Область техники, к которой относится изобретение
Настоящее изобретение относится к способу получения rhPTH (1-34), также известного как терипаратид, путем создания нового нуклеотида как фрагмента NcoI/XhoI, как указано в Seq. ID №1, кодирующего химерный гибридный белок, как указано в Seq. ID №2, содержащий слитый (химерный) партнер (fusion partner), состоящий из 41 аминокислоты, принадлежащих гену β-галактозидазы Escherichia coli, сайт эндонуклеазного расщепления, фрагмент гена hPTH (1-34), клонирование указанного нуклеотида в вектор экспрессии под контролем промотора Т7, трансформацию Escherichia coli указанным вектором и экспрессирование химерного гибридного белка в способе периодической ферментации с добавлением субстрата. Настоящее изобретение дополнительно относится к индуцированию лактозой с низкой скоростью подачи (питания) для оптимизированной экспрессии rhPTH (1-34) в Escherichia coli. Кроме того, настоящее изобретение дополнительно относится к уникальному, новому двухстадийному ортогональному способу очистки rhPTH (1-34), включающему катионообменную хроматографию необязательно с последующей препаративной хроматографией, выбранной из HIC (хроматография гидрофобного взаимодействия) или RT-HPLC с выходом целевого белка ≥99%. Настоящее изобретение относится к простому, рентабельному, благоприятному с экологической точки зрения способу получения высокочистого rhPTH (1-34).
Предшествующий уровень техники
Паратиреоидный гормон представляет собой пептид, состоящий из 84 аминокислот, вырабатываемый паращитовидной железой. Его физиологическая роль заключается в поддерживании кальция в сыворотке и в реконструировании костей (Dempstor D.W. et al., Endocrine Reviews, 1993, 14, 690-709). Реконструирование кости обычно представляет собой способ продолжительностью от 3 до 6 месяцев, в котором имеется сочетание резорбции кости и костеобразования. Известно, что эстрогены, витамин D, бисфосфонаты ингибируют резорбцию кости, тогда как rhPTH (1-34), анаболическое вещество, увеличивает костную массу. Остеопороз представляет собой болезнь, которая делает кость подверженной переломам, напротив rhPTH (1-34) представляет собой пептидное лекарственное средство, которое произвело революцию в лечении остеопороза. При отдельном применении RhPTH (1-34) стимулирует образование костной массы и вызывает чистое увеличение костной массы в каждом цикле реконструирования, в котором минеральная плотность костной ткани увеличивается, как правило, до 10% в год в области поясничного отдела позвоночника.
Паратиреоидный гормон (РТН) выделяется паращитовидными железами в ответ на снижение концентрации кальция в плазме и обладает некоторыми эффектами, которые восстанавливают нормальные уровни кальция. Показано, что РТН обладает и анаболическим и катаболическим действием на скелет. Длительное повышение РТН вызывает увеличение резорбции кости, тогда как периодически введенный РТН приводит к усиленному формированию кости (Canalis, E., Hock, J.M., и Raisz, L.G. (1994) in The Parathyroidds: Basic и Clinical Concepts, ed by Bilezikian, J.P., Marcus,R., и Levine, M., Raven Press Ltd., NY), хотя клеточный механизм этого двойного действия еще не ясен.
РТН биосинтезируется в виде состоящего из 115-аминокислот предшественника, препропаратиреоидного гормона (препроРТН). Несмотря на то что анаболическая активность РТН (1-84) известна, фактическое структурно-необходимое условие для наличия полной биологической активности заключено в остатках с 1 по 34 на N-конце молекулы (PNAS 68, 63-67, 1971; Endocrinology 93, 1349-1353, 1973).
РТН является чрезвычайно чувствительным к окислению, особенно при остатках метионина Met8 и Met18, и для сохранения его биоактивности нужна интактная N-концевая последовательность. Frelinger, A.L., et al., (J. Biol. Chem., (1984) 259 (9), 5507-5513) разработали способ разделения РТН с окисленным метионином от нативной молекулы и показали, что действенность окисленной молекулы значительно меньше, чем нативного РТН. Для сравнимой биоактивности с РТН полной длины РТН (1-34) нужны обе N и C-концевая спиральная конформация, как показано Jin. L. et al., (J Biol. Chem., (2000) 275 (35), 27238-44. Их модель предполагает наличие рецептор-связывающего кармана для N-конца PTH (1-34) и гидрофобную область взаимодействия с рецептором для С-конца PTH (1-34). Pellegrini M, et al., J.Biol.Chem. (1998) 273 (17), 10420-427 объясняет структуры hPTH (1-34) с высоким разрешением в водном растворе при изучении влияния рН и концентрации соли на вторичную и третичную структуры с помощью КД и ЯМР. Содержание спирали PTH (1-34), исходя из спектра CD, увеличивается в присутствии кислого буфера по сравнению с доброкачественной (benign) водой. Исследования с помощью ЯМР подтверждают наличие спиральной структуры, расположенной на N- и C-концевой части hPTH (1-34). Данная молекула (вне контекста рецепторного взаимодействия) является слишком гибкой, чтобы отдавать предпочтение какому-либо определенному вторичному или третичному свертыванию, при этом экспериментальное ограничение дистанции до 0,06Å дает форму «гибкого стебля» без рецепторного взаимодействия и "U" форму для белка при взаимодействии с рецептором. Было показано, что периодическая (прерывистая) экспозиция лиганда с его рецептором имеет предпочтительное анаболическое действие.
Клонирование и экспрессия терапевтического белка ставит в начале сложную задачу, заключающуюся в наличии подходящего генетического материала. Олигонуклеотидные праймеры, используемые для клонирования, не должны быть вырожденными, должны иметь сравнимые значения tm и в большинстве случаев располагаться в единственном местоположении в представляющем интерес гене. Обнаружено, что РНК-транскрипт гена РТН (1-84) является положительным в печени, почке, мозге и плацентарной ткани человека. Ген-специфические праймеры, используемые при полимеразной цепной реакции, пригодны в RT-PCR реакции для клонирования в кДНК-фрагмент специфической последовательности. Рекомендуется to fully sequence the insert (дополнить последовательность вставкой) до реклонирования гена. В данной области техники хорошо известно о многочисленных векторах экспрессии Escherichia coli, доступных для лигирования клонированного фрагмента, например, НВ101, JM109, BL21(DE3), ТОР10 для высокоуровневой гетерологичной экспрессии рекомбинантных белков. Пары праймеров, используемые для клонирования в tag-нуклеотидную последовательность, могут быть сконструированы так, чтобы включать сайты рестрикции для последовательных шагов клонирования, которые содержат представляющий интерес ген с сайтом протеазного разрезания и аффинную метку.
US 5496801 сообщает о препаратах hPTH, которые демонстрируют стабильность при хранении в отношении состава и стабильности гормонов. US5496801 поддерживает лиофилизацию hPTH (1-84) с маннитолом в качестве криозащитного вещества и нелетучим цитратным буфером при значении рН в пределах от 3,5 до 6,5 с получением стабильной, готовой к использованию жидкой композиции для парентерального введения.
US4086196 впервые раскрывает, что все фрагменты РТН, больше чем 1-27 hPTH, и (Ala1)-hPTH (1-27) обладают полезными биологическими свойствами, US4086196 по существу заявляет hPTH (1-Х) и Ala1 hPTH (1-Х), в которых Х представляет собой Ser или Ala.
US4086196 также сообщает о синтезе hPTH (1-34) посредством твердофазного пептидного синтеза (SPPS) с помощью хорошо известных в данной области техники способов. К тому же US4086196 раскрывает восстановление биоактивного hPTH (1-34) с помощью гель- фильтрации с последующей ионообменной хроматографией на Whatman-СМ-52 и элюированием, выполняемым вслед за линейным градиентом, использующим ионы аммония в качестве противоионов. Основными ограничениями гель-фильтрационной хроматографии (GFC) являются медленное разделение и низкий пик разрешения, приписываемые некоторым факторам, включая неподходящий выбор матрицы, длину колонки, высокую скорость течения и большие мертвые зоны, захваченные в колонку. Однако GFC является действительно привлекательным вариантом для обессоливания и буферного обмена во время очистки пептидов и белков. К тому же химический синтез часто касается высокого риска и стоимости, и хотя, как ожидается, производство с помощью рекомбинантной генетической технологии заменит этот способ, выход продукта до сих пор является недостаточным. Более того, синтез требует использования методов, которые нуждаются в высоком уровне мастерства и компетенции. Таким образом, желательным является производство hPTH с применением методов рекомбинантных ДНК.
Вследствие простоты культивирования, низкой стоимости и высокого потенциала производства, Escherichia coli представляет собой предпочтительного хозяина для того, чтобы экспрессировать и очищать фармацевтически важные белки.
hPTH вследствие естественной неустойчивости, связанной с прямой экспрессией белка (Morelle et al., 1988, Biochim. Biophys. Acta 950, 459-462), используют стратегию химерного белка. Обычно используемые для экспрессии и нисходящей очистки данного белка слитые (химерные) партнеры включают β-галактозидазу, cro-β-галактозидазу, hGH, Trx и фосфорибулокиназу (Suzuki, Y et al, Appl.Env.Micro 1998, 64, 526-529, Wingender E et al., J. Biol. Chem 264, 4367-4373, Gardella TJ et al., J. Biol. Chem 26, 15854-15859, Xiang Yang Fu et al., Biotechnol. Prog 2005, 21, 1429-1435, WO/1999/005277, C12N15/62). К тому же количество аминокислот, которые были использованы из β-галактозидазы в качестве слитого партнера, было разным для разных белков. В случае инсулина и проинсулина (Shen, S-H. (1984), Proc. Natl. Acad. Sci. USA 81: 4627-4631, Guo, L. (1984), Gene 29: 251-25) β-Gal слитый партнер был намного длиннее, чем РТН, на одинаковой задней части промотора (Т7 and lac promoter, Massayuki, Y et al., (1997), US5670340). Большое количество литературы сообщает об экспрессии химерного белка в виде нерастворимых агрегатов при проведении серии стадий денатурирования и повторного свертывания (рефолдинга). В случаях, когда β-галактозидазу использовали в качестве слитого партнера для РТН (1-34), сообщалось об окончательных выходах в пределах от 20 мг/л до 500 мг/л при использовании изопропил-бета-галактозида (IPTG) в качестве индуктора при концентрации 1 мМ. Однако использование IPTG для крупномасштабного производства рекомбинантных белков нежелательно из-за его высокой стоимости и токсичности (Donovan et al., 1996; Figge et al., 1988; Gombert и Kilikian, 1998; Ksinski et al., 1992). Oldenburg K.R., et al., (Protein Exp. Purification 5(3), 278-284. (1994)) описывает способ высокоуровневой экспрессии rPTH (1-34) в E.coli с помощью полигистидин лидерного пептида и восьми копий гена РТН. Oldenburg K.R., et al., также сообщает о «захвате» химерного белка с помощью никель-хелатной (Ni) хроматографии с последующим расщеплением бромцианом (CNBr) и очисткой с помощью RT-HPLC с конечным выходом 300 мг/л высокоочищенного биологически активного hPTH (1-34). Использование бромциана представляет собой экологическую опасность в отношении безопасного обращения и устранения, что жестко ограничивает его использование.
ЕР 794255 раскрывает очистку rhPTH (1-34) с использованием вырезания Кех2 из его химеры. Suzuki. Y, et al. (Applied и Environmental Microbiology 64(2), 526-529 (1998)) получили 0,5 г hPTH (1-34) с чистотой >99% из одного литра культуры Escherichia coli с использованием линкеров β-галактозидазы разной длины вместе с His-tag слитым партнером. Suzuki Y., et al. раскрывают выделение телец включения, солюбилизацию в 8М мочевине, разбавление до концентрации мочевины 3М с последующим Kex2 расщеплением, а затем промежуточную очистку с помощью ионообменной хроматографии, и наконец доочистку с помощью двух стадий обращенно-фазовой хроматографии. Suzuki Y. et al. сообщает, что Кех2, секреторный тип протеазы Кех2 из дрожжей, используемый для ферментативного расщепления, находится под значительным влиянием мочевины в концентрации 3М, необходимой для сохранения химерного белка в растворимой форме, даже добавление 2,5 мМ CaCl2, чтобы подавить инактивацию, ведет к осаждению химерного белка, которое приводит к использованию молярного отношения 1:2000 фермента к субстрату. Suzuki Y. et al., таким образом все-таки сообщает о высоком выходе целевого белка, но применение Kex2 тем не менее несет ответственность за слишком большую часть первичной стоимости способа производства. Suzuki Y. et al., также подробно сообщает, что использование 97, 117 и 139 аминокислотных фрагментов β-галактозидазы как слитого партнера, разделенного линкером, имеющим SVKKR в качестве сайта расщепления для Kex2, отвечает за высокий выход химерного белка.
Jin. L., et al., J Biol. Chem. (2000), 275(35), 27238-27244) раскрывает способ очистки LY 333334 молекулы rhPTH (1-34) с помощью солюбилизации в 7М мочевине и очистке на обращенно-фазовой колонке, с последующей очисткой на FF SP катионообменной колонке с применением градиента NaCl, а затем методом RT-HPLC, в котором очищенное вещество в 20 мМ глициновом буфере с рН 9 высушивают сублимацией. Он представляет собой метод про-растворителя (pro-solvent method) и может быть опасным в промышленном масштабе. Fu, Xiang-Yang et al, (Biotechnology Progress (2005), 21(5), 1429-1435) сообщает способ очистки тиоредоксин-слияния РТН (1-34) из клеток BL21(DE3) посредством использования Тритона-Х 100 и термической денатурации, вызывающей частичную очистку осаждением. Нагревание проводят при 80°С в течение 15 минут. В данном способе белок обрабатывают при высокой температуре, которая является в целом нежелательной для протеинов и пептидов. CN 1417231 описывает способ восстановления рекомбинантното PTH (1-34) ферментацией, с последующей очисткой телец включения, ренатурацией, расщеплением тромбином и очисткой во время катионообменной хроматографии. CN 1424325 описывает GST химерный РТН (1-34) пептид (с GSP в качестве сайта расщепления) расщепленный тромбином, очистку на аффинной колонке с химотрипсином, расщепление с помощью пролинэндопептидазы и дополнительную хроматографическую очистку. Это трудоемкий способ, а использование двух протеаз увеличивает затраты на способ очистки, ограничивая его коммерческое использование. Biochem. Biophys. Res.Commun., 166, 50-60 (1990) documents а кДНК approach for the synthesis of hPTH.
Chen, J.Y. et al., ((2004), 40 (1), 58-65) раскрывает очистку РТН (1-34) путем экспрессии РТН (1-34) как химеры с целлюлоза-связывающим доменом, расщепления РТН (1-34) с помощью Фактора Ха, очистки с помощью целлюлозной смолы и RT-HPLC с выходом 3 мг/л, который представляет собой очень малопродуктивный способ, нежизнеспособный с коммерческой точки зрения. GST-химерная технология для производства РТН (1-34) также хорошо известна в данной области техники. Gram Hermann et al., (Bio/ Technology (1994), 12(10), 1017-23) описывает способ очистки РТН (1-34) с использованием дипептидилпептидазы IV. Wingender E., et al., (J Biol. Chem. (1989), 264(8), 4367-4373) экспрессировали РТН в E.coli как cro-β-галактозидаза-hPTH химерный белок. Выход из 1 л культуры составлял около 250 мг химерного белка, который они солюбилизировали в мочевине и затем использовали обработку кислотой для высвобождения РТН. Кислые условия расщепления досаждают ограничениями из-за образования дезаминированных или окисленных побочных белковых примесей, которые трудно отделить от некоторых белков и, кроме того, кислое значение рН представляет собой жесткие условия, вредные для биоактивности белка.
ЕР 0483509 В1 относится к кодон-оптимизированному синтетическому гену, производящему hPTH, соответствующий аминокислотной последовательности hPTH, содержащей его ДНК, клетке-хозяину, трансформированной данной DNA и способу получения hPTH с использованием трансформанта в E.coli с IPTG индукцией. ЕР0483509 В1 раскрывает очистку экспрессированного РТН с помощью RT-HPLC. Применение органических растворителей в качестве элюантов в RT-HPLC может повредить белок, что представляет собой масштабную проблему. US 5208041 сообщает о выработке по существу чистого hPTH, характеризуемого единичным пиком переноса при анализе с помощью капиллярного электрофореза при 214 нм, и величиной ЕС50 (как определено в UMR исследовании на аденилатциклазе со 106 основаниями) не больше чем 2 нМ при очистке сырого hPTH с помощью RT-HPLC с катионным агентом для образования пары ионов, например, таким как триэтиламинфосфат. US 5208041 также раскрывает обработку hPTH, полученного или из ткани млекопитающего, из микробных источников РТН или из синтетических источников, по меньшей мере одной стадией фракционирования на колонке до RT-HPLC. US 5208041 приводит пример способа очистки путем обработки всего бульона при рН от 4,0 до 8,0 ледяной уксусной кислотой, очищения центрифугированием, с последующей загрузкой в ионообменную хроматографическую колонку из S-сефарозы, дополнительной обработки элюата промежуточной стадией очистки путем загрузки в HIC-колонку из фенил-сефарозы и в заключение очистки с помощью RT-HPLC с использованием С18 колонки с триэтиламинфосфатом в качестве агента для образования пары ионов. US 5208041 упоминает обнаружение с помощью капиллярного электрофореза 4 раньше необнаруженных незначительных пиков, элюирующих впереди пика РТН, и нескольких замыкающих пиков, которые не были определены при использовании трифторуксусной кислоты (TFA) или гептафтормасляной кислоты (HFBA) в качестве агента для образования пары ионов, таким образом приводя к восстановлению по существу чистого hPTH. US 5208041 не сообщает об использовании каким-либо образом HIC в качестве окончательной стадии очистки для того, чтобы получить очищенный hPTH (1-34) с чистотой ≥99%, которая находится в фокусе настоящего изобретения. Патент США 5457047 относится к последовательностям ДНК, кодирующим варианты РТН, векторам экспрессии, бактериальным хозяевам, применениям и терапевтическим композициям, US 5457047 раскрывает очистку hPTH при помощи СМ-целлюлозы в периодическом способе с последующей RT-HPLC из cro-β-галактозидаза-hPTH химерного белка.
US 6590081 сообщает о синтезе чистой кристаллической формы терипаратида и способах получения и очистки фрагментированного РТН. US 6590081 подробно сообщает о преимуществе кристаллической формы гормона для чистоты продукта и стабильности хранения. Также, чтобы создать более эффективные и пригодные для орального применения аналоги РТН, подробная информация о структуре пептида должна способствовать описанию молекулярных взаимодействий между лигандом и рецептором. US 6590081 также раскрывает, что кристаллический РТН к тому же может быть включен в состав других композиций, таких как, например, таблетки, капсулы или суппозитории, например, то же самое легко растворяется в стерильном растворе в пузырьках. US 6590081 заявляет кубические, гексагональные и пластинчатые кристаллы hPTH (1-34) и способ очистки РТН для получения того же самого.
Liu, Q., et al., in Protein Expr Purif., 2007 Aug., 54(2): 212-9, раскрывает крупномасштабный способ получения hPTH (1-84) из E.coli с использованием стратегии растворимого химерного белка, путем конструирования кодон-оптимизированного синтетического гена, кодирующего hPTH (1-84), и клонирования того же самого в рЕТ32а (+) вектор, экспрессируемый в клетках Е.coli BL21 (DE3) как растворимый His(6)-тиоредоксин-hPTH (1-84) химерный белок. Liu, Q., et al., придерживается продолжения стадий очистки «захвата» путем аффинной хроматографии с использованием иммобилизованных металлов, с последующим энтерокиназным расщеплением, и в конце подвергая то же самое воздействию эксклюзионной хроматографии размеров с количественным выходом 300 мг/л с чистотой 99% после изъятия (сбора) растворимого химерного белка.
Замысел ортогонального способа основывается на полностью контролируемых способах очистки (Gagnon P. The secrets of Orthogonal Process Development. Validated Biosystems, 2006:www.validated.com/revalbio/pdffiles/orthopd.pdf). Идея заключается в том, что сочетание стадий с наибольшей комплементарностью должно обеспечить самую лучшую полную очистку. Сильнейший вариант осуществления идеи обычно достигается, когда соответствующие стадии основаны на разных механизмах разделения. Двухстадийный способ, который включает одну стадию фракционирования на основе размера продукта, и другую на заряде продукта может быть рассмотрен как ортогональный; подобным образом, способ с одной стадией на основе заряда продукта и другой на гидрофобности также может быть ортогональным. Важной особенностью дизайна ортогонального способа является то, что возможность очистки любой одной стадии измеряется только в рамках контекста его потенциального партнера.
Настоящее изобретение содержит ортогональный способ очистки rhPTH (1-34), сочетающий катионообменную хроматографию с HIC для того, чтобы добиться чистоты ≥99% с хорошим выходом.
Крупномасштабное производство терапевтических белков сталкивается с некоторыми проблемами, такими как короткие временные рамки развития, учет стоимости и даже увеличение требований к уровню качества. Уровень техники методов выделения и очистки, конкретно принципы ортогонального разделения, имеют огромное значение для того, чтобы ускорить развитие, уменьшить продолжительность обработки и сократить издержки производства. Препаративная хроматография значительно развита в отношении стабильности матрикса или наличия селективности и предоставляет главную технологию в очистке биомолекул до чистоты свыше 95%. Хроматография гидрофобного взаимодействия (HIC) приобретает популярность в последние годы, так как она предлагает возможности ортогонального разделения для широко используемых методов очистки на основе ионных взаимодействий. HIC часто является отличным выбором вслед за ионообменной хроматографией в процедуре очистки белка. Оба метода имеют чрезвычайно широкую применимость и являются каноническими подходами относительно друг друга (т.е. разделение в соответствии с гидрофобностью и зарядом, соответственно). Более того, вещество, элюируемое с солевым градиентом при ионообменном разделении, требует минимумальной обработки образца. С другой стороны, несмотря на проявление сходной селективности, HIC является менее денатурирующей по сравнению с обращенно-фазовой хроматографией, использующей более гидрофобные лиганды и органические растворители. В промышленном масштабе требуются взрывобезопасные производственные модули для обращения с токсическими органическими растворителями, размещение больших количеств которых дорого. Следовательно, ортогональный подход сочетания ионообменной хроматографии с последующей HIC является рентабельной, благоприятной с экологической точки зрения технологической платформой для крупномасштабного производства рекомбинантных химерных белков.
Вышеупомянутый предшествующий уровень техники ни сообщает точно ортогональный подход очистки hPTH (1-34) путем сочетания катионообменной хроматографии, как промежуточной стадии очистки с HIC в качестве конечной стратегии очистки, ни сообщает о применении нового способа индуцирования лактозой с хорошим выходом hPTH (1-34), который находится в главном фокусе настоящего изобретения.
Настоящее изобретение включает уникальный новый признак применения ортогонального подхода двухстадийного способа очистки, состоящего из катионообменной хроматографии необязательно с последующей препаративной хроматографией, выбранной из HIC или RT-HPLC. Настоящее изобретение раскрывает простой, рентабельный, благоприятный с экологической точки зрения способ получения высокочистого hPTH (1-34). Другим уникальным признаком настоящего изобретения является стратегия индуцирования лактозой при периодическом добавлении для получения оптимизированной экспрессии hPTH (1-34) у прокариотического хозяина. Другим признаком настоящего изобретения является создание гибридного химерного белка, состоящего из слитого (химерного) партнера, состоящего из 41 аминокислоты гена β-галактозидазы (LacZ) Escherichia coli, сайта эндонуклеазного расщепления, фрагмента гена rhPTH, в котором выбран слитый партнер, являющийся присущим гену β-галактозидазы Escherichia coli, с высоким содержанием % GC, соответствующей вторичной структурой пептида, the secondary structure of ribonucleotide translating the same и pI химерного фрагмента в качестве помощи, способствующей технологии производства.
Цели изобретения
Первым аспектом настоящего изобретения является способ синтеза rhPTH (1-34), включающий:
i. выделение общей РНК из тканевого источника,
ii. конструирование кДНК, кодирующей химерный нуклеотид ORF как NcoI/XhoI фрагмент, как указано в SEQ. ID NO.:1, путем амплификации кДНК с помощью RT-PCR с использованием ген-специфических праймеров, выбранных из SEQ. ID NO.:3, SEQ. ID NO.:4, SEQ. ID NO.:5, SEQ. ID NO.:6, SEQ. ID NO.:7 и SEQ. ID NO.:8, кодирующих rhPTH (1-34) химерный гибридный белок, как указано в SEQ. ID NO. 2,
iii. трансформацию Escherichia coli вектором экспрессии, содержащим химерный нуклеотид ORF как NcoI/XhoI фрагмент, как указано в SEQ. ID NO.:1, кодирующей rhPTH (1-34) гибридный химерный белок, как указано в SEQ. ID NO.:2, в которой гибридный химерный белок состоит из аффинного конца (affinity handle), слитого (химерного) партнера, сайта эндонуклеазного расщепления и пептида rhPTH (1-34),
iv. культивирование трансформированной Escherichia coli от стадии iii для экспрессии гибридного химерного белка в присутствии индуктора,
v. выделение гибридного химерного белка из культуры в форме телец включения,
vi. захват rhPTH (1-34) гибридного химерного белка стадии v,
vii. расщепление rhPTH (1-34) гибридного химерного белка стадии vi,
viii. очистку rhPTH (1-34), полученного посредством стадии vii с помощью ортогонального способа.
Второй аспект настоящего изобретения раскрывает применение индуцирования лактозой с низкой скоростью подачи для экспрессии гибридного химерного белка rhPTH (1-34), в котором индуцирование лактозой находится в пределах примерно от 10% до 30% общего белка.
Третий аспект настоящего изобретения представляет собой ортогональный способ очистки rhPTH (1-34), включающий:
i. захват солюбилизированного гибридного химерного белка, как указано в SEQ. ID NO.:2, с помощью хроматографии ЕВА (адсорбция на слой вспученного адсорбента),
ii. расщепление элюированного гибридного химерного белка стадии i с помощью эндопептидазы с выходом целевого белка,
iii. очистку целевого белка с помощью катионообменной хроматографии до чистоты ≥98%,
iv. необязательно доочистку целевого белка с помощью HIC или RT-HPLC до чистоты ≥99%.
Четвертый аспект настоящего изобретения раскрывает ортогональный способ очистки rhPTH (1-34), в котором солюбилизированный гибридный химерный белок улавливают на проточной хелатной сефарозной колонке при денатурирующих условиях, захваченный гибридный химерный белок расщепляют энтерокиназой при концентрации мочевины в пределах от 500 мМ до 4000 мМ в течение периода от 4 до 6 часов, расщепленный целевой белок очищают до чистоты ≥98% на SP-XL катионообменной колонке и элюированный целевой протеин необязательно доочищают с помощью HIC (хроматографии гидрофобного взаимодействия) на фенилсефарозе до чистоты ≥99%.
Пятый аспект настоящего изобретения представляет собой rhPTH (1-34) в жидкой форме, в котором rhPTH (1-34) не находится ни в кристаллической форме, ни в аморфной форме.
Другие аспекты и преимущества настоящего изобретения будут видны после рассмотрения следующего подробного описания, которое включает многочисленные иллюстративные примеры осуществления на практике данного изобретения.
Краткое описание чертежей
Способ, с помощью которого могут быть достигнуты цели и преимущества изобретения, будет понятен более полно из подробного описания и сопроводительных чертежей, которые изложены ниже:
фиг.1 показывает анализ амплифицированного ПНР-продукта в 1% агарозном геле, где фрагмент 281 п.о., покрывающий lacZ часть ORF, и область 141 п.о., которая содержит соответствующие нуклеотиды аминокислот 1-34, показана на Дорожке 1. Эти фрагменты были дополнительно расщеплены до 281 п.о. с помощью NcoI/SalI (Дорожка 2) и до 141 п.о. с помощью SalI/XhoI (Дорожка 3). Дорожка 4: pET19b вектор после расщепления с помощью NcoI/XhoI и гель-очистка для применения клонирования, Дорожка 5: 100 п.о. DNA маркер (promega);
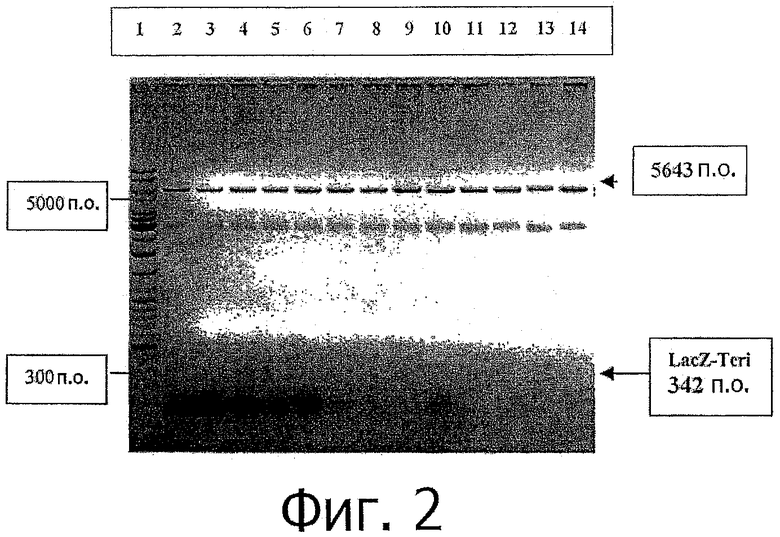
фиг.2 показывает минипрепарат ДНК (Miniprep DNA), расщепленной с помощью NcoI/XhoI, показывающий наличие позитивных клонов со вставкой представляющего для нас интерес фрагмента β-галактозидаза-hPTH (1-34) (LacZ). Дорожки 2-12 и 14: показывают полосу вставки 342 п.о. фрагмента 4) β-галактозидаза-hPTH (1-34) (показана стрелкой). Фрагмент вектора 5643 п.о. pET19b показан наверху (показан острием стрелки). Дорожка 13 подтверждает размер фрагмента вектора расщепленного NcoI/XhoI, который показывает отсутствие полосы вставки 342 и.о.;
фиг.3 показывает карту вставки рЕТ-β-галактозидаза-hPTH (1-34) в вектор рЕТ-19b;
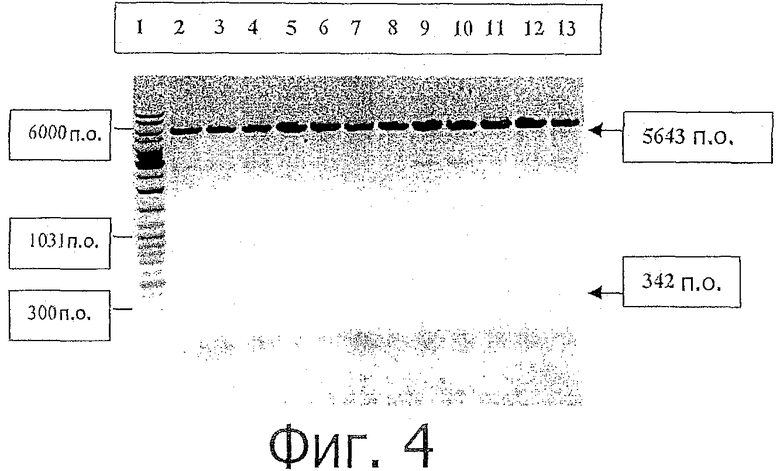
фиг.4 показывает RE анализ с помощью NcoI/XhoI на позитивные клоны-трансформанты BL21(DE3), демонстрируя наличие расщепленного фрагмента, 342 п.о. (показан стрелкой). Дорожка 1: Gene roller ladder mix, Дорожка 2-13: Клон # 48 и 49. То же самое было сделано с другими позитивными клонами, упомянутыми на фиг.3;
фиг.5 показывает RE анализ с помощью NcoI/XhoI на позитивные клоны-трансформанты BL21(DE3), демонстрируя наличие расщепленного фрагмента, 342 п.о. (показан стрелкой). Дорожка I: Gene roller ladder mix. Дорожка 2-6: Клон #48 и 49. То же самое было сделано с другими позитивными клонами, упомянутыми на фиг.4;
фиг.6 показывает профиль питания и ферментации клона rhPTH (1-34);
фиг.7 показывает ферментный анализ экспрессии генов химерного rhPTH (1-34). Дорожка 1: маркер, Дорожка 2: неиндуцированный, Дорожка 3: индуцированный лактозой;
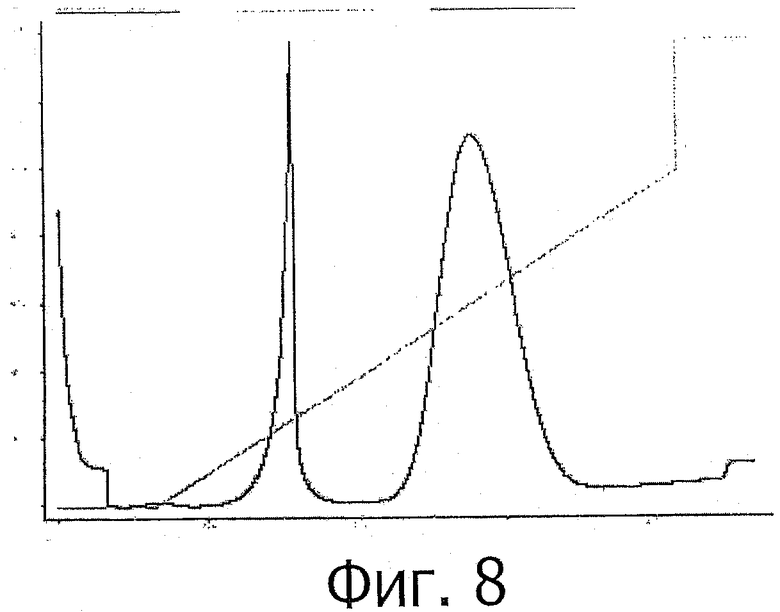
фиг.8 показывает ЕВА-хроматограмму с проточной хелатной сефарозой;
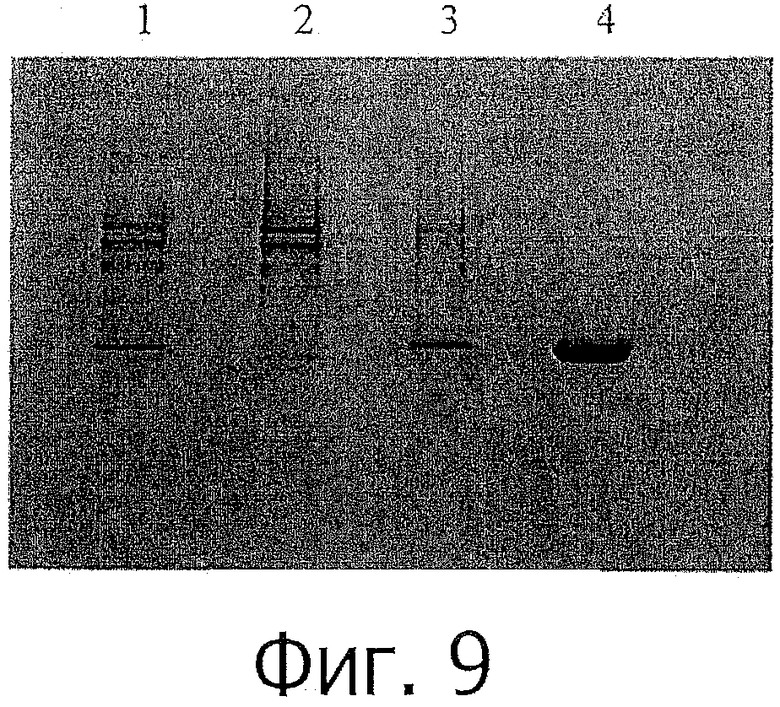
фиг.9 показывает Кумасси окрашенный SDS-PAGE профиль химерного rhPTH (1-34) с помощью проточной хелатной сефарозы на ЕВА. Дорожка 1: исходный материал, Дорожка 2: несвязанный (проскок), Дорожка 3: смыв. Дорожка 4: элюирование;
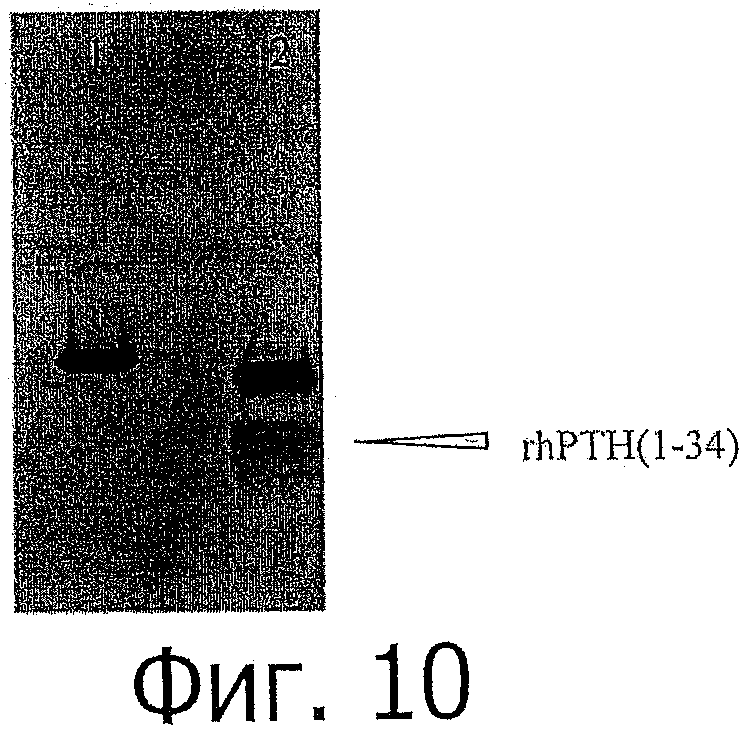
фиг.10 показывает Кумасси окрашенный SDS-PAGE анализ EK расщепления химерного rhPTH (1-34). Дорожка 1: нерасщепленный. Дорожка 2: EK расщепленный образец;
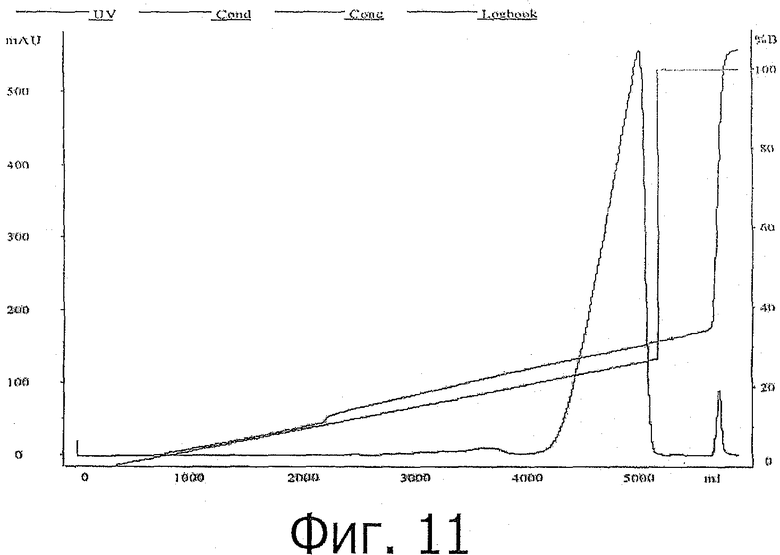
фиг.11 показывает ионообменную хроматограмму с использованием SP-XL колонки;
фиг.12 показывает анализ фракций ионообменной хроматографии с помощью гель-электрофореза. Дорожка 1: образец нагружен на IEX, Дорожка 2: FT, Дорожка 3: IEX элюирование;
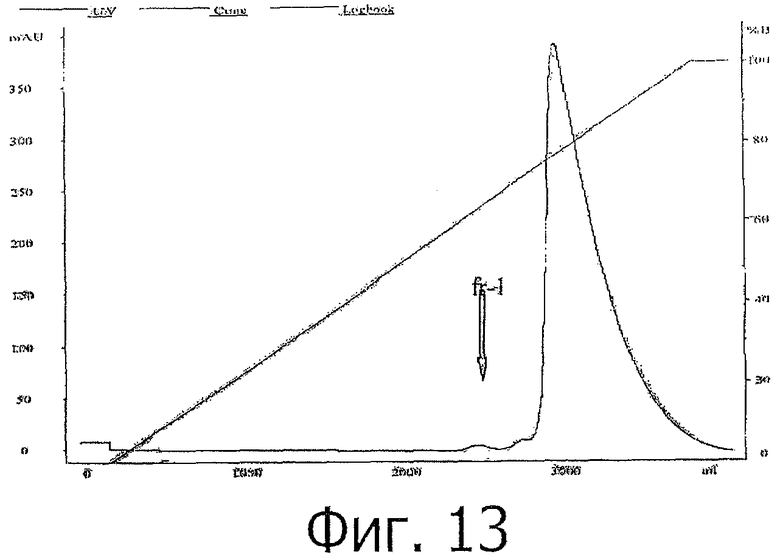
фиг.13 показывает хроматограмму очистки rhPTH (1-34) с помощью HIC;
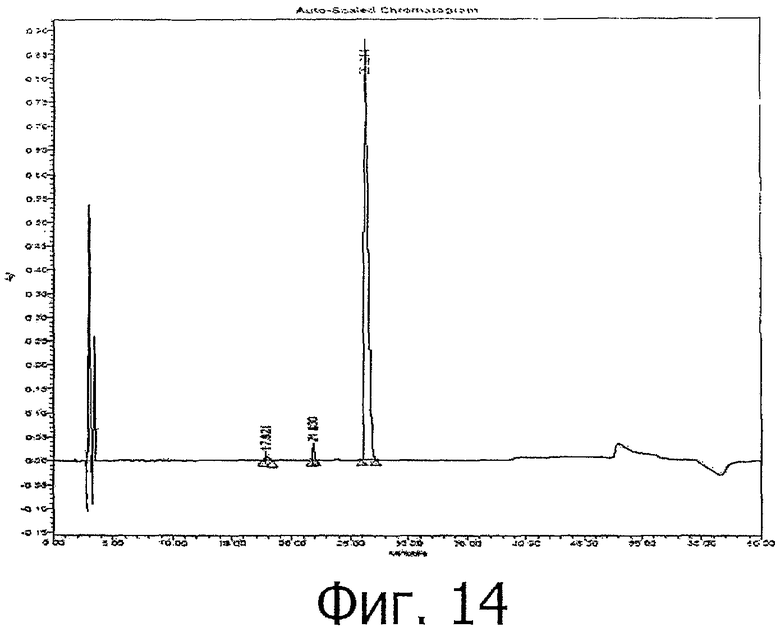
фиг.14 показывает RT-HPLC анализ фракции 1 хроматографии гидрофобного взаимодействия (HIC);
фиг.15 показывает хроматографический обращено-фазовый профиль rhPTH (1-34);
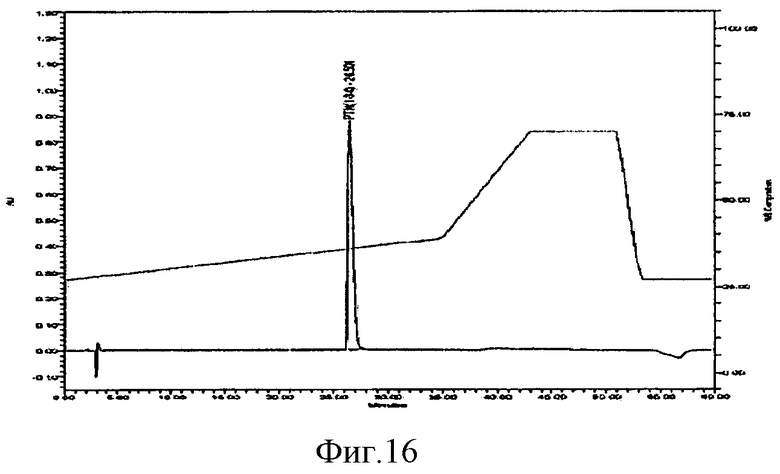
фиг.16 показывает анализ на чистоту rhPTH (1-34) с помощью RT-HPLC.
Подробное описание изобретения
Один вариант осуществления настоящего изобретения касается способа синтеза rhPTH (1-34), включающего:
i. выделение общей РНК из тканевого источника,
ii. конструирование кДНК, кодирующей химерный нуклеотид ORF как NcoI/XhoI фрагмент, как указано в SEQ. ID NO.:1, путем амплификации кДНК с помощью RT-PCR с использованием ген-специфических праймеров, выбранных из SEQ. ID NO.:3, SEQ. ID NO.:4, SEQ. ID NO.:5, SEQ. ID NO.:6, SEQ. ID NO.:7 и SEQ. ID NO.:8, кодирующих rhPTH (1-34) химерный гибридный белок, как указано в SEQ. ID NO. 2,
iii. трансформацию Escherichia coli вектором экспрессии, содержащим химерный нуклеотид ORF как NcoI/XhoI фрагмент, как указано в SEQ. ID NO.:1, кодирующей rhPTH (1-34) гибридный химерный белок, как указано в SEQ. ID NO.:2, в которой гибридный химерный белок состоит из аффинного конца, слитого (химерного) партнера, сайта эндонуклеазного расщепления и пептида rhPTH (1-34),
iv. культивирование трансформированной Escherichia coli от стадии iii для экспрессии гибридного химерного белка в присутствии индуктора,
v. выделение гибридного химерного белка из культуры в форме телец включения,
vi. захват rhPTH (1-34) гибридного химерного белка стадии v,
vii. расщепление rhPTH (1-34) гибридного химерного белка стадии vi,
viii. очистку rhPTH (1-34), полученного посредством стадии vii с помощью ортогонального способа.
Другой вариант осуществления настоящего изобретения касается применения индуцирования лактозой с низкой скоростью подачи для экспрессии гибридного химерного белка rhPTH (1-34), в котором индуцирование лактозой находится в пределах примерно от 10% до 30% общего белка.
Другой вариант осуществления настоящего изобретения касается ортогонального способа очистки rhPTH (1-34), включающего:
i. захват солюбилизированного гибридного химерного белка, как указано в SEQ. ID NO.:2, с помощью хроматографии ЕВА (адсорбция на слой вспученного адсорбента),
ii. расщепление элюированного гибридного химерного белка стадии i с помощью эндопептидазы с выходом целевого белка,
iii. очистку целевого белка с помощью катионообменной хроматографии до чистоты ≥98%,
iv. необязательно доочистку целевого белка с помощью HIC или RT-HPLC до чистоты ≥99%.
Еще один вариант осуществления настоящего изобретения касается ортогонального способа очистки rhPTH (1-34), в котором солюбилизированный гибридный химерный белок улавливают на проточной хелатной сефарозной колонке при денатурирующих условиях, захваченный гибридный химерный белок расщепляют энтерокиназой при концентрации мочевины в пределах от 500 мМ до 4000 мМ в течение периода от 4 до 6 часов, расщепленный целевой белок очищают до чистоты ≥98% на SP-XL катионообменной колонке и элюированный целевой протеин необязательно доочищают с помощью HIC (хроматографии гидрофобного взаимодействия) на фенил-сефарозе до чистоты ≥99%.
Еще один вариант осуществления настоящего изобретения касается rhPTH (1-34) в жидкой форме, в которой rhPTH (1-34) не находится ни в кристаллической форме, ни в аморфной форме.
Термин "кДНК" или комплементарная ДНК при использовании здесь относится к синтетической ДНК, транскрибированной со специфической РНК с помощью фермента обратная транскриптаза.
Термин "ORF" или открытая рамка считывания при использовании здесь относится к части генома организма, которая содержит последовательность оснований, потенциально кодирующих белок.
Термин "β-галактозидаза" и "LacZ" используются здесь как синонимичные термины.
Фраза "ген-специфические праймеры" имеет в виду праймеры, полностью комплементарные для гибридизации с целевым полинуклеотидом для синтеза удлиненного продукта праймера, который является комплементарным целевому полинуклеотиду.
Хроматография гидрофобного взаимодействия (HIC) наносит минимальное структурное повреждение биомолекулам, и их биологическая активность сохраняется благодаря более слабому взаимодействию, чем при аффинной, ионообменной или обращенно-фазовой хроматографии (ОФХ) (Fausnaugh et al., 1984; Regnier, 1987). HIC представляет собой альтернативный путь использования гидрофобных свойств протеинов при работе в более полярной и менее денатурирующей окружающей среде, чем при ОФХ, так как данная методика требует использования неполярных растворителей для элюирования белков из-за сильной связи с адсорбентом (E1 Rassi, 1996).
РТН представляет собой одноцепочечный пептид, состоящий из 84 аминокислот, у которого структурные требования в отношении полной биологической активности удовлетворяются первыми 34 NH2-концевыми аминокислотами. Устранение некоторых аминокислот или из NH2 или СООН конца активного фрагмента РТН (1-34) приводит к прогрессирующему спаду биологической активности, так что область непрерывной последовательности 2-26 определяется как минимальная последовательность, необходимая для биологической активности, как установлено посредством активации почечной аденилатциклазы. Rossenblatt, M. et al., подробно сообщает, что синтетический аналогичный (Nle-8, Nle-18, Tyr-34)bPTH-(3-34) амид, который включает модификации, показанные с bPTH (1-34) и для усиления биологической активности и придания устойчивости к окислению, будет ингибировать bPTH (1-84) активность, когда присутствует в эквимолярных количествах с нативным гормоном. Известные виды биологической активности РТН представлены во фрагменте, который включает только первые 34 аминокислоты (Tregear, G.W., Rietschoten, J.V., Greene. E., Keutmann, H.T., Niall, H.D, Reit, B., Parsons, J.A., и Potts, J.T., Jr.(1973) Endocrinology 93, 1349-1353)), a функция 50 остатков на карбоксильном конце неизвестна. Взаимодействие 1-34 РТН с его рецепторами изменяется и при окислении остатка метионина в положениях 8 и 18 (Tashjian, A.H., Ontjec, D.A., и Munson, P.L. (1964) Biochemistry 3, 1175-1182; Frelinger, A.L., III, и Zull, J.E. (1984) J.Biol.Chem. 259, 5507-5513; Frelinger, A.L., III, и Zull, J.E. (1986) Arch. Biochem. Biophys. 244, 641-649) и при устранении аминокислот на амино-конце гормона (Martin, K.J., Bellorin-Font, E., Freitag, J., Rosenblatt, М., и Slatopolsky, E. (1981) Endocrinology 109, 956-959; Mckee, R.L., Goldman, M.E., Caulfield, M.P., deHaven, P.A., Lave, J.J., Nutt, R.F., и Rosenblatt, М. (1988) Endocrinology 122, 3008-3010; Goldman, M.E., McKee, R.L., Caulfield, M.P., Reagen, J.E., Levy, J.J., Gay, C.T., DeHaven, P.A., Rosenblatt, М., и Chorev, M. (1988) Endocrinology 123, 2597-2599). Окисленные пептиды являются полными агонистами со сниженной аффинностью (сродством) (Frelinger, A.L., III, и Zull, J.E. (1984) J. Biol.Chem. 259, 5507-5513; Frelinger, A.L., III, и Zull, J.E. (1986) Arch. Biochem. Biophys. 244, 641-649), а пептиды с делециями на амино-конце являются частичными агонистами или антагонистами со сниженной аффинностью (Martin, K.J., Bellorin-Font, E., Freitag, J., Rosenblatt, М., и Slatopolsky, E. (1981) Endocrinology 109, 956-959; Mckee, R.L., Goldman, M.E., Caulfield, M.P., deHaven, P.A., Lave, J.J., Nutt, R.F., и Rosenblatt, М. (1988) Endocrinology 122, 3008-3010). Окисление остатка 8 имеет самое большое влияние на аффинность гормона, и делеция (устранение) остатков 1 и 2 вызывает самые драматические эффекты на биологическую активность. Таким образом, 3-34 РТН является очень слабым агонистом, а 7-34 фрагмент является антагонистом. Представляется, что или измененные остатки прямо вовлекаются в рецепторное связывание, или что окисление или устранение вызывает вторичные или третичные структурные изменения в пептиде, так что рецепторное связывание или активация становится дефектной. Следовательно, уникальный чувствительный ортогональный способ синтеза N и С концевого интактного полной длины rhPTH (1-34), который проявляет биоактивность, является существенным признаком данного изобретения, в котором отделение rhPTH (1-34) от других примесей выполняется с чистотой ≥99% с хорошим выходом продукта. Окисленные примеси часто получаются во время водной очистки rhPTH (1-34) при воздействии воздуха на пептид. Поэтому в литературе сообщается о применении RT-HPLC в качестве завершающей стадии доочистки, в которой органические растворители, использованные для элюирования, предотвращают окисление воздухом. Взамен преимуществ, предлагаемых HIC, в отношении затрат, которые уменьшаются при отказе от использования органических растворителей HPLC-класса, и дополнительных затрат, понесенных при удалении следов органических растворителей до желательных пределов, а также при избавлении и, главным образом, обработке летучих растворителей в крупном масштабе, изобретатели разработали простой, благоприятный с экологической точки зрения, рентабельный ортогональный способ сочетания катионообменной хроматографии с HIC с получением rhPTH (1-34) с чистотой ≥99%. Настоящее изобретение, таким образом, избегает образования каких-либо окисленных примесей и других сопутствующих примесей, как, например, дезаминированные пептиды, при использовании водной среды для элюирования и производства N и С концевого интактного rhPTH (1-34). В настоящем изобретении промежуточная стадия очистки катионообменной хроматографией, с использованием колонки SP-XL, давала rhPTH (1-34) с чистотой ≥98% с повышенным выходом в пределах 300-400 мг/л. Таким образом в фокусе настоящего изобретения находится синтез высоко очищенного стабильного rhPTH (1-34) в жидкой форме.
Широкий спектр коммерчески важных белков производится с помощью Escherichia coli. Поэтому были сделаны значительные усилия для оптимизации объемного выхода рекомбинантных белков с целью снижения производственных затрат (Lee, 1996).
Важной характеристикой промоторов, используемых в этих системах, является их индуцибельность, в смысле простоты и экономической эффективности. Применение лактозы в качестве индуктора для оптимизации при периодическом добавлении субстрата экспрессии химерного белка также представляет собой существенный признак настоящего изобретения.
В настоящем изобретении мы описываем способ клонирования РТН (1-34), посредством амплификации кДНК, кодирующей человеческий РТН, состоящий из 1-84 аминокислот, с использованием RT-PCR. кДНК клонировали в клонирующий вектор на основе UC-18, который использовали как матрицу для клонирования области, кодирующей 1-34 аминокислоты hPTH. Целевая конструкция была разработана способом, когда 41 новая аминокислота β-галактозидазы (LacZ) (со 124й аминокислоты по 164ю аминокислоту) были взяты в качестве химерного пептида, для того чтобы повысить уровень экспрессии РТН в 1 л встряхиваемой колбе, а также в увеличенной ферментации. Пептид добавляет преимущество р1 для интактного рекомбинантного белка, и был выбран, исходя из вторичной структуры пептида и свободной энергии рибонуклеотида. Сайт протеазного разрезания, например, энтерокиназой, был включен на конце lacZ слияния для эффективного удаления слитого партнера из представляющего интерес белка. Полная ORF находится под контролем промотора Т7, индуцируемого ампициллином как селективный маркер.
Клеточный осадок, полученный из ферментативного бульона, суспендировали в буфере; для того, чтобы вызвать лизис клеток, клеточную суспензию подвергали воздействию повторных циклов гомогенизации с помощью высокого давления. К этой лизированной клеточной суспензии добавляли кристаллы мочевины до конечной концентрации 4-8М и перемешивали в течение 8-12 часов для того, чтобы солюбилизировать клеточные белки. Затем раствор центрифугировали или подвергали микрофильтрованию, чтобы удалить обломки клеток.
Затем представляющий интерес гибридный белок очищали от растворимого общего белка с помощью проточной хелатной сефарозной колонки. Затем смыв из проточной хелатной колонки, содержащий химерный белок, или обессоливали с использованием G-25 или непосредственно разбавляли, чтобы снизить конечную концентрацию соли. К этому раствору добавили рекомбинантную энтерокиназу в концентрации 1-10 единиц на 20-100 мкг белка и держали в течение 4-12 часов в присутствии мочевины в концентрации 1М. Одновременно проводили обессоливание и подготовку к ферментативной стадии.
Значение рН раствора этого расщепленного образца было отрегулировано до рН 5-7 и проведено связывание на ионообменнике, предпочтительно катионообменнике. Катионообменник может быть или полимерной бусиной, или бусиной на основе сефарозы, содержащей сульфопропиловую, метилсульфонатную или карбоксиметильную группу, присоединенную к ним. После загрузки образца колонку промывали ацетатным буфером с низкой концентрацией около 20-50 мМ, и связанный белок элюировали с помощью градиента буфера В, содержащего 0,5-1М NaCl. Контролировали оптическую плотность или при 254 нм или при 280 нм для проверки профиля элюции. Основной пик включает РТН (1-34), который был взят для дополнительной очистки.
Завершающую очистку проводили с использованием HIC или RPC. Для HIC, сульфат аммония или NaCl добавили к IEX элюированию до конечной концентрации 1-2М и затем образец загружали в HIC колонку. Колонка HIC может являться фенил, бутил, изопропил или другой группой, присоединенной или к полимерной бусине, или к бусине на основе сефарозы/агарозы. В настоящем изобретении могут быть использованы оба источника фенил и фенилсефароза. Градиентное элюирование связанного белка из высокосолевого до низкосолевого в, или водном, или ацетат/цитратном буфере 20-50 мМ дает повышение чистоты РТН (1-34) до >99%. Альтернативный не-водный способ может являться стадией ОФХ с использованием колонки С4 с двуокисью кремния, так же как и полимерной обращенно-фазовой колонки. В подвижной фазе А можно использовать воду, содержащую 0,01%-0,1% TFA или натрий-ацетатный буфер рН 4,0; а мобильная фаза В может быть, или ацетонитрилом, или смесью ацетонитрила с метанолом. Градиентное элюирование с подвижной фазой В может прочно разделить сопутствующие примеси и дать повышение чистоты фракций PTH (1-34) до >99%. Следует понимать, что последующие описанные здесь примеры представлены только с иллюстративной целью, и что различные модификации или изменения, которые в свете этого будут предложены специалистами в данной области техники, включаются в сущность и область действия данной заявки и в рамки прилагаемой формулы изобретения.
Сведения, подтверждающие возможность осуществления изобретения
Пример 1. Выделение общей РНК и синтез первой цепи кДНК
Общая РНК была выделена при помощи "RNAgents For Total RNA Isolation Systems" (Promega, Cat # Z5110). Было подвергнуто обработке около 2 г ткани. Согласно выбранной стратегии, полноразмерную кДНК РТН выделяли с использованием общей РНК из нескольких типов тканей, например, печеночной, почечной, мозговой и плацентарной. Осадок общей РНК ресуспендировали в 1 мл воды, не содержащей нуклеазы, и хранили при -70°С. Концентрация общей РНК составляла около 4 нг/мл. Полиаденилированная РНК была объединена при помощи Oligo (dT)8+12 бусин (частиц) из общей РНК, как описано выше. Для каждой реакции было использовано 10 нг общей РНК. Полиаденилированная РНК была подвергнута обратной транскрипции в одноцепочечную комплементарную ДНК (1я-цепь кДНК матрицы) с использованием обратной транскриптазы AMV при 42°С в течение 1 ч. Было получено около 20 нг/мкл одноцепочечной кДНК. Качество 1-й цепи было проверено посредством амплификации пар праймеров бета-актина.
Пример 2. PCR-амплификация предполагаемой кДНК РТН, содержащей 1-84 аминокислоты, из тканевых источников
Исходя из полной длины 1-84 аминокислот, была синтезирована ген-специфическая пара праймеров для построения первых цепей методом OT-PCR. Все вышеупомянутые тканевые источники были использованы для амплификации и оказались положительными по РТН. Окончательно для клонирования 1-34 аминокислотного домена РТН в Т7 Е.coli экспрессионную векторную систему была выбрана ткань печени. кДНК, длиной 476 пн, кодирующая 1-84 аминокислоты, была клонирована и служила матрицей для дальнейшей амплификации домена, содержавшего аминокислоты 1-34.
RT-PCR была проведена на пуле мРНК с использованием ген-специфичесой пары праймеров (SEQ ID. NO.3 & 4), которая была обнаружена на последовательности кДНК РТН. Эти пары использовали для амплификации полноразмерной кДНК последовательности РТН длиной 476 п.о. (Фрагмент 1).
Пример 3. Лигирование и трансформация
Смесь для лигирования, содержащую 20 нг-50 нг ДНК фрагмента 1 (476 пн), отдельно лигировали с pUC-18, основанном на ТА-векторе клонирования, и рЕТ-19b вектором в JM109 и BL21(DE3), соответственно. Для увеличения числа трансформированных клеток чашки инкубировали при 4°С в течение ночи. Для подтверждения того же самого был проведен рестрикционный анализ и анализ последовательности позитивных клонов. Были получены несколько клонов, положительных для РТН (1-84).
Пример 4. Клонирование РТН (1-34 аминокислот) человека посредством PCR-амплификации
На следующем этапе была проведена PCR-амплификация, как упомянуто ниже, для амплифицирования 1-34 РТН домена с использованием вышеупомянутого конструкта ДНК РТН, содержавшего аминокислотные остатки 1-84. Целевой конструкт был создан так, что часть β-галактозидазы (LacZ) была использована как химерный пептид. Сайт разрезания протеазой, например энтерокиназой, был вставлен методами PCR в конец lacZ для его энзиматического удаления из зрелого 1-34 АА, целевого белка.
Нижеуказанная последовательность:
VDNVDESWLQEGQTRIIFDGVNSAFHLWGRWVGYGQDSRLP является аминокислотной последовательностью β-галактозидазы, химерной для rhPTH (1-34) с сайтом разрезания EK DDDDK.
Затем пара праймеров (SEQ. ID NO.5 & 6) была использована для амплифицирования 5'-конца гибридной химеры, lacZ с His-tag и сайтом разрезания ЕК на 3'-конце. Этот PCR-амплифицированный продукт длиной 241 п.о. (Фрагмент 2), имеет два RE сайта, NcoI и SalI, которые были пригодны во время клонирования вместе с 1-34 аминокислотами в вектор рЕТ-19b.
3'-конец rhPTH (1-34) был амплифицирован с использованием другого набора пар праймеров (SEQ ID NO. 7 & 8), который был использован на полноразмерной кДНК РТН (1-84) для амплифицирования фрагмента длиной 141 п.о. с RE сайтами SalI и XhoI для клонирования непосредственно после сайта разрезания EK.
Пример 5. Создание конечного конструкта рЕТ-lacZ-rhPTH (1-34)
Как показано на фиг.3, конечный конструкт содержит 5-upstream (предшествующий) старт-сайт с сайтом NcoI наряду с последовательностью lacZ, сайтом расщепления ЕК и непосредственным старт-сайтом последовательности rhPTH (1-34), расположенным после сайта расщепления ЕК и до ТСТ GTG…ТАА ТАА с двумя стоп-кодонами XhoI сайт СТС GAG. Затем культуры этих клонов выращивали в течение ночи при 37°С и хранили при -70°С. Полная ORF, которая транслируется до стоп-кодонов на 3'-конце непосредственно после 108 п.о. (кодирует 34 аминокислоты rhPTH (1-34)). Приведена зрелая кДНК, кодирующая 34 аминокислоты, включая два стоп-кодона (фиг.16).
Затем эта ORF была клонирована в экспрессионный вектор рЕТ-19b Е.coli на сайте NcoI/XhoI. Последовательные шаги показаны ниже. Для заключительного клонирования (фиг.2) и PCR-амплификации rhPTH (1-34) на 5'-конец был прикреплен химерный tag (lacZ). Полная химера в векторе для клонирования содержит 6 Х His-lacZ-EK-(1-34) без какого-либо сигнального пептида.
Очистка целевых фрагментов 2 и 3:
Гель-очищенная кДНК РТН была лигирована с вектором рЕТ-19b по RE сайтам NcoI/XhoI.
На дорожке 2, PCR-продукт длиной 281 п.о, (Фрагмент 2, фиг.1) показывает 5'-конец lacZ части с NcoI и SalI сайтом, который принадлежит части 6 Х His-lacZ-EK сайта. На дорожке 3, PCR-продукт демонстрирует начало зрелой ORF предполагаемого РТН после сигнального пептида, содержащего аминокислоты 1-34 (rhPTH (1-34)) с двумя стоп-кодонами. Этот фрагмент 141 п.о. (Фрагмент 3, фиг.1) содержит сайты SalI и XhoI для лигирования с вышележащими сайтами lacZ и EK. Лигирование расщепленных фрагментов 2 и 3 по сайту SalI дает в результате lacZ-rhPTH (1-34) вставку фрагмента длиной 342 п.о. (Фрагмент 4, фиг.1), который был лигирован с вектором по сайту NcoI на 5'-конце и сайту XhoI на 3'-конце. Лигирование, приводящее к образованию финального конструкта (фиг.3), и процедуры трансформации были проведены согласно примеру 3 и были подвергнуты скринингу на наличие положительных трансформантов посредством расщепления выделенной минипрепаративным способом ДНК с помощью NcoI/XhoI, чтобы получить лигированные фрагменты.
Пример 6. Подтверждение рестрикционной карты
RE анализ был проведен с использованием NcoI/XhoI и BglII/XhoI, было проведено двойное расщепление для подтверждения этих положительных клонов дополнительно после трансформации BL21(DE3) с предварительно идентифицированными положительными клонами с использованием 50-100 нг ДНК. RE анализ также был проведен с использованием SalI, которая имеет два сайта в конструкте pET-lacZ-rhPTH (1-34). Фрагмент длиной 141 п.о. (Фрагмент 3) был виден при ферментативном анализе (данные не показаны).
Несколько положительных клонов мы получили на основе штамма BL21(DE3), которые были проанализированы с помощью двойного расщепления сочетанием ферментов NcoI/XhoI. Результаты продемонстрировали правильный размер вставленного фрагмента (Фрагмент 4), равный 342 п.о. Затем, до проведения анализа последовательности (секвенирования), данные клоны были проверены с использованием BglII/XhoI и некоторых других сочетаний рестрикционных ферментов, например, BamHI/SalI, SalI (фиг.4 и 5).
Пример 7
Затем минипрепарат ДНК (Miniprep DNA) положительных клонов был выделен для очистки с последующей ПЭГ-преципитацией для анализа последовательности (фиг.2). Последовательность была подтверждена с использованием соответствующих прямых и обратных праймеров. После подтверждения последовательности, данные клоны выращивали в течение ночи при 37°С и хранили при -70°С. Около 10 клонов были признаны, утверждены как положительные посредством анализа последовательности, Секвенирование было проведено с использованием ABI-Prism.
Среди 10 клонов, основываясь на подтверждении уровня экспрессии, был выбран конечный клон для культивирования в 1 л ферментере для протекания далее технологического способа.
Пример 8. Подготовка высевания культуры
Штамм Escherichia coli BL21, трансформированный для экспрессирования rhPTH (1-34), очищали и хранили как глицериновый стоковый раствор (сток). Аликвоту глицеринового стока переносили рассевающим штрихом на 2,5% YE чашку (2,5% дрожжевой экстракт; 0,5% хлорид натрия рН 7,4; 1,5% агар-агар), содержащую 50 мкг/мл ампициллина, и инкубировали при 37°С в течение 24 ч для получения изолированных колоний. Одиночную колонию высевали в 10 мл 2,5% YE жидкой среды (2,5% дрожжевой экстракт; 0,5% хлорид натрия рН 7,4) с добавлением 50 мкл/мл ампициллина на пробирку «falcon» и инкубировали при 37°С в течение периода времени от 8 до 16 ч. 5 мл культуры из пробирки пересевали в коническую колбу с объемом 500 мл, содержавшую 100 мл основной питательной (базальной) среды. Колбу инкубировали при 37°С на роторном шейкере при 200 об/мин в течение 8 ч.
Пример 9. Периодическая ферментация с добавлением субстрата
Вышеупомянутую культуру из конической колбы использовали для инокулирования в 2 л Jar ферментер (реактор), содержащий 600 мл основной питательной среды. Ферментацию осуществляли при 37°С, поддерживали значение рН 7 с помощью 12,5% раствора аммония. Перемешивание осуществляли при 1200 об/мин с подачей воздуха 10 л в минуту, поддерживаемой на всем протяжении ферментации. Питательную среду закачивали в ферментер, следуя стратегии питания, как указано на фиг.6. Индукцию проводили через 18 ч после начала ферментации с помощью 20% раствора лактозного субстрата при скорости 0,25 мл/мин. Раствор пеногасителя на основе силикона использовали, чтобы контролировать излишнее вспенивание. Ферментацию проводили в течение 24 ч, в течение которых отбирали образцы для определения оптической плотности и продукции rhPTH (1-34). Выход в различные моменты времени оценивали при помощи сканирования SDS-PAGE гелей, окрашенных Кумасси синим (таблица 1). Анализ полученных образцов ферментационной среды показал, что средний выход химерного белка, представляющего интерес, составляет приблизительно 3 gms/л после первой аффинной очистки. В таблице 2 приведена скорость подачи (питания), использованная в способе ферментации, описанной на фиг.7.
Пример 10. Состав базальной среды
Основная среда, использованная в способе ферментации, содержала растворы BS1, BS2, стоковый раствор MgSO4, раствор следовых элементов (TES) и стоковый раствор антибиотика.
Раствор BS1 приготавливали, растворяя 2,5-15 г источника углерода, такого как глюкоза или глицерин, и 2,5-15 г источника азота, такого как дрожжевой экстракт или соевый пептон, в 400-600 мл RO воды.
Раствор BS2 приготавливали, растворяя 0,5-4 г сульфата аммония, 0,8-3,2 г KH2PO4 и 3,3-13,2 г Na2HPO4.2H2O и 0,45-1,8 г хлорида натрия в 50-200 мл RO воды.
Стоковый раствор MgSO4 приготавливали, растворяя 61,7-246,5 г MgSO4.2H2O в 1000 мл RO воды, и автоклавировали.
Раствор следовых элементов TES приготавливали, растворяя 0,13-1,24 г Н3ВО3, 0,88-0,322 г CoCl2·6H2O, 0,025-0,1 г NaMoO4·2H2O, 0,088-0,352 г CaCl2·2H2O, 0,125-0,5 г MnSO4·2H2O, 2,1-8,35 г FeCl3 и 0,0125-0,05 г CuSO4·5H2O и 0,05-0,2 г ZnSO4·7H2O в 500 мл RO воды. Раствор стерилизовали фильтрованием. Стоковый раствор Антибиотика состоял из простерилизованного фильтрованием ампициллина (50 мг/мл).
Базальная среда была приготовлена следующим образом:
Перед автоклавированием смешивали 400-600 мл BS1 с 100-1000 микролитрами раствора пеногасителя (пеногаситель Dow coming 1510). Для получения основной среды к данному раствору добавляли смесь из 50-200 мл раствора BS2, 1-5 мл стокового раствора MgSO4, 1-5 мл раствора TES и 1-1,5 мл стокового раствора антибиотика. Концентрация источников углерода и азота в основной среде находилась в пределах 0,25-1,5 об./вес. и 0,25-1,5 об./вес., соответственно.
Пример 11. Состав питательной среды
Питательная среда, использованная в способе ферментации, содержала растворы FS1, FS2, FS3 и TES. Раствор FS1 получали растворением 100-200 г источника углерода, такого как глюкоза или глицерин, в 200-300 мл RO воды. Раствор FS2 получали растворением 100-200 г источника азота, такого как дрожжевой экстракт или соевый пептон, в 250-350 мл RO. Раствор FS3 получали растворением 8-10 г KH2PO4, 6-9 г Na2HPO4·2H2O и 6-9 г K2HPO4 в 25-75 мл RO воды. Все вышеуказанные растворы стерилизовали автоклавированием. Питательную среду приготавливали, смешивая 200-300 мл раствора FS1, 250-350 мл раствора FS2, 25-75 мл раствора FS3 и 18-25 мл раствора TES. Концентрация источника углерода, источника азота и неорганических фосфатов в питательной среде составляла 10-30% об/вес и 10-30% об/вес и 2,5-4,25% об/вес, соответственно. Клетки, полученные после ферментации (осуществленной согласно примеру 9), осаждали и подвергали дальнейшей очистке.
Пример 12. Очистка клеток (от посторонних частиц), лизис и растворение
Ферментационный бульон центрифугировали для получения клеточного осадка, который суспендировали в лизирующем буферном растворе (50 мМ Tris; pH 8,0; 150 мМ NaCl) и гомогенизировали. После охлаждения гомогенизированного раствора до ~10 град клетки лизировали с помощью разрушения давлением при 850 бар и проводили три цикла лизирования. К полученному раствору добавляли 8М мочевину и перемешивали, чтобы солюбилизировать белки. После солюбилизации образец (фиг.7) фильтровали через 0,45 мкм мембрану, что помогало избежать channeling (просачивания) при первоначальной ЕВА хроматографии.
Пример 13. Первичный «захват» целевого белка
Отфильтрованный раствор солюбилизированных белков был нанесен на вспученный слой (expanded bed) проточной хелатной сефарозной колонки после уравновешивания колонки буферным раствором А, содержащим 20 мМ Tris pH 8,0, 150 мМ NaCl, 8М мочевины, и буферным раствором В, содержащим 20 мМ Tris pH 8,0, 150 мМ NaCl, 8М мочевины, 250 мМ имидазола.
После нанесения образца колонку промывали буферным раствором А, и затем связанный белок элюировали буферным раствором В. Хроматограмма и анализ SDS-PAGE показаны на фиг.8 и фиг.9, соответственно.
Пример 14. Расщепление химерного белка энтерокиназой
ЕВА элюат разбавили 20 мМ Tris pH 8,0 в восемь раз до получения окончательной концентрации мочевины ~1М и добавили энтерокиназу в отношении 1:100 (фермент:субстрат) при перемешивании при комнатной температуре в течение 1 ч. Расщепленный образец анализировали в геле Трис-Трицин (показан на фиг.10).
Пример 15. Катионообменная хроматография расщепленного образца
После EK расщепления в течение 1 часа, значение pH образца доводили до 7,0 при помощи HCl, а затем образец наносили на катионообменную колонку (SP-XL), которая была уравновешена буферным раствором А до нанесения образца. После нанесения образца колонку промывали буферным раствором А, содержащим 20 мМ ацетата натрия рН 5,5, и элюировали связанный белок градиентом NaCl в буфере В: 20 мМ ацетата натрия рН 5,5, 1 М NaCl. Значение рН образца доводили до 7,0 перед нанесением на колонку, поскольку было рассчитано, что заряд химерного tagged (помеченного) РТН (нерасщепленного) и химерного tag должен быть отрицательным, а РТН положительно заряжен, следовательно, ожидалось, что только РТН будет связываться с катионообменником, и были получены сходные результаты.
На фиг.11 показаны катионообменная хроматограмма и анализ фракций с помощью геля Трис-Трицин.
Пример 16а. Хроматография гидрофобных взаимодействий для окончательной очистки
К вышеуказанному IEX-элюату добавили сульфат аммония до конечной концентрации 1,75М. После тщательного перемешивания образец наносили на колонку с фенил-сефарозой, до нанесения образца уравновешенную буфером А, содержащим 20 мМ ацетата натрия рН 5,5, 1,75 М сульфата аммония. После нанесения образца колонку промывали буфером А и затем для окончательной очистки проводили градиентную элюцию с буфером В, содержащим 20 мМ ацетата натрия рН 5,5. Согласно RP-HPLC анализу основной пик (см. фиг.12) содержит >99% чистого PTH (1-34). Загрязнения, элюированные до основного пика, обозначены на хроматограмме как 'fr-1'. Результат анализа fr-1 с помощью RP-HPLC приведен ниже (фиг.14).
Пример 16б. Обращенно-фазовая хроматография для окончательной очистки
Альтернативно была опробована окончательная очистка IEX-элюата с использованием препаративной обращенно-фазовой С4 кремниевой колонки. В данном случае IEX-элюат наносили на С4 колонку, уравновешенную буфером А (0,01% TFA в воде). Градиент вплоть до 30% буфера В (0,1% TFA в 90% ацетонитриле) в 5 объемах колонки (CV), затем изократическое промывание при 30% В 5 объемами колонки и затем резкое увеличение до 100% в 5 объемах колонки приводило к очень хорошему выходу PTH (1-34) с чистотой >99%. Фиг.15 демонстрирует профиль очистки rhPTH (1-34) с помощью обращенно-фазовой хроматографии.
Анализ окончательно очищенного PTH (1-34) проводили с помощью RP-HPLC, используя Bachem PTH (1-34) или Forteo в качестве контроля, и было подтверждено, что чистота постоянно составляет >99% (фиг.16).
PTH (1-34), очищенный вышеописанным способом, был исследован с помощью следующих методов:
Масса, равная 4117 Da, была определена с помощью MALDI-TOF.
Первыми пятью аминокислотами согласно данным N-концевого секвенирования являются Ser Val Ser Glu Iso.
Биологическое исследование с использованием метода цАМФ на клетках линии UMR106 доказало, что полученный нами РТН (1-34) является биологически активным.
Последовательности для "Нового ортогонального способа очистки паратиреоидного гормона человека (hPTH) (1-34)"
SEQ. ID NO:1
Длина: 297 по
Тип: ДНК
Организм: Человек
Признак:
Другая информация: Гибридный нуклеотид ORF как NcoI/XhoI фрагмент hPTH (1-34)
Длина: 97 по
Тип: Белок
Организм:-
Признак:
Другая информация: hPTH (1-34) химерный белок
MGGAHHHHHHVDNVDESWLQEGQTRIIFDGVNSAFHLWINGRWV
GYGQDSRLPDVGAVDDDDKSVSEIQLMHNLGKHLN
SMERVEWLRKKLQDVHNF
SEQ ID NO.3
Длина: 31 по
Тип: ДНК
Организм: Искусственная последовательность
Признак:
Другая информация: Синтетический праймер
5'-CTTTTAGTGACCATGGTACCTGCAAAAGACA-3'
SEQ ID NO.4
Длина: 32 по
Тип: ДНК
Организм: Искусственная последовательность
Признак:
Другая информация: Синтетический праймер
5'-TCTGTTCTCGAGTTCACTGGGATTTAGCTTTA-3'
SEQ ID NO.5
Длина: 30 по
Тип: ДНК
Организм: Искусственная последовательность
Признак:
Другая информация: Синтетический праймер
5'-CACTATAGGGGAATTGTGAGCGGATAACAA-3'
SEQ ID NO.6
Длина: 37 по
Тип: ДНК
Организм: Искусственная последовательность
Признак:
Другая информация: Синтетический праймер
5'-TATTTCACTCACAGACTTGTCGTCGTCGTCGACCGCA-3'
SEQ ID NO.7
Длина: 41 по
Тип: ДНК
Организм: Искусственная последовательность
Признак:
Другая информация: Синтетический праймер
5'-GTCGACGACGACGACAAGTCTGTGAGTGAAATACAGCTTAT-3'
SEQ ID NO.8
Длина:45 по
Тип: ДНК
Организм: Искусственная последовательность
Признак:
Другая информация: Синтетический праймер
5'-TAGAGGAGCTCGAGGTTATTAAAAATTGTGCACATCCTGCAGCTT-3'








Изобретение относится к области биотехнологии, конкретно к получению рекомбинантных белков в Escherichia coli, и может быть использовано для синтеза паратиреоидного гормона человека (rhPTH (1-34)). Конструируют кДНК, кодирующую химерный нуклеотид ORF под контролем индуцибельного промотора, выбранного из группы, состоящей из araBAD, trp, T7, lac, Pho и trc, путем амплификации кДНК с помощью RT-PCR с использованием ген-специфических праймеров, кодирующих rhPTH (1-34) химерный гибридный белок, в котором химерный гибридный партнер по существу состоит из 41 аминокислот, полученных из 124-164 аминокислоты пептида бета-галактозидазы (LacZ) Escherichia coli. Затем экспрессируют химерный гибридный белок в присутствии лактозы в качестве индуктора в пределах около 10-30% общего белка на основе массы. Изобретение позволяет упростить и удешевить синтез rhPTH (1-34) за счет оптимизации его экспрессии в Escherichia coli. 11 з.п. ф-лы, 15 ил., 2 табл.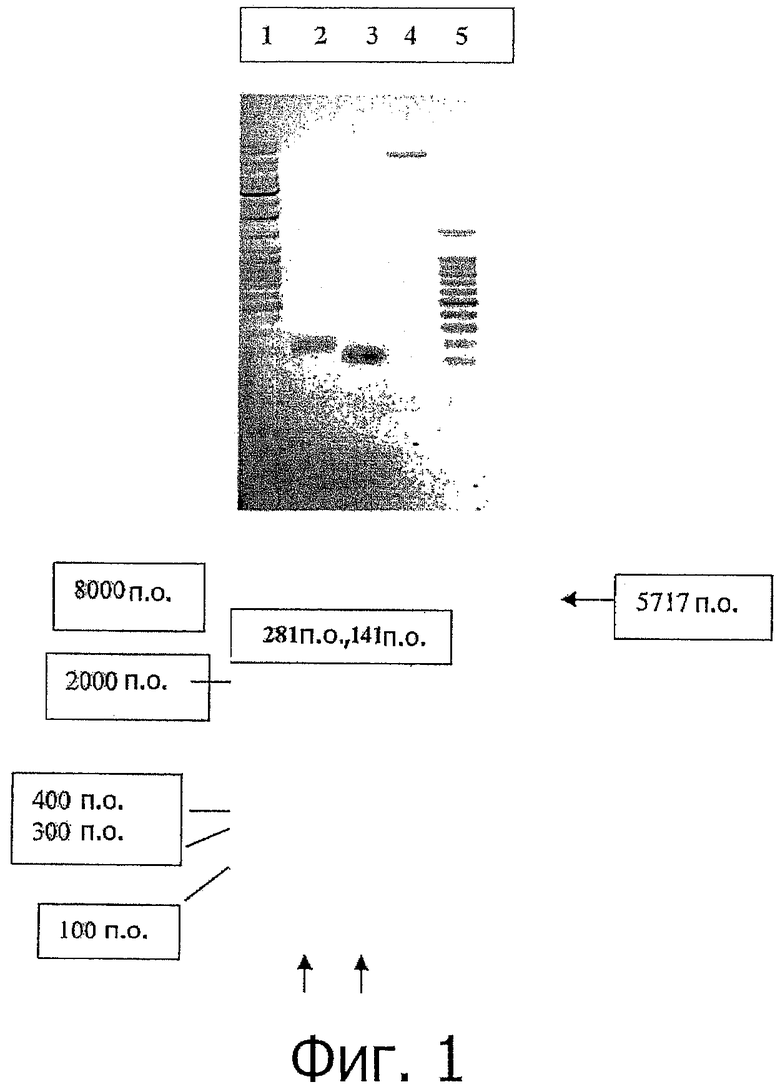
1. Способ синтеза rhPTH (1-34), включающий:
i) конструирование кДНК, кодирующей химерный нуклеотид ORF, как указано в SEQ ID NO:1 под контролем индуцибельного промотора, выбранного из группы, состоящей из araBAD, trp, T7, lac, Pho и trc, путем амплификации кДНК с помощью RT-PCR с использованием ген-специфических праймеров, выбранных из SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 и SEQ ID NO:8, кодирующих rhPTH (1-34) химерный гибридный белок, как указано в SEQ ID NO:2, в котором химерный гибридный партнер, по существу, состоит из 41 аминокислот, полученных из 124-164 аминокислоты пептида Р-галактозидазы (LacZ) Escherichia coli,
ii) экспрессирование химерного гибридного белка в присутствии лактозы в качестве индуктора в пределах около 10-30% общего белка на основе массы.
2. Способ по п.1, в котором вектор экспрессии, содержащий нуклеотид ORF как указано в SEQ ID NO:1, выбирают из группы, состоящей из pLMab, pRA, pET19b, в котором вектор представляет собой рЕТ19b.
3. Способ по п.1, в котором химерный гибридный белок, как указано в SEQ ID NO:2, состоит из аффинного конца, выбранного из группы, состоящей из полигистидин-tag, GST tag, связывающего мальтозу белка и стафилококкового белка, в котором аффинным концом является полигистидин-tag.
4. Способ синтеза rhPTH (1-34) по п.1, в котором полученный rhPTH (1-34) дополнительно очищают с помощью сочетания катионообменной хроматографии с хроматографией гидрофобного взаимодействия (HIC) или обращенно-фазовой ВЭЖХ (RP-HPLC) до степени очистки ≥99,5%, где содержание отдельной максимальной примеси составляет не более 0,2%.
5. Способ по п.4, в котором катионообменную хроматографию проводят на колонке, выбранной из группы, состоящей из SP-XL, SP-FF, Source-30S и Source-15S, в котором колонка представляет собой SP-XL.
6. Способ по п.4, в котором rhPTH (1-34) связывается с катионообменной колонкой, и колонку промывают, используя промывной буфер, имеющий рН в пределах от 3,5 до 5,5 и проводимость в пределах от 1 mS/см до 3 mS/см.
7. Способ по п.4, в котором rhPTH (1-34) элюирует из катионообменной хроматографической колонки при значении рН в пределах от 4,0 до 5,5 и проводимости в пределах от 15 mS/см до 35 mS/см.
8. Способ по п.4, в котором на стадии HIC используют колонку, состоящую из сшитых агарозных бусин с лигандами, выбранными из группы, состоящей из этила, пропила, изопропила, бутила, изобутила, третичного бутила, пентила, изопентила, гексила, фенила, изопропанола, изобутанола, С4-С6 эфира, в котором колонка представляет собой сефарозу, а лиганд представляет собой фенил.
9. Способ по п.4, в котором HIC-колонку промывают раствором промывного буфера, содержащим 20 мМ ацетатного буфера и от 1000 до 2000 мМ сульфата аммония с рН 5,5.
10. Способ по п.4, в котором HIC-колонку промывают буфером для элюирования, содержащим от 10 до 50 мМ ацетатного буфера с рН 5,5 в градиенте или изократическим способом, или их комбинацией.
11. Способ по п.1, в котором RP-HPLC колонку выбирают из группы, состоящей из С4, С8 и С18, в котором колонка представляет собой С4.
12. Способ по п.1, в котором rhPTH (1-34) имеет массу около 4117 Да.
| YUJI SUZUKI et al., High-Level Production of Recombinant Human Parathyroid Hormone 1-34, Appl | |||
| Environ | |||
| Microbiol., 1998, v | |||
| Нефтяной конвертер | 1922 |
|
SU64A1 |
| WO 03100022 A2, 04.12.2003 | |||
| DING BANG et al., Fusion protein Trx-PTH (1-34) of thioredoxin and parathyroid hormone PTH1-34, preparation method and use thereof, Database CA/STN N 145:286072, CN 1807456 | |||

