ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к генам, вовлеченным в споруляцию микобактерий, в областях техники иммунологии и медицины.
УРОВЕНЬ ТЕХНИКИ
Род Mycobacterium включает высокоэффективные патогены, например, Mycobacterium tuberculosis и Mycobacterium leprae, возбудители туберкулеза и лепры, соответственно, M. tuberculosis (Mtb) является одним из наиболее эффективных патогенов человека, и оценки позволяют предполагать, что одна треть популяции человека является носителем этих бактерий. Другими видами Mycobacterium, заслуживающими упоминания, являются Mycobacterium marinum (Mm), Mycobacterium avium и Mycobacterium ulcerans.
В настоящее время приблизительно у восьми миллионов человек в год развивается активное заболевание и два миллиона умирают. Патогенные микобактерии обладают способностью персистировать долгое время в хозяине без вызывания каких-либо симптомов. Эта латентность представляет собой дополнительную скрытую угрозу здоровью человека, но еще не ясно понятую. Микобактерии могут колонизировать своих хозяев без проявления каких-либо неблагоприятных признаков. Например, миллиарды людей по всему миру являются инфицированными M. tuberculosis, но никогда об этом не узнают, поскольку у них не развивается симптомов.
Общеизвестно, что микобактериальные инфекции трудно лечить. Организмы являются жизнестойкими из-за их клеточной стенки, которая не является ни истинно грамотрицательной, ни истинно грамположительной, и, что уникально для семейства, они являются от природы устойчивыми к ряду антибиотиков, действующих посредством разрушения клеточных стенок, таких как пенициллин. Большинство микобактерий являются чувствительными к антибиотикам, например, кларитромицину, изониазиду и рифамицину, но известно, что существуют устойчивые к антибиотикам штаммы, и возникновение штаммов с множественной устойчивостью представляет собой увеличивающуюся проблему.
Ослабленный штамм Mycobacterium bovis в настоящее время является наиболее широко используемой и наиболее безопасной ослабленной вакциной для человека, однако, возникли разногласия относительно его эффективности.
Следовательно, существует необходимость в улучшенных способах и лекарственных средствах для профилактики и лечения заболеваний, вызванных микобактериями.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящее время впервые показано, что микобактерии обладают способностью образовывать споры. Это противоречит описаниям микобактерий в учебниках как не спорулирующих, грамположительных видов. Это вносит новые перспективы в образ жизни патогенных микобактерий и их потенциал к размножению, также как к персистентности, микобактериальных инфекций. Следовательно, настоящее изобретение относится к способам и соединениям, применимым для лечения и/или профилактики заболеваний, вызванных микобактериями, таких как туберкулез. В частности, изобретение относится к лечению и/или профилактике таких заболеваний, при которых микобактерии спорулируют или могут спорулировать. Такие способы и соединения могут являться основанными на активной или пассивной иммунизации, где активную иммунизацию проводят введением вакцины, а пассивную иммунизацию проводят введением антитела.
В одном аспекте изобретение относится к способам профилактики или лечения заболевания, вызванного микобактериями. Такие заболевания представляют собой, например, туберкулез, лепру и заболевания, вызванные нетуберкулезными микобактериями (NTM). Включены также болезнь Крона и заболевание скота болезнь Джона. Хорошо известно, что индивидуумы с нарушенной иммунной системой, такие как пациенты со СПИД, и пациенты, которым вводят иммуносупрессирующие лекарственные средства, являются чувствительными к микобактериальным инфекциям. Для таких пациентов может являться благоприятным лечение согласно этому изобретению.
Изобретение относится также к соединениям, которые могут являться применимыми для лечения вышеупомянутых заболеваний, либо в форме вакцины, либо в форме антител. Предусмотрены также связывающие молекулы, отличные от антител.
Способы лечения таких заболеваний вызывают индукцию у пациента иммунного ответа против пептидного компонента из одного из продуктов генов, вовлеченных в споруляцию. Такая индукция может являться активной посредством введения иммуногена (такого как вакцина), или пассивной посредством введения антитела или активного фрагмента или производного антитела. В некоторых способах у пациента отсутствуют симптомы. В некоторых способах лечение является профилактическим.
Для лечения пациентов, страдающих заболеванием, вызванным микобактериями, один из режимов лечения включает введение пациенту дозы пептида, полученного из белкового компонента оболочки споры, для индукции иммунного ответа. В некоторых способах пептид оболочки споры вводят с адъювантом, усиливающим иммунный ответ на пептид оболочки споры. Примеры адъювантов включают квасцы и MPL. Доза пептида оболочки споры, вводимого пациенту, как правило, составляет по меньшей мере 1 или 10 мкг при введении с адъювантом, и по меньшей мере 50 мкг при введении без адъюванта. В некоторых способах доза составляет по меньшей мере 100 мкг.
В некоторых способах пептид оболочки споры представляет собой фрагмент продуктов генов, показанных в таблицах 1-4.
В некоторых способах лекарственное средство представляет собой эффективную дозу нуклеиновой кислоты, кодирующей пептид оболочки споры или его активный фрагмент или производное. Нуклеиновую кислоту, кодирующую оболочку споры или ее фрагмент, экспрессируют у пациента для получения пептида оболочки споры или его активного фрагмента, индуцирующего иммунный ответ. В некоторых таких способах нуклеиновую кислоту вводят через кожу, необязательно посредством пластыря. В некоторых способах лекарственное средство идентифицируют скринингом библиотеки соединений для идентификации соединения, связывающегося с антителами против пептида оболочки споры, и введением соединения пациенту для индукции иммунного ответа.
В некоторых способах иммунный ответ включает T-клетки, связывающие пептид оболочки споры в комплексе с MHCI или MHCII на CD8 или CD4 клетках. В других способах иммунный ответ индуцируют введением антитела против пептида оболочки споры пациенту. В некоторых способах иммунный ответ индуцируют взятием T-клеток от пациента, приведением T-клеток в контакт с пептидом оболочки споры в условиях, в которых T-клетки являются примированными, и замещением T-клеток у пациента.
Изобретение относится также к молекулам, отличным от антител, способным связывать пептид оболочки споры. Вместе молекулы, например антитела и другие молекулы, способные связывать пептид оболочки споры, называют «связывающие молекулы». Связывающие молекулы по изобретению представляют собой предпочтительно антитела, как определено в настоящем документе. Термин «антитело» относится к интактному антителу, или его связывающему фрагменту. Антитело может включать полную молекулу антитела (включая поликлональные, моноклональные, химерные, гуманизированные, или человеческие варианты, имеющие полноразмерные тяжелые и/или легкие цепи) или включать его антигенсвязывающий фрагмент. Фрагменты антител включают F(ab')2, Fab, Fab', Fv, Fc и Fd фрагменты, и могут быть включены в однодоменные антитела, одноцепочечные антитела, макси-антитела, мини-антитела, интраантитела, диатела, триатела, тетратела, v-NAR и бис-scFv (смотри, например, Hollinger and Hudson, 2005, Nature Biotechnology, 23, 9, 1126-1136). Полипептиды антител описаны также в патенте США № 6703199, включая монотела полипептида фибронектина. Другие полипептиды антител, которые представляют собой одноцепочечные полипептиды, описаны в публикации патента США 2005/0238646.
Антигенсвязывающие фрагменты, происходящие из антитела, можно получать, например, протеолитическим гидролизом антитела, например, расщеплением пепсином или папаином полноразмерных антител общепринятыми способами. В качестве примера, фрагменты антитела можно получать ферментативным расщеплением антител пепсином для получения 5S-фрагмента, названного F(ab')2. Этот фрагмент можно далее расщеплять с использованием восстанавливающего тиолового средства для получения моновалентных фрагментов 3,5S Fab'. Необязательно, реакцию расщепления можно проводить с использованием блокирующей группы для сульфгидрильных групп, образующихся в результате расщепления дисульфидных связей. В качестве альтернативы, ферментативным расщеплением с использованием папаина напрямую получают два моновалентных Fab фрагмента и Fc фрагмент. Эти способы описаны, например, в Goldenberg, патент США № 4331647, Nisonoff et al., Arch. Biochem. Biophys. 89:230, 1960; Porter, Biochem. J. 73:119, 1959; Edelman et al., in Methods in Enzymology 1:422 (Academic Press 1967); и в Andrews, S.M. and Titus, J.A. in Current Protocols in Immunology (Coligan J.E., et al., eds), John Wiley & Sons, New York (2003), страницы 2.8.1-2.8.10 и 2.10A.1-2.10A.5. Другие способы расщепления антител, такие как разделение тяжелых цепей для получения моновалентных фрагментов легкая-тяжелая цепь (Fd), дополнительное расщепление фрагментов, или другие ферментативные, химические или генетические способы также можно использовать, при условии, что фрагменты связывают антиген, узнаваемый интактным антителом.
Фрагмент антитела может представлять собой также синтетический или полученный способами генной инженерии белок. Например, фрагменты антитела включают выделенные фрагменты, состоящие из вариабельной области легкой цепи, «Fv» фрагменты, состоящие из вариабельных областей тяжелой и легких цепей, рекомбинантные одноцепочечные полипептидные молекулы, в которых легкие и тяжелые вариабельные области соединены пептидным линкером (scFv-белки).
Другой формой фрагмента антитела является пептид, содержащий одну или несколько определяющих комплементарность областей (CDR) антитела. CDR (называемые также «минимальные узнающие единицы» или «гипервариабельная область») можно получать конструированием полинуклеотидов, кодирующих интересующую CDR. Такие полинуклеотиды можно получать, например, с использованием полимеразной цепной реакции для синтеза вариабельной области с использованием мРНК антитело-продуцирующих клеток в качестве матрицы (смотри, например, Larrick et al., Methods: A Companion to Methods in Enzymology 2:106, 1991; Courtenay-Luck, «Genetic Manipulation of Monoclonal Antibodies», in Monoclonal Antibodies: Production, Engineering and Clinical Application, Ritter et al. (eds.), page 166 (Cambridge University Press 1995); и Ward et al., «Genetic Manipulation and Expression of Antibodies», in Monoclonal Antibodies: Principles and Applications, Birch et al., (eds.), page 137 (Wiley-Liss, Inc. 1995)).
Таким образом, в одном варианте осуществления связывающая молекула содержит по меньшей мере одну CDR, как описано в настоящем документе. Связывающая молекула может содержать по меньшей мере две, три, четыре, пять или шесть CDR, как описано в настоящем документе. Связывающая молекула дополнительно может содержать по меньшей мере один домен вариабельной области антитела, описанного в настоящем документе. Домен вариабельной области может обладать любым размером или аминокислотным составом и может, как правило, включать по меньшей мере одну последовательность CDR, ответственную за связывание с формирующим споры пептидом, например CDR-H1, CDR-H2, CDR-H3 и/или CDR легкой цепи, конкретно описанные в настоящем документе и являющиеся соседними или находящимися в одной рамке считывания с одной или несколькими каркасными последовательностями. В общих чертах, домен вариабельной (V) области может представлять собой любое подходящее расположение вариабельных доменов тяжелой (VH) и/или легкой (VL) цепи иммуноглобулина. Таким образом, например, домен V-области может являться мономерным и представлять собой домен VH или VL, способный независимо связывать формирующий споры пептид с аффинностью, по меньшей мере равной 1×10-7M или меньшей, как описано ниже. Альтернативно домен V-области может являться димерным и содержать димеры VH-VH, VH-VL или VL-VL. Димер V-области содержит по меньшей мере одну VH и по меньшей мере одну VL цепь, которые могут являться нековалентно связанными (далее в настоящем документе обозначен Fv). Если желательно, цепи можно связывать ковалентно напрямую, например, посредством дисульфидной связи между двумя вариабельными доменами, или посредством линкера, например, пептидного линкера, для получения одноцепочечного Fv (scFv).
Домен вариабельной области может представлять собой любой природный вариабельный домен или его сконструированный вариант. Под сконструированным вариантом понимают домен вариабельной области, полученный с использованием способов конструирования рекомбинантной ДНК. Такие сконструированные варианты включают варианты, полученные, например, из вариабельной области специфического антитела посредством вставок, делеций или изменений в аминокислотных последовательностях специфического антитела. Конкретные примеры включают сконструированные домены вариабельной области, содержащие по меньшей мере одну CDR и необязательно одну или несколько каркасных аминокислот из первого антитела и остаток домена вариабельной области из второго антитела.
Другие связывающие молекулы, которые можно использовать, включают, например, молекулы, описанные в WO03/080653, камелиды, аффитела и диатела.
Домен вариабельной области можно ковалентно присоединять к C-концевой аминокислоте по меньшей мере одного другого домена антитела или его фрагмента. Таким образом, например, VH домен, который присутствует в домене вариабельной области, может являться связанным с доменом иммуноглобулина CH1 или его фрагментом. Подобным образом, VL домен может являться связанным с доменом CK или его фрагментом. Следовательно, например, антитело может представлять собой Fab-фрагмент, где антигенсвязывающий домен содержит связанные VH и VL домены, ковалентно присоединенные по их C-концам к CH1 и CK домену, соответственно. CH1 домен можно расширять дополнительными аминокислотами, например, для предоставления шарнирной области или части домена шарнирной области, как обнаружено в Fab'-фрагменте, или для предоставления дальнейших доменов, таких как домены CH2 и CH3 антитела.
Как описано в настоящем документе, связывающие молекулы содержат по меньшей мере одну из этих CDR. Например, одну или несколько CDR можно включать в известные каркасные области антитела (IgG1, IgG2 и т.д.) или конъюгировать с подходящим носителем для увеличения времени ее полужизни. Пригодные носители включают в качестве неограничивающих примеров Fc, полиэтиленгликоль (PEG), альбумин, трансферрин и т.п. Эти и другие пригодные носители известны в данной области. Такие конъюгированные пептиды CDR могут существовать в мономерной, димерной, тетрамерной или другой форме. В одном варианте осуществления один или несколько водорастворимых полимеров можно присоединять в одном или нескольких специфических положениях, например, на N-конце, связывающей молекулы.
В конкретных предпочтительных вариантах осуществления связывающая молекула содержит одно или несколько присоединений водорастворимого полимера, включая в качестве неограничивающих примеров полиэтиленгликоль, полиоксиэтиленгликоль или полипропиленгликоль. Смотри, например, патенты США №№ 4640835, 4496689, 4301144, 4670417, 4791192 и 4179337. В конкретных вариантах осуществления производное связывающей молекулы включает одно или несколько из монометоксиполиэтиленгликоля, декстрана, целлюлозы или других полимеров на основе углеводов, поли-(N-винилпирролидон)-полиэтиленгликоля, гомополимеров пропиленгликоля, сополимера полипропиленоксида/этиленоксида, полиоксиэтилированных полиолов (например, глицерина) и поливинилового спирта, также как смеси таких полимеров. В конкретных вариантах осуществления один или несколько водорастворимых полимеров случайным образом присоединяют к одной или нескольким боковым цепям. В конкретных вариантах осуществления PEG может действовать для улучшения терапевтической способности связывающей молекулы, такой как антитело. Конкретные такие способы обсуждают, например, в патенте США № 6133426, содержание которого таким образом приведено в качестве ссылки для любых целей.
Следует понимать, что связывающая молекула по настоящему изобретению может обладать по меньшей мере одной аминокислотной заменой, при условии, что связывающая молекула сохраняет специфичность связывания. Таким образом, модификации структур связывающих молекул включены в объем изобретения. Они могут включать аминокислотные замены, которые могут являться консервативными или неконсервативными, которые не нарушают способности пептида оболочки споры связываться со связывающей молекулой. Консервативные аминокислотные замены могут включать неприродные аминокислотные остатки, которые, как правило, включают химическим синтезом пептида, а не синтезом в биологических системах. Они включают пептидомиметики и другие обращенные или инвертированные формы аминокислотных групп. Консервативная аминокислотная замена может также включать замену природного аминокислотного остатка на нормативный остаток, так что присутствует небольшой эффект или отсутствует эффект на полярность или заряд аминокислотного остатка в этом положении.
Связывающие молекулы по этому изобретению можно использовать также в качестве диагностического инструмента для детекции присутствия, или отсутствия, микобактерий в форме спор у животного, например, у людей или у скота. Детекцию предпочтительно проводят в образце от указанного субъекта - животного или человека, который, как подозревают, содержит споры, происходящие из микобактерий, таком как жидкости организма, например, кровь, сыворотка, жидкость бронхоальвеолярного лаважа, слюна, моча, фекалии, слезы и т.п. Предусмотрена также недиагностическая детекция спор, происходящих из микобактерий, в пищевых продуктах, таких как молоко, сыр, масло, овощи и мясо, или в продуктах, предназначенных для кормления животных. Связывающие вещества по изобретению можно также использовать для детекции микобактерий в образцах из окружающей среды для окружающей среды, подозреваемой на контаминацию спорами, происходящими из микобактерий, например, рыбных ферм и птицеферм. Способы такой детекции являются хорошо известными и включают, например, анализы ELISA и другие иммунологические анализы.
Лекарственное средство или связывающую молекулу можно, как правило, вводить перорально, интраназально, внутрикожно, подкожно, внутримышечно, местно или внутривенно. В некоторых способах пациента мониторируют после введения для оценки иммунного ответа. Если мониторирование указывает на уменьшение иммунного ответа с течением времени, пациенту можно вводить одну или несколько дополнительных доз средства.
В другом аспекте изобретение относится к фармацевтическим композициям, содержащим пептид оболочки споры и наполнитель, пригодный для перорального и других способов введения. Изобретение относится также к фармацевтическим композициям, содержащим средство, эффективное для индукции иммуногенного ответа против пептида оболочки споры у животного, и фармацевтически приемлемый адъювант. В некоторых таких композициях средство представляет собой пептид оболочки споры или его активный фрагмент. В некоторых композициях адъювант содержит квасцы.
В некоторых композициях адъювант содержит эмульсию масло-в-воде. В некоторых композициях пептид оболочки споры или его активный фрагмент является компонентом сополимера полилактида и полигликолида (PLPG) или другой частицы. Кроме того, изобретение относится к композициям, содержащим пептид оболочки споры или активный фрагмент, связанный с молекулой конъюгата, которая способствует доставке пептида оболочки споры в кровоток пациента и/или стимулирует иммунный ответ против пептида оболочки споры. Например, конъюгат может служить для стимуляции иммунного ответа против пептида оболочки споры. В некоторых композициях конъюгат представляет собой холерный токсин. В некоторых композициях конъюгат представляет собой иммуноглобулин. В некоторых композициях конъюгат представляет собой ослабленный дифтерийный токсин CRM 197 (Gupta, Vaccine 15, 1341-3 (1997)).
Изобретение относится также к фармацевтическим композициям, содержащим средство, эффективное для индукции иммуногенного ответа против пептида оболочки споры у пациента, при условии, что композиция является свободной от полного адъюванта Фрейнда. Изобретение относится также к композициям, содержащим вирусный вектор, кодирующий пептид оболочки споры или его активный фрагмент, эффективный для индукции иммунного ответа против пептида оболочки споры. Пригодные вирусные векторы включают вирус герпеса, аденовирус, аденоассоциированный вирус, ретровирус, вирус Синдбис, вирус леса Семлики, вирус осповакцины или вирус оспы птиц.
Кроме того, изобретение относится к способам профилактики или лечения заболевания, вызванного микобактериями. В таких способах эффективную дозу пептида оболочки споры можно вводить пациенту. Кроме того, изобретение относится к применению пептида оболочки споры или антитела против него, для изготовления лекарственного средства для профилактики или лечения заболевания, вызванного микобактериями.
В другом аспекте изобретение относится к способам оценки эффективности способа лечения, описанного выше, для пациента. В этих способах фоновое количество антитела, специфического для пептида оболочки споры, определяют в образце ткани пациента до лечения средством. Количество антитела, специфического для пептида оболочки споры, в образце ткани от пациента после лечения средством сравнивают с фоновым количеством специфического для пептида оболочки споры антитела. Количество специфического для пептида оболочки споры антитела, измеренное после лечения, которое значительно превышает фоновое количество специфического для пептида оболочки споры антитела, указывает на положительный исход лечения.
В других способах оценки эффективности способа лечения для пациента фоновое количество антитела, специфического для пептида оболочки споры, определяют в образце ткани пациента до лечения средством. Количество антитела, специфического для пептида оболочки споры, в образце ткани от субъекта после лечения средством сравнивают с фоновым количеством антитела, специфического для пептида оболочки споры. Уменьшение или отсутствие значительной разницы между количеством специфического для пептида оболочки споры антитела, измеренным после лечения, по сравнению с фоновым количеством антитела, специфического для пептида оболочки споры, указывает на отрицательный исход лечения.
В других способах оценки эффективности способа лечения для пациента контрольное количество антитела, специфического для пептида оболочки споры, определяют в образцах ткани из контрольной популяции. Количество антитела, специфического для пептида оболочки споры, в образце ткани от пациента после введения средства сравнивают с контрольным количеством антитела, специфического для пептида оболочки споры. Количество антитела, специфического для пептида оболочки споры, измеренное после лечения, которое значительно превышает контрольное количество антитела, специфического для пептида оболочки споры, указывает на положительный исход лечения.
В других способах оценки эффективности способа лечения для пациента контрольное количество антитела, специфического для пептида оболочки споры, определяют в образцах ткани из контрольной популяции. Количество антитела, специфического для пептида оболочки споры, в образце ткани пациента после введения средства сравнивают с контрольным количеством антитела, специфического для пептида оболочки споры. Отсутствие значительной разницы между количеством антитела, специфического для пептида оболочки споры, измеренным после начала указанного лечения, по сравнению с контрольным количеством антитела, специфического для пептида оболочки споры, указывает на отрицательный исход лечения.
Другие способы мониторирования заболевания или чувствительности к нему у пациента включают детекцию иммунного ответа против пептида оболочки споры в образце от пациента. В некоторых таких способах пациенту вводят средство, эффективное для лечения или профилактики заболеваний, вызванных микобактериями, и уровень ответа определяет будущий режим лечения пациента.
В других способах оценки эффективности способа лечения для пациента определяют значение количества антитела, специфического для пептида оболочки споры, в образце ткани пациента, которого лечили средством. Значение сравнивают с контрольным значением, определенным для популяции пациентов, испытывающих облегчение или освобождение от симптомов микобактериального заболевания благодаря лечению средством. Значение для пациента, по меньшей мере равное контрольному значению, указывает на положительный ответ на лечение.
Кроме того, изобретение относится к диагностическим наборам для осуществления вышеуказанных способов. Такие наборы, как правило, содержат реагент, специфически связывающийся с антителами против пептида оболочки споры или стимулирующий пролиферацию T-клеток, узнающих пептид оболочки споры.
ОПРЕДЕЛЕНИЯ
Белки, которые являются важными для образования спор, включают белки, кодируемые: a) генами для формирования оболочки, кортекса и внешнего слоя споры; b) генами, вовлеченными в расхождение хромосом и транслокацию ДНК от материнской клетки к споре; и c) факторами транскрипции, регулирующими предположительные гены споруляции. Такие белки в настоящей заявке называют «относящиеся к спорам белки» или «относящиеся к спорам пептиды».
Термин «относящиеся к спорам пептиды» включает пептиды, кодируемые одним из генов, перечисленных в таблицах 1-4, или их фрагменты. Предпочтительные такие пептиды представляют собой, например, CotA (MM1618), CotD (MM4853), CotT (MM4208), SpoVK, CotSA (MM4226), YrbC (MM2098), SpoVE, Soj и SpoIIIE. Формирующий споры пептид или его фрагмент должен предпочтительно являться иммуногенным средством. Наиболее предпочтительными являются CotA (MM1618), CotD (MM4853), CotT (MM4208), CotSA (MM4226) и YrbC (MM2098).
Термин «формирующий споры пептид» обозначает относящийся к спорам пептид, который формирует часть споры. Подобным образом, «пептид оболочки споры» обозначает относящийся к спорам пептид, который формирует часть оболочки споры. Группа «пептиды оболочки споры», таким образом, является подгруппой группы «формирующие споры пептиды», которая, в свою очередь, является подгруппой «относящихся к спорам пептидов». При применении относящегося к спорам пептида по настоящему изобретению, применение формирующих споры пептидов и особенно пептида оболочки споры представляет собой предпочтительные варианты осуществления, если конкретно не указано иначе.
Термин «пациент» предназначен, чтобы включать животных, таких как человек, и скот, такой как коровы, овцы, свиньи, собаки, куры, козы, кенгуру, страусы и буйволы, и домашних животных, таких как кошки, собаки, рыбы и птицы. Включены также дикие животные, такие как олень, северный олень, лось, кабан.
Термин «значительная идентичность» обозначает, что две пептидные последовательности, при оптимальном выравнивании, таком как с помощью программы GAP или BESTFIT с использованием стоимости пропусков по умолчанию, разделяют по меньшей мере 65-процентную идентичность последовательностей, предпочтительно, по меньшей мере 80- или 90-процентную идентичность последовательностей, более предпочтительно по меньшей мере 95-процентную идентичность последовательностей или более (например, 99-процентную идентичность последовательностей или выше). Предпочтительно положения остатков, которые не являются идентичными, отличаются консервативными аминокислотными заменами.
Термины «связывающая молекула» и «лекарственное средство» можно использовать взаимозаменяемо.
При сравнении последовательностей, как правило, одна последовательность действует как контрольная последовательность, с которой сравнивают тестируемые последовательности.
При использовании алгоритма сравнения последовательностей, тестируемую и контрольную последовательности вводят в компьютер, назначают координаты подпоследовательностей, если необходимо, и назначают алгоритм последовательности параметров программы. Алгоритм сравнения последовательностей, рассчитывающий процентную идентичность последовательностей для тестируемой последовательности (последовательностей) относительно контрольной последовательности, основан на назначенных параметрах программы.
Оптимальное выравнивание последовательностей для сравнения можно проводить, например, по алгоритму локальной гомологии Smith & Waterman, Adv. Appl. Math. 2:482 (1981), по алгоритму выравнивания гомологии Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), по способу поиска сходства Pearson & Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444 (1988), с помощью компьютеризованных осуществлений этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Madison, WI) или по визуальному контролю (смотри в общем Ausubel et al., выше). Одним примером алгоритма, подходящего для определения процентной идентичности последовательностей и сходства последовательностей, является алгоритм BLAST, описанный в Altschul et al., J. Mol. Biol. 215:403-410 (1990). Программное обеспечение для проведения анализов BLAST является публично доступным через Национальный центр биотехнологической информации (http://www.ncbi.nlm.nih.gov/). Как правило, параметры программы по умолчанию можно использовать для проведения сравнения последовательностей, хотя можно также использовать разработанные пользователем параметры. Для аминокислотных последовательностей в программе BLASTP используют по умолчанию длину слова (W) 3, ожидание (E) 10 и оценочную матрицу BLOSUM62 (смотри Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA 89, 10915 (1989)). Для целей классификации аминокислотных замен, как консервативных или неконсервативных, аминокислоты группируют следующим образом: Группа I (гидрофобные боковые цепи): норлейцин, met, ala, val, leu, ile; Группа II (нейтральные гидрофильные боковые цепи): cys, ser, thr; Группа III (кислые боковые цепи): asp, glu; Группа IV (основные боковые цепи): asn, gln, his, lys, arg; Группа V (остатки, влияющие на ориентацию цепи): gly, pro; и Группа VI (ароматические боковые цепи): trp, tyr, phe.
Консервативные замены включают замены между аминокислотами одного и того же класса. Неконсервативные замены состоят в замене члена одного из этих классов на член другого.
Лекарственные средства по изобретению, как правило, являются по существу чистыми. Это означает, что средство обладает, как правило, по меньшей мере приблизительно 50% масс./масс. (масса/масса) чистоты, также как является по существу свободным от создающих помехи белков и загрязнений.
Средства могут обладать чистотой по меньшей мере приблизительно 80% масс./масс. и более предпочтительно по меньшей мере 90% масс./масс. или приблизительно 95% масс./масс. Однако с использованием общепринятых способов очистки белка можно получать гомогенные пептиды по меньшей мере 99% масс./масс.
Аффинность между двумя молекулами означает аффинность по меньшей мере 106, 107, 108, 109 M-1 или 1010 M-1. Аффинности более 108 M-1 являются предпочтительными.
Термин «антитело» используют, чтобы включать интактные антитела и их связывающие фрагменты. Как правило, фрагменты конкурируют с интактным антителом, из которого они происходят, за специфическое связывание с антигеном. Необязательно, антитела или их связывающие фрагменты можно химически конъюгировать с другими белками или экспрессировать как слитые белки с ними.
Термин «эпитоп» или «антигенная детерминанта» относится к участку антигена, на который отвечают B- и/или T-клетки. Эпитопы B-клеток могут являться сформированными или из соседних аминокислот, или из не соседних аминокислот, располагающихся рядом при третичном сворачивании белка. Эпитопы, сформированные из соседних аминокислот, как правило, сохраняются при воздействии денатурирующих растворителей, в то время как эпитопы, сформированные посредством третичного сворачивания, как правило, теряются при обработке денатурирующими растворителями. Эпитоп, как правило, включает по меньшей мере 3, и более обычно, по меньшей мере 5 или 8-10 аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов включают, например, рентгеновскую кристаллографию и 2-мерный ядерный магнитный резонанс. Смотри, например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, Glenn E. Morris, Ed. (1996).
Антитела, узнающие один и тот же эпитоп, можно идентифицировать простым иммуноанализом, показывающим способность одного антитела блокировать связывание другого антитела с антигеном-мишенью. T-клетки узнают непрерывные эпитопы приблизительно из девяти аминокислот для CD8 клеток или приблизительно 13-15 аминокислот для CD4 клеток. T-клетки, узнающие эпитоп, можно идентифицировать в анализах in vitro, измеряющих зависимую от антигена пролиферацию, как определяют по включению 3H-тимидина в примированные T-клетки в ответ на эпитоп (Burke et al., J. Inf. Dis. 170, 1110-19 (1994)), по зависимому от антигена убиванию (анализ цитотоксических T-лимфоцитов, Tigges et al., J. Immunol. 156, 3901-3910) или по секреции цитокинов.
Термин «иммунологический» или «иммунный» ответ представляет собой развитие благоприятного гуморального (опосредованного антителом) и/или клеточного (опосредованного антигенспецифическими T-клетками или продуктами их секреции) ответа, направленного против пептида оболочки споры у пациента-реципиента. Такой ответ может представлять собой активный ответ, индуцированный введением иммуногена, или пассивный ответ, индуцированный введением антитела или примированных T-клеток. Клеточный иммунный ответ вызывают представлением полипептидных эпитопов совместно с молекулами MHC класса I или класса II для активации антигенспецифических CD4+ T-хелперных клеток и/или CD8+ цитотоксических T-клеток. Ответ может также вовлекать активацию моноцитов, макрофагов, NK-клеток, базофилов, дендритных клеток, астроцитов, клеток микроглии, эозинофилов или других компонентов врожденного иммунитета.
Присутствие опосредованного клетками иммунологического ответа можно определять анализами пролиферации (CD4+ T-клетки) или анализами CTL (цитотоксических T-лимфоцитов) (смотри Burke, выше; Tigges, выше). Относительные вклады гуморального и клеточного ответов в защитный или терапевтический эффект иммуногена можно различить отдельным выделением IgG и T-клеток от иммунизированного сингенного животного и измерением защитного или терапевтического эффекта у второго субъекта.
Термин «специфический для» указывает на то, что вариабельные области антител, или связывающие молекулы, узнают и связывают исключительно пептиды оболочки споры по изобретению (т.е. способны отличить пептид оболочки споры от других сходных полипептидов, несмотря на идентичность, гомологию или сходство последовательности, обнаруженные в семействе полипептидов), но могут также взаимодействовать с другими белками (например, белком A S. aureus или другими антителами в способах ELISA) посредством взаимодействий с последовательностями за пределами вариабельной области антитела, и в частности, в константной области молекулы. Анализы скрининга, в которых можно определять специфичность связывания антитела против пептида оболочки споры, хорошо известны и являются общепринятой практикой в данной области (Chapter 6, Antibodies A Laboratory Manual, Eds. Harlow, et al., Cold Spring Harbor Laboratory; Cold Spring Harbor, NY (1988), полное содержание которого приведено в настоящем документе в качестве ссылки).
«Иммуногенное средство» или «иммуноген» является способным индуцировать иммунологический ответ против себя при введении пациенту, необязательно совместно с адъювантом.
Термин «голый полинуклеотид» относится к полинуклеотиду, не образовавшему комплекс с коллоидными веществами. Голые полинуклеотиды иногда клонируют в плазмидный вектор.
Термин «адъювант» относится к соединению, которое при введении совместно с антигеном усиливает иммунный ответ на антиген, но при введении отдельно не вызывает иммунного ответа на антиген.
Адъюванты могут усиливать иммунный ответ посредством нескольких механизмов, включая рекрутирование лимфоцитов, стимуляцию B- и/или T-клеток и стимуляцию макрофагов.
Термин «пациент» включает субъектов - людей и других млекопитающих, подвергающихся либо профилактическому, либо терапевтическому лечению.
Композиции или способы, «включающие» один или несколько перечисленных элементов, могут включать другие элементы, конкретно не перечисленные.
Например, композиция содержащая пептид оболочки споры, включает как выделенный пептид оболочки споры, так и пептид оболочки споры в качестве компонента более длинной полипептидной последовательности.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1
Профили проточной цитометрии, покрывающие полный жизненный цикл M. marinum. Старую исходную культуру M. marinum использовали для инокуляции свежей среды, и за прогрессом культуры до стационарной фазы через экспоненциальный рост следили посредством проточной цитометрии. Исходную культуру в возрасте 2 месяцев высевали на чашки с агаром 7H10 с необходимыми добавками; клетки собирали в различные периоды времени в течение периода 2 месяцев. Собранные клетки фиксировали в 70% этаноле, промывали и ресуспендировали в 0,1 объеме TM (10 мМ Tris-HCl pH 7,8; 10 мМ MgCl2), окрашивали митрамицином A и бромидом этидия и проводили обработку в проточном цитометре (смотри Методы). Профили представляют собой гистограммы числа клеток, нанесенные на график против содержания ДНК в них (столбец A: флуоресценция, откалиброванная в эквивалентах числа хромосом) или размера (столбец B: светорассеяние, измеренное в единицах условной шкалы, поддерживаемое одинаковым для всех гистограмм, не показано). Возраст всех культур, в час(ах) после инокуляции в свежую среду, показан на левой стороне каждого ряда. Профили в верхнем ряду, показывающие культуру в стационарной фазе лабораторного штамма грамотрицательной бактерии Escherichia coli K12 (цилиндры 1-3 мкм×1 мкм, содержащие 4,6 М п.о. ДНК на хромосому), использовали в качестве стандарта калибровки для определения содержания ДНК и размера клеток в профилях M. marinum.
Фигура 2
Флуоресцентная микроскопия M. marinum на различных стадиях роста от свежего инокулята до стационарной фазы. Исходную культуру в возрасте 2-месяцев высевали на чашки, и аликвоты фиксировали в этаноле, как описано в подписи к фиг. 1. Фиксированные клетки промывали в PBS, наслаивали на тонкие пленки агарозы (1% в 0,9% NaCl), содержащие 0,5 мкг/мл DAPI на предметном стекле, и обследовали под микроскопом Zeiss (Axioplan 2) с помощью камеры CCD (смотри Методы). На панелях a-g показаны клетки, собранные через 0 час, 6 час, 12 час, 72 час (3 суток), 120 час (5 суток), 168 час (1 неделя) и 336 час (2 недели) после инокуляции в свежую среду. Линейка: 5 мкм.
Фигура 3
Морфология поверхности и внутренние структуры клеток M. marinum на различных стадиях споруляции по сканирующей электронной микроскопии (SEM) и трансмиссионной электронной микроскопии на тонких срезах (TEM), соответственно. Клетки в различное время после инокуляции в свежую среду получали для SEM и TEM, как описано в Методах.
(A) Изображения SEM: (a) выделенные споры; (b) клетки при прорастании; (c) предположительные эндоспоры (5 суток после инокуляции) и (d) спора с вегетативной клеткой (культура в возрасте 7 суток). Линейка: 1 мкм.
(B) Изображения TEM: (a) выделенные споры; (b, c) зрелые споры; (d-f) различные стадии прорастания спор (6 час после инокуляции); (g) предспора слева и зрелая эндоспора справа (культура в возрасте 5 суток). Увеличения: (a) 6000×; (b,c) 60000×; (d-f) 40000×; (g) 30000×.
Фигура 4
Биохимические свойства поверхности и термостабильность предположительных спор M. marinum
(a) Специфическое для спор дифференциальное окрашивание клеток M. marinum из культур экспоненциальной и стационарной фазы. Клетки, выращенные и собранные на различных стадиях роста, фиксировали нагреванием, подвергали окрашиванию и контрастному окрашиванию малахитовым зеленым и сафранином, соответственно (смотри Методы), и исследовали под 100× объективом микроскопа Olympus CH30RF200.
(b) Способность к образованию колоний после обработки нагреванием. Чашки, содержащие клетки в экспоненциальной (12 час после инокуляции) и стационарной (после 14 суток роста) фазе с подверганием и без подвергания обработке влажностью-нагреванием (15 мин, 65°C).
(c) Скорость убивания влажностью-нагреванием клеток M. marinum с экспоненциальной и стационарной фазах. На клетки в экспоненциальной фазе (12 час, ромбы) и стационарной фазе (14 суток, квадраты) воздействовали влажностью-нагреванием в течение различных периодов времени, выжившие колонии выражали как проценты относительно идентичной, не подвергавшейся воздействию популяции клеток, и наносили на график как функцию периодов времени воздействия. Каждая точка на графике выживаемости представляет среднее по меньшей мере из трех измерений (таблица 5) с различиями между экспериментами, указанными планками погрешностей.
Фигура 5
Относительные уровни экспрессии предположительных генов споруляции из генома M. marinum на различных стадиях жизненного цикла. Уровни мРНК 9 генов из генома M. marinum, гомологов известных генов споруляции из Bacillus subtilis и Streptomyces coelicolor, сравнивали с использованием дот-блот гибридизации при прогрессировании культуры от экспоненциальной до стационарной фазы. Специфические олигонуклеотидные зонды (последовательности в таблице 6) использовали для каждой мРНК-кандидата. Относительные интенсивности точек нормализовали с использованием соответствующих сигналов 5S рРНК в качестве внутренних стандартов и наносили на график в условных единицах (Y-ось) как сигналы мРНК M. marinum (определенные как их гомологи, X-ось) из культур различного возраста (столбцы различного рисунка). Все образцы анализировали по меньшей мере три раза, и определенные различия между экспериментами указаны планками погрешностей. Числовые значения (таблица 7) получены посредством анализа сигналов дот-блоттинга на аппарате для визуализации фосфора (Image Quant, Molecular Dynamics 400).
Фигура 6
Присутствие споровых частиц в культуре Mycobacterium bovis (BCG).
(a) Фазовое флуоресцентное изображение клеток M. bovis из культуры в возрасте шести месяцев. Линейка: 5мкм.
(b) Дифференциально окрашенные очищенные споровые частицы из культуры BCG.
(c) Изображение SEM очищенных спор BCG. Линейка: 1 мкм.
(d) Изображение TEM очищенных спор BCG на тонких срезах. Увеличение: 60000×.
Фигура 7.
Морфологическое сравнение колоний M. marinum из культур, пророщенных из отдельных очищенных (смотри Способы) независимых споровых частиц (a-d) с колониями из экспоненциально растущей культуры, инокулированной из старой исходной культуры (e). Линейка: 1 см.
Фигура 8.
Последовательности гена РНК РНКазы P из независимых клонов, происходящих из выделенных отдельных споровых частиц. Выравнивание остатков 89-315 гена РНК РНКазы P из хромосом выделенных клонированием колоний. (1-4): Последовательности из геномной ДНК, выделенной из отдельных колоний, выращенных из отдельных выделенных споровых частиц. (5) Последовательность гена РНК РНКазы P M. marinum, клонированная в плазмиде pIGn. (6) Последовательность гена РНК РНКазы P M. marinum, полученная поиском в базе данных Singer 24 с использованием последовательности гена РНК РНКазы P M. tuberculosis H37Rv (http://www.sanger.ac.uk./Projects/M_tuberculosis) в качестве контрольной. Последовательности 1-5 являются идентичными последовательностям, опубликованным в базе данных, за исключением двух положений, 93 и 158. Это может являться обусловленным различиями между лабораторным штаммом (ATCC 927) и штаммом, используемым для построения базы данных (ATCC BAA-535), или ошибкой в базе данных. Олигонуклеотиды, используемые для ПЦР-амплификации области 89-315, представляли собой:
Прямой: 5'-ATCGAGGAAAGTCCGGACTTCACA-3' (содержащий область 65-88)
Обратный: 5'-ACCCGCAGACTCGGGCGAGCAG-3' (комплементарный области 294-315). Прямой олигонуклеотид использовали для получения последовательностей 1-5.
Фигура 9.
Жизненный цикл M. marinum из очищенных спор по микроскопии. a, чистая популяция спор, 0 час. b, прорастающие споры, 6 час. c, прорастающие клетки, 8 час. d, прорастающие клетки, 24 час (1 сутки). e, то же, что и d, 2 суток. f, то же, что и e, 3 суток. g и h, внешний вид эндоспор, 5 d. i, внешний вид зрелых спор, 7 суток. j, зрелые споры с клеточным дебрисом, 14 суток. Линейки: 5 мкм.
Фигура 10.
Жизненный цикл M. marinum из очищенных спор по проточной цитометрии. Культуру M. marinum начинали с суспензии очищенных спор и следили за прорастанием, экспоненциальным ростом до споруляции в поздней стационарной фазе по проточной цитометрии. В столбцах (A) и (B) показано содержание ДНК (флуоресценция откалибрована по числу хромосом) и распределение размера клеток (светорассеяние в условных единицах), соответственно. Верхние панели относятся к культурам E. coli в стационарной фазе в LB, используемым в качестве калибровочных стандартов для содержания ДНК и размера клеток. Девять нижних панелей соответствуют образцам M. marinum, взятым через: 0, 6, 8, 24, 48, 72, 120, 168 и 336 часов после суспендирования очищенных спор в среде для роста. Способы фиксации и окрашивания являлись такими, как описано в Методах и в подписи фигуры 1.
Узкое распределение размеров и сдвиг пика в целом с небольшим расширением соответствует синхронному переходу спор в вегетативные клетки. На поздней стационарной стадии (через 120 часов роста) распределение размера клеток соответствует возникновению эндоспор по сужению пика и его сдвигу в сторону меньшего размера (столбец B: 168 час и 336 час). Профили флуоресценции показывают начало синтеза ДНК к 6 час после добавления среды для роста; репликация продолжается в течение 72 час, после чего после нерепликативного деления клеток следует споруляция.
ПОДРОБНОЕ ОПИСАНИЕ
Изобретение относится к фармацевтическим композициям и способам профилактического и терапевтического лечения заболеваний, вызванных инфекцией микобактерией. Более того, изобретение относится к таким композициям и способам для лечения таких заболеваний, где микобактерия формирует споры.
Род Mycobacterium включает непатогенные бактерии из окружающей среды, так же как высокоэффективные патогены, например, Mycobacterium tuberculosis (Mtb) и Mycobacterium leprae, возбудители туберкулеза и лепры, соответственно. В то время как Mtb является специфической для человека, близкородственная Mycobacterium bovis может инфицировать как людей, так и животных, так и приводить к туберкулезу вне зависимости от хозяина. Более того, микобактерия Mycobacterium avium подвид paratuberculosis (MAP), возбудитель хронической диареи у жвачных (болезнь Джона), может присутствовать в пищевых продуктах, происходящих от скота, и обсуждают, что MAP является возбудителем болезни Крона у человека.
Другая близкородственная микобактерия, Mycobacterium marinum, вызывает системную, подобную туберкулезу инфекцию и заболевание у холоднокровных хозяев, например, рыб и лягушек. По сравнению с Mtb, M. marinum растет с относительно коротким временем удвоения, и рост является ограниченным при более высоких температурах (смотри ниже), что делает M. marinum более легкой для манипуляций, чем, например, Mtb. M. marinum недавно стала модельной системой для идентификации и изучения факторов, важных для микобактериальной инфекции, развития заболевания и его персистентности. Патогенные микобактерии являются инвазивными медленно растущими бактериями, которые могут расти и размножаться внутри макрофагов. Более того, большинство из них могут вызывать долговременные инфекции, которые могут вызывать острую или хроническую стадию заболевания или стадию, которая является клинически бессимптомной. В последней ситуации бактерии обладают способностью возобновлять рост через несколько десятков лет состояния покоя, что приводит к активному заболеванию.
Врожденная иммунная система является необходимой для борьбы против бактериальных инфекций. Макрофаги являются важными участниками этого процесса, и они играют важную роль на передовой линии защиты. Патогенные микобактерии, например Mtb, поглощаются посредством фагоцитоза альвеолярными макрофагами в участке вхождения, и внутри макрофага бактерии находятся внутри связанной с мембраной цитоплазматической вакуоли, обозначаемой как фагосома Mycobacterium. Патогенные микобактерии избегают фаголизосомальной деградации посредством: 1) блокирования ацидификации фагосомы, 2) изменения содержания белка вакуоли и 3) профилактики взаимодействия с другими эндосомальными компартментами. Это обеспечивает выживание бактерий внутри макрофага и последующее распространение патогена к другим клеткам, например, дендритным клеткам и нейтрофилам.
Полный жизненный цикл M. marinum: размер клеток и распределение содержания ДНК
За изменениями размера клеток и распределением содержания ДНК культур M. marinum на чашках на протяжении полного жизненного цикла следили посредством проточной цитометрии, Maisnier-Patin, S., Dasgupta, S., Krabbe, M., Nordstrom, K. Conversion to bidirectional replication after unidirectional initiation from Rl plasmid origin integrated at oriC in Escherichia coli. Mol. Microbiol. 30, 1067-1079 (1998). На фигуре 1 показаны профили проточной цитометрии для распределения содержания ДНК и размера клеток в культуре от 2 час до 2 месяцев после инокуляции. Измерения проводили каждые 2 час в течение 12 час, через 24 час, каждые 24 час до 96 час, через 196 час и через 2 месяца. Для ночной стационарной культуры Escherichia coli K12, используемой в качестве калибровочного стандарта, показали однородную популяцию с узким распределением размеров (отдельный острый симметричный пик с нормальным распределением; верхний ряд, столбец B), содержащую либо одну, либо две полноразмерные хромосомы (два острых пика, указанные номерами 1 и 2 на гистограмме флуоресценции, предварительно откалиброванной в отношении размера генома с использованием хромосомы E. coli в качестве стандарта; смотри верхний ряд, столбец A). Профили проточной цитометрии клеток M. marinum из исходной культуры в возрасте 2 месяцев остаются неизменными от 0 час (не показано) вплоть до 4 час после инокуляции в свежую среду. Для них показали два отдельных класса популяций клеток: большинство, состоящее из клеток очень малого размера с флуоресценцией, соответствующей одному геному M. marinum (6,6 М п.о., http://www.sanger.ac.uk/Projects/M_marinum/) или менее, и субпопуляция с флуоресценцией, соответствующий ДНК, эквивалентной 2 геномам (показанной линейками, отмеченными 1 и 2, на профиле содержания ДНК для 2 месяцев). Через 6 часов пик светорассеяния, представляющий размер клеток, начал расширяться, позволяя предполагать увеличение размера клеток, сопровождающихся содержанием ДНК, эквивалентным более 2 геномам в некоторых клетках (начало синтеза ДНК). С увеличением времени распределения размера клеток и содержания ДНК подверглись значительным изменениям - отдельные пики, представляющие очень маленькие и большие клетки, заменились отдельным, более широким пиком, соответствующим непрерывному распределению размеров экспоненциально растущей популяции клеток (10 час после инокуляции). Пик флуоресценции, соответствующий эквиваленту 2 хромосом ДНК, плавно расширился до 2-4 или более хромосом, указывая на непрерывный синтез ДНК. В соответствии с этим, пик для 1 хромосомы постепенно исчез, когда большее количество клеток вошли в репликативную фазу из подобного споре состояния «с отсутствием репликации, небольшим размером» через 10 час роста в свежей среде. Очень маленькие клетки с одним геномным эквивалентом ДНК персистировали до приблизительно 6 час после инокуляции в свежую среду, и затем оба вида исчезали с заменой клетками большего размера, содержащими 2-4 хромосомных эквивалента ДНК (смотри панели для 10 час). Считают, что очень маленький размер клеток представляет не реплицирующие подобные спорам частицы. При дальнейшей инкубации распределение размера клеток достигает его наивысшего среднего значения с наиболее широким распределением (24 час) и затем разделяется на меньшие, более однородные распределения размеров; содержание ДНК указывает на то, что продолжающийся синтез ДНК постепенно заканчивается, и начинают появляться клетки с 2 и 4 хромосомными эквивалентами ДНК (24 час). Репликация в субпопуляции продолжалась, в то время как нерепликативное деление являлось очевидным по увеличивающейся доле 2-хромосомных клеток через 96 час роста (смотри 48 час, 72 час и 96 час). Приблизительно через неделю почти все клетки входили в отдельное распределение с меньшим размером с 1 либо 2 полноразмерными хромосомными эквивалентами ДНК: через два месяца большинство клеток перешли в форму с очень малым размером с 1 хромосомой с малой субпопуляцией более крупных клеток, содержащих 2 хромосомных эквивалента ДНК, сходной с исходной культурой в возрасте 2 месяцев через 2 час (сравни профили проточной цитометрии для 2 час и 2 месяцев на фиг. 1).
Чтобы прояснить природу очень маленьких клеток без репликации на ранних и поздних стадиях роста (2 час-8 час и через 192 час), популяцию клеток исследовали на различных стадиях роста от старой исходной культуры до стационарной через экспоненциальную фазу под фазовым флуоресцентным микроскопом. Фиксированные этанолом клетки окрашивали 4,6-диамидино-2-фенилиндолом (DAPI) для визуализации ДНК. На фигуре 2 показаны фазовые флуоресцентные микрофотографии клеток из: исходной культуры в возрасте 2 месяцев (через 0 час роста), через 6 час, 12 час, 3 суток, 5 суток, 7 суток и 14 суток роста (a-g, соответственно) после инокуляции в свежую среду. Для старой исходной культуры (фиг. 2a) показали маленькие эллипсоидальные частицы с ярким отражением на фазовых изображениях, характеризующим споры, Hitchins, A. D., Kahn, A. J., Slepecky, R. A. Interference contrast and phase contrast microscopy of sporulation and germination of Bacillus megaterium. J. Bacteriol. 96, 1811-1817 (1968), возможно, соответствующие маленьким клеткам без репликации на профилях проточной цитометрии (смотри 2 час, фиг. 1). Эти клетки, хотя и содержат один геномный эквивалент ДНК, обладают более низкой флуоресценцией DAPI по сравнению с вегетативными клетками (например, 12 час, фиг. 2), что позволяет предполагать слабое поглощение красителя; это объясняет также слабую флуоресценцию популяции маленьких клеток на профилях проточной цитометрии (смотри фиг. 1: 2 час-6 час и 2 месяца). После высевания на свежую среду наблюдали возникновение цилиндрических вегетативных клеток из спор с серединой прорастания через 6 час инкубации (фиг. 2b), т.е. через время приблизительно одного поколения. Флуоресценция от окрашенной DAPI ДНК являлась видимой в удлиненной части. Через 12 час почти все клетки находились в состоянии вегетативного роста и выглядели подобно цилиндрическим бактериям с явно видимыми окрашенными DAPI нуклеоидами (фиг. 2c), и это продолжалось вплоть до 3 суток (фиг. 2d). На сутки 5 наблюдали подобные эндоспорам виды (фиг. 2e), со светящимся пятном внутри клетки около одного полюса, с перетяжкой, отделяющей яркое пятно от остальной части клетки. Через неделю роста начали появляться споры, как видно по ярким фазовым изображениям и затуханию флуоресценции (фиг. 2f). Формирование спор продолжалось с прогрессированием возраста культуры (фиг. 2g, через 2 недели роста). По объединенным данным проточной цитометрии и флуоресцентной микроскопии, вероятно, M. marinum начинает спорулировать через некоторое время после установления стационарной фазы, хотя точное время начала и ареста споруляции еще предстоит определить (смотри фиг. 1 и 2). Однако, по настоящим данным, споруляция начинается поздно в жизненном цикле культуры M. marinum, наиболее вероятно, через несколько часов после начала стационарной фазы. С другой стороны, прорастание или выход из споруляции, по-видимому, начинается, как только споры подвергают воздействию свежей среды.
Отдельная спора развивается в отдельную колонию M. marinum
Для исключения возможности того, что эти подобные спорам частицы представляли собой контаминацию от другого источника образования спор, выделяли чистые споры, свободные от вегетативных клеток (Методы, фиг. 9a). Партию чистых спор серийно разводили так, чтобы в каждой разведенной фракции присутствовала одна или менее жизнеспособная спора (смотри Методы). Образцы из 10 повторов разведения высевали. Через неделю после инокуляции на четырех из десяти чашек показали отдельную колонию, которая прорастала и росла. Каждая из этих колоний, происходящих из четырех независимых спор, являлась морфологически идентичной колонии M. marinum (фиг. 7). Для верификации того, что эти колонии действительно являлись M. marinum, часть гена РНК РНКазы P амплифицировали с помощью ПЦР с использованием тотальной ДНК, выделенной из каждой из них. Секвенированием ДНК (фиг. 8) подтвердили, что колонии представляют собой M. marinum. Выделенные споры M. marinum также высевали на свежие чашки, и за их жизненным циклом следили с помощью фазовой флуоресцентной микроскопии и проточной цитометрии (фиг. 9 и 10). Наблюдали, что они прорастали до вегетативных клеток, которые снова спорулировали в поздней стационарной фазе. Таким образом, очищенные споровые частицы прорастали в клетки M. marinum и, таким образом, не являлись контаминантами.
Визуализация споровых частиц, споруляции и прорастания с помощью электронной микроскопии
Очень различающиеся размеры и морфологию клеток в состояниях споруляции и прорастания исследовали по сканирующей электронной микрофотографии (SEM), Lackner P. et al. Scanning electron microscopy of the neuropathology of murine cerebral malaria. Malar J. 5, 116 (2006). На фигуре 3 (панель A; a-d) показаны изображения SEM M. marinum на различных стадиях споруляции. Наблюдаемые различные типы клеток представляли собой: a) выделенные споры; b) вегетативные клетки, прорастающие из споры (6 час); c) клетки с возможными эндоспорами, обладающие выпуклостями около одного полюса (сутки 5); и d) спору с вегетативной клеткой (сутки 7). Внутренние структуры спор исследовали трансмиссионной электронной микроскопией (TEM) на тонких срезах фиксированных, выделенных спор (фиг. 3, панель B; a-c) и клеток M. marinum из культур на различных стадиях (фиг. 3 панель, B; d-g). Посредством сравнения изображений TEM хорошо охарактеризованных спор Bacillus, Wang, L., Perpich, J., Driks, A., and Kroos, L. Maintaining the transcription factor SpoIIID level late during sporulation causes spore defects in Bacillus subtilis. J. Bacteriol. 189, 7302-7309 (2007), с изображениями выделенных спор M. marinum ясно идентифицировали внешнюю оболочку, внутреннюю оболочку, кортекс и сердцевину зрелой споры (фиг. 3, панель B; b и c). На изображении TEM клеток через 6 часов после инокуляции спор в свежую среду показали вегетативные клетки, постепенно прорастающие из пустых спор с нарушенной поверхностью (фиг. 3, панель B; d-f). На изображениях TEM на сутки 5 клеток, для которых показали подобные эндоспорам виды при фазовой флуоресцентной микроскопии (фиг. 2e), наблюдали структуры, выглядящие подобно эндоспорам на различных стадиях их созревания (фиг. 3, панель B; g, предспора и зрелая эндоспора, рядом). Они являются морфологически очень сходными с опубликованными изображениями эндоспор Bacillus spp., Erlendsson, L. S., Möller, M., and Hederstedt, L. Bacillus subtilis StoA is a thiol-disulfide oxidoreductase important for spore cortex synthesis. J. Bacteriol. 186, 6230-6308 (2004). Не наблюдали таких структур на изображениях TEM партии вегетативно растущих клеток в экспоненциальной фазе (не показано). Эти данные позволяют предполагать, что M. marinum действительно формирует споры, и что путь споруляции проходит через формирование эндоспоры.
Физические и биохимические свойства споровых частиц M. marinum и присутствие геномной информации, необходимой для споруляции у микобактерий
Если «подобные спорам» частицы, визуализируемые микроскопически в стационарной культуре M. marinum, являются истинными спорами, они должны обладать: (i) определенными биохимическими свойствами, позволяющими окрашивание специфическими для спор красителями; (ii) высокой устойчивостью к физическим и химическим стрессам; и (iii) присутствием генетического аппарата, необходимого для входа в состояние споры и выхода из него. Дифференциальное окрашивание спор и тест устойчивости к нагреванию, соответственно, проводили для проверки первых двух требований, и биоинформатический анализ для третьего.
Классическое дифференциальное окрашивание спор проводили с использованием малахитового зеленого в качестве первичного красителя, воды в качестве обесцвечивающего вещества и сафранина в качестве контрастного красителя Cappuccino, J. G., Sherman, N. in Microbiology, a Laboratory Manual (Benjamin Cummings, San Fransisco, sixth edition, 2001). Как показано на фиг. 4a, вегетативные клетки, по-видимому, теряют первичный краситель и поглощают контрастный краситель и выглядят красными. В отличие от этого, споры сохраняют первичный краситель и выглядят зелеными, указывая на то, что поверхность подобных спорам частиц в культурах M. marinum обладает биохимическими свойствами, сходными с хорошо известными спорами B. subtilis.
Затем сравнивали эффект обработки влажностью-нагреванием (65°C, 15 минут) экспоненциально растущих клеток (12 час) и клеток в стационарной фазе (14 суток), Chakrabarty, K. et al., Infection and Immunity. 74, 4430-4438 (2006). В то время как экспоненциально растущие клетки являлись почти полностью убитыми после 10 мин обработки нагреванием, около 40% клеток в стационарной фазе выживали даже после 30 мин обработки (фиг. 4, b и c). Это является количественно пропорциональным популяции спор, наблюдаемой в стационарных культурах. Показали также, что споры являются более устойчивыми к обработке глутаральдегидом по сравнению с клетками, растущими в экспоненциальной фазе (не показано).
Наконец, геном микобактерий сканировали для поиска генов, ответственных за формирование спор. Гены, необходимые для споруляции, идентифицированы в грамположительных бактериях, например, B. subtilis, http://genolist.pasteur.fr/SubtiList/, и Streptomyces coelicolor, http://streptomyces.org.uk/sco/index.html. Использовали подход обратимого наилучшего совпадения (RBH), Rivera, M. C, Jain, R., Moore, J. E. & Lake, J. A. Genomic evidence for two functionally distinct gene classes. Proc. Natl Acad. Sd. USA 95, 6239-6244 (1998), для идентификации предположительных ортологов хорошо известных генов споруляции в геномах M. marinum, ftp://ftp.sanger.ac.uk/pub/pathogens/mm/, и M. tuberculosis, http://cmr.tigr.org/tigr-scripts/CMR/CrnrHomePage.cgi, (таблицы 1-4). На основании функциональной аннотации генов B. subtilis их разделяли на три класса и они включали: a) гены для формирования оболочки, кортекса и внешнего слоя споры; b) гены, вовлеченные в расхождение хромосом и транслокацию ДНК от материнской клетки к споре; и c) факторы транскрипции, регулирующие предположительные гены споруляции. Некоторые из этих генов, из всех трех классов, выбрали для проверки относительных уровней экспрессии их мРНК посредством дот-блот-анализа, Sambrook, J., Fritsch, E.F., Maniatis, T. Molecular Cloning, a Laboratory Manual. (Cold Spring Harbor Laboratory Press, New York, third edition, 2001). Как показано на фигуре 5, выбранные гены транскрибировались в M. marinum. В значительной степени, экспрессия гомологов генов споруляции зависела от возраста культуры. Для уровней мРНК гомологов, http://genolist.pasteur.fr/SubtiList/, SpoVK (нарушение этого гена приводит к формированию незрелых спор, стадия V), CotSA (белок оболочки споры), YrbC (сходный с белком оболочки споры) и SpoVE (необходим для синтеза пептидогликана кортекса споры, стадия V) показали резкое увеличение с суток 5, продолжающееся до суток 7. Для гомологов Soj (функция, подобная центромере, вовлечен в расхождение хромосом предспоры/отрицательную регуляцию инициации споруляции) и SpoIIIE (транслоказа ДНК, необходимая для расхождения хромосом через септу в предспору) показали сходную картину, но более умеренное увеличение по сравнению с вышеупомянутыми четырьмя генами. Некоторые более слабые совпадения получены в биоинформатическом поиске для некоторых главных факторов транскрипции, и некоторые из них исследованы, например, для SpoOA (главный регулятор для перехода к споруляции) резкое увеличение с суток 5, которое продолжается до суток 7. Сигнал SpoOA с наилучшей суммой баллов в штамме Mtb CDC1 551 представляет собой ген trcR, кодирующий ДНК-связывающий регулятор ответа, Haydel, S. E., Clark-Curtiss, J. E. The Mycobacterium tuberculosis TrcR response regulator represses transcription of the intracellularly expressed Rv1057 gene. J.Bacteriol. 188, 150-159 (2006), (TrcR на фиг. 5). Для других факторов SigF (сигма-фактор специфической для поздней споруляции РНК-полимеразы) показали умеренное увеличение на сутки 7, в то время как для гомолога анти-SigF, SpoIIAB, показали умеренное увеличение на сутки 5 и 7. мРНК сигма-фактора «домашнего хозяйства», SigA (σ70), Gruber, T. M., Gross, C. A. Multiple sigma subunits and the partitioning of bacterial transcription space. Annu. Rev. Microbiol. 57, 441-466 (2003), M. marinum не играет какой-либо непосредственной роли в споруляции, Piggot, P. J., Hubert, D. W. Sporulation of Bacillus subtilis. Current Opinion in Microbiology. 7, 579 (2004), и уровень его мРНК уменьшается, когда культура входит в стационарную фазу и прогрессирует через нее. Эти данные поддерживают присутствие молекулярного пути для споруляции в M. marinum и, предположительно, также в Mtb. Экспрессию этих генов на уровне белка, и их дальнейшую характеризацию, такую как характер фосфорилирования/дефосфорилирования, еще предстоит исследовать. Тот факт, что не обнаружили сильных сигналов для главного фактора транскрипции, специфического для споруляции, может указывать либо на то, что: a) соответствующие гены отличаются по их последовательностям при сохранении их функций, либо на то, что b) эти факторы являются уникальными для микобактерий и их еще предстоит идентифицировать с использованием других способов. Следует отметить что образцы РНК, полученные из клеток на сутки 7, представляют гены, экспрессированые в смешанной популяции вегетативных клеток и спор (фиг. 2f и 3, панель A; d).
Споровые частицы в старых культурах штамма M. bovis BCG
Авторы настоящего изобретения наблюдали также подобные спорам виды в поздних стационарных культурах штамма Mycobacterium bovis BCG. При фазовой флуоресцентной микроскопии для клеток BCG с чашки в возрасте шести месяцев показали яркие, рефракционные частицы (фиг. 6a), сходные со спорами M. marinum. Эти частицы очищали (смотри Методы) и подвергали специфическому для спор окрашиванию (фиг. 6b) и электронной микроскопии (как SEM, так и TEM, фиг. 6c и d, соответственно). На основании этих данных, по-видимому, эти частицы являются сходными со спорами M. marinum. Таким образом, эти данные предоставляют экспериментальные свидетельства в пользу возможности того, что споруляция является более общим свойством микобактерий и не ограничена M. marinum. Следовательно, споруляцию можно рассматривать как общую стратегию выживания при стрессе.
Лекарственные средства для применения по настоящему изобретению индуцируют иммунный ответ против пептида оболочки споры, или других белков или пептидов, вовлеченных в формирование спор. Эти средства включают собственно пептид оболочки споры и его варианты, аналоги и миметики пептида оболочки споры, которые индуцируют антитела против пептида оболочки споры и/или перекрестно реагируют с ними, и антитела или T-клетки, узнающие пептид оболочки споры. Индукция иммунного ответа может являться активной, как при введении иммуногена для индукции антител или T-клеток, узнающих пептид оболочки споры, у пациента, или пассивной, как при введении антитела, которое само связывает пептид оболочки споры у пациента.
Лекарственное средство, применяемое в заявленных способах, может представлять собой любую из природных форм белка, вовлеченного в формирование спор, такого как белки, полученные с генов в таблицах 1-4. Такой ген включает в частности: a) гены для формирования оболочки, кортекса и внешнего слоя споры; b) гены, вовлеченные в расхождение хромосом и транслокацию ДНК от материнской клетки к споре; и c) факторы транскрипции, регулирующие предположительные гены споруляции. Ряд этих генов выбрали из всех трех классов (a, b или c) для проверки относительных уровней экспрессии их мРНК посредством дот-блот-анализа. Выбранные гены могут транскрибироваться в M. marinum. Экспрессия гомологов генов споруляции зависела от возраста культуры. Для уровней мРНК гомологов SpoVK (нарушение этого гена приводит к формированию незрелых спор, стадия V), CotSA (белок оболочки споры), YrbC (белок оболочки споры) и SpoVE (необходим для синтеза пептидогликана кортекса споры, стадия V) показали резкое увеличение с суток 5, продолжающееся до суток 7. Гомологи Soj (функция, подобная центромере, вовлечен в расхождение хромосом предспоры/отрицательную регуляцию инициации споруляции) и SpoIIIE (транслоказа ДНК, необходимая для расхождение хромосом через септу в предспору)
Лекарственное средство может представлять собой иммуногенный пептид, фрагмент или аналог белка, кодируемого геном, показанным в таблицах 1-4.
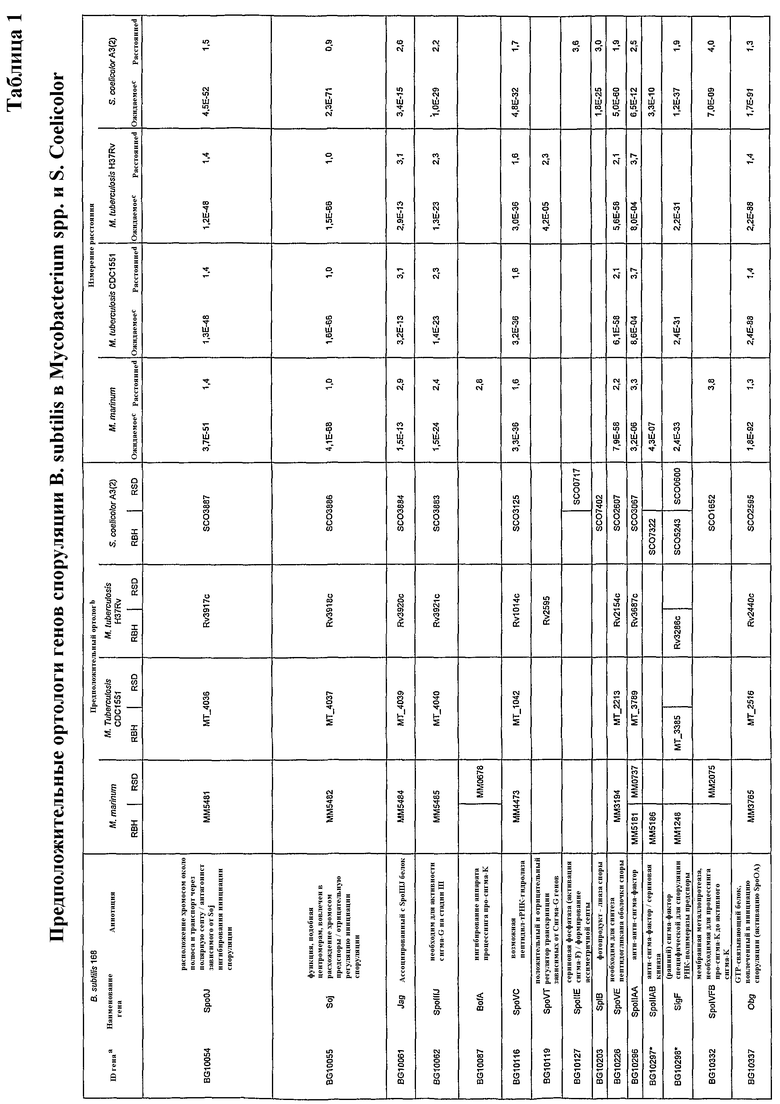
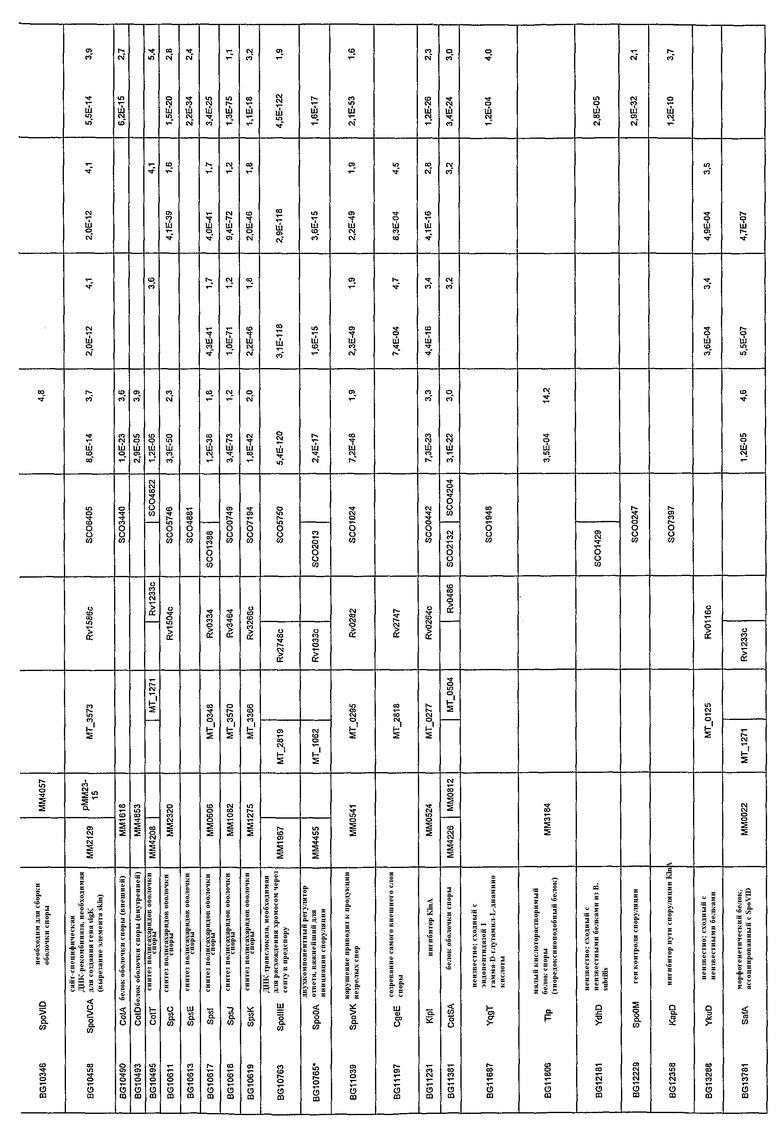

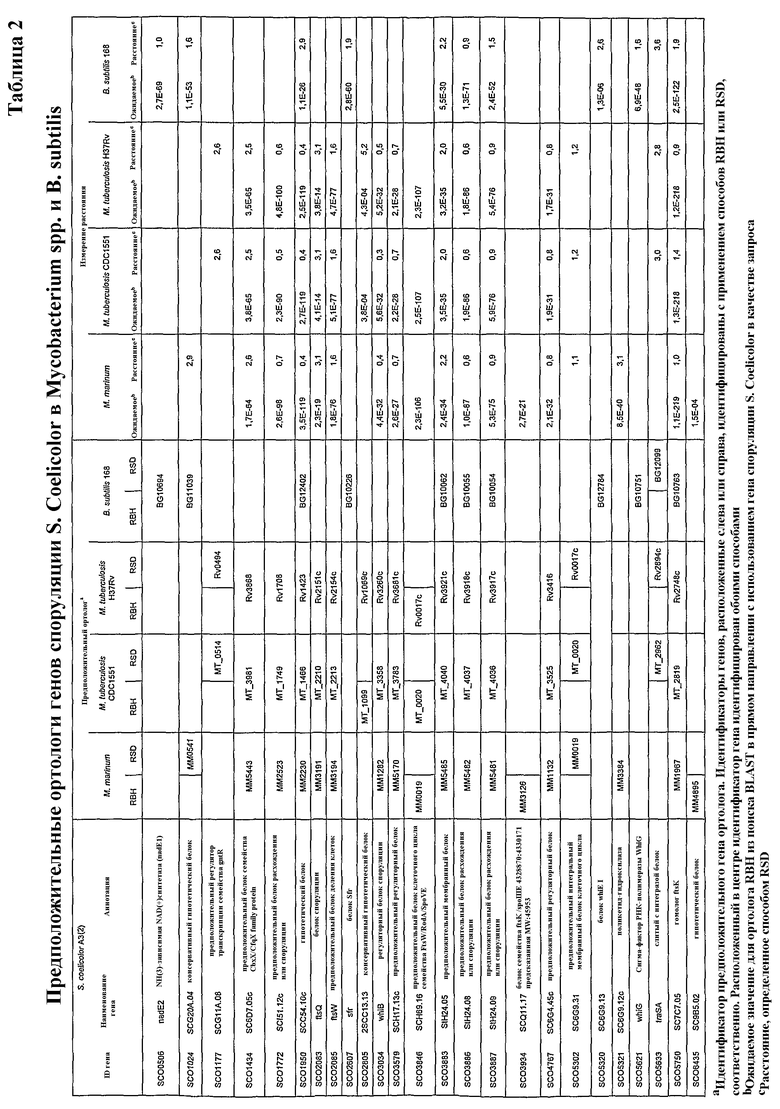

Предусматривают также, что иммуногенные пептиды могут происходить из гомологов вышеупомянутых белков из других микобактерий, например, M. tuberculosis, M. leprae, M. bovis, M. avium, M. microti, M. intracellulare, M. paratuberculosis, M. ulcerans или M. africanum.
Иммуногенные фрагменты, как правило, обладают последовательностью по меньшей мере из 3, 5, 6, 10 или 20 непрерывных аминокислот из природного пептида.
Аналоги включают аллельные, видовые и индуцированные варианты. Аналоги, как правило, отличаются от природных пептидов в одном или нескольких положениях, часто благодаря консервативным заменам. Аналоги, как правило, обладают по меньшей мере 80% или 90% идентичностью последовательности с природными пептидами. Некоторые аналоги включают также неприродные аминокислоты или модификации N- или C-концевых аминокислот. Примеры неприродных аминокислот представляют собой a,a-дизамещенные аминокислоты, N-алкиламинокислоты, молочную кислоту, 4-гидроксипролин, карбоксиглутамат, e-N,N,N-триметиллизин, e-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, N-метиларгинин.
Можно проводить скрининг фрагментов и аналогов по профилактической или терапевтической эффективности в моделях заболевания, как описано ниже.
Белок, выбранный из одного из белков, кодируемых одним из генов в таблицах 1-4, их фрагментов, аналогов и формирующих споры пептидов, можно синтезировать твердофазным пептидным синтезом или рекомбинантной экспрессией, или можно получать из природных источников. Автоматические синтезаторы пептидов являются коммерчески доступными от различных поставщиков, таких как Applied Biosystems, Foster City, California. Рекомбинантная экспрессия может происходить в бактериях, таких как E. coli, дрожжах, клетках насекомых или клетках млекопитающих. Способы рекомбинантной экспрессии описаны в Sambrook et al., Molecular Cloning: A Laboratory Manual (C. S. H. P. Press, NY 2d ed., 1989).
Лекарственные средства включают также более длинные полипептиды, которые включают, например, формирующий споры пептид, его активный фрагмент или аналог вместе с другими аминокислотами. Лекарственные средства включают также мультимеры из мономерных иммуногенных агентов.
В дополнительном варианте иммуногенный пептид можно представлять в форме вирусной или бактериальной вакцины. Нуклеиновую кислоту, кодирующую иммуногенный пептид, включают в геном или эписому вируса или бактерии. Необязательно нуклеиновую кислоту включают таким образом, чтобы иммуногенный пептид экспрессировался как секретированный белок или как слитый белок с белком внешней поверхности вируса или трансмембранным белком бактерий, так что пептид является экспонированным. Вирусы или бактерии, применяемые в таких способах, должны являться непатогенными или ослабленными. Пригодные вирусы включают аденовирус, HSV, вирус осповакцины и оспы птиц. Слияние иммуногенного пептида с HBsAg HBV является особенно пригодным.
Можно также проводить скрининг случайных библиотек пептидов или других соединений по пригодности. Комбинаторные библиотеки можно получать для множества типов соединений, которые можно синтезировать ступенчатым способом. Такие соединения включают полипептиды, миметики бета-поворота, полисахариды, фосфолипиды, гормоны, простагландины, стероиды, ароматические соединения, гетероциклические соединения, бензодиазепины, олигомерные N-замещенные глицины и олигокарбаматы. Большие комбинаторные библиотеки соединений можно конструировать способом кодируемых синтетических библиотек (ESL), описанных в Affymax, WO 95/12608, Affymax, WO 93/06121, Columbia University, WO 94/08051, Pharmacopeia, WO 95/35503 и Scripps, WO 95/30642 (содержание каждого из которых приведено в качестве ссылки для всех целей). Пептидные библиотеки можно также получать способами фагового дисплея. Смотри, например, Devlin, WO 91/18980.
Проводят первоначальный скрининг комбинаторных библиотек и других соединений по пригодности посредством определения их способности связывать антитела или лимфоциты (B или T), известные как специфические для формирующих споры пептидов. Например, первоначальный скрининг можно проводить с любой поликлональной сывороткой или с моноклональным антителом против формирующего споры пептида.
Соединения, идентифицированные такими скринингами, затем анализируют по способности индуцировать антитела или лимфоциты, узнающие формирующий споры пептид. Например, множественные разведения сыворотки можно тестировать в микропланшетах для титрования, предварительно покрытых формирующим споры пептидом, и общепринятый ELISA можно проводить для тестирования антител, узнающих формирующий споры пептид. Затем соединения можно тестировать по профилактической и терапевтической эффективности в моделях на животных.
Лекарственные средства по изобретению включают также антитела, специфически связывающие формирующий споры пептид. Такие антитела могут являться моноклональными или поликлональными. Продукцию не относящихся к человеку моноклональных антител, например, мыши или крысы, можно осуществлять, например, посредством иммунизации животного формирующим споры пептидом. Смотри Harlow & Lane, Antibodies, A Laboratory Manual (CSHP NY, 1988) (приведенный в качестве ссылки для всех целей). Такой иммуноген можно получать из природного источника, посредством синтеза пептидов или посредством рекомбинантной экспрессии.
Гуманизированные формы мышиных антител можно получать соединением областей CDR не относящихся к человеку антител с человеческими константными областями посредством способов рекомбинантной ДНК. Смотри Queen et al., Proc. Natl. Acad. Sci. USA 86,10029-10033 (1989) и WO 90/07861 (приведен в качестве ссылки для всех целей).
Человеческие антитела можно получать с использованием способов фагового дисплея. Смотри, например, Dower et al., WO 91/17271; McCafferty et al., WO 92/01047. В этих способах получают библиотеки фагов, члены которых экспонируют различные антитела на своих внешних поверхностях. Антитела обычно экспонированы в виде Fv или Fab фрагментов. Экспонированные на фагах антитела с желательной специфичностью отбирают по обогащению аффинности против формирующего споры пептида или его фрагментов. Человеческие антитела против формирующего споры пептида можно также получать из не относящихся к человеку трансгенных млекопитающих, обладающих трансгеном, кодирующим по меньшей мере фрагмент локуса иммуноглобулинов человека, и инактивированным эндогенным локусом иммуноглобулинов. Смотри, например, Lonberg et al., W093/12227 (1993); Kucherlapati, WO 91/10741 (1991) (полное содержание каждого из которых приведено в настоящем документе в качестве ссылок для всех целей). В экспериментах по конкурентному связыванию, или иным образом, можно отбирать человеческие антитела, обладающие такой же эпитопной специфичностью, как и конкретное мышиное антитело. Такие антитела, вероятно, разделяют полезные функциональные свойства мышиных антител.
Человеческие поликлональные антитела можно также предоставлять в форме сыворотки от людей, иммунизированных иммуногенным средством.
Необязательно такие поликлональные антитела можно концентрировать аффинной очисткой с использованием формирующего споры пептида в качестве аффинного реагента.
Человеческие или гуманизированные антитела можно конструировать, чтобы они обладали константными областями IgG, IgD, IgA и IgE и любым изотипом, включая IgG1, IgG2, IgG3 и IgG4. Антитела можно экспрессировать в форме тетрамеров, содержащих две легкие и две тяжелые цепи, в форме отдельных тяжелых цепей, легких цепей, в форме Fab, Fab'F (ab')2 и Fv, или в форме одноцепочечных антител, в которых вариабельные домены тяжелой и легкой цепи соединены посредством спейсера.
Лекарственные средства для применения в настоящих способах включают также T-клетки, связывающие формирующий споры пептид. Например, T-клетки можно активировать против формирующего споры пептида посредством экспрессии гена MHC класса I человека и гена β-2-микроглобулина человека из линии клеток насекомых, в результате чего пустой комплекс формируется на поверхности клеток и может связывать формирующий споры пептид. T-клетки, контактирующие с линией клеток, становятся специфически активированными против пептида. Смотри Peterson et al., US 5314813. Линии клеток насекомых, экспрессирующие антиген MHC класса II, можно сходным образом использовать для активации CD4 T-клеток.
Некоторые средства для индукции иммунного ответа содержат подходящий эпитоп для индукции иммунного ответа против пептида оболочки споры, но являются слишком маленькими, чтобы являться иммуногенными. В этой ситуации пептидный иммуноген можно связывать с подходящим носителем, чтобы помочь вызывать иммунный ответ. Пригодные носители включают сывороточные альбумины, гемоцианин морского блюдечка, молекулы иммуноглобулинов, тиреоглобулин, овальбумин, токсоид столбняка или токсоид из других патогенных бактерий, таких как возбудитель дифтерии, E. coli, возбудитель холеры или H. pylori, или ослабленное производное токсина. Другие носители для стимуляции или усиления иммунного ответа включают цитокины, такие как IL-1, IL-1a и пептиды, IL-2, zINF, IL-10, GM-CSF и хемокины, такие как MIP1a и 0 и RANTES. Иммуногенные средства можно также связывать с пептидами, усиливающими транспорт через ткани, как описано в O'Mahony, WO 97/17613 и WO 97/17614.
Иммуногенные средства можно связывать с носителями посредством химической сшивки. Способы связывания иммуногена с носителем включают формирование дисульфидных связей с использованием N-сукцинимидил-3-(2-пиридилтио)пропионата (SPDP) и сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилата (SMCC) (если у пептида отсутствует сульфгидрильная группа, ее можно предоставить добавлением остатка цистеина). Эти реагенты создают дисульфидную связь между ними и пептидными цистеиновыми остатками в белке и амидную связь между ε-амино на лизине, или другими свободными аминогруппами в других аминокислотах. Множество таких дисульфид/амид-формирующих средств описаны в Immun. Rev. 62, 185 (1982). Другие бифункциональные связывающие средства формируют тиоэфирную, а не дисульфидную связь. Многие из этих формирующих тиоэфир средств являются коммерчески доступными и включают реакционно-способные эфиры 6-малеимидокапроновой кислоты, 2-бромуксусной кислоты и 2-йодуксусной кислоты, 4-(N-малеимидометил)циклогексан-1-карбоновой кислоты.
Карбоксильные группы можно активировать, объединяя их с сукцинимидом или l-гидроксил-2-нитро-4-сульфоновой кислотой, натриевой солью.
Иммуногенные пептиды можно также экспрессировать в форме слитых с носителями белков. Иммуногенный пептид можно присоединять к носителю на N-конце, на C-конце или во внутренней части. Необязательно, множественные повторы иммуногенного пептида могут присутствовать в слитом белке.
Иммунные ответы против формирующих споры пептидов можно также индуцировать введением нуклеиновых кислот, кодирующих формирующий споры пептид. Такие нуклеиновые кислоты могут представлять собой ДНК или РНК. Фрагмент нуклеиновой кислоты, кодирующий иммуноген, как правило, связывают с регуляторными элементами, такими как промотор и энхансер, позволяющими экспрессию фрагмента ДНК в намеченной клетке-мишени пациента. Для экспрессии в клетках крови, как это желательно для индукции иммунного ответа, промоторные и энхансерные элементы из генов легкой или тяжелой цепи иммуноглобулинов или главный промежуточный ранний промотор и энхансер CMV являются пригодными для управления экспрессией. Связанные регуляторные элементы и кодирующие последовательности часто клонируют в вектор.
Ряд вирусных векторных систем являются доступными, включая ретровирусные системы (смотри, например, Lawrie and Tumin, Cur. Opin. Genet. Develop. 3, 102-109 (1993)); аденовирусные векторы (смотри, например, Bett et al., J. Virol. 67, 5911 (1993)); аденоассоциированные вирусные векторы (смотри, например, Zhou et al., J. Exp. Med. 179, 1867 (1994)), вирусные векторы из семейства оспы, включая вирус осповакцины и вирусы оспы птиц, вирусные векторы из рода альфа-вирусов, такие как полученные из вируса Синдбис и вируса леса Семлики (смотри, например, Dubensky et al., J. Virol. 70, 508-519 (1996)), и вирусов папилломы (Ohe et al., Human Gene Therapy 6, 325-333 (1995); Woo et al., WO 94/12629 и Xiao & Brandsma, Nucleic Acids. Res. 24, 2630-2622 (1996)).
ДНК, кодирующую иммуноген, или вектор, содержащий ДНК, можно упаковывать в липосомы. Подходящие липиды и родственные аналоги описаны в US5208036, US5264618, US5279833 и US5283185. Векторы и ДНК, кодирующие иммуноген, можно также адсорбировать на конкретных носителях или связывать с конкретными носителями, примеры которых включают полиметилметакрилатные полимеры и полилактиды и сополимеры полилактида и полигликолида, смотри, например, McGee et al., J. Micro Encap. (1996).
Векторы для генотерапии или голую ДНК можно доставлять in vivo посредством введения индивидуальному пациенту, как правило, посредством системного введения (например, внутривенного, внутрибрюшинного, назального введения, введения в желудок, внутрикожного, внутримышечного, подкожного введения или интракраниальной инфузии) или местного введения (смотри, например, US 5399346). ДНК можно вводить также с использованием генной пушки. Смотри Xiao & Brandsma, выше. ДНК, кодирующую иммуноген, преципитируют на поверхности микроскопических частиц металла. Летящие микрочастицы усиливают ударной волной или расширяющимся газом гелием, и они проникают в ткани на глубину нескольких слоев клеток. Например, является доступным устройство для доставки генов AccelTM, изготовленное в Agacetus, Inc. Middleton WI. Альтернативно голую ДНК можно пропускать через кожу в кровоток просто нанесением пятен ДНК на кожу с химическим или механическим раздражением (смотри WO 95/05853).
Пациенты, подлежащие лечению, включают индивидуумов, подверженных риску заболевания, но не проявляющих симптомов, также как пациентов, в настоящее время проявляющих симптомы.
Таким образом, настоящие способы можно применять профилактически к общей популяции без какой-либо оценки риска для рассматриваемого пациента. Настоящие способы являются особенно применимыми для индивидуумов, действительно обладающих известным риском являться носителями инфекции микобактериями. Такие индивидуумы включают индивидуумов, обладающих ослабленной иммунной системой.
При профилактических применениях фармацевтические композиции или лекарственные средства вводят пациенту, чувствительному к конкретному заболеванию или иным образом подверженному риску конкретного заболевания, в количестве, достаточном для исключения или уменьшения риска или задержки дебюта заболевания. При терапевтических применениях композиции или лекарственные средства вводят пациенту, у которого подозревают такое заболевание, или уже страдающему таким заболеванием, в количестве, достаточном для излечения, или по меньшей мере, частичного ареста симптомов заболевания и его осложнений. Количество, адекватное для осуществления этого, определяют как терапевтически или фармацевтически эффективную дозу. Как в профилактическом, так и в терапевтическом режимах средства обычно вводят в нескольких дозах, пока не достигнут достаточного иммунного ответа. Как правило, иммунный ответ мониторируют, и повторные дозы вводят, если иммунный ответ начинает затухать.
Эффективные дозы композиций по настоящему изобретению для лечения вышеописанных состояний могут меняться в зависимости от многих различных факторов, включая способы введения, намеченный участок, физиологическое состояние пациента, является ли пациент человеком или животным, другие вводимые лекарственные средства, и является ли лечение профилактическим или терапевтическим. Как правило, пациент является человеком, но пациент может являться не относящимся к человеку животным, таким как бык. Дозы для лечения необходимо титровать для оптимизации безопасности и эффективности. Количество иммуногена зависит от того, вводят ли также адъювант, где более высокие дозы необходимы в отсутствие адъюванта. Количество иммуногена для введения может меняться от 1 мкг-500 мкг на пациента и, более обычно, 5-500 мкг на инъекцию для введения человеку. Иногда можно использовать более высокую дозу 0,5-5 мг на инъекцию. Как правило, приблизительно 10, 20, 50 или 100 мкг используют для каждой инъекции человеку. Сроки инъекций могут значительно меняться от одного раза в сутки, до одного раза в год, до одного раза в десять лет. В любые данные сутки, когда вводят дозу иммуногена, доза превышает 1 мкг/пациента и обычно превышает 10 мкг/пациента, если также вводят адъювант, и превышает 10 мкг/пациента и обычно превышает 100 мкг/пациента в отсутствие адъюванта. Типичный режим состоит в иммунизации с последующими бустер-инъекциями с 6-недельными интервалами. Другой режим состоит в иммунизации с последующими бустер-инъекциями через 1, 2 и 12 месяцев. Другой режим включает инъекцию каждые два месяца в течение жизни. Альтернативно бустер-инъекции могут происходить на регулярной или нерегулярной основе, как показано по мониторированию иммунного ответа.
Для пассивной иммунизации антителом доза может лежать в диапазоне приблизительно от 0,0001 до 100 мг/кг и более обычно 0,01-5 мг/кг массы тела хозяина. Дозы нуклеиновых кислот, кодирующих иммуногены, могут лежать в диапазоне приблизительно от 10 нг до 1 г, 100 нг-100 мг, 1 мкг-10 мг или 30-300 мкг ДНК на пациента. Дозы для инфекционных вирусных векторов могут меняться от 10-100 вирионов или более на дозу.
Средства для индукции иммунного ответа можно вводить парентеральными, местными, внутривенными, пероральными, подкожными, внутрибрюшинными, интраназальными или внутримышечными способами для профилактического и/или терапевтического лечения. Наиболее типичным способом введения является подкожный, хотя другие могут являться в равной степени эффективными. Следующей из наиболее общепринятых является внутримышечная инъекция. Этот тип инъекции наиболее обычно проводят в мускулы руки или ноги. Внутривенные инъекции, также как внутрибрюшинные инъекции, внутриартериальные, интракраниальные или внутрикожные инъекции, также являются эффективными для получения иммунного ответа. В некоторых способах средства инъецируют напрямую в конкретную ткань, где накапливаются депозиты.
Иммуногенные средства по изобретению, такие как пептиды, иногда вводят в сочетании с адъювантом. Множество адъювантов можно использовать в сочетании с формирующим споры пептидом, чтобы вызывать иммунный ответ. Предпочтительные адъюванты усиливают истинный ответ на иммуноген, не вызывая конформационных изменений иммуногена, влияющих на качественную форму ответа. Предпочтительные адъюванты включают квасцы, 3-де-O-ацилированный монофосфориллипид A (MPL) (смотри GB 2220211). QS21 представляет собой тритерпеновый гликозид или сапонин, выделенный из коры дерева Quillaja Saponaria Molina, обнаруженного в Южной Америке (смотри Kensil et al., in Vaccine Design: The Subunit and Ajuvant Approach (eds. Powell & Newman, Plenum Press, NY, 1995); патент США № 5057540). Формирующие споры пептиды по изобретению сами по себе являются гидрофобными, и их также можно использовать в качестве адъювантов.
Другие адъюванты представляют собой эмульсии масло-в-воде (такие как сквален или арахисовое масло), необязательно в сочетании с иммуностимуляторами, такими как монофосфориллипид A (смотри Stoute et al., N. Engl. J. Med. 336, 86-91 (1997)). Другой адъювант представляет собой CpG (Bioworld Today, Nov. 15, 1998). Альтернативно формирующий споры пептид можно присоединять к адъюванту. Например, липопептидный вариант формирующего споры пептида можно получать присоединением пальмитиновой кислоты или других липидов непосредственно на N-конце формирующего споры пептида, как описано для вакцинации антигеном гепатита B (Livingston, J. Immunol. 159, 1383-1392 (1997)). Однако такое присоединение не должно по существу изменять конформацию формирующего споры пептида, так чтобы не влиять на природу иммунного ответа на него. Адъюванты можно вводить в качестве компонента терапевтической композиции с активным средством, или можно вводить отдельно, до, во время или после введения лекарственного средства.
Предпочтительным классом адъювантов являются соли алюминия (квасцы), такие как гидроксид алюминия, фосфат алюминия, сульфат алюминия. Такие адъюванты можно использовать в присутствии или в отсутствие других иммуностимулирующих средств, таких как MPL или 3-DMP, QS21, полимерные или мономерные аминокислоты, такие как полиглутаминовая кислота или полилизин. Другой класс адъювантов представляет собой эмульсионные составы масло-в-воде. Такие адъюванты можно использовать в присутствии или в отсутствие других специфических иммуностимулирующих средств, таких как мурамилпептиды (например, N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетил-нормурамил-L-аланил-D- изоглутамин (nor-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)-этиламин (MTP-PE), N-ацетилглюкозаминил-N-ацетилмурамил-L-Al-D-изоglu-L-Ala-дипальмитоксипропиламид (DTP-DPP) терамидTM), или другие компоненты клеточной стенки бактерий. Эмульсии масло-в-воде включают (a) MF59 (WO 90/14837), содержащую 5% сквален, 0,5% Tween 80 и 0,5% Span 85 (необязательно содержащие различные количества MTP-PE), составленные в субмикронные частицы с использованием микрофлюидизатора, такого как микрофлюидизатор модели HOY (Microfluidics, Newton MA), (b) SAF, содержащие 10% сквалан, 0,4% Tween 80,5% плюроновый блокполимер L121 и thr-MDP, или микрофлюидизированные в субмикронную эмульсию или обработанные на вортексе для получения эмульсии с более крупным размером частиц, и (c) систему адъюванта Ribi (RAS), (Ribi Immunochem, Hamilton, MT), содержащую 2% сквален, 0,2% Tween 80 и один или несколько компонентов клеточной стенки бактерий из группы, состоящей из монофосфориллипида A (MPL), димиколата трегалозы (TDM) и каркаса клеточной стенки (CWS), предпочтительно MPL + CWS (Detox).
Другим классом предпочтительных адъювантов являются сапониновые адъюванты, такие как Stimulons (QS21, Aquila, Worcester, MA) или полученные из них частицы, такие как ISCOM (иммуностимулирующие комплексы) и ISCOMATRIX. Другие адъюванты включают полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (IFA). Другие адъюванты включают цитокины, такие как интерлейкины (IL-1, IL-2 и IL-12), макрофагальный колониестимулирующий фактор (M-CSF), фактор некроза опухоли (TNF).
Адъювант можно вводить с иммуногеном в форме одной композиции, или его можно вводить до, во время или после введения иммуногена. Иммуноген и адъювант можно упаковывать и поставлять в одном и том же флаконе, или их можно упаковывать в отдельные флаконы и смешивать перед использованием. Иммуноген и адъювант, как правило, упакованы с этикеткой, указывающей на предполагаемое терапевтическое применение. Если иммуноген и адъювант упакованы отдельно, упаковка, как правило, включает инструкции для смешивания перед использованием. Выбор адъюванта и/или носителя зависит от стабильности вакцины, содержащей адъювант, способа введения, расписания дозирования, эффективности адъюванта для видов, подлежащих вакцинации, и, для человека, фармацевтически приемлемый адъювант является таким, который одобрен или может быть одобрен для введения человеку соответствующими регулирующими органами. Например, полный адъювант Фрейнда не подходит для введения человеку. Квасцы, MPL и QS21 являются предпочтительными. Необязательно два или несколько различных адъювантов можно использовать одновременно. Предпочтительные сочетания включают квасцы с MPL, квасцы с QS21, MPL с QS21, и квасцы, QS21 и MPL вместе. Можно также использовать неполный адъювант Фрейнда, (Chang et al, Advanced Drug Delivery Reviews 32, 173-186 (1998)), необязательно в сочетании с любыми квасцами, QS21 и MPL и все их сочетания.
Средства по изобретению можно вводить в форме фармацевтических композиций, содержащих активное лекарственное средство, т.е. и множество других фармацевтически приемлемых компонентов. Смотри Remington's Pharmaceutical Science (15th ed., Mack Publishing Company, Easton, Pennsylvania, 1980). Предпочтительная форма зависит от намеченного способа введения и терапевтического применения. Композиции могут также включать, в зависимости от желательного состава, фармацевтически приемлемые, нетоксичные носители или разбавители, определенные как носители, общепринятые для составления фармацевтической композиции для введения человеку или животному. Разбавитель выбирают так, чтобы не влиять на биологическую активность сочетания. Примеры таких разбавителей включают дистиллированную воду, физиологический фосфатно-солевой буфер, раствор Рингера, раствор декстрозы и раствор Хэнкса. Кроме того, фармацевтическая композиция или состав может включать также другие носители, адъюванты или нетоксичные, не терапевтические, неиммуногенные стабилизаторы и т.п. Однако некоторые реагенты, пригодные для введения животным, такие как полный адъювант Фрейнда, как правило, не включают в композиции для применения для человека.
Фармацевтические композиции могут включать также большие, медленно поддающиеся метаболизму макромолекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты и сополимеры (такие как функционализированные латексом сефароза, агароза, целлюлоза и т.п.), полимерные аминокислоты, сополимеры аминокислот и липидные агрегаты (такие как масляные капли или липосомы). Кроме того, эти носители могут функционировать как иммуностимулирующие средства (т.е. адъюванты).
Для парентерального введения средства по изобретению можно вводить в форме пригодных для инъекции доз раствора или суспензии вещества в физиологически приемлемом разбавителе с фармацевтическим носителем, который может представлять собой стерильную жидкость, такую как вода-масло, солевой раствор, глицерин или этанол.
Кроме того, вспомогательные вещества, такие как увлажняющие вещества или эмульгаторы, поверхностно-активные вещества, забуферивающие pH вещества и т.п., могут присутствовать в композициях. Другие компоненты фармацевтических композиций представляют собой компоненты, происходящие из нефти, животных, растений, или синтетического происхождения, например, арахисовое масло, соевое масло и минеральное масло. Как правило, гликоли, такие как пропиленгликоль или полиэтиленгликоль, являются предпочтительными жидкими носителями, в частности, для пригодных для инъекции растворов.
Как правило, композиции можно получать в форме пригодных для инъекции композиций, в форме либо жидких растворов, либо суспензий; можно получать также твердые формы, пригодные для растворения или суспендирования в жидких носителях перед инъекцией. Препарат можно также эмульгировать или инкапсулировать в липосомы или микрочастицы, такие как полилактид, полигликолид или сополимер для усиления адъювантного эффекта, как обсуждают выше (смотри Langer, Science 249, 1527 (1990) и Hanes, Advanced Drug Delivery Reviews 28, 97-119 (1997). Средства по изобретению можно вводить в форме инъекции-депо или препарата-имплантата, которые можно составлять таким образом, чтобы позволять замедленное или пульсирующее высвобождение активного ингредиента.
Дополнительные составы, пригодные для других способов введения, включают пероральный, интраназальный и пульмональный составы, суппозитории и чрескожные применения.
Для суппозиториев, связующие вещества и носители включают, например, полиалкиленгликоли или триглицериды; такие суппозитории можно составлять из смесей, содержащих активный ингредиент в диапазоне 0,5k-10k, предпочтительно, 1%-2%. Пероральные составы включают наполнители, такие как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлозы и карбонат магния фармацевтического качества. Эти композиции могут принимать форму растворов, суспензий, таблеток, пилюль, капсул, составов или порошков с замедленным высвобождением и содержать 10%-95% активного ингредиента, предпочтительно 25%-70%.
Местное применение может приводить к чрескожной или внутрикожной доставке. Местное введение можно облегчать совместным введением средства с токсином холеры или его детоксифицированными производными или субъединицами или другими сходными бактериальными токсинами (смотри Glenn et al., Nature 391, 851 (1998)). Совместное введение можно осуществлять с использованием компонентов в форме смеси или в форме связанных молекул, полученных химическим сшиванием или экспрессией в форме слитого белка.
Альтернативно чрескожную доставку можно осуществлять с использованием кожного пластыря или с использованием трансферосом (Paul et al., Eur. J. Immunol. 25, 3521-24 (1995); Cevc et al., Biochem. Biophys. Acta 1368, 201-15 (1998)).
Изобретение относится к способам детекции иммунного ответа против формирующего споры пептида у пациента, страдающего заболеванием, вызванным микобактериями, или чувствительного к нему. Способы являются, в частности, применимыми для мониторирования курса лечения, введенного пациенту. Способы можно использовать для мониторирования как терапевтического лечения проявляющих симптомы пациентов, так и профилактического лечения бессимптомных пациентов.
Некоторые способы включают определение фонового уровня иммунного ответа у пациента до введения дозы средства, и сравнение этого со значением для иммунного ответа после лечения. Значительное увеличение (т.е. больше, чем обычная граница ошибки эксперимента при повторных измерениях одного и того же образца, выраженная как одно стандартное отклонение от среднего таких измерений) величины иммунного ответа указывает на положительный исход лечения (т.е. на то, что введением средства вызывают или усиливают иммунный ответ). Если величина иммунного ответа не изменяется значительно или уменьшается, это указывает на отрицательный исход лечения. Как правило, для пациентов, подвергаемых исходному курсу терапии средством, ожидают увеличения иммунного ответа при последовательных дозированиях, которое, в конечном счете, достигает плато. Введение средства, как правило, продолжают пока иммунный ответ увеличивается.
Достижение плато является показателем того, что введение лекарственного средства можно прекратить или уменьшить его дозу или частоту.
В других способах контрольное значение (т.е. среднее и стандартное отклонение) иммунного ответа определяют для контрольной популяции. Как правило, индивидуумов в контрольной популяции не подвергают предварительному лечению. Измеренные значения иммунного ответа у пациента после введения лекарственного средства можно затем сравнивать с контрольным значением. Значительное увеличение по сравнению с контрольным значением (например, более одного стандартного отклонения от среднего) указывает на положительный исход лечения. Отсутствие значительного увеличения или уменьшение указывает на отрицательный исход лечения.
Введение средства, как правило, продолжают, пока иммунный ответ увеличивается относительно контрольного значения.
Как и ранее, достижение плато относительно контрольных значений является показателем того, что введение лекарственного средства можно прекратить, или уменьшить его дозу или частоту.
В других способах контрольное значение иммунного ответа (например, среднее и стандартное отклонение) можно определять для контрольной популяции индивидуумов, подвергавшихся лечению с помощью лекарственного средства, и иммунные ответы которых в ответ на лечение достигли плато. Измеренные значения иммунного ответа у пациента сравнивают с контрольным значением. Если измеренный уровень у пациента значительно не отличается (например, более одного стандартного отклонения) от контрольного значения, лечение можно прекратить. Если уровень у пациента значительно ниже контрольного значения, продолжение введения средства является обоснованным. Если уровень у пациента сохраняется ниже контрольного значения, тогда может являться показанным изменение режима лечения, например, применение другого адъюванта.
В других способах у пациента, не получающего лечения в настоящее время, но подвергавшегося предшествующему курсу лечения, мониторируют иммунный ответ для определения, необходимо ли возобновление лечения. Измеренное значение иммунного ответа у пациента можно сравнивать со значением иммунного ответа, ранее достигаемого у пациента после предшествующего курса лечения. Значительное уменьшение относительно предшествующего измерения (т.е. больше, чем обычная граница ошибки при повторных измерениях одного и того же образца) является показателем того, что лечение можно возобновить. Альтернативно значение, измеренное у пациента, можно сравнивать с контрольным значением (среднее плюс стандартное отклонение), определенным в популяции пациентов после проведения курса терапии.
Альтернативно значение, измеренное у пациента, можно сравнивать с контрольным значением в популяциях пациентов после профилактического лечения, остающихся свободными от симптомов заболевания, или в популяциях пациентов после терапевтического лечения, для которых показали улучшение характеристик заболевания. Во всех этих случаях значительное уменьшение относительно контрольного уровня (т.е. более стандартного отклонения) является показателем того, что лечение следует возобновить для пациента.
Образцы ткани для анализа, как правило, представляют собой кровь, плазму, сыворотку, слизь или спинномозговую жидкость от пациента. Образец анализируют по показателям иммунного ответа на любую форму формирующего споры пептида, как правило, SpoVK, CotSA, YrbC, SpoVE, Soj и SpoIIIE. Иммунный ответ можно определять по присутствию, например, антител или T-клеток, специфически связывающих такой пептид. Способы ELISA детекции антител, специфических для формирующего споры пептида, описаны в разделе Примеры. Способы детекции узнающих T-клеток описаны выше (смотри Определения).
Кроме того, изобретение относится к диагностическим наборам для осуществления диагностических способов, описанных выше. Такие диагностические способы включают детекцию присутствия микобактерий у пациентов, в продуктах питания и в окружающей среде, например на рыбных фермах или птицефермах. Как правило, наборы для такой диагностики или детекции содержат средство, специфически связывающее антитела против формирующего споры пептида или реагирующее с T-клетками, специфическими для формирующего споры пептида. Набор может также включать метку. Для детекции антител против формирующего споры пептида, метка, как правило, присутствует в форме меченых антиидиотипических антител. Для детекции антител средство можно поставлять предварительно связанным с твердой фазой, такой как лунки планшета для микротитрования. Для детекции реакционноспособных T-клеток, метку можно поставлять в форме 3H-тимидина для измерения пролиферативного ответа. Наборы также, как правило, содержат этикетку, предоставляющую указания для применения набора. Этикетка может также включать схему или другую систему соответствия для корреляции уровней измеряемой метки с уровнями антител против формирующего споры пептида или T-клеток, реагирующих на формирующий споры пептид. Термин «этикетка» относится к любому записанному или документированному материалу, приложенному к набору или другим образом сопровождающему набор в любое время в ходе его изготовления, транспорта, продажи или применения. Например, термин этикетка относится к рекламным листовкам и брошюрам, упаковочным материалам, инструкциям, аудио или видеокассетам, компьютерным дискам, также как к надписям, нанесенным непосредственно на наборы.
Связывающие молекулы, описанные в настоящем документе, можно использовать в качестве диагностических инструментов в области медицины и ветеринарии.
В итоге, изобретение относится к:
связывающей молекуле, способной связывать пептид, выбранный из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24 и 26, для применения в качестве лекарственного средства.
В одном варианте осуществления изобретения связывающая молекула предназначена для применения для лечения, диагностики или профилактики заболевания, вызванного микобактерией.
В одном варианте осуществления изобретения связывающая молекула предназначена для применения для лечения, диагностики или профилактики заболевания, вызванного микобактериями, где заболевание представляет собой туберкулез, лепру, болезнь Джона, болезнь Крона или заболевание, вызванное нетуберкулезной микобактерией.
Более того, изобретение относится к средству, эффективному для индукции иммуногенного ответа против формирующего споры пептида, или антитела против него, для применения в качестве лекарственного средства.
В одном варианте осуществления изобретения средство, эффективное для индукции иммуногенного ответа против формирующего споры пептида, или антитела против него, представляет собой пептид, выбранный из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24 и 26, или связывающую молекулу, способную связывать его.
В одном варианте осуществления изобретения средство, эффективное для индукции иммуногенного ответа против формирующего споры пептида, или антитела против него, предназначено для лечения или профилактики заболевания, вызванного микобактерией.
В одном варианте осуществления изобретения средство, эффективное для индукции иммуногенного ответа против формирующего споры пептида, или антитела против него, предназначено для лечения или профилактики заболевания, выбранного из туберкулеза, лепры, болезни Джона, болезни Крона или заболевания, вызванного нетуберкулезной микобактерией.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей средство, эффективное для индукции иммуногенного ответа против формирующего споры пептида или антитела против него у пациента, и фармацевтически приемлемый адъювант.
Более того, изобретение относится к фармацевтической композиции, согласно вышеописанному, где средство включает формирующий споры пептид или его активный фрагмент.
Более того, изобретение относится к фармацевтической композиции, как определено в настоящем документе выше, где средство выбрано из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24 и 26, или антитела против него.
Более того, изобретение относится к фармацевтической композиции, как определено в настоящем документе выше, где формирующий споры пептид выбран из группы, содержащей CotA, CotD, CotT, CotSA и YrbC.
Более того, изобретение относится к фармацевтической композиции согласно вышеописанному, для применения в качестве лекарственного средства.
В другом аспекте изобретение относится к фармацевтической композиции согласно вышеописанному, для лечения или профилактики заболевания, вызванного микобактерией.
В другом аспекте изобретение относится к способу профилактики или лечения микобактериальной инфекции у пациента, включающему введение средства, эффективного для индукции иммунного ответа против пептидного компонента оболочки споры микобактерии у млекопитающего.
Более того, изобретение относится к способу, согласно вышеописанному, где пациент представляет собой человека.
Более того, изобретение относится к способу согласно вышеописанному, где заболевание представляет собой туберкулез, лепру, болезнь Джона, болезнь Крона или заболевание, вызванное нетуберкулезной микобактерией.
Более того, изобретение относится к способу согласно вышеописанному, где средство содержит формирующий споры пептид или его активный фрагмент.
Более того, изобретение относится к способу согласно вышеописанному, где формирующий споры пептид выбран из группы, содержащей CotA, CotD, CotT, CotSA, YrbC.
Более того, изобретение относится к способу согласно вышеописанному, где иммунный ответ включает антитела, связывающие формирующий споры пептид.
В другом аспекте изобретение относится к набору, содержащему связывающую молекулу, способную связывать пептид, выбранный из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24 и 26, и инструкции для применения.
Набор можно использовать для детекции присутствия микобактерий в различных образцах, таких как образцы, взятые от животных, или из внешней среды, окружающей животных, например, рыбных ферм или птицеферм.
ПРИМЕРЫ
Штаммы, среды и условия роста
Mycobacterium marinum T CCUG 20998 (ATCC 927) выращивали при 30°C на чашках с агаром 7H10, дополненным 0,5% глицерином, 10% OADC (комплекс олеиновая кислота-альбумин-декстроза) и 0,01% циклогексимидом. Mycobacterium bovis BCG выращивали при 37°C на чашках с агаром 7H10, дополненным 0,5% Tween 80, 10% OADC и 0,01% циклогексимидом. Escherichia coli K12 штамм MG1655 (Maisnier-Patin, S., Dasgupta, S., Krabbe, M., Nordstrom, K. Conversion to bidirectional replication after unidirectional initiation from R1 plasmid origin integrated at oriC in Escherichia coli. Mol. Microbiol. 30, 1067-1079 (1998)) выращивали в среде M9, дополненной 0,2% глюкозой и 0,2% казаминокислотами при 37°C.
Проточная цитометрия
Клетки собирали в различные периоды роста, фиксировали, окрашивали и проводили обработку в проточном цитометре. Данные анализировали, как описано ранее (Maisnier-Patin, S., Dasgupta, S., Krabbe, M., Nordstrom, K. Conversion to bidirectional replication after unidirectional initiation from Rl plasmid origin integrated at oriC in Escherichia coli. Mol. Microbiol. 30, 1067-1079 (1998)). Кратко, клетки фиксировали в 70% (конечная концентрация) ледяном этаноле и сохраняли при 4°C. Для проточной цитометрии фиксированные клетки промывали свободным от частиц (пропущенным через нитроцеллюлозные фильтры 0,2 микрон) 10 мМ Tris-HCl (pH 7,9) буфер, дополненный 10 мМ MgCl2. Затем клетки ресуспендировали в 0,1 × объема такого же буфера. Промытые клетки окрашивали бромидом этидия (0,4 мкг/мл, конечная концентрация) и митрамицином A (16 мкг/мл, конечная концентрация) и сохраняли на льду в течение 30 мин перед проточной цитометрией (установка для анализа Compact A440, Apogee Flow Systems, на фиг. 10 использовали BioRad Bryte HS). По меньшей мере 20000 клеток подсчитывали для каждой проточной гистограммы.
Флуоресцентная и фазовоконтрастная микроскопия
Клетки собирали и фиксировали, как описано выше, промывали фосфатно-солевым буфером (PBS) и наносили пятнами на тонкий слой 1% агарозы в 0,9% солевом растворе, содержащем 0,5 мкг/мл 4,6-диамидино-2-фенилиндола (DAPI) на предметном стекле. Для всей микроскопии использовали микроскоп Zeiss (Axioplan 2) с камерой CCD (AxioCam, Zeiss), соединенной с компьютеризованной системой анализа изображения (программа: Axiovision 4.3). Фильтр, использованный для флуоресценции DAPI, представлял собой D360/40 (360±20 нм). Фазовые и флуоресцентные изображения совмещали с использованием программы Axiovision 4.3.
Выделение чистых спор
Чистые споры M. marinum выделяли, как описано в предшествующих способах, с небольшими изменениями (Lewis P.J., & Errington, J. Mol. Microbiol, 25, 945-954 (1997), Long S.K., & Williams, O. B. J. Bacteriol. 76, 332 (1958), Powers, EM., Appl. Microbiol. 16, 180-181 (1968)). Клетки объединяли и собирали, как описано выше, с чашек в возрасте двух недель (M. marinum) и шести месяцев (M. bovis BCG), промывали в 0,9% NaCl, и затем ресуспендировали в TE-буфере (10 мМ Tris-HCl pH 7,9 и 1 мМ ЭДТА). Бусины из оксида кремния (0,1 мм, Q-Biogene, Lysing matrix B) добавляли к клеткам и энергично встряхивали. Две суспензии сохраняли при 4°C в течение 5 минут, чтобы позволить осаждение стеклянных бусин. Супернатанты собирали и затем фильтровали через стерильный фильтр 2 микрон (Millipore). Фильтрат центрифугировали при 10000 об/мин в течение 10 минут при 4°C. Осадок ресуспендировали в TE-буфере и добавляли лизоцим до конечной концентрации 3 мг/мл и инкубировали при 37°C в течение 1 часа. Затем добавляли SDS до 6,25% (об./об., конечная концентрация) и инкубировали при 37°C в течение по меньшей мере 30 мин. Лизат центрифугировали при 10000 об/мин в течение 10 мин при 4°C. Осадок промывали 5 раз в стерильной воде при 4°C и, наконец, ресуспендировали в стерильной воде. Присутствие спор верифицировали микроскопически (фиг. 6 и 9).
Серийные разведения чистых спор для выделения отдельной споровой частицы
Идея серийного разведения для выделения отдельной споры основана на предшествующем сообщении (Lister, J., Transactions of the Pathological Society of London 29, 425-467 (1878)). Партию выделенных спор разделяли на две фракции; одну высевали на свежие чашки и через неделю роста подсчитывали колонии для оценки числа жизнеспособных споровых частиц в препарате спор. Это число составляло порядка 103/мл. Другую фракцию серийно разводили в 104 раз, так чтобы оставалось не более одной жизнеспособной споры в каждой пробирке. Полученные таким образом разведения спор высевали на чашки с 7H10 с необходимыми добавками (смотри выше), и чашки инкубировали при 30°C.
Сканирующая электронная микроскопия (SEM)
Образцы для SEM получали, как описано ранее (Lackner P. et al., Malar J. 5, 116 (2006)). Кратко, клетки выращивали и собирали, как описано выше, фиксировали 2,5% глутаральдегидом, промывали 3× в 0,1 M PBS с последующей фиксацией в 1% тетраоксиде осмия. Фиксированные клетки промывали 3× в 0,1 M PBS, дегидратировали в этаноле (последовательно в 50%, 70%, 90%, 95% и абсолютном этаноле), инъецировали через фильтры Millipore (2 микрон) и высушивали. Фильтры с образцами закрепляли на 12-мм столбиках Cambridge с использованием углеродной ленты и опыляли золотом. Исследования образцов проводили с использованием Zeiss Supra 35-VP Field Emission SEM при 4кВ.
Трансмиссионная электронная микроскопия (TEM)
Образцы для TEM получали, как описано ранее (Reynolds, E. S., 1963. J. Cell Biol. 17, 208-212). Кратко, клетки выращивали и собирали, как описано выше, фиксировали в 2,5% глутаральдегиде, промывали в буфере 0,1 M какодилате натрия pH 7,2, и фиксировали в 1% тетраоксиде осмия в буфере какодилате натрия. Фиксированные клетки дегидратировали в этаноле (последовательно в 70%, 95% и абсолютном этаноле), обрабатывали пропиленоксидом (промежуточный растворитель), инфильтрировали в смесь пропиленоксида и смолы (Epon) и затем погружали в чистую смесь смолы. За этим следовало отверждение при 60°C. Тонкие срезы по 50 нм получали с использованием ультратома LKB 2088 V (V=5), помещали в медную решетку с прорезями и окрашивали по отдельности 5% ацетатом уранила и 3% цитратом свинца. Изображения исследовали и фотографировали на TEM Hitachi H-7100 при 75 кВ.
Дифференциальное окрашивание спор и вегетативных клеток M. marinum
Это осуществляли, как описано ранее (Cappuccino, J.G., Sherman, N. Microbiology, a Laboratory Manual (Benjamin Cummings, San Fransisco, sixth edition, 2001)). Клетки выращивали, как описано выше, и собирали на разных стадиях роста. Клетки ресуспендировали в воде, фиксировали нагреванием на предметных стеклах, окрашивали при нагревании малахитовым зеленым (5% масс./об. в воде), промывали водой и затем подвергали контрастному окрашиванию сафранином (2,5% масс./об. в этаноле). После контрастного окрашивания образцы промывали водой, и клетки визуализировали с помощью микроскопа Olympus CH30RF200. Фотографии получали с помощью камеры FujiFilm FinePix 2400Z.
Обработка клеток влажностью-нагреванием
Это осуществляли, как описано ранее (Chakrabarty, K. et al., Infection and Immunity. 74, 4430-4438 (2006)). Клетки выращивали, как описано выше, и собирали в разные периоды времени во время роста, ресуспендировали в воде и разделяли на две равные аликвоты. Одну аликвоту подвергали нагреванию, при 65°C, в течение различных периодов времени и затем высевали на чашки с агаром 7H10 с необходимыми добавками (смотри выше), и выращивали при 30°C в течение 5 суток; другую содержали без воздействия нагревания и высевали в качестве контроля.


Выделение тотальной РНК из M. marinum
Тотальную РНК выделяли, как описано ранее (Raynaud, C. et al, Mol. Microbiol. 45, 203 (2002)). Клетки разрушали в 1 мл раствора тризола (Invitrogen) и 0,3 мл хлороформа с 0,1 мм бусинами из оксида кремния (Q-Biogene, Lysing matrix B) с использованием аппарата BIO101/Savant FastPrep FP120 (скорость 6,5, пульс 30 с, четыре цикла). РНК выделяли с помощью 300 мкл хлороформа. Через 10 мин центрифугирования при 13000g водную фазу повторно экстрагировали с помощью 300 мкл хлороформа и затем переносили в пробирку, содержащую 250 мкл изопропанола. Тотальную РНК преципитировали в течение ночи при 4°C и промывали 1 мл 75% этанола перед ресуспендированием в свободной от РНКазы воде. Контаминирующую ДНК удаляли расщеплением ДНКазой RQl (Ambion) согласно инструкциям производителя.
Дот-блоттинг РНК
Дот-блоттинг и гибридизацию с зондом РНК выполняли, как описано ранее (Sambrook, J., Fritsch, E.F., Maniatis, T. Molecular Cloning, a Laboratory Manual. (Cold Spring Harbor Laboratory Press, New York, third edition, 2001)). Обработанные ДНКазой образцы РНК, полученные из клеток M. marinum в возрасте 1, 3, 5 и 7 суток, денатурировали при 90°C в течение 5 мин и затем наносили пятнами (10 мкг в каждом пятне) на нейлоновые мембраны Hybond-N+ (Amersham Biosciences) и высушивали на воздухе. РНК иммобилизовали пришивкой ультрафиолетом с использованием аппарата для пришивки Bio-Rad, с последующей гибридизацией со специфическими олигонуклеотидами с меченным 32P-5'-концом при 42°C. Мембраны промывали, как описано ранее (Sambrook, J., Fritsch, E.F., Maniatis, T. Molecular Cloning, a Laboratory Manual (Cold Spring Harbor Laboratory Press, New York, third edition, 2001)). После гибридизации с зондами для мРНК, мембраны отмывали горячим 0,01% SDS и затем повторно гибридизовали с зондами для 5S рРНК, и промывали, как выше. Сигналы гибридизации анализировали с использованием аппарата для визуализации фосфора (ImageQuant, Molecular Dynamics). Образцы РНК E. coli K12 в стационарной фазе также наносили пятнами на те же самые мембраны и гибридизовали с теми же самыми олигонуклеотидами для расчета фоновых сигналов для каждого пятна, возникающих из-за неспецифического связывания, которые являлись очень низкими в условиях гибридизации. Фоновые значения вычитали из соответствующих значений сигналов для мРНК и 5S рРНК. Олигонуклеотидные зонды метили по 5'-концам с помощью γ32P-ATP с использованием T4 полинуклеотидкиназы (PNK) согласно общепринятым способам. Нуклеотидные последовательности олигонуклеотидных зондов перечислены в таблице 6. Для гибридизации с 5S рРНК M. marinum использовали олигонуклеотид 5'-GCTGACAGGCTTAGCTTCCG.
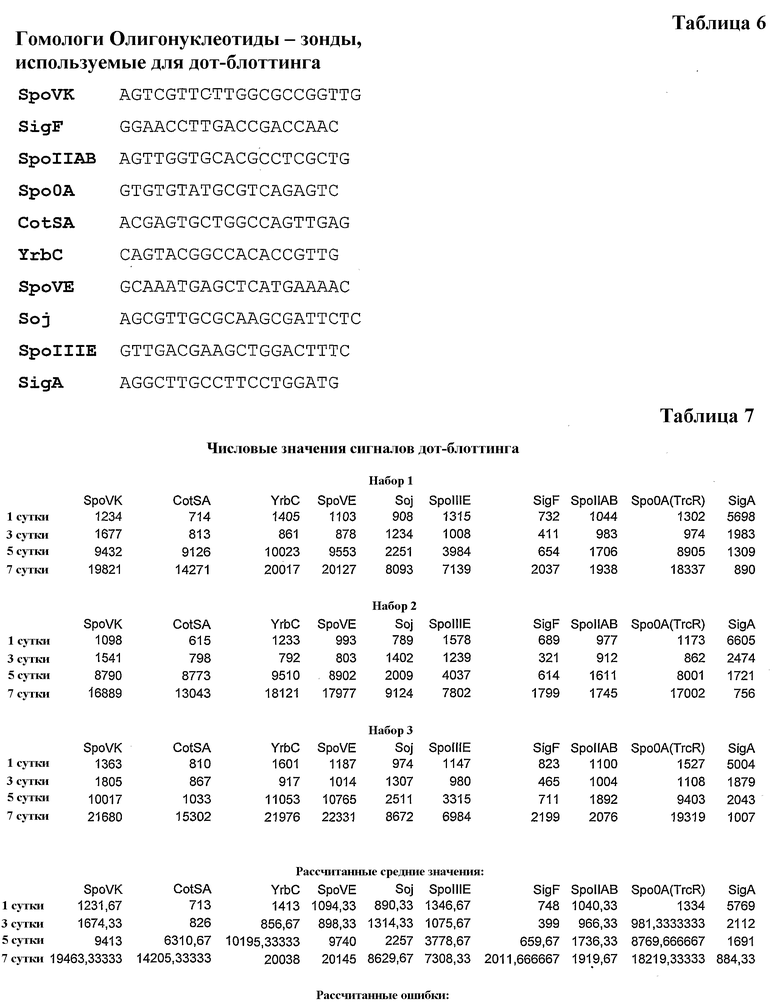
БИОИНФОРМАТИКА
Данные последовательности
Последовательности белков из Bacillus subtilis загружали с интернет-сервера SubtiList, http://genolist.pasteur.fr/SubtiList/ (Moszer, L, Glaser P., & Danchin, A., Microbiol. 141, 261-268 (1995), Moszer, L, FEBS Letters 430, 28-36 (1998)).
Специфические для споруляции белки получены согласно функциональной категории 1.8 в SubtiList. Кроме того, загружали также последовательности белков для связанных со споруляцией факторов SigE, SigF, SigG, SpoOA (TrcR на фиг. 5) и SpoVT. Аминокислотные последовательности идентифицированных открытых рамок считывания из M. tuberculosis CDC 1551, M. tuberculosis H37Rv и S. coelicolor A3 (2) загружали из TIGR Comprehensive Microbial Resource (TIGR-CMR, http://cmr.tigr.org/tigr-scripts/CMR/CmrHomePage.cgi) Peterson, J.D., Umayam, L.A., Dickinson, T.M., Hickey E.K., & White, O., Nucleic Acids Res. 29, 123-125 (2001), Cole S. T. et al,. Nature 393, 537-544 (1998), Fleischmann, R. D. Whole-Genome Comparison of Mycobacterium tuberculosis Clinical and Laboratory Strains. J. Bacteriol. 184, 5479-5490 (2002), http://cmr.tigr.org/tigr-scripts/CMR/CmrHomePage.cgi). Аминокислотные последовательности исходных предсказанных генов для M. marinum загружали с ftp-сервера Sanger Institute, ftp://ftp.sanger.ac.uk/pub/patogenes/mm/. Белки, вовлеченные в споруляцию S. coelicolor, получали по запросу в базе данных ScoDB (http://streptomyces.org.uk/sco/index.html) для генов с аннотациями, содержащими любое из слов «споруляция», «whiA», «whiB», «whiD», «whiE», «whiG», «whiH», «whiI» или «whiJ», также как для генов с названиями генов, совпадающими с «whi*» ('*' соответствует любой букве). Кроме того, включен ген S. coelicolor для sigF.
Поиск ортологов
Пакет программного обеспечения Washington University BLAST (WU-BLAST) (версия 2.0, дата выпуска 10 мая 2005 г., Gish, W. [1996-2004] http://blast.wustl.edu) использовали в способе двустороннего наилучшего совпадения (RBH) (Rivera, M. C, Jain, R., Moore, J. E. & Lake, J. A. Genomic evidence for two functionally distinct gene classes. Proc. Natl Acad. Sci. USA 95, 6239-6244 (1998)) для идентификации предположительных ортологов для связанных со споруляцией белков B. subtilis и S. coelicolor в видах микобактерий, перечисленных выше. Пару генов, a и b из геномов A и B, соответственно, принимали в качестве предположительных ортологов, если поиск с помощью a против B идентифицировал b как наиболее значительное совпадение, и поиск с помощью b против A приводил в результате к a, как к наиболее значительному совпадению. Использовали программу blastp с параметрами по умолчанию и отсечением ожидаемого при 0,001. Предположительные ортологи перечислены в таблицах 1-4. Кроме того, использовали совпадения с наибольшим количеством баллов для факторов транскрипции B. subtilis SpoOA, SpoIIAB и SigF. Эти совпадения не возвращали исходный запрашиваемый белок B. subtilis в качестве совпадения с наибольшим количеством баллов в поиске RBH, и таким образом, их рассматривали как слабые совпадения. В качестве дополнения к способу RBH, предположительные ортологи идентифицировали с помощью алгоритма двустороннего наименьшего расстояния (RSD), реализованного в пакете программного обеспечения, доступном для загрузки с http://cbi.med.harvard.edu/RSD.tar.gz (Wall, D. P., Fraser, H. B. & Hirsh, A. E. Detecting putative orthologs. Bioinformatics 19 (13), 1710-1711 (2003)). Программу запускали с установками по умолчанию, за исключением порога E-значения 0,001 и порога расхождения 0,5. Результаты перечислены в таблицах 1-4.
Измерение титров антител
Измерение титров антител у мышей можно проводить следующим образом. У мышей забирали кровь от четырех до семи суток после каждой иммунизации, начиная со второй иммунизации, всего пять заборов крови. Титры антител можно измерять как связывающее формирующий споры пептид антитело с использованием сэндвич-ELISA с помощью пластиковых мультилуночных планшетов, покрытых формирующим споры пептидом.
Зависимую от формирующего споры пептида лимфопролиферацию можно измерять с использованием клеток селезенки, собранных приблизительно через одну неделю после последней, шестой, иммунизации. Свежесобранные клетки, 105 на лунку, можно, например, культивировать в течение 5 суток в присутствии формирующего споры пептида в концентрации 5 мкм для стимуляции.
Получение поликлональных антител для пассивной защиты можно проводить следующим образом. Двадцать нетрансгенных мышей можно иммунизировать формирующим споры пептидом или другим иммуногеном, необязательно плюс адъювант, и затем подвергать эвтаназии через 4-5 месяцев. Затем у иммунизированных мышей можно забирать кровь. Необязательно IgG отделяют от других компонентов крови. Антитело, специфическое для иммуногена, можно частично очищать аффинной хроматографией. В среднем можно получать приблизительно 0,5-1 мг специфического для иммуногена антитела на мышь, получая всего 5-10 мг.
Пассивную иммунизацию антителами против формирующего споры пептида можно проводить следующим образом. В группах мышей в возрасте 7-9 месяцев каждой можно инъецировать в PBS 0,5 мг поликлональных антител против формирующего споры пептида или специфических моноклональных антител против формирующего споры пептида, как показано ниже. Все препараты антител следует очищать до низких уровней эндотоксинов. Моноклональные антитела против фрагмента можно получать инъекцией мыши фрагмента или более длинной формы формирующего споры пептида, получением гибридом и скринингом гибридом по антителу, специфически связывающему желательный фрагмент формирующего споры пептида без связывания с другими, не перекрывающимися, фрагментами формирующего споры пептида.
Мышам можно вводить внутрибрюшинные (i.p.) инъекции, по необходимости, в течение периода 4 месяцев для поддержания титра циркулирующего антитела, измеренного посредством ELIZA, более 1/1000, определенного посредством ELIZA против CotSA или другого иммуногена по этому изобретению. Титры можно мониторировать, как выше, и мышей можно подвергать эвтаназии в конце 4 месяцев инъекций.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2648791C2 |
| АКТИВНАЯ ИММУНИЗАЦИЯ ДЛЯ СОЗДАНИЯ АНТИТЕЛ К РАСТВОРИМОМУ А-БЕТА | 2004 |
|
RU2390350C2 |
| СПОСОБ БЫСТРОГО ОТБОРА ВАРИАНТОВ GP-120 ВИЧ | 2012 |
|
RU2603732C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2721274C2 |
| Иммуногенная композиция, включающая ее вакцина, набор для приготовления вышеуказанной композиции и способ лечения заболеваний, связанных с патологией секреции гастрина | 2014 |
|
RU2664197C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2385163C2 |
| КОМПОЗИЦИЯ МИКОБАКТЕРИАЛЬНЫХ АНТИГЕНОВ | 2011 |
|
RU2576007C2 |
| ИММУНОРЕГУЛЯТОРНАЯ ВАКЦИНА | 2013 |
|
RU2650709C2 |
| КОМПОЗИЦИИ И СПОСОБЫ WT1-СПЕЦИФИЧНОЙ ИММУНОТЕРАПИИ | 1999 |
|
RU2237674C2 |
| ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ (VLP) ИЗ ГЛИКОПРОТЕИНА ВИРУСА БЕШЕНСТВА | 2011 |
|
RU2575800C2 |






















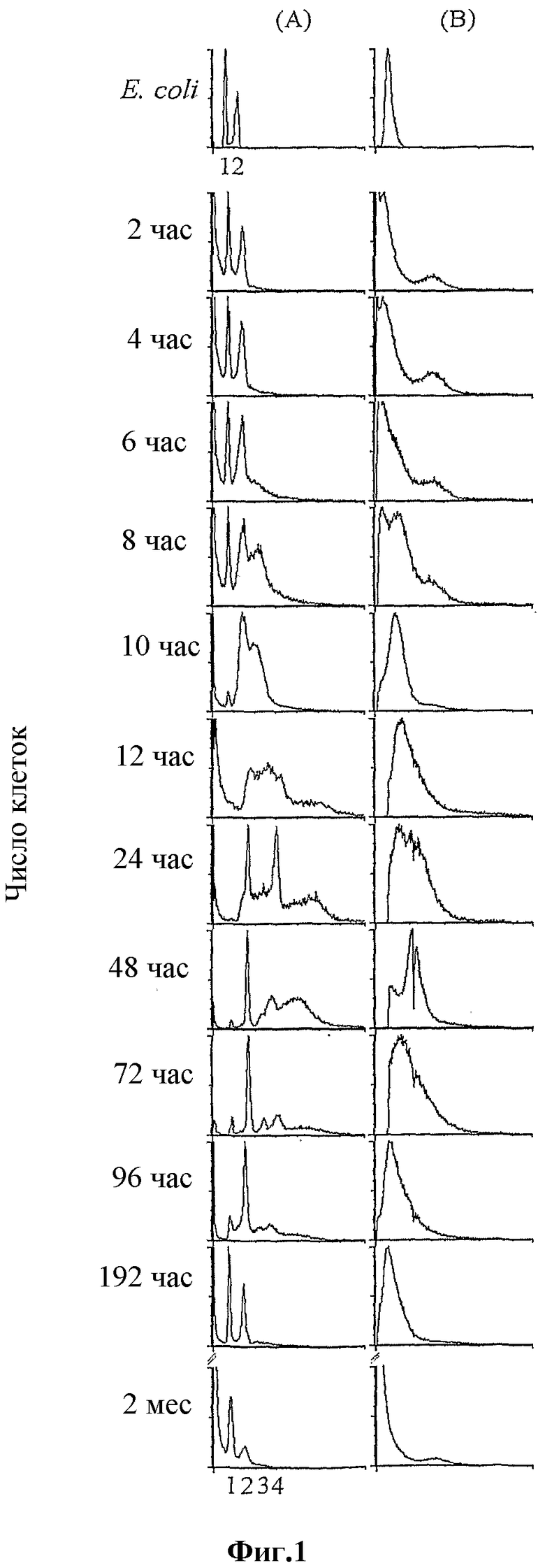
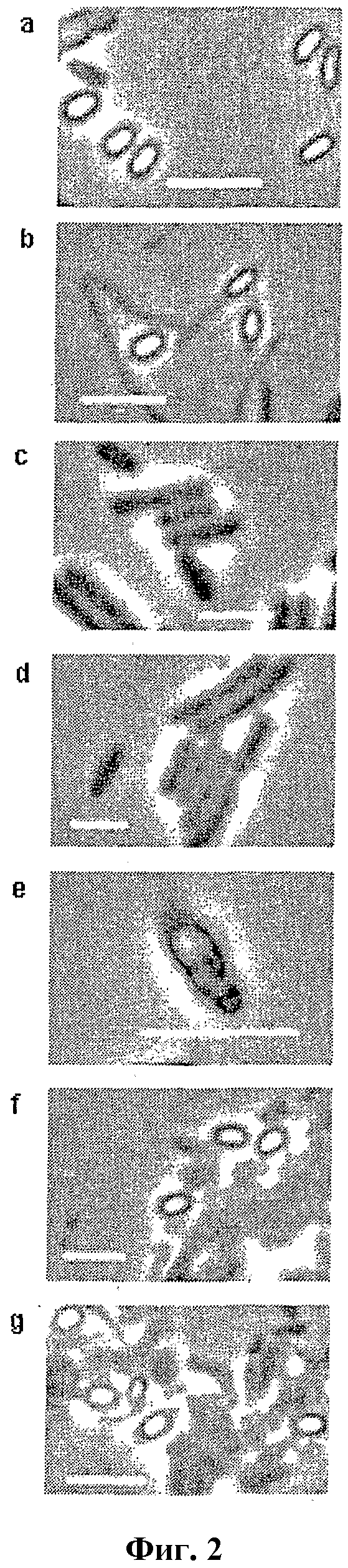
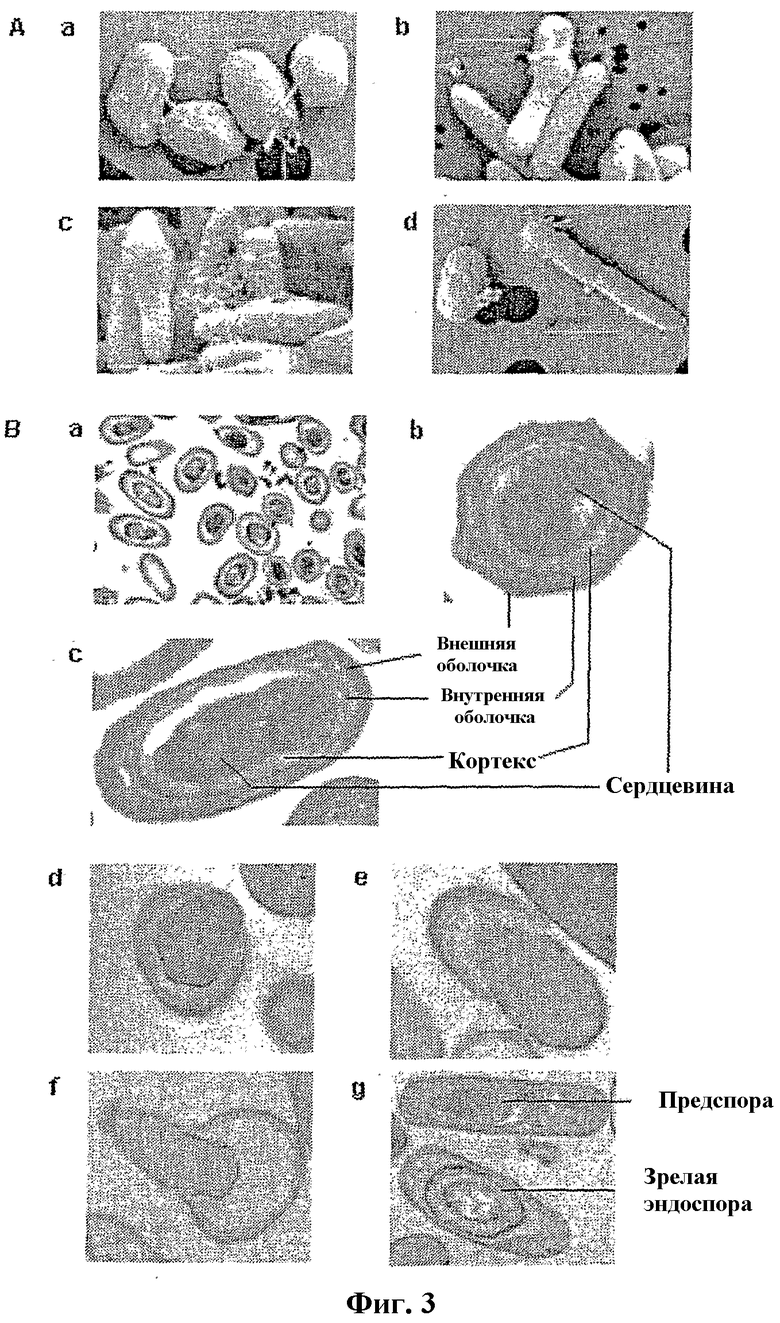
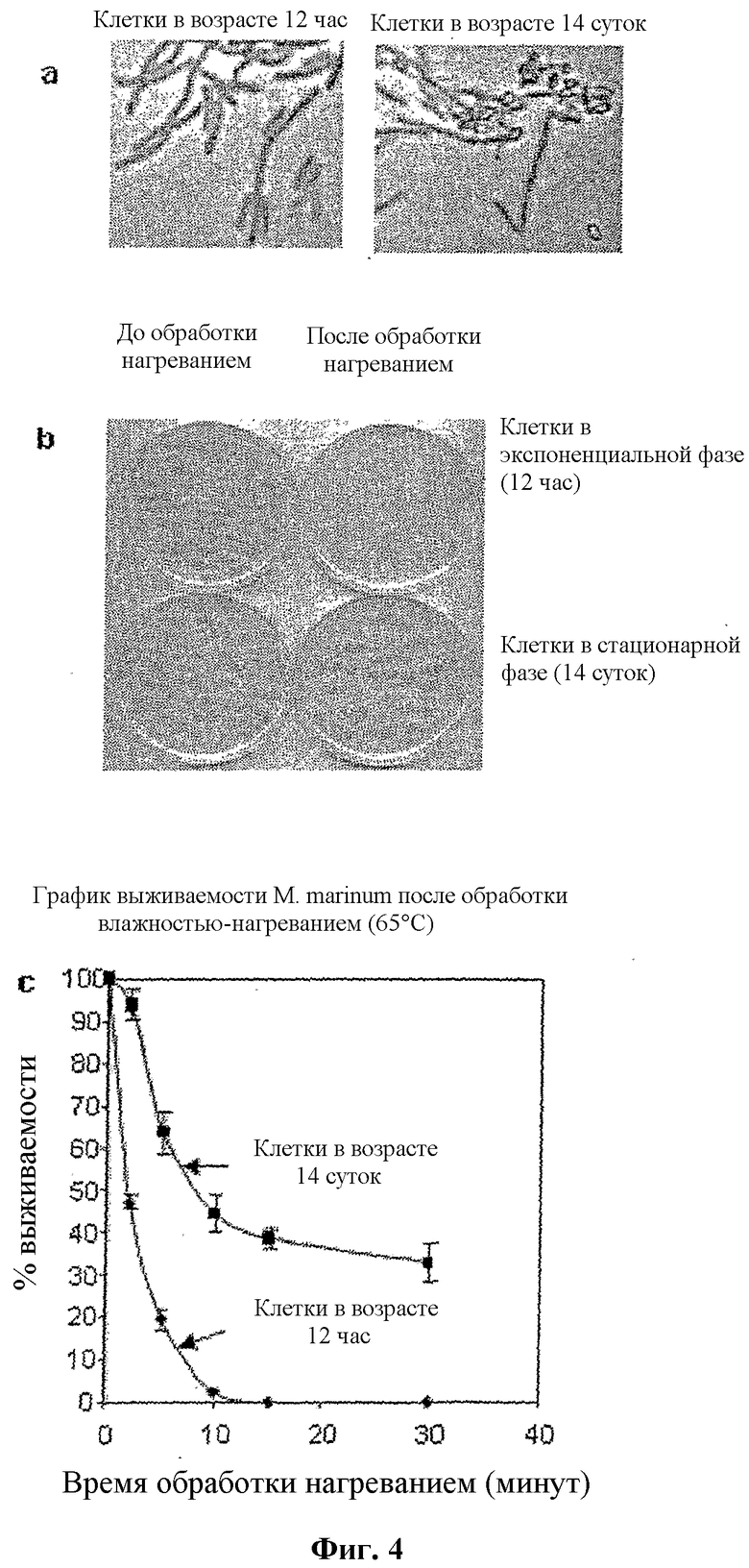
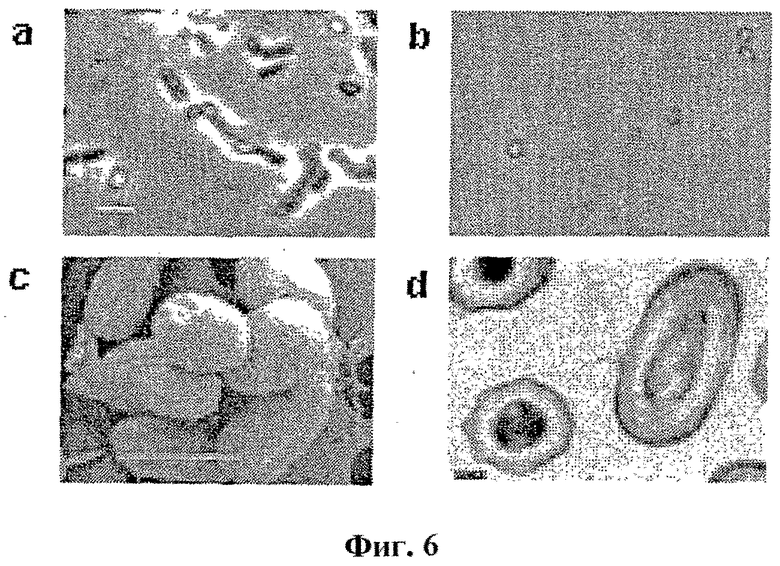


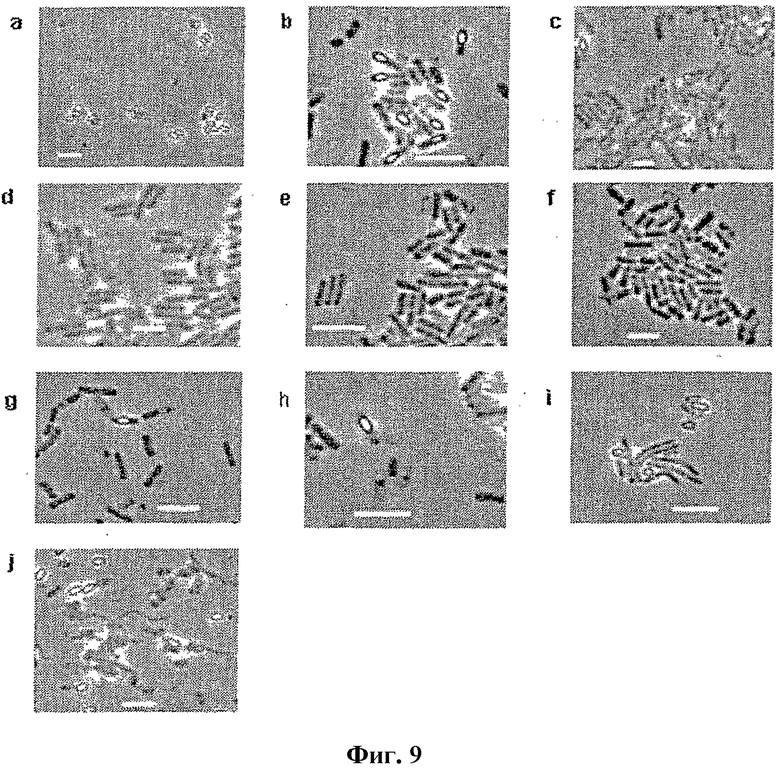
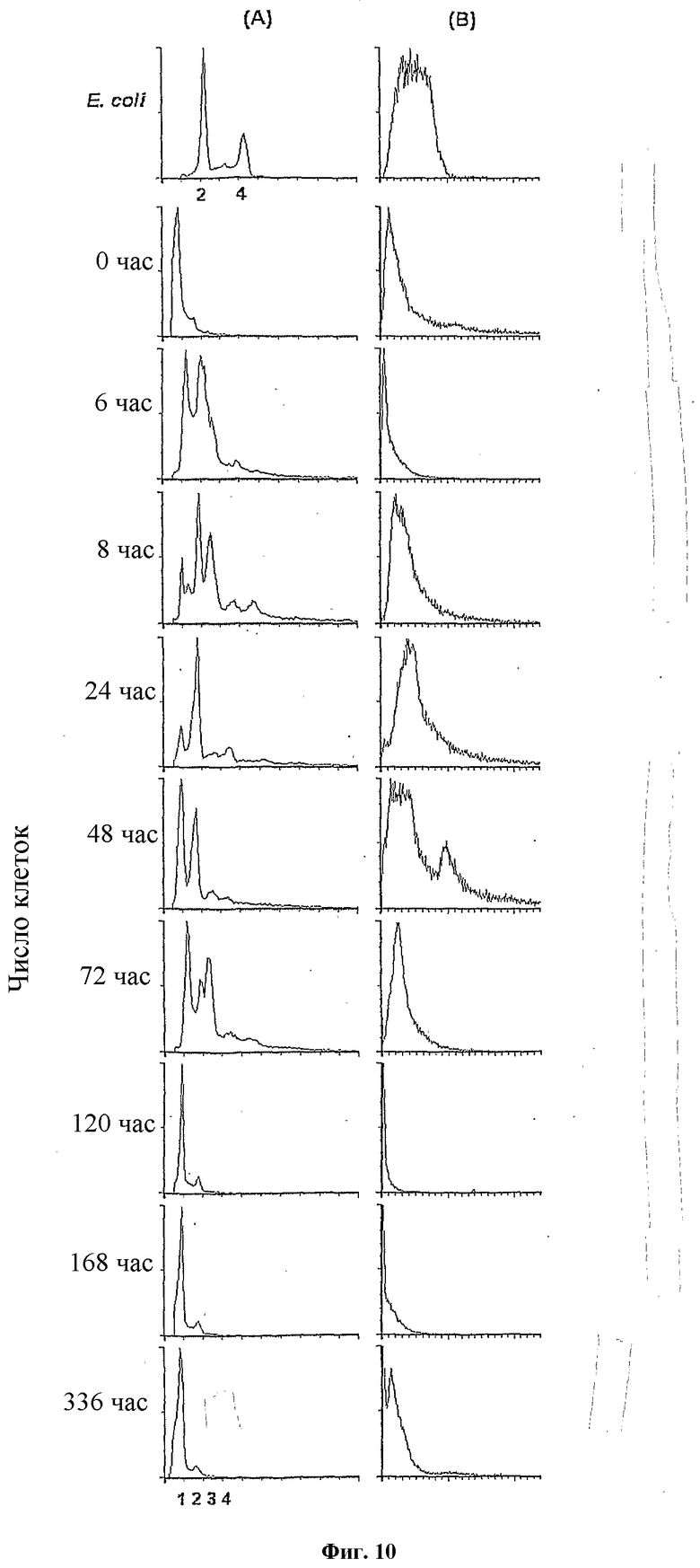
Изобретение относится к области микробиологии, а именно к набору для детекции спор, происходящих из микобактерий в образце. Набор содержит антитело, которое специфически связывает относящийся к спорам пептид из микобактерий, где относящийся к спорам пептид из микобактерий выбран из группы, состоящей из CotA, CotD, CotT, SpoVK, CotSA, YrbC, SpoVE, Soj, SpoIIIE и SEQ ID NO: 2, 4, 6, 8, 10, 12, 16, 18, 20, 22, 24 и 26, где указанное антитело получают иммунизацией человека или организма, не являющегося человеком, указанным относящимся к спорам пептидом. Набор дополнительно содержит молекулу, специфически связывающую комплекс между указанным антителом и указанным относящимся к спорам пептидом из микобактерий. Использование заявленного набора позволяет эффективно осуществить детекцию спор микобактерий в образцах из окружающей среды. 1 з.п. ф-лы, 7 табл., 10 ил., 1 пр.
1. Набор для детекции спор, происходящих из микобактерий, в образце, где набор содержит антитело, которое специфически связывает относящийся к спорам пептид из микобактерий, где относящийся к спорам пептид из микобактерий выбран из группы, состоящей из CotA, CotD, CotT, SpoVK, CotSA, YrbC, SpoVE, Soj, SpoIIIE, и SEQ ID NO: 2, 4, 6, 8, 10, 12, 16, 18, 20, 22, 24 и 26, где указанное антитело получают иммунизацией человека или организма, не являющегося человеком, указанным относящимся к спорам пептидом, причем набор дополнительно содержит молекулу, специфически связывающую комплекс между указанным антителом и указанным относящимся к спорам пептидом из микобактерий.
2. Набор по п. 1, где по меньшей мере одно из антитела и молекулы дополнительно содержит детектируемую группу.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Красильников А.П | |||
| Микробиологический словарь справочник | |||
| Минск | |||
| "АСАР" | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| Телефонно-осведомительный аппарат | 1921 |
|
SU306A1 |

