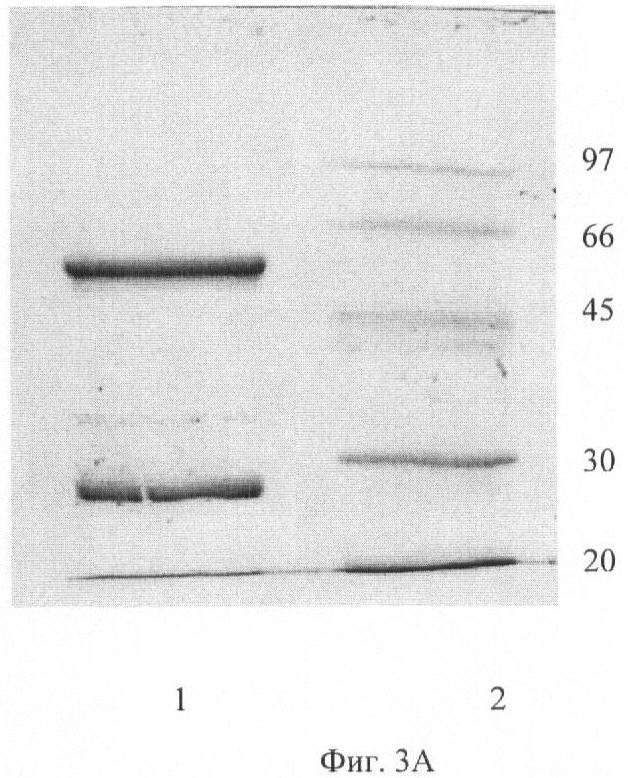
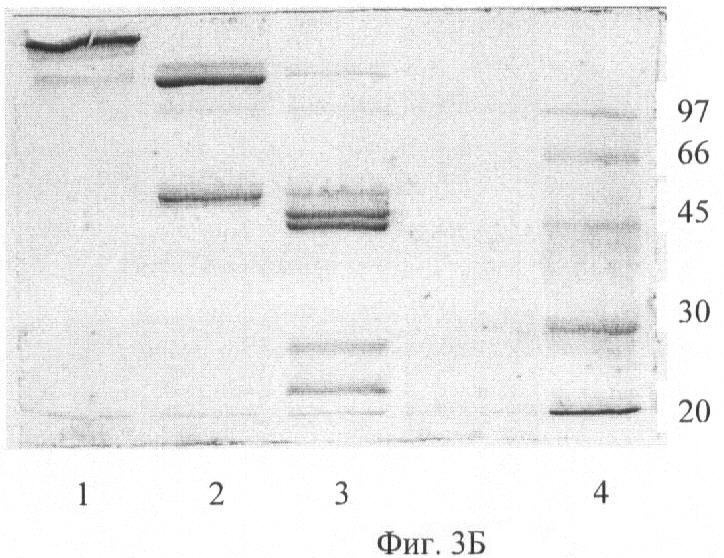
На Фиг.3А показан электрофорез МоАт из 1С5 в 12% SDS-ПААГ в присутстствии ДТТ. Дорожка 1 - МоАт из 1С5, дорожка 2 - маркеры молекулярной массы, кДа. На Фиг.3Б показан электрофорез МоАт и Fab-фрагментов из 1С5 10% SDS-ПААГ. Дорожка 1 - 1С5 IgG, дорожка 2 - 1С5 F(ab)'2, дорожка 3 - 1С5 Fab', дорожка 4 - Маркеры молекулярной массы, кДа.
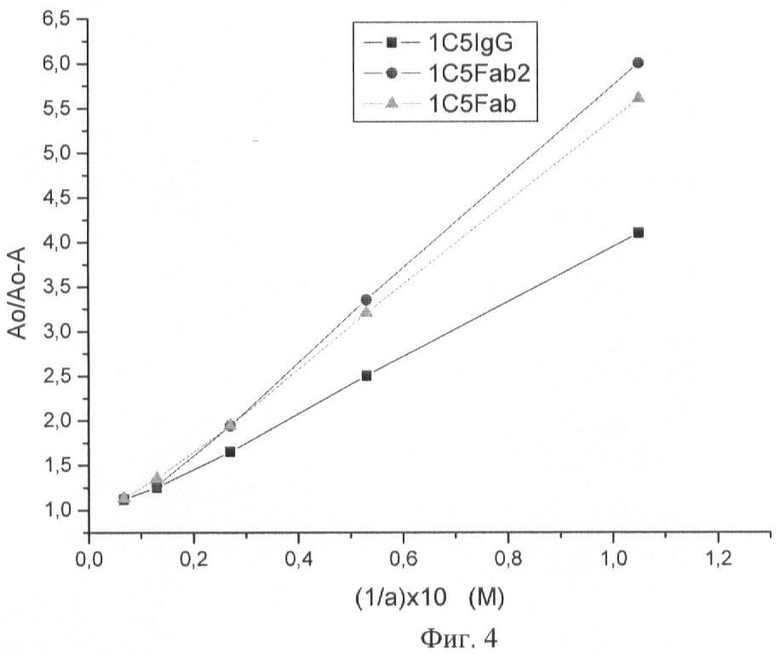
На Фиг.4 показаны кривые Клотца связывания вируса бешенства антителом 1С5 и фрагментами 1С5 Fab2 и 1С5 Fab.
На Фиг.5 показано сравнение мышиной, консенсусной и гуманизованной аминокислотных последовательностей вариабельного домена тяжелой цепи антитела против вируса бешенства.
На Фиг.6 показано сравнение мышиной, консенсусной и гуманизованной аминокислотных последовательностей вариабельного домена легкой цепи антитела против вируса бешенства.
На Фиг.7 показаны карты плазмид pPICZαA-AR-human-opt-H (А) и pPICZαA-ΔPmeI-AR-human-opt-L (В).
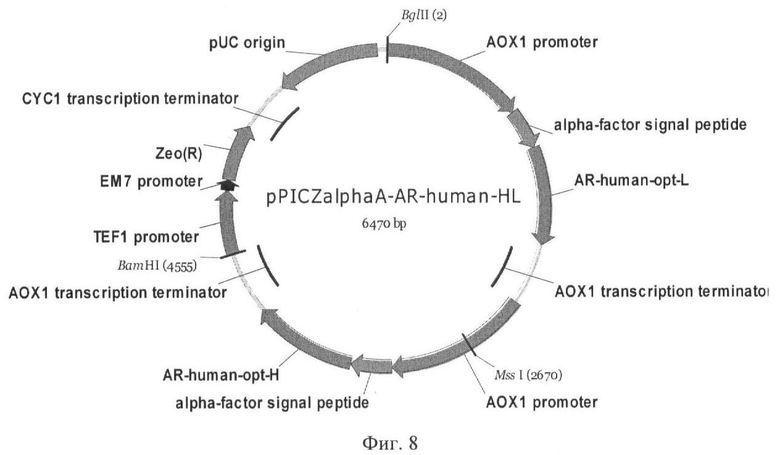
На Фиг.8 показана схематическая карта плазмиды pPICZαA-AR-human-opt-HL.
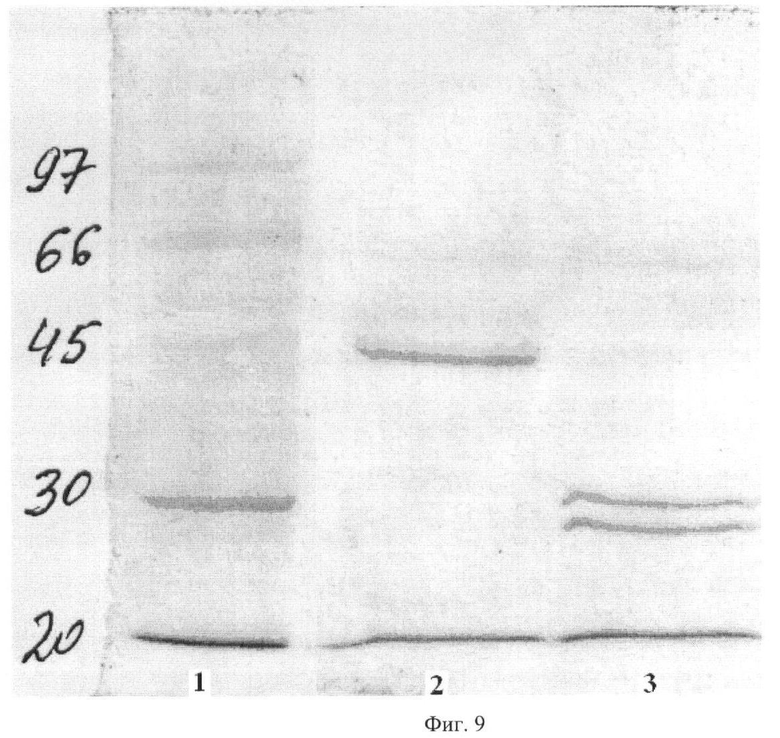
На Фиг.9 показан электрофорез рекомбинантного Fab фрагмента в 2% SDS-ПААГ с окрашиванием Coomassie. Дорожка 1 - маркеры молекулярной массы, кДа, дорожка 2 - рекомбинантный Fab фрагмент в невосстанавливающих условиях, дорожка 3 - рекомбинантный Fab фрагмент в восстанавливающих условиях.
На Фиг.10 показан Вестерн блот для определения специфичности и степени гуманизации полученных рекомбинантных Fab-фрагментов. ЭФ гликопротеида ВБ в невосстанавливающих (дорожки 1-3) или в восстанавливающих (дорожки 4-6) условиях. Мембраны после переноса инкубировали с гуманизированным Fab-фрагментом в концентрации 5 мкг/мл. Последующую инкубацию проводили либо с антителами против каппа-цепей иммуноглобулинов человека (дорожки 1 и 4), либо с антителами против гистидин-6 тага (дорожки 2 и 5), либо с антителами против каппа-цепей иммуноглобулинов мыши (дорожки 3 и 6).
Подробное описание настоящего изобретения
Антитела обычно состоят из двух тяжелых цепей, связанных между собой дисульфидными связями, и легких цепей, ассоциированных с N-концом каждой из тяжелых цепей. Каждая тяжелая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Каждая легкая цепь также содержит на N-конце вариабельный домен с константным доменом. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий участок. Вариабельные домены легкой и тяжелой цепей обладают похожей общей структурой, и каждый домен включает каркас из четырех участков, последовательности которых являются относительно консервативными, связанных посредством трех участков, определяющих комплементарность (complementarity determining regions, CDRs). Четыре каркасных участка формируют конформацию типа бета-складчатого слоя, а CDRs образуют петли, связывающие конструкцию из бета-складчатого слоя. Участки CDRs расположены в близком соседстве друг с другом благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDRs и каркасные участки антител могут быть определены путем ссылки на нумерационную систему Кабата (Kabat numbering system, Kabat et al., (1987) "Sequences of Proteins of Immunological Interest", US Dept. of Health and Human Services, US Government Printing Office) в сочетании с данными рентгеновской кристаллографии, как описано в патентной заявке РСТ WO 91/09967.
Антитела, полученные против определенных белков или мимотопов, могут обладать некоторыми преимуществами, поскольку такие антитела не сильно загрязнены антителами против других соединений, что могло бы, в противном случае, повлиять на точность диагностического метода. Способы получения таких антител известны из предшествующего уровня техники и подробно описаны Harlow и др. (Antibodies, a Laboratory Manual, Cold Spring Harbor Labs Press, 1988), и включают в себя иммунизацию животных с целью получения препаратов антител, которые выделяют из, например, плазмы или асцитной жидкости и очищают способами, известными из уровня техники, с получением препаратов, дающих реакцию с антигеном. Множество видов имеют белки с близкими последовательностями и поэтому могут возникнуть сложности в ходе использования стандартных протоколов иммунизации при получении антител, которые узнают белок только одного вида. Поэтому также стоит рассмотреть модификации стандартных методов получения антител, таких как, например, разностные (вычитательные) гибридизационные методики. Такие модификации могут быть известны специалисту в данной области техники, также они описаны в настоящем описании. В других методах антитела, которые могут быть использованы в рамках настоящего изобретения, получают с использованием рекомбинантных методов, описанных Sambrook и др. (Molecular Cloning: A Laboratory Manual, (Cold Spring Harbor Labs Press, 1989)).
Существуют также "дисплей" - технологии, при которых различные микроорганизмы, включая фаги, дрожжи, бактерии и вирусы, используются для представления репертуара одноцепочечных вариабельных фрагментов антител (scFvs), антигенсвязывающих фрагментов (Fabs) или доменных антител (Dabs) на их поверхности, откуда их можно обогащать и выделять путем повторяющихся циклов отбора (panning) (Carter, P.J. Potent antibody therapeutics by design. // Nat. Rev. Immunol. - 2006 - V.6 - P.343-357; Hoogenboom, H.R. Selecting and screening recombinant antibody libraries. // Nat. Biotechnol. - 2005 - V.23 - P.1105-1116).
Ранее авторы настоящего изобретения получили панель мышиных моноклональных антител к вакцинному штамму вируса бешенства (ВБ) Внуково-32 (Грибенча С.В., Василенко О.В., Фуралев В.А., Клюшник С.Ю., Кузьмицкая Т.М., Свешников П.Г., Баринский И.Ф. 1991. Получение и характеристика гибридом, продуцирующих моноклональные антитела к структурным белкам вируса бешенства, штамм Внуково-32 // Вопросы вирусологии. №4. 318-321). В результате детального изучения свойств антител (Грибенча С.В., Василенко О.В., Кузьмицкая Т.М., Фуралев В.А., Свешников П.Г., Татаров А.Г., Колотвина П.В., Баринский И.Ф. 1991. Взаимодействие моноклональных антител к структурным белкам вакцинного вируса Внуково-32 с другими вирусами группы бешенства // Вопросы вирусологии. №5. 399-402; Грибенча С.В., Василенко О.В., Фуралев В.А., Клюшник С.Ю., Кузьмицкая Т.М., Свешников П.Г., Баринский И.Ф. 1991. Получение и характеристика гибридом, продуцирующих моноклональные антитела к структурным белкам вируса бешенства, штамм Внуково-32 // Вопросы вирусологии. №4. 318-321) было отобрано антитело 1С5.
Указанное антитело 1С5 обладает следующими характеристиками: 1) Взаимодействует с гликопротеидом ВБ; 2) Взаимодействует со всеми исследованными штаммами ВБ. Всего было исследовано 33 штамма, среди которых все основные штаммы ВБ, встречающиеся на территории России, а также штаммы из Центральной Европы, Украины, США и Африки. 3) Обладает вируснейтрализующей активностью порядка 107 LD50 как по отношению к вакцинному штамму Внуково-32, так и по отношению к диким штаммам ВБ, тогда как аналогичный показатель для коммерческого лошадиного иммуноглобулина составил 6,6·102 LD50. 4) Обладает 100% лечебным действием: мыши, зараженные сверхлетальными дозами ВБ (8-10 LD50), выживали без развития симптомов бешенства, если в течение 2-48 часов после заражения им был введен препарат на основе антитела 1С5. Коммерческий лошадиный иммуноглобулин, «Антирабический иммуноглобулин из сыворотки лошади» (АИГ) (пр-во ЗАО «БИОЛЕК», Украина) защищал только 16% зараженных животных.
Затем авторы настоящего изобретения клонировали гены, кодирующие тяжелую и легкую цепи Fab-фрагмента мышиного антитела против вируса бешенства, и определили их последовательности (SEQ ID NO: 5 и 7 соответственно). Такое антитело содержит последовательность аминокислот SEQ ID NO: 6 в качестве вариабельного участка тяжелой цепи (VH) антитела, последовательность аминокислот SEQ ID NO: 8 в качестве вариабельного участка легкой цепи (VL) антитела и консервативные участки обеих цепей, необходимые для функционирования антитела. Полученные Fab-фрагменты моноклонального мышиного антитела продемонстрировали высокую специфичность и эффективность связывания антигена с вирусом бешенства.
Затем были осуществлены дизайн и гуманизация мышиного антитела против вируса бешенства. Сначала были определены границы каркасных участков согласно Kabat, E.A., Wu, Т.Т., Reid-Miller, M., Perry, H.M., and Gottesman, К.S. (Sequences of Proteins of Immunological Interest, (1987) U.S. Department of Health and Human Services, NIH, Bethesda, MD). Для каждого каркасного участка было найдено 50 ближайших гомологов в базах данных мышиных и человеческих антительных последовательностей из клеток зародышевой линии. Несколько систематических различий было найдено при сравнении мышиной и человеческих выборок. Аминокислоты в таких позициях рассматривались в качестве кандидатов на гуманизацию (Staelens S, Desmet J, Ngo TH, Vauterin S, Pareyn I, Barbeaux P, Van Rompaey I, Stassen JM, Deckmyn H, Vanhoorelbeke К Humanization by variable domain resurfacing and grafting on a human IgG4, using a new approach for determination of non-human like surface accessible framework residues based on homology modelling of variable domains. Mol Immunol. 2006 Mar; 43(8):1243-57).
При анализе последовательностей вариабельных доменов мышиного антитела против вируса бешенства были идентифицированы возможные ключевые аминокислотные остатки в определении связывания с антигеном. А именно, были определены аминокислотные остатки, которые являются частью канонических структур конформаций CDR, предложенных в (Chothia, С., Lesk A. M., Tramontano, A., Levitt, М., Smith-Gill, S.J., Air, G., Sheriff, S., Padlan, E. A., Davies, D., Tulip, W. R., Colman, P. M., Spinelli, S., Alzari, P.M., and Poljak, R. J. Conformations of immunoglobulin hypervariable regions. (1989) Nature. 342(6252):877-83. Review), остатки, которые потенциально участвуют в укладке VL-VH, как описано там же, а также были найдены аминокислоты, редко встречающиеся в последовательностях мышиных антител (Kolbinger, F., Saldanha, J., Hardman, N. and Bendig, M. "Humanization of a mouse anti-human IgE antibody: a potential therapeutic for IgE-mediated allergies" Prot. Engng. (1993) 6:971-980). При анализе пространственной модели Fv-фрагмента мышиного антитела против вируса бешенства были определены аминокислотные остатки, которые находятся в радиусе 5 ангстрем от CDR и наиболее вероятно связываются с антигеном.
Все вышеперечисленные позиции были исключены из числа кандидатов на гуманизацию. В итоге была получена консенсусная последовательность гуманизованного вариабельного домена антитела против вируса бешенства, состоящая из аминокислот, наиболее часто встречающихся в человеческих гомологах мышиного антитела против вируса бешенства, а также аминокислот мышиного антитела против вируса бешенства, определяющих связывание с антигеном.
Из полученных последовательностей были удалены любые потенциальные сайты гликозилирования. Кроме этого, были сделаны обратные замены на мышиные аминокислоты из-за опасности нарушить конформацию Fv-фрагмента (3 для тяжелой цепи и 1 для легкой). Таким образом, была получена последовательность гуманизованного вариабельного домена антитела против вируса бешенства (SEQ ID NO: 2 и 4).
Термин "CDR" или "участок, определяющий комплементарность" относится к тем частям тяжелой и легкой цепи антитела, которые расположены в непосредственной близости друг от друга в трехмерном пространстве и формируют связывающую поверхность антитела.
Так было выполнено настоящее изобретение.
В частности, антигенсвязывающим фрагментом (Fab) согласно настоящему изобретению является изолированный Fab, который селективно связывается с вирусом бешенства и который включает в себя вариабельный участок тяжелой цепи (VH), соединенный с доменом CH1 иммуноглобулина человека, с последовательностью аминокислот SEQ ID NO: 2 и вариабельный участок легкой цепи (VL), соединенный с доменом Сæ иммуноглобулина человека с последовательностью аминокислот SEQ ID NO: 4. Домен CH1 тяжелой цепи человеческого иммуноглобулина IgG, IgM, IgA, IgD или IgE может быть использован в качестве домена CH1 тяжелой цепи Fab согласно настоящему изобретению. Домен Сæ легкой цепи человеческого иммуноглобулина каппа или лямбда может быть использован в качестве домена Сæ легкой цепи Fab согласно настоящему изобретению. Такой Fab согласно настоящему изобретению представлен, но не ограничивается Fab, содержащим последовательности аминокислот SEQ ID NO: 2 (тяжелая цепь) и SEQ ID NO: 4 (легкая цепь).
Термин "гуманизированный Fab" используется для описания антитела, содержащего по крайней мере один, а предпочтительно два или три, участка CDR в одном или обоих вариабельных участках, полученных из антитела из первого вида животного, это понимается как то, что он может содержать определенный выбранный каркасный участок из аминокислот, соединенный с определенной гипервариабельной последовательностью аминокислот. Оставшиеся части Fab, полученные из иммуноглобулина Ig, получают из одного или нескольких различных антител. Вариабельные домены могут быть получены с использованием техники рекомбинантных ДНК или пептидным синтезом.
Гуманизированные Fab согласно настоящему изобретению содержат последовательности аминокислот, включающие в себя все или части CDR, полученные главным образом из моноклонального антитела, обладающего специфичностью к вирусу бешенства. В наиболее предпочтительном варианте осуществления изобретения таким моноклональным антителом является мышиное по происхождению антитело. Аминокислотные последовательности каркасных участков вариабельных доменов антитела или их части являются главным образом человеческими по происхождению в наиболее предпочтительном варианте осуществления изобретения и, следовательно, "гуманизированными Fab". Эта "гуманизация" считается полезной в снижении иммуногенности указанного антитела при терапевтическом назначении пациентам. Определенные выбранные остатки в каркасных участках остаются мышиными, а не человеческими. Считается, что это необходимо для достижения нужной трехмерной структуры молекулы, и таким образом для повышения связывающей специфичности и аффинности к вирусу бешенства.
Любая часть гуманизированных Fab (и следовательно, в широком смысле определения термина антитела), полученная в соответствии с настоящим описанием, не выходит за рамки настоящего изобретения при условии, что специфичность связывания и аффинность к вирусу бешенства сохраняется. Таким образом, связывающие белки, полученные из указанных антител, без сомнений в рамках настоящего изобретения, как и другие фрагменты, которые способствуют проявлению из способности, по крайней мере в степени необходимой для терапевтического использования, как описано ниже.
В настоящем изобретении фраза "Fab против вируса бешенства" означает молекулу, которая селективно связывается с вирусом бешенства и образует стабильный комплекс. Стабильным комплексом является комплекс, в котором связывание между партнерами происходит на период времени, достаточный для того, чтобы произвести детектирование указанного комплекса с использованием описанных здесь методов. Термин "селективно связывает вирус бешенства" означает способность указанной молекулы предпочтительно связываться с вирусом бешенства в отличие от связывания с белками, не имеющими отношения к вирусу бешенства, или связывания с небелковыми компонентами, присутствующими в образце. Fab, который предпочтительно связывается с вирусом бешенства, является Fab, который связывается с вирусом бешенства, но не связывается в существенной степени с другими молекулами или компонентами, которые могут присутствовать в образце. Существенное связывание предполагает, например, связывание антитела, связывающегося с вирусом бешенства, с молекулой, не имеющей отношения к вирусу бешенства, с аффиностью или силой, достаточной для того, чтобы помешать способности антитела, связывающегося с вирусом бешенства, определить уровень вируса бешенства в образце. Примерами таких молекул и компонентов, которые могут присутствовать в образце, являются, но не ограничиваются ими, белки, не имеющие отношение к вирусу бешенства, липиды и углеводы.
Способность Fab к связыванию с антигеном может быть определена специалистом в данной области с использованием методов, включающих, но не ограничивающихся методом ELISA и равновесным диализом. Методы определения аффинности и силы связывания хорошо известны специалисту в данной области техники, подробно описаны Janeway и др. (Immunobiology: The Immune System in Health and Disease (Garland Publishing Company, 1996)).
Fab, пригодный для осуществления настоящего изобретения, - это Fab, обладающий способностью к связыванию с вирусом бешенства, когда его концентрация составляет от около 10 нг/мл и около 1 г/мл. В частности, пригодные в рамках настоящего изобретения антитела связывают вирус бешенства, когда его концентрация составляет 10 нг/мл, около 1 нг/мл или менее, предпочтительно 100 пг/мл. Такой Fab описан в сопутствующих Примерах.
Изолированным фрагментом ДНК, кодирующим Fab согласно настоящему изобретению, является экспрессирующийся фрагмент ДНК, содержащий промотор, сигнальную последовательность, последовательность нуклеотидов, кодирующую структурные части тяжелой и легкой цепей Fab, участок терминации транскрипции.
В частности, нуклеотидные последовательности, кодирующие структурные части тяжелой и легкой цепей Fab согласно настоящему изобретению, содержат фрагмент ДНК, кодирующий вариабельный участок тяжелой цепи (VH) (SEQ ID NO: 1) и вариабельный участок легкой цепи (VL) (SEQ ID NO: 3), связанный с фрагментом ДНК, кодирующим консервативные участки обеих цепей, необходимые для функционирования указанного Fab. Фрагмент ДНК, кодирующий Fab согласно настоящему изобретению, может быть получен любым методом, известным специалисту в данной области техники, включая ПЦР с использованием набора перекрывающихся праймеров, синтез по технологии Slonogene™ (Sloning Biotechnology GmbH), химическим способом и т.д.
Ввиду вырожденности трансляционного кода могут быть различия в последовательности ДНК. Фрагменты ДНК согласно настоящему изобретению не ограничены фрагментами, показанными в SEQ ID NO: 1 или 3, при условии, что они кодируют участки цепей Fab с аминокислотной последовательностью, показанной в SEQ ID NO: 2 или 4.
Клетками дрожжей согласно настоящему изобретению являются клетки дрожжей, трансформированные фрагментом ДНК, описанным выше, и обладающие способностью к продукции Fab против вируса бешенства.
Фраза «клетки дрожжей, трансформированные фрагментом ДНК» означают, что желаемый фрагмент ДНК был введен в клетку дрожжей с использованием методов, известных специалисту в данной области техники. Трансформация клетки дрожжей фрагментом ДНК приводит к увеличению экспрессии фрагмента ДНК, кодирующего антитело согласно настоящему изобретению. Присутствие сигнальной последовательности α-фактора приводит к секреции произведенного антитела в культуральную жидкость. Методы трансформации клеток дрожжей включают в себя все известные методы, например модифицированная версия процедуры, описанной для S. cerevisiae (Gietz and Schiestl, 1996).
Способом согласно настоящему изобретению является способ получения Fab против вируса бешенства, включающий выращивание дрожжей в питательной среде и выделение полученного Fab из культуральной жидкости.
Примером клетки дрожжей, пригодных для продукции Fab согласно настоящему изобретению, являются, но не ограничивается ими, клетки дрожжей Pichia pastoris. Фраза "дрожжи Pichia pastoris" означает, что указанные дрожжи классифицируют как Pichia pastoris (Р. pastoris) в соответствии с классификацией, известной специалисту в данной области микробиологии. Примерами дрожжей Р. pastoris, применимых в рамках настоящего изобретения, являются, но не ограничиваются ими, дрожжи Р.pastoris GS115 (Invitrogen).
В настоящем изобретении выращивание, накопление и очистка Fab из культуральной жидкости и других жидкостей может быть осуществлена методом, сходным с традиционными методами ферментации, когда некий белок производится с использованием микроорганизма.
Питательная среда для выращивания может быть как синтетической, так и натуральной при условии, что указанная среда содержит источник углерода, источник азота, минералы и, если это необходимо, подходящее количество питательных веществ, в которых нуждаются дрожжи для их роста. К источникам углерода относятся углеводы, такие как глюкоза и сахароза, и различные органические кислоты. В зависимости от способа ассимиляции у указанного микроорганизма может быть использован спирт, включая метанол, этанол, глицерин. В качестве источника азота могут быть использованы различные соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, источники природного азота, такие как пептон, гидролизат соевых бобов расщепленные ферментированные микроорганизмы. В качестве источника минералов могут быть использованы монофосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут быть использованы тиамин, дрожжевой экстракт и подобные им соединения.
Выращивание предпочтительно осуществляют в аэробных условиях, таких как культивирование с перемешиванием, взбалтывание с аэрацией, при температуре от 20 до 40°С, предпочтительно от 28 до 30°С. рН среды обычно поддерживают в диапазоне от 2 до 9, предпочтительно в диапазоне от 6 до 7.5. рН среды может быть скорректирован с помощью аммиака, карбоната кальция, различных оснований и буферов. Обычно выращивание в течение от 1 до 7 дней приводит к накоплению Fab в культуральной жидкости.
После выращивания твердые компоненты, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрации с использованием мембраны, а затем антитела или Fab могут быть выделены и очищены методом осаждения с солями, с использованием сульфата натрия или сульфата аммония, аффинной хроматографии, ионообменной хроматографии и т.п.
Для терапии, применяемой к людям, необходимо использовать человеческие изотипы для того, чтобы минимизировать антиглобулинный ответ в течение этой терапии. Последовательности ДНК человеческих константных доменов, предпочтительно в сочетании с каркасными участками вариабельных доменов, могут быть получены в соответствии с хорошо известными методиками. Примером такой методики является методика CAMPATH 1H, доступная у компании Burroughs Wellcome Ltd.
Антитело, обладающее способностью к связыванию вируса бешенства, использованное в настоящем изобретении, может содержаться в составе медицинской рецептуры. Например, антитело может быть объединено с буфером, в котором указанное антитело растворено, и/или с неким носителем. Буферы и носители, пригодные для этого, известны специалистам в данной области техники. Примерами таких буферов являются буферы, в которых указанное антитело может функционировать, селективно связывая вирус бешенства, такие как, но не ограничивающиеся солевым фосфатным буфером, водой, салином, фосфатным буфером, буфером HEPES (солевой буфер N-2-гидроксиэтилпиперазин-N'-2-этансльноновой кислоты), буфером TES (солевым буфером Трис-ЭДТА), буфером Трис и буфером ТАЕ (Трис-ацетат-ЭДТА). Примерами носителей являются, но не ограничены полимерными матрицами, токсоидами, сывороточными альбуминами, такие как бычий сывороточный альбумин. Носители могут быть комбинированы с антителами и конъюгированными (то есть, присоединенными) антителами так, что они несущественно влияют на способность антитела к селективному связыванию вируса бешенства.
Методы получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации, подбора олигонуклеотидов в качестве праймеров и подобные методы могут быть стандартными методами, хорошо известными специалисту в данной области техники. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т. ("Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Последующие примеры приведены для целей объяснения и не ограничивают каким-либо образом рамки настоящего изобретения.
Пример 1. Клонирование генов, кодирующих Fab-фрагменты тяжелых и легких цепей мышиных моноклональных антител против вируса бешенства.
1. Выделение РНК.
В качестве объекта исследования использовали мышиные моноклональные антитела (гибридома 1С5), полученные ранее. Указанная гибридома является генетически стабильной и моноклональной. Гибридома 1С5 индуцирует образование асцита при внутрибрюшинном введении мышам линии Balb/c с содержанием МоАт не менее 5 мг/мл. Одноступенчатая аффинная хроматография асцитной жидкости на белок G-Сефарозе приводила к получению препарата МоАт с чистотой более 95%. Полученные МоАт узнают антиген - гликопротеид вируса бешенства - с высоким аффинитетом, так как конечная точка титрования соответствует концентрации МоАт 1 нг/мл и менее. Также было однозначно показано, что в результате пептического гидролиза получаются функционально активные Fab'-фрагменты МоАт 1С5, обладающие высоким аффинитетом, сравнимым с аффинитетом исходных IgG. (Грибенча С.В., Василенко О.В., Фуралев В.А., Клюшник С.Ю., Кузьмицкая Т.М., Свешников П.Г., Баринский И.Ф. Получение и характеристика гибридом, продуцирующих моноклональные антитела к структурным белкам вируса бешенства, штамм Внуково-32. // Вопр. вирусол. - 1991 - №4 - С.318-321; Грибенча С.В., Василенко О.В., Кузьмицкая Т.М., Фуралев В.А., Свешников П.Г., Татаров А.Г., Колотвина П.В., Баринский И.Ф. Взаимодействие моноклональных антител к структурным белкам вакцинного вируса Внуково-32 с другими вирусами группы бешенства. // Вопр. вирусол. - 1991 - №5 - С.399-402.)
Суммарную РНК выделяли из 5×106 клеток гибридомы 1С5 по стандартной методике. Выход по РНК составлял 0.05 мкг РНК на мкл гибридомы 1С5. В результате проведенных экспериментов наработали 2,5 мкг суммарной РНК, которую затем использовали для получения кДНК.
2. Синтез кДНК.
кДНК получали из выделенной РНК с использованием набора для синтеза RevertAid™ H Minus First Strand cDNA Synthesis Kit (Fermentas®) по "протоколу синтеза кДНК, пригодной для ПЦР".
3. Выбор праймеров для клонирования гена тяжелой цепи Fab-фрагмента.
Для определения пары праймеров, необходимых для клонирования гена тяжелой цепи Fab-фрагмента, были проведены ПЦР с Tag DNA Polymerase. кДНК использована как матрица. В качестве прямого праймера был выбран комплекс из 16 вырожденных праймеров, узнающих более 95% последовательностей сигнальных пептидов тяжелой цепи, представленных в базе данных Kabat (Kabat E.A., Wu T.T., Perry H., Gottesman K. and Foeller, C. Sequences of Proteins of Immunological Interest // Fifth Edition. NIH Publication. 1991. P 3242). Электрофорез продуктов ПЦР в агарозном геле показал, что использование пары праймеров B11 (SEQ ID NO: 9) и А12 (SEQ ID NO: 10) приводит к образованию ПЦР-фрагмента длиной ≈ 450 п.н., размер которого совпадает с ожидаемым результатом на основании анализа последовательностей кДНК генов тяжелых цепей иммуноглобулинов IgG Mus musculus из Gene-Bank и IMGT-GENE-DB.
4. Выбор праймеров для клонирования гена легкой цепи Fab-фрагмента.
Так как изначально было известно, что легкие цепи относятся к каппа-типу, то в качестве прямого праймера был использован комплекс из 14 вырожденных праймеров, узнающих более 95% последовательностей сигнальных пептидов каппа цепи, представленных в базе данных Kabat (см. выше). Электрофорез продуктов ПЦР в агарозном геле показал, что использование праймера A15 (SEQ ID NO: 11) в паре с обратным праймером L-m-r (SEQ ID NO: 12) приводит к образованию ПЦР-фрагмента длиной ≈700 п.н., размер которого совпадает с ожидаемым результатом на основании анализа последовательностей кДНК генов тяжелых цепей иммуноглобулинов IgG Mus musculus из Gene-Bank и IMGT-GENE-DB.
5. Клонирование генов Fab-фрагмента антитела против вируса бешенства.
Для уменьшения вероятности ошибок при клонировании генов Fab-фрагмента использовали Pfu ДНК полимеразу, которая делает в 12 раз меньше ошибок, чем Tag ДНК полимераза. ПЦР-фрагменты из независимых ПЦР (по три независимых фрагмента для каждой цепи) были элюированы из агарозного геля, фосфорилированы с использованием Т4 ДНК полинуклеотидкиназы и клонированы в плазмидный вектор pUC18, расщепленный по сайту рестрикции SmaI. Отбирали рекомбинантные плазмиды с генами легкой и тяжелой цепей Fab-фрагмента антитела против вируса бешенства, названные соответственно pUC18-AR-L и pUC18-AR-H.
Пример 2. Секвенирование вариабельных частей Н- и L-цепей мышиного антитела 1С5.
Рекомбинантные плазмиды с генами цепей Fab-фрагмента антитела против вируса бешенства (три плазмиды, содержащие независимые ПЦР-фрагменты тяжелой цепи (pUC18-AR-H1-3, pUC18-AR-H2-3, pUC18-AR-H3-3), и три плазмиды, содержащие независимые ПЦР-фрагменты легкой цепи (pUC18-AR-L1-4, pUC18-AR-L2-4, pUC18-AR-L3-4)) были отобраны и клонированные фрагменты были секвенированы с обоих концов каждый. В качестве праймеров для секвенирования были использованы стандартные праймеры SEQ ID NO: 13 и 14.
Нуклеотидные последовательности тяжелой (Н) и легкой (L) цепей Fab-фрагмента мышиного антитела 1С5 против вируса бешенства приведены в Перечне последовательностей под номерами SEQ ID NO: 5 и 7 соответственно и представлены на Фиг.1 и 2. Аминокислотная последовательность была определена путем трансляции нуклеотидной последовательности клонированных генов, кодирующих последовательности тяжелой (Н) и легкой (L) цепей Fab-фрагмента мышиного антитела 1С5 против вируса бешенства в аминокислотную. Аминокислотные последовательности тяжелой (Н) и легкой (L) цепей Fab-фрагмента мышиного антитела 1С5 против вируса бешенства приведены в Перечне последовательностей под номерами SEQ ID NO: 6 и 8 соответственно.
Пример 3. Очистка и характеристика моноклональных антител 1С5 из асцитной жидкости и получение и характеристика функциональных Fab-фрагментов против гликопротеидов вируса бешенства.
Очистка моноклональных антител
МоАт выделяли из асцитной жидкости, используя аффинную хроматографию на белок G-сефарозе, и переводили диализом в фосфатно-солевой буфер (ФСБ). Концентрацию определяли на спектрофотометре при длине волны 280 нм. Коэффициент экстинции (Е) для 1 мг/мл иммуноглобулинов равен 1.4. Выход МоАт составил 5 мг/мл асцита. Суб-изотип МоАт - IgG2a.
Степень чистоты препаратов МоАт контролировали с помощью электрофореза в 12% полиакриламидном геле в присутствии SDS и ДТТ в ступенчатой буферной системе Лэмлли, используя Mini PROTEAN 3 Electrophoresis System (BIO-RAD). Одноступенчатая аффинная хроматография асцитной жидкости на белок G-Сефарозе приводит к получению препарата МоАт с чистотой более 95% (Фиг.3).
Характеристика моноклональных антител в иммуноферментном анализе с разделением компонентов (непрямой ИФА)
Сорбцию антигена проводили при температуре 4°C в течение ночи. Антиген сорбировали в концентрации 2,5 мкг/мл в ФСБ на 96-луночные планшеты с высокой связывающей способностью производства Corning-Costar (Нидерланды). Результаты приведены в таблице 1.
Как видно из приведенных данных, МоАт узнают антиген с высоким аффинитетом, так как конечная точка титрования соответствует концентрации МоАт 1 нг/мл и менее.
Получение функционально активных Fab
Изотипы тяжелой и легкой цепей МоАт 1С5 определяли методом непрямого ИФА с использованием типирующих сывороток из набора ISO-2 (Sigma, USA). Установлено, что изотип тяжелой цепи - IgG2a, изотип легкой цепи - κ.
Последующий протеолиз проводили следующим образом: МоАт диализовали против 0,2М ацетата натрия рН 4,7 и к полученному препарату добавляли пепсин, предварительно растворенный в том же буфере (10 мг/мл) в соотношении 1:33. Протеолиз проводили при 37°С в течение 48 часов. Реакцию останавливали, повышая рН до 7.5, 1М трисом. Известно, что пепсин отщепляет от IgG фрагмент Fab'2, что позволило ожидать образования аналогичного продукта в данном эксперименте. Полученные Fab'2 диализовали против 0,55М трис-HCl буфера, рН 7,8. К отдиализованному препарату добавляли 2-меркаптоэтиламин до 10 мМ концентрации и инкубировали в течение 1 часа при 37°С для восстановления S-S связей. Для остановки реакции рН понижали до 6,5 и добавляли 10 мМ N-этилмалеимида. Температуру понижают до 0°С. Полученные Fab диализом переводили в ФСБ.
Чистоту полученных препаратов контролировали при помощи электрофореза в 10% полиакриламидном геле в присутствии SDS в невосстановленных условиях, в ступенчатой буферной системе Лэмлли, используя Mini PROTEAN 3 Electrophoresis System (BIO-RAD, Catalog N 165-3301) (Фиг.3Б).
Характеристика МоАт из 1С5, F(ab)'2 и Fab'-фрагментов в иммуноферментном анализе с разделением компонентов (непрямой ИФА)
Сорбцию антигена проводили при температуре 4°С в течение ночи. Гликопротеид вируса бешенства сорбировали в концентрации 2,5 мкг/мл в ФБР на 96-луночные планшеты с высокой связывающей способностью производства Corning-Costar (Нидерланды). Результаты приведены в таблице 2.
Представленные результаты однозначно свидетельствуют о том, что в результате пептического гидролиза получены функционально активные Fab'-фрагменты МоАт 1С5, обладающие высоким аффинитетом, сравнимым с аффинитетом исходных IgG.
Определение константы диссоциации (Kd)
В качестве антигена использовали вирус бешенства. Константу диссоциации (Kd) измеряли по методике, описанной в статье: Friguet В и др. (Measurements of the True Affinity Constant in Solution of Antigen-Antiojdy Complex by Enzyme-Linked Immunosorbent Assay. Journal of Immunological Methods, 77 (1985) 305-319).
Константу диссоциации (Kd) рассчитывали по уравнению Клотца:
Ao/Ao-A=1+1/a Kd,
где Ao - оптическая плотность, измеренная для антител в отсутствии антигена, А - оптическая плотность, измеренная для свободных антител в смеси антиген-антитело, а - концентрация антигена.
Константа диссоциации Kd полноразмерного мышиного антитела 1С5 IgG составила 1,92×10-9 М, Kd фрагмента 1С5 Fab'2 составила 2,1×10-9 М, Kd фрагмента 1С5 Fab' составила 2,7×10-9 М.
Пример 4. Дизайн и гуманизация мышиного антитела 1С5 против вируса бешенства.
Дизайн и гуманизацию мышиного антитела против вируса бешенства проводили следующим образом.
Сначала определяли границы каркасных участков согласно Kabat, E.A., Wu, Т.Т., Reid-Miller, M., Perry, H.M., and Gottesman, K. S. (Sequences of Proteins of Immunological Interest, (1987) U.S. Department of Health and Human Services, NIH, Bethesda, MD). Для каждого каркасного участка было найдено 50 ближайших гомологов в базах данных мышиных и человеческих антительных последовательностей из клеток зародышевой линии. Несколько систематических различий было найдено при сравнении мышиной и человеческих выборок. Аминокислоты в таких позициях рассматривались в качестве кандидатов на гуманизацию (Staelens S, Desmet J, Ngo TH, Vauterin S, Pareyn I, Barbeaux P, Van Rompaey I, Stassen JM, Deckmyn H, Vanhoorelbeke K Humanization by variable domain resurfacing and grafting on a human IgG4, using a new approach for determination of non-human like surface accessible framework residues based on homology modelling of variable domains. Mol Immunol. 2006 Mar; 43(8):1243-57).
При анализе последовательностей вариабельных доменов мышиного антитела против вируса бешенства идентифицировали возможные ключевые аминокислотные остатки в определении связывания с антигеном. А именно, были определены аминокислотные остатки, которые являются частью канонических структур конформаций CDR, предложенных в (Chothia, С., Lesk A. M., Tramontano, A., Levitt, M., Smith-Gill, S.J., Air, G., Sheriff, S., Padlan, E.A., Davies, D., Tulip, W.R., Colman, P.M., Spinelli, S., Alzari, P.M., and Poljak, R.J. Conformations of immunoglobulin hypervariable regions. (1989) Nature. 342(6252): 877-83. Review.), остатки, которые потенциально участвуют в укладке VL-VH, как описано там же, а также были найдены аминокислоты, редко встречающиеся в последовательностях мышиных антител (Kolbinger, F., Saldanha, J., Hardman, N. and Bendig, M. "Humanization of a mouse anti-human IgE antibody: a potential therapeutic for IgE-mediated allergies" Prot. Engng. (1993) 6:971-980). При анализе пространственной модели Fv-фрагмента мышиного антитела против вируса бешенства были определены аминокислотные остатки, которые находятся в радиусе 5 ангстрем от CDR и наиболее вероятно связываются с антигеном.
Все вышеперечисленные позиции исключали из числа кандидатов на гуманизацию. В итоге получили консенсусную последовательность гуманизованного вариабельного домена антитела против вируса бешенства, состоящую из аминокислот, наиболее часто встречающихся в человеческих гомологах мышиного антитела против вируса бешенства, а также аминокислот мышиного антитела против вируса бешенства, определяющих связывание с антигеном.
Из полученных последовательностей удалили любые потенциальные сайты гликозилирования. Кроме этого, сделали обратные замены на мышиные аминокислоты из-за опасности нарушения конформаций Fv-фрагмента (3 для тяжелой цепи и 1 - для легкой). Таким образом, получили последовательность гуманизованного вариабельного домена антитела против вируса бешенства (Фиг.4 и 5).
Пример 5. Конструирование и синтез гибридных генов тяжелой и легкой цепей, кодирующих фрагмент Fab против вируса бешенства, состоящий из гуманизированных вариабельных областей и константных областей IgG человека.
Аминокислотную последовательность цепей гуманизованного Fab-фрагмента получали путем слияния аминокислотной последовательности гуманизованных VH и VL с аминокислотной последовательностью человеческих константных доменов IgG1 CH1 и Сæ соответственно (Tan P, Mitchell DA, Buss TN, Holmes MA, Anasetti C, Foote J. "Superhumanized" antibodies: reduction of immunogenic potential by complementarity-determining region grafting with human germline sequences: application to an anti-CD28. J Immunol. 2002 Jul 15; 169(2):1119-25). Нуклеотидную последовательность цепей гуманизованного Fab-фрагмента составляли из наиболее часто встречающихся в Pichia pastoris кодонов (SEQ ID NO: 2 и 4).
Гены легкой и тяжелой цепей Fab-фрагмента гуманизованного антитела против вируса бешенства синтезировали из набора перекрывающихся олигонуклеотидов с помощью ПЦР.
ДНК-фрагменты, кодирующие тяжелые и легкие цепи Fab-фрагмента гуманизованного антитела против вируса бешенства, были клонированы по сайтам XbaI и XhoI в векторы pBluescript II SK ΔApaI и pBluescript II SK соответственно. После секвенирования были отобраны плазмиды pBluescript II SK ΔApaI-AR-human-opt-H и pBluescript II SK-AR-human-opt-L с генами тяжелой и легкой цепей Fab-фрагмента гуманизованного антитела против вируса бешенства соответственно.
Пример 6. Конструирование экспрессионной системы для получения фрагмента Fab гуманизированного антитела против вируса бешенства и получение указанного фрагмента Fab с использованием дрожжей.
Фрагмент ДНК (SEQ ID NO: 1), содержащий ген, кодирующий фрагмент VH Fab против вируса бешенства, соединенный с доменом CH1 иммуноглобулина человека IgG1, содержащий сайты рестрикции XhoI и XbaI на концах, клонировали из плазмиды pBluescript II SK ΔApaI-AR-human-opt-H в вектор pPICZalpha A (Invitrogen). Для клонирования использовались сайты XhoI и XbaI. В результате была получена плазмида pPICZalphaA-AR-human-opt-H (Фиг.7А).
Фрагмент ДНК (SEQ ID NO: 3), содержащий ген, кодирующий фрагмент VL Fab против вируса бешенства, соединенный с доменом Сæ иммуноглобулина человека каппа, содержащий сайты рестрикции XhoI и XbaI на концах, клонировали из плазмиды pBluescript II SK-AR-human-opt-L в модифицированную плазмиду pPICZalpha А-ΔPmeI. Для клонирования использовались сайты XhoI и XbaI. В результате была получена плазмида pPICZalpha A-ΔPmeI-AR-human-opt-L (Фиг.7Б).
Для получения плазмиды pPICZalpha A-ΔPmeI плазмиду pPICZalpha A модифицировали путем замены сайта рестрикции PmeI на сайт рестрикции KspAI с помощью ПЦР. В качестве праймеров использовали синтетические олигонуклеотиды SEQ ID NO: 15-18. Матрицей послужила плазмида pPICZalpha А. Полученный ПЦР-фрагмент обрабатывали рестриктазами SacI и Mph1103I и клонировали в плазмиду pPICZalpha А, обработанную теми же рестриктазами. После секвенирования была получена плазмида pPICZalpha А-ΔPmeI.
Сборку обоих генов, кодирующих цепи фрагмента Fab, на одной плазмиде осуществляли лигированием фрагмента BglII-BamHI из вектора pPICZalpha A-ΔPmeI-AR-human-opt-L, содержащего легкую цепь, в сайт рестрикции BglII вектора pPICZalphaA-AR-human-opt-H, содержащего тяжелую цепь. В результате была получена плазмида pPICZalphaA-AR-human-opt-HL (Фиг.8), кодирующая обе цепи Fab гуманизированного антитела против вируса бешенства, каждый из которых был непосредственно соединен в одной рамке считывания с сигнальной последовательностью α-фактора из Saccharomyces cerevisiae под контролем промотора АОХ1.
Пример 7. Получение трансформантов дрожжей и анализ продукции фрагмента Fab.
Экспрессионную плазмиду pPICZalphaA-AR-human-opt-HL, линеаризованную с помощью рестриктазы PmeI, вводили в клетки GS115 в соответствии с протоколом производителя INVITROGEN с помощью методики, использующей LiCl. Трансформанты отбирали на питательной среде YPDS (Invitrogen), содержащей глюкозу и Zeocin (100 мкг/мл). Интеграцию линеаризованной плазмиды в хромосому подтверждали методом ПЦР с использованием ген-специфичных праймеров SEQ ID NO: 19 и 20 для тяжелой цепи, SEQ ID NO: 21 и 22 для легкой цепи и геномной ДНК трансформантов в качестве матрицы.
Отобранные трансформанты GS115/AR-human-Fab выращивали на полной среде с глицерином (BMGY), переносили на полную среду с метанолом (BMMY) для индукции экспрессии генов Fab-фрагмента гуманизованного антитела против вируса бешенства и выращивали в течение 45 часов при 30°С и рН 6.0 (конечная оптическая плотность - OD 40). Культуральные жидкости трансформантов анализировали с помощью ELISA. По результатам анализа был отобран штамм GS115/AR-human-Fab 7-22-1 с максимальной продукцией Fab-фрагментов гуманизованного антитела против вируса бешенства, составившей 21 мг/л.
Пример 8. Характеристика аффинности фрагмента Fab гуманизированного антитела против вируса бешенства, произведенного дрожжами.
Степень чистоты рекомбинантного человеческого Fab контролировали при помощи электрофореза в 10% полиакриламидном геле в присутствии SDS, в невостановленных и восстановленных условиях в ступенчатой буферной системе Лэмлли (Laemmli U.K. 1970. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4 //Nature. 227. 680-685). Степень чистоты полученного Fab составила не менее 95%.
Fab-фрагменты при окрашивании SDS-ПААГ геля с помощью Coomassie дают одиночную полосу с мобильностью, соответствующей предполагаемой массе Fab-фрагмента 50 кДа. Электрофорез в восстанавливающих условиях дает полосы, соответствующие предполагаемым массам свободных тяжелых и легких цепей (Фиг.9).
Специфичность Fab определяли методом вестерн-блот (Towbin H, Staehelin Т, Gordon J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications // Proc Natl Acad Sci USA. 76. N9. 4350-4354), используя гликопротеид вируса бешенства. Полученный Fab-фрагмент связывает гликопротеид вируса бешенства как в восстанавливающих, так и в невосстанавливающих условиях (Фиг.10). На высокую степень гуманизации указывает факт, что поликлональные антитела, специфичные к каппа-цепям иммуноглобулинов человека, узнают гуманизированный Fab-фрагмент, тогда как антитела против каппа-цепей иммуноглобулинов мыши гуманизированный Fab-фрагмент практически не связывают.
Определение констант диссоциации Fab.
Определение констант связывания антител проводили, как описано в методике (Klotz, 1953) с модификациями (Friguet et al., 1985). Было установлено, что константа диссоциации Kd рекомбинантного гуманизированного Fab-фрагмента 1С5 IgG составляет 1,15×10-9 М. Эти результаты указывают на то, что гуманизированные Fab-фрагменты обладают аффинностью, не меньшей, чем у исходного полноразмерного антитела 1С5.
Анализ ELISA для фрагмента Fab в культуральной жидкости:
- Сорбция антитела 2А11, разбавленного 1/1000 в 20 мМ Na2HPO4 (pH 7.2), 100 мкл на лунку, 2 часа при комнатной температуре. Антитела 2А11 связывают константные домены фрагмента Fab человеческого IgG1.
- Промывка лунок три раза буфером PBSt (0.1% Tween).
- Блокировка 0.5% БСА в PBSt (200 мкл на лунку) в течение 30 минут.
- Добавление образцов питательной среды (СМ), 100 мкл на лунку (20 мкл CM + 80 мкл человеческого IgG1, 25 нг/мл в 0.05% БСА), 2 часа при комнатной температуре. Образцы фрагмента Fab человеческого IgG1, 20 нг/мл в 0.05% БСА, использовали в качестве стандарта.
- Промывка лунок три раза буфером PBSt (0.1% Tween).
- Добавление меченых пероксидазой хрена анти-Fc антител, разбавленных 1/7000 в 0.05% БСА в PBS, 100 мкл на лунку, и инкубирование 90 минут при комнатной температуре.
- Промывка лунок шесть раз буфером PBSt (0.1% Tween).
- Добавление о-фенилендиамина 100 мкл (4 мг/10 мл) с Н2О2 в буфере рН 5.0.
- Остановка реакции добавлением 100 мкл 10% H2SO4.
Хотя настоящее изобретение было подробно описано со ссылкой на предпочтительные варианты его осуществления, для специалиста в данной области техники ясно, что могут быть сделаны различные замены и применены эквиваленты, которые не выходят за рамки настоящего изобретения. Все процитированные здесь документы являются частью настоящей заявки, включенные путем ссылки.













| название | год | авторы | номер документа |
|---|---|---|---|
| УЧАСТОК СВЯЗЫВАНИЯ АНТИГЕНА (Fab), В ТОМ ЧИСЛЕ ГУМАНИЗИРОВАННЫЙ Fab, ПРОТИВ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА С (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ Fab С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ, СПОСОБ И НАБОР ДЛЯ ДЕТЕКЦИИ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА С | 2016 |
|
RU2623157C1 |
| ПРЕПАРАТ НА ОСНОВЕ НЕЙТРАЛИЗУЮЩИХ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СВЯЗЫВАЮЩИХСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА БЕШЕНСТВА | 2018 |
|
RU2711553C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |
| НЕЙТРАЛИЗУЮЩЕЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ G ВИРУСА БЕШЕНСТВА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2017 |
|
RU2718835C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА И Fab, СВЯЗЫВАЮЩИЕСЯ С АНТИГЕНОМ F1 ИЗ Yersinia pestis, И СПОСОБ ИХ ПОЛУЧЕНИЯ С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2009 |
|
RU2420587C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА ЭБОЛА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2015 |
|
RU2630304C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА ЭБОЛА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2015 |
|
RU2639533C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА ЭБОЛА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2016 |
|
RU2644334C2 |
| Вариабельные домены лёгкой и тяжёлой цепи мышиного моноклонального антитела против интерферона альфа (IFN-α) человека, антигенсвязывающий фрагмент (Fab) против IFN-α человека, содержащий указанные домены | 2018 |
|
RU2694672C1 |
| ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ЛЕГКОЙ И ТЯЖЕЛОЙ ЦЕПИ МЫШИНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ АЛЬФА (ФНО-α) ЧЕЛОВЕКА (ВАРИАНТЫ), АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab) ПРОТИВ ФНО-α ЧЕЛОВЕКА, СОДЕРЖАЩИЙ УКАЗАННЫЕ ДОМЕНЫ (ВАРИАНТЫ) | 2012 |
|
RU2499000C1 |





Изобретение относится к биотехнологии и представляет собой гуманизированный антигенсвязывающий фрагмент (Fab) против вируса бешенства, изолированный фрагмент ДНК, кодирующий указанный Fab, клетку дрожжей, трансформированную этим фрагментом ДНК, способ получения указанного Fab с использованием этих клеток дрожжей. Изобретение может эффективно использоваться против вируса бешенства. 4 н. и 1 з.п. ф-лы, 10 ил., 2 табл.
1. Гуманизированный антигенсвязывающий фрагмент (Fab) против вируса бешенства, который содержит вариабельный участок тяжелой цепи (VH), соединенный с доменом СН1 человеческого иммуноглобулина IgGl, с последовательностью аминокислот SEQ ID NO: 2 и вариабельный участок легкой цепи (VL), соединенный с доменом Сæ человеческого иммуноглобулина каппа, с последовательностью аминокислот SEQ ID NO: 4.
2. Изолированный фрагмент ДНК, кодирующий антигенсвязывающий фрагмент (Fab) против вируса бешенства по п.1.
3. Клетка дрожжей, трансформированная фрагментом ДНК по п.2, обладающая способностью к продукции антигенсвязывающего фрагмента (Fab) против вируса бешенства по п.1.
4. Клетка дрожжей по п.3, отличающаяся тем, что указанными дрожжами являются дрожжи Pichia pastoris.
5. Способ получения антигенсвязывающего фрагмента (Fab) против вируса бешенства по п.1, включающий стадии выращивания клеток дрожжей по п.4 в питательной среде и выделения указанного Fab из культуральной жидкости.
| Cheung S.C | |||
| et al | |||
| A recombinant human Fab expressed in Escherichia coli neutralizes rabies virus // J Virol | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| YAMASHIRO TETSU, et al | |||
| He synthesis of a panel of human monoclonal antibodies with neutralizing activities against viral infectious diseases // Tropical Medicine and Health, Vol.35 (2007), No.2 p.47 | |||
| HOUIMEL M | |||

