ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Иммунная система включает в себя широкий диапазон определенных типов клеток, в том числе лимфоциты. Лимфоциты определяют специфичность иммунной реакции в организме и включают в себя два класса: B-лимфоциты, которые являются предшественниками антителопродуцирующих клеток, и T-лимфоциты, которые необходимы для определенных регуляторных функций, таких как развитие специфичного иммунного ответа.
Зрелые T-клетки могут быть активированы, в частности, антигеном или другими стимулами и могут продуцировать, например, цитокины, молекулы биохимической передачи сигнала или рецепторы, которые далее влияют на судьбу популяции T-клеток.
Активация B-клеток может происходить через рецепторы на их клеточной поверхности, включая B-клеточный рецептор, и другие вспомогательные молекулы для осуществления вспомогательных клеточных функций, таких как продукция цитокинов.
Активация моноцитов/макрофагов и T-клеток может происходить через рецепторы на их клеточной поверхности, и они играют центральную роль в иммунном ответе посредством представления антигена лимфоцитам, а также выступают в качестве вспомогательных клеток для лимфоцитов посредством секреции ряда цитокинов.
Естественные киллерные (NK) клетки имеют общую с T-клетками и B-клетками клетку-предшественник и участвуют в иммунологическом надзоре. NK-клетки, которые составляют вплоть до 15% лимфоцитов крови, не экспрессируют рецепторы для антигенов и, таким образом, не используют распознавание MHC как необходимый фактор для связывания клетки-мишени. NK-клетки вовлечены в распознавание и уничтожение определенных опухолевых клеток и инфицированных вирусом клеток. Полагают, что in vivo NK-клеткам необходима активация, однако было показано, что in vitro NK-клетки вызывают гибель некоторых типов опухолевых клеток баз активации.
Интерлейкины представляют собой семейство цитокинов, опосредующих иммунологический ответ, в том числе воспаление. Интерлейкины опосредуют разнообразные воспалительные патологии. Основную роль в иммунном ответе играют T-клетки, которые продуцируют множество цитокинов и осуществляют адаптивный иммунитет против антигенов. Цитокины, продуцируемые T-клетками, классифицируют как цитокины 1 типа и 2 типа (Kelso, A. Immun. Cell Biol. 76:300-317, 1998). Цитокины 1 типа включают в себя IL-2, IFN-γ, LT-α, и они вовлечены в воспалительный ответ, вирусный иммунитет, иммунитет против внутриклеточных паразитов и отторжение аллотрансплантата. Цитокины 2 типа включают в себя IL-4, IL-5, IL-6, IL-10 и IL-13, и они вовлечены в гуморальный ответ, иммунитет против гельминтов и аллергический ответ. Общие для 1 и 2 типа цитокины включают в себя IL-3, GM-CSF и TNF-α. Существуют некоторые данные, подтверждающие, что продуцирующие 1 тип и 2 тип популяции T-клеток преимущественно мигрируют в различные типы воспаленной ткани.
Кожа играет важную роль в иммунной системе и состоит из слоев. Эпидермис представляет собой поверхностный слой. Под эпидермисом расположена дерма, слой соединительной ткани. Под дермой расположена гиподерма, слой из большого количества жировой ткани. Циркулирующие T-лимфоциты мигрируют в кожу в норме и в условиях воспаления. Кожный лимфоцитарный антиген (CLA) считают "хоминг"-рецептором для T-клеток, тропных к коже. Santamaria-Babi, L., Eur. J. Dermatol. 14:13-18, 2004.
Известно, что при некоторых заболеваниях кожи выявляются высокие уровни CLA+ T-клеток, включая атопический дерматит, контактный дерматит, индуцируемые лекарственным средством аллергические реакции, тропные к коже вирусы и ассоциированный с вирусом зуд, витилиго, кожную T-клеточную лимфому, очаговую алопецию, красные угри, обыкновенные угри, узловатую почесуху и буллезный пемфигоид. Существует необходимость в лечении таких кожных опосредуемых T-клетками заболеваний.
Демонстрируемые in vivo виды активности семейства цитокинов иллюстрируют огромный клинический потенциал и необходимость других цитокинов, агонистов цитокинов и антагонистов цитокинов. IL-31 представляет собой недавно идентифицированный цитокин. При сверхэкспрессии у мышей IL-31 приводит к подобным дерматиту симптомам. В патологию кожных заболеваний у человека вовлечены как направленные в кожу T-клетки, так и эпидермальные кератиноциты. Настоящее изобретение удовлетворяет эти потребности, предоставляя антагонисты провоспалительного цитокина IL-31. Такие антагонисты согласно изобретению, которые могут блокировать, ингибировать, снижать, оказывать антагонистическое воздействие или нейтрализовать активность IL-31, включают в себя растворимые рецепторы IL-31RA и нейтрализующие антитела против IL-31. Кроме того, это изобретение относится к их применению при воспалительном заболевании, а также к связанным с ним композициям и способам.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед подробным изложением этого изобретения, для его понимания может быть целесообразным определение следующих терминов:
Здесь термин "антитела" включает в себя поликлональные антитела, очищенные аффинными способами поликлональные антитела, моноклональные антитела и антигенсвязывающие фрагменты, такие как протеолитические фрагменты F(ab')2 и Fab. Также определение включает в себя полученные способами генетической инженерии целые антитела или фрагменты, такие как химерные антитела, Fv-фрагменты, одноцепочечные антитела и т.п., а также синтетические антигенсвязывающие пептиды и полипептиды. Не являющиеся человеческими антитела могут быть гуманизированы посредством пересадки не являющихся человеческими CDR в каркасную область и константные участки человека, или посредством встраивания целых не являющихся человеческими вариабельных доменов (необязательно "маскируя" их подобной человеческой поверхностью посредством замены экспонированных остатков с получением в результате "гиперхимеризованного" антитела). В некоторых случаях в гуманизированных антителах могут быть сохранены не являющиеся человеческими остатки в каркасных доменах вариабельного участка человека для усиления необходимых свойств, касающихся связывания. С помощью гуманизации антител может быть повышено биологическое время полужизни, и при введении человеку возможность неблагоприятных иммунных реакций снижается.
Термин "химерное антитело" или "химерные антитела" относится к антителам, гены легкой и тяжелой цепей которых были сконструированы, главным образом, с помощью генетической инженерии, из генов вариабельного и константного участка иммуноглобулинов, принадлежащих другому виду. Например, вариабельные сегменты генов из моноклонального антитела мыши могут быть присоединены к константным сегментам человека, таким как гамма 1 и гамма 3. Таким образом, типичное терапевтическое химерное антитело представляет собой гибридный белок, состоящий из вариабельного или антигенсвязывающего домена из антитела мыши и константного домена из антитела человека, хотя могут использоваться другие виды млекопитающих.
Здесь термин "иммуноглобулин" относится к белку, состоящему из одного или нескольких полипептидов, главным образом, кодируемых генами иммуноглобулинов. Одна форма иммуноглобулина представляет собой основной структурный элемент антитела. Эта форма представляет собой тетрамер и состоит из двух идентичных пар цепей иммуноглобулинов, где каждая пара обладает одной легкой и одной тяжелой цепями. В каждой паре вариабельные участки легкой и тяжелой цепей совместно отвечают за связывание с антигеном, и константные участки отвечают за эффекторные функции антитела.
Полноразмерные "легкие цепи" иммуноглобулина (приблизительно 25 кДа или 214 аминокислот) кодируются геном вариабельного участка на NH2-конце (приблизительно 110 аминокислот) и геном константного участка каппа или лямбда на COOH-конце. Полноразмерные "тяжелые цепи" иммуноглобулина (приблизительно 50 кДа или 446 аминокислот), аналогично кодируются геном вариабельного участка (приблизительно 116 аминокислот) и одним из других вышеупомянутых генов константных участков (приблизительно 330 аминокислот). Тяжелые цепи классифицируют как гамма, мю, альфа, дельта или эпсилон, и они определяют изотип антитела IgG, IgM, IgA, IgD и IgE, соответственно. В легкой и тяжелой цепях вариабельные и константные участки соединены участком "J" приблизительно из 12 или более аминокислот, а тяжелая цепь также включает в себя участок "D" приблизительно из 10 или более аминокислот. (Смотрите, главным образом, Fundamental Immunology (Paul, W., ed., 2nd ed. Raven Press, N.Y., 1989), Ch. 7 (включенная в качестве ссылки в полном объеме для любых целей).
Вариабельный участок легкой или тяжелой цепи иммуноглобулина состоит из "каркасного" участка, прерывающегося тремя гипервариабельными участками. Таким образом, термин "гипервариабельный участок" относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельный участок содержит аминокислотные остатки из "определяющего комплементарность участка" или "CDR" (т.е. остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)) и/или эти остатки из "гипервариабельной петли" (т.е. остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (H1), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia and Lesk, 1987, J. Mol. Biol. 196: 901-917) (обе из которых включены в настоящее описание в качестве ссылок). Остатки "каркасной области" или "FR" представляют собой те остатки вариабельного домена, которые отличаются от остатков гипервариабельного участка, как определено в настоящем описании. Последовательности каркасных областей различных легких и тяжелых цепей являются относительно консервативными в пределах вида. Таким образом, "каркасная область человека" представляет собой каркасную область, которая является по существу идентичной (приблизительно на 85% или более, как правило, на 90-95% или более) каркасной области встречающегося в природе иммуноглобулина человека. Каркасная область антитела, которая представляет собой комбинированные каркасные области из составляющих ее легкой и тяжелой цепей, служит для локализации и выравнивания CDR. CDR, главным образом, отвечают за связывание с эпитопом антигена.
Таким образом, термин "гуманизированный" иммуноглобулин относится к иммуноглобулину, содержащему каркасную область человека и один или несколько CDR из иммуноглобулина, не являющегося человеческим (как правило, мыши или крысы). Не являющийся человеческим иммуноглобулин, предоставляющий CDR, называют "донором", а иммуноглобулин человека, предоставляющий каркасную область, называют "акцептором". Нет необходимости в наличии константных участков, однако в случае их наличия они должны быть по существу идентичными константным участкам иммуноглобулина человека, т.е. по меньшей мере приблизительно на 85-90%, предпочтительно приблизительно на 95% или более идентичными. Таким образом, все части гуманизированного иммуноглобулина, за исключением, возможно, CDR, являются по существу идентичными соответствующим частям природных последовательностей иммуноглобулина человека. "Гуманизированное антитело" представляет собой антитело, содержащее иммуноглобулин с гуманизированной легкой цепью и гуманизированной тяжелой цепью. Например, гуманизированное антитело не включает в себя типичное химерное антитело, как определено выше, например, вследствие того, что целый вариабельный участок химерного антитела является нечеловеческим.
Термин "генетически измененные антитела" означает антитела, где аминокислотная последовательность отличается от последовательности природного антитела. Вследствие значения способов рекомбинантных ДНК при получении антител, нет необходимости в ограничении последовательностями аминокислот, встречающимися в природных антителах; антитела можно переконструировать для получения требуемых характеристик. Возможные изменения многочисленны и варьируют от изменения только одной или нескольких аминокислот до полного переконструирования, например, вариабельного или константного участка. Изменения в константном участке, как правило, проводят в целях улучшения или изменения характеристик, таких как связывание комплемента, взаимодействие с мембранами и другие эффекторные функции. Изменения в вариабельном участке проводят в целях улучшения характеристик, касающихся связывания антигена.
В дополнение к антителам, иммуноглобулины могут существовать во множестве других форм, включая, например, одноцепочечные антитела или Fv, Fab и (Fab')2, а также димеры, линейные антитела, поливалентные или полиспецифичные гибридные антитела (как описано выше и подробнее в: Lanzavecchia et al., Eur. J. Immunol. 17, 105 (1987)) и в форме отдельных цепей (например, Huston et al., Proc. Natl. Acad. Sci. U.S.A., 85, 5879-5883 (1988), и Bird et al., Science, 242, 423-426 (1988), которые включены в настоящее описание в качестве ссылок). (Смотрите, главным образом, Hood et al., "Immunology", Benjamin, N.Y., 2nd ed. (1984), и Hunkapiller and Hood, Nature, 323, 15-16 (1986), которые включены в настоящее описание в качестве ссылок).
Здесь термины "одноцепочечные Fv", "одноцепочечные антитела", "Fv" или "scFv" относятся к фрагментам антител, которые содержат вариабельные участки как из тяжелой, так и из легкой цепей, однако лишены константных участков, но в пределах одной полипептидной цепи. Как правило, одноцепочечное антитело между доменами VH и VL дополнительно содержит полипептидный линкер, который позволяет им формировать требуемую структуру, которая обеспечивает возможность связывания антигена. Одноцепочечные антитела подробно описаны Pluckthun в The Pharmacology of Monoclonal Antobodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994); смотрите также публикацию международной патентной заявки № WO 88/01649 и патенты США №№ 4946778 и 5260203, описания которых включены в качестве ссылок для любой цели. В конкретных вариантах осуществления, одноцепочечные антитела также могут быть биспецифичными и/или гуманизированными.
"Fab-фрагмент" содержит одну легкую цепь и C.sub.H1 и вариабельные участки одной тяжелой цепи. Тяжелая цепь молекулы Fab не может образовывать дисульфидную связь с другой молекулой тяжелой цепи.
"Fab'-фрагмент" содержит одну легкую цепь и одну тяжелую цепь, которая между доменами CН1 и CH2 содержит большую часть константного участка, так что между двумя цепями может образоваться межцепочечная дисульфидная связь с образованием молекулы F(ab')2.
"F(ab')2-фрагмент" содержит две легкие цепи и две тяжелые цепи, содержащие часть константного участка между доменами CH1 и CH2, так что между двумя тяжелыми цепями образуется межцепочечная дисульфидная связь.
Понятно, что молекулярные массы и длины полимеров, определенные неточными аналитическими способами (например, гель-электрофорез), представляют собой приблизительные значения. Когда такое значение выражают как "приблизительно" X или "приближенно" X, следует понимать, что указанное значение X является точным на ±10%.
Все приведенные в настоящем описании ссылки включены в качестве ссылок в полном объеме.
В основе настоящего изобретение отчасти лежит открытие, связанное с применением антител в качестве антагонистов IL-31, ингибирующих, таким образом, воспаление в целом и симптомы дерматита и связанных с зудом заболеваний. Это изобретение относится к применению моноклональных антител для ингибирования, уменьшения, предотвращения или минимизации эффектов дерматита и связанных с зудом заболеваний, как описано далее в настоящем описании. В одном варианте осуществления дерматит представляет собой атопический дерматит. В другом варианте осуществления дерматит представляет собой узловатую почесуху. В другом варианте осуществления дерматит представляет собой экзему. IL-31 представляет собой недавно открытый T-клеточный цитокин, который при сверхэкспрессии у мышей приводит к подобным дерматиту симптомам. Смотрите также Dillon, et al., Nature Immunol. 5:752-760, 2004. В патологию кожных заболеваний у человека вовлечены как направленные в кожу T-клетки, так и эпидермальные кератиноциты. Экспрессия мРНК и белка IL-31 как у пациентов с атопическим дерматитом (AD), так и у здоровых индивидуумов ограничивается направленной в кожу популяцией CLA+ T-клеток, в то время как анализ рецептора для IL-31, IL-31RA, посредством иммуногистохимии (IHC) подтверждает несколько более высокие уровни экспрессии IL-31RA на кератиноцитах кожи в биоптатах кожи пациентов, страдающих острым и хроническим AD, по сравнению со здоровыми индивидуумами.
IL-31 также имеет название HUGO, которое было присвоено цитокину, который ранее был описан как Zcyto17rlig в опубликованной патентной заявке США (смотрите публикацию номер 20030224487, Sprecher, Cindy et al., 2003, включенную в качестве ссылки). Смотрите также Dillon, et al., Nature Immunol., выше. Гетеродимерный рецептор для IL-31 также описан в 20030224487 в качестве zcytor17 (название HUGO, IL-31RA), который образуют гетеродимер с бета-рецептором онкостатина М (OSMRbeta). IL-31 был выделен из библиотеки кДНК, полученной из активированных клеток периферической крови человека (hPBC), среди которых была проведена селекция на CD3. CD3 представляет собой клеточный поверхностный маркер клеток исключительно лимфоидного происхождения, в частности, T-клеток. Полинуклеотидная и полипептидная последовательности IL-31 человека представлены в SEQ ID NO:1 и 2, соответственно. Полинуклеотидная и полипептидная последовательности IL-31 мыши представлены в SEQ ID NO:10 и 11, соответственно. Как используют в настоящем описании, термин IL-31 означает IL-31, как используют в патентной публикации США номер 20030224487, как показано выше. Секреторная сигнальная последовательность IL-31 содержит аминокислотные остатки от 1 (Met) до 23 (Ala), и зрелый полипептид содержит аминокислотные остатки от 24 (Ser) до 164 (Thr) (как показано в SEQ ID NO:2). Дополнительный анализ с N-концевым секвенированием очищенного IL-31 из T-клеток 293 показал локализацию N-конца в положении 27 (Leu), как представлено в SEQ ID NO:2, и зрелый полипептид содержал аминокислотные остатки от 27 (Leu) до 164 (Thr) (как показано в SEQ ID NO:2).
Полипептидная последовательность IL-31RA (рецептор для IL-31) представлена в SEQ ID NO:5, и полипептидная последовательность бета-рецептора онкостатина М (OSMRbeta) представлена в SEQ ID NO:7.
Рецепторы IL-31RA и OSMRbeta принадлежат классу I подсемейства рецепторов цитокинов, который включает в себя, но не ограничивается ими, рецепторы для IL-2, IL-4, IL-7, Lif, IL-12, IL-15, EPO, TPO, GM-CSF и G-CSF (для обзора смотрите Cosman, "The Hematopoietin Receptor Superfamily" в Cytokine 5(2): 95-106, 1993). Субъединица IL-31RA полностью описана в совместной патентной заявке PCT № US01/20484 (публикация WIPO № WO 02/00721). Анализ распределения мРНК субъединицы IL-31RA в тканях выявил экспрессию в активированных T-клетках подтипов CD4+ и CD8+, CD14+ моноцитах, и более слабую экспрессию в CD19+ B-клетках. Более того, мРНК была представлена как в покоящихся, так и в активированных моноцитарных клеточных линиях THP-1 (ATCC № TIB-202), U937 (ATCC № CRL-1593.2) и HL60 (ATCC № CCL-240).
Ингибирование, нейтрализацию, блокирование передачи сигнала посредством антагонистов IL-31, описанных в настоящем описании, можно определить с помощью ряда способов анализа, известных специалисту в данной области. Например, анализы для определения снижения пролиферации включают в себя анализы восстановления красителя, такого как AlamarBlue™ (AccuMed International, Inc. Westlake, Ohio), бромид 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия (Mosman, J. Immunol. Meth. 65: 55-63, 1983); 3,(4,5-диметилтиазол-2-ил)-5-3-карбоксиметоксифенил-2H-тетразолий; гидроксид 2,3-бис(2-метокси-4-нитро-5-сульфофенил)-5-[(фениламино)карбонил]-2H-тетразолия; и хлорид цианодитолилтетразолия (которые являются коммерчески доступными от Polysciences, Inc., Warrington, PA); анализы митогенеза, такие как измерение встраивания 3H-тимидина; анализы вытеснения красителя с использованием, например, нафталинового черного или трипанового синего; захват красителя с использованием диацетилфлуоресцеина; и высвобождение хрома. Смотрите, главным образом, Freshney, Culture of Animal Cells: A Manual of Basic Technique, 3rd ed., Wiley-Liss, 1994, которая включена в настоящее описание в качестве ссылки. В дополнение к указанным выше смотрите опубликованную патентную публикацию США номер 20030224487 (Sprecher, Cindy et al., 2003) в качестве примера клеток BaF3, экспрессирующих IL-31RA и полноразмерный OSMRbeta.
Как правило, цитокины предположительно обладают структурой из четырех альфа-спиралей, где спирали A, C и D являются наиболее важными для взаимодействия лиганд-рецептор и наиболее высоко консервативными среди членов семейства. Указание на аминокислотную последовательность IL-31 человека, представленную в SEQ ID NO:2, выравнивание IL-31 человека, IL-3 человека и аминокислотные последовательности цитокинов человека предполагает, что спираль A в IL-31 определяется аминокислотными остатками 38-52; спираль B определяется аминокислотными остатками 83-98; спираль C определяется аминокислотными остатками 104-117; и спираль D определяется аминокислотными остатками 137-152, как представлено в SEQ ID NO:2. Структурный анализ позволяет предположить, что петля A/B является длинной, петля B/C является короткой и петля C/D является длинной. Эта структура петель приводит к организации спирали вверх-вверх-вниз-вниз. Исходя из структуры глобулы из 4 спиралей, остатки цистеина в IL-31, которые являются консервативными, соответствуют аминокислотным остаткам 72, 133, и 147 в SEQ ID NO:2; и 74, 137 и 151 в SEQ ID NO:11, как описано в настоящем описании. Соответствующее расположение цистеина является дополнительным подтверждением структуры глобулы из четырех спиралей. Также в IL-31 высоко консервативным является остаток Glu, как представлено в SEQ ID NO:2, в положении 43. Эти спирали IL-31 могут представлять собой специфичные мишени для ингибирования, снижения или нейтрализации посредством антител, описанных в настоящем описании, для блокирования эффектов передачи сигнала IL-31 через узнающий его рецептор.
Исходя из сравнения последовательностей IL-31 человека и мыши, обнаружили консервативные остатки в участках, предположительно кодирующих альфа-спирали C и D. Соответствующие полинуклеотиды, кодирующие полипептидные участки, домены, мотивы, остатки и последовательности IL-31 человека, описанные в настоящем описании, представлены в SEQ ID NO:1. Эти спирали IL-31 могут представлять собой специфичные мишени для ингибирования, снижения или нейтрализации с помощью антител, описанных в настоящем описании, для блокирования эффектов передачи сигнала IL-31 через узнающий его рецептор.
В то время как спираль D является относительно консервативной в IL-31 человека и мыши, спираль C является наиболее консервативной. Несмотря на то, что у обоих видов в этом участке преобладают кислые аминокислоты, их различия могут быть причиной видовой специфичности во взаимодействии между IL-31 и его рецептором, IL-31RA, включающим в себя мономерные, гетеродимерные или мультимерные рецепторы. Петля A/B и спираль B IL-31 являются мало консервативными, и спираль C с петлей C/D и спиралью D являются наиболее консервативными между видами; консервативность в этом участке позволяет предположить, что он является функционально значимым. D-спирали IL-31 человека и мыши также являются консервативными. Антагонисты рецептора IL-31RA можно сконструировать посредством мутаций в D-спирали IL-31. Они могут включать в себя укорочение белка от остатка Thr156 (SEQ ID NO:2), или сохранение остатков, которые обеспечивают связывание лиганда с рецептором, но уменьшают активность передачи сигнала.
Способы получения полинуклеотидов, кодирующих антитела, описанные в настоящем описании (включая ДНК и РНК), хорошо известны в данной области. Тотальную РНК можно получать с использованием экстракции с изотиоцианатом гуанидиния с последующим выделением центрифугированием в градиенте CsCl (Chirgwin et al., Biochemistry 18:52-94, 1979). Поли-(A)+ РНК получают из тотальной РНК с использованием способа Aviv and Leder (Proc. Natl. Acad. Sci. USA 69:1408-12, 1972). Комплементарную ДНК (кДНК) получают из поли(A)+ РНК с использованием известных способов. Альтернативно, можно выделять геномную ДНК. Полинуклеотиды, кодирующие антитела против IL-31, затем идентифицируют и выделяют, например, посредством гибридизации или ПЦР.
Полинуклеотидная последовательность мышиного ортолога IL-31 была идентифицирована и представлена в SEQ ID NO:3. Зрелая последовательность IL-31 мыши предположительно начинается с Met1; как представлено в SEQ ID NO:4, который соответствует Met1, как представлено в SEQ ID NO:2, в последовательности человека. Тканевой анализ выявил, что экспрессия IL-31 мыши обнаруживается в семенниках, головном мозге, клетках CD90+, клетках простаты, слюнных железах и коже. Дополнительный анализ с N-концевым секвенированием очищенного IL-31 из T-клеток 293 показал расположение N-конца в остатке 31 (Ala), как представлено в SEQ ID NO:4, и зрелый полипептид содержит аминокислотные остатки от 31 (Ala) до 163 (Cys) (как представлено в SEQ ID NO:4).
Можно получать профиль гидрофильности Хоппа/Вудса для белковой последовательности IL-31, как представлено в SEQ ID NO:2 (Hopp et al., Proc. Natl. Acad. Sci.78:3824-3828, 1981; Hopp, J. Immun. Meth. 88:1-18, 1986 и Triquier et al., Protein Engineering JU: 153-169, 1998). В основе профиля лежит перемещающееся окно из шести остатков. Углубленные остатки G, S и T и экспонированные остатки H, Y и W не учитывают. Например, в IL-31 человека, гидрофобные участки включают в себя аминокислотные остатки 54-59 в SEQ ID NO:2, аминокислотные остатки 129-134 в SEQ ID NO:2, аминокислотные остатки 53-58 в SEQ ID NO:2, аминокислотные остатки 35-40 в SEQ ID NO:2 и аминокислотные остатки 33-38 в SEQ ID NO:2. Например, в IL-31 мыши гидрофобные участки включают в себя аминокислотные остатки 34-39 в SEQ ID NO:11, аминокислотные остатки 46-51 в SEQ ID NO:11, аминокислотные остатки 131-136 в SEQ ID NO:11, аминокислотные остатки 158-163 в SEQ ID NO:11 и аминокислотные остатки 157-162 в SEQ ID NO:11.
Специалисты в данной области поймут, что гидрофильность или гидрофобность будут учтены при разработке модификаций в аминокислотной последовательности полипептида IL-31 так, чтобы не нарушать общий структурный и биологический профили. Особый интерес для замещения представляют собой гидрофобные остатки, выбранные из группы, состоящей из Val, Leu и Ile, или группы, состоящей из Met, Gly, Ser, Ala, Tyr и Trp. Например, остатки, допускающие замену, могут включать в себя Val, Leu и Ile или группу, состоящую из остатков Met, Gly, Ser, Ala, Tyr и Trp, как представлено в SEQ ID NO:2. Консервативные остатки цистеина в положениях в SEQ ID NO:2 и SEQ ID NO:11 являются относительно не допускающими замену.
Настоящее изобретение также относится к антителам, которые связывают функциональные фрагменты полипептидов IL-31, и к молекулам нуклеиновых кислот, кодирующим такие функциональные фрагменты. "Функциональный" IL-31 или его фрагмент, как определено в настоящем описании, характеризуется его активностью в отношении пролиферации или дифференцировки, его способностью индуцировать или ингибировать специализированные клеточные функции, или его способностью специфически связываться с антителом против IL-31 или с IL-31RA или с антителом или гетеродимерами IL-31RA/OSMRbeta этих рецепторов (либо растворимыми, либо иммобилизованными). Как описано в настоящем описании выше, IL-31 характеризуется структурой глобулы с четырьмя спиралями, содержащей спираль A (аминокислотные остатки 38-52), спираль B (аминокислотные остатки 83-98), спираль C (аминокислотные остатки 104-117) и спираль D (аминокислотные остатки 137-152), как представлено в SEQ ID NO:2. Таким образом, настоящее изобретение далее относится к слитым белкам, включающим в себя: (a) полипептидные молекулы, содержащие одну или несколько спиралей, как описано выше; и (b) функциональные фрагменты, содержащие одну или несколько из этих спиралей. Другая полипептидная часть слитого белка может быть представлена другим цитокином в виде глобулы с четырьмя спиралями, таким как IL-15, IL-2, IL-4 и GM-CSF, или неприродным и/или неродственным секреторным сигнальным пептидом, который облегчает секрецию слитого белка.
Настоящее изобретение также относится к антителам, которые связываются с полипептидными фрагментами или пептидами, содержащими часть полипептида IL-31 с эпитопом, описанного в настоящем описании. Такие фрагменты или пептиды могут содержать "иммуногенный эпитоп", который является частью белка, который вызывает антительный ответ, когда в качестве иммуногена используют целый белок. Иммуногенные обладающие эпитопом пептиды можно идентифицировать с использованием стандартных способов (смотрите, например, Geysen et al., Proc. Nat'l Acad. Sci. USA 81:3998 (1983)). Связывание антител с этими функциональными фрагментами приводит к ингибированию, блокированию, нейтрализации и/или снижению передачи сигнала IL-31 на узнающий его рецептор.
В противоположность этому полипептидные фрагменты или пептиды могут содержать "антигенный эпитоп", который представляет собой участок молекулы белка, с которой может специфически связываться антитело. Определенные эпитопы состоят из линейных или непрерывных участков из аминокислот, и антигенность такого эпитопа не нарушается денатурирующими веществами. В данной области известно, что для стимуляции продукции антител против белка можно использовать относительно короткие синтетические пептиды, которые могут имитировать эпитопы белка (смотрите, например, Sutcliffe et al., Science 219:660 (1983)). Таким образом, антигенные обладающие эпитопом пептиды и полипептиды согласно изобретению являются пригодными для индукции антител (например, нейтрализующих антител), которые связываются с полипептидами, описанными в настоящем описании. Для определения участков, которые обладают наибольшим антигенным потенциалом, можно использовать профили гидрофильности Хоппа/Вудса (Hopp et al., 1981, там же, и Hopp, 1986, там же). Например, в IL-31 человека, гидрофильные участки включают в себя аминокислотные остатки 54-59 в SEQ ID NO:2, аминокислотные остатки 129-134 в SEQ ID NO:2, аминокислотные остатки 53-58 в SEQ ID NO:2, аминокислотные остатки 35-40 в SEQ ID NO:2 и аминокислотные остатки 33-38 в SEQ ID NO:2. Например, в IL-31 мыши гидрофильные участки включают в себя аминокислотные остатки 34-39 в SEQ ID NO:11, аминокислотные остатки 46-51 в SEQ ID NO:11, аминокислотные остатки 131-136 в SEQ ID NO:11, аминокислотные остатки 158-163 в SEQ ID NO:11 и аминокислотные остатки 157-162 в SEQ ID NO:11.
Имеющие эпитоп антигенные пептиды и полипептиды предпочтительно содержат по меньшей мере от четырех до десяти аминокислот, по меньшей мере от десяти до четырнадцати аминокислот или приблизительно от четырнадцати до приблизительно тридцати аминокислот SEQ ID NO:2 или SEQ ID NO:4. Такие обладающие эпитопом пептиды и полипептиды можно получать посредством фрагментации полипептида IL-31 или химического синтеза пептидов, как описано в настоящем описании. Более того, отбор эпитопов можно проводить с помощью фагового дисплея библиотек случайных пептидов (смотрите, например, Lane and Stephen, Curr. Opin. Immunol. 5:268 (1993); и Cortese et al, Curr. Opin. Biotechnol. 7:616 (1996)). Стандартные способы идентификации эпитопов и получения антител на основе небольших пептидов, которые содержат эпитоп, описаны, например, Mole, "Epitope Mapping," в Methods in Molecular Biology, Vol. 10, Manson (ed.), страницы 105-116 (The Humana Press, Inc. 1992); Price, "Production and Characterization of Synthetic Peptide-Derived Antibodies", в Monoclonal Antibodies: Production, Engineering, and Clinical Application, Ritter и Ladyman (eds.), страницы 60-84 (Cambridge University Press 1995), и Coligan et al. (eds.), Current Protocols in Immunology, страницы 9.3.1-9.3.5 и страницы 9.4.1-9.4.11 (John Wiley & Sons 1997).
Активность антител, как описано в настоящем описании, можно определять по их способности ингибировать или снижать пролиферацию с использованием множества способов анализа, в которых определяют пролиферацию и/или связывание с клетками, экспрессирующими рецептор IL-31RA. Особый интерес представляют изменения в IL-31-зависимых клетках. Пригодные клеточные линии для создания IL-31-зависимых клеточных линий включают в себя IL-3-зависимую клеточную линию BaF3 (Palacios and Steinmetz, Cell 41: 727-734, 1985; Mathey-Prevot et al., Mol Cell. Biol. 6: 4133-4135, 1986), FDC-P1 (Hapel et al., Blood 64: 786-790, 1984), и MO7e (Kiss et al., Leukemia 7: 235-240, 1993). Зависимые от фактора роста клеточные линии можно получать в соответствии с опубликованными способами (например, Greenberger et al., Leukemia Res. 8: 363-375, 1984; Dexter et al., Baum et al. Eds., Experimental Hematology Today, 8th Ann. Mtg. Int. Soc. Exp. Hematol. 1979, 145-156, 1980).
Активность антител против IL-31, описанных в настоящем описании, можно определять с помощью биосенсорного микрофизиометра на основе силикона, который измеряет скорость внеклеточного закисления или экскрецию протонов, ассоциированную со связыванием рецептора и последующими физиологическими клеточными ответами. Иллюстративное устройство представляет собой микрофизиометр Cytosensor™, производимый Molecular Devices, Sunnyvale, CA. Этим способом можно определить множество клеточных ответов, таких как пролиферация клеток, транспорт ионов, выработка энергии, воспалительный ответ, активация регулятора и рецептора, и т.п. Смотрите, например, McConnell, H.M. et al., Science 257:1906- 1912, 1992; Pitchford, S. et al., Meth. Enzymol. 228:84-108, 1997; Arimilli, S. et al., L Immunol. Meth. 212:49-59, 1998; Van Liefde, I. et al., Eur. J. Pharmacol. 346:87-95, 1998.
Также в качестве реагентов для исследования с целью охарактеризации участков взаимодействия лиганд-рецептор пригодны антагонисты. Антагонисты пригодны для ингибирования размножения, пролиферации, активации и/или дифференцировки клеток, вовлеченных в регуляцию гемопоэза. Ингибиторы активности IL-31 (антагонисты IL-31) включают в себя антитела против IL-31 и растворимые рецепторы для IL-31, а также другие пептидные и непептидные вещества (включая рибозимы).
Ингибирование активности IL-31 можно определять с помощью ряда способов анализа. В дополнение к способам анализа, описанным в настоящем описании, образцы можно тестировать на ингибирование активности IL-31 с помощью множества анализов, разработанных для определения связывания рецептора, стимуляции/ингибирования IL-31-зависимых клеточных ответов или пролиферации экспрессирующих рецептор IL-31RA клеток.
IL-31-связывающий полипептид также можно использовать для очистки лиганда. Полипептид иммобилизуют на твердой подложке, такой как гранулы из агарозы, поперечносшитой агарозы, стекла, целлюлозных смол, смол на основе диоксида кремния, полистирола, поперечносшитого полиакриламида или сходных материалов, которые являются стабильными в условиях применения. Способы связывания полипептидов с твердыми подложками известны в данной области и включают в себя химические группы аминов, активацию цианоген-бромидом, активацию N-гидроксисукцинимидом, активацию эпоксидом, сульфгидрильную активацию и активацию гидразидом. Полученному материалу, как правило, придают форму колонки, и жидкости, содержащие лиганд, пропускают через колонку один или несколько раз для обеспечения связывания лиганда с рецепторным полипептидом. Затем лиганд элюируют с использованием изменения концентрации соли, хаотропных веществ (гуанидин-HCl), или pH для нарушения связывания лиганд-рецептор.
Преимущественно может быть использована система анализа, в которой используют связывающий лиганд рецептор (или антитело, один член пары комплемент/антикомплемент) или его связывающий фрагмент, и коммерчески доступное биосенсорное устройство (BIAcore, Pharmacia Biosensor, Piscataway, NJ). Такой рецептор, антитело, член пары комплемент/антикомплемент или фрагмент иммобилизуют на поверхности рецепторного чипа. Применение этого устройства описано в Karlsson, J. Immunol. Methods 145:229-40, 1991 и Cunningham and Wells, J. Mol. Biol. 234:554-63, 1993. Рецептор, антитело, член или фрагмент ковалентно присоединяют, с использованием химической группы амина или сульфгидрила, к декстрановым волокнам, которые прикрепляют к пленке из золота на проточной ячейке. Тестируемый образец пропускают через ячейку. В случае наличия лиганда, эпитопа или противоположного члена пары комплемент/антикомплемент в образце, он будет связываться с иммобилизованным рецептором, антителом или членом, соответственно, вызывая изменения коэффициента рефракции среды, который определяют по изменению поверхностного плазмонного резонанса пленки из золота. Эта система дает возможность определения скоростей ассоциации и диссоциации, из которых можно вычислять аффинность связывания, и оценки стехиометрии связывания. Альтернативно связывание лиганд/рецептор можно анализировать с использованием технологии SELDI™ (Ciphergen, Inc., Palo Alto, CA).
Антитела против IL-31 можно использовать для блокирования биологического действия провоспалительного IL-31, и они являются пригодными в качестве противовоспалительных лекарственных средств от множества заболеваний, как описано в настоящем описании. Специалист в данной области поймет, что антигенные обладающие эпитопом полипептиды содержат последовательность по меньшей мере от 6, предпочтительно по меньшей мере от 9 и более предпочтительно по меньшей мере от 15 до приблизительно 30 последовательных аминокислотных остатков полипептида IL-31 (например, SEQ ID NO:2). Также в их число входят полипептиды, содержащие фрагмент полипептида IL-31 большего размера, т.е. от 30-100 остатков вплоть до полноразмерной аминокислотной последовательности. Антигены или иммуногенные эпитопы также могут включать в себя присоединенные к ним метки, адъюванты, вещества для доставки и носители, как описано в настоящем описании. Пригодные антигены включают в себя полипептид IL-31, кодируемый SEQ ID NO:2 от аминокислоты номер 24 до аминокислоты номер 164, или его непрерывный фрагмент из от 9 до 141 аминокислоты. Другие пригодные антигены включают в себя полноразмерный и зрелый IL-31, спирали A-D и отдельные или составные спирали A, B, C и D структуры глобулы из четырех спиралей IL-31, как описано в настоящем описании. Предпочтительные для применения в качестве антигенов пептиды представляют собой гидрофильные пептиды, такие как пептиды, предсказанные специалистом в данной области, исходя из графика гидрофобности, как описано в настоящем описании, например, аминокислотные остатки 114-119, 101-105, 126-131, 113-118 и 158-162 в SEQ ID NO:2; и аминокислотные остатки 34-39, 46-51, 131-136, 158-163 и 157-162 в SEQ ID NO:11. Более того, в качестве предпочтительных антигенов выступают антигенные эпитопы IL-31, предсказанные с помощью графика Джеймсона-Вольфа, например, с использованием программы DNASTAR Protean (DNASTAR, Inc., Madison, WI), и их легко определит специалист в данной области.
Антитела вследствие иммунного ответа, индуцированного посредством инокуляции животному этих антигенов, можно выделять и очищать, как описано в настоящем описании. Способы получения и выделения поликлональных и моноклональных антител хорошо известны в данной области. Смотрите, например, Current Protocols in Immunology, Cooligan, et al. (eds.), National Institutes of Health, John Wiley and Sons, Inc., 1995; Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, NY, 1989; и Hurrell, J. G. R., Ed., Monoclonal Hybridoma Antibodies: Techniques and Applications. CRC Press, Inc., Boca Raton, FL, 1982.
Как будет очевидно среднему специалисту в данной области, поликлональные антитела можно получать посредством инокуляции разнообразным теплокровным животным, таким как лошади, коровы, козы, овцы, собаки, куры, кролики, мыши и крысы, полипептида IL-31 или его фрагмента. Иммуногенность полипептида IL-31 можно повышать посредством применения адъюванта, такого как квасцы (гидроксид алюминия) или полный и неполный адъювант Фрейнда. Полипептиды, пригодные для иммунизации, также включают в себя слитые полипептиды, такие как слитый полипептид из IL-31 или его части с полипептидом иммуноглобулина или со связывающим мальтозу белком. Полипептидный иммуноген может представлять собой полноразмерную молекулу или ее часть. Если полипептидная часть является "подобной гаптену", то для иммунизации такая часть преимущественно может быть соединена или связана с макромолекулярным носителем (таким как гемоцианин лимфы улитки (KLH), бычий сывороточный альбумин (BSA) или столбнячный токсоид).
Антитела считают специфически связывающимися, если: 1) они обладают пороговым уровнем связывающей активности, и 2) они не реагируют перекрестно в значительной степени со сходными полипептидными молекулами. Пороговый уровень связывания определяют, если антитела против IL-31, описанные в настоящем описании, связываются с полипептидом, пептидом или эпитопом IL-31 с аффинностью, по меньшей мере в 10 раз превышающей аффинность связывания с контрольным (не IL-31) полипептидом. Предпочтительно, чтобы антитела обладали аффинностью связывания (Ka) 106 M-1 или более, предпочтительно 107 M-1 или более, более предпочтительно 108 M-1 или более и наиболее предпочтительно 109 M-1 или более. Аффинность связывания антитела может легко определить средний специалист в данной области, например, посредством анализа Скэтчарда (Scatchard, G., Ann. NY Acad. Sci. 51: 660-672, 1949).
Отсутствие значительного перекрестного реагирования антител против IL-31 с родственными молекулами показывают посредством выявления с помощью антитела полипептида IL-31, но не родственных ему полипептидов, с использованием стандартного вестерн-блот анализа (Ausubel et al., там же). Примеры известных родственных полипептидов представляют собой полипептиды, описанные в предшествующем уровне техники, такие как ортологи и паралоги, и сходные известные члены белкового семейства. Также можно проводить скрининг с использованием не являющегося человеческим IL-31 и мутантных полипептидов IL-31. Более того, для антител можно проводить "скрининг против" известных сходных полипептидов, для выделения группы, которая специфически связывается с полипептидами IL-31. Например, антитела, индуцированные против IL-31, адсорбируют на родственные полипептиды, присоединенные к нерастворимой матрице; антитела, специфичные к IL-31, проходят через матрицу при надлежащих условиях буфера. Скрининг дает возможность выделения поликлональных и моноклональных антител, не реагирующих перекрестно с известными близко родственными полипептидами (Antibodies: A Laboratory Manual, Harlow and Lane (eds.), Cold Spring Harbor Laboratory Press, 1988; Current Protocols in Immunology, Cooligan, et al. (eds.), National Institutes of Health, John Wiley and Sons, Inc., 1995). Скрининг и выделение специфичных антител хорошо известны в данной области. Смотрите Fundamental Immunology, Paul (eds.), Raven Press, 1993; Getzoff et al., Adv. in Immunol. 43: 1-98, 1988; Monoclonal Antibodies: Principles and Practice, Goding, J.W. (eds.), Academic Press Ltd., 1996; Benjamin et al., Ann. Rev. Immunol. 2: 67-101, 1984. Специфичное связывание антител против IL-31 можно выявлять рядом способов данной области и способов, описанных ниже.
Моноклональные антитела можно получать, например, посредством иммунизации крыс Sprague-Dawley (Charles River Laboratories, Wilmington, MA) очищенным зрелым рекомбинантным полипептидом IL-31 человека (аминокислотные остатки от 27 (Leu) до 167 (Thr) в SEQ ID NO:2) или ортологом мыши, полученным из экспрессирующих систем, описанных в настоящем описании. Крысам проводят внутрибрюшинную (IP) инъекцию 100 мкг очищенного рекомбинантного белка IL-31 человека в полном адъюванте Фрейнда (Pierce, Rockford, IL) с последующими вторичными IP инъекциями 50 мкг очищенного рекомбинантного белка в неполном адъюванте Фрейнда каждые две недели. От семи до десяти суток после введения третьей вторичной инъекции, у животных проводят забор крови и собирают сыворотку.
Охарактеризацию образцов сыворотки крыс, специфичной к IL-31 человека, проводят посредством ELISA на титр для специфичного нацеливания антитела на биотинилированный IL-31 человека.
От 2 крыс с высоким титром собирают спленоциты и клетки лимфатических узлов и проводят слияние с клетками миеломы SP2/0 (мыши) с использованием PEG 1500 в двух отдельных процессах слияния (соотношение для слияния спленоцитов и клеток миеломы 4:1, "Antibodies A Laboratory Manual, E. Harlow and D.Lane, Cold Spring Harbor Press). После 10 суток выращивания после слияния, специфичные антителопродуцирующие гибридомные пулы идентифицируют посредством ELISA. Гибридомные пулы, положительные в обоих протоколах ELISA, далее анализируют на их способность блокировать или снижать активность очищенного рекомбинантного IL-31 в отношении связывания рецептора ("анализ нейтрализации").
Гибридомные пулы, для которых получены положительные результаты посредством только ELISA или посредством ELISA и "анализа нейтрализации", клонируют по меньшей мере два раза посредством лимитирующего разведения.
Моноклональные антитела, очищенные из среды культуры ткани, охарактеризовывают в отношении их пригодности в ELISA для количественного определения рекомбинантного и природного IL-31 человека. Антитела отбирают и проводят количественный анализ.
Моноклональные антитела, очищенные из среды культуры ткани, охарактеризовывают по их способности блокировать или снижать активность в отношении связывания ("анализ нейтрализации") очищенного рекомбинантного huIL-31 на клетках BaF3/MPL-IL-31. Таким способом идентифицируют ряд "нейтрализующих" моноклональных антител. Затем гибридомы, экспрессирующие описанные нейтрализующие моноклональные антитела против IL-31 человека, можно депонировать в патентную коллекцию Американской коллекции типовых тканевых культур (ATCC; Manassas VA) в качестве первоначальных депонируемых образцов согласно Будапештскому договору.
Моноклональные антитела можно получать посредством иммунизации крыс Lewis (Rockland Immunochemicals, Gilbertsville, PA), расщепленным и очищенным рекомбинантным слитым белком IL-31. Каждой крысе проводят внутрибрюшинную (IP) инъекцию 100 мкг очищенного рекомбинантного слитого белка в полном адъюванте Фрейнда (Pierce, Rockford, IL) с последующими вторичными IP инъекциями 50 мкг очищенного рекомбинантного белка в неполном адъюванте Фрейнда каждые две недели в течение четырех недель. После первых четырех недель иммунизации проводят вспомогательные вторичные внутрибрюшинные инъекции 50 мкг расщепленного очищенного рекомбинантного белка, присоединенного к белку-носителю, представляющему собой гемоцианин лимфы улитки (KLH, Pierce, Rockford, IL), в неполном адъюванте Фрейнда каждые две недели в течение четырех недель. От семи до десяти суток после введения четвертой вторичной инъекции, у животных отбирают кровь и собирают сыворотку. Охарактеризацию образцов сыворотки крысы, специфичной в отношении IL-31, проводят посредством ELISA с использованием очищенного рекомбинантного слитого белка IL-31-Fc в качестве специфичной мишени для антитела и неродственного слитого белка в качестве неспецифичной мишени для антитела в концентрации 500 нг/мл. Спленоциты отбирают от одной или нескольких крыс с высокими титрами и проводят слияние с клетками миеломы SP2/0 (мыши) в оптимизированном опосредуемом PEG протоколе слияния (Rockland Immunochemicals). После 12 суток выращивания после слияния, специфичные антителопродуцирующие гибридомные пулы идентифицируют посредством ELISA с использованием очищенного рекомбинантного слитого белка IL-31 в качестве специфичной мишени для антитела и неродственного слитого белка в качестве неспецифичной мишени для антитела, где концентрация каждого из них составляет 500 нг/мл. Гибридомные пулы, положительные только в отношении специфичной мишени для антитела, далее анализируют на их способность блокировать или снижать активность в отношении связывания рецептора ("анализ нейтрализации") рекомбинантного huIL-31 на клетках BaF3/MPL-IL-31 и на способность к связыванию в анализе FACS в качестве мишени для антитела. Гибридомные пулы, для которых получают положительные результаты в анализе ELISA и положительные результаты либо в FACS, либо в "анализе нейтрализации", клонируют по меньшей мере два раза посредством лимитирующего разведения.
Аналогичным образом получили пять гибридом крысы с противомышиными антителами и им присвоили следующие обозначения клонов: клон 271.9.4.2.6, клон 271.26.6.6.1, клон 271.33.1.2.2, клон 271.33.3.2.1 и клон 271.39.4.6.5. Смотрите пример 1. Моноклональные антитела, продуцируемые этими клонами, охарактеризовывали рядом способов, включая сортировку (т.е. определение способности каждого антитела ингибировать связывание при каком-либо другом связывании), относительную аффинность и нейтрализацию. Оказалось, что моноклональные антитела распределились на две отдельные серии, где клон 271.33.3.2.1 связывается эпитопом, отличающимся от эпитопов других четырех клонов. Относительная аффинность связывания для четырех из этих моноклональных антител описана в примере 14.
Можно достигать аффинности связывания моноклональных антител. Специфичное к IgG-Fc-гамма крысы антитело козы (Jackson) иммобилизуют на чипе CM5 Biacore. Анализ оптимизируют для связывания каждого mAb с поверхностью для улавливания с антителами против антител крысы, а затем серии концентраций IL-31 пропускают через mAb для определения ассоциации (Ka) и диссоциации (Kd). После каждого анализа поверхность регенерируют до антитела против антител крысы посредством 2 инъекций 20 мМ HCl. Данные получают для каждого образца и для оценки кинетики связывания антитела против IL-31 с белком IL-31 используют оценочное программное обеспечение (BIAevaluation software version 3,2, Pharmacia BIAcore, Uppsala, Sweden).
mAb мыши против IL-31 человека можно получать следующим образом. Мышей с нокаутом IL-31 в возрасте от шести до двенадцати недель иммунизируют посредством внутрибрюшинной инъекции 25-50 мкг растворимого белка IL-31 человека-muFc, смешанного 1:1 (об.:об.) с адъювантом Ribi (Sigma) по двухнедельной схеме. От семи до десяти суток после третьей иммунизации, посредством ретроорбитального забора крови получают образцы крови, сыворотку собирают и оценивают ее на способность ингибировать связывание IL-31 в анализах нейтрализации (например, описанных в настоящем описании) и окрашивать трансфицированные IL-31 клетки 293 относительно нетрансфицированных клеток в анализе окрашивания FACS. Иммунизацию мышей продолжают и образцы крови отбирают и оценивают, как описано выше, до достижения нейтрализующими титрами плато. Затем мышам с наиболее высокими нейтрализующими титрами проводят внутрисосудистую инъекцию 25-50 мкг растворимого белка IL-31-Fc в PBS. Через три дня селезенку и лимфатические узлы из этих мышей собирают и используют для получения гибридомы, например, с использованием клеток миеломы мыши (P3-X63-Ag8.653.3.12.11) или других пригодных для данной области клеточных линий, с использованием стандартных способов, известных в данной области (например, смотрите Kearney, J.F. et al., J Immunol. 123:1548-50, 1979; и Lane, R.D. J Immunol Methods 81:223-8, 1985).
На 8-10 сутки после слияния в супернатантах гибридом можно проводить первичный скрининг на их способность связывать белок IL-31-muFc посредством ELISA с использованием вторичных стоп-реагентов в виде конъюгированных с HRP антител козы против легкой цепи каппа и лямбда мыши для идентификации связавшихся антител мыши.
Биохимическое подтверждение того, что молекула-мишень, IL-31, распознанная предполагаемыми mAb против IL-31, действительно представляет собой IL-31, проводят стандартной иммунопреципитацией с последующим анализом SDS-PAGE или способами вестерн-блоттинга, в обоих из которых используют растворимые препараты мембраны из трансфицированных посредством IL-31 клеток Baf3 относительно нетрансфицированных клеток. mAb тестируют на их способность к специфичной иммунопреципитации или к вестерн-блоттингу с растворимым белком IL-31-muFc.
Моноклональные антитела, очищенные из среды культуры ткани, охарактеризовывали на их способность блокировать или ингибировать способность IL-31 связываться с его рецептором в анализе нейтрализации. Таким способом было идентифицировано двадцать "нейтрализующих" моноклональных антител. Десять из них было идентифицировано как "хорошие нейтрализаторы" после первого цикла клонирования: клон 292.72.3, клон 292.118.6, клон 292.63.5, клон 292.64.6, клон 292.84.1, клон 292.109.4, клон 292.12.3, клон 292.51.5, клон 292.39.5 и клон 292.105.4. Другие десять получили следующие обозначения клонов после первого цикла клонирования: клон 294.35.2, клон 294.146.5, клон 292.152.4, клон 292.154.4, клон 294.154.5, клон 294.35.3, клон 291.78.4, клон 294.155.6, клон 294.158.5, клон 294.163.2 и клон 294.144.3.
Для указанных моноклональных антител проводили второй цикл клонирования и снова охарактеризовывали на их способность блокировать или ингибировать способность IL-31 связываться с его рецептором в анализе нейтрализации. Обозначения клона после второго цикла клонирования для "хороших нейтрализаторов" представляли собой: клон 292.12.3.1, клон 292.63.5.3, клон 292.72.3.1, клон 292.84.1.6, клон 292.118.6.4, клон 292.64.6.5.5, клон 292.39.5.3, клон 292.51.5.2, клон 292.109.4.4 и клон 292.105.4.1. Шесть из других десяти обладают следующими обозначениями клонов: клон 292.152.4.1, клон 294.158.5.2, клон 294.32.2.6.3, клон 294.144.3.5, клон 294.154.5.3 и клон 294.163.2.1.
Гибридомы, экспрессирующие нейтрализующие моноклональные антитела против IL-31 человека, описанные выше, были депонированы в патентной коллекции Американской коллекции типовых тканевых культур (ATCC; Manassas VA) в качестве первоначальных депонированных образцов согласно Будапештскому договору и им были присвоены следующие регистрационные номера ATCC No: клон 292.12.3.1 (Патентный номер депозита в ATCC PTA-6815); клон 292.72.3.1 (Патентный номер депозита в ATCC PTA-6816); клон 292.63.5.3 (Патентный номер депозита в ATCC PTA-6829); клон 292.118.6.4 (Патентный номер депозита в ATCC PTA-6830); клон 294.163.2.1 (Патентный номер депозита в ATCC PTA-6831); клон 292.84.1.6 (Патентный номер депозита в ATCC PTA-6871); клон 294.35.2.6.3 (Патентный номер депозита в ATCC PTA-6872); клон 294.154.5.6 (Патентный номер депозита в ATCC PTA-6875); и клон 294.144.3.5 (Патентный номер депозита в ATCC PTA-6873).
Гибридома, экспрессирующая нейтрализующие моноклональные антитела против IL-31 мыши, описанные в настоящем описании, была депонирована в патентной коллекции Американской коллекции типовых тканевых культур (ATCC; Manassas VA) в качестве первоначального образца согласно Будапештскому договору, и ей был присвоен регистрационный номер ATCC No: клон 271.26.6.6.1 (Патентный номер депозита в ATCC PTA-6874).
Моноклональные антитела, продуцируемые этими клонами гибридом, можно культивировать в среде для выращивания из 90% среды Дульбекко, модифицированной по способу Исков, с 2 мМ L-глутамином, пенициллином в концентрации 100 мкг/мл и сульфатом стрептомицина в концентрации 100 мкг/мл, и 10% эмбриональной сыворотки клона I (Hyclone Laboratories). Размножение клонов можно проводить, начиная культивирование с 2·105 клеток/мл с поддержанием между 1·105 и 5·105 клеток/мл при 37°C и 5-6% CO. При последующих переносах клетки можно адаптировать к условиям без сыворотки. Замороженные клетки хранят в 90% сыворотке, 10% DMSO и их хранят в газообразной фазе в устройстве для замораживания с жидким азотом.
Моноклональные антитела в среде культуры ткани охарактеризовывают по их способности блокировать, ингибировать, предотвращать или снижать связывание рецептора при выращивании в присутствии очищенных рекомбинантных белков IL-31 человека. Например, моноклональные антитела, продуцируемые этими клонами, были охарактеризованы рядом способов, включая сортировку (т.е. определение способность каждого антител ингибировать связывание при каком-либо другом связывании), относительную аффинность и нейтрализацию. Оказалось, что десять хороших нейтрализующих антител относятся к одной серии, а другие моноклональные антитела распределяются на три отдельные серии. Кроме того, восемь из хороших нейтрализующих антител представляют собой изотип IgG1 и другие два представляют собой изотип IgG2a.
Моноклональные антитела из супернатантов гибридом извлекали с использованием pAb козы против Fc мыши и определяли кажущуюся аффинность связывания (EC50) биотинилированного IL-31. В этих условиях анализа, хорошие нейтрализаторы обладают наименьшими и сравнимыми значениями EC50 ~4 нг/мл Bt-IL31. Кажущаяся аффинность слабых нейтрализаторов находится в диапазоне от ~10 нг/мл до 236 нг/мл Bt-IL31.
Моноклональные антитела, полученные описанными в настоящем описании способами, можно тестировать на нейтрализацию множеством способов. Например, можно использовать анализ люциферазы, как описано в опубликованной патентной заявке США (Смотрите публикацию номер 20030224487, Sprecher, Cindy et al., 2003). Кроме того, нейтрализацию можно тестировать посредством измерения снижения продукции провоспалительных хемокинов, таких как TARC и MDC, культурами кератиноцитов в присутствии лиганда и моноклонального антитела. Нейтрализацию также можно определять посредством моделей in vivo, как описано в настоящем описании.
В одном варианте осуществления антитела согласно изобретению представляют собой антигенсвязывающие фрагменты антител человека согласно изобретению и включают в себя, но не ограничиваются ими, Fab, Fab' и F(ab')2, Fd, одноцепочечные Fv (scFv), одноцепочечные антитела, связанные дисульфидными связями Fv (sdFv), и фрагменты, содержащие либо домен VL, либо домен VH. Антигенсвязывающие фрагменты антитела, включающие в себя одноцепочечные антитела, могут содержать вариабельный участок(и) отдельно или в сочетании с всеми или частью из: шарнирной области, доменов CH1, CН2 и CН3. Также это изобретение относится к антигенсвязывающим фрагментам, содержащим также любое сочетание из вариабельного участка(ов) с шарнирной областью, доменами CH1, CH2 и CH3.
В другом варианте осуществления антитела согласно изобретению могут быть моноспецифичными, биспецифичными, триспецифичными или более полиспецифичными. Полиспецифичные антитела могут быть специфичными к различным эпитопам полипептида согласно изобретению или могут быть специфичными как к полипептиду согласно изобретению, так и к гетерологичному эпитопу, такому как гетерологичный полипептид или материал твердой подложки. Смотрите, например, публикации PCT WO 93/17715; WO 92/08802; WO 91/00360; WO 92/05793; Tutt, et al., J. Immunol. 147:60-69 (1991); патенты США №№ 4474893; 4714681; 4925648; 5573920; 5601819; Kostelny et al., J. Immunol. 148:1547-1553 (1992).
Настоящее изобретение также относится к генетически измененным антителам, которые функционально эквивалентны описанным выше антителам. Предпочтительными являются модифицированные антитела, обеспечивающие повышенную стабильность и/или терапевтическую эффективность. Примеры модифицированных антител включают в себя антитела с консервативными заменами аминокислотных остатков, и одну или несколько делеций или вставок аминокислот, которые не приводят к значительному отрицательному изменению свойства в отношении связывания антигена. Замены могут варьировать от изменения или модификации одного или нескольких аминокислотных остатков до полного переконструирования участка, при условии сохранения пригодности в терапевтических целях. Антитела согласно изобретению могут быть модифицированы посттрансляционно (например, ацетилирование и фосфорилирование), или они могут быть модифицированы синтетически (например, присоединение метки).
Генетически измененные антитела также включают в себя химерные антитела, полученные из антител против IL-31. Химерные антитела содержат вариабельный участок мыши или крысы и константный участок человека, так что химерное антитело обладает более продолжительным временем полужизни и является менее иммуногенным при введении человеку. Способ получения химерных антител известен в данной области. Вариабельные участки этих антител можно связывать с константным участком IgG человека с получением требуемого химерного антитела.
Генетически измененные антитела против IL-31, используемые согласно изобретению, включают в себя гуманизированный вариант антител, описанных в настоящем описании. Гуманизированные антитела содержат CDR донорного иммуноглобулина мыши и каркасные области тяжелой цепи и легкой цепи акцепторного иммуноглобулина человека. Способ получения гуманизированного антитела описан в патентах США №№ 5301101; 5585089; 5693762 и 6180370 (все из которых включены в настоящее описание в качестве ссылок в полном объеме). Затем CDR этих антител можно пересаживать в любые выбранные каркасные области человека, которые известны в данной области, с получением требуемого гуманизированного антитела.
Антитела согласно изобретению могут быть описаны или охарактеризованы с точки зрения эпитопа(ов) или части(ей) полипептида согласно изобретению, который они распознают или специфически связывают. Эпитоп(ы) или полипептидная часть(и) могут быть охарактеризованы, как описано в настоящем описании, например, по N-концевому и C-концевому положению, по размеру в смежных аминокислотных остатках, или представлены в таблицах и последовательностях. Антитела, которые специфически связывают любой эпитоп или полипептид согласно изобретению, могут быть также исключены. Таким образом, настоящее изобретение относится к антителам, которые специфически связывают полипептиды согласно изобретению, и предусматривает их исключение.
Это изобретение также относится к антителам, которые конкурентно ингибируют связывание моноклонального антитела с полипептидом согласно изобретению, предпочтительно полипептидом SEQ ID NO:2 или SEQ ID NO:4. Конкурентное ингибирование можно определять любым способом, известным в данной области, например, с использованием конкурентных анализов связывания, описанных в настоящем описании. В предпочтительных вариантах осуществления антитело конкурентно ингибирует связывание моноклонального антитела согласно изобретению по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60% или по меньшей мере на 50% с полипептидом SEQ ID NO:2 или SEQ ID NO:4.
Это изобретение также относится к антителам, которые конкурентно ингибируют связывание антитела с эпитопом согласно изобретению, как определяют любым способом, известным в данной области, для определения конкурентного связывания, например, способами иммунологического анализа, описанными в настоящем описании. В предпочтительных вариантах осуществления антитело конкурентно ингибирует связывание с эпитопом по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60% или по меньшей мере на 50%.
Антитела согласно изобретению включают в себя производные, которые являются модифицированными, а именно, посредством ковалентного присоединения к антителу молекулы любого типа, так чтобы ковалентное присоединение не препятствовало формированию антителом антиидиотипического ответа. В качестве неограничивающего примера, производные антитела включают в себя антитела, модифицированные, например, посредством гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидации, образования производных с помощью известных защитных/блокирующих групп, протеолитического расщепления, связывания с клеточным лигандом или другим белком, и т.д. Любую из множества химических модификаций можно проводить известными способами, включая, но не ограничиваясь ими, специфичное химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д. Кроме того, производное может содержать одну или несколько неклассических аминокислот.
Антитела согласно изобретению также включают в себя антитела или их фрагменты, которые обладают временем полужизни (например, временем полужизни в сыворотке) у млекопитающего, предпочтительно у человека, более 15 суток, предпочтительно более 20 суток, более 25 суток, более 30 суток, более 35 суток, более 40 суток, более 45 суток, более 2 месяцев, более 3 месяцев, более 4 месяцев или более 5 месяцев. Повышенное время полужизни антител согласно изобретению или их фрагментов у млекопитающего, предпочтительно у человека, приводит к более высокому сывороточному титру указанных антител или фрагментов антител у млекопитающего и, таким образом, к снижению частоты введения указанных антител или фрагментов антител и/или к снижению концентрации указанных антител или фрагментов антител, подлежащих введению.
Антитела или их фрагменты, обладающие повышенным временем полужизни in vivo, можно получать способами, известными специалистам в данной области. Например, антитела или их фрагменты с повышенным временем полужизни in vivo можно получать модификацией (например, заменой, делецией или вставкой) аминокислотных остатков, которые идентифицированы, как вовлеченные во взаимодействие между Fc-доменом и FcRn-рецептором (смотрите, например, международные публикации №№ WO 97/34631 и WO 02/060919, которые включены в настоящее описание в качестве ссылок в полном объеме). Антитела или их фрагменты с повышенным временем полужизни in vivo можно получать посредством присоединения к указанным антителам или фрагментам антител молекулы полимера, такого как высокомолекулярный полиэтиленгликоль (PEG). PEG можно присоединять к антителам или фрагментам антител при помощи многофункционального линкера, или без него, либо посредством сайт-специфичной конъюгации PEG с N- или C-концом указанных антител или фрагментов антител, либо через эпсилон-аминогруппы, находящиеся на остатках лизина. Используют получение производных с линейным или разветвленным полимером, которые вызывают минимальную потерю биологической активности. Для того чтобы убедиться в надлежащей конъюгации молекул PEG, с антителами проводят тщательный мониторинг уровня конъюгации посредством SDS-PAGE и масс-спектрометрии. Не вступивший в реакцию PEG можно отделять от конъюгатов антитело-PEG, например, посредством гель-фильтрации или ионообменной хроматографии.
Понятно, что гуманизированные антитела, сконструированные посредством данного способа, могут обладать дополнительными консервативными аминокислотными заменами, которые по существу не оказывают эффекта на связывание антигена или другие функции иммуноглобулина. Под консервативными заменами подразумевают сочетания, такие как gly, ala; val, ile, leu; asp, glu; asn, gln; ser, thr; lys, arg; и phe, tyr.
Способы гуманизации не являющихся человеческими антител хорошо известны в данной области. Как правило, гуманизированные иммуноглобулины, включая гуманизированные антитела, конструируют способами генетической инженерии. Большинство гуманизированных иммуноглобулинов, которые описаны ранее (Jones et al., ук. соч.; Verhoeyen et al., ук. соч.; Riechmann et al., ук. соч.), содержат каркасную область, которая является идентичной каркасной области конкретной цепи иммуноглобулина человека, акцептор, и три CDR из не являющейся человеческой донорной цепи иммуноглобулина. Конкретно, гуманизированные антитела представляют собой молекулы антител из антител вида, не относящегося к человеку, которые связывают требуемый антиген, обладающие одним или несколькими определяющими комплементарность участками (CDR) вида, не относящегося к человеку, и каркасными областями из молекулы иммуноглобулина человека.
Настоящее изобретение относится к критериям, по которым выбирают ограниченное число аминокислот в каркасной области цепи гуманизированного иммуноглобулина, чтобы они представляли собой те же аминокислоты в тех же положениях, что и в доноре, а не в акцепторе, в целях повышения аффинности антитела, содержащего гуманизированную цепь иммуноглобулина.
Антитела согласно изобретению получают отчасти на основе модели, которая в предшествующих способах продукции антитела (с использованием, например, антител мыши в качестве источника CDR) приводит к двум причинам потери аффинности:
(1) При объединении CDR мыши с каркасной областью человека происходит замена мышиных аминокислот в каркасной области, расположенных близко к CDR, человеческими аминокислотами. Без связи с какой-либо теорией, эти измененные аминокислоты могут немного деформировать CDR вследствие того, что они создают электростатические или гидрофобные силы, отличающиеся от донорного антитела мыши, и деформированные CDR могут не обеспечивать такие же эффективные контакты с антигеном, какие эти CDR обеспечивали в донорном антителе.
(2) Аминокислоты в исходном антителе мыши, которые расположены близко к CDR, но не являются их частью (т.е. все еще являются частью каркасной области), могут обеспечивать контакты с антигеном, которые могут способствовать аффинности. Эти аминокислоты утрачиваются при гуманизации антитела, поскольку все аминокислоты каркасной области являются человеческими.
Для избежания этих проблем и для получения гуманизированных антител, которые обладают очень высокой аффинностью к требуемому антигену, применяют один или несколько из следующих принципов согласно изобретению для конструирования гуманизированных иммуноглобулинов. Кроме того, критерии можно использовать по отдельности, или, при необходимости, в сочетании, для достижения требуемой аффинности или других характеристик.
Принцип состоит в том, что в качестве акцептора используют каркасную область из конкретного иммуноглобулина человека, который является высоко гомологичным донорному иммуноглобулину, подлежащему гуманизации, или используют консенсусную каркасную область из множества антител человека. Например, сравнение последовательности вариабельного участка тяжелой (или легкой) цепи мыши и вариабельных участков тяжелой (или легкой) цепи человека в базе данных (например, Ресурс идентификации белков национального биомедецинского исследовательского учреждения) показывает, что степень его гомологии с различными участками человека значительно варьирует, как правило, от приблизительно 40% до приблизительно 60-70%. Выбирая в качестве акцепторного иммуноглобулина один из вариабельных участков тяжелых (соответственно легких) цепей человека, который является наиболее гомологичным вариабельному участку тяжелой (соответственно легкой) цепи донорного иммуноглобулина, заменяют меньшее количество аминокислот при преобразовании донорного иммуноглобулина в гуманизированный иммуноглобулин. Таким образом, и также без связи с какой-либо теорией, полагают, что существует небольшая вероятность замены аминокислоты вблизи CDR, которая изменяет их конформацию. Более того, точная общая конфигурация гуманизированного антитела, содержащего гуманизированную цепь иммуноглобулина, может обладать более близким сходством с конфигурацией донорного антитела, что также снижает вероятность деформации CDR.
Как правило, для получения каркасной области тяжелой цепи, и аналогично для легкой цепи, в качестве акцептора выбирают одну из 3-5 наиболее гомологичных последовательностей вариабельных участков тяжелой цепи в репрезентативной коллекции по меньшей мере приблизительно из от 10 до 20 различных тяжелых цепей человека. Предпочтительно, используют один из 1-3 наиболее гомологичных вариабельных участков. Выбранная акцепторная цепь иммуноглобулина, наиболее предпочтительно, обладает по меньшей мере приблизительно 65% гомологией каркасной области с донорным иммуноглобулином.
Во многих случаях считают предпочтительным применение легких и тяжелых цепей из того же антитела человека, что и акцепторные последовательности, чтобы убедиться в приемлемом контактировании гуманизированных легких и тяжелых цепей друг с другом. В этом случае донорные легкие и тяжелые цепи сравнивают только с цепями из антител человека с известной полной последовательностью, например, с антителами Eu, Lay, Pom, Wol, Sie, Gal, Ou и WEA антитела (Kabat et al., ук. соч.; иногда последние несколько аминокислот цепи человека неизвестны, и их необходимо устанавливать по гомологии с другими антителами человека). Выбирают антитело человека, в котором последовательности вариабельных участков легкой и тяжелой цепей, взятые вместе, являются, в целом, наиболее гомологичными донорным последовательностям вариабельного участка легкой и тяжелой цепей. Иногда большее значение придают последовательности тяжелой цепи. Затем выбранное антитело человека предоставляет акцепторные последовательности как легкой, так и тяжелой цепи. На практике часто обнаруживают, что эту роль выполняет антитело человека Eu.
В соответствии с так называемым способом "наилучшего соответствия" проводят скрининг последовательности вариабельного домена антитела грызуна против целой библиотеки известных последовательностей вариабельных доменов человека. Последовательность человека, которая является наиболее сходной с последовательностью грызуна, затем принимают в качестве каркасной области (FR) человека для гуманизированного антитела (Sims et al., J. Immunol., 151: 2296 (1993); Chothia et al., J. Mol. Biol., 196: 901 (1987)). В другом способе используют конкретную каркасную область, образованную консенсусной последовательностью всех антител человека конкретной подгруппы легкой или тяжелой цепи. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci, USA, 89: 4285 (1992); Presta et al., J. Immnol., 151: 2623 (1993)).
Независимо от того, каким образом выбирают акцепторный иммуноглобулин, более высокой активности можно достигать, выбирая небольшое количество аминокислот в каркасной области гуманизированной цепи иммуноглобулина, которые являются такими же аминокислотами в тех же положениях в доноре, а не в акцепторе. Часто остатки каркасной области в каркасных областях человека заменяют соответствующим остатком из CDR донорного антитела для изменения, предпочтительно улучшения, связывания антитела. Эти замены в каркасной области идентифицируют способами, хорошо известными в данной области, например, посредством моделирования взаимодействия остатков CDR и каркасной области для выявления остатков каркасной области, важных для связывания антигена, и сравнения последовательностей для идентификации редких остатков каркасной области в конкретных положениях. (Смотрите, например, Queen et al., патент США № 5585089; Riechmann et al., Nature 332:323 (1988), которые включены в настоящее описание в качестве ссылок в полном объеме). Антитела можно гуманизировать с использованием различных способов, известных в данной области, включая, например, пересадку CDR (EP 239400; публикация PCT WO 91/09967; патенты США №№ 5225539; 5530101 и 5585089), гиперхимеризацию или изменение поверхности (EP 592106; EP 519596; Padlan, Molecular Immunology 28(4/5):489-498 (1991); Studnicka et al., Protein Engineering 7(6):805-814 (1994); Roguska. et al., PNAS 91:969-973 (1994)), и перестановку цепей (патент США № 5565332). Таким образом, такие "гуманизированные" антитела представляют собой химерные антитела (патент США № 4816567), где по существу менее чем целый вариабельный домен человека заменен соответствующей последовательностью не относящегося к человеку вида.
Второй принцип состоит в том, что указанные ниже категории определяют, какие аминокислоты можно выбирать из донора. Предпочтительно, во многих или во всех аминокислотных положениях в одной из этих категорий, в действительности выбирают донорную аминокислоту.
Категория 1: Аминокислотное положение находится в CDR, как определяют посредством Kabat et al., ук. соч.
Категория 2: Если аминокислота в каркасной области акцепторного иммуноглобулина человека является необычной (т.е. "редкой", что, как используют в настоящем описании, указывает на аминокислоту, встречающуюся в этом положении менее чем приблизительно в 20%, однако, как правило, менее чем приблизительно в 10% последовательностей V-участка тяжелой (соответственно легкой) цепи в репрезентативной базе данных), и если донорная аминокислота в этом положении является типичной для последовательностей человека (т.е. "общей", что, как используют в настоящем описании, указывает на аминокислоту, встречающуюся более чем приблизительно в 25%, однако, как правило, более чем приблизительно в 50% последовательностей в репрезентативной базе данных), тогда можно выбирать аминокислоту донора, а не акцептора. Этот критерий помогает обеспечить то, чтобы нетипичная аминокислота в каркасной области человека не нарушала структуры антитела. Более того, заменяя редкую аминокислоту аминокислотой из донорного антитела, которая является типичной для антител человека, можно получать менее иммуногенное гуманизированное антитело. Все последовательности вариабельных участков легкой и тяжелой цепей человека соответственно группируют в "подгруппы" последовательностей, которые являются особенно гомологичными друг другу и обладают одинаковыми аминокислотами в определенных критических положениях (Kabat et al., ук. соч.). При определении того, что аминокислота в акцепторной последовательности человека является "редкой" или "общей" среди последовательностей человека, часто является предпочтительным рассмотрение только тех последовательностей человека, которые находятся в той же подгруппе, что и акцепторная последовательность.
Категория 3: В положениях, расположенных непосредственно рядом с одним или несколькими из 3 CDR в первичной последовательности цепи гуманизированного иммуноглобулина, можно выбирать аминокислоту(ы) донора, а не акцептора. Особенно вероятно, что эти аминокислоты взаимодействуют с аминокислотами в CDR и, если выбраны из акцептора, деформируют донорные CDR и снижают аффинность. Более того, соседние аминокислоты могут взаимодействовать непосредственно с антигеном (Amit et al., Science, 233, 747-753 (1986), которая включена в настоящее описание в качестве ссылки) и может быть желательным выбор этих аминокислот из донора для сохранения всех контактов с антигеном, которые обеспечивают аффинность исходного антитела.
Категория 4: 3-мерная модель, главным образом, исходного донорного антитела показывает, что определенные аминокислоты вне CDR расположены близко к CDR и обладают высокой вероятностью взаимодействия с аминокислотами в CDR посредством образования водородных связей, ван-дер-ваальсовых сил, гидрофобных взаимодействий и т.д. В этих аминокислотных положениях можно выбирать аминокислоту донорного иммуноглобулина, а не акцепторного иммуноглобулина. Аминокислоты в соответствии с этим критерием, как правило, обладают атомом боковой цепи в пределах приблизительно 3 ангстрем от некоторого атома в CDR, и они должны содержать атом, который может взаимодействовать с атомами CDR в соответствии с широко известными химическими силами, такими как перечисленные выше.
В случае атомов, которые могут образовывать водородную связь, величина, равная 3 ангстремам, определяется между их ядрами, однако для атомов, которые не образуют связь, величина, равная 3 ангстремам, определяется между их ван-дер-ваальсовыми поверхностями. Таким образом, в последнем случае для атомов, которые следует считать способными к взаимодействию, ядра должны находиться в пределах приблизительно 6 ангстрем (3+сумма ван-дер-ваальсовых радиусов). Во многих случаях ядра расположены на расстоянии от 4 или 5 до 6 ангстрем. При определении возможности взаимодействия аминокислоты с CDR, предпочтительно не рассматривать последние 8 аминокислот CDR 2 тяжелой цепи, как часть CDR, поскольку, с точки зрения структуры, эти 8 аминокислот обладают свойствами части каркасной области.
Аминокислоты в каркасной области, которые способны взаимодействовать с аминокислотами в CDR и которые, таким образом, относятся к категории 4, можно выявить другим способом. Вычисляют доступную для растворителя площадь поверхности каждой аминокислоты каркасной области двумя способами: (1) в целом антителе и (2) в гипотетической молекуле, состоящей из антитела с удаленными CDR. Значительное различие между этими значениями, составляющее приблизительно 10 кв. ангстрем или более, показывает, что доступ растворителя к аминокислоте каркасной области по меньшей мере частично блокирован CDR, и, таким образом, что аминокислота контактирует с CDR. Доступную для растворителя площадь поверхности аминокислоты можно вычислять на основе 3-мерной модели антитела с использованием алгоритмов, известных в данной области (например, Connolly, J. Appl. Cryst. 16, 548 (1983) и Lee and Richards, J. Mol. Biol. 55, 379 (1971), оба из которых включены в настоящее описание в качестве ссылок). Аминокислоты каркасной области также иногда могут взаимодействовать с CDR непрямым образом, посредством влияния на конформацию других аминокислот каркасной области, которые в свою очередь контактируют с CDR.
Известно, что аминокислоты в некоторых положениях в каркасной области способны взаимодействовать с CDR во многих антителах (Chothia and Lesk, J. Mol. Biol. 196, 901 (1987), Chothia et al., Nature 342, 877 (1989), и Tramontano et al., J. Mol. Biol. 215, 175 (1990), все из которых включены в настоящее описание в качестве ссылок), особенно в положениях 2, 48, 64 и 71 легкой цепи и 26-30, 71 и 94 тяжелой цепи (нумерация в соответствии с Kabat, ук. соч.), и, таким образом, эти аминокислоты, как правило, относятся к категории 4. Как правило, гуманизированные иммуноглобулины согласно изобретению включают в себя донорные аминокислоты (где отличаются) из категории 4, в дополнение к этим аминокислотам. Аминокислоты в положениях 35 легкой цепи и 93 и 103 тяжелой цепи, вероятно, также взаимодействуют с CDR. Во всех этих перечисленных положениях предпочтительным является выбор для гуманизированного иммуноглобулина донорной аминокислоты вместо акцепторной аминокислоты (если они отличаются). С другой стороны, определенные положения, которые могут относиться к категории 4, такие как первые 5 аминокислот легкой цепи, иногда можно выбирать из акцепторного иммуноглобулина без утраты аффинности в гуманизированном иммуноглобулине. Chothia and Lesk (ук. соч.) определяют CDR иным образом, чем Kabat et al. (ук. соч.). В частности, CDR1 определяют, как включающий остатки 26-32. Таким образом, Riechmann et al. (ук. соч.) выбирают эти аминокислоты из донорных иммуноглобулинов.
Также важно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и других положительных биологических свойств. Для достижения этой цели, в соответствии с предпочтительным способом гуманизрованные антитела получают посредством процесса анализа исходных последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Доступны компьютерные программы, которые иллюстрируют и отображают вероятные трехмерные конформационные структуры выбранных в качестве кандидатов последовательностей иммуноглобулинов. Исследование этих изображений позволяет анализировать вероятную роль остатков в функционировании последовательности иммуноглобулина-кандидата, т.е. анализировать остатки, которые влияют на способность иммуноглобулина-кандидата связывать свой антиген. Таким образом, остатки FR можно выбирать и объединять из реципиентных и импортных последовательностей, чтобы достигать требуемого свойства антитела, такого как повышенная аффинность к антигену(ам)-мишени. Как правило, остатки CDR непосредственно и наиболее существенно вовлечены во влияние на связывание антигена. Компьютерные программы для создания моделей белков, таких как антитела, широко доступны и хорошо известны специалистам в данной области (смотрите Levy et al., Biochemistry, 28, 7168-7175 (1989); Bruccoleri et al., Nature, 335, 564-568 (1988); Chothia et al., Science, 233, 755-758 (1986), все из которых включены в настоящее описание в качестве ссылок). Они не являются частью этого изобретения. Действительно, вследствие того, что все антитела обладают сходными структурами, известные структуры антитела, которые являются доступными из Белковой базы данных Брукхевена, можно использовать, если необходимо, в качестве приближенных моделей других антител. Коммерчески доступные компьютерные программы можно использовать для выведения изображения этих моделей на монитор компьютера, для вычисления расстояния между атомами и для оценки вероятности взаимодействия различных аминокислот (смотрите Ferrin et al., J. Mol. Graphics, 6, 13-27 (1988)).
В дополнение к указанным выше категориям, которые определяют, когда аминокислоту в гуманизированном иммуноглобулине можно выбирать от донора, определенные аминокислоты в гуманизированном иммуноглобулине можно выбирать ни из донора, ни из акцептора, если они относятся к:
Категории 5: Если аминокислота в данном положении донорного иммуноглобулина является "редкой" для последовательностей человека, и аминокислота в этом положении в акцепторном иммуноглобулине также является "редкой" для последовательностей человека, как определено выше, тогда можно выбирать аминокислоту в этом положении гуманизированного иммуноглобулина, которая представляет собой некоторую аминокислоту, "типичную" для последовательностей человека. Предпочтительным выбором является аминокислота, которая наиболее часто встречается в этом положении в известных последовательностях человека, принадлежащих к той же подгруппе, что и акцепторная последовательность.
Гуманизированные антитела, как правило, обладают по меньшей мере тремя потенциальными преимуществами по сравнению с антителами мыши или, в некоторых случаях, с химерными антителами, для применения при лечении человека:
(1) Вследствие того, что эффекторная часть является человеческой, она может лучше взаимодействовать с другими частями иммунной системы человека (например, более эффективно разрушать клетки-мишени посредством комплементзависимой цитотоксичности (CDC) или антителозависимой клеточной цитотоксичности (ADCC)).
(2) Иммунная система человека не должна распознавать каркасную область или константный участок гуманизированного антитела как чужеродные, и, таким образом, антительный ответ против такого инъецированного антитела должен быть меньшим, чем против полностью чужеродного антитела мыши или частично чужеродного химерного антитела.
(3) Было описано, что инъецированные антитела мыши обладают временем полужизни в кровотоке человека, значительно более коротким, чем время полужизни нормальных антител (D. Shaw et al., J. Immunol., 138, 4534-4538 (1987)). Инъецированные гуманизированные антитела, предположительно, обладают временем полужизни, более сходным с встречающимися в природе антителами человека, позволяя введение меньших и менее частых доз.
В одном аспекте настоящее изобретение относится к конструированию гуманизированных антител, которые получают посредством экспрессии рекомбинантных сегментов ДНК, кодирующих CDR тяжелой и легкой цепей из донорного иммуноглобулина, способного связываться с требуемым антигеном, таким как IL-31, присоединенных к сегментам ДНК, кодирующим акцепторные каркасные области человека.
Сегменты ДНК, как правило, дополнительно включают в себя последовательности контроля экспрессии ДНК, функционально связанные с кодирующими гуманизированный иммуноглобулин последовательностями, включающие в себя ассоциированные с ними в природе или гетерологичные промоторные участки. Последовательности контроля экспрессии представляют собой эукариотические промоторные системы в векторах, способных к трансформации или трансфекции в эукариотические клетки-хозяева, однако также можно использовать контролирующие последовательности для прокариотических хозяев. После встраивания вектора в соответствующего хозяина, хозяина содержат в условиях, пригодных для экспрессии нуклеотидных последовательностей на высоком уровне, и, если желательно, затем можно выделять и очищать гуманизированные легкие цепи, тяжелые цепи, димеры легкая/тяжелая цепь или целые антитела, связывающие фрагменты или другие формы иммуноглобулина (смотрите S. Beychok, Cells of Immunoglobulin Synthesis, Academic Press, N.Y., (1979), которая включена в настоящее описание в качестве ссылки).
Последовательности ДНК константного участка человека можно выделять в соответствии с хорошо известными способами из разнообразных клеток человека, но предпочтительно из иммортализованных B-клеток (смотрите Kabat ук. соч. и WP87/02671). Источником CDR для получения иммуноглобулинов согласно изобретению аналогично являются моноклональные антитела, способные связываться с определенным антигеном, таким как IL-31, и получаемые хорошо известными способами из любого пригодного источника, представляющего собой млекопитающего, включая мышей, крыс, кроликов, или из других позвоночных, способных продуцировать антитела. Пригодные клетки, являющиеся источником последовательностей ДНК константного участка и каркасной области, и клетки-хозяева для экспрессии и секреции иммуноглобулина, можно получать из множества источников, таких как Американская коллекция типовых культур ("Catalogue of Cell Lines and Hybridomas", шестое издание (1988) Rockville, Md. U.S.A., который включен в настоящее описание в качестве ссылки).
В дополнение к гуманизированным иммуноглобулинам, конкретно описанным в настоящем описании, можно легко конструировать и изготавливать другие "по существу гомологичные" природной последовательности модифицированные иммуноглобулины с использованием различных способов рекомбинантных ДНК, хорошо известных специалистам в данной области. В качестве основы гуманизированных иммуноглобулинов согласно изобретению можно использовать множество различных каркасных областей человека, отдельно или в сочетании. Как правило, модификации генов можно легко проводить посредством множества хорошо известных способов, таких как сайт-направленный мутагенез (смотрите Gillman and Smith, Gene, 8, 81-97 (1979) и S. Roberts et al., Nature, 328, 731-734 (1987), обе из которых включены в настоящее описание в качестве ссылок).
Гуманизированные антитела согласно изобретению включают в себя фрагменты, а также целые антитела. Как правило, эти фрагменты конкурируют с целым антителом, из которого они получены, за связывание антигена. Фрагменты, как правило, связываются с аффинностью по меньшей мере 107 M-1, и более конкретно 108 или 109 M-1 (т.е. в том же диапазоне, что и целое антитело). Фрагменты гуманизированного антитела включают в себя отдельные тяжелые цепи, легкие цепи, Fab, Fab', F(ab')2 и Fv. Фрагменты получают способами рекомбинантных ДНК, или посредством ферментативного или химического расщепления целых иммуноглобулинов.
Для более подробного описания гуманизации антител смотрите европейские патенты №№ EP 239400, EP 592106 и EP 519596; международные публикации №№ WO 91/09967 и WO 93/17105; патенты США №№ 5225539, 5530101, 5565332, 5585089, 5766886 и 6407213; и Padlan, 1991, Molecular Immunology 28(4/5): 489 498; Studnicka et al., 1994, Protein Engineering 7(6): 805 814; Roguska et al., 1994, PNAS 91: 969 973; Tan et al., 2002, J. Immunol. 169: 1119 25; Caldas et al., 2000, Белок Eng. 13: 353 60; Morea et al., 2000, Methods 20: 267 79; Baca et al., 1997, J. Biol. Chem. 272: 10678 84; Roguska et al., 1996, Protein Eng. 9: 895 904; Couto et al., 1995, Cancer Res. 55 (23 Supp): 5973s 5977s; Couto et al., 1995, Cancer Res. 55: 1717 22; Sandhu, 1994, Gene 150: 409 10; Pedersen et al., 1994, J. Mol. Biol. 235: 959 73; Jones et al., 1986, Nature 321: 522-525; Reichmann et al., 1988, Nature 332: 323-329; и Presta, 1992, Curr. Op. Struct. Biol. 2: 593-596.
Для получения фрагментов антител разработаны различные способы. Традиционно эти фрагменты получают посредством протеолитического расщепления целых антител (смотрите, например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24: 107-117 (1992) и Brennan et al., Science, 229:81 (1985)). Однако в настоящее время эти фрагменты можно получать с помощью рекомбинантных клеток-хозяев. Например, фрагменты антител можно выделять из фаговых библиотек антител, описанных выше. Альтернативно фрагменты Fab'-SH можно непосредственно выделять из E. coli и химически соединять с образованием фрагментов F(ab').sub.2 (Carter et al., Bio/Technology 10: 163-167 (1992)). В соответствии с другим подходом фрагменты F(ab').sub.2 можно выделять непосредственно из культур рекомбинантных клеток-хозяев. Другие способы получения фрагментов антител понятны квалифицированному специалисту. Кроме того, примеры способов, которые можно использовать для получения одноцепочечных Fv и антител, включают в себя способы, описанные в патентах США 4946778 и 5258498; Huston et al., Methods in Enzymology 203:46-88 (1991); Shu et al., PNAS 90:7995-7999 (1993) и Skerra et al., Science 240:1038-1040 (1988).
Настоящее изобретение относится к антителам, слитым рекомбинантными способами или химически конъюгированным (включая как ковалентную, так и нековалентную конъюгацию) с полипептидом (или его частью, предпочтительно по меньшей мере с 10, 20 или 50 аминокислотами полипептида) согласно изобретению с получением слитых белков. Таким образом, это изобретение также относится к иммуноконъюгатам, содержащим антитело, описанное в настоящем описании, конъюгированное с цитотоксическим веществом, таким как химиотерапевтическое средство, токсин (например, ферментативно активный токсин бактерий, грибов, растений или животных или его фрагменты), или радиоактивный изотоп (т.е. радиоконъюгат).
Настоящее изобретение относится к антителам, слитым рекомбинантными способами или химически конъюгированным (включая как ковалентную, так и нековалентную конъюгацию) с полипептидом (или его частью, предпочтительно по меньшей мере с 10, 20 или 50 аминокислотами полипептида) согласно изобретению с получением слитых белков. Слияние не обязательно должно быть прямым, а может быть проведено с помощью линкерных последовательностей. Антитела могут быть специфичными в отношении антигенов, отличных от полипептидов (или их частей, предпочтительно по меньшей мере 10, 20 или 50 аминокислот полипептида) согласно изобретению. Например, антитела можно использовать для нацеливания полипептидов согласно изобретению на конкретные типы клеток либо in vitro, либо in vivo, посредством проведения слияния или конъюгации полипептидов согласно изобретению с антителами, специфичными в отношении конкретных рецепторов клеточной поверхности. Антитела, слитые или конъюгированные с полипептидами согласно изобретению, также можно использовать в иммунологических анализах in vitro и способах очистки с использованием известных в данной области способов. Смотрите, например, Harbor et al., выше, и публикацию PCT WO 93/21232; EP 439095; Naramura et al., Immunol. Lett. 39:91-99 (1994); патент США № 5474981; Gillies et al., PNAS 89:1428-1432 (1992); Fell et al., J. Immunol. 146:2446-2452(1991), которые включены в качестве ссылок в полном объеме.
Кроме того, настоящее изобретение относится к композициям, содержащим полипептиды согласно изобретению (например, полипептиды, содержащие иммуногенный или антигенный эпитоп), слитые или конъюгированные с гетерологичными полипептидными последовательностями (например, доменами антитела, отличными от вариабельных участков). Например, полипептиды согласно изобретению могут быть слитыми или конъюгированными с Fc-участком антитела или его частью. Например, полипептиды согласно изобретению (включая их фрагменты или варианты) могут быть слитыми с константным доменом иммуноглобулинов (IgA, IgE, IgG, IgM), или их частями (CH1, CH2, CH3 или любым их сочетанием или их частью), с получением химерных полипептидов. Часть антитела, слитая с полипептидом согласно изобретению, может содержать константный участок, шарнирную область, домен CH1, домен CH2 и домен CH3 или любое сочетание целых доменов или их частей. Полипептиды также могут быть слитыми или конъюгированными с указанными выше частями антитела с образованием мультимеров. Например, Fc-участки, слитые с полипептидами согласно изобретению, могут формировать димеры посредством образования дисульфидных связей между Fc-участками. Более мультимерные формы можно получать посредством слияния полипептидов с частями IgA и IgM. Способы слияния или конъюгации полипептидов согласно изобретению с частями антитела известны в данной области. Смотрите, например, патенты США №№ 5336603; 5622929; 5359046; 5349053; 5447851; 5112946; EP 307434; EP 367166; публикации PCT WO 96/04388; WO 91/06570; Ashkenazi et ah, Proc. Natl. Acad. Sci. USA 88:10535-10539 (1991); Zheng et al., J. Immunol. 154:5590-5600 (1995); и Vil et al., Proc. Natl. Acad. Sci. USA 89:11337-11341 (1992) (указанные ссылки включены в качестве ссылок в полном объеме). В качестве другого неограничивающего примера, полипептиды и/или антитела согласно изобретению (включая их фрагменты или варианты) могут быть слитыми с альбумином (включая, но не ограничиваясь ими, рекомбинантный сывороточный альбумин человека или его фрагменты или варианты (смотрите, например, патент США № 5876969, выданный 2 марта 1999 года, патент EP 0413622 и патент США № 5766883, выданный 16 июня 1998 года, включенные в настоящее описание в качестве ссылок в полном объеме). Полипептиды и/или антитела согласно изобретению (включая их фрагменты или варианты) могут быть слитыми либо с N-концом, либо с C-концом гетерологичного белка (например, Fc-полипептида иммуноглобулина или полипептида сывороточного альбумина человека). Также к этому изобретению относятся полинуклеотиды, кодирующие слитые белки согласно изобретению.
Как описано выше, полипептиды согласно изобретению могут быть слитыми или конъюгированными с указанными выше частями антитела для повышения времени полужизни полипептидов in vivo или для применения в иммунологических анализах с использованием известных в данной области способов. Кроме того, полипептиды согласно изобретению могут быть слитыми или конъюгированными с указанными выше частями антител для упрощения очистки. В одном опубликованном примере описаны химерные белки, состоящие из первых двух доменов CD4-полипептида человека и различных доменов константных участков тяжелых или легких цепей иммуноглобулинов млекопитающих. (Смотрите, например, EP 394827; Traunecker et al., Nature 331:84-86 (1988)). Повышенная доставка антигена через эпителиальный барьер к иммунной системе показана для антигенов (например, инсулина), конъюгированных с партнером для связывания FcRn, таким как IgG или Fc-фрагменты (смотрите, например, публикации PCT WO 96/22024 и WO 99/04813). Полипептиды согласно изобретению, слитые или конъюгированные с антителом, обладающим связанными дисульфидными связями димерными структурами (вследствие IgG), также могут быть более эффективными в отношении связывания и нейтрализации других молекул, чем мономерный секретируемый белок или фрагмент белка отдельно (Fountoulakis et al., J. Biochem. 270:3958-3964 (1995)). Во многих случаях Fc-часть в слитом белке является пригодной для лечения и диагностики и, таким образом, может приводить, например, к улучшенным фармакокинетическим свойствам (EP A 232262). Альтернативно является желательным удаление Fc-части после экспрессии, выявления и очистки слитого белка. Например, Fc-участок может препятствовать лечению и диагностике, если слитый белок используют в качестве антигена для иммунизаций. При разработке лекарственных средств, например, проводили слияние белков человека, таких как hIL-5, с Fc-участками, в целях высокопроизводительных скрининговых анализов для идентификации антагонистов hIL-5. (Смотрите D. Bennett et al., J. Molecular Recognition 8:52-58 (1995); K. Johanson et al., J. Biol. Chem. 270:9459-9471 (1995)). Такие способы также включают в себя, но не ограничиваются ими, применение бифункциональных веществ для конъюгации (смотрите, например, патенты США №№ 5756065; 5714631; 5696239; 5652361; 5505931; 5489425; 5435990; 5428139; 5342604; 5274119; 4994560 и 5808003; содержание каждого из которых включено в настоящее описание в качестве ссылки в полном объеме).
Более того, можно проводить слияние полипептидов согласно изобретению (например, антител или их фрагментов) с маркерными последовательностями, такими как пептид, для упрощения их очистки. В следующем варианте осуществления также можно проводить рекомбинацию нуклеиновых кислот, кодирующих полипептиды согласно изобретению (включая, но не ограничиваясь ими, нуклеиновые кислоты, кодирующие иммуногенные и/или антигенные эпитопы), с представляющим интерес геном, таким как маркер эпитопа (например, гемагглютининовый маркер ("HA") или маркер flag), для упрощения детекции и очистки экспрессированного полипептида. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид, такой как маркер, представленный в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, Calif., 91311), помимо прочих маркеров, многие из которых являются коммерчески доступными. Как описано в Gentz et al., Proc. Natl. Acad. Sci. USA 86:821-824 (1989), например, гексагистидин обеспечивает удобную очистку слитого белка. Другие пептидные маркеры, пригодные для очистки, включают в себя, но не ограничиваются ими, маркер "HA", который соответствует эпитопу из белка гемагглютинина вируса гриппа (Wilson et al., Cell 37:767 (1984)) и маркер "flag".
Кроме того, настоящее изобретение относится к антителам или их фрагментам, конъюгированным с диагностическим или лекарственным средством. Антитела можно использовать в диагностических целях, например, для мониторинга развития или прогрессии опухоли, как часть процесса клинического тестирования, например, для определения эффективности назначенной схемы лечения, диагностики, выявления и/или профилактики. Выявление можно упрощать посредством присоединения антитела к поддающемуся детекции веществу. Примеры поддающихся детекции веществ включают в себя различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества, радиоактивные вещества, позитронно-активные металлы, используемые в различных способах позитронно-эмиссионной томографии, и нерадиоактивные ионы металлов. Смотрите, например, патент США № 4741900, для ионов металлов, которые можно конъюгировать с антителами для применения в диагностике в соответствии с настоящим изобретением. Примеры пригодных ферментов включают в себя пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; примеры пригодных комплексов простетических групп включают в себя стрептавидин/биотин и авидин/биотин; примеры пригодных флуоресцентных веществ включают в себя умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин-флуоресцеин, дансилхлорид или фикоэритрин; пример люминесцентного вещества включает в себя люминол; примеры биолюминесцентных веществ включают в себя люциферазу, люциферин и экворин; и примеры пригодного радиоактивного вещества включают в себя 125I, 131I, 111In или 99Tc.
Кроме того, антитело или его фрагмент можно конъюгировать с терапевтической группой, такой как цитотоксин, например, цитостатическое или цитоцидное вещество, лекарственное средство или ион радиоактивного металла. Цитотоксин или цитотоксическое вещество представляет собой вещество, вызывающее гибель клетки. Примеры включают в себя паклитаксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопзид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, татракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи. Лекарственные средства включают в себя, но не ограничиваются ими, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацилдекарбазин), алкилирующие средства (например, мехлорэтамин, тиотепу, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бисульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиамин-платину (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)) и антимитотические срества (например, винкристин и винбластин).
Конъюгаты согласно изобретению можно использовать для модификации данного биологического ответа, лекарственное средство или группу лекарственного средства не следует понимать, как ограниченные классическими лекарственными средствами. Например, группа лекарственного средства может представлять собой белок или полипептид, обладающий требуемой биологической активностью. Такие белки могут включать в себя, например, токсин, такой как абрин, рицин A, экзотоксин pseudomonas или дифтерийный токсин; белок, такой как фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, тромбоцитатный фактор роста, тканевой активатор плазминогена, тромботическое вещество или антиангиогенное вещество, например, ангиостатин или эндостатин; или модификаторы биологического ответа, такие как, например, лимфокины, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF"), гранулоцитарный колониестимулирующий фактор ("G-CSF") или другие факторы роста.
Антитела также могут быть связаны с твердыми подложками, которые являются особенно пригодными для иммунологических анализов или очистки антигена-мишени. Такие твердые подложки включают в себя, но не ограничиваются ими, стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
Способы конъюгации таких терапевтических групп с антителами хорошо известны, смотрите, например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", в Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", в Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", в Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", в Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev. 62:119-58 (1982).
Альтернативно антитело можно конъюгировать со вторым антителом с образованием гетероконъюгата антитела, как описано Segal в патенте США № 4676980, который включен в настоящее описание в качестве ссылки в полном объеме.
Антитело с конъюгированной с ним терапевтической группой, или без нее, вводимое отдельно или в сочетании с цитотоксическим фактором(ами) и/или цитокином(ами), можно использовать в качестве лекарственного средства.
В другом варианте осуществления антитело может быть конъюгированным с "рецептором" (таким как стрептавидин) для применения в предварительном нацеливании на опухоль, где конъюгат антитело-рецептор вводят пациенту, а затем с использованием вещества для выведения удаляют из кровотока несвязанный конъюгат, с последующим введением "лиганда" (например, авидина), который конъюгирован с цитотоксическим веществом (например, радионуклидом).
Для выявления антител, которые связываются с белками или полипептидами IL-31, можно использовать множество способов, известных специалистам в данной области. Иллюстративные способы анализа подробно описаны в Antibodies: A Laboratory Manual, Harlow and Lane (Eds.), Cold Spring Harbor Laboratory Press, 1988. Репрезентативные примеры таких анализов включают в себя: совмещенный иммуноэлектрофорез, радиоиммунный анализ, радиоиммунопреципитацию, твердофазный иммуноферментный анализ (ELISA), дот-блот или вестерн-блот анализ, ингибиторный или конкурентный анализ и "сэндвич"-анализ. Кроме того, можно проводить скрининг антител на связывание с белком или полипептидом IL-31 дикого типа относительно мутантной формы.
Антитела против IL-31 можно использовать для маркирования клеток, которые экспрессируют IL-31; для выделения IL-31 посредством аффинной очистки; для диагностических анализов для определения уровней полипептидов IL-31 в кровотоке; для выявления или количественного определения растворимого IL-31 в качестве маркера связанной с ним патологии или заболевания; в аналитических способах с использованием способов FACS; для скрининга экспрессирующих библиотек; для получения антиидиотипических антител; и в качестве нейтрализующих антител или в качестве антагонистов для блокирования активности IL-31 in vitro и in vivo. Пригодные прямые маркеры или метки включают в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и т.п.; характерной чертой непрямых маркеров может быть применение пар биотин-авидин или других пар комплемент/антикомплемент в качестве промежуточных соединений. Описанные в настоящем описании антитела также можно прямо или непрямо конъюгировать с лекарственными средствами, токсинами, радионуклидами и т.п., и эти конъюгаты можно использовать для диагностических или терапевтических способов применения in vivo. Более того, антитела против IL-31 или их фрагменты можно использовать in vitro для выявления денатурированного IL-31 или его фрагментов в анализах, например, в вестерн-блотах или других анализах, известных в данной области.
Пригодные поддающиеся детекции молекулы могут быть прямо или непрямо присоединены к полипептиду или антителу и могут включать в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и т.п. Пригодные цитотоксические молекулы могут быть прямо или непрямо присоединены к полипептиду или антителу и могут включать в себя бактериальные или растительные токсины (например, дифтерийный токсин, сапорин, экзотоксин Pseudomonas, рицин, абрин и т.п.), а также терапевтические радионуклиды, такие как йод-131, рений-188 или иттрий-90 (либо прямо присоединенные к полипептиду или антителу, либо непрямо присоединенные, например, посредством хелатирующей группы). Полипептиды или антитела также могут быть конъюгированы с цитотоксическими лекарственными средствами, такими как адриамицин. Для непрямого присоединения поддающейся детекции или цитотоксической молекулы, поддающуюся детекции или цитотоксическую молекулу можно конъюгировать с членом пары комплементарное вещество/антикомплементарное вещество, где другой член связывают с полипептидом или частью антитела. Для этих целей иллюстративной парой комплементарное вещество/антикомплементарное вещество является пара биотин/стрептавидин.
Связывающиеся полипептиды также могут действовать как "антагонисты" IL-31 для блокирования связывания IL-31 и передачи сигнала in vitro и in vivo. Эти связывающиеся полипептиды против IL-31 могут быть пригодными для ингибирования активности IL-31 или связывания белка.
Слитые белки полипептид-токсин и слитые белки антитело-токсин также можно использовать для ингибирования или разрушения клеток-мишеней или тканей-мишеней (например, для воздействия на злокачественные клетки или ткани). Альтернативно, если полипептид обладает различными функциональными доменами (т.е. активирующим доменом или связывающим рецептор доменом, плюс домен для нацеливания), то слитый белок, включающий в себя только домен для нацеливания, может быть пригодным для направления поддающейся детекции молекулы, цитотоксической молекулы или комплементарной молекулы в представляющий интерес тип клетки или ткани. В случаях, когда слитый белок только с одним доменом включает в себя комплементарную молекулу, антикомплементарную молекулу можно конъюгировать с поддающейся детекции или цитотоксической молекулой. Такие слитые белки домен-комплементарная молекула, таким образом, представляют собой типичный носитель для нацеливания или носитель для специфичной в отношении клетки/ткани доставки типичных конъюгатов антикомплементарной поддающейся детекции/цитотоксической молекулы.
В другом варианте осуществления можно использовать слитые белки антитело против IL-31-цитокин для уничтожения in vivo тканей-мишеней (например, тканей лейкоза, лимфомы, рака легкого, рака толстого кишечника, меланомы, рака поджелудочной железы, рака яичника, злокачественных опухолей кожи, крови и костного мозга, или других злокачественных опухолей, где экспрессируются рецепторы для IL-31) (Смотрите, главным образом, Hornick et al., Blood 89:4437-47, 1997). Описанные слитые белки осуществляют нацеливание антитела в требуемую область действия, обеспечивая посредством этого повышенную локальную концентрацию антитела. Пригодные полипептиды IL-31 или антитела против IL-31 направлены на нежелательную клетку или ткань (т.е. опухоль или лейкоз), и они повышают опосредуемый слитым с ними цитокином лизис клетки-мишени эффекторными клетками. Пригодные для этих целей цитокины включают в себя, например, интерлейкин-2 и гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF).
В другом варианте осуществления, если полипептид IL-31, или антитело против IL-31, нацелен на клетки или ткани сосудов, такой полипептид или антитело можно конъюгировать с радионуклидом и, в частности, с испускающим бета-излучение радионуклидом, для снижения рестеноза.
Моноклональные антитела согласно изобретению можно определять по их способности ингибировать, блокировать или нейтрализовать лиганд IL-31, как определяют посредством различных моделей in vivo, известных в данной области и описанных в настоящем описании, включая, но не ограничиваясь ими, модель NC/Nga, накожную модель Ova, модель хронической гиперчувствительности и хроническую модель с гаптеном.
Как направленные в кожу T-клетки, так и эпидермальные кератиноциты, вовлечены в патологию кожных заболеваний у человека. Экспрессия мРНК и белка IL-31 у человека ограничивается направленной в кожу популяцией T-клеток CLA+. По существу, антагонист IL-31, включая антитело или антагонист рецептора, будет пригодным для лечения заболеваний кожи и эпидермиса, при которых выявляются T-клетки CLA+. Такие заболевания включают в себя, например, атопический дерматит, контактный дерматит, индуцируемые лекарственным средством аллергические реакции, тропные к коже вирусы и ассоциированный с вирусом зуд, витилиго, кожную T-клеточную лимфому, очаговую алопецию, красные угри, обыкновенные угри, узловатую почесуху и буллезный пемфигоид. Для определения эффекта нейтрализующего моноклонального антитела против IL-31 являются пригодными хемокиновые маркеры, такие как TARC и MDC. Как представлено в примере 15, ингибиторные эффекты лечения антителами против IL-31, описанными в настоящем описании, можно определять посредством мониторинга уровней TARC и MDC.
Контактный дерматит
Аллергический контактный дерматит определяют как опосредуемую T-клетками иммунную реакцию на антиген, который контактирует с кожей. Считают, что популяция T-клеток CLA+ вовлечена в запуск дерматита, поскольку зависимый от аллергена T-клеточный ответ в основном ограничивается популяцией клеток CLA+ (Смотрите Santamaria-Babi, L.F., et al., J Exp Med: 181, 1935, (1995)). Последние данные показали, что только клетки памяти (CD45RО+) CD4+ CLA+, но не CD8+ T-клетки, в ответ на никель, распространенный аллерген контактной гиперчувствительности, пролифирируют и продуцируют цитокины как 1 типа (IFN-), так 2 типа (IL-5). Более того, количество клеток, экспрессирующих CLA в сочетании с CD4, CD45RO (памяти) или CD69, возрастает после специфичной стимуляции никелем, и они экспрессируют рецепторы для хемокинов CXCR3, CCR4, CCR10, но не CCR6. Смотрите Moed H., et al., Br J Dermatol:51, 32, (2004).
В моделях на животных было показано, что аллергический контактный дерматит является зависимым от T-клеток и что отвечающие за аллергическую реакцию T-клетки мигрируют в область нанесения аллергена. Смотрите главным образом: Engeman T.M., et al., J Immunol: 164, 5207, (2000); Ferguson T.A. & Kupper T.S. J Immunol: 150, 1172, (1993); и Gorbachev A.V. & Fairchild R.L. Crit Rev Immunol: 21, 451(2001). Поскольку T-клетки CLA+ продуцируют IL-31, и стимуляция кератиноцитов кожи посредством IL-31 может индуцировать провоспалительные хемокины, такие как TARC и MDC, IL-31 может быть вовлечен в патофизиологию контактного дерматита. Нейтрализующие антитела против IL-31 применяли в модели контактной гиперчувствительности на мышах. Смотрите пример 8.
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем описании, можно применять для улучшения клинического исхода контактной гиперчувствительности посредством ингибирования, снижения, предотвращения или блокирования воспаления и/или расчесывания, связанных с заболеванием.
Атопический дерматит
Атопический дерматит (AD) представляет собой хроническое рецидивирующее воспалительное заболевание кожи, и его частота в течение последних десятилетий резко возросла. Клинически AD характеризуется сильно зудящими часто шелушащимися бляшками и папулами, которые отражают хроническое рецидивирующее течение. Диагноз AD, главным образом, основан на главных и второстепенных клинических признаках. Смотрите Hanifin J.M., Arch Dermatol: 135, 1551 (1999). При гистопатологии выявляют спонгиоз, гиперкератоз и фокальный паракератоз в острых очагах повреждения, в то время как выраженная гиперплазия эпидермиса с гиперкератозом и паракератозом, акантозом/гипергранулезом и околососудистой инфильтрацией дермы лимфоцитами и изобилием тучных клеток являются признаками хронических очагов повреждения.
T-клетки играют центральную роль в запуске локальных иммунных ответов в тканях, и данные подтверждают, что инфильтрирующие кожу T-клетки, в частности, могут играть ключевую роль в запуске и поддержании иммунных ответов с нарушенной регуляцией в коже. Приблизительно 90% инфильтрированных T-клеток в воспалительных областях кожи экспрессируют кожный ассоциированный с лимфоцитами Ag (CLA+), который связывает E-селектин, индуцибельную молекулу адгезии на эндотелии (описан в Santamaria-Babi L.F., et al., Eur J Dermatol: 14, 13, (2004)). Документировано значительное повышение циркулирующих T-клеток CLA+ среди пациентов с AD по сравнению с контрольными индивидуумами (Смотрите Teraki Y., et al., Br J Dermatol: 143, 373 (2000)), в то время как в других источниках показано, что на экстракт аллергена предпочтительно отвечают T-клетки памяти CLA+ пациентов с AD по сравнению с популяцией CLA- (Смотрите Santamaria-Babi, L.F., et al., J Exp Med:181, 1935, (1995)). У человека патогенез атопического нарушения кожи ассоциирован с повышением количества T-клеток CLA+, которые экспрессируют повышенные уровни цитокинов типа Th-2, таких как IL-5 и IL-13, 9, 10. Смотрите Akdis M., et al., Eur J Immunol: 30, 3533 (2000); и Hamid Q., et al., J Allergy Clin Immunol: 98, 225 (1996).
У мышей NC/Nga спонтанно развиваются подобные AD очаги повреждения, которые соответствуют AD человека во многих аспектах, включая клиническое течение и признаки, гистопатологию и иммунопатологию, при содержании в условиях без неспецифичного патогена (не-SPF) в возрасте приблизительно 6-8 недель. В противоположность, у мышей NC/Nga, которых содержат в условиях SPF, кожные очаги повреждения не развиваются. Однако начало спонтанных повреждений кожи и поведения с расчесыванием можно синхронизировать у мышей NC/Nga, содержащихся в условиях SPF, посредством недельной внутрикожной инъекции неочищенного антигена пылевого клеща. Смотрите Matsuoka H., et al., Allergy: 58, 139 (2003). Таким образом, развитие AD у NC/Nga является приемлемой моделью для оценки новых лекарственных средств для лечения AD.
В дополнение к модели спонтанного AD у NC/Nga, накожную сенсибилизацию мышей с использованием OVA также можно использовать в качестве модели для индукции антиген-зависимого утолщения эпидермы и дермы с инфильтратом мононуклеаров в коже сенсибилизированных мышей. Это, как правило, совпадает с повышением сывороточных уровней общих и специфичных IgE, однако в норме в этой модели не происходит дисфункции кожного барьера или зуда. Смотрите Spergel J.M., et al., J Clin Invest, 101: 1614, (1998). Этот протокол можно модифицировать в целях индукции дисрегуляции кожного барьера и зуда посредством сенсибилзации трансгенных мышей DO11.10 OVA TCR посредством OVA. Повышенное количество антиген-специфичных T-клеток, которые могут распознавать сенсибилизирующий антиген, может повышать уровень воспаления в коже с индукцией видимого расчесывания и лихенизации/шелушения кожи.
Как модель спонтанного AD на NC/Nga, так и накожную модель с OVA в DO11.10, используют для изучения экспрессии IL-31 и IL-31RA при AD, а также способности антител, описанных в настоящем описании, ингибировать, снижать или нейтрализовать эффекты IL-31. Смотрите примеры 9 и 10 настоящего описания. В одном исследовании с использованием модели NC/Nga in vivo (пример 10), введение антител крысы против IL-31 мыши показало уменьшение расчесывания, но не уменьшение дерматита. Антитела, описанные в настоящем описании, являются пригодными для ингибирования расчесывания, обусловленного дерматитом и связанных с зудом заболеваний, включая атопический дерматит, узловатую почесуху и экзему. Ингибирование, уменьшение или предотвращение расчесывания отдельно может быть эффективным для лечения связанных с зудом заболеваний, включая, но не ограничиваясь ими, атопический дерматит, узловатую почесуху и экзему, поскольку прекращение расчесывания остановит прогрессирование дерматита, развитие которого зависит от расчесывания.
Дополнительные модели для определения ингибиторных эффектов антител против IL-31, описанных в настоящем описании, описаны в Umeuchi, H. et al., European Journal of Pharmacology, 518: 133-139, 2005; и в Yoo, J. et al., J. Experimental Medicine, 202:541-549, 2005.
Таким образом, нейтрализацию IL-31 с помощью антител, описанных в настоящем описании, можно использовать для улучшения клинического исхода дерматита и связанных с зудом заболеваний, включая атопический дерматит, узловатую почесуху и экзему, посредством ингибирования, снижения, предотвращения или блокирования воспаления и/или расчесывания, ассоциированных с заболеванием.
Индуцированные лекарственным средством кожные аллергические реакции замедленного типа
Индуцированные лекарственным средством кожные аллергические реакции замедленного типа являются высоко гетерогенными и могут отражать множество различных патофизиологических процессов. Смотрите Brockow K., et al., Allergy: 57, 45 (2002). Было показано, что иммунологические механизмы, вовлеченные в эти реакции, либо опосредуются антителами, либо являются клеточно-опосредуемыми. При аллергии на лекарственное средство немедленного типа, IgE-опосредуемая антительная реакция может быть показана с помощью инъекционной кожной пробы и/или интрадермального теста через 20 мин, в то время как не являющиеся немедленными реакции на лекарственные средства могут происходить более чем через один час после последнего приема лекарственного средства и часто опосредованы T-клетками. Не являющиеся немедленными опосредуемые T-клетками реакции замедленного типа могут происходить у пациентов с побочными реакциями лекарственного средства, например, на пенициллины. Было показано, что у пациентов с аллергией на пенициллин пролиферативные T-клеточные ответы на пенициллины ограничиваются субпопуляцией T-клеток памяти (CD45RO+) CLA+, в то время как в подгруппе CD45RO+ CLA- показано отсутствие пролиферативного ответа. Смотрите Blanca M., Leyva L., et al., Blood Cells Mol Dis:31, 15 (2003). Реакции гиперчувствительности замедленного типа (ГЗТ) можно искусственно воспроизводить у мышей, что позволяет проводить оценку факторов, которые могут быть вовлечены в инициацию и поддержание ответа ГЗТ. Нейтрализующее антитело против IL-31 может быть эффективным при реакции гиперчувствительности замедленного типа. Смотрите пример 51.
Токсический эпидермальный некролиз (TEN) представляет собой очень редкую, но крайне тяжелую реакцию на лекарственное средство, характеризующуюся распространенным апоптозом эпидермиса с обширными волдырями. В исследованиях было показано, что лимфоциты, инфильтрирующие волдыри, представляют собой T-клетки CLA+, и они могут проявлять цитотоксичность в отношении кератиноцитов эпидермиса. Смотрите Leyva L., et al.,. J Allergy Clin Immunol: 105, 157 (2000); и Nassif A., Bensussan A., et al., J Allergy Clin Immunol:114, 1209 (2004). Система с трансгенными мышами, в которой OVA экспрессируется под контролем промотора кератина-5 (K5) в кератиноцитах эпидермиса и волосяного фолликула мышей, была создана для получения модели TEN на животных. Специфичные в отношении OVA CD8+ T-клетки при адоптивном переносе в мышей K5-OVA подвергаются активации и пролиферации в дренирующих кожу лимфатических узлах и оказывают воздействие на кожу мышей K5-OVA, что приводит к развитию кожных очагов повреждения, которые напоминают TEN. Смотрите Azukizawa H., et al., Eur J Immunol: 33, 1879 (2003).
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем описании, можно использовать для улучшения клинического исхода TEN посредством ингибирования, уменьшения, предотвращения или блокирования воспаления и/или расчесывания, обусловленного заболеванием.
Буллезный пемфигоид
Буллезный пемфигоид представляет собой субэпидермальное нарушение, которое проявляется в виде субэпидермальных волдырей с инфильтратом в дерме нейтрофилов и эозинофилов. Диагноз характеризуется наличием антиген-специфичных антител против определенных белков адгезии эпидермиса и области перехода дерма-эпидерма. Смотрите Jordon R.E., et al., JAMA: 200, 751 (1967). В исследованиях по анализу роли T-клеток в патогенезе буллезного пемфигоида посредством анализа PBL и T-клеток волдырей кожи было выявлено преобладание T-клеток CLA+, экспрессирующих повышенные уровни Th2-цитокинов, таких как IL-4 и IL-13. Смотрите Terald Y., et si., J Invest Dermatol: 111, 1097 (2001). У пациентов с буллезным пемфигоидом после лечения системными кортикостероидами, встречаемость клеток CLA+, но не CLA-, продуцирующих интерлейкин-13, значительно снижается. Уменьшение количества клеток CLA+ после лечения кортикостероидами ассоциировано с клиническим улучшением. Смотрите Teraki, там же.
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем описании, можно использовать для улучшения клинического исхода буллезного пемфигоида посредством ингибирования, уменьшения, предотвращения или блокирования воспаления и/или расчесывания, обусловленных заболеванием.
Очаговая алопеция
Под очаговой алопецией (AA) понимают ограниченное тканью аутоиммунное заболевание волосяных фолликулов, в которых прекращена активность фолликула вследствие непрерывной активности лимфатических инфильтратов. AA приводит к областям полной потери волос по всему телу, хотя в действительности не происходит утраты волосяных фолликулов, даже в областях, в которых волосы отсутствуют. Несмотря на отсутствие клинических признаков воспаления, в биоптатах кожи из областей активного заболевания показано перифолликулярное лимфоцитарное воспаление, главным образом, из CD4+ клеток, совместно с внутрифолликулярным инфильтратом CD8+. Смотрите Kalish R.S. & Gilhar A. J Investig Dermatol Symp Proc: 8, 164 (2003).
В исследованиях было показано, что инфильтрирующие кожу черепа CD4+ или CD8+ лимфоциты экспрессируют CLA, и в периферической крови индивидуумов с AA процентное содержание CD4+ или CD8+ лимфоцитов CLA+ значительно выше, чем в периферической крови у здоровых контрольных доноров. Более того, было показано, что пациенты с тяжелой или прогрессирующей AA являются значительно более CLA-положительными по сравнению с пациентами, выздоравливающими от заболевания, и снижение процентного содержания клеток CLA+ соответствует хорошему клиническому течению. Смотрите Yano S., et al., Acta Derm Venereal: 82, 82 (2002). Таким образом, эти исследования подтверждают, что лимфоциты CLA+ могут играть важную роль в AA. Модели с ксенотрансплантацией показали, что активированные T-клетки, вероятно, участвуют в патогенезе AA. В поврежденной коже черепа пациентов с AA, трансплантированной мышам nude, происходило восстановление роста волос, совпадающее с утратой инфильтрирующих лимфоцитов в трансплантате, и перенос активированных T-клеток из очагов повреждения мышам SCID может передавать потерю волос эксплантатам кожи черепа человека на мышах SCID. Смотрите Kalish R.S. & Gilhar A. J Investig Dermatol Symp Proc: 8, 164 (2003).
Разнообразные иммуномодулирующие лекарственные средства составляют часть традиционного лечения этого нарушения, однако ни одно из этих лекарственных средств не оказывает постоянного эффекта. Смотрите Tang L., et al., J Invest Dermatol: 120, 400 (2003); Tang L., et al., (2004); и Tang L., et al., J Am Acad Dermatol: 49, 1013 (2003). Тем не менее, их применение в пригодных моделях на животных является инструментом для анализа молекулярных механизмов терапевтических эффектов. Смотрите Shapiro J., et al., J Investig Dermatol Symp Proc: 4, 239 (1999); Tang L., et al., Old wine in new bottles: reviving old therapies for alopecia areata using rodent models (2003); и Verma D.D., et al., Eur J Dermatol:14, 332 (2004).
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем изобретении, можно использовать для улучшения клинического исхода очаговой алопеции посредством ингибирования, уменьшения, профилактики или блокирования воспаления и/или расчесывания, обусловленных заболеванием.
Красные угри/Обыкновенные угри
Обыкновенные угри, нарушение аппарата волосяных фолликулов и сальных желез, представляют собой наиболее распространенную кожную проблему подростков. Полагают, что аномалии кератинизации фолликула вызывают угревую сыпь. Красные угри отличаются от обыкновенных наличием красных папул, пустул и пузырей и обширных телеангиэктозий, но отсутствием комедонов (белых головок). Повышенное выделение кожного сала из сальных желез является основным фактором патофизиологии обыкновенных угрей. С развитием угрей ассоциированы также другие функции сальных желез, включая провоспалительные липиды сальных желез; различные продуцируемые локально цитокины; околожелезистые пептиды и нейропептиды, такие как рилизинг-фактор кортикотропина, который продуцируется себоцитами; и вещество P, которое экспрессируется в нервных окончаниях вблизи кажущихся здоровыми железами пациентов с угревой сыпью. Смотрите Zouboulis CC. Clin Dermatol: 22, 360 (2004).
Несмотря на то что патофизиология обыкновенных угрей и красных угрей остается неизвестной, клинические наблюдения и гистопатологические исследования позволяют предположить, что воспаление волосяного фолликула и сальной железы может быть центральным звеном патогенеза красных угрей и обыкновенных угрей. Предшествующие исследования с анализом подгрупп T-клеток, инфильтрирующих очаги повреждения при красных угрях, указывают на то, что большинство T-клеток экспрессируют CD4. Смотрите Rufli T. & Buchner S.A. Demiatologica: 169, 1 (1984).
CD4+ T-клетки продуцируют IL-31, и анализ IHC кожи на экспрессию IL-31 подтверждает, что IL-31 экспрессируется в сальных и потовых железах. Стимуляция посредством IL-31 кератиноцитов эпидермиса индуцирует экспрессию хемокинов, которая, вероятно, приводит к клеточной инфильтрации, подтверждая, что IL-31 может приводить к провоспалительному ответу в коже. Смотрите Dillon S.R., et al., Nat Immunol: 5, 752 (2004). Таким образом, IL-31 может участвовать в патофизиологии красных угрей и обыкновенных угрей.
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем описании, можно использовать для улучшения клинического исхода обыкновенных угрей посредством ингибирования, уменьшения, профилактики или блокирования воспаления и/или расчесывания, обусловленных заболеванием.
Узловатая почесуха
Узловатая почесуха представляет собой высыпание лихенизированных или экскориированных узелков, вызванное некупируемым зудом, который трудно лечить. Несмотря на то что постоянное растирание приводит к лихенизации и расчесывание к продольным ссадинам, у индивидуумов, ковыряющих и выдавливающих их зудящую раздраженную кожу, имеется тенденция к появлению значительно утолщенных папул, известных как узелки пруриго. Несмотря на то что узловатая почесуха не является характерной для атопического дерматита, многие пациенты с этими узелками также имеют атопическую реакцию, которая проявляется как аллергический ринит, астма или пищевая аллергия. T-клетки представляют собой большую часть инфильтрирующих клеток в очагах при почесухе, и эти очаги повреждения часто представляют собой наиболее зудящие очаги повреждения кожи у пациентов с атопией.
Было показано, что местное лечение узловатой почесухи капсаицином, алкалоидом против зуда, который препятствует ощущению зуда и боли посредством истощения нейропептидов, таких как вещество P, в малых чувствительных нервах кожи, представляет собой эффективную и безопасную схему, приводящую к устранению очагов повреждения кожи. Смотрите Stander S., et al., J Am Acad Dermatol: 44, 471 (2001). Исследования связанного с зудом ответа у мышей NC/Nga с применением введения капсаицина показали практически полное предотвращение спонтанного развития очагов повреждения, связанных с дерматитом. Более того, происходило значительное подавление повышения сывороточных уровней IgE и количество инфильтрирующих эозинофилов и тучных клеток в поврежденной коже мышей, которым вводили капсаицин, снижалось. Смотрите Mihara K., et al., Br J Dermatol: 151, 335 (2004). Наблюдения этой группы подтвердили, что расчесывание может участвовать в развитии дерматита посредством усиления различных иммунологических ответов, подразумевая, таким образом, что предотвращение ощущения зуда и/или обусловленного зудом связанного с расчесыванием поведения может представлять собой эффективный способ лечения AD. Смотрите Mihara K., et al., Br J Dermatol: 151, 335 (2004). Таким образом, антитела против IL-31, описанные в настоящем описании, являются пригодными для минимизации эффектов AD, узловатой почесухи и других связанных с зудом заболеваний, поскольку было показано, что они снижают степень расчесывания у мышей NC/Nga.
Постоянная доставка IL-31 вызывает зуд и алопецию у мышей, с последующим развитием очагов повреждения кожи, напоминающих дерматит, подтверждая, что IL-31 может вызывать зуд. Смотрите Dillon S.R., et al., Nat Immunol: 5, 752 (2004). Участие IL-31 в индукции связанного с зудом ответа тестировали двумя способами (i) посредством введения капсаицина мышам, которым вводили IL-31, и (ii) посредством введения IL-31 мышам с нокаутом Tac1, у которых значительно снижен ноцицептивный болевой ответ вследствие отсутствия экспрессии нейропептидов, в примере 52. Кроме того, возможность посредством нейтрализации IL-31 у мышей, которым вводили IL-31, предотвращать зуд и алопецию тестировали в примере 12.
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем описании, можно использовать для улучшения клинического исхода узловатой почесухи посредством ингибирования, уменьшения, профилактики или блокирования воспаления и/или расчесывания, обусловленных заболеванием.
Тропные к коже вирусы и ассоциированный с вирусом зуд
Специфичные к вирусу простого герпеса (HSV) CD8+ T-клетки в периферической крови и HSV-специфичные CD8+ T-клетки, выделенные из герпетических очагов повреждения, экспрессируют высокие уровни CLA, а в нетропных к коже ассоциированных с вирусом простого герпеса CD8+ T-клетках экспрессия CLA отсутствует. Смотрите Koelle D.M., et al., J Clin Invest: 110, 537 (2002). Реакционно-способные в отношении HSV-2 CD4+ T-лимфоциты также экспрессируют CLA, однако на уровнях, которые ниже ранее наблюдаемых уровней дл CD8+ T-лимфоцитов. Смотрите Gonzalez J.C., et al., J Infect Dis: 191, 243 (2005). Зуд также часто ассоциирован с инфекциями вирусом герпеса (Смотрите Hung K. Y., et al., Blood Purif: 16, 147 (1998), хотя другие вирусные заболевания, такие как ВИЧ, также ассоциированы с зудящими очагами повреждения кожи. Тяжелый некупируемый зуд, часто ассоциированный с эритематозно-папулярными очагами повреждения кожи и гиперэозинофилией, представляет собой состояние, наблюдаемое у некоторых инфицированных ВИЧ пациентов без атопии 36. Смотрите Singh F. & Rudikoff D, Am J Clin Dermatol; 4, 177 (2003); и Milazzo R, Piconi S., et al., Allergy: 54, 266 (1999).
Ассоциация тропных к коже вирусов с зудом и T-клетками CLA+ позволяет предположить, что продуцирующие IL-31 T-клетки могут участвовать в патофизиологии вирусных инфекций.
Таким образом, нейтрализацию IL-31 посредством антител, описанных в настоящем описании, можно использовать для улучшения клинического исхода зуда, ассоциированного с тропными к коже вирусами, посредством ингибирования, уменьшения, профилактики или блокирования воспаления и/или расчесывания, обусловленных заболеванием.
Более того, воспаление представляет собой защитный ответ организма для борьбы с вторгшимся агентом. Воспаление представляет собой каскадный процесс, в который вовлечено множество клеточных и гуморальных медиаторов. С одной стороны, подавление воспалительного ответа может привести к ослаблению иммунитета у хозяина; однако, если его не контролировать, воспаление может привести к тяжелым осложнениям, включая хронические воспалительные заболевания (например, ревматоидный артрит, рассеянный склероз, воспалительное заболевание кишечника и т.п.), септический шок и полиорганную недостаточность. Важно, что эти различные болезненные состояния имеют общие медиаторы. Совокупность заболеваний, которые характеризуются воспалением, оказывает значительное влияние на болезненность и смертность человека. Таким образом, очевидно, что противовоспалительные антитела и связывающие полипептиды, такие как антитела против IL-31 и связывающие полипептиды, описанные в настоящем изобретении, могут обладать значительным терапевтическим потенциалом для широкого класса заболеваний человека и животных, от астмы и аллергии до аутоиммунных заболеваний и септического шока. По существу, применение противовоспалительных антител против IL-31 и связывающих полипептидов, описанных в настоящем описании, можно применять терапевтически в качестве антагонистов IL-31, описанных в настоящем описании, в частности, при заболеваниях, таких как артрит, эндотоксемия, воспалительное заболевание кишечника, псориаз, связанные с ними заболевания и т.п.
1. Артрит
Артрит, включая остеоартрит, ревматоидный артрит, артритические суставы как результат повреждения, и т.п., представляет собой распространенное воспалительное состояние, на которое может оказать положительное воздействие терапевтическое применение противовоспалительных антител и связывающих полипептидов, таких как антитела против IL-31 и связывающие полипептиды согласно изобретению. Например, ревматоидный артрит (RA) представляет собой системное заболевание, которое поражает весь организм и является одной из наиболее распространенных форм артрита. Он характеризуется воспалением покрывающей сустав оболочки, которое вызывает боль, неподвижность, повышение температуры, покраснение и припухлость. Воспалительные клетки высвобождают ферменты, которые могут расщеплять кость и хрящ. В результате ревматоидного артрита покрывающая сустав оболочка, синовиальная мембрана, может поражать и повреждать кость и хрящ, что, помимо прочих физиологических эффектов, приводит к повреждению сустава и тяжелой боли. Пораженный сустав может утрачивать свою форму и положение, вызывая боль и утрату подвижности.
Ревматоидный артрит (RA) представляет собой иммуноопосредуемое заболевание, в частности, характеризующееся воспалением и последующим повреждением ткани, приводящим к тяжелой нетрудоспособности и повышенной смертности. В суставах при ревматоидном артрите локально продуцируется множество цитокинов. В ряде исследований было показано, что IL-1 и TNF-альфа, два прототипных провоспалительных цитокина, играют важную роль в механизмах, вовлеченных в воспаление синовиальной оболочки, и в прогрессирующем разрушении сустава. Действительно, введение ингибиторов TNF-альфа и IL-1 у пациентов с RA приводит к значительному улучшению клинических и биологических признаков воспаления и к снижению рентгенологических признаков изъязвления кости и разрушения хряща. Однако, несмотря на эти обнадеживающие результаты, значительная процентная доля пациентов не отвечает на эти средства, подтверждая, что в патофизиологию артрита также вовлечены другие медиаторы (Gabay, Expert. Opin. Biol. Ther. 2(2): 135-149, 2002). Одним из этих медиаторов может быть IL-31, и, по существу, молекула, которая связывает или ингибирует IL-31, такая как антитела против IL-31 или связывающие партнеры, может выступать в качестве пригодного терапевтического средства для снижения воспаления при ревматоидном артрите и других артритических заболеваниях.
В данной области известно несколько моделей ревматоидного артрита на животных. Например, в модели индуцируемого коллагеном артрита (CIA) у мышей развивается хронический воспалительный артрит, который является близко сходным с ревматоидным артритом человека. Поскольку CIA обладает общими иммунологическими и патологическими признаками с RA, это привело к тому, что он представляет собой идеальную модель для скрининга потенциальных противовоспалительных соединений человека. Модель CIA представляет собой хорошо известную модель на мышах, которая зависит как от иммунного ответа, так и от воспалительного ответа, возникающих в этом порядке. Иммунный ответ включает в себя взаимодействие B-клеток и CD4+ T-клеток в ответ на коллаген, который вводят в качестве антигена, и приводит к продукции антител против коллагена. Воспалительная фаза является результатом тканевых ответов посредством медиаторов воспаления, являющихся следствием того, что некоторые из этих антител перекрестно реагируют с собственным коллагеном мыши и активируют каскад комплемента. Преимущество использования модели CIA состоит в том, что основные механизмы его патогенеза известны. Соответствующие эпитопы для T-клеток и B-клеток на коллагене II типа идентифицированы, и определены различные иммунологические (например, гиперчувствительность замедленного типа и антитело против коллагена) и воспалительные (например, цитокины, хемокины и деградирующие матрикс ферменты) параметры, связанные с иммуноопосредуемым артритом, и, таким образом, их можно использовать для оценки эффективности тестируемого соединения в модели CIA (Wooley, Curr. Opin. Rheum. 3:407-20, 1999; Williams et al, Immunol. 89:9784-788, 1992; Myers et al., Life Sci. 61:1861-78, 1997; и Wang et al., Immunol. 92:8955-959, 1995).
Введение растворимого IL-31RA, содержащего полипептиды (включая гетеродимерные и мультимерные рецепторы, описанные в настоящем описании), такие как IL-31RA-Fc4, или других растворимых и слитых белков IL-31RA мышам в этой модели CIA использовали для оценки применения IL-31RA для смягчения симптомов и изменения течения заболевания. Поскольку молекула, которая модулирует иммунный и воспалительный ответ, IL-31, может индуцировать продукцию SAA, который вовлечен в патогенез ревматоидного артрита, антагонисты IL-31 могут снижать активность SAA in vitro и in vivo, и системное или местное введение антагонистов IL-31, таких как антитела против IL-31 или связывающие партнеры, IL-31RA, содержащий полипептиды (включая гетеродимерные и мультимерные рецепторы, описанные в настоящем описании), такие как IL-31RA-Fc4, или другие растворимые и слитые белки IL-31RA, потенциально может подавлять воспалительный ответ при RA. Другие потенциальные лекарственные средства включают в себя полипептиды IL-31RA, растворимые гетеродимерные и полимерные полипептиды рецептора, или антитела против IL-31 или связывающие партнеры согласно изобретению, и т.п.
2. Эндотоксемия
Эндотоксемия представляет собой тяжелое состояние, обычно возникающее вследствие инфекционных агентов, таких как бактерии и другие агенты инфекционных заболеваний, сепсиса, синдрома токсического шока, или у пациентов с ослабленным иммунитетом, подвергшихся оппортунистическим инфекциям, и т.п. Терапевтически пригодные противовоспалительные антитела и связывающие полипептиды, такие как антитела против IL-31 и связывающие полипептиды согласно изобретению, могут облегчать профилактику и лечение эндотоксемии у человека и животных. Другие потенциальные лекарственные средства, включающие в себя полипептиды IL-31RA, растворимые гетеродимерные и мультимерные полипептиды рецептора, или антитела против IL-31 или связывающие партнеры согласно изобретению, и т.п., могут выступать в качестве пригодного лекарственного средства для снижения воспаления и патологических эффектов эндотоксемии.
В индуцированную липополисахаридом (LPS) эндотоксемию вовлечено множество провоспалительных медиаторов, которые оказывают патологические эффекты при инфекционных заболеваниях, и индуцированная LPS эндотоксемия у грызунов представляет собой широко используемую и приемлемую модель для изучения фармакологических эффектов потенциальных провоспалительных или иммуномодулирующих средств. LPS, продуцируемый в грамотрицательных бактериях, представляет собой основной этиологический фактор в патогенезе септического шока (Glausner et al., Lancet 338:732, 1991). Подобное шоку состояние действительно можно индуцировать экспериментально посредством однократной инъекции LPS животным. Молекулы, продуцируемые клетками, отвечающими на LPS, могут оказывать воздействие на патогены прямо или косвенно. Хотя эти биологические ответы защищают хозяина от вторгшихся патогенов, они также могут приносить вред. Таким образом, массивная стимуляция врожденного иммунитета, происходящая вследствие тяжелой инфекции грамотрицательными бактериями, приводит к избыточной продукции цитокинов и других молекул и к развитию смертельного синдрома, синдрома токсического шока, который характеризуется лихорадкой, гипотензией, диссеминированным внутрисосудистым свертыванием крови и полиорганной недостаточностью (Dumitru et al. Cell 103:1071-1083, 2000).
Эти токсические эффекты LPS, главным образом, связаны с активацией макрофагов, приводящей к высвобождению множества медиаторов воспаления. Среди этих медиаторов, TNF, по-видимому, играет ключевую роль, как показано посредством предотвращения связанной с LPS токсичности введением нейтрализующих антител против TNF (Beutler et al., Science 229:869, 1985). Определено, что инъекция 1 мкг LPS E. coli в мышь C57B1/6 приводит к значительному повышению циркулирующих IL-6, TNF-альфа, IL-1 и белков острой фазы (например, SAA) приблизительно через 2 часа после инъекции. Токсичность LPS, по-видимому, опосредуется этими цитокинами, поскольку пассивная иммунизация против этих медиаторов может приводить к снижению смертности (Beutler et al., Science 229:869, 1985). Потенциальные стратегии с вмешательством в иммунную систему для профилактики и/или лечения септического шока включают в себя mAb против TNF, антагонист рецептора для IL-1, LIF, IL-10 и G-CSF. Поскольку LPS индуцирует продукцию провоспалительных факторов, вероятно, приводящих к патологии эндотоксемии, нейтрализацию активности IL-31, SAA или других провоспалительных факторов посредством антагонистического воздействия на полипептид IL-31 можно использовать для уменьшения симптомов эндотоксемии, таких как выявляют при эндотоксиновом шоке. Другие потенциальные лекарственные средства включают в себя полипептиды IL-31RA, растворимые гетеродимерные и мультимерные полипептиды рецептора, или антитела против IL-31 или связывающие партнеры согласно изобретению, и т.п.
3. Воспалительное заболевание кишечника. IBD
В США приблизительно 500000 человек страдают воспалительным заболеванием кишечника (IBD), которое может поражать либо толстую и прямую кишку (язвенный колит), либо и тонкий и толстый кишечник (болезнь Крона). Патогенез этих заболеваний остается неясным, однако в них вовлечено хроническое воспаление пораженных тканей. Потенциальные лекарственные средства включают в себя полипептиды IL-31RA, растворимые гетеродимерные и мультимерные полипептиды рецептора, или антитела против IL-31 или связывающие партнеры согласно изобретению, и т.п., которые могут служить в качестве лекарственного средства для снижения воспаления и патологических эффектов при IBD и связанных с ним заболеваний.
Язвенный колит (UC) представляет собой воспалительное заболевание толстого кишечника, обычно называемого ободочной кишкой, характеризующееся воспалением и изъязвлением слизистой или наиболее глубокой оболочки кишечника. Это воспаление вызывает частое опорожнение толстого кишечника, что приводит к диарее. Симптомы включают в себя ослабленный стул и связанные с ним аномальные спазмы, жар и снижение массы тела. Несмотря на то что точная причина UC остается неизвестной, недавние исследования позволили предположить, что естественная защита организма действует против белков организма, которые организм воспринимает как чужеродные ("аутоиммунная реакция"). Возможно вследствие того, что они являются сходными с бактериальными белками кишечника, эти белки могут либо инициировать, либо стимулировать воспалительный процесс, который начинает разрушать выстилку кишечника. По мере разрушения выстилки формируются язвы с выделением слизи, гноя и крови. Заболевание, как правило, начинается в области прямой кишки и может в конечном итоге распространиться по всему толстому кишечнику. Повторяющиеся эпизоды воспаления приводят к утолщению стенки кишечника и прямой кишки с рубцовой тканью. В случае тяжелого заболевания может происходить гибель ткани толстого кишечника или сепсис. Симптомы язвенного колита варьируют по тяжести, и их начало может быть постепенным или внезапным. Приступы могут провоцировать многие факторы, включая инфекции дыхательных путей или стресс.
Несмотря на то что в настоящее время не существует доступного лечения UC, способы лечения сосредоточены на подавлении аномального воспалительного процесса в выстилке кишечника. Для лечения заболевания доступны лекарственные средства, включающие в себя кортикостероиды, иммунодепрессивные средства (например, азатиоприн, меркаптопурин и метотрексат) и аминосалицилаты. Однако длительное применение иммунодепрессантов, таких как кортикостероиды и азатиоприн, может приводить к тяжелым побочным эффектам, включая истончение костей, катаракту, инфекцию и эффекты на печень и костный мозг. У пациентов, у которых современные лекарственные средства оказываются безуспешными, возможно проведение хирургической операции. Хирургическая операция включает в себя удаление всего толстого кишечника и прямой кишки.
Существует несколько моделей на животных, которые могут практически имитировать хронический язвенный колит. Наиболее широко используемой моделью является модель колита, индуцируемого 2,4,6-тринитробензолсульфоновой кислотой/этанолом (TNBS), которые индуцируют хроническое воспаление и изъязвление в толстом кишечнике. При введении TNBS в толстый кишечник чувствительным мышам посредством интраректальной инстилляции происходит индукция опосредуемого T-клетками иммунного ответа в слизистой оболочке толстого кишечника, приводя, в этом случае, к массивному воспалению слизистой оболочки, характеризующемуся плотной инфильтрацией T-клеток и макрофагов через всю стенку толстого кишечника. Более того, эта гистопатологическая картина сопровождается клинической картиной прогрессирующего снижения массы тела (исхудания), геморрагической диареей, выпадением прямой кишки и утолщением стенки толстого кишечника (Neurath et al. Intern. Rev. Immunol. 19:51-62, 2000).
В другой модели колита используют сульфат декстрана натрия (DSS), который индуцирует острый колит, проявляющийся геморрагической диареей, снижением массы тела, укорочением толстого кишечника и изъязвлением слизистой оболочки с инфильтрацией нейтрофилов. Индуцированный посредством DSS колит гистологически характеризуется инфильтрацией воспалительных клеток в собственную пластинку с гиперплазией лимфоидной ткани, фокальным разрушением крипт и изъязвлением эпителия. Полагают, что эти изменения развиваются вследствие токсического эффекта DSS на эпителий, и фагоцитоза клеток собственной пластинки и продукции TNF-альфа и IFN-гамма. Несмотря на широкое применение, несколько вопросов, касающихся соответствию механизмов DSS заболеванию у человека, остаются нерешенными. DSS рассматривают как независимую от T-клеток модель, вследствие того, что она наблюдается у дефицитных по T-клеткам животных, таких как мыши SCID.
Введение антител против IL-31 или связывающих партнеров, растворимых IL-31RA, содержащих полипептиды (включая гетеродимерные и мультимерные рецепторы), такие как IL-31RA-Fc4, или других растворимых и слитых белков IL-31RA в этих моделях с TNBS или DSS можно использовать для оценки применения антагонистов IL-31 для смягчения симптомов и изменения течения желудочно-кишечного заболевания. IL-31 может участвовать в воспалительном ответе при колите, и нейтрализация активности IL-31 посредством введения антагонистов IL-31 представляет собой потенциальный терапевтический подход при IBD. Другие потенциальные лекарственные средства включают в себя полипептиды IL-31RA, растворимые гетеродимерные и мультимерные полипептиды рецептора, или антитела против IL-31 или связывающие партнеры согласно изобретению, и т.п.
4. Псориаз
Псориаз представляет собой хроническое кожное состояние, которым страдают семь миллионов американцев. Псориаз происходит при аномальном росте клеток кожи, что приводит к воспаленным, припухлым и чешуеобразным участкам кожи, где старая кожа недостаточно быстро отшелушивается. Бляшковидный псориаз, наиболее распространенная форма, характеризуется воспаленными участками кожи ("очагами повреждения"), покрытыми серебристо-белыми чешуйками. Псориаз может ограничиваться несколькими бляшками или вовлекать от умеренной до обширной области кожи, чаще всего появляясь на коже черепа, колен, локтей и туловища. Несмотря на то что он является хорошо заметным, псориаз не является контагиозным заболеванием. Патогенез заболевания включает в себя хроническое воспаление пораженных тканей. Полипептиды IL-31RA, растворимые гетеродимерные и мультимерные полипептиды рецептора, или антитела против IL-31 или связывающие партнеры согласно изобретению, и т.п., могут служить в качестве пригодного лекарственного средства для снижения воспаления и патологических эффектов при псориазе, других воспалительных заболеваниях кожи, аллергии кожи и слизистой оболочки, и связанных с ними заболеваниях.
Псориаз представляет собой опосредуемое T-клетками воспалительное нарушение, которое может вызывать значительный дискомфорт. Он представляет собой заболевание, от которого не существует лечения и которое поражает людей всех возрастов. Псориазом страдают приблизительно два процента населения Европы и Северной Америки. Несмотря на то что пациенты с мягким течением псориаза часто могут осуществлять контроль их заболевания посредством веществ для местного применения, более чем одному миллиону пациентов по всему миру необходимо лечение ультрафиолетом и системными иммунодепрессантами. К сожалению, неудобство и риск облучения ультрафиолетом и токсичность многих лекарственных средств ограничивают их длительное применение. Более того, у пациентов, как правило, происходит рецидив псориаза, и в некоторых случаях
IL-31 был выделен из ткани, о которой известно, что она обладает важной иммунологической функцией и которая содержит клетки, которые играют роль в иммунной системе. IL-31 экспрессируется в прошедших селекцию по CD3+ активированных клетках периферической крови, и было показано, что экспрессия IL-31 возрастает после активации T-клеток. Более того, результаты экспериментов, описанных в разделе примеров настоящего описания, подтверждают, что полипептиды согласно изобретению могут оказывать эффект на рост/размножение моноцитов/макрофагов, T-клеток, B-клеток, NK-клеток и/или на уровень дифференцировки моноцитов/макрофагов, T-клеток, B-клеток, NK-клеток или предшественников этих клеток. Факторы, которые как стимулируют пролиферацию гемопоэтических предшественников, так и активируют зрелые клетки, главным образом, известны, однако для пролиферации и активации также могут быть необходимы дополнительные факторы роста. Например, было показано, что IL-7 и фактор Стила (лиганд c-kit) необходимы для образования колоний предшественников NK-клеток. IL-15 + IL-2 в сочетании с IL-7 и фактором Стила были наиболее эффективными (Mrozek et al., Blood 87:2632-2640, 1996). Однако для пролиферации определенных подтипов NK-клеток и/или предшественников NK-клеток могут быть необходимы неидентифицированные цитокины (Robertson et. al., Blood 76:2451-2438, 1990). Аналогично, IL-31 может действовать отдельно, или совместно, или синергично с другими цитокинами, усиливая рост, пролиферативную экспансию и модификацию дифференцировки моноцитов/макрофагов, T-клеток, B-клеток или NK-клеток.
Настоящее изобретение относится к способу ингибирования активации или дифференцировки моноцитов/макрофагов. Моноциты представляют собой клетки с незавершенной дифференцировкой, которые мигрируют в ткани, где они созревают и превращаются в макрофаги. Макрофаги играют центральную роль в иммунном ответе, представляя антиген лимфоцитам, и играют вспомогательную роль в качестве вспомогательных клеток для лимфоцитов, секретируя ряд цитокинов. Макрофаги могут интернализовывать внеклеточные молекулы и при активации обладают повышенной способностью к уничтожению внутриклеточных микроорганизмов и клеток опухоли. Активированные макрофаги также вовлечены в стимуляцию острого или местного воспаления.
Тканевое распределение рецепторов для данного цитокина является строгим показателем потенциальных областей воздействия этого цитокина. Экспрессию IL-31RA наблюдали в моноцитах и B-клетках, со значительным повышением экспрессии при активации CD3+, CD4+ и CD8+ T-клеток. Кроме того, также в отношении экспрессии IL-31RA были положительными две моноцитарные клеточные линии, THP-1 (Tsuchiya et al., Int. J. Cancer 26:171-176, 1980) и U937 (Sundstrom et al., Int. J. Cancer 17:565-577, 1976).
Описано, что экспрессия OSMR является очень широкой (Mosley et al, JBC 271:32635-32643, 1996). Это распределение IL-31RA и рецепторов для OSM подтверждает роль IL-31 в иммунных ответах, особенно в экспансии T-клеток при активации, или роль в моноцитарном/макрофагальном звене иммунной системы.
Таким образом, конкретные варианты осуществления настоящего изобретения направлены на применение растворимых гетеродимеров IL-31RA/OSMR в качестве антагонистов при воспалительных и иммунных заболеваниях или состояниях, таких как панкреатит, диабет I типа (IDDM), рак поджелудочной железы, панкреатит, болезнь Грэйвса, воспалительное заболевание кишечника (IBD), болезнь Крона, злокачественная опухоль толстого кишечника и интестинальная злокачественная опухоль, дивертикулез, аутоиммунное заболевание, сепсис, трансплантация органа или костного мозга; воспаление вследствие травмы, хирургической операции или инфекции; амилоидоз; спленомегалия; реакция трансплантат против хозяина; и при ингибировании воспаления, подавлении иммунной системы, снижении пролиферации гемопэтических, иммунных, воспалительных или лимфоидных клеток, макрофагов, T-клеток (включая Th1 и Th2-клетки, CD4+ и CD8+ клетки), подавлении иммунного ответа на патоген или антиген. Более того, наличие экспрессии IL-31RA в активированных клетках иммунной системы, таких как активированные CD4+ и CD19+ клетки, показывают, что рецептор IL-31RA может быть вовлечен в иммунные защитные реакции организма против чужеродных агентов: таких как микроорганизмы и клеточный дебрис, и может участвовать в иммунных ответах в процессе воспаления и образования злокачественной опухоли. По существу, антитела и связывающие партнеры согласно изобретению, которые являются агонистами или антагонистами функции рецептора IL-31RA, такие как IL-31, можно использовать для модификации иммунного ответа и воспаления.
IL-31 может быть пригодным для подавления иммунной системы, как, например, лечение аутоиммунных заболеваний, включая ревматоидный артрит, рассеянный склероз, сахарный диабет, воспалительное заболевание кишечника, болезнь Крона и т.д. Подавление иммунной системы также можно использовать для уменьшения отторжения трансплантированных или пересаженных тканей или органов и для лечения T-клеточных, B-клеточных или специфичных в отношении моноцитов лейкозов или лимфом и других злокачественных опухолей, посредством ингибирования пролиферации поврежденного типа клеток. Более того, IL-31 можно применять для выявления моноцитов, макрофагов и активированных T-клеток и облегчения диагностики такого аутоиммунного заболевания, в частности, при болезненном состоянии, где происходит повышение количества или активация моноцитов.
Полипептиды, пептиды, антитела против IL-31 и т.п. также можно применять в диагностических системах для определения циркулирующих уровней IL-31. В сходном варианте осуществления антитела или другие вещества, которые специфически связываются с полипептидами IL-31, можно применять для выявления циркулирующих полипептидов IL-31. Повышенные или сниженные уровни полипептидов-лигандов могут указывать на патологические состояния, включая злокачественную опухоль. Полипептиды IL-31 могут участвовать в патологических процессах и могут быть косвенными маркерами связанного с ними заболевания.
Более того, специалист в данной области поймет, что являются пригодными антагонисты IL-31. Например, в атеросклеротических бляшках одной из первых аномалий является локализация моноцитов/макрофагов среди клеток эндотелия. Эти очаги повреждения можно предотвращать посредством применения антагонистов IL-31. Антитела против IL-31 (например, нейтрализующее антитело против IL-31), растворимые рецепторы IL-31RA, гетеродимеры и мультимеры, и связывающие IL-31 партнеры также можно использовать в качестве антагонистов IL-31. Более того, монобластный лейкоз ассоциирован с различными клиническими аномалиями, которые отражают высвобождение биологических продуктов макрофагов, их примеры включают в себя высокие уровни лизозима в сыворотке и моче и высокую лихорадку. Более того, при таких лейкозах происходит аномальное повышение моноцитарных клеток. Эти эффекты, вероятно, можно предотвратить посредством антагонистов IL-31, таких как описаны в настоящем описании. Более того, антитела против IL-31 можно конъюгировать с молекулами, такими как токсические группы и цитокины, как описано в настоящем описании, для нацеливания на уничтожение лейкозных моноцитарных клеток.
Поскольку характер экспрессии IL-31 является специфичным в отношении T-клеток, макрофагов и моноцитов, и эти заболевания включают в себя аномалии моноцитарных клеток, такие как пролиферация, функционирование, локализация и активация, полинуклеотиды, полипептиды и антитела согласно изобретению можно использовать в качестве диагностических средств для выявления таких аномалий моноцитарных клеток, и показателя наличия заболеваний. Такие способы включают в себя забор биологического образца пациента, такого как кровь, слюна или биоптат, и сравнение их с нормальным контрольным образцом. Для определения относительных уровней или локализации IL-31 или клеток, экспрессирующих IL-31, т.е. моноцитов, в образце пациента по сравнению с нормальным контролем можно использовать гистологические, цитологические способы, способы проточной цитометрии, биохимические и другие способы. Изменение уровня (повышение или снижение) экспрессии IL-31, или изменение количества или локализации моноцитов (например, повышение количества или инфильтрация моноцитарных клеток в тканях, где в норме они не присутствуют) по сравнению с контролем, является показателем наличия заболевания. Такие диагностические способы также могут включать в себя применение радиометрических, флуоресцентных и колориметрических меток, присоединенных к полинуклеотидам, полипептидам или антителам согласно изобретению. Такие способы хорошо известны в данной области и описаны в настоящем описании.
Было показано, что IL-31 экспрессируется активированными мононуклеарными клетками, и он может быть вовлечен в регуляцию воспаления. По существу, полипептиды согласно изобретению можно оценивать и применять, исходя из их способности модифицировать воспаление, или их можно использовать в качестве маркера воспаления. Способы определения провоспалительных и противовоспалительных свойств IL-31 известны в данной области и описаны в настоящем описании. Более того, он может быть вовлечен в положительную регуляцию продукции реактантов острой фазы, таких как сывороточный амилоид A (SAA), α1-антихимотрипсин и гаптоглобин, и эта экспрессия лиганда рецептора IL-31RA может возрастать при инъекции липополисахарида (LPS) in vivo, которые вовлечены в воспалительный ответ (Dumoutier, L. et al., Proc. Nat'l. Acad. Sci. 97:10144-10149, 2000). Полагают, что продукция белков острой фазы, таких как SAA, является кратковременным механизмом выживания, где воспаление оказывает положительное воздействие; однако, поддержание белков острой фазы в течение более длительных периодов приводит к хроническому воспалению и может быть опасным для здоровья человека. Для обзора смотрите Uhlar, CM and Whitehead, AS, Eur. J. Biochem. 265:501-523, 1999, и Baumann H. and Gauldie, J. Immunology Today 15: 74-80, 1994. Более того, белок острой фазы SAA вовлечен в патогенез нескольких хронических воспалительных заболеваний, он вовлечен в атеросклероз и ревматоидный артрит и является предшественником белка амилоида A, депонируемого при амилоидозе (Uhlar, CM and Whitehead, выше). Таким образом, в случае, когда лиганд, такой как IL-31, выступает в качестве провоспалительной молекулы и индуцирует продукцию SAA, антагонисты могут быть пригодны для лечения воспалительного заболевания и другого заболевания, ассоциированного с ответом белков острой фазы, индуцируемым лигандом. Настоящее изобретение относится к таким антагонистам. Например, способ уменьшения воспаления включает в себя введение млекопитающему с воспалением количества композиции IL-31, или антитела против IL-31 (например, нейтрализующего антитела), которое является достаточным для уменьшения воспаления. Более того, способ подавления воспалительного ответа у млекопитающего с воспалением может включать в себя: (1) определение уровня сывороточного белка амилоида A; (2) введение композиции, содержащей полипептид IL-31 или антитело против IL-31, как описано в настоящем описании, в приемлемом фармацевтическом носителе; (3) определения уровня сывороточного белка амилоида A после введения; (4) сравнения уровня сывороточного белка амилоида A стадии (1) с уровнем сывороточного белка амилоида A стадии (3), где отсутствие повышения или снижения уровня сывороточного белка амилоида A указывает на подавление воспалительного ответа.
Подобно IL-31, анализ тканевого распределения мРНК, соответствующей кДНК его рецептора IL-31RA, показал, что уровень мРНК был наивысшим в моноцитах и клетках простаты и повышен в активированных моноцитах и активированных CD4+, активированных CD8+ и активированных CD3+ клетках. Таким образом, рецептор IL-31RA также вовлечен в индукцию воспалительного и иммунного ответа. Таким образом, конкретные варианты осуществления настоящего изобретения относятся к применению антител против IL-31, и IL-31, а также растворимых гетеродимеров рецептора IL-31RA, в качестве антагонистов при воспалительных и иммунных заболеваниях или состояниях, таких как панкреатит, диабет I типа (IDDM), рак поджелудочной железы, панкреатит, болезнь Грэйвса, воспалительное заболевание кишечника (IBD), болезнь Крона, рак толстого кишечника и интестинальный рак, дивертикулез, аутоиммунное заболевание, сепсис, трансплантация органа или костного мозга; воспаление вследствие травмы, хирургической операции или инфекции; амилоидоз; спленомегалия; реакция трансплантат против хозяина; и при ингибировании воспаления, подавлении иммунной системы, снижении пролиферации гемопоэтических, иммунных, воспалительных или лимфоидных клеток, макрофагов, T-клеток (включая клетки Th1 и Th2, CD4+ и CD8+ клетки), подавлении иммунного ответа на патоген или антиген. Более того, наличие рецептора IL-31RA и экспрессия IL-31 в активированных клетках иммунной системы, таких как активированные CD3+, моноциты, CD4+ и CD19+ клетки, показывают, что рецептор IL-31RA может быть вовлечен в иммунные защитные реакции организма против чужеродных агентов: таких как микроорганизмы и клеточный дебрис, и может участвовать в иммунных ответах в процессе воспаления и образования злокачественной опухоли. По существу, IL-31 и антитела против IL-31 согласно изобретению, которые являются агонистами или антагонистами функции рецептора IL-31RA, можно использовать для модификации иммунного ответа и воспаления.
Полипептиды IL-31, которые связывают полипептиды рецептора IL-31RA, и антитела к ним являются пригодными для:
1) Антагонизма или блокирования передачи сигнала через содержащие IL-31RA рецепторы при лечении острого воспаления, воспаления в результате травмы, повреждения тканей, хирургической операции, сепсиса или инфекции, и хронических воспалительных заболеваний, таких как астма, воспалительное заболевание кишечника (IBD), хронический колит, спленомегалия, ревматоидный артрит, рецидивирующие острые воспалительные эпизоды (например, туберкулез), и при лечении амилоидоза и атеросклероза, болезни Кастлмана, астмы и других заболеваний, связанных с индукцией ответа острой фазы.
2) Антагонизма или блокирования передачи сигнала через рецепторы IL-31RA при лечении аутоиммунных заболеваний, таких как IDDM, рассеянный склероз (MS), системная красная волчанка (SLE), миастения, ревматоидный артрит и IBD, для предотвращения или ингибирования передачи сигнала в иммунных клетках (например, лимфоцитах, моноцитах, лейкоцитах) через рецептор IL-31RA (Hughes C et al., J. Immunol 153: 3319-3325, 1994). Альтернативно антитела, такие как моноклональные антитела (MAb) против IL-31, также можно использовать в качестве антагониста для уменьшения количества нежелательных иммунных клеток для лечения аутоиммунного заболевания. Астму, аллергию и другие заболевания можно лечить посредством MAb против, например, антител против IL-31, растворимых рецепторов IL-3IRA или гетеродимеров IL-31RA/CRF2-4, для ингибирования иммунного ответа или снижения количества поврежденных клеток. Блокирование или ингибирование передачи сигнала через IL-31RA с использованием полипептидов и антител согласно изобретению, также может быть пригодным в случае заболеваний поджелудочной железы, почек, гипофиза и нейрональных клеток. Положительный эффект можно быть оказан на IDDM, NIDDM, панкреатит и карциному поджелудочной железы. IL-31RA может выступать в качестве мишени для лечения посредством MAb злокачественной опухоли, где антагонистическое MAb ингибирует рост злокачественной опухоли и направляет опосредуемое иммунной системой уничтожение клеток (Holliger P, и Hoogenboom, H: Nature Biotech. 16: 1015-1016, 1998). Mab против растворимых мономеров, гомодимеров, гетеродимеров и мультимеров рецептора IL-31RA также могут быть пригодными для лечения нефропатий, таких как гломерулосклероз, мембранозная нефропатия, амилоидоз (который, помимо других тканей, также поражает почки), почечный артериосклероз, гломерулонефрит различного происхождения, фибропролиферативные заболевания почки, а также дисфункция почки, ассоциированная с SLE, IDDM, диабетом II типа (NIDDM), опухолями почек и другими заболеваниями.
3) Антагонизма или запуска передачи сигала через рецепторы IL-31RA при лечении аутоиммунных заболеваний, таких как IDDM, MS, SLE, миастения, ревматоидный артрит и IBD. IL-31 может передавать сигнал лимфоцитам или другим иммунным клеткам на дифференцировку, изменение пролиферации, или смену продукции цитокинов или белков клеточной поверхности, которые снижают аутоиммунитет. Конкретно, модулирование ответа T-хелперных клеток на измененный характер секреции цитокинов может отклонять аутоиммунный ответ в сторону смягчения заболевания (Smith JA et al., J. Immunol. 160:4841-4849, 1998). Аналогично, IL-31 можно использовать для передачи сигнала, уменьшения количества и отклонения иммунных клеток, вовлеченных в астму, аллергию и атопическое заболевание. Передача сигнала через рецептор IL-31RA также может оказывать положительный эффект при заболеваниях поджелудочной железы, почки, гипофиза и нейрональных клеток. Положительный эффект можно быть оказан на IDDM, NIDDM, панкреатит и карциному поджелудочной железы. IL-31RA может выступать в качестве мишени для терапии посредством MAb рака поджелудочной железы, где передача сигнала посредством MAb ингибирует рост злокачественной опухоли и нацеливает на опосредуемое иммунной системой уничтожение клеток (Tutt, AL et al., J Immunol. 161: 3175-3185, 1998). Аналогично специфичные в отношении T-клеток лейкозы, лимфомы, дискразию плазматических клеток (например, множественную миелому) и карциному можно лечить посредством моноклональных антител (например, нейтрализующего антитела) против содержащих IL-31RA растворимых рецепторов согласно изобретению.
Антитела против IL-31, растворимые мономерные, гомодимерные, гетеродимерные и мультимерные полипептиды рецептора IL-31RA, описанные в настоящем описании, можно использовать для нейтрализации/блокирования активности лиганда рецептора IL-31RA при лечении аутоиммунного заболевания, атопического заболевания, NIDDM, панкреатита и дисфункции почек, как описано выше.
Антитела против IL-31 и растворимые содержащие IL-31RA рецепторы являются пригодными в качестве антагонистов IL-31. Такие антагонистические эффекты могут быть достигнуты посредством непосредственной нейтрализации или связывания их природного лиганда. В дополнение к применению в качестве антагонистов, растворимые рецепторы могут связывать IL-31 и выступать в качестве носителя или белков-переносчиков, в целях транспорта IL-31 в различные ткани, органы и клетки в организме. По существу, растворимые рецепторы могут быть слитыми или связанными с молекулами, полипептидами или химическими группами, которые направляют комплекс растворимый рецептор-лиганд в конкретную область, такую как ткань, конкретная иммунная клетка, моноциты или опухоль. Например, при острой инфекции или некоторых злокачественных опухолях, положительный эффект может появлятся вследствие индукции воспаления и локального ответа белков острой фазы. Таким образом, растворимые рецепторы, описанные в настоящем описании, или антитела согласно изобретению можно использовать для специфичного направления воздействия провоспалительного лиганда IL-31. Смотрите Cosman, D. Cytokine 5: 95-106, 1993; и Fernandez-Botran, R. Exp. Qpin. Invest. Drugs 9:497-513, 2000.
Как правило, дозировка вводимого полипептида IL-31 (или аналога или слитого белка IL-31RA) варьирует, в зависимости от таких факторов, как возраст пациента, масса тела, рост, пол, общее медицинское состояние и предшествующий медицинский анамнез. Как правило, желательно предоставление реципиенту дозировки полипептида IL-31, которая находится в диапазоне от приблизительно 1 пг/кг до 10 мг/кг (количество средства/масса тела пациента), хотя в определенных случаях также можно вводить более низкие или более высокие дозы. Специалист в данной области может легко определить такие дозировки и их изменение с использованием известных в данной области способов.
Введение полипептида IL-31 субъекту может быть местным, ингаляционным, внутривенным, внутриартериальным, внутрибрюшинным, внутримышечным, подкожным, интраплевральным, интратекальным, посредством перфузии через местный катетер, или посредством прямой инъекции в очаг повреждения. При введении терапевтических белков посредством инъекции введение можно проводить посредством непрерывной инфузии или посредством однократной или многократных болюсных инъекций.
Дополнительные способы введения включают в себя пероральное введение, введение на слизистую оболочку, легочное введение и чрескожное введение. Пероральная доставка является пригодной для микросфер на основе полиэфира, микросфер на основе зеина, протеоидных микросфер, микросфер на основе полицианоакрилата и систем на основе липидов (смотрите, например, DiBase and Morrel, "Oral Delivery of Microencapsulated Proteins," in Protein Delivery: Physical Systems, Sanders and Hendren (eds.), страницы 255-288 (Plenum Press 1997)). Пригодность интраназальной доставки проиллюстрирована посредством такого способа введения инсулина (смотрите, например, Hinchcliffe and Hlum, Adv. Drug Deliv. Rev. 35:199 (1999)). Сухие или жидкие частицы, содержащие IL-31, можно получать и ингалировать с помощью диспергаторов сухих порошков, генераторов жидких аэрозолей или небулайзеров (например, Pettit and Gombotz, TIBTECH 16:343 (1998); Patton et al, Adv. Drug Deliv. Rev. 35:235 (1999)). Этот подход проиллюстрирован системой контроля диабета AERX, которая представляет собой карманный электронный ингалятор, который доставляет инсулин в виде аэрозоля в легкие. В исследованиях была показана доставка белков размером 48000 кДа через кожу в терапевтических концентрациях с помощью ультразвуковой обработки с низкой частотой, что иллюстрирует пригодность чрескожного введения (Mitragotri et al, Science 269:850 (1995)). Чрескожная доставка с использованием электропорации представляет собой другой способ введения молекулы, обладающей активностью в отношении связывания IL-31 (Potts et al, Pharm. Biotechnol. 10:213 (1997)).
Фармацевтическую композицию, содержащую белок, полипептид, или пептид, обладающий активностью в отношении связывания IL-31, можно составлять в соответствии с известными способами с получением фармацевтически приемлемой композиции, где терапевтические белки объединяют в смесь с фармацевтически приемлемым носителем. Говорят, что композиция является "фармацевтически приемлемым носителем", если ее введение может переносить являющийся реципиентом пациент. Одним примером фармацевтически приемлемого носителя является стерильный фосфатно-солевой буфер. Другие пригодные носители хорошо известны в данной области. Смотрите, например, Gennaro (ed.), Remington's Pharmaceutical Sciences, 19th Edition (Mack Publishing Company 1995).
Для целей терапии молекулы, обладающие активностью в отношении связывания IL-31, и фармацевтически приемлемый носитель вводят пациенту в терапевтически эффективном количестве. Говорят, что сочетание белка, полипептида или пептида, обладающего активностью в отношении связывания IL-31, и фармацевтически приемлемого носителя вводят в "терапевтически эффективном количестве", если вводимое количество является физиологически значимым. Вещество является физиологически значимым, если его наличие приводит к поддающемуся выявлению изменению физиологии являющегося реципиентом пациента. Например, средство, используемое для лечения воспаления, является физиологически значимым, если его присутствие ослабляет по меньшей мере часть воспалительного ответа.
Фармацевтическая композиция, содержащая IL-31 (или аналог или слитый белок IL-31), может быть представлена в жидкой форме, в форме аэрозоля или в твердой форме. Примером жидких форм являются инъецируемые растворы, аэрозоли, капли, топологические растворы и пероральные суспензии. Иллюстративные твердые формы включают в себя капсулы, таблетки и формы с контролируемым высвобождением. Иллюстративным примером последней формы является миниосмотические насосы и имплантаты (Bremer et al, Pharm. Biotechnol. 10:239 (1997); Ranade, "Implants in Drug Delivery", в Drug Delivery Systems, Ranade and Hollinger (eds.), страницы 95-123 (CRC Press 1995); Bremer et al, "Protein Delivery with Infusion Pumps", в Protein Delivery: Physical Systems, Sanders and Hendren (eds.), страницы 239-254 (Plenum Press 1997); Yewey et al, "Delivery of Proteins from a Controlled Release Injectable Implant", в Protein Delivery: Physical Systems, Sanders and Hendren (eds.), страницы 93-117 (Plenum Press 1997)). Другие твердые формы включают в себя кремы, пасты, другие средства для топологического применения и т.п.
Антитела против IL-31, описанные в настоящем описании, также могут быть составлены в качестве иммунолипосом. Липосомы, содержащие антитело, получают способами, известными в данной области, такими как описано в Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA, 77: 4030 (1980); и патентах США №№ 4485045 и 4544545. Липосомы с повышенным временем циркуляции описаны в патенте США № 5013556.
Особенно пригодные липосомы можно получать способом выпаривания с обращенной фазой композиции липидов, содержащей фосфатидилхолин, холестерин и образованное с помощью PEG производное фосфатидилэтаноламина (PEG-PE). Липосомы экструдируют через фильтры с определенным размером пор с получением липосом с требуемым диаметром. Fab'-фрагменты антитела согласно изобретению можно конъюгировать с липосомами, как описано в Martin et al. J. Biol. Chem. 257: 286-288 (1982), посредством реакции дисульфидного обмена. В липосоме необязательно содержится химиотерапевтическое средство (такое как доксорубицин). Смотрите Gabizon et al. J. National Cancer Inst. 81(19) 1484 (1989).
Для хранения терапевтические составы антитела получают посредством смешивания антитела, обладающего требуемой степенью чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Phermaceuticals Sciences 16th edition, Osol, A. Ed. (1980)), в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают в себя буферы, такие как фосфатный, цитратный буфер и буфер на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензэтония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как Tween™, Pluronics™ или полиэтиленгликоль (PEG).
Описанный в настоящем описании состав также может содержать более одного активного соединения, если необходимо в случае конкретных показаний для лечения, предпочтительно активные соединения с дополнительными видами активности, которые не оказывают отрицательного воздействия друг на друга. Например, также может быть желательным предоставление антител, которые связываются с IL-31, в одном составе. Альтернативно или дополнительно, композиция может содержать химиотерапевтическое средство или цитокин. Такие молекулы в сочетании соответственно представлены в количествах, которые являются эффективными для предполагаемой цели.
Активные ингредиенты также могут быть заключены в микрокапсулы, полученные, например, посредством способов коацервации или межповерхностной полимеризации, например, микрокапсулы на основе гидроксиметилцеллюлозы или желатина и микрокапсулы на основе полиметилметакрилата, соответственно, в коллоидных системах для доставки лекарственного средства (например, липосомах, микросферах альбумина, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие способы описаны в Remington's Pharmaceutical Sciences, 16 издание, Osol, A. Ed. (1980).
Составы, предназначенные для введения in vivo, должны быть стерильными. Этого легко добиваются посредством фильтрации через стерильные фильтрационные мембраны.
Можно получать препараты с замедленным высвобождением. Пригодные примеры препаратов с замедленным высвобождением включают в себя полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антитело, где матрицы находятся в форме имеющих определенную форму изделий, например, пленок, или микрокапсул. Примеры матриц с замедленным высвобождением включают в себя полиэфиры, гидрогели (например, поли-2-гидроксиэтилметакрилат или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, недеградируемый этилен-винилацетат, деградируемые сополимеры молочной кислоты и гликолевой кислоты, такие как Lupron Depot™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как полимеры, такие как этилен-винилацетат и молочная кислота-гликолевая кислота обеспечивают высвобождение молекул в течение 100 суток, определенные гидрогели высвобождают белки в течение более коротких периодов времени. Когда инкапсулированные антитела остаются в организме в течение длительного времени, они могут денатурировать или образовывать агрегаты в результате воздействия влажности при 37°C, приводящего к потере биологической активности и возможным изменениям иммуногенности. Для стабилизации можно разрабатывать целесообразные стратегии, в зависимости от вовлеченного механизма. Например, если выявлено, что механизм агрегации представляет собой межмолекулярное образование связи S-S через тио-дисульфидный обмен, стабилизации можно достигать посредством модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контроля степени влажности, с использованием соответствующих добавок и разработки определенных композиций полимерных матриц.
Одним способом доставки терапевтических полипептидов субъекту внутривенно, внутрибрюшинно, интратекально, внутримышечно, подкожно или посредством перорального введения, ингаляции или интраназального введения, являются липосомы. Липосомы представляют собой микроскопические везикулы, которые состоят из одного или нескольких липидных бислоев, окруженных водными камерами (смотрите, главным образом, Bakker-Woudenberg et al, Eur. J. Clin. Microbiol. Infect. Dis. 12 (Suppl. 1): S61 (1993), Kim, Drugs 46:618 (1993), и Ranade, "Site-Specific Drug Delivery Using Liposomes as Carriers", в Drug Delivery Systems, Ranade and Hollinger (eds.), стр 3-24 (CRC Press 1995)). Состав липосом является сходным с составом мембран, и в результате этого возможно безопасное введение липосом, и они являются биологически деградируемыми. В зависимости от способа получения, липосомы могут быть однослойными или многослойными, и размер диаметра липосом может варьировать от 0,02 мкм до более чем 10 мкм. В липосомы можно инкапсулировать множество веществ: гидрофобные вещества распределяются в бислоях, а гидрофильные вещества распределяются во внутреннем водном пространстве(ах) (смотрите, например, Machy et al, Liposomes In Cell Biology And Pharmacology (John Libbey 1987), и Ostro et al, American J. Hosp. Pharm. 46:1516 (1989)). Более того, терапевтическую доступность инкапсулированного вещества можно контролировать, варьируя размер липосом, количество бислоев, липидный состав, а также заряд и поверхностные свойства липосом.
Липосомы могут адсорбироватся на клетки практически любого типа, а затем медленно высвобождать инкапсулированное вещество. Альтернативно может происходить эндоцитоз адсорбированных липосом клетками, которые являются фагоцитарными. После эндоцитоза происходит интрализосомальная деградация липосомальных липидов и высвобождение инкапсулированных веществ (Scherphof et al, Ann. N.Y. Acad. Sci. 446:368 (1985)). После внутривенного введения небольшие липосомы (от 0,1 до 1,0 мкм), как правило, захватываются клетками ретикулоэндотелиальной системы, расположенными, главным образом, в печени и селезенке, в то время как липосомы более 3,0 мкм депонируются в легком. Предпочтительный захват липосом меньших размеров клетками ретикулоэндотелиальной системы использовали для доставки химиотерапевтических средств в макрофаги и в опухоли печени.
Ретикулоэндотелиальную систему можно обходить несколькими способами, включая насыщение частиц липосом большими дозами или селективную инактивацию макрофагов фармакологическими способами (Claassen et al, Biochim. Biophys. Acta 502:428 (1984)). Кроме того, было показано, что встраивание образованных с помощью гликолипидов или полиэтиленгликоля производных фосфолипидов в мембраны липосом приводит к значительному снижению захвата ретикулоэндотелиальной системой (Allen et al, Biochim. Biophys. Acta 1068:133 (1991); Allen et al, Biochim. Biophys. Acta 1150:9 (1993)).
Липосомы также можно получать для нацеливания на конкретные клетки или органы, варьируя фосфолипидный состав или встраивая в липосомы рецепторы или лиганды. Например, липосомы, изготовленные с высоким содержанием неионного поверхностно-активного вещества, использовали для нацеливания на печень (Hayakawa et al., японский патент 04-244018; Kato et al, Biol. Pharm. Bull. 16:960 (1993)). Эти составы получали смешиванием фосфатидилхолина сои, α-токоферола и этоксилированного гидрогенизированного касторового масла (HCO-60) в метаноле, концентрации смеси в вакууме и последующим разбавлением смеси водой. Также было показано, что состав липосом из дипальмитоилфосфатидилхолина (DPPC) с полученной из сои смесью стерилглюкозида (SG) и холестерина (Ch) направлен на печень (Shimizu et al, Biol Pharm. Bull 20:881 (1997)).
Альтернативно, с поверхностью липосомы можно связывать различные лиганды для нацеливания, такие как антитела, фрагменты антител, углеводы, витамины и транспортные белки. Например, для нацеливания на рецепторы для асиалогликопротеина (галактозы), которые экспрессируются исключительно на поверхности клеток печени, липосомы можно модифицировать с помощью производных галактозиллипида разветвленного типа (Kato and Sugiyama, Grit. Rev. Ther. Drug Carrier Syst. 14:287 (1997); Murahashi et al, Biol. Pharro. Bull. 20:259 (1997)). Аналогично, было показано, Wu et al, Hepatology 27:772 (1998), что липосомы, меченные асиалофетуином, приводят к укороченному периоду полужизни липосом в плазме и к значительно усиленному захвату меченных асиалофетуином липосом гепатоцитами. С другой стороны, накопление в печени липосом, содержащих производные галактозиллипида разветвленного типа, можно ингибировать посредством предварительной инъекции асиалофетуина (Murahashi et al, Biol. Pharm. Bull. 20:259 (1997)). Полиаконитилированные липосомы на основе сывороточного альбумина человека представляют собой другой подход для нацеливания липосом в клетки печени (Kamps et al, Proc. Nat'l Acad. Sci. USA 94:11681 (1997)). Более того, Geho, et al., патент США № 4603044, описывают направленную в гепатоциты систему доставки липосомальных везикул, которая обладает специфичностью в отношения гепатобилиарных рецепторов, связанных со специализированными метаболическими клетками печени.
Полипептиды, обладающие активностью в отношении связывания IL-31, можно инкапсулировать в липосомы с использованием стандартных способов микроинкапсулирования белков (смотрите, например, Anderson et al, Infect. Immun. 31:1099 (1981), Anderson et al, Cancer Res. 50:1853 (1990), и Cohen et al, Biochim. Biophys. Acta 1063:95 (1991), Alving et al "Preparation and Use of Liposomes in Immunological Studies", в Liposome Technology, 2 издание, Vol. III, Gregoriadis (ed.), страница 317 (CRC Press 1993), Wassef et al, Meth. Enzymol. 149:124 (1987)). Как указано выше, терапевтически приемлемые липосомы могут содержать множество компонентов. Например, липосомы могут содержать липидные производные полиэтиленгликоля (Allen et al, Biochim. Biophys. Acta 1150:9 (1993)).
Для поддержания высоких системных уровней терапевтических белков разработаны деградируемые полимерные микросферы. Микросферы получают из деградируемых полимеров, таких как сополимер лактида и гликолида (PLG), полиангидриды, полиортоэфиры, биологически недеградируемые полимеры этилвинилацетата, в которых в полимере заключены белки (Gombotz и Pettit, Bioconiugate Chem. 6:332 (1995); Ranade, "Role of Polymers in Drug Delivery", в Drug Delivery Systems, Ranade and Hollinger (eds.), страницы 51-93 (CRC Press 1995); Roskos and Maskiewicz, "Degradable Controlled Release Systems Useful for Белок Delivery," в Protein Delivery: Physical Systems, Sanders and Hendren (eds.), страницы 45-92 (Plenum Press 1997); Bartus et al, Science 281:1161 (1998); Putney and Burke, Nature Biotechnology 16:153 (1998); Putney, Curr. Opin. Chem. Biol. 2:548 (1998)). Покрытые полиэтиленгликолем (PEG) наносферы также могут представлять собой носители для внутривенного введения терапевтических белков (смотрите, например, Gref et al, Pharm. Biotechnol. 10: 167 (1997)).
Специалисты в данной области могут разработать другие дозированные формы, как показано, например, Ansel and Popovich, Pharmaceutical Dosage Forms and Drug Delivery Systems, 5 издание (Lea & Febiger 1990), Gennaro (ed.), Remington's Pharmaceutical Sciences, 19 издание (Mack Publishing Company 1995), и Ranade and Hollinger, Drug Delivery Systems (CRC Press 1996).
В качестве примера, фармацевтические композиции могут поставляться в качестве набора, содержащего контейнер, который содержит полипептид IL-31 или антагонист IL-31 (например, антитело или фрагмент антитела, которое связывает полипептид IL-31). Терапевтические полипептиды могут быть предоставлены в форме инъецируемого раствора для однократных или многократных доз, или в качестве стерильного порошка, который разбавляют перед инъекцией. Альтернативно такой набор может включать в себя диспергатор сухого порошка, генератор жидкого аэрозоля или небулайзер для введения терапевтического полипептида.
В одном аспекте это изобретение относится к моноклональному антителу, содержащему моноклональное антитело, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6815; b) патентного номера депозита в ATCC PTA-6816; c) патентного номера депозита в ATCC PTA-6829; d) патентного номера депозита в ATCC PTA-6830; e) патентного номера депозита в ATCC PTA-6831; f) патентного номера депозита в ATCC PTA-6871; g) патентного номера депозита в ATCC PTA-6872; h) патентного номера депозита в ATCC PTA-6875 и i) патентного номера депозита в ATCC PTA-6873. В этом варианте осуществления антитело нейтрализует взаимодействие IL-31 (SEQ ID NO:2) с IL-31RA (SEQ ID NO:5). В другом варианте осуществления антитело представляет собой (a) моноклональное антитело мыши, (b) гуманизированное антитело, образованное из (a), или (c) фрагмент антитела, или (d) моноклональное антитело человека. В другом варианте осуществления, антитело дополнительно содержит радионуклид, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидный маркер, магнитную частицу или токсин. В другом варианте осуществления антитело дополнительно обладает пегилированием. В другом варианте осуществления антитело представляет собой нейтрализующее антитело. В другом варианте осуществления введение антитела млекопитающему ингибирует, предотвращает, снижает или блокирует провоспалительную активность полипептида. В другом варианте осуществления введение антитела млекопитающему ингибирует, предотвращает, снижает или блокирует расчесывание и дерматит, обусловленные провоспалительной активностью полипептида.
В другом аспекте это изобретение относится к моноклональному антителу, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6815; b) патентного номера депозита в ATCC PTA-6816; c) патентного номера депозита в ATCC PTA-6871; d) патентного номера депозита в ATCC PTA-6829 и e) патентного номера депозита в ATCC PTA-6830.
В другом аспекте это изобретение относится к моноклональному антителу, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ED NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6815; b) патентного номера депозита в ATCC PTA-6816 и c) патентного номера депозита в ATCC PTA-6871.
В другом аспекте это изобретение относится к моноклональному антителу, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6829 и b) патентного номера депозита в ATCC PTA-6830.
В другом аспекте предусмотрено моноклональное антитело, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6872 и b) патентного номера депозита в ATCC PTA-6875.
В другом аспекте предусмотрено моноклональное антитело, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC PTA-6874. В этом варианте осуществления антитело представляет собой нейтрализующее антитело. В этом варианте осуществления введение антитела млекопитающему ингибирует, предотвращает, снижает или блокирует провоспалительную активность полипептида. В этом варианте осуществления введение антитела млекопитающему ингибирует, предотвращает, снижает или блокирует расчесывание и дерматит, обусловленные провоспалительной активностью полипептида.
В другом аспекте это изобретение относится к способу уменьшения воспаления у млекопитающего, включающему в себя введение млекопитающему количества антитела против IL-31, описанного в настоящем описании, посредством которого снижается воспаление.
В другом аспекте это изобретение относится к способу лечения млекопитающего, страдающего воспалительным заболеванием, в котором участвует IL-31, включающему в себя: введение антагониста IL-31 млекопитающему так, чтобы воспаление снижалось, где антагонист содержит (i) антитело, фрагмент антитела или связывающий полипептид, который специфически связывает полипептид или фрагмент полипептида IL-31RA или (ii) полипептид или фрагмент полипептида IL-31RA; и где воспалительная активность IL-31 снижается. В этом варианте осуществления заболевание представляет собой воспалительное заболевание, включающее в себя связанные с зудом заболевания. В этом варианте осуществления заболевание представляет собой атопический дерматит. В этом варианте осуществления заболевание представляет собой узловатую почесуху.
В другом аспекте это изобретение относится к способу уменьшения, ингибирования или предотвращения эффекта зуда у млекопитающего, в котором участвует IL-31, включающему в себя: введение антагониста IL-31 млекопитающему так, чтобы зуд снижался, где антагонист содержит (i) антитело, фрагмент антитела или связывающий полипептид, который специфически связывает полипептид или полипептидный фрагмент полипептида, содержащего аминокислотную последовательность SEQ ID NO:2, или ее фрагмент; и где происходит снижение связанной с зудом активности IL-31. В этом варианте осуществления заболевание млекопитающего представляет собой атопический дерматит. В этом варианте осуществления заболевание млекопитающего представляет собой дерматит. В этом варианте осуществления антитело представляет собой антитело, как описано в настоящем описании.
В другом аспекте это изобретение относится к способу снижения, ингибирования или предотвращения эффекта зуда у млекопитающего, в котором участвует IL-31, включающему в себя: введение антагониста IL-31 млекопитающему так, чтобы происходило снижение зуда у млекопитающего, где антагонист содержит (i) антитело, фрагмент антитела или связывающий полипептид, который специфически связывает полипептид или полипептидный фрагмент полипептида, содержащего аминокислотную последовательность SEQ ID NO:2, или ее фрагмент; и где посредством этого происходит снижение активности IL-31 в отношении расчесывания. В этом варианте осуществления заболевание млекопитающего представляет собой атопический дерматит. В этом варианте осуществления заболевание млекопитающего представляет собой дерматит. В этом варианте осуществления антитело представляет собой антитело, как описано в настоящем описании.
В другом аспекте это изобретение относится к моноклональному антителу, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое клоном гибридомы, обозначаемым номером, выбранным из: a) клона 292.12.3.1 (патентный номер депозита в ATCC PTA-6815); b) клона 292.63.5.3 (патентный номер депозита в ATCC PTA-6829); c) клона 292.72.3.1 (патентный номер депозита в ATCC PTA-6816); d) клона 292.84.1.6 (патентный номер депозита в ATCC PTA-6871) и e) клона 292.118.6.4 (патентный номер депозита в ATCC PTA-6830).
В другом аспекте это изобретение относится к моноклональному антителу, которое специфически связывает полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое клоном гибридомы, обозначаемым номером, выбранным из: a) клона 294.35.2.6.3 (патентный номер депозита в ATCC PTA-6872); b) клона 294.144.3.5 (патентный номер депозита в ATCC PTA-6873); c) клона 294.154.5.6 (патентный номер депозита в ATCC PTA-6875) и d) клона 294.163.2.1 (патентный номер депозита в ATCC PTA-6831).
В другом аспекте это изобретение относится к моноклональному антителу, содержащему моноклональное антитело, способное конкурировать за связывание полипептида, содержащего аминокислотную последовательность SEQ ID NO:2, где полипептид способен связывать моноклональное антитело, продуцируемое гибридомой, депонированной в Американской коллекции типовых культур, обладающей патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6815; b) патентного номера депозита в ATCC PTA-6816; c) патентного номера депозита в ATCC PTA-6829; d) патентного номера депозита в ATCC PTA-6830; e) патентного номера депозита в ATCC PTA-6831; f) патентного номера депозита в ATCC PTA-6871; g) патентного номера депозита в ATCC PTA-6872; h) патентного номера депозита в ATCC PTA-6875 и i) патентного номера депозита в ATCC PTA-6873.
В другом аспекте предусмотрена гибридома, где гибридома депонирована в Американской коллекции типовых культур с патентным обозначением образца ATCC, выбранным из: a) патентного номера депозита в ATCC PTA-6815; b) патентного номера депозита в ATCC PTA-6816; c) патентного номера депозита в ATCC PTA-6829; d) патентного номера депозита в ATCC PTA-6830; e) патентного номера депозита в ATCC PTA-6831; f) патентного номера депозита в ATCC PTA-6871; g) патентного номера депозита в ATCC PTA-6872; h) патентного номера депозита в ATCC PTA-6875 и i) патентного номера депозита в ATCC PTA-6873.
В другом аспекте настоящее изобретение относится к способу получения антитела против полипептида IL-31, включающему в себя: инокуляцию животному полипептида, выбранного из группы, состоящей из: (a) полипептида, состоящего из от 9 до 141 аминокислоты, где полипептид является идентичным непрерывной последовательности аминокислотных остатков в SEQ ID NO:2, от аминокислоты номер 24 (Ser) до аминокислоты номер 164 (Thr); (b) полипептида, как описано выше; (c) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 из аминокислот номер 38-52; (d) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 из аминокислот номер 83-98; (e) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 из аминокислот номер 104-117; (f) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 из аминокислот номер 137-152; (g) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 из аминокислот номер 38-152; (h) полипептида, содержащего аминокислотную последовательность SEQ 3D NO:2 из аминокислот номер 24-164; (c) полипептида, содержащего аминокислотную последовательность SEQ ID NO:11, из аминокислот номер 38-52; (d) полипептида, содержащего аминокислотную последовательность SEQ ID NO:11 из аминокислот номер 85-98; (e) полипептида, содержащего аминокислотную последовательность SEQ ID NO:11 из аминокислот номер 104-118; (f) полипептида, содержащего аминокислотную последовательность SEQ ID NO:11 из аминокислот номер 141-157; (g) полипептида, содержащего аминокислотную последовательность SEQ ID NO:11 из аминокислот номер 38-157; (h) полипептида, содержащего аминокислотную последовательность SEQ ID NO:11 из аминокислот номер 24-163; (i) полипептида, содержащего антигенный эпитоп в соответствии с профилем гидрофильности Хоппа/Вудса SEQ ID NO:2 или SEQ ID NO:11, где профиль основан на продвижении окна из шести остатков, углубленные остатки G, S и T и экспонированные остатки H, Y и W не учитывают; и где полипептид вызывает иммунный ответ у животного с продукцией антитела; и выделение антитела из животного.
В другом аспекте настоящее изобретение относится к антителу (например, нейтрализующему антителу), продуцируемому способом, как описано выше, где антитело связывается с полипептидом SEQ IDN NO:2 или SEQ ID NO:11. В одном варианте осуществления антитело, описанное выше, специфически связывается с полипептидом, представленным в SEQ ID NO:2 или SEQ ID NO:11.
В другом аспекте настоящее изобретение относится к способу выявления наличия IL-31 в биологическом образце, включающему в себя стадии: (a) контактирования биологического образца с антителом или фрагментом антитела, как описано выше, где контактирование проводят в условиях, которые обеспечивают связывание антитела или фрагмента антитела с биологическим образцом, и (b) выявления либо связавшегося антитела, либо связавшегося фрагмента антитела.
В другом аспекте настоящее изобретение относится к способу уничтожения клеток злокачественной опухоли, включающему в себя получения ex vivo ткани или биологического образца, содержащего злокачественные клетки пациента, или идентификацию злокачественных клеток in vivo; продукцию полипептида способом, как описано в настоящем описании; составления полипептида в фармацевтически приемлемом носителе; и введения пациенту или воздействия полипептида на злокачественные клетки; где полипептид вызывает гибель клеток. В одном варианте осуществления способ уничтожения злокачественных клеток является таким, как описано выше, где полипептид дополнительно конъюгируют с токсином. В одном варианте осуществления антитело является таким, как описано выше, где антитело выбрано из группы, состоящей из: (a) поликлонального антитела, (b) моноклонального антитела мыши, (c) гуманизированного антитела, образованного из (b), (d) фрагмента антитела и (e) моноклонального антитела человека.
В другом аспекте настоящее изобретение относится к антителу или фрагменту антитела, которое специфически связывается с полипептидом, содержащим последовательность из аминокислотных остатков, выбранную из группы, состоящей из: (a) полипептида, представленного от остатка 38 (Val) до 152 (Leu), как показано в SEQ ID NO:2; (b) полипептида, представленного от остатка 27 (Leu) до 164 (Thr), как показано в SEQ ID NO:2; (c) полипептида, представленного от остатка 24 (Thr) до 164 (Thr), как показано в SEQ ID NO:2; и (d) полипептида, представленного от остатка 1 (Met) до 164 (Thr), как показано в SEQ ID NO:2. В другом варианте осуществления антитело является таким, как описано выше, где антитело дополнительно содержит радионуклид, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидную метку, магнитную частицу, лекарственное средство или токсин.
В другом аспекте настоящее изобретение относится к способу ингибирования индуцируемой IL-31 пролиферации или дифференцировки кроветворных клеток и гемопоэтических клеток-предшественников, включающему в себя культивирование клеток костного мозга или периферической крови с композицией, содержащей количество антитела, как описано в настоящем описании, достаточное для снижения пролиферации или дифференцировки кроветворных клеток в клетках костного мозга или периферической крови по сравнению с клетками костного мозга или периферической крови, культивируемыми в отсутствие растворимого рецептора для цитокина. В одном варианте осуществления способ ингибирования индуцируемой IL-31 пролиферации или дифференцировки кроветворных клеток и гемопоэтических клеток-предшественников является таким, как описано выше, где кроветворные клетки и гемопоэтические клетки-предшественники представляют собой лимфоидные клетки. В другом варианте осуществления способ ингибирования индуцируемой IL-31 пролиферации или дифференцировки кроветворных клеток и гемопоэтических клеток-предшественников является таким, как описано выше, где лимфоидные клетки представляют собой макрофаги или T-клетки.
В другом аспекте настоящее изобретение относится к способу уменьшения индуцированного IL-31 воспаления, включающему в себя введение млекопитающему с воспалением количества композиции антитела, как описано в настоящем описании, достаточного для уменьшения воспаления.
В другом аспекте настоящее изобретение относится к способу подавления воспалительного ответа у млекопитающего с воспалением, включающему в себя: (1) определение уровня воспалительной молекулы; (2) введение композиции, содержащей антитело, как описано в настоящем описании, в приемлемом фармацевтическом носителе; (3) определение уровня воспалительной молекулы после введения; (4) сравнение уровня воспалительной молекулы стадии (1) с уровнем воспалительной молекулы стадии (3), где отсутствие повышения или снижения уровня воспалительной молекулы указывает на подавление воспалительного ответа. В одном варианте осуществления антитело является таким, как описано выше, где антитело дополнительно содержит радионуклид, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидную метку, магнитную частицу, лекарственное средство или токсин.
В другом аспекте настоящее изобретение относится к способу ингибирования индуцированной IL-31 пролиферации или дифференцировки кроветворных клеток и гемопоэтических клеток-предшественников, включающему в себя культивирование клеток костного мозга или периферической крови с композицией, содержащей количество антитела, как описано в настоящем описании, достаточное для уменьшения пролиферации или дифференцировки кроветворных клеток в костном мозге или клетках периферической крови по сравнению с клетками костного мозга или периферической крови, культивированными в отсутствие растворимого рецептора для цитокина. В одном варианте осуществления способ ингибирования индуцированной IL-31 пролиферации или дифференцировки кроветворных клеток и гемопоэтических клеток-предшественников является таким, как описано выше, где кроветворные клетки и гемопоэтические клети-предшественники представляют собой лимфоидные клетки. В другом варианте осуществления способ ингибирования индуцированной IL-31 пролиферации или дифференцировки кроветворных клеток и гемопоэтических клеток-предшественников является таким, как описано выше, где лимфоидные клетки представляют собой макрофаги или T-клетки.
В другом аспекте настоящее изобретение относится к способу уменьшения индуцированного IL-31 воспаления, включающему в себя введение млекопитающему с воспалением количества композиции антитела, как описано в настоящем описании, достаточного для уменьшения воспаления.
В другом аспекте настоящее изобретение относится к способу подавления воспалительного ответа у млекопитающего с воспалением, включающему в себя: (1) определение уровня воспалительной молекулы; (2) введение композиции, содержащей антитело, как описано в настоящем описании, в приемлемом фармацевтическом носителе; (3) определение уровня воспалительной молекулы после введения; (4) сравнение уровня воспалительной молекулы стадии (1) с уровнем воспалительной молекулы стадии (3), где отсутствие повышения или снижения воспалительной молекулы указывает на подавление воспалительного ответа.
В другом аспекте настоящее изобретение относится к способу лечения млекопитающего, страдающего воспалительным заболеванием, в котором участвует IL-31, включающему в себя: введение антагониста IL-31 млекопитающему так, чтобы происходило уменьшение воспаления, где антагонист выбран из группы, состоящей из антитела или связывающего полипептида, который специфически связывает полипептид или фрагмент полипептида IL-31 (SEQ ID NO:2). В одном варианте осуществления, способ лечения млекопитающего, страдающего воспалительным заболеванием, является таким, как описано выше, где заболевание представляет собой хроническое воспалительное заболевание. В другом варианте осуществления способ лечения млекопитающего, страдающего воспалительным заболеванием, является таким, как описано выше, где заболевание представляет собой хроническое воспалительное заболевание, выбранное из группы, состоящей из: воспалительного заболевания кишечника; язвенного колита; болезни Крона; атопического дерматита; экземы и псориаза. В другом варианте осуществления способ лечения млекопитающего, страдающего воспалительным заболеванием, является таким, как описано выше, где заболевание представляет собой острое воспалительное заболевание. В другом варианте осуществления способ лечения млекопитающего, страдающего воспалительным заболеванием, является таким, как описано выше, где заболевание представляет собой острое воспалительное заболевание, выбранное из группы, состоящей из: эндотоксемии; септицемии; синдрома токсического шока и инфекционного заболевания. В другом варианте осуществления способ лечения млекопитающего, страдающего воспалительным заболеванием, является таким, как описано выше, где антитело дополнительно содержит радионуклид, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидную метку, магнитную частицу, лекарственное средство или токсин.
В другом аспекте настоящее изобретение относится к способу выявления воспаления у пациента, включающему в себя: получение ткани или биологического образца пациента; инкубацию ткани или биологического образца с антителом, как описано в настоящем описании, в условиях, где антитело связывается с комплементарным ему полипептидом в ткани или биологическом образце; визуализации антитела, связавшегося с тканью или биологическим образцом; и сравнения уровней антитела, связавшегося с тканью или биологическим образцом пациента, с нормальной контрольной тканью или биологическим образцом, где повышение уровня антитела, связавшегося с тканью или биологическим образцом пациента, относительно нормальной контрольной ткани или биологического образца указывает на воспаление у пациента.
В другом аспекте настоящее изобретение относится к способу выявления воспаления у пациента, включающему в себя: получение ткани или биологического образца пациента; мечение полинуклеотида, содержащего по меньшей мере 14 последовательных нуклеотидов SEQ ID NO:1 или комплементарной SEQ ID NO:1 последовательности; инкубации ткани или биологического образца с полинуклеотидом в условиях, при которых полинуклеотид гибридизуется с комплементарной полинуклеотидной последовательностью; визуализацию меченого полинуклеотида в ткани или биологическом образце; и сравнение уровня гибридизации меченого полинуклеотида в ткани или биологическом образце пациента с нормальной здоровой тканью или биологическим образцом, где усиление гибридизации меченого полинуклеотида с тканью или биологическим образом пациента относительно нормальной здоровой ткани или биологического образца указывает на наличие у пациента воспаления.
Это изобретение далее проиллюстрировано посредством следующих неограничивающих примеров.
ПРИМЕРЫ
Пример 1
Получение MAb крысы против IL-31 мыши
A. Иммунизация и скрининг сыворотки крыс
Четырех самок крыс Sprague-Dawley в возрасте 3 месяца (Charles River Laboratories, Wilmington, MA) иммунизировали посредством IL-31 мыши. Крысам проводили первичную иммунизацию посредством внутрибрюшинной инъекции ~290 мкг очищенного рекомбинантного IL-31 мыши (продуцируемого клетками BHK), слитого на C-конце с пептидом, состоящим из последовательности EYMPME (SEQ ID NO:7), (в дальнейшем в настоящем описании обозначаемого как mIL31-CEE) в сочетании с полным адъювантом Фрейнда (Pierce, Rockford, IL) в соответствии с инструкциями изготовителя. После первичной иммунизации каждой крысе вводили дополнительно 150 мкг mIL-31-CEE в неполном адъюванте Фрейнда посредством внутрибрюшинного способа каждые две недели в течение шести недель. Через семь суток после третьей и четвертой иммунизаций у крыс через ретроорбитальное сплетение отбирали кровь и из крови выделяли сыворотку для анализа ее способности связываться с mIL-31-CEE в растворе и ингибировать (как определяют по нейтрализации) стимулирующую активность mIL-31-CEE в отношении клеточной линии, трансфицированной IL-31RA мыши.
Способность антител крысы против IL-31 мыши в антисыворотке связываться с mIL-31-CEE оценивали с использованием анализа ELISA по типу "антигенной ловушки". В этом анализе, лунки 96-луночных планшетов для ELISA из полистирола сначала покрывали Fc-специфичным антителом козы против IgG крысы (Jackson Immunoresearch) в концентрации 500 нг/мл в количестве 100 мкл/лунка в 0,1 М Na2CO3, pH 9,6. Планшеты инкубировали в течение ночи при 4°C, после чего несвязанное антитело аспирировали, и планшеты два раза промывали посредством PBS-Tween в количестве 300 мкл/лунка (0,137 М NaCl, 0,0027 М KCl, 0,0072 М Na2HPO4, 0,0015 М KH2PO4, 0,05% об./об. полисорбат 20, pH 7,2). Лунки блокировали посредством SuperBlock (Pierce, Rockford, IL) в количестве 200 мкл/лунка в течение 5 минут при комнатной температуре (RT), SuperBlock стряхивали с планшета и блокирование повторяли еще раз, после чего планшеты два раза промывали посредством PBS-Tween. Получали серийные 10-кратные разведения сыворотки (в PBS-Tween плюс 1% мас./об. бычий сывороточный альбумин (BSA) и 0,05% об./об. Proclin 300, pH 7,2 = буфер для разведения), начиная с первоначального разведения 1:1000 и до 1:1000000. По три образца каждого разведения затем переносили в планшет для анализа, 100 мкл/лунка, для связывания IgG крысы в сыворотке для анализа планшета через Fc-участок молекулы. Нормальная сыворотка крысы служила в качестве отрицательного контроля. После 1 часа инкубации при RT, лунки аспирировали и планшеты два раза промывали, как описано выше. Затем в лунки добавляли биотинилированный mIL-31-CEE (молекулярное соотношение биотин:белок 12:1) в концентрации 500 нг/мл, 100 мкл/лунка. После 1 часа инкубации при RT несвязанный биотинилированный mIL-31-CEE аспирировали из лунок и планшеты промывали два раза. Затем в каждую лунку добавляли меченный пероксидазой хрена стрептавидин (Pierce, Rockford, IL) в концентрации 500 нг/мл, 100 мкл/лунка, и планшеты инкубировали при RT в течение 1 часа. После удаления несвязанного HRP-SA, планшеты промывали 5 раз, в каждую лунку добавляли тетраметилбензидин (TMB) (BioFX Laboratories, Owings Mills, MD) в количестве 100 мкл/лунка и планшеты инкубировали в течение 5 минут при RT. Образование цвета останавливали посредством добавления стоп-реагента TMB для 450 нм в количестве 100 мкл/лунка (BioFX Laboratories, Owings Mills, MD) и значение поглощения для лунок определяли на устройстве Molecular Devices Spectra MAX 340 при 450 нм.
Способность антител крысы против IL-31 мыши в антисыворотке ингибировать (как определяют посредством нейтрализации) стимулирующую активность IL-31 мыши через распознающий его рецептор оценивали с использованием клеточного анализа нейтрализации, в котором использовали трансфицированную с помощью mIL-31RA/OSMRbeta клеточную линию Baf3 с люциферазной репортерной системой (Baf3/KZ134/mcytor17/mOSMRbeta), которую можно активировать посредством стимуляции клеток с помощью IL-31 мыши. Для этого анализа получали исходное разведение 1:250 каждой антисыворотки в среде для анализа (RPMI 1640, 10% ЭТС, 2 мМ L-глутамин, 1 мМ пируват натрия, 1× пенициллин-стрептомицин-неомицин (Lavitrogen, Carlsbad, CA)), а затем для каждой из них проводили серийные двукратные разведения до конечного разведения 1:32000. К каждому разведению антисыворотки добавляли равный объем mIL-31-CEE, разбавленного до 4 нг/мл в буфере для анализа, до конечного объема в лунках 100 мкл. Смеси антитело/цитокин инкубировали при RT в течение 0,5 часа, после чего клетки-мишени отмывали от их среды для роста, доводили до плотности 300000 клеток/мл и добавляли к смесям антитело/цитокин, 100 мкл/лунка. Среда для анализа, содержащая 2 нг/мл mIL-31 и не содержащая антисыворотки, служила в качестве отрицательного контроля и показывала максимальную стимуляцию, которой можно достичь в этом анализе. После инкубации при 37°C, 5% CO2, в течение 16-24 часов планшеты центрифугировали при 1500 об/мин в течение 5 минут, среду осторожно стряхивали и в каждую лунку добавляли 25 мкл 1× буфера для лизиса клеток (Promega, Madison, WI). Планшеты осторожно встряхивали в течение 10 минут при RT для обеспечения полного лизиса клеток, а затем лизаты переносили в сплошные белые 96-луночные планшеты (Corning/Costar 3917, Acton, MA). В каждую лунку добавляли 40 мкл субстрата для анализа люциферазы (Promega). Затем лунки оценивали на люциферазную активность (отражающую активацию посредством IL-31 репортерной конструкции STAT) на люминометре с использованием 5-секундного интеграционного интервала.
Как анализ ELISA по типу "антигенной ловушки", так и клеточный анализ нейтрализации, показали, что у всех четырех крыс развивался высокий антительный ответ на mIL-31, хотя у одной крысы отчетливо развился более сильный ответ, чем у других. В общем, ответ, измеренный с помощью ELISA по типу "антигенной ловушки", в значительной степени соответствовал ответу, выявленному в клеточном анализе нейтрализации, подтверждая, что антитела класса IgG отвечают, главным образом, за ингибирование mIL-31.
B. Слияние
Через четыре недели после последней внутрибрюшинной иммунизации, крыс с наиболее значимым титром нейтрализации mIL-31 иммунизировали последний раз посредством приблизительно 150 мкг mIL-31-CEE в PBS посредством внутрисосудистой инъекции. Через пять суток селезенку и лимфатические узлы этих крыс собирали, превращали в суспензию отдельных клеток и проводили слияние с клеточной линией миеломы мыши Sp2/0 (Shulman, M. et al., Nature 276:269-270, 1976) в соотношении лимфоидные клетки:клетки миеломы 4:1 с помощью PEG 1500 с использованием стандартных способов, известных в данной области (Harlow, E. and Lane, D. 1988. "Antibodies A Laboratory Manual", Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). Смесь для слияния распределяли в ряд 96-луночных планшетов с плоским дном совместно с тимоцитами BALB/c в качестве фидерного слоя (Oi, V.T. and Herzenberg, L.A.. 1980. "Selected Methods in Cellular Immunology", B.B. Mishell and S.M. Shiigi, eds., pp. 351-372, Freeman, San Francisco). Подпитку в лунках планшетов для слияния проводили один раз посредством замены 70% среды и тимоцитов через 4 суток и снова через двое суток посредством замены только среды. Лунки анализировали через восемь суток после культивирования подлежащих слиянию клеток.
C. Скрининг после слияния
Для первоначального скрининга использовали ELISA по типу "антигенной ловушки" для mIL-31, как описано выше, за исключением того, что супернатанты гибридом тестировали без разбавления и наносили в качестве реплик на планшеты для анализа из культуральных планшетов. Было выявлено приблизительно 170 положительных лунок. Затем супернатанты из каждой из этих лунок, а также нескольких отрицательных лунок, оценивали на их способность ингибировать mIL-31 в клеточном анализе нейтрализации, описанном ранее. Каждый из них тестировали в конечном разведении 1:8 в среде для анализа. В этом разведении уровень неспецифичных стимулирующих компонентов в супернатантах гибридом был достаточно снижен, чтобы вносить минимальный вклад в определяемый индекс стимуляции. Оказалось, что большая часть супернатантов нейтрализует в некоторой степени mIL-31, и приблизительно десять из них демонстрирует по существу полную нейтрализацию, а остальные показывают диапазон ингибирования вплоть до уровня, где приблизительно другие десять, как оказалось, обладают очень низким, если обладают им, нейтрализующим потенциалом. Клетки гибридомы приблизительно в 150 из лунок, положительных в ELISA по типу "антигенной ловушки", были успешно выращены в культуры в 24-луночных планшетах. Когда плотность культур 24-луночных планшетов составляла приблизительно 4-6·105 клеток/мл, супернатант (приблизительно 1,5 мл) собирали и хранили для каждой лунки по отдельности и клетки из каждой лунки замораживали.
Каждый из супернатантов из 24-луночных планшетов повторно анализировали как в ELISA по типу "антигенной ловушки", так и в клеточных анализах нейтрализации IL-31, используемых в скрининге после слияния. Кроме того, эти супернатанты также тестировали в ELISA по типу "антигенной ловушки" с использованием человеческого гомолога IL-31, также сконструированного с маркером EYMPME (SEQ ID NO:7), слитым с C-концом молекулы IL-31, и продуцированного в клетках BHK. Результаты указывают на то, что после экспансии большая часть основных супернатантов в лунках сохраняет их способность распознавать IL-31 мыши в растворе. Среди этих положительных в анализе по типу "антигенной ловушки" лунок, ингибиторная активность в отношении mIL-31 варьировала от практически полного ингибирования для приблизительно 20 супернатантов до по существу отсутствия ингибирования для 10-15 супернатантов. В анализе по типу "антигенной ловушки" не было выявлено перекрестной реактивности в отношении IL-31-CEE человека, указывая на то, что ни одно из антител крысы против mIL-31 не реагирует перекрестно с человеческим гомологом IL31 или не распознают маркер CEE, слитый с C-концом молекулы mIL-31-CEE.
D. Селекция и клонирование гибридом, продуцирующих нейтрализующие MAb против mIL31
Клетки из семи из десяти основных лунок с наиболее высокой нейтрализацией IL-31 (271.5.7, 271.9.4, 271.26.6, 271.27.4, 271.33.1, 271.33.3 и 271.39.4) клонировали в целях выделения клонированной гибридомы, продуцирующей представляющие интерес нейтрализующие mAb. Клетки клонировали в присутствии тимоцитов в 96-луночных титрационных микропланшетах для культивирования клеток с использованием стандартного подхода с разведением до низкой плотности (менее 1 клетки на лунку) и перед анализом оценивали моноклональность посредством микроскопического исследования лунок на единичные очаги роста. Через шесть суток выращивания проводили скрининг всех лунок на планшетах посредством ELISA по типу "антигенной ловушки" на IgG, специфичные в отношении mIL-31. Супернатанты из 6-10 лунок, которые были положительными в отношении специфичных mAb, также были получены из лунок только с одной колонией роста гибридомы, собирали из каждой клонированной серии и проводили повторный скрининг при различных разведениях в ELISA по типу "антигенной ловушки", а также в клеточном анализе нейтрализации, для идентификации "наилучшего" продуцирующего mAb клона. Результаты этих тестов показали, что соответствующие продуцирующие нейтрализующие mAb клоны гибридом были получены в сериях 271.9.4, 271.26.6, 271.33.1, 271.33.3 и 271.39.4. "Наилучший" клон в каждой из этих серий повторно клонировали и проводили скрининг субклонов, как описано, с получением конечных линий гибридом 271.9.4.2.6, 271.26.6.6.1, 271.33.1.2.2, 271.33.3.2.1 и 271.39.4.6.5. Изотип mAb IgG крысы, продуцируемого каждой из этих гибридом, определяли с использованием ELISA с помощью биотинилированных mAb мыши, специфичных в отношении изотипа IgG крысы. Было выявлено, что все пять mAb относятся к подклассу IgG1. Гибридома 271.26.6.6.1 была депонирована в патентной коллекции Американской коллекции типовых культур (ATCC, Manassas, VA) в качестве первоначального образца согласно Будапештскому договору и ей был присвоен регистрационный номер ATCC № PTA-6874.
Каждую из полностью клонированных гибридом, продуцирующих mAb против IL-31 мыши, адаптировали для роста в гибридомной не содержащей сыворотки среде (Hybridoma-SFM, Invitrogen). Супернатант от культур с высокой плотностью, выращенных в Hybridoma-SFM, центрифугировали для удаления клеток и клеточного дебриса, фильтровали через 0,2-мкм фильтр и очищали mAb посредством хроматографии с белком G с использованием стандартных способов, известных в данной области.
E. Оценка специфичной нейтрализующей активности MAb in vitro
Для оценки in vitro специфичной нейтрализующей активности каждого из пяти mAb крысы против IL-31 мыши, каждое из очищенных mAb тестировали в клеточном анализе нейтрализации, описанном ранее, за исключением того, что сначала к клеткам добавляли mIL-31, а затем эту смесь добавляли к антителам. В дополнение к нейтрализующей активности в отношении mIL-31-CEE, mAb также оценивали в отношении другой формы IL-31 мыши, обозначаемой как c108smIL-31, в которой остаток цистеина в положении 108 превращен в остаток серина. Последний материал продуцировали в E. coli без пептида C-EE. Обе формы mIL-31 использовали в конечной концентрации 1 и 5 нг/мл. mAb доводили до концентрации 20 мкг/мл в среде для анализа и тестировали по два образца в виде серийных 10-кратных разведений в диапазоне конечной концентрации в анализируемой смеси от 10 мкг/мл до 0,00001 мкг/мл. Значения IC50 определяли с использованием программного обеспечения Softmax (Molecular Devices). Результаты представлены в таблице 1 и показывают, что mAb 271.26.6.6.1, 271.33.1.2.2 и 271.39.4.6.5 представляют собой наиболее эффективные нейтрализующие mAb и обладают сходной эффективностью в этом тесте in vitro. Интересно, что mAb 271.33.3.2.1 было значительно менее эффективным против варианта c108smIL-31 mIL-31, чем другие mAb, подтверждая выводы исследований по сортировке по эпитопам, что это антитело по всей вероятности обладает отличающейся от других четырех mAb специфичностью в отношении эпитопа. Тот факт, что это mAb было также значительно менее эффективным в отношении варианта c108smIL-31 mIL-31 по сравнению с вариантом mIL-31-CEE подтверждает, что эпитоп, распознаваемый 271.33.3.2.1, может частично включать в себя участок гликозилирования на молекуле IL-31.
Активность MAb крысы против IL-31 мыши в клеточной нейтрализации in vitro
Пример 2
Получение mAb мыши против IL-31 человека
A. Иммунизация и скрининг сыворотки мышей
Две группы самок мышей BALB/c в возрасте от шести до восьми недель, по пять животных в каждой, иммунизировали посредством IL-31 человека. Мышей в группе 1 иммунизировали посредством внутрибрюшинной инъекции очищенного рекомбинантного IL-31 человека, слитого на C-конце с пептидом, состоящим из последовательности EYMPME (SEQ ID NO:7), (в дальнейшем в настоящем описании обозначаемым как IL-31-CEE) в сочетании с адъювантом Ribi в соответствии с инструкциями изготовителя. Этот белок продуцировали в клетках BHK. Мышей в группе 2 аналогичным образом иммунизировали очищенной рекомбинантной мутантной формой IL-31 человека, в которой остаток цистеина в положении 108 превращен в остаток серина (в дальнейшем в настоящем описании обозначаемой как c108sIL-31). Этот материал продуцировали в E. coli без пептида C-EE. Всем мышам вводили 50 мкг белка каждые две недели в течение 10 недель. От семи до десяти суток после третьей и четвертой иммунизаций, у мышей через ретроорбитальное сплетение отбирали кровь и от крови отделяли сыворотку для анализа ее способности ингибировать связывание и последующую стимулирующую активность IL-31 человека в отношении клеточной линии, трансфицированной IL-RA человека.
Способность отдельных антисывороток ингибировать IL-31 человека оценивали с использованием люциферазного анализа нейтрализации с использованием IL-31-CEE или c108sIL-31 и клеток BaF3/KZ134/IL-31RA/OSMRbeta (смотрите примеры 3 и 4) со следующими модификациями. Отдельные антисыворотки титровали по два образца посредством серийных 4-кратных разведений в 96-луночном белом планшете с плоским дном из полистирола (Corning/Costar 3917), начиная с исходного разведения антисывороток 1:250 в буфере для анализа (RPMI 1640, 10% ЭТС, 1 мМ пируват натрия, 2 мМ L-глутамин, 100 Ед/мл пенициллина G натрия, 100 мкг/мл стрептомицина сульфата). В целях нормализации до некоторой степени стимулирующего эффекта сыворотки мыши в этом анализе, все разведения, за исключением исходного разведения 1:250 проводили в буфере для анализа с 0,2% нормальной сывороткой мышей BALB/c. Объем разведенных антисывороток в каждой лунке составлял 100 мкл. Клетки промывали 1,5 раза в буфере для анализа, доводили до концентрации 3·106/мл, объединяли либо с IL-31-CEE (100 пг/мл), либо с c108sIL-31 (30 пг/мл), а затем смесь добавляли в планшеты, 100 мкл/лунка. Общий объем для анализа составлял 200 мкл/лунка и состоял из 30000 клеток, IL-31 в конечной концентрации 50 пг/мл (IL-31-CEE) или 15 пг/мл (c108sIL-31) и антисывороток в конечном разведении 1:500, 1:2000, 1:8000, 1:32000, 1:128000, 1:512000, 1:2048000 и 1:8192000. Планшеты инкубировали при 37°C, 5% CO2, в течение 16-24 часов, после чего их центрифугировали при 1250 об/мин в течение 5 минут, среду осторожно стряхивали и в каждую лунку добавляли 25 мкл 1× буфера для лизиса клеток. Планшеты осторожно встряхивали в течение 10 минут для обеспечения лизиса клеток, после чего в каждую лунку добавляли 40 мкл субстрата для анализа люцифиразы. Затем лунки оценивали на люциферазную активность (отражающую активацию посредством IL-31 репортерной конструкции STAT) на люминометре с использованием 4-секундного интеграционного интервала.
Главным образом, было выявлено, что в этом анализе антисыворотка мышей, иммунизированных посредством IL-31-CEE, эффективно ингибирует стимулирующую активность как IL-31-CEE, так и c108sIL-31, и наоборот. По этой причине, дальнейшие анализы нейтрализации проводили только с IL-31-CEE.
Мыши с наиболее высокими титрами нейтрализации использовали для серии из трех слияний, нацеленной на получение гибридом, продуцирующих моноклональные антитела, которые могут с высокой эффективностью нейтрализовать взаимодействие IL-31 с узнающим его рецептором.
B. Слияние
Слияние 291
В первом слиянии использовали клетки лимфатических узлов одной мыши, иммунизированной посредством IL-31-CEE, и одной мыши, иммунизированной посредством c108sIL-31. Каждого из этих животных иммунизировали в шестой раз посредством 15 мкг соответствующего иммуногена, разбавленного в PBS и вводимого посредством внутрисосудистой инъекции через три недели после их последней иммунизации. Через трое суток, селезенку и лимфатические узлы от этих мышей собирали, объединяли, превращали в суспензию отдельных клеток (всего 4,69·108 клеток), а затем проводили их слияние с клоном клеточной линии миеломы мыши P3-X63-Ag8.653 (Kearney, J.F. et al., J Immunol. 123:1548-50, 1979) (обозначаемым как P3-X63-Ag8.653.3.12.11) в соотношении лимфоидные клетки:клетки миеломы 2,3:1 посредством 3 мл PEG 1500 в течение 2 минут 55 секунд с использованием стандартных способов, известных в данной области (Lane, R.D. J Immunol Methods 81:223-8, 1985). Смесь для слияния распределяли в серию 96-луночных планшетов с плоским дном, подпитывали один раз посредством замены 70% среды через 4 суток и анализировали через 6 суток.
Слияние 292
Для второго слияния использовали лимфоидные клетки одной мыши, иммунизированной посредством IL-31-CEE. В дополнение к пяти внутрибрюшинным иммунизациям, упомянутым выше, этой мыши проводили внутрисосудистую инъекцию 15 мкг IL-31-CEE в PBS через три недели после внутрибрюшинной инъекции и аналогичную инъекцию через три недели после первой внутрисосудистой иммунизации. Через трое суток селезенку и лимфатические узлы подвергали обработке (всего 2,27·108 клеток) и проводили слияние с P3-X63-Ag8.653.3.12.11 в соотношении лимфоидные клетки:клетки миеломы 2:1 посредством 1,8 мл PEG 1500 в течение 2 минут 55 секунд, как описано выше. Смесь для слияния распределяли в серию 96-луночных планшетов с плоским дном, подпитывали один раз посредством замены 70% среды через 4 суток и анализировали через 5 суток.
Слияние 294
Для последнего слияния использовали лимфоидные клетки от одной мыши, иммунизированной только посредством c108sIL-31. В дополнение к пяти внутрибрюшинным иммунизациям антигеном, описанным ранее, этой мыши проводили внутрисосудистую инъекцию 15 мкг c108sIL-31 в PBS через шесть недель после последней внутрибрюшинной инъекции, а затем снова через две недели после первой внутрисосудистой инъекции. Через трое суток селезенку и лимфатические узлы подвергали обработке (всего 1,586·108 клеток) и проводили слияние с P3-X63-Ag8.653.3.12.11 в соотношении лимфоидные клетки:клетки миеломы 2:1 посредством 1,5 мл PEG 1500 в течение 2 минут 55 секунд, как описано выше. Смесь для слияния распределяли в серию 96-луночных планшетов с плоским дном, дважды проводили подпитку посредством замены 70% среды через 4 и 6 суток и анализировали через 3 суток после проведения подпитки.
C. Скрининг после слияния
После всех трех слияний проводили скрининг с использованием клеточного анализа нейтрализации IL-31, описанного выше, с двумя модификациями. Во-первых, вместо разведенной антисыворотки в планшетах для анализа, из каждой лунки планшетов для слияния на планшеты для анализа высевали 100 мкл супернатанта в качестве реплик. Во-вторых, конечная концентрация IL-31-CEE в смеси для анализа составляла 150 пг/мл.
В дополнение к анализу нейтрализации проводили скрининг нескольких планшетов после каждого слияния, выбранных случайным образом, на антитела IgG, которые могут связываться с IL-31-CEE, который предварительно адсорбировали на планшеты для ELISA из полистирола. Целью этого анализа была идентификация основных лунок, содержащих гибридому, продуцирующую антитело против IL31, которое либо плохо нейтрализует IL-31, либо совсем не нейтрализует этот цитокин. В этом анализе, лунки 96-луночных планшетов для ELISA из полистирола сначала покрывали IL31-CEE в концентрации 200 нг/мл в количестве 50 мкл/лунка в 0,1 M Na2CO3, pH 9,6. Планшеты инкубировали в течение ночи при 4°C, после чего несвязанный антиген аспирировали и планшеты дважды промывали посредством PBS-Tween (0,137 М NaCl, 0,0027 М KCl, 0,0072 М Na2HPO4, 0,0015 М KH2PO4, 0,05% об./об. полисорбат 20, pH 7,2) в количестве 300 мкл/лунка. Лунки блокировали посредством PBS-Tween + 1% BSA в количестве 200 мкл/лунка в течение 1 часа при комнатной температуре (RT). Затем блокирующий раствор стряхивали с планшетов и проводили посев 50 мкл кондиционированной среды из каждой лунки в качестве реплик в лунки планшета для анализа. Планшеты инкубировали в течение 1 часа при RT, после чего несвязанное антитело аспирировали и планшеты дважды промывали посредством PBS-Tween в количестве 300 мкл/лунка. Fc-специфичную антисыворотку с конъюгированными с HRP антителами козы против IgG мыши (Jackson Immunoresearch) разбавляли 1:5000 в PBS-Tween + 1% BSA и добавляли в лунки планшетов для анализа, 100 мкл/лунка. После инкубации в течение 1 часа при RT, несвязанное антитело второй стадии аспирировали из лунок и планшеты промывали 5 раз. Затем в каждую лунку добавляли тетраметилбензидина (TMB) (BioFX Laboratories, Owings Mills, MD) в количестве 100 мкл/лунка и планшеты инкубировали в течение 5 минут при RT. Образование цвета останавливали посредством добавления стоп-реагента TMB для 450 нм (BioFX Laboratories, Owings Mills, MD) в количестве 100 мкл/лунка и значения поглощения для лунок определяли на устройстве Molecular Devices Spectra MAX 340 при 450 нм.
Результаты анализов нейтрализации показали, что 52 и 139 лунок после слияния 291 и 292, соответственно, содержали антитело, которое ингибирует по меньшей мере 40% стимулирующей активности IL-31. Более точное определение позитивности проводили после слияния 294, где было выявлено, что 131 лунка содержала антитело, которое ингибировало по меньшей мере 70% активности IL-31. Результаты анализов ELISA по выявлению mAb, которые связываются с IL-31-CEE, показали, что в большинстве случаев, если было выявлено, что основная лунка содержит антитело IgG, которое связывается с IL-31-CEE, то она также содержит антитело, которое ингибирует IL-31-CEE в клеточном анализе нейтрализации. Однако некоторые лунки были положительными в анализе ELISA, но демонстрировали относительно слабую ингибиторную способность в клеточном анализе нейтрализации и их сохраняли для последующего анализа, как описано ниже.
Клетки гибридомы, выращенные в положительных основных лунках, выращивали в культуру в 24-луночных планшетах. Когда плотность культур из 24-луночных планшетов составляла приблизительно 4-6·105 клеток/мл, супернатант (приблизительно 1,5 мл) собирали и хранили отдельно для каждой лунки и клетки из каждой лунки замораживали.
D. Селекция и клонирование основных лунок для выделения гибридом, продуцирующих эффективные MAb, нейтрализующие IL-31
Каждый из новых супернатантов из 24 лунок повторно анализировали на способность в отношении нейтрализации IL-31 с использованием клеточного анализа нейтрализации IL-31, используемого в скрининге после слияния. Каждый супернатант анализировали неразведенным и по два образца. Результаты указывают на то, что после выращивания большая часть супернатантов из основных лунок сохранила ингибиторную активность, равную активности или лучшую чем активность, выявленная при скрининге исходных основных лунок. Для того, чтобы лучше определить, какой из новых супернатантов из основных лунок обладает наибольшей ингибиторной способностью, те супернатанты, для которых было показано приблизительно 90% или лучшее ингибирование IL-31-CEE в этом анализе, повторно анализировали посредством тестирования разведений супернатантов в клеточном анализе нейтрализации. Супернатанты титровали посредством серийных 3-кратных разведений в среде для слияния в планшетах для анализа с получением 100 мкл в каждой лунке следующих разведений: без разведения, 1:3, 1:9, 1:27, 1:81 и 1:243, и анализ проводили, как описано выше для скрининга после слияния. Несмотря на то, что в этом анализе были отчетливо идентифицированы супернатанты с наибольшей ингибиторной способностью, было невозможно определить, которые из них обладают наивысшей специфичной активностью, поскольку концентрация антитела против IL31 в супернатантах была неизвестна.
Для решения проблемы концентрации специфичного антитела, все супернатанты тестировали посредством титрования, с использованием либо серийных 2-кратных, либо 4-кратых разведений с помощью IL-31-CEE в прямом анализе ELISA, описанном выше. После нанесения на график данных в Microsoft EXCEL, определяли разведение супернатанта, приводящее к половине наблюдаемой в анализе максимальной оптической плотности (OD), которая, в свою очередь, представляет собой приблизительный показатель относительной концентрации антитела против IL-31 в супернатанте.
Объединяя данные анализа нейтрализации с данными анализа ELISA, определяли относительную специфичную активность каждого супернатанта и идентифицировали двадцать основных лунок, обладающих наивысшей специфичной активностью в отношении нейтрализации IL-31. Интересно, что все наиболее эффективные нейтрализующие IL-31 основные лунки были получены после слияния 292. В данных нейтрализации для этих основных лунок было отмечено, что, несмотря на то, что максимальное ингибирование IL-31 достигалось для каждого супернатанта без разведения и в разведении 1:3, абсолютная степень нейтрализации была немного выше для некоторых супернатантов относительно других, что отражалось в немного более низких относительных величинах люциферазных единиц. Это интерпретировали, как незначительно отличающиеся степени "полной" нейтрализации IL-31. Объединяя это наблюдение с относительной специфичной активностью в отношении нейтрализации для каждой основной лунки, список наиболее эффективных нейтрализующих основных лунок был сокращен до предполагаемых наилучших десяти. Они включали в себя 292.12, 292.39, 292.51, 292.63, 292.64, 292.72, 292.84, 292.105, 292.109 и 292.118.
Клетки из каждой из этих наилучших десяти нейтрализующих IL-31 основных лунок клонировали в целях выделения клонированной гибридомы, продуцирующей представляющие интерес нейтрализующие mAb. Клетки клонировали в 96-луночных титрационных микропланшетах для культивирования клеток с использованием подхода со стандартным разведением до низкой плотности (менее 1 клетки на лунку) и перед анализом оценивали моноклональность посредством микроскопического исследования лунок на единичные очаги роста. Среда для клонирования состояла из среды для слияния, в которой отсутствовал компонент HAT (IMDM, 10% сыворотка FC1, 2 мМ L-глутамин, 1× пенициллин/стрептомицин, 10% фактор клонирования гибридомы (Roche Applied Science)). От шести до восьми суток после посева проводили скрининг супернатантов во всех лунках посредством ELISA со связанным на планшете IL-31-CEE. Клетки из 4-6 лунок в каждой группе, в которых супернатант был высоко положительным в отношении специфичных mAb и где происходил рост только одной колонии гибридомы, выращивали в культуры в 24-луночных планшетах и полученные супернатанты повторно тестировали посредством ELISA со связанным на планшете IL-31-CEE и посредством клеточного анализа нейтрализации IL-31. Исходя из этих результатов, предполагаемый наиболее эффективный нейтрализующий клон в каждой группе повторно клонировали и проводили его скрининг посредством ELISA, как описано выше. Если 95% или более лунок с положительным ростом также были положительными в отношении специфичного mAb, дальнейшие попытки субклонирования считали необязательными и гибридомы признавали клональными. Среди полученных клонов, только для одной основной лунки, 292.64, проводили второй цикл субклонирования, необходимый для достижения требуемой процентной позитивности в отношении специфичных mAb. Клетки из 5-6 лунок в каждой конечной группе субклонов, в которых супернатант был высоко положительным в отношении специфичного mAb и где происходил рост только одной колонии гибридомы, выращивали в культуры 24-луночных планшетах. Затем каждый клон гибридомы адаптировали для роста в среде, с отсутствием фактора клонирования гибридом (IMDM, 10% сыворотка FC1, 2 мМ L-глутамин, 1× пенициллин/стрептомицин) посредством распределения клеток в последней среде с приемлемой плотностью клеток. После адаптации собирали супернатанты субклонов каждой группы и проводили их скрининг посредством ELISA со связанным на планшете IL-31-CEE. Исходя из титра с учетом клеточной плотности во время сбора супернатантов, выбирали "наилучший" конечный клон, что привело к отбору следующей группы конечных клонов: 292.12.3.1; 292.39.5.3; 292.51.5.2; 292.63.5.3; 292.64.6.5.5; 292.72.3.1; 292.84.1.6; 292.105.4.1; 292.109.4.4 и 292.118.6.4.
Изотип IgG mAb мыши, продуцируемого каждой из этих гибридом, определяли с использованием теста с моноклональным антителом мыши IsoStrip (Roche Applied Science). Было выявлено, что все mAb относятся к подклассу IgG1, за исключением 292.72.3.1 и 292.84.1.6, которые, как было показано, относятся к подклассу IgG2a.
E. Оценка in vitro специфичной нейтрализующей активности эффективных MAb, нейтрализующих IL-31
Для оценки специфичной активности эффективных mAb мыши против IL-31 человека в отношении нейтрализации in vitro, истощенный супернатант от каждого клона первого цикла повторно тестировали в клеточном анализе нейтрализации, описанном ранее. Во-первых, определяли количество IgG в каждом супернатанте посредством анализа ВЭЖХ в колонке с белком G с использованием mAb с известной концентрацией в качестве стандарта. Затем каждый супернатант разбавляли до концентрации IgG 1 мкг/мл в той же среде, что использовали для получения истощенного супернатанта. Затем разбавленные супернатанты титровали посредством 10-кратного серийного разведения в среде с получением диапазона концентраций от 1 мкг/мл до 0,0001 мкг/мл и тестировали по три образца в описанном ранее клеточном анализе нейтрализации с конечной концентрацией IL-31-CEE 300 пг/мл (18,27 пМ). Результаты анализа представлены в таблице 2, где mAb представлены в порядке от наиболее эффективного до наименее эффективного. MAb 292.84.1 и 292.12.3 были наиболее эффективными нейтрализаторами, за ними следуют mAb 292.72.3 и 292.63.5, которые оказались приблизительно в два раза менее эффективными, чем первые два. Для остальных mAb было показано, что они приблизительно в 6-9 раз менее эффективны, чем два наилучших mAb.
Оценка in vitro эффективных нейтрализующих MAb против IL31
по активности в отношении специфичной нейтрализации
Исходя из приведенных выше результатов, гибридомы 292.84.1.6, 292.12.3.1, 292.72.3.1, 292.63.5.3 и 292.118.6.4 были выбраны, как отражающие различные уровни специфичной нейтрализующей способности, для депонирования в патентной коллекции Американской коллекции типовых культур (ATCC, Manassas, VA) в качестве первоначальных образцов согласно Будапештскому договору, и им были присвоены следующие регистрационные номера ATCC №№: клон 292.12.3.1 (Патентный номер депозита в ATCC PTA-6815); клон 292.72.3.1 (Патентный номер депозита в ATCC PTA-6816); клон 292.63.5.3. (Патентный номер депозита в ATCC PTA-6829); клон 292.118.6.4 (Патентный номер депозита в ATCC PTA-6830) и клон 292.84.1.6 (Патентный номер депозита в ATCC PTA-6871).
F. Селекция и клонирование основных лунок для выделения гибридом, продуцирующих MAb со слабой нейтрализацией IL-31
При получении mAb против IL-31 человека представляло интерес выделение пар mAb, которые можно использовать в "сэндвич"-анализе для количественного определения количества IL-31 в различных жидкостях. Такие пары mAb, как правило, обладают специфичностью в отношении различных эпитопов на молекуле-мишени и предпочтительно не конкурируют перекрестно за связывание с этой молекулой. Для выделения пар-кандидатов таких mAb, была выбрана стратегия формирования пары из mAb с превосходной нейтрализацией IL-31 и mAb, обладающего небольшой, если обладающей, способностью в отношении нейтрализации, предполагая, что два таких различных функциональных антитела вероятнее всего не связывают не перекрывающиеся (пространственно разделенные) эпитопы.
Как указано ранее для скрининга после слияния, после каждого слияния для идентификации положительных в отношении антитела против IL-31 супернатантов основных лунок проводили скрининг нескольких планшетов посредством ELISA со связанным на планшете IL-31-CEE. Сравнение результатов ELISA с результатами клеточного анализа нейтрализации показало основные лунки, которые содержали антитело, которое хорошо связывается с IL-31-CEE, но не обладает высокой нейтрализацией этой молекулы в клеточном анализе нейтрализации. Как проводили с наиболее эффективными нейтрализующими основными лунками, клетки гибридомы, выращиваемые в этих основных лунках, выращивали в культуру в 24-луночных планшетах. Когда плотность культур в 24-луночных планшетах составляла приблизительно 4-6·105 клеток/мл, супернатант (приблизительно 1,5 мл) собирали и хранили для каждой лунки по отдельности и клетки из каждой лунки замораживали.
Эти новые супернатанты тестировали с использованием клеточного анализа нейтрализации (серийные 3-кратные разведения, начиная с супернатанта без разведения и продолжая до конечного разведения 1:243), а также с использованием анализа ELISA по типу "антигенной ловушки", сходного с анализом, используемым при разработке mAb крысы против IL-31 мыши со следующими отличиями: 1) Fc-специфичное антитело овцы против IgG мыши (Jackson Immunoresearch) (1 мкг/мл) в качестве антитела для покрытия планшета, 2) планшеты блокировали один раз посредством PBS-Tween + 1% BSA в течение 1 часа, 3) каждый супернатант добавляли в лунки по одному титрованию (серийные 3-кратные разведения, начиная с супернатанта без разведения и продолжая до конечного разведения 1:2187), и 4) добавление биотинилированного IL-31-CEE (молярное соотношение биотин:белок 3:1), 200 нг/мл. Полагали, что этот анализ является более пригодным, чем связывание антитела со связанным на планшете IL-31-CEE, поскольку в нем оценивали способность антитела связываться с IL-31-CEE в растворе, что является свойством, необходимым для хорошего антитела для ELISA по типу "сэндвич". В действительности было выявлено, что ряд супернатантов основных лунок, содержащих антитело, которое хорошо связывается со связанным с планшетом IL-31-CEE, слабо распознавали эту молекулу в растворе.
Для выбора лунок, из которых клонировали соответствующие секретирующие антитело гибридомы, было идентифицировано 10 лунок (291.78, 292.152, 292.154, 294.35, 294.144, 294.146, 294.154, 294.155, 294.158 и 294.163), супернатанты которых слабо нейтрализовали IL-31-CEE в анализе нейтрализации (менее чем 50%, но предпочтительно только 1-20%), а также относительно хорошо связывались с IL-31-CEE в растворе (как определено по OD выше чем 2,0 при концентрации супернатанта без разведения). Клонирование, скрининг клонов и селекцию наилучшего клона первого цикла проводили, как описано выше для лунок с эффективной нейтрализацией.
Для снижения количества гибридом, выбранных для дальнейшего субклонирования, проводили два новых анализа с истощенным супернатантом каждого из выбранных клонов первого цикла. Первым способом оценки была попытка разделить mAb на группы на основе специфичности к эпитопу ("сортировки") с использованием устройства для поверхностного плазмонного резонанса Biacore 3000. Один из супернатантов с хорошим нейтрализующим mAb (292.64.6) и 10 более слабых супернатанов клонов первого цикла анализировали друг относительно друга, что привело к идентификации четырех предполагаемых "серий". Серия 1 состояла только из хорошего нейтрализующего антитела. Серия 2 состояла из 292.152.4, 292.154.4, 294.35.2, 294.146.5 и 294.154.5. Серия 3 включала в себя 291.78.4, 294.155.6, 294.158.5 и 294.163.2. Серия 4 содержала только 294.144.3. Второй анализ включал в себя повторение ELISA по типу "антигенной ловушки", только в этот раз, вследствие более высокой концентрации специфичного антитела в супернатантах по сравнению с исходными супернатантами основных культур в 24-луночных планшетах, получили более полную кривую титрования и имели возможность определения EC50 для связывания антитела с IL-31-CEE. Результаты представлены в таблице 3 (совместно с результатами сортировки и % ингибированием активности в отношении IL-31-CEE в клеточном анализе нейтрализации), и они указывают на широкий диапазон значений EC50 (и, как следствие, аффинности mAb для IL-31-CEE) для различных mAb.
Суммарные данные для супернатанта культуры клона первого цикла из менее эффективно нейтрализующих IL-31 гибридом
С учетом селекции гибридом, продуцирующих mAb, которые 1) находились в сериях по эпитопам, отличающихся от серии mAb с хорошей нейтрализацией (292.64.6) и 2) обладали наивысшей аффинностью, как показано по низкой EC50 в анализе по типу "антигенной ловушки", только клонам первого цикла 294.35.2, 294.144.3, 294.154.5 и 294.163.2 был присвоен статус конечного клона, как описано выше для более эффективных нейтрализующих mAb. Эта дальнейшая работа по субклонированию привела к селекции следующей группы конечных клонов: 294.35.2.6.3; 294.144.3.5; 294.154.5.6 и 294.163.2.1.
Изотип IgG mAb мыши, продуцируемого каждой из этих гибридом, определяли с использованием теста моноклонального антитела мыши IsoStrip (Roche Applied Science). Было выявлено, что все четыре mAb относятся к подклассу IgG1. Образцы каждой из четырех гибридом были депонированы в патентной коллекции Американской коллекции типовых культур (ATCC, Manassas, VA) в качестве первоначальных образцов согласно Будапештскому договору, и им были присвоены следующие регистрационные номера ATCC №№: клон 294.163.2.1 (Патентный номер депозита в ATCC PTA-6831); клон 294.35.2.6.3 (Патентный номер депозита в ATCC PTA-6872); клон 294.154.5.6 (Патентный номер депозита в ATCC PTA-6875) и клон 294.144.3.5 (Патентный номер депозита в ATCC PTA-6873).
G. Перекрестная реакция mAb мыши против IL-31 человека с IL-31 мыши
Все супернатанты клонов первого цикла для десяти эффективных нейтрализующих mAb, а также десяти менее эффективных нейтрализующих mAb тестировали на их способность распознавать IL-31 мыши посредством ELISA со связанными на планшете mIL-31-CEE. Не было выявлено перекрестной реактивности с какими-либо mAb, что указывает на то, что mAb, по-видимому, обладают специфичностью в отношении IL-31 человека. Более того, эти результаты показали, что ни одно из антител не распознавало пептидный маркер CEE, общий для C-конца рекомбинантных молекул IL-31 как человека, так и мыши, используемых в этих исследованиях, и это определение не проводили для супернатантов исходных лунок.
Пример 3
Люциферазный анализ IL-31RA/рецептор OSMRbeta
Плазмиду KZ134 конструировали с комплементарными олигонуклеотидами, которые содержат связывающие транскрипционный фактор элементы STAT из 4 генов, которые включают в себя модифицированный индуцибельный элемент Sis c-fos (m67SIE или hSIE) (Sadowski, H. et al., Science 261:1739-1744, 1993), p21 SIE1 из гена p21 WAF1 (Chin, Y. et al., Science 272:719-722, 1996), отвечающий элемент молочной железы гена β-казеина (Schmitt-Ney, M. et al., Mol. Cell. Biol. 11:3745-3755, 1991) и индуцибельный элемент STAT гена Fcg RI (Seidel, H. et al., Proc. Natl. Acad. Sci. 92:3041-3045, 1995). Эти олигонуклеотиды содержат совместимые с Asp718-XhoI концы, и их лигировали с использованием стандартных способов в реципиентный репортерный вектор с люциферазой светляка с промотором c-fos (Poulsen, L.K. et al., J. Biol. Chem. 273:6229-6232, 199S), расщепленным указанными ферментами и содержащим селективный маркер неомицина. Плазмиду KZ134 использовали для стабильной трансфекции клеток BaF3 с использованием стандартных способов трансфекции и селекции с получением клеточной линии BaF3/KZ134.
Получали стабильную индикаторную клеточную линию BaF3/KZ134, экспрессирующую полноразмерный IL-31RA или IL-31RA/рецептор OSMRbeta. Клоны разбавляли, высевали и проводили селекцию с использованием стандартных способов. Скрининг клонов проводили посредством анализа люциферазы (смотрите B, ниже) с использованием кондиционированной среды с IL-31 человека или очищенного белка IL-31 в качестве индуктора. Отбирали клоны с наивысшим люциферазным ответом (через люцифнеразу STAT) и с наименьшим фоном. Отбирали стабильно трансфицированные клеточные линии. Клеточные линии назвали BaF3/KZ134/IL-31RA или BaF3/KZ134/IL-31RA/OSMRbeta в зависимости от рецепторов, трансфицированных в клеточную линию.
Аналогично, также получали клеточные линии BHK с использованием способа, описанного в настоящем описании, и их использовали в анализах люциферазы, описанных в настоящем описании. Клеточные линии назвали BHK/KZ134/IL-31RA или BHK/KZ134/IL-31RA/OSMRbeta в зависимости от рецепторов, трансфицированных в клеточную линию.
Клетки BaF3/KZ134/IL-31RA и BaF3/KZ134/IL-31RA/OSMRbeta центрифугировали и промывали не содержащей mIL-3 средой. Клетки центрифугировали и промывали 3 раза, чтобы обеспечить удаление mIL-3. Затем клетки подсчитывали на гемоцитометре. Клетки высевали в 96-луночный планшет в количестве 30000 клеток на лунку в объеме 100 мкл на лунку с использованием не содержащей mIL-3 среды. Тот же процесс использовали для нетрансфицированных клеток BaF3/KZ134 для применения в качестве контроля в последующем анализе. Клетки BHK/KZ134/IL-31RA или BHK/KZ134/IL-31RA/OSMRbeta высевали в 96-луночный планшет в количестве 15000 клеток на лунку в 100 мкл среды. Родительские клетки BHK/KZ134 использовали в качестве контроля.
Активацию STAT клеток BaF3/KZ134/IL-31RA, BaF3/KZ134/IL-31RA/OSMRbeta, BHK/KZ134/IL-31RA или BHK/KZ134/IL-31RA/OSMRbeta оценивают с использованием кондиционированной среды или очищенного белка. К клеткам BaF3/KZ134/IL-31RA, BaF3/KZ134/IL-31RA/OSMRbeta, BHK/KZ134/IL-31RA или BHK/KZ134/IL-31RA/OSMRbeta добавляют сто микролитров разведенной кондиционированной среды или белка. Анализ с использованием кондиционированной среды одновременно проводят на нетрансфицированных клетках BaF3/KZ134 или BHK/KZ134 в качестве контроля. Общий анализируемый объем составляет 200 мкл. Планшеты для анализа инкубируют при 37°C, 5% CO2 в течение 24 часов, а затем клетки BaF3 осаждают центрифугированием при 2000 об/мин в течение 10 мин, и среду аспирируют и добавляют 25 мкл буфера для лизиса (Promega). Для клеточных линий BHK, стадия центрифугирования не является необходимой, поскольку они представляют собой прикрепляющиеся клеточные линии. Через 10 минут при комнатной температуре в планшетах измеряют активацию репортерной конструкции STAT посредством считывания на люминометре (Labsystems Luminoskan, модель RS), в котором происходит добавление 40 мкл субстрата для анализа люциферазы (Promega) с интеграцией в течение пяти секунд.
Пример 4
Анализ люциферазы на трансформированных эпителиальных клеточных линиях человека посредством временной инфекции аденовирусным репортерным геном STAT/SRE
Ингибирование, снижение и/или нейтрализацию активности IL-31 можно определять посредством анализа люциферазы. Например, можно проводить посев трансформированных клеточных линий человека в 96-луночные планшеты с плоским дном в количестве 10000 клеток/лунка в обычной среде для роста, как описано для каждого типа клеток. На следующие сутки клетки инфицируют репортерной конструкцией аденовируса, KZ136, при множественности заражении 5000. В дополнение к отвечающему элементу сыворотки репортер KZ136 содержит элементы STAT. Общий объем составляет 100 мкл/лунка с использованием DMEM, дополненной 2 мМ L-глутамином (GibcoBRL), 1 мМ пируватом натрия (GibcoBRL) и 1× добавкой инсулин-трансферрин-селен (GibcoBRL) (в дальнейшем в настоящем описании называемой не содержащей сыворотки средой). Клетки культивируют в течение ночи.
На следующие сутки среду удаляют и заменяют 100 мкл среды для индукции. Среда для индукции представляет собой IL-31 человека, разведенный в не содержащей сыворотки среде в концентрации 100 нг/мл, 50 нг/мл, 25 нг/мл, 12,5 нг/мл, 6,25 нг/мл, 3,125 нг/мл и 1,56 нг/мл. Положительный контроль в виде 20% ЭТС используют для проверки анализа и для подтверждения того, что инфекция аденовирусом является успешной. Клетки индуцируют в течение 5 часов, после чего среду аспирируют. Затем клетки промывают в PBS в количестве 50 мкл/лунка, а затем лизируют в 1× буфере для лизиса клеток (Promega) в количестве 30 мкл/лунка. После инкубации в течение 10 минут при комнатной температуре, лизат в количестве 25 мкл/лунка переносят в непрозрачные белые 96-луночные планшеты. Затем проводят считывание планшетов на люминометре с использованием 5-секундной интеграции с инъекцией субстрата люциферазы (Promega) в количестве 40 мкл/лунка.
Пример 5
Ингибирование продукции цитокинов посредством антител против IL-31
Было показано, что IL-31 стимулирует продукцию IL-6 в DU145 (клеточные линии при заболевании и нормальные клеточные линии); а также стимулирует клеточные линии A549 (при заболевании и нормальные) со стимуляцией продукции IL-8, и снижает продукцию IL-8 в клеточных линиях U2OS (при заболевании и нормальных). Смотрите опубликованную патентную заявку США (смотрите публикацию номер 20030224487, Sprecher, Cindy et al, 2003). По существу, активность моноклональных антител против IL-31, описанных в настоящем описании, можно измерить по ингибированию, снижению и/или нейтрализации активности IL-31 в этих эпителиальных клеточных линиях с заболеванием.
A. Ингибирование продукции цитокинов эпителиальных клеточных линий с заболеванием, культивируемых с IL-31 человека
Клетки высевают в 6-луночный планшет (Costar) с плотностью 4,5·105 клеток на лунку и культивируют в соответствующей среде для роста. Клетки культивируют с тестируемыми реагентами; 100 нг/мл IL-31 (с и без антитела), 10 нг/мл интерферона-гамма (IFN-гамма) (R&D Systems, Minneapolis, MN), 10 нг/мл фактора некроза опухоли альфа (TNF-альфа) (R&D Systems, Minneapolis, MN), 10 нг/мл IL-1beta (R&D Systems, Minneapolis, MN) или 100 мкг/мл липополисахарида (LPS) (Sigma). Супернатанты собирают через 24 и 48 часов и оценивают на цитокины: GM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор), IL-1b, IL-6, IL-8, MCP-1 (Хемоаттрактантный белок макрофагов-1) и TNFa. Для измерения цитокинов в образцах используют мультиплексные наборы антител с гранулами из BioSource International (Camarillo, CA). Считывание для анализа проводят на устройстве Luminex-100 (Luminex, Austin, TX) и данные анализируют с использованием программного обеспечения MasterPlex (MiraiBio, Alameda, CA). Определяют продукцию цитокинов (пг/мл) для каждой клеточной линии в образцах после 24 часов культивирования.
B. Ингибирование продукции цитокинов нормальными эпителиальными клеточными линиями человека, культивируемыми с IL-31 человека
В 24-луночный планшет высевают клетки с плотностью 1·105 клеток на лунку и культивируют с тестируемыми реагентами; 1000 нг/мл, 100 нг/мл и 10 нг/мл IL-31 (с антителом и без него), 10 нг/мл TNFa, 10 нг/мл OSM, 10 нг/мл IFNa, 10 нг/мл TGFb или 10 нг/мл лимфотактина. Супернатанты собирают через 24 и 48 часов и оценивают на цитокины: IL-6, IL-8, MCP-1, MIP-1a, RANTES и эотаксин. Цитокины анализируют, как описано ранее. Определяют продукцию цитокинов (пг/мл) для каждой клеточной линии в образцах после 48 часов культивирования.
C. Ингибирование продукции цитокинов клеточными линиями человека с заболеванием совместно культивируемыми с IL-31 и IFN-гамма человека
Клетки высевают в 24-луночный планшет с плотностью 2·105 клеток на лунку и совместно культивируют с 10 нг/мл IFN-гамма +/- IL-31 в концентрации 100 нг/мл, 10 нг/мл или 1 нг/мл. Супернатанты собирают через 24 и 48 часов и оценивают f (с антителом и без него), или IL-8 и MCP-1. Определяют продукцию цитокинов (пг/мл) для каждой линии в образцах после 24 часов культивирования.
Пример 6
Ингибирование эффектов IL-31 на адгезию моноцитов U937 к монослою трансформированных эндотелиальных клеток костного мозга (TRBMEC)
Было показано, что IL-31 может влиять на исходную прикрепляемость клеток U937 к эндотелиальным монослоям. В частности, IL-31 обладает синергичным эффектом с TNF-альфа и дополнительно усиливает адгезию U937. Смотрите опубликованную патентную заявку США (Смотрите публикацию номер 20030224487, Sprecher, Cindy et al., 2003). Таким образом, ингибирование IL-31 антителами против IL-31, описанными в настоящем описании, можно определять с помощью следующего анализа.
Трансформированные эндотелиальные клетки костного мозга (TRBMEC) высевают в 96-луночные тканевые кластеры (Falcon) с плотностью 25000/лунка в среде M131 (Cascade Biologies), дополненной добавкой для роста микрососудов (MVGS) (Cascade Biologies). При слиянии монослоя (через 24 часа), клетки переключают на M199 (Gibco-Life Technologies), дополненную 1% эмбриональной телячьей сывороткой (Hyclone). Добавляют рекомбинантный IL-31 человека (тестируемый реагент) в различных концентрациях (от 0,4 до 10 нг/мл) с антителом или без него, для тестирования эффекта белка на взаимодействие иммунные клетки-эндотелиальные клетки, приводящее к адгезии. В некоторые лунки добавляли 0,3 нг/мл фактора некроза опухоли (TNF-альфа R&D Systems), известного провоспалительного цитокина, в дополнение к IL-31, для тестирования эффекта белка на эндотелиальные клетки в условиях воспаления. TNF-альфа в концентрации 0,3 нг/мл отдельно использовали в качестве положительного контроля и используемая концентрация отражает приблизительно 70% от максимального эффекта TNF-альфа в этой системе, т.е. она не индуцирует максимального прикрепления клеток U937 (подобная моноцитам человека клеточная линия) к эндотелию. По этой причине, в этих условиях можно выявлять как положительную, так и отрицательную регуляцию эффектов TNF-альфа. Базальные уровни адгезии как с TNF-альфа, так и без него, используют в качестве исходного уровня для оценки эффектов тестируемых реагентов.
После инкубации клеток эндотелия с тестируемыми реагентами в течение ночи, клетки U937, окрашенные посредством 5 мкМ флуоресцентного маркера кальцеин-AM (Molecular Probes), суспендируют в RPMI 1640 (без фенолового красного), дополненной 1% ЭТС, и высевают в количестве 100000 клеток/лунка на промытый монослой TRBMEC. Через 30 минут измеряют уровни флуоресценции при длинах волн возбуждения/испускания 485/538 нм (устройство для считывания микропланшетов Molecular Devices, CytoFluor application), до и после промывания лунки три раза теплой RPMI 1640 (без фенолового красного), для удаления неприкрепившихся U937. Флуоресценцию до промывания (общую) и после промывания (специфичную для прикрепления) используют для определения процентного прикрепления (значение для прикрепившихся/значение для общих · 100 = % прикрепление).
Пример 7
Протокол биологического анализа IL-31
Клетки BAF3, трансфицированные посредством hzCYTOR17 (IL-31RA), hOSMRB и KZ134 выращивают до 5·105 и 1·106 клеток/мл. Клетки промывают средой для анализа (RPMI 1640, 10% ЭТС, L-глутамин, пируват натрия, и Pen/Strep (все от Gibco)) и ресуспендируют в количестве 3·105 клеток/мл в среде для анализа. В 96-луночном непрозрачном планшете титруют стандарты hIL-31 по два образца от 600 пг/мл до 9,38 пг/мл в среде для анализа посредством серийных разведений 1:2, 100 мкл/лунка. В планшет добавляют стандарты для контроля качества по два образца в концентрации 350 пг/мл и 35 пг/мл в 100 мкл. Тестируемые образцы многократно разбавляют 1:2 или 1:4 и добавляют по два образца в лунки с образцами. В каждую лунку добавляют 100 мкл промытых клеток BAF3 до конечной концентрации 3·104 клеток/лунка. Планшет инкубируют в течение 16-24 часов при +37°C в инкубаторе с 5% CO2. Планшет центрифугируют при 1200 об/мин в течение 5 минут, среду стряхивают и в каждую лунку добавляют буфер для лизиса в количестве 25 мкл/лунка (Promega). Через 10 минут проводят считывание планшета на люминометре (Berthold). Люминометр добавляет смесь субстрата для люциферазы в количестве 40 мкл/лунка (Promega) и интегрирует люминесценцию в течение 4 секунд. Показатели люминесценции передаются в электронную таблицу, где происходит их анализ и перевод в пикограммы IL-31 на 106 клеток на мл объема.
Пример 8
Участие IL-31 в запуске и поддержании контактной гиперчувствительности in vivo
Способ I
Выбритую средне-заднюю часть спины мышей BALB/c окрашивают посредством 25 мкл 0,5% DNFB (2,4-динитрофторбензол, Sigma, St. Louis MO), растворенным в растворе ацетон:оливковое масло (4:1) с использованием пипеттора. В контрольной группе с носителем вводят только 25 мкл смеси ацетон:оливковое масло. Через 5 суток мышам проводят анестезию посредством изофлурана в ингаляционной камере и ушные раковины как экспериментальных, так и контрольных животных измеряют посредством инженерного микрометра (Mitutoyo) с получением исходных значений. Затем мышей иммунизируют посредством нанесения 10 мкл 0,25% DNFB в смеси ацетон:оливковое масло (4:1) на обе стороны каждого уха всех мышей. Контактную гиперчувствительность измеряют через 24 ч и 48 ч как разницу между правым ухом (иммунизированным) и левым ухом (неиммунизированным). Все измерения проводят с помощью инженерного микрометра. Фоновые значения определяют из разницы в увеличении уха между иммунизированными и неиммунизированными ушами не подвергавшихся воздействию мышей.
Перед умерщвлениями проводят забор цельной крови и сыворотки для анализа FACS и/или ELISA и уши забирают для гистологии.
Способ II (Индукция Th2-ответов)
Выбритую средне-заднюю часть спины мышей BALB/c окрашивают посредством 100 мкл 0,5% FITC (флуоресцеин-изотиоцианат) в растворе ацетон/дибутилфталат 1:1 (доступный MSDS) с использованием пипеттора на 1, 2 и 8 сутки. На 13 сутки мышам проводят анестезию посредством изофлуорана в ингаляционной камере и обе ушные раковины экспериментальных животных измеряют посредством инженерного микрометра (Mitutoyo) с получением исходных значений. Мышей иммунизируют посредством нанесения 25 мкл 0,5% FITC (в смеси ацетон/дибутилфталат 1:1) на дорсальную поверхность каждого уха. Контактную гиперчувствительность определяют через 24 ч и 48 ч, в качестве различия между правым ухом (иммунизированным) и левым ухом (неиммунизированным). Все измерения проводят с помощью инженерного микрометра. Фоновые значения определяют по различию в увеличении уха между иммунизированными и неиммунизированными ушами не подвергавшихся воздействию мышей. Перед умерщвлениями проводят забор цельной крови и сыворотки для анализа FACS и/или ELISA и уши забирают для гистологии.
Способ III (индукция Th1-ответов)
Выбритую средне-заднюю часть спины мышей BALB/c окрашивают посредством 25 мкл 2% оксазалона (в смеси ацетон/оливковое масло 4:1) с использованием пипеттора. На 7 сутки мышам проводят анестезию посредством изофлуорана в ингаляционной камере и ушные раковины как экспериментальных, так и контрольных животных измеряют посредством инженерного микрометра (Mitutoyo) с получением исходных значений. Мышей иммунизируют посредством нанесения 8 мкл оксазалона на дорсальную поверхность каждого уха. Контактную гиперчувствительность измеряют через 24 ч и 48 ч как разницу между правым ухом (иммунизированным) и левым ухом (неиммунизированным). Все измерения проводят с помощью инженерного микрометра. Фоновые значения определяют из разницы в увеличении уха между иммунизированными и неиммунизированными ушами не подвергавшихся воздействию мышей. Перед умерщвлением проводят забор цельной крови и сыворотки для анализа FACS и/или ELISA и уши забирают для гистологии.
Участие IL-31 в запуске и поддержании контактной гиперчувствительности тестируют с использованием антител, описанных в настоящем описании, против IL-31, как в фазу эксперимента сенсибилизации, так и иммунизации.
Пример 9
Участие IL-31 в атопическом дерматите in vivo
Способы I (Сенсибилизация мышей NC/Nga)
Самцов мышей NC/Nga приобретали из CRL Japan. При доставке возраст мышей составлял 4 недели и их помещали в условия карантина SPF на 4 недели для акклиматизации. К началу сенсибилизации антигеном возраст мышей составлял приблизительно 10-11 недель. Мышам проводили анестезию посредством изофлуорана и их спины выбривали посредством электростригальных машин. Приблизительно 10 мкг экстракта Dermatophagoides pteronyssinus (Dp) (Indoor Biotechnologies, специальный заказ) инъецировали подкожно в заднюю часть шеи 3 раза в неделю в течение от 5 до 6 недель до тех пор, пока у мышей не развились повреждения на коже. Контрольным животным проводили внутрикожные инъекции 10 мкл PBS 3 раза в неделю. Экстракт Dp получали в соответствии со способом Matsuoka и коллег. Matsuoka H., et al., Allergy: 58, 139 (2003). В кратком изложении, 595 мг лиофилизированного экстракта истощенной культуры Dp растворяли в 12 мл стерильного PBS (Gibco). Dp перемешивали в 50-мл пробирке Falcon на качалке для встряхивания в течение 30 минут. Экстракт центрифугировали в течение 10 минут при 2000 об/мин и супернатант собирали и распределяли на аликвоты в 1-мл криопробирки и хранили при -20°C.
Способы II (сенсибилизация мышей DO11.10)
Трансгенных мышей DO11.10 выводили из собственной колонии, и к началу сенсибилизации антигеном их возраст составлял между 9,5 и 14 неделями. За 24 часа до накожной сенсибилизации мышам проводили анестезию изофлуораном и все туловище (спину и живот) мышей выбривали с помощью электростригальной машины. Затем мышам обнажали полосу кожи посредством киперной ленты Elastin (Johnson and Johnson) на спине. 1 см2 стерильных марлевых лоскутов увлажняли либо посредством 500 мкг овальбумина (Calbiochem 32467), либо стерильного PBS (Gibco) и прикрепляли к задней части мышей слева посредством повязки DuoDerm Extra Thin Dressing (ConvaTec 187932). Затем лоскут и повязку покрывали манжетой из киперной ленты Elastin, так что мыши не могли удалить или повредить лоскуты. Лоскуты держали в течение 7 суток и удаляли. Мышей повторно тестировали за две недели до начала следующего цикла накожной сенсибилизации. Всего мышам проводили три сенсибилизации в течение одной недели.
Результаты
Иммуногистохимический анализ экспрессии IL-31RA в поврежденной и неповрежденной коже из сенсибилизированных частицами пылевого клеща NC/Nga и сенсибилизированных OVA животных DO11.10 показал, что у мышей IL-31RA экспрессируется кератиноцитами эпидермиса, однако нельзя выявить значимого различия в уровнях экспрессии между сенсибилизированными антигеном животных по сравнению с сенсибилизированными посредством PBS животными.
Пример 10
Ингибирование зуда посредством введения антител крысы против IL-31 мыши
В другом исследовании на мышах NC/Nga, к самцам мышей NC/Nga (SLC, Tokyo) в возрасте 4 недель обеспечивали доступ мышей NC/Nga, у которых уже проявлялся дерматит, от мягкого до умеренного. Через семь недель мышей подразделяли на три группы и проводили следующие введения. Каждые пятые сутки в течение семи недель мышам в одной группе вводили антитела крысы против IL-31 мыши в количестве 10 мг/кг; в одной группе проводили инъекции сывороточного альбумина мыши; и в третьей группе не проводили никакого введения. Мышей оценивали клинически дважды в неделю на тяжесть повреждений кожи. Конкретно, оценивали поражение кожи (0= нет поражения; 1= поражение спины; 2= спины и морды; 3= спины, морды и ушей) и тяжесть повреждений (0= нормальная кожа; l= отрубевидная и сухая; 2= узелковые повреждения; 3= геморрагические повреждения). Кроме того, оценивали расчесывание. Расчесывание у мышей пальцами их задних лап автоматически определяли и объективно оценивали с использованием Micro Act (Neuroscience, Tokyo, Japan). Небольшой покрытый тефлоном магнит имплантировали подкожно в дорзальную сторону обеих задних лап мышей при анастезии посредством смеси кеталар/ксилазин. Мышей с магнитами помещали в камеру для наблюдения, окруженную круглыми катушками. Электрический ток, индуцированый посредством движения магнитов, усиливался и записывался. В программе для анализа использовали следующие условия для регистрации расчесывания: Порог (В) 0,1; интервал между событиями (с) 0,2; максимальная частота (Гц) 20,0; минимальная частота (Гц) 2,0 и минимальная длительность (с) 1,5. Следует отметить: в модели на мышах NC/Nga, у мышей со специфичным для дерматита ощущением зуда связано продолжительное (>1,5 с), но не кратковременное расчесывание (0,3-1,5 с). В целом, основными измеряемыми показателями были расчесывание, дерматит и масса тела.
Результаты
Учитывая весь период, лечение антителом крысы против IL-31 мыши не изменило основных показателей. Однако в дополнительном анализе было выявлено значительное снижение расчесывания посредством введения антитела против IL-31 в период времени между 22-43 сутками. В этот период времени, как и для всего периода, введение антител против IL-31 не оказало влияния на развитие очагов повреждения, подобных очагам при атопическом дерматите, и не нормализовало набора массы тела у больных животных, возможно вследствие замедленного начала клинических проявлений или нейтрализующих аутоантител в конце лечения. В этом эксперименте, расчесывание, но не дерматит, было уже повышенным среди большинства животных к моменту времени, когда начинали введение антитела.
Пример 11
Участие IL-31 в ГЗТ in vivo
Способы
Для получения ответа ГЗТ, мышей сенсибилизировали к антигену на 0 сутки посредством подкожной иммунизации в основание чвостапосредством 100 мкг овальбумина (OVA) в полном адъюванте Фрейнда (CFA, общий объем 50-100 мкл). Через одну неделю мышам проводили анестезию изофлуораном в ингаляционной камере и обе ушные раковины экспериментальных и контрольных животных измеряли посредством инженерного микрометра (Mitutoyo) с получением исходных измерений. Мышей иммунизировали внутрикожно посредством 10 мкг OVA в PBS в общем объеме 10 мкл в левую ушную раковину, непосредственно под кожу, не попадая в какие-либо вены. В качестве контроля мышам также инъецировали 10 мкл PBS в правую ушную раковину. В некоторых случаях, в отдельной контрольной группе, в которой проводили внутрикожную инъекцию OVA в ухо, также могли вводить местные кортикостероиды в качестве положительного контроля ингибирования реакции. Через 24 и 48 ч после иммунизации мышам проводили анестезию и измеряли толщину уха. Результаты выражали в виде: специфичное увеличение уха = (измерение, соответствующее 24 ч, - измерение, соответствующее 0 ч) для экспериментального уха - (измерение, соответствующее 24 ч, - измерение, соответствующее 0 ч) для уха отрицательного контроля. Индурация, признак ГЗТ, выявляется через 18 часов после инъекции сенсибилизирующего антигена и является максимальной через 24-48 часов. Латентный период при появлении пальпируемой индурации является причиной названия этого ответа ответом "замедленного типа".
Результаты
Трансгенных мышей по IL-31 тестировали в отношении ГЗТ, однако вследствие повышения толщины уха неиммунизированных трансгенных по IL-31 животных, нельзя было выявить статистически значимых различий в ГЗТ между Tg по IL-31 животными и контролями дикого типа. Животные с нокаутом рецептора IL-31 также тестировали в ответе ГЗТ, и нельзя было выявить значимых отличий в ответе ГЗТ между животными с нокаутом рецепторов и животными дикого типа.
Пример 12
Участие IL-31 в индукции ответа в виде зуда
Способы I (Введение капсаицина мышам после введения IL-31)
Животным BALB/c в возрасте десяти недель (CRL) проводили анестезию и инъецировали анальгезирующее вещество продолжительного действия, бупранорфина гидрохлорид, подкожно в количестве 0,1 мг/кг, а затем инъецировали 0,25 мл раствора капсаицина в концентрации 4 мг/мл в 10% этаноле + 10% Tween-80 в физиологическом растворе подкожно в заднюю часть шеи. Анестезия продолжалась у животных в течение по меньшей мере 30 мин после введения нейротоксина. Через сорок восемь часов подкожно имплантировали 14-суточные осмотические насосы для непрерывной доставки IL-31 в количестве 20 мкг/сутки в течение 14 суток. Мышам проводили мониторинг каждые сутки в течение 6 суток на алопецию и зуд с использованием следующих критериев: 0 = нет расчесывания, животное выглядит нормально, 1 = истончение шерсти на небольших областях, отмечается расчесывание, 2 = небольшая потеря шерсти (небольшие участки), расчесывание, 3 = умеренная потеря шерсти, расчесывание и 4 = тяжелая потеря шерсти, чрезмерное расчесывание.
Результаты показали, что в то время как у мышей, которым не вводили капсаицин, было показано среднее значение расчесывания/потери шерсти 2,625 через трое суток доставки IL-31, у мышей, которым вводили капсаицин, было показано значительно более низкое значение, составляющее 1. Таким образом, у мышей, которым вводили капсаицин перед доставкой IL-31, было показано как замедление встречаемости расчесывания и потери волос, так и более низкое значение интенсивности расчесывания и потери волос через шесть суток эксперимента. Эти данные позволяют предположить, что IL-31 индуцирует некоторый нейрональный компонент, который приводит к алопеции и зуду, индуцируемому IL-31. Таким образом, нейтрализация IL-31 может снизить встречаемость и интенсивность зуда, и, таким образом, дерматита, у пациентов, страдающих нарушениями кожи, в которые вовлечен зуд.
Способы II
Мыши, нуль-гомозиготные по гену Tac1, не экспрессируют вещество P или нейрокинин A. Эти мыши обладают значительно уменьшенными ноцицептивными ответами на боль со смягчением интенсивных стимулов и, таким образом, они являются пригодным инструментом для исследования вклада пептидов тахикинина в развитие боли/зуда и состояний воспалительного заболевания. Мышам с нокаутом Tac1 в возрасте двенадцать недель имплантировали 14-суточные осмотические насосы, доставляющие белок IL-31 в количестве 1 мкг/сутки, и наблюдали каждые сутки на алопецию и зуд с использованием следующих критериев: 0 = нет расчесывания, животное выглядит нормально, 1 = истончение шерсти на небольших областях, отмечается расчесывание, 2 = небольшая потеря шерсти (небольшие участки), расчесывание, 3 = умеренная потеря шерсти, расчесывание и 4 = тяжелая потеря шерсти, чрезмерное расчесывание.
Результаты этого исследования показывают, что дефектные по Tac1 мыши были менее восприимчивыми в отношении индуцируемого посредством IL-31 расчесывания/потери шерсти по сравнению с контрольными мышами дикого типа. В то время как у 100% (10/10) мышей дикого типа развивались признаки расчесывания и потери шерсти на 6 сутки введения IL-31, только у 33,3% (2/6) дефицитных по Tac1 мышей были выявлены признаки расчесывания и потери шерсти в тот же момент времени. Эти данные показывают, что IL-31 индуцирует нейрональный компонент, который приводит к фенотипу с расчесыванием/потерей шерсти у мышей, которым вводили IL-31, и нейтрализация IL-31 может снизить частоту и интенсивность расчесывания, связанного с дерматитом.
Способы III (Введение нейтрализующего антитела против IL-31)
Нормальным самкам мышей BALB/c (CRL) в возрасте приблизительно от 8 до 12 недель имплантировали подкожно 14-суточные осмотические насосы (Alzet, #2002), доставляющие mIL-31 в количестве 1 мкг/сутки. Группам мышей проводили внутрибрюшинные (i.p.) инъекции моноклонального антитела крысы против IL-31 мыши в количестве 10 мг/кг (200 мкг/мыши) два раза в неделю, начиная за 1 неделю перед доставкой IL-31. В контрольных группах мышей проводили внутрибрюшинные инъекции носителя (PBS/0,1%BSA) с идентичными схемами дозирования. Мышей каждые сутки оценивали на алопецию и зуд с использованием следующих критериев: 0 = нет расчесывания, животное выглядит нормально, 1 = истончение шерсти на небольших областях, отмечается расчесывание, 2 = небольшая потеря шерсти (небольшие участки), расчесывание, 3 = умеренная потеря шерсти, расчесывание и 4 = тяжелая потеря шерсти, чрезмерное расчесывание.
Во всех экспериментах, у мышей, которым вводили mAb крысы против mIL-31, было замедлено проявление симптомов приблизительно на 5-7 суток, и они обладали более низкими общими показателями алопеции и зуда. Во всех группах мышей, которым вводили mAb (независимо от частоты дозирования или концентрации дозы), развивались алопеция и зуд, сходные с контрольными мышами к 13 суткам исследования. Эти данные подтверждают, что нейтрализация IL-31 может замедлить начало ответа в виде расчесывания/потери шерсти, индуцируемого IL-31.
Пример 13
Охарактеризация моноклональных антител мыши против IL-31 человека
Получали панель из 21 клональной гибридомы, продуцирующей моноклональные антитела (mAb), специфичные в отношении (рекомбинантного) интерлейкина 31 (IL31) человека. Выделяли десять гибридом, продуцирующие mAb с высокой нейтрализацией. Их распределяли на две подсерии, исходя из исследований конкурентного связывания. Выделяли одиннадцать гибридом, продуцирующих mAb со слабой нейтрализацией. Их распределяли на 4 дополнительные серии, исходя из исследований конкурентного связывания. Одно mAb хорошо работало в вестерн-блоттинге, и оно также хорошо работало в иммуногистохимии. Было идентифицировано три mAb, пригодных для применения в качестве конкурентных антагонистов. Также идентифицировали моноклональные антитела, пригодные для применения в иммунологических анализах по типу "сэндвич".
A. Сортировка по эпитопу посредством Biacore
Материалы: иммобилизованное антитело представляло собой Fc-γ-специфичное антитело козы против IgG мыши (Jackson #115-005-071); блокирующее антитело представляло собой Fc-фрагмент поликлонального IgG (Jackson Labs.); антиген: rIL31, продуцированный в BHK с аффинным маркером CEE; Biacore 1000; NHS-активированный чип Biacore CM5 (Biacore, PN BR-1000-14).
Способ: Исследования проводили с помощью системы Biacore 1000™. Для способов программного анализа использовали BIAlogue v. 1,2. Антитело козы против Fc мыши ковалентно иммобилизовывали на чипе Biacore CM5 через остатки лизина с формированием активной связывающей поверхности для исследования. Супернатанты кондиционированных сред клонов каждого из 21 mAb мыши против huIL31 сначала инъецировали на поверхность с антителом козы против Fc мыши так, чтобы первичные mAb, подлежащие тестированию, могли связываться с поверхностью антитела против Fc мыши в подходящей ориентации. Затем остальные участки для связывания блокировали посредством инъекции незначимых Fc-фрагментов поликлонального IgG. Затем антиген инъецировали (rIL31) и иммобилизовывали на поверхности с первичным антителом. Затем проводили другую инъекцию одного из супернатантов кондиционированных сред клонов каждого из 21 mAb мыши против huIL31 для тестирования связывания вторичного антитела. Если первичное и вторичное mAb конкурировали за один и тот же участок связывания на антигене, то второе mAb не связывалось. Если два mAb не конкурировали за один и тот же участок связывания на антигене, то происходило связывание второго mAb.
Каждый супернатант тестировали в качестве первичного mAb в сочетании со всем набором mAb. Контрольные циклы проводили для того, чтобы показать отсутствие ответа вторичного mAb в отсутствие первичного mAb или антигена. Каждое mAb, тестируемое против его самого, использовали в качестве отрицательного контроля для установления уровня фонового сигнала. Данные объединяли с использованием программного обеспечения BioEvaluation 3,2 RCI, а затем загружали в Excel™ для обработки данных. Между каждыми циклами тестирования пары моноклональных антител, поверхность с антителами козы против Fc мыши регенерировали посредством промываний 2·30 с посредством 50 мМ HCl.
mAb с высокой нейтрализацией (10) и mAb со слабой нейтрализацией (11) исследовали в двух отдельных панелях, за исключением того, что при исследовании mAb со слабой нейтралзиацией в исследование включали mAb из гибридомы 292.64.6 в качестве mAb, представляющего собой mAb с высокой нейтралзиацией. Кроме того, после изучения показателей относительной аффинности и данных нейтрализации, учитывая серии по эпитопам, для первых двух панелей для сортировки, проводили третью панель сортировки для 8 "выбранных" mAb, включая нейтрализующие mAb как с высокой, так и с низкой нейтрализацией.
Результаты: Было проведено тестирование спаривания трех панелей моноклональных антител с анализом mAb как с высокой нейтрализацией, так и со слабой нейтрализацией. В первой панели тестировали все mAb с высокой нейтрализацией друг против друга и во второй панели тестировали все mAb со слабой нейтрализацией (и репрезентативное mAb с высокой нейтрализацией из гибридомы 292.64.6) друг против друга. Третью панель проводили для прямой оценки спаривания ряда mAb как с высокой нейтрализацией, так и со слабой нейтрализацией, выбранных для дальнейшей оценки.
В дополнение к варьированию абсолютного ответа (RU), определенного для различных пар mAb, несколько пар mAb работало по разному при изменении ориентации пар (т.е. одно mAb из пары использовали в качестве первичного, а не вторичного mAb). Такое поведение наблюдалось часто и, главным образом, является свойственным mAb, предположительно имеющим перекрывающиеся эпитопы.
При завершении анализа первых двух панелей было получено всего четыре отличающиеся серии. Для первой панели, все антитела с высокой нейтрализацией объединяли в группу, чтобы они совместно составляли одну серию (серия 1: 292.12.3, 292.39.5, 292.51.5, 292.63.5, 292.64.6, 292.72.3, 292.84.1, 292.105.4, 292.109.4, 292.118.6). Во второй панели, антитела со слабой нейтрализацией группировали в 3 дополнительные основные серии (серия 2: 294.35.2, 294.146.5, 292.152.4, 292.152.4, 294.154.5; серия 3: 294.35.3, 291.78.4, 294.158.5, 294.155.6, 294.163.2; серия 4: 294.144.3), которые отличались от единственного представителя серии #1 (292.64.6). Кроме того, множество пар mAb, оцениваемых в панели 2, показало отчетливую асимметрию ответа в зависимости от ориентации, в которой тестировали mAb, указывая на некоторое перекрывание участков связывания. Для двух mAb из серии 3, 294.35.3 и 291.78.4, были показаны признаки конкуренции с mAb из серии 2, при тестировании в качестве вторичного mAb. Также в панели 2, для антитела с высокой нейтрализацией 292.64.6 (серия 1) показана конкуренция с mAb в серии 3, при тестировании в качестве первичного антитела.
После завершения анализа исходных двух панелей для сортировки, для дальнейшей оценки отбирали группу mAb с наиболее высокой нейтрализацией и mAb с наивысшей аффинностью со слабой нейтрализацией, и их тестировали друг против друга в панели 3. Результаты для третьей панели полностью согласовывались с предшествующими анализами, показывая, что антитела c высокой нейтрализацией были отсортированы совместно и антитела со слабой нейтрализацией разделились в определенные множественные серии. Кроме того, в третьей панели было показано частичное перекрывание связывания двух mAb из серии 2 (294.35.2 и 292.154.4) с подгруппой mAb, приписанных к серии 1 (292.12.3, 292.72.3, 292.84). Это привело к выделению подсерий 1A и 1B в серии 1. В панели 3, mAb из гибридомы 292.163.2 (серия 3) и 292.144.3 (серия 4) были отсортированы отдельно.
B. Сортировка по эпитопу в титрационном микроплашете (конкурентный ELISA без метки)
Материалы: Иммобилизованное антитело представляло собой антитело козы против IgG мыши (Fc-γ-специфичный) Jackson #115-005-071; блокирующее антитело представляло собой IgG1 мыши, (ZymoGenetics); для этого исследования использовали супернатанты кондиционированных сред гибридом; антиген представлял собой rIL31, продуцированный в BHK с аффинным маркером CEE, биотинилированным с использованием набора сульфо-NHS-биотин (Pierce, Rockford, IL); 96-луночные планшеты Nunc Maxisorp (Nalge Nunc, Rochester, NY); ELISA B (PBS, 0,1% Tween 20, 1% BSA); стрептавидин-HRP (Pierce, Rockford, IL); субстрат TMB (BioFX Laboratories, Owings Mills, MD)
Способ: этот способ является сходным с конкурентным ELISA без метки (LFC-ELISA), описанным by Nagata et al (2004). В этом способе для сортировки по эпитопу используют биотинилированный rIL31. Микропланшеты для титрования покрывали Fc-γ-специфичным антителом козы против IgG мыши (Jackson ImmunoResearch #115-005-071) в концентрации 1 мкг/мл в количестве 100 мкл/лунка, разведенном в ELISA B (PBS, 0,1% Tween 20, 1% BSA). После связывания этого антитела для покрытия в течение 3 часов при температуре окружающей среды, каждую содержащую mAb кондиционированную среду разбавляли в ELISA B с получением приблизительной концентрации mAb 0,5 мкг/мл и обеспечивали связывание с покрытыми антителом козы против IgG мыши планшетами в течение ночи при 4°C (mAb#1). Аналогично, второй набор кондиционированных сред (mAb#2) разбавляли в пробирках для тестирования из полистирола до концентрации mAb приблизительно 0,5 мкг/мл в ELISA B, смешивали с биотинилированным антигеном rIL31 в концентрации 50 нг/мл и инкубировали в течение ночи при 4°C. После инкубации mAb#1 с антителом для покрытия, планшеты блокировали неродственным антителом для насыщения незанятых участков связывания на планшете. В планшет добавляли смеси mAb#2-биотин-rIL31 и обеспечивали связывание. В качестве контроля в анализе (неконкурентного) добавляли биотинилированный rIL31 в концентрации 50 нг/мл непосредственно (без предварительной инкубации с mAb#2) в лунки, содержащие иммобилизованное mAb#1. После инкубации с комплексом биотинилированный IL31-mAb#2, в планшет добавляли стрептавидин-HRP (Pierce, Rockford, IL) в концентрации 0,5 мкг/мл. Планшеты проявляли посредством субстрата TMB (BioFX Laboratories, Owings Mills, MD), и величину поглощения в отдельных лунках при 450 нм измеряли посредством устройства для считывания планшетов (Molecular Devices SpectraMax340, Sunnyvale, CA). Если mAb#1 распознавал эпитоп, отличный от эпитопа mAb#2, то комплекс биотин-rIL31-mAb#2 связывался с планшетом, что приводило к считыванию с высоким поглощением. Если mAb#1 распознавал тот же эпитоп, что и mAb#2, тот комплекс биотин-rIL31-MAb#2 не связывался с планшетом, что приводило к считыванию с низким поглощением.
Результаты: полную панель из 21 mAb тестировали против них самих одновременно в серии из шести 96-луночных микропланшетов для титрования. Величину поглощения при 450 нм (A450 нм) записывали для каждого сочетания и ориентации и объединяли. Во всех случаях, для сочетаний mAb с этим же mAb получали низкие значения поглощения. В дополнение к контролю в виде этого же mAb, тестировали положительный контроль, представляющий собой rIL31-биотин (без конкурентного mAb#2) в концентрации 50 нг/мл с использованием каждого первичного mAb на каждом планшете. Для каждого первичного mAb получали два положительных контрольных образца, и показатели дублированных образцов были сходными. Величины поглощения, полученные для лунок с положительным контролем (без mAb#2) для 9 из 11 mAb со слабой нейтрализацией (из серий 2, 3 или 4), были меньше, чем величины поглощения, измеренные для контрольных образцов для остальных mAb. Вероятно, это было следствием более низкой аффинности (более высокой EC50) этих mAb. Для упрощения интерпретации результатов, значения поглощения нормализовали таким образом, что максимальному измеренному поглощению для любого сочетания mAb в одном ряду (иммобилизованное mAb (mAb#1) оставалось постоянным) приписывали значение 100%. Нормализованные значения относились к двум различным группам: одна большая группа значений оказалось ниже значения 32%, что соответствует конкуренции, и вторая большая группа значений оказалась выше 32%, что соответствует отсутствию конкуренции. 32% пороговое значение использовали для определения каждого сочетания и ориентации mAb в одну или две различные категории (неконкурентную и конкурентную). С использованием сгруппированных данных, автоматически проводили сортировку, исходя из алгоритма иерархической группировки. Как было выявлено в процессе экспериментов по сортировке с использованием Biacore, некоторые пары mAb группировались по-разному в зависимости от того, какое mAb использовали в качестве первичного mAb. Высокий уровень строгости, требующий конкуренции в обоих направлениях, использовали для определения mAb в основные серии. Затем менее строгие критерии, требующие конкуренции только в одном направлении, использовали для упрощения определения mAb в подсерии, исходя из небольших различий в характеристиках. Нормализованные данные группировали в соответствии с определением в 5 основных серий, исходя из критериев высокой строгости. Кроме того, сочетания и ориентации mAb кодировали цветом (белый для конкурентных и темный для неконкурентных) для упрощения визуализации взаимодействий, которые определяют mAb в подсерии.
Результаты сортировки посредством конкурентного ELISA согласуются с определениями серий на основе результатов исследований с использованием Biacore. С использованием строгих условий сортировки были определены следующие серии:
Серия #1 (содержащая все mAb с высокой нейтрализацией): 292.72.3, 292.64.6, 292.63.5, 292.51.5, 292.39.5, 292.12.3. 292.118.6, 292.109.4, 292.105.4, 292.84.1
Серия #2: 294.35.2, 294.154.5, 294.146.5, 292.154.4, 292.152.4
Серия #3: 294.35.3, 294.163.2, 294.158.5, 294.155.6
Серия #4: 294.144.3
Серия #5: 291.78.4
Рассматривая серию #1, для mAb из 3 гибридом серии #1 (292.72.3, 292.12.3, 292.84.1) была показана конкуренция с 3 mAb из серии #2, при использовании в качестве вторичного mAb (mAb#2). Исходя из этих характеристик, серию 1 разделили на две подсерии; #1A и #1B, где серия #1B содержала mAb, которые конкурируют с mAb серии 2 за связывание. Серия #1A содержит 292.64.6, 292.63.5, 292.51.5, 292.39.5, 292.118.6, 292.109.4, 292.105.4. Серия #1B содержит 292.72.3, 292.12.3 и 292.84.1.
Рассматривая серию #2, наблюдали три различных взаимодействия, которые использовали для разделения серии #2 на 3 подсерии. Серия #2A содержит mAb из гибридомы 294.146.5, которое конкурирует со всеми mAb из обеих серий #1 и #2. Серия #2B содержит mAb из гибридом 294.35.2 и 294.154.5, которые конкурируют с (вторичными) mAb из серии #1. Серия #2C содержит mAb из гибридом 292.154.4 и 292.152.4, которые конкурируют с mAb из серии #4, когда его используют в качестве вторичного mAb.
Рассматривая серии #3, #4 и #5: mAb серии #3 (из гибридом 294.35.3, 294.163.2.1 и 294.155.6) конкурируют с mAb из обеих серий #4 и #5, когда их используют в качестве вторичных mAb. mAb в сериях #4 (из гибридомы 294.144.3.5) и #5, главным образом, отличаются по их взаимодействию с mAb в подсерии #2C (и по своеобразным свойствам связывания mAb из гибридомы 294.144.3 на вестерн-блотах).
Когда полную панель из 21 mAb тестировали одновременно с использованием LFC-ELISA, были отчетливо идентифицированы серия 1 и подсерии #1A и #1B. Также было выявлено частичное перекрывание серий #2 и #3, когда полную панель mAb тестировали одновременно, и серия #2 была разделена на 3 подсерии. Одним небольшим отклонением от определения серий на основе данных Biacore было определение mAb из гибридом 291.78.4 и 294.144.3 в две различные серии, когда использовали строгие критерии в отношении данных LFC-ELISA. Исходя из исследований Biacore, mAb из гибридомы 291.78.4 было определено в серию #3.
C. Измерение аффинности на планшетах
Материалы: Иммобилизованное поликлональное антитело представляло собой антитело козы против IgG мыши (Fc-γ-специфичное) (Jackson ImmunoResearch West Grove, Pennsylvania #115-005-071); блокирующее поликлональное антитело представляло собой антитело мыши против IgG человека (Jackson ImmunoResearch, #209-005-082); для этого исследования использовали супернатанты кондиционированных сред гибридом; антиген представлял собой rIL31, продуцированный в BHK с аффинным маркером CEE, биотинилированным с использованием набора Сульфо-NHS-Биотин (Pierce, Rockford, JJL); 96-луночные планшеты Nunc Maxisorp (Nalge Nunc, Rochester, NY); ELISA B (PBS, 0,1% Tween 20, 1% BSA); стрептавидин-HRP (Pierce, Rockford, IL); субстрат TMB (BioFX Laboratories, Owings Mills, MD).
Способ: Этот способ является сходным со способом, описанным van Heyningen (1986). Титрационные микропланшеты покрывали Fc-γ-специфичным антителом козы против IgG мыши (Jackson ImmunoResearch #115-005-071) в концентрации 1 мкг/мл в количестве 100 мкл/лунка, разбавленным в ELISA B (PBS, 0,1% Tween 20, 1% BSA). После связывания этого антитела для покрытия в течение 3 часов при температуре окружающей среды, каждый очищенный супернатант с моноклональным антителом разбавляли в ELISA B с получением приблизительной концентрации mAb 1 мкг/мл и обеспечивали связывание с планшетом в течение 1 часа при температуре окружающей среды. Получали серийные разведения от 500 нг/мл до 0 нг/мл биотинилированного антигена rIL31 в ELISA B и добавляли в лунки. После инкубации с биотинилированным антигеном, в планшет добавляли стрептавидин-HRP (Pierce, Rockford, IL) в концентрации 0,5 мкг/мл. Планшеты проявляли посредством субстрата TMB (BioFX Laboratories, Owings Mills, MD) и измеряли поглощение для отдельных лунок при 450 нм с помощью устройства для считывания планшетов (Molecular Devices SpectraMax340, Sunnyvale, CA). Значения для дублей усредняли и данные анализировали посредством четырехпараметрической подгонки. Значение "C" при 4-параметрической подгонке известно как кажущаяся EC50 (нг/мл), и она представляет собой концентрацию биотин-rIL31, которая вызывает половину максимального ответа в анализе.
Результаты: Четырехпараметрическую подгонку получали из экспериментальных кривых для всех 21 mAb. Концентрация биотин-rIL31, вызывающая половину максимального ответа (EC50) в анализе, варьировала от 3,3 до 236 нг/мл, где все 10 mAb с высокой нейтрализацией проявляли низкие и сравнимые значения EC50 (3,3-4,4 нг/мл).
D. Вестерн-блоттинг
Материалы: антиген представлял собой rIL31, продуцированный в BHK с аффинным маркером CEE; 4-12% гели бис-Трис NuPAGE (Invitrogen, Carlsbad, CA); нередуцирующий буфер для образца (Invitrogen, Carlsbad, CA); Стандарты молекулярной массы представляли собой SeeBlue (Invitrogen); 1× буфер для анализа MES (Invitrogen); Буфер A для вестерн-блоттинга (50 мМ Трис pH 7,4, 5 мМ ЭДТА, 150 мМ NaCl, 0,05% игепал, 0,25% желатин); 0,2-мкм нитроцеллюлозные мембраны (Invitrogen); антитела овцы против IgG мыши-HRP (Amersham, Piscataway, NJ); Очищенное аффинными способам pAb кролика, специфичное в отношении rIL31 (ZymoGenetics); антитело осла против Ig кролика-HRP (Amersham); хемилюминесцентный реагент Lumi-Light Plus (Roche, Mannheim, Germany); Lumi-Imager (Mannheim-Boehringer).
Способ: В этом исследовании оценивали способность mAb выявлять денатурированный/восстановленный rIL31 на вестерн-блоте. Антиген rIL31 смешивали либо с редуцирующим, либо с нередуцирующим буфером для образца, нагревали при 70°C в течение 10 мин, а затем наносили на 4-12% бис-Трис гели NuPAGE в количестве 100 нг/дорожка. Стандарты молекулярной массы представляли собой SeeBlue (Invitrogen), и электрофорез поводили в 1× буфере для анализа MES (Invitrogen). Полосы белков в гелях переносили на 0,2-мкм нитроцеллюлозные мембраны (Invitrogen) и блокировали в течение ночи в 2,5% обезжиренном сухом молоке в буфере A для вестерн-блоттинга (50 мМ Трис pH 7,4, 5 мМ ЭДТА, 150 мМ NaCl, 0,05% игепал, 0,25% желатин). На нитроцеллюлозные мембраны наносили каждое моноклональное антитело в приблизительной концентрации mAb 0,1 мкг/мл. Затем на блоты наносили вторичное антитело, конъюгированное с пероксидазой хрена (антитело овцы против IgG мыши-HRP; Amersham, Piscataway, NJ). В качестве положительного контроля на отдельный вестерн-блот наносили поликлональное антитело кролика, специфичного в отношении IL-31 (ZymoGenetics) в концентрации 0,1 мкг/мл, и вторичное антитело, конъюгированное с пероксидазой хрена (антитело осла против Ig кролика-HRP; Amersham). Полосы на вестерн-блотах выявляли посредством реагента Lumi-Light Plus (Roche, Mannheim, Germany) и регистрировали хемилюминесценцию посредством Lumi-Imager (Mannheim-Boehringer).
Результаты: Вестерн-блот анализ mAb против IL31 показал, что mAb только трех из 21 гибридомы выявляли денатурированный антиген rIL31. Наиболее сильный сигнал был получен от mAb, продуцируемого гибридомой 294.144.3. Это mAb также выявляло восстановленный/денатурированный rIL31 более эффективно, чем невосстановленный/денатурированный rIL31. Интенсивность сигнала, наблюдаемого для этого mAb, была сходной с интенсивностью, полученной для контроля в виде поликлонального антитела. Следует отметить, что mAb из гибридомы 294.144.3 относится к серии, отличающейся от серий других оцениваемых mAb. Моноклональные антитела из двух других гибридом (292.84.1 и 292.64.6) выявляли невосстановленный/денатурированный rIL31 очень слабо, однако не выявляли восстановленный/денатурированный rIL31. Этот слабый сигнал не воспроизводился на сканированных блотах. Оба эти mAb представляют собой нейтрализующие антитела серии #1.
Пример 14
Относительная аффинность связывания моноклональных антител крысы против антител мыши в отношении лиганда IL-31
Относительную аффинность связывания четырех MAb крысы против лиганда Ms-IL-31 определяли следующим образом. Оценивали клон 271.26.6.6.1, клон 271.33.3.2.1, клон 271.33.1.2.2 и клон 271.39.4.6.5. Fc-γ-специфичное антитело козы против IgG крысы (Jackson) иммобилизовывали на чип Biacore CM5. После предварительного тестирования анализ дополнительно оптимизировали, связывая каждое MAb с поверхностью для улавливания противокрысиных антител, а затем через MAb пропускали серии концентраций лиганда IL-31 для определения ассоциации и диссоциации. После каждого анализа поверхность регенерировали до иммобилизованного антитела против антител крысы посредством 2 инъекций 30 мМ HCl. Получали данные для каждого MAb и для оценки использовали программное обеспечение для определения относительных значений кинетики.
Относительные данные кинетики, полученные посредством оценки кривых связывания MAb-антиген, представлены в таблице 4.
Пример 15
Снижение TARC и MDC в ответ на антитело против IL-31 в модели AD на мышах
Способ I
Самцов мышей NC/Nga в возрасте шесть недель (CRL Japan) сенсибилизировали внутрикожно посредством 50 мкг экстракта клеща домашней пыли (D. pteronyssinus, Indoor Biotechnologies) три раза в неделю в спину и оценивали подобные AD очаги повреждения. Через 5 недель сенсибилизации, мышей умерщвляли и правые уши вырезали и помещали в отдельную лунку 48-луночной чашки для культивирования (Corning), с добавлением RPMI+2%ЭТС (GIBCO Invitrogen). Планшеты помещали в инкубаторы с 5% CO2 и контролируемой влажностью. Через 24 часа собирали супернатанты и замораживали при -20°C до проведения дальнейшего анализа.
Способ II
Самок мышей NC/Nga в возрасте двенадцать недель (CRL Japan) сенсибилизировали внутрикожно посредством 10 мкг SEB (Toxin Technology) в ухо и в спину три раза в неделю. Мышей оценивали на подобные AD очаги повреждения. Через 5 недель сенсибилизации мышей умерщвляли и 6 мм пункционной биопсии забирали из подвергавшегося инъекциям уха каждой мыши и помещали в отдельную лунку 48-луночной культуральной чашки, дополненной RPMI+2%ЭТС. Планшеты помещали в инкубаторы с 5% CO2 и контролируемой влажностью. Через 24 часа собирали супернатанты и замораживали при -20°C до проведения дальнейшего аналзиа.
В группах мышей в обоих исследованиях вводили либо моноклональное антитело крысы против IL-31 мыши в концентрации 10 мг/кг, либо носитель, внутрибрюшинно два раза каждую неделю, начиная от 1 до 2 недели сенсибилизации.
Концентрации TARC и MDC в образцах супернатантов через 24 часа измеряли посредством общепринятого ELISA (R&D Systems).
Концентрации TARC и MDC были ниже в супернатантах ушей из мышей, которым вводили антитело против IL-31 по сравнению с контрольными мышами в обоих исследованиях, однако, эти результаты не были статистически значимыми при анализе посредством ANOVA, возможно вследствие малого объема выборки. При объединении и анализе всех данных из обоих экспериментов, выявлено статистически значимое различие между группами по введению.
Пример 16
Введение нейтрализующего IL-31 антитела
Нормальным самкам мышей BALB/c (CRL) в возрасте приблизительно от 8 до 12 недель имплантировали подкожно 14-суточные осмотические насосы (Alzet, #2002), доставляющие mIL-31 в количестве 1 мкг/сутки. В группах мышей проводили внутрибрюшинные (i.p.) инъекции моноклональнго антитела крысы против IL-31 мыши в количестве 10 мг/кг (200 мкг/мышь) два раза в неделю, начиная за 1 неделю до доставки IL-31. В контрольных группах мышей проводили внутрибрюшинные инъекции носителя (PBS/0,1%BSA) с идентичными схемами дозирования. Мышей каждые сутки оценивали на алопецию и зуд с использованием следующих критериев: 0 = нет расчесывания, животное выглядит нормально, 1 = истончение шерсти на небольших областях, отмечается расчесывание, 2 = небольшая потеря шерсти (небольшие участки), расчесывание, 3 = умеренная потеря шерсти, расчесывание и 4 = тяжелая потеря шерсти, чрезмерное расчесывание.
Во всех экспериментах мыши, которым вводили mAb крысы против mIL-31, обладали замедленным проявлением симптомов на приблизительно от 5 до 7 суток и более низкие общие показатели для алопеции и зуда. Во всех группах мышей, в которых вводили mAb (независимо от частоты дозирования и концентрации дозы), развивалась алопеция и зуд, сходные с контрольными мышами, к 13 суткам исследования. Эти данные подтверждают, что нейтрализация IL-31 может замедлить появление ответа в виде расчесывания/потери шерсти, индуцируемого посредством IL-31.
Исходя из вышесказанного, следует учитывать, что, несмотря на то, что в целях иллюстрации в настоящем описании описаны конкретные варианты осуществления этого изобретения, можно проводить различные модификации без отклонения от сущности и объема этого изобретения. Таким образом, это изобретение не ограничивается ничем, кроме прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ И ВОСПАЛЕНИЯ В НЕЙРОННОЙ ТКАНИ С ПРИМЕНЕНИЕМ АНТАГОНИСТОВ IL-31 | 2007 |
|
RU2440130C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ВОСПАЛЕНИЯ В НЕЙРОННОЙ ТКАНИ С ПРИМЕНЕНИЕМ МОНОКЛОНАЛЬНОГО АНТИТЕЛА ИЛИ ЕГО СВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2007 |
|
RU2565391C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| СКОНСТРУИРОВАННОЕ АНТИТЕЛО ПРОТИВ TSLP | 2010 |
|
RU2575039C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ИНТЕРФЕРОНА-АЛЬФА И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2431638C2 |
| ЛЕЧЕНИЕ ИНТЕРСТИЦИАЛЬНОГО ЦИСТИТА | 2008 |
|
RU2570559C2 |
| АНТИТЕЛА, ОБЛАДАЮЩИЕ СПЕЦИФИЧНОСТЬЮ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2794996C2 |
| ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО ОТ ЗУДА | 2008 |
|
RU2541780C2 |
| АНТИТЕЛА К РОДСТВЕННОЙ РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ МОЛЕКУЛЕ КЛЕТОЧНОЙ АДГЕЗИИ (СЕАСАМ) | 2012 |
|
RU2650869C2 |
| НОВЫЕ АНТИ-IL 13 АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2004 |
|
RU2487887C2 |
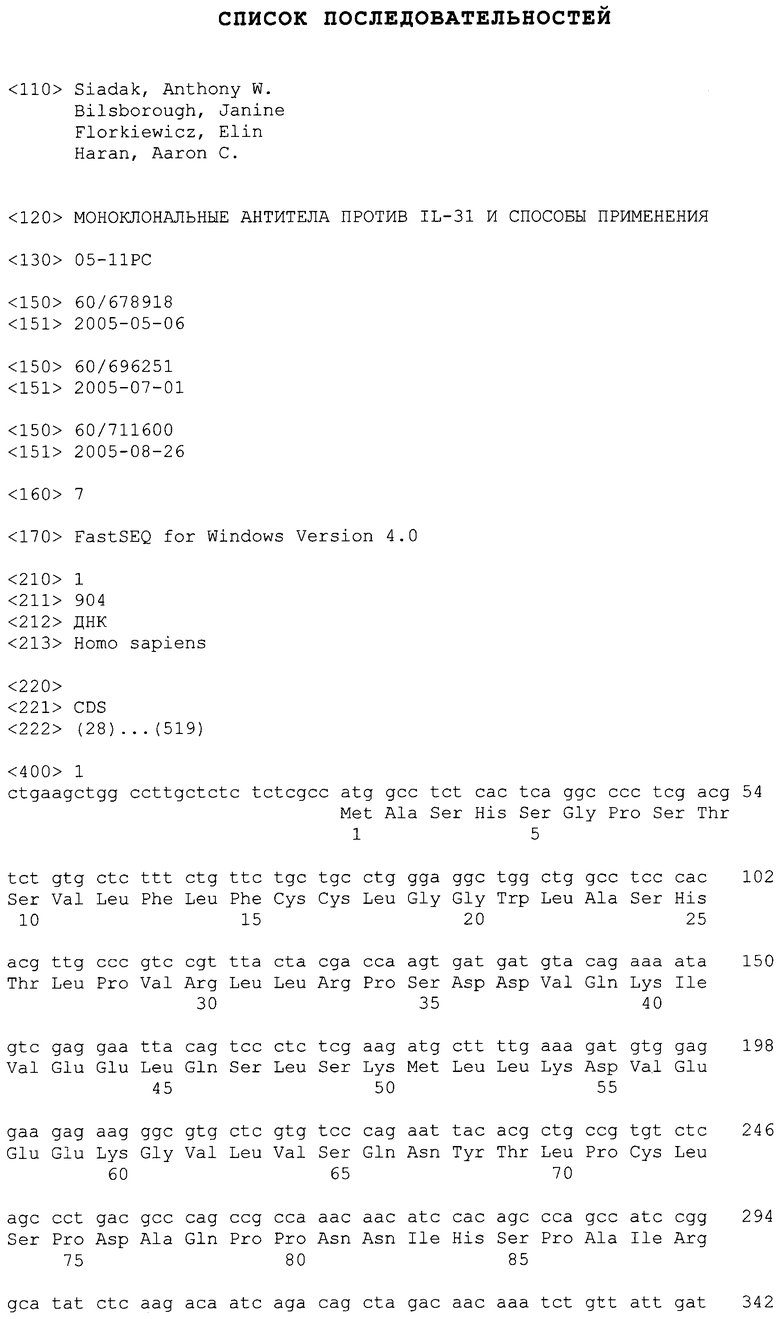







Изобретение относится к биотехнологии. Описано моноклональное антитело против IL-31. Предложен способ лечения млекопитающего, страдающего воспалительным заболеванием, в котором участвует IL-31, включающий введение описанного антитела. Предложены способы снижения, ингибирования или минимизации зуда у млекопитающего, включающие введение описанного антитела. Изобретение расширяет арсенал противовоспалительных средств. 13 н. и 26 з.п. ф-лы, 4 табл., 16 пр.
1. Моноклональное антитело против IL-31 или его функциональный фрагмент, где моноклональное антитело продуцируется гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС, выбранный из
a) патентного номера депозита в АТСС РТА-6815;
b) патентного номера депозита в АТСС РТА-6816;
c) патентного номера депозита в АТСС РТА-6829;
d) патентного номера депозита в АТСС РТА-6830;
e) патентного номера депозита в АТСС РТА-6831;
f) патентного номера депозита в АТСС РТА-6871;
g) патентного номера депозита в АТСС РТА-6872;
h) патентного номера депозита в АТСС РТА-6875; и
i) патентного номера депозита в АТСС РТА-6873, и специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
2. Моноклональное антитело против IL-31 или его функциональный фрагмент, где моноклональное антитело продуцируется гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС, выбранный из
a) патентного номера депозита в АТСС РТА-6815;
b) патентного номера депозита в АТСС РТА-6816;
c) патентного номера депозита в АТСС РТА-6871;
d) патентного номера депозита в АТСС РТА-6829; и
e) патентного номера депозита в АТСС РТА-6830,
и специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
3. Моноклональное антитело против IL-31 или его функциональный фрагмент, где моноклональное антитело продуцируется гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС, выбранный из
a) патентного номера депозита в АТСС РТА-6815;
b) патентного номера депозита в АТСС РТА-6816; и
c) патентного номера депозита в АТСС РТА-6871,
и специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
4. Моноклональное антитело против IL-31 или его функциональный фрагмент, где моноклональное антитело продуцируется гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС, выбранный из
a) патентного номера депозита в АТСС РТА-6829; и
b) патентного номера депозита в АТСС РТА-6830,
и специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
5. Моноклональное антитело против IL-31 или его функциональный фрагмент, где моноклональное антитело продуцируется гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС, выбранный из
a) патентного номера депозита в АТСС РТА-6872; и
b) патентного номера депозита в АТСС РТА-6875, и
специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
6. Антитело или фрагмент антитела по пп.1 или 2-5, где антитело или фрагмент антитела нейтрализуют взаимодействие IL-31 (SEQ ID NO:2) с IL-31RA (SEQ ID NO:5).
7. Антитело или фрагмент антитела по пп.1 или 2-5, где антитело или фрагмент антитела представляют собой (а) моноклональное антитело мыши, (b) гуманизированное антитело, полученное из (а), или (с) фрагмент антитела, или (d) моноклональное антитело человека.
8. Антитело или фрагмент антитела по пп.1-5, где антитело или фрагмент антитела содержат радионуклид, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидный маркер, магнитную частицу или токсин.
9. Антитело или фрагмент антитела по пп.1-5, где антитело или фрагмент антитела содержат пегилирование.
10. Антитело или фрагмент антитела по пп.1-5, где антитело или фрагмент антитела являются нейтрализующими.
11. Антитело или фрагмент антитела по пп.1-5, где введение антитела или фрагмента антитела млекопитающему ингибирует, снижает или блокирует провоспалительную активность полипептида.
12. Антитело или фрагмент антитела по пп.1-5, где введение антитела или фрагмента антитела млекопитающему ингибирует, снижает или блокирует расчесывание и дерматит, обусловленные провоспалительной активностью полипептида.
13. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6815.
14. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6816.
15. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6829.
16. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6830.
17. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6831.
18. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6871.
19. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6872.
20. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6875.
21. Моноклональное антитело по п.1, где моноклональное антитело или фрагмент антитела продуцируются гибридомой, депонированной в Американской коллекции типовых культур, имеющей патентный номер депозита в АТСС 6873.
22. Антитело или фрагмент антитела по пп.1 или 2-5 для ингибирования, снижения или блокирования очаговой алопеции или дерматита.
23. Антитело или фрагмент антитела по пп.1 или 2-5, где введение антитела млекопитающему ингибирует, снижает или блокирует продукцию провоспалительных хемокинов.
24. Антитело или фрагмент антитела по п.23, где провоспалительные хемокины представляют собой TARC или MDC.
25. Антитело или фрагмент антитела по пп.1 или 2-5, где введение антитела млекопитающему ингибирует, снижает или блокирует у млекопитающего желание чесаться.
26. Антитело или фрагмент антитела по пп.1 или 2-5, где антитело или фрагмент антитела представляют собой молекулу Fab или молекулу (Fab′)2.
27. Антитело или фрагмент антитела по пп.1 или 2-5, где антитело или фрагмент антитела представляют собой одноцепочечную молекулу.
28. Способ уменьшения воспаления у млекопитающего, включающий в себя введение млекопитающему количества антитела IL-31 по п.1, посредством которого происходит уменьшение воспаления.
29. Способ по п.28, где воспаление является симптомом витилиго, кожной Т-клеточной лимфомы, очаговой алопеции, красных угрей, обыкновенных угрей, узловатой почесухи или буллезного пемфигоида, и где происходит уменьшение воспаления.
30. Способ лечения млекопитающего, страдающего воспалительным заболеванием, в котором участвует IL-31, включающий в себя
введение антагониста IL-31 млекопитающему с тем, чтобы уменьшить воспаление, причем антагонист содержит антитело или фрагмент антитела по п.1; и
при этом происходит снижение воспалительной активности IL-31.
31. Способ снижения, ингибирования или минимизации зуда у млекопитающего, в котором участвует IL-31, включающий в себя
введение антагониста IL-31 млекопитающему с тем, чтобы происходило снижение зуда, причем антагонист содержит антитело или фрагмент антитела по п.1;
и при этом происходит снижение вызывающей зуд активности IL-31.
32. Способ снижения, ингибирования или минимизации зуда у млекопитающего, в котором участвует IL-31, включающий в себя введение антагониста IL-31 млекопитающему, причем антагонист содержит антитело или фрагмент антитела по п.1, в силу чего введение антагониста IL-31 снижает у животного желание чесаться.
33. Способ по пп.28-32, где заболевание представляет собой воспалительное заболевание, включающее в себя связанные с зудом заболевания.
34. Способ по пп.28-32, где заболевание представляет собой атопический дерматит.
35. Способ по пп.28-32, где заболевание представляет собой узловатую почесуху.
36. Способ по пп.28-32, где антителом является антитело по пп.1-5.
37. Моноклональное антитело или его функциональный фрагмент, где моноклональное антитело продуцируется клоном гибридомы, обозначенным номером, выбранным из
a) клона 292.12.3.1 (патентный номер депозита в АТСС РТА-6815);
b) клона 292.63.5.3 (патентный номер депозита в АТСС РТА-6829);
c) клона 292.72.3.1 (патентный номер депозита в АТСС РТА-6816);
d) клона 292.84.1.6 (патентный номер депозита в АТСС РТА-6871); и
e) клона 292.118.6.4 (патентный номер депозита в АТСС РТА-6830),
и специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
38. Моноклональное антитело или его функциональный фрагмент, где моноклональное антитело продуцируется клоном гибридомы, обозначенным номером, выбранным из
a)клона 294.35.2.6.3 (патентный номер депозита в АТСС РТА-6872);
b) клона 294.144.3.5 (патентный номер депозита в АТСС РТА-6873);
c) клона 294.154.5.6 (патентный номер депозита в АТСС РТА-6875); и
d) клона 294.163.2.1 (патентный номер депозита в АТСС РТА-6831),
и специфически связывает полипептид, состоящий из аминокислотной последовательности SEQ ID NO:2.
39. Гибридома, продуцирующая антитело против IL-31, депонированная в Американской коллекции типовых культур, имеющая патентный номер депозита в АТСС РТА-6815.
| US 20050063945 А1, 24.03.2005 | |||
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М.А., ИГНАТЬЕВА Г.А., СТАРЧЕУС А.П | |||
| Моноклональные антитела и гибридомы | |||
| - М.: ВАСХНИЛ, 1989. |

