Изобретение относится к анатомическим методам исследованиям и может быть использовано в фармокогнозии и селекции.
Известны способы морфологического изучения трихом [2], микроскопической диагностики листьев лекарственного сырья [1, 3, 5, 8, 9], выражающиеся в подсчете железок на 1 мм2 на свежем растительном материале [4] или с помощью сканера «EPSON perfection 1660 photo» при большой разрешающей способности (1200 dpi) [3], и определения стадии развития пельтатных железок [10, 11], не подразумевающие определение их функциональности.
По мнению ряда авторов (В.Д.Работягова [4], Т.Г.Фурст [8], G.Heinrich [11]) на продуктивность влияет количество пельтатных железок на единицу площади у полностью сформировавшихся листьев. Их количество на единицу площади листа может сильно отличаться у разных популяций одного и того же вида. Этот фактор оказывает существенное влияние на содержание эфирного масла в сырье эфироносов и соответственно хозяйственную ценность образца [7]. Некоторые авторы указывают на прямую зависимость между количеством пельтатных железок на единицу площади листа и выходом эфирного масла из сырья [6, 11].
Задачей предлагаемого изобретения является оценка функциональной активности пельтатных железок путем расчета коэффициента интенсивности синтеза эфирных масел.
Способ реализуют следующим образом.
- Собирают материал для исследования.
- Определяют концентрацию пельтатных железок путем подсчета количества пельтатных железок (n, шт./мм2) в 1 площади поля зрения (S, мкм2) (расчет площади производят путем умножения числовых показателей увеличений указанных на объективе и окуляре) не менее чем в 25 полях зрения в средней части листовой пластинки.
- Определяют площади пельтатных железок и их субкутикулярных полостей по формулам:
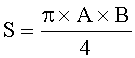 ;
; 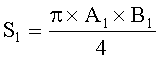 ,
,
где S - площадь пельтатной железки; S1 - площадь субкутикулярной полости, А и В - максимальные поперечные и продольные габаритные размеры пельтатной железки, В1 и A1 - максимальные поперечные и продольные габаритные размеры субкутикулярной полости пельтатной железки. Габаритные размеры определяют с помощью светового микроскопа. На фиг.1 показаны максимальные продольные и поперечные габаритные размеры пельтатных железок (А) и ее субкутикулярной полости (Б).
- Определяют коэффициент эксцентричности, отражающий наполненность железок секретом в случае визуально определяемой потери эластичности кутикулы или ее опадения.
- Для оценки формы каждой пельтатной железки и ее субкутикулярной полости высчитывается эксцентриситет:
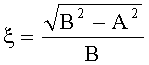 ;
; 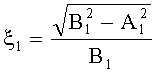 ,
,
где ξ - коэффициент эксцентричности пельтатной железки; ξ1 - коэффициент эксцентричности субкутикулярной полости.
Анализ показателя коэффициента эксцентричности пельтатной железки в динамике позволяет оценить степень ее наполненности.
- Оценку функциональной активности железок проводят путем:
а) анализа данных, полученных в результате расчета коэффициента интенсивности синтеза эфирных масел:
K=S1×n×10-3,
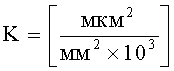 ,
,
где S1 - площадь субкутикулярной полости, в предсекреторной стадии пельтатной железки S1=0; n - число пельтатных железок на 1 мм2, позволяющего рассчитать синтетическую активность пельтатных железок на единицу площади листа;
б) оценки наполненности секретом пельтатных железок. Более наполненные железки имеют округлую форму, эксцентриситет приближается к нулю.
Технический результат
Предлагаемый способ позволяет: а) определить функциональную активность пельтатных железок на единицу площади листовой пластинки с учетом стадии созревания пельтатных железок, их площади; б) определить фазу вегетации, наиболее подходящую для сбора эфиромасличного сырья; в) выделить наиболее продуктивные формы среди популяций растений; г) определить топографию высокопродуктивных органов (например, адрес листа) на эфиромасличных растениях, которые могут идти на сырье.
Пример осуществления способа оценки функциональной активности пельтатных железок у смородины американской (Ribes americanum Mill.).
Объектом исследования являлись интродуцированные растения смородины американской и гибриды от свободного опыления отборных форм смородины американской (семья 2, 3, 4, 5), гибриды смородины американской от свободного опыления сорта Плутон (семья 1) 2002 года посадки, полученные в условиях культивирования, а также тетраплоид смородины американской. Растения высажены по схеме 3×1 м. Контролем послужил наиболее близкий в систематическом отношении вид R. nigrum ssp. sibirikum (Egb. Wolf.) Pavl., относящийся к тому же подроду Eucoreosma, что и экспериментальный образец, и давно интродуцированный в Белгородской области в виде сортов.
Эфирные масла у смородины американской синтезируются пельтатными железками, которые располагаются на адаксиальной и абаксиальной поверхностях и черешке листа.
Время исследования
Для исследований выбраны критические периоды в вегетационный период: цветение, формирование плодов, полная спелость.
Сбор материала
Для исследований отбирались листья с годичных приростов (4-й от основания прироста) с учетом их морфологического адреса, из верхнего яруса кроны куста, т.к. листья данного местоположения являются наиболее типичными и обладают максимальной фотосинтетической поверхностью. Листья отбирались в утренние часы, с одного яруса.
Определение концентрации пельтатных железок
Для определения концентрации пельтатных железок необязательно отделять эпидермис. Микроскопирование объектов проводили на бинокулярном МБС 10.
У смородины американской и ее гибридов концентрация пельтатных железок находится в пределах 8,19±1,47 (сем. 3, во время цветения) до 30,81±2,25 шт. на 1 мм2 площади листа (тетраплоид, полная спелость).
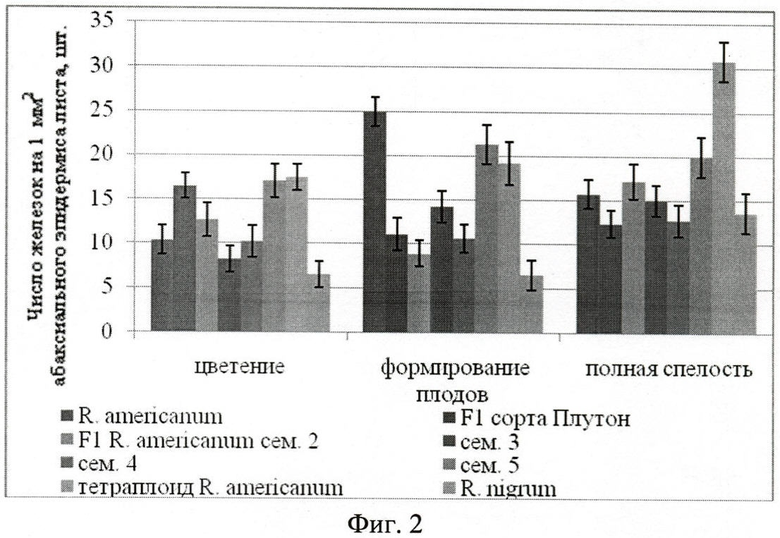
Концентрация железок на единицу поверхности абаксиальной стороны листа увеличивается к периоду формировании плодов, а после уменьшается в силу высыхания и отрывания железок. На фиг.2. отражена зависимость числа пельтатных железок на 1 мм2 абаксиального эпидермиса листа от фазы вегетации.
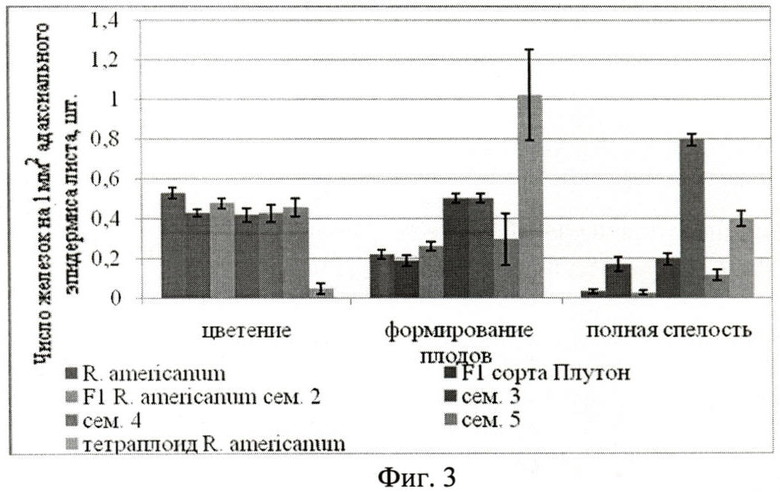
У представителей гибридов сорта Плутон (16,48±1,45 шт./мм2), гибридов 5-й семьи (17,09±1,89 шт./мм2) во время цветения число железок на адаксиальном эпидермисе достоверно превышает их число у смородины американской (10,37±1,60 шт./мм2) (Фиг.3. Число пельтатных железок на 1 мм2 адаксиального эпидермиса листа Ribes americanum Mill.) Во время формирования плодов их концентрация ниже (11,11±1,84 шт./мм2 сорт Плутон, 21,33±12,24 шт./мм2), чем у исходной смородины американской (24,92±1,65). К концу вегетации у представителей двух семей (3-й и 5-й) концентрация железок достоверно выше (20,74±1,76 для 3-й семьи, 26,91±1,3 0 шт./мм2 для 5-й семьи), чем у смородины американской (15,75±1,60 шт./мм2). У них она растет прямолинейно на протяжении всего периода вегетации, у остальных же семей концентрация несколько снижается. В период полного плодоношения наблюдается высыхание и отмирание железок.
Тетраплоид смородины американской имеет достоверно большую концентрацию пельтатных железок на верхней поверхности листа во время формирования плодов и полной спелости (1,02±0,230 формирование плодов, 0,398±0,038 шт./мм2 полная спелость), чем у смородины американской (0,219±0,022 формирование плодов, 0,031±0,012 шт./мм2 полная спелость).
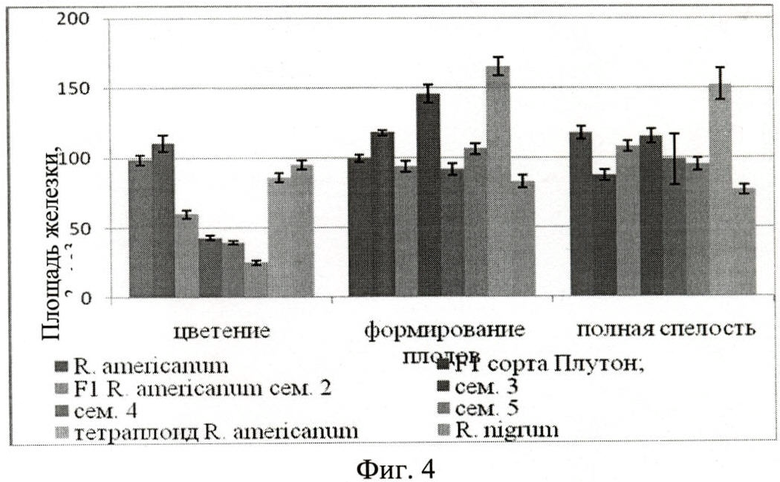
Определение площадей пельтатных железок и их субкутикулярных полостей Пельтатная железка и ее субкутикулярная полость имеют овальную форму. Измеряется их длинная и кроткая ось, вычисляется площадь железки и ее субкутикулярной полости по выведенным формулам. Наибольшая площадь пельтатных железок у большинства исследуемых гибридов нарастает к периоду формирования плодов, к концу вегетации происходит их уменьшение за счет высыхания (Фиг.4. Площадь железок абаксиального эпидермиса, мкм2×103).У смородины американской площадь железок нарастает в весь период вегетации (от 98,50±3,39×103 до 117,62±4,69×103 мкм2), данная закономерность наблюдается у представителей гибридов 2-й семьи (от 59,47±2,86×103 до 108,08±3,99×103 мкм2). Для смородины черной площадь пельтатных железок достоверно меньше (82,57±4,68×103, 76,62±3,81×103 мкм2), чем у смородины американской на протяжении всего периода вегетации, весь период вегетации этот показатель у смородины черной стабилен.
Во время цветения площадь железок гибридов достоверно меньше (от 24,47±1,69×103 до 59,47±2,86×103 мкм2), чем у смородины американской (98,50±3,39×103 мкм2). Гибриды сорта Плутон во время цветения и формирования плодов имеют более крупные железки (110,71±5,93×103 мкм2 (цветение), 117,89±2,04×103 мкм2 (формирование плодов)), чем у смородины американской (98,50±3,39×103 мкм2 (цветение), 99,56±2,45×103 мкм2 (формирование плодов)). Площадь железок тетраплоида во время формирования плодов (165,20±6,58×103 мкм2) и полной спелости 152,23±11,44 достоверно превышают эти значения у смородины американской (117,62±4,69×103 мкм2).
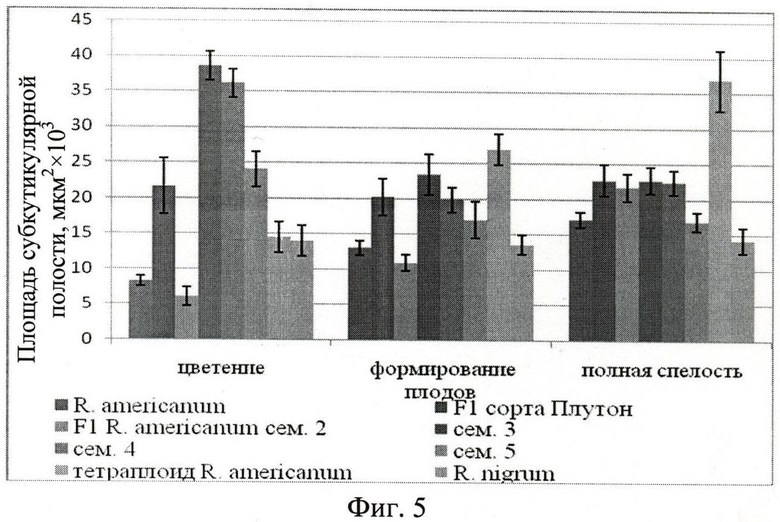
Выявлено, что площадь субкутикулярной полости пельтатных железок тем больше, чем больше секрета в них, характеризует активность синтеза эфирных масел (Фиг.5. Площадь субкутикулярной полости железок абаксиальной стороны листа, мкм2×103).
Во время цветения площадь субкутикулярной полости большинства гибридов от свободного опыления отборных форм смородины американской превышает ее у растений смородины американской, при этом площадь самой железки у гибридов меньше, чем у растений смородины американской. Этот факт свидетельствует о большой интенсивности испарения эфирных масел с поверхности кутикулы и высокой интенсивности синтеза секрета. У представителей гибридов 3-й, 4-й, 5-й семей наблюдается уменьшение интенсивности синтеза эфирных масел к периоду формирования плодов.
Площадь субкутикулярной полости у исходной смородины американской (в 2 раза), ее тетраплоида (в 2,5 раза) и у гибридов 2-й семьи (в 3,5 раза) к концу вегетации возрастает. Это говорит о большей устойчивости железок к неблагоприятным условиям смородины американской, ее тетраплоида и гибридов 2-й семьи. Для гибрида сорта Плутон интенсивность синтеза эфирных масел в период вегетации остается примерно на одном уровне.
К концу вегетации площадь железок отборных форм смородины американской от свободного опыления увеличивается в 1,21, а субкутикулярной полости в 2,14 раз. Такая же закономерность наблюдается и у тетраплоида смородины американской. Это можно интерпретировать тем, что секрет испаряется с поверхности железки, а интенсивность синтеза секрета несколько увеличивается.
Определение коэффициента эксцентричности
Установлено, что коэффициент эксцентричности пельтатных железок в течение вегетации смородины американской и гибридов сорта Плутон существенно не меняется.
Наименьший эксцентриситет в фазу формирования плодов, что совпадает с наибольшей площадью пельтатных железкок. Для гибридов семей 2, 3, 4, 5 наибольший эксцентриситет в фазу цветения (0,758±0,033 сем.2, 0,922±0,0061 сем. 3, 0,944±0,004 сем. 4, 0,964±0,0044 сем. 5). Для гибридных семей 3, 5 эксцентриситет наименьший в фазу формирования плодов (0,346±0,032 сем.3, 0,347±0,020 сем. 5). Тетраплоид смородины американской имеет коэффициент эксцентричности 0,408±0,028 в фазу цветения, 0,283±0,037 в фазу формирование плодов, 0,470±0,038 в фазу полной спелости.
Коэффициент эксцентричности субкутикулярных полостей пельтатных железок смородины американской уменьшается к концу вегетации, что совпадает с увеличением их площади (0,567±0,26 (цветение), 0,546±0,034 (формирование плодов), 0,480±0,037 (полная спелость)). Для тетраплоида эксцентриситет субкутикулярных полостей пельтатных железок существенно не изменяется в течение вегетационного периода. Коэффициент эксцентричности гибрида сорта Плутон к концу вегетации не существенно увеличивается: 0,409±0,056 (цветение) до 0,479±0,042 (полная спелость). У растений гибридной семьи 2 эксцентриситет уменьшается к концу вегетации, а площадь ее увеличивается.
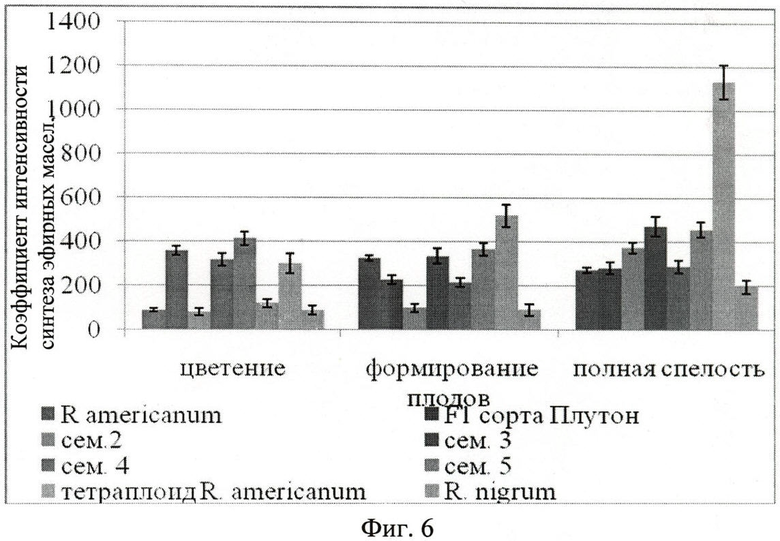
Определение коэффициента интенсивности пельтатных железок на единицу поверхности листа. Для оценки интенсивности синтеза эфирных масел на единицу поверхности листа находился коэффициент, учитывающий концентрацию пельтатных железок, площадь субкутикулярных полостей пельтатных железок (Фиг.6. Коэффициент интенсивности синтеза эфирных масел на единицу абаксиальной поверхности листа).
Для большинства гибридов (1-й, 3-й, 4-й, 5-й семей) интенсивность синтеза эфирных масел на единицу поверхности листа во время вегетации существенно не меняется. Для смородины американской, представителей гибридов 2-й семьи и тетраплоида интенсивность синтеза увеличивается к концу вегетации. Тетраплоид во время формирования плодов и полной спелости существенно отличается повышенной интенсивностью синтеза эфирных масел. Смородина черная имеет наименьшую интенсивность синтеза из всех исследуемых образцов. Площадь пельтатных железок и эксцентриситет имеют обратную зависимость. При увеличении наполненности железок секретом железка и ее субкутикулярная полость принимает более округлую форму, эксцентриситет уменьшается (r=-0,8). 0≤ξ≤1. Чем более округлая железка, тем ξ, ближе к 0, чем менее округлая железка, тем ξ, ближе к 1.
Таким образом, предлагаемый способ позволил сделать выводы.
Наибольшая интенсивность синтеза эфирных масел у тетраплоида R. americanum и гибридов (сем. 3, 5) в фазу полной спелости. У тетраплоида нарастание происходит не только за счет числа пельтатных железок на ед. абаксиальной поверхности листа, что определяется способом-прототипом [2], но и за счет увеличения площади железки и ее субкутикулярной полости, что учитывается заявляемым способом.
К концу вегетации происходит формирование небольших листьев с большей плотностью пельтатных железок. Несмотря на большую концентрацию пельтатных железок к концу вегетации (что расценивалось бы способом-прототипом как максимум биопродуктивности эфиромасличности) для гибридов сорта Плутон и семьи 4 смородины американской наибольшая интенсивность синтеза эфирных масел выявлена в фазу цветения. Площадь субкутикулярных полостей пельтатных железок наибольшая в фазу цветения, что отражает наибольшую синтетическую активность железок.
Сбор листьев рекомендуем производить из верхней кроны куста, т.к. верхний ярус имеет больше годичных приростов и, следовательно, листьев.
Для получения стабильного выхода эфирного масла из сырья листьев смородины американской, основываясь на нашем способе, рекомендуем использовать чередование сборов со следующих форм:
- гибриды сорта Плутон и семьи 4 смородины во время цветения;
- тетраплоид R. americanum и гибриды сем. 5 в период формирования плодов;
- тетраплоид R. americanum и гибридов (сем. 3, 5) в фазу полной спелости.
Список литературы
1. Корчашкина Н.В. Биологические особенности роста и развития видов рода Монарда (Monarda L.) в условиях нечерноземной зоны Российской Федерации / Н.В.Корчашкина. Автореферат диссертация на соискание ученой степени кандидата биологических наук. Москва, 2009, 23 с.
2. Паушева З.П. Практикум по цитологии растений / З.П.Паушева. - М.: «Колос», 1970, 256 с.
3. Практикум по фармакогнозии: Учебное пособие для студентов вузов / В.Н.Ковалев, Н.В.Попова, B.C.Кисличенко и др.; Под общей ред. В.Н.Ковалева. - Xарьков: Изд-во НФаУ; Золотые страницы, 2003. - 512 с.
4. Работягов В.Д. Изучение генетических закономерностей накопления эфирного масла у межвидовых гибридов лаванды // Частная генетика растений. - Киев, - 1989. - Т.2 - С.47-48.
5. Работягов В.Д., Машанов В.И., Андреева А.Ф Интродукция эфиромасличных и пряных растений. - Ялта: ГНБС, 1999. - 32 с.
6. Работягов В.Д., Федорчук М.И., Гридин А.Е. Анатомоморфологическое исследование шалфея лекарственного // Материалы XII Международной конференции «Вивчення онтогенезу рослин природних i культурних флор у ботанiчних закладах i дендрарках Евразii». - Полтава, 2000. - С.269-271.
7. Танасиенко Ф.С. Эфирные масла, содержание и состав в растениях / Ф.С.Танасиенко. - Киев: Наук. Думка, 1985, 264 с.
8. Фурст Т.Г. Методы анатомогистохимического исследования тканей. М.: Наука, 1979, 155 с.
9. Ascensao L., Pais M.S. The leaf capitate trichomes of Leonotis leonorus: histochemistry, ultastructure and secretion // Ann Bot, - 1998. - Vol 81. - P.277-286.
10. Croteau R., Gershenson J. Genetic control of monoterpene biosynthesis in mint (Mentha: Lamiaceae) // Recent Adv Phytochem. - 1994. - Vol.28. - P.193-229.
11. Heinrich G. Entwiklung Feinbau und Olgehalt der Drusenschuppfen von Monarda fistulosa // Planta medica. - 1973. - Vol.23; N2. - P.154-166.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ЭФИРНОГО МАСЛА В ЛИСТЬЯХ РАСТЕНИЙ МЯТЫ ПО СУММАРНОМУ ОБЪЕМУ СЕКРЕТОРНЫХ ЖЕЛЕЗ | 2012 |
|
RU2490866C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ОБЪЕМА ФРАКЦИИ ЛЕГКОЛЕТУЧИХ СОЕДИНЕНИЙ В ЭФИРНОМ МАСЛЕ ЛИСТА МЯТЫ | 2012 |
|
RU2490867C1 |
| Способ отбора высокопродуктивных форм лаванды | 1987 |
|
SU1421279A1 |
| СПОСОБ ВОЗДЕЛЫВАНИЯ ПЕРЦА СЛАДКОГО, ПРЕИМУЩЕСТВЕННО В СИСТЕМЕ КАПЕЛЬНОГО ОРОШЕНИЯ | 2009 |
|
RU2415534C2 |
| СПОСОБ ВОЗДЕЛЫВАНИЯ ОГУРЦА, ПРЕИМУЩЕСТВЕННО В СИСТЕМЕ КАПЕЛЬНОГО ОРОШЕНИЯ | 2007 |
|
RU2354094C1 |
| Способ возделывания подсолнечника | 2022 |
|
RU2791876C1 |
| Способ повышения урожайности подсолнечника | 2023 |
|
RU2808542C1 |
| Способ подбора гибридов томата для промышленных теплиц зоны Урала | 2020 |
|
RU2740106C1 |
| Фиточай из 77 трав | 2016 |
|
RU2664459C2 |
| СПОСОБ ВОЗДЕЛЫВАНИЯ ОВОЩНЫХ КУЛЬТУР, ПРЕИМУЩЕСТВЕННО ТОМАТОВ, ПРИ КАПЕЛЬНОМ ОРОШЕНИИ | 2010 |
|
RU2432728C1 |
Изобретение относится к области анатомических методов исследования, к фармокогнозии и селекции. Способ включает сбор материала, определение концентрации пельтатных железок путем подсчета количества пельтатных железок в единице площади поля зрения не менее чем в 25 полях зрения в средней части листовой пластинки. Определяют площади пельтатных железок и их субкутикулярных полостей по формулам 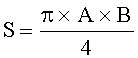 ;
; 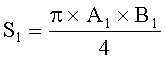 , где S - площадь пельтатной железки; S1 - площадь субкутикулярной полости, А и В - максимальные поперечные и продольные габаритные размеры пельтатной железки, A1 и B1 - максимальные поперечные и продольные габаритные размеры субкутикулярной полости пельтатной железки. Определяют коэффициент эксцентричности по формуле
, где S - площадь пельтатной железки; S1 - площадь субкутикулярной полости, А и В - максимальные поперечные и продольные габаритные размеры пельтатной железки, A1 и B1 - максимальные поперечные и продольные габаритные размеры субкутикулярной полости пельтатной железки. Определяют коэффициент эксцентричности по формуле 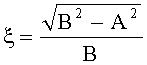 ;
; 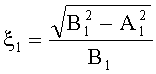 , где ξ - коэффициент эксцентричности пельтатной железки; ξ1 - коэффициент эксцентричности субкутикулярной полости. Причем оценку функциональной активности пельтатных железок ведут путем анализа данных, полученных в результате расчета коэффициента интенсивности синтеза эфирных масел по формуле K=S1×n×10-3,
, где ξ - коэффициент эксцентричности пельтатной железки; ξ1 - коэффициент эксцентричности субкутикулярной полости. Причем оценку функциональной активности пельтатных железок ведут путем анализа данных, полученных в результате расчета коэффициента интенсивности синтеза эфирных масел по формуле K=S1×n×10-3, 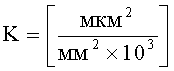 , где S1 - площадь субкутикулярной полости, в предсекреторной стадии пельтатной железки S1=0; n - число пельтатных железок на 1 мм2, что позволяет рассчитать синтетическую активность пельтатных железок на единицу площади листа и оценить наполненность секретом - более наполненные пельтатные железки имеют округлую форму, эксцентриситет приближается к нулю. Способ позволяет выделить наиболее продуктивные формы среди популяций растений. 6 ил., 1 пр.
, где S1 - площадь субкутикулярной полости, в предсекреторной стадии пельтатной железки S1=0; n - число пельтатных железок на 1 мм2, что позволяет рассчитать синтетическую активность пельтатных железок на единицу площади листа и оценить наполненность секретом - более наполненные пельтатные железки имеют округлую форму, эксцентриситет приближается к нулю. Способ позволяет выделить наиболее продуктивные формы среди популяций растений. 6 ил., 1 пр.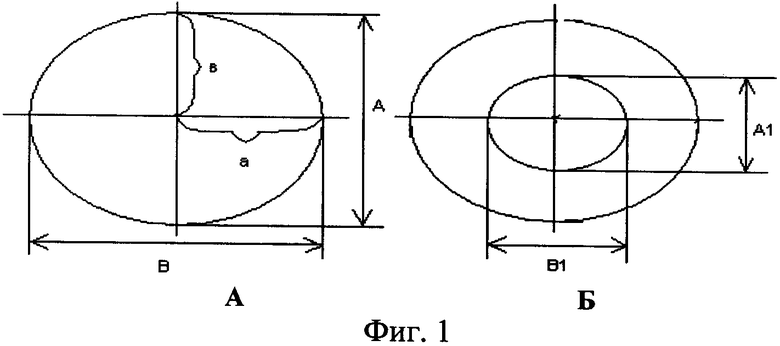
Способ оценки функциональной активности пельтатных железок, включающий: сбор материала, определение концентрации пельтатных железок путем подсчета количества пельтатных железок в 1 площади поля зрения не менее чем в 25 полях зрения в средней части листовой пластинки, определение площади пельтатных железок и их субкутикулярных полостей по формулам 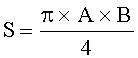 ;
; 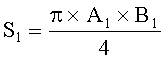 , где S - площадь пельтатной железки; S1 - площадь субкутикулярной полости, А и В - максимальные поперечные и продольные габаритные размеры пельтатной железки, A1 и B1 - максимальные поперечные и продольные габаритные размеры субкутикулярной полости пельтатной железки; определение коэффициента эксцентричности по формуле
, где S - площадь пельтатной железки; S1 - площадь субкутикулярной полости, А и В - максимальные поперечные и продольные габаритные размеры пельтатной железки, A1 и B1 - максимальные поперечные и продольные габаритные размеры субкутикулярной полости пельтатной железки; определение коэффициента эксцентричности по формуле 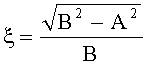 ;
; 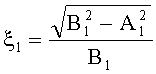 , где ξ - коэффициент эксцентричности пельтатной железки; ξ1 - коэффициент эксцентричности субкутикулярной полости, причем оценку функциональной активности пельтатных железок ведут путем анализа данных, полученных в результате расчета коэффициента интенсивности синтеза эфирных масел по формуле K=S1×n×10-3,
, где ξ - коэффициент эксцентричности пельтатной железки; ξ1 - коэффициент эксцентричности субкутикулярной полости, причем оценку функциональной активности пельтатных железок ведут путем анализа данных, полученных в результате расчета коэффициента интенсивности синтеза эфирных масел по формуле K=S1×n×10-3, 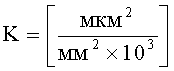 , где S1 - площадь субкутикулярной полости, в предсекреторной стадии пельтатной железки S1=0; n - число пельтатных железок на 1 мм2, что позволяет рассчитать синтетическую активность пельтатных железок на единицу площади листа и оценить наполненность секретом - более наполненные пельтатные железки имеют округлую форму, эксцентриситет приближается к нулю.
, где S1 - площадь субкутикулярной полости, в предсекреторной стадии пельтатной железки S1=0; n - число пельтатных железок на 1 мм2, что позволяет рассчитать синтетическую активность пельтатных железок на единицу площади листа и оценить наполненность секретом - более наполненные пельтатные железки имеют округлую форму, эксцентриситет приближается к нулю.
| РЕЗАНОВА Т.А | |||
| Морфо-анатомические и экологические особенности RIBES AMERICANUM MILL | |||
| при интродукции на юге Среднерусской возвышенности | |||
| Автореф | |||
| дисс | |||
| на соиск | |||
| уч | |||
| ст | |||
| к.б.н | |||
| Саратов, 2010, автореф | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| ЕГОРОВА А.В | |||
| и др | |||
| Анатомо-морфологическое исследование плодов Ribes nigrum (Grossulariaceae) // Известия | |||

