Область техники, к которой относится изобретение
Данное изобретение относится к реакциям амплификации нуклеиновых кислот и анализам с использованием полимеразной цепной реакции (ПЦР), включая гомогенные анализы, как в режиме реального времени, так и конечных результатов.
Уровень техники
Хорошо известна амплификация нуклеиновых кислот с использованием полимеразной цепной реакции (ПЦР), также как анализы, которые включают ПЦР-амплификацию. См. патенты США 4683202, 4683195 и 4965188 и в основном монографию PCR Protocols, a guide to Methods и Applications (Протоколы ПЦР, руководство по методам и применению) под ред. Innis et al., Academic Press (San Diego CA (USA) 1990). Кроме того, хорошо известны гомогенные ПЦР-анализы, в которых не требуется промывание для удаления несвязанных детекторных реагентов или зондов, и которые, таким образом, можно проводить, не открывая емкости для реакции амплификации. Однородные ПЦР-анализы включают как анализы конечных результатов, в которых амплифицированный продукт определяют в конце реакции амплификации, так и анализы в режиме реального времени, в которых амплифицированный продукт определяют во время некоторых или всех температурных циклов по мере протекания реакции. См. патенты США 5994056, 5487972, 5925517 и 6150097.
Как правило, разрабатывают симметричные реакции ПЦР-амплификации, что означает создание двунитевых ампликонов при использовании прямого праймера и обратного праймера, которые "составляют пару", что означает, что они имеют температуры плавления, которые настолько близки, насколько это возможно, и их вводят в реакцию в эквимолярных концентрациях. Методика, которая нашла ограниченное применение для получения однонитевой ДНК непосредственно в реакции ПЦР, представляет собой "асимметричную ПЦР". См. статью Gyllensten и Erlich, "Generation of Single-Stranded ДНК by the Polymerase Chain Reaction and Its Application to Direct Sequencing of the HLA-DQA Locus" (Генерация однонитевой ДНК посредством полимеразной цепной реакции и ее применение для прямого секвенирования локуса HLA-DQA), Proc. Natl. Acad. Sci. (USA) 85:7652-7656 (1988) и патент США 5066584. Асимметричная ПЦР отличается от симметричной ПЦР тем, что один из праймеров вводят в ограничивающем количестве, как правило, 1-20% от концентрации другого праймера.
Совсем недавно авторы разработали способ асимметричной ПЦР-амплификации, известный как "Линейная после экспоненциальной" ПЦР или сокращенно "ЛПЭ-ПЦР" ("LATE-PCR"). См. статьи Sanchez et al. (2004) PNAS 101: 1933-1938, Pierce et al. (2005) PNAS 102: 8609-8614 и опубликованную патентную заявку WO 03/054233 (3 июля 2003), которая включена в данном контексте в виде ссылки во всей полноте. При ЛПЭ-ПЦР принимают во внимание, что действительные температуры плавления ПЦР-праймеров в начале амплификации, обозначаемые как Tm[o]-Tm[o], можно определить эмпирически, что является необходимым, когда используют неприродные нуклеотиды или рассчитать согласно методу "ближайшего соседа" (см. статьи Santa Lucia J. (1998) PNAS (USA) 95: 1460-1465 и Allawi, H.T. и Santa Lucia J. (1997) Biochem. 36: 10581-10594) с использованием подведения концентрации солей. В данной работе авторы используют концентрацию одновалентной соли 0,07 М.
Нежелательной характеристикой ПЦР-амплификаций, сниженной в случае ЛПЭ-ПЦР, является разброс копий. После экспоненциальной фазы амплификации следующие в режиме реального времени амплификации копий расходятся и выходят на плато на различных уровнях. Разброс показывает, что копии не имеют одинаковой кинетики реакции и уменьшают точность. Это представляет собой проблему для ПЦР-анализов в общем, но в особенности в анализах конечных результатов и анализах, которые зависят от крутизны сигнала во время линейной фазы.
Другой существенной проблемой для ПЦР-амплификаций является ошибочный старт, который, как считают авторы, обнаружен по меньшей мере трех типов: Тип 1 - ошибочный старт, который происходит во время приготовления реакционной смеси перед началом амплификации; Тип 2 - ошибочный старт, который происходит во время амплификации, если температуры цикла включают какую-либо температуру, существенно ниже температуры плавления праймера, и Тип 3 - ошибочный старт, который происходит на поздних стадиях ПЦР-амплификаций, которая продолжается после того, как получена высокая концентрация ампликона. Использованы различные подходы, касающиеся первого типа ошибочного старта. Одним из подходов является модифицировать полимеразу химическим путем, чтобы она была неактивной до нагревания до высокой температуры, такой как 95°C. См. патенты США 5677152 и 5773258. Другим подходом является связывание антитела с полимеразой, чтобы ингибировать полимеразу до нагревания реакции до высокой температуры, такой как 95°C, чтобы необратимо денатурировать антитело. См. патент США 5338671. Еще одним подходом является включение аптамера в реакционную смесь. См. статью Doug и Jayasena (1996), J. Mol. Biol. 264: 268-278 и патент США 6020130. Аптамер представляет собой однонитевый олигонуклеотид длиной приблизительно 30 нуклеотидов, который связывается с полимеразой и ингибирует ее способность к удлинению углубленного 3'-конца при низких температурах. Аптамеры необратимо денатурируются при 95°C, типичной самой высокой температуре для цикла ПЦР. Kainz et al. (2000) Biotechniques 28: 278-282 показали, что добавление к реакционным смесям для ПЦР фрагментов двунитевой ДНК длиной 16-21 нуклеотидов в определенных количествах ингибирует полимеразы при температурах ниже типичных температур ПЦР-удлинения и подавляет синтез неспецифических продуктов. ДНК-фрагменты не денатурируются необратимым образом во время циклинга ПЦР. Фирма Eppendorf-5 Prime, Inc. выпускает на рынок патентованный лиганд, который, как полагают, связывается с полимеразой Taq температурозависимым образом и ингибирует ее связывание с двунитевой ДНК при температурах ниже приблизительно 50°C. Несмотря на данные многочисленные попытки ошибочный старт остается проблемой, присущей ПЦР-амплификации.
Другое проявление ошибочного старта в процессе ПЦР-амплификации известно как образование и амплификация димера праймера. Согласно данному феномену, один праймер гибридизуется с другим праймером или сам с собой и затем подвергается удлинению 3'-конца с генерацией маленького двунитевого ампликона, который затем может амплифицироваться дальше или может мультимеризоваться и амплифицироваться дальше. Формирование димера праймера может происходить в отсутствие мишени.
Количественный анализ ПЦР-амплификаций осуществляют методами детекции в режиме реального времени, поскольку цикл ПЦР, в котором флуоресцентный сигнал становится видимым выше порогового цикла или СТ реакций, является показателем исходных концентрацией мишени. Анализы конечных результатов являются в лучшем случае полуколичественными, отчасти вследствие разброса копий, поскольку реакция выходит из рамок экспоненциальной амплификации. Электрофоретический анализ двунитевых ампликонов является полуколичественным, и в нем можно использовать праймеры с флуоресцентной меткой. Анализ конечных результатов с использованием зондов с флуоресцентной меткой, либо аллель-дискриминирующих зондов, либо зондов, толерантных к ошибочному спариванию, в лучшем случае также является полуколичественным. Вследствие уменьшения разброса и образования однонитевого продукта ЛПЭ-ПЦР дает существенное усовершенствование анализа конечных результатов, но разброс копий часто устраняется неполностью, делая количественную многократную детекцию менее точной, чем требуется.
Аспектом данного изобретения является класс дополнительных реагентов для повышения специфичности продукта и устранения эффектов ошибочного старта в реакциях ПЦР-амплификации. Данные дополнительные компоненты позволяют усовершенствовать имеющиеся методики "горячего старта" во всех типах ПЦР и могут быть использованы для предупреждения накопления нежелательных продуктов, включая димеры праймеров и ошибочно запущенные ампликоны, как на ранних стадиях, так и во время реакций ЛПЭ-ПЦР с многочисленными циклами (как правило, 60 и более циклов).
Другой аспект изобретения представляет собой амплификацию и способы анализа с использованием ПЦР, как симметричной ПЦР, так и асимметричной ПЦР, включая, но без ограничения перечисленным ЛПЭ-ПЦР, а также наборы, частичные наборы и группы олигонуклеотидов, которые включают указанные дополнительные реагенты.
Раскрытие изобретения
Реагенты, соответствующие данному изобретению, представляют собой дополнительные компоненты, которые способны предотвращать одно или более проявлений ошибочного старта в по меньшей мере некоторых ПЦР-амплификациях. Под выражением "предотвращать проявление" авторы понимают, что продукт или продукты ошибочного старта не определяются в конце реакции методиками, описанными в данном контексте, а именно флуоресцентным окрашиванием ДНК, гель-электрофорезом, секвенированием ДНК и анализом температуры плавления. Реагенты, соответствующие данному изобретению, могут быть включены в смеси для ПЦР-амплификаций перед началом амплификации в относительно низкой концентрации, меньше, чем 1 микромоль (мкМ), что означает 1000 нМ, предпочтительно не больше, чем 650 наномоль (нМ), более предпочтительно не больше, чем 300 нм, и наиболее-предпочтительно 50-250 нМ, даже при использовании полимераз, которые обладают как активностью полимеризации, так и 5'-3'-экзонуклеазной активностью. Реагенты, соответствующие данному изобретению, представляют собой модифицированные однонитевые олигонуклеотиды. Олигонуклеотиды, которые можно использовать для конструирования реагентов, соответствующих данному изобретению, являются олигонуклеотидами в широком смысле слова. Они могут представлять собой ДНК, РНК или смешанную ДНК-РНК. Они могут включать модифицированные нукпеотиды, неприродные нуклеотиды, например, 2'0-метилрибонуклеотиды, неприродные межнукпеотидные связи, ненуклеотидные линкеры, ПНК, ЛНК и введенные химические группы, такие как нуклеотиды с кэпом, описанные Glenn Research. Реагенты, соответствующие данному изобретению, представляют собой однонитевые олигонуклеотиды, которые образуют структуру типа ствол-петля, обычно называемую структурой "шпильки" ("шпилечной" структурой), в реакционной смеси для ПЦР-амплификации, хотя они могут также состоять из структуры типа ствол-петля, в которой петля не состоит из нуклеотидов. В данной структуре центральная часть молекулы остается однонитевой (негибридизованной) (петлей), а концы гибридизуются друг с другом с образованием стволаствола. Ствол может быть с тупыми концами или одна нить может выходить за пределы другой. Ствол может включать непрерывный двунитевый участок, или он может включать внутреннее ошибочное спаривание, приводящее к образованию выступа. Окончание стволаствола, образованное концами олигонуклеотида, стабилизируют, чтобы они были более тесно связаны, чем гибрид ДНК-ДНК. Авторы характеризуют ствол реагента, соответствующего изобретению, по его температуре плавления или Тm. Температура плавления представляет собой такую температуру в градусах по шкале Цельсия, при которой 50% комплементарных последовательностей ствола негибридизованы (открытая конфигурация) и 50% комплементарных последовательностей гибридизованы или частично гибридизованы сами с собой (закрытая конфигурация). В данной заявке выражение "рассчитанная Тт" ствола означает рассчитанную температуру плавления части ствола соответствующего нестабилизированного олигонуклеотида, полностью состоящего из ДНК, полученную при использовании программы M-fold: см. статью Zucker М. (2003), "Mfold Web Server for Nucleic Acid Folding и Hybridization Prediction" (Веб-сервер Mfold для предсказания укладки и гибридизации нуклеиновых кислот), Nucl. Acids Res. 31: 3406-3415, предполагающей концентрацию натрия 70 миллимоль (мМ) и концентрацию магния 3 мМ. В случае предпочтительных вариантов осуществления с наличием добавленных пар гасителей, предпочтительно нефлуоресцентных гасителей, таких как Дабцил и гасители Black Hole, которые поглощают свет, но испускают поглощенную энергию в виде тепла, Тт представляет собой рассчитанную Тт шпильки ДНК без гасителей. В случае вариантов осуществления с наличием нуклеотидов с кэпом на концах Тт представляет собой рассчитанную Тт шпильки ДНК, включающей эквивалентные нуклеотиды без кэпа. В случае включений 2'-0-метил рибонуклеотидов на конце ствола Тт представляет собой рассчитанную Тт шпильки, полностью состоящей из ДНК, включающей дезоксирибонуклеотидные аналоги 2'-0-метилрибонуклеотидов. Авторы используют данную методику из практических соображений, заключающихся в том, что трудно получить действительную температуру плавления стабилизированной шпильки, признавая, что действительная температура плавления будет на несколько градусов выше, чем рассчитанная ТТ вследствие стабилизирующей модификации. Реагенты, соответствующие данному изобретению, имеют рассчитанную ТТ ствола, которая не превышает 94°C и предпочтительно лежит в интервале 50-85°C. Для ряда вариантов осуществления предпочитают рассчитанную ТТ ствола, которая превышает температуру отжига праймера (как правило, 55-72°C для реакции амплификации, т.е. в интервале 72-85°C), но в других вариантах осуществления предпочитают рассчитанную ТТ ствола, которая ниже, чем температура отжига праймера, т.е. лежит в интервале 50-71°C. Комплементарные последовательности щели ствола становятся однонитевыми при 95°C, температуре, используемой для плавления нити во время циклинга ПЦР. Кроме того, ствол реагентов, соответствующих данному изобретению, имеет длину, превышающую шесть нукпеотидов при температурах достаточно низких для формирования закрытой самогибридизующейся конформации. В настоящее время предпочтительные варианты осуществления формируют стволы, которые имеют длину 9-12 пар оснований, предпочтительно 9-11 пар оснований в замкнутом состоянии, более предпочтительно 9-12, наиболее предпочтительно 9-11, непрерывных комплементарных пар оснований без внутренних ошибочных спариваний, более предпочтительно стволы с тупыми концами, которые имеют длину 9-12 пар оснований и в полной мере комплементарны. 3'-конец реагента не удлиняется ДНК-полимеразой в реакции амплификации, вследствие этого реагент не является ПЦР-праймером. Любой свешивающийся 3'-конец блокирован, чтобы предупредить удлинение, например, введением фосфата или какой-либо блокирующей химической группы. Длина петли может значительно варьировать. Если петля состоит из нуклеотидов и межнуклеотидных связей, ее длина составляет по меньшей мере три нуклеотида. Кроме того, если ее длина составляет ровно три нуклеотида, она включает остаток тимидина. Представленные авторами в настоящее время предпочтительные нуклеотидные петли имеют длину в интервале 3-22 нуклеотидов. Петли могут также представлять собой ненукпеотидные химические линкеры, например, алкиленовые цепи. Для линкеров в виде углеродных цепей авторы предпочитают длину 3-6 атомов углерода и наиболее предпочтительную длину 3 атома углерода. В алкиленовых углеродных цепях электроны остальных двух валентностей каждого члена углеродной цепи также участвуют в ковалентных связях. Данные связи могут быть с атомами водорода, с алкильными или алкиленовыми группами с короткой цепью, с заместителями для связывания реагента с твердой поверхностью. Неолигонуклеотидные петли, включающие химические линкеры, не ограничены углеводородными цепями и могут включать гетеро- (отличные от углерода и водорода) атомы. Предпочитают, чтобы химические линкеры были электронейтральными, т.е. не связывались с полимеразой. Поскольку активность реагента зависит от замкнутой конформации ствола, предпочтительно, чтобы состав петли для реагентов с петлями из больше, чем 5 нуклеотидов, не обладал значительной комплементарностью в отношении любой другой последовательности или последовательностей, участвующих в реакции или генерированных посредством реакции. Реагент, соответствующий данному изобретению, является гибридизационным зондом для какого-либо продукта реакции амплификации и не дает сигнала о накоплении продукта.
Как указано, реагенты, соответствующие данному изобретению, включают стабилизацию конца ствола, удаленного от петли, так что конец ствола связан более тесно, чем конец гибрида ДНК-ДНК. В представленных наиболее предпочтительных вариантах осуществления ствол имеет тупой конец, и конец ствола модифицируют таким образом, чтобы он ингибировал частичное разделение нитей по сравнению с естественным гибридом ДНК-ДНК. Ниже авторы демонстрируют эффект введения стабилизирующих интерактивных химических групп в концы ствола, как пары групп Дабцила, ковалентно присоединенных к 3'- и 5'-нуклеотидам ствола посредством коммерческого линкера, так и пары коммерчески доступных гасителей Black Hole™ (патентованных гасителей, представляемых на рынке фирмой Biosearch Technologies, Novato, СА, U.S.A.). Авторы демонстрируют также эффект применения сильно связывающих неприродных нуклеотидов, 2'О-метилрибонуклеотидов, на конце ствола. Предпочтительные стабилизирующие группы являются нефлуоресцентными, так что реагенты не вносят вклад в фоновую флуоресценцию при амплификациях и анализах амплификации. Нестабилизированные олигонуклеотидные ДНК-шпильки реагентов, соответствующих данному изобретению, вводимые в концентрациях ниже 1 мкМ, не обязательно в существенной мере уменьшают ошибочный старт, если они не модифицированы, как описано, с целью стабилизации связанного с водородом конца ствола, и во многих случаях не осуществляют этого. Таким образом стабилизация важна в реагентах, соответствующих данному изобретению.
Реагенты в виде шпильки, соответствующие данному изобретению, можно описать как молекулы, которые включают два комплементарных нуклеотидных олигомера, которые удерживают вместе так, чтобы они могли сформировать ствола из больше, чем 6 пар оснований, и открытый конец ствола химически модифицируют одним или другим образом, чтобы подавить тенденцию ствола к раскручиванию или "расплетанию" открытого конца. Хотя полное понимание молекулярного механизма действия предполагает проведение дальнейшего анализа на основе кинетики фермента в присутствии и в отсутствие реагентов, соответствующих данному изобретению, а также структурного анализа взаимодействия данных реагентов с полимеразой с помощью электронной микроскопии, ядерного магнитного резонанса и рентгенокристаллографии, частичное понимание механизма можно предположить на основе информации, представленной в данном материале и научной литературе, В литературе описано, что полимераза Taq, подобно другим ДНК-полимеразам, состоит из синтетического домена и 5'-экзонуклеолитического домена. 5'-экзонукпеолитический домен не присутствует во фрагменте Стоффеля. Кроме того, в литературе описано, что синтетический домен осуществляет полимеризацию ДНК путем первичного связывания и скольжения вдоль двунитевой ДНК. Синтетический домен имеет форму, напоминающую "раскрытую кисть руки", с которым в присутствии дНТФ происходит изменение формы на полипептидном уровне при контакте с двунитевой ДНК. В результате "пальцы руки сжимаются" вокруг молекулы двунитевой ДНК. Ошибочно спаренные последовательности в двунитевой молекуле приводят к тому, что полимераза восстанавливает прежнее состояние некоторых нуклеотидов и заменяет правильным основанием, используя функцию З'-редактора синтетического домена. Синтез новой нитиДНКосуществляется, поскольку фермент считывает матрицу и удлиняет З'-конец комплементарного праймера. Если фермент встречает 5'-конецолигонуклеотида, уже связанный с матричной нитью, полимераза может войти на участок связанного олигонуклеотида на 1-2 нуклеотида и отщепить полученный 5'-конец с помощью 5'-экзонуклеазного домена фермента.
Основываясь на данной информации, можно предположить, что двунитевый ствол реагента, соответствующего данному изобретению, действует, по меньшей мере частично, связывая синтетический домен полимеразы в открытой конформации и вызывая его закрытие. Следовательно происходит ингибирование синтетической активности полимеразы. Таким образом, хотя и не желая быть связанными какой-либо теорией, можно допустить, что Первым способом действия является действие в качестве температурозависимого ингибитора синтетической активности полимеразы. Факты, представленные ниже, показывают, что по меньшей мере некоторые варианты реагентов не высвобождаются ферментом при температуре плавления их двунитевых участков, например, стволов, но остаются связанными при более высоких температурах, особенно включая типичные температуры ПЦР-удлинения. Однако во время стадии денатурации (или плавления нити) цикла ПЦР реагенты, соответствующие данному изобретению, перестают быть связанными с ДНК-полимеразой. Во время стадии денатурации циклов ПЦР циклов (обозначаемой также как стадия плавления нити), как правило, при температуре свыше 90°C, стволы шпилечных реагентов, соответствующих данному изобретению, плавятся отдельно, и они снова связываются с полимеразой только если и когда температура падает достаточно для преобразования двунитевого участка. Вследствие этого можно выбрать конструкцию реагента, соответствующего данному изобретению, который является двунитевым только до первой денатурации, т.е. является двунитевым при добавлении в реакционную смесь для ПЦР перед амплификацией (обычно при комнатной температуре), но после этого его поддерживают при температуре выше Тт его ствола. Для данного варианта осуществления температура отжига праймера всех циклов амплификации и любая температура низкотемпературной детекции ЛПЭ-ПЦР выше, чем рассчитанная Тт ствола, предпочтительно по меньшей мере на 5°C. Альтернативно можно выбрать конструкцию реагента, соответствующего данному изобретению, который позднее снова становится двунитевым во время амплификации. Это можно осуществить, используя Тт ствола (рассчитанную как описано), которая выше, чем температура отжига праймера, используемая во всех или некоторых циклах амплификации. Альтернативно в ЛПЭ-ПЦР-амплификации это можно осуществить при использовании Тт ствола, которая ниже температуры отжига праймера, но выше, чем температура низкотемпературной детекции. Конструкция первого типа, как правило, не будет ингибировать полимеризацию, как показывают по задержке в Ст, тогда как использование температуры отжига праймера ниже Тт ствола обычно будет давать в результате задержку Ст 1-3 циклов амплификации.
Кроме того, хотя и не желая быть связанными какой-либо теорией, можно допустить, что Вторым способом действия реагентов, соответствующих данному изобретению, является действие в качестве температурозависимого ингибитора 5'-экзонуклеолитической активности полимеразы. Ниже показывают, что реагенты, соответствующие данному изобретению, ингибируют 5'-3' экзонуклеазную активность фермента полимеразы при температурах до по меньшей мере 55°C и осуществляют это без чрезмерной супрессии способности фермента удлинять нить ДНК путем полимеризации с ее З'-концом (как ПЦР-праймеры удлиняются при ПЦР-амплификациях).
Кроме того, хотя и не желая быть связанными какой-либо теорией, можно допустить, что Третьим способом действия реагентов, соответствующих данному изобретению, является действие в качестве температурозависимого лиганда, который связывает полимеразу, вызывая изменение формы полипептидов полимеразы. После высвобождения лиганда из фермента данный фермент сохраняет измененную форму в течение некоторого времени перед тем, как вернуться в прежнюю форму. При этом в измененной форме фермент преимущественно связывается с полностью комплементарными гибридами праймер-матрица по сравнению с частично ошибочно спаренными гибридами праймер-матрица.
Основываясь на данной модели, можно предположить, что способность ствола реагентов, соответствующих данному изобретению, изменять форму синтетического домена полимеразы отличается от способности реагентов, соответствующих данному изобретению, ингибировать либо функцию синтеза полимеразы (Первый способ, см. выше), либо функцию 5'-экзонуклеазы (Второй способ, см. выше). Оба, Первый способ и Второй способ действия, в действительности зависят от скорости, с которой связанный реагент высвобождается полимеразой. Реагенты, соответствующие данному изобретению, которые высвобождаются быстро, преимущественно действуют третьим способом без существенного ингибирования синтетической активности полимеразы, даже в высокой концентрации. Напротив, реагенты, соответствующие данному изобретению, которые высвобождаются из полимеразы медленно, по-видимому, являются ингибирующими в высокой концентрации. Например, как соединение 12-C3DD, состоящее из ствола и петли углеродного линкера из трех метиленовых (-СН2-) групп, так и соединение 12-C3C3DD, состоящее из ствола и петли углеродного линкера из шести метиленовых (-СН2-) групп, только частично ингибируют ПЦР-амплификацию в концентрациях 3000 нМ. Напротив, реагенты, соответствующие данному изобретению, имеющие петли, состоящие из нуклеотидов, как правило, ингибируют ПЦР-амплификацию полностью в концентрации <1000 нМ и иногда в концентрациях меньше, чем <500 нМ в анализе в Примере 1.
Как будут иметь в виду специалисты, знакомые с взаимодействиями типа белок-нуклеиновая кислота, действие реагента, соответствующего данному изобретению, может выходить за рамки Способов 1-3, которые не являются взаимоисключающими, но являются близкими. Как описано ниже, авторами разработан количественный тест (см. Пример 14), который можно использовать для определения уровня, до которого определенная молекулярная структура, связывающая два олигонуклеотида, включающих ствол, обусловливает действие целого соединения как ингибитора фермента (посредством Первого способа или Второго способа) или в основном как агента, усиливающего специфичность фермента, Третий способ действия. Основываясь на вышеописанной модели, можно далее предположить, что одна или более молекул полимеразы могут быть связаны с равным числом молекул реагента, соответствующего изобретению, который, в свою очередь, ковалентно связан с более крупной структурой, такой как гранула, частица или материал, сделанный из твердого вещества. Твердое вещество можно, таким образом, считать "нагруженным" полимеразой. Связь реагентов, соответствующих данному изобретению с твердым веществом может быть временной или может быть расщепляемой рядом средств, известных в области техники. Расщепление связи приводило бы к выходу реагента или комплекса реагент-полимераза в раствор (впоследствии реагент мог бы высвободиться из полимеразы).
Создание реагентов, соответствующих данному изобретению, входит в компетенцию специалистов в данной области. Следует иметь в виду, что температуру плавления ствола можно подобрать в существенной мере путем варьирования содержания в нем G-C. Например, стволы двух нижеописанных предпочтительных вариантов осуществления в обоих случаях имеют длину девять нуклеотидов, но они имеют рассчитанные Тт, которые отличаются на 25°C (81°C и 56°C). Обычно используют компьютерную программу M-fold, приведенную выше для расчета Тт ствола в сочетании с одной или более возможными последовательностями петли. Обнаружено, что последовательность петли в других случаях несущественна в реагентах против ошибочного старта, соответствующих данному изобретению. При конструировании авторы используют последовательность, которая формирует идеальный ствола с тупыми концами, т.е. ствол без внутренних ошибочных спариваний и без концевого свивающего конца. Затем можно путем простого испытания оценить эффект интродукции внутреннего ошибочного спаривания (концевое ошибочное спаривание является дестабилизирующим и неприемлемо, как показано в Примере 1) или короткого удлинения в виде одного, максимум двух нуклеотидов после двунитевого участка.
Конструирование реагентов, соответствующих данному изобретению, находится в компетенции специалистов в области техники. Например, олигонуклеотидные последовательности можно получить в синтезаторе олигонуклеотидов. Стабилизирующие группы можно включить при использовании известных методов. Например, Дабцил легко можно ввести, начиная с дабцилированной колонки (фирмы Glen Research) и завершая синтез Дабцил-модифицированным нуклеотидом. Неприродные нуклеотиды можно использовать в качестве первого и предпоследнего, а также последнего нуклеотидов в синтезе.
После разработки и конструирования потенциального реагента температуру плавления его ствола можно в ряде случаев приблизительно определить эмпирически путем введения флуоресцентного ДНК-связывающего красителя и проведения анализа плавления при стимуляции красителя (учитывая, что сам краситель воздействует на Тт). Крометого, практически полезную информацию, касающуюся действительной температуры плавления стабилизированного ствола реагента, соответствующего данному изобретению, можно во многих случаях вывести посредством проведения ПЦР-амплификаций в режиме реального времени, используя различные температуры отжига. При необходимости можно осуществить регуляцию длины ствола или содержания G-C с целью достижения требующейся Тт. Затем оценивают эффекты потенциального реагента на различные проявления ошибочного старта и эффективность амплификации посредством определения его действия в жесткой ПЦР-амплификации, такой как описана ниже в Примере 1. Потенциальный реагент считают реагентом, соответствующим данному изобретению, если при включении в реакционную смесь, представленную в Примере 1, в какой-либо концентрации ниже 1000 нМ, предпочтительно не больше, чем 650 нМ, получают чистый ампликон (См. Фиг.1). Анализы потенциальных реагентов оценивают, используя анализ, представленный в Примере 1, за исключением предусмотренной мишени и предусмотренных праймеров. Однако амплификации и анализы (амплификация в сочетании с детекцией), соответствующие изобретению, не ограничены условиями или способами, представленными в Примере 1. Хотя реагент, соответствующий данному изобретению, должен ингибировать по меньшей мере одно проявление ошибочного старта при введении в концентрации ниже 1000 нМ относительно концентрации полимеразы (см. Пример 1) 1,25 единиц/25 мкл реакционной смеси, реагент можно использовать в любой концентрации относительно концентрации полимеразы, при которой он эффективен, т.е. в любой концентрации (или относительной концентрации), при которой он предотвращает ошибочный старт, но в существенной мере не препятствует амплификации. Использование в оценке предусмотренных праймеров будет обнаруживать непредусмотренные последствия, такие как неспособность блокировать 3'-конец реагента или невыявленная комплементарность.
Данное изобретение включает реакции ПЦР-амплификаций и анализы, которые включают реакции ПЦР-амплификаций, в том числе реакции, в которых реакционная смесь для амплификации включает термостабильную ДНК-полимеразу, обладающую как полимеризационной активностью, так и 5'-3'-экзонуклеазной активностью, такую как ДНК-полимераза Taq, и в которых по меньшей мере один реагент, соответствующий данному изобретению, включен в реакционную смесь для амплификации. Многие реагенты, соответствующие данному изобретению, ингибируют активность полимеразы, и их добавляют в концентрации относительно концентрации полимеразы в вышеописанных интервалах перед или во время термического цикпинга, т.е. не больше, чем 1000 нМ, предпочтительно не больше, чем 650 нМ для реакции, содержащей 1,25 единиц ДНК-полимеразы/25 мкл объема реакционной смеси. Некоторые реагенты, соответствующие данному изобретению, хотя эффективны в концентрациях ниже 1000 нМ и, предпочтительно, в концентрациях ниже 650 нМ, ингибируют активность полимеразы в меньшей степени и могут быть добавлены в концентрациях до 1500 нМ или даже 3000 нМ для данной концентрации полимеразы. В анализах, включающих более одного реагента, соответствующего изобретению, каждый реагент может иметь ствол со своей собственной Тт и может быть добавлен в своей собственной концентрации, так что различные реагенты действуют в различныхфрагментахстадий процесса амплификации. Если реагент, соответствующий данному изобретению, добавляют во время термического циклинга, его следует добавлять без аспирации реакционной смеси в рабочую область. Данные реакции амплификации включают симметричные ПЦР-амплификации, асимметричные ПЦР-амплификации и ЛПЭ-ПЦР-амплификации, любая из которых может далее включать обратную транскрипцию, если участвуют РНК-мишени. ПЦР-амплификации можно использовать для получения амплифицированного продукта для любой цели, например, как исходного материала для расшифровки последовательности ДНК. ПЦР-амплификации можно комбинировать с детекцией амплифицированного продукта в анализе, включая в высокой степени гомогенные анализы с использованием меченых праймеров, меченых зондов или флуоресцентных ДНК-связывающих красителей, таких как SYBR Green или бромид этидия. Анализ может представлять собой анализ в режиме реального времени, в котором считывание показаний для детекции проводят во время множества циклов амплификации, или анализ конечных результатов, в котором детекцию проводят после завершения амплификации. Он может быть качественным или количественным, включая, но без ограничения перечисленным, количественный анализ конечных результатов. Может быть разработан анализ для амплификации одного двунитевого или однонитевого продукта либо более, чем одного двунитевого продукта без или вместе с родственными однонитевыми продуктами.
Как используют в данной заявке, "ЛПЭ-ПЦР" означает асимметричную амплификацию ДНК с помощью процесса полимеразной цепной реакции (ПЦР) при использовании одного олигонуклеотидного праймера ("Избыточный праймер") в по меньшей мере пятикратном избытке относительно другого праймера ("Ограничивающий праймер"), который используют в низкой концентрации, до 200 нМ, так чтобы он израсходовался в циклах ПЦР, примерно достаточных для получения флуоресцентно определяемого двунитевого ампликона, причем подобранная для концентрации температура плавления ограничивающего праймера в начале амплификации, Тm[0], выше, чем или не больше, чем на 5°C ниже подобранной для концентрации температуры плавления избыточного праймера в начале амплификации, Тm[0] X, предпочтительно на 3-10°C выше, и при этом термический циклинг продолжают в течение многочисленных циклов после израсходования ограничивающего праймера, чтобы получить однонитевый продукт, а именно, продукт удлинения избыточного праймера. Анализы ЛПЭ-ПЦР могут включать стадию низкотемпературной детекции, в которой температуру снижают ниже температуры отжига праймера на протяжении по меньшей мере нескольких циклов линейной амплификации. Предпочтительно, когда данная стадия имеет место после удлинения перед плавлением нити.
Данное изобретение включает также полные наборы для ПЦР, неполные наборы и группы олигонуклеотидов. Полный набор для ПЦР-амплификации включает по меньшей мере все реагенты для осуществления ПЦР-амплификации или анализа, в том числе по меньшей мере одну пару ГЩР-праймеров, дНТФ, реакционный буфер, термостабильную ДНК-полимеразу, предпочтительно полимеразу, обладающую 5'-3'-экзонуклеазной активностью, и по меньшей мере один реагент, соответствующий изобретению. Полный набор для гомогенного ПЦР-анализа далее включает любые дополнительные реагенты для детекции, которые необходимы, например, флуоресцентный краситель ДНК или зонды с флуоресцентной меткой. Полный набор любого типа предпочтительно включает реагенты для получения образца и может включать в некоторых вариантах осуществления обратную транскриптазу. Неполные наборы, соответствующие изобретению, не включают по меньшей мере некоторые ингредиенты полного набора, но включают по меньшей мере термостабильную ДНК-полимеразу (и, если требуется, обратную транскриптазу) и по меньшей мере один реагент, соответствующий изобретению. Например, продукты, коммерчески известные как "главная смесь" или "основной набор", как правило, не включают ПЦР-праймеры и зонды. Предпочтительный неполный набор включает все реагенты, необходимые для амплификации или анализа за исключением получения образца. Олигонуклеотидные группы, соответствующие изобретению, включают по меньшей мере одну пару ПЦР-праймеров и по меньшей мере один реагент, соответствующий изобретению. Они могут далее включать олигонуклеотидные зонды или праймеры для секвенирования.
Детали одного или более вариантов осуществления изобретения приведены на сопровождающих фигурах и в представленном ниже описании. Другие признаки, объекты и преимущества изобретения будут очевидны из описания и фигуры, а также из формулы изобретения.
Краткое описание чертежей
На Фиг.1 показан анализ при помощи гель-электрофореза образцов, полученных при качественном точном ПЦР-анализе ошибок, связанных с ошибочным стартом, разработанный для тестирования ингибирующей активности реагентов, описанных в данном изобретении.
На Фиг.2 показано, что реагенты, соответствующие данному изобретению, делают возможным проведение основанных на ПЦР анализов в режиме реального времени и конечных результатов.
На Фиг.3 показана хроматограмма последовательности ДНК, полученной из однонитевых продуктов ДНК, генерированных после 70 циклов ЛПЭ-ПЦР-амплификации при отсутствии ошибок, связанных с ошибочным стартом, с использованием реагентов, описанных в данном изобретении.
На Фиг.4, 5, 6, 7 показано влияние соединения 9-3 DD на пятикратную ЛПЭ-ПЦР-амплификацию.
На Фиг.8 и 9 приведены данные о том, что после того как реагенты, соответствующие изобретению, связывают полимеразу Taq, они не высвобождаются из полимеразы до тех пор, пока температуры циклинга ПЦР не достигнут стадии денатурации.
На Фиг.10, 11, 12 показано прямое сравнение реагентов, соответствующих данному изобретению, по их эффекту на активность ДНК-полимеразы при 25°C.
На Фиг.13 проиллюстрирован анализ, направленный на оценку влияния реагентов, соответствующих данному изобретению, на экзонуклеазную активность полимеразы в отсутствие полимеразной активности.
На Фиг.14 и 15 проиллюстрирована способность реагента, соответствующего данному изобретению, препятствовать ошибочному старту в симметричном ПЦР-анализе.
На Фиг.16 проиллюстрирован анализ, направленный на количественную оценку ингибирования экзонуклеазы.
На Фиг.17 показан дозозависимый эффект реагента, соответствующего данному изобретению, на формирование димеров праймера и олигомеров праймера в ЛПЭ-ПЦР-амплификации.
На Фиг.18 показано влияние низкой концентрации реагента 9-22 DD на двойные ЛПЭ-ПЦР-амплификации двух целевых последовательностей сдвумя парами праймеров.
На Фиг.19 показано влияние варьирования композиции ствола реагента 9-3 DD на двойную ЛПЭ-ПЦР-амплификацию двух целевых последовательностей с двумя парами праймеров.
На Фиг.20 показано влияние варьирования концентрации ствола реагента 9-3b DD на двойную ЛПЭ-ПЦР-амплификацию двух целевых последовательностей с двумя парами праймеров.
На Фиг.21 показано влияние повышения концентраций реагента 9-3 DD на эффективность ЛПЭ-ПЦР амплификации в показателях знамений СТ.
На Фиг.22 показано влияние повышения концентраций реагента 9-3 DD на эффективность ЛПЭ-ПЦР амплификации в показателях наклона кривой флуоресцентного сигнала и конечной флуоресценции.
На Фиг.23 представлены продукты многофакторной реакции, проводимой с использованием или без использования смеси соединений, соответствующих изобретению.
На Фиг.24 показаны результаты теста на действие соединения 12-3DD.
На Фиг.25 показаны результаты теста на действие соединения 12-C3DD.
Одинаковые ссылочные обозначения на различных фигурах указывают на одинаковые элементы.
Осуществление изобретения
Как указано выше, выявлен эффект, который, как полагают, бывает трех различных типов и вызывает ошибки, связанные с ошибочным стартом. Для оценки ошибочного старта всех трех типов разработан точный анализ, приведенный ниже в Примере 1. Амплификация начинается с расщепленной ДНК, которая способствует ошибочному старту. Физические манипуляции, например, пипетирование ДНК, способствуют расщеплению ДНК. Кроме того, поскольку только ошибочный старт Типа 1 можно предотвратить с помощью реагентов и методов "горячего старта", в анализ включают много температурных циклов (по меньшей мере 60). В дополнение включают стадию низкотемпературной детекции, используемую в ряде ЛПЭ-ПЦР - анализов. Кроме того, формирование димеров праймеров и их дальнейшую олигомеризацию и амплификацию считают также дополнительным типом ошибочного старта. Формирование и амплификация димера праймера может происходить как в присутствии, так и в отсутствие добавленных ДНК-матриц, и авторы разработали также анализы данных феноменов.
Поскольку ферменты горячего старта эффективны только перед началом амплификации вследствие необратимых изменений, которые происходят при нагревании до высокой температуры, например, 95°C, реагенты против ошибочного старта, соответствующие данному изобретению, не денатурируются необратимым образом и могли бы оказывать положительные или отрицательные эффекты либо оба из них позднее в реакции, если в нее включена низкотемпературная стадия.
Можно создать стволы, имеющие температуры плавления, Тт, выше или ниже температур определенных циклов с различными воздействиями на амплификацию. При создании стволов предпочтительно учитывать, что плавление представляет собой динамический феномен, происходящий в температурном интервале, причем Тт определяет температуру, при которой пятьдесят процентов молекул находится в двунитевой форме и пятьдесят процентов находится в однонитевой форме. Считают, что связывание некоторых закрытых стволов с полимеразой сдвигает равновесие среди молекул, которые остаются несвязанными с полимеразой в сторону увеличения молекул с закрытым стволом. Если требуется, чтобы подавляющее большинство стволов были закрытыми при температуре определенного цикла, такой как температура отжига праймера, начинают, как правило, с немодифицированного олигонуклеотидного ствола, имеющего рассчитанную Тт по меньшей мере на 5°С выше данной температуры, учитывая, что стабилизирующая модификация ствола повысит действительную Тт на несколько градусов. Напротив, если требуется, чтобы подавляющее большинство стволов было открытыми при температуре определенного цикла, начинают, как правило, с немодифицированного олигонуклеотидного ствола, имеющего рассчитанную Тт по меньшей мере на 5°С ниже, предпочтительно по меньшей мере приблизительно на 10°С ниже, чем температура определенного цикла, снова учитывая, что стабилизирующая модификация ствола, вероятно, повысит действительную Тт на несколько градусов.
Использование определенного реагента, соответствующего данному изобретению, с различными температурами отжига праймера может давать различные эффекты в отношении эффективности амплификации, как демонстрируют ниже в Примере 5. Один из представленных в настоящее время рассчитанных вариантов осуществления (соединение 9-22 DD), обсуждаемых ниже, имеет рассчитанную Тт 81°С. Она будет близкой во время каждого типичного термического цикла ПЦР, поскольку температура снижается от температуры плавления нити (например, 95°С) до температуры отжига праймера (например, 55°С). Реагент с данным стволом будет оказывать воздействие во время каждого цикла амплификации. Другой из представленных в настоящее время предпочтительных вариантов осуществления (соединение 9-3 DD) имеет рассчитанную Тт 56°С. Когда смеси для ПЦР-амплификаций готовят при комнатной температуре, данный реагент имеет закрытый ствол и связывается с полимеразой. Он остается связанным по мере повышения температуры от начального момента амплификации. Связанные с полимеразой молекулы 9-3 DD становятся несвязанными и открытыми во время исходной высокотемпературной стадии и не связываются с полимеразой снова до тех пор, пока температура не снижается достаточно для того, чтобы ствол несвязанных молекул закрылся. Таким образом, при поддержании самой низкой температуры цикла 65°С ствол не реорганизуется. В данных условиях этот вариант осуществления будет действовать в данной амплификации как реагент горячего старта для предупреждения ошибочного старта, который происходит перед началом амплификации. Однако, если включена стадия n низкотемпературной детекции, например, инкубирование при 40°C после удлинения в цикле ЛПЭ-ПЦР с целью обеспечения возможности гибридизации низкотемпературного молекулярного сигнала или другого меченого зонда с одной нитью, накапливающейся во время линейной фазы амплификация, ствол, имеющий рассчитанную температуру плавления 56°C будет закрыт, давая возможность регенту снова связываться с полимеразой до тех пор, пока не будет достигнута высокая температура плавления ствола.
В Примере 1 ниже описан точный анализ ЛПЭ-ПЦР, который используют для оценки действия реагентов, соответствующих данному изобретению. На Фиг.1 показывают, что ошибки, связанные с ошибочным стартом, имеются не только при использовании "нормальных" ДНК-полимераз Taq, т.е. термостабильных ДНК-полимеразс 5'-3'-экзонуклеазной активностью, но без модификации горячего старта (дорожки b и дорожки c), но также при использовании полимераз Taq горячего старта (дорожки d и дорожки e). Кроме того, на Фиг.1 показывают удаление неспецифических продуктов, образованных вследствие ошибок, связанных с ошибочным стартом, и присутствие только желательного специфического продукта, полученного путем добавления реагента 9-22 DD в концентрации 300 нМ не только при амплификации с полимеразами горячего старта (дорожки h и дорожки i), но также при амплификации с нормальными ДНК-полимеразами Taq (дорожки f и дорожки д). Эффективность соединений 9-22 DD и 9-3 DD в супрессии ошибочного старта оценивают с использованием анализа, описанного в Примере 1. Обнаружено, что оба соединения подавляют ошибочный старт Типа 1. Низкая рассчитанная Тт ствола 9-3 DD (56°C) позволяет использовать его просто в качестве эффективного реагента горячего старта при использовании протокола ПЦР-амплификации с температурами всех циклов 60°C и выше. Находясь при температуре выше температуры плавления ствола, данный ствол не реорганизуется по мере прохождения амплификации. Соединение 9-3 DD не обладает ингибирующим действием на эффективность полимеризации при использовании такого рода. Нижеприведенный Пример 5 является наглядным в этом отношении. В нем сравнивают амплификации с использованием реагента 9-3 DD и температур отжига 65°C и 55°C. При использовании 65°C не обнаруживают никакого эффекта на полимеризацию, что отражено в значении Ст. Однако при использовании 55°С происходит задержка Ст. Это свидетельствует о том, что, если температуру снизить достаточно для закрытия ствола и вовлечения в процесс полимеразы, ее включение продолжается при повышении температуры для удлинения праймера, даже если температура удлинения праймера превышает Тт ствола. С другой стороны, высокая Тт ствола 9-22 DD (81°С) делает возможной реорганизацию ствола в шпильку на стадии отжига праймера каждого цикла ПЦР. Это препятствует проявлению не только ошибочного старта Типа 1, но также ошибочного старта Типа 2, ошибочного старта Типа 3 и образованию димера праймера. Тем не менее это действительно определенным образом воздействует на эффективность полимеризации, что доказано путем задержки 1,5 циклов в пороговом цикле (Ст) ПЦР-амплификации, идущей в режиме реального времени, с помощью SYBR Green 1 или мишень-специфического молекулярного сигнального зонда при использовании в концентрации 100 нМ.
В Таблице I Примера 1 представляют применение точного анализа серии дополнительных компонентов и, кроме того, приводят минимальную концентрацию, необходимую для того, чтобы достигнуть предотвращения проявлений ошибочного старта, т.е. избежания определяемых уровней неспецифических продуктов, образованных вследствие ошибок, связанных с ошибочным стартом, и присутствия только требующегося специфического продукта амплификации, такого как показан на дорожках f и g Фиг.1, при использовании нормальной ДНК-полимеразы Taq, а не варианта для горячего старта. В Таблице I соединения идентифицируют согласно номенклатурной системе авторов. Немодифицированные шпилечные молекулы идентифицируют по длине ствола-петли. Например, соединение 6-22 имеет ствол длиной шесть нуклеотидов и петлю длиной двадцать два нуклеотида. Когда идентификатор петли начинается с числа или представляет собой только число ("22" или "3b"), петля представляет собой нуклеотиды. Ненуклеотидные петли определяют далее. Соединение 12-СЗСЗ имеет ствол длиной двенадцать нуклеотидов и петлю, которая представляет собой цепь из шести атомов углерода, т.е. цепь из шести метиленовых (CH2) групп. Делают различные модификации на одном или обоих концах базовых шпилек. Их обозначают индексами, где 3D представляет собой Дабцил (5'-диметокситритилокси-5-[(М-4'-карбокси-4-(диметиламино)-азобензол)-аминогексил-3-акриламидо]-2'-дезоксиуридин-3'-[(2-цианоэтил)-(М, 1\1-ди изопропил)]-фосфорамидит)> добавленный к З'-концевому нуклеотиду, 5D представляет собой Дабцил, добавленный к 5'-концевому нуклеотиду, DD представляет собой Дабцил, добавленный к обоим концевым нуклеотидам, BHQBHQ означает гаситель Black Hole, добавленный к обоим концевым нуклеотидам, FF представляет собой флуорофор (в данном случае FAM), добавленный к обоим концевым нуклеотидам, AA представляет собой аденозин (А), добавленный к обоим концевым нуклеотидам, и ТТ представляет собой тимидин (Т), добавленный к обоим концевым нуклеотидам.
Кроме добавлений, на базовых стволах делают ряд модификаций. Индекс 2'ОМ4 показывает, что четыре концевых нуклеотида, два последних на каждом олигонуклеотиде или плече ствола, изменены с дезоксирибонуклеотидов на 2'-0-метилнуклеотиды. Для изменения температуры плавления стволов изменяют содержание G-C. Соединения, в которых изменена последовательность ствола, но не длина ствола, обозначают указателем в виде строчной буквы, предшествующей индексу концевого модификатора, например, 9-3b DD или 9-3i DD. Нуклеотидные последовательности, имеющие также петли из углеродных цепей, используемые для различных соединений в Таблице I, представлены в Таблице II. Таблица I является информативной в нескольких отношениях. В ней показано, например, что в данном анализе немодифицированные олигонуклеотиды шпильки ДНК либо неэффективны, либо эффективны только в высокой концентрации, 1000 нМ или выше. Олигомер 9-22 (ствол длиной 9 нуклеотидов, петля длиной 22 нуклеотида) эффективен только в концентрации 3000 нМ, в количестве, описанном для супрессии ошибочного старта с помощью двунитевой ДНК. Олигомеры 6-22 (ствол длиной 6 нуклеотидов, петля длиной 22 нуклеотида) и 9-5 (ствол длиной 9 нуклеотидов, петля длиной 5 нуклеотидов) неэффективны даже в концентрации 3000 нМ. Однако, когда олигомер 9-22 модифицируют введением групп Дабцила в оба конца ствола с целью создания соединения 9-22 DD, предпочтительный реагент, соответствующий изобретению, требуется в концентрации только 50 нМ для предотвращения проявлений ошибочного старта. При аналогичной модификации олигомера 9-3 требуется концентрация только 100 нМ.
В Таблице I показан эффект длины ствола. Хотя некоторые олигомеры, имеющие стволы из девяти и двенадцати нуклеотидов устраняют ошибочный старт в низкой концентрации при модификации парой взаимодействующих гасителей Дабцил, олигомер 6-22 не реагирует таким образом на модификацию: композиция 6-22 DD, не являющаяся реагентом, соответствующим изобретению, которая эффективна только при высокой (3000 нМ) концентрации. Реагенты, соответствующие данному изобретению, в большинстве случаев имеют стволы длиной более шести нуклеотидов и короче четырнадцати пар оснований. Длины короче четырнадцати пар оснований в настоящее время предпочтительны.
В Таблице I также показывают эффективность других модификаций, которые повышают силу связывания на конце ствола. Введение пары интерактивных гасителей Black Hole™ в ствола олигомера 9-5 дает реагент 9-5 BHQBHQ, который требуется в концентрации только 100 нМ для исключения неспецифических продуктов, образованных вследствие ошибок, связанных с ошибочным стартом, и присутствия только желательного специфического продукта амплификации. Замена двух 2'-0-метилрибонуклеотидов на каждом конце олигомера 9-5 дает реагент 9-5 2'ОМ4, для которого требуется концентрация только 50 нМ.
В Таблице I показывают, что длина олигонуклеотидной петли может значительно варьировать. Все длины петель, составляющие три нуклеотида (реагент 9-3 DD), пять нуклеотидов (реагент 9-5 DD) и двадцать два нуклеотида (реагент 9-22 DD), приводят в результате к образованию реагентов, соответствующих данному изобретению, которые препятствуют ошибочному старту в концентрации 100 нМ или меньше. В то же самое время в Таблице I показывают, что для оптимизации требуются некоторые исследования и коррекция ошибок. Если пару 2'-O-метилрибонуклеотидов заменяют гасителями Дабцил в стабилизированной модификации олигомера 9-5 (сравните реагент 9-5 2'ОМ4 с реагентом 9-5 DD), уровень эффективности не изменяется: требуется концентрация только 50 нМ. Однако при осуществлении такого же изменения у модифицированного олигомера 9-3 (сравните реагент 9-3 2'ОМ4 с реагентом 9-3 DD) уровень эффективности снижается, хотя и остается достаточно высоким: требуется концентрация 300 нМ, а не концентрация 50 нМ. Эти же данные показывают, что увеличение длины петли реагента 9-3 2'ОМ4 на два нуклеотида для создания реагента 9-5 2'ОМ4 повышает эффективность: требуется концентрация только 50 нМ, а не концентрация 300 нМ. Потенциальные реагенты можно рутинно оценить и скорректировать с помощью точного анализа, такого как описан в Примере 1.
В Таблице I также показывают, что соединения, состоящие из двух комплементарных олигонуклеотидов, удерживаемые вместе при противоположной полярности посредством ненуклеотидного мостика (петли), также активны, когда свободные 3'- и 5'-концы олигонуклеотидов модифицированы согласно изобретению (в тестированных вариантах осуществления с помощью групп Дабцила). В Таблице 1 показывают, что размер ненуклеотидного мостика неважен. Мостики, описанные в Таблице I, представляют собой химические линкеры, состоящие из линейных цепей из 3-6 атомов углерода, но компетентные специалисты в данной области могли бы сконструировать много других вариантов ненуклеотидных мостиков (петель). Данные, приведенные в Таблице I, позволяют предположить, что мостик из цепи из 3 атомов углерода лучше, чем мостик из цепи из 6 атомов углерода для одного и того же олигонуклеотидного ствола (активен при более низкой концентрации) и, хотя точный анализ не проведен, предполагают, что данное различие отражает относительную гибкость мостиков и их последующий эффект или способность повышать или снижать температуру плавления ствола. Авторы используют химически нейтральные линкеры (незаряженные), которые предпочтительны.
Различные применения реагентов и способов, соответствующих изобретению, проиллюстрированы ниже в Примерах 2-4. Пример 2 описывает применение реагента 9-22 DD в ЛПЭ-ПЦР-анализе, который представляет собой анализ в режиме реального времени, но, как показано, пригоден также для анализа конечных результатов. В анализе, приведенном в Примере 2, используют способ детекции, который является объектом одновременно поданной теми же авторами предварительной патентной заявки Соединенных Штатов Америки # 60/619654 под названием Праймеры, зонды и способы амплификации нуклеиновой кислоты, которая включена в данном контексте в виде ссылки во всей полноте. Данный способ детекции включает добавление как флуоресцентного красителя ДНК, такого как SYBR Gold, который флуоресцирует, когда связан с двунитевой ДНК, так и меченного флуорофором гибридизационного зонда, комплементарного однонитевому ампликону, образованному в ЛПЭ-ПЦР-амплификации после израсходования ограничивающего праймера, причем флурофор стимулируется эмиссией из красителя, стимуляцию красителя, детекцию эмиссий как из красителя, так и флуорофора и вычисление соотношения сигнала флуорофора и сигнала красителя. Пример 2, таким образом, показывает совместимость реагентов, соответствующих данному изобретению, с флуоресцентными зондами и флуоресцентными красителями, и эффективное предотвращение ошибочного старта при использовании одного из данных компонентов или обоих.
В Примере 4 описывают ряд многофакторных ЛПЭ-ПЦР-амплификаций с использованием двух, трех, четырех и даже пяти пар праймеров и показывают амплификацию корректных ампликонов во всех случаях, когда реагент 9-3 DD вводят в реакционную смесь. Хотя амплифицированные продукты анализируют гель-электрофорезом для целей Примера 4, многофакторный способ также применим в многофакторных анализах, в том числе качественных и количественных анализах, таких как анализы в режиме реального времени с использованием других средств детекции, таких как флуоресцентные гибридизационные зонды для соответствующих ампликонов. Для ЛПЭ-ПЦР-анализов в режиме реального времени предпочитают использовать стадию низкотемпературной детекции после стадии удлинения праймера и низкотемпературные зонды, как проиллюстрировано, например, в Примере 3.
Пример 13 иллюстрирует, что можно также использовать реакции амплификациисамплификационными реакционными смесями, содержащими больше одного варианта реагентов, соответствующих данному изобретению. Например, амплификационная смесь может включать оба, соединение, соответствующее данному изобретению с высокой температурой плавления и другое соединение, соответствующее данному изобретению, с низкой температурой плавления, каждое из которых добавляют в своей собственной оптимизированной концентрации. Ствол соединения 9-22DD с рассчитанной Тт 81°C находится в закрытой конформации в начале реакции и вскоре после снижения температуры во время каждого термического цикла, тогда как соединение 12-3DD с рассчитанной Тт 58°C находится в закрытом состоянии только при относительно низких температурах. Вследствие этого оно действует как "горячий старт" для предотвращения ошибочного старта, когда реакционную смесь готовят при комнатной температуре, но во время последующихтермическихциклов ПЦР-амплификаций, имеющихтемпературу отжига 60°C или выше, не закрывается и связывается с полимеразой. Смеси реагентов, в частности, используют для конструирования многофакторных реакций, в которых множество пар праймеров, охватывающих интервал температур отжига, комбинируют для амплификации множества ампликонов. Смесь реагентов, используемых в Примере 13, лучше, чем каждый реагент в отдельности.
Пример 3 описывает ЛПЭ-ПЦР-амплификацию с использованием реагента 9-3 DD для получения амплифицированного однонитевого продукта в достаточном количестве и в достаточной степени свободного от неспецифических продуктов, возникающих при ошибочном старте, и подходящего для применения в качестве исходного материала для секвенирования. Амплификацию продолжают до семидесяти циклов, чтобы обеспечить достаточно исходного материала. Обнаруживают, что некоторые ампликоны более подвержены ошибкам, связанным с ошибочным стартом Типа 2 и Типа, чем другие ампликоны. При амплификациях, в которых данный ошибочный старт более вероятен, предпочтительно, чтобы ствол реагента, соответствующего данному изобретению, был закрыт, когда температура цикпинга падает от стадии плавления нити до стадии отжига праймера. Это можно обеспечить подбором Тт ствола, температуры отжига или обоих факторов. В близкой работе обнаруживают, что разведение продукта представляет собой простой способ очистки, предназначенный для применения при секвенировании, как описано в вышеуказанной одновременно поданной предварительной патентной заявке Соединенных Штатов Америки.
Исследовано действие и эффективность вариантов осуществления реагентов, соответствующих данному изобретению, по сравнению с немодифицированными олигомерами ДНК, образующими шпильку, и друг с другом. Разработан анализ полимеризации ДНК для установления удлинения меченого праймера ДНК-полимеразой Taq в присутствии тест-реагента. В анализе используют праймеры, которые помечены флуорофорами, которые возбуждаются эмиссией из флуоресцентного красителя, как описано в одновременно поданной теми же авторами вышеуказанной предварительной патентной заявке Соединенных Штатов Америки. Смесь для анализа включает синтетический олигонуклеотид ДНК-матрицы в концентрации 0,5 мкМ, ДНК-праймер, комплементарный 3'-концу матрицы и меченый Су5 на 5'-конце в концентрации 1,5 мкМ, буфер для ПЦР, MgCl2, ДНК-полимеразу Taq, флуоресцентный краситель ДНК SYBR Green в разведении 1:40000 и тест-реагент. Контролируемую инициацию реакции осуществляют добавлением дНТФ. Реакция является изотермической: она протекает при заданной температуре в течение заданного периода времени. Для оценки эффекта тест-соединения на 5'-3'-экзонуклеазную активность ДНК-полимеразы к реакционной смеси добавляют
"блокатор", а именно олигонуклеотид, который комплементарен 5'-концу матрицы, блокированной по 3'-концу фосфатом, и который помечен ROX на 5'-конце. Для оценки эффекта тест-соединения на полимеризационную активность ДНК-полимеразы блокатор не используют. Для облегчения анализа продуктов реакции с помощью анализа кривой плавления разрабатывают матрицу, праймер и блокатор, чтобы последующие гибриды были легко отличимыми по своим температурам плавления: праймер-матрица, блокатор-матрица, продукт удлинения полной длины-матрица и неполный продукт удлинения (до блокатора) - матрица. Анализ в режиме реального времени осуществляют путем периодического возбуждения красителя и периодического мониторирования эмиссии флуоресценции из красителя и двух флуорофоров, оба из которых возбуждаются косвенным образом путем эмиссии из красителя. Повышение уровня флуоресценции SYBR Green I указывает на полимеризацию. Снижение флуоресценции ROX указывает на разложение блокатора. Анализ конечных результатов осуществляют остановкой реакции добавлением 12 мМ ЭДТА (этилендиаминтетрауксусной кислоты), доведением разведения SYBR Green I до 1:14200 и доведением концентрации тест-соединения до 800 нМ и последующим проведением анализа стандартной кривой плавления при стимуляции красителя и мониторировании флуоресценции каждого из двух флуорофоров.
Соединения 9-22, 9-22 DD и 9-3 DD оценивают при инкубированиях при 55°C в течение 60 минут. Включают также контроль, не содержащий никакого тест-соединения. Результаты, полученные в контроле, инкубируемом в присутствии блокатора, но без тест-соединения, показывают, что ДНК-полимераза Taq удлиняет праймер через участок блокатора, хотя в данных условиях не полностью, приводя к образованию продукта с практически одним пиком плавления согласно флуоресценции Су5. Таким образом фермент проявляет как полимеризационную активность, так и 5'-3'-экзонуклеазную активность. Результаты с соединением 9-22 DD, присутствующим в концентрации 300 нМ или в концентрации 1000 нМ, показывают образование продукта неполного удлинения и дозозависимый эффект на количество остаточного блокатора.
Таким образом соединение 9-22 DD ингибирует 5'-3'-экзонуклеазную активность полимеразы при 55°C. Имеется снижение образования продукта полной длины, особенно при концентрации 1000 нМ, позволяющее предположить, что высокая концентрация соединения 9-22 DD начинает ингибировать полимеризацию.
Другой анализ, который используют для исследования свойств реагентов, соответствующих данному изобретению, описан ниже в Примере 6. Данный анализ определяет способность тест-реагента ингибировать удлинение с помощью ДНК-полимеразы Taq при 25С. Результаты, приведенные в Примере 7, показывают, что, хотя немодифицированные шпилечные олигонуклеотиды 9-22 и 9-3 не обладают эффектом в отношении контроля, реагенты 9-22 DD и 9-3 DD ингибируют удлинение праймера дозозависимым образом. Так, даже при температуре, достаточно низкой, чтобы реагенты, соответствующие данному изобретению ингибировали полимеризационную активность в низкой концентрации, их немодифицированные аналоги не обладают данным свойством при таких же концентрациях.
Для исследования ингибирующегоэффекта реагентов, соответствующих данному изобретению, на 5'-3'-экзонуклеазную активность ДНК-полимеразы используют два анализа. Один анализ описан ниже в Примере 7. Анализ измеряет ингибирование б'-З'-экзонуклеазной активности во время термического циклинга. На Фиг.13 показано, что реагент 9-22 DD ингибирует данную экзонуклеазную активность дозозависимым образом при низких концентрациях, лежащих в интервале от 50 нМ до 300 нМ.
Второй анализ описан ниже в Примере 9. Он количественно измеряет ингибирование б'-З'-экзонуклеазной активности при 25°C. На Фиг.16 показывают дозозависимое ингибирование реагентом 9-22 DD в концентрации от 50 нМ до 1000 нМ.
Еще один анализ, описанный в Примере 6, измеряет ингибирование реагентом удлинения праймера, осуществляемого фрагментом Стоффеля ДНК-полимеразы Taq. Как в случае с интактной полимеразой Taq, ингибирование удлинения требует присутствия модифицированных концов на олигонуклеотидных стволах, и на него не оказывает существенного воздействия длина петли между стволом. Однако, в отличие от случая интактной полимеразы Taq, фрагмент Стоффеля ингибируется соединением 6-22DD, реагентом, имеющим ствол только из шести нуклеотидов. Заключают, что фрагмент Стоффеля не взаимодействует с реагентом тем же образом, что интактная полимераза.
Реагенты, соответствующие данному изобретению, используют для предотвращения ошибочного старта в симметричных ПЦР-анализах, как демонстрируют в Примере 8. Анализ измеряет амплификацию последовательности в гене кистозного фиброза при использовании эквимолярных пар праймеров, имеющих очень близкие значения Тт.
ПЦР-амплификаций и анализы, используемые в настоящее время, включают получение реакционных смесей для амплификации при комнатной температуре, таким образом требуется включение реагентов, соответствующихданному изобретению, в исходнуюсмесь для ингибирования ошибочного старта Типа 1. Однако, это необязательно. Некоторые автоматизированные методы, например, микрожидкостные методы, позволяют добавлять полимеразу в реакционную смесь при повышенной температуре для горячего старта, чтобы обойти таким образом ошибочный старт Типа 1. В данных способах реагенты, соответствующие данному изобретению, можно добавлять после начала амплификации.
Формирование димера праймера является проявлением ошибочного старта, который имеет место в завершенных реакциях, т.е. тех, которые включают все компоненты, требующиеся для амплификации, и исходные целевые последовательности, а также в реакциях, которые не включают целевые последовательности. Димеры ираймеров, как правило, представляют собой короткие двунитевые последовательности ДНК, сформированные в реакции путем ошибочного старта одного или более праймеров посредством гибридизации и удлинения либо другой копии того же однонитевого праймера, либо какого-либо другого праймера, присутствующего в реакции. Формирование димера праймера в присутствии целевой последовательности наблюдают как накопление димеров праймера в дополнение к накоплению ожидаемого ампликона. Формирование димера праймера в отсутствие целевой последовательности наблюдают как накопление димеров праймера без накопления специфического ампликона. Димеры праймера могут также формировать олигомеры с более длинной последовательностью, чем базовый димер праймера, поскольку они включают дополнительные связанные копии одного или обоих праймеров. Процесс олигомеризации до конца непонятен, но легко определяется с помощью гель-электрофореза или анализа температуры плавления продуктов реакции. Вследствие формирования димера праймера количество ожидаемого ампликона, образующегося при заданном числе термических циклов уменьшается, потому что один или оба праймера расходуются на генерацию и накопление димеров праймера. Вследствие этого элиминация димеров праймера необходима, поскольку она повышает как специфичность, так и выход корректного продукта. Данные свойства димеров праймера, а также их элиминация путем добавления реагентов, описанных в данном изобретении, иллюстрируют в Примере 10.
Вероятность формирования димера праймера повышается при гомологии последовательностей 3'-конца одного праймера и внутренних последовательностей этого же праймера или последовательностей другого праймера в дополнение ко многим другим факторам, например, повышению концентрации праймера, повышению концентрации магния, увеличению длины праймера, снижению температуры отжига термического цикла и увеличению общего числа праймеров, включенных в реакцию. Кроме того, в области техники хорошо известно, что, возможно, уровень формирования димера праймера значительно выше в реакциях с использованием полимераз негорячего старта по сравнению с реакциями с использованием полимераз горячего старта. Это обусловлено тем, что гибридизация типа праймер-праймер и удлинение могут происходить при относительно низких температурах, когда смешивают компоненты реакции. Ферменты горячего старта уменьшают, но полностью не устраняют генерацию и накопление димеров праймера. Данные свойства димеров праймера, а также их элиминация посредством добавления реагентов, описанных в данном изобретении, иллюстрируют в Примере 11.
Формирование димеров праймеров может также оказывать тонкое воздействие на кинетику реакции и их элиминацию посредством добавления реагентов, описанных в данном контексте, которые можно наблюдать и оптимизировать кинетическим путем. В Примере 12 описывают ЛПЭ-ПЦР-амплификацию, включающую две пары праймеров и генерирующую два однонитевых продукта. Кинетическое накопление одного из данных продуктов определяют по гибридизации с флуоресцентным зондом и представляют на Фиг.20 и Фиг.21. Данные демонстрируют, что на линейность кинетики воздействуют оба фактора, точная композиция ствола реагента вследствие его влияния на температуру плавления и концентрация реагента. Более того, оптимальное и субоптимальное применение реагентов может изменить скорость линейной амплификации в ЛПЭ-ПЦР и, таким образом, усилить сигнал в конце реакции без воздействия на значение Ст реакции. На Фиг.22 и Фиг.23 демонстрируют, что эффективность амплификации является такой же, как предполагают на основании значений Ст относительно интервала исходных концентраций матрицы, когда соединение 9-3DD используют в концентрации 150 и 300 нМ, но величина сигналов, идущих от исходных количеств матрицы 100 и 1000 в присутствии 300 нМ 9-3DD, выше, чем полученная при использовании 150 нМ 9-3DD, вероятно, вследствие отсутствия формирования димера праймера.
ПРИМЕРЫ
Пример 1. Точный анализ для оценки ошибок, связанных с ошибочным стартом
С целью оценки ошибочного старта Типа 1, Типа 2 и Типа 3 в реакциях ПЦР-амплификации проводят ЛПЭ-ПЦР-амплификацию с использованием расщепленной геномной ДНК и исследуют продукты с помощью анализа температуры плавления и гель-электрофореза. Конкретная амплификация, которую используют, состоит в следующем:
A. Субстрат: 10-10000 геномных эквивалентов расщепленной геномной ДНК. ДНК приобретают коммерческим путем и подвергают замораживанию и оттаиванию геномную ДНК или обрабатывают любым другим аналогичным методом, известным компетентным специалистам в области техники для расщепления ДНК.
B. Основа смеси для ПЦР-амплификации (см. статью Sanchez et al. (1994) PNAS 101: 1933-1938):
Субстрат: 2000 геномов расщепленной геномной ДНК
IX буфер для ПЦР
Mg+2: в концентрации 3 миллимоль (мМ)
дНТФ: концентрация 250 микромоль (мкМ) каждого из четырех дНТФ
Избыточный праймер: последовательность 5' CTTTGATGACGCTTCTGTATCTA 3'(SEQ ID NO: 13) в концентрации 1000 наномоль (нМ)
Ограничивающий праймер: последовательность 5'
CCTGGATTATGCCTGGCACCAT 3' (SEQ ID NO: 14) в концентрации 50 нМ
ДНК-полимераза: 1,25 единицы/25 микролитров (мкл) реакционной смеси.
C. Протокол амплификации:
Инкубирование при низкой температуре: 35 мин при комнатной температуре.
Протокол ПЦР-амплификации: выдерживание при высокой температуре 95°C в течение 15 минут; 10 циклов при 95°C в течение 10 сек, 55°C в течение 30 сек и 70°C в течение 30 сек с последующими 70 циклами 95°C в течение 10 сек, 50°C в течение 30 сек и 70°C в течение 30 сек.
Исходный уровень ошибочного старта устанавливают путем амплификации с использованием двух термостабильных ДНК-полимераз, Promega Taq и Invitrogen Taq без использования какого-либо метода горячего старта. Дополнительные амплификации проводят с использованием двух различных коммерческих ДНК-полимераз для горячего старта, Qiagen Hot Star Taq и Platinum Taq (Invitrogen). Наконец, дополнительные амплификации проводят с добавлением 300 нМ в настоящее время предпочтительного варианта осуществления реагента, соответствующего изобретению. Один вариант осуществления, используемый в данном примере, представляет собой реагент, который обозначают как композиция 9-22 DD (по номенклатуре авторов "9" означает длину ствола, "22" означает длину петли и "DD" обозначает стабилизирующую модификацию, в данном случае пару гасителей Дабцил). Соединение 9-22 DD имеет рассчитанную Тт 81°C. Оно имеет последовательность, представленную в Таблице II, которая модифицирована введением 5'-концевой и З'-концевой групп Дабцила.
Результаты гель-электрофореза различных амплификации с окрашиванием бромидом этидия показаны на Фиг.1, которая включает маркеры размера {с разницей 100 пар оснований) в немеченых крайних колонках. Делают повторности, поэтому представлены две дорожки для продукта каждой ЛПЭ-ПЦР-амплификации. Дорожки а - исходный уровень, в котором отсутствует ДНК; дорожки b - исходный уровень, в котором ДНК-полимераза является Promega Taq; дорожки с - исходный уровень, в котором ДНК-полимераза является Invitrogen Taq; дорожки d - замена ферментом горячего старта Qiagen Hot Star Taq; дорожки e - замена ферментом горячего старта Platinum Taq; дорожки 1 - амплификация с использованием полимеразы Promega Taq с добавлением соединения 9-22 DD; дорожки g - амплификация с использованием полимеразы Invitrogen Taq и соединения 9-22 DD; дорожки h - амплификация с использованием ДНК-полимеразы Qiagen Hot Star Taq и соединения 9-22 DD; дорожки I - амплификация с использованием полимеразы Platinum Taq и соединения 9-22DD.
На Фиг.1 можно видеть, что нормальные ДНК-полимеразы Taq, которые обладают как полимеризационной активностью, так и 5'-3'-экзонуклеазной активностью (дорожки b и дорожки с) дают ряд размеров продукта, среди которых почти отсутствует требующийся ампликон, определяемый парой праймеров, который представляет собой продукт, наблюдаемый на дорожках f-i. Переключение на модифицированные полимеразы "горячего старта" (дорожки d-e) помогает, но еще не элиминирует неспецифические продукты, сформированные вследствие ошибок, связанных с ошибочным стартом. Однако, когда соединение 9-22 DD присутствует в концентрации 300 нМ, требующийся ампликон получают без неспецифических продуктов, сформированных вследствие ошибок, связанных с ошибочным стартом, в обоих случаях, когда фермент представляет собой нормальную ДНК-полимеразу Taq (дорожки f-g) и когда фермент представляет собой ДНК-полимеразу Taq (дорожки h-i). Используя точный анализ данного примера, сравнивают число немодифицированных и модифицированных шпилечных молекул ДНК без концевой стабилизации, с концевой дестабилизацией, как получают при введении идентичных нуклеотидов в конец каждого плеча, с концевой стабилизацией, соответствующей изобретению, и с нестабилизирующими концевыми вставками. Чтобы сделать сравнение количественным, проводят тесты ответа на дозу с помощью анализа каждого соединения, которое проявляет активность супрессии ошибочного старта, с целью определения минимальной концентрации, необходимой для получения элиминации неспецифических продуктов, формирующихся вследствие ошибок, связанных с ошибочным стартом, и присутствия только требующегося специфического продукта, полученного, как показано на Фиг.1, с использованием нормальной ДНК-полимеразы Taq. Результаты представлены в Таблице I, которая включает расчеты Тт ствола для каждого тестируемого реагента. Последовательности различных соединений, представленных в Таблице I, приведены в Таблице II.

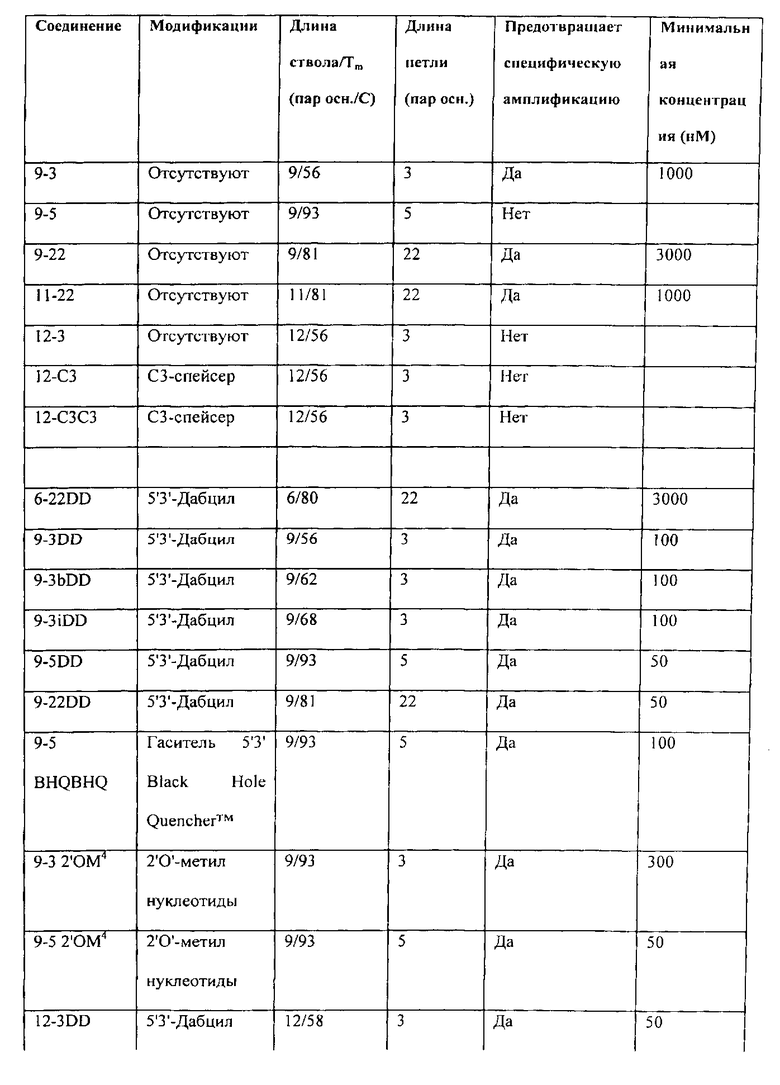
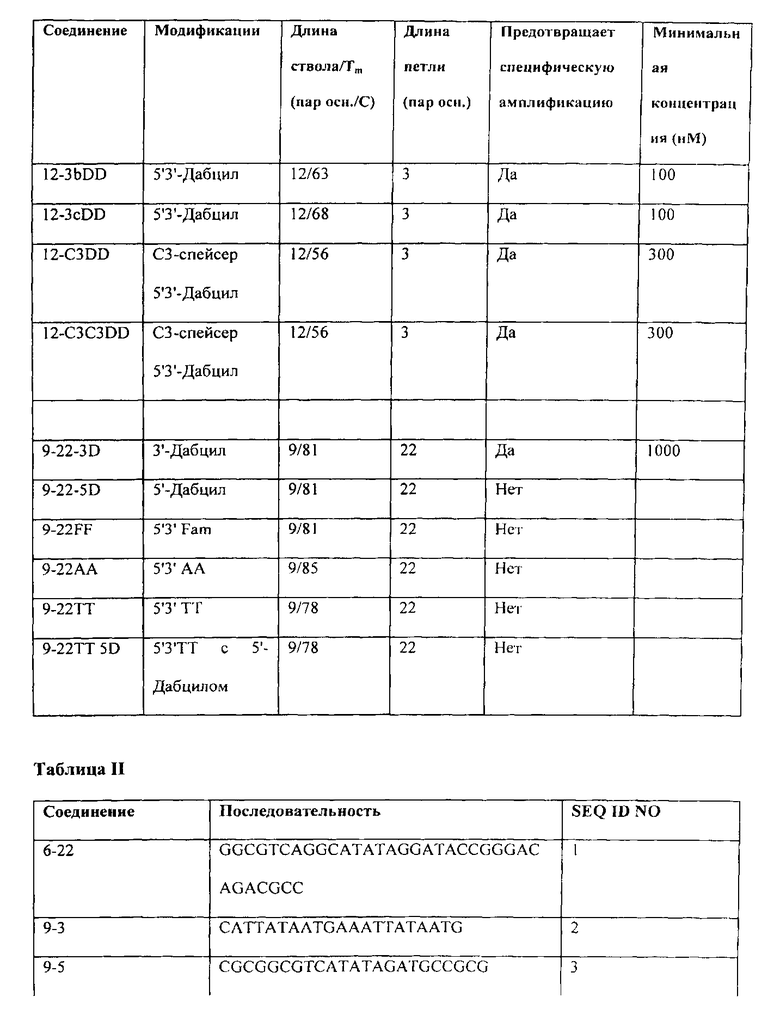

В Таблице I сравнивают ряд шпилек ДНК с модификациями и без модификаций и идентифицируют ряд стабилизированных на конце шпилечных молекул, которые являются реагентами, соответствующими данному изобретению (чистый ампликон без продуктов ошибочного старта в концентрации меньше, чем 1000 нМ) или которые являются предпочтительными реагентами, соответствующими данному изобретению (чистый ампликон без продуктов ошибочного старта в концентрации не более 650 нМ). Немодифицированные шпильки идентифицируют по двум числам, первое является длиной ствола и второе представляет собой длину петли. Соединения, соответствующие изобретению, показанные в Таблице I, имеют ряд стабилизирующих модификаций: 5' гаситель Дабцил и 3' гаситель Дабцил, ковалентно присоединенные к концевым нуклеотидам с помощью коммерческого линкера (обозначают окончанием "DD"), 3' гаситель Black Hole™ и 5' гаситель Black Hole™, присоединенный аналогичным образом (обозначаемый окончанием "BHQBHQ") и два 3'2'-О-метилрибонуклеотида и два 5'2'-О-метилрибонуклеотида, замещающие дезоксирибонуклеотиды на концах ствола (обозначают "2'ОМ4"). Дестабилизирующие модификации концов ствола включают замену пары А или Т на 3'- и 5'-концах молекулы. Введение пары флуорофоров FAM, которые, как полагают, не взаимодействуют друге другом стабилизирующим ствол образом, является дестабилизирующим, как и введение одного 5'-Дабцила. Однако введение одного 3'-Дабцила немного стабилизирует шпильку 9-22, как показано по уменьшению требующейся концентрации. Шпилька 9-22 супрессирует ошибочный старт только в высокой (3000 нМ) концентрации, но 9-22 DD делает это в низкой концентрации (50 нМ).
Другая из предпочтительных в настоящее время шпилек, соединение 9-3, супрессирует ошибочный старт только в высокой (1000 нМ) концентрации, но 9-3 DD и 9-3 2'ОМ4 делают это в гораздо более низкой концентрации (100-300 нМ). Еще один вариант осуществления, 9-5 DD, 9-5 BHQBHQ и 9-5 2'ОМ4 делает это также в низких концентрациях (50-100 нМ). Олигонукпеотид 9-5 имеет такой же ствол, как олигонукпеотид 9-22, но более короткую петлю, нуклеотиды которой представляют собой А и Т. Рассчитанная Тт ствола олигонукпеотида 9-5 составляет 93°C, примерно на 12°C выше, чем рассчитанная Тm ствола олигонуклеотида 9-22 вследствие влияния петель.
Аналогичные результаты получают с даже более длинным стволом. Соединение 12-3 не подавляет ошибочный старт, но соединение 12-3DD эффективно только в концентрации 50 нМ.
Таблица I показывает, что стабилизированные стволы можно модифицировать, чтобы изменить рассчитанную температуру плавления ствола (Тт). Например, соединение 9-3DD, Тт 56°C, эффективно в концентрации 100 нМ. Изменением содержания G-C в стволе создают соединения 9-3bDD (Tm 62°C) и 9-3iDD (Tm 68°C). Оба также являются эффективными в концентрациях 100 нМ. Соединение 12-3DD, Тт 58°C, эффективно в концентрации 50 нМ. Соединения с измененными стволами, а именно, соединение 12-3bDD, Tm 63°C, и соединение 12-3cDD, Tm 68°C, также весьма эффективны в концентрации 100 нМ.
Пример 2. ЛПЭ-ПЦР-анализ конечных результатов
Случайное изменение в кинетике реакции среди образцов копий приводит к разбросу сигналов от гибридизационных зондов и затрудняет анализ конечных результатов. ЛПЭ-ПЦР существенно снижает данную проблему. Применение реагента, соответствующего данному изобретению, кроме того, повышает возможность использовать детекцию конечных результатов. На Фиг.2 представляют результаты, полученные при амплификации ампликона, который включает аллель G269 гена гексозаминидазы А, ответственного за болезньТея-Сакса. Реакционная смесь включает 0,6 мкМ гибридизационного зонда и либо 1000 геномов гомозиготной ДНК-мишени дикого типа (+/+), либо гетерозиготную ДНК-мишень (G269/+). Образцы копий каждого типа амплифицируют следующим образом: 1 × буфер для ПЦР, 3 мМ MgCl2, 250 мкМ каждого дНТФ, 25 нМ ограничивающего праймера, имеющего последовательность 5' CGAGGTCATTGAATACGCACGGCTCC 3' (SEQ ID NO: 15), 1000 нМ избыточного праймера, имеющего последовательность 5' TAACAAGCAGAGTCCCTCTGGT 3' (SEQ ID NO: 16), 1,25 единицы полимеразы Taq Platinum (горячего старта), 0,6 мкМ меченного Су5 гибидизационного зонда, имеющего последовательность 5' Cy5-GGGACCAGGTAAGAA - фосфат 3' (SEQ ID NO: 17) и разведение 1:40000 SYBR Gold I в присутствии или в отсутствие 100 нМ варианта осуществления 9-22 DD в реакциях объемом 25 мкл. Используют следующие параметры цикла ПЦР: 95°C в течение 3 минут; 25 циклов при 95°C в течение 10 сек, 65°C в течение 20 сек и 72°C в течение 20 сек и 30 циклов при 95°C в течение 10 сек, 65°C в течение 20 сек, 72°C в течение 20 сек, 55°C в течение 20 сек и 45°C в течение 20 сек при обнаружении флуоресценции при 72°C для SYBR Gold и обнаружении флуоресценции при 55°C и 45°C для Су5. Для коррекции вариаций кинетики реакции в различных пробирках гибридизационные сигналы нормализуют по соотношению сигналов Су5 при 55°C и сигналов SYBR Gold при 72°C. Одно из последствий данной нормализации состоит в том, что маскируется любая задержка СТ.
На Фиг.2 линии, обозначенные кругом 21, и линии, обозначенные кругом 22, показывают результаты для множества копий без добавления реагента, соответствующего данному изобретению, но с использованием полимеразы горячего старта. Гомозиготные копии (круг 21) и гетерозиготные копии (круг 22) можно разделить по наклону их линейных графиков. Разброс среди копий и ошибки, связанные с ошибочным стартом, в данных образцах, подтверждаемые падением сигнала гибридизационного зонда ("эффект крюка") после приблизительно 40 циклов, препятствуют идентификации данных образцов по конечному результату. На Фиг.2 линии, идентифицированные кругами 23-24, показывают результаты с добавлением реагента, соответствующего изобретению, а именно, 100 нМ 9-22 DD. В данных условиях ошибки, связанные с ошибочным стартом, не происходят, линейная кинетика сохраняется в ходе анализа, разброс уменьшается и устраняется эффект крюка, и идентификация по конечному результату гомозиготных и гетерозиготных образцов возможна за 50 циклов.
Пример 3. Получение образцов ДНК для дидезоксисеквенирования
ЛПЭ-ПЦР представляет собой способ асимметричной амплификации, который потенциально генерирует большие количества однонитевой ДНК, пригодной для секвенирования ДНК, когда реакцию амплификации проводят за 60 и более термических циклов. Однако такие большие количества циклов амплификации способствуют появлению продуктов ошибочного старта, включающих, как полагают, формирование ошибок, связанных с ошибочным стартом Типа 2 и Типа 3, которые проявляются в виде накопления определяемых неспецифических продуктов, которые в конечном счете становятся преобладающими молекулами в реакции. Реагенты, соответствующие данному изобретению, обеспечивают возможность синтеза больших количеств однонитевой ДНК, подходящей для секвенирования ДНК в отсутствие неспецифических продуктов, при использовании 70 и более циклов ЛПЭ-ПЦР-амплификации. Образцы, подходящие для дидезоксисеквенирования посредством капиллярного гель-электрофореза получают следующим образом. Реакционную смесь для ЛПЭ-ПЦР сначала готовят в 25 мкл, содержащих IX буфер для ПЦР, 3 мМ MgCl2 1000 нМ избыточного праймера, имеющего последовательность 5' GCCAGGGGTTCCACTACGTAGA 3' (SEQ ID NO: 18), 25 нМ ограничивающего праймера, имеющего последовательность 5' CCGCCCTTCTCTCTGCCCCCTGGT 3' (SEQ ID NO: 19), 1,25 единицы полимеразы Taq Platinum, 600 нМ реагента 9-3 DD и 250 мкМ каждого дНТФ. Соединение 9-3 DD включает гаситель Дабцил, введенный в каждый концевой нуклеотид олигонуклеотида 9-3. Амплификацию осуществляют при 95°C в течение 3 минут; 10 циклов при 95°C в течение 10 сек, 65°C в течение 20 сек и 72°C в течение 20 сек; и 60 циклов при 95°C в течение 10 сек, 65°C в течение 20 сек, 72°C в течение 20 сек и 45°C в течение 20 сек. Параллельные реакции проводят либо с SYBR Green в разведении 1:40000, либо 2,4 мкМ молекулярного сигнала с низкой Тm, имеющего последовательность 5' FAM-CGTGCGCTCTGGTAAG GGTTTGCACG-Дабцил 3' (SEQ ID NO: 20) с целью раздельного мониторирования синтеза двунитевой и однонитевой ДНК. Каждый образец включает 6 нг (приблизительно 1000 геномов) человеческой ДНК. Кроме того, проводят контроль без добавления соединения 9-3 DD.
Результаты реакций амплификации показывают, что контроль без ДНК-полимеразы Taq горячего старта, но не соединение 9-3 DD проявляет существенный ошибочный старт, о чем свидетельствует повторное появление сигнала SYBR Green во время последних стадий амплификации после израсходования ограничивающего праймера. Поскольку продукт представляет собой данную смесь, невозможно секвенировать контроль. С другой стороны, результаты амплификации с добавлением соединения 9-3 DD показывают присутствие только требующегося специфического продукта без задержанного появления сигнала SYBR Green через 70 циклов. На основании концентрации ограничивающего праймера и допускаемой эффективности линейной амплификации 50% рассчитывают, что реакция генерирует 250 фмоль/мкл чистого продукта. Данный продукт представляют в отдел дидезоксисеквенирования Университета Брандейса для секвенирования.
Реакцию секвенирования проводят, используя стандартный протокол, например, капиллярный электрофорез с использованием ДНК-секвенатора Beckman CEQ 2000. Реакцию секвенирования проводят с 1/5 аликвоты амплифицированного продукта, что означает только с 50 фмоль продукта. Последовательность, полученная автоматизированным путем, представлена на Фиг.3, где показана последовательность, идущая вдоль верхней части схемы в виде ясных однозначных пиков продукта, из которого выделена последовательность, и в нижней части показан очень низкий уровень фонового сигнала. Правильность последовательности, полученной автоматизированным путем, подтверждают, сравнивая ее с известной последовательностью ампликона, имеющегося в GenBank, Регистрационный No. NT 010235.
Пример 4. Многофакторная ПЦР-амплификация
Супрессия ошибок, связанных с ошибочным стартом, особенно важна во многофакторных реакциях, когда одновременно амплифицируют множество продуктов. Присутствие нескольких пар праймеров и продуктов амплификации повышает вероятность взаимодействий ошибочного старта между и среди данных реакционных молекул. Многофакторные реакции, состоящие из увеличивающихся чисел ампликонов, проводят в отсутствие или в присутствии соединения 9-3 DD. Реакции ЛПЭ-ПЦР объемом 25 мкл, включающие образец, IX буфер для ПЦР, 3 мМ MgCl2, 100 мкМ каждого дНТФ, 1000 нМ избыточного праймера, 50 нМ ограничивающего праймера и 1,25 единицы полимеразы Taq Promega негорячего старта, подготавливают в отсутствие или в присутствии 300 нМ реагента 9-3 DD для различных комбинаций пар праймеров, как указано ниже. Образцы амплифицируют с использованием следующего профиля термического циклинга: 95°C в течение 3 минут; 10 циклов при 95°C в течение 10 сек, 65°C в течение 30 сек и 70°C в течение 30 сек; и 40 циклов при 95°C в течение 10 сек, 50°C в течение 30 сек, 70°C в течение 30 сек. В конце реакции образцы анализируют гель-электрофорезом в 3,5% агарозном геле в 0,5 × ТВЕ. Целевыми ампликонами и тестируемыми парами праймеров являются следующие:
Реакция A: два участка TSD 1278+TSD 1421 гена Нех-А, ассоциированного с болезнью Тея-Сакса.
Избыточный праймер TSD 1278: 5' GCCAGGGGTTCCACTACGTAGA 3' (SEQ ID NO: 21)
Ограничивающий праймер TSD 1278: 5' CCGCCCTTCTCTCTGCCCCCTGGT 3' (SEQ ID NO: 22)
Избыточный праймер TSD 1421: 5' CCGGGTCTCTAAGGGAGAACTCCT 3' (SEQ ID NO: 23)
Ограничивающий праймер TSD 1421: 5' CCGGCCGACAACACAAACCTGG TCC 3' (SEQ ID NO: 24)
Реакция B: ампликон TSD 1278, ампликон TSD 1421 и ампликон участка гена CFTR, CF экзон 10
Данная реакция включает те же пары праймеров, что и реакция A плюс
Избыточный праймер CF экзон 10: 5' GCTTTGATGACGCTTCTGTATCTA 3' (SEQ ID NO: 25)
Ограничивающий праймер CF экзон 10: 5' CAGTTTTCCTGGATTATGCCTG GCACCAT 3' (SEQ ID NO: 26)
Реакция C: ампликон TSD 1278, ампликон TSD 1421, ампликон CF экзон 10 и другой участок гена CFTR, ампликон CF экзон 11. Данная реакция включает те же пары праймеров, что и реакция B плюс
Избыточный праймер CF экзон 11: 5' TCGAAGTTTGCAGAGAAAGACAAT 3' (SEQ ID NO: 27)
Ограничивающий праймер CF экзон 11:5' TGACGTTTACAGCGAATGCTTG CTAGACCAAT 3' (SEQ ID NO: 28)
Реакция D: ампликон TSD 1278, ампликон TSD 1421, ампликон CF экзон 10, ампликон CF ex 11 и участок гена β-глобина человека.
Данная реакция включает те же пары праймеров, что и реакция С плюс
Избыточный праймер β-глобина: 5' TGGGTTTCTGATACGCACTGACTCTCTC 3' (SEQ ID NO: 29)
Ограничивающий праймер β-глобина: 5' GGCCATCACTAAAGGCACC GAGCACT 3' (SEQ ID NO: 30)
Гели, полученные при электрофорезе, окрашивают бромидом этидия для детекции двунитевых продуктов, образованных во время экспоненциальной фазы реакции ЛПЭ-ПЦР. На Фиг.4, 5, 6, 7 показывают подходящие части гелей продуктов, полученных в многофакторных реакциях A-D. Стрелки указывают на специфические двунитевые продукты амплификации, не обозначенные номерами полосы соответствуют специфическим однонитевым продуктам ДНК с вторичной структурой, и звездочки обозначают неспецифические продукты. Пары гелей A-D соотносятся с реакциями A-D, соответственно. Каждый гель, помеченный знаком "+", представляет собой продукт амплификации, включающей соединение 9-3 DD. Каждый гель, помеченный знаком представляет собой продукт амплификации без соединения 9-3 DD.
Пара гелей A показывает, что в отсутствие соединения 9-3 DD участок TSD 1278 (стрелка 41) не амплифицируется, но участок TSD 1421 (стрелка 42) амплифицируется вместе с большим числом неспецифических продуктов, преимущественно или полностью двунитевых, что проявляется в геле в виде пятна. Добавление соединения 9-3 DD уменьшает фон неспецифических продуктов и позволяет амплифицировать как TSD 1278 (стрелка 41), так и TSD 1421 (стрелка 42), амплифицируемых с большей чистотой, показывая предотвращение ряда проявлений ошибочного старта.
Пара гелей B показывает, что в отсутствие соединения 9-3 DD участки TSD 1278, TSD 1421 и CF экзон 10 (стрелка 43) амплифицируются вместе с большим числом неспецифических продуктов, которые проявляются в геле в виде пятна. Добавление соединения 9-3 DD устраняет фоновые несиецифические продукты, и получают все три ожидаемых ампликона.
Пара гелей C показывает, что в отсутствие соединения 9-3 DD участки TSD 1278, TSD 1421, CF экзон 10 и CF экзон 11 (стрелка 44) амплифицируются вместе с неспецифическим продуктом, указанным звездочкой. Добавление соединения 9-3 DD исключает синтез неспецифического продукта, и получают все четыре ожидаемых ампликона.
Пара гелей D показывает, что в отсутствие соединения 9-3 DD выбранный участок гена β-глобина (стрелка 45) не амплифицируется, но амплифицируются TSD 1278, TSD 1421, CF экзон 10 и CF экзон 11 вместе с неспецифическим продуктом, указанным звездочкой. Добавление соединения 9-3 DD исключает синтез неспецифического продукта, и дает возможность амплификации всех пяти ожидаемых ампликонов.
Демонстрируют также использование соединения, соответствующего изобретению, в многофакторных реакциях амплификации и детекции, когда только одна мишень присутствует в каком-либо заданном тест-образце, например, как это может иметь место при скрининге инфекционного патогена. В таком случае присутствует множество пар праймеров (используют множество пар избыточных праймеров и ограничивающих праймеров для ЛПЭ-ПЦР-амплификации), но только одна пара праймеров будет активной в отношении конкретного образца. С помощью флуоресценции SYBR Green и электрофоретического анализа после амплификации показано, что соединения, соответствующие настоящему изобретению эффективно ингибируютошибочныйстартв многофакторных реакциях, когда присутствуют в меньшем количестве, чем все субстраты.
Пример 5. Связь между Тх ствола и температурой отжига
Два разных протокола ЛПЭ-ПЦР-амплификации сравнивают, используя одну смесь для амплификации, включающую мишень. Каждую амплификацию проводят с 600 нМ соединения 9-3 DD и без соединения 9-3 DD. Как указано ранее, Тт ствола соединения 9-3 DD (т.е. рассчитанная температура плавления немодифицированного олигонукпеотида 9-3) составляет 56°C. Используют ДНК-полимеразу Taq Platinum для горячего старта. Две амплификации различаются в основном по используемой температуре отжига праймера, либо 65°C (выше Тт ствола), либо 55°C (ниже Тт ствола). Параметрами цикла амплификации являются:
Профиль A: 95°C в течение 3 минут; десять циклов 95°C в течение 10 сек, 65°C в течение 20 сек, 72°C в течение 20 сек; шестьдесят циклов 95°C в течение 15 сек, 55°C в течение 20 сек, 72°C в течение 20 сек, 50°C в течение 20 сек.
Профиль B: так же, как профиль A за исключением того, что последние шестьдесят циклов представляют собой 95°C в течение 10 сек, 65°C в течение 20 сек, 72°C в течение 20 сек, 45°C в течение 20 сек.
Реакции амплификации мониторируют в режиме реального времени, используя флуоресцентный краситель SYBR Green I для мониторирования синтеза двунитевого продукта, который выходит на плато (стабилизируется) в реакции ЛПЭ-ПЦР после израсходования ограничивающего праймера, если ошибочный старт не вызывает вариацию в последней части амплификации. Считывания проводят во время низкотемпературной стадии детекции.
Результаты представляют на Фиг.8 и 9. Панель A показывает считывания флуоресценции образцов копий Профиля A, когда используют температуру отжига 55°C после десяти циклов удлинения, как с включением (круг 52), так и без (круг 51) включения соединения 9-3 DD. Панель B показывает считывания флуоресценции образцов копий Профиля B, когда используют температуру отжига 65°C после десяти циклов удлинения, как с включением (круг 54), так и без (круг 53) включения соединения 9-3 DD.
На Фиг.8 и 9 показывают, что, когда используют температуру отжига праймера ниже Тт ствола реагента, соответствующего данному изобретению (Панель A), наблюдают задержку нескольких циклов в пороговом цикле, Ст. Однако когда поддерживают температуру отжига праймера существенно выше Тт ствола на протяжении экспоненциальной фазы ПЦР (Панель В), наблюдают незначительную, если она имеет место, задержку Ст. Отмечают, что для конкретного ампликона, используемого в данной амплификации, не наблюдают ошибочный старт Типа 2Дипа 3 с добавлением или без добавления соединения 9-3 DD, когда используют Taq горячего старта, показывая непредсказуемость появления ошибок, связанных с данным ошибочным стартом. Данный пример демонстрирует, что ствол реагента 9-3 DD закрыт, когда температура реакции падает до 55°C и что реагент остается связанным с полимеразой во время стадии удлинения реакции с более высокой температурой.
Пример 6. Влияние на активность ДНК-полимеразы при 25°C
Серию экспериментов по удлинению праймера проводят при 25°C с целью оценки влияния реагентов, соответствующих данному изобретению, и их немодифицированных олигонукпеотидных аналогов. Каждая реакционная смесь включает матрицу, праймер и ДНК-полимеразуТад. Реакцию проводят в течение двух часов в каждом случае. Праймер метят флуоресцентной меткой Су5. В реакционную смесь добавляют флуоресцентный краситель SYBR Green. После реакций удлинения строят кривые плавления, в которых стимулируют краситель и считывают эмиссию флуорофора, обусловленную FRET (переносом энергии резонансной флуоресценции) - переносом с красителя, какописано в одновременно поданной предварительной патентной заявке США под названием "Праймеры, зонды и способы амплификации нуклеиновых кислот". Результаты представляют на Фиг.10, 11, 12.
Фиг.10, включает кривую плавления 61, полученную, когда удлинению препятствуют, исключая дНТФ из реакционной смеси. Кривая 61 представляет собой, таким образом, кривую плавления гибрида праймер-мишень, показывающую Tm 58°C. Панель A также включает ряд контролей, которые содержат дНТФ, либо в виде монокомпонента, либо с немодифицированным олигонуклеотидом 9-3 (300 нМ, 1000 нМ) или 9-22 (50 нМ, 100 нМ, 300 нМ). Кривые, выделенные кругом 62, для контролей показывают удлинение во всех случаях, т.е. пик плавления праймера исчезает при 58°C. На Фиг.11, показывают дозозависимый эффект включения реагента 9-22 DD в реакционную смесь: кривая 63, 50 нМ; кривая 64, 100 нМ; кривая 65, 300 нМ. Чем больше пик при 58°C, тем больше эффект предотвращения удлинения посредством полимеризации в изотермических условиях теста. На Фиг.12, показывают дозозависимый эффект включения реагента 9-3 DD в реакционную смесь: кривая 66, 300 нМ; кривая 67, 1000 нМ. Снова, чем больше пик при 58С, тем сильнее эффект предотвращения удлинения посредством полимеризации в условиях теста.
Исследуют также влияние на фрагмент Стоффеля (см. статью Lawyer et al. (1993) PCR Methods и Applications 2:275-287) ДНК-полимераз с отсутствием активности 5'-3'-экзонуклеазы. Сравнивают различные соединения, перечисленные в Таблице 1, с отрицательным контролем, включающим только матрицу, праймер с флуоресцентной меткой, фрагмент Стоффеля и дНТФ. В данной серии удлинение осуществляют при 40°C и считывания SYBR Green проводят каждые 20 секунд в течение более, чем 30 минут. Кривые сравнивают визуально, это надежно, поскольку либо полученная кривая почти точно следует за контролем, либо, в противном случае, полученная кривая значительно отличается от контроля. Тест-соединения добавляют в концентрации 50 нМ, 100 нМ и 300 нМ. Немодифицированные шпильки 6-22, 9-3, 9-5 и 9-22 не подавляют элонгацию фрагментом Стоффеля в любой концентрации. Соединение 6-22DD подавляет элонгацию во всех трех концентрациях, как и соединения 9-3DD, 9-5DD и 9-22DD. Следующие соединения не подавляют элонгацию в концентрациях 50 М или 100 нМ, но подавляют элонгацию в концентрации 300 нМ: 9-22-5D, 9-22-3D, 9-5 2'ОМ4, 9-5BHBHQ и 9-32'ОМ4.
Пример 7. Влияние на активность экзонуклеазы Tag в отсутствие активности полимеразы
Разработан анализ, который демонстрирует ингибирование 5'-3'-экзонуклеазной активности в отсутствие синтеза ДНК. В анализе в качестве субстрата используют специфическую двунитевую молекулу ДНК, в которой одна нить помечена флуорофором FAM на 5'-конце и другая нить помечена на 3'-конце гасителем Дабцил. Флуорофор находится в тесной близости с гасителем, и флуоресценция отсутствует при стимуляции флуорофора. Денатурация и отжиг нити при нагревании и охлаждении вызывает расщепление нити, меченной FAM, и высвобождение флуорофора. Считают, что циклинг каким-то образом генерирует субстрат для 5'-3'-экзонуклеазной активности полимеразы Taq. Вследствие этого возрастание флуоресценции представляет меру 5'-3'-экзонуклеазной активности активности полимеразы Taq.
25 мкл реакционной смеси включает 300 нМ двунитевой ДНК-матрицы в IX буфере для ПЦР, 3 мМ MgCl2, 1,25 единицы, (U), полимеразы Taq и присутствие или отсутствие соответствующей концентрации соединения 9-22 DD. Реакции не включают ни дНТФ, ни другую нуклеиновую кислоту-мишень кроме двунитевой ДНК. Последовательности комплементарных нитей двунитевой ДНК-матрицы представляют собой 5' FAM-AGTGTGATGA
TGGTGAGG-фосфат 3' (SEQ ID NO: 31) и 5'- ACTTTCAACTCTGTCT 3'-Дабцил (SEQ ID NO: 32). Последовательности денатурируют при 95°C в течение 3 минут и затем подвергают циклам: 95°C в течение 10 сек, 67°C в течение 30 сек, 72°C в течение 30 сек и 45°C в течение 20 сек. Флуоресценцию обнаруживают во время каждого цикла при 45°C.
Результаты считывания флуоресценции при проведении анализа на нескольких образцах представлены на Фиг.13, где результаты считывания нормализованы, начиная с одинаковой фоновой флуоресценции. Кривая 76 представляет собой контроль, показывающий флуоресценцию без присутствия полимеразы Taq. Полимеразу Taq включают в остальные анализы. Кривая 71 показывает возрастание флуоресценции, происходящее, когда не добавляют реагент 9-22 DD. Флуоресценция постоянно возрастает на протяжении тридцати циклов. Кривые 72-75 показывают возрастание флуоресценции с реагентом 9-22 DD, включенным в концентрации 50 нМ, 100 нМ, 200 нМ и 300 нМ, соответственно. Добавление соединения 9-22 DD к вышеуказанной реакции снижает наблюдаемую флуоресценцию дозозависимым образом.
Пример 8. Симметричная ПЦР-амплификация
Проводят анализ ПЦР-амплификаций в режиме реального времени, используя расщепленную геномную ДНК и пару праймеров с близкой Тm в эквимолярной концентрации. Осуществляют анализы копий с добавлением и без добавления 400 нМ реагента 9-3 DD. Действия реагента оценивают на основе кинетики накопления суммарной ДНК путем окрашивания SYBR Green, а также с помощью гель-электрофореза при окрашивании бромидом этидия.
Намеченной мишенью амплификации является аллель гена CFTR, которая находится в интроне 19 в виде транзиции одного основания (GenBank регистрационый No. АС000061). 25 мкл реакционной смеси включают в субстрат в виде 120 пикограмм (пг) расщепленной геномной ДНК человека, 1 × буфер для ПЦР, 5 мМ MgCl2, 250 мкМ каждого дНТФ, SYBR Green в разведении 1:40000, 1000 нМ прямого праймера: последовательность 5' TAATTACAAGAGTCTTCCAT 3' (SEQ ID NO: 33), Tm 56,6°C и 1000 нМ обратного праймера: последовательность 5' CATGAATAGAACATTTCCTT 3' (SEQ ID NO: 34), Tm 56,3°C, и 1,25 единицы полимеразы Taq негорячего старта Invitrogen в отсутствие или в присутствии 400 нМ реагента 9-3 DD. Образцы амплифицируют с использованием профиля термического циклинга 95°C в течение 3 минут; 60 циклов при 95°C в течение 10 сек, 55°C в течение 30 сек и 72°C в течение 30 сек с мониторированием флуоресценции SYBR Green I при 72°C и в конце градиентом температуры от 54°C до 96°C при повышении на 1°C в течение 30 сек. В конце реакции несколько случайно выбранных образцов из каждой серии реакций анализируют с помощью гель-электрофореза в 3,0% агарозном геле в растворе 0,5 × Трис, борной кислоты, ЭДТА (ТВЕ).
На Фиг.14 и 15 представляют полученные результаты. Схема амплификации на Фиг.14 показывает флуоресценцию SYBR Green I на протяжении 60 циклов с кривыми, обозначенными кругом 81, которые представляют собой образцы копий, которые не включают реагент 9-3 DD, и кривыми, обозначенными кругом 82, представляющими собой образцы копий при содержании 400 нМ 9-3 DD. Кинетика флуоресцентных сигналов, возникающих в двух сериях образцов, демонстрирует, что суммарная двунитевая ДНК накапливается быстрее в отсутствие реагента 9-3 DD, чем в присутствии реагента, но общее количество двунитевой ДНК, накапливающееся на протяжении 60 циклов, в действительности идентично в двух сериях реакций.
На Фиг.15 показывают электрофоретические гели при окрашивании бромидом этидия. Слева дорожка а представляет собой маркер размера (разница 50 пар оснований), и дорожки b-i представляют собой примеры без реагента 9-3 DD. Справа дорожки j-q представляют собой примеры с включением 400 нМ 9-3 DD, и дорожка г является маркером размера. Результаты показывают, что реакции, которые не включают реагент, соответствующий данному изобретению, генерируют правильный продукт, а также много неспецифических более высокомолекулярных продуктов, тогда как реакции, которые включают реагент 9-3 DD, генерируют только правильный продукт. Более того, реакции, которые включают реагент 9-3 DD, генерируют приблизительно в два раза больше правильного продукта, чем не включающие его реакции, что оценивают по относительным интенсивностям полосы правильного продукта в гелях.
Сочетание кинетического анализа и электрофоретического анализа показывает, что реакции 81, которые генерируют неспецифические продукты, образуют флуоресцентные сигналы раньше, чем реакции 82, которые генерируют только правильный продукт. Краситель SYBR Green интеркалирует в двунитевую ДНК безотносительно специфичности последовательности. Таким образом сигмоидальная кинетика реакций 82 по сравнению с более прямолинейной кинетикой реакций 81 может быть использована для оценки того, накапливает или не накапливает симметричная реакция только правильный продукт.
Пример 9. Количественная оценка ингибированин экзонуклеазы
Данный пример описывает точный анализ, направленный на измерение ингибирующего действия реагента, соответствующего данному изобретению, на 5'-3'-экзонуклеазную активность ДНК-полимеразы Taq. В данном анализе используют систему праймер-матрица, аналогичную описанной в статье М.W. Kaiser, N. Lyamicheva, W. Ma, С. Miller, В. Neri, L. Fors, and V.I. Lyamicheva в J. Bio. Chem., 274, стр.21387-21394 (1999), за исключением того, что матрицу метят Су5 на 5'-конце и анализ проводят в присутствии SYBR Green I, красителя ДНК, который флуоресцирует при связывании с двунитевой ДНК. Матрица для данного анализа состоит из олигонуклеотидной последовательности 5'-Су5-AAAACGCTGTCTCGCTGAAAGCGAGACAGCGA
AAGACGCTCGT-3' (SEQ ID NO: 35), которую отжигают с праймером нуклеотидной последовательности: 5'-ACGAGCGTCTTTC-3' (SEQ ID NO: 36). Более длинный олигонуклеотид образует структуру шпильки с двумя однонитевыми хвостами различной длины. Более короткий 5'-хвост матрицы включает флуорофор Су5 и состоит из 4 остатков аденозина; более длинный 3'-хвост матрицы служит мишенью для олигонуклеотида праймера, так что праймер сидит непосредственно впереди 5'-хвоста матрицы с перекрыванием в одну пару оснований. Добавление SYBR Green I к данному комплексу матрица-праймер приводит в результате к связыванию SYBR Green I с двунитевыми участками ДНК комплекса. При возбуждении SYBR Green I лазером с длиной волны 480 нм энергия флуоресценции связанного красителя ДНК полностью поглощается Су5, что, как считают, происходит вследствие переноса энергии резонансной флуоресценции, и определяемая флуоресценция отсутствует при длине волны максимума эмиссии SYBR Green (длина волны максимальной эмиссии: 521 нм). Гидролиз 5'-хвоста посредством 5'-3'-экзонуклеазной активности полимеразы Taq приводит к удалению группы Су5 из комплекса матрица-праймер и восстановлению определяемой флуоресценции SYBR Green I. Кинетика восстановления флуоресценции SYBR Green I в присутствии ДНК-полимеразы Taq обеспечивает количественное измерение 5'-3'-экзонуклеазной активности. Реагенты, соответствующие данному изобретению, ингибируют 5'-3'-экзонуклеазргую активность полимеразы Taq и замедляют рост флуоресценции SYBR Green I.
Анализ состоит из 0,5 мкМ вышеуказанной матрицы и 1,5 мкМ вышеуказанного праймера, смешанных в 1X буфере для ПЦР, 3 мМ MgCl2, коммерческом исходном SYBR Green I в разведении 1:40000 (Molecular Probes, Eugene, OR), 1,25 ед. полимеразы Taq (Invitrogen, Calrsbad, CA) в отсутствие или в присутствии соединения 9-22 DD в концентрации 50 нМ, 100 нМ, 300 нМ или 1000 нМ в объеме 25 микролитров (мкл). В реакционную смесь не добавляют дНТФ, поскольку анализ не основан на ДНК-полимеразной активности. Реакционную смесь без матрицы и праймера выдерживают при 25°C для стимуляции взаимодействия соединения 9-22 DD и ДНК-полимеразы Taq. Затем образец помещают на лед, вносят матрицу и праймеры и хранят на льду до начала реакции. Отрицательные контроли не содержат ДНК-полимеразу Taq. Реакцию инициируют инкубированием образца при 25°C в Детекторе последовательностей 7700 ABI Prism в течение 60 минут со сбором флуоресценции в канале SYBR Green I каждые 30 секунд. Для каждого контроля и каждой концентрации реагента 9-22 DD берут среднее по трем различным испытаниям.
На Фиг.16 представляют результаты анализа. Определяемая флуоресценция SYBR Green I обеспечивает измерение степени 5'-3'-экзонуклеазной активности. Контроль "Без Taq" показывает отсутствие сигнала флуоресценции, согласующееся с полным поглощением флуоресценции SYBR Green I посредством Су5 и полное отсутствие 5'-3'-экзонуклеазной активности (линия 96). Добавление полимеразы Taq восстанавливает флуоресценцию SYBR Green I и дает исходный уровень для максимальных уровней 5'-3'-экзонуклеазной активности в условиях анализа (линия 91). Добавление соединения 9-22 DD к реакционной смеси замедляет восстановление флуоресценции SYBR Green I дозозависимым образом вследствие ингибирующего эффекта 9-22 DD на 5'-3'-экзонуклеазную активность полимеразы Taq (линия 92, 50 нМ; линия 93, 100 нМ; линия 94, 300 нМ; линия 95, 1000 нМ). В Таблице III ниже приводят количественные данные по проценту ингибирования 5'-3'-экзонуклеазной активности различными концентрациями соединения 9-22 DD на основе относительных уровней флуоресценции SYBR Green I, достигнутых через 60 минут инкубирования при 25°C.
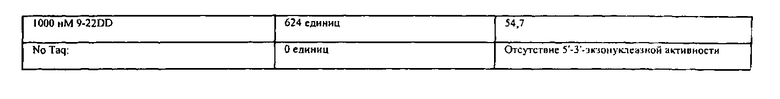
Пример 10. Предотвращение формирования димера праймера и олигомеризации в присутствии и в отсутствие ДНК-мишени
Серию реакций ЛПЭ-ПЦР-амплификации проводят в присутствии или в отсутствие 100 геномов плацентарной ДНК человека (Sigma, St Louis, МО) в конечном объеме 25 мкл. В данном эксперименте реакции ЛПЭ-ПЦР-амплификации состоят из 1X буфера для ПЦР (Invitrogen, Carlsbad, СА), 3 мМ MgCl2, 0,25 мМ дНТФ, 1000 нМ избыточного праймера, 50 нМ ограничивающего праймера, 0,25 мкМ меченной FAM нити зонда, 0,3 мкМ меченной Дабцилом обратной комплементарной нити, 1,25 единиц ДНК-полимеразы Taq Platinum (Invitrogen, Carlsbad, СА). Используют следующие последовательности праймеров и зондов:
Избыточный праймер: 5' GTTTCTTTGCTGCCGTGTTC 3' (SEQ ID NO: 37)
Ограничивающий праймер: 5' CCCCAGAGACCCCAGTTGCTAACCAGAC 3' (SEQ ID NO: 38)
Меченная FAM нить зонда: 5' [TET]AGACAGAGTTGAAAGTCAGG[Phos] 3' (SEQ ID NO: 39)
Меченная Дабцилом обратная комплементарная нить: 5' ACTTTCAACTCTGTCT [Дабцил] 3' (SEQ ID NO: 40)
Данные праймеры и зонды амплифицируют и детектируют ампликон из 488 пар оснований (пар осн.), который охватывает экзоны 5 и 6 гена р53 человека.
Амплификацию проводят в детекторе последовательностей 7700 ABI Prism (Applied Biosystems, СА) с термическими профилями, которые состоят из 1 цикла при 950°C в течение 3 минут; 25 циклов при 95°C в течение 10 сек, 64°C в течение 30 сек, 75°C в течение 30 сек и 35 циклов при 95°C в течение 10 сек, 64°C в течение 30 сек, 75°C в течение 30 сек, 45°C в течение 20 сек при детекции флуоресценции в канале ТЕТ во время стадии 45°C.
Полученные в результате продуты амплификации анализируют гель-электрофорезом в 3% агарозном геле в 0,5X буфере ТВЕ в течение 2 часов и окрашивают бромидом этидия. Результаты представляют на Фиг.17. Образцы на дорожках 3, 5, 7, 9, 11 получены без геномной ДНК. Образцы на дорожках 2, 4, 6, 8, 10 получены с геномной ДНК. Дорожка 1 включает электрофоретические маркеры размера с шагом 100 пар основания. Реагент 9-22 DD добавляют в исходные реакции следующим образом: дорожки 2 и 3 - 0 нМ; дорожки 4 и 5 - 50 нМ; дорожки 6 и 7 - 100 нМ; дорожки 8 и 9 - 200 нМ; дорожки 10 и 11 - 300 нМ.
Сбоку с правой стороны геля отмечают размер правильного продукта, а также размеры, которые, как полагают, являются димерами праймера и олигомерами праймера. Результаты, представленные на Фиг.17, демонстрируют, что в реакциях, инициируемых геномной ДНК, повышение концентрации реагента 9-22 DD действует дозозависимым образом в плане предотвращения проявления неспецифических продуктов, включая димеры праймера и олигомеры праймера и, таким образом, повышая как специфичность, так и выход правильных продуктов. Повышение концентрации реагента 9-22 DD также предотвращает проявление димеров праймера и олигомеров праймера в реакциях, которые не включают геномную ДНК.
Пример 11. Предотвращение формирования димера праймера и в двойной реакции
Серию реакций ЛПЭ-ПЦР-амплификации проводят в присутствии или в отсутствие 100 геномов плацентарной ДНК человека (Sigma, St Louis, МО) в конечном объеме 25 мкл. В данном эксперименте реакции ЛПЭ-ПЦР-амплификации состоят из 1X буфера для ПЦР (Invitrogen, Carlsbad, СА), 3 мМ MgCl2, 0,20 мМ дНТФ. Полимеразу Taq используют в концентрации 1,25 единиц в каждом образце. Первая целевая последовательность амплификации (Продукт 1) представляет собой часть экзона 11 гена кистозного фиброза, и ее амплифицируют с ограничивающим праймером: 5' GACGTTTACAGCGAATGCTTGCTAGACCAAT 3' (SEQ ID NO: 41) в концентрации 100 нМ и избыточным праймером: 5' TCCAAGTTTGCAGAGAAAGACAAT 3' (SEQ ID NO: 42) в концентрации 2000 нМ. Вторая целевая последовательность амплификации (Продукт 2) представляет собой часть экзона 10 гена кистозного фиброза, и ее амплифицируют с ограничивающим праймером: 5' CAGTTTTCCTGGATTATGCCTGGCACCAT 3' (SEQ ID NO: 43) в концентрации 50 нМ и избыточным праймером: 5' GCTTTGATGACGCTTCTGTATCTA 3' (SEQ ID NO: 44) в концентрации 1000 нМ.
Амплификацию проводят в Детекторе последовательностей 7700 ABI Prism (Applied Biosystems, СА) 2 мин при 95°C с последующими 25 циклами при 95°C в течение 10 сек, 56°C в течение 15 сек, 70°C в течение 20 сек, с последующими 50 циклами при 95°C в течение 10 сек, 56°C в течение 15 сек, 70°C в течение 20 сек и 45°C в течение 30 сек с обнаружением флуоресценции.
Продукты, полученные в результате амплификации, анализируют гель-электрофорезом в 3% агарозном геле с 0,5X буфером ТВЕ в течение 2 часов и окрашивают бромидом этидия. Результаты показывают на Фиг.18. В реакциях, анализируемых на дорожках 1-6, используют полимеразу Taq негорячего старта, тогда как тогда как в реакциях, анализируемых на дорожках 7-12, используют полимераза Taq с антителом горячего старта. Реагент 9-22 DD добавляют к анализируемым на дорожках реакциям 4-6 и 10-12 в концентрации 100 нМ.
Сбоку справа от геля прилагают интерпретацию идентичности продуктов амплификации, включая однонитевые ампликоны (ssДHК) и двунитевые ампликоны (dsДНK) Продукта 1 и Продукта 2, а также более короткие продукты, которые, как полагают, являются димерами праймера. Другие неспецифические продукты отмечают звездочкой. Амплификации с полимеразой негорячего старта и без реагента, соответствующего изобретению, дорожки 1-3, дают димеры праймера и другие неспецифические продукты. Амплификации с полимеразой горячего старта и без реагента, соответствующего изобретению, дорожки 7-9, в целом до некоторой степени чище, но, те не менее, показывают проявление ошибочного старта. Добавление реагента, соответствующего данному изобретению, к амплификациям с горячим стартом, дорожки 10-12, дают только целевые продукты. Добавление реагента, соответствующего данному изобретению, к амплификациям негорячего старта, дорожки 4-6, исключает димеры праймера и в двух из трех повторностей (дорожка 4 и дорожка 6) также все другие неспецифические продукты, причем только одна повторность (дорожка 5) проявляет неспецифический продукт. Результаты, представленные на Фиг.18, демонстрируют, что низкая концентрация (100 нМ) реагента 9-22 DD достаточна для предотвращения формирования димеров праймера при использовании либо полимеразы Taq негорячего старта, либо полимеразы Taq горячего старта, кроме того, то, что низкая концентрация реагента 9-22 DD почти достаточна для предотвращения всех проявлений ошибочного старта даже при использовании с полимеразой Taq негорячего старта в амплификации двух мишеней с двумя парами праймеров.
Пример 12. Оптимизация кинетики двойной ПЦР в режиме реального времени и ПЦР в режиме реального времени путем предотвращения формирования димера праймера
Разрабатывают анализ двойной ЛПЭ-ПЦР в режиме реального времени для одновременной амплификации последовательностей внутри экзонов генов Oct4 и Xist мыши (GenBank Accession No NM_013633 и L04961, соответственно). Каждую реакцию проводят в конечном объеме 50 мкл, и она включает следующие реагенты: 1X буфер для ПЦР (Invitrogen, Carlsbad, СА), состоящий из 20 мМ Трис-HCl, pH 8,4 и 50 мМ KCl, 3 мМ MgCl2, 0,4 мМ каждого дНТФ, 50 нМ ограничивающего праймера Oct4, имеющего последовательность 5' TGGCTGGACACCTGGCTTCAGACT 3' (SEQ ID NO: 45), 2 мкМ избыточного праймера Oct4, имеющего последовательность 5' CAACTTGGGGGACTAGGC 3' (SEQ ID NO: 46), 100 нМ ограничивающего праймера Xist, имеющего последовательность 5' GGTCGTACAGGAAA
AGATGGCGGCTCAA 3' (SEQ ID NO: 47), 2 мкМ избыточного праймера Xist, имеющего последовательность 5' TGAAAGAAACCACTAGAGGGCA 3' (SEQ ID NO: 48), 1 мкМ молекулярного сигнала Oct4 с низкой Тm, имеющего последовательность 5' TET-CCG ССТ GGG ATG GCA ТАС TGT GGA AGG CGG-Дабцил 3' (SEQ ID NO: 49) и 2 единицы комплексированной с антителом ДНК-полимеразы Taq Platinum® (Invitrogen, Carlsbad, СА). Либо соединение 9-3DD, либо соединение 9-3bDD также включают в смесь для ПЦР в концентрации, определенной ниже. Молекулярный сигнал для детекции ампликонов Xist в данном примере не добавляют. Каждый анализ содержит также реагенты, необходимые для лизиса клеток и обратной транскрипции согласно протоколу PurAmp (см. статью Hartshorn et al. (2005) ВМС Biotechnol 5:2) в объеме 2,5 мкл. В данной двойной ЛПЭ-ПЦР конечные концентрации данных реагентов являются следующими: 2,5 мМ Трис-ацетата, pH 8,4, 3,75 мМ ацетата калия и 0,4 мМ ацетата магния (разведенный буфер для синтеза кДНК, ThermoScript™ Система ОТ-ПЦР, Invitrogen, Carlsbad, СА), дополнительные 50 мкМ каждого дНТФ, 0,13 мМ изотиоцианата гуанидина, 6,7 мкМ β-меркаптоэтанола, 0,7 мкМ цитрата натрия, pH 7,0, 0,7×10-4% (об./об.) диметилсульфоксида и 0,2×10-4% саркозила.
Геномную ДНК мыши (Sigma, St Louis, МО) также добавляют к каждому анализу, и она обеспечивает матрицы для ПЦР-амплификации. Число геномов, добавленных в каждую пробирку, рассчитывают, основываясь на 6 пг/размер генома (см. статью Vendrely and Yendrely (1949) Experientia 5: 327-329).
Амплификацию проводят в детекторе последовательностей 7700 ABI Prism (Applied Biosystems, СА) с термическим профилем, состоящим из 1 цикла при 95°C в течение 5 минут; 15 циклов при 95°C в течение 10 сек., 63°C в течение 20 сек. и 72°C в течение 30 сек. и 40 циклов при 95°C в течение 15 сек., 55°C в течение 25 сек., 72°C в течение 35 сек. и 45°C в течение 30 сек. при детекции флуоресценции в канале ТЕТ при 45°C.
В конце ПЦР продукты амплификации анализируют гель-электрофорезом в 3% агарозном геле в 0,5X буфере ТВЕ в течение 2 часов и окрашивают бромидом этидия. Как двунитевые, так и однонитевые продукты ожидаемых размеров визуально определяют для двух соамплифицируемых генов, свидетельствуя об эффективной двойной ЛПЭ-ПЦР.
Фиг.19 иллюстрирует эффект варьирования композиции ствол реагента 9-3DD на кинетику вышеописанной двойной ЛПЭ-ПЦР, включающей два набора праймеров для амплификации двух негомологичных последовательностей внутри генов Oct4 и Xist. На Фигуре показывают флуоресцентные сигналы, генерируемые накоплением ампликонов Oct4, посредством гибридизации с молекулярным сигналом TET-Oct4. Результаты демонстрируют, что значения Ст очень близки в присутствии 300 нМ 9-3DD (линии с треугольниками) или его модифицированной формы 9-3bDD в той же концентрации (жирные линии без треугольников) для каждой анализируемой концентрации геномов (10 геномов, круг 122; 100 геномов, круг 123; 1000 геномов, круг 124. Композиция ствола реагента, напротив, влияет на кинетику флуоресцентных сигналов в режиме реального времени. Соединение 9-3bDD имеет более высокую Тт ствола по сравнению с 9-3DD, таким образом, как полагают, оптимально предотвращая формирование димера праймера, что, в свою очередь, дает в результате очень линейные и параллельные сигналы при всех тестируемых концентрациях генома. В присутствии менее жесткого соединения 9-3DD, напротив, некоторые флуоресцентные сигналы имеют более крутой наклон, но другие рано выходят на плато в реакции, указывая на случайное формирование димеров праймера. Сигналы, генерируемые при различных концентрациях матрицы, с соединением 9-3DD менее параллельны, чем получаемые в присутствии соединения 9-3bDD (с модифицированным стволом). Линейная амплификация, генерирующая совершенно параллельные сигналы (имеющие постоянный наклон), требуется в идеале и особенно подходит для типов анализа конечных результатов.
Анализ в агарозном геле образцов, содержащих 9-3DD, показывает полосы, особенно заметные при повышенных количествах матриц, которые могут представлять собой димеры праймера, и очень маленькие полосы при более низких количествах матриц, которые соответствуют праймерам. Данные полосы не появляются при анализе образцов, включающих 9-3bDD.
Фиг.20 иллюстрирует эффект варьирования концентрации реагента 9-3bDD в двойной ЛПЭ-ПЦР, описанной в данном примере, а также используемой для Фиг.19. В данном случае снижение концентрации реагента 9-3bDD от 300 нМ (жирные линии без треугольников) до 200 нМ (линии с треугольниками) снова воздействует на наклон линейной амплификации вследствие формирования димера праймера, как подтверждают анализом в агарозном геле. В результате некоторые образцы, включающие низкое исходное количество матриц (10 геномов, круг 132), имеют более высокую флуоресценцию в последнем цикле ("конечный результат"), чем образцы, содержащие большее исходное количество матриц (100 геномов, круг 133 и 1000 геномов, круг 134).
Влияние концентрации реагентов, описанных в данной патентной заявке, на эффективность ПЦР тестируют также в ЛПЭ-ПЦР, амплифицируя одну матрицу с одной парой праймеров. Амплифицированная матрица представляет собой ту же последовательность Oct4 мыши, которую используют в двойном анализе, описанном на Фиг.19 и Фиг.20, и праймеры, а также последовательности молекулярного сигнала такие же, как в той реакции.
Каждую реакцию проводят в конечном объеме 100 мкл, и она содержит следующие реагенты: 1X буфер для ПЦР (Invitrogen, Carlsbad, СА), состоящий из 20 мМ Трис-HCl, pH 8,4, и 50 мМ KCl, 3 мМ MgCl2, 0,25 мМ каждого дНТФ, 50 нМ ограничивающего праймера Oct4, 2 мкМ избыточного праймера Oct4, 1 мкМ молекулярного сигнала TET-Oct4 с низкой Тm и 2 единицы комплексированной с антителом ДНК полимеразы Taq Platinum® (Invitrogen, Carlsbad, CA). Соединение 9-3DD также включают в смесь для ПЦР в концентрациях 150 или 300 либо 450 нМ. Как в случае вышеупомянутой двойной реакции (см. Фиг.19 и Фиг.20), каждый анализ включает также реагенты, необходимые для лизиса клеток и обратной транскрипции согласно протоколу PurAmp в объеме 10,5 мкл. В данной ЛНЭ-НЦР конечные концентрации данных реагентов являются следующими: 5 мМ Трис-ацетата, pH 8,4, 7,5 мМ ацетата калия и 0,8 мМ ацетата магния (разведенный буфер для синтеза кДНК), 1 нг/мкл случайных гексамеров и дополнительно 100 мкМ каждого дНТФ (все компоненты системы RT-ПЦР ThermoScript™, Invitrogen, Carlsbad, CA), 0,4 мМ изотиоцианата гуанидина, 20 мкМ β-меркаптоэтанола, 2 мкМ цитрата натрия, pH 7,0, 2×10-4% (об./об.) диметилсульфоксида и 0,5×10-4% саркозила.
Геномную ДНК мыши также добавляют к каждому анализу, и она обеспечивает матрицы для ПЦР-амплификаций, как определено для двойной ЛПЭ-ПЦР, описанной в данном примере. Амплификацию снова проводят в детекторе последовательностей 7700 ABI Prism с тем же термическим профилем, детально описанным для двойной реакции. В конце ПЦР продукты амплификации анализируют гель-электрофорезом в 3% агарозном геле в 0,5Х буфере ТВЕ в течение 2 часов и окрашивают бромидом этидия. Как двунитевые, так и однонитевые продукты видны в геле и имеют ожидаемые размеры, указывая на эффективную амплификацию посредством ЛПЭ-ПЦР.
Тестируют также влияние повышения концентрации соединения 9-3DD на эффективность амплификации матицы Oct4. Значения СТ получают из ПЦР-анализов в режиме реального времени, включающих 5 геномов (среднее по двум повторностям для каждой тестируемой концентрации 9-3DD), 20 геномов (среднее но двум повторностям для каждой тестируемой концентрации 9-3DD), 100 геномов и 1000 геномов, проведенных с каждой из тестируемых концентраций 9-3DD: 150 нМ, 300 нМ, 450 нМ. Линейная регрессия для каждой серии точек СТ в каждой концентрации представлена на Фиг.21, где треугольники означают концентрацию 450 нМ, белые квадраты означают 300 нМ и черные квадраты означают 150 нМ. Из данного графика непосредственно очевидно, что 450 нМ 9-3DD сильно задерживает появление флуоресцентных сигналов, превышающих пороговый уровень, изменяя также зависимость ПЦР-амплификации от дозы (см. точки 10 копий гена и 40 копий гена, отстоящие от линии регрессии). Значения СТ, полученные с использованием 150 нМ или 300 нМ 9-3DD, с другой стороны, очень близки и демонстрируют линейную связь между числом копий матрицы и первым появлением флуоресцентного сигнала.
Однако, анализ образцов флуоресценции в режиме реального времени подчеркивает разницу между двумя условиями (концентрациями), как иллюстрируют на Фиг.22. Наклон кривых, генерированных либо 100 геномами, круг 151, либо 1000 геномами, круг 152, более крутой при использовании 300 нМ 9-3DD (жирные линии без треугольников), чем при использовании 150 нМ 9-3DD (линии с треугольниками), позволяя предположить, что более жесткие условия 300 нМ, элиминируют димеры праймера, повышая, таким образом, эффективность амплификации. Пример 13. Применение многочисленных реагентов, соответствующих данному изобретению.
Реакционные смеси для ЛПЭ-ПЦР получают, чтобы амплифицировать исходный материал пяти различных последовательностей-мишеней геномной ДНК человека и амплифицировать все пять совместно в многофакторной реакции. Для каждой последовательности-мишени требуется своя собственная пара ограничивающего и избыточного праймеров. Целевые последовательности представляют собой (1) аллель глобина из 191 пары оснований, (2) аллель G269 Тея-Сакса из 312 пар оснований, (3) аллель 1278 и 1421 Тея-Сакса из 452 пар оснований, (4) сегмент митохондриального гипервариабельного участка 1 из 549 пар оснований и (5) сегмент, включающий экзоны 7 и 8 гена р53 из 611 пар оснований. 25 мкл реакционных смесей включают 1х буфер для ПЦР (Invitrogen), 0,4 мМ дНТФ, 3 мМ Mg++, 0,24 × SYBR Green и 1,5 единицы ДНК-полимеразы Taq (Invitrogen). Праймеры добавляют в концентрации 50 нМ для ограничивающего праймера и 1000 нМ для избыточного праймера. Все пять пар праймеров добавляют к многофакторным реакциям. Термический циклинг составляет сорок пять циклов при 95°C в течение 10 сек., 64°C в течение 20 сек. и 72°C в течение 1 минуты.
Обе реакционные смеси с индивидуальными парами праймеров и индивидуальными мишенями и реакционные смеси со всеми пятью парами праймеров и всеми пятью мишенями амплифицируют в присутствии комбинации 25 нМ соединения 9-22DD и 100 нМ соединения 12-3DD. Пятифакторную реакционную смесь амплифицируют также без добавления какого-либо реагента, соответствующего изобретению. Продукты реакции исследуют с помощью гель-электрофореза. Результаты гель-электрофореза различных амплификации показаны на Фиг.23, которая включает маркеры размера (разница 100 пар оснований) на дорожке r. Центральные дорожки g-k представляют собой продукты индивидуальных амплификации последовательностей-мишеней (1)-(5), соответственно. Дорожки 1-q представляют собой продукты пятифакторных амплификации в шести повторностях, которая включает смесь соединения 9-22DD и соединения 12-3DD. Дорожки a-f представляют собой продукты пятифакторных амплификации в шести повторностях, которые не включают реагент, соответствующий изобретению. На Фиг.23 показывают, что смесь соединений усиливает амплификацию (больше требующихся ампликонов, и они чище) по сравнению с многофакторной реакцией без добавления какого-либо соединения, демонстрируя, что смеси реагентов можно использовать в наборах, амплификациях и анализах, соответствующих данному изобретению.
Пример 14. Тестирование способов действия
Разрабатывают тест для количественного исследования снижения уровня ошибочного старта (повышения специфичности) и ингибирующих полимеразу действий реагентов, соответствующих данному изобретению, которые, как полагают, отличает Первый способ действия от Второго способа действия. Тест представляет собой анализ ПЦР-амплификации, в основном аналогичный анализу, описанному в Примере 1, со следующими исключениями: реакционная смесь для амплификации включает половину (1000 геномов) разрезанной геномной ДНК, и конечную часть термического циклинга сокращают от 70 циклов до 40 циклов. Амплификации проводят при добавлении различных количеств реагента, соответствующего изобретению, как правило, 25 нМ, 50 нМ, 100 нМ, 300 нМ, 1500 нМ и 3000 нМ. Тест включает два анализа: первый (на специфичность) - кривая плавления амплифицированного продукта и второй (на ингибирование) - кривая флуоресценции синтезируемого двунитевого продукта в режиме реального времени.
На Фиг.24-25 представляют фрагменты результатов анализа для двух реагентов, соответствующих данному изобретению, соединения 12-3DD и соединения 12-C3DD, соответственно. На каждой фигуре колонка I представляет кривые плавления для некоторых выбранных концентраций реагента, как указано. На каждой фигуре в колонке II представляют кривые флуоресценции в режиме реального времени для тех же концентраций, как указано. Относительно Фиг.24, в колонке I кривые плавления показывают, что соединение 12-3DD достигает высокой специфичности (избегая ошибочного старта) в концентрациях 100 нМ и выше (самая низкая концентрация, 100 нМ, незначительно выше показанной для реакции, описанной в Примере 1). Относительно Фиг.24, в колонке II кривые флуоресценции в режиме реального времени показывают, что ингибирование полимеразы постепенно возрастает по мере роста концентрации соединения 12-3DD выше 100 нМ до тех пор, пока реакция в основном не останавливается при концентрациях выше 1000 нМ. Ингибирование выражается в уменьшенном количестве генерируемого суммарного двунитевого продукта(ов) (включая требующийся специфический продукт и неспецифические продукты ошибочного старта), показанном как плато уровня флуоресценции и замедление роста значений СТ (19,6 для 100 нМ, 21,1 для 300 нМ и неувеличивающихся, т.е. по меньшей мере 40 для 1500 нМ и 3000 нМ). Относительно Фиг.25, в колонке I кривые плавления показывают, что соединение 12-3DD достигает высокой специфичности при концентрациях 300 нМ и выше (самая низкая концентрация, 300 нМ, такая же, как показана для реакции, описанной в Примере 1). Относительно Фиг.25, в колонке II кривые флуоресценции в режиме реального времени показывают, что ингибирование полимеразы постепенно возрастает по мере роста концентрации соединения 12-3DD выше 300 нМ (значения СТ составляют 18,8 для 100 нМ, 18,8 для 300 нМ, 21,1 для 1500 нМ и 24,5 для 3000 нМ), но что реакция не блокируется даже в концентрации 3000 нМ.
При сравнении Фиг.25 можно видеть, что действие соединения 12-C3DD в целом меньше зависит от концентрации, чем действие соединения 12-3DD в целом. Таким образом, для данной конкретной реакции амплификации задержка СТ и снижение плато флуоресценции, происходят, когда концентрация соединения 12-C3DD растет от 300 нМ до 1500 нМ, примерно так же, как они происходят, когда концентрация соединения 12-3DD растет от 100 нМ до 300 нМ.
Описан ряд вариантов осуществления изобретения. Тем не менее, следует иметь в виду, что могут быть сделаны различные модификации, не выходящие за рамки сущности и объема изобретения. В соответствии с этим другие варианты осуществления входят в объем следующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ГОМОГЕННОЙ ДЕТЕКЦИИ ПО МЕНЬШЕЙ МЕРЕ ОДНОГО ПРОДУКТА ОДНОЦЕПОЧЕЧНОЙ АМПЛИФИКАЦИИ | 2005 |
|
RU2460804C2 |
| ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ ДЛЯ ГЕНОТИПИРОВАНИЯ B.mallei МЕТОДОМ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2012 |
|
RU2478714C1 |
| ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ ДЛЯ ГЕНОТИПИРОВАНИЯ B. mallei МЕТОДОМ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2012 |
|
RU2474618C1 |
| ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ ДЛЯ ГЕНОТИПИРОВАНИЯ B. mallei МЕТОДОМ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2012 |
|
RU2474619C1 |
| ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ ДЛЯ ГЕНОТИПИРОВАНИЯ B.mallei МЕТОДОМ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2012 |
|
RU2478715C1 |
| КОМПОЗИЦИИ ДЛЯ РЕАКЦИИ ОБРАТНОЙ ТРАНСКРИПЦИИ С ГОРЯЧИМ СТАРТОМ ИЛИ ДЛЯ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ С ОБРАТНОЙ ТРАНСКРИПЦИЕЙ С ГОРЯЧИМ СТАРТОМ | 2013 |
|
RU2630999C2 |
| Способ детекции последовательностей нуклеотидов методом полимеразной цепной реакции в реальном времени с универсальным зондом | 2017 |
|
RU2675378C1 |
| ДИАГНОСТИЧЕСКАЯ СИСТЕМА ДЛЯ ВЫЯВЛЕНИЯ ДНК ПРОВИРУСОВ ЛЕЙКОЗА И ИММУНОДЕФИЦИТА КРУПНОГО РОГАТОГО СКОТА МЕТОДОМ МУЛЬТИПЛЕКСНОЙ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2015 |
|
RU2615465C2 |
| Тест-система для обнаружения генома возбудителя ротовируса типа А у сельскохозяйственных животных с помощью мультиплексной полимеразной цепной реакции с флуоресцентной детекцией в режиме реального времени | 2018 |
|
RU2694501C1 |
| НАБОР СИНТЕТИЧЕСКИХ ОЛИГОНУКЛЕОТИДНЫХ ПРАЙМЕРОВ И СПОСОБ ЕГО ПРИМЕНЕНИЯ ДЛЯ ИНДИКАЦИИ ВИРУСОВ ИММУНОДЕФИЦИТА И ЛЕЙКЕМИИ КОШЕК В КЛИНИЧЕСКОМ МАТЕРИАЛЕ МЕТОДОМ МУЛЬТИПЛЕКСНОЙ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2015 |
|
RU2586504C1 |









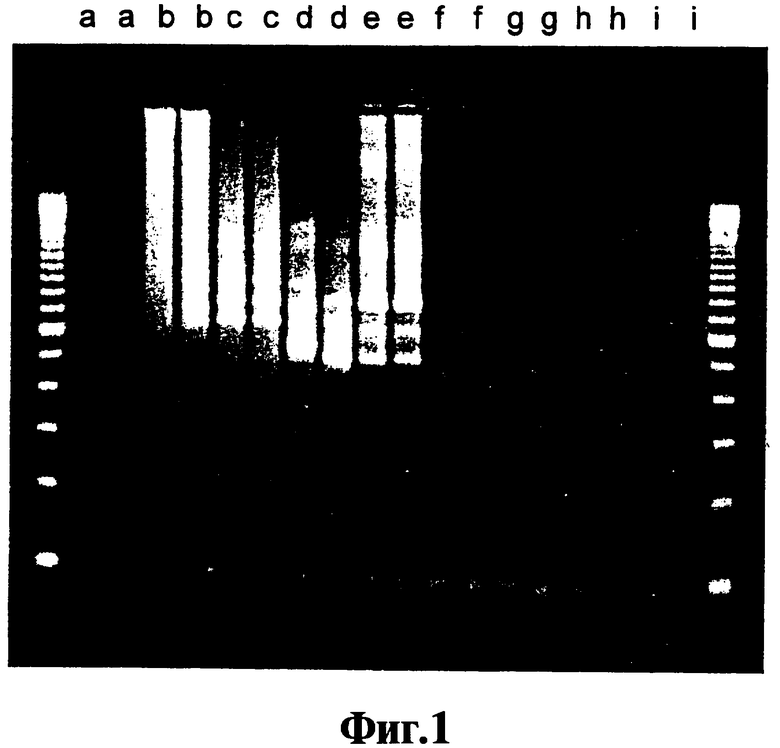
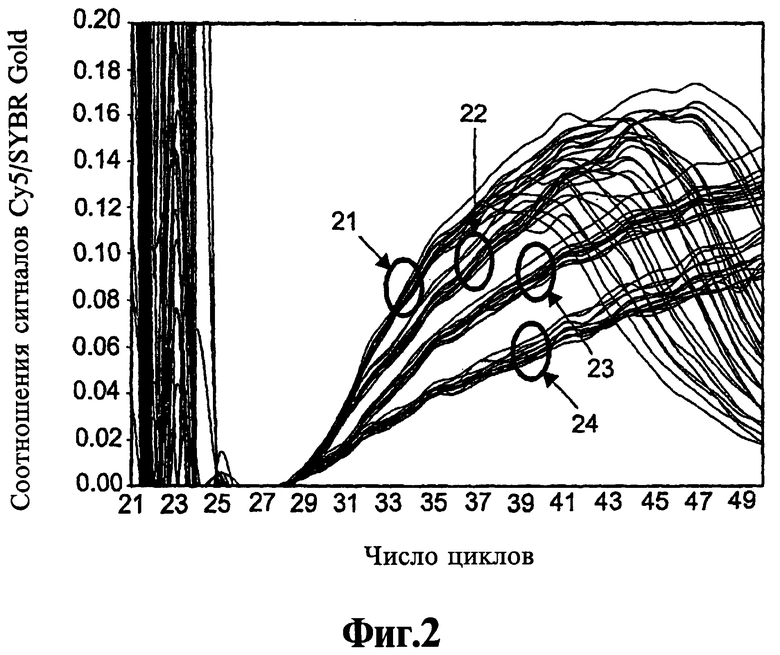
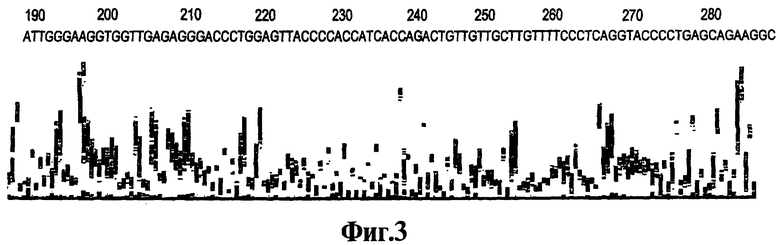
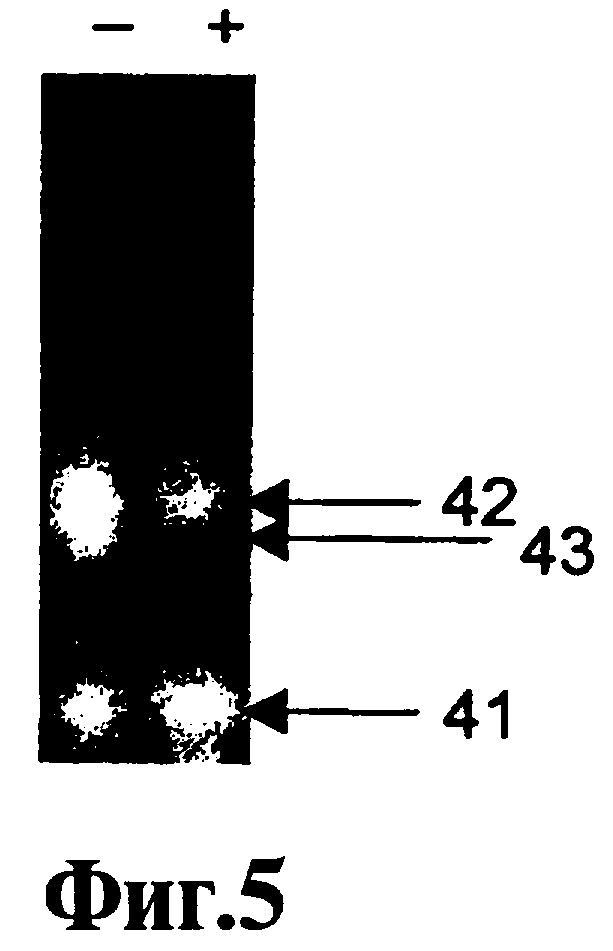
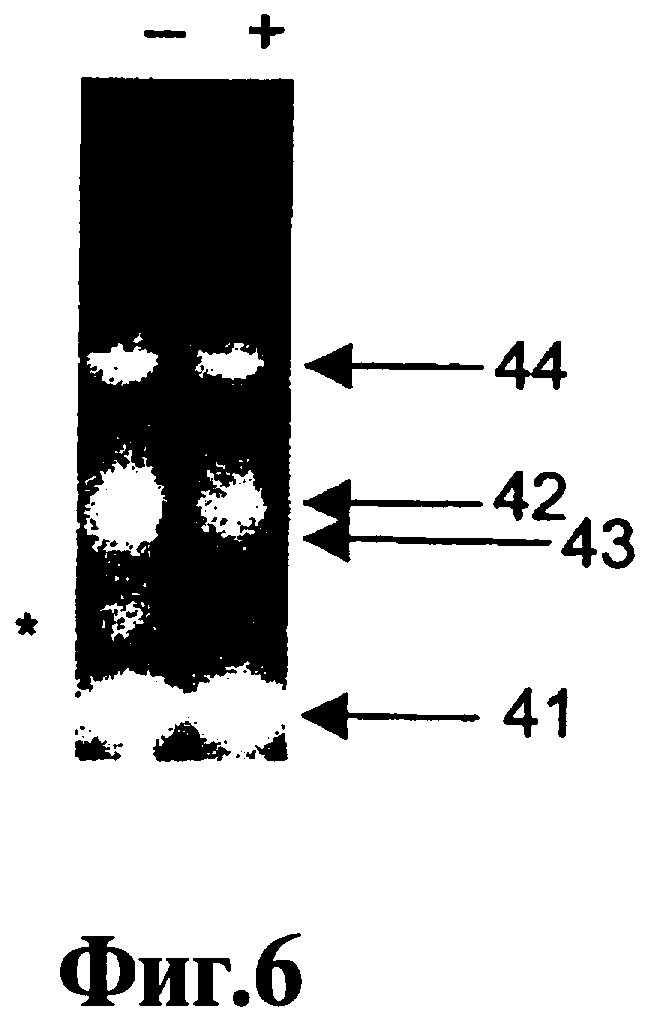

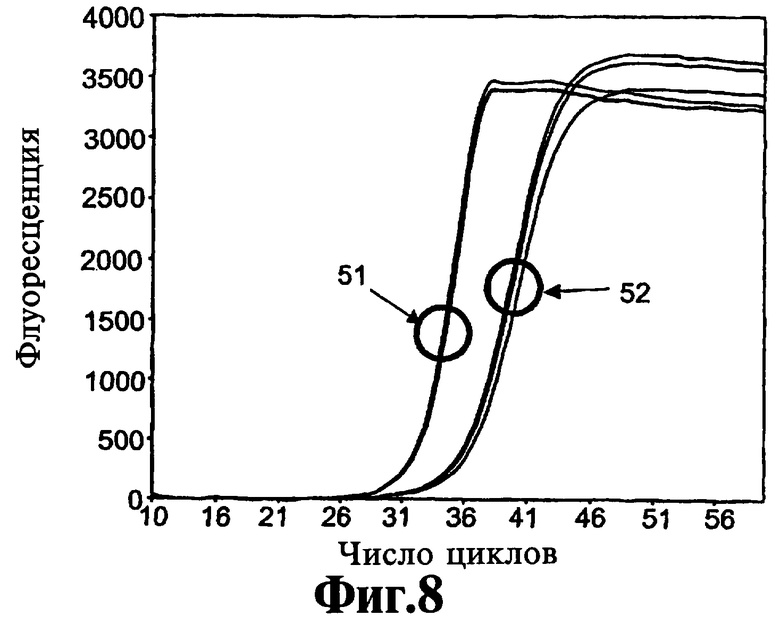
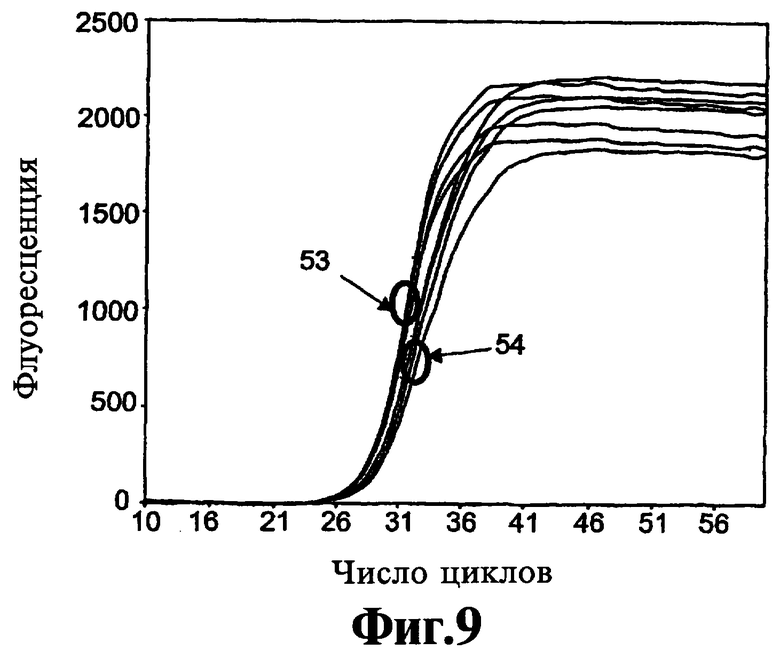
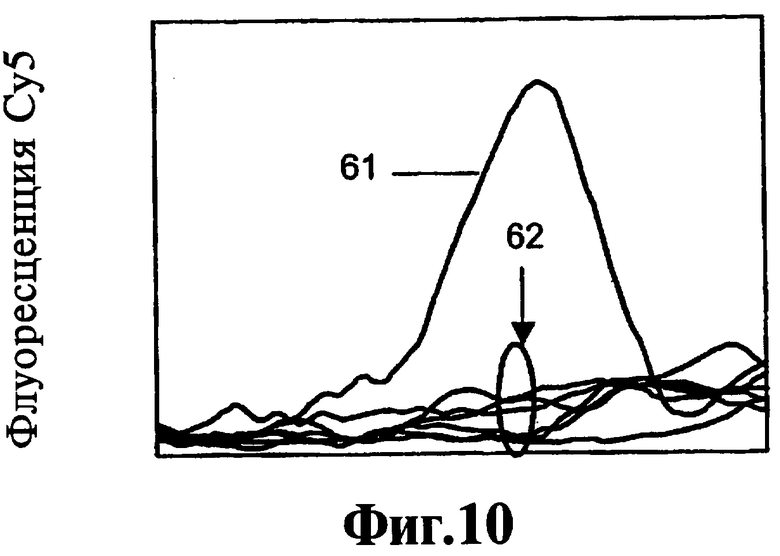
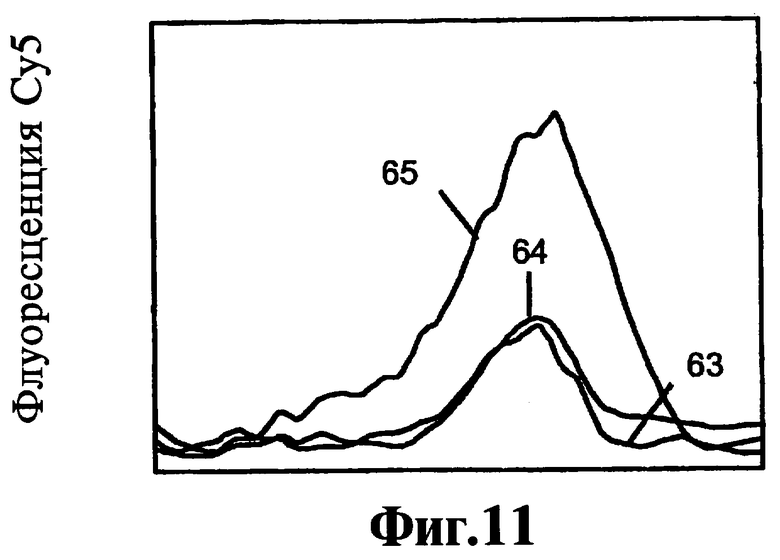
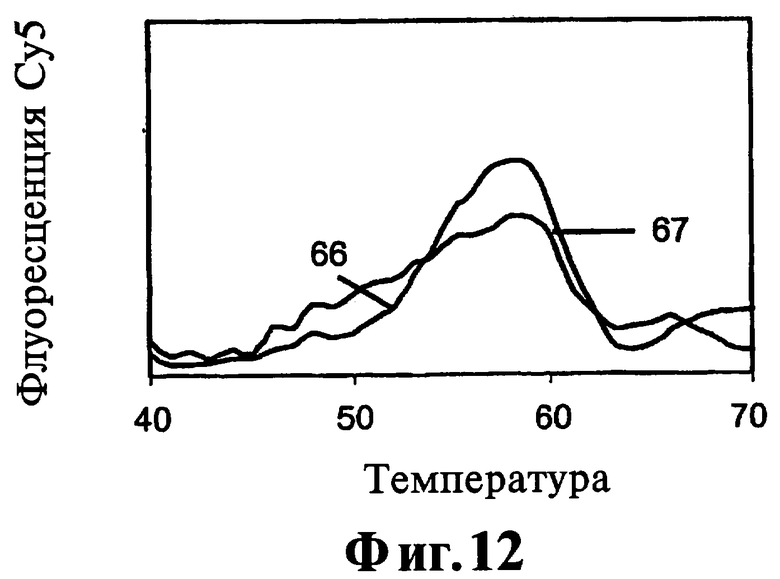
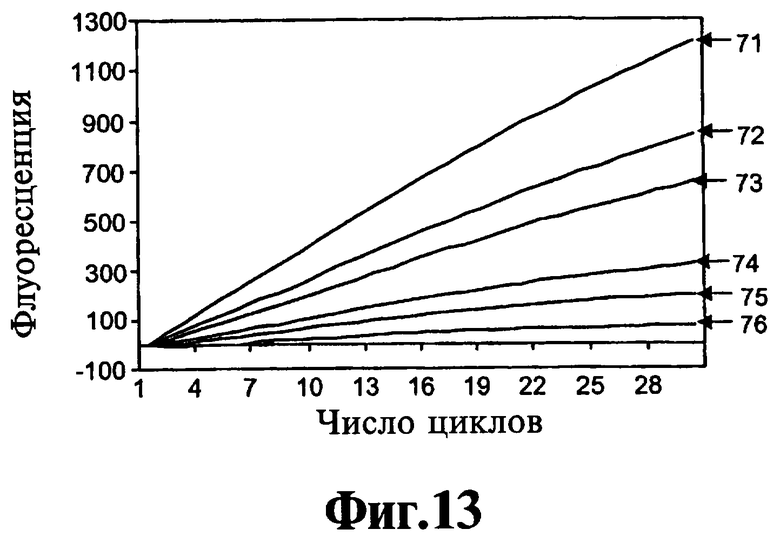
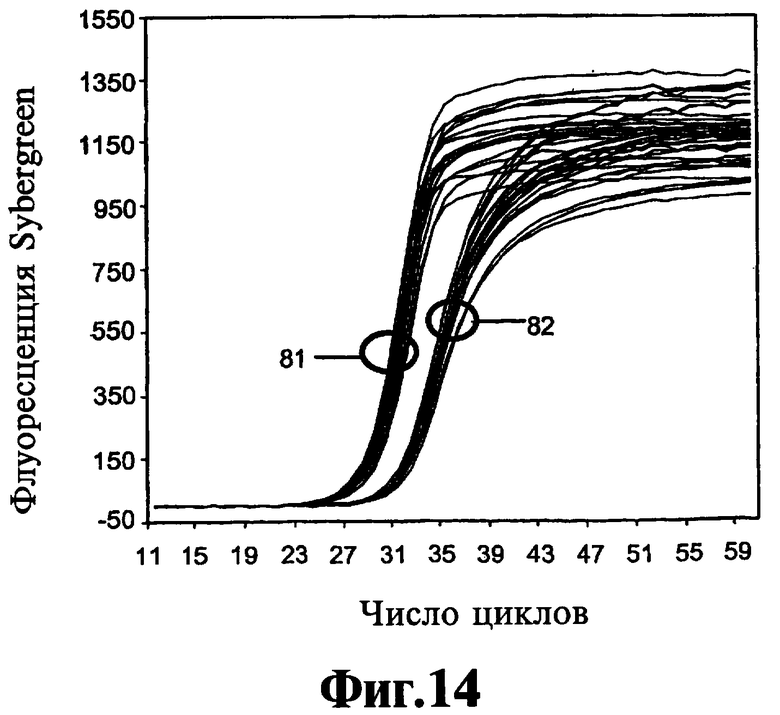
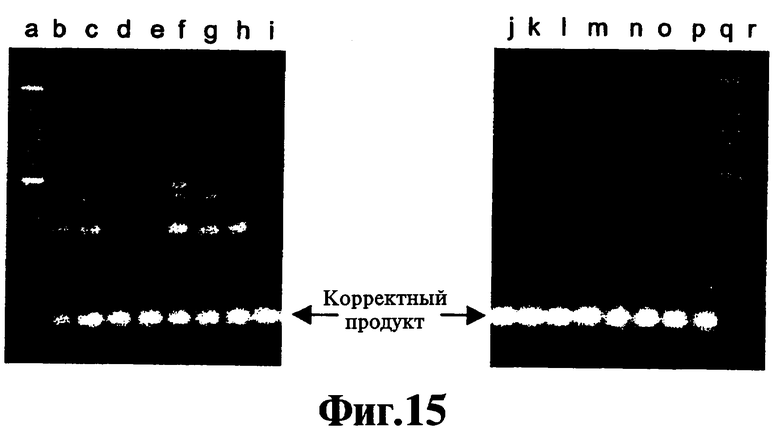

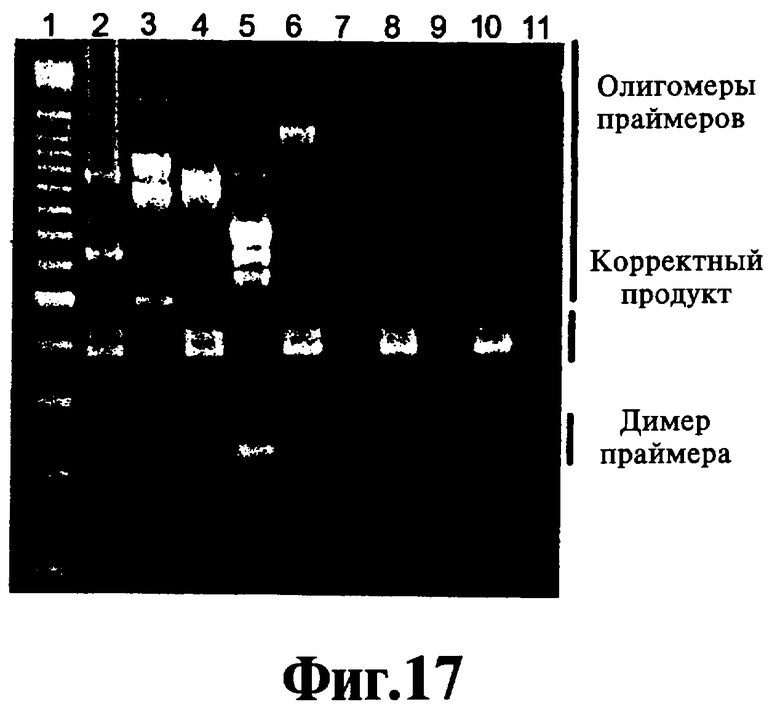
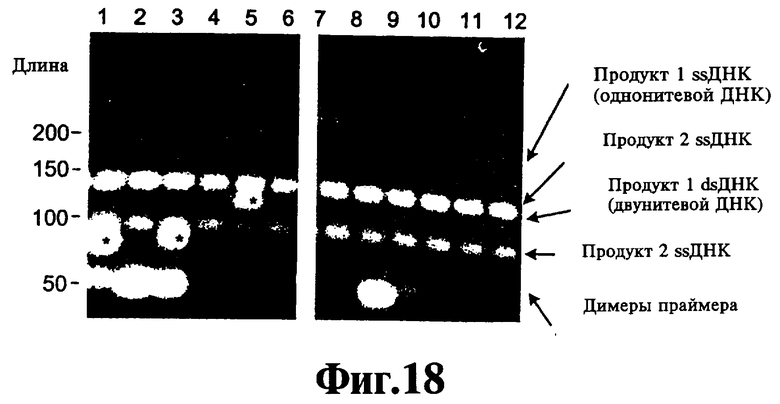
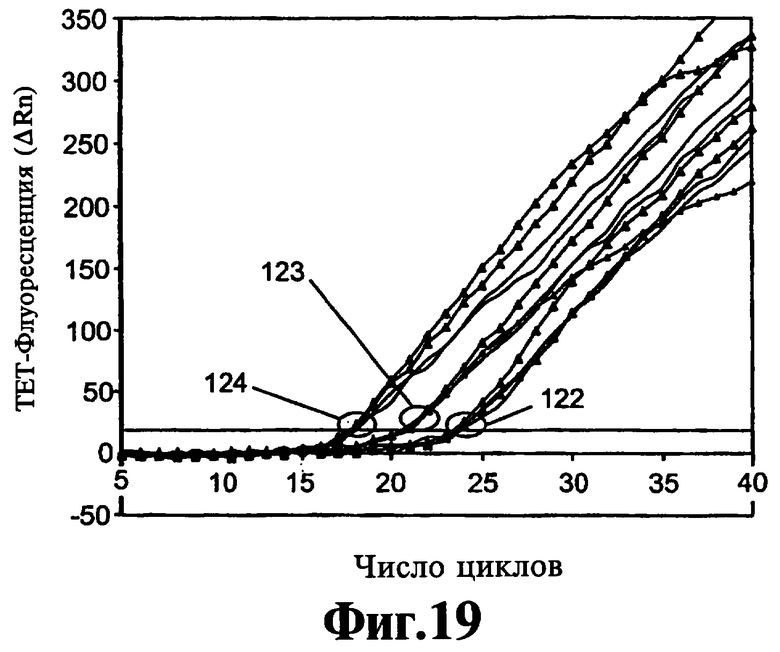
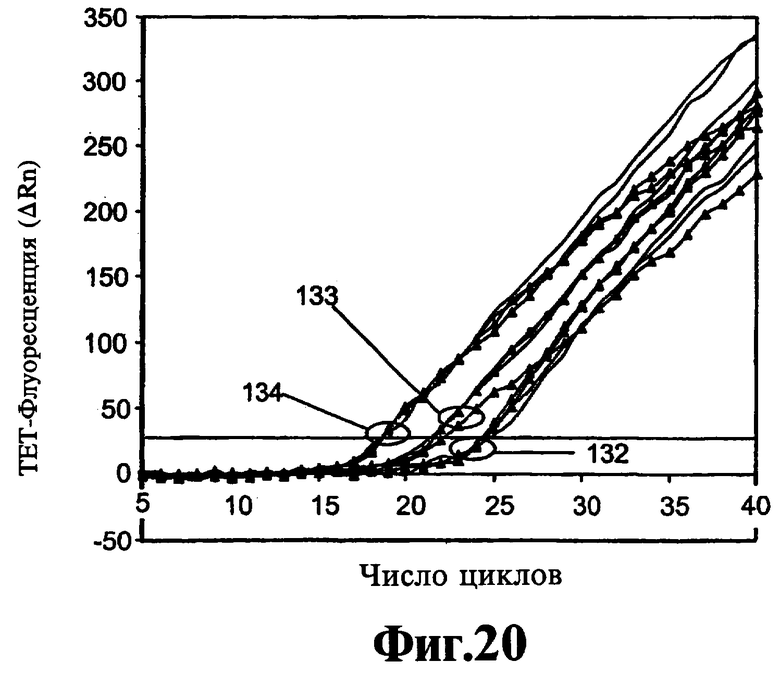
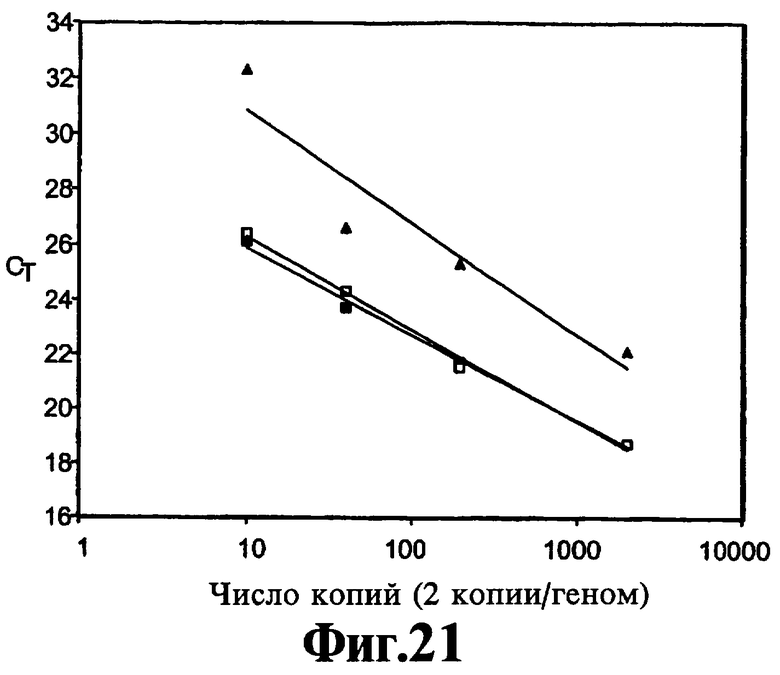
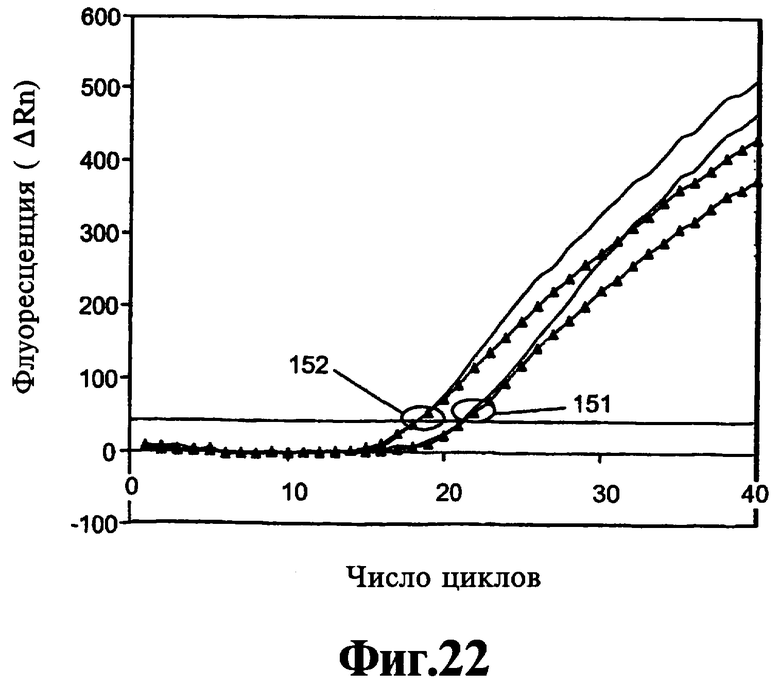
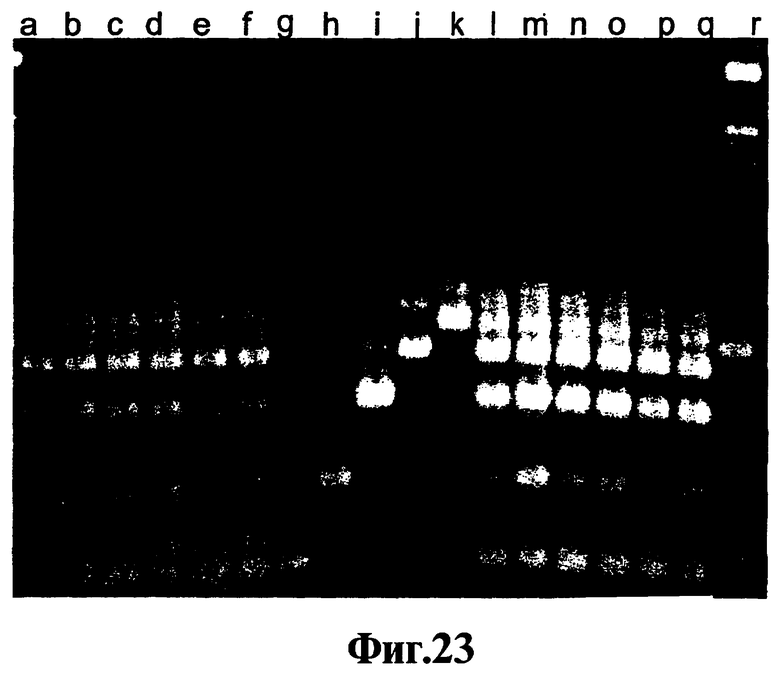
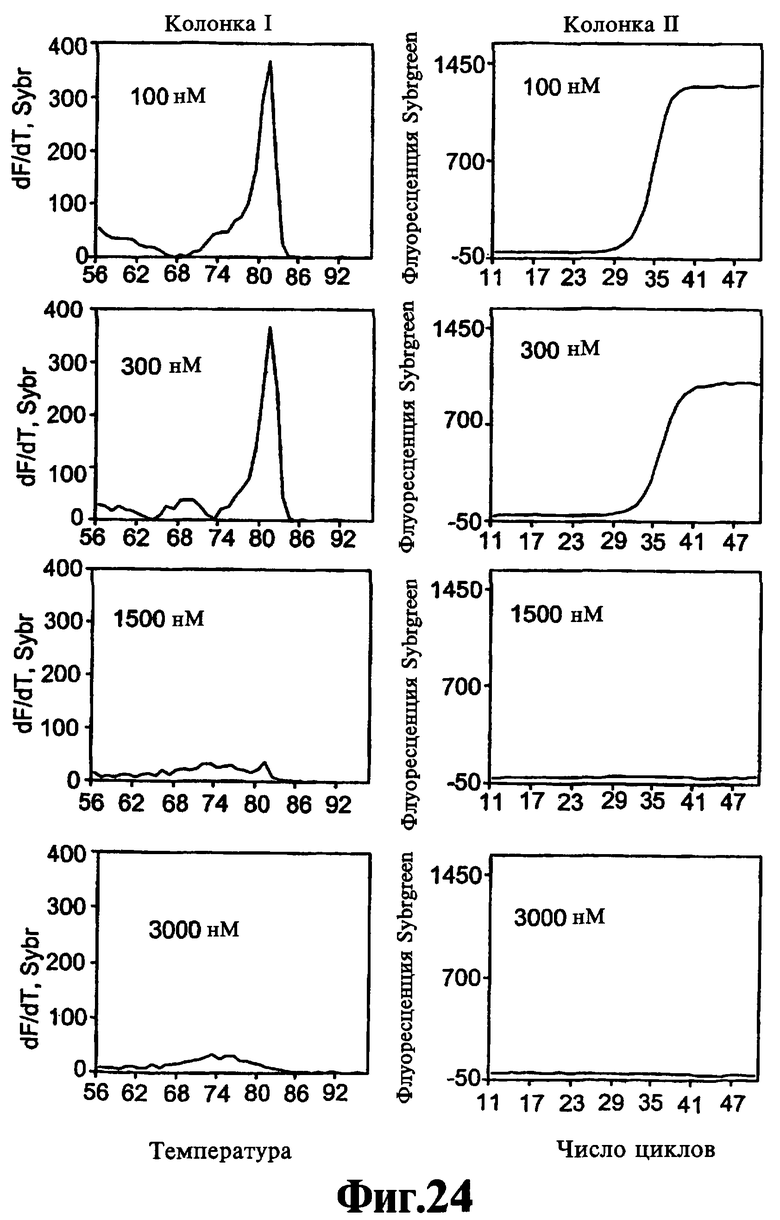
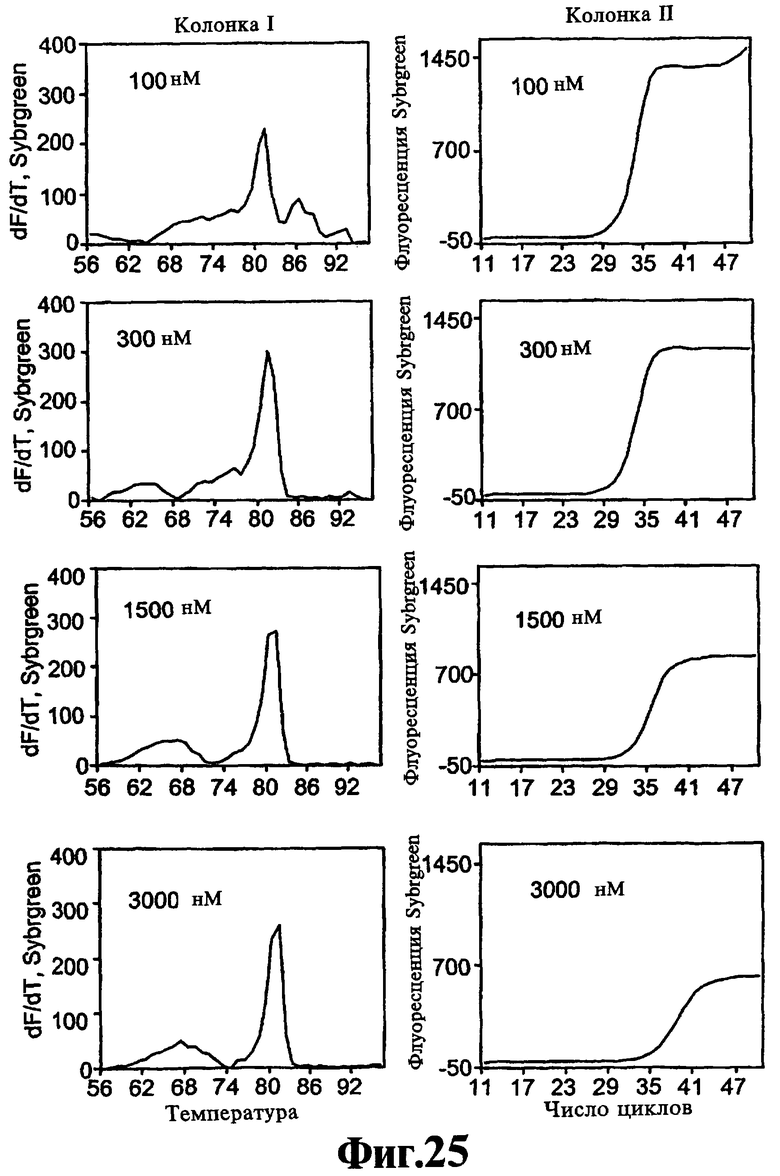
Изобретение относится к области биотехнологии, а именно к реагенту для предотвращения по меньшей мере одного ошибочного старта в амплификации полимеразной цепной реакции (ПЦР), способу предотвращения по меньшей мере одного ошибочного старта в амплификации полимеразной цепной реакции (ПЦР-амплификация) и наборам, которые включают данный реагент. Реагент используется при полимеразной цепной реакции (ПЦР), проводимой в присутствии 1,25 единиц термостабильной ДНК-полимеразы на 25 мкл реакционной смеси при концентрации не более 650 нМ. Реагент представляет собой не удлиняющийся ДНК-полимеразой олигонуклеотид, имеющий структуру ствол-петля, и не является гибридизационым зондом для указанного амплифицированного продукта ДНК. Ствол имеет длину более чем шесть нуклеотидов, стабилизирован на конце, отдаленном от петли, нефлуоресцентными веществами, выбранными из группы: гаситель флуоресценции и нуклеотидный аналог, позволяющими достичь более тесной связи между олигонуклеотидами в стволе, чем гибрид ДНК-ДНК. Температура плавления ствола (Tm) ниже 94°C. Петля представляет собой олигонуклеотидный и/или углеродный линкер. Предложенное изобретение позволяет предотвращать ошибочный старт (ненамеренный отжиг) в амплификациях и анализах с использованием полимеразной цепной реакции (ПЦР). 4 н. и 16 з.п. ф-лы, 25 ил., 3 табл., 14 пр.
1. Реагент для предотвращения по меньшей мере одного ошибочного старта в амплификации полимеразной цепной реакции (ПЦР), проводимой в присутствии 1,25 единиц термостабильной ДНК-полимеразы на 25 мкл реакционной смеси при концентрации не более 650 нМ для получения по меньшей мере одного амплифицированного продукта ДНК, характеризующийся тем, что указанный реагент представляет собой неудлиняющийся ДНК-полимеразой олигонуклеотид, который имеет структуру ствол-петля и не является гибридизационым зондом для указанного по меньшей мере одного амплифицированного продукта ДНК, имеет ствол длиной более чем шесть нуклеотидов, стабилизирован на конце, отдаленном от петли, нефлуоресцентнами веществами, выбранными из группы: гаситель флуоресценции и нуклеотидный аналог, позволяющими достичь более тесной связи между олигонуклеотидами в стволе, чем гибрид ДНК-ДНК, и который имеет рассчитанную температуру плавления ствола (Tm) ниже 94°C, причем указанная петля представляет собой петлю из олигонуклеотида и/или углеродного линкера.
2. Реагент по п.1, в котором петля представляет собой олигонуклеотид, включающий по меньшей мере три нуклеотида.
3. Реагент по п.1, в котором петля представляет собой ненуклеотидпый химический линкер.
4. Реагент по п.1, в котором ствол стабилизирован на конце посредством химических групп, которые ковалентно присоединены к 3- и 5-нуклеотидам ствола.
5. Реагент по п.4, в котором химические группы обладают свойствами, отличными от флуоресцентных.
6. Реагент по п.1, в котором часть ствола, являющаяся двупитевой при температурах ниже его Tm, имеет длину 9-12 пар оснований.
7. Реагент по пп.1-6, в котором рассчитанная температура плавления ствола находится в интервале 72-85°C.
8. Реагент по пп.1-6, в котором рассчитанная температура плавления ствола лежит в интервале 50-71°C.
9. Способ предотвращения по меньшей мере одного ошибочного старта в амплификации полимеразной цепной реакции (ПЦР-амплификация), проводимой для получения по меньшей мере одного амплифицированного продукта ДНК из реакционной смеси для амплификации, которая включает дезоксинуклеотидфосфаты (дНТФ) и термостабильную ДНК-полимеразу, характеризующийся в добавлении в данную реакционную смесь по меньшей мере одного реагента по любому из пп.1-8.
10. Способ по п.9, в котором реагент имеет олигонуклеотидную петлю, а концентрация указанного реагента не превышает 300 нМ.
11. Способ по п.9 в котором реагент имеет петлю, которая представляет собой ненуклеотидный химический линкер, а реагент добавляют к реакционной смеси в концентрации, не превышающей 1000 нМ.
12. Способ по п.9, в котором амплификация представляет собой анализ на основе линейной после экспоненциальной ПЦР (ЛПЭ-ПЦР), включающий низкотемпературную стадию детекции, проводимую при температуре ниже, чем указанная Tm.
13. Способ по п.12, в котором амплификация представляет собой анализ на основе симметричной ПЦР.
14. Способ по пп.9-13, в котором после амплификации следует детекция конечного результата.
15. Набор реагентов для осуществления амплификации посредством полимеразной цепной реакции (ПЦР), включающий термостабильную ДНК-полимеразу, по меньшей мере одну пару ПЦР-праймеров, деоксинуклеотид фосфатов и по меньшей мере один реагент по любому из пп.1-8.
16. Набор по п.15, дополнительно включающий реагенты для выделения нуклеиновой кислоты.
17. Набор олигонуклеотидов для осуществления амплификации посредством полимеразной цепной реакции (ПЦР), включающая по меньшей мере одну пару ПЦР-праймеров и по меньшей мере один реагент по любому из пп.1-8.
18. Набор по п.17, дополнительно включающий по меньшей мере один зонд с флуоресцентной меткой, который гибридизуется с амплифицированным продуктом, определяемым указанной по меньшей мере одной парой ПЦР-праймеров.
19. Набор по п.18, в котором по меньшей мере одна пара праймеров включает ограничивающий праймер и избыточный праймер в соотношении, подходящем для проведения амплификации посредством линейной после экспоненциальной ПЦР (ЛПЭ-ПЦР).
20. Набор по пп.17-19, в котором указанный амплифицированный продукт является однонитевым.
| Молотильный аппарат для клещевины | 1954 |
|
SU102559A1 |
| US 6183967 B1, 06.02.2001 | |||
| SANCHEZ J.А | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| US 5866336, 02.02.1999 | |||
| US | |||

