Перекрестные ссылки к родственным изобретениям
Настоящая заявка утверждает приоритет и преимущество заявки США на временное применение, серийный №60/670541, заполненной 11 апреля 2005 года, раскрытие которой включено сюда посредством ссылки.
Область изобретения
Настоящее изобретение касается генетически модифицированных белков, имеющих уриколитическую активность. Более конкретно, изобретение касается белков, содержащих усеченный вариант урат-оксидазы, и методов их получения.
Предпосылки к созданию изобретения
Термин урат-оксидаза и уриказа используются здесь взаимозаменяемо. Урат-оксидаза (уриказа; E.C. 1.7.3.3) являются ферментами, катализирующими окисление мочевой кислоты до более растворимого продукта, аллантоина, пуринового метаболита, подвергающегося более легкой экскреции. В человеческом организме не образуется энзиматически активной уриказы, что является результатом нескольких мутаций в гене уриказы, приобретенных во время эволюции высших приматов. Wu, X. et al., (1992) J Mol Evol 34: 78-84, включенная сюда во всем объеме посредством ссылки. В результате этого у восприимчивых индивидуумов повышенная концентрация мочевой кислоты в крови (гиперурикемия) может привести к болезненным артритам (подагре), обезображивающим отложениям мочевины (отложений) и почечной недостаточности. Hande, KR, et al., (1984) Am J Med 76: 47-56; Fam, AG, (1990) Bailliere's Clin Rheumatol 4: 177-192, каждое из которого включено сюда во всем объеме посредством ссылки. Инъекция уриказы может ослабить гиперурикемию и гиперурикозурия, по меньшей мере временно. Поскольку уриказа является чужеродным белком для человеческого организма, даже первая инъекция немодифицированного белка, полученного из Aspergillus flavus, приводит к анафилактическим реакциям у нескольких процентов пациентов (Pui, C-H, et al., (1997) Leukemia 11: 1813-1816, включенная сюда во всем объеме посредством ссылки), и имунная реакция ограничивает его использование для хроничекой или периодической терапии. Donadio, D, et al., (1981) Nouv Presse Med 10: 711-712; Leaustic, M, et al., (1983) Rev Rhum Mal Osteoartic 50: 553-554, каждое из которых включено сюда во всем объеме посредством ссылки.
Настоящее изобретение касается мутантной рекомбинантной уриказы, имеющей усечения и увеличенную структурную стабильность.
Сущность изобретения
Объектом настоящего изобретения является представление новых рекомбинантных белков уриказ. Белки рассматриваемого изобретения будут усеченными и будут иметь мутированные аминокислоты по отношению к встречающимся в природе белкам уриказам.
Рассматриваемое изобретение предоставляет мутантную рекомбинантную уриказу, содержащую последовательность аминокислот №8 (SEQ ID NO. 8). Также предоставлена мутантная рекомбинантная уриказа, содержащая последовательность аминокислот №13 (SEQ ID NO. 13).
Дополнительным объектом настоящего изобретения является предоставление способов метаболизирования мочевой кислоты, включающих новый рекомбинантный белок уриказу, имеющую уриколитическую активность. Уриколитическая активность понимается здесь как способность осуществлять энзиматическое превращение мочевой кислоты в аллантоин.
В варианте осуществления, уриказа содержит последовательность аминокислот с позиции 8 по позицию 287 последовательности №7 (SEQ ID NO. 7) или последовательности №12 (SEQ ID NO. 12). Также предоставлены уриказы, содержащие последовательности аминокислот №8 (SEQ ID NO. 8) или №13 (SEQ ID NO. 13). В варианте осуществления, уриказа содержит N-концевой аминокислотный остаток, являющийся остатком аланина, глицина, пролина, серина или треонина. В отдельном варианте осуществления, N-концевой остаток является остатком метионина.
Также предоставлены отдельные нуклеотидные последовательности, содержащие последовательности нуклеотидов, последовательность нуклеотидов, кодирующих уриказу изобретения. В отдельном варианте осуществления последовательность нуклеотидов, кодирующая уриказу, оперативно связана с гетерологичным промотором, например промотором osmB. Также предоставлены нуклеиновокислотные векторы, содержащие кодирующую уриказу нуклеотидную последовательность, клетки-хозяева, содержащие такие векторы, и способы получения уриказы, включающие стадии, культивирования таких клеток-хозяев при условиях, в которых последовательность нуклеотидов экспрессируется в клетках-хозяевах, и выделение экспрессируемой уриказы.
Краткое описание чертежей
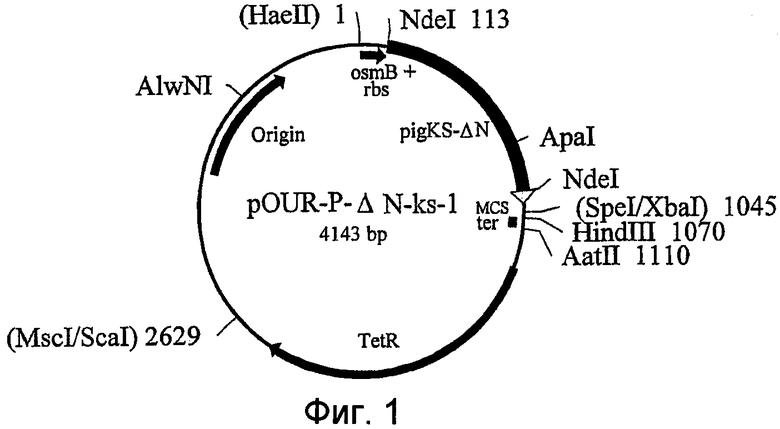
Фиг.1 иллюстрирует структуру плазмиды pOUR-P-ΔN-ks-1. Цифры возле сайтов рестрикции обозначают номера нуклеотидов относительно сайта HaeII, принятого за 1. В скобках обозначены сайты рестрикции, исчезающие во время процедуры клонирования.
Фиг.2 изображает последовательность ДНК и транслированную последовательность аминокислот уриказы Pig-KS-ΔN (последовательность №9 [SEQ ID NO. 9] и №7 [SEQ ID NO. 7] соответственно). Номера аминокислотных остатков на Фиг.2 даны относительно полной последовательности уриказы свиньи. Следующий за первым метионином треонин соответствует замещаемому аспартату в положении 7 последовательности уриказы свиньи. Обозначены используемые на различных стадиях субклонирования сайты рестрикции. 3'-нетранслируемая область выделена строчными буквами. Стоп-кодон трансляции обозначен звездочкой.
Фиг.3 демонстрирует выравнивание транслированных последовательностей аминокислот различных уриказ: рекомбинантной свиной (последовательность №11 [SEQ ID NO. 11]), PBC-ΔNC (последовательность №12 [SEQ ID NO. 12]) и Pig-KS-ΔN (последовательность №7 [SEQ ID NO. 7]). Звездочками отмечены положения, в которых имеются различия в аминокислотной последовательности pig-KS-ΔN и опубликованной последовательности уриказы свиньи; кружками обозначены положения, в которых имеются различия в аминокислотной последовательности pig-KS-ΔN и PBC-ΔN. Пунктирными линиями обозначены отсутствующие аминокислоты.
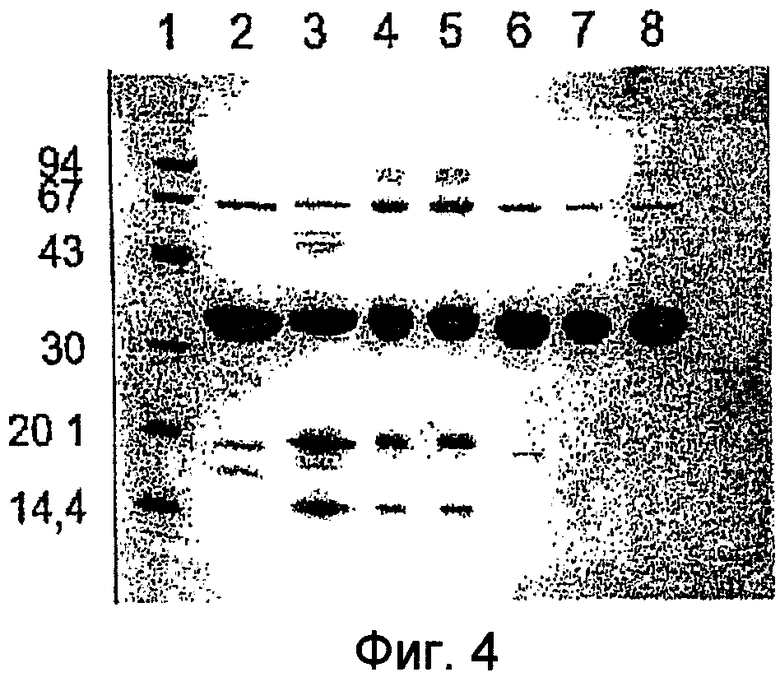
Фиг.4 демонстрирует СДС-ПААГ (разделение в денатурирующем полиакриламидном геле) уриказы свиньи и высоко очищенных вариантов уриказы, полученных согласно Примерам 1-3. Дата получения (месяц/год) и соответствующий номер дорожки для каждой пробы даны в нижеследующем ключе. По оси Y отмечены молекулярные массы белков маркера, номера дорожек обозначены наверху чертежа. На дорожки нанесены: дорожка 1 - маркер молекулярной массы; дорожка 2 - Pig KS-ΔN (7/98); дорожка 3 - Pig (9/98): дорожка 4 - Pig KS (6/99); дорожка 5 - Pig KS (6/99); дорожка 6 - Pig-ΔN (6/99); дорожка 7 - Pig KS-ΔN (7/99); дорожка 8 - Pig KS-ΔN (8/99).
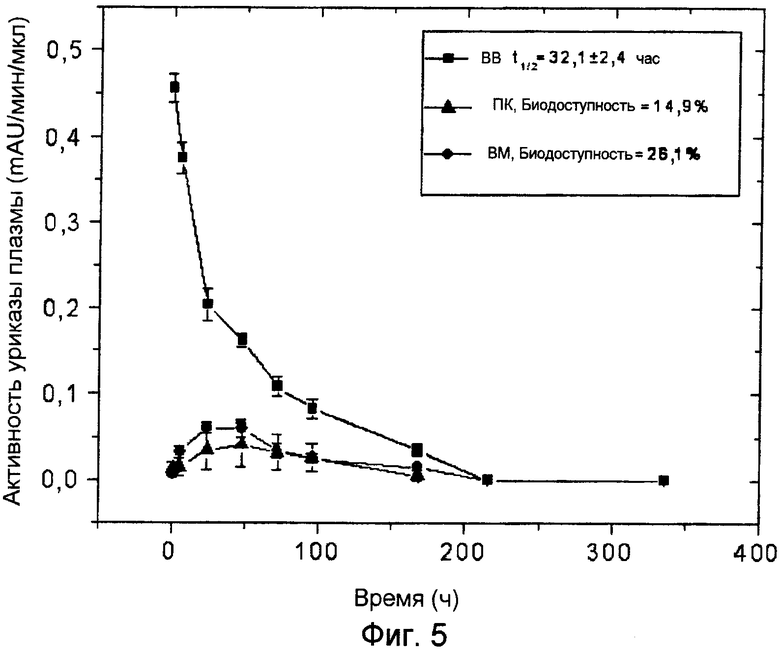
Фиг.5 иллюстрирует фармакокинетические профили ПЭГ-конъюгированной (9×10 кДа) Pig-KS-ΔN уриказы у крыс после ВМ (внутримышечного), ПК (подкожного) и ВВ (внутривенного) введения, определенные с помощью мониторинга энзиматической активности в пробах крови. Активность уриказы в пробах плазмы, отобранных в указанные временные точки, определяется с помощью колориметрии. Значения активности (mAU = тысячная доля единицы абсорбции) представляют скорость энзиматической реакции на 1 мкл плазмы крови. Биодоступность (количество вещества, попавшего в кровь по отношению к внутривенной инъекции) введенной уриказы была рассчитана, исходя из площади кривой на графике.
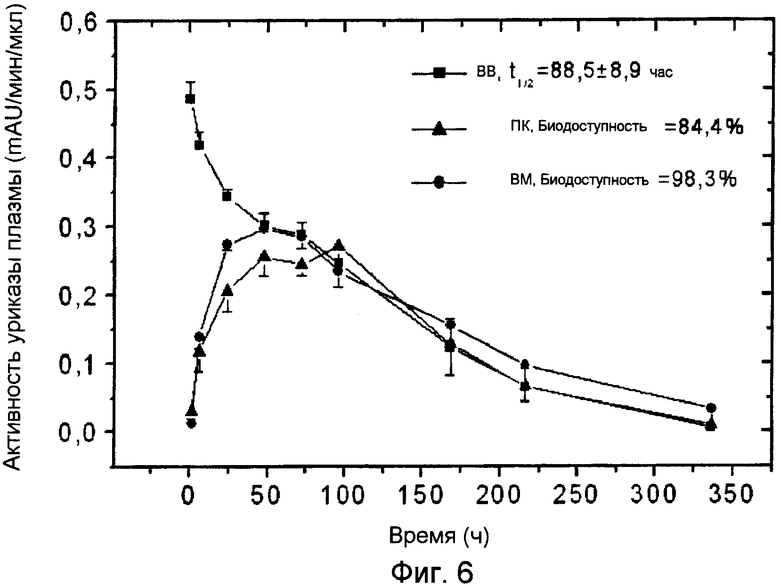
Фиг.6 иллюстрирует фармакокинетические профили ПЭГ-конъюгированной (9×10 кДа) Pig-KS-ΔN уриказы у кроликов после ВМ (внутримышечного), ПК (подкожного) и ВВ (внутривенного) введения, определенные с помощью мониторинга энзиматической активности в пробах крови. Активность уриказы в пробах плазмы, отобранных в указанные временные точки, определяется с помощью колориметрии. Значения активности (mAU = тысячная доля единицы абсорбции) представляют скорость энзиматической реакции на 1 мкл плазмы крови. Биодоступность (количество вещества, попавшего в кровь по отношению к внутривенной инъекции) введенной уриказы была рассчитана, исходя из площади кривой на графике.
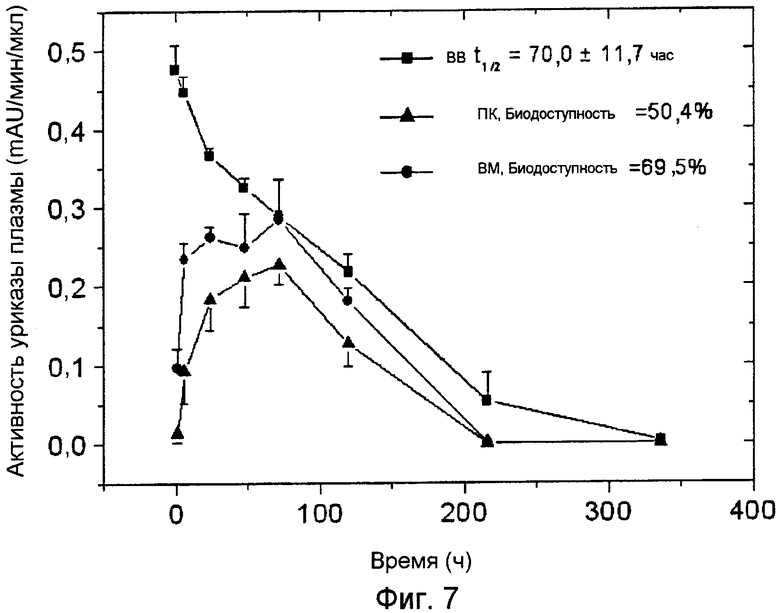
Фиг.7 иллюстрирует фармакокинетические профили ПЭГ-конъюгированной (9×10 кДа) Pig-KS-ΔN уриказы у собак после ВМ (внутримышечного), ПК (подкожного) и ВВ (внутривенного) введения, определенные с помощью мониторинга энзиматической активности в пробах крови. Активность уриказы в пробах плазмы, отбираемых в указанные временные точки, определяется с помощью колориметрии. Значения активности (mAU = тысячная доля единицы абсорбции) представляют скорость энзиматической реакции на 1 мкл плазмы крови. Биодоступность (количество вещества, попавшего в кровь по отношению к внутривенной инъекции) введенной уриказы была рассчитана, исходя из площади кривой на графике.
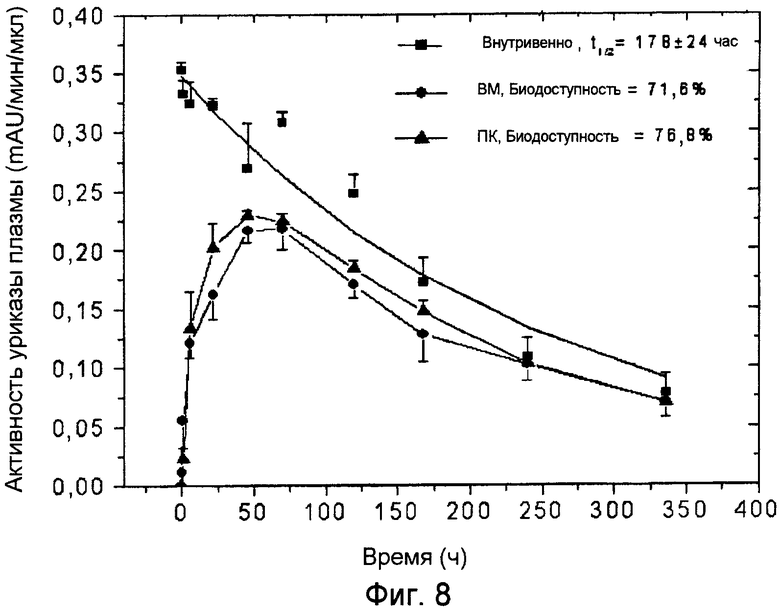
Фиг.8 иллюстрирует фармакокинетические профили ПЭГ-конъюгированной (9×10 кДа) Pig-KS-ΔN уриказы у свиней после ВМ (внутримышечного), ПК (подкожного) и ВВ (внутривенного) введения, определенные с помощью мониторинга энзиматической активности в пробах крови. Активность уриказы в пробах плазмы, отбираемых в указанные временные точки, определяется с помощью колориметрии. Значения активности (mAU = тысячная доля единицы абсорбции) представляют скорость энзиматической реакции на 1 мкл плазмы крови. Биодоступность (количество вещества, попавшего в кровь по отношению к внутривенной инъекции) введенной уриказы была рассчитана, исходя из площади под кривой на графике.
Подробное описание изобретения
Из предыдущих исследований известно, что хотя значительное снижение иммуногенности и/или антигенности уриказы достигается при конъюгировании с ПЭГ (полиэтиленгликоль), это ассоциировано с существенной потерей уриколитической активности. В безопасность, удобство и экономическую эффективность биофармацевтических препаратов вносит отрицательный вклад снижение их действенности и, как следствие, необходимость увеличения применяемой дозы. Таким образом, существует потребность в безопасных и эффективных альтернативных способах снижения повышенного уровня мочевой кислоты в жидкостях тела, включая кровь. Настоящее изобретение предоставляет мутантную рекомбинантную уриказу, содержащую аминокислотные последовательности №7, 8, 12 или 13 (SEQ ID NO. 7, SEQ ID NO. 8, SEQ ID NO 12 или SEQ ID NO. 13).
Уриказа, как здесь использовано, включает индивидуальные субъединицы, так же как тетрамер, если не обозначено другое.
В отдельном варианте осуществления уриказа имеет N-концевой метионин. Этот метионин может быть изъят при посттрансляционных модификациях. В отдельном варианте осуществления N-концевой метионин изымается после получения уриказы. В отдельном варианте осуществления, метионин изымается эндогенной бактериальной аминопептидазой. Предпоследней аминокислотой может быть та, которая позволяет изъятие N-концевого метионина бактериальной метионин-аминопептидазой (MAP). Аминокислотами, позволяющими наиболее полное изъятие N-концевого метионин, являются аланин, глицин, пролин, серин и треонин. В отдельном варианте осуществления, уриказа содержит две N-концевых аминокислоты, где две N-концевых аминокислоты являются метионином с последующей аминокислотой, выбираемой из группы, состоящей из аланина, глицина, пролина, серина и треонина.
Рассматриваемое изобретение предоставляет последовательность нуклеотидов, кодирующую уриказу.
Рассматриваемое изобретение предоставляет вектор, содержащий эту последовательность нуклеотидов.
В отдельном варианте осуществления уриказу выделяют. В отдельном варианте осуществления уриказу очищают. В отдельном варианте осуществления уриказу выделяют и очищают.
Рассматриваемое изобретение предоставляет клетки-хозяева, содержащие вектор, содержащий кодирующую уриказу последовательность нуклеотидов.
Рассматриваемое изобретение предоставляет способ получения кодирующей уриказу последовательности нуклеотидов, включающий модификацию кодирующей уриказу последовательности нуклеотидов методом ПЦР (полимеразной цепной реакции). Специалист в технологии понимает, что желаемая последовательность нуклеотидов получается в ПЦР при использовании синтетических олигонуклеотидных праймеров, комплементарных участку ДНК (один для каждой цепи), который должен быть амплифицирован. Праймеры добавляют кДНК-мишени (которая не должна быть чистой) в присутствии избытка дезоксинуклеотидов и Taq-полимеразы, устойчивой к нагреванию полимеразы. В серии (обычно 30) температурных циклов повторяется денатурация ДНК-мишени (примерно 90°С), отжига праймеров (обычно примерно 50-60°С) и удлинения дочерних цепей с праймеров (72°С). Поскольку дочерние цепи сами по себе являются матрицами для последующих циклов, фрагменты ДНК, соответствующие обоим праймерам, амплифицируются скорее экспоненциально, чем линейно.
Рассматриваемое изобретение предоставляет способ получения мутантной рекомбинантной уриказы, включающий трансфекцию клеток-хозяев вектором, где клетки-хозяева экспрессируют уриказу, выделение мутантной рекомбинантной уриказы из клеток-хозяев, выделение чистой мутантной рекомбинантной уриказы, например, используя подходы хроматографии, и очистку мутантной рекомбинантной уриказы. Например, уриказа может быть получена в соответствии с методом, описанным в международной патентной публикации № WO 00/08196 и заявке США №60/095489, включенных сюда во всем объеме посредством ссылок.
В предпочтительном варианте осуществления клетки-хозяева обрабатывают таким образом, что это приводит к экспрессии мутантной рекомбинантной уриказы. Специалист в технологии осведомлен, что трансфекция клеток вектором обычно совершается с использованием ДНК, осажденной с ионами кальция, хотя различные другие методы могут быть использованы (например, электропорация).
Уриказа может быть выделена и/или очищена любым методом, известным специалисту в технологии. Экспрессируемые полипептиды данного изобретения обычно выделяются в достаточно чистой форме. Предпочтительно, полипептиды выделяются до чистоты как минимум 80% по массе, более предпочтительно - до чистоты как минимум 95% по массе, и более предпочтительно до чистоты как минимум 99% по массе. В общем, такая очистка может быть достигнута, например, стандартными техниками высаливания сульфатом аммония, СДС-ПААГ и афинной хроматографией. Уриказу предпочтительно изолируют с использованием катионного сурфактанта, например хлорид цетил-пиридина (CPC), в соответствии с методом, описанным в рассматриваемых одновременно заявке на патент США, заполненной 11 апреля 2005 года, имеющей номер 60/670520 и номер 103864.146644 по картотеке патентного поверенного, озаглавленным «Очистка белков с катионными сурфактантами», включенным сюда во всем объеме посредством ссылки.
В варианте осуществления изобретения экспрессия последовательности с вектора находится под контролем промотора, чувствительного к осмотическому стрессу. Промотор - это участок ДНК, с которым связывается РНК-полимераза перед инициацией транскрипции ДНК в РНК. Промотор, чувствительный к осмотическому стрессу, инициирует транскрипцию в результате ощущаемого клеткой осмотического давления.
Уриказа изобретения включает уриказу, конъюгированную с полимером, например, уриказу, конъюгированную с полиэтиленгликолем, т.е. ПЭГ-конъюгированную уриказу.
В варианте осуществления изобретения предоставлена фармацевтическая композиция, содержащая уриказу. В одном варианте осуществления композиция является раствором уриказы. В предпочтительном варианте осуществления раствор стерилен и пригоден для инъекции. В одном варианте осуществления такая композиция содержит уриказу в качестве раствора в фосфатно-солевом буфере. В одном варианте осуществления композиция поставляется в пузырьках, имеющих необязательно резиновый инъекционный стопор. В отдельном варианте осуществления композиция содержит уриказу в растворе в концентрации от 2 до 16 миллиграммов уриказы на миллилитр раствора, от 4 до 12 миллиграммов на миллилитр или от 6 до 10 миллиграммов на миллилитр. В предпочтительном варианте осуществления композиция содержит уриказу в концентрации 8 миллиграммов на миллилитр. Предпочтительно, масса уриказы измеряется с учетом массы белка.
Эффективный режим применения композиции изобретения может быть определен специалистом в технологии. Подходящие индикаторы для оценки эффективности данного режима известны специалисту в технологии. Примеры таких индикаторов включают нормализацию или понижение уровня мочевой кислоты в плазме (PUA) и понижение или поддержание PUA на уровне 6,8 мг/мл или ниже, предпочтительно 6 мг/мл или ниже. В предпочтительном варианте осуществления субъект, проходящий лечение с использованием данной композиции, имеет PUA на уровне 6 мг/мл или ниже в течение как минимум 70%, как минимум 80% или как минимум 90% общего времени лечения. Например, для периода времени в 24 недели, субъект предпочтительно имеет PUA на уровне 6 мг/мл или ниже в течение как минимум 80% 24-недельного периода времени, т.е. в течение как минимум времени, равного 134,4 дням (24 недели × 7 дней/неделю × 0,8 = 134,4 дня).
В отдельном варианте осуществления от 0,5 до 24 мг растворенной уриказы вводят раз в каждые 2-4 недели. Уриказа может быть введена любым подходящим способом, известным специалисту в технологии, например внутривенно, внутримышечно или подкожно. Предпочтительно, в случае внутривенного введения, вводят от 0,5 до 12 мг уриказы. Предпочтительно, в случае подкожного введения, вводят от 4 до 24 мг уриказы. В предпочтительном варианте осуществления введение уриказы при внутривенной инфузии осуществляется в течение периода времени от 30 до 240 минут. В одном варианте осуществления 8 мг уриказы вводят раз в каждые две недели. В отдельном варианте осуществления инфузия может быть осуществлена с использованием от 100 до 500 мл раствора. В предпочтительном варианте осуществления 8 мг раствора уриказы вводят в течение 120 минут раз в каждые 2 недели или каждые 4 недели; предпочтительно уриказу растворяют в 250 мл солевого раствора для инфузии (физиологического раствора). В отдельном варианте осуществления лечение с применением уриказы осуществляют в течение 3 месяцев, 6 месяцев, 8 месяцев или 12 месяцев. В другом варианте осуществления, период лечения составляет 12 недель, 24 недели, 36 недель или 48 недель. В отдельном варианте осуществления период лечения составляет более продолжительное время, например 2 года или дольше, вплоть до всей продолжительности жизни субъекта. В довершение множественные периоды лечения могут быть прерываемы периодами без лечения, например, 6 месяцев лечения с последующими 3 месяцами без лечения, затем 6 месяцев лечения и т.д.
В определенном варианте осуществления противовоспалительные соединения могут быть применены для элиминирования или уменьшения реакций на инфузию из-за применения уриказы. В одном варианте осуществления как минимум один кортикостероид, как минимум один антигистамин, как минимум один NSAID (нестероидное антивоспалительное лекарство) или их комбинации применяют таким образом. Подходящие кортикостероиды включают бетаметазон, будесонид, кортизон, дексаметазон, гидрокортизон, метилпреднизолон преднизолон, преднизон и триамцинолон. Подходящие NSAID включают ибупрофен, индометацин, напроксен, аспирин, ацетоминофен, целекоксиб и валдекоксиб. Подходящие антигистамины включают азатадин, бромфенирамин, цетиризин, хлорфенирамин, клемастин, ципрогептадин, деслоратадин, дексохлорфенирамин, дименгидринат, дифенгидрамин, доксиламин, фексофенадин, гидроксизин, лоратадин и фениндамин.
В предпочтительном варианте осуществления антигистамином является фексофенадин, NSAID является ацетаминофен и кортикостероидом является гидрокортизон и/или преднизон. Предпочтительно, комбинация всех трех компонентов (необязательно одновременно) применяется перед инфузией раствора уриказы. В предпочтительном варианте осуществления NSAID и антигистамин применяются перорально за 1-4 часа перед инфузией уриказы. Подходящая доза фексофенадина включает количества от примерно 30 до примерно 180 мг, от примерно 40 до примерно 150 мг, от примерно 50 до примерно 120 мг, от примерно 60 до примерно 90 мг, примерно 60 мг, предпочтительно 60 мг. Подходящая доза ацетоминофена включает количества от примерно 500 до примерно 1500 мг, от примерно 700 до примерно 1200 мг, от примерно 800 до примерно 1100 мг, примерно 1000 мг, предпочтительно 1000 мг. Подходящая доза гидрокортизона включает количества от примерно 100 до примерно 500 мг, от примерно 150 до примерно 300 мг, примерно 200 мг, предпочтительно 200 мг. В одном варианте осуществления антигистамин не является дифенгидрамином. В другом варианте осуществления, NSAID не является ацетоминофеном. В предпочтительном варианте осуществления 60 мг фексофенадина применяется перорально за ночь до инфузии уриказы, 60 мг фексофенадина и 1000 мг ацетаминофена применяются перорально на следующее утро и, наконец, 200 мг гидрокортизона применяются непосредственно инфузией раствора уриказы. В одном варианте осуществления преднизон применяется за день, предпочтительно вечером, до применения уриказы. Подходящая доза преднизона включает количества от 5 до 50 мг, предпочтительно 20 мг. В определенном варианте осуществления эти профилактические процедуры для элиминирования или уменьшения случаев реакций на инфузию применяются по отношению к пациентам, получающим или готовым получить уриказу, в том числе ПЭГ-конъюгированную и ПЭГ-неконъюгированную уриказы. В отдельном варианте осуществления эти профилактические процедуры применяются по отношению к пациентам, получающим или готовым получить терапевтические пептиды, другие чем уриказа, в случает ПЭГ-конъюгированных или ПЭГ-неконъюгированных терапевтических пептидов.
В варианте осуществления фармацевтическая композиция содержит конъюгат уриказы с полимером, и модифицированная уриказа сохраняет уриколитическую активность. В предпочтительном варианте осуществления уриказа является ПЭГ-конъюгированной уриказой.
В варианте осуществления фармацевтическая композиция содержит уриказу, подвергшуюся модификации при конъюгировании с полимером, и модифицированная уриказа сохраняет уриколитическую активность. В отдельном варианте осуществления, конъюгат полимера и уриказы готовят в соответствии с описанием, приведенным в международной патентной публикации № WO 01/59078 и заявке США №09/501730, включенных сюда во всем объеме посредством ссылок.
В варианте осуществления полимер выбирают из группы, содержащей полиэтиленгликоль, декстран, полипропиленгликоль, гидроксипропилметилцеллюлозу, карбоксиметилцеллюлозу, поливинилпирролидон и поливиниловый спирт. В предпочтительном варианте осуществления полимером является полиэтиленгликоль и уриказа является ПЭГ-конъюгированной уриказой.
В варианте осуществления композиция содержит 2-12 молекул полимера на одну субъединицу уриказы, предпочтительно от 3 до 10 молекул полимера на одну субъединицу уриказы. В варианте осуществления каждая молекула полимера имеет молекулярный вес между примерно 1 и примерно 100 кДа.
В другом варианте осуществления изобретения каждая молекула полимера имеет молекулярный вес между примерно 1 и примерно 50 кДа. В предпочтительном варианте осуществления изобретения каждая молекула полимера имеет молекулярный вес между примерно 5 и примерно 20 кДа, примерно 8 и примерно 15 кДа, примерно 10 и примерно 12 кДА, предпочтительно примерно 10 кДа. В предпочтительном варианте осуществления каждая молекула полимера имеет молекулярный вес от примерно 5 до примерно 20 кДа. В особенно предпочтительном варианте осуществления изобретения каждая молекула полимера имеет молекулярный вес примерно 10 кДа.
В варианте осуществления изобретения композиция является подходящей для повторяемого введения композиции.
Объект изобретения предоставляет способы метаболизации мочевой кислоты с использованием уриказы.
Объект изобретения предоставляет использование соединения уриказы для уменьшения уровня мочевой кислоты в биологической жидкости.
В варианте осуществления изобретения композиция уриказы используется для уменьшения уровня мочевой кислоты в биологической жидкости, включая кровь.
Также предоставлены новые молекулы нуклеиновых кислот, кодирующих уриказу изобретения. Манипуляции, результатом которых является их производство, хорошо известны специалисту в технологии. Например, нуклеотидная последовательность уриказы может быть модифицирована с помощью любой из множества стратегий, известных в технологии (Maniatis T., 1990, Molecular Cloning, A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.). Последовательность может быть расщеплена по подходящим сайтам с помощью эндонуклеаз(ы) рестрикции, с последующей энзиматической модификацией, если желательно, изолирована и лигирована in vitro. При производстве гена, кодирующего уриказу, особое внимание следует уделить тому, чтобы модифицированный ген остался внутри подходящей рамки считывания, непрерываемой стоп-сигналом трансляции.
Последовательность нуклеотидов, кодирующая белок-уриказу, может быть вставлена в подходящий экспрессионный вектор, т.е. вектор, содержащий необходимые элементы для транскрипции и трансляции встроенной белок-кодирующей последовательности. Разнообразные системы векторов-носителей могут быть использованы для экспрессии белок-кодирующей последовательности. Они включают, но не только, линии клеток млекопитающих, инфицированных вирусом (например, вирусом коровьей оспы, аденовирусами и т.д.); линии клеток насекомых, инфицированных вирусами (например, бакуловирусами); микроорганизмы, такие как дрожжи, содержащие дрожжевые векторы или бактерии, трансформированные ДНК бактериофага, плазмидной ДНК или космидной ДНК. Экспрессионные элементы этих векторов различаются по силе и специфичности. В зависимости от используемой системы вектора-хозяина может быть использован любой из списка подходящих элементов для транскрипции и трансляции.
Любой из известных методов для введения фрагментов ДНК в вектор может быть использован для конструирования экспрессионного вектора, содержащего химерный ген, состоящий из подходящего элемента контроля транскрипции/трансляции и последовательности, кодирующей белок. Эти методы могут включать техники синтетические и in vitro рекомбинантной ДНК, а также in vivo рекомбинации (генетической рекомбинации). Экспрессия последовательностей нуклеиновых кислот, кодирующих белок-уриказу, может быть регулируема с помощью второй последовательности нуклеотидов таким образом, что белок-уриказа будет экспрессироваться в хозяине, трансформированном рекомбинантной молекулой ДНК. Например, экспрессия уриказы может быть контролируема любым промоторным/энхансерным элементом, известным в технологии. Промоторы, которые могут быть использованы для контроля экспрессии уриказы, включают, но не только, ранний промотор вируса SV40 (Bernoist and Chambon, 1981, Nature 290: 304-310), промотор, содержащийся в 3'-длинном концевом повторе вируса саркомы Рауса (Yamamoto, et al., 1980, Cell 22: 787-797), промотор тимидин киназы вируса герпеса (Wagner et al., 1981, Proc. Natl. Acad. Sci. U.S.A. 78: 144-1445), регуляторную область гена металлотионина (Brinster et al., 1982, Nature 296: 39-42); прокариотические экспрессионные векторы, такие как промотор β-лактамазы (Villa-Kamaroff, et al., 1978, Proc. Natl. Acad. Sci. U.S.A. 75:3727-3731), промотор tac (DeBoer, et al., 1983, Proc. Natl. Acad. Sci. U.S.A. 80: 21-25) и osmB промотор. В отдельном варианте осуществления нуклеиновая кислота, содержащая последовательность нуклеотидов, кодирующую уриказу, оперативно связана с гетерологичным промотором.
Когда данная рекомбинантная молекула ДНК, содержащая кодирующую последовательность нуклеотидов приготовлена и выделена, несколько известных в технологии методов могут быть использованы для ее размножения. Когда имеются налаженные система-хозяин и условия для роста, рекомбинантный экспрессионный вектор может быть размножен и приготовлен количественно. Как объяснено ранее, экспрессионные векторы, которые могут быть использованы, включают, но не только, следующие векторы или их производные: вирусы человека и животных, такие как вирусы коровьей оспы; вирусы насекомых, такие как бакуловирус; дрожжевые векторы; векторы бактериофагов (например, лямбда), и плазмидные и космидные ДНК-векторы, а также другие.
В добавление, могут быть выбраны линии клеток-хозяев, модулирующие экспрессию введенной последовательности, или модифицирующие и осуществляющие созревание продукта гена специфическим желательным образом. Экспрессия с определенных промоторов может быть увеличена в присутствии определенных добавок; так, можно контролировать экспрессию белка-уриказы, полученного с помощью генной инженерии. Более того, различные клетки-хозяева имеют характеристики и специфические механизмы для трансляционного и пост-трансляционного созревания и модификаций (например, гликозилирования, расщепления) белков. Подходящие линии клеток или систем-хозяев могут быть выбраны для обеспечения желаемых модификаций и созревания чужеродного экспрессируемого белка. Различные векторы/экспрессионные системы-хозяева могут по-разному влиять на реакции созревания, такие как протеолитическое расщепление.
В отдельном варианте осуществления изобретения экспрессия уриказы в E.coli предпочтительно осуществляется с использованием вектора, содержащего osmB промотор.
ПРИМЕРЫ
Пример 1. Создание гена и экспрессионной плазмиды для экспрессии уриказы
Рекомбинантную уриказу (урат-оксидазу) свиньи, Pig-KS-ΔN (усеченный с N-конца белок уриказы свиньи с замененными в положениях 291 и 301 аминокислотами на лизин и серин соответственно) экспрессировали в штамме E.coli K-12 W3110 F- Результатом создания серии плазмид была конструкция pOUR-P-ΔN-ks-1, с которой осуществлялась эффективная экспрессия уриказы при трансформации в клетки-хозяева E.coli.
Выделение и субклонирование кДНК уриказы из печени свиньи и бабуина
кДНК уриказы была приготовлена из печени свиньи и бабуина посредством выделения и субклонирования соответствующей РНК. Тотальная клеточная РНК была экстрагирована из печени свиньи и бабуина (Erlrich, H.A. (1989). PCR technology; Principles and Application for DNA Amplification; Sambrook, J., et al. (1989). Molecular cloning: A Laboratory Manual, 2nd edition; Ausubel, F.M. et al. (1998). Current protocols in molecular Biology), после чего была использована для обратной транскрипции с использованием First-Strand cDNA Synthesis Kit (Pharmacia Biotech). ПЦР-амплификацию проводили с использованием Taq ДНК-полимеразы (Gibco BRL, Life Technologies).
Последовательности синтетических олигонуклеотидов-праймеров, использованные для ПЦР-амплификации urate oxidase (уриказы) свиньи и бабуина, приведены в Таблице 1.
Последовательности нуклеотидов, выделенные строчными буквами в Таблице 1, являются введенными в последовательности праймеров рестрикционными сайтами: для рестриктаз EcoRI и NcoI (смысловой для свиньи и бабуина) и NcoI, HindIII и XbaI (антисмысловой для свиньи), XbaI и NcoI (антисмысловой для бабуина). Для смыслового праймера для бабуина, третий кодон GAC (кодирующий аспартат) в последовательности уриказы бабуина, был заменен на CAC (кодирующий гистидин) - кодон, присутствующий в данном положении в кодирующей последовательности псевдогена урат-оксидазы человека. Рекомбинантная конструкция уриказы бабуина, полученная с помощью этих праймеров, получила название D3H уриказа бабуина.
ПЦР-продукт, полученный для уриказы свиньи, был расщеплен с EcoRI и HindIII и клонирован в pUC18 для производства pUC18 - Pig Uricase. ПЦР-продукт, полученный для D3H уриказы бабуина, был клонирован непосредственно в pCR™II-вектор, используя TA Cloning™ (Invitrogen, Carlsbad, CA), для создания вектора pCR™II - D3H Baboon Uricase.
Лигированная ДНК была использована для трансформации штамма E.coli XL1-Blue (Stratagene, La Jolla, CA). Плазмидная ДНК, содержащая клонированную кДНК уриказы, была получена, клоны, содержащие опубликованную последовательность ДНК, кодирующую уриказу (кроме D3H замены в уриказе бабуина, указанной в таблице 1), были отобраны и выделены. В отобранном клоне pCR™II-D3H Baboon Uricase последовательности pCR™II оказались рядом со стоп-кодоном уриказы, получающимся при делеции последовательностей, введенным методом ПЦР. Как следствие, рестрикционные сайты XbaI и NcoI на 3'-конце были элиминированы, что сделало возможным прямое клонирование с использованием NcoI на 5'-конце ПЦР-продукта и BamHI, присутствующего в pCR™II векторе.
Субклонирование кДНК уриказы в экспрессионный вектор pET
Субклонирование уриказы бабуина
D3H кДНК бабуина, содержащая полноразмерную последовательность, кодирующую уриказу, была введена в экспрессионный вектор pET-3d (Novagen, Madison, WI). Конструкция pCR™II-D3H Baboon Uricase была расщеплена рестриктазами NcoI и BamHI, после чего был выделен продукт длиной 960 пар нуклеотидов. Экспрессионная плазмида pET-3d была расщеплена рестриктазами NcoI и BamHI, после чего был выделен фрагмент длиной 4600 пар оснований. Два фрагмента были лигированы для получения конструкции pET-3d-D3H-Baboon.
Субклонирование химерной уриказы свиньи-бабуина
Химерная уриказа свиньи-бабуина (PBC) была получена для достижения более высокого уровня экспрессии, стабильности и активности рекомбинантного гена. Конструкция PBC была получена посредством выделения фрагмента NcoI-ApaI длиной 4936 пар оснований из клона pET-3d-D3H-Baboon и лигирования его с выделенным фрагментом NcoI-ApaI длиной 624 пары оснований из pUC18-Pig Uricase, что привело к получению конструкции pET-3d-PBC. кДНК уриказы PBC состоит из кодонов 1-225 уриказы свиньи, соединенных в одной рамке с кодонами 226-304 уриказы бабуина.
Субклонирование уриказы Pig-KS
Уриказа Pig-KS была сконструирована для добавления одного остатка лизина, который может обеспечить дополнительный сайт конъюгации с ПЭГ. KS относится к введению аминокислоты лизина в последовательность уриказы свиньи в позицию 291, на место аргинина (R291K). В добавление, треонин в позиции 301 был замещен серином (T301S). Плазмида, несущая уриказу PigKS, была сконструирована из фрагмента NcoI-NdeI длиной 4696 пар оснований, выделенного из pET-3d-D3H-Baboon, лигированного с фрагментом NcoI-NdeI длиной 864 пары оснований, выделенного из pUC18 - Pig Uricase, что привело к образованию pET-3d-PigKS. Полученная последовательность уриказы PigKS состоит из кодонов 1-288 уриказы свиньи, соединенных в одной рамке с кодонами 289-304 уриказы бабуина.
Субклонирование последовательности уриказы под контролем промотора osmB
Ген уриказы был субклонирован в экспрессионный вектор, содержащий промотор osmB (согласно указаниям в Патенте Соединенных Штатов №5795776, включенном сюда во всем объеме посредством ссылки). Этот вектор позволяет индуцировать экспрессию белка в ответ на осмотическое давление или истощения среды. Экспрессионная плазмида pMFOA-18 содержала промотор osmB, последовательность связывания рибосомой (rbs) и последовательность терминатора транскрипции (ter). Она несет также ген устойчивости к ампициллину (AmpR) и экспрессирует человеческую рекомбинантную ацетилхолинэстеразу (AChE).
Субклонирование D3H уриказы бабуина
Плазмида pMFOA-18 была расщеплена рестриктазами NcoI и BamHI, после чего был выделен большой фрагмент. Конструкция pET-3d-D3H-Baboon была расщеплена рестриктазами NcoI и BamHI, после чего был выделен фрагмент длиной 960 пар оснований, включающий ген D3H уриказы бабуина. Два фрагмента были лигированы для получения pMFOU18.
Экспрессионная плазмида pMFXT113 содержала промотор osmB, rbs (deo опреон E.coli), ter (E.coli TrypA), рекомбинантный полипептид ингибитор-фактор Xa (FxaI), а также она несет ген устойчивости к тетрациклину (TetR). Ген уриказы бабуина был встроен в эту плазмиду вместо гена устойчивости к антибиотику. Плазмида pMFOU18 была рестрицирована NcoI, достроена, затем рестрицирована с XhoI, после чего был выделен фрагмент длиной 1030 пар оснований. Плазмида pMFXT133 была рестрицирована NdeI, достроена, затем рестрицирована XhoI, после чего был выделен большой фрагмент. Два фрагмента были лигированы для создания вектора pURBA16, экспрессирующего уриказу бабуина.
Субклонирование химерной уриказы свиньи бабуина
Плазмида pUBRA16 была рестрицирована ApaI и AlwNI, после чего был выделен фрагмент длиной 2320 пар оснований. Плазмида pMFXT133 была рестрицирована NdeI, достроена, затем рестрицирована AlwNI, после чего был выделен фрагмент длиной 620 пар оснований. Конструкция pET-3d-PBC была рестрицирована XbaI, достроена, затем была рестрицирована ApaI, после чего был выделен фрагмент длиной 710 пар оснований. Три фрагмента были лигированы для создания pUR-PB, плазмиды, экспрессирующей уриказу PBC под контролем промотора osmB и rbs, так же как T7 rbs, полученного из вектора pET-3d.
T7 rbs вырезали дополнительно. Конструкцию pUR-PB была рестрицирована NcoI, достраивали, затем рестрицировали AlwNI, после чего был выделен фрагмент длиной 620 пар оснований. Два фрагмента лигировали для получения конструкции pDUR-PB, экспрессирующей PBC под контролем промотора osmB.
Создание конструкции pOUR-PB-ΔNC
В кДНК уриказа были введены несколько изменений, что привело к существенному увеличению стабильности рекомбинантного фермента. Была сконструирована плазмида pOUR-PBC-ΔNC, в которой были изъяты последовательности, кодирующие N-концевой пептид созревания, а также С-концевой трипептид, необходимый для локализации фермента в пероксисомах. Это было достигнуто при использовании ПЦР с последовательности PBC в плазмиде pDUR-PB и специфических олигонуклеотидов-праймеров, приведенных в Таблице 2:
Последовательности нуклеотидов, выделенные жирным шрифтом в Таблице 2, являются введенными в последовательности праймеров рестрикционными сайтами, строчными буквами показаны некодирующие участки. Сайт NdeI - в смысловом праймере, XbaI - в антисмысловом. Anti-sence праймер был также использован для элиминирования внутреннего сайта рестрикции NdeI посредством введения точечной мутации (подчеркнуто), которая не изменяла аминокислотную последовательность и, таким образом, облегчала субклонирование с помощью NdeI.
Полученный с помощью ПЦР-амплификации фрагмент длиной 900 пар оснований был разрезан с NdeI и XbaI и выделен. Полученный фрагмент затем был введен в deo-экспрессирующую плазмиду pDBAST-RAT-N, несущую промотор deo-P1P2 и полученный из E.coli rbs и постоянно экспрессирующую рекомбинантный предшественник инсулина человека. Плазмиду расщепляли NdeI и XbaI, после чего фрагмент длиной 4035 пар оснований был выделен и лигирован с ПЦР-продуктом, полученным с уриказы PBC. Полученная конструкция, pDUR-PB-ΔNC, была использована для трансформации клеток E.coli K-12 Sφ733 (F-cytR strA), которые имели высокий уровень экспрессии активной усеченной уриказы.
Последовательность дважды усеченной уриказы PBC-ΔNC также была экспрессирована под контролем промотора osmB. Плазмида pDUR-PB-ΔNC была расщеплена AlwNI-NdeI, после чего был выделен фрагмент длиной 3459 пар оснований. Вышеописанная плазмида pMFXT133 была расщеплена NdeI-AlwNI, после чего был выделен фрагмент длиной 660 пар оснований. Фрагменты были лигированы для получения pOUR-PB-ΔNC, которая была введена в клетки E.coli K-12 штамм W3110 F-, которые имели высокий уровень экспрессии активной усеченной уриказы.
Получение экспрессионной плазмиды для уриказы pOUR-P-ΔN-ks-1
Плазмида была сконструирована для увеличения активности и стабильности рекомбинантного фермента. Уриказа Pig-KS-ΔN была усечена только c N-конца (ΔN), где был удален N-концевой пептид созревания длиной 6 остатков, и содержала мутации S46T, R291K и T301S. В положении 46 в результате консервативной мутации, случившейся во время ПЦР и клонирования, вместо серина был треонин. В положении 291 аргинин был замещен лизином, и в положении 301 серин был введен вместо треонина. Обе были получены из последовательности уриказы бабуина. Модификации R291K и T301S были названы KS, как обсуждалось выше. Экстрализин обеспечил дополнительный возможный сайт конъюгации с ПЭГ.
Конструкция pOUR-P-ΔN-ks-1 (Фиг.1), плазмида pOUR-PB-ΔNC была расщеплена ApaI-XbaI, после чего был выделен фрагмент длиной 3873 пары оснований. Плазмида pET-3d-PKS (конструкция показана на Фиг.4) была расщеплена ApaI-SpeI, после чего был выделен фрагмент длиной 270 пар оснований. Расщепление с SpeI оставило «липкий конец» 5' CTAG, который был эффективно лигирован к фрагменту ДНК, полученному с помощью XbaI. Два фрагмента были лигированы для получения pOUR-P-ΔN-ks-1. После лигирования сайты узнавания SpeI и XbaI были утеряны (эти сайты указаны в скобках на Фиг.9). Конструкция pOUR-P-ΔN-ks-1 была введена в клетки E.coli K-12 штамм W3110 F-, прототрофный, ATCC #27325. Получившаяся уриказа Pig-KS-ΔN, экспрессируемая под контролем промотора osmB, имела болльшой выход рекомбинантного белка с превосходной активностью и стабильностью.
На Фиг.1 показана конструкция плазмиды pOUR-P-ΔN-ks-1. Цифрами рядом с сайтами рестрикции обозначено их положение относительно сайта рестрикции HaeII, принятого за 1; рестрикционные сайты, в скобках обозначены сайты рестрикции, исчезающие во время процедуры клонирования. Плазмида pOUR-P-ΔN-ks-1, кодирующая уриказу Pig-KS-ΔN, имеет длину 4143 пары оснований (п.о.) и содержит следующие элементы:
1. Фрагмент ДНК длиной 113 п.о., от находящийся между 1-м нуклеотидом и сайтом рестрикции NdeI (в положении 113), включающий промотор osmB и сайт связывания рибосомы (rbs).
2. Фрагмент ДНК длиной 932 п.о., находящийся между сайтом рестрикции NdeI (в положении 113) и объединением SpeI/XbaI (в положении 1045), включающий: участок длиной 900 п.о., кодирующий Pig-KS-ΔN (нуклеотидную последовательность усеченного с N-конца белка уриказы свиньи, в которой были заменены аминокислоты в положениях 291 и 301 на лизин и серин соответственно) и фланкирующая последовательность длиной 32 п.о., полученная из pCR™ II, из ТА-сайта клонирования перед SpeI/XbaI рестрикционным сайтом.
3. Последовательность сайта множественного клонирования (MCS) длиной 25 п.о. от SpeI/XbaI (в положении 1045) до HindIII (в положении 1070).
4. Синтетический олигонуклеотид длиной 40 п.о., содержащий TrpA терминатор транскрипции (ter) с концами: HindIII (в положении 1070) и AatII (в положении 1110).
5. Фрагмент ДНК длиной 1519 п.о., находящийся между сайтами AatII (в положении) и MscI/ScaI (в положении 2629) на pBR322, который включает ген устойчивости к тетрациклину (TetR).
6. Фрагмент ДНК длиной 1514 п.о., находящийся между сайтами ScaI (в положении 2629) и HaeII (в положении 4143) на pBR322, который включает ориджин ДНК репликации.
Фиг.2 изображает ДНК и получаемую аминокислотную последовательность уриказы Pig-KS-ΔN. В этой фигуре номера аминокислот соответствуют полной последовательности уриказы. Следующий за инициаторным метионином треонин был введен вместо аспартата в последовательность уриказы свиньи. Этот остаток треонина делает возможным удаление метионина с помощью бактериальных аминопептидаз. Брешь в аминокислотной последовательности указывает на удаленный N-концевой пептид созревания. Указаны рестрикционные сайты, которые были использованы для различных шагов субклонирования различных последовательностей уриказы (ApaI, NdeI, BamHI, EcoRI и SpeI). 3'-нетранслируемая последовательность, выделенная строчными буквами, была получена из последовательности pCR™II. Стоп-кодон трансляции отмечен звездочкой.
На Фиг.3 приведено выравнивание последовательностей аминокислот различных рекомбинантных уриказ. Верхняя строка представляет уриказу свиньи, включающую полную последовательность аминокислот. Вторая строка является последовательностью дважды усеченной химерной уриказы свиньи-бабуина (PBC-ΔNC). В третьей строке показана последовательность уриказы Pig-KS-ΔN, усеченной только с N-конца и содержащей мутации S46T и замены аминокислот R291K и T301S, обе отражающие происхождение С-конца кодирующей уриказу последовательности от уриказы бабуина. Звездочки отмечают положения, в которых есть различия в аминокислотах между Pig-KS-ΔN и опубликованной последовательностью уриказы свиньи; кружки указывают положения, в которых существуют различия между Pig-KS-ΔN и PBC-ΔN, химерной уриказой свиньи-бабуина; пунктирные линии отмечают делецию аминокислот.
кДНК нативных уриказ бабуина, свиньи и кролика с мутацией Y97H и химера уриказы свиньи/бабуина (PBC) были сконструированы для клонирования в E.coli. Клоны с высоким уровнем экспрессии вариантов уриказы были сконструированы и отобраны таким образом, что все являлись клонами W3110F- E.coli, и экспрессия регулировалась промотором osmB. Плазмидная ДНК была выделена, проверена ДНК-секвенированием и рестрикционным анализом, и клетки были культивированы.
Конструирование усеченных уриказ, включая pig-ΔN и Pig-KS-ΔN, было осуществлено с помощью перекрестного лигирования между PBC-ΔNC и Pig-KS с последующей рестрикцией эндонуклеазами ApaI и XbaI и ApaI плюс SpeI, соответственно. Логично, что усеченные мутанты сохранят активность, поскольку N-концевые 6 остатков, «пептид созревания» (1-2), и C-концевой трипептид, «сигнал локализации в пероксисомах» (3-5), не имеют функций, которые существенно влияют на энзиматическую активность, и возможно, что эти последовательности могут быть иммуногенными. Были отобраны клоны, экспрессирующие очень высокий уровень вариантов уриказы.
Пример 2. Трансформация экспрессионной плазмиды в бактериальную клетку-хозяина
Экспрессионная плазмида, pOUR-P-ΔN-ks-1, была введена в клетки E.coli K-12 штамм W3110 F-. Бактериальные клетки были приготовлены для трансформации, включая рост до середины лог-фазы в среде Лурье (LB), затем клетки подвергались центрифугированию, были промыты в холодной воде и суспендированы в 10% глицероле, в воде, в концентрации 3×1010 клеток на мл. Клетки хранили в аликвотах при -70°С. Плазмидную ДНК осаждали этанолом и растворяли в воде.
Бактериальные клетки и плазмидную ДНК смешивали и трансформацию проводили методом электропорации при высоком напряжении, используя Gene Pulser II от BIO-RAD (Trevors et al (1992). Electrotransformation of bacteria by plasmid DNA, in Guide to Electroporation and Electrofusion (D.C.Chang, B.M.Chassy, J.A. Saunders and A.E.Sowers, eds.), pp.265-290, Academic Press Inc., San Diego, Hanahan et al (1991) Meth. Enzymol., 204, 63-113). Трансформированные клетки были ресуспендированы в среде SOC (2% триптона, 0,5% дрожжевого экстракта, 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза), инкубировали при 37°С в течение 1 часа и отобраны по устойчивости к тетрациклину. Клоны с высоким уровнем экспрессии были отобраны.
Пример 3. Получение рекомбинантной уриказы
Бактерии, такие как эти трансформированные (см. выше), культивировали в среде, содержащей глюкозу; рН поддерживали на уровне 7,2±0,2, при примерно 37°С.
Перед последними 5-6 часами культивирования в среду был добавлен KCl до конечной концентрации 0,3 М. Культивирование было продолжено для накопления уриказы.
Рекомбинантная уриказа накапливалась в бактериальных клетках в качестве нерастворимого преципитата в тельцах включения (ТВ). Суспензию клеток промывали центрифугированием и ресуспендировали в 50 мМ Трис-буфере, рН 8,0 и 10 мМ ЭДТА, доводили объем до 40 раз по отношению к сухой массе клеток.
Содержащие рекомбинантную уриказу ТВ выделяли центрифугированием после разрушения бактериальных клеток с помощью лизоцима и высокого давления. Обработку лизоцимом (2000-3000 единиц/мл) проводили в течение 16-20 часов при рН 8,0 и 7±3°С при перемешивании. Осадок промывали водой и хранили при -20°С до использования.
Обогащенную фракцию ТВ далее обрабатывали 50 мМ NaHCO3-буфером рН 10,3±0,1. Суспензию инкубировали в течение ночи при комнатной температуре для растворения и последующей очистки центрифугированием уриказы, содержащейся в ТВ.
Дальнейшую очистку уриказы проводили хроматографией в несколько этапов. Вначале хроматографию проводили на Q-Sepharose FF колонке. Нагруженную колонку промывали бикарбонатным буфером, содержащим 150 мМ NaCl, после чего уриказу элюировали бикарбонатным буфером, содержащим 250 мМ NaCl. Затем xanthine-agarose смола (Sigma) была использована для удаления минорных примесей в препарате уриказы. Элюат с Q-Sepharose FF был разведен 50 мМ глициновым буфером, рН 10,3±0,1 до концентрации белка примерно 0,25 мг/мл и нанесен на колонку. Колонку дважды промыли бикарбонатным буфером, рН 10,3±0,1 содержащим 100 мМ NaCl, и элюировали уриказу тем же буфером, содержащим 60 мкМ ксантин. На этом этапе уриказу доочищали хроматографией на Q-Sepharose для удаления агрегированных форм.
Чистота каждого препарата уриказы составляет более 95%, что было определено гель-фильтрационной хроматографией. Менее чем 0,5% агрегированных форм было детектировано в каждом препарате с использованием колонки Sephadex 200.
В Таблице 3 обобщены данные по очистке уриказы Pig-KS-ΔN из ТВ, полученных из 25 л ферментативного бульона.
Очистка уриказы Pig-KS-ΔN
Пример 4. Характеристика рекомбинантной уриказы
СДС-ПААГ
СДС-ПААГ анализ тонко очищенных вариантов уриказы (Фиг.4) выявил различающиеся картины. Образцы хранили при 4°С в карбонатном буфере, рН 10,3, до нескольких месяцев. В образцах полноразмерных вариантов: Pig, Pig-KS и PBC было детектировано накопление продуктов деградации, имеющих молекулярную массу примерно 20 и 15 кДа. Данное наблюдение указывает на то, что как минимум один разрыв разделяет молекулу субъединицы уриказы. Другая картина деградации детектирована в клонах, укороченных с N-конца, а также для уриказы кролика. N-конец белка кролика имеет сходство с таковыми у укороченных клонов. Была определена аминокислотная последовательность фрагментов уриказы, генерированных во время очистки и хранения.
Секвенирование пептидов
N-концевое секвенирование всего препарата уриказы было проведено методом деградации по Эдману. Было осуществлено десять циклов. Рекомбинантная уриказа Pig (полноразмерный клон) производила большее количество фрагментов деградации, чем Pig-KS-ΔN. Определенные сайты расщепления, приводящего к образованию фрагментов деградации, следующие:
1) Мажорный сайт в положении 168, имеющий последовательность:
-QSG↓FEGFI-
2) Минорный сайт в положении 142, имеющий последовательность:
-IRN↓GPPVI-
Приведенные последовательности не соответствуют ни одному из известных механизмов протеолитического расщепления. Тем не менее, расщепление может быть результатом либо протеолитической, либо химической реакции. Неожиданно, что уриказы, усеченные с N-конца, более стабильны, чем уриказы, не усеченные с N-конца. PBC-ΔN также имела стабильность, сходную с другими ΔN-молекулами и меньшую, чем PBC без N-концевого усечения.
Эффективность
Активность уриказы была измерена УФ-методом. Скорость энзиматической реакции определяли, измеряя уменьшение абсорбции при 292 нм, происходящей из-за окисления мочевой кислоты до аллантоина. Одна единица активности определяется как количество уриказы, необходимое для окисления одного микромоля мочевой кислоты за минуту при 25°С, при определенных условиях. Эффективность уриказы обозначается в единицах активности на мг белка (Ед/мг).
Коэффициент экстинкции 1мМ мочевой кислоты при 292 нм равен 12,2 мМ-1 см-1. Таким образом, окисление 1 микромоля мочевой кислоты в 1 мл реакционной смеси приводит к уменьшению абсорбции, равному 12,2 мА292. Изменение абсорбции со временем (ΔА292 в минуту) было определено на линейном участке кривой.
Концентрацию белка определяли, используя модифицированный метод Брэдфорд (Macart and Gerbaut (1982) Clin Chim Acta 122:93-101). Удельную активность (эффективность) уриказы считали делением активности в Ед/мл на концентрацию белка мг/мл. Результаты по энзиматической активности для различных вариантов уриказы обобщены в Таблице 4. Результаты для коммерческих препаратов включены в эту таблицу в качестве референтных значений. Из этих результатов очевидно, что усечение белков уриказ не имеет существенного эффекта на их энзиматическую активность.
Краткая характеристика кинетических параметров рекомбинантной
и нативной уриказ
Замечания к Таблице 4:
(1) Концентрацию белка определяли по величине поглощения при 278 нм, используя коэффициент экстинкции, равный 11,3, для раствора уриказы с концентрацией 10 мг/мл (Mahler, 1963).
(2) 1 единица активности уриказы определяется как количество фермента, необходимого для окисления 1 мкмоль мочевой кислоты до аллантоина за минуту при 25°С.
(3) Значения удельной активности были получены из графика Лайнуивера-Барка, при концентрации субстрата эквивалентной 60 мкМ.
(4) Реакционные смеси состояли из различных комбинаций следующих маточных растворов:
100 мМ натрий-боратный буфер, рН 9,2;
300 мкМ мочевая кислота в 50 мМ натрий-боратном буфере, рН 9,2;
1 мг/мл БСА в 50 мМ натрий-боратном буфере, рН 9,2.
(5) Kcat рассчитывали делением Vmax (рассчитанным из соответствующего графика Лайнуивера-Барка) на концентрацию уриказы в реакционной смеси (выраженной в моль эквивалентах, основываясь на молекулярном весе для четвертичной структуры уриказы).
Пример 5. Конъюгация уриказы с м-ПЭГ (ПЭГ-конъюгация)
Уриказу Pig-KS-ΔN конъюгировали, используя м-ПЭГ-НФК (монометокси-поли(этиленгликоль)-нитрофенил карбонат). Были созданы условия, при которых образовывались 2-12 цепей 5, 10 или 20 кДа ПЭГ на единицу уриказы. м-ПЭГ-НФК постепенно добавляли к раствору белка. После добавления ПЭГ реакционную смесь уриказа/м-ПЭГ-НФК инкубировали при 2-8°С в течение 16-18 часов, пока максимальное количество несвязанных м-ПЭГ цепей не было конъюгировано с уриказой.
Количество цепей ПЭГ на мономер ПЭГ-уриказа определяли с помощью Superose 6 гель-фильтрационной хроматографией (SEC) с использованием ПЭГ и стандартов уриказы. Количество связанных ПЭГ-цепей определяли с помощью следующего уравнения:
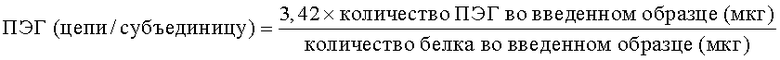
Концентрация компонентов ПЭГ и уриказы в пробе ПЭГ-уриказы была определена последовательно организованными size exclusion хроматографией (SEC), ультрафиолетом (УФ) и согласно индексу преломления (ИП) (разработано Kunitani, et al., 1991). Были сгенерированы три калибровочные кривые: кривая для белка (абсорбцию измеряли при 220 нм), кривая для белка (измеряли ИП) и кривая для ПЭГ (измеряли ИП). Затем образцы ПЭГ-уриказы анализировали, используя такую же систему. Получившиеся для экспериментальных образцов максимальные значения площади для УФ и ИП были использованы для расчета концентрации ПЭГ и белка относительно калибровочной кривой. Индекс 3,42 является отношением между молекулярной массой мономера уриказы (34192 Да) к таковой для 10 кДа ПЭГ.
Присоединенный ПЭГ увеличил растворимость уриказы в растворах, имеющих физиологические значения рН. В Таблице 5 продемонстрированы различия между препаратами ПЭГ-конъюгированной уриказы Pig-KS-ΔN. В общем, существует обратное соотношение между количеством присоединенных цепей ПЭГ и сохраняющейся удельной активностью (УА) фермента.
Ферментативная активность ПЭГ-конъюгированной уриказы Pig-KS-ΔN
Пример 6. ПЭГ-конъюгирование уриказы с 1000 Да и 100000 Да ПЭГ
Уриказу Pig-KS-ΔN конъюгировали, используя 1000 Да и 100000 Да м-ПЭГ-НФК, как было описано в Примере 5. Были созданы условия, при которых образовывались 2-12 цепей ПЭГ на единицу уриказы. После добавления ПЭГ реакционную смесь уриказа/м-ПЭГ-НФК инкубировали при 2-8°С в течение 16-18 часов, пока максимальное количество несвязанных м-ПЭГ цепей не было конъюгировано с уриказой.
Количество цепей ПЭГ на мономер ПЭГ-уриказа определяли как описано выше.
Присоединенный ПЭГ увеличил растворимость уриказы в растворах, имеющих физиологические значения рН.
Пример 7. Фармакокинетика уриказы Pig-KS-ΔN, конъюгированной с ПЭГ
Были проведены биологические эксперименты для определения оптимальной протяженности и размера ПЭГ-конъюгирования, необходимые для обеспечения терапевтической пользы.
Фармакокинетические исследования на крысах с использованием внутривенной инъекции немодифицированной уриказы 0,4 мг (2 ед) на кг массы тела, вводимые на 1 и 8 день, имели время полужизни в крови примерно 10 минут. Однако исследования на крысах скорости выведения уриказы 2-11×10 кДа ПЭГ-Pig-KS-ΔN после 9 недельных инъекций показали, что выведение не зависит от количества цепей ПЭГ (внутри данного разброса) и остается относительно стабильным в течение времени исследований (см. Таблицу 6; с временем полужизни примерно 30 часов). Различия между неделями не превышают экспериментальную ошибку. Такая же картина наблюдается после девяти инъекций конъюгатов 10×5 кДа ПЭГ и 10×20 кДа ПЭГ-уриказа. Результаты указывают на то, что вне зависимости от степени ПЭГ-конъюгирования уриказы, внутри данного разброса, сходные биологические эффекты наблюдаются при использовании крыс в качестве моделей.
Время полужизни препаратов ПЭГ-конъюгированной уриказы Pig-KS-ΔN в крысах
±1,7 (5)
±3,4 (5)
±3,1 (5)
±3,9 (5)
±4,3 (5)
±4,3 (5)
±1,5 (5)
±3,0 (5)
±1,6 (5)
±3,8 (5)
±2,6 (5)
±11,7 (5)
±11,1 (5)
±4,7 (5)
±3,3 (5)
±2,7 (5)
±5,3 (5)
±2,2 (4)
±5,6 (4)
±1,7 (5)
±2,7 (5)
±3,9 (4)
±3,0 (5)
±3,6 (5)
±1,3 (5)
±2,5 (5)
±3,1 (14)
±3,6 (13)
±2,9 (13)
±4,9 (5)
±4,3 (5)
±2,7 (5)
±2,5 (5)
±4,0 (15)
±2,7 (15)
±2,4 (13)
±3,4 (13)
±2,6 (13)
±1,4 (15)
±0,5 (15)
Замечания к Таблице 6: Результаты приведены в часах ± ошибка среднего. Цифры в скобках обозначают количество животных в эксперименте.
Еженедельно крысам внутривенно вводили препарат уриказы Pig-KS-ΔN, модифицированной так, как указано в таблице, из расчета 0,4 мг на кг массы тела. Исходно каждая группа содержала 15 крыс, которые поочередно были поделены на подгруппы по 5 животных. Несколько крыс умерло из-за наркоза в процессе эксперимента. Время полужизни определяли, измеряя активность уриказы (колориметрическим методом) в пробах плазмы крови, отбираемых через 5 минут и 6, 24 и 48 часов после инъекции.
В Таблице 5 приведены описания препаратов, использованных в этих исследованиях.
Исследования по биодоступности с препаратом уриказы 6×5 ПЭГ-Pig-KS-ΔN на кроликах продемонстрировали, что после первой инъекции время полужизни составляло 98,2±1,8 часов (внутривенно, вв) и биодоступность после внутримышечного (вм) и подкожного (пк) введений составляла 71% и 52%, соответственно. Однако у всех кроликов был определен высокий титр антител на уриказу после вторичного вм и пк введений, скорость выведения также увеличилась. При инъекции того же препарата крысам было детектировано время полужизни, равное 26±1,6 часов (вв), биодоступность после вм и пк введений составляла 33% и 22%, соответственно.
Исследования на крысах с препаратом уриказы 9×10 кДа ПЭГ-Pig-KS-ΔN продемонстрировали, что время полужизни после первой инъекции составляло 42,4 часа (внутривенно, вв) и биодоступность после внутримышечного (вм) и подкожного (пк) введений составляла 28,9% и 14,5%, соответственно (см. Фиг.5 и Таблицу 7). После четвертой инъекции время полужизни в крови составляло 32,1±2,4 часа и биодоступность после вм и пк введений составляла 26,1% и 14,9%, соответственно.
Сходные исследования фармакокинетики на кроликах с препаратом уриказы 9×10 кДа ПЭГ-Pig-KS-ΔN продемонстрировали, что после инъекций этого конъюгата не наблюдалось увеличенная скорость выведения (4 инъекции дважды в неделю). В этих животных время полужизни составляло 88,5 часов (вв), биодоступность после вм и пк инъекций составляла 98,3% и 84,4, соответственно (см. Фиг.6 и Таблицу 7). После четвертой инъекции время полужизни в крови составляло 141,1±15,4 часа, биодоступность после вм и пк введений составляла 85% и 83%, соответственно.
Сходные исследования по биодоступности 9×10 кДа ПЭГ-Pig-KS-ΔN были проведены на гончих собаках (2 самца и 2 самки в каждой группе). После первого введения зарегистрированное время полужизни в крови составляло 70±11,7 часов, биодоступность после вм и пк введений составляла 69,5% и 50,4%, соответственно (см. Фиг.7 и Таблицу 7).
Исследования препарата 9×10 кДа ПЭГ-Pig-KS-ΔN были проведены на свиньях. Три животных в каждой группе были использованы для вв, пк и вм введений. После первой инъекции время полужизни в крови составляло 178±24 часа, биодоступность после вм и пк введений составляла 71,6% и 76,8%, соответственно (см. Фиг.8 и Таблицу 7).
Исследования фармакокинетики препарата уриказы 9×10 кДа ПЭГ-Pig-KS-ΔN
Исследования абсорбции, распределения, метаболизма и выделения (ADME) были проведены после йодирования уриказы 9×10 кДа ПЭГ-Pig-KS-ΔN методом Болтона-Хантера с 125I. Меченный конъюгат вводили 7 группам животных, по 4 животных в каждой (2 самца и 2 самки). Распределение радиоактивности анализировали после 1 часа и каждые 24 часа в течение 7 дней (за исключением дня 5). Каждая группа, в свою очередь, была умерщвлена и различные органы вырезаны и проанализированы. Седьмая группа содержалась в метаболической камере, где собирали мочу и фекалии. Распределение материала в теле животного оценивали, измеряя общую радиоактивность каждого органа и доступную для осаждения с ТХУ (т.е. связанную с белком, нормализованную на размер органа) меченную фракцию (почки, печень, легкие и селезенка). Ни один из вырезанных органов не продемонстрировал более высокую специфическую радиоактивность по сравнению с другими, что свидетельствует об отсутствии существенного накопления, например, в печени или почках. К 7-му дню 70% радиоактивности было выведено из организма.
Пример 8. Клинические исследования
Для исследований реакцией на урат, фармакокинетического профиля и профиля безопасности ПЭГ-уриказы (Puricase®, Savient Pharmaceuticals) на пациентах с гиперурикемией и тяжелой подагрой, невосприимчивых или толерантных к традиционной терапии, были проведены рэндомизированные случайно распределенные исследования открытого типа в параллельных группах. Средняя продолжительность заболевания составляла 14 лет, 70 процентов подопытных в этих исследованиях имели один или более отложений.
В этих исследованиях 41 пациент (средний возраст 58,1 год) были случайно отобраны для 12-недельного лечения с помощью внутривенного введения ПЭГ-уриказы в одном из следующих режимов: 4 мг каждые две недели (7 пациентов), 8 мг каждые две недели (8 пациентов), 8 мг каждые четыре недели (13 пациентов) или 12 мг каждые четыре недели (13 пациентов). Активность уриказы в плазме крови и уровень мочевины измеряли с определенными интервалами. Параметры фармакокинетики, средняя концентрация мочевины в плазме и доля времени, когда концентрация была равной или ниже 6 мг/дл, определяли из анализа активности уриказы и уровня мочевины.
Пациенты, принимавшие 8 мг ПЭГ-уриказы каждые две недели, имели наиболее сильное уменьшение в PUA при уровне ниже 6 мг/мл в 92% времени лечения (мочевина в крови перед лечением - 9,1 мг/дл против среднего уровня мочевины в крови 1,4 мг/дл после 12 недель).
Существенное и постоянное уменьшение уровня мочевины в плазме крови также наблюдалось в других группах, принимающих ПЭГ-конъюгированную уриказу: PUA ниже 6 мг/мл 86% времени лечения группе 8 мг каждые четыре недели (перед лечением мочевина в крови 9,1 мг/дл против 2,6 мг/дл после 12 недель); PUA ниже 6 мг/мл 84% времени лечения группе 12 мг каждые четыре недели (перед лечением мочевина в крови 8,5 мг/дл против 2,6 мг/дл после 12 недель); PUA ниже 6 мг/мл 73% времени лечения группе 4 мг каждые две недели (перед лечением мочевина в крови 7,6 мг/дл против 4,2 мг/дл после 12 недель).
Максимальный процент снижения мочевой кислоты в плазме с контрольной линии в течение 24 часов после дозы ПЭГ-уриказы составил 72% для субъектов, принимавших 4 мг/2 недели (р равно 0,0002); 94% для субъектов, принимавших 8 мг/2 недели (р меньше 0,0001); 87% для субъектов, принимавших 8 мг/4 недели (р меньше 0,0001); 93% для субъектов, принимавших 12 мг/4 недели (р меньше 0,0001).
Процент снижения мочевой кислоты в плазме после 12 недельного лечения составил 38% для субъектов, принимавших 4 мг/2 недели (р равно 0,0002); 86% для субъектов, принимавших 8 мг/2 недели (р меньше 0,0001); 58% для субъектов, принимавших 8 мг/4 недели (р равно 0,0003); 67% для субъектов, принимавших 12 мг/4 недели (р меньше 0,0001).
Неожиданно, несколько субъектов, получавших ПЭГ-уриказу, имели неблагоприятные эффекты, связанные с лечением, т.е. реакцию на введение. Эти реакции наблюдались в 14% от общего числа введений.
Все ссылки, цитированные здесь, включены сюда в полном объеме посредством ссылок и для всех приложений настолько же, как если каждая индивидуальная публикация или патент или патентная заявка была специфически или индивидуально включена во всей полноте посредством ссылки для всех приложений.
Много модификаций и вариантов настоящего изобретения может быть сделано без уклонения от его смысла и содержания, что будет очевидно специалистам в технологии. Специфические варианты осуществления, описанные здесь, предлагаются только в качестве примеров, и изобретение ограничивается только терминами прилагаемых формул изобретения вместе с полным объемом эквивалентов, которым данные формулы озаглавлены.













Изобретение относится к области биохимии. Представлены варианты уриказы, содержащие последовательности, приведенные в описании. Также представлена выделенная нуклеиновая кислота, содержащая последовательность нуклеотидов, которая кодирует указанную уриказу, и указанная нуклеиновая кислота оперативно связанная с гетерологичным промотором. Описан экспрессирующий вектор, содержащий указанную нуклеиновую кислоту и клетка-хозяин, содержащая этот вектор и являющаяся продуцентом уриказы. Описаны способ получения уриказы, включающий стадии культивирования указанных клеток-хозяев при условиях, когда последовательность нуклеиновой кислоты экспрессируется клетками-хозяевами, и выделение уриказы. Изобретение позволяет ослабить гиперурикемию и гиперурикозурию. 12 н. и 1 з.п. ф-лы, 8 ил., 7 табл.
1. Выделенная уриказа, содержащая последовательность SEQ ID NO. 7.
2. Выделенная уриказа, содержащая последовательность SEQ ID NO. 8.
3. Выделенная уриказа, содержащая последовательность SEQ ID NO. 7 и N-концевую аминокислоту, где N-концевой аминокислотой является аланин, глицин, пролин, серии, треонин или метионин.
4. Выделенная уриказа, содержащая последовательность SEQ ID NO. 12.
5. Выделенная уриказа, содержащая последовательность аминокислот SEQ ID NO. 13.
6. Выделенная уриказа, содержащая последовательность SEQ ID NO. 12 и N-концевую аминокислоту, где N-концевая аминокислота является аланином, глицином, пролином, серином, треонином или метионином.
7. Конъюгат, содержащий уриказу по любому из пп.1-6 и полиэтиленгликоль (ПЭГ).
8. Выделенная нуклеиновая кислота, содержащая последовательность нуклеотидов, которая кодирует уриказу, выбранную из группы, состоящей из: уриказы по п.1, уриказы по п.2, уриказы по п.3, уриказы по п.4, уриказы по п.5, и уриказы по п.6.
9. Выделенная нуклеиновая кислота, содержащая последовательность нуклеотидов, которая кодирует уриказу, выбранную из группы, состоящей из: уриказы по п.1, уриказы по п.2, уриказы по п.3, уриказы по п.4, уриказы по п.5, и уриказы по п.6, где последовательность нуклеотидов оперативно связана с гетерологичным промотором.
10. Нуклеиновая кислота по п.9, где промотор является промотором osmB.
11. Нуклеиновокислотный экспрессирующий вектор, содержащий нуклеиновую кислоту по п.9.
12. Клетка-хозяин, содержащая вектор по п.11, являющаяся продуцентом уриказы.
13. Способ получения уриказы, включающий стадии культивирования клеток-хозяев по п.12 при условиях, когда последовательность нуклеиновой кислоты экспрессируется клетками-хозяевами, и выделение экспрессированной уриказы.
| US 6576235, 10.07.2003 | |||
| WO 00/08196 A2, 17.02.2000 | |||
| RU 2001103144 A, 20.10.2003 | |||
| RU 2002120486 A, 20.08.2004 | |||
| КРЕТОВИЧ В.Л | |||
| Введение в энзимологию | |||
| - М.: Наука, 1986, с.154-158. |

