Область техники
Настоящее изобретение относится к антителам, в частности к участкам связывания антигена (Fab), в том числе гуманизированным Fab, против ботулинического нейротоксина С, способу получения указанных Fab с использованием дрожжей, применению указанных Fab для детекции ботулинического нейротоксина С и к набору для указанного применения, содержащему указанные Fab.
Предшествующий уровень техники
Антитела обычно состоят из двух тяжелых цепей, связанных между собой дисульфидными связями, и легких цепей, ассоциированных с N-концом каждой из тяжелых цепей. Каждая тяжелая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Каждая легкая цепь также содержит на N-конце вариабельный домен с константным доменом. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий участок. Вариабельные домены легкой и тяжелой цепей обладают похожей общей структурой, и каждый домен включает каркас из четырех участков, последовательности которых являются относительно консервативными, связанных посредством трех участков, определяющих комплементарность (complementarity determining regions, CDRs). Четыре каркасных участка формируют конформацию типа бета-складчатого слоя, a CDRs образуют петли, связывающие конструкцию из бета-складчатого слоя. Участки CDRs расположены в непосредственной близости друг от друга благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDRs и каркасные участки антител могут быть определены путем ссылки на нумерационную систем у Кабата (Kabat numbering system, Kabat et al., (1987) "Sequences of Proteins of Immunological Interest", US Dept. of Health and Human Services, US Government Printing Office) в сочетании in conjunction with x-ray crystallography, as set forth in WO 91/09967.
Для получения антитела, которое может связываться с каким-либо специфическим антигеном, обычно используют методику Kohler и Milstein (Kohler et al., (1976) Nature 256:495-497). Она включает в себя главным образом иммунизацию мыши антигеном, слияние клеток селезенки из иммунизированной мыши с клетками миеломы мыши и селекцию среди полученных таким образом гибридом одной или нескольких гибридом, которые секретируют моноклональное антитело, специфическое для целевого антигена.
Ботулотоксин (ботулинический токсин, токсин ботулизма) - нейротоксин белковой природы, вырабатываемый бактериями Clostridium botulinum. Ботулотоксин является сильнейшим ядом из известных науке органических токсинов и веществ в целом (Супотницкий М.В. Глава 1.5. Токсины патогенных бактерий // Микроорганизмы, токсины и эпидемии. М., 2000. 376 с.). Попадание ботулотоксина в организм вызывает тяжелое токсическое поражение - ботулизм, который в естественных условиях встречается у людей, лошадей, птиц, реже - у крупного рогатого скота, пушных зверей. Ботулотоксин по серовару возбудителя делят на типы А, В, C1, С2, D, Е, F, G, из них наиболее часто встречается тип А. Летальная доза LD50 у различных сероваров в среднем составляет 0,001 мкг/кг веса, наиболее ядовитым является тип D (0,0004 мкг/кг веса). Ботулотоксин является одним из самых сложных белков, синтезируемых живым организмом. Его масса составляет около 150 тысяч атомных единиц масс, что в три раза превосходит типичный размер белковой цепи, немногие белки (например, титин) превышают этот средний размер.
Clostridium botulinum, будучи строгими анаэробами, способны размножаться только в условиях полного отсутствия кислорода. Как правило, это консервированные и колбасные изделия (особенно консервированные жареные грибы и заготовленные большими кусками мясо и рыба с повреждениями на поверхности). Вырабатываемый бактериями при размножении экзотоксин попадает в организм вместе с пищей, всасываясь в желудочно-кишечном тракте и воздействуя при этом на нервную систему, вызывает нарушения в работе черепных нервов, скелетной мускулатуры, нервных центров сердца. Характерны глазная симптоматика (туман, мушки перед глазами, мидриаз и анизокория зрачков, косоглазие), позднее присоединяются бульбарные симптомы (нарушение речи и глотания, маскообразное лицо). Смерть наступает от гипоксии, вызванной нарушением обменных процессов кислорода, асфиксией дыхательных путей, параличом дыхательной мускулатуры и сердечной мышцы.
Ботулотоксин не имеет вкуса, цвета и запаха (изредка пораженный продукт приобретает запах прогорклого масла). Разрушается при кипячении в течение 25-30 минут, при автоклавировании в течение 10 мин при температуре 120°С, при замачивании в растворе 1% пищевой соды в течение часа. Токсин не разрушается при взаимодействии с пепсином и с соляной кислотой желудочного сока.
Биохимические исследования показали, что нейротоксины обладают цинк-зависимой эндопептидазной активностью и энзиматически расщепляют ряд белков нервных окончаний, важных для нормального высвобождения нейротрансмиттеров. Ботулотоксины А и Ε расщепляют белок SNAP-25, ботулотоксин С - НРС-1 (синтаксин), токсины D и F расщепляют изоформу VAMP-1 (синаптобревин-1), токсин В - VAMP-2 (синаптобревин-2) и т.д. Протеолитическое расщепление этих специфических белков в нервных окончаниях приводит к угнетению нормального высвобождения ацетилхолина и, в конечном счете, к появлению основных признаков ботулизма.
Известно применение специфических антител для диагностики ботулинической инфекций (патент РФ 2152036). В патенте описан способ определения ботулотоксинов А и В на основе специфических F(ab)2-фрагментов антител. Предложены Fab-фрагменты рекомбинантных антител, специфичных к ботулотоксинам типов А и В, однако их структура, последовательность аминокислот и свойства не раскрыты.
Известны гибридомы, продуцирующие мышиные моноклональные антитела к ботулиническому нейротоксину типа В, характеризующиеся стабильностью продукции антител, на основе которых разработан ряд высокочувствительных тест-систем в формате иммуноферментного анализа (Stanker, L.H. A Monoclonal Antibody Based Capture ELISA for Botulinum Neurotoxin Serotype B: Toxin Detection in Food / L.H. Stanker, M.C. Scotcher, L. Cheng, K. Ching, J. McGarvey, D. Hodge, and R. Hnasko // Toxins (Basel). 2013. V. 5. P. 2212-2226).
Описан штамм гибридных клеток Mus. musculus 3F11, продуцирующих моноклональные антитела, специфичные к ботулиническому токсину типа В (патент РФ №2566553). Указанный штамм депонирован в коллекцию микроорганизмов Федерального государственного учреждения науки Государственного научного центра прикладной микробиологии и биотехнологии (ФБУН ГНЦ ПМБ) под коллекционным номером Н-45. Структура антител, их последовательность аминокислот и свойства не раскрыты.
Наиболее близким аналогом настоящего изобретения являются Fab-фрагменты антител, которые могут использоваться в качестве иммуносенсоров для обнаружения ботулотоксинов в продуктах питания и для других нужд здравоохранения и министерства обороны (патент США №5932449).
Таким образом, разработка реагентов и экспресс-методов для детекции ботулинических нейротоксинов, в частности ботулинического нейротоксина С, остается актуальной проблемой как для медицины, так и для органов государственного санитарно-эпидемиологического контроля вследствие экстремальной токсичности и самой высокой летальности при пищевых отравлениях. Кроме того, в последнее время внимание к данной категории токсинов возросло во всем мире из-за угрозы биотерроризма.
Краткое описание изобретения
Целью настоящего изобретения было получение реагентов для детекции ботулинического нейротоксина С (BoNT/C). В качестве указанных реагентов для детекции BoNT/C предпочтительно использование рекомбинантных высокоаффинных антител против BoNT/C, которые могут применяться в экспресс-тестах для определения содержания BoNT/C в объектах окружающей среды, продуктах питания и кормах.
Указанная цель была достигнута путем выделения мышиных антител, обладающих способностью к связыванию с BoNT/C, определения последовательности аминокислот их участков связывания антигена (Fab), последующей гуманизации полученных Fab и демонстрацией их высокой специфичности в связывании BoNT/C. Также указанная цель была достигнута путем создания способа получения указанного функционального Fab с использованием клеток дрожжей.
Настоящее изобретение предоставляет новые изолированные фрагменты с участками связывания антигена (fragment antigen binding, Fab) моноклонального мышиного антитела, которое селективно связывается с BoNT/C и включает в себя вариабельный участок тяжелой цепи (VH) с последовательностью аминокислот SEQ ID NO: 1 или 5, и вариабельный участок легкой цепи (VL) с последовательностью аминокислот SEQ ID NO: 2 или 6.
Также настоящее изобретение предоставляет изолированные фрагменты ДНК, кодирующие фрагмент с антиген-связывающим участком (fragment antigen binding, Fab), описанный выше.
Также настоящее изобретение предоставляет гуманизированные моноклональные Fab, которые селективно связываются с BoNT/C. Последовательности аминокислот тяжелых цепей указанных гуманизированных Fab приведены в Перечне последовательностей под номерами SEQ ID NO: 9 и 10, последовательности аминокислот легких цепей указанных гуманизированных Fab приведены в Перечне последовательностей под номерами SEQ ID NO: 13 и 14.
Также настоящее изобретение предоставляет изолированные фрагменты ДНК, кодирующие гуманизированные Fab, описанные выше.
Также настоящее изобретение предоставляет клетку дрожжей, трансформированную описанным выше фрагментом ДНК, обладающую способностью к продукции указанных гуманизированных Fab.
Также настоящее изобретение предоставляет клетку дрожжей, описанную выше, где указанной клеткой дрожжей является Pichia pastoris.
Также настоящее изобретение предоставляет способ получения гуманизированных Fab, описанных выше, включающий выращивание указанных дрожжей в питательной среде и выделение указанных Fab из культуральной жидкости.
Также настоящее изобретение предоставляет способ детекции BoNT/C, включающий: (а) получение образца; (b) взаимодействие указанного образца с Fab, описанными выше; (с) детектирование комплекса, образованного указанным Fab и BoNT/C; и, необязательно, (d) определение количества BoNT/C, присутствующего в указанном образце на основании детектированного количества комплекса, образованного Fab и BoNT/C.
Также настоящее изобретение предоставляет набор для детекции BoNT/C, содержащего указанные выше изолированные Fab, которые селективно связываются с BoNT/C, и средства, позволяющие детектировать указанный BoNT/C с использованием указанных Fab.
Краткое описание чертежей
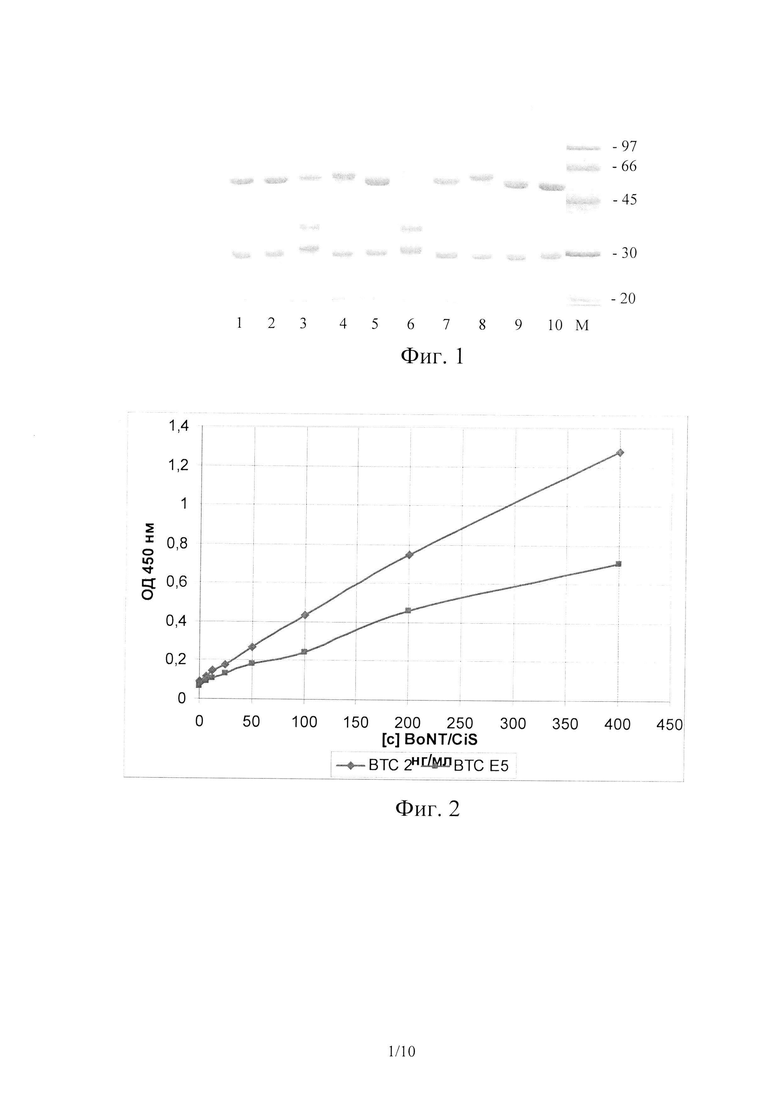
На Фиг. 1 показан 12% SDS-PAGE панели моноклональных антител в восстанавливающих условиях. Дорожка 1 - В ТС 1; Дорожка 2 - ВТС D4; Дорожка 3 - ВТС D41-; Дорожка 4 - ВТС DIO; Дорожка 5 - ВТС Е5; Дорожка 6 - ВТС Е8; Дорожка 7 - ВТС В2Е12; Дорожка 8 - ВТС F5; Дорожка 9 - ВТС G1; Дорожка 10 - ВТС 2; M - стандарты молекулярной массы.
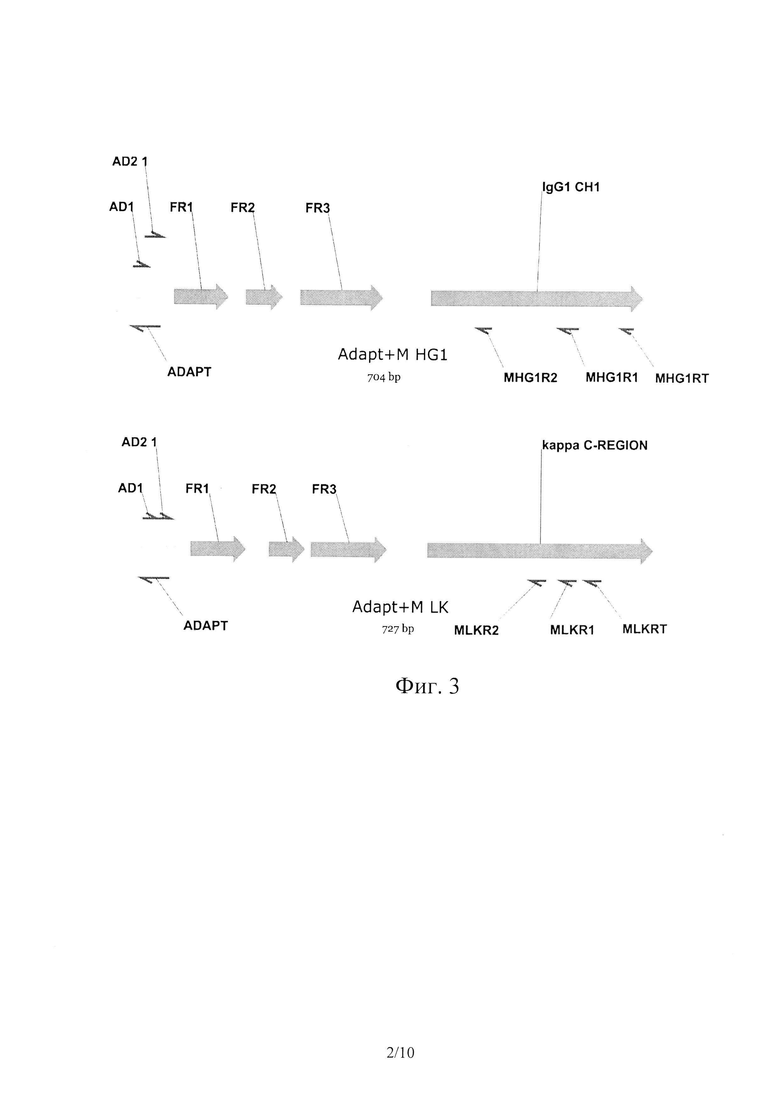
На Фиг. 2 показаны калибровочные кривые для определения BoNT/Cis методом «сэндвич» ИФА.
На Фиг. 3 показана схема расположения адаптера ADAPT, праймеров для обратной транскрипции (M…RT), первичной (AD1, M…R1) и вторичной (AD2_1, M…R2) ПЦР.
На Фиг. 4 показаны нуклеотидные и транслированные аминокислотные последовательности VL:VH генов иммуноглобулинов мыши, а, в) гены тяжелых цепей клеточных линий ВТС1 и ВТС2, соответственно; б, г) гены легких цепей клеточных линий ВТС1 и ВТС2, соответственно. Указаны «framework regions» (FR) и «complementarity determining regions» (CDR).
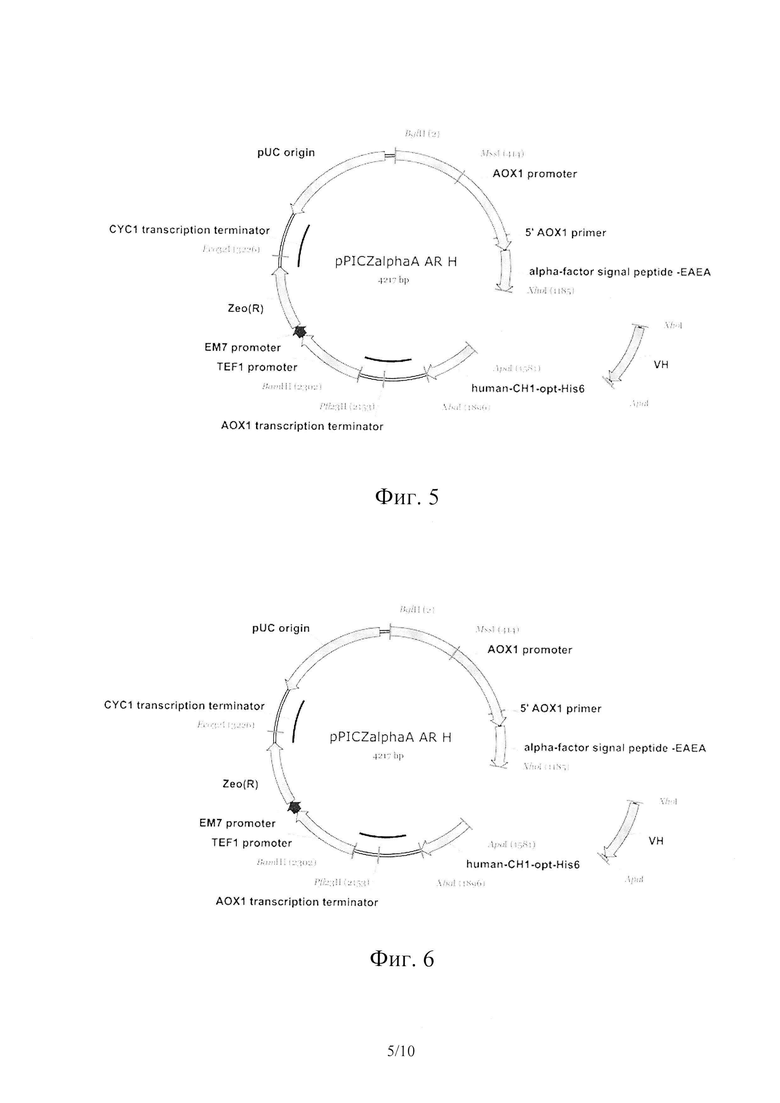
На Фиг. 5 показана схема клонирования ПЦР-фрагментов вариабельных доменов генов тяжелых цепей в плазмиду pPICZαA-AR-H. Указаны сайты действия эндонуклеаз, промоторы и терминаторы генов, кодирующие области сигнального пептида, вариабельного и константного домена, гена устойчивости к антибиотику, а также область, ответственная за репликацию плазмиды в Е.coli.
На Фиг. 6 показана схема клонирования ПЦР-фрагментов вариабельных доменов генов легких цепей в плазмиду pPICZαA-ΔPmeI-AR-L. Обозначения аналогичны обозначениям на Фиг. 5.
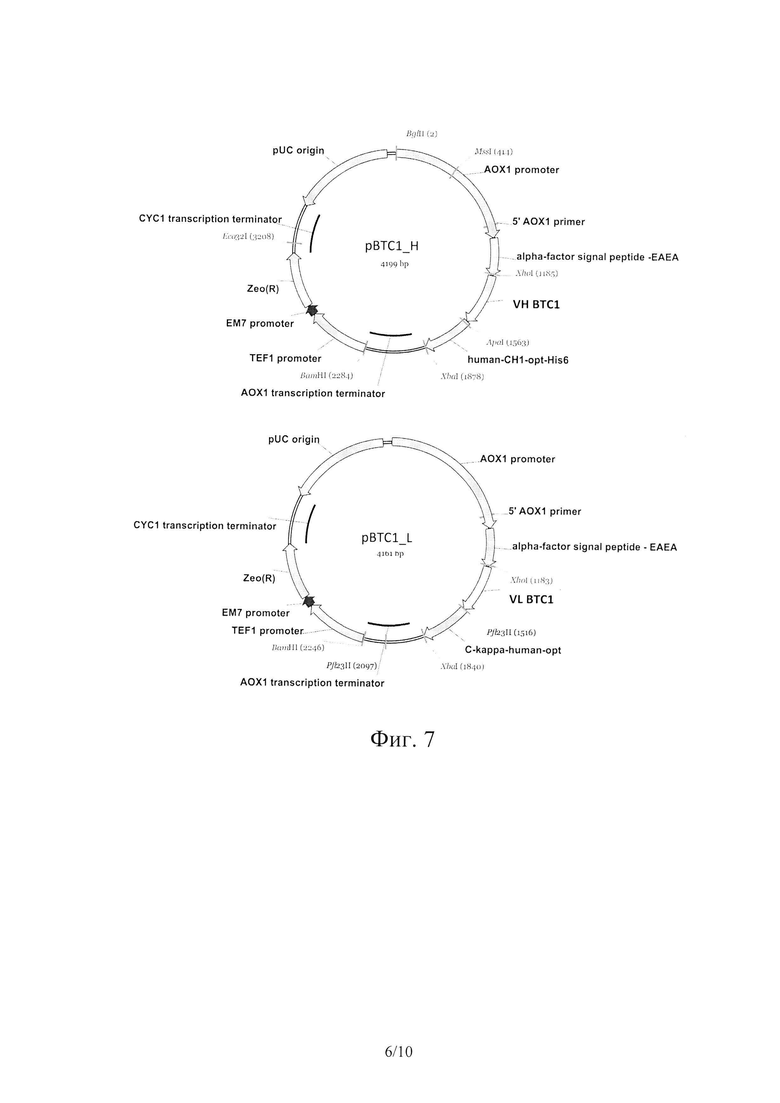
На Фиг. 7 показаны карты экспрессионных плазмид pBTC1_H и pBTC1_L.
На Фиг. 8 показаны карты экспрессионных плазмид рВТС2_Н и pBTC2_L.
На Фиг. 9 показаны карты экспрессионных плазмид pBTC1 и рВТС2.
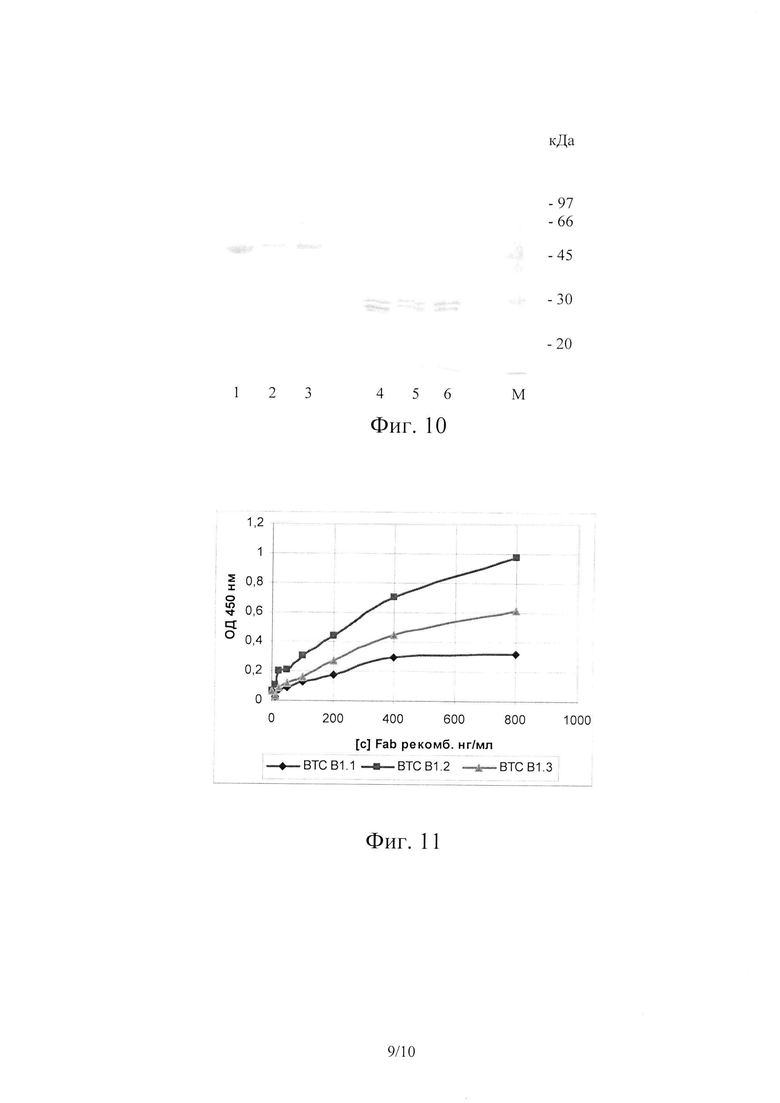
На Фиг. 10 показан 12% SDS-PAGE рекомбинантных гуманизированных Fab ВТС1.1, ВТС1.2, ВТС1.3 в невосстанавливающих (дорожки 1-3) и восстанавливающих (дорожки 4-6) условиях. Дорожки 1, 4 - ВТС1.1; дорожки 2, 5 - ВТС1.2; дорожка 3, 6 - ВТС1.3; дорожка M - стандарты молекулярной массы.
На Фиг. 11 показаны кривые титрования рекомбинантных гуманизированных Fab в непрямом ИФА с иммобилизованным BoNT/CiS.
На Фиг. 12 показаны результаты иммуноблоттинга рекомбинантных Fab фрагментов против BoNT/CiS в восстанавливающих (дорожки 1-5) и невосстанавливающих (дорожки 6-10) условиях после 12% SDS-PAGE: А - МоАт против Сκ, Б - МоАт против His6tag. Дорожки 1,5 - 1С5 Fab фрагмент (положительныйконтроль); дорожки 2, 6 - рекомбинантный Fab фрагмент ВТС2; дорожки 3, 7 - рекомбинантный Fab фрагмент ВТС1 (серия ВТС 1.3); дорожки 4, 8 - рекомбинантный Fab фрагмент ВТС1 (серия ВТС 1.2); дорожки 5, 10 - рекомбинантный Fab фрагмент ВТС1 (серия ВТС 1.1).
Подробное описание настоящего изобретения
Подходящий способ для выделения антител, эффективных для использования в рамках настоящего изобретения, включает: (а) назначение животному эффективного количества белка, пептида или его мимотопа с целью получения антител и (b) выделение указанных антител. В данном конкретном случае для иммунизации животных был использован нетоксичный аналог BoNT/C (BoNT/CiS), предоставленный компанией fzmb GmbH (Бад-Лагензальца, Германия). Затем были проведены эксперименты, подтверждающие, что антитела против нетоксичного аналога BoNT/CiS также эффективно связываются с BoNT/C.
Антитела, полученные против определенных белков или мимотопов, могут обладать некоторыми преимуществами, поскольку такие антитела не сильно загрязнены антителами против других соединений, что могло бы, в противном случае, повлиять на точность диагностического метода. Способы получения таких антител известны из предшествующего уровня техники и подробно описаны Harlow и др. (Antibodies, a Laboratory Manual, Cold Spring Harbor Labs Press, 1988) и включают в себя иммунизацию животных с целью получения препаратов антител, которые выделяют из, например, плазмы или асцитной жидкости и очищают способами, известными из уровня техники, с получением препаратов, дающих реакцию с антигеном. Множество видов имеют белки с близкими последовательностями и поэтому могут возникнуть сложности в ходе использования стандартных протоколов иммунизации при получении антител, которые узнают белок только одного вида. Поэтому также стоит рассмотреть модификации стандартных методов получения антител, таких как, например, разностные (вычитательные) гибридизационные методики. Такие модификации могут быть известны специалисту в данной области техники, также они описаны в настоящем описании. В других методах антитела, которые могут быть использованы в рамках настоящего изобретения, получают с использованием рекомбинантных методов, описанных Sambrook и др. (Molecular Cloning: A Laboratory Manual, (Cold Spring Harbor Labs Press, 1989)).
Вкратце, моноклональные антитела получают путем слияния клеток селезенки из иммунизированного животного и клеток миеломы с получением гибридомы. Гибридомы могут быть проверены на способность к продукции нужного антитела, затем гибридомы могут быть выращены, из них могут быть выделены указанные антитела. Термин «выращенные клетки», использованный в настоящем описании, означает гибридомы или другие линии клеток, которые производят антитела. Методы получения и проверки таких выращенных клеток описаны Harlow и др. (см. выше). Получение материала, использующегося в качестве антигена, для инъекций животных включают в себя методики, хорошо известные из уровня техники, например использование полноразмерного белка, использование пептида, выбранного из иммуногенных участков белка, модифицирование антигена такими способами, как, например, связывание с динитрофенолом, связывание с арсаниловой кислотой, денатурация антигена, связывание антигена с белком-переносчиком, таким как, например, keyhole limpet hemacyanin, с пептидами, содержащими участки связывания с рецепторами Т-клеток класса II, с бусинами, а также любыми другими методами, известными из уровня техники. См. Harlow и др. (см. выше).
В частности, Fab в соответствии с настоящим изобретением являются Fab мышиных моноклональных антител против BoNT/C. Указанные Fab являются Fab мышиных моноклональных антител, содержащих вариабельный участок тяжелой цепи антитела (VH) с аминокислотной последовательностью SEQ ID NO: 1 или 5, и вариабельный участок легкой цепи антитела (VL) с аминокислотной последовательностью SEQ ID NO: 2 или 6. Указанные новые Fab мышиных моноклональных антител селективно связываются с BoNT/C.
Далее, гуманизированными Fab в соответствии с настоящим изобретением также являются гуманизированные Fab против BoNT/C, которые были получены на основе описанных выше Fab мышиных антител путем их «гуманизации». Окончательные последовательности аминокислот тяжелых цепей указанных гуманизированных Fab против BoNT/C приведены в Перечне последовательностей под номерами SEQ ID NO: 9 и 10, последовательности аминокислот легких цепей указанных гуманизированных Fab против BoNT/C приведены в Перечне последовательностей под номерами SEQ ID NO: 13 и 14. Указанные новые гуманизированные Fab селективно связываются с BoNT/C.
Любая часть гуманизированных антител (и, следовательно, в широком смысле определения термина антитела), полученная в соответствии с настоящим описанием, не выходит за рамки настоящего изобретения, при условии, что специфичность связывания и аффинность к BoNT/C сохраняется. Таким образом, связывающие белки, полученные на основе указанных Fab, в частности полноразмерные антитела, содержащие указанные Fab, без сомнений находятся в рамках настоящего изобретения, как и другие фрагменты, которые способствуют проявлению из способности, по крайней мере, в степени, необходимой для терапевтического использования, как описано ниже.
В настоящем изобретении фраза «антитело или Fab, обладающий способностью к связыванию с BoNT/C» означает молекулу, которая связывается с BoNT/C и образует стабильный комплекс. Стабильным комплексом является комплекс, в котором связывание между партнерами происходит на период времени, достаточный для того, чтобы произвести детектирование указанного комплекса с использованием описанных здесь методов. Термин «селективно связывает BoNT/C» означает способность указанной молекулы предпочтительно связываться с BoNT/C в отличие от связывания с белками, не имеющими отношения к BoNT/C, или связывания с небелковыми компонентами, присутствующими в образце. Антителом или Fab, которое предпочтительно связывается с BoNT/C, является антитело или Fab, которое связывается с BoNT/C, но не связывается в существенной степени с другими молекулами или компонентами, которые могут присутствовать в образце. Существенная степень связывания предполагает, например, связывание антитела, связывающегося с BoNT/C, с молекулой, не являющейся в существенной степени, с аффиностью или силой, достаточной для того, чтобы помешать способности антитела, связывающегося с BoNT/C, определить уровень BoNT/C в образце. Примерами таких молекул и компонентов, которые могут присутствовать в образце, являются, но не ограничиваются ими, белки, не являющиеся BoNT/C, липиды и углеводы.
Способность антитела или Fab к связыванию с антигеном может быть определена специалистом в данной области с использованием методов, включающих, но не ограничивающимися методом ELISA и равновесным диализом. Методы определения аффинности и силы связывания хорошо известны специалисту в данной области техники, подробно описаны Janeway и др. (Immunobiology: The Immune System in Health and Disease (Garland Publishing Company, 1996)).
Fab, пригодный для осуществления настоящего изобретения, - это Fab, обладающий способностью к связыванию BoNT/C, когда концентрация BoNT/C составляет от около 10 нг/мл и около 10 пг/мл. В частности, пригодные в рамках настоящего изобретения антитела связывают антиген BoNT/C, когда концентрация BoNT/C составляет 10 нг/мл, около 1 нг/мл или менее, предпочтительно 100 пг/мл. Такие антитела и Fab описаны в сопутствующих Примерах.
Изолированным фрагментом ДНК, кодирующим гуманизированный Fab согласно настоящему изобретению, является экспрессирующийся фрагмент ДНК, содержащий промотор, сигнальную последовательность, последовательность нуклеотидов, кодирующую структурные части тяжелой и легкой цепей антитела, участок терминации транскрипции.
В частности, нуклеотидные последовательности, кодирующие структурные части тяжелой и легкой цепей Fab согласно настоящему изобретению, содержат фрагмент ДНК, кодирующий вариабельный участок тяжелой цепи (VH) (SEQ ID NO: 3 или 7) и вариабельный участок легкой цепи (VL) (SEQ ID NO: 4 или 8, соответственно), связанный с фрагментом ДНК, кодирующим консервативные участки обеих цепей, необходимые для функционирования указанного антитела.
Также, нуклеотидные последовательности, кодирующие Fab согласно настоящему изобретению, содержат фрагмент ДНК, кодирующий вариабельный участок тяжелой цепи (VH), например SEQ ID NO: 3 или 7, и фрагмент ДНК, кодирующий вариабельный участок легкой цепи (VL), например SEQ ID NO: 4 или 8.
Также, нуклеотидные последовательности, кодирующие структурные части тяжелой и легкой цепей гуманизированного Fab согласно настоящему изобретению, содержат фрагмент ДНК, кодирующий тяжелые цепи (VH), например SEQ ID NO: 9 или 10, и фрагмент ДНК, кодирующий легкие цепи (VL), например SEQ ID NO: 13 или 14.
Ввиду вырожденности трансляционного кода могут быть различия в последовательности ДНК. Фрагменты ДНК согласно настоящему изобретению не ограничены фрагментами, показанными в SEQ ID NO: 3 или 7, 4 или 8, 11 или 15, и 12 или 16, при условии, что они кодируют участки цепей антитела с аминокислотной последовательностью, показанной в SEQ ID NO: 1 или 5, 2 или 6, 9 или 13, и 10 или 14 соответственно.
Фрагмент ДНК, кодирующий антитело или Fab согласно настоящему изобретению, может быть получен любым методом, известным специалисту в данной области техники, включая ПЦР с использованием набора перекрывающихся праймеров, синтез по технологии Slonogene (Sloning Biotechnology GmbH), химическим способом и т.д.
Клетками дрожжей согласно настоящему изобретению являются клетки дрожжей, трансформированные фрагментом ДНК, описанным выше, и обладающие способностью к продукции Fab против BoNT/C.
Фраза «клетки дрожжей, трансформированные фрагментом ДНК» означает, что желаемый фрагмент ДНК был введен в клетку дрожжей с использованием методов, известных специалисту в данной области техники. Трансформация клетки дрожжей фрагментом ДНК приводит к увеличению экспрессии фрагмента ДНК, кодирующего антитело согласно настоящему изобретению. Присутствие сигнальной последовательности α-фактора приводит к секреции произведенного антитела в культуральную жидкость. Методы трансформации клеток дрожжей включают в себя все известные методы, например модифицированная версия процедуры, описанной для S.cerevisiae (Gietz and Schiesti, 1996).
Способом согласно настоящему изобретению является способ получения гуманизированных Fab против BoNT/C, включающий выращивание дрожжей в питательной среде и выделение полученных Fab из культуральной жидкости.
Примером клетки дрожжей, пригодной для продукции гуманизированных Fab согласно настоящему изобретению, являются клетки дрожжей Pichia pastoris, но не ограничивается ими. Фраза «дрожжи Pichia pastoris» означает, что указанные дрожжи классифицируют как Pichia pastoris (P.pastoris) в соответствии с классификацией, известной специалисту в данной области микробиологии. Примерами дрожжей Р.pastoris, применимых в рамках настоящего изобретения, являются дрожжи P.pastoris GS115 (Invitrogen), но не ограничиваются ими.
В настоящем изобретении выращивание, накопление и очистка Fab из культуральной жидкости и других жидкостей может быть осуществлена методом, сходным с традиционными методами ферментации, когда некий белок производится с использованием микроорганизма.
Питательная среда для выращивания может быть как синтетической, так и натуральной при условии, что указанная среда содержит источник углерода, источник азота, минералы и, если это необходимо, подходящее количество питательных веществ, в которых нуждаются дрожжи для их роста. К источникам углерода относятся углеводы, такие как глюкоза и сахароза, и различные органические кислоты. В зависимости от способа ассимиляции у указанного микроорганизма может быть использован спирт, включая метанол, этанол, глицерин. В качестве источника азота могут быть использованы различные соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, источники природного азота, такие как пептон, гидролизат соевых бобов, расщепленные ферментированные микроорганизмы. В качестве источника минералов могут быть использованы монофосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут быть использованы тиамин, дрожжевой экстракт и подобные им соединения.
Выращивание предпочтительно осуществляют в аэробных условиях, таких как культивирование с перемешиванием, взбалтывание с аэрацией, при температуре от 20 до 40°С, предпочтительно от 28 до 30°С. рН среды обычно поддерживают в диапазоне от 2 до 9, предпочтительно в диапазоне от 6 до 7.5. рН среды может быть скорректирован с помощью аммиака, карбоната кальция, различных оснований и буферов. Обычно выращивание в течение от 1 до 7 дней приводит к накоплению Fab в культуральной жидкости.
После выращивания твердые компоненты, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрации с использованием мембраны, а затем антитела или Fab могут быть выделены и очищены методом осаждения с солями, с использованием сульфата натрия или сульфата аммония, аффинной хроматографии, ионообменной хроматографии и т.п.
Моноклональные мышиные антитела в соответствии с настоящим изобретением, которые селективно связываются с BoNT/C, также могут быть получены с использованием подходящей гибридомы. Гибридомы могут быть выращены в питательной среде и накопленные антитела могут быть выделены. Здесь и далее термин "выращиваемые клетки" относится к гибридомам или другим линиям клеток, которые производят антитела. Способы получения и проверки таких выращиваемых клеток описаны Harlow и др. (см. выше). Методы получения антигенного материала для инъекций в животное включают в себя методы, известные из уровня техники, например использование полноразмерного белка, пептидов, выбранных из иммуногенного участка этого белка, модифицирование антигена такими способами, как, например, связывание с динитрофенолом, связывание с арсаниловой кислотой, денатурация антигена, связывание антигена с белком-переносчиком, таким как, например, keyhole limpet hemacyanin, с пептидами, содержащими участки связывания с рецепторами Т-клеток класса II, с бусинами, а также любыми другими методам, известными из уровня техники. Смотри Harlow и др. (см. выше).
Другим способом согласно настоящему изобретению является способ детекции BoNT/C. Указанный способ включает в себя: (а) получение образца; (b) взаимодействие указанного образца с Fab, описанными выше; (с) детектирование комплекса, образованного указанным Fab и BoNT/C; и, необязательно, (d) определение количества BoNT/C, присутствующего в указанном образце на основании детектированного количества комплекса, образованного Fab и BoNT/C. Присутствие в образце любого количества BoNT/C является свидетельством наличия токсина.
В одном из вариантов осуществления настоящего изобретения образец получают или выделяют из объектов окружающей среды, продуктов питания или кормов для того, чтобы протестировать на наличие BoNT/C. Специалист в данной области может без труда выбрать подходящие образцы.
Хотя это не является необходимым в рамках настоящего изобретения, образец может быть подвергнут желаемой предварительной обработке. Например, образец может быть нормирован до достижения желаемой степени специфической активности. Нормирование образца с использованием подходящего метода разведения, известного из уровня техники, позволяет определить количественное содержание BoNT/C независимо от начальной концентрации (например, удельного веса) в образце.
После получения образца определяют уровень BoNT/C в данном образце. Термины «определяют», «определяют уровень BoNT/C», «определяют количество BoNT/C» и подобные им, использованные здесь, используются для того, чтобы очертить круг методов, которые могут быть использованы для определения или измерения количества BoNT/C в образце. Такие методы могут дать качественные или количественные результаты. Уровень содержания BoNT/C может быть определен по содержанию целого белка BoNT/C или по содержанию фрагментов, продуктов деградации или продуктов реакции BoNT/C.
Антитела или Fab, связывающиеся с BoNT/C, пригодные в способах согласно настоящему изобретению, могут включать в себя многофункциональные антитела, например бифункциональные антитела или Fab, содержащие, по крайней мере, одну функциональную часть, которая специфически связывается с BoNT/C. Такие многофункциональные антитела или Fab могут включать в себя, например, химерные молекулы, включающие в себя молекулу, которая связывается с BoNT/C, и вторую часть, которая дает возможность данной химерной молекуле связываться с субстратом или позволяет детектировать ее способом, при котором связывание с BoNT/C не ухудшается. Примерами таких вторых частей являются фрагменты молекулы иммуноглобулина, флуоресцентный белок или фермент, но не ограничиваются ими.
Гибриды или слитые белки антител или Fab, которые сохраняют способность связываться с BoNT/C, также могут быть использованы. В таких гибридах часть, связывающая BoNT/C, может быть связана со второй частью, которая может связываться с субстратом или может быть детектирована. Примеры таких вторых частей включают в себя фрагменты молекул иммуноглобулина, эпитопы, флуоресцентные белки или ферменты, но не ограничиваются ими.
Антитело или Fab, обладающее способностью к связыванию с BoNT/C, использованное в рамках настоящего изобретения, может содержаться в составе для детекции. Например, антитело или Fab может содержаться в буфере, в котором это антитело или Fab растворено, и/или быть объединено с носителем. Подходящие буферы и носители известны для специалиста в данной области техники. Примеры подходящих буферов включают в себя любой буфер, в котором антитело или Fab может функционировать с селективным связыванием BoNT/C, такой как фосфатный солевой буфер, вода, буфер HEPES (буферная соль N-2-гидроксиэтилпиперазин-N'-2-этансульфоновой кислоты), буфер TES (солевой буфер Трис-ЭДТА), буфер Трис и буфер ТАЭ (Трис-ацетат-ЭДТА), но не ограничиваются ими. Примеры носителей включают в себя полимерные матрицы, токсоиды, сывороточные альбумины, такой как бычий сывороточный альбумин, но не ограничиваются ими. Носители могут быть комбинированы с антителом или Fab, или конъюгированы (присоединены) к антителу или Fab способом, который существенно не изменяет способность антитела или Fab к связыванию BoNT/C.
Термин «взаимодействие», использованный здесь, означает введение, добавление образца, предположительно содержащего BoNT/C, к антителу или Fab, связывающему BoNT/C, например, путем объединения или смешения образца с антителом или Fab. В случае, если BoNT/C присутствует в образце, образуется комплекс BoNT/C с антителом или Fab; образование такого комплекса означает способность антитела или Fab селективно связываться с BoNT/C с образованием стабильного комплекса, который может быть детектирован. Детектирование может быть качественным, количественным или полуколичественным. Связывание BoNT/C из образца с антителом или Fab происходит в условиях, подходящих для формирования комплекса. Такие условия (например, подходящие концентрации, буферы, температура, время реакции), а также методы оптимизации таких условий хорошо известны специалисту в данной области техники. Связывание может быть обнаружено и измерено с использованием множества методов, являющихся стандартными в данной области техники, включая ферментативные методы иммунохимии (например, ELISA), иммунопреципитации, методы иммуноблотинга и другие методы иммунохимии, описанные, например, Sambrook и др. (см. выше) и Harlow и др. (см. выше), но не ограничиваются ими. Эти ссылки также описывают примеры условий, в которых образуется комплекс.
В рамках одного из воплощений настоящего изобретения комплекс BoNT/C и антитела или Fab может образовываться в растворе. В рамках другого воплощения настоящего изобретения может образовываться комплекс BoNT/C с антителом или Fab, в котором антитело или Fab иммобилизовано на подложке (например, нанесено на подложку). Методики иммобилизации хорошо известны специалисту в данной области техники. Подходящие материалы для подложки включают в себя, но не ограничиваются пластмассой, стеклом, гелем, целлулоидом, тканями, бумагой и другими материалами. Примеры материалов подложки включают в себя, но не ограничиваются латексом, полистиролом, нейлоном, нитроцеллюлозой, агарозой, хлопком, PVDF (поливинилиденфторидом) и магнитными смолами. Подходящие формы материала подложки включают в себя, но не ограничиваются углублениями (например, плашки для микротитрования), чашки для микротитрования, стержни, полоски, бусины, аппараты с ответвляющимся потоком, мембраны, фильтры, трубки, чашки, целлулоидный матрикс, магнитные частицы, другие макрочастицы. В частности, предпочтительными подложками являются, например, пластинки для ELISA, стержни, полоски для иммуноанализа, радиоиммунологические чашки, агарозные бусины, пластиковые бусины, бусины из латекса, гибки, хлопковые нити, пластиковые чипы, мембраны для иммуноблотинга, бумага для иммуноблотинга и проточные мембраны. В одном из вариантов воплощения изобретения подложка, указанная выше, может содержать маркер для детекции. Описание примеров материалов для подложки смотри, например, в Kemeny, D.M. (1991) A Practical Guide to ELISA, Pergamon Press, Elmsford, N.Y. pp 33-44, and Price, С.) и Newman, D. eds. (Principles and Practice of Immunoassay, 2nd edition (1997) Stockton Press, NY, N.Y.).
Антитело или Fab может быть иммобилизовано на подложке, такой как плашка для микротитрования, стержне, полоске для иммуноанализа, аппараты с ответвляющимся потоком. Образец наносят на подложку и инкубируют в условиях, подходящих (то есть, достаточных) для образования комплекса BoNT/C с антителом, связанным с подложкой.
В соответствии с настоящим изобретением комплекс BoNT/C с антителом или Fab детектируют. Термин «детектирование образования комплекса», использованный здесь, означает идентификацию присутствия антитела или Fab, связанного с BoNT/C. Если комплекс образуется, то количество образованного комплекса может быть оценено количественно, но это не является обязательным. Образование комплекса или селективное связывание между предполагаемым в образце BoNT/C с антителом или Fab может быть измерено (то есть, детектировано, определено) с использованием множества методов, являющихся стандартными в данной области (см., например, Sambrook и др., выше), примеры которых описаны здесь. Комплекс может быть детектирован множеством методов, включающих в себя, но не ограниченных использованием одного или нескольких методов: ферментоподобным иммунометодом, конкурентным ферментоподобным иммунометодом, радиоиммунным, флуоресцентным иммунометодом, хемилюминесцентным иммунометодом, методом с использованием аппаратов с ответвляющимся потоком, проточным методом, методом агглютинации, методом с использованием макрочастиц (например, с использованием частиц, таких как, но не ограниченных магнитными частицами или пластиковыми полимерами, такими как латекс или полистироловые бусины), методом иммунопреципитации, методом BioCore™ (например, с использованием коллоидного золота), методом иммунодота (например, Heska AG's Immunodot System, Фрибург, Швейцария) и методом иммуноблотинга (например, Вестерн-блотинг), методом фосфоресценции, проточным методом, методом хроматографии, методом с использованием полиакриламидного геля, методом поверхностного резонанса плазмонов, методом спектрофотометрии, методом с использованием частиц, методом электронных сенсоров. Указанные способы хорошо известны специалисту в данной области техники.
Эти методы могут быть использованы для получения количественных или качественных результатов в зависимости от того, как эти методы будут использованы. Результаты методов могут базироваться на детектировании нативной молекулы BoNT/C или его фрагментов, продуктов деградации или продуктов реакции BoNT/C. Некоторые методы, такие как агглютинация, разделение с использованием макрочастиц и иммунопреципитация, могут дать визуальный результат (например, как наблюдение невооруженным глазом или с использованием устройств, таких как денситометр или спектрофотометр), не требующий применения маркера для детектирования.
В других методах конъюгация (то есть, присоединение) антитела или Fab с маркером для детектирования или с реагентом, который селективно связывается с антителом (например, антитело на антитело или Fab, связывающее BoNT/C), помогает в детектировании образования комплекса. Такие соединения и реагенты являются примерами молекул для детектирования (то есть молекул, способных детектировать комплекс антитела с BoNT/C). Маркер для детектирования может быть присоединен к части молекулы, которая не влияет на способность этой молекулы связываться с BoNT/C. Методы конъюгации хорошо известны специалисту в данной области техники. Примеры маркеров для детектирования включают в себя, но не ограничиваются радиоактивными, флуоресцентными, хемилюминесцентными, хромофорными, ферментативными, фосфоресцентными, электронными маркерами; метками из золей металлов, цветных бусин, физическими метками или лигандами. Бусинами считаются макрочастицы, состоящие из материала в виде матрикса, такого как латекс или полистирол, который может быть ковалентно или нековалентно связан с молекулой для детекции. К лигандам относятся молекулы, которые селективно связываются с другой молекулой. Предпочтительные маркеры для детектирования включают в себя, но не ограничиваются флуоресцеином, радиоактивными изотопами, фосфатазой (например, щелочной фосфатазой), биотином, авидином, пероксидазой (например, пероксидазой хрена), β-галактозидазой, соединениями, подобными биотину или авидину (например, стрептовидин или ImmunoPure® NeutrAvidin). Например, в наиболее предпочтительном варианте осуществления изобретения антитела, связывающие BoNT/C, конъюгируют с бусинами их латекса.
Настоящее изобретение также может включать в себя один или несколько слоев и/или типов вторичных молекул или других связывающих молекул, способных детектировать присутствие комплекса антитела или Fab с BoNT/C. Например, немодифицированные (то есть не связанные с молекулой для детектирования) вторичные антитела, которые селективно связываются с антителом или Fab, связывающим BoNT/C, могут связываться модифицированным (то есть связанным с молекулой для детектирования) третичным антителом, которое селективно связывает вторичные антитела, что позволяет детектировать BoNT/C. Подходящие вторичные антитела, третичные антитела и другие вторичные и третичные молекулы могут быть легко выбраны специалистом в данной области техники. Предпочтительные третичные молекулы также могут быть выбраны специалистом в данной области техники на основании характеристик вторичной молекулы. Подобная стратегия может быть применена для последовательных слоев.
Также могут быть использованы не иммунологические методы. Для того чтобы детектировать BoNT/C, могут быть использованы методы, такие как предварительное концентрирование образца, для повышения концентрации BoNT/C с целью повышения чувствительности к белку. Такие не иммунологические методы также включают в себя, например, электрофорез, где детектирование BoNT/C может быть произведено методами, известными из уровня техники, например окрашивание белка. В другом варианте осуществления изобретения тест на наличие BoNT/C, основанный на использовании белков, может быть использован для определения BoNT/C в предварительно концентрированном образце.
Как только уровень BoNT/C измерен, может быть сделано заключение о наличии токсина в образце. Заключение о наличии токсина в образце означает сравнение уровня BoNT/C в образце для тестирования с уровнем, обнаруженным в чистом образце. В рамках настоящего изобретения заключение о наличии токсина делается, когда детектируется любое количество BoNT/C.
Настоящее изобретение также включает в себя набор, подходящий для детекции (детектирования) BoNT/C, использующий способ, описанный выше. Термин «подходящий» означает, что детекция включает в себя методы, описанные здесь, использующие антитела или Fab, связывающие BoNT/C, в соответствии с настоящим изобретением. Такие антитела или Fab могут быть конъюгированы с маркером для детектирования. В другом варианте осуществления изобретения набор может содержать немеченые антитела или Fab в соответствии с настоящим изобретением, также как и меченые антитела с или без маркеров для детекции, обладающие способностью к детектированию BoNT/C. Набор также может содержать сопутствующие компоненты, такие как, но не ограниченные буферами, метками, контейнерами, вставками, тюбингами, флаконами, шприцами и подобными предметами.
Для терапии, применяемой к людям, необходимо использовать человеческие изотипы для того, чтобы минимизировать антиглобулинный ответ в течение этой терапии. Последовательности ДНК человеческих константных доменов, предпочтительно в сочетании с каркасными участками вариабельных доменов могут быть получены в соответствии с хорошо известными методиками. Примером такой методики является методика CAMPΑΤΗ 1Η, доступная у компании Burroughs Wellcome Ltd.
Антитело или Fab, обладающее способностью к связыванию с BoNT/C, согласно настоящему изобретению, может содержаться в составе медицинской рецептуры. Например, антитело или Fab может быть объединено с буфером, в котором указанное антитело или Fab растворено, и/или с неким носителем. Буферы и носители, пригодные для этого, известны специалистам в данной области техники. Примерами таких буферов являются буферы, в которых указанное антитело или Fab может функционировать, селективно связывая BoNT/C, такие как, но не ограничивающиеся солевым фосфатным буфером, водой, салином, фосфатным буфером, буфером HEPES (солевой буфер N-2-гидроксиэтилпиперазин-N'-2-этансульфоновой кислоты), буфером TES (солевым буфером Трис-ЭДТА), буфером Трис и буфером ТАЕ (Трис-ацетат-ЭДТА). Примерами носителей являются полимерные матрицы, токсоиды, сывороточные альбумины, такие как бычий сывороточный альбумин, но не ограничены ими. Носители могут быть комбинированы с антителами или Fab и конъюгированными (то есть, присоединенными) с антителами или Fab так, что они несущественно влияют на способность антитела или Fab к селективному связыванию с BoNT/C.
Методы получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации, подбора олигонуклеотидов в качестве праймеров и подобные методы могут быть стандартными методами, хорошо известными специалисту в данной области техники. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, T., ("Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)).
Последующие примеры приведены для целей объяснения и не ограничивают каким-либо образом рамки настоящего изобретения.
Пример 1. Получение мышиных моноклональных антител против BoNT/Cis.
Иммунизация
Мыши линии Balb/c в возрасте 8 недель были дважды с интервалом в 2 недели иммунизированы в подушечки задних лап иммуногеном следующего состава: нетоксичный аналог ботулинического токсина С (BoNT/Cis) 20 мкг в фосфатно-солевом буфере (ФСБ) эмульгировали с эквивалентным объемом адъюванта Фройнда. Для первой иммунизации использовали смесь 1:1 антигена и полным адъювантом Фройнда (complete Freund's adjuvant) в форме гомогенной эмульсии. Антиген для второй иммунизации был приготовлен похожим образом, но только с неполным адъювантом Фройнда.
Получение гибридом
Через 3 дня после второй иммунизации сыворотки иммунных мышей тестировали методом непрямого иммуноферментного анализа на содержание антител к BoNT/Cis. В тот же день выделяли иммунные лимфоциты из подколенных лимфоузлов и гибридизовали их с клетками мышиной миеломы sp2/0 по стандартной методике с использованием ПЭГ 4000.
На 7-й день после гибридизации супернатанты гибридом тестировали методом непрямого ИФА с использованием нетоксичного аналога BoNT/Cis. Позитивные клоны клонировали 2-4 раза методом предельных разведений для получения стабильных продуцентов антител. Антитела нарабатывали в асцитных жидкостях мышей. Всего было получено 10 моноклональных антител (МоАт).
Очистка моноклональных антител против BoNT/Cis
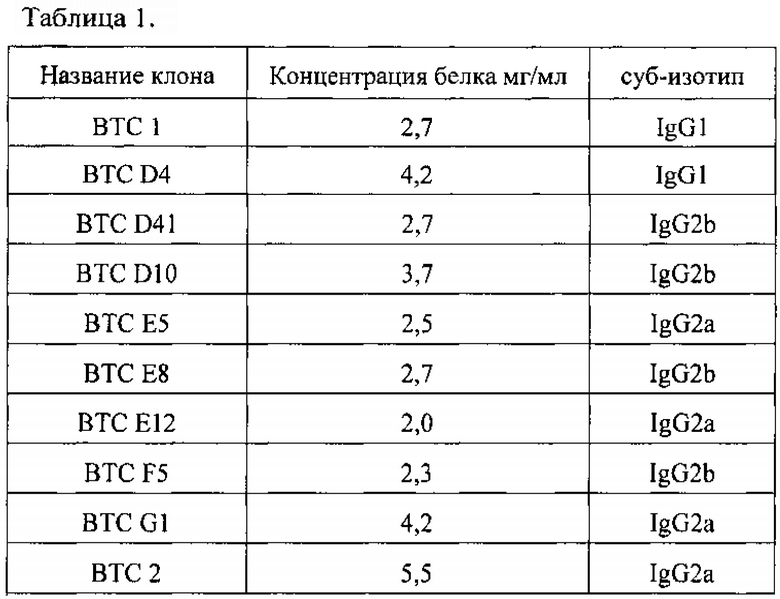
Для иммунохимической характеристики все МоАт были выделены из соответствующих асцитных жидкостей методом аффинной хроматографии на колонке с белок G - сефарозой и переведены диализом в ФСБ (Andrew S.M. Purification of Immunoglobulin G / S.M. Andrew, J.A. Titus // Current Protocols in Immunology. Chapter 2. Unit 2. 2001). Концентрацию антител определяли на спектрофотометре при длине волны 280 нм, коэффициент экстинкции для 1 мг/мл иммуноглобулинов равен 1.4 (Howard G.C. Making and Using Antibodies: A Practical Handbook / G.C. Howard, M.R. Kaser // A Practical Handbook. Second Edition, CRC Press. 2013). Данные по характеристики моноклональных антител приведены в Таблице 1. Суб-изотипы тяжелых цепей мышиных антител определяли при помощи набора ISO-2 (Sigma, США).
Степень чистоты препаратов МоАт контролировали при помощи электрофореза в 12% полиакриламидном геле в присутствии SDS и DTT в ступенчатой буферной системе Лэммли (Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4 / U.K. Laemmli // Nature. 1970. V. 227. P. 680-685). Использовали систему для вертикального электрофореза Bio-Rad Mini-Protean 3 cell (Bio-RAD, США). На каждую дорожку наносили по 3 мкг исследуемого образца (по белку), окрашивание белковых полос проводили Coomassie brilliant blue R250. Результаты электрофореза приведены на Фиг. 1.
Как видно из приведенных данных, полученные антитела имеют изотип G, их чистота составила не менее 95%.
Пример 2. Характеристика моноклональных антител в иммуноферментном анализе с разделением компонентов (непрямой ИФА).
BoNT/CiS сорбировали в концентрации 2 мкг/мл при температуре 4°С в течение ночи в ФСБ (0.01М KH2PO4, 0,1М NaCl), рН 7.2-7.4 на 96-луночные планшеты с высокой связывающей способностью. Все антитела титровали в диапазоне концентраций от 3000 нг/мл до 100 нг/мл в ФСБ-AT (0,01М KH2PO4, 0,1М NaCl; 0,2% бычий сывороточный альбумин; 0,1% Твин 20). В качестве конъюгата использовали антивидовой пероксидазный конъюгат антител против IgG мыши в ФСБ-АТ. Результаты приведены в Таблице 2.
Как видно из приведенных данных, антитела ВТС1, D10, Е5, F5 и G1 узнают сорбированный на твердой фазе токсин, что вполне соответствует данным об изменении конформации BoNT/C при сорбции.
Пример 3. Характеристика моноклональных антител методом захвата по биотинилированным BoNT/CiS.
Все МоАт сорбировали на планшет в концентрации 2,5 мкг/мл в 0,1М карбонатном буфере рН 9,3. BoNT/CiS - биотин разводили и титровали в буфере PBS - AT в диапазоне концентраций от 300 нг/мл до 10 нг/мл. В качестве конъюгата использовали коммерческий препарат СА-ПХ. Конъюгат разводили в буфере PBS-AT до концентрации 150 нг/мл. Все инкубации проводились при комнатной температуре. Результаты приведены в Таблице 3.

Как видно из приведенных данных, антитела ВТС1, D4, D10, Е5, F5, G1, ВТС2 реагируют с BoNT/CiS - биотин и могут быть использованы для конструирования тест-системы.
Пример 4. Разработка и оптимизация сэндвич ИФА для определения BoNT/CiS.
Протокол ИФА для построения калибровочной кривой.
По результатам ИФА, периодатным методом, моноклональные антитела ВТС 1, ВТС D10, ВТС F5, ВТС G1 были конъюгированы с пероксидазой хрена (Tijssen Р. Practice and theory of enzyme immunoassays / P. Tijssen, Elsevier, 1985). Все антитела сорбировали на планшет с концентрацией 2,5 мкг/мл в 0,1М карбонатном буфере в течение ночи при 4°С, и все конъюгаты были раститрованы от 1000 с шагом 3. Концентрация BoNT/Cis в данном эксперименте была 50 нг/мл. Результаты «спаривания» МоАт приведены в Таблице 4.

При разработке сэндвича была проведена его оптимизация по нескольким параметрам (подбор буфера для сорбции МоАт, концентрация МоАт для сорбции, разведение конъюгата, были выбраны оптимальная температура и время проведения анализа). В результате проведенных экспериментов было выбрано 6 пар антител.
Протокол ИФА для построения калибровочной кривой
а. Поликлональные антитела козы против антител мыши, истощенные на Fab2 фрагментах антител ВТС В2 сорбировали в концентрации 10 мкг/мл при температуре 4°С в течение ночи в ФСБ (0,01М KH2PO4, 0,1М NaCl), рН 7,2-7,4 на 96-луночные планшеты с высокой связывающей способностью, производства Corning-Costar, Нидерланды, кат. номер 9018.
б. Планшет отмывали три раза, ФСБ-Т (0,01M KH2PO4, 0,1М NaCl; 0,1% Твин 20), 200 мкл в лунку.
в. Для блокировки, в каждую лунку добавляли по 200 мкл 1%-го бычьего сывороточного альбумина в ФСБ. Инкубировали 1 час при комнатной температуре на шейкере.
г. Планшет отмывали три раза, ФСБ-Т (0,01М KH2PO4, 0,1М NaCl; 0,1% Твин 20), 200 мкл в лунку.
д. МоАт ВТС Е5 или ВТС 2 вносили в лунки в концентрации 5 мкг/мл в ФСБ-AT (0,01M KH2PO4, 0,1М NaCl; 0,2% бычий сывороточный альбумин; 0,1% Твин 20). Инкубировали 1 час при комнатной температуре на шейкере.
е. BoNT/CiS титровали в диапазоне концентраций от 400 нг/мл до 6,25 нг/мл в ФСБ-AT (0,01М KH2PO4,0,1М NaCl; 0,2% бычий сывороточный альбумин; 0,1% Твин 20). Инкубировали 1 час при комнатной температуре на шейкере.
ж. Планшет отмывали три раза, ФСБ-Т (0,01М KH2PO4, 0,1М NaCl; 0,1% Твин 20) 200 мкл в лунку.
з. В каждую лунку добавляли пероксидазный конъюгат антител ВТС1 Fab2 в ФСБ-АТ. Инкубировали 1 час.
и. Планшет отмывали три раза, ФСБ-Т (0,1% Твин 20), 200 мкл в лунку.
к. В каждую лунку добавляли по 100 мкл ТМБ субстрат (3,3,5,5-Тетраметил-бензидин). Инкубировали 30 мину при комнатной температуре на шейкере.
л. Реакцию останавливали добавлением 0,5М серной кислоты по 100 мкл в лунку.
м. Оптическую плотность измеряли при 450 нм.
Результаты ИФА показаны в Таблице 5 и на Фиг. 2.
Всего было обнаружено 6 пар антител, составляющих «сэндвич» в иммуноферментном анализе, следовательно, существует по меньшей мере две антигенные детерминанты, к которым получены антитела.
Вся панель антител против BoNT/CiS, за исключением антитела Е12, была протестирована на предмет узнавания BoNT/C. Для тестирования был использован «сэндвич» ИФА, где в качестве сорбированных на твердую фазу антител брали поликлональную гипериммунную сыворотку лошади (fzmb GmbH, Бад-Лангензальца, Германия). Результаты показали, что все протестированные антитела эффективно узнают BoNT/C.
Для дальнейшей работы были взяты МоАт ВТС1 и ВТС2, составляющие наиболее чувствительную пару в «сэндвич» ИФА.
Пример 5. Амплификация VH:VL генов клеточных линий ВТС1 и ВТС2, продуцирующих антитела класса IgG против BoNT/C, и определение их последовательностей аминокислот.
Лизис клеток
Около 300 клеток линии ВТС1 и линии ВТС2, продуцирующих антитела класса IgG, осаждали центрифугированием при 600 g в течение 5 мин в пробирках типа «Эппендорф» объемом 1,5 мл. Супернатант удаляли пипеткой, после чего пробирки помещали в жидкий азот, что приводило к лизису клеток.
Обратная транскрипция
Пробирки размораживали в ледяной бане в течение 3 мин, после чего клеточный лизат растворяли в 20 мкл реакционной смеси следующего состава:
Реакцию обратной транскрипции проводили при 52°С в течение 20 мин.
Специфичные праймеры на константные домены типа G1 тяжелой цепи и kappa легкой цепи, выбранные с помощью программного обеспечения Vector NTI® Advance software, Version 10.0, 2008 Invitrogen Corporation, приведены в Перечне последовательностей под номерами SEQ ID NO: 17 (праймер MHG1RT) и 18 (праймер MLKRT).
Очистка препарата на колонках QIAquick
Полученная после обратной транскрипции кДНК была очищена от олигонуклеотидов и дезокситрифосфатов с использованием колонок QIAquick.
Для этого в пробирку с 20 мкл кДНК добавляли 60 мкл буфера QG. Добавляли 20 мкл изопропанола, после чего раствор перемешивали пипетированием. Колонку QIAquick помещали в 2 мл пробирку. На колонку наносили 100 мкл образца и центрифугировали 1 мин при 12000 g. Из 2 мл пробирки удаляли «проскок», после чего наносили на колонку 0.75 мл буфера РЕ и центрифугировали 1 мин при 12000 g. Из 2 мл пробирки удаляли «проскок», после чего наносили на колонку 0.4 мл буфера РЕ и центрифугировали 1 мин при 12000 g. Из 2 мл пробирки удаляли «проскок», и центрифугировали 1 мин при 12000 g. Перемещали колонку QIAquick в чистую 1.5 мл пробирку. Для элюции к ДНК, в центр колонки наносили 25 мкл воды, свободной от нуклеаз, и центрифугировали 1 мин при 12000 g. Пробирку с элюатом закрывали крышкой и помещали в ледяную баню, после чего проводили дефосфорилирование кДНК.
Дефосфорилирование
Для этого, 11.5 мкл препарата очищенной кДНК помещали в 500 мкл пробирку и добавляли 1.4 мкл 10х буфера «В» и 1 мкл щелочной фосфатазы. Реакцию дефосфорилирования проводили в течение 10 мин при 37°С, после чего останавливали нагреванием до 75°С в течение 5 минут.
Лигирование с адаптером
Полученную дефосфорилированную кДНК лигировали с 5'-фосфорилированным олигонуклеотидом ADAPT (SEQ ID NO: 19).
Для этого к 14 мкл препарата дефосфорилированной кДНК добавляли 6 мкл реакционной смеси следующего состава:
Реакционную смесь инкубировали 30 мин при 37°С, после чего проводили очистку препарата лигированной с олигонуклеотидом кДНК от солей, олигонуклеотидов и коротких ДНК-фрагментов на колонках MiniSpin S-400 HR.
Очистка препарата на колонках MiniSpin S-400 HR
Для этого объем лигазной смеси доводили до 30 мкл водой, свободной от РНКаз. Ресуспендировали матрикс в колонке MiniSpin S-400 HR с помощью «вортекса». Откручивали крышку колонки на !4 оборота и обламывали кончик колонки. Колонку помещали в 2 мл пробирку и центрифугировали 1 мин при 735 g. Перемещали колонку MiniSpin S-400 HR в чистую 1.5 мл пробирку. Осторожно наносили 30 мкл образца в центр матрикса колонки, не нарушая его структуру. Центрифугировали 2 мин при 735 g. Пробирку с очищенным образцом закрывали крышкой и помещали в ледяную баню.
Первичная ПЦР
Очищенный препарат был использован в качестве матрицы для первичной ПЦР, в которой одновременно амплифицировались фрагменты легкой и тяжелой цепей ДНК, имеющие сайты отжига прямого праймера AD1 (SEQ ID NO: 20), комплементарного адаптеру ADAPT, и обратных праймеров MLKR1 (SEQ ID NO: 21) и MHG1R1 (SEQ ID NO: 22), комплементарных константным доменам легкой и тяжелой цепей ДНК генов иммуноглобулинов соответственно.
Реакционная смесь одной пробы имела следующий состав:
Амплификацию проводили на приборе Bio-Rad S1000 Thermal Cycler CFX96 в 96-луночных низкопрофильных плашках Bio-Rad MLL9651.
Использовали следующую программу ПЦР:
23 цикла:
Пробы из первичной ПЦР разводили добавлением 130 мкл Н2О. По 2 мкл из каждой пробы использовали в качестве образца для вторичной ПЦР легкой и тяжелой цепи.
Вторичная ПЦР
Для вторичной ПЦР тяжелой цепи использовали прямой праймер AD21 (SEQ ID NO: 23), комплементарный адаптеру ADAPT и обратный праймер MHG1R2 (SEQ ID NO: 24), комплементарный константному домену тяжелой цепей ДНК генов иммуноглобулинов. Реакционная смесь одной пробы для тяжелой цепи имела следующий состав:
Для вторичной ПЦР легкой цепи использовали праймер AD2_1 (SEQ ID NO: 23) и обратный праймер MLKR2 (SEQ ID NO: 25), комплементарный константному домену kappa генов иммуноглобулинов.
Реакционная смесь одной пробы для легкой цепи имела следующий состав:
Для вторичной ПЦР использовали следующую программу:
40 циклов:
Схема расположения адаптера ADAPT, праймеров для обратной транскрипции (M…RT), первичной (AD1, M…R1) и вторичной (AD2_1, M…R2) ПЦР приведены на Фиг. 3.
Секвенирование.
После проведения вторичной ПЦР образцы анализировались методом разделения ДНК электрофорезом в агарозном геле.
Электрофорез в агарозном геле.
К пробам вторичного ПЦР добавляли 3 мкл «буфера для нанесения», после чего пробы наносили на 1,1% агарозный гель. Также наносили 3 мкл маркера длины ДНК. Разделение проводили при напряжении 6 В/см в буфере ТАЕ. Расчетный размер полной вариабельной области тяжелой цепи составляет более 480 п.н., легкой цепи более 530 п.н. Фрагменты ДНК в этой области (по 3 фрагмента для каждого гена) элюировали из геля с помощью «Gel extraction Kit» (QIAGEN).
Элюция ПЦР-фрагментов из агарозного геля.
Фрагмент агарозы, содержащий интересующую полосу ДНК, вырезали из геля скальпелем, после чего взвешивали, переносили в 1.5 мл пробирку и добавляли 3 объема буфера QG. Пробирку инкубировали при 50°С, до полного растворения агарозы, периодически перемешивая. Колонку QIAquick помещали в 2 мл пробирку. В центр колонки наносили образец, после чего центрифугировали 1 мин при 12000 g. Из 2 мл пробирки удаляли «проскок», наносили на колонку 0.75 мл буфера РЕ и центрифугировали 1 мин при 12000 g. Удаляли «проскок» и центрифугировали 1 мин при 12000 g. Перемещали колонку QIAquick в чистую 1.5 мл пробирку. Для элюции ДНК, в центр колонки наносили 15 мкл Н2О, и центрифугировали 1 мин при 12000 g. Пробирку с элюатом закрывали крышкой и помещали в ледяную баню.
Подготовка образцов для секвенирования
4 мкл элюата наносили на агарозный гель, и после электрофореза определяли концентрацию ДНК визуально. К 10 нг ПЦР-фрагментов ДНК легкой и тяжелой цепей добавляли 3.2 пкмоль праймеров MHG1R2 (SEQ ID NO: 24) или MLKR2 (SEQ ID NO: 25), соответственно, после чего образцы высушивали в термостате при 65°С, и передавали в Центр коллективного пользования «ГЕНОМ» Института молекулярной биологии им. В.А. Энгельгардта для секвенирования.
Анализ результатов секвенирования.
Полученные хроматограммы анализировали с помощью программного обеспечения Chromas ver. 1.45. По результатам секвенирования были определены нуклеотидные последовательности VL:VH фрагментов генов иммуноглобулинов. Результаты представлены на Фиг. 4. Последовательность аминокислот вариабельного домена тяжелой цепи (VH) антитела ВТС1 приведена в Перечне последовательностей под номером SEQ ID NO: 1. Последовательность аминокислот вариабельного домена легкой цепи (VH) антитела ВТС1 приведена в Перечне последовательностей под номером SEQ ID NO: 2. Нуклеотидные последовательности, кодирующие указанные домены, приведены под номерами SEQ ID NO: 3 и 4, соответственно. Последовательность аминокислот вариабельного домена тяжелой цепи (VH) антитела ВТС2 приведена в Перечне последовательностей под номером SEQ ID NO: 5. Последовательность аминокислот вариабельного домена легкой цепи (VH) антитела ВТС2 приведена в Перечне последовательностей под номером SEQ ID NO: 6. Нуклеотидные последовательности, кодирующие указанные домены, приведены под номерами SEQ ID NO: 7 и 8, соответственно.
Пример 6. Создание экспрессионных кассет, содержащих легкие и тяжелые цепи рекомбинантных гуманизированных Fab-фрагментов против BoNT/C.
Для экспрессии генов, кодирующих гуманизированные Fab-фрагменты против ботулинического нейротоксина типа С, использовали конструкции на основе плазмид pPICZαA-AR-H и pPICZαA-ΔPmeI-AR-L (Вестник Московского Университета. Химия 2010, том 51, №3, стр. 185-190), несущих гуманизированные вариабельные домены мышиных антител против вируса бешенства, слитые с оптимизированными, для экспрессии в Pichia pastoris, константными доменами СН1 и С-каппа иммуноглобулинов IgG1 Homo sapiens в экспрессионном векторе pPICZαA (https://tools.thermofisher.com/content/sfs/manuals/ppiczalpha_man.pdf).
Плазмида pPICZαA-AR-H содержит вариабельный домен тяжелой цепи антитела, который может «вырезаться» по сайтам рестрикции XhoI (в конце последовательности, кодирующей сигнальный пептид) и ApaI (в начале последовательности, кодирующей оптимизированный СН1-домен H.sapiens). Кроме того, в конце константного домена присутствует область, кодирующая последовательность полигистидинов (6xHis) tag.
Плазмида pPICZαA-AR-L содержит вариабельный домен легкой цепи антитела, который может «вырезаться» по сайтам рестрикции XhoI (в конце последовательности, кодирующей сигнальный пептид) и Pfl23II (на стыке вариабельного домена и оптимизированного С-каппа домена Н.sapiens). Кроме того, в промоторе АОХ1 удален сайт рестрикции MssI.
Синтезировали следующие олигонуклеотидные праймеры: MHG1XhoD (SEQ ID NO: 26), MHG1ApaR (SEQ ID NO: 27), MHG2AXhoD (SEQ ID NO: 28), MHG2AApaR (SEQ ID NO: 29), MLKG1XhoD (SEQ ID NO: 30), MLKG1PflR (SEQ ID NO: 31), MLKG2AXhoD (SEQ ID NO: 32), MLKG2APflR (SEQ ID NO: 33).
Прямой праймер MHG1XhoD (SEQ ID NO: 26) содержит последовательность, 3'-область которой комплементарна началу вариабельного домена тяжелой цепи антитела против BoNT/C ВТС1, а 5'-область комплементарна концу последовательности, кодирующей сигнальный пептид экспрессионного вектора и содержит сайт рестрикции XhoI.
Обратный праймер MHG1ApaR (SEQ ID NO: 27) содержит последовательность, 3'-область которой обратно комплементарна концу вариабельного домена тяжелой цепи антитела против BoNT/C ВТС1, а 5'-область обратно комплементарна началу последовательности, кодирующей оптимизированный СН1-домен Н.sapiens и содержит сайт рестрикции ApaI.
Прямой праймер MLKG1XhoD (SEQ ID NO: 30) содержит последовательность, 3'-область которой комплементарна началу вариабельного домена легкой цепи антитела против BoNT/C ВТС1, а 5'-область комплементарна концу последовательности, кодирующей сигнальный пептид экспрессионного вектора и содержит сайт рестрикции XhoI.
Обратный праймер MLKG1PflR (SEQ ID NO: 31) содержит последовательность, 3'-область которой обратно комплементарна концу вариабельного домена легкой цепи антитела против BoNT/C ВТС1, а 5'-область содержит сайт рестрикции Pfl23II.
Прямой праймер MHG2AXhoD (SEQ ID NO: 28) содержит последовательность, 3'-область которой комплементарна началу вариабельного домена тяжелой цепи антитела против BoNT/C ВТС2, а 5'-область комплементарна концу последовательности, кодирующей сигнальный пептид экспрессионного вектора и содержит сайт рестрикции XhoI.
Обратный праймер MHG2AApaR (SEQ ID NO: 29) содержит последовательность, 3'-область которой обратно комплементарна концу вариабельного домена тяжелой цепи антитела против BoNT/C ВТС2, а 5'-область обратно комплементарна началу последовательности, кодирующей оптимизированный СН1-домен Н.sapiens и содержит сайт рестрикции ApaI.
Прямой праймер MLKG2AXhoD (SEQ ID NO: 32) содержит последовательность, 3'-область которой комплементарна началу вариабельного домена легкой цепи антитела против BoNT/C ВТС2, а 5'-область комплементарна концу последовательности, кодирующей сигнальный пептид экспрессионного вектора и содержит сайт рестрикции XhoI.
Обратный праймер MLKG2APAR (SEQ ID NO: 33) содержит последовательность, 3'-область которой обратно комплементарна концу вариабельного домена легкой цепи антитела против BoNT/C ВТС2, а 5'-область содержит сайт рестрикции Pfl23II.
Пары праймеров на вариабельные домены легких и тяжелых цепей были использованы для проведения высокоточной ПЦР, в качестве матриц для которых использовались ПЦР-фрагменты генов вариабельных доменов, полученных в Примере 5.
Использовали следующую программу ПЦР:
25 циклов:
К пробам ПЦР добавляли 3 мкл «буфера для нанесения», после чего пробы наносили на 1,2% агарозный гель. Также наносили 3 мкл маркера длины ДНК. Разделение проводили при напряжении 6 В/см в буфере ТАЕ. Фрагменты ДНК элюировали из геля с помощью «Gel extraction Kit» (QIAGEN) в соответствии с рекомендациями производителя.
Полученные ПЦР-фрагменты клонировали в плазмиды pPICZαA-AR-H и pPICZαA-ΔPmeI-AR-L вместо фрагментов плазмид, несущих вариабельные домены антител против вируса бешенства. Схема клонирования показана на Фиг. 5 и 6.
Для клонирования ПЦР-фрагментов тяжелых цепей, 1 мкг ДНК плазмиды pPICZαA-AR-H обрабатывали эндонуклеазами рестрикции XhoI и ApaI в 5х избытке, после чего рестрикционную смесь наносили на 1% агарозный гель и полосу ДНК, размером около 3,5 т.п.н., несущую фрагмент плазмиды без последовательности, кодирующей вариабельный домен тяжелой цепи элюировали из геля с помощью «Gel Extraction Kit» (QIAGEN).
По 0,5 мкг ДНК ПЦР-фрагментов генов вариабельных доменов тяжелых цепей иммуноглобулинов ВТС1 и ВТС2, несущих на своих концах последовательности, необходимые для слияния вариабельных доменов с сигнальным пептидом и с константным доменом, обрабатывали этими же эндонуклеазами в 3х избытке, после чего рестрикционную смесь очищали от солей и низкомолекулярной ДНК, возникшей при рестрикции концов ПЦР-фрагментов, используя «Gel Extraction Kit» (QIAGEN).
Полученные фрагменты ДНК использовали в реакции лигирования.
Реакционная смесь имела следующий состав:
Контрольная проба не содержала ДНК ПЦР фрагмента.
Лигирование проводили в течение ночи при температуре 14°С.
Для клонирования ПЦР-фрагментов легких цепей, 1 мкг ДНК плазмиды pPICZαA-ΔPmeI-AR-L обрабатывали эндонуклеазами рестрикции XhoI и XbaI в 5х избытке, после чего рестрикционную смесь наносили на 1% агарозный гель и полосу ДНК, размером около 3,3 т.п.н., несущую фрагмент плазмиды без последовательности, кодирующей вариабельный домен легкой цепи и константный домен, элюировали из геля с помощью «Gel Extraction Kit» (QIAGEN).
Около 3 мкг ДНК плазмиды pPICZαA-ΔPmeI-AR-L обрабатывали эндонуклеазами рестрикции Pfl23II и XbaI в 3х избытке, после чего рестрикционную смесь наносили на 1,3% агарозный гель, и полосу ДНК, размером около 320 п.н., несущую фрагмент плазмиды, кодирующий константный С-каппа домен, элюировали из геля с помощью «Gel Extraction Kit» (QIAGEN).
По 0,5 мкг ДНК ПЦР-фрагментов генов вариабельных доменов легких цепей иммуноглобулинов ВТС1 и ВТС2, несущих на своих концах последовательности, необходимые для слияния вариабельных доменов с сигнальным пептидом и с константным доменом, обрабатывали эндонуклеазами XhoI и Pfl23II в 3х избытке, после чего рестрикционную смесь очищали от солей и низкомолекулярной ДНК, возникшей при рестрикции концов ПЦР-фрагментов, используя «Gel Extraction Kit» (QIAGEN).
Полученные фрагменты ДНК использовали в реакции лигирования.
Реакционная смесь имела следующий состав:
Контрольная проба не содержала ДНК ПЦР фрагмента и фрагмента плазмиды, кодирующего константный С-каппа домен.
Лигирование проводили в течение ночи при температуре 14°С.
По 4 мкл лигазных смесей трансформировали в клетки штамма DH5α Ε.coli (ттт), как описано в Molecular cloning: a laboratory manual (4th ed. Michael R. Green, Joseph Sambrook. Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press, 2012), после чего клетки высевали на чашки с агаризованной средой Low Salt LB (https://tools.thermofisher.com/content/sfs/manuals/ppiczalpha_man.pdf), содержащей селективный антибиотик Zeocin™ в концентрации 25 мкг/мл.
По 4 клона из каждой трансформации выращивали в 10 мл жидкой среды Low Salt LB в присутствии Zeocin™ в концентрации 25 мкг/мл, после чего, выделяли плазмидную ДНК щелочным методом (Molecular cloning: a laboratory manual. 4th ed. Michael R. Green, Joseph Sambrook. Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press, c2012).
Полученные плазмидные ДНК проверяли рестрикцией на восстановление сайтов клонирования. Плазмиды, с клонированными тяжелыми цепями проверяли рестрикцией по сайтам XhoI и ApaI. Плазмиды, с клонированными легкими цепями проверяли рестрикцией по сайтам XhoI-XbaI, и рестрикцией по сайтам Pfl23II-XbaI.
По одной плазмиде для каждого гена вариабельного домена выбирали для секвенирования с праймера 5`AΟΧ1 (SEQ ID NO: 34) на промотор гена АОХ1.
Анализ показал отсутствие ошибок в клонированных ПЦР-фрагментах ДНК вариабельных доменов тяжелых и легких цепей Fab иммуноглобулинов ВТС1 и ВТС2, а также правильность сборки экспрессионных векторов.
Пример 7. Получение экспрессионных конструкций, несущих одновременно легкие и тяжелые цепи рекомбинантных гуманизированных Fab против BoNT/C.
Выбранные секвенированные плазмиды рВТС1_Н, pBTC1_L, рВТС2_Н и pBTC2_L использовали для сборки экспрессионных конструкций, содержащих одновременно легкие и тяжелые цепи рекомбинантных гуманизированных Fab против BoNT/C. Карты плазмид представлены на Фиг. 7 и 8.
ДНК 0,5 мкг плазмид pBTC1_L и pBTC2_L, несущих гены легких цепей Fab против BoNT/C, линеаризовали рестрикцией 5х избытком эндонуклеазы BamHI, после чего в рестрикционную смесь добавляли 1 мкл (1 ед.) щелочной фосфатазы FastAP. Смесь инкубировали 15 минут при 37°С, после чего инактивировали фосфатазу нагреванием в течение 5 минут при 75°С.
К реакционной смеси добавляли 1/5 объема 6х-го «буфера для нанесения», после чего пробы наносили на 1% агарозный гель. Линеаризованную дефосфорилированную плазмидную ДНК элюировали из геля с помощью «Gel Extraction Kit».
По 1,5 мкг ДНК плазмид рВТС1_Н и рВТС2_Н, несущих гены тяжелых цепей Fab против BoNT/C обрабатывали 3х кратным избытком эндонуклеаз BglII, BamHI и Eco32I. Рестрикционную смесь наносили вместе с «буфером для нанесения» на 1% агарозный гель и после проведения электрофореза элюировали BglII-BamHI фрагмент плазмиды размером около 2500 п.н., несущий ген тяжелой цепи Fab-фрагмента с сигнальной последовательностью под контролем промотора АОХ1 и терминатор этого гена.
Полученные фрагменты ДНК использовались в реакции лигирования.
Реакционная смесь имела следующий состав:
Контрольная проба не содержала фрагмента ДНК плазмид рВТС1_Н и РВТС2_Н.
Лигирование проводили в течение ночи при температуре 14°С.
По 4 мкл лигазных смесей трансформировали в клетки штамма DH5α Е.coli, после чего клетки высевали на чашки с агаризованной средой Low Salt LB, содержащей селективный антибиотик Zeocin™ в концентрации 25 мкг/мл.
По 8 клонов из каждой трансформации выращивали в 10 мл жидкой среды Low Salt LB в присутствии Zeocin™ в концентрации 25 мкг/мл, после чего выделяли плазмидную ДНК щелочным методом.
Полученные плазмидные ДНК проверяли рестрикцией на восстановление сайта BamHI, с одновременным контролем размера плазмидной ДНК.
Были отобраны по 5 плазмидных ДНК для каждого Fab, в которых определяли ориентацию гена тяжелой цепи рестрикцией по сайту XbaI.
По одной плазмиде для каждого Fab (ВТС1 и ВТС2), с однонаправленными генами легкой и тяжелой цепей были выбраны для трансформации в P. pastoris. Карты плазмид pBTC1 и рВТС2 представлены на Фиг. 9.
Пример 8. Получение трансформантов дрожжей и анализ продукции антител.
Экспрессионные плазмиды pBTC1 и рВТС2, линеаризованные с помощью рестриктазы Mssl, вводили в клетки GS115 P.pastoris в соответствии с протоколом производителя INVITROGEN с помощью методики трансформации электропорацией (прибор MicroPulser фирмы Bio-Rad). Трансформанты отбирали на питательной среде YPDS (Invitrogen), содержащей глюкозу и Zeocin™ (400 мкг/мл). Интеграцию линеаризованной плазмиды в хромосому подтверждали методом ПЦР с использованием ген-специфичных праймеров для легкой цепи (SEQ ID NO: 32 и 33) и для тяжелой цепи (SEQ ID NO: 32 и 29) в случае плазмиды рВТС1 и ген-специфичных праймеров для легкой церии (SEQ ID NO: 30 и 31) и для тяжелой цепи (SEQ ID NO: 26 и 27) в случае плазмиды рВТС2 и геномной ДНК трансформантов в качестве матрицы.
Отобранные трансформанты переносили на питательную среду BMMY (Invitrogen), содержащую метанол, для индукции экспрессии генов, кодирующих антитело против BoNT/C. Клоны штаммов GS115/ pBTC1 и GS115/ рВТС2 выращивали в пробирках (V=4 мл) в течение 45 часов при 30°С и рН 6.0 (конечная оптическая плотность - OD 40). Измерение количества антител в культуральной жидкости осуществляли методом ELISA. В соответствии с результатами анализа были выбраны штаммы, продуцирующие до 150 мг/л гуманизированного Fab против BoNT/C.
Пример 9. Очистка и иммунохимическая характеристика рекомбинантного гуманизированного Fab ВТС1 против BoNT-C.
Полученные экспериментальные партии гуманизированных Fab ВТС1.1, ВТС1.2, ВТС1.3 характеризовали по параметрам чистоты и специфической активности. Электрофорез ВТС 1.1, ВТС 1.2, ВТС 1.3 проводили в 12% SDS-PAGE в невосстанавливающих и восстанавливающих условиях в ступенчатой буферной системе Лэммли [Laemmli UK (August 1970). "Cleavage of structural proteins during the assembly of the head of bacteriophage T4". Nature 227 (5259): 680-685]. Использовали систему для вертикального электрофореза Bio-Rad Mini-Protean 3 cell (Bio-RAD, США). На каждую дорожку наносили по 5 мкг исследуемого образца (по белку), окрашивание белковых полос проводили Coomassie brilliant blue R250. Результаты электрофореза приведены на Фиг. 10.
Как следует из представленных данных, все серии экспериментальных партий в невосстанавливающих условиях имеют одну мажорную полосу с подвижностью около 48 кДа, что соответствует молекулярной массе мономерной формы Fab фрагмента и доказывает подлинность целевого продукта. По данным 12% SDS-PAGE чистота экспериментальных образцов составляет не менее 80%.
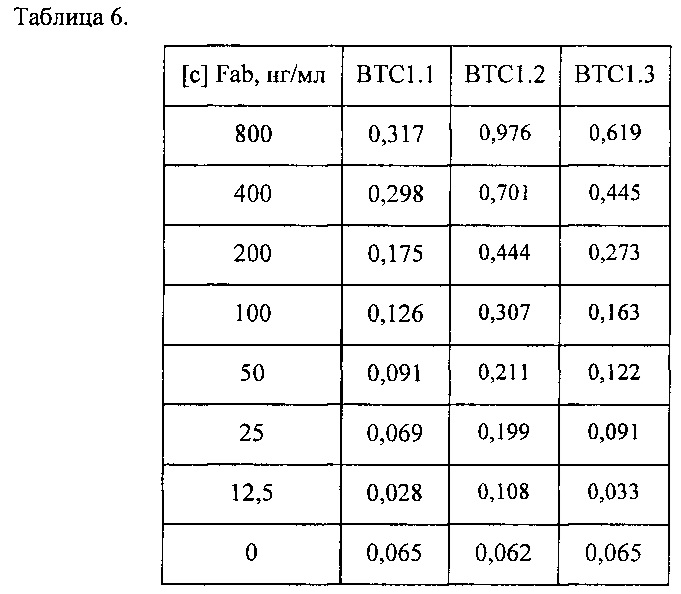
Для оценки специфичности полученных рекомбинантных гуманизированных Fab проводили непрямой ИФА с иммобилизованным BoNT/CiS. Сорбцию Fab проводили при температуре 4°С в течение ночи. BoNT/CiS сорбировали в концентрации 1 мкг/мл в ФСБ на 96-луночные планшеты с высокой связывающей способностью производства Corning-Costar, Нидерланды, кат. номер 9018. Результаты приведены в Таблице 6 и на Фиг. 11.
Для подтверждения подлинности рекомбинантных Fab фрагментов против BoNT/CiS проводили 12% SDS-PAGE в невосстанавливающих (дорожки 1-5) и восстанавливающих (дорожки 6-10) условиях с последующим иммуноблотом с пероксидазными конъюгатами МоАт против каппа-цепи Ig человека и метки из шести остатков гистидина (Фиг. 12). Известно, что Fab-фрагмент состоит из тяжелой (VH+СН1) и легкой (VL + Сκ) цепей, имеющих практически одинаковую молекулярную массу около 25 кДа и соединенных межцепочечной дисульфидной связью. Таким образом, используемые для проявления иммуноблота МоАт должны выявлять одну полосу в районе 25 кДа в восстанавливающих условиях и одну полосу в районе 50 кДа в невосстанавливающих условиях. Результаты иммуноблота, приведенные на Фиг. 12, подтверждают подлинность полученных рекомбинантных Fab фрагментов против BoNT/CiS согласно настоящему изобретению.
Хотя настоящее изобретение было подробно описано со ссылкой на предпочтительные варианты его осуществления, для специалиста в данной области техники ясно, что могут быть сделаны различные замены и применены эквиваленты, которые не выходят за рамки настоящего изобретения. Все процитированные здесь документы являются частью настоящей заявки, включенные путем ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ (Fab) ПРОТИВ ВИРУСА БЕШЕНСТВА, ИЗОЛИРОВАННЫЙ ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА, КЛЕТКА ДРОЖЖЕЙ, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2010 |
|
RU2440412C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА И Fab, СВЯЗЫВАЮЩИЕСЯ С АНТИГЕНОМ F1 ИЗ Yersinia pestis, И СПОСОБ ИХ ПОЛУЧЕНИЯ С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2009 |
|
RU2420587C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К ФНО-α, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab) И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2556815C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКОВ В ДВУХЦЕПОЧЕЧНОЙ ФОРМЕ С ДИСУЛЬФИДНЫМ МОСТИКОМ | 2006 |
|
RU2412253C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА ТИПА А НА ОСНОВЕ ИММУНОДЕТЕКЦИИ, СОПРЯЖЕННОЙ С ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИЕЙ | 2013 |
|
RU2549463C1 |
| СРЕДСТВА И СПОСОБЫ ДЛЯ ОПРЕДЕЛЕНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДОВ НЕЙРОТОКСИНА В КЛЕТКАХ | 2014 |
|
RU2704808C2 |
| Однодоменное антитело и его модификации, специфически связывающиеся с ботулиническим нейротоксином типа А, и способ их применения для терапии или экстренной профилактики интоксикации, вызванной ботулиническим нейротоксином типа А. | 2021 |
|
RU2766348C1 |
| ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ЛЕГКОЙ И ТЯЖЕЛОЙ ЦЕПИ МЫШИНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ АЛЬФА (ФНО-α) ЧЕЛОВЕКА (ВАРИАНТЫ), АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab) ПРОТИВ ФНО-α ЧЕЛОВЕКА, СОДЕРЖАЩИЙ УКАЗАННЫЕ ДОМЕНЫ (ВАРИАНТЫ) | 2012 |
|
RU2499000C1 |
| НЕЙРОТОКСИНЫ, ПРОЯВЛЯЮЩИЕ УКОРОЧЕННУЮ БИОЛОГИЧЕСКУЮ АКТИВНОСТЬ | 2012 |
|
RU2646110C2 |




















Изобретение относится к области биохимии. Описаны антиген-связывающие фрагменты (Fab), селективно связывающиеся с ботулиническим нейротоксином С, в частности гуманизированные Fab. Представлены клетки дрожжей, продуцирующие указанные гуманизированные Fab. Предложен способ получения описанных гуманизированных Fab. Также предложены способ детекции ботулинического нейротоксина С и набор для детекции ботулинического нейротоксина С. Изобретение позволяет получить реагенты для детекции ботулинического нейротоксина С, обладающие высокой специфичностью связывания с указанным нейротоксином. 8 н. и 1 з.п. ф-лы, 12 ил., 6 табл., 9 пр.
1. Антиген-связывающий фрагмент (Fab), селективно связывающийся с ботулиническим нейротоксином С, который включает в себя вариабельный домен тяжелой цепи (VH) с последовательностью аминокислот SEQ ID NO: l или 5 и вариабельный домен легкой цепи (VL) с последовательностью аминокислот SEQ ID NO: 2 или 6 соответственно.
2. Изолированный фрагмент ДНК, кодирующий антигенсвязывающий фрагмент (Fab) по п. 1.
3. Гуманизированный Fab, селективно связывающийся с ботулиническим нейротоксином С, отличающееся тем, что тяжелая цепь указанного Fab включает в себя последовательность аминокислот SEQ ID NO: 9 или 13, а легкая цепь указанного Fab включает в себя последовательность аминокислот SEQ ID NO: 10 или 14 соответственно.
4. Изолированный фрагмент ДНК, кодирующий гуманизированный Fab по п. 3.
5. Клетка дрожжей, трансформированная фрагментом ДНК по п. 4 и обладающая способностью к продукции гуманизированного Fab по п. 3.
6. Клетка дрожжей по п. 5, отличающаяся тем, что указанными дрожжами являются дрожжи Pichia pastoris.
7. Способ получения гуманизированного Fab по п. 3, включающий в себя выращивание клеток дрожжей по п. 5 в питательной среде и выделение указанного Fab из культуральной жидкости.
8. Способ детекции ботулинического нейротоксина С, включающий в себя: (а) получение образца; (b) взаимодействие указанного образца с Fab по п. 1 или 3; (с) детектирование комплекса, образованного указанным Fab, и ботулиническим нейротоксином С; и, необязательно, (d) определение количества ботулинического нейротоксина С, присутствующего в указанном образце, на основании детектированного количества комплекса, образованного указанным Fab и ботулиническим нейротоксином С.
9. Набор для детекции ботулинического нейротоксина С, содержащий изолированные Fab по п. 1 или 3, которые селективно связываются с ботулиническим нейротоксином С, и средства, позволяющие детектировать указанный ботулинический нейротоксин С с использованием указанного Fab.
| US 5932449 A, 03.08.1999 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА ТИПА А НА ОСНОВЕ ИММУНОДЕТЕКЦИИ, СОПРЯЖЕННОЙ С ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИЕЙ | 2013 |
|
RU2549463C1 |
| ШТАММ ГИБРИДНЫХ КЛЕТОК ЖИВОТНЫХ Mus musculus 3F11 - ПРОДУЦЕНТ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СПЕЦИФИЧНЫХ К БОТУЛИНИЧЕСКОМУ ТОКСИНУ ТИПА B | 2014 |
|
RU2566553C1 |
| КОЛЕСНИКОВ А.В | |||
| и др., Современные подходы к разработке специфической терапии особо опасных токсикоинфекций, ВЕСТНИК РАМН, 2015, Том 70, N 4, с.428-434 | |||
| HUNTER B.F., ROSEN M.N., Detection of Clostridium botulinum Type C Cells and Toxin by the Fluorescent Antibody Technique, Avian Diseases, 1967, Vol | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

