ОБЛАСТЬ ТЕХНИКИ
[0001] Данная заявка испрашивает приоритет на основании предварительной заявки на патент США № 61/357774, поданной 23 июня 2010 года, полное содержание которой включено в настоящее описание посредством ссылки.
[0002] Варианты реализации изобретения включают олигонуклеотиды, модулирующие экспрессию и/или функцию гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и связанных с ним молекул.
УРОВЕНЬ ТЕХНИКИ
[0003] Гибридизация ДНК-РНК и РНК-РНК важна для многих аспектов функционирования нуклеиновых кислот, включая репликацию ДНК, транскрипцию и трансляцию. Процесс гибридизации также является основным во множестве технологий, которые позволяют либо обнаруживать конкретную нуклеиновую кислоту, либо изменять ее экспрессию. Антисмысловые нуклеотиды, например, нарушают экспрессию генов за счет гибридизации с РНК-мишенью, таким образом препятствуя сплайсингу РНК, транскрипции, трансляции и репликации. Антисмысловая ДНК обладает дополнительной особенностью, заключающейся в том, что гибриды ДНК-РНК выступают в качестве субстрата для ферментативного расщепления рибонуклеазой Н, активность которой присутствует в большинстве типов клеток. Антисмысловые молекулы могут быть доставлены в клетки, как в случае для олигодезоксинуклеотидов (ОДН), или они могут представлять собой продукты экспрессии эндогенных генов в виде молекул РНК. Недавно Федеральное агентство по контролю за лекарственными средствами США (FDA) одобрило антисмысловое лекарственное средство, Витравен (VITRAVENETM) (для лечения цитомегаловирусного ретинита), что отражает возможность применения антисмысловых соединений для терапевтических целей.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0004] Настоящее Краткое описание предлагает краткое описание настоящего изобретения, предназначенное для того, чтобы в сжатой форме обозначить предмет и сущность изобретения. Краткое описание представляют с пониманием того, что его не следует применять для толкования или ограничения объема или смыслового содержания формулы изобретения.
[0005] В одном варианте реализации согласно изобретению предложены способы для ингибирования действия природного антисмыслового транскрипта путем применения антисмыслового олигонуклеотида (олигонуклеотидов), нацеленного на любую область природного антисмыслового транскрипта, приводящего к увеличению экспрессии соответствующего смыслового гена. Согласно настоящему изобретению также предполагается, что ингибирование природного антисмыслового транскрипта может быть достигнуто с помощью малых интерферирующих РНК (ммРНК), рибозимов и малых молекул, рассматриваемых в рамках настоящего изобретения.
[0006] Согласно одному варианту реализации предложен способ модулирования функции и/или экспрессии полинуклеотида SCNA в клетках или тканях пациента in vivo или in vitro, включающий осуществление контакта указанных клеток или тканей с антисмысловым олигонуклеотидом длиной от 5 до 30 нуклеотидов, при этом указанный олигонуклеотид имеет по меньшей мере 50 % идентичность последовательности с последовательностью, которая представляет собой обратный комплемент полинуклеотида, содержащего от 5 до 30 последовательных нуклеотидов в пределах нуклеотидов 1-1123, последовательности SEQ ID NO: 12, и 1-2352, последовательности SEQ ID NO:13, 1-267, последовательности SEQ ID NO: 14, 1-1080, последовательности SEQ ID NO: 15, 1-173, последовательности SEQ ID NO: 16, 1-618, последовательности SEQ ID NO: 17, 1-871, последовательности SEQ ID NO: 18, 1-304, последовательности SEQ ID NO: 19, 1-293, последовательности SEQ ID NO: 20, 1-892, последовательности SEQ ID NO: 21, 1-260, последовательности SEQ ID NO: 22, 1-982, последовательности SEQ ID NO: 23, 1-906, последовательности SEQ ID NO: 24, 1-476, последовательности SEQ ID NO: 25, 1-285, последовательности SEQ ID NO: 26, 1-162, последовательности SEQ ID NO: 27, и 1-94, последовательности SEQ ID NO: 28, что приводит к модулированию функции и/или экспрессии полинуклеотида SCNA в клетках или тканях пациента in vivo или in vitro.
[0007] В одном варианте реализации олигонуклеотид нацелен на природную антисмысловую последовательность для полинуклеотидов SCNA, например, на нуклеотиды, указанные в последовательностях, соответствующих SEQ ID NOS: 12-28, и любые их варианты, аллели, гомологи, мутанты, производные, фрагменты и последовательности, которые представляют собой их комплемент. Примеры антисмысловых олигонуклеотидов указаны в виде последовательностей, соответствующих SEQ ID NOS: 29-94.
[0008] Согласно другому варианту реализации предложен способ модулирования функции и/или экспрессии полинуклеотида SCNA в клетках или тканях пациента т vivo или in vitro, включающий осуществление контакта указанных клеток или тканей с антисмысловым олигонуклеотидом длиной от 5 до 30 нуклеотидов, при этом указанный олигонуклеотид имеет по меньшей мере 50% идентичность последовательности с последовательностью, которая представляет собой обратный комплемент антисмысловой последовательности для полинуклеотида SCNA; что приводит к модулированию функции и/или экспрессии полинуклеотида SCNA в клетках или тканях пациента in vivo или in vitro.
[0009] Согласно другому варианту реализации предложен способ модулирования функции и/или экспрессии полинуклеотида SCNA в клетках или тканях пациента in vivo или in vitro, включающий осуществление контакта указанных клеток или тканей с антисмысловым олигонуклеотидом длиной от 5 до 30 нуклеотидов, при этом указанный олигонуклеотид имеет по меньшей мере 50% идентичность последовательности с антисмысловым олигонуклеотидом для антисмыслового полинуклеотида гена SCNA; что приводит к модулированию функции и/или экспрессии полинуклеотида SCNA в клетках или тканях пациента in vivo или in vitro.
[0010] В одном варианте реализации композиция содержит один или более антисмысловых олигонуклеотидов, которые связываются со смысловыми и/или антисмысловыми полинуклеотидами гена SCNA, при этом указанные полинуклеотиды выбраны из группы, состоящей из SCNA-SCN12A и их вариантов. В предпочтительном варианте реализации полинуклеотид-мишень выбран из SCNA.
[0011] В одном варианте реализации указанные олигонуклеотиды содержат один или более модифицированных или замещенных нуклеотидов.
[0012] В другом предпочтительном варианте реализации указанные олигонуклеотиды содержат одну или более модифицированных связей.
[0013] В еще одном варианте реализации указанные модифицированные нуклеотиды содержат модифицированные основания, включая молекулы фосфотиоатных, метилфосфонатных производных нуклеиновых кислот, пептидо-нуклеиновых кислот, 2'-О-метил-, фтор- или углерод-, метилен- или других закрытых нуклеиновых кислот (ЗНК; locked nucleic acids, LNA). Предпочтительно, указанные модифицированные нуклеотиды представляют собой молекулы закрытых нуклеиновых кислот, включая ЗНК с α-L-рибо-конфигурацией (α-L-ЗНК).
[0014] В другом предпочтительном варианте реализации указанные олигонуклеотиды вводят пациенту подкожно, внутримышечно, внутривенно или внутрибрюшинно.
[0015] В другом предпочтительном варианте реализации указанные олигонуклеотиды вводят в составе фармацевтической композиции. Схема лечения включает по меньшей мере однократное введение указанных антисмысловых соединений пациенту; однако, это лечение может быть модифицировано до введения многократных доз в течение некоторого периода времени. Указанное лечение может быть объединено с одним или более другими типами терапии.
[0016] В другом предпочтительном варианте реализации указанные олигонуклеотиды инкапсулируют в липосому или присоединяют к молекуле-носителю (например, холестерину, ТАТ-пептиду).
[0017] Другие аспекты описаны ниже.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0018] Фигура 1 представляет собой графическое изображение результатов полимеразной цепной реакции (ПНР) в режиме реального времени, иллюстрирующее кратность изменения ± стандартное отклонение числа копий мРНК гена SCN1A после обработки клеток HepG2 фосфотиоатными олигонуклеотидами, вводимыми с применением агента для трансфекции Липофектамин 2000 (Lipofectamine 2000), по сравнению с контролем. Результаты ПЦР в режиме реального времени свидетельствуют о том, что уровни мРНК гена SCN1A в клетках HepG2 достоверно повышаются через 48 ч после обработки одним из антисмысловых олигонуклеотидов, нацеленным на антисмысловую последовательность BG724147 для гена SCN1A. Столбцы диаграммы, обозначенные как CUR-1624 - CUR-1627, соответствуют образцам, обработанным олигонуклеотидами SEQ ID NOS: 30-33, соответственно.
[0019] Фигура 2 представляет собой графическое изображение результатов ПЦР в режиме реального времени, иллюстрирующее кратность изменения ± стандартное отклонение числа копий мРНК гена SCN1A после обработки клеток HepG2 фосфотиоатными олигонуклеотидами, вводимыми с применением Липофектамина 2000, по сравнению с контролем. Столбцы диаграммы, обозначенные как CUR-1628 - CUR-1631, соответствуют образцам, обработанным олигонуклеотидами SEQ ID NOS: 34-37, соответственно.
[0020] Фигура 3 представляет собой графическое изображение результатов ПЦР в режиме реального времени, иллюстрирующее кратность изменения ± стандартное отклонение числа копий мРНК гена SCN1A после обработки клеток HepG2 фосфотиоатными олигонуклеотидами, вводимыми с применением Липофектамина 2000, по сравнению с контролем. Столбцы диаграммы, обозначенные как CUR-1632 - CUR-1636, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 38-42, соответственно.
[0021] Фигура 4 иллюстрирует дозозависимое увеличение экспрессии мРНК гена SCN1A в первичных фибробластах кожи человека, несущих мутацию, ассоциированную с синдромом Драве. Обозначения CUR-1916, CUR-1740, CUR-1764 и CUR-1770 соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 70, 45, 52 и 57, соответственно.
[0022] Фигура 5 иллюстрирует дозозависимое увеличение экспрессии мРНК гена SCN1A в клетках SK-N-AS. Обозначения CUR-1916, CUR-1740, CUR-1764 и CUR-1770 соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 70, 45, 52 и 57, соответственно.
[0023] Фигура 6 иллюстрирует дозозависимое увеличение экспрессии мРНК гена SCN1A в клетках Vero 76. Обозначения CUR-1916, CUR-1740, CUR-1764 и CUR-1770 соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 70, 45, 52 и 57, соответственно.
[0024] Фигура 7 иллюстрирует, что увеличение экспрессии мРНК гена SCN1A не вызвано неспецифической токсичностью антисмысловых олигонуклеотидов. А - увеличение экспрессии при обработке олигонуклеотидом CUR-1916; В - увеличение экспрессии при обработке олигонуклеотидом CUR-1770. Олигонуклеотид CUR-1462 представляет собой неактивный контрольный олигонуклеотид, характеризующийся сходной химической структурой.
[0025] Фигура 8 иллюстрирует, что обработка антисмысловыми олигонуклеотидами, нацеленными на природный антисмысловой транскрипт гена SCN1A, не оказывает существенного влияния на экспрессию каналов SCN8A и SCN9A в фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве. А - обработка олигонуклеотидом CUR-1770; В - обработка олигонуклеотидом CUR-1916.
[0026] Фигура 9 иллюстрирует стабильность антисмысловых олигонуклеотидов, нацеленных на SCN1A-специфичный природный антисмысловой транскрипт. Клетки Vero 76 обрабатывали в соответствии с описанием, приведенным в Примере 2, двумя разными партиями олигонуклеотида CUR-1916, синтезированного в августе 2010 года и марте 2011 года. Олигонуклеотид, синтезированный в августе 2010 года, хранили в виде водного раствора с концентрацией 1 мМ при температуре 4°С. Олигонуклеотид, синтезированный в марте 2011 года, был отправлен в лиофилизированной форме и проверен сразу после доставки.
[0027] Фигура 10 иллюстрирует увеличение экспрессии белка SCN1A в фибробластах, несущих мутацию, ассоциированную с синдромом Драве, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A. Фибробласты выращивали в 24-луночных планшетах и обрабатывали антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрации 20 нМ (панель с: CUR-1740; d: CUR-1770; e: CUR-1916) и 0 нМ (b). Клетки окрашивали на белок SCN1A (b-е) посредством непрямого иммуногистохимического метода с применением антител к белку SCN1A (Abeam cat#ab24820) и окрашивания/усиления сигнала вторичными антителами, основанного на взаимодействии авидин/биотин (Vector Laboratories cat#SP-2001; Vector Laboratories cat#PK-6101; Vector Laboratories cat#SK-4105); панель а - отрицательный контроль, кроличьи антимышиные антитела применяли в качестве первичных антител с той же последующей процедурой окрашивания, как и на панелях b-е.
[0028] Фигура 11 иллюстрирует увеличение экспрессии белка SCN1A в клетках SK-N-AS, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A. Клетки SK-N-AS выращивали в 24-луночных планшетах и обрабатывали олигонуклеотидами в концентрации 20 нМ (с: CUR-1740; d: CUR-1764; e: CUR-1770; f: CUR-1916) и (b: 0 нМ). Клетки SK-N-AS окрашивали на белок SCN1A (b-f) посредством непрямого иммуногистохимического метода с применением антител к белку SCN1A (Abeam cat#ab24820) и окрашивания/усиления сигнала вторичными антителами, основанного на взаимодействии авидин/биотин (Vector Laboratories cat#SP-2001; Vector Laboratories cat#PK-6101; Vector Laboratories cat#SK-4105); в качестве отрицательного контроля применяли кроличьи антимышиные антитела в качестве первичных антител с той же последующей процедурой окрашивания, как и на панелях b-f (панель а).
[0029] Фигура 12 иллюстрирует увеличение экспрессии белка SCN1A в клетках Vero 76, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A. Клетки Vero 76 выращивали в 24-луночных планшетах и обрабатывали антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрации 20 нМ (с: CUR-1740; d: CUR-1945; e: CUR-1770; f: CUR-1916; g: CUR-1924) и 0 нМ (b). Клетки Vero 76 окрашивали на белок SCN1A (b-g) посредством непрямого иммуногистохимического метода с применением антител к белку SCN1A (Abeam cat#ab24820) и окрашивания/усиления сигнала вторичными антителами, основанного на взаимодействии авидин/биотин (Vector Laboratories cat#SP-2001; Vector Laboratories cat# РК-6101; Vector Laboratories cat#SK-4105); панель а - в качестве отрицательного контроля применяли кроличьи антимышиные антитела в качестве первичных антител с той же последующей процедурой окрашивания, как и на панелях b-g.
[0030] Фигура 13 иллюстрирует, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, увеличивающие экспрессию мРНК гена SCN1A, не увеличивают экспрессию гена актина в клетках Vero 76. Те же самые антисмысловые олигонуклеотиды (CUR-1740, CUR-1838, CUR-1924), нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано в Примерах 5 и 12, увеличивают экспрессию мРНК и белка SCN1A, исследовали на их влияние на экспрессию мРНК гена бета-актина в клетках Vero 76. Данные эксперимента подтверждают, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, не увеличивают экспрессию неродственного гена, такого как ген актина. Столбцы диаграммы, обозначенные как CUR-1740, CUR-1838 и CUR-1924, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 45, 62 и 78, соответственно.
[0031] Фигура 14 иллюстрирует, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCN1A, не увеличивают экспрессию гена актина в фибробластах, несущих мутацию, ассоциированную с синдромом Драве. Олигонуклеотиды (CUR-1916, CUR-1945), нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано в Примерах 2 и 7, увеличивают экспрессию мРНК и белка SCN1A, исследовали на их влияние на экспрессию мРНК гена актина в фибробластах, несущих мутацию, ассоциированную с синдромом Драве. Приведенные ниже данные подтверждают, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, не увеличивают экспрессию неродственного гена, такого как ген актина. Столбцы диаграммы, обозначенные как CUR-1916 и CUR-1945, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 70 и 93, соответственно.
[0032] Фигура 15 иллюстрирует, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCN1A, не увеличивают экспрессию гена актина в клетках SK-N-AS. Те же самые антисмысловые олигонуклеотиды (CUR-1740, CUR-1770, CUR-1916, CUR-1764, CUR-1838), которые, как было показано в Примерах, увеличивают экспрессию мРНК и белка SCN1A, исследовали на их влияние на экспрессию мРНК гена актина в клетках SK-N-AS. Данные эксперимента подтверждают, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, не увеличивают экспрессию неродственного гена, такого как ген актина. Столбцы диаграммы, обозначенные как CUR-1740, CUR-1770, CUR-1916, CUR-1764, CUR-1838, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 45, 57, 70, 52 и 62, соответственно.
[0033] Фигура 16 иллюстрирует окрашивание белка актина в клетках SK-N-AS, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A. Клетки SK-N-AS выращивали в 24-луночных планшетах и обрабатывали олигонуклеотидами в концентрации 20 нМ (b: CUR-1740; с: CUR-1764; d: CUR-1770; e: CUR-1916) и 0 нМ (а). Клетки SK-N-AS окрашивали на актин (a-е) посредством непрямого иммуногистохимического метода с применением антител к актину (Abeam cat#ab1801) и окрашивания/усиления сигнала вторичными антителами, основанного на взаимодействии авидин/биотин (Vector Laboratories cat#SP-2001; Vector Laboratories cat#PK-6101; Vector Laboratories cat#SK-4105).
[0034] Фигура 17 иллюстрирует окрашивание белка актина в клетках Vero 76, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A. Клетки Vero 76 выращивали в 24-луночных планшетах и обрабатывали антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрации 20 нМ (b: CUR-1740; с: CUR-1770; d: CUR-1916; e: CUR-1924; f: CUR-1945) и 0 нМ (а). Клетки Vero 76 окрашивали на актин (b-f) посредством непрямого иммуногистохимического метода с применением антител к актину (Abeam cat#abl801) и окрашивания/усиления сигнала вторичными антителами, основанного на взаимодействии авидин/биотин (Vector Laboratories cat#SP-2001; Vector Laboratories cat# PK-6101; Vector Laboratories cat#SK-4105); панель а - отрицательный контроль, кроличьи антимышиные антитела применяли в качестве первичных антител с той же последующей процедурой окрашивания, как и на панелях b-g.
[0035] Фигура 18 иллюстрирует увеличение экспрессии белка актина в фибробластах, несущих мутацию, ассоциированную с синдромом Драве, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A. Фибробласты выращивали в 24-луночных планшетах и обрабатывали антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрации 20 нМ (панель b: CUR-1740; с: CUR-1764; d: CUR-1770; e: CUR-1838 и f: CUR-1916) и 0 нМ (а). Клетки окрашивали на актин (a-f) посредством непрямого иммуногистохимического метода с применением антител к актину (Abeam cat#ab1801) и окрашивания/усиления сигнала вторичными антителами, основанного на взаимодействии авидин/биотин (Vector Laboratories cat#SP-2001; Vector Laboratories cat#PK-6101; Vector Laboratories cat#SK-4105).
[0036] Фигура 19 иллюстрирует увеличение экспрессии белка SCN1A в фибробластах, несущих мутацию, ассоциированную с синдромом Драве, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, количественно определенное с применением иммуноферментного анализа (ИФА; enzyme-linked immunosorbent assay, ELISA). Фибробласты обрабатывали олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрациях 0 или 80 нМ. Через 48 ч клетки переносили в 96-луночный планшет на 24 ч перед фиксацией и исследованием с помощью ИФА-систем для определения белка SCN1A и актина. Показания оптической плотности (ОП) для сигнала белка SCN1A нормализовывали к сигналу актина для одних и тех же условий проведения эксперимента. Нормализованный сигнал белка SCN1A в клетках, обработанных олигонуклеотидами в концентрации 0 нМ, применяли в качестве точки отсчета (100%). Столбцы диаграммы, обозначенные как CUR-1740, CUR-1770 и CUR-1916, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 45, 57 и 70, соответственно.
[0037] Фигура 20 иллюстрирует увеличение экспрессии белка SCN1A в клетках Vero 76, обработанных антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, количественно определенное с применением ИФА. Клетки Vero 76 обрабатывали антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрациях 0 или 80 нМ. Через 48 ч клетки переносили в 96-луночный планшет на 24 ч перед фиксацией и исследованием с помощью ИФА-систем для определения белка SCN1A и актина. Показания ОП для сигнала белка SCN1A нормализовывали к сигналу актина для одних и тех же условий проведения эксперимента. Нормализованный сигнал белка SCN1A в клетках, обработанных олигонуклеотидами в концентрации 0 нМ, применяли в качестве точки отсчета (100 %). Столбцы диаграммы, обозначенные как CUR-1740, CUR-1770, CUR-1916, CUR-1924, CUR-1945, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 45, 57, 70, 78 и 93, соответственно.
[0038] Фигура 21 иллюстрирует увеличение экспрессии белка SCN1A в клетках SK-N-AS, обработанных олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, количественно определенное с применением ИФА. Клетки SK-N-AS обрабатывали антисмысловыми олигонуклеотидами, комплементарными природной антисмысловой последовательности для гена SCN1A, в концентрациях 0 или 20 нМ. Через 48 ч клетки переносили в 96-луночный планшет на 24 ч перед фиксацией и исследованием с помощью ИФА-систем для определения белка SCN1A и актина. Показания ОП для сигнала белка SCN1A нормализовывали к сигналу актина для одних и тех же условий проведения эксперимента. Нормализованный сигнал белка SCN1A в клетках, обработанных олигонуклеотидами в концентрации 0 нМ, применяли в качестве точки отсчета (100 %). Столбцы диаграммы, обозначенные как CUR-1740, CUR-1770, CUR-1924, CUR-1945, соответствуют образцам, обработанным олигонуклеотидами, соответствующими SEQ ID NOS: 45, 57, 78 и 93, соответственно.
[0039] На Фигуре 22 показаны продукты после второго раунда ПЦР с быстрой амплификацией 3'-концевого фрагмента кДНК (3'-rapid amplification of cDNA ends, 3'-RACE) природного антисмыслового транскрипта BG724147 гена SCN1A. 3'-RACE была проведена с применением: а) тотальной РНК из клеток HepG2 с добавлением аденозина; b - РНК, содержащей поли(А)-последовательность, выделенной из клеток HepG2; с - с применением тотальной РНК из первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве, с добавлением аденозина; d - с применением РНК, содержащей поли(А)-последовательность, выделенной из первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве. Фигура представляет собой негатив электрофореграммы, полученной в результате проведения электрофореза в 1% агарозном геле/lx буфере ТАЕ (смесь трис/уксусная кислота/ЭДТА) с окрашиванием геля красителем GelRed (GenScript, cat#M00120). Стрелки указывают на зоны, общие для клеток HepG2 и первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве, что свидетельствует о наличии природного антисмыслового транскрипта BG724147 в этих клетках.
[0040] Описание Перечня последовательностей - SEQ ID NO: 1: Альфа-субъединица потенциалзависимого натриевого канала типа I (SCN1A) человека, транскриптный вариант 1, мРНК (номер последовательности в базе данных NCBI (Национального центра биотехнологической информации США): NM_001165963); SEQ ID NO: 2: Альфа-субъединица потенциалзависимого натриевого канала типа II (SCN2A) человека, транскриптный вариант 1, мРНК (номер последовательности в базе данных NCBI: NM_021007); SEQ ID NO: 3: Альфа-субъединица потенциалзависимого натриевого канала типа III (SCN3A) человека, транскриптный вариант 1, мРНК (номер последовательности в базе данных NCBI: NM_006922); SEQ ID NO: 4: Альфа-субъединица потенциалзависимого натриевого канала типа IV (SCN4A) человека, мРНК (номер последовательности в базе данных NCBI: NM_000334); SEQ ID NO: 5: Альфа-субъединица потенциалзависимого натриевого канала типа V (SCN5A) человека, транскриптный вариант 1, мРНК (номер последовательности в базе данных NCBI: NM_198056); SEQ ID NO: 6: Альфа-субъединица потенциалзависимого натриевого канала типа VII (SCN7A) человека, мРНК (номер последовательности в базе данных NCBI: NM_002976); SEQ ID NO: 7: Альфа-субъединица потенциалзависимого натриевого канала типа VIII (SCN8A) человека, транскриптный вариант 1, мРНК (номер последовательности в базе данных NCBI: NM_014191); SEQ ID NO: 8: Альфа-субъединица потенциалзависимого натриевого канала типа IX (SCN9A) человека, мРНК (номер последовательности в базе данных NCBI: NM_002977); SEQ ID NO: 9: Альфа-субъединица потенциалзависимого натриевого канала типа Х (SCN10A) человека, мРНК (номер последовательности в базе данных NCBI: NM_006514); SEQ ID NO: 10: Альфа-субъединица потенциалзависимого натриевого канала типа XI (SCN11A) человека, мРНК (номер последовательности в базе данных NCBI: NM_014139); SEQ ID NO: 11: Альфа-субъединица потенциалзависимого натриевого канала SCN12A (SCN12A) человека, мРНК, полная кодирующая последовательность (номер последовательности в базе данных NCBI: AF109737); SEQ ID NO: 12: Природная антисмысловая последовательность для гена SCN1A (BG724147 расширенная); SEQ ID NO: 13: Природная антисмысловая последовательность для гена SCN1A (Hs.662210); SEQ ID NO: 14: Природная антисмысловая последовательность для гена SCN1A (АА383040); SEQ ID NO: 15: Природная антисмысловая последовательность для гена SCN1A (ВС029452); SEQ ID NO: 16: Природная антисмысловая последовательность для гена SCN1A (АА630035); SEQ ID NO: 17: Природная антисмысловая последовательность для гена SCN1A (ВЕ566126); SEQ ID NO: 18: Природная антисмысловая последовательность для гена SCN1A (BF673100); SEQ ID NO: 19: Природная антисмысловая последовательность для гена SCN1A (BG181807); SEQ ID NO: 20: Природная антисмысловая последовательность для гена SCN1A (BG183871); SEQ ID NO: 21: Природная антисмысловая последовательность для гена SCN1A (BG215777); SEQ ID NO:22: Природная антисмысловая последовательность для гена SCN1A (BG227970); SEQ ID NO: 23: Природная антисмысловая последовательность для гена SCN1A (ВМ905527); SEQ ID NO: 24: Природная антисмысловая последовательность для гена SCN1A (BU180772); SEQ ID NO: 25: Природная антисмысловая последовательность для гена SCN1A мыши (BG724147 ExtMouse); SEQ ID NO: 26: Природная антисмысловая последовательность для гена SCN1A мыши (Hs.662210mouseAS1); SEQ ID NO: 27: Природная антисмысловая последовательность для гена SCN1A мыши (Hs.662210mouseAS2); SEQ ID NO: 28: Природная антисмысловая последовательность для гена SCN1A мыши (Hs.662210mouseAS3); SEQ ID NOS: 29-94: Антисмысловые олигонуклеотиды. SEQ ID NO: 95 и 96 являются последовательностями, представляющими собой обратные комплементы антисмысловых олигонуклеотидов, соответствующих SEQ ID N0:58 и 59, соответственно. Символ "*" обозначает фосфотиоатную связь, "+" обозначает ЗНК, "r" обозначает РНК, а "m" указывает на наличие метильной группы на 2'-атоме кислорода в обозначенном сахарном остатке олигонуклеотида.
ПОДРОБНОЕ ОПИСАНИЕ
[0041] Некоторые аспекты изобретения описаны ниже со ссылкой на примеры применения для иллюстрации. Следует понимать, что многочисленные конкретные детали, связи и способы приведены с целью обеспечения полного понимания изобретения. Однако для специалиста в данной области техники будет очевидно, что изобретение может быть применено на практике без одной или более конкретных деталей или с применением других способов. Настоящее изобретение не ограничивается указанным порядком действий или процессов, так как некоторые действия могут быть осуществлены в разных порядках и/или одновременно с другими действиями или процессами. Кроме того, не все представленные в качестве примеров действия или процессы необходимы для реализации методологии в соответствии с настоящим изобретением.
[0042] Все гены, названия генов и продукты генов, описанные в настоящей заявке, соответствуют гомологам из организмов любых видов, для которых применимы композиции и способы, предложенные в настоящей заявке. Таким образом, данные термины включают, но не ограничиваются перечисленными, гены и генные продукты из организмов людей и мышей. Следует понимать, что, когда предложен ген или продукт гена из организма конкретного вида, этот элемент изобретения представлен исключительно в качестве примера, и его не следует истолковывать как ограничение, если контекст, в котором он появляется, ясно не предписывает иное. Соответственно, например, гены, описанные в настоящей заявке, которые в некоторых вариантах реализации относятся к нуклеотидным и аминокислотным последовательностям млекопитающих, охватывают гомологичные и/или ортологичные гены и продукты генов из организмов других животных, включая, но не ограничиваясь перечисленными, других млекопитающих, рыб, земноводных, пресмыкающихся и птиц. В предпочтительных вариантах реализации гены или нуклеотидные последовательности представляют собой гены или последовательности человека.
Определения
[0043] Терминология, применяемая в настоящей заявке, служит исключительно для описания конкретных вариантов реализации и не ограничивает изобретение. В настоящей заявке неопределенная и определенная формы единственного числа также включают множественные формы, если контекст ясно не предписывает иное. Кроме того, в той мере, в какой термины "включающий", "включает", "имеющий", "имеет", "с" или их варианты применяют в подробном описании и/или формуле изобретения, такие термины подразумевают включение в себя подобно термину "содержащий".
[0044] Термин "примерно" или "приблизительно" обозначает нахождение внутри диапазона приемлемой погрешности для конкретной величины, определенного специалистом в данной области техники, который будет зависеть отчасти от того, каким образом величину измеряют или определяют, т.е. от ограничений измерительной системы. Например, термин "примерно" может означать, что величина находится в пределах 1 или более чем 1 стандартного отклонения, как показывает практика в данной области техники. Альтернативно, термин "примерно" может означать диапазон погрешности, составляющий вплоть до 20%, предпочтительно вплоть до 10%, более предпочтительно вплоть до 5% и еще более предпочтительно вплоть до 1% от заданной величины. Альтернативно, применительно к биологическим системам или процессам, термин может означать нахождение в пределах порядка величины, предпочтительно в пределах пятикратного значения и более предпочтительно в пределах двукратного значения величины. В тех случаях, когда в заявке и формуле изобретения приведены конкретные значения, то, если не указано иное, следует считать, что термин "примерно" обозначает нахождение внутри диапазона приемлемой погрешности для конкретной величины.
[0045] В настоящей заявке термин "мРНК" обозначает известный на настоящий момент мРНК-транскрипт(ы) гена-мишени и любые дополнительные транскрипты, происхождение которых может быть объяснено.
[0046] С помощью терминов "антисмысловые олигонуклеотиды" или "антисмысловое соединение" обозначают молекулу РНК или ДНК, которая связывается с другой РНК или ДНК (РНК-, ДНК-мишенью). Например, если антисмысловое соединение представляет собой олигорибонуклеотид, оно связывается с другой РНК-мишенью за счет РНК-РНК-взаимодействий и изменяет активность РНК-мишени. Антисмысловой олигонуклеотид может увеличивать или подавлять экспрессию и/или функцию конкретного полинуклеотида. Определение включает любую чужеродную молекулу ДНК или РНК, подходящую с терапевтической, диагностической или другой точки зрения. Такие молекулы включают, например, молекулы антисмысловых ДНК или РНК, молекулы интерферирующих РНК (РНКи), микроРНК, ложных РНК, ммРНК, ферментативные РНК, РНК, применяемые в исправляющей терапии, и РНК, имеющие свойства агонистов и антагонистов, антисмысловые олигомерные соединения, антисмысловые олигонуклеотиды, олигонуклеотиды внешней направляющей последовательности (external guide sequence, EGS), сплайс-варианты, праймеры, зонды и другие олигомерные соединения, которые гибридизуются с по меньшей мере частью нуклеиновой кислоты-мишени. В связи с этим данные соединения могут быть введены в форме одноцепочечных, двухцепочечных, частично одноцепочечных или кольцевых олигомерных соединений.
[0047] В контексте настоящего изобретения термин "олигонуклеотид" относится к олигомеру или полимеру рибонуклеиновой кислоты (РНК) или дезоксирибонуклеиновой кислоты (ДНК) или их миметикам. Термин "олигонуклеотид" также включает линейные или кольцевые олигомеры природных и/или модифицированных мономеров или связей, включая дезоксирибонуклеозиды, рибонуклеозиды, их замещенные и альфа-аномерные формы, пептидо-нуклеиновые кислоты (ПНК), закрытые нуклеиновые кислоты (ЗНК; locked nucleic acids, LNA), фосфотиоатные, метилфосфонатные производные и т.п. Олигонуклеотиды способны к специфическому связыванию с полинуклеотидом-мишенью за счет регулярных взаимодействий между мономерными звеньями, такими как Уотсон-Криковский тип спаривания оснований, Хугстиновский или обратный Хугстиновский типы спаривания оснований и т.п.
[0048] Олигонуклеотид может быть "химерным", то есть составленным из разных областей. В контексте настоящего изобретения "химерные" соединения представляют собой олигонуклеотиды, которые содержат две или более химически отличные области, например, область (области) ДНК, область (области) РНК, область (области) ПНК и т.д. Каждая химически отличная область состоит из по меньшей мере одной мономерной единицы, т.е. нуклеотида в случае олигонуклеотидного соединения. Эти олигонуклеотиды, как правило, содержат по меньшей мере одну область, в которой олигонуклеотид модифицирован с целью проявления одного или более желаемых свойств. Желаемые свойства олигонуклеотида включают, но не ограничиваются перечисленными, например, повышенную устойчивость к деградации нуклеазами, повышенное клеточное поглощение и/или повышенную способность к связыванию с нуклеиновой кислотой-мишенью. Разные области олигонуклеотида могут, таким образом, обладать разными свойствами. Химерные олигонуклеотиды согласно настоящему изобретению могут быть образованы в виде смешанных структур из двух или более олигонуклеотидов, модифицированных олигонуклеотидов, олигонуклеозидов и/или аналогов олигонуклеотидов, описанных выше.
[0049] Олигонуклеотид может быть составлен из областей, которые могут быть соединены в "ряд", то есть когда мономеры соединены последовательно, как в нативной ДНК, или соединены посредством спейсеров. Спейсеры формируют ковалентный "мостик" между областями и имеют в предпочтительных случаях длину, не превышающую примерно 100 углеродных атомов. Спейсеры могут иметь разные функциональные характеристики, например, нести положительный или отрицательный заряд, обладать свойствами специфичного связывания нуклеиновых кислот (интеркаляторы, вещества, связывающиеся с бороздками, токсины, флуорофоры и т.д.), быть липофильными, индуцировать образование специфических вторичных структур, как, например, аланинсодержащие пептиды, которые индуцируют образование альфа-спиралей.
[0050] В настоящей заявке термины "ген SCN1A" и "альфа-субъединица потенциалзависимото натриевого канала типа I" включают все члены семейства, мутанты, аллели, фрагменты, виды, кодирующие и некодирующие последовательности, смысловые и антисмысловые полинуклеотидные цепи и т.д. Аналогично, гены SCN2A-SCN12A включают все мутанты, аллели, фрагменты и т.д.
[0051] В настоящем описании слова "альфа-субъединица потенциалзависимого натриевого канала типа I", SCN1A, FEB3, FEB3A, GEFSP2, HBSCI, NaCl, Nav1.1, SCN1, SMEI, альфа-субъединица белка натриевого канала мозга I, альфа-субъединица белка натриевого канала типа 1, альфа-субъединица белка натриевого канала типа I и альфа-субъединица потенциалзависимого натриевого канала Nav1.1 рассматривают как имеющие одно и то же значение в литературе и применяют взаимозаменяемо в настоящей заявке.
[0052] В настоящей заявке термин "олигонуклеотид, специфичный к" или "олигонуклеотид, нацеленный на" относится к олигонуклеотиду, содержащему последовательность, (i) способную образовывать стабильный комплекс с частью гена-мишени или (ii) способную образовывать стабильный дуплекс с частью мРНК-транскрипта гена-мишени. Стабильность комплексов и дуплексов может быть определена посредством теоретических расчетов и/или анализов in vitro. Типовые анализы для определения стабильности гибридизационных комплексов и дуплексов описаны в Примерах ниже.
[0053] В настоящей заявке термин "нуклеиновая кислота-мишень" охватывает ДНК, РНК (содержащую пре-мРНК и мРНК), транскрибируемую с такой ДНК, и также кДНК, получаемую на матрице такой РНК, кодирующие, некодирующие последовательности, смысловые или антисмысловые полинуклеотиды. Специфическая гибридизация олигомерного соединения с его нуклеиновой кислотой-мишенью препятствует нормальной функции нуклеиновой кислоты. Это модулирование функции нуклеиновой кислоты-мишени с помощью соединений, которые специфически гибридизуются с последней, в общем случае называют "антисенс-технологией". Затрагиваемые функции ДНК включают, например, репликацию и транскрипцию. Затрагиваемые функции РНК включают все жизненно важные функции, такие как, например, транслокация РНК к месту трансляции белков, трансляция белков на матрице РНК, сплайсинг РНК с образованием одного или более видов мРНК и каталитическая активность, которая может присутствовать у РНК или которой РНК может способствовать. Суммарный эффект такого препятствия функции нуклеиновой кислоты-мишени заключается в модулировании экспрессии кодируемого продукта или олигонуклеотидов.
[0054] РНК-интерференция "РНКи" опосредована молекулами двухцепочечной РНК (дцРНК), которые имеют последовательности, гомологичные их нуклеотидным последовательностям-"мишеням". В некоторых вариантах реализации настоящего изобретения посредники представляют собой дуплексы "малых интерферирующих" РНК (ммРНК) длиной 5-25 нуклеотидов. ммРНК образуются в результате разрезания дцРНК ферментом-РНКазой, известным как Дайсер (Dicer). Дуплексные продукты ммРНК включаются в состав мультибелкового комплекса, называемого РНК-индуцируемый комплекс сайленсинга (RNA induced silencing complex, RISC). Без привязки к какой-либо конкретной теории, комплекс RISC, как полагают, нацелен на нуклеиновую кислоту-мишень (на роль которой подходит мРНК), при этом дуплекс ммРНК взаимодействует с мРНК за счет комплементарности их последовательностей, опосредуя расщепление мРНК каталитическим путем. Малые интерферирующие РНК, которые могут быть применены согласно настоящему изобретению, могут быть синтезированы и применены в соответствии со способами, которые хорошо известны в данной области техники и которые будут хорошо знакомы специалисту в данной области техники. Малые интерферирующие РНК для применения в соответствии со способами согласно настоящему изобретению подходящим образом содержат от примерно 1 до примерно 50 нуклеотидов (нт). В примерах неограничивающих вариантов реализации ммРНК могут содержать от примерно 5 до примерно 40 нт, от примерно 5 до примерно 30 нт, от примерно 10 до примерно 30 нт, от примерно 15 до примерно 25 нт или примерно 20-25 нуклеотидов.
[0055] Отбор подходящих олигонуклеотидов облегчают путем применения компьютерных программ, которые автоматически выравнивают нуклеотидные последовательности и отмечают идентичные или гомологичные области. Такие программы применяют для сравнения нуклеотидных последовательностей, полученных, например, в результате поиска в базах данных, таких как GenBank, или посредством секвенирования ПНР-продуктов. Сравнение нуклеотидных последовательностей из организмов ряда видов позволяет отобрать нуклеотидные последовательности, которые обнаруживают подходящую степень межвидовой идентичности. В случае генов, которые не были просеквенированы, проводят блоттинги по Саузерну с целью определения степени идентичности между генами вида-мишени и других видов. Путем проведения блоттингов по Саузерну при различных степенях строгости условий гибридизации, как хорошо известно в данной области техники, можно получить приблизительную меру идентичности. Эти процедуры позволяют отобрать олигонуклеотиды, которые показывают высокую степень комплементарности нуклеотидным последовательностям-мишеням субъекта, подлежащего контролю, и меньшую степень комплементарности соответствующим нуклеотидным последовательностям других видов. Специалист в данной области техники обнаружит, что существует значительная свобода при выборе подходящих областей генов для применения согласно настоящему изобретению.
[0056] С помощью термина "ферментативная РНК" обозначают молекулу РНК, обладающую ферментативной активностью (Cech, (1988) J. American. Med. Assoc. 260, 3030-3035). Ферментативные нуклеиновые кислоты (рибозимы) действуют за счет, во-первых, связывания с РНК-мишенью. Такое связывание происходит через участок ферментативной нуклеиновой кислоты, связывающий РНК-мишень, который находится в непосредственной близости к ферментативному участку молекулы, катализирующему расщепление РНК-мишени. Таким образом, ферментативная нуклеиновая кислота сперва распознает и далее связывает РНК-мишень за счет спаривания оснований, и, как только происходит связывание с нужным сайтом, проявляет ферментативную активность, разрезая РНК-мишень.
[0057] С помощью термина "ложная РНК" обозначают молекулу РНК, которая имитирует природный связывающий домен для лиганда. Ложная РНК, таким образом, конкурирует с природной связывающей мишенью за связывание конкретного лиганда. Например, было показано, что сверхэкспрессия РНК-элемента трансактивационного ответа (trans-activation response, TAR) ВИЧ может действовать как "ложная" и эффективно связывает белок tat (транскрипционный трансактиватор) ВИЧ, таким образом препятствуя связыванию этого белка с последовательностями TAR, кодируемыми РНК ВИЧ. Подразумевают, что приведенная информация представляет собой конкретный пример. Специалистам в данной области техники будет очевидно, что это всего лишь один пример, и другие варианты реализации могут быть легко разработаны с применением методик, общеизвестных в данной области техники.
[0058] В настоящей заявке термин "мономеры", как правило, обозначает мономеры, соединенные посредством фосфодиэфирных связей или их аналогов с образованием олигонуклеотидов, размеры которых изменяются в пределах от нескольких мономерных единиц, например, от примерно 3-4, до примерно нескольких сотен мономерных единиц. Аналоги фосфодиэфирных связей включают: фосфотиоатные, фосфодитиоатные, метилфосфонатные, фосфоселеноатные, фосфорамидатные связи и т.п., как более подробно описано ниже.
[0059] Термин "нуклеотид" распространяется на встречающиеся в природе нуклеотиды, а также не встречающиеся в природе нуклеотиды. Специалисту в данной области техники должно быть ясно, что различные нуклеотиды, которые ранее считали "не встречающимися в природе", были впоследствии найдены в природе. Таким образом, термин "нуклеотиды" включает не только известные молекулы, содержащие пуриновые и пиримидиновые гетероциклы, но также и их гетероциклические аналоги и таутомеры. Иллюстративные примеры других типов нуклеотидов представляют собой молекулы, содержащие аденин, гуанин, тимин, цитозин, урацил, пурин, ксантин, диаминопурин, 8-оксо-N6-метиладенин, 7-деазаксантин, 7-деазагуанин, N4,N4-этаноцитозин, Н6,N6-этано-2,6-диаминопурин, 5-метилцитозин, 5-(С3-С6)-алкинилцитозин, 5-фторурацил, 5-бромурацил, псевдоизоцитозин, 2-гидрокси-5-метил-4-триазолпиридин, изоцитозин, изогуанин, инозин и "не встречающиеся в природе" нуклеотиды, описанные в патенте США №5432272, Benner et al. Термин "нуклеотид" распространяется на каждый и все из этих примеров, а также их аналоги и таутомеры. Особенно интересными нуклеотидами являются те из них, которые содержат аденин, гуанин, тимин, цитозин и урацил, которые рассматривают как встречающиеся в природе нуклеотиды относительно терапевтического и диагностического применения у людей. Нуклеотиды содержат природные 2'-дезокси- и 2'-гидроксисахара, например, описанные в Komberg and Baker, DNA Replication, 2nd Ed. (Freeman, San Francisco, 1992), a также их аналоги.
[0060] Термин "аналоги" в применении к нуклеотидам включает синтетические нуклеотиды, содержащие модифицированные основания и/или модифицированные сахара (см., например, аналоги, описанные, в основном, в Scheit, Nucleotide Analogs, John Wiley, New York, 1980; Freier & Altmann, (1997) Nucl. Acid. Res., 25(22), 4429-4443, Toulrne, J.J, (2001) Nature Biotechnology 19: 17-18; Manoharan M., (1999) Biochemica et Biophysica Acta 1489: 117-139; Freier S.M., (1997) Nucleic Acid Research, 25: 4429-4443, Uhlman, E., (2000) Drug Discovery & Development, 3: 203-213, Herdewin P., (2000) Antisense & Nucleic Acid Drug Dev., 10: 297-310); 2'-O, 3'-С-соединенные [3.2.0]бициклоарабинонуклеозиды. Такие аналоги включают синтетические нуклеотиды, направленные на усиление связывающих свойств, например, стабильности дуплексов или триплексов, специфичности и т.п.
[0061] В настоящей заявке термин "гибридизация" обозначает спаривание практически комплементарных цепей олигомерных соединений. Один механизм спаривания задействует образование водородных связей, которые могут представлять собой Уотсон-Криковские, Хугстиновские или обратные Хугстиновские водородные связи между комплементарными основаниями нуклеозидов или нуклеотидов (нуклеотидами) цепей олигомерных соединений.
Например, аденин и тимин являются комплементарными нуклеотидами, которые спариваются за счет образования водородных связей. Гибридизация может происходить при различных условиях.
[0062] Антисмысловое соединение является "специфически гибридизуемым", если связывание соединения с нуклеиновой кислотой-мишенью препятствует нормальной функции нуклеиновой кислоты-мишени, модулируя функцию и/или активность последней, и обеспечена достаточная степень комплементарности во избежание неспецифического связывания антисмыслового соединения с нуклеотидными последовательностями, не являющимися мишенями, при условиях, в которых желательно специфическое связывание, т. е. при физиологических условиях в случае анализов in vivo или терапевтического лечения и при условиях, в которых проводят анализы в случае анализов in vitro.
[0063] В настоящей заявке фраза "строгие условия гибридизации" или "строгие условия" относится к условиям, при которых соединение согласно изобретению будет гибридизоваться со своей последовательностью-мишенью, но с минимальным количеством других последовательностей. Строгие условия зависят от последовательности и будут различаться в разных обстоятельствах, и, в контексте настоящего изобретения, "строгие условия", при которых олигомерные соединения гибридизуются с последовательностью-мишенью, определяются природой и составом олигомерных соединений и видами анализов, посредством которых их исследуют. В общем случае, строгие условия гибридизации включают низкие концентрации (<0,15 М) солей, содержащих неорганические катионы, такие как Na++ или К++ (т.е. низкую ионную силу), температуру выше точки, расположенной на 20°С-25°С ниже Тпл комплекса олигомерное соединение: последовательность-мишень, и присутствие денатурантов, таких как формамид, диметилформамид, диметилсульфоксид, или детергента додецилсульфата натрия (ДСН). Например, скорость гибридизации снижается на 1,1% на каждый добавленный 1% формамида. Примером условий гибридизации с высокой степенью строгости является гибридизация с применением 0,1X буфера хлорид натрия-цитрат натрия (SSC, стандартный цитратно-солевой)/0,1% (масс./об.) ДСН при 60°С в течение 30 минут.
[0064] В настоящей заявке термин "комплементарный" относится к способности точного спаривания между двумя нуклеотидами на одной или двух олигомерных цепях. Например, если нуклеиновое основание в определенном положении цепи антисмыслового соединения способно к образованию водородных связей с нуклеиновым основанием в определенном положении цепи нуклеиновой кислоты-мишени, причем, указанная нуклеиновая кислота-мишень представляет собой молекулу ДНК, РНК или олигонуклеотида, то положение, в котором происходит образование водородных связей между олигонуклеотидом и нуклеиновой кислотой-мишенью, считают комплементарным положением. Олигомерное соединение и дополнительная молекула ДНК, РНК или олигонуклеотида комплементарны друг другу, когда достаточное число комплементарных положений в каждой молекуле заняты нуклеотидами, способными образовывать водородные связи друг с другом. Таким образом, термины "специфически гибридизуемый" и "комплементарный" применяют для обозначения достаточной степени точного спаривания, или комплементарности, в пределах достаточного числа нуклеотидов, так что между олигомерным соединением и нуклеиновой кислотой-мишенью осуществляется стабильное и специфичное связывание.
[0065] Как понятно специалистам в данной области техники, последовательность олигомерного соединения не обязательно должна быть на 100% комплементарна последовательности его нуклеиновой кислоты-мишени для того, чтобы соединение было специфически гибридизуемым. Кроме того, олигонуклеотид может гибридизоваться поверх одного или более сегментов, так что промежуточные или соседние сегменты оказываются не вовлеченными в процесс гибридизации (например, петлевая структура, отсутствие комплементарности или шпилечная структура). Олигомерные соединения согласно настоящему изобретению содержат последовательность, комплементарную на по меньшей мере примерно 70%, или по меньшей мере примерно 75%, или по меньшей мере примерно 80%, или по меньшей мере примерно 85%, или по меньшей мере примерно 90%, или по меньшей мере примерно 95%, или по меньшей мере примерно 99% области-мишени в пределах нуклеотидной последовательности-мишени, на которую эти соединения нацелены. Например, антисмысловое соединение, в котором 18 из 20 нуклеотидов антисмыслового соединения комплементарны области-мишени и, следовательно, специфически гибридизовались бы с ним, характеризовалось бы 90% комплементарностью. В данном примере остальные некомплементарные нуклеотиды могут располагаться в кластерах комплементарных нуклеотидов или чередоваться с комплементарными нуклеотидами и не обязательно должны примыкать друг к другу или к комплементарным нуклеотидам. По этой причине антисмысловое соединение длиной 18 нуклеотидов, содержащее 4 (четыре) некомплементарных нуклеотида, которые фланкированы двумя областями, полностью комплементарными нуклеиновой кислоте-мишени, в целом характеризовалось бы 77,8% комплементарностью с нуклеиновой кислотой-мишенью и попадало бы, таким образом, в рамки настоящего изобретения. Процент комплементарности антисмыслового соединения с областью нуклеиновой кислоты-мишени может быть определен обычным способом с применением программ BLAST (средства поиска основного локального выравнивания) и программ PowerBLAST, известных в данной области техники. Процент гомологии, идентичности последовательностей или комплементарности может быть определен с помощью, например, программы Gap (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, Madison Wis.) с применением параметров, установленных по умолчанию, которая использует алгоритм Смита-Ватермана (Adv. Appl. Math, (1981) 2, 482-489).
[0066] В настоящей заявке термин "температура плавления (Тпл)" относится к температуре, в условиях определенной ионной силы, рН и концентрации нуклеиновой кислоты, при которой 50 % олигонуклеотидов, комплементарных последовательности-мишени, гибридизуются с последовательностью-мишенью в состоянии равновесия. Как правило, строгие условия гибридизации будут отличаться тем, что концентрация соли составляет по меньшей мере примерно от 0,01 до 1,0 М концентрации ионов Na (или других солей) при рН 7,0-8,3 и температура равна по меньшей мере примерно 30°С для коротких олигонуклеотидов (например, длиной от 10 до 50 нуклеотидов). Строгие условия могут также быть достигнуты путем добавления дестабилизирующих агентов, таких как формамид.
[0067] В настоящей заявке термин "модулирование" обозначает либо увеличение (стимулирование), либо ослабление (ингибирование) экспрессии гена.
[0068] Термин "вариант", применительно к последовательности полинуклеотида, может охватывать последовательность полинуклеотида, относящегося к гену дикого типа. Это определение может также включать, например, "аллельные", "сплайс-", "видовые" или "полиморфные" варианты. Сплайс-вариант может быть в значительной степени идентичным исходной молекуле, но будет в общем случае содержать большее или меньшее число полинуклеотидов вследствие альтернативного сплайсинга экзонов во время процессинга мРНК. Соответствующий полипептид может обладать дополнительными функциональными доменами или отличаться отсутствием доменов. Видовые варианты представляют собой последовательности полинуклеотидов, которые варьируют от одного вида к другому. Особенно полезными согласно настоящему изобретению являются варианты продуктов генов дикого типа. Варианты могут возникнуть в результате по меньшей мере одной мутации в нуклеотидной последовательности и могут приводить к появлению измененных мРНК или полипептидов, структура или функция которых могут быть или могут не быть изменены. Любой отдельно взятый природный или рекомбинантный ген может не иметь, иметь одну или много аллельных форм. Обычные мутационные изменения, вызывающие возникновение вариантов, в общем случае приписывают природным делециям, вставкам или заменам нуклеотидов. Каждый из этих типов изменений может произойти отдельно или в комбинации с остальными типами, один или более раз в данной последовательности.
[0069] Итоговые полипептиды в общем случае будут характеризоваться значительной идентичностью аминокислотных последовательностей относительно друг друга.
Полиморфный вариант представляет собой вариацию в полинуклеотидной последовательности конкретного гена между отдельными особями данного вида. Полиморфные варианты также могут охватывать "одиночные нуклеотидные полиморфизмы" (ОНП; single nucleotide polymorphisms, SNPs), или мутации, затрагивающие одно основание, при которых полинуклеотидная последовательность изменяется на одно основание. Наличие ОНП может указывать на, например, некоторую популяцию со склонностью к болезненному состоянию, что отражает восприимчивость против устойчивости.
[0070] Производные полинуклеотиды включают нуклеиновые кислоты, подвергнутые химической модификации, например, замещению водорода на алкил, ацил или аминогруппу. Производные, например, производные олигонуклеотиды, могут содержать не встречающиеся в природе фрагменты, такие как измененные остатки сахаров или связи между остатками сахаров. Типовыми среди них являются фосфотиоатные и другие содержащие серу виды связей, которые известны в данной области техники. Производные нуклеиновые кислоты могут также содержать метки, включая радионуклеотиды, ферменты, флуоресцентные агенты, хемилюминесцентные агенты, хромогенные агенты, субстраты, кофакторы, ингибиторы, магнитные частицы и т.п.
[0071] "Производным" полипептидом или пептидом является тот, который модифицирован, например, посредством гликозилирования, пегилирования, фосфорилирования, сульфатирования, восстановления/алкилирования, ацилирования, химического сочетания или мягкой обработки формалином. Производное может также быть модифицировано добавлением детектируемой метки, либо напрямую, либо косвенно, включая, но не ограничиваясь перечисленными, радиоизотопную, флуоресцентную и ферментативную метку.
[0072] В настоящей заявке термин "животное" или "пациент" включает, например, людей, овцу, лосей, оленя, чернохвостого оленя, норок, млекопитающих, обезьян, лошадей, крупный рогатый скот, свиней, коз, собак, кошек, крыс, мышей, птиц, курицу, пресмыкающихся, рыбу, насекомых и паукообразных.
[0073] Термин "млекопитающее" распространяется на теплокровных млекопитающих, которым, как правило, оказывают медицинскую помощь (например, люди и одомашненные животные). Примеры включают кошачьих, псовых, лошадиных, бычьих и человека, а также только человека.
[0074] Термин "лечение" распространяется на лечение болезненного состояния у млекопитающего и включает: (а) предотвращение возникновения болезненного состояния у млекопитающего, в частности, когда такое млекопитающее предрасположено к болезненному состоянию, но последнее еще не было диагностировано у субъекта; (b) ингибирование болезненного состояния, например, остановку его развития; и/или (с) облегчение болезненного состояния, например, индукцию ремиссии болезненного состояния, до тех пор, пока не будет достигнут желаемый результат. Лечение также включает уменьшение интенсивности симптома заболевания (например, уменьшение боли или дискомфорта), при этом такое уменьшение интенсивности симптомов может напрямую влиять или не влиять на заболевание (например, причину, передачу, проявление и т.д.).
[0075] В настоящей заявке термин "неврологическое заболевание или расстройство" относится к любому заболеванию или расстройству нервной системы и/или зрительной системы. "Неврологическое заболевание или расстройство" включает заболевание или расстройства, которые затрагивают центральную нервную систему (мозг, ствол мозга и мозжечок), периферическую нервную систему (включая черепные нервы) и вегетативную нервную систему (части которой находятся и в центральной, и в периферической нервной системе). Неврологическое заболевание или расстройство включает, но не ограничивается перечисленными, приобретенную эпилептиформную афазию; острый диссеминированный энцефаломиелит; адренолейкодистрофию; возрастную макулярную дегенерацию; агенезию мозолистого тела; агнозию; синдром Айкарди; болезнь Александера; болезнь Альпера; альтернирующую гемиплегию; болезнь Альцгеймера; сосудистую деменцию; боковой амиотрофический склероз; анэнцефалию; синдром Ангельмана; ангиоматоз; аноксию; афазию; апраксию; арахноидальные кисты; арахноидит; мальформацию Арнольда-Киари; артериовенозную мальформацию; синдром Аспергера; атаксию-телеангиэктазию; синдром дефицита внимания и гиперактивности; аутизм; вегетативную дисфункцию; боль в спине; болезнь Баттена; болезнь Бехчета; паралич Белла; доброкачественный эссенциальный блефароспазм; доброкачественную фокальную эпилепсию; амиотрофию; доброкачественную внутричерепную гипертензию; болезнь Бинсвангера; блефароспазм; синдром Блоха-Сульцбергера; повреждение плечевого сплетения; абсцесс мозга; травму головного мозга; опухоли мозга (включая мультиформную глиобластому); опухоль спинного мозга; синдром Броун-Секара; болезнь Канавана; синдром запястного канала; каузалгию; центральный болевой синдром; центральный понтинный миелинолиз; цефалический синдром; аневризму сосудов головного мозга; церебральный артериосклероз; церебральную атрофию; церебральный гигантизм; церебральный паралич; болезнь Шарко-Мари-Тута; невропатию и невропатическую боль, вызванные химиотерапией; мальформацию Киари; хорею; хроническую воспалительную демиелинизирующую полинейропатию; хроническую боль; хронический региональный болевой синдром; синдром Коффина-Лоури; кому, включающую устойчивое вегетативное состояние; врождённую лицевую диплегию; кортикобазальную дегенерацию; височный артериит; краниосиностоз; болезнь Крейтцфельдта-Якоба; кумулятивные травматические расстройства; синдром Кушинга; инклюзионную цитомегалию; цитомегаловирусную инфекцию; синдром танцующих глаз-танцующих ног; синдром Денди-Уокера; болезнь Доусона; синдром Де Морсье; паралич Дежерин-Клюмпке; деменцию; дерматомиозит; диабетическую нейропатию; диффузный склероз; вегетативную дистонию; дисграфию; дислексию; дистонии; раннюю инфантильную эпилептическую энцефалопатию; синдром пустого турецкого седла; энцефалит; энцефалоцеле; энцефалотригеминальный ангиоматоз; эпилепсию; паралич Эрба; эссенциальный тремор; болезнь Фабри; синдром Фара; потерю сознания; семейный спастический паралич; фебрильные судороги; синдром Фишера; атаксию Фридрейха; лобно-височную деменцию и другие "таупатии"; болезнь Гоше; синдром Герстманна; гигантоклеточный артериит; гигантоклеточную инклюзионную болезнь; глобоидно-клеточную лейкодистрофию; синдром Гийена-Барре; HTLV-1-ассоциированную миелопатию (миелопатию, ассоциированную с вирусом Т-клеточного лейкоза человека); болезнь Галлервордена-Шпатца; травму головы; головную боль; гемифациальный спазм; наследственную спастическую параплегию; наследственную полиневропатическую атаксию; синдром коленчатого узла; опоясывающий лишай; синдром Хираямы; ВИЧ-ассоциированные деменцию и нейропатию (также неврологические проявления СПИДа); голопрозэнцефалию; болезнь Хантингтона и другие заболевания, обусловленные наличием полиглутаминовых повторов; гидроанэнцефалию; гидроцефалию; гиперкортицизм; гипоксию; иммуноопосредованный энцефаломиелит; миозит с включениями телец; недержание пигмента; инфантильную форму болезни накопления фитановой кислоты; инфантильную болезнь Рефсума; инфантильные спазмы; воспалительную миопатию; внутричерепную кисту; внутричерепную гипертензию; синдром Жубера; синдром Кернса-Сейра; болезнь Кеннеди; синдром Kinsboume; синдром Клиппеля-Фейля; болезнь Краббе; болезнь Кугельберга-Веландер; куру; болезнь Лафора; миастенический синдром Ламберта-Итона; синдром Ландау-Клеффнера; боковой медуллярный синдром (синдром Валленберга); необучаемость; болезнь Лея; синдром Леннокса-Гасто; синдром Леша-Нихена; лейкодистрофию; деменцию с тельцами Леви; лиссэнцефалию; синдром окружения; болезнь Лу Герига (например, болезнь двигательных нейронов или боковой амиотрофический склероз); заболевание поясничных дисков; неврологические осложнения, ассоциированные с болезнью Лайма; болезнь Мачадо-Джозефа; макроэнцефалию; мегацефалию; синдром Мелькерссона-Розенталя; болезнь Меньера; менингит; болезнь Менкеса; метахроматическую лейкодистрофию; микроцефалию; мигрень; синдром Миллера-Фишера; мини-инсульты; митохондриальные миопатии; синдромом Мебиуса; мономелическую амиотрофию; болезнь двигательных нейронов; болезнь мойя-мойя; мукополисахаридозы; мультиинфарктную деменцию; моторную мультифокальную невропатию; рассеянный склероз и другие демиелинизирующие расстройства; множественную системную атрофию с ортостатической гипотензией; мышечную дистрофию; тяжелую псевдопаралитическую миастению; диффузный миелинокластический склероз; раннюю миоклоническую энцефалопатию; миоклонус; миопатию; врожденную миотонию; нарколепсию; нейрофиброматоз; злокачественный нейролептический синдром; неврологические проявления СПИДа; неврологические осложнения после волчанки; нейромиотонию; нейрональный цероидный липофусциноз; нарушения миграции нейронов; болезнь Ниманна-Пика; синдром О'Салливана-МакЛауда; затылочную невралгию; скрытую дизрафию спинного мозга; синдром Охтахара; оливопонтоцеребеллярную атрофию; опсоклонус-миоклонус синдром; неврит зрительного нерва; ортостатическую гипотензию; синдром перегрузки; парестезию; нейродегенеративное заболевание или расстройство (болезнь Паркинсона, болезнь Хантингтона, болезнь Альцгеймера, боковой амиотрофический склероз (БАС), деменцию, рассеянный склероз и другие заболевания и расстройства, ассоциированные с гибелью нейронов); врожденную парамиотонию; паранеопластические заболевания; пароксизмальные приступы; синдром Парри-Ромберга; болезнь Пелицеуса-Мерцбахера; периодические параличи; периферическую нейропатию; болезненную нейропатию и нейропатическую боль; устойчивое вегетативное состояние; первазивные расстройства развития; световой рефлекс чихания; болезнь накопления фитановой кислоты; болезнь Пика; защемление нерва; опухоли гипофиза; полимиозит; порэнцефалию; постполиомиелитный синдром; постгерпетическую невралгию; постинфекционный энцефаломиелит; ортостатическую гипотензию; синдром Прадера-Вилли; первичный боковой склероз; прионные заболевания; прогрессирующую гемифациальную атрофию; прогрессирующую многоочаговую лейкоэнцефалопатию; прогрессирующую склерозирующую полиодистрофию; прогрессирующий надъядерный паралич; доброкачественную внутричерепную гипертензию; синдром Рамсея-Ханта (типы I и II); энцефалит Расмуссена; синдром рефлекторной симпатической дистрофии; болезнь Рефсума; повторяющиеся расстройства движения; туннельный синдром; синдром беспокойных ног; миелопатию, ассоциированную с ретровирусной инфекцией; синдром Ретта; синдром Рейе; пляску Святого Витта; болезнь Сандхоффа; болезнь Шидьдера; шизэнцефалию; септо-оптическую дисплазию; синдром детского сотрясения; опоясывающий лишай; синдром Шая-Дрейджера; синдром Шегрена; апноэ во сне; синдром Сотоса; спастичность; расщепленный позвоночник; повреждение спинного мозга; опухоли спинного мозга; спинальную мышечную атрофию; синдром мышечной скованности; инсульт; синдром Стерджа-Вебера; подострый склерозирующий панэнцефалит; подкорковую атеросклеротическую энцефалопатию; хорею Сиденгама; обмороки; сирингомиелию; позднюю дискинезию; болезнь Тея-Сакса; височный артериит; синдром фиксированного спинного мозга; болезнь Томсена; синдром верхней апертуры грудной клетки; невралгию тройничного нерва; паралич Тодда; синдром Туретта; транзиторную ишемическую атаку; трансмиссивные спонгиоформные энцефалопатии; поперечный миелит; черепно-мозговую травму; тремор; тригеминальную невралгию; тропический спастический парапарез; туберозный склероз; сосудистую деменцию (мультиинфарктную деменцию); васкулит, в том числе височный артериит; болезнь Гиппеля-Линдау; синдром Валленберга; болезнь Верднига-Гоффмана; синдром Веста; хлыстовую травму; синдром Вильямса; болезнь Вильсона; и синдром Цельвегера и другие неврологические расстройства, перечисленные в настоящей заявке.
[0076] Сердечно-сосудистое заболевание или расстройство включает такие расстройства, которые либо могут привести к ишемии, либо вызваны реперфузией сердца. Примеры включают, но не ограничиваются перечисленными, атеросклероз, ишемическую болезнь сердца, гранулематозный миокардит, хронический миокардит (негранулематозный), первичную гипертрофическую кардиомиопатию, болезнь периферических артерий (БПА), заболевание периферических сосудов, венозную тромбоэмболию, эмболию легочной артерии, инсульт, стенокардию, инфаркт миокарда, повреждения сердечно-сосудистых тканей, вызванные остановкой сердца, повреждения сердечно-сосудистых тканей, вызванные шунтированием сердца, кардиогенный шок и сходные состояния, известные специалистам в данной области техники или связанные с дисфункцией или повреждением тканей сердца или сосудов, в частности, но не ограничиваясь перечисленными, повреждение тканей, связанное с активацией гена SCNA. Заболевания сердечно-сосудистой системы включают, но не ограничиваются перечисленными, атеросклероз, гранулематозный миокардит, инфаркт миокарда, вторичный фиброз миокарда в результате порока клапана сердца, фиброз миокарда без инфаркта, первичную гипертрофическую кардиомиопатию и хронический миокардит (негранулематозный).
[0077] Примеры заболеваний или расстройств, ассоциированных с дисфункцией натриевого канала, включают, но не ограничиваются перечисленными, злокачественную гипертермию, миастению, эпизодическую атаксию, нейропатическую и воспалительную боль, болезнь Альцгеймера, болезнь Паркинсона, шизофрению, гиперэкплексию, миотонии, такие как гипо- и гиперкалиемический периодический паралич, врожденную парамиотонию и обостряемую действием калия миотонию, а также сердечные аритмии, такие как синдром удлиненного интервала QT.
Полинуклеотидные и олигонуклеотидные композиции и молекулы
[0078] Мишени: В одном варианте реализации мишени включают нуклеотидные последовательности гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA), включая, без ограничения, смысловые и/или антисмысловые некодирующие и/или кодирующие последовательности, ассоциированные с геном SCNA.
[0079] Потенциалчувствительные ионные каналы представляют собой класс трансмембранных белков, которые обеспечивают основу для клеточной возбудимости и способности передавать информацию посредством создаваемых ионами мембранных потенциалов. В ответ на изменения мембранных потенциалов эти молекулы опосредуют быстрый поток ионов через селективные каналы в клеточной мембране. Если плотность канала достаточно высока, возникает регенеративная деполяризация, которую называют потенциалом действия.
[0080] Потенциалзависимый натриевый канал отвечает за генерацию и распространение потенциалов действия в большинстве электрически возбудимых клеток, включая нейроны, клетки сердца и мышцы. Электрическую активность вызывает деполяризация мембраны, открывающая каналы через мембрану, которые являются высокоселективными для ионов натрия. Далее ионы перемещаются внутрь клетки через открытые каналы под действием электрохимического градиента. Несмотря на то, что потенциалы действия, обусловленные потоками ионов натрия, одинаковы в разных тканях, электрофизиологические исследования показали, что существуют несколько структурно и функционально различных натриевых каналов, и были клонированы многочисленные гены, кодирующие натриевые каналы. Ген SCNA принадлежит к семейству генов потенциалзависимых натриевых каналов.
[0081] Потенциалзависимые натриевые каналы могут быть названы в соответствии со стандартизованной формой номенклатуры, изложенной в Goldin et al. (2000) Neuron 28: 365-368. Согласно этой системе, потенциалзависимые натриевые каналы группируют в одно семейство, из которого девять изоформ натриевых каналов млекопитающих были идентифицированы и экспрессированы. Данные девять изоформ получили названия Nav1.1-Nav1.9. Кроме того, сплайс-варианты различных изоформ обозначают с помощью строчных букв, следующих за номерами (например, "Nav1.1a").
[0082] Потенциалзависимые натриевые каналы играют важную роль в генерации потенциала действия в нервных клетках и мышцах. Альфа-субъединица (SCNA) представляет собой основной компонент канала, и ее наличия было бы достаточно для создания эффективного канала при экспрессии в клетках in vitro. В свою очередь, бета-1- и бета-2-субъединицы нуждаются в присутствии альфа-субъединицы для образования эффективного канала. Роль этих субъединиц заключается, возможно, в изменении кинетических свойств канала, главным образом за счет быстрой инактивации натриевых токов. Мутация, обнаруженная при синдроме генерализованной эпилепсии с фебрильными припадками (СГЭФП; generalized epilepsy with febrile seizures, GEFS) в гене SCN1B, как было показано, замедляет быструю инактивацию натриевых каналов по сравнению с нормальным геном SCNB1 при совместной экспрессии с альфа-субъединицей.
[0083] В одном варианте реализации антисмысловые олигонуклеотиды применяют для предотвращения или лечения заболеваний или расстройств, ассоциированных с членами семейства генов SCNA. Типовые заболевания и расстройства, опосредованные альфа-субъединицей потенциалзависимого натриевого канала (SCNA), которые можно лечить с помощью клеток/тканей, регенерированных из стволовых клеток, полученных с применением антисмысловых соединений, включают: заболевание или расстройство, ассоциированное с аномальной функцией и/или экспрессией гена SCNA, неврологическое заболевание или расстройство, конвульсии, боль (включая хроническую боль), нарушенную электрическую возбудимость, сопряженную с дисфункцией натриевого канала, заболевание или расстройство, ассоциированное с дисфункцией натриевого канала, заболевание или расстройство, ассоциированное с неправильной регуляцией активности альфа-субъединицы потенциалзависимого натриевого канала (например, паралич, гиперкалиемический периодический паралич, врожденную парамиотонию, обостряемую действием калия миотонию, синдром 3 удлиненного интервала QT, заболевание двигательных замыкательных пластинок, атаксию и т.д.), заболевание желудочно-кишечного тракта, возникшее вследствие дисфункции энтеральной нервной системы (например, колит, илеит, синдром воспаленного кишечника и т.д.), сердечно-сосудистое заболевание или расстройство (например, гипертензию, застойную сердечную недостаточность и т.д.); заболевание или расстройство мочеполовой системы с вовлечением симпатической и парасимпатической иннервации (например, доброкачественную гиперплазию предстательной железы, импотенцию); заболевание или расстройство, ассоциированное с нервно-мышечной системой (например, мышечную дистрофию, рассеянный склероз, эпилепсию, аутизм, мигрень (например, спорадические и семейные гемиплегические мигрени и т.д.), тяжелую миоклоническую эпилепсию младенчества (severe myoclonic epilepsy of infancy, SMEI; или синдром Драве), генерализованную эпилепсию с фебрильными припадками плюс (ГЭФП+; GEFS+) и т.д.), и ассоциированную с геном SCNA эпилепсию.
[0084] Настоящее изобретение также относится к фармацевтической композиции, содержащей по меньшей мере один из олигонуклеотидов, нацеленных на природный антисмысловой транскрипт по меньшей мере одной или более из мишеней, выбранных из группы, состоящей из генов или мРНК SCN1A-SCN12A или их изоформ или вариантов. Настоящее изобретение также относится к способу лечения неврологического заболевания или расстройства, включающему введение олигонуклеотида, нацеленного на природный антисмысловой транскрипт по меньшей мере одной или более из мишеней, выбранных из группы, состоящей из мРНК SCN1A, SCN2A, SCN3A, SCN4A, SCN5A, SCN6A, SCN7A, SCN8A, SCN9A, SCN10A, SCN11A и SCN12A или их вариантов. В предпочтительном варианте реализации олигонуклеотиды выбраны так, чтобы увеличивать экспрессию полнофункционального продукта экспрессии указанного семейства генов SCNA. В предпочтительном варианте реализации олигонуклеотиды согласно настоящему изобретению увеличивают транскрипцию и/или трансляцию любой из мРНК семейства генов SCNXA с обеспечением синтеза полнофункциональных натриевых каналов у пациента, нуждающегося в таком лечении. У пациентов, имеющих заболевание или расстройство, ассоциированное с мутантным вариантом потенциалзависимого натриевого канала, в предпочтительном варианте реализации введение или лечение с применением фармацевтической композиции, содержащей олигонуклеотид, нацеленный на природный антисмысловой транскрипт гена альфа-субъединицы потенциалзависимого натриевого канала или мРНК такого гена, увеличивает экспрессию полнофункционального продукта экспрессии в большей степени, чем продукта экспрессии мутантной формы гена. В другом варианте реализации настоящее изобретение относится к комбинации олигонуклеотидов, нацеленных на по меньшей мере один природный антисмысловой транскрипт по меньшей мере двух членов семейства генов SCNXA, где Х выбран из 1-12. Например, при лечении синдрома Драве может быть применена комбинация олигонуклеотидов для увеличения экспрессии продуктов экспрессии, например, генов SCN1A и SCN9A. В другом варианте реализации по меньшей мере один олигонуклеотид может быть выбран для нацеливания на природный антисмысловой транскрипт по меньшей мере двух генов, выбранных из любого из генов SCN1A-SCN12A. Предпочтительные олигонуклеотиды согласно изобретению имеют длину от примерно 5 до примерно 30 нуклеотидов и комплементарны на по меньшей мере 50% сегменту природного антисмыслового транскрипта (natural antisense transcript, NAT) длиной от 5 до примерно 30 нуклеотидов. Предпочтительными NAT любого из генов SCNA или продуктов их транскрипции являются те, которые, при выборе их в качестве мишени для олигонуклеотида согласно изобретению, взаимодействуют с олигонуклеотидом, что приводит к модулированию экспрессии мРНК и/или продукта трансляции указанной мРНК. В предпочтительном варианте реализации олигонуклеотиды увеличивают экспрессию функционального белка мишени для лечения или смягчения последствий ассоциированного с геном SCNA заболевания. В предпочтительном варианте реализации это "увеличение экспрессии" не связано с вызыванием или развитием заболевания, такого как рак.
[0085] Изменения в гене SCNA могут включать или охватывать многие или все формы генных мутаций, включая вставки, делеции, перестановки и/или точечные мутации в кодирующих и/или некодирующих областях гена. Делеции могут представлять собой делеции целого гена или части гена. Точечные мутации могут приводить к аминокислотным заменам, сдвигам рамки считывания или появлению стоп-кодонов. Точечные мутации могут также возникать в регуляторной области гена SCNA, такой как промотор, в результате чего происходит потеря или понижение экспрессии мРНК или может произойти некорректный процессинг такой мРНК, приводящий к уменьшению стабильности мРНК или эффективности трансляции. Такие изменения у людей могут приводить к различным формам заболевания, и существуют много публикаций, в которых описана связь между изменением в гене SCNA и, например, эпилепсией или синдромом Драве (SMEI). Такие изменения могут возникать "de novo" или могут быть унаследованы. Настоящее изобретение не ограничивается лечением заболеваний, ассоциированных с изменениями в гене SCNA, и также включает лечение ассоциированного с геном SCNA заболевания или состояния, при котором пациент не имеет или не обязательно имеет изменение или мутацию в гене SCNA. Как полагают, любое модулирование или увеличение экспрессии функциональных продуктов экспрессии потенциалзависимых натриевых каналов приведет к смягчению последствий или лечению ассоциированного с геном SCNA заболевания или состояния у пациента, нуждающегося в лечении указанного заболевания или состояния. Такое смягчение последствий также может включать по меньшей мере один измеряемый признак клинического улучшения, в том числе меньшее количество припадков, менее частые припадки, менее тяжелые припадки, развитие меньшего количества типов припадков, улучшение неврологического развития или любая другая польза от лечения.
[0086] В одном варианте реализации модулирование гена SCNA с помощью одного или более антисмысловых олигонуклеотидов представляет собой введение последних пациенту, нуждающемуся в этом, для предотвращения или лечения любого заболевания или расстройства, связанного с аномальными экспрессией, функцией, активностью белка SCNA по сравнению с нормальным контролем.
[0087] В одном варианте реализации олигонуклеотиды специфичны для полинуклеотидов SCNA, которые содержат, без ограничения, некодирующие области. Мишени гена SCNA включают варианты гена SCNA; мутанты гена SCNA, в том числе ОНП; некодирующие последовательности гена SCNA; аллели, фрагменты и т.п. Предпочтительно, олигонуклеотид представляет собой молекулу антисмысловой РНК.
[0088] В соответствии с вариантами реализации изобретения, молекула нуклеиновой кислоты-мишени не ограничивается исключительно полинуклеотидами SCNA, но распространяется на любые из изоформ, рецепторов, гомологов, некодирующих областей гена SCNA и т.п.
[0089] В одном варианте реализации олигонуклеотид нацелен на природную антисмысловую последовательность (природную антисмысловую последовательность для кодирующих и некодирующих областей) для SCNA-мишеней, включая, без ограничения, их варианты, аллели, гомологи, мутанты, производные, фрагменты и последовательности, которые представляют собой их комплемент. Предпочтительно, олигонуклеотид представляет собой молекулу антисмысловой РНК или ДНК.
[0090] В одном варианте реализации олигомерные соединения согласно настоящему изобретению также включают варианты, в которых разные основания находятся в одном или более нуклеотидных положениях в соединении. Например, если первый нуклеотид представляет собой аденин, то могут быть получены варианты, содержащие тимидин, гуанозин, цитидин или другие природные или неприродные нуклеотиды в этом положении. Это может быть выполнено в любой из позиций антисмыслового соединения. Эти соединения далее исследуют применяя способы, описанные в настоящей, заявке, с целью определения их способности ингибировать экспрессию нуклеиновой кислоты-мишени.
[0091] В некоторых вариантах реализации гомология, идентичность последовательностей или комплементарность между антисмысловым соединением и мишенью составляет от примерно 50% до примерно 60%. В некоторых вариантах реализации гомология, идентичность последовательностей или комплементарность составляет от примерно 60% до примерно 70%. В некоторых вариантах реализации гомология, идентичность последовательностей или комплементарность составляет от примерно 70% до примерно 80%. В некоторых вариантах реализации гомология, идентичность последовательностей или комплементарность составляет от примерно 80% до примерно 90%. В некоторых вариантах реализации гомология, идентичность последовательностей или комплементарность составляет примерно 90%, примерно 92%, примерно 94%, примерно 95%, примерно 96%, примерно 97%, примерно 98%, примерно 99% или примерно 100%.
[0092] Антисмысловое соединение является специфически гибридизуемым, когда связывание соединения с нуклеиновой кислотой-мишенью препятствует нормальной функции нуклеиновой кислоты-мишени, вызывая потерю активности, и обеспечена достаточная степень комплементарности во избежание неспецифического связывания антисмыслового соединения с нуклеотидными последовательностями, не являющимися мишенями, при условиях, в которых желательно специфическое связывание. Такие условия включают, например, физиологические условия в случае анализов in vivo или терапевтического лечения и условия, в которых проводят анализы в случае анализов т vitro.
[0093] Антисмысловое соединения, будь то ДНК, РНК, химерное соединение, замещенное соединение и т.д., является специфически гибридизуемым, когда связывание соединения с молекулой ДНК- или РНК-мишени препятствует нормальной функции ДНК- или РНК-мишени, вызывая потерю функции, и обеспечена достаточная степень комплементарности во избежание неспецифического связывания антисмыслового соединения с последовательностями, не являющимися мишенями, при условиях, в которых желательно специфическое связывание, т.е. при физиологических условиях в случае анализов in vivo или терапевтического лечения и, в случае анализов in vitro, при условиях, в которых проводят указанные анализы.
[0094] В одном варианте реализации нацеленное воздействие на ген SCNA, включающее, без ограничения, антисмысловые последовательности, идентифицируемые и раскрываемые с применением, например, ПЦР, гибридизации и т.д., одну или более из последовательностей, соответствующих SEQ ID NOS: 12-28, и т.п., модулирует экспрессию или функцию гена SCNA. В одном варианте реализации экспрессия или функция увеличивается по сравнению с контролем. В одном варианте реализации экспрессия или функция подавляется по сравнению с контролем.
[0095] В одном варианте реализации олигонуклеотиды содержат нуклеотидные последовательности, соответствующие SEQ ID NOS: 29-94, включая антисмысловые последовательности, идентифицируемые и раскрываемые с применением, например, ПЦР, гибридизации и т.д. Эти олигонуклеотиды могут содержать один или более модифицированных нуклеотидов, более короткие или более длинные фрагменты, модифицированные связи и т.п. Примеры модифицированных связей или межнуклеотидных связей включают фосфотиоатные, фосфодитиоатные связи и т.п. В одном варианте реализации нуклеотиды содержат фосфатное производное. Указанное фосфатное производное (или модифицированная фосфатная группа), которое может быть присоединено к остатку сахара или аналогу остатка сахара в модифицированных олигонуклеотидах согласно настоящему изобретению, может представлять собой монофосфат, дифосфат, трифосфат, алкилфосфат, алканфосфат, фосфотиоат и т.п. Получение упомянутых выше аналогов фосфата и их включение в состав нуклеотидов, модифицированных нуклеотидов и олигонуклеотидов, по сути, также известно и не обязательно должно быть описано в настоящей заявке.
[0096] Специфичность и чувствительность антисмысловой последовательности также используются специалистами в данной области техники для терапевтического применения. Антисмысловые олигонуклеотиды применяли в качестве терапевтических агентов для лечения болезненных состояний у животных и человека. Осуществили безопасное и эффективное введение антисмысловых олигонуклеотидов людям, и многочисленные клинические испытания в настоящее время находятся в стадии реализации. Таким образом, установлено, что олигонуклеотиды могут представлять собой подходящие терапевтические средства, которые могут быть сконфигурированы так, чтобы подходить к режимам лечения для обработки клеток, тканей и животных, особенно людей.
[0097] В вариантах реализации настоящего изобретения олигомерные антисмысловые соединения, в частности, олигонуклеотиды, связываются с молекулами нуклеиновых кислот-мишеней и модулируют экспрессию и/или функцию молекул, кодируемых геном-мишенью. Затрагиваемые функции ДНК включают, например, репликацию и транскрипцию. Затрагиваемые функции РНК включают все жизненно важные функции, такие как, например, транслокация РНК к месту трансляции белков, трансляция белков на матрице РНК, сплайсинг РНК с образованием одного или более видов мРНК и каталитическая активность, которая может присутствовать у РНК или которой РНК может способствовать. Функции могут быть усилены или ингибированы в зависимости от желаемых функций.
[0098] Антисмысловые соединения включают антисмысловые олигомерные соединения, антисмысловые олигонуклеотиды, олигонуклеотиды внешней направляющей последовательности (EGS), сплайс-варианты, праймеры, зонды и другие олигомерные соединения, которые гибридизуются с по меньшей мере частью нуклеиновой кислоты-мишени. Сами по себе эти соединения могут быть введены в форме одноцепочечных, двухцепочечных, частично одноцепочечных или кольцевых олигомерных соединений.
[0099] Нацеливание антисмыслового соединения на молекулу конкретной нуклеиновой кислоты, в контексте настоящего изобретения, может представлять собой многоступенчатый процесс. Процесс обычно начинается с идентификации нуклеиновой кислоты-мишени, функция которой подлежит модулированию. Эта нуклеиновая кислота-мишень может являться, например, клеточным геном (или мРНК, транскрибируемой на матрице гена), экспрессия которого ассоциирована с конкретным расстройством или болезненным состоянием, или молекулой нуклеиновой кислоты из инфекционного агента. Согласно настоящему изобретению, нуклеиновая кислота-мишень кодирует альфа-субъединицу потенциалзависимого натриевого канала (SCNA).
[00100] Процесс нацеливания обычно также включает определение по меньшей мере одной области-мишени, сегмента или сайта в пределах нуклеиновой кислоты-мишени для того, чтобы антисмысловое взаимодействие могло произойти, так, что желаемый эффект, например, модулирование экспрессии, будет достигнут. В контексте настоящего изобретения термин "область" определяют как часть нуклеиновой кислоты-мишени, имеющую по меньшей мере одну поддающуюся идентификации структуру, функцию или характеристику. Внутри областей нуклеиновых кислот-мишеней находятся сегменты. "Сегменты" определяют как меньшие области или субчасти областей в пределах нуклеиновой кислоты-мишени. "Сайты" в настоящем изобретении определяют как позиции в пределах нуклеиновой кислоты-мишени.
[00101] В одном варианте реализации антисмысловые олигонуклеотиды связываются с природными антисмысловыми последовательностями гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и модулируют экспрессию и/или функцию гена SCNA (соответствующего SEQ ID NO: 1-11). Примеры природных антисмысловых последовательностей включают последовательности, соответствующие SEQ ID NOS: 12-28. Примеры антисмысловых олигонуклеотидов включают последовательности, соответствующие SEQ ID NOS: 29-94.
[00102] В одном варианте реализации антисмысловые олигонуклеотиды связываются с одним или более сегментами полинуклеотидов гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и модулируют экспрессию и/или функцию гена SCNA. Сегменты содержат по меньшей мере пять последовательных нуклеотидов смысловых или антисмысловых полинуклеотидов гена SCNA.
[00103] В одном варианте реализации антисмысловые олигонуклеотиды специфичны к природным антисмысловым последовательностям для гена SCNA, при этом связывание олигонуклеотидов с природными антисмысловыми последовательностями для гена SCNA модулирует экспрессию и/или функцию гена SCNA.
[00104] В одном варианте реализации олигонуклеотидные соединения содержат последовательности, соответствующие SEQ ID NOS: 29-94, антисмысловые последовательности, идентифицируемые и раскрываемые с применением, например, ПЦР, гибридизации и т.д. Эти олигонуклеотиды могут содержать один или более модифицированных нуклеотидов, более короткие или более длинные фрагменты, модифицированные связи и т.п. Примеры модифицированных связей или межнуклеотидных связей включают фосфотиоатные, фосфодитиоатные связи и т.п. В одном варианте реализации нуклеотиды содержат фосфатное производное. Фосфатное производное (или модифицированная фосфатная группа), которое может быть присоединено к остатку сахара или аналогу остатка сахара в модифицированных олигонуклеотидах согласно настоящему изобретению, может представлять собой монофосфат, дифосфат, трифосфат, алкилфосфат, алканфосфат, фосфотиоат и т.п. Получение указанных выше аналогов фосфата и их включение в состав нуклеотидов, модифицированных нуклеотидов и олигонуклеотидов, по сути, также известно и не обязательно должно быть описано в настоящей заявке.
[00105] Поскольку, как известно в данной области техники, инициаторный кодон представляет собой, как правило, 5'-AUG (в транскрибируемых молекулах мРНК; 5'-ATG в соответствующей молекуле ДНК), инициаторный кодон также часто называют "AUG кодон", "старт-кодон" или "AUG старт-кодон". Меньшая часть генов имеет инициаторный кодон, содержащий последовательность РНК 5'-GUG, 5'-UUG или 5'-CUG; и 5'-AUA, 5'-ACG и 5'-CUG, как было показано, функционируют т vivo. Таким образом, термин "инициаторный кодон" и "старт-кодон" может охватывать множество последовательностей кодонов, хотя инициаторная аминокислота в каждом случае представляет собой, как правило, метионин (у эукариот) или формилметионин (у прокариот). Эукариотические и прокариотические гены могут содержать два или более альтернативных старт-кодона, любой из которых может быть предпочтительно использован для инициации трансляции в конкретном типе клеток или ткани, или при конкретном наборе условий. В контексте настоящего изобретения, "старт-кодон" и "инициаторный кодон" относятся к кодону или кодонам, которые используются in vivo для инициации трансляции мРНК, транскрибируемой на матрице гена, кодирующего альфа-субъединицу потенциалзависимого натриевого канала (SCNA), независимо от последовательности (последовательностей) таких кодонов. Кодон терминации трансляции (или "стоп-кодон") гена может иметь одну из трех последовательностей, т.е. 5'-UAA, 5'-UAG и 5'-UGA (соответствующими последовательностями ДНК являются 5'-ТАА, 5'-TAG и 5'-TGA, соответственно).
[00106] Термины "область старт-кодона" и "область инициаторного кодона" относятся к части такой мРНК или гена, которая охватывает от примерно 25 до примерно 50 последовательных нуклеотидов в любом направлении (т.е. 5' или 3') от инициаторного кодона. Аналогично, термины "область стоп-кодона" и "область кодона терминации трансляции" относятся к части такой мРНК или гена, охватывает от примерно 25 до примерно 50 последовательных нуклеотидов в любом направлении (т.е. 5' или 3') от кодона терминации трансляции. Следовательно, "область старт-кодона" (или "область инициаторного кодона") и "область стоп-кодона" (или "область кодона терминации трансляции") обе являются областями, на которые могут быть эффективно нацелены антисмысловые соединения согласно настоящему изобретению.
[00107] Открытая рамка считывания (ОРС) или "кодирующая область", которая, как известно в данной области техники, относится к области между инициаторным кодоном и кодоном терминации трансляции, также представляет собой область, на которую может быть эффективно направлено воздействие. В контексте настоящего изобретения область-мишень является внутригенной областью, включающей инициаторный кодон или кодон терминации трансляции открытой рамки считывания (ОРС) гена.
[00108] Другая область-мишень включает 5'-нетранслируемую область (5'-НТО), обозначающую в данной области техники часть мРНК в 5'-направлении от инициаторного кодона и, таким образом, содержащую нуклеотиды между 5'-сайтом кэпирования и инициаторным кодоном мРНК (или соответствующие нуклеотиды гена). Еще одна область-мишень включает 3'-нетранслируемую область (3'-НТО), обозначающую в данной области техники часть мРНК в 3'-направлении от кодона терминации трансляции и, таким образом, содержащую нуклеотиды между кодоном терминации трансляции и 3'-концом мРНК (или соответствующие нуклеотиды гена). 5'-сайт кэпирования мРНК содержит N7-метилированный остаток гуанозина, присоединенный к самому крайнему 5'-остатку мРНК посредством 5'-5'-трифосфатной связи. Область 5'-кэпа мРНК, как считают, включает собственно структуру 5'-кэпа, а также первых 50 нуклеотидов, прилегающие к сайту кэпирования. Другая область-мишень согласно настоящему изобретению представляет собой область 5'-кэпа.
[00109] Несмотря на то, что некоторые эукариотические мРНК-транскрипты прямо транслируются, многие из них содержат одну или более областей, известных как "интроны", которые вырезаются из транскрипта, прежде чем он подвергается трансляции. Остальные (и, таким образом, транслируемые) области известны как "экзоны", и они соединяются друг с другом с образованием непрерывной последовательности мРНК. В одном варианте реализации нацеливание на сайты сплайсинга, т.е. на места соединения интрон-экзон или места соединения экзон-интрон, особенно подходит в случаях, в которых аберрантный сплайсинг вовлечен в развитие заболевания или в которых сверхсинтез конкретного продукта сплайсинга вовлечен в развитие заболевания. Соединение в результате аберрантного слияния вследствие перестройки или делеции представляет собой другой вариант реализации сайта-мишени. мРНК-транскрипты, полученные в результате процесса сплайсинга двух (или более) мРНК из различных источников гена известны как "транскрипты слияния". Нитроны могут быть эффективно задействованы в качестве мишени с применением антисмысловых соединений, нацеленных на, например, ДНК или пре-мРНК.
[00110] В одном варианте реализации антисмысловые олигонуклеотиды связываются с кодирующими и/или некодирующими областями полинуклеотида-мишени и модулируют экспрессию и/или функцию молекулы-мишени.
[00111] В одном варианте реализации антисмысловые олигонуклеотиды связываются с природными антисмысловыми полинуклеотидами и модулируют экспрессию и/или функцию молекулы-мишени.
[00112] В одном варианте реализации антисмысловые олигонуклеотиды связываются со смысловыми полинуклеотидами и модулируют экспрессию и/или функцию молекулы-мишени.
[00113] Альтернативные РНК-транскрипты могут быть получены из одной и той же геномной области ДНК. Эти альтернативные транскрипты в общем случае известны как "варианты". Конкретнее, "варианты пре-мРНК" представляют собой транскрипты, получаемые из одной и той же геномной ДНК, которые отличаются от других транскриптов, получаемых из этой же геномной ДНК, положением их либо старт-, либо стоп-кодона и содержат как интронные, так и экзонные последовательности.
[00114] После удаления одной или более экзонной или интроной областей или их частей в процессе сплайсинга варианты пре-мРНК становятся источниками меньших "вариантов мРНК". Следовательно, варианты мРНК являются процессированными вариантами пре-мРНК, и каждый уникальный вариант пре-мРНК должен всегда давать уникальный вариант мРНК в результате сплайсинга. Эти варианты мРНК также известны как "альтернативные сплайс-варианты". Если сплайсинга варианта пре-мРНК не происходит, то вариант пре-мРНК идентичен варианту мРНК.
[00115] Варианты могут быть получены путем применения альтернативных сигналов для запуска или остановки транскрипции. Пре-мРНК и мРНК могут иметь более одного старт-кодона или стоп-кодона. Варианты, которые происходят из пре-мРНК или мРНК, использующих альтернативные старт-кодоны, известны как "альтернативные старт-варианты" этих пре-мРНК или мРНК. Те транскрипты, которые используют альтернативный стоп-кодон, известны как "альтернативные стоп-варианты" этих пре-мРНК или мРНК. Один конкретный тип альтернативного стоп-варианта представляет собой "полиА-вариант", в котором множественные получаемые транскрипты являются результатом альтернативного выбора одного из "полиА-стоп-сигналов" транскрипционным аппаратом, что приводит к получению транскриптов, которые заканчиваются на уникальных полиА-сайтах. В контексте настоящего изобретения типы вариантов, описанные в настоящей заявке, также представляют собой варианты реализации нуклеиновых кислот-мишеней.
[00116] Области в целевой нуклеиновой кислоте, с которыми гибридизуются антисмысловые соединения, определяют как часть области-мишени длиной по меньшей мере 5 нуклеотидов, на которую нацелено активное антисмысловое соединение.
[00117] В то время как специфические последовательности некоторых типовых сегментов-мишеней приведены в настоящей заявке, как будет обнаружено специалистом в данной области техники, эти последовательности служат для иллюстрации и описания конкретных вариантов реализации в рамках настоящего изобретения. Дополнительные сегменты-мишени легко поддаются идентификации специалистом в данной области техники в свете настоящего описания.
[00118] Сегменты-мишени длиной 5-100 нуклеотидов, содержащие цепь из по меньшей мере пяти (5) последовательных нуклеотидов, выбранных из нуклеотидов в пределах иллюстративных предпочтительных сегментов-мишеней, как полагают, также являются подходящими для нацеливания.
[00119] Сегменты-мишени могут включать последовательности ДНК или РНК, которые содержат по меньшей мере 5 последовательных нуклеотидов с 5'-конца одного из иллюстративных предпочтительных сегментов-мишеней (остальные нуклеотиды представляют собой последовательную цепь той же самой ДНК или РНК, начинающуюся непосредственно выше от 5'-конца сегмента-мишени и продолжающуюся в пределах от примерно 5 до примерно 100 нуклеотидов). Аналогично предпочтительные сегменты-мишени представлены последовательностями ДНК или РНК, которые содержат по меньшей мере 5 последовательных нуклеотидов с 3'-конца одного из иллюстративных предпочтительных сегментов-мишеней (остальные нуклеотиды представляют собой последовательную цепь той же самой ДНК или РНК, начинающуюся непосредственно ниже 3'-конца сегмента-мишени и продолжающуюся в пределах от примерно 5 до примерно 100 нуклеотидов). Специалист в данной области техники, имеющий в наличии сегменты-мишени, проиллюстрированные в настоящей заявке, будет способен без проведения лишних экспериментов идентифицировать далее предпочтительные сегменты-мишени.
[00120] Как только одну или более областей-мишеней, сегментов-мишеней или сайтов-мишеней идентифицируют, выбирают антисмысловые соединения, которые в достаточной степени комплементарны мишени, т.е. гибридизуются достаточно хорошо и с достаточной специфичностью с получением желаемого эффекта.
[00121] В вариантах реализации настоящего изобретения олигонуклеотиды связываются с антисмысловой цепью конкретной мишени. Олигонуклеотиды имеют длину по меньшей мере 5 нуклеотидов и могут быть синтезированы таким образом, что каждый олигонуклеотид нацелен на перекрывающиеся последовательности, так что олигонуклеотиды синтезируют для покрытия всей длины полинуклеотида-мишени. Указанные мишени также включают и кодирующие, и некодирующие области.
[00122] В одном варианте реализации предпочитают воздействовать на конкретные нуклеиновые кислоты антисмысловыми олигонуклеотидами. Нацеливание антисмыслового соединения на конкретную нуклеиновую кислоту представляет собой многоступенчатый процесс. Процесс обычно начинается с идентификации последовательности нуклеиновой кислоты, функция которой подлежит модулированию. Эта последовательность может представлять собой, например, клеточный ген (или мРНК, транскрибируемую на матрице этого гена), экспрессия которого связана с конкретным расстройством или болезненным состоянием, или некодирующий полинуклеотид, такой как, например, некодирующая РНК (нкРНК).
[00123] Молекулы РНК могут быть классифицированы на (1) матричные РНК (мРНК), которые транслируются в белки, и (2) РНК, не кодирующие белки (нкРНК). нкРНК включают микроРНК, антисмысловые транскрипты и другие единицы транскрипции (ЕТ), содержащие высокую плотность стоп-кодонов и не имеющие каких-либо протяженных "открытых рамок считывания". Многие нкРНК, по-видимому, начинаются с сайтов инициации в 3'-нетранслируемых областях (3'-НТО) кодирующих белок локусов. нкРНК, как правило, редки и по меньшей мере половина из нкРНК, которые были просеквенированы Международным консорциумом по анализу РНК (FANTOM), по-видимому, не являются полиаденилированными. Большинство исследователей по очевидным причинам сфокусировали внимание на полиаденилированных мРНК, которые подвергаются процессингу и экспорту в цитоплазму. Недавно было показано, что набор неполиаденилированных ядерных РНК может быть очень большим и что многие такие транскрипты появляются из так называемых межгенных областей. Механизм, посредством которого нкРНК могут регулировать экспрессию генов, заключается в спаривании оснований с транскриптами-мишенями. РНК, которые функционируют за счет спаривания оснований, могут быть сгруппированы в (1) цис-кодируемые РНК, которые кодируются в одном и том же участке гена, но на противоположной цепи по отношению к РНК, на которые они воздействуют, показывая, таким образом, совершенную комплементарность их мишени, и (2) транс-кодируемые РНК, которые кодируются в участках хромосом, отличных от участков для РНК, на которые они воздействуют, и не показывают в совершенной степени потенциального спаривания оснований с их мишенями.
[00124] Без привязки к какой-либо конкретной теории, модификация антисмыслового полинуклеотида антисмысловыми олигонуклеотидами, описанными в настоящей заявке, может изменять экспрессию соответствующих смысловых информационных РНК. Однако такая регуляция может быть либо несогласованной (антисмысловой "нокдаун" приводит к повышению уровня информационной РНК), либо согласованной (антисмысловой "нокдаун" приводит к сопутствующему понижению уровня информационной РНК). В этих случаях антисмысловые олигонуклеотиды могут быть нацелены на перекрывающиеся или неперекрывающиеся части антисмыслового транскрипта, в результате чего происходит его "нокдаун" или секвестрирование. Кодирующие, а также некодирующие антисмысловые {молекулы} можно нацеливать одинаковым образом, и обе категории способны регулировать соответствующие смысловые транскрипты - либо согласованным, либо несогласованным образом. Стратегии, которые применяют для выявления новых олигонуклеотидов для взаимодействия с мишенями, могут быть основаны на "нокдауне" антисмысловых РНК-транскриптов антисмысловыми олигонуклеотидами или на любых других способах модулирования желаемой мишени.
[00125] Стратегия 1: В случае несогласованной регуляции "нокдаун" антисмыслового транскрипта повышает экспрессию обычного (смыслового) гена. В случае, если этот ген кодирует известную или предполагаемую мишень для лекарственного средства, то "нокдаун" его антисмыслового эквивалента может предположительно имитировать действие агониста рецептора или стимулятора фермента.
[00126] Стратегия 2: В случае согласованной регуляции можно одновременно осуществить "нокдаун" как антисмыслового, так и смыслового транскриптов и тем самым добиться синергетического уменьшения экспрессии обычного (смыслового) гена. Если, например, антисмысловой олигонуклеотид применяют для достижения "нокдауна", тогда эта стратегия может быть применена для нацеленного воздействия одним антисмысловым олигонуклеотидом на смысловой транскрипт и другим антисмысловым олигонуклеотидом на соответствующий антисмысловой транскрипт или единственным энергетически симметричным антисмысловым олигонуклеотидом, одновременно нацеленным на перекрывающиеся смысловые и антисмысловые транскрипты.
[00127] Согласно настоящему изобретению, антисмысловые соединения включают антисмысловые олигонуклеотиды, рибозимы, олигонуклеотиды внешней направляющей последовательности (EGS), ммРНК соединения, одно- или двухцепочечные РНК-интерферирующие (РНКи) соединения, такие как ммРНК соединения, и другие олигомерные соединения, которые гибридизуются с по меньшей мере частью нуклеиновой кислоты-мишени и модулируют ее функцию. Соответственно, они могут представлять собой ДНК, РНК, ДНК-подобные соединения, РНК-подобные соединения или их смеси или могут представлять собой миметики одного или более из указанных соединений. Такие соединения могут представлять собой одноцепочечные, двухцепочечные, кольцевые или шпилькообразные олигомерные соединения и могут содержать структурные элементы, такие как внутренние или концевые выпетливания, некомплементарные положения или петли. Антисмысловые соединения обычно получают в виде линейных соединений, но они могут быть соединены или получены иным способом для получения кольцевых и/или разветвленных соединений. Антисмысловые соединения могут включать такие конструкты, как, например, две гибридизованных цепи, образующих полностью или частично двухцепочечное соединение или одиночную цепь с самокомплементарностью, достаточной для гибридизации и образования полностью или частично двухцепочечного соединения. Указанные две цепи могут быть связаны внутренними связями, имея свободные 3'- или 5'-концы, или могут быть связаны с образованием непрерывной шпилькообразной структуры или петли. Указанная шпилькообразная структура может содержать липкий конец либо на 5'-, либо на 3'-конце, образующий удлинение одноцепочечного характера. Указанные двухцепочечные соединения необязательно могут содержать липкие концы. Другие модификации могут включать конъюгированные группы, присоединенные к одному из концов, нуклеотидам в определенных положениях, сахарам в определенных положениях или к одной из межнуклеозидных связей. Альтернативно, указанные две цепи могут быть связаны посредством не-нуклеинового фрагмента или линкерной группы. Образованная только одиночной цепью дцРНК может принимать форму самокомплементарной шпилькообразной молекулы, складывающейся с самой собой с образованием дуплекса. Таким образом, указанные дцРНК могут быть полностью или частично двухцепочечными. Специфическое модулирование экспрессии генов может быть достигнуто за счет стабильной экспрессии шпилек дцРНК в трансгенных клеточных линиях, однако, в некоторых вариантах реализации указанные экспрессия или функция генов увеличиваются. При образовании из двух цепей или одиночной цепи, принимающей форму самокомплементарной шпилькообразной молекулы, складывающейся с самой собой с образованием дуплекса, указанные две цепи (или образующие дуплекс области одиночной цепи) представляют собой комплементарные цепи РНК, которые спариваются основаниями по Уотсону-Крику.
[00128] При попадании в систему, соединения согласно настоящему изобретению могут вызывать действие одного или нескольких ферментов или структурных белков по расщеплению или другим модификациям нуклеиновой кислоты-мишени или могут работать по механизму "занятия положения" (occupancy-based mechanism). В общем случае, нуклеиновые кислоты (в том числе олигонуклеотиды) могут быть описаны как "ДНК-подобные" (т.е. содержащие, как правило, один или более 2'-дезоксисахаров и, как правило, преимущественно Т, а не U основания) или "РНК-подобные" (т.е. содержащие, как правило, один или более 2'-гидрокси- или 2'-модифицированных сахаров и, как правило, преимущественно U, а не Т основания). Спирали нуклеиновых кислот могут быть представлены структурами более чем одного типа, чаще всего А- и В-формами. Считают, что, в общем случае, олигонуклеотиды, которые имеют структуру, подобную В-форме, являются "ДНК-подобными", а олигонуклеотиды, которые имеют структуру, подобную А-форме, являются "РНК-подобными". В некоторых (химерных) вариантах реализации антисмысловое соединение может содержать области как А-, так и В-формы.
[00129] В одном варианте реализации желаемые олигонуклеотиды или антисмысловые соединения содержат по меньшей мере одно из следующего: антисмысловую РНК, антисмысловую ДНК, химерные антисмысловые олигонуклеотиды, антисмысловые олигонуклеотиды, содержащие модифицированные связи, интерферирующие РНК (РНКи), малые интерферирующие РНК (ммРНК); микро интерферирующие РНК (микро-РНК); малые временные РНК (мвРНК); или короткие шпилечные РНК (кшРНК); малые РНК индуцированной активации гена (РНКа); малые активирующие РНК (маРНК) или их комбинации.
[00130] дцРНК может также активировать экспрессию генов, механизм, который получил название "малые РНК-индуцированной активации генов" или РНКа. дцРНК, нацеленные на промоторы генов, вызывают мощную транскрипционную активацию ассоциированных генов. РНКа была продемонстрирована в клетках человека с помощью синтетических дцРНК, называемых "малые активирующие РНК" (маРНК). В настоящее время неизвестно, консервативна ли РНКа в других организмах.
[00131] Было обнаружено, что малые двухцепочечные РНК (дцРНК), такие как малые интерферирующие РНК (ммРНК) и микроРНК (микро-РНК), запускают эволюционно консервативный механизм, известный как РНК-интерференция (РНКи). РНКи неизбежно ведет к выключению (сайленсингу) генов за счет ремоделирования хроматина, что приводит к подавлению транскрипции, деградации комплементарной мРНК или блокированию трансляции белка. Тем не менее, в примерах, подробно описанных в последующей экспериментальной части, показано, что олигонуклеотиды вызывают увеличение экспрессии и/или функции полинуклеотидов гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и кодируемых ими продуктов. дцРНК может также выступать в качестве малых активирующих РНК (маРНК). Без привязки к какой-либо конкретной теории, посредством нацеленного действия на последовательности в промоторах генов-мишеней маРНК индуцируют экспрессию гена-мишени в результате явления, называемого дцРНК-индуцированной транскрипционной активацией (РНКа).
[00132] Согласно другому варианту реализации "предпочтительные сегменты-мишени", определенные в настоящей заявке, можно применять для отбора дополнительных соединений, которые модулируют экспрессию полинуклеотидов гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA). "Модуляторы" представляют собой соединения, понижающие или повышающие экспрессию молекулы нуклеиновой кислоты, кодирующей SCNA, и содержащие по меньшей мере 5-нуклеотидный участок, комплементарный предпочтительному сегменту-мишени. Способ отбора включает этапы контактирования предпочтительного сегмента-мишени молекулы нуклеиновой кислоты, кодирующей смысловые или природные антисмысловые полинуклеотиды SCNA с одним или несколькими потенциальными модуляторами, и отбор одного или более потенциального модулятора, понижающего или повышающего экспрессию молекулы нуклеиновой кислоты, кодирующей полинуклеотиды SCNA, например, SEQ ID NOS: 29-94. После того, как подтверждена способность потенциального(ых) модулятора(ов) к модулированию (например, понижению или повышению) экспрессии молекулы нуклеиновой кислоты, кодирующей полинуклеотиды SCNA, указанный модулятор можно применять для дальнейших исследований функций полинуклеотидов SCNA, или для применения в качестве агента для исследований, диагностики или лечения согласно настоящему изобретению.
[00133] Нацеленное воздействие при помощи природной антисмысловой последовательности предпочтительно модулирует функцию гена-мишени. Например, гена SCNA (в частности, под номерами доступа NM_001165963, NM_021007, NM_006922, NM_000334, NMJ98056, NM_002976, NM_014191, NM_002977, NM_006514, NM_014139, AF109737). Согласно одному из вариантов реализации мишень представляет собой антисмысловой полинуклеотид гена SCNA. Согласно одному из вариантов реализации антисмысловой олигонуклеотид нацелен на смысловые и/или природные антисмысловые последовательности полинуклеотидов SCNA (в частности, под номерами доступа NM_001165963, NM_021007, NM_006922, NM_000334, NM_198056, NM_002976, NM_014191, NM_002977, NM_006514, NM_014139, AF109737), их варианты, аллели, изоформы, гомологи, мутанты, производные, фрагменты и комплементарные последовательности. Предпочтительно, указанный олигонуклеотид представляет собой антисмысловую молекулу, и указанные мишени включают кодирующие и некодирующие участки антисмысловых и/или смысловых полинуклеотидов SCNA.
[00134] Предпочтительные целевые сегменты согласно настоящему изобретению могут также быть скомбинированы с соответствующими комплементарными антисмысловыми соединениями, предложенными в настоящем изобретении, с формированием стабилизированных двухцепочечных (дуплексных) олигонуклеотидов.
[00135] В данной области техники показано, что такие двухцепочечные олигонуклеотидные фрагменты модулируют экспрессию мишеней и регулируют трансляцию, а также процессинг РНК, посредством антисмыслового механизма. Кроме того, указанные двухцепочечные фрагменты могут подвергаться химическим модификациям. Например, показано, что такие двухцепочечные фрагменты ингибируют мишень посредством классической гибридизации антисмысловой цепи указанного дуплекса с мишенью, запуская тем самым ферментное расщепление мишени.
[00136] Согласно одному из вариантов реализации антисмысловой олигонуклеотид нацелен на полинуклеотиды гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) (в частности, под номерами доступа NM_001165963, NM_021007, NM_006922, NM_000334, NM_198056, NM_002976, NM_014191, NM_002977, NM_006514, NM_014139, AF109737), их варианты, аллели, изоформы, гомологи, мутанты, производные, фрагменты и комплементарные последовательности. Предпочтительно, указанный олигонуклеотид представляет собой антисмысловую молекулу.
[00137] Согласно вариантам реализации настоящего изобретения целевая молекула нуклеиновой кислоты не ограничивается собственно SCNA и может быть представлена любыми изоформами, рецепторами, гомологами и т.п. молекулами SCNA.
[00138] Согласно одному из вариантов реализации олигонуклеотид нацелен на природную антисмысловую последовательность полинуклеотидов SCNA, например, полинуклеотидов, представленных в последовательностях SEQ ID NOS: 12-28, и любых их вариантов, аллелей, гомологов, мутантов, производных, фрагментов и комплементарных последовательностей. Примеры антисмысловых олигонуклеотидов представлены последовательностями SEQ ID NOS: 29-94.
[00139] Согласно одному из вариантов реализации указанные олигонуклеотиды комплементарны или связываются с нуклеиновыми кислотами антисмысловых последовательностей SCNA, включая, не ограничиваясь указанными, некодирующие смысловые и/или антисмысловые последовательности, связанные с полинуклеотидами SCNA, и модулируют экспрессию и/или функцию молекул SCNA.
[00140] Согласно одному варианту реализации указанные олигонуклеотиды комплементарны природным антисмысловым последовательностям нуклеиновых кислот SCNA, представленным в последовательностях SEQ ID NOS: 12-28, или связываются с ними, и модулируют экспрессию и/или функцию молекул SCNA.
[00141] Согласно одному из вариантов реализации олигонуклеотиды содержат последовательности, состоящие по меньшей мере из 5 последовательных нуклеотидов последовательностей SEQ ID NOS: 29-94, и модулируют экспрессию и/или функцию молекул SCNA.
[00142] Указанные полинуклеотидные мишени включают SCNA, в том числе представителей этого семейства, варианты SCNA; мутанты SCNA, в том числе ОНП; некодирующие последовательности SCNA; аллели SCNA; видовые варианты, фрагменты и т.п. Предпочтительно, указанный олигонуклеотид представляет собой антисмысловую молекулу.
[00143] В одном из вариантов реализации олигонуклеотид, который нацелен на полинуклеотиды SCNA, включает: антисмысловую РНК, интерферирующую РНК (мРНК), малую интерферирующую РНК (ммРНК); микро интерферирующую РНК (микроРНК), малую, временную РНК (мвРНК), или короткую шпилечную РНК (кшРНК); малую РНК-индуцированной активации гена (РНКа), или малую активирующую РНК (маРНК).
[00144] Согласно одному из вариантов реализации нацеленное воздействие на полинуклеотиды гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA), например, SEQ ID NOS: 1-11, модулирует экспрессию или функцию таких мишеней. Согласно одному из вариантов реализации экспрессия или функция регулируются повышающе по сравнению с контролем. Согласно одному из вариантов реализации экспрессия или функция регулируются понижающе по сравнению с контролем. В еще одном варианте реализации нацеленное воздействие на природные антисмысловые транскрипты (например, соответствующие SEQ ID NOS: 12-28), а также любые другие NAT-мишени таких целевых полинуклеотидов приводит к усилению экспрессии указанной целевой мРНК и соответствующего белка.
[00145] Согласно одному из вариантов реализации антисмысловые соединения включают последовательности, представленные в последовательностях SEQ ID NOS: 29-94. Такие олигонуклеотиды могут содержать один или более модифицированный нуклеотид, более короткие или более длинные фрагменты, модифицированные связи и т.п.
[00146] Согласно одному из вариантов реализации последовательности SEQ ID NOS: 29-94 содержат один или более ЗНК-нуклеотид. В Таблице 1 приведены типовые антисмысловые олигонуклеотиды, подходящие для применения в способах согласно настоящему изобретению.
Таблица 1:
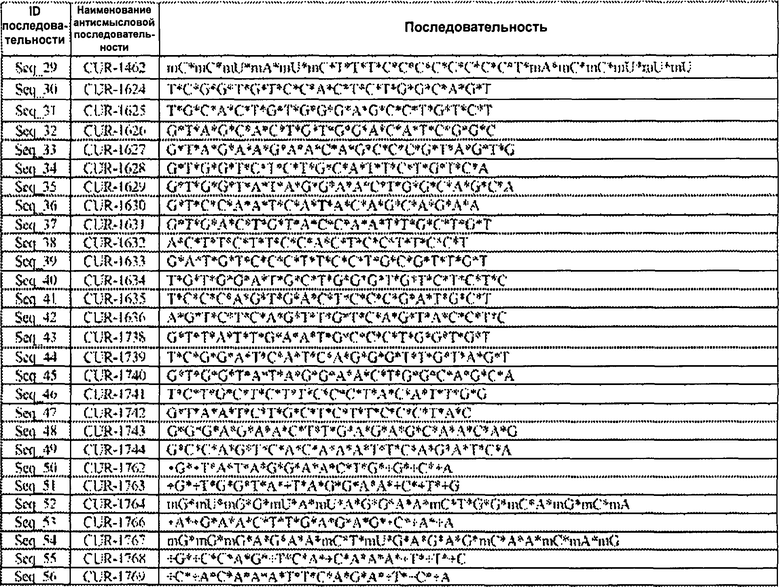
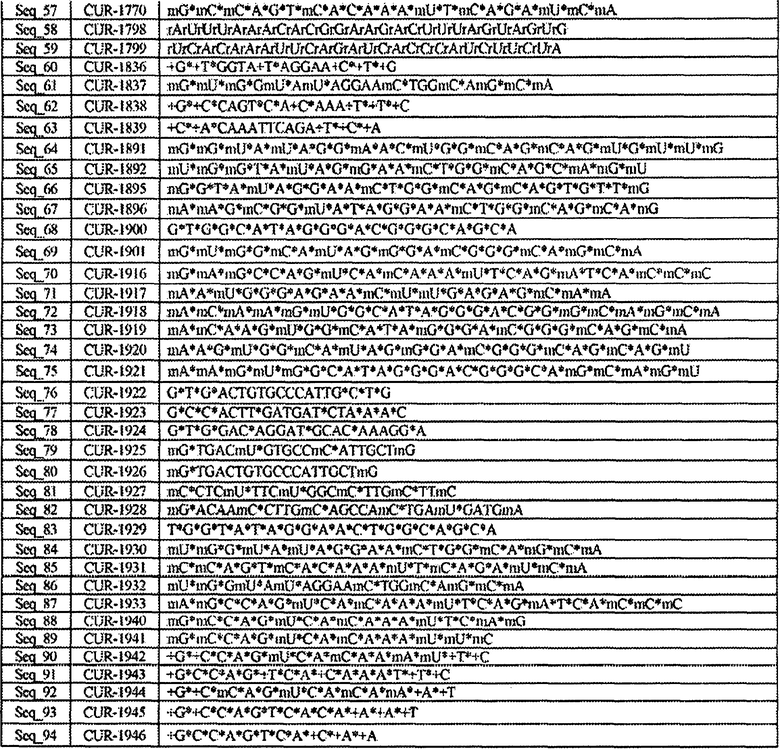
[00147] Символ "*" обозначает фосфотиоатную связь, "+" обозначает ЗНК, "r" обозначает РНК, а "m" указывает на наличие метильной группы на 2'-атоме кислорода в обозначенном сахарном остатке олигонуклеотида. Во избежание неясности, эта ЗНК имеет формулу:
где В представляет собой конкретное обозначенное основание.
[00148] Таблица 2: Относительная экспрессия мРНК гена SCN1A клетках, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCN1A-специфичный природный антисмысловой транскрипт. Ср - средняя кратность изменения экспрессии SCN1A по сравнению с ложнотрансфицированным контролем; СО - стандартное отклонение, Р - вероятность того, что обработанные образцы не отличаются от ложнотрансфицированного контроля. N - общее число повторностей.
[00149] Модулирование нужной целевой нуклеиновой кислоты может быть осуществлена несколькими известными в данной области техники способами. Например, антисмысловые олигонуклеотиды, ммРНК и т.д. Молекулы ферментативной нуклеиновой кислоты (например, рибозимы) представляют собой молекулы нуклеиновой кислоты, способные катализировать одну или несколько разнообразных реакций, включая способность многократно расщеплять другие отдельные молекулы нуклеиновой кислоты чувствительным к последовательности нуклеиновых оснований сиквенс-специфичным образом. Такие молекулы ферментативной нуклеиновой кислоты могут применяться, например, для нацеленного действия на практически любой РНК-транскрипт.
[00150] Из-за сиквенс-специфичности транс-расщепляющие молекулы ферментативной нуклеиновой кислоты являются многообещающими потенциальными терапевтическими агентами для лечения заболеваний человека. Ферментативные молекулы нуклеиновых кислот могут быть сконструированы для расщепления конкретных РНК мишеней в общем пуле клеточной РНК. Такое расщепление делает указанные мРНК нефункциональными и отменяет экспрессию белка с этой РНК. Таким образом, синтез белка, связанного с болезненным состоянием может быть избирательно заблокирована.
[00151] В общем случае, ферментативные нуклеиновые кислоты с активностью по расщеплению РНК действуют, во-первых, за счет связывания с РНК-мишенью. Такое связывание происходит посредством мишень-связывающего фрагмента ферментативной нуклеиновой кислоты, который располагается в непосредственной близости к ферментативному фрагменту указанной молекулы, расщепляющему целевую РНК. Таким образом, указанная ферментативная нуклеиновая кислота сначала распознает целевую РНК, а потом связывается с ней посредством комплементарного спаривания оснований, и после связывания с соответствующим участком ферментативно расщепляет целевую РНК. Стратегическое расщепление такой целевой РНК уничтожит ее способность к прямому синтезу кодируемого белка. После того, как ферментативная нуклеиновая кислота связала и расщепила целевую РНК, она отсоединяется от этой РНК, освобождается для другой мишени и может повторно связывать и расщеплять новые мишени.
[00152] Ряд подходов, таких как т vitro стратегии отбора (извлечения) (Orgel, (1979) Proc. R. Soc. London, В 205, 435), был использован для получения новых нуклеиновых катализаторов, способных катализировать различные реакции, такие как расщепление и лигирование фосфодиэфирных связей и амидных связей.
[00153] Получение рибозимов, обладающих оптимальной каталитической активностью, внесет существенный вклад в любую стратегию, включающую применение РНК-расщепляющих рибозимов для регулирования генной экспрессии. Рибозим типа «головка молотка», например, функционирует со скоростью каталитической реакции (kcat) приблизительно 1 мин-1 в присутствии насыщающих (10 мМ) концентраций кофактора Mg2+. Показано, что синтетический «РНК-лигазный» рибозим катализирует соответствующую реакцию самоизменения со скоростью приблизительно 100 мин-1. Кроме того, известно, что определенные модифицированные рибозимы типа «головка молота», имеющие субстрат-связывающие ДНК-»ручки», катализируют расщепление РНК со скоростями оборотов, достигающими 100 мин-1. Наконец, замещение специфического остатка каталитического ядра «головки молота» определенными нуклеотидными аналогами дает модифицированные рибозимы, демонстрирующие повышение скорости каталитической реакции вплоть до 10-кратного. Эти результаты показывают, что рибозимы могут обеспечивать химические преобразования с каталитическими скоростями, значительно превышающими таковые, демонстрируемые in vitro большинством природных саморасщепляющих рибозимов. В этом случае возможно, что структуры некоторых саморасщепляющих рибозимов можно оптимизировать, чтобы получить максимальную каталитическую активность, или что могут быть получены совершенно новые РНК мотивы, демонстрирующие значительно увеличенные скорости фосфодиэфирного расщепления РНК.
[00154] Внутримолекулярное расщепление РНК-субстрата РНК катализатором, соответствующим модели «головка молота», впервые было показано в 1987 (Uhlenbeck, О.С. (1987) Nature, 328: 596-600). Указанный РНК катализатор был выделен и вступал в реакцию с различными РНК-молекулами, подтверждая, что он действительно является катализатором.
[00155] Каталитические РНК, сконструированные на основе мотива «головка молота», применялись для расщепления специфических целевых последовательностей осуществлением подходящего спаривания оснований в каталитической РНК для обеспечения необходимого спаривания оснований с целевыми последовательностями. Это позволило применить указанную каталитическую РНК для расщепления специфических целевых последовательностей и показать, что каталитические РНК, сконструированные согласно модели «головка молота», предположительно способны расщеплять специфические субстратные РНК in vivo.
[00156] РНК-интерференция (РНКи) стала мощным инструментом модулирования генной экспрессии у млекопитающих и в клетках млекопитающих. Этот подход предполагает получение малых интерферирующих РНК (ммРНК) в виде самой РНК или в виде ДНК, с применением экспрессионной плазмиды или вируса и кодирующей последовательности для малых шпилечных РНК, которые процессируются в ммРНК. Эта система обеспечивает эффективный транспорт пре-ммРНК в цитоплазму, где они проявляют активность, и позволяет применение регулируемых и тканеспецифичных промоторов генной экспрессии
[00157] Согласно одному из вариантов реализации олигонуклеотид или антисмысловое соединение содержит олигомер или полимер рибонуклеиновой кислоты (РНК) и/или дезоксирибонуклеиновой кислоты (ДНК), или их миметик, химеру, аналог или гомолог. Этот термин включает олигонуклеотиды, состоящие из встречающихся в природе нуклеотидов, сахаров и ковалентных межнуклеозидных (скелетных) связей, а также олигонуклеотиды, содержащие не встречающиеся в природе фрагменты, функционирующие сходным образом. Такие модифицированные или замещенные олигонуклеотиды часто предпочтительнее нативных форм, так как они обладают необходимыми свойствами, такими как, например, увеличенное поглощение клетками, увеличенная связывающая способность в отношении целевой нуклеиновой кислоты и повышенная стабильность в присутствии нуклеаз.
[00158] Согласно настоящему изобретению указанные олигонуклеотиды или «антисмысловые соединения» включают антисмысловые олигонуклеотиды (например, РНК, ДНК, их миметики, химеры, аналоги или гомологи), рибозимы, олигонуклеотиды внешних вспомогательных последовательностей (EGS), ммРНК соединения, одно- или двухцепочечные РНК-интерферирующие (РНКи) соединения, такие как ммРНК соединения, каРНК, аРНК, и другие олигомерные соединения, которые гибридизуются по меньшей мере с частью целевой нуклеиновой кислоты и модулируют ее функцию. Соответственно, они могут представлять собой ДНК, РНК, ДНК-подобные соединения, РНК-подобные соединения или их смеси, или могут представлять собой миметики одного или более из указанных соединений. Такие соединения могут представлять собой одноцепочечные, двухцепочечные, кольцевые или шпилькообразные олигомерные соединения и могут содержать структурные элементы, такие как внутренние или концевые выпетливания, некомплементарные положения или петли. Антисмысловые соединения обычно получают в виде линейных соединений, но они могут быть объединены или получены иным способом для получения кольцевых или и/или разветвленных соединений. Антисмысловые соединения могут включать такие конструкты, как, например, две гибридизованных цепи, формирующих полностью или частично двухцепочечное соединение, или одиночная цепь с достаточной самокомплементарностью для гибридизации и формирования полностью или частично двухцепочечного соединения. Указанные две цепи могут быть связаны внутренними связями, имея свободные 3' или 5' концы, или могут связываться с образованием непрерывной шпилькообразной структуры или петли. Указанная шпилькообразная структура может содержать липкий конец на 5' или на 3' конце, образующий удлинение одноцепочечного характера. Указанные двухцепочечные соединения в некоторых случаях могут содержать липкие концы. Другие модификации могут включать конъюгированные группы, присоединенные к одному из концов, нуклеотидам в определенных положениях, сахарам в определенных положениях или к одной из межнуклеозидных связей. Как вариант, указанные две цепи могут быть связаны посредством не-нуклеинового фрагмента или линкерной группы. Сформированная одиночной цепью дцРНК может принимать форму самокомплементарной шпилькообразной молекулы, складывающейся с самой собой с образованием дуплекса. Таким образом, указанные дцРНК могут быть полностью или частично двухцепочечными. Специфическое модулирование генной экспрессии может быть достигнуто стабильной экспрессией дцРНК-шпилек в трансгенных клеточных линиях. При формировании из двух цепей или одиночной цепи, принимающей форму самокомплементарной шпилькообразной молекулы, складывающейся с самой собой с формированием дуплекса, указанные две цепи (или формирующие дуплекс участки одиночной цепи) представляют собой комплементарные цепи РНК, которые спариваются по Уотсону-Крику.
[00159] При введении в систему соединения, предложенные в настоящем изобретении, могут приводить к активации одного или более ферментов или структурных белков, влияя на расщепление или иные модификации целевой нуклеиновой кислоты, либо могут действовать через механизмы «заполнения». Как правило, нуклеиновые кислоты (включая олигонуклеотиды) могут быть названы «ДНК-подобными» (т.е., как правило, содержащими один или более 2'-дезокси-сахар и, как правило, преимущественно Т, а не U, основания) или «РНК-подобными» (т.е., как правило, содержащими один или более 2'- гидроксил или 2'-модифицированные сахара и, как правило, преимущественно Т, а не U, основания). Спирали нуклеиновых кислот могут быть представлены структурами более чем одного типа, чаще всего А- и В-формами. Предполагают, что, как правило, олигонуклеотиды, имеющие структуру типа В-формы, «ДНК-подобны», а имеющие структуру типа А-формы, «РНК-подобны». Согласно некоторым вариантам реализации (с химерами) антисмысловое соединение может содержать участки как А-, так и В-формы.
[00160] Антисмысловые соединения согласно настоящему изобретению могут содержать антисмысловой фрагмент приблизительно от 5 до приблизительно 80 нуклеотидов (т.е. приблизительно от 5 до приблизительно 80 связанных нуклеозидов) длиной. Это относится к длине указанной антисмысловой цепи или части указанного антисмыслового соединения. Иными словами, одноцепочечное антисмысловое соединение, предложенное в настоящем изобретении, содержит от 5 до приблизительно 80 нуклеотидов, а двухцепочечное антисмысловое соединение, предложенное в настоящем изобретении (такие как, например, дцРНК), содержит смысловую и антисмысловую цепь или фрагменты от 5 до приблизительно 80 нуклеотидов длиной. Специалисту в данной области техники будет ясно, что под этим понимаются антисмысловые фрагменты, составляющие в длину 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79 или 80 нуклеотидов, или принадлежащие любому диапазону в пределах указанного.
[00161] Согласно одному из вариантов реализации антисмысловые соединения, предложенные в настоящем изобретении, содержат антисмысловые фрагменты 10-50 нуклеотидов длиной. Специалисту в данной области техники будет ясно, что сюда включены олигонуклеотиды, содержащие антисмысловые фрагменты, составляющие в длину 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, или 50 нуклеотидов, или принадлежащие любому диапазону в пределах указанного. Согласно некоторым вариантам реализации длина указанных олигонуклеотидов составляет 15 нуклеотидов.
[00162] Согласно одному из вариантов реализации антисмысловые или олигонуклеотидные соединения, предложенные в настоящем изобретении, содержат антисмысловые фрагменты, составляющие от 12 или 13 до 30 нуклеотидов в длину. Специалисту в данной области техники будет ясно, что сюда включены антисмысловые соединения, содержащие антисмысловые фрагменты, составляющие в длину 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидов, или принадлежащие любому диапазону в пределах указанного.
[00163] Согласно одному из вариантов реализации олигомерные соединения, предложенные в настоящем изобретении, также включают варианты, в которых в одном или нескольких положениях нуклеотидов указанного соединения присутствуют различающиеся основания. Например, если первый нуклеотид представляет собой аденозин, могут быть получены варианты, которые содержат тимидин, гуанозин или цитидин в этом положении. Это может быть осуществлено в любом положении указанного антисмыслового или дцРНК-соединения. Такие соединения затем тестируют с применением способов, описанных в настоящей заявке, для определения их способности ингибировать экспрессию целевой нуклеиновой кислоты.
[00164] Согласно некоторым вариантам реализации степень гомологии, идентичности последовательностей или комплементарности между указанным антисмысловым соединением и мишенью составляет от приблизительно 40% до приблизительно 60%. Согласно некоторым вариантам реализации степень гомологии, идентичности последовательностей или комплементарности составляют приблизительно от 60% до приблизительно 70%. Согласно некоторым вариантам реализации степень гомологии, идентичности последовательностей или комплементарности составляют приблизительно от 70% до приблизительно 80%. Согласно некоторым вариантам реализации степень гомологии, идентичности последовательностей или комплементарности составляют приблизительно от 80% до приблизительно 90%. Согласно некоторым вариантам реализации гомология, идентичность последовательностей или комплементарность составляют приблизительно 90%, приблизительно 92%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или приблизительно 100%.
[00165] Согласно одному из вариантов реализации антисмысловые олигонуклеотиды, такие как, например, молекулы нуклеиновой кислоты, представленные в последовательностях SEQ ID NOS: 29-94, включают одну или более чем одну замену или модификацию. Согласно одному из вариантов реализации указанные нуклеотиды заменены закрытыми нуклеиновыми кислотами (ЗНК).
[00166] Согласно одному из вариантов реализации указанные олигонуклеотиды нацеленно взаимодействуют с одним или более участком молекул нуклеиновой кислоты смысловых и/или антисмысловых кодирующих и/или некодирующих последовательностей, связанных с SCNA, и последовательностей, представленных в последовательностях SEQ ID NOS: 1-28. Олигонуклеотиды также нацелены на перекрывающиеся участки, соответствующие SEQ ID NOS: 1-28.
[00167] Некоторые предпочтительные олигонуклеотиды, предложенные в настоящем изобретении, представляют собой химерные олигонуклеотиды. «Химерные олигонуклеотиды» или «химеры» в контексте настоящего изобретения представляют собой олигонуклеотиды, которые содержат два или более химически отличных участка, каждый из которых состоит по меньшей мере из одного нуклеотида. Такие олигонуклеотиды, как правило, содержат по меньшей мере один участок модифицированных нуклеотидов, обеспечивающий одно или более полезное свойство (такое как, например, увеличенная устойчивость к нуклеазам, увеличенное поглощение клетками, повышенная связывающая способность по отношению к мишени) и участок, который представляет собой субстрат для ферментов, способных к расщеплению РНК:ДНК или РНК: РНК гибридов. Как пример, РНКаза Н представляет собой клеточную эндонуклеазу, которая расщепляет РНК-цепь РНК: ДНК дуплекса. Активация РНКазы Н, таким образом, приводит к расщеплению целевой РНК, тем самым значительно повышая эффективность антисмыслового модулирования генной экспрессии. Соответственно, при использовании химерных олигонуклеотидов сопоставимые результаты часто могут быть получены с более короткими олигонуклеотидами, по сравнению с фосфоротиоатными дезоксиолигонуклеотидами, гибридизующимися с тем же целевым участком. Расщепление целевой РНК может быть определено стандартным способом посредством гель-электрофореза и, при необходимости, сопутствующих методик гибридизации нуклеиновых кислот, известных в данной области техники. Согласно одному из вариантов реализации химерный олигонуклеотид содержит по меньшей мере один участок, модифицированный для увеличения способности связываться с мишенью, и, как правило, участок, функционирующий как субстрат РНКазы Н. Связывающая способность олигонуклеотида по отношению к мишени (в данном случае, нуклеиновой кислоте, кодирующей ras) определяется стандартным способом измерением температуры плавления пары олигонуклеотид/мишень, представляющим собой температуру, при которой олигонуклеотид и мишень диссоциируют; диссоциацию определяют спектрофотометрически. Чем выше температура плавления, тем больше связывающая способность указанного олигонуклеотида по отношению к мишени.
[00168] Химерные антисмысловые соединения, предложенные в настоящем изобретении, могут быть получены в виде составных структур из двух или более олигонуклеотидов, модифицированных олигонуклеотидов, олигонуклеозидов и/или миметиков олигонуклеотидов согласно приведенным выше описаниям. Такие соединения также известны в данной области техники как гибриды или гапмеры. Примеры патентов США, в которых описано получение таких гибридных структур, включают, не ограничиваясь перечисленными, патенты США 5013830; 5149797; 5220007; 5256775; 5366878; 5403711; 5491133; 5565350; 5623065; 5652355; 5652356; и 5700922, каждый из которых включен в настоящую заявку посредством ссылки.
[00169] Согласно одному из вариантов реализации модифицируемый участок указанного олигонуклеотида содержит по меньшей мере один нуклеотид, модифицированный по 2' положению сахара, наиболее предпочтительно 2'-O-алкил, 2'-O-алкил-O-алкил или 2'-фтор-модифицированный нуклеотид. Согласно другому варианту реализации РНК модификации включают 2'-фтор, 2'-амино и 2'-O-метильные модификации рибозы пиримидинов, лишенные азотистого основания остатки или обратное основание на 3'-конце указанной РНК. Такие модификации олигонуклеотидов осуществляют стандартными способами; показано, что такие олигонуклеотиды имеют более высокую температуру плавления (т.е. большую способность связываться с мишенью), чем 2'-дезоксиолигонуклеотиды, относительно определенной мишени. В результате такой повышенной связывающей способности значительно усиливается ингибирование генной экспрессии РНКи-олигонуклеотидом. РНКаза Н представляет собой клеточную эндонуклеазу, расщепляющую РНК цепь РНК:ДНК дуплексов; активация этого фермента, таким образом, приводит к расщеплению целевой РНК, и, следовательно, может значительно увеличивать эффективность РНКи ингибирования. Расщепление целевой РНК может быть подтверждено обычным способом посредством гель-электрофореза. Согласно одному из вариантов реализации указанный химерный олигонуклеотид также модифицирован для увеличения устойчивости к нуклеазе. Клетки содержат множество экзо- и эндонуклеаз, способных разлагать нуклеиновые кислоты. Показано, что некоторые нуклеотидные и нуклеозидные модификации придают олигонуклеотиду, в состав которого они входят, большую устойчивость к нуклеазному расщеплению по сравнению с нативным олигодезоксинуклеотидом. Устойчивость к нуклеазам измеряют обычным способом, инкубируя олигонуклеотиды с клеточными экстрактами или растворами с выделенными нуклеазами и измеряя содержание интактных олигонуклеотидов спустя определенное время, как правило, посредством гель-электрофореза. Олигонуклеотиды, модифицированные для увеличения устойчивости к нуклеазам, остаются интактными на протяжении более длительного времени по сравнению с немодифицированными олигонуклеотидами. Показано, что ряд модификаций олигонуклеотидов усиливают или придают им устойчивость к нуклеазам. Олигонуклеотиды, которые содержат по меньшей мере одну фосфоротиоатную модификацию, на настоящий момент более предпочтительны. В некоторых случаях модификации олигонуклеотидов, усиливающие способность связываться с мишенью также, независимым образом, способны повышать устойчивость к нуклеазам.
[00170] Специфические примеры некоторых предпочтительных олигонуклеотидов, предусмотренных настоящим изобретением, включают содержащие модифицированные остовы, например, фосфоротиоаты, фосфотриэфиры, метилфосфонаты, алкильные с короткой цепью или циклоалкильные связи между сахарами или гетероатомные с короткой цепью или гетероциклические связи между сахарами. Наиболее предпочтительными являются олигонуклеотиды с фосфоротиоатными остовами и с гетероатомными остовами, в частности, СН2-NH-O-CH2, CH, -N(CH3)-O-CH2 [известный как метилен(метилимино) или MMI остов], СН2-O-N(СН3)-СН2, СН2-N(CH3) -N(СН3)-СН2 и O-N (СН3)-СН2 -СН2 остовы, где нативный фосфодиэфирный остов представлен в виде O-Р-О-СН,). Амидные остовы, описанные в De Mesmaeker et al. (1995) Асе. Chem. Res. 28: 366-374, также являются предпочтительными. Также предпочтительными являются олигонуклеотиды, содержащие морфолиновые остовы (Summerton and Weller, патент США 5034506). Согласно другому варианту реализации, такому как остов на основе пептидной нуклеиновой кислоты (ПНК), фосфодиэфирный остов олигонуклеотида заменен полиамидным остовом, нуклеотиды связаны прямо или непрямо с азотными атомами азагрупп полиамидного остова. Олигонуклеотиды могут также содержать один или более фрагмент замещенного сахара. Предпочтительные олигонуклеотиды содержат один из следующих заместителей в положении 2': ОН, SH, SCH3, F, OCN, ОСН3 ОСН3, ОСН3 O(CH2)n CH3, O(CH2)n NH2 или O(СН2)n CH3, где n принимает значения от 1 до приблизительно 10; С1-С10 низшие алкилы, алкоксиалкокси, замещенные низшие алкилы, алкарил или аралкил; Cl; Br; CN; CF3; OCF3; O-, S-, или N-алкил; O-, S-, или N-алкенил; SOCH3; SO2 CH3; ONO2; NO2; N3; NH2; гетероциклоалкил; гетероциклоалкарил; аминоалкиламино; полиалкиламино; замещенные силилы; РНК расщепляющую группу; репортерную группу; интеркалятор; группу, улучшающую фармакокинетические свойства олигонуклеотида; или группу, улучшающую фармакодинамические свойства олигонуклеотида, и другие заместители, обладающие сходными свойствами. Предпочтительная модификация включает 2'-метоксиэтокси [2'-O-СН2 СН2 ОСН3, также известный как 2'-O-(2-метоксиэтил)]. Другие предпочтительные модификации включают 2'-метокси (2'-O-СН3), 2'-пропокси (2'-ОСН2 СН2СН3) и 2'-фтор (2'-F). Сходные модификации могут быть также осуществлены в других положениях олигонуклеотида, в частности, по 3' положению сахара 3' концевого нуклеотида и 5' положению 5' концевого нуклеотида. Олигонуклеотиды могут также содержать миметики сахаров, такие как циклобутилы, вместо пентофуранозильной группы.
[00171] Олигонуклеотиды могут также содержать, дополнительно или альтернативно, модификации или замены нуклеинового основания (часто называемого в данной области техники просто «основанием»). Используемые в настоящей заявке термины «немодифицированные» или «природные» нуклеотиды включают аденин (А), гуанин (G), тимин (Т), цитозин (С) и урацил (U). Модифицированные нуклеотиды включают нуклеотиды, обнаруживаемые в природных нуклеиновых кислотах редко или временно, например, гипоксантин, 6-метиладенин, 5-Ме пиримидины, в частности, 5-метилцитозин (также называемый 5-метил-2' дезоксицитозин и часто упоминаемый в данной области техники как 5-Ме-С), 5-гидроксиметилцитозин (ГМЦ), гликозил ГМЦ и гентобиозил ГМЦ, а также синтетические нуклеотиды, например, 2-аминоаденин, 2-(метиламино)аденин, 2-(имидазолилалкил)аденин, 2-(аминоалкиламино)аденин или другие гетерозамещенные алкиладенины, 2-тиоурацил, 2-тиотимин, 5-бромурацил, 5-гидроксиметилурацил, 8-азагуанин, 7-деазагуанин, N6 (6-аминогексил)аденин и 2,6-диаминопурин. «Универсальное» основание из известных в данной области техники, например, инозин, может также быть включено. Показано, что 5-Ме-С замещения увеличивают стабильность дуплексов нуклеиновых кислот на 0,6-1,2°С и на настоящий момент являются предпочтительными замещениями оснований.
[00172] Другая модификация олигонуклеотидов, предложенных в настоящем изобретении, включает химическое связывание с указанным олигонуклеотидом одного или нескольких фрагментов или конъюгатов, увеличивающих активность или поглощение клетками указанного олигонуклеотида. Такие фрагменты включают, не ограничиваясь перечисленными, липидные фрагменты, такие как фрагменты холестерина, фрагмент холестерила, алифатические цепи, например, додекандиольные или ундецильные остатки, цепь полиамина или полиэтиленгликоля, или адамантан-уксусную кислоту. Олигонуклеотиды, содержащие липофильные фрагменты, и способы получения таких олигонуклеотидов известны в данной области техники, см., например, патенты США 5138045, 5218105 и 5459255.
[00173] Необязательно, чтобы все положения конкретного нуклеотида были модифицированы единообразно, и на самом деле в одном олигонуклеотиде, или даже в одном нуклеозиде в составе олигонуклеотида может содержаться более одной из упомянутых выше модификаций. Настоящее изобретение также включает Олигонуклеотиды, которые представляют собой химерные Олигонуклеотиды согласно данному выше определению.
[00174] Согласно другому варианту реализации молекула нуклеиновой кислоты, предложенная в настоящем изобретении, конъюгирована с другим фрагментом, включая, но не ограничиваясь перечисленными, лишенные оснований нуклеотиды, полиэфиры, полиамины, полиамиды, пептиды, углеводороды, липиды или полиуглеводородные соединения. Специалистам в данной области техники будет ясно, что такие молекулы могут быть связаны с любым одним или более чем одним нуклеотидом, содержащим молекулу нуклеиновой кислоты, в нескольких положениях на сахаре, основании или фосфатной группе.
[00175] Олигонуклеотиды, применяемые согласно настоящему изобретению, могут быть удобно и просто получены с применением общеизвестной техники твердофазного синтеза. Оборудование для такого синтеза коммерчески доступно от ряда поставщиков, включая Applied Biosystems. Также можно применять любые другие способы такого синтеза; фактический синтез указанного олигонуклеотида не представляет трудностей для среднего специалиста в данной области техники. Также общеизвестна возможность использования сходных методик для получения других олигонуклеотидов, таких как фосфоротиоатные и алкилатные производные. Также общеизвестна возможность использования сходных техник и коммерчески доступных модифицированных амидитов и продуктов на основе стекла с контролируемым размером пор (CPG), таких как биотин-, флуоресцеин-, акридин- или псорален-модифицированные амидиты и/или CPG (доступные от Glen Research, Sterling VA) для синтеза флуоресцентно меченых, биотинилированных или других модифицированных олигонуклеотидов, таких как холестерин-модифицированные олигонуклеотиды.
[00176] В соответствии с настоящим изобретением подразумевается применение модификаций, например, применение мономеров ЗНК для усиления силы, специфичности и продолжительности действия и расширение диапазона путей введения олигонуклеотидов, состоящих из конкретных компонентов, таких как МОЭ, АНК, ФАНК, ФТ и т.д. Это может быть достигнуто замещением некоторых мономеров в конкретных олигонуклеотидах ЗНК-мономерами. ЗНК-модифицированный олигонуклеотид может иметь размер, соответствующий исходному соединению, или может быть больше, либо, предпочтительно, меньше. Предпочтительно, чтобы такие ЗНК-модифицированные олигонуклеотиды содержали менее чем приблизительно 70%, более предпочтительно менее чем приблизительно 60%, наиболее предпочтительно менее чем приблизительно 50% мономеров ЗНК, и чтобы их размеры составляли приблизительно от 5 до 25 нуклеотидов, более предпочтительно приблизительно от 12 до 20 нуклеотидов.
[00177] Предпочтительные модифицированные олигонуклеотидные остовы включают, не ограничиваясь перечисленными: фосфоротиоаты, хиральные фосфоротиоаты, фосфородитиоаты, фосфотриэфиры, аминоалкилфосфотриэфиры, метил- и другие алкилфосфонаты, включая 3'алкиленфосфонаты и хиральные фосфонаты, фосфинаты, фосфорамидаты, включая 3'-амино фосфорамидат и аминоалкилфосфорамидаты, тионофосфорамидаты, тионоалкилфосфонаты, тионоалкилфосфотриэфиры, и боранофосфаты, включающие нормальные 3'-5' связи, 2'-5' связанные аналоги, и имеющие обращенную полярность, где соседние пары нуклеозидных единиц связаны 3'-5' к 5'-3' или 2'-5' к 5'-2'. Различные соли, смешанные соли и свободные кислоты также включены.
[00178] Примеры патентов США, раскрывающих получение вышеприведенных фосфорсодержащих связей, включают, не ограничиваясь перечисленными, патенты США 3687808; 4469863; 4476301;5023243; 5177196; 5188897; 5264423; 5276019; 5278302; 5286717; 5321131; 5399676; 5405939; 5453496; 5455233; 5466677; 5476925; 5519126; 5536821; 5541306; 5550111; 5563253; 5571799; 5587361; и 5625050, каждый из которых включен в настоящую заявку посредством ссылки.
[00179] Предпочтительные модифицированные олигонуклеотидные остовы, не включающие атомы фосфора, образованы алкильными с короткой цепью или циклоалкильными межнуклеозидными связями, смешанными гетероатомными и алкильными или циклоалкильными межнуклеозидными связями, или одной или более гетероатомными с короткой цепью или гетероциклическими межнуклеозидными связями. Они включают остовы с морфолиновыми связями (образованными частично сахаром нуклеозида); силоксановые остовы; сульфидные, сульфоксидные и сульфоновые остовы; формацетильные и тиоформацетильные остовы; метиленформацетильные и тиоформацетильные остовы; алкенсодержащие остовы; сульфаматные остовы; метилениминовые и метиленгидразиновые остовы; сульфонатные и сульфонамидные остовы; амидные остовы; и другие, содержащие смешанные N, О, S и СН2-компоненты.
[00180] Примеры патентов США, содержащие указание по получению описанных выше олигонуклеотидов, включают, не ограничиваясь перечисленными, патенты США 5034506; 5166315; 5185444; 5214134; 5216141; 5235033; 5264562; 5264564; 5405938; 5434257; 5466677; 5470967; 5489677; 5541307; 5561225; 5596086; 5602240; 5610289; 5602240; 5608046; 5610289; 5618704; 5623070; 5663312; 5633360; 5677437; и 5677439, каждый из которых включен в настоящую заявку посредством ссылки.
[00181] В других предпочтительных миметиках олигонуклеотидов и сахара, и межнуклеозидная связь, т.е. остов, нуклеотидных единиц, заменены новыми группами. Основания сохраняют для гибридизации с подходящим целевым соединением нуклеиновой кислоты. Одно из таких олигомерных соединений, олигонуклеотидный миметик, продемонстрировавший прекрасную способность к гибридизации, называют пептидной нуклеиновой кислотой (ПНК). В ПНК-соединениях сахарный остов олигонуклеотида заменен амидсодержащим остовом, например, аминоэтилглициновым остовом. Основания нуклеиновых кислот сохранены и связаны прямо или непрямо с азотными атомами азагрупп амидной части остова. Примеры патентов США, в которых описано получение ПНК-соединений, включают, не ограничиваясь перечисленными, патенты США 5539082; 5714331; и 5719262, каждый из которых включен в настоящую заявку посредством ссылки. Дальнейшее описание ПНК-соединений можно найти у Nielsen, et al. (1991) Science 254, 1497-1500.
[00182] Согласно варианту реализации изобретения олигонуклеотиды с фосфоротиоатными остовами и олигонуклеозиды с гетероатомными остовами, и в частности -CH2-NH-O-CH2-, -CH2-N (СН3)-O-СН2-, известным как метилен (метилимино) или MMI остов, -CH2-O-N(СН3)-СН2-, -CH2N(CH3)-N(CH3) СН2- и -O-N(CH3)-CH2-CH2-, где нативный фосфодиэфирный остов представлен как -O-Р-O-СН2- согласно упомянутому выше патенту США 5489677, и амидные остовы согласно упомянутому выше патенту США 5602240. Также предпочтительными являются олигонуклеотиды, содержащие морфолиновые остовы согласно вышеупомянутому патенту США 5034506.
[00183] Модифицированные олигонуклеотиды могут также содержать один или более фрагмент замещенного сахара. Предпочтительные олигонуклеотиды содержат один из следующих заместителей по положению 2': ОН; F; O-, S-, или N-алкил; O-, S-, или N-алкенил; O-, S- или N-алкинил; или О алкил-О-алкил, где указанные алкил, алкенил и алкинил могут представлять собой замещенный или незамещенный С→СО алкил или С2→СО алкенил и алкинил. В частности, предпочтительными являются O(СН2)n OmCH3, O(CH2)n, OCH3, O(CH2)nNH2, O(CH2)nCH3, O(CH2)nONH2 и O(CH2)n ON(CH2nCH3)2, где n и m могут принимать значения от 1 до приблизительно 10. Другие предпочтительные олигонуклеотиды содержат один из следующих заместителей по положению 2': С→СО, (низшие алкилы, замещенные низшие алкилы, алкарил, аралкил, O-алкарил или O-аралкил, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2, гетероциклоалкил, гетероциклоалкарил, аминоалкиламино, полиалкиламино, замещенные силилы, РНК расщепляющую группу, репортерную группу, интеркалятор, группу, улучшающую фармакокинетические свойства олигонуклеотида, или группу, улучшающую фармакодинамические свойства олигонуклеотида, и другие заместители, обладающие сходными свойствами. Предпочтительная модификация включает 2'-метоксиэтокси (2'-O-CH2CH2OCH3, также известную как 2'-O-(2-метоксиэтил) или 2'-МОЭ), т.е. алкоксиалкокси группу. Еще одна предпочтительная модификация включает 2'-диметиламинооксиэтокси, т.е., O(CH2)2ON(CH3)2 группу, также известную как 2'-ДМАОЭ, согласно описанию в приведенных ниже в настоящей заявке примерах, и 2'- диметиламиноэтоксиэтокси (также известной в данной области техники как 2'-O-диметиламиноэтоксиэтил или 2'-ДМАЭОЭ), т.е., 2'-O-CH2-O-CH2-N(CH2)2.
[00184] Другие предпочтительные модификации включают 2'-метокси (2'-O СН3), 2'-аминопропокси (2'-O CH2CH2CH2NH2) и 2'-фтор (2'-F). Сходные модификации могут быть также осуществлены в других положениях олигонуклеотида, в частности, по 3' положению сахара 3' концевого нуклеотида или в 2'-5'связанных олигонуклеотидах и 5' положению 5' концевого нуклеотида. Олигонуклеотиды могут также содержать миметики сахаров, например, циклобутильные фрагменты вместо пентофуранозильного сахара. Примеры патентов США, в которых описано получение таких модифицированных сахарных структур, включают, не ограничиваясь перечисленными, патенты США 4981957; 5118800; 5319080; 5359044; 5393878; 5446137; 5466786; 5514785; 5519134; 5567811; 5576427; 5591722; 5597909; 5610300; 5627053; 5639873; 5646265; 5658873; 5670633; и 5700920, каждый из которых включен в настоящую заявку посредством ссылки.
[00185] Олигонуклеотиды могут также содержать модифицированные или замещенные нуклеиновые основания (часто называемое в данной области техники просто «основания»). В контексте настоящей заявки «немодифицированные» или «природные» нуклеотиды включают пуриновые основания аденин (А) и гуанин (G), и пиримидиновые основания тимин (Т), цитозин (С) и урацил (U). Модифицированные нуклеотиды включают другие синтетические и природные нуклеотиды, такие как 5-метилцитозин (5-те-С), 5-гидроксимегил цитозин, ксантин, гипоксантин, 2-аминоаденин, 6-метил и другие алкильные производные аденина и гуанина, 2-пропил и другие алкильные производные аденина и гуанина, 2-тиоурацил, 2-тиотимин и 2-тиоцитозин, 5-галогенурацил и цитозин, 5-пропинил урацил и цитозин, 6-азо урацил, цитозин и тимин, 5-урацил (псевдо-урацил), 4-тиоурацил, 8-галоген, 8-амино, 8-тиол, 8-тиоалкил, 8-гидроксил и другие 8-замещенные аденины и гуанины, 5-галоген, в частности, 5-бром, 5-трифторметил и другие 5-замещенные урацилы и цитозины, 7-метилгуанин и 7-метиладенин, 8-азагуанин и 8-азааденин, 7-деазагуанин и 7-деазааденин и 3-деазагуанин и 3-деазааденин.
[00186] Далее, нуклеотиды включают описанные в патенте США 3687808, описанные в 'The Concise Encyclopedia of Polymer Science And Engineering', стр.858-859, Kroschwitz, J.I., ed. John Wiley & Sons, 1990, описанные у Englisch et al., 'Angewandle Chemie, International Edition', 1991, 30, стр.613, и описанные у Sanghvi, Y.S., Chapter 15, 'Antisense Research and Applications', стр.289-302, Crooke, S.T. и Lebleu, B. ea., CRC Press, 1993. Некоторые из этих нуклеотидов подходят, в частности, для увеличения связывающей способности олигомерных соединений, предложенных в настоящем изобретении. Такие включают 5-замещенные пиримидины, 6- азапиримидины и N-2, N-6 и 0-6 замещенные пурины, в том числе 2-аминопропиладенин, 5-пропинилурацил и 5-пропинилцитозин. 5-метилцитозиновые замещения, как было показано, повышают стабильность дуплексов нуклеиновых кислот на 0,6-1,2°С (Sanghvi, Y.S., Crooke, S.T., Lebleu, В., eds, 'Antisense Research and Applications', CRC Press, Boca Raton, 1993, pp.276-278) и на настоящий момент являются предпочтительными замещениями оснований, в частности, в комбинации с 2'-O-метоксиэтиловыми модификациями сахаров.
[00187] Примеры патентов США, в которых описано получение вышеупомянутых модифицированных нуклеотидов, а также других модифицированных нуклеотидов, включают, не ограничиваясь перечисленными, патенты США 3687808, а также 4845205; 5130302; 5134066; 5175273; 5367066; 5432272; 5457187; 5459255; 5484908; 5502177; 5525711; 5552540; 5587469; 5596091; 5614617; 5750692 и 5681941, каждый из которых включен в настоящую заявку посредством ссылки.
[00188] Другая модификация олигонуклеотидов, предложенных в настоящем изобретении, подразумевает химическое связывание с олигонуклеотидом одного или более фрагмента или конъюгата, стимулирующего активность, распределение в клетках или поглощение клетками указанного олигонуклеотида.
[00189] Такие фрагменты содержат, не ограничиваясь перечисленными, липидные фрагменты, такие как фрагменты холестерина, холевая кислота, тиоэфир, например, гексил-S-тритилтиол, тиохолестерин, алифатические цепи, например, додекандиольные или ундецильные остатки, фосфолипид, например, ди-гексадецил-rac-глицерин или триэтиламмоний 1,2-ди-О-гексадецил-rac-глицеро-3-Н-фосфонат, цепь полиамина или полиэтиленгликоля, или адамантан-уксусную кислоту, пальмитиловый фрагмент, или октадециламиновый или гексиламино-карбонил-t оксихолестериновый фрагмент.
[00190] Примеры патентов США, в которых описано получение таких олигонуклеотидных конъюгатов, включают, не ограничиваясь перечисленными, патенты США 4828979; 4948882; 5218105; 5525465; 5541313; 5545730; 5552 538; 5578717, 5580731; 5580731; 5591584; 5109124; 5118802; 5138045; 5414077; 5486 603; 5512439; 5578718; 5608046; 4587044; 4605735; 4667025; 4762 779; 4789737; 4824941; 4835263; 4876335; 4904582; 4958013; 5082 830; 5112963; 5214136; 5082830; 5112963; 5214136; 5 245022; 5254469; 5258506; 5262536; 5272250; 5292873; 5317098; 5371241, 5391723; 5416203, 5451463; 5510475; 5512667; 5514785; 5 565552; 5567810; 5574142; 5585481; 5587371; 5595726; 5597696; 5599923; 5599 928 и 5688941, каждый из которых включен в настоящую заявку посредством ссылки.
[00191] Поиск новых лекарственных средств: Соединения, предложенные в настоящем изобретении, могут также применяться для поиска новых лекарств и валидации мишеней. Настоящее изобретение включает применение описанных в настоящей заявке соединений и предпочтительных целевых сегментов при поиске новых лекарственных средств для выяснения взаимосвязей, существующих между полинуклеотидами гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и болезнью, фенотипом или состоянием. Такие способы включают определение или модуляцию полинуклеотидов SCNA, включая контактирование образца, ткани, клетки или организма с соединениями, предложенными в настоящем изобретении, измерение уровня нуклеиновой кислоты или белка полинуклеотидов SCNA и/или оценка соответствующего фенотипического или химического конечного состояния через некоторое время после лечения, и, в некоторых случаях, сравнение измеряемой величины с необработанными образцами или образцами, обработанными другими предложенными в настоящем изобретении соединениями. Эти способы могут применяться параллельно или в комбинации с другими экспериментами по определению функций неизвестных генов в процессе валидации мишеней или для определения обоснованности применения конкретного генного продукта в качестве мишени для лечения или предотвращения конкретного заболевания, состояния или фенотипа.
Оценка повышающего регулирования или ингибирования экспрессии генов:
[00192] Перенос экзогенной нуклеиновой кислоты в клетку-хозяина или в организм-хозяина может быть оценен прямым определением присутствия указанной нуклеиновой кислоты в указанных клетке или организме. Такое определение может быть проведено несколькими известными специалистам в данной области техники способами. Например, присутствие экзогенной нуклеиновой кислоты может быть определено посредством саузерн-блоттинга или полимеразной цепной реакции (ПЦР) с применением праймеров, специфически амплифицирующих нуклеотидные последовательности, связанные с указанной нуклеиновой кислотой. Экспрессия указанных экзогенных нуклеиновых кислот также может быть измерена с применением общепринятых способов, в том числе анализа генной экспрессии. Например, мРНК, полученная из экзогенной нуклеиновой кислоты, может быть обнаружена и количественно определена с применением нозерн-блоттинга и ПЦР с обратной транскрипцией (ОТ-ПЦР).
[00193] Экспрессия РНК с указанной экзогенной нуклеиновой кислоты может также быть определена измерением ферментативной активности или активности репортерного белка. Например, антисмысловая модулирующая активность может быть непрямо измерена через понижение или повышение экспрессии целевой нуклеиновой кислоты в качестве показателя того, что экзогенная нуклеиновая кислота производит эффекторную РНК. Используя консервативность последовательностей, возможно сконструировать праймеры и использовать для амплификации кодирующих участков указанных генов-мишеней. Сначала может быть использована наиболее активно экспрессируемая кодирующая область каждого гена для построения модели контрольного гена, хотя может быть использована любая кодирующая или некодирующая область. Каждый контрольный ген получен встраиванием каждой кодирующей области между кодирующей репортер областью и ее поли(А)-сигналом. Эти плазмиды производят мРНК с репортерным геном в вышележащей части гена и потенциальной мишенью РНКи в 3' некодирующей области. Эффективность индивидуальных антисмысловых олигонуклеотидов может быть проанализирована по модулированию репортерного гена. Репортерные гены, подходящие для применения в способах, предложенных в настоящем изобретении, включают гены синтазы ацетогидроксикислот (АГКС), щелочной фосфатазы (ЩФ), бетагалактозидазы (LacZ), бета глюкоронидазы (GUS), хлорамфеникол ацетилтрансферазы (ХАТ), зеленого флуоресцентного белка (GFP), красного флуоресцентного белка (RFP), желтого флуоресцентного белка (YFP), голубого флуоресцентного белка (CFP), пероксидазы хрена (ПОХ), люциферазы (Luc), нопалин синтазы (NOS), октопин синтазы (OCS) и их производных. Доступны различные селективные маркеры, придающие устойчивость к ампициллину, блеомицину, хлорамфениколу, гентамицину, гигромицину, канамицину, линкомицину, метотрексату, фосфинотрицину, пуромицину и тетрациклину Способы определения модулирования репортерного гена хорошо известны в данной области техники, и включают, не ограничиваясь перечисленными: флуорометрические способы (например, флуоресцентная спектроскопия, сортировка флуоресцентно-активированных клеток (FACS), флуоресцентная микроскопия), определение устойчивости к антибиотику.
[00194] Сегменты нуклеиновой кислоты-мишени могут также быть обнаружены посредством анализов, основанных на применении клеток. Проводят эксперименты с целью обнаружения природной антисмысловой последовательности BG724147 для гена Scnia в клетках HepG2, в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве, и также в семенниках человека. В случае клеток HepG2, а также первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве, клетки выращивают, и РНК экстрагируют. В случае семенников человека приобретают и применяют выделенную поли(А)-РНК. Этот эксперимент называют RACE (быстрая амплификация концевых фрагментов кДНК), и применяют специфические праймеры для РНК-транскрипта BG724147.
[00195] Был обнаружен очень похожий ПЦР-продукт в выделенной поли(А)-РНК из клеток HepG2 и выделенной поли(А)-РНК из первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве, но этот продукт не был обнаружен в выделенной поли(А)-РНК из семенников человека. Кроме того, указанный ПЦР-продукт не был обнаружен (или был обнаружен в крайне малых количествах) в тотальной РНК из клеток HepG2 и тотальной РНК из первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве. Результаты позволяют предположить, что природная антисмысловая последовательность для гена Scnia, названная BG724147, присутствует в клетках HepG2 и первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве, но не в семенниках человека.
[00196] Экспрессию белка и мРНК SCNA можно проанализировать с применением известных специалистам в данной области техники способов, описанных в любом из разделов настоящей заявки. Например, для измерения уровней белка могут применяться методы иммуноанализа, такие как ELISA. Наборы ELISA для анализа SCNA коммерчески доступны, например, от R&D Systems (Minneapolis, MN).
[00197] Согласно вариантам реализации экспрессия SCNA (например, мРНК или белка) в образце (например, в клетках или тканях in vivo или in vitro), обработанном с применением антисмыслового олигонуклеотида, предложенного в настоящем изобретении, оценивается посредством сравнения с экспрессией SCNA в контрольном образце. Например, экспрессия белка или нуклеиновой кислоты может сравниваться с применением известных специалистам в данной области техники способов с таковой в плацебо-образце или в необработанном образце. Как вариант, может быть проведено сравнение с образцом, обработанным контрольным антисмысловым олигонуклеотидом (например, имеющим измененную или иную последовательность), в зависимости от того, какую информацию необходимо получить. Согласно другому варианту реализации различие в экспрессии белка или нуклеиновой кислоты SCNA между необработанным и обработанным образцами может сравниваться с различием в экспрессии различных нуклеиновых кислот (включая любые стандартные, признанные исследователем подходящими, например, конститутивный ген) в обработанном и необработанном образцах.
[00198] Наблюдаемые различия могут быть выражены в удобной форме, например, в виде пропорции или доли, для сравнения с контролем. Согласно вариантам реализации уровень мРНК или белка SCNA в образце, обработанном антисмысловым олигонуклеотидом, предложенным в настоящем изобретении, повышается или снижается приблизительно 1,25-кратно-10-кратно или более относительно необработанного образца или образца, обработанного контрольной нуклеиновой кислотой. Согласно вариантам реализации уровень мРНК или белка SCNA повышается или снижается по меньшей мере приблизительно 1,25-кратно, по меньшей мере приблизительно 1,3-кратно, по меньшей мере приблизительно 1,4-кратно, по меньшей мере приблизительно 1,5-кратно, по меньшей мере приблизительно 1,6-кратно, по меньшей мере приблизительно 1,7-кратно, по меньшей мере приблизительно 1,8-кратно, по меньшей мере приблизительно 2-кратно, по меньшей мере приблизительно 2,5-кратно, по меньшей мере приблизительно 3-кратно, по меньшей мере приблизительно 3,5-кратно, по меньшей мере приблизительно 4-кратно, по меньшей мере приблизительно 4,5-кратно, по меньшей мере приблизительно 5-кратно, по меньшей мере приблизительно 5,5-кратно, по меньшей мере приблизительно 6-кратно, по меньшей мере приблизительно 6,5-кратно, по меньшей мере приблизительно 7-кратно, по меньшей мере приблизительно 7,5-кратно, по меньшей мере приблизительно 8-кратно, по меньшей мере приблизительно 8,5-кратно, по меньшей мере приблизительно 9-кратно, по меньшей мере приблизительно 9,5-кратно, или по меньшей мере приблизительно 10-кратно или более.
Наборы, реагенты для исследования, диагностика и терапия
[00199] Соединения, предложенные в настоящем изобретении, можно применять для диагностики, лечения и профилактики, а также в качестве реагентов для исследований и компонентов наборов. Кроме того, антисмысловые олигонуклеотиды, способные ингибировать генную экспрессию с высокой специфичностью, часто используются специалистами в данной области техники для выяснения функций отдельных генов или распознавания функций различных компонентов биологического пути.
[00200] Для применения в наборах и для диагностики, и в различных биологических системах, соединения, предложенные в настоящем изобретении, по отдельности или в комбинации с другими соединениями или терапевтическими средствами подходят в качестве инструмента дифференциального и/или комбинаторного анализа для определения паттернов экспрессии части или всего набора генов, экспрессируемых в клетках и тканях.
[00201] Используемый в настоящей заявке термин «биологическая система» или «система» определен как любой организм, клетка, клеточная культура или ткань, экспрессирующая, или с приобретенной способностью экспрессировать, продукты генов альфа-субъединицы потенциалзависимого натриевого канала (SCNA). Биологические системы включают, но не ограничиваются перечисленными, человека, трансгенных животных, клетки, культуры клеток, ткани, ксенотрансплантаты, трансплантаты и их комбинации.
[00202] Согласно одному из неограничивающих примеров паттерны экспрессии в клетках или тканях, обработанных одним или несколькими антисмысловыми соединениями, сравнивают с контрольными клетками или тканями, не обработанными антисмысловыми соединениями, и исходя из полученных паттернов анализируют дифференциальные уровни генной экспрессии, так как они связаны, например, с болезнью, сигнальными путями, локализацией в клетках, уровнями экспрессии, размером, структурой или функцией изучаемых генов. Эти исследования могут быть выполнены на стимулированных или не стимулированных клетках в присутствии или в отсутствие других соединений, которые влияют на паттерны экспрессии.
[00203] Примеры способов анализа генной экспрессии, известных в данной области техники, включают анализ на ДНК-чипах или на микрочипах, SAGE (серийный анализ экспрессии генов), READS (рестриктазная амплификация расщепляемых кДНК), TOGA (полный анализ генной экспрессии), белковые чипы и протеомику, секвенирование меток экспрессируемой последовательности (EST), вычитающий РНК фингерпринтинг (SuRF), вычитающее клонирование, дифференциальный дисплей (DD), сравнительная геномная гибридизация, FISH (флуоресцентная гибридизация in situ) и масс-спектрометрические методы.
[00204] Соединения, предложенные в настоящем изобретении, подходят для исследований и диагностики, поскольку такие соединения гибридизуются с нуклеиновыми кислотами, кодирующими альфа-субъединицу потенциалзависимого натриевого канала (SCNA). Например, олигонуклеотиды, гибридизующиеся с эффективностью и в условиях, описанных в настоящей заявке, являясь эффективными модуляторами SCNA, представляют собой эффективные праймеры или зонды в условиях, благоприятных для амплификации или определения гена, соответственно. Такие праймеры и зонды подходят для способов, требующих специфического определения молекул нуклеиновой кислоты, кодирующих SCNA, и для амплификации указанных молекул нуклеиновой кислоты для определения или для применения в дальнейших исследованиях SCNA. Гибридизация указанных антисмысловых олигонуклеотидов, в частности, праймеров и зондов, предложенных в настоящем изобретении, с нуклеиновой кислотой, кодирующей SCNA, может быть определена при помощи известных в данной области техники способов. Такие способы могут включать конъюгирование фермента с указанным олигонуклеотидом, радиоактивное мечение указанного олигонуклеотида и любые другие подходящие способы детекции. Могут также быть подготовлены наборы для определения уровней SCNA в образце при помощи таких способов определения.
[00205] Специфичность и чувствительность антисмысловых соединений также используются специалистами в данной области техники для терапевтических целей. Антисмысловые соединения применялись в качестве терапевтических компонентов при лечении болезненных состояний у животных, в том числе человека. Лекарственные средства на основе антисмысловых олигонуклеотидов безопасно и эффективно вводили людям, и в настоящее время проводятся многочисленные клинические испытания. Таким образом, установлено, что антисмысловые соединения могут применяться в качестве терапевтических средств, которые могут быть модифицированы под терапевтические схемы для лечения клеток, тканей и животных, в особенности человека.
[00206] В случае терапевтических средств животное, предпочтительно человек, у которого, предположительно, имеется заболевание или расстройство, лечение которого может проводиться модулированием экспрессии полинуклеотидов SCNA, получает лечение в виде введения антисмысловых соединений в соответствии с настоящим изобретением. Например, согласно одному из неограничивающих вариантов реализации указанные способы включают этап введения животному, нуждающемуся в лечении, терапевтически эффективного количества модулятора SCNA. Модуляторы SCNA, предложенные в настоящем изобретении, эффективно модулируют активность SCNA или модулируют экспрессию белка SCNA. Согласно одному из вариантов реализации указанные активность или экспрессия SCNA у животного подавлены приблизительно на 10% по сравнению с контролем. Предпочтительно, указанные активность или экспрессия SCNA у животного подавлены приблизительно на 30%. Более предпочтительно, указанные активность или экспрессия SCNA у животного подавлены приблизительно на 50% или более. Таким образом, указанные олигомерные соединения модулируют экспрессию мРНК альфа-субъединицы потенциалзависимого натриевого канала (SCNA) по меньшей мере на 10%, по меньшей мере на 50%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98%, по меньшей мере на 99%, или на 100% по сравнению с контролем.
[00207] Согласно одному из вариантов реализации указанные активность и/или экспрессия гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) у животного повышены приблизительно на 10% по сравнению с контролем. Предпочтительно, указанные активность или экспрессия SCNA у животного повышены приблизительно на 30%. Более предпочтительно, указанные активность или экспрессия SCNA у животного повышены на 50% или более. Таким образом, указанные олигомерные соединения изменяют экспрессию мРНК SCNA по меньшей мере на 10%, по меньшей мере на 50%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98%, по меньшей мере на 99%, или на 100% относительно контроля.
[00208] Например, снижение экспрессии альфа-субъединицы потенциалзависимого натриевого канала (SCNA) может быть определено в сыворотке, крови, жировой ткани, печени или любой другой жидкости организма, ткани или органе указанного животного. Предпочтительно, клетки, содержащиеся в указанных анализируемых жидкостях, тканях или органах, содержат молекулу нуклеиновой кислоты, кодирующую пептиды SCNA и/или собственно белок SCNA.
[00209] Соединения, предложенные в настоящем изобретении, можно применять для получения фармацевтических композиций посредством добавления эффективного количества соединения к подходящему фармацевтически приемлемому разбавителю или носителю. Применение соединений и способов, предложенных в настоящем изобретении, может также подходить для профилактики.
Конъюгаты
[00210] Другая модификация олигонуклеотидов, предложенных в настоящем изобретении, включает химическое связывание с указанным олигонуклеотидом одного или нескольких фрагментов или конъюгатов, которые повышают активность, усиливают клеточное распределение или поглощение клетками указанного олигонуклеотида. Такие фрагменты или конъюгаты могут включать конъюгированные группы, ковалентно связанные с функциональными группами, такими как первичные или вторичные гидроксильные группы. Конъюгированные группы, предложенные в настоящем изобретении, включают интеркаляторы, репортерные молекулы, полиамины, полиамиды, полиэтиленгликоли, простые полиэфиры, группы, улучшающие фармакодинамические свойства олигомеров, и группы, улучшающие фармакокинетические свойства олигомеров. Типовые конъюгированные группы включают холестерины, липиды, фосфолипиды, биотин, феназин, фолат, фенантридин, антрахинон, акридин, флуоресцеины, родамины, кумарины и красители. Группы, усиливающие фармакодинамические свойства, в контексте настоящего изобретения включают группы, улучшающие поглощение, повышающие сопротивление разрушению и/или усиливающие сиквенс-специфичную гибридизацию с целевой нуклеиновой кислотой. Группы, улучшающие фармакокинетические свойства, в контексте настоящего изобретения включают группы, улучшающие поглощение, распределение, метаболизм или выведение соединений, предложенных в настоящем изобретении. Типовые конъюгированные группы описаны в международной заявке на патент PCT/US92/09196, поданной 23 октября 1992, и патенте США 6287860, включенных в настоящую заявку посредством ссылки. Конъюгированные фрагменты включают, не ограничиваясь перечисленными: липидные фрагменты, такие как фрагменты холестерина, холевую кислоту, тиоэфир, например, гексил-5-тритилтиол, тиохолестерин, алифатические цепи, например, додекандиольные или ундецильные остатки, фосфолипид, например, ди-гексадецил-rac-глицерин или триэтиламмоний 1,2-ди-О-гексадецил-rac-глицеро-3-Нфосфонат, цепь полиамина или полиэтиленгликоля, или адамантан-уксусную кислоту, пальмитиловый фрагмент, октадециламин или фрагмент гексиламино-карбонил-оксихолестерина. Олигонуклеотиды, предложенные в настоящем изобретении, могут быть также конъюгированы с активными лекарственными веществами, например, аспирином, варфарином, фенилбутазоном, ибупрофеном, супрофеном, фенбуфеном, кетопрофеном, (S)-(+)-пранопрофеном, карпрофеном, дансилсаркозином, 2,3,5-трийодбензойной кислотой, флуфенамовой кислотой, фолиновой кислотой, бензотиадиазидом, хлортиазидом, диазепином, индометицином, барбитуратом, цефалоспорином, сульфамидным препаратом, антидиабетическим средством, антибактериальным веществом или антибиотиком.
[00211] Примеры патентов США, в которых описано получение таких олигонуклеотидных конъюгатов, включают, не ограничиваясь перечисленными: патенты США 4828979; 4948882; 5218105; 5525465; 5541313; 5545730; 5552538; 5578717, 5580731; 5580731; 5591584; 5109124; 5118802; 5138045; 5414077; 5486603; 5512439; 5578718; 5608046; 4587044; 4605735; 4667025; 4762779; 4789737; 4824941; 4835263; 4876335; 4904582; 4958013; 5082830; 5112963; 5214136; 5082830; 5112963; 5214136; 5245022; 5254469; 5258506; 5262536; 5272250; 5292873; 5317098; 5371241, 5391723; 5416203, 5451463; 5510475; 5512667; 5514785; 5565552; 5567810; 5574142; 5585481; 5587371; 5595726; 5597696; 5599923; 5599928 и 5688941.
Составы
[00212] Соединения, предложенные в настоящем изобретении, также могут быть смешаны, инкапсулированы, конъюгированы или иным образом связаны с другими молекулами, молекулярными структурами или смесями соединений, например, липосомами, молекулами-мишенями рецепторов, составами для перорального, ректального, местного или иного пути введения, для способствования поглощению, распределению и/или абсорбции. Примеры патентов США, в которых описано получение таких способствующих поглощению, распределению и/или абсорбции составов, включают, не ограничиваясь перечисленными: патенты США 5108921; 5354844; 5416016; 5459127; 5521291; 5543165; 5547932; 5583020; 5591721; 4426330; 4534899; 5013556; 5108921; 5213804; 5227170; 5264221; 5356633; 5395619; 5416016; 5417978; 5462854; 5469854; 5512295; 5527528; 5534259; 5543152; 5556948; 5580575; и 5595756, каждый из которых включен в настоящую заявку посредством ссылки.
[00213] Хотя указанные антисмысловые олигонуклеотиды не обязательно должны вводиться при помощи вектора, чтобы модулировать экспрессию и/или функцию мишени, варианты реализации настоящего изобретения включают векторные конструкции для экспрессии антисмысловых олигонуклеотидов, включая промоторы, последовательности генов составных промоторов и обладающие выраженной конститутивной промоторной активностью, или промоторной активностью, которая может быть индуцирована при необходимости.
[00214] Согласно одному из вариантов реализации осуществление изобретения включает введение по меньшей мере одного из вышеуказанных антисмысловых олигонуклеотидов при помощи подходящей системы доставки на основе нуклеиновой кислоты. Согласно одному из вариантов реализации такая система включает невирусный вектор, функционально связанный с указанным полинуклеотидом. Примеры таких невирусных векторов включают собственно олигонуклеотид (например, любая(ые) из последовательностей SEQ ID NOS: 29-94) или его комбинацию с подходящим белковым, полисахаридным или липидным составом.
[00215] Дополнительно, подходящие системы доставки нуклеиновой кислоты включают вирусные векторы, как правило, на основе последовательности одного или нескольких аденовирусов, аденоассоциированного вируса (ААВ), хелпер-зависимого аденовируса, ретровируса, или липосомального комплекса с японским гемагглютинирующим вирусом (HVJ). Предпочтительно, указанный вирусный вектор содержит сильный эукариотический промотор, функционально связанный с указанным полинуклеотидом, например, промотор цитомегаловируса (ЦМВ).
[00216] Дополнительные предпочтительные векторы включают вирусные векторы, белки слияния и химические конъюгаты. Ретровирусные векторы включают вирусы мышиного лейкоза Молони и ВИЧ-вирусы. Один из предпочтительных ВИЧ-вирусных векторов содержит по меньшей мере два вектора, где гены gag и pol происходят из генома ВИЧ, а ген env - из другого вируса. Предпочтительными являются ДНК-вирусные векторы. Такие векторы включают рох-векторы, такие как ортопокс- или авипокс-векторы, герпесвирусные векторы, такие как векторы на основе вируса простого герпеса (HSV) I, аденовирусные векторы и векторы на основе аденоассоциированного вируса.
[00217] Антисмысловые соединения, предложенные в настоящем изобретении, включают любые фармацевтически приемлемые соли, сложные эфиры или соли таких сложных эфиров, или любое другое соединение, которое при введении животному, включая человека, способно обеспечить получение (прямо или непрямо) биологически активного метаболита или его компонента.
[00218] Термин «фармацевтически приемлемые соли» относится к физиологически и фармацевтически приемлемым солям соединений, предложенных в настоящем изобретении: т.е. солям, сохраняющим необходимую биологическую активность исходного соединения и, кроме того, не дающих нежелательных токсических эффектов. Предпочтительные примеры фармацевтически приемлемых солей олигонуклеотидов и их применение подробнее описаны в патенте США 6287860, включенном в настоящую заявку посредством ссылки.
[00219] Настоящее изобретение также включает фармацевтические композиции и составы, содержащие антисмысловые соединения, предложенные в настоящем изобретении. Фармацевтические композиции, предложенные в настоящем изобретении, могут вводиться различными способами в зависимости от того, требуется ли местное или системное лечение, и от того, на какую область необходимо воздействовать. Введение может быть местным (в том числе через глаза и через слизистые оболочки, включая вагинальное и ректальное введение), через легкие, например, посредством вдыхания или вдувания порошков или аэрозолей, в том числе при помощи небулайзера; интратрахеально, интраназально, эпидермально и трансдермально), пероральным или парентеральным. Парентеральное введение включает внутривенные, внутриартериальные, подкожные, внутрибрюшинные или внутримышечные инъекцию или инфузию; или внутричерепное, например, интратекальное или интравентрикулярное, введение.
[00220] При лечении тканей центральной нервной системы введение может осуществляться, например, инъекцией или инфузией в спинномозговую жидкость. Введение антисмысловой РНК в спинномозговую жидкость описано, например, в заявке на патент США, опубликованной под номером 2007/0117772, "Methods for slowing familial ALS disease progression» («Способы замедления развития наследственного заболевания БАС»), включенной в настоящую заявку посредством ссылки во всей полноте.
[00221] Если предполагается введение антисмыслового олигонуклеотида, предложенного в настоящем изобретении, в клетки центральной нервной системы, введение может осуществляться совместно с одним или несколькими агентами, способными обеспечить проникновение указанного антисмыслового олигонуклеотида через гематоэнцефалический барьер. Инъекция может быть сделана, например, в энторинальную область коры или гиппокамп. Доставка нейротрофических факторов введением аденовирусного вектора в двигательные нейроны мышечной ткани описана, например, в патенте США 6632427, "Adenoviral-vector-mediated gene transfer into medullary motor neurons» («Перенос генов в двигательные нейроны мозга, опосредованный аденовирусным вектором»), включенном в настоящую заявку посредством ссылки. Доставка векторов непосредственно в мозг, например, в стриатум, таламус, гиппокамп или черную субстанцию известна в данной области техники и описана, например, в патенте США 6756523, «Adenovirus vectors for the transfer of foreign genes into cells of the central nervous system particularly in brain» («Аденовирусные векторы для переноса чужеродных генов в клетки центральной нервной системы, в частности, клетки мозга»), включенном в настоящую заявку посредством ссылки. Введение может быть быстрым в случае инъекции или осуществляться в течение некоторого промежутка времени в случае медленной инфузии или введения составов с замедленным высвобождением.
[00222] Рассматриваемые антисмысловые олигонуклеотиды могут быть связаны или конъюгированы с агентами, обеспечивающими необходимые фармацевтические или фармакокинетические свойства. Например, указанный антисмысловой олигонуклеотид может сочетаться с любым веществом, которое, как известно в данной области техники, способствует проникновению или транспорту через гематоэнцефалический барьер, таким как антитело к рецептору трансферрина, и вводимому путем внутривенной инъекции. Указанное антисмысловое соединение может быть связано с вирусным вектором, например, придающим указанному антисмысловому соединению большую эффективность и/или усиливающим транспорт указанного антисмыслового соединения через гематоэнцефалический барьер. Осмотическое преодоление гематоэнцефалического барьера может также быть достигнуто, например, инфузией сахаров, включая, но не ограничиваясь перечисленными, мезоэритрит, ксилит, D(+) галактозу, D(+) лактозу, D(+) ксилозу, дульцит, миоинозитол, L(-) фруктозу, D(-) маннит, D(+) глюкозу, D(+) арабинозу, D(-) арабинозу, целлобиозу, D(+) мальтозу, D(+) раффинозу, L(+) рамнозу, D(+) мелибиозу, D(-) рибозу, адонит, D(+) арабит, L(-) арабит, D(+) фукозу, L(-) фукозу, D(-) ликсозу, L(+) ликсозу и L(-) ликсозу, или аминокислот, включая, но не ограничиваясь перечисленными, глутамин, лизин, аргинин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глицин, гистидин, лейцин, метионин, фенилаланин, пролин, серии, треонин, тирозин, валин и таурин. Способы и материалы для повышения проникновения через гематоэнцефалический барьер описаны, например, в патенте США 4866042, «Method for the delivery of genetic material across the blood brain barrier» («Способы доставки генетического материала через гематоэнцефалический барьер»), патенте США 6294520, «Material for passage through the blood-brain barrier» («Материал для прохождения через гематоэнцефалический барьер») и патенте США 6936589, «Parenteral delivery systems» («Парентеральные системы доставки»), включенных в настоящую заявку посредством ссылки во всей полноте.
[00223] Рассматриваемые антисмысловые соединения могут быть смешаны, инкапсулированы, конъюгированы или иным образом связаны с другими молекулами, молекулярными структурами или смесями соединений, например, липосомами, молекулами-мишенями рецепторов, составами для перорального, ректального, местного или иного пути введения, для способствования поглощению, распределению и/или абсорбции. Например, катионные липиды могут также быть включены в указанный состав для облегчения поглощения олигонуклеотидов. Одной из таких композиций, как показано, способствующей поглощению, является LIPOFECTIN (доступный у GIBCO-BRL, Bethesda, MD).
[00224] Олигонуклеотиды, содержащие по меньшей мере одну 2'-O-метоксиэтильную модификацию, предположительно подходят, в частности, для перорального введения. Фармацевтические композиции и составы для местного применения могут включать трансдермальные пластыри, мази, лосьоны, кремы, гели, капли, суппозитории, спреи, жидкости и порошки. Обычные фармакологические носители, водные, порошковые или масляные основы, загустители и т.п. могут быть необходимы или желательны. Подходящие применения включают также нанесение в качестве покрытия на презервативы, перчатки и т.п.
[00225] Фармацевтические составы, предложенные в настоящем изобретении, которые могут быть представлены в виде удобной единичной дозированной формы, могут быть получены согласно общепринятым методикам, хорошо известным в области фармацевтической индустрии. Такие техники включают этап соединения активных ингредиентов с фармацевтическими носителями и вспомогательными веществами. Как правило, указанные составы получают равномерным глубоким соединением активных ингредиентов с жидкими носителями или тонкомолотыми твердыми носителями, или с ними обоими, и затем, при необходимости, придают продукту форму.
[00226] Композиции, предложенные в настоящем изобретении, могут быть получены в виде любой из многих возможных лекарственных форм, включая, но не ограничиваясь перечисленными, таблетки, капсулы, гелевые капсулы, жидкие сиропы, мягкие гели, суппозитории и клизмы. Композиции, предложенные в настоящем изобретении, могут также быть получены в виде суспензий на основе водных, неводных или смешанных сред. Водные суспензии могут дополнительно содержать вещества, увеличивающие вязкость суспензии, включая, например, натрия карбоксиметилцеллюлозу, сорбит и/или декстран. Указанная суспензия может также содержать стабилизаторы.
[00227] Фармацевтические композиции, предложенные в настоящем изобретении, включают, не ограничиваясь перечисленными: растворы, эмульсии, пены и содержащие липосомы составы. Фармацевтические композиции и составы, предложенные в настоящем изобретении, могут содержать один или более усилители проникновения, носители, вспомогательные вещества или другие активные или неактивные ингредиенты.
[00228] Эмульсии представляют собой, как правило, гетерогенные системы, где одна жидкость распределена в другой в виде капель, как правило, более 0,1 мкм в диаметре. Эмульсии могут содержать дополнительные компоненты помимо дисперсных фаз, и активное лекарственное вещество может присутствовать в растворе в водной фазе, масляной фазе или быть представлено отдельной фазой. Микроэмульсии также относятся к вариантам реализации настоящего изобретения. Эмульсии и их применение хорошо известны в данной области техники и подробнее описаны в патенте США 6287860.
[00229] Составы, предложенные в настоящем изобретении, включают липосомальные составы. В контексте настоящего изобретения термин «липосома» означает пузырьки, состоящие из амфифильных липидов, образующих один или более чем один сферический бислой. Липосомы представляют собой однослойные или многослойные пузырьки с мембраной, образованной липофильными веществами и водной внутренней фазой, где содержится композиция, которую необходимо доставить. Катионные липосомы представляют собой положительно заряженные липосомы, которые предположительно взаимодействуют с отрицательно заряженными молекулами ДНК с образованием стабильного комплекса. рН-чувствительные или отрицательно заряженные липосомы, предположительно, скорее захватывают ДНК, чем образуют с ней комплексы. Как катионные, так и некатионные липосомы применялись для доставки ДНК в клетки.
[00230] Липосомы также включают «стерически стабилизированные» липосомы, что при использовании в настоящей заявке относится к липосомам, содержащим один или более специализированный липид. После включения в липосомы эти специализированные липиды образуют липосомы с увеличенной продолжительностью существования по сравнению с липосомами, не содержащими таких специализированных липидов. Примеры стерически стабилизированных липосом являются такие, в которых часть образующего пузырек липидного компонента липосомы содержит один или более гликолипид или дериватизирован одним или несколькими гидрофильными полимерами, таких как фрагмент полиэтиленгликоля (ПЭГ). Липосомы и их применение подробнее описаны в патенте США 6287860.
[00231] Фармацевтические составы и композиции, предложенные в настоящем изобретении, могут также включать поверхностно-активные вещества. Применение поверхностно-активных веществ в лекарственных продуктах, составах и эмульсиях хорошо известно в данной области техники. Поверхностно-активные вещества и их применение подробнее описаны в патенте США 6287860, включенном в настоящую заявку посредством ссылки.
[00232] Согласно одному из вариантов реализации настоящее изобретение включает различные усилители проникновения, способствующие эффективной доставке нуклеиновых кислот, в частности, олигонуклеотидов. Кроме способствования диффузии нелипофильных препаратов через клеточные мембраны, усилители проникновения повышают проникающую способность липофильных препаратов. Усилители проникновения могут быть разделены на пять больших категорий, а именно, поверхностно-активные вещества, жирные кислоты, соли желчных кислот, хелатирующие агенты, и нехелатирующие не-ПАВ-вещества. Усилители проникновения и их применение подробнее описаны в патенте США 6287860, включенном в настоящую заявку посредством ссылки.
[00233] Специалисту в данной области техники будет понятно, что составы получают согласно стандартным методикам в соответствии с их назначением, т.е. способом введения.
[00234] Предпочтительные составы для местного применения включают такие, в которых олигонуклеотиды, предложенные в настоящем изобретении, смешаны с агентами для местного введения, такими как липиды, липосомы, жирные кислоты, сложные эфиры жирных кислот, стероиды, хелатирующие агенты и поверхностно-активные вещества. Предпочтительные липиды и липосомы включают нейтральные (например, диолеилфосфатидил DOPE этаноламин, димиристоилфосфатидил холин DMPC, дистеаролфосфатидилхолин) отрицательно заряженные (например, димиристоилфосфатидил глицерина DMPG) и катионные (например, диолеилтетраметиламинопропил DOTAP и диолеилфосфатидил этаноламин DOTMA).
[00235] Для местного или другого применения олигонуклеотиды, предложенные в настоящем изобретении, могут быть инкапсулированы в липосомы или входить в состав липосомальных комплексов, в частности, с катионными липосомами. Как вариант, олигонуклеотиды могут входить в состав липидных комплексов, в частности с катионными липидами. Предпочтительные жирные кислоты и сложные эфиры, их фармацевтически приемлемые соли, и их применение подробнее описаны в патенте США 6287860.
[00236] Композиции и составы для перорального введения включают порошки или гранулы, содержащие микрочастицы, наночастицы, суспензии или растворы в воде или неводных средах, капсулы, гелевые капсулы, порционные упаковки, таблетки или минитаблетки. Присутствие загустителей, вкусоароматических агентов, разбавителей, эмульгаторов, диспергирующих агентов или связующих веществ может быть желательным. Предпочтительными составами для перорального приема являются такие, в которых олигонуклеотиды, предложенные в настоящем изобретении, вводятся совместно с одним или несколькими усилителями проникновения, поверхностно-активными веществами и хелатирующими веществами. Предпочтительные поверхностно-активные вещества включают жирные кислоты и/или сложные эфиры или их соли, жёлчные кислоты и/или их соли. Предпочтительные жёлчные кислоты/соли и жирные кислоты и их применение подробнее описаны в патенте США 6287860, включенном в настоящую заявку посредством ссылки. Также предпочтительными являются комбинации усилителей проникновения, например, жирные кислоты/соли в комбинации с жёлчными кислотами/солями. В частности, предпочтительной является комбинация натриевой соли лауриновой кислоты, каприновой кислоты и УДХК. Другие усилители проникновения включают полиоксиэтилен-9-лауриловый эфир, полиоксиэтилен-20-цетиловый эфир. Олигонуклеотиды, предложенные в настоящем изобретении, могут быть введены перорально, в гранулированной форме, включая высушенные распылением частицы, или в составе комплексов микро- или наночастиц. Комплексообразующие агенты для олигонуклеотидов и их применение подробнее описаны в патенте США 6287860, включенном в настоящую заявку посредством ссылки.
[00237] Композиции и составы для парентерального, интратекального или интравентрикулярного введения могут включать стерильные водные растворы, которые могут также содержать буферы, разбавители и другие подходящие добавки, включая, но не ограничиваясь перечисленными, усилители проникновения, переносчики и другие фармацевтически приемлемые носители или вспомогательные вещества.
[00238] Определенные варианты реализации настоящего изобретения включают фармацевтические композиции, содержащие одно или более олигомерное соединение и один или более другой химиотерапевтический агент, действующий с помощью не-антисмыслового механизма. Примеры таких химиотерапевтических агентов включают, не ограничиваясь перечисленными, химиотерапевтические лекарственные средства для лечения рака, такие как даунорубицин, дауномицин, дактиномицин, доксорубицин, эпирубицин, идарубицин, эзорубицин, блеомицин, мафосфамид, ифосфамид, цитозина арабинозид, бис-хлорэтил-нитрозомочевина, бусульфан, митомицин С, актиномицин D, митрамицин, преднизон, гидроксипрогестерон, тестостерон, тамоксифен, дакарбазин, прокарбазин, гексаметилмеламин, пентаметилмеламин, митоксантрон, амсакрин, хлорамбуцил, метилциклогексилнитрозомочевина, мустарген, мелфалан, циклофосфамид, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5- азацитидин, гидроксимочевина, дезоксикоформицин, 4-гидроксипероксицикло-фосфорамид, 5-фторурацил (5-FU), 5-фтордезоксиуридин (5-FUdR), метотрексат (МТХ), колхицин, таксол, винкристин, винбластин, этопозид (VP-16), триметрексат, иринотекан, топотекан, гемцитабин, тенипозид, цисплатин и диэтилстильбэстрол(DES). При применении с соединениями, предложенными в настоящем изобретении, такие химиотерапевтические агенты могут применяться индивидуально (например, 5-FU и олигонуклеотид), последовательно (например, 5-FU и олигонуклеотид в течение некоторого времени, затем МТХ и олигонуклеотид), или в комбинации с одним или несколькими другими такими химиотерапевтическими агентами (например, 5-FU, МТХ и олигонуклеотид, или 5-FU, лучевая терапия и олигонуклеотид). Противовоспалительные лекарственные средства, включая, но не ограничиваясь перечисленными, нестероидные противовоспалительные лекарственные средства и кортикостероиды, и противовирусные лекарственные средства, включая, но не ограничиваясь перечисленными, рибавирин, видарабин, ацикловир и ганцикловир, могут также быть скомбинированы в композициях, предложенных в настоящем изобретении. Комбинации антисмысловых соединений и других не-антисмысловых лекарственных средств также входят в объем настоящего изобретения. Два или более скомбинированных соединения могут применяться совместно или последовательно.
[00239] Согласно другому сходному варианту осуществления композиции, предложенные в настоящем изобретении, могут содержать одно или несколько антисмысловых соединений, в частности, олигонуклеотидов, нацеленных на первую нуклеиновую кислоту, и одно или несколько дополнительных антисмысловых соединений, нацеленных на вторую нуклеиновую кислоту. Например, первой мишенью может быть конкретная антисмысловая последовательность для гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA), а второй мишенью может быть участок другой нуклеотидной последовательности.
Как вариант, композиции, предложенных в настоящем изобретении, могут содержать два антисмысловых соединения или более, нацеленных на разные участки одной и той же целевой нуклеиновой кислоты альфа-субъединицы потенциалзависимого натриевого канала (SCNA). Многочисленные примеры антисмысловых соединений приведены в настоящей заявке, другие могут быть выбраны из подходящих соединений, известных в данной области техники. Два или более скомбинированных соединения могут применяться совместно или последовательно.
Дозирование:
[00240] Получение фармацевтических композиций и их последующее введение (дозирование) должны быть знакомы специалистам в данной области техники. Дозы зависят от тяжести и чувствительности к лечению болезненного состояния-мишени терапии, курс лечения может длиться от нескольких дней до нескольких месяцев, или до излечения, или до облегчения болезненного состояния. Оптимальный режим дозирования может быть рассчитан на основании измерений накопления препарата в организме пациента. Специалист без труда определит оптимальные дозировки, методологию дозировок и частоту повторения. Оптимальные дозы могут варьировать в зависимости от относительной эффективности конкретных олигонуклеотидов, и, как правило, могут быть рассчитаны исходя из ЕС50, эффективных для животных моделей in vitro и in vivo. Как правило, доза составляет от 0,01 мкг до 100 г на кг массы тела, и может вводиться однократно или чаще ежедневно, еженедельно, ежемесячно или ежегодно, или даже каждые 2-20 лет. Специалист без труда может рассчитать частоту введения доз, основанную на времени удержания и концентрациях лекарства в жидкостях или тканях организма. После проведения эффективного лечения может быть желательно применение поддерживающей терапии у пациента для предотвращения рецидива заболевания, при этом указанный олигонуклеотид вводится в поддерживающих дозах, варьирующих от 0,01 мкг до 100 г на кг массы тела, от однократного или более частого ежедневного приема до однократного приема один раз в 20 лет.
[00241] Согласно вариантам реализации пациент получает дозу лекарственного средства, составляющую по меньшей мере приблизительно 1, по меньшей мере приблизительно 2, по меньшей мере приблизительно 3, по меньшей мере приблизительно 4, по меньшей мере приблизительно 5, по меньшей мере приблизительно 6, по меньшей мере приблизительно 7, по меньшей мере приблизительно 8, по меньшей мере приблизительно 9, по меньшей мере приблизительно 10, по меньшей мере приблизительно 15, по меньшей мере приблизительно 20, по меньшей мере приблизительно 25, по меньшей мере приблизительно 30, по меньшей мере приблизительно 35, по меньшей мере приблизительно 40, по меньшей мере приблизительно 45, по меньшей мере приблизительно 50, по меньшей мере приблизительно 60, по меньшей мере приблизительно 70, по меньшей мере приблизительно 80, по меньшей мере приблизительно 90, или по меньшей мере приблизительно 100 мг/кг массы тела. Некоторые вводимые дозировки антисмысловых олигонуклеотидов описаны, например, в патенте США 7563884, «Antisense modulation of PTP1B expression» («Антисмысловое модулирование экспрессии PTP1B»), включенном в настоящую заявку посредством ссылки во всей полноте.
[00242] Хотя выше и были описаны различные варианты реализации настоящего изобретения, следует понимать, что они представлены только в качестве примеров, а не для ограничения. Многочисленные модификации раскрытых вариантов реализации могут быть осуществлены согласно описаниям в настоящей заявке, не выходя за рамки сущности и объёма настоящего изобретения. Таким образом, объем и сущность настоящего изобретения не ограничены какими-либо из приведенных выше вариантов реализации.
[00243] Все упоминаемые здесь документы включены в настоящую заявку посредством ссылок. Все публикации и патентные документы, упомянутые в настоящей заявке, включены посредством ссылок в том же объеме, как если бы каждая публикация или каждый патентный документ были упомянуты в индивидуальном порядке. Цитируя различные источники в данном документе, заявители не признают, что какой-либо из источников представляет «предшествующий уровень техники» относительно настоящего изобретения. Варианты реализации предложенных в изобретении композиций и способов проиллюстрированы приведенными ниже примерами.
ПРИМЕРЫ
[00244] Следующие неограничивающие Примеры предназначены для иллюстрации некоторых вариантов реализации настоящего изобретения. Следует иметь в виду, что вариации содержания и вариантов элементов описанных компонентов очевидны специалистам в данной области и входят в объем настоящего изобретения.
Пример 1: Конструирование антисмысловых олигонуклеотидов, специфичных к молекуле нуклеиновой кислоты, антисмысловой для гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и/или смысловой цепи полинуклеотида SCNA
[00245] Как указано выше, термин «олигонуклеотид, специфичный для» или «олигонуклеотид с нацеленным действием» относится к олигонуклеотиду, имеющему последовательность, (i) способную формировать стабильный комплекс с фрагментом гена-мишени или (ii) способную формировать стабильный дуплекс с фрагментом мРНК транскрипта гена-мишени.
[00246] Отбор подходящих олигонуклеотидов упрощается при применении компьютерных программ (например, IDT AntiSense Design, IDT OligoAnalyzer), автоматически идентифицирующих в каждой конкретной последовательности субпоследовательности из 19-25 нуклеотидов, способных образовывать гибриды с целевой полинуклеотидной последовательностью, имеющие требуемую температуру плавления (как правило, 50-60°С) и не образующие гомодимеров или других комплексных вторичных структур.
[00247] Отбор подходящих олигонуклеотидов выполняют с применением компьютерных программ, которые автоматически выравнивают последовательности нуклеиновых кислот и определяют участки идентичности или гомологии. Такие программы применяют для сравнения последовательностей нуклеиновых кислот, полученных, например, из баз данных, таких как GenBank, или секвенированием продуктов ПЦР. Сравнение последовательностей нуклеиновых кислот различных генов и межгенных областей конкретного генома позволяет отобрать последовательности нуклеиновых кислот, которым свойственна необходимая степень специфичности к нужному гену. Такие процедуры позволяют провести отбор олигонуклеотидов, проявляющих значительную степень комплементарности целевым последовательностям нуклеиновых кислот и более низкую степень комплементарности другим последовательностям нуклеиновых кислот из конкретного генома. Специалисту в данной области техники будет ясно, что существует значительная свобода выбора подходящих участков генов для применения согласно настоящему изобретению.
[00248] Антисмысловое соединение является «специфически гибридизуемым», если связывание указанного соединения с целевой нуклеиновой кислотой нарушает нормальное функционирование целевой нуклеиновой кислоты, приводя к модулированию функции и/или активности, и степень комплементарности достаточна, чтобы избежать неспецифичного связывания указанного антисмыслового соединения с нецелевыми последовательностями нуклеиновой кислоты в условиях, при которых необходимо специфическое связывание, т.е., в физиологических условиях в случае in vivo методик анализа или терапевтического воздействия, и в условиях, при которых проводится анализ, в случае исследований in vitro.
[00249] Характеристики гибридизации олигонуклеотидов, описанных в настоящей заявке, могут быть определены с применением одного или более in vitro способа анализа из известных специалистам в данной области техники. Например, указанные характеристики олигонуклеотидов, описанных в настоящей заявке, могут быть получены путем определения силы связывания целевых природных антисмысловых последовательностей и молекул потенциального лекарственного средства с применением анализа кривых плавления.
[00250] Сила связывания целевых природных антисмысловых последовательностей и молекулы потенциального лекарственного средства (далее - Молекулы) может быть оценена с применением любых стандартных способов измерения силы межмолекулярных взаимодействий, например, анализа кривых плавления.
[00251] Анализ кривых плавления определяет температуру, при которой происходит быстрый переход от двухцепочечной к одноцепочечной конформации комплекса природной антисмысловой последовательности/Молекулы. Эта температура широко используется как надежная мера силы взаимодействия между двумя молекулами.
[00252] Анализ кривых плавления может быть выполнен с применением кДНК-копии актуальной природной антисмысловой молекулы РНК или синтетического нуклеотида ДНК или РНК, соответствующего связывающему сайту Молекулы. Доступны многочисленные наборы, включающие все необходимые реагенты для проведения анализа (например, набор MeltDoctor от Applied Biosystems Inc.). Такие наборы содержат подходящий буферный раствор, содержащий один из связывающих двухцепочечную ДНК (дцДНК) красителей (таких как красители HRM от ABI, SYBR Green, SYTO и т.д.). Свойства дцДНК красителей таковы, что они практически не флуоресцируют в свободной форме, но очень выражение флуоресцируют при связывании с дцДНК.
[00253] Для проведения анализа указанную кДНК или соответствующий олигонуклеотид смешивают с Молекулой в концентрациях, определенных согласно протоколам конкретных производителей. Смесь подогревают до 95°С для диссоциации пре-формированных комплексов дцДНК, затем медленно охлаждают до комнатной температуры или другой более низкой температуры, указанной производителем набора, для ренатурации молекул ДНК. Вновь сформировавшиеся комплексы затем медленно нагревают до 95°С, одновременно непрерывно измеряя интенсивность флуоресценции в результате реакции. Интенсивность флуоресценции обратно пропорциональна количеству присутствующих в реакции дцДНК. Информация может быть получена с применением инструмента на основе ПЦР в реальном времени, совместимого с набором (например, StepOne Plus Real Time PCR System от ABI или инструмент lightTyper от Roche Diagnostics, Lewes, UK).
[00254] Пики плавления получают построением графика зависимости отрицательной производной флуоресценции по температуре (-d(Флуоресценция)/dT) по оси y) от температуры (ось х) с применением подходящего программного обеспечения (например, lightTyper (Roche) или SDS Dissociation Curve, ABI). Информацию анализируют с целью определить температуру быстрого перехода комплекса дцДНК в одноцепочечные молекулы. Эту температуру называют температурой плавления и она прямо пропорциональна силе взаимодействия между двумя молекулами. Как правило, температура плавления превышает 40°С.
Пример 2: Модулирование полинуклеотидов SCNA
Обработка клеток HepG2 антисмысловыми олигонуклеотидами
[00255] Клетки HepG2 от АТСС (cat# HB-8065) выращивали на ростовой среде (MEM/EBSS (Hyclone cat #SH30024 либо Mediatech cat # MT-10-OIO-CV) с добавлением 10 % эмбриональной бычьей сыворотки (ЭБС; FBS) (Mediatech cat#МТ35-011-CV) и пенициллина/стрептомицина (Mediatech cat#MT30-002-CI)) при 37°С и 5 % CO2. За один день до эксперимента клетки пересевали в концентрации 1,5×105/мл в 6-луночные планшеты и инкубировали при 37°С и 5 % СО2. В день эксперимента среду в 6-луночных планшетах заменяли на свежую ростовую среду. Все антисмысловые олигонуклеотиды разводили до концентрации 20 мкМ. Два мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл агента для трансфекции Липофектамина 2000 (Lipofectamine 2000; Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили в каждую лунку б-луночных планшетов с клетками HepG2. Сходную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Через 3-18 ч инкубации при 37°С и 5% СО2 среду заменяли на свежую ростовую среду. Через 48 ч после добавления антисмысловых олигонуклеотидов среду удаляли и выделяли РНК из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105) или набора RNeasy Total RNA Isolation kit от Qiagen (cat#74181), следуя инструкциям производителя. 600 нг выделенной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Verso cDNAkit от Thermo Scientific (cat#AB1453B) или набора High Capacity cDNA Reverse Transcription Kit (cat#4368813) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (Applied Biosystems Taqman Gene Expression Assay: Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCNA человека) от Applied Biosystems Inc., Foster City CA). Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems).
[00256] Кратность изменения генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Результаты: Результаты ПЦР в режиме реального времени показывают, что уровни мРНК SCN1A в клетках HepG2 достоверно повышаются через 48 ч после обработки антисмысловыми олигонуклеотидами, нацеленными на антисмысловую последовательность BG724147 для гена SCN1A (Фиг.1, 4). Другие олигонуклеотиды, нацеленные на антисмысловые последовательности BG724147 и Hs.662210 для гена SCN1A, не повышали уровни SCN1A (Фиг.2, 3).
Пример 3: Увеличение экспрессии мРНК SCNA в разных клеточных линиях путем обработки антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00257] В Примере 3 был проведен скрининг антисмысловых олигонуклеотидов с различными химическими структурами, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, с применением набора различных клеточных линий в конечной концентрации, составляющей 20 нМ. Применяемые клеточные линии происходят из разных органов и разных видов животных. Приведенные ниже данные подтверждают, что увеличение экспрессии мРНК/белка SCN1A посредством модулирования функции SCN1A-специфичного природного антисмыслового транскрипта не ограничивается одним олигонуклеотидом, тканью или видом и, следовательно, представляет собой общее явление.
Материалы и методы
[00258] Первичные фибробласты человека, несущие мутацию, ассоциированную с синдромом Драве. Первичные фибробласты кожи человека, несущие мутацию, ассоциированную с синдромом Драве, Е1099Х, введенные в культуру Dr. N.Kenyon (University of Miami), выращивали на ростовой среде, состоящей из а-МЕМ (Gibco, cat: 12561-056) с добавлением 10 % ЭБС (Mediatech, cat: 35-015 CV) и 1% противогрибкового и антибактериального агента Antimycotic-Antibiotic (Gibco, cat: 15240-062) при 37°С и 5 % СО2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 2×105/лунка в 6-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 6-луночных планшетах заменяли на свежую ростовую среду (1,5 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для всех олигонуклеотидов приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку 2 мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 6-луночного планшета с клетками. Аналогичную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Кроме того, неактивный олигонуклеотид CUR-1462 применяли в той же концентрации в качестве контроля. После примерно 18 ч инкубации при 37°С и 5% СО2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли, и РНК выделяли из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105), следуя инструкциям производителя. Шестьсот нанограммов очищенной тотальной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Superscript VILO cDNA Synthesis Kit от Invitrogen (cat#11754-250) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (зонды Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCN1A человека). Результаты, полученные с применением всех трех зондов, были очень похожими. Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems). Система для анализа 18S была произведена ABI (cat#4319413E). Изменение кратности генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Согласно альтернативному способу "Измерение в этот же день", все процедуры были выполнены аналогично, но клетки обрабатывали антисмысловыми олигонуклеотидами в первый день, сразу после того как рассеивали их в 6-луночные планшеты.
[00259] Клеточная линия SK-N-AS. Клетки нейробластомы человека SK-N-AS от АТСС (cat#CRL-2137) выращивали на ростовой среде (DMEM. (Mediatech cat#10-013-CV) с добавлением 10% ЭБС (Mediatech cat#MT35-011-CV), пенициллина/стрептомицина (Mediatech cat#MT30-002-CI) и не являющихся незаменимыми аминокислот (non-essential amino acids, NEAA) (HyClone SH30238.01)) при 37°С и 5% CO2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 3×105/лунка в 6-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% СО2 в течение ночи. На следующий день среду в 6-луночных планшетах заменяли на свежую ростовую среду (1,5 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для всех олигонуклеотидов приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку 2 мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 6-луночного планшета с клетками. Аналогичную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Кроме того, неактивный олигонуклеотид CUR-1462 применяли в той же концентрации в качестве контроля. После примерно 18ч инкубации при 37°С и 5% CO2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли, и РНК выделяли из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105), следуя инструкциям производителя. Шестьсот нанограммов очищенной тотальной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Superscript VILO cDNA Synthesis Kit от Invitrogen (cat#11754-250) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (зонды Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCN1A человека). Результаты, полученные с применением всех трех зондов, были очень похожими. Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems). Система для анализа 18S была произведена ABI (cat#4319413E). Изменение кратности генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Согласно альтернативному способу "Измерение в этот же день", все процедуры были выполнены аналогично, но клетки обрабатывали антисмысловыми олигонуклеотидами в первый день, сразу после того как рассеивали их в 6-луночные планшеты.
[00260] Клеточная линия СНР-212. Клетки нейробластомы человека СНР-212 от АТСС (cat#CRL-2273) выращивали на ростовой среде (1: смесь MEM и F-12 (АТСС cat#30-2003 и Mediatech cat#10-080-CV, соответственно) с добавлением 10% ЭБС (Mediatech cat#MT35-011-CV) и пенициллина/стрептомицина (Mediatech cat#MT30-002-CI) при 37°С и 5% CO2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 2×1l05/лунка в 6-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 6-луночыых планшетах заменяли на свежую ростовую среду (1,5 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для всех олигонуклеотидов приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку 2 мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 6-луночного планшета с клетками. Аналогичную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Кроме того, неактивный олигонуклеотид CUR-1462 применяли в той же концентрации в качестве контроля. После примерно 18 ч инкубации при 37°С и 5% CO2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли, и РНК выделяли из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105), следуя инструкциям производителя. Шестьсот нанограммов очищенной тотальной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Superscript VILO cDNA Synthesis Kit от Invitrogen (cat#11754-250) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (зонды Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCN1A человека). Результаты, получение с применением всех трех зондов, были очень похожими. Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems). Система для анализа 18S была произведена ABI (cat#4319413E). Изменение кратности генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Согласно альтернативному способу "Измерение в этот же день", все процедуры были выполнены аналогично, но клетки обрабатывали антисмысловыми олигонуклеотидами в первый день, сразу после того как рассеивали их на 6-луночные планшеты.
[00261] Клеточная линия Vero76. Эмбриональные почечные клетки африканской зеленой мартышки Vero76 от АТСС (cat#CRL-1587) выращивали на ростовой среде (модифицированная по способу Дульбекко среда Игла; DMEM (Cellgrow 10-013-CV) с добавлением 5% ЭБС (Mediatech cat#MT35-011-CV) и пенициллина/стрептомицина (Mediatech cat#MT30-002-CI)) при 37°С и 5% CO2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 105/лунка в 6-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 6-луночных планшетах заменяли на свежую ростовую среду (1,5 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для всех олигонуклеотидов приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку 2 мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 6-луночного планшета с клетками. Аналогичную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Кроме того, неактивный олигонуклеотид CUR-1462 применяли в той же концентрации в качестве контроля. После примерно 18 ч инкубации при 37°С и 5% CO2 среду заменяли на свежую ростовую среду.
Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли, и РНК выделяли из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105), следуя инструкциям производителя. Шестьсот нанограммов очищенной тотальной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Superscript VILO cDNA Synthesis Kit от Invitrogen (cat#11754-250) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (зонды Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCN1A человека). Результаты, полученные с применением всех трех зондов, были очень похожими. Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems). Система для анализа 18S была произведена ABI (cat#4319413E). Изменение кратности генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Согласно альтернативному способу "Измерение в этот же день", все процедуры были выполнены аналогично, но клетки обрабатывали антисмысловыми олигонуклеотидами в первый день, сразу после того как рассеивали их в 6-луночные планшеты.
[00262] Клеточная линия ЗТЗ. Эмбриональные фибробласты мыши ЗТЗ от АТСС (cat#CRL-1658) выращивали на ростовой среде (модифицированная по способу Дульбекко среда Игла; DMEM (Cellgrow 10-013-CV) с добавлением 10% эмбриональной телячьей сыворотки (ЭТС; PCS) (Cellgrow 35-22-CV) и пенициллина/стрептомицина (Mediatech cat#MT30-002-CI)) при 37°С и 5% СО2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 105/лунка в 6-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% СО2 в течение ночи. На следующий день среду в 6-луночных планшетах заменяли на свежую ростовую среду (1,5 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для всех олигонуклеотидов приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку 2 мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 6-луночного планшета с клетками. Аналогичную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Кроме того, неактивный олигонуклеотид CUR-1462 применяли в той же концентрации в качестве контроля. После примерно 18 ч инкубации при 37°С и 5% СО2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли, и РНК выделяли из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105), следуя инструкциям производителя. Шестьсот нанограммов очищенной тотальной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Superscript VILO cDNA Synthesis Kit от Invitrogen (cat#11754-250) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (зонды Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCN1A человека). Результаты, полученные с применением всех трех зондов, были очень похожими. Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems). Система для анализа 18S была произведена ABI (cat#4319413E). Изменение кратности генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Согласно альтернативному способу "Измерение в этот же день", все процедуры были выполнены аналогично, но клетки обрабатывали антисмысловыми олигонуклеотидами в первый день, сразу после того как рассеивали их в 6-луночные планшеты.
[00263] Клеточная линия HepG2. Клетки гепатоцеллюлярной карциномы человека HepG2 от АТСС (cat#HB-8065) выращивали на ростовой среде (MEM/EBSS (Hyclone cat#SH30024 либо Mediatech cat # MT-10-010-CV) с добавлением 10% ЭБС (Mediatech cat# MT35-011-CV) и пенициллина/стрептомицина (Mediatech cat#MT30-002-CI)) при 37°С и 5% СО2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 3×105/лунка в 6-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 6-луночных планшетах заменяли на свежую ростовую среду (1,5 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для всех олигонуклеотидов приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку 2 мкл этого раствора инкубировали с 400 мкл среды Opti-MEM (Gibco cat#31985-070) и 4 мкл Липофектамина 2000 (Invitrogen cat# 11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 6-луночного планшета с клетками. Аналогичную смесь, содержащую 2 мкл воды вместо раствора олигонуклеотида, применяли для ложнотрансфицированных контролей. Кроме того, неактивный олигонуклеотид CUR-1462 применяли в той же концентрации в качестве контроля. После примерно 18ч инкубации при 37°С и 5% CO2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли, и РНК выделяли из клеток с применением SV Total RNA Isolation System от Promega (cat#Z3105), следуя инструкциям производителя. Шестьсот нанограммов очищенной тотальной РНК вводили в реакцию обратной транскрипции, проводимую с применением кДНК-набора Superscript VILO cDNA Synthesis Kit от Invitrogen (cat#11754-250) согласно описанию в протоколе производителя. кДНК, полученные в результате этой реакции обратной транскрипции, применяли для мониторинга генной экспрессии посредством ПЦР в режиме реального времени с применением ABI Taqman Gene Expression Mix (cat#4369510) и праймеров/зондов, сконструированных ABI (зонды Hs00374696_ml, Hs00897350_ml или Hs00897341_ml для SCN1A человека). Результаты, полученные с применением всех трех зондов, были очень похожими. Применяли следующий цикл ПЦР: 50°С в течение 2 мин, 95°С в течение 10 мин, 40 циклов при (95°С в течение 15 секунд, 60°С в течение 1 мин) с применением аппарата для ПЦР в режиме реального времени StepOne Plus Real Time PCR Machine (Applied Biosystems). Система для анализа 18S была произведена ABI (cat#4319413Е). Изменение кратности генной экспрессии после обработки антисмысловыми олигонуклеотидами вычисляли на основании разности значений dCt, нормализованных по 18S, между обработанными и ложнотрансфицированными образцами. Согласно альтернативному способу "Измерение в этот же день", все процедуры были выполнены аналогично, но клетки обрабатывали антисмысловыми олигонуклеотидами в первый день, сразу после того как рассеивали их в 6-луночные планшеты.
[00264] Результаты: Уровни мРНК SCN1A в разных клеточных линиях после обработки антисмысловыми олигонуклеотидами в концентрации 20 нМ по сравнению с ложнотрансфицированным контролем приведены в Таблице 2. Как видно из представленных данных, некоторые из олигонуклеотидов при применении их в концентрации 20 нМ были высокоактивными в повышении уровней мРНК SCN1A и неизменно демонстрировали увеличение экспрессии у нескольких видов (человека, африканской зеленой мартышки и мыши) в клеточных линиях, полученных из разных органов/типов клеток (печени, почки, мозга, эмбриональных фибробластов), и первичных фибробластах кожи, несущих мутацию в гене SCN1A. Увеличение экспрессии белка SCN1A в клетках, несущих мутацию, ассоциированную с синдромом Драве, свидетельствует в пользу пригодности способа для лечения заболеваний, ассоциированных с мутациями в гене SCN1A. Некоторые из олигонуклеотидов, нацеленных на природную антисмысловую последовательность, не влияли или лишь незначительно влияли на уровни мРНК SCN1A во всех или некоторых из исследованных клеточных линий. Эти различия согласуются с литературными данными, которые указывают на то, что связывание олигонуклеотидов может зависеть от вторичной и третичной структур последовательности-мишени для олигонуклеотида. Примечательно, что уровни SCN1A в клетках, обработанных олигонуклеотидами, не имеющими гомологии с природной антисмысловой последовательностью для гена SCN1A, но характеризующиеся сходной химической структурой (CUR-1462), значимо не отличаются от ложнотрансфицированного контроля, и данный факт подтверждает, что эффекты нацеленных олигонуклеотидов не зависят от неспецифической токсичности этих молекул.
Пример 4: Зависимость увеличения экспрессии мРНК SCNA от дозы в разных клеточных линиях при обработке антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00265] В Примере 4 антисмысловые олигонуклеотиды с различными химическими структурами, нацеленные на SCNA-специфичный природный антисмысловой транскрипт, были проскринированы с использованием набора различных клеточных линий в конечных концентрациях, находящихся в диапазоне от 5 до 80 нМ. Применяемые клеточные линии происходят из разных органов и разных видов животных. Приведенные ниже данные подтверждают, что степень увеличения экспрессии мРНК SCNA посредством модулирования функции SCNA-специфичного природного антисмыслового транскрипта может быть изменена путем применения различных количеств активных олигонуклеотидов.
[00266] Материалы и методы: Клетки SK-N-AS, Vero 76 и первичные фибробласты человека, несущие мутацию, ассоциированную с синдромом Драве, обрабатывали антисмысловыми олигонуклеотидами, как описано в Примере 2, за исключением концентраций олигонуклеотидов и Липофектамина 2000, выбранных для добавления в каждую лунку. Концентрации олигонуклеотидов и Липофектамина 2000 подводили таким образом, чтобы обеспечить конечные концентрации олигонуклеотидов, составляющие 5, 10, 20, 40 и 80 нМ, и отношение Липофектамина 2000 к стоковому раствору олигонуклеотида с концентрацией 20 мкМ, равное 2:1 (об./об.).
[00267] Результаты: Результаты экспериментов по исследованию зависимости между дозой и эффектом подтвердили, что антисмысловые олигонуклеотиды, нацеленные на SCNA1-специфичную природную антисмысловую РНК, могут вызвать дозозависимое увеличение экспрессии мРНК SCN1A (Фиг.1-3). В некоторых случаях это увеличение экспрессии было очень большим (вплоть до 60 раз) при более высоких дозах (Фиг.1-3). Степень увеличения экспрессии, индуцированного одним и тем же нуклеотидом в разных клеточных линиях, как оказалось, различается, например, увеличение экспрессии, достигаемое в первичных фибробластах при концентрации 40 нМ, было на уровне 10-40 раз, в то время как увеличение экспрессии в клетках Vero 76 теми же олигонуклеотидами при тех же концентрациях составляло 2-6 раз (Фиг.1 по сравнению с Фиг.3). Эти различия могут быть обусловлены разной эффективностью трансфекции для разных клеточных линий и/или различными путями, действующими по принципу обратной связи, экспрессируемыми ими. Эффект большинства олигонуклеотидов достигал плато при концентрации, составляющей примерно 40 нМ, за исключением олигонуклеотидов CUR-1764 и CUR-1770 в SCN1A-фибробластах и всех олигонуклеотидов, исследованных на клетках Vero 76, где плато не было достигнуто при самых высоких проверяемых концентрациях (Фиг.1-3).
Пример 5; Специфичность к последовательности увеличения экспрессии мРНК SCNA путем применения антисмысловых олигонуклеотидов, нацеленных на SCNA-специфичный природный антисмысловой транскрипт
[00268] В Примере 5 антисмысловые олигонуклеотиды, нацеленные на SCNA-специфичный природный антисмысловой транскрипт, проверили в экспериментах, направленных на подтверждение независимости увеличения экспрессии гена SCN1A, вызываемого действием олигонуклеотидов, от неспецифической токсичности, связанной с химической структурой применяемых олигонуклеотидов. Приведенные ниже данные подтверждают, что степень увеличения экспрессии мРНК SCN1A посредством модулирования функции SCN1A-специфичного природного антисмыслового транскрипта зависит только от количеств активных олигонуклеотидов, а не от общего количества молекул, имеющих сходную химическую структуру.
[00269] Материалы и методы: Клетки Vero 76 и первичные фибробласты человека, несущие мутацию, ассоциированную с синдромом Драве, обрабатывали антисмысловыми олигонуклеотидами, как описано в Примере 2, за исключением концентраций олигонуклеотидов, выбранных для добавления в каждую лунку. Активный олигонуклеотид вводили совместно с неактивным олигонуклеотидом, характеризующимся сходной химической структурой, но не имеющим известной мишени в геноме человека (CUR-1462) и не влияющим на экспрессию нескольких исследованных генов (данные не показаны). Общее количество олигонуклеотидов, как и количество Липофектамина 2000, сохраняли постоянным, в то время как долю активного олигонуклеотида в смеси изменяли. Концентрации олигонуклеотидов подводили таким образом, чтобы обеспечить конечные концентрации активных олигонуклеотидов, составляющие 5, 10, 20, и 40 нМ, и общую концентрацию олигонуклеотидов (активный + неактивный), составляющую 40 нМ.
[00270] Как видно из приведенных данных (Фиг.7), дозозависимый эффект олигонуклеотидов, нацеленных на природную антисмысловую последовательность для гена SCN1A, не был вызван потенциально связанной с такими молекулами неспецифической токсичностью. Уровни мРНК SCN1A зависели от дозы активного олигонуклеотида, применяемого для их увеличения (Фиг.7).
Пример 6: Специфичность к мишени увеличения экспрессии мРНК SCNA путем применения антисмысловых олигонуклеотидов, нацеленных на SCNA-специфичный природный антисмысловой транскрипт
[00271] В Примере 6 антисмысловые олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, проверили в экспериментах, направленных на подтверждение специфичности олигонуклеотидов к их мишени, т.е. SCN1A. Приведенные ниже данные подтверждают, что увеличение экспрессии мРНК SCN1A посредством модулирования функции SCN1A-специфичного природного антисмыслового транскрипта ограничивалось мРНК SCN1A, и олигонуклеотиды не влияли на родственные натриевые каналы SCN9A, SCN8A, SCN7A, SCN3A и SCN2A.
[00272] Материалы и методы: Клетки Vero 76 и первичные фибробласты человека, несущие мутацию, ассоциированную с синдромом Драве, обрабатывали антисмысловыми олигонуклеотидами, как описано в Примере 3. После обработки выполняли анализ выделенной РНК, как описано в Примере 2, за исключением того, что при проведении ПЦР в режиме реального времени применяли системы для анализа экспрессии генов Taqman, позволяющие обнаружить мРНК для каналов SCN9A, SCN8A, SCN7A, SCN3A и SCN2A. Системы для анализа альфа-субъединиц каналов SCN9A, SCN8A, SCN7A, SCN3A и SCN2A человека были получены от ABI Inc. (cat#Hs00161567_ml, Hs00274075_ml, Hs00161546_ml, Hs00366902_ml и Hs00221379_ml, соответственно).
[00273] Результаты: Как показано на Фиг.8, обработка олигонуклеотидами CUR-1916 и CUR-1770 значимо не влияет на экспрессию каналов SCN8A и SCN9A в фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве. Экспрессия каналов SCN7A, SCN3A и SCN2A была необнаружима в этих клетках до или после обработки (данные не показаны). Приведенные данные подтверждают специфичность модулирования экспрессии генов с применением олигонуклеотидов, нацеленных на природную антисмысловую РНК для данного гена.
Пример 7: Стабильность антисмысловых олигонуклеотидов, нацеленных на SCNA-специфичный природный антисмысловой транскрипт
[00274] В Примере 7 две партии антисмыслового олигонуклеотида, нацеленного на SCNA-специфичный природный антисмысловой транскрипт, исследовали в экспериментах, направленных на проверку его стабильности после хранения в разбавленном (1 мМ) водном растворе при температуре 4°С. Приведенные ниже данные показывают, что олигонуклеотиды могут быть стабильными в этих условиях в течение периода времени, составляющего по меньшей мере 6 месяцев, без существенной потери активности.
[00275] Материалы и методы: Клетки Vero 76 обрабатывали двумя различными партиями антисмыслового олигонуклеотида, как описано в Примере 2. Партии были синтезированы в августе 2010 года и марте 2011 года. Олигонуклеотид, синтезированный в августе 2010 года, хранили в виде водного раствора с концентрацией 1 мМ при температуре 4°С. Олигонуклеотид, синтезированный в марте 2011 года, был отправлен в лиофилизированной форме в течение 3 дней после синтеза и проверен сразу после доставки.
[00276] Результаты: Как показано на Фиг.9, не наблюдали значительной потери биологической активности олигонуклеотидов после длительного хранения в течение 6 месяцев в водном растворе при температуре 4°C.
Пример 8: Увеличение экспрессии белка SCNA в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00277] Цель этого эксперимента заключалась в упорядочивании антисмысловых олигонуклеотидов CUR-1740, CUR-1770 и CUR-1916 в соответствии с их способностью увеличивать экспрессию белка SCNA в фибробластах, несущих мутацию, ассоциированную с синдромом Драве.
[00278] Материалы и методы: Фибробласты, несущие мутацию, ассоциированную с синдромом Драве, введенные в культуру Dr. N.Kenyon (University of Miami), выращивали на ростовой среде, состоящей из а-МЕМ (Gibco, cat: 12561-056) с добавлением 10% ЭБС (Mediatech, cat: 35-015 CV) и 1 % противогрибкового и антибактериального агента Antimycotic-Antibiotic (Gibco, cat: 15240-062) при 37°С и 5% CO2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 4×104/лунка в 24-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 24-луночных планшетах заменяли на свежую ростовую среду (1 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами CUR-1740, CUR-1770 и CUR-1916. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для олигонуклеотидов CUR-1740, CUR-1770 и CUR-1916 приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку до конечной концентрации, составляющей 20 нМ, 1 мкл стокового раствора олигонуклеотида с концентрацией 20 мкМ инкубировали с 200 мкл среды Opti-MEM (Gibco cat#31985-070) и 2 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 24-луночного планшета с клетками. Для получения конечных концентраций, составляющих 5, 10, 40 и 80 нМ, объемы стоков олигонуклеотидов с концентрацией 20 мкМ подводили соответственно. Отношение стокового раствора олигонуклеотида с концентрацией 20 мкМ к Липофектамину 2000 составляло 1:2 (об./об.). Аналогичные смеси, содержащие 8 мкл воды вместо раствора олигонуклеотида и соответствующий объем Липофектамина 2000, применяли для ложнотрансфицированных контролей. После примерно 18 ч инкубации при температуре 37°С и 5% CO2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли и клетки промывали 3 раза фосфатно-солевым буфером Дульбекко, не содержащим кальция и магния (ФСБ; PBS) (Mediatech cat#21-031-CV). Затем ФСБ удаляли, а клетки фиксировали в 24-луночном планшете с применением 300 мкл 100 % метанола в течение 15 мин при температуре -20°С. После удаления метанола и промывки ФСБ клетки инкубировали с 3% перекисью водорода (Fisher Chemical cat#11325-100) в течение 5 мин при температуре 21°С. Клетки промывали ФСБ три раза в течение 5 мин, затем инкубировали с 300 мкл бычьего сывороточного альбумина (БСА; BSA) (Sigma cat#A-9647) в ФСБ с концентрацией 0,1% в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза в течение 5 мин, затем инкубировали с 300 мкл раствора авидина (Vector Laboratories cat#SP-2001) в течение 30 мин при температуре 21°С. Клетки быстро промывали ФСБ три раза, затем инкубировали с раствором биотина (Vector Laboratories cat# SP-2001) в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза и затем инкубировали в течение ночи при температуре 4°С с 300 мкл на лунку антител кролика против белка SCN1A человека (Abeam cat#ab24820), разведенных 1:250 в ФСБ/БСА 0,1%. После уравновешивания планшета течение 5 мин при температуре 21°С, клетки промывали ФСБ три раза по 5 мин каждый, затем инкубировали с козьими антикроличьими антителами, разведенными 1:200 в ФСБ/БСА 0,1%, в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза 5 мин и затем инкубировали с 300 мкл смеси Vectastain Elite ABC reagent A+В solution (Vector Laboratories cat# PK-6101) в течение 30 мин; смесь Vectastain Elite ABC reagent A+В solution готовили при температуре 21°С за 30 минут до инкубации с клетками путем добавления и смешивания последовательно 2 капель реагента А в 5 мл ФСБ и затем 2 капель реагента В. Клетки промывали ФСБ три раза по 5 мин каждый при температуре 21°С и далее инкубировали с субстратным раствором для пероксидазы на основе диаминобензидина (ДАБ; DAB) (Vector Laboratories cat#SK-4105) до тех пор, пока клетки не окрашивались; субстратный раствор для пероксидазы на основе ДАБ восстанавливали перед добавлением к клеткам путем смешивания 1 мл разбавителя для ДАБ ImmPACTTMDAB Diluent с 30 мкл концентрированного раствора ДАБ ImmPACTTMAB Chromogen concentrate. В это время клетки быстро промывали ФСБ три раза и оставляли 300 мкл ФСБ в каждой лунке. Окрашивание клеток анализировали непосредственно внутри лунок 24-луночного планшета с применением инвертированного микроскопа Nikon Eclipse TS100, снабженного камерой Nikon DS-Ri1 в сочетании с оборудованием Nikon Digital-Sight на экране портативного компьютера Dell Latitude D630. Фотографии отдельных клеток были сделаны с применением программного обеспечения NIS-Elements D 3.0, поставляемого вместе с камерой Nikon.
[00279] Результаты: Все исследуемые антисмысловые олигонуклеотиды эффективно увеличивали экспрессию белка SCN1A, причем, олигонуклеотиды CUR-1770 и CUR-1916 представляли собой два лучших препарата (Фиг.10).
Пример 9: Увеличение экспрессии белка SCNA в клетках SK-N-AS, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00280] Цель этого эксперимента заключалась в упорядочивании антисмысловых олигонуклеотидов CUR-1740, CUR-1764, CUR-1770 и CUR-1916 в соответствии с их способностью увеличивать экспрессию белка SCN1A в клетках SK-N-AS. Клетки SK-N-AS представляют собой линию клеток нейробластомы человека.
[00281] Материалы и методы: Клетки нейробластомы человека SK-N-AS от АТСС (cat#CRL-2137) выращивали на ростовой среде (DMEM (Mediatech cat#10-013-CV) с добавлением 10% ЭБС (Mediatech cat#MT35-011-CV), пенициллина/стрептомицина (Mediatech cat# MT30-002-CI) и не являющихся незаменимыми аминокислот (NEAA) (HyClone SH30238.01)) при 37°С и 5% CO2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 5×104/лунка в 24-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 24-луночных планшетах заменяли на свежую ростовую среду (1 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами CUR-1740, CUR-1764, CUR-1770 и CUR-1916. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc. (Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для олигонуклеотидов CUR-1740, CUR-1764, CUR-1770 и CUR-1916 приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку до конечной концентрации, составляющей 20 нМ, 1 мкл стокового раствора олигонуклеотида с концентрацией 20 мкМ инкубировали с 200 мкл среды Opti-MEM (Gibco cat#31985-070) и 2 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 24-луночного планшета с клетками. Для получения конечных концентраций, составляющих 5, 10, 40 и 80 нМ, объемы стоков олигонуклеотидов с концентрацией 20 мкМ подводили соответственно. Отношение стокового раствора олигонуклеотида с концентрацией 20 мкМ к Липофектамину 2000 составляло 1:2 (об./об.). Аналогичные смеси, содержащие 8 мкл воды вместо раствора олигонуклеотида и соответствующий объем Липофектамина 2000, применяли для ложнотрансфицированных контролей. После примерно 18 ч инкубации при температуре 37°С и 5% CO2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли и клетки промывали 3 раза фосфатно-солевым буфером Дульбекко, не содержащим кальция и магния (ФСБ; PBS) (Mediatech cat#21-031-CV). Затем ФСБ удаляли, а клетки фиксировали в 24-луночном планшете с применением 300 мкл 100 % метанола в течение 15 мин при температуре -20°С. После удаления метанола и промывки ФСБ клетки инкубировали с 3 % перекисью водорода (Fisher Chemical cat#11325-100) в течение 5 мин при температуре 21°С. Клетки промывали ФСБ три раза в течение 5 мин, затем инкубировали с 300 мкл бычьего сывороточного альбумина (БСА; BSA) (Sigma cat#A-9647) в ФСБ с концентрацией 0,1% в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза в течение 5 мин, затем инкубировали с 300 мкл раствора авидина (Vector Laboratories cat# SP-2001) в течение 30 мин при температуре 21°С. Клетки быстро промывали ФСБ три раза, затем инкубировали с раствором биотина (Vector Laboratories cat#SP-2001) в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза и затем инкубировали в течение ночи при температуре 4°С с 300 мкл на лунку антител кролика против белка SCN1A человека (Abeam cat#ab24820), разведенных 1:250 в ФСБ/БСА 0,1%. После уравновешивания планшета течение 5 мин при температуре 21°С, клетки промывали ФСБ три раза по 5 мин каждый, затем инкубировали с козьими антикроличьими антителами, разведенными 1:200 в ФСБ/БСА 0,1 %, в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза 5 мин и затем инкубировали с 300 мкл смеси Vectastain Elite ABC reagent A+В solution (Vector Laboratories cat# PK-6101) в течение 30 мин; смесь Vectastain Elite ABC reagent A+В solution готовили при температуре 21°С за 30 минут до инкубации с клетками путем добавления и смешивания последовательно 2 капель реагента А в 5 мл ФСБ и затем 2 капель реагента В. Клетки промывали ФСБ три раза по 5 мин каждый при температуре 21°С и далее инкубировали с субстратным раствором для пероксидазы на основе диаминобензидина (ДАБ; DAB) (Vector Laboratories cat#SK-4105) до тех пор, пока клетки не окрашивались; субстратный раствор для пероксидазы на основе ДАБ восстанавливали перед добавлением к клеткам путем смешивания 1 мл разбавителя для ДАБ ImmPACTTMDAB Diluent с 30 мкл концентрированного раствора ДАБ ImmPACTTMDAB Chromogen concentrate. В это время клетки быстро промывали ФСБ три раза и оставляли 300 мкл ФСБ в каждой лунке. Окрашивание клеток анализировали непосредственно внутри лунок 24-луночного планшета с применением инвертированного микроскопа Nikon Eclipse TS100, снабженного камерой Nikon DS-Ri1 в сочетании с оборудованием Nikon Digital-Sight на экране портативного компьютера Dell Latitude D630. Фотографии отдельных клеток были сделаны с применением программного обеспечения NIS-Elements D 3.0, поставляемого вместе с камерой Nikon.
[00282] Результаты: Все исследуемые антисмысловые олигонуклеотиды увеличивали экспрессию белка SCN1A, причем, олигонуклеотиды CUR-1764 и CUR-1770 представляли собой два лучших препарата (Фиг.11).
Пример 10: Увеличение экспрессии белка SCNA в клетках Vero 76, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00283] Цель этого эксперимента заключалась в упорядочивании антисмысловых олигонуклеотидов CUR-1740, CUR-1770, CUR-1916, CUR-1924 и CUR-1945 в соответствии с их способностью увеличивать экспрессию белка SCN1A в клетках Vero 76. Клетки Vero76 представляют собой линию почечных клеток Cercopithecus aethiops (зеленой мартышки, или африканской зеленой мартышки).
[00284] Материалы и методы: Эмбриональные почечные клетки африканской зеленой мартышки Vero76 от АТСС (cat#CRL-1587) выращивали на ростовой среде (модифицированная по способу Дульбекко среда Игла; DMEM (Cellgrow 10-013-CV) с добавлением 5% ЭБС (Mediatech cat#MT35-011-CV) и пенициллина/стрептомицина (Mediatech cat# MT30-002-CI)) при 37°С и 5% CO2. Клетки обрабатывали антисмысловыми олигонуклеотидами с применением одного из следующих способов. Согласно способу "Измерение на следующий день", за один день до эксперимента клетки пересеивали при плотности приблизительно 4×104/лунка в 24-луночные планшеты в ростовую среду и инкубировали при 37°С и 5% CO2 в течение ночи. На следующий день среду в 24-луночных планшетах заменяли на свежую ростовую среду (1 мл/лунка), и клетки обрабатывали антисмысловыми олигонуклеотидами CUR-1740, CUR-1770 и CUR-1916. Все антисмысловые олигонуклеотиды были изготовлены в IDT Inc.
(Coralville, IA) или Exiqon (Vedback, Denmark). Последовательности для олигонуклеотидов CUR-1740, CUR-1770, CUR-1916, CUR-1924 и CUR-1945 приведены в Таблице 1. Стоковые растворы олигонуклеотидов разводили до концентрации 20 мкМ в стерильной воде, не содержащей ДНКаз/РНКаз. Для добавления раствора олигонуклеотида в одну лунку до конечной концентрации, составляющей 20 нМ, 1 мкл стокового раствора олигонуклеотида с концентрацией 20 мкМ инкубировали с 200 мкл среды Opti-MEM (Gibco cat#31985-070) и 2 мкл Липофектамина 2000 (Invitrogen cat#11668019) при комнатной температуре в течение 20 мин и вносили по каплям в одну лунку 24-луночного планшета с клетками. Для получения конечных концентраций, составляющих 5, 10, 40 и 80 нМ, объемы стоков олигонуклеотидов с концентрацией 20 мкМ подводили соответственно. Отношение стокового раствора олигонуклеотида с концентрацией 20 мкМ к Липофектамину 2000 составляло 1:2 (об./об.). Аналогичные смеси, содержащие 8 мкл воды вместо раствора олигонуклеотида и соответствующий объем Липофектамина 2000, применяли для ложнотрансфицированных контролей. После примерно 18ч инкубации при температуре 37°С и 5% CO2 среду заменяли на свежую ростовую среду. Через сорок восемь часов после добавления антисмысловых олигонуклеотидов среду удаляли и клетки промывали 3 раза фосфатно-солевым буфером Дульбекко, не содержащим кальция и магния (ФСБ; PBS) (Mediatech cat#21-031-CV). Затем ФСБ удаляли, а клетки фиксировали в 24-луночном планшете с применением 300 мкл 100 % метанола в течение 15 мин при температуре -20°С. После удаления метанола и промывки ФСБ клетки инкубировали с 3 % перекисью водорода (Fisher Chemical cat#11325-100) в течение 5 мин при температуре 21°С. Клетки промывали ФСБ три раза в течение 5 мин, затем инкубировали с 300 мкл бычьего сывороточного альбумина (БСА; BSA) (Sigma cat#A-9647) в ФСБ с концентрацией 0,1 % в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза в течение 5 мин, затем инкубировали с 300 мкл раствора авидина (Vector Laboratories cat#SP-2001) в течение 30 мин при температуре 21°С. Клетки быстро промывали ФСБ три раза, затем инкубировали с раствором биотина (Vector Laboratories cat# SP-2001) в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза и затем инкубировали в течение ночи при температуре 4°С с 300 мкл на лунку антител кролика против белка SCN1A человека (Abeam cat#ab24820), разведенных 1:250 в ФСБ/БСА 0,1%. После уравновешивания планшета течение 5 мин при температуре 21°С, клетки промывали ФСБ три раза по 5 мин каждый, затем инкубировали с козьими антикроличьими антителами, разведенными 1:200 в ФСБ/БСА 0,1%, в течение 30 мин при температуре 21°С. Клетки промывали ФСБ три раза 5 мин и затем инкубировали с 300 мкл смеси Vectastain Elite ABC reagent A+В solution (Vector Laboratories cat#PK-6101) в течение 30 мин; смесь Vectastain Elite ABC reagent A+В solution готовили при температуре 21°С за 30 минут до инкубации с клетками путем добавления и смешивания последовательно 2 капель реагента А в 5 мл ФСБ и затем 2 капель реагента В. Клетки промывали ФСБ три раза по 5 мин каждый при температуре 21°С и далее инкубировали с субстратным раствором для пероксидазы на основе диаминобензидина (ДАБ; DAB) (Vector Laboratories cat#SK-4105) до тех пор, пока клетки не окрашивались; субстратный раствор для пероксидазы на основе ДАБ восстанавливали перед добавлением к клеткам путем смешивания 1 мл разбавителя для ДАБ ImmPACTTMDAB Diluent с 30 мкл концентрированного раствора ДАБ ImmPACTTMDAB Chromogen concentrate. В это время клетки быстро промывали ФСБ три раза и оставляли 300 мкл ФСБ в каждой лунке. Окрашивание клеток анализировали непосредственно внутри лунок 24-луночного планшета с применением инвертированного микроскопа Nikon Eclipse TS100, снабженного камерой Nikon DS-Ri1 в сочетании с оборудованием Nikon Digital-Sight на экране портативного компьютера Dell Latitude D630. Фотографии отдельных клеток были сделаны с применением программного обеспечения NIS-Elements D 3.0, поставляемого вместе с камерой Nikon.
[00285] Результаты: Все исследуемые антисмысловые олигонуклеотиды увеличивали экспрессию белка SCN1A, причем, олигонуклеотиды CUR-1764 и CUR-1770 обеспечивали наибольшую степень увеличения экспрессии (Фиг.12).
Пример 11: Олигонуклеотиды, нацеленные на SCNA-специфичный природный антисмысловой транскрипт, обладающие способностью увеличивать экспрессию мРНК SCNA, не увеличивают экспрессию мРНК актина в клетках Vero76
[00286] Цель данного эксперимента заключалась в проверке того, способны ли антисмысловые олигонуклеотиды (CUR-1924, CUR-1740, CUR-1838), нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCN1A, регулировать экспрессию мРНК других неродственных генов, таких как ген актина, в эмбриональных почечных клетках африканской зеленой мартышки Vero76.
[00287] Материалы и методы: Эмбриональные почечные клетки африканской зеленой мартышки линии Vero76 от АТСС (cat#CRL-1587) обрабатывали олигонуклеотидами в тех же условиях, как описано в Примере 2. мРНК актина определяли количественно посредством ПЦР в режиме реального времени, как описано в Примере 2, за исключением того, что на этот раз праймеры/зонды, сконструированные ABI, были специфичны для актина (cat#Hs99999903_ml). Данные представлены на Фигуре 13.
[00288] Результаты: Как показано на Фигуре 13, олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1924, CUR-1740, CUR-1838), которые, как было показано в Примерах 3 и 10, увеличивают экспрессию мРНК и белка SCN1A в клетках Vero76, были исследованы на их действие в отношении экспрессии мРНК актина в клетках Vero 76. Данные, представленные на Фигуре 13, подтверждают, что олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, не увеличивают экспрессию неродственного гена, такого как ген актина. Таким образом, эти олигонуклеотиды являются специфичными в отношении увеличения экспрессии гена SCN1A.
Пример 12: Олигонуклеотиды, нацеленные на SCNA-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCNA, не увеличивают экспрессию мРНК актина в первичных фибробластах, несущих мутацию, ассоциированную с синдромом Драве
[00289] Цель данного эксперимента заключалась в проверке того, способны ли антисмысловые олигонуклеотиды (CUR-1916, CUR-1945), нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCN1A, регулировать экспрессию мРНК других неродственных генов, таких как ген актина, в первичных фибробластах кожи человека, несущих мутацию, ассоциированную с синдромом Драве, Е1099Х.
[00290] Материалы и методы: Первичные фибробласты кожи человека, несущие мутацию, ассоциированную с синдромом Драве, Е1099Х, введенные в культуру Dr. N.Kenyon (University of Miami), обрабатывали олигонуклеотидами в тех же условиях, как описано в Примере 3. мРНК актина определяли количественно посредством ПЦР в режиме реального времени, как описано в Примере 3, за исключением того, что на этот раз праймеры/зонды, сконструированные ABI, были специфичны для актина (cat#Hs99999903_ml). Данные представлены на Фигуре 14.
[00291] Результаты: Как показано на Фигуре 14, олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, не увеличивают экспрессию неродственного гена, такого как ген актина. Таким образом, эти олигонуклеотиды являются специфичными в отношении увеличения экспрессии гена SCN1A.
Пример 13: Олигонуклеотиды, нацеленные на SCNA-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCNA, не увеличивают экспрессию мРНК актина в клетках SK-N-AS
[00292] Цель данного эксперимента заключалась в проверке того, способны ли антисмысловые олигонуклеотиды (CUR-1740, CUR-1764, CUR-1770, CUR-1838, CUR-1916), нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, которые, как было показано, увеличивают экспрессию мРНК и белка SCN1A, регулировать экспрессию мРНК других неродственных генов, таких как ген актина, в клетках нейробластомы человека SK-N-AS.
[00293] Материалы и методы: Клетки нейробластомы человека SK-N-AS от АТСС (cat#CRL-2137) обрабатывали олигонуклеотидами в тех же условиях, как описано в Примере 2. мРНК актина определяли количественно посредством ПЦР в режиме реального времени, как описано в Примере 2, за исключением того, что на этот раз праймеры/зонды, сконструированные ABI, были специфичны для актина (cat#Hs99999903_ml). Данные представлены на Фигуре 15.
[00294] Результаты: Как показано на Фигуре 15, олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт, не увеличивают экспрессию неродственного гена, такого как ген актина. Таким образом, эти олигонуклеотиды являются специфичными в отношении увеличения экспрессии гена SCN1A.
Пример 14: Экспрессия белка актина не увеличивается в клетках SK-N-AS, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00295] Цель данного эксперимента заключалась в определении того, могут ли олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1740, CUR-1764, CUR-1770 и CUR-1916) и способные увеличивать экспрессию белка SCN1A, также регулировать экспрессию неродственных белков, таких как актин, в клетках SK-N-AS. Клетки SK-N-AS представляют собой линию клеток нейробластомы человека.
[00296] Материалы и методы: Клетки нейробластомы человека SK-N-AS от АТСС (cat#CRL-2137) выращивали в тех же условиях, как описано в Примере 9. Клетки фиксировали и окрашивали точно в таких же условиях, как описано в Примере 8, за исключением того, что первые антитела представляли собой кроличьи антитела к актину (Abeam cat#ab1801), применяемые в разведении 1:500. Окрашивание клеток анализировали непосредственно внутри лунок 24-луночного планшета, применяя ту же самую методику, как описано в Примере 9.
[00297] Результаты: Как показано на Фигуре 16, ни один из исследованных антисмысловых олигонуклеотидов не увеличивал экспрессию белка актина. Таким образом, эти олигонуклеотиды являются специфичными в отношении увеличения экспрессии белка SCN1A.
Пример 15: Экспрессия белка актина не увеличивается в клетках Vero 76, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00298] Цель данного эксперимента заключалась в определении того, могут ли специфические антисмысловые олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1740, CUR-1770, CUR-1916, CUR-1924 и CUR-1945) и способные увеличивать экспрессию белка SCN1A, также регулировать экспрессию белков неродственных генов, таких как актин, в клетках Vero76. Клетки Vero76 представляют собой линию почечных клеток Cercopithecus aethiops (зеленой мартышки, или африканской зеленой мартышки).
[00299] Материалы и методы: Эмбриональные почечные клетки африканской зеленой мартышки линии Vero76 от АТСС (cat#CRL-1587) выращивали в тех же условиях, как описано в Примере 10. Клетки фиксировали и окрашивали точно в таких же условиях, как описано в Примере 10, за исключением того, что первые антитела представляли собой кроличьи антитела к актину (Abeam cat#ab1801), применяемые в разведении 1:500. Окрашивание клеток анализировали непосредственно внутри лунок 24-луночного планшета, применяя ту же самую методику, как описано в Примере 10.
[00300] Результаты: Как показано на Фигуре 17, ни один из исследованных антисмысловых олигонуклеотидов не увеличивал экспрессию белка актина. Таким образом, эти олигонуклеотиды являются специфичными в отношении увеличения экспрессии белка SCN1A.
Пример 16: Экспрессия белка актина не увеличивается в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве, обработанных антисмысловыми олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00301] Цель данного эксперимента заключалась в определении того, могут ли олигонуклеотиды, нацеленные на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1740, CUR-1764, CUR-1770, CUR-1838 и CUR-1916) и способные увеличивать экспрессию белка SCNA, также регулировать экспрессию белков неродственных генов, таких как актин, в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве.
[00302] Материалы и методы: Фибробласты, несущие мутацию, ассоциированную с синдромом Драве, введенные в культуру Dr. N.Kenyon (University of Miami), выращивали в тех же условиях, как описано в Примере 8. Клетки фиксировали и окрашивали точно в таких же условиях, как описано в Примере 8, за исключением того, что первые антитела представляли собой кроличьи антитела к актину (Abeam cat#ab1801), применяемые в разведении 1:500. Окрашивание клеток анализировали непосредственно внутри лунок 24-луночного планшета, применяя ту же самую методику, как описано в Примере 8.
[00303] Результаты: Как показано на Фигуре 18, ни один из исследованных антисмысловых олигонуклеотидов не увеличивал экспрессию белка актина. Таким образом, эти олигонуклеотиды являются специфичными в отношении увеличения экспрессии белка SCN1A.
Пример 17: Количественное определение белка SCNA с применением ИФА в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве, обработанных олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00304] Цель этого эксперимента состояла в количественном определении посредством ИФА уровня увеличения экспрессии белка SCN1A, происходящего в результате обработки олигонуклеотидами, нацеленными на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1740, CUR-1770 и CUR-1916), в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве.
[00305] Материалы и методы: Фибробласты, несущие мутацию, ассоциированную с синдромом Драве, введенные в культуру Dr. N.Kenyon (University of Miami), выращивали в тех же условиях, как описано в Примере 8, но обработку олигонуклеотидами осуществляли только в концентрациях, составляющих 0 и 80 нМ. Затем клетки подсчитывали и повторно высевали в 96-луночные планшеты. Через 24 часа клетки фиксировали точно в таких же условиях, как описано в Примерах 8 и 16, за исключением того, что все объемы, равные 300 мкл, уменьшили до 100 мкл. Повторные лунки окрашивали антителами к актину и белку SCN1A, как описано в Примере 8, за исключением того, что все реакции проводили в объемах, равных 100 мкл. Разведение антител к актину составляло 1:500, разведение антител к белку SCN1A составляло 1:250, а разведение антимышиных антител составляло 1:250. Кроме того, вместо субстратного раствора для пероксидазы на основе диаминобензидина (ДАБ), применяли субстратный раствор для пероксидазы на основе тетраметилбензидина (ТМБ; ТМВ) (Thermo Scientific cat#N301). После того, как супернатант окрашивался в синий цвет, его переносили в новый 96-луночный планшет (Greiner bio one cat#65121) и добавляли 1 М серную кислоту. Считывали поглощение при 450 нм с применением спектрофотометра Multiskan Spectrum (Thermo Scientific). Фоновый сигнал (считываемый в лунках, окрашенных антимышиными антителами в качестве первичных антител) вычитали из всех показаний для SCN1A и актина. Далее сигнал SCN1A нормализовывали к сигналу актина для каждого условия.
[00306] Результаты: Как показано на Фигуре 19, все исследованные антисмысловые олигонуклеотиды (CUR-1740, CUR-1770 и CUR-1916) были эффективны с точки зрения увеличения экспрессии белка SCN1A вплоть до 40%.
Пример 18: Количественное определение белка SCNA с применением ИФА в клетках Vero76, обработанных олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00307] Цель этого эксперимента состояла в количественном определении посредством ИФА уровня увеличения экспрессии белка SCN1A, происходящего в результате обработки олигонуклеотидами, нацеленными на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1740, CUR-1770, CUR-1916, CUR-1924 и CUR-1945), в первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве.
[00308] Материалы и методы: Эмбриональные почечные клетки африканской зеленой мартышки Vero76 выращивали в тех же условиях, как описано в Примере 10, но обработку олигонуклеотидами осуществляли только в концентрациях, составляющих 0 и 80 нМ. Затем клетки подсчитывали и повторно высевали в 96-луночные планшеты. Через 24 часа клетки фиксировали точно в таких же условиях, как описано в Примере 8, за исключением того, что все объемы, равные 300 мкл, уменьшили до 100 мкл. Повторные лунки окрашивали антителами к актину и белку SCN1A, как описано в Примерах 10 и 15, за исключением того, что все реакции проводили в объемах, равных 100 мкл, разведение антител к актину составляло 1:500, разведение антител к белку SCN1A составляло 1:250, а разведение антимышиных антител составляло 1:250. Кроме того, вместо субстратного раствора для пероксидазы на основе диаминобензидина (ДАБ), применяли субстратный раствор для пероксидазы на основе тетраметилбензидина (ТМБ; ТМВ) (Thermo Scientific cat#N301). После того, как супернатант окрашивался в синий цвет, его переносили в новый 96-луночный планшет (Greiner bio one cat#651201) и добавляли 1 М серную кислоту. Считывали поглощение при 450 нм с применением спектрофотометра Multiskan Spectrum (Thermo Scientific). Фоновый сигнал (считываемый в лунках, окрашенных антимышиными антителами в качестве первичных антител) вычитали из всех показаний для SCN1A и актина. Далее сигнал SCN1A нормализовывали к сигналу актина для каждого условия.
[00309] Результаты: Как показано на Фигуре 20, все исследованные антисмысловые олигонуклеотиды (CUR-1740, CUR-1770, CUR-1916, CUR-1924, CUR-1945) были эффективны с точки зрения увеличения экспрессии белка SCN1A вплоть до 300 %.
Пример 19: Количественное определение белка SCNA с применением ИФА в клетках SK-N-AS, обработанных олигонуклеотидами, нацеленными на SCNA-специфичный природный антисмысловой транскрипт
[00310] Цель этого эксперимента состояла в количественном определении уровня увеличения экспрессии белка SCN1A, происходящего в результате обработки олигонуклеотидами, нацеленными на SCN1A-специфичный природный антисмысловой транскрипт (CUR-1740, CUR-1770, CUR-1924 и CUR-1945), в клетках SK-N-AS.
[00311] Материалы и методы: Клетки нейробластомы человека SK-N-AS от АТСС (cat#CRL-2137) выращивали в тех же условиях, как описанные в Примере 10, но обработку олигонуклеотидами осуществляли только в концентрациях, составляющих 0 и 20 нМ. Затем клетки подсчитывали и повторно высевали в 96-луночные планшеты. Через 24 часа клетки фиксировали точно в таких же условиях, как описано в Примере 9, за исключением того, что все объемы, равные 300 мкл, уменьшили до 100 мкл. Повторные лунки окрашивали антителами к актину и белку SCN1A, как описано в Примерах 9 и 13, за исключением того, что все реакции проводили в объемах, равных 100 мкл, разведение антител к актину составляло 1: 500, разведение антител к белку SCN1A составляло 1:250, а разведение антимышиных антител составляло 1: 250. Кроме того, вместо субстратного раствора для пероксидазы на основе диаминобензидина (ДАБ), применяли субстратный раствор для пероксидазы на основе тетраметилбензидина (ТМБ; ТМВ) (Thermo Scientific cat#N301). После того, как супернатант окрашивался в синий цвет, его переносили в новый 96-луночный планшет (Greiner bio one cat#651201) и добавляли 1 М серную кислоту. Считывали поглощение при 450 нм с применением спектрофотометра Multiskan Spectrum (Thermo Scientific). Фоновый сигнал (считываемый в лунках, окрашенных антимышиными антителами в качестве первичных антител) вычитали из всех показаний для SCN1A и актина. Далее сигнал SCN1A нормализовывали к сигналу актина для каждого условия.
[00312] Результаты: Как показано на Фигуре 21, все исследованные антисмысловые олигонуклеотиды (CUR-1740, CUR-1770, CUR-1924 и CUR-1945) были эффективны с точки зрения увеличения экспрессии белка SCN1A в клетках SK-N-AS вплоть до 500%.
Пример 20: Обнаружение природной антисмысловой последовательности BG724147 в клетках HepG2 и первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве
[00313] Цель этого эксперимента состояла в определении того, присутствует ли природная антисмысловая последовательность BG724147 в линии клеток гепатоцеллюлярной карциномы человека HepG2 и первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве. Для достижения этого применяли два разных вида РНК (РНК, содержащую поли(А)-последовательность, (поли(А)-РНК) и тотальную РНК), выделенные из каждого типа клеток. ПНР-продукты, полученные после двух последовательных циклов ПЦР для обоих типов клеток, анализировали посредством электрофореза в геле. Амплификация нуклеотидных последовательностей, дающих зоны одинакового размера при использовании ВС724147-специфичного праймера, подтвердила присутствие последовательности BG724147 в обоих типах клеток.
Материалы и методы
[00314] Выделение тотальной РНК. Клетки HepG2 или первичные фибробласты человека, несущие мутацию, ассоциированную с синдромом Драве, выращенные в культуральных флаконах с площадью поверхности 75 см2 до 80% слияния, дважды промыли ФСБ от AccuGENE IX (Lonza Rockeland Inc., Rockeland, ME). После удаления ФСБ к этим клеткам добавили 5 мл буфера RLT с р-меркаптоэтанолом (QIAGEN Inc. - USA, Valencia, CA), и клеточный лизат хранили в аликвотах объемом 1 мл в микроцентрифужных пробирках при температуре -80°С до выделения тотальной РНК. Выделение тотальной РНК из этих клеток выполняли с применением набора для выделения РНК RNeasy midi kit (Qiagen Inc. - USA, Valencia, CA), следуя протоколу производителя. Кратко, клеточный лизат центрифугировали при 3000 × g в течение 5 мин для очистки лизата и удаления какого-либо осадка. Очищенный клеточный лизат центрифугировали через колонки QIAshredder (внутри микроцентрифужных пробирок объемом 2 мл) при 14800 × g, и полученный гомогенизированный лизат смешали с равным объемом 70 % этанола. Клеточный лизат, смешанный с этанолом, нанесли на колонки RNeasy midi columns (внутри конической пробирки объемом 15 мл) и центрифугировали в течение 5 мин при 3000 × g. Колонку промыли один раз 4 мл буфера RW1 и затем прямо на колонке подвергали лизат расщеплению ДНКазой в течение 15 мин путем добавления 140 мкл раствора ДНКазы, не содержащего РНКаз, в буфере RDD. Расщепление ДНКазой останавливали добавлением 4 мл буфера RW1 и центрифугированием колонки при 3000 × g. Колонку дважды промыли буфером RPE, и тотальную РНК, связанную с фильтром, элюировали 150 мкл воды, не содержащей ДНКаз и РНКаз. Тотальную РНК хранили при -80°С до следующей стадии.
[00315] Выделение РНК, содержащей поли(А)-последовательность, (поли(А)-РНК) из тотальной РНК клеток HepG2. Выделение поли(А)-РНК из тотальной РНК клеток HepG2 и первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве, выполняли с применением набора для выделения поли(А)-РНК, в состав которого входят магнитные частицы, от Ambion (Applied Biosystems/Ambion, Austin, TX), следуя протоколу производителя. В общем, 100 мкг тотальной РНК ресуспендировали в воде, не содержащей ДНКаз и РНКаз, до конечной концентрации, составляющей 600 мкг/мл, и добавили равный объем 2Х раствора для связывания. В течение этого времени, 10 мкл магнитных частиц, содержащих олиго(dT), поместили в микроцентрифужную пробирку, захватили путем помещения этой пробирки на магнитную подставку и удалили буфер для хранения частиц. К частицам добавили 50 мкл Промывочного раствора 1, и пробирку сняли с магнитной подставки, и промывочный раствор удалили. В это время тотальную РНК из клеток HepG2 в IX Связывающем буфере смешали с магнитными частицами и нагревали при 70°С в течение 5 мин, затем инкубировали в течение 60 мин при комнатной температуре с осторожным перемешиванием. Поли(А)-РНК, связанную с магнитными частицами, захватывали с помощью магнитной подставки в течение 5 мин. Супернатант удалили. Магнитные частицы, содержащие олиго(dT), дважды промыли Промывочным раствором 1 и один раз - Промывочным раствором 2 для удаления неспецифически связанной РНК. Магнитные частицы захватили с помощью магнитной подставки и добавили к ним 200 мкл теплого раствора для хранения РНК (предварительно нагреваемого при температуре 70°С в течение 5 мин). Магнитные частицы захватили с помощью магнитной подставки, супернатант сохранили (первый элюат поли(А)-РНК). Далее к частицам добавили вторые 200 мкл теплого раствора для хранения РНК (предварительно нагреваемого при температуре 70°С в течение 5 мин). Второй элюат поли(А)-РНК добавили к первому элюату. На данном этапе элюированную РНК осаждали с применением 5 М ацетата аммония, гликогена и 100% этанола при температуре -20°С в течение ночи. Поли(А)-РНК центрифугировали при 14800 × g в течение 30 мин при 4°С. Супернатант удалили, а осадок РНК промывали три раза добавлением 1 мл 70% этанола, при этом осадок РНК каждый раз собирали посредством центрифугирования в течение 10 мин при 4°С. Наконец, осадок поли(А)-РНК ресуспендировали в растворе для хранения РНК, нагретом до 70°C, для лучшего растворения РНК. Поли(А)-РНК хранили при -80°С.
[00316] Добавление аденозинов к 3'-концу РНК-транскрипта. Тотальную РНК (40 мкг) из клеток HepG2 или первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве, смешали с 2 единицами РНК-поли(А)-полимеразы так, что конечный объем реакционной смеси составлял 100 мкл (Ambion, Applied Biosystems, St. Austin TX). АТФ, применяемая для реакции полиаденилирования, была приобретена у Invitrogen. После процесса полиаденилирования РНК очищали с применением фенол-хлороформного метода с последующим осаждением гликогеном/ацетом натрия. Эту РНК ресуспендировали в 40 мкл воды, не содержащей ДНКаз и РНКаз, и применяли для реакции быстрой амплификации 3'-концевого фрагмента кДНК (3'-RACE) (набор FirstChoice RLM-RACE kit от Ambion, Applied Biosystems, St. Austin TX).
[00317] 3'-Достройка природного антисмыслового транскрипта BG724147 гена SCN1A. Проводили два разных ряда реакций 3'-быстрой амплификации концевых фрагментов кДНК (RACE) с применением набора FirstChoice RLM-RACE kit от Ambion, Applied Biosystems (St. Austin, TX). Для реакций одного ряда применяли поли(А)-РНК, а для другого - тотальную РНК с одним добавленным аденозином из клеток HepG2 или первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве. Проводили два последовательных цикла ПЦР. Первый цикл ПЦР был осуществлен с применением 3'-внешнего праймера, поставляемого в наборе, и 5'-праймера, специфичного для последовательности BG724147, сконструированного ОРКО CURNA (5'-GATTCTCCTACAGCAATTGGTA-3'). Второй цикл ПЦР был проведен с применением 3'-внешнего праймера, поставляемого в наборе, и другого 5'-праймера, специфичного для последовательности BG724147, сконструированного ОРКО CURNA (5'-GACATGTAATCACTTTCATCAA-3'). Продукты второй реакции ПЦР подвергли анализу методом электрофореза в 1% агарозном геле-lx буфере ТАЕ (трис/уксусная кислота/ЭДТА).
[00318] Результаты: Фигура 22 иллюстрирует продукты из второго цикла реакций ПЦР, проведенных в ходе 3'-КАСЕ-экспериментов с применением поли(А)-РНК и тотальной РНК с добавленным аденозином из клеток HepG2 и поли(А)-РНК и тотальной РНК с добавленным аденозином из первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве. Идентичная зона наблюдается в случае поли(А)-РНК из клеток HepG2 и первичных фибробластов человека, несущих мутацию, ассоциированную с синдромом Драве.
[00319] Вывод: ПЦР-амплификация с применением праймеров, специфичных для природного антисмыслового транскрипта BG724147 гена SCN1A, привела к получению общего ПЦР-продукта в двух разных видах клеток (клетках HepG2 и первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве). Кроме того, антисмысловые олигонуклеотиды, нацеленные на природную антисмысловую последовательность BG724147 для гена SCN1A, как было показано, увеличивали экспрессию мРНК и белка SCN1A в этих клетках, как проиллюстрировано в Примерах 2, 7 и 16. Приведенные данные указывают на то, что последовательность BG724147 действительно присутствует в этих двух видах клеток (клетках HepG2 и первичных фибробластах человека, несущих мутацию, ассоциированную с синдромом Драве).
Пример 21: Расширение природной антисмысловой последовательности BG724147 для гена SCNA
[00320] Цель этого эксперимента состояла в расширении известной последовательности природной антисмысловой последовательности BG724147 для гена SCN1A путем секвенирования всей ее последовательности. Оригинальный РНК-транскрипт последовательности BG724147 был получен из семенников человека, доставленных Miklos Palkovits. Библиотека кДНК была получена в векторе pBluescriptR, разработанном Michael J. Brownstein (в Национальном научно-исследовательском институте генома человека; NHGRI), Shiraki Toshiyuki и Piero Caminci (в Институте физико-химических исследований; RIKEN). Библиотека кДНК была упорядочена I.M.A.G.E. Consortium (или LLNL), и клоны были просеквенированы в Incyte Genomics, Inc. в мае 2001 года. Клон BG724147 доступен у Open Biosystems (Open Biosystems Products, Huntsville, AL). В 2001 году кДНК-вставка в клоне BG724147 не была просеквенирована полностью. Компания OPKO-CURNA получила клон BG724147 и выполнила секвенирование полной вставки. Для достижения этой цели бактериальный клон, содержащий плазмиду со вставкой BG724147, был приобретен у Open Biosystems и посеян на чашку, содержащую среду Luria Bertani (LB)-агар с ампициллином, для выделения индивидуальных колоний. Затем колонии растили в 5 мл бульона LB. Далее плазмиду, содержащую вставку BG724147, выделяли из этих бактерий и отправляли для секвенирования в Davis Sequencing (Davis, CA).
Материалы и методы
[00321] Выделение и секвенирование плазмиды, содержащей кДНК для природной антисмысловой последовательности BG724147 для гена SCNA. Суспензия замороженных бактерий, содержащих плазмиду BG724147, была приобретена у Open Biosystems (Open Biosystems Products, cat#4829512), разбавлена в 1:10, 1:100, 1:1000, 1:10000, 1:100000 раз, затем посеяна на чашку (Falcon, cat#351005), содержащую среду Luria Bertani (LB) (BD, cat#244520)-агар с добавлением 100 мкг/мл ампициллина (Calbiochem, cat#171254). Через 15 ч выделили 20 индивидуальных бактериальных колоний из чашки с разведением 1: 100000 и вырастили их отдельно в 5 мл бульона LB (Fisher Scientific, cat#BP1426-2) в течение 15 ч - 24 ч. На данном этапе бактерии осаждали, и плазмиду (содержащую кДНК из РНК-транскрипта BG724147) выделяли с применением набора PureYieldTM Plasmid Miniprep System kit от Promega (Promega, cat#A1222), следуя протоколу производителя. Выделенную ДНК разводили до концентрации, составляющей 200 нг/мл, и 12 мкл плазмиды из каждой колонии отправляли для секвенирования в Davis Sequencing (Davis, CA).
[00322] Результаты: Последовательности, полученные от Davis Sequencing, обеспечивают составление расширенной последовательности BG724147 (SEQ ID NO: 12).
[00323] Вывод: Успешное расширение известной последовательности BG724147 на 403 нуклеотида послужило основой для разработки антисмысловых олигонуклеотидов, нацеленных на природный антисмысловой транскрипт BG724147 гена SCN1A. Хотя настоящее изобретение было иллюстрировано и описано посредством ссылок на один или несколько вариантов реализации, эквивалентные изменения и модификации будут очевидны для специалистов после прочтения и понимания настоящего описания и прилагаемых чертежей. Кроме того, хотя конкретные свойства согласно настоящему изобретению могут раскрываться только в одном из нескольких вариантов реализации, такие свойства могут быть скомбинированы с одним или несколькими другими свойствами других вариантов реализации, что может быть необходимо и полезно для любого или частного случая применения.
[00324] Реферат настоящего изобретения позволит читателю быстро понять техническую суть изобретения. Подразумевается, что он не будет использован для целей толкования или ограничения сути и объема приведенной ниже формулы изобретения.
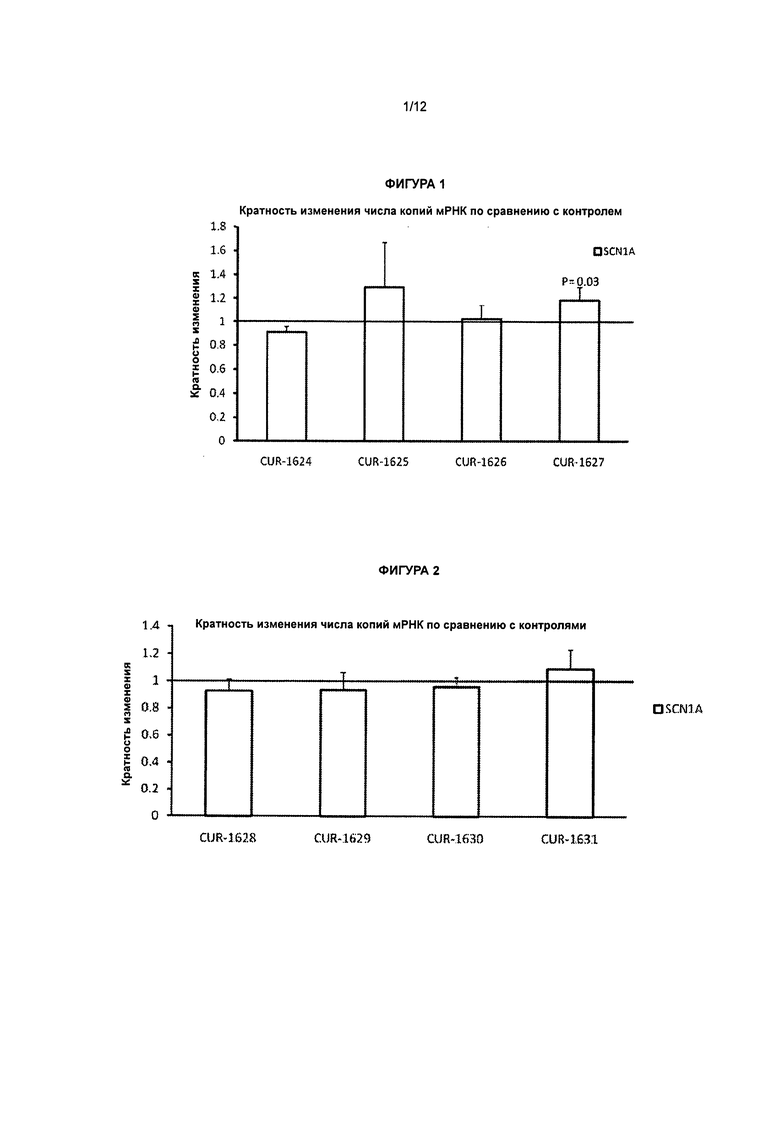
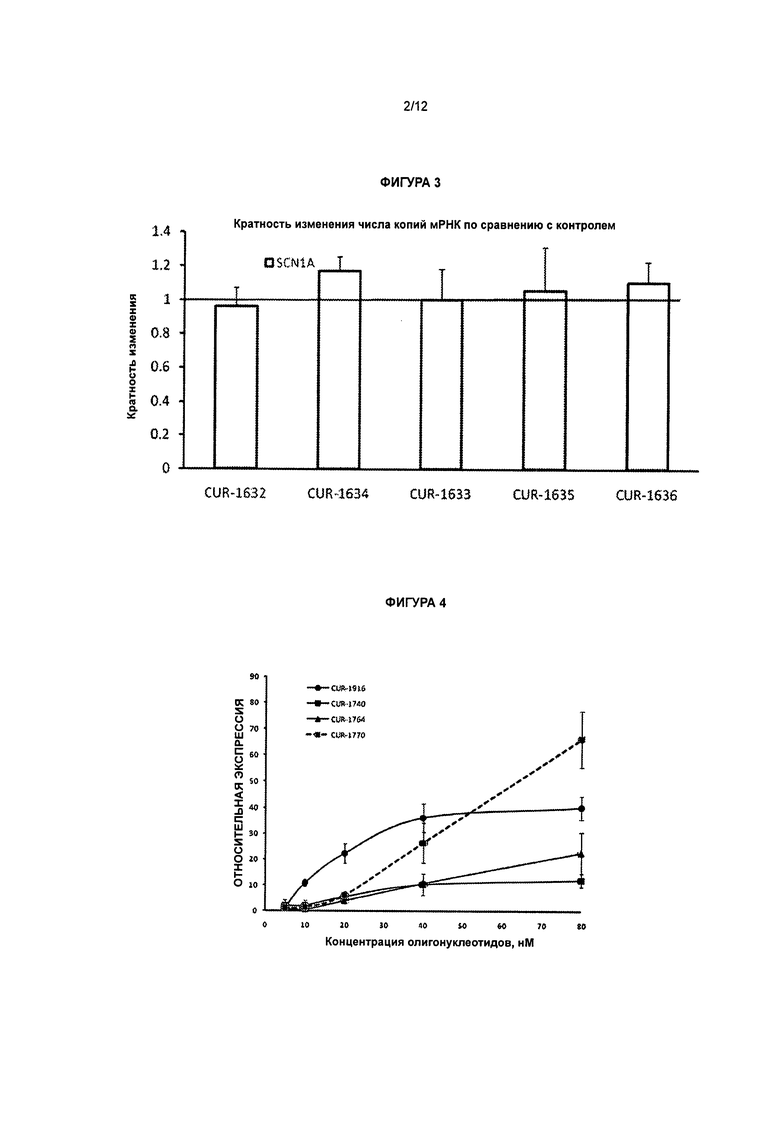
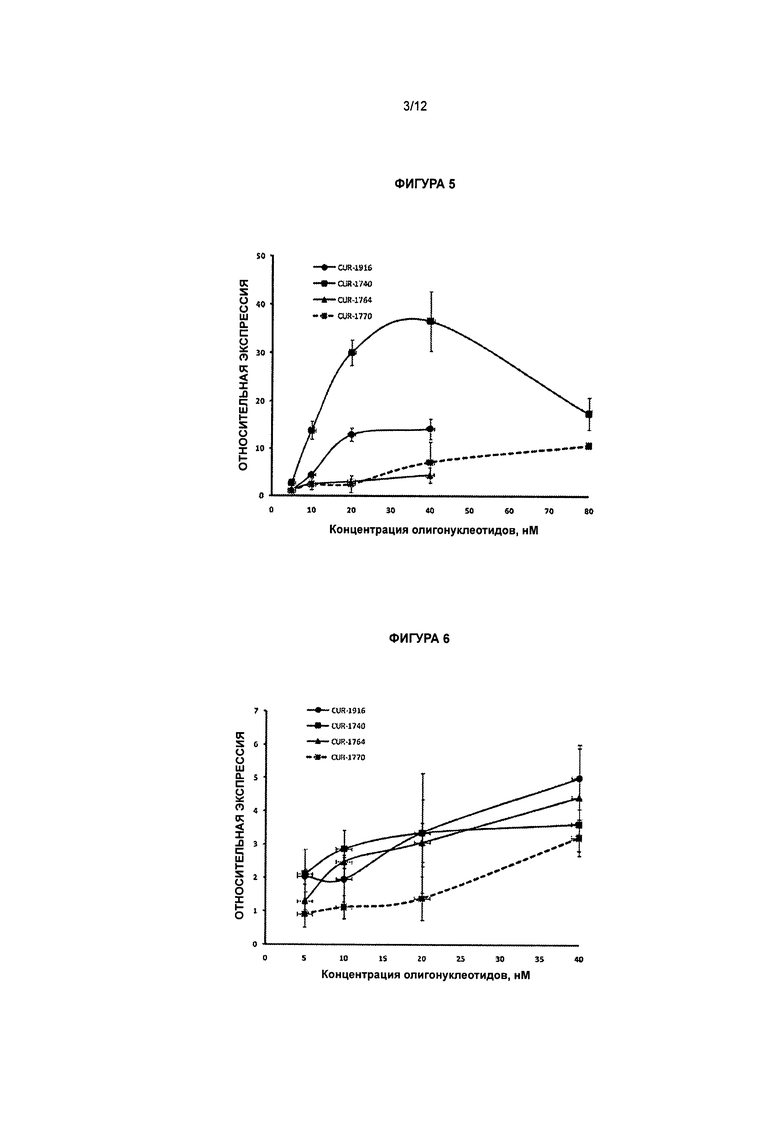
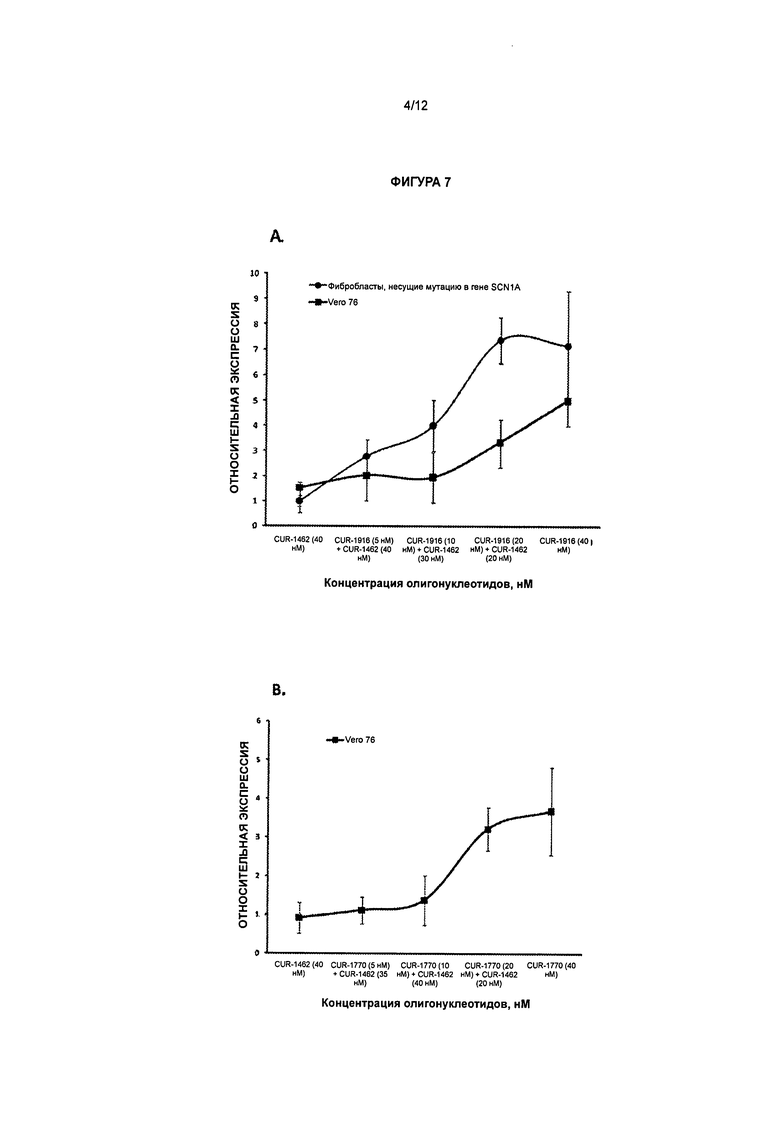
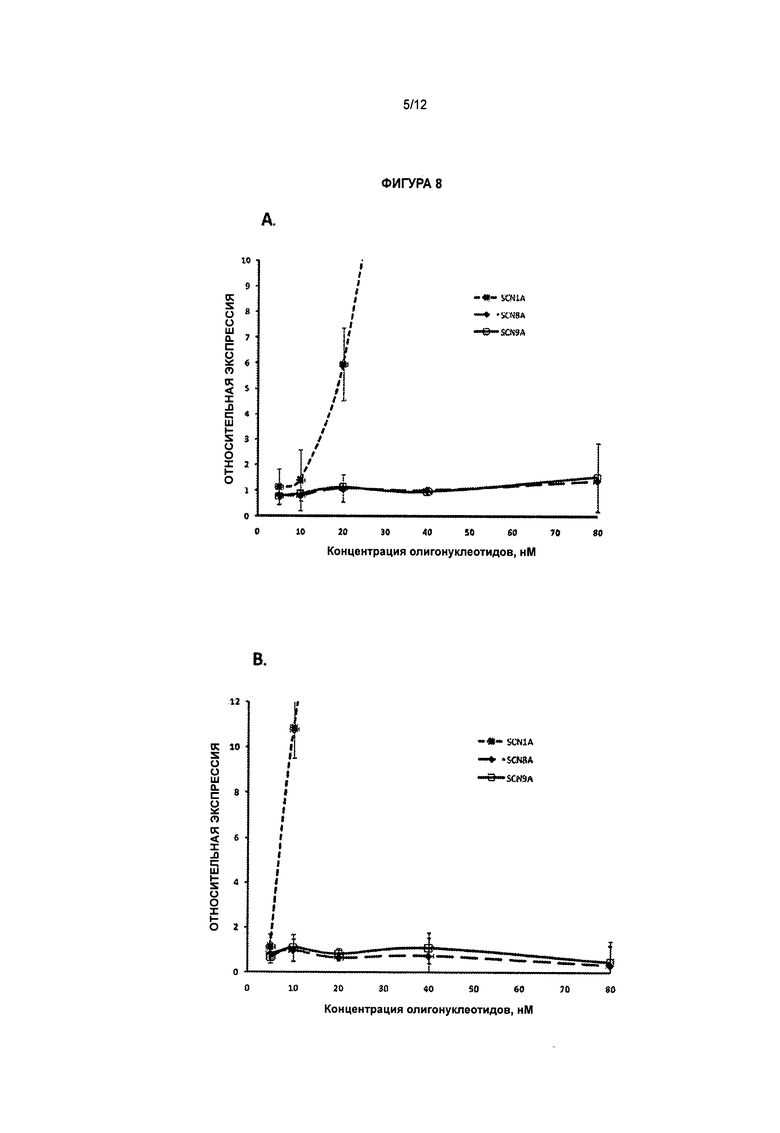
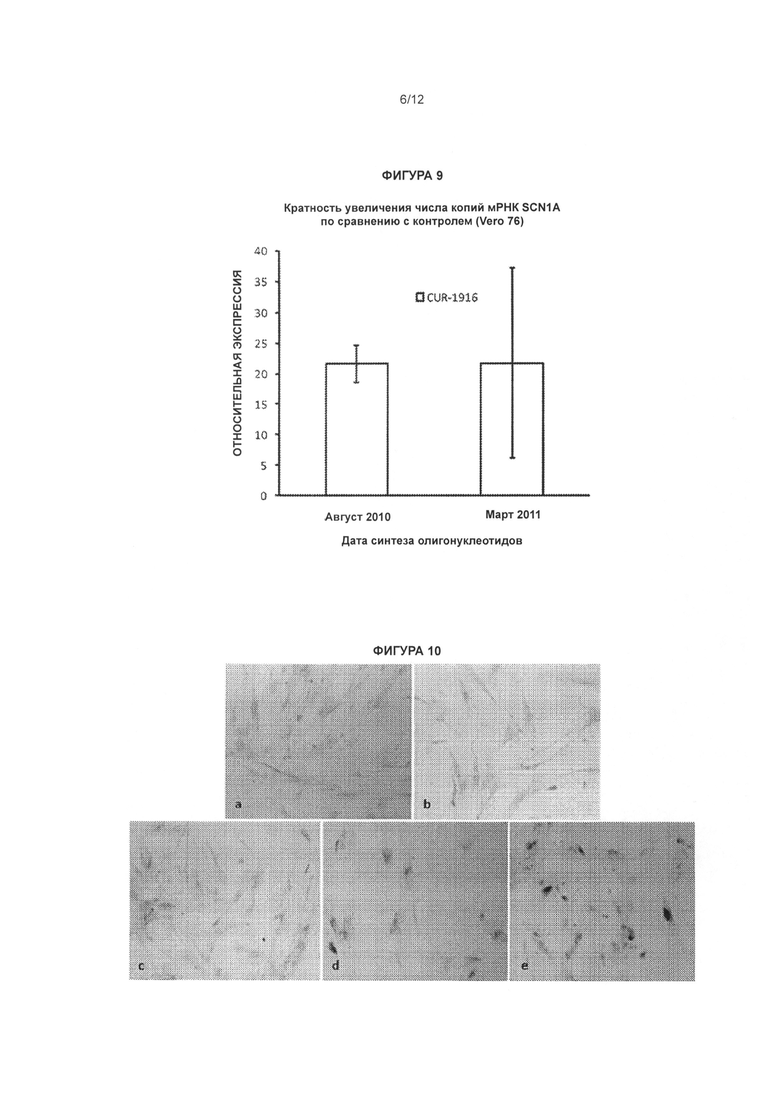
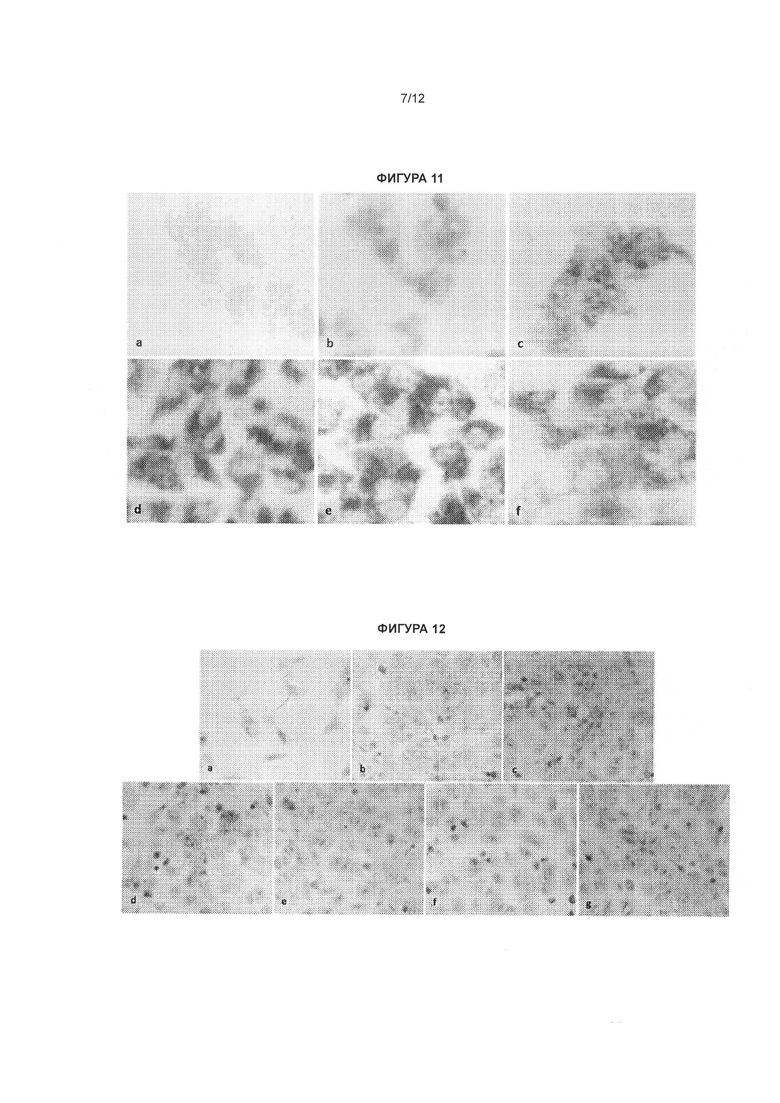
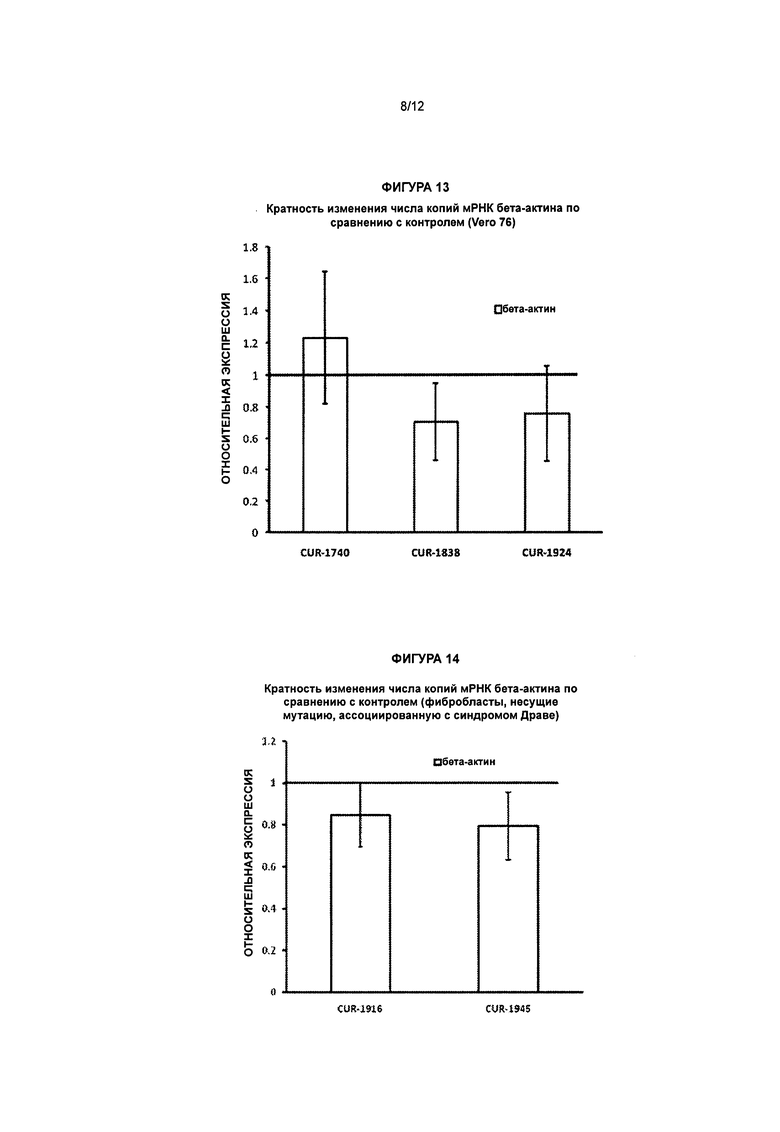
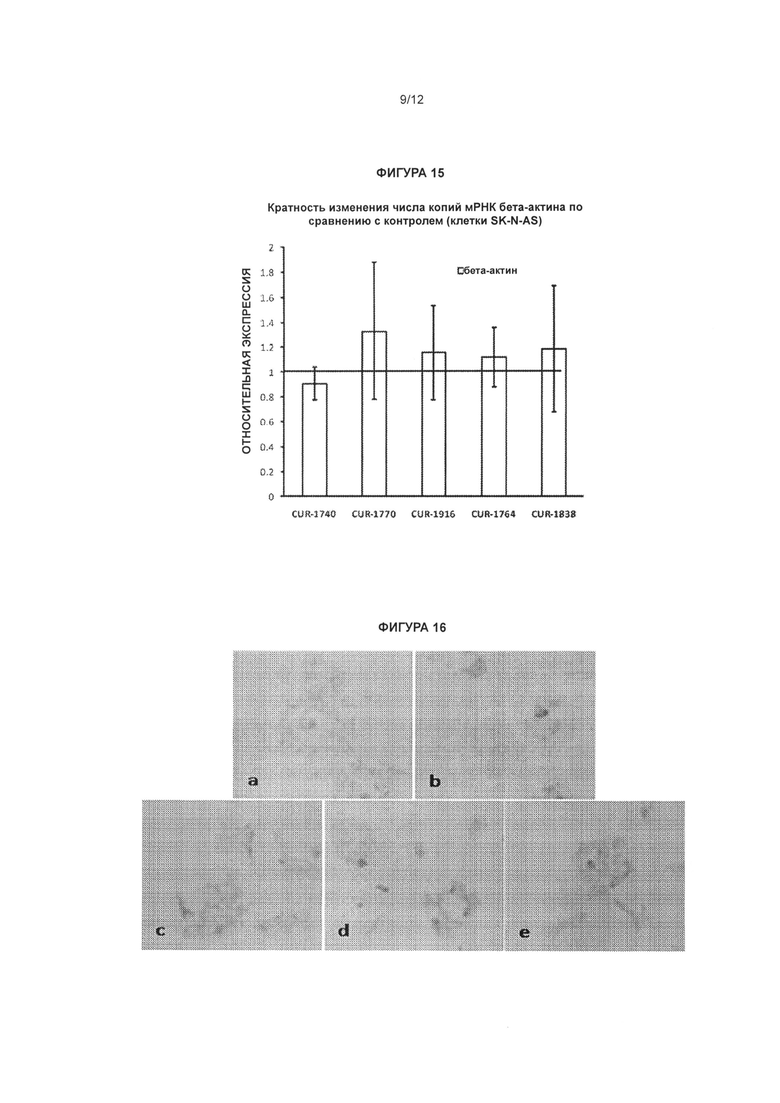
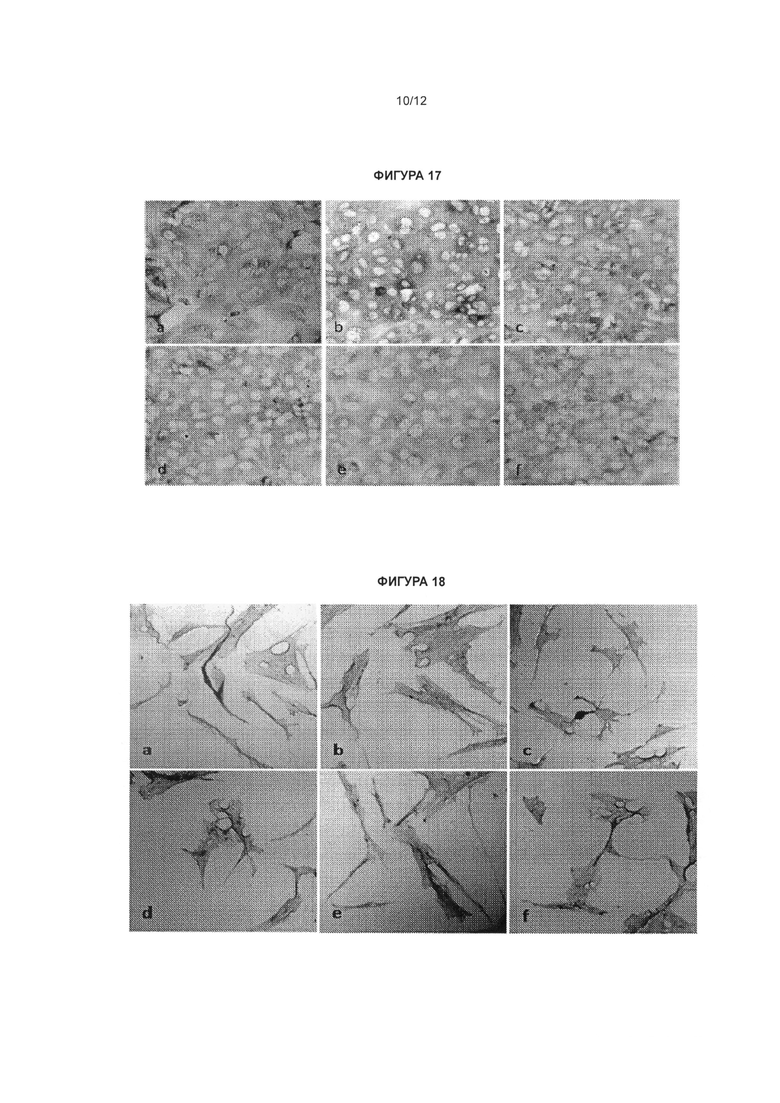
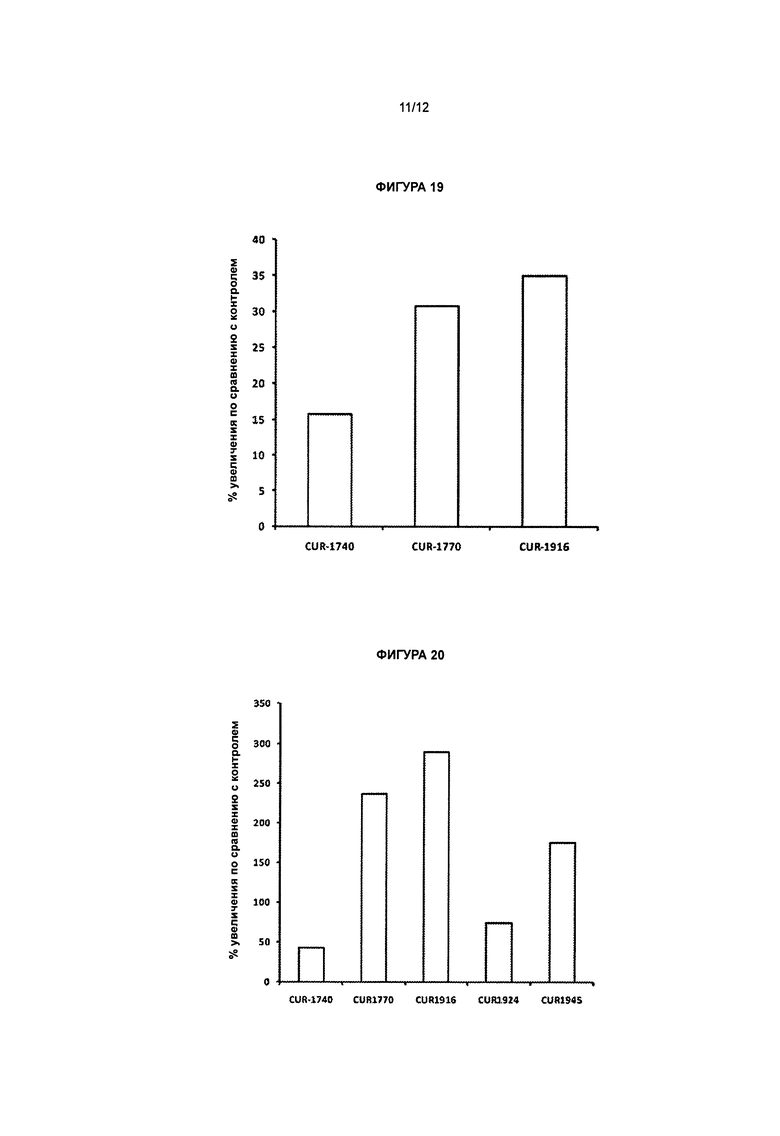
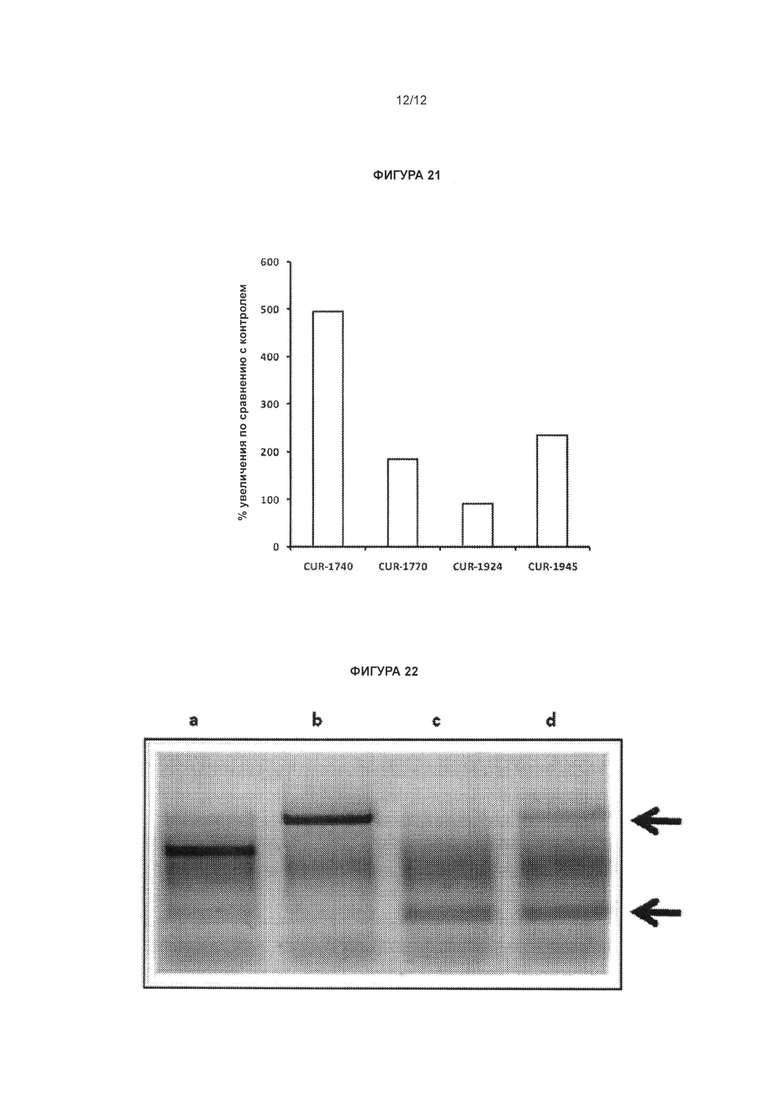
































































Изобретение относится к биохимии. Описаны антисмысловые олигонуклеотиды, которые увеличивают экспрессию указанного гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) in vivo или in vitro по сравнению с нормальным контролем. Олигонуклеотиды представляют собой фосфотиоатные аналоги длиной от 11 до 30 нуклеотидов, содержащие по меньшей мере одну модификацию, выбранную из: модифицированного остатка сахара; модифицированной межнуклеотидной связи; модифицированного нуклеотида и их комбинаций; при этом указанные олигонуклеотиды представляют собой соединения, которые специфически гибридизуются с комплементарной областью природного антисмыслового олигонуклеотида гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA). Изобретение также относится к применению указанных олигонуклеотидов для лечения заболеваний и расстройств, ассоциированных с экспрессией гена SCNA. 6 н. и 27 з.п. ф-лы, 22 ил., 2 табл., 21 пр.
1. Способ увеличения экспрессии полинуклеотида, кодирующего альфа-субъединицу потенциалзависимого натриевого канала (SCNA) в биологической системе, включающий: осуществление контакта указанной системы с по меньшей мере одним олигонуклеотидом, представляющим собой фосфотиоатный аналог, длиной от 11 до 30 нуклеотидов, при этом указанный по меньшей мере один олигонуклеотид, представляющий собой фосфотиоатный аналог, имеет по меньшей мере 90% комплементарность с 11-30 последовательными нуклеотидами в природной антисмысловой последовательности для полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA), что приводит к увеличению экспрессии полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA).
2. Способ увеличения экспрессии полинуклеотида, кодирующего альфа-субъединицу потенциалзависимого натриевого канала типа I (SCN1A) в биологической системе по п. 1, включающий: осуществление контакта указанной биологической системы с по меньшей мере одним олигонуклеотидом, представляющим собой фосфотиоатный аналог, длиной от 11 до 30 нуклеотидов, при этом указанный по меньшей мере один олигонуклеотид, представляющий собой фосфотиоатный аналог, по меньшей мере на 90% комплементарен 11-30 последовательным нуклеотидам в полинуклеотиде, содержащем от 11 до 30 последовательных нуклеотидов в пределах нуклеотидов природного антисмыслового транскрипта: 1-1123 последовательности SEQ ID NO: 12 и 1-2352 последовательности SEQ ID NO: 13, 1-267 последовательности SEQ ID NO: 14, 1-1080 последовательности SEQ ID NO: 15, 1-173 последовательности SEQ ID NO: 16, 1-618 последовательности SEQ ID NO: 17, 1-871 последовательности SEQ ID NO: 18, 1-304 последовательности SEQ ID NO: 19, 1-293 последовательности SEQ ID NO: 20, 1-892 последовательности SEQ ID NO: 21, 1-260 последовательности SEQ ID NO: 22, 1-982 последовательности SEQ ID NO: 23, 1-906 последовательности SEQ ID NO: 24, 1-476 последовательности SEQ ID NO: 25, 1-285 последовательности SEQ ID NO: 26, 1-162 последовательности SEQ ID NO: 27 и 1-94 последовательности SEQ ID NO: 28, что приводит к увеличению экспрессии полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала типа I (SCN1A).
3. Способ увеличения экспрессии полинуклеотида, кодирующего альфа-субъединицу потенциалзависимого натриевого канала (SCNA) в биологической системе, включающий: осуществление контакта указанной системы с по меньшей мере одним антисмысловым олигонуклеотидом, представляющим собой фосфотиоатный аналог и имеющим длину от 11 до 30 нуклеотидов, который нацелен на и специфично гибридизуется с участком природного антисмыслового олигонуклеотида для полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA), что приводит к увеличению экспрессии полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA).
4. Способ по п. 3, отличающийся тем, что экспрессия полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала типа I (SCN1A) увеличивается in vivo или in vitro по сравнению с контролем.
5. Способ по п. 3, отличающийся тем, что указанный по меньшей мере один антисмысловой олигонуклеотид, представляющий собой фосфотиоатный аналог, нацелен на природный антисмысловой олигонуклеотид для полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала типа I (SCN1A), при этом указанный природный антисмысловой олигонуклеотид выбран из группы, состоящей из 11-30 последовательных нуклеотидов в пределах нуклеотидов природных антисмысловых транскриптов: 1-1123 последовательности SEQ ID NO: 12 и 1-2352 последовательности SEQ ID NO: 13, 1-267 последовательности SEQ ID NO: 14, 1-1080 последовательности SEQ ID NO: 15, 1-173 последовательности SEQ ID NO: 16, 1-618 последовательности SEQ ID NO: 17, 1-871 последовательности SEQ ID NO: 18, 1-304 последовательности SEQ ID NO: 19, 1-293 последовательности SEQ ID NO: 20, 1-892 последовательности SEQ ID NO: 21, 1-260 последовательности SEQ ID NO: 22, 1-982 последовательности SEQ ID NO: 23, 1-906 последовательности SEQ ID NO: 24, 1-476 последовательности SEQ ID NO: 25, 1-285 последовательности SEQ ID NO: 26, 1-162 последовательности SEQ ID NO: 27 и 1-94 последовательности SEQ ID NO: 28.
6. Способ по п. 3, отличающийся тем, что указанный по меньшей мере один олигонуклеотид, представляющий собой фосфотиоатный аналог, нацелен на нуклеотидную последовательность, содержащую кодирующие и/или некодирующие нуклеотидные последовательности полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала типа I (SCN1A).
7. Способ по п. 3, отличающийся тем, что указанный по меньшей мере один олигонуклеотид, представляющий собой фосфотиоатный аналог, нацелен на перекрывающиеся или неперекрывающиеся последовательности полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала типа I (SCN1A).
8. Способ по п. 3, отличающийся тем, что указанный по меньшей мере один олигонуклеотид, представляющий собой фосфотиоатный аналог, содержит одну или более модификаций, выбранных из: по меньшей мере одного модифицированного остатка сахара, по меньшей мере одной модифицированной межнуклеозидной связи, по меньшей мере одного модифицированного нуклеотида и их комбинаций.
9. Способ по п. 8, отличающийся тем, что указанные одна или более модификаций включают по меньшей мере один модифицированный остаток сахара, выбранный из: 2′-O-метоксиэтил-модифицированного остатка сахара, 2′-метокси-модифицированного остатка сахара, 2′-O-алкил-модифицированного остатка сахара, бициклического остатка сахара и их комбинаций.
10. Способ по п. 8, отличающийся тем, что указанные одна или более модификаций включают по меньшей мере одну модифицированную межнуклеозидную связь, выбранную из: фосфотиоата, 2′-O-метоксиэтила (МОЭ), 2′-фтор, алкилфосфоната, фосфодитиоата, алкилфосфонотиоата, фосфорамидата, карбамата, карбоната, триэфирфосфата, ацетамидата, карбоксиметилового эфира и их комбинаций.
11. Способ по п. 8, отличающийся тем, что указанные одна или более модификаций включают по меньшей мере один модифицированный нуклеотид, выбранный из: пептидо-нуклеиновой кислоты, закрытой нуклеиновой кислоты, арабинонуклеиновой кислоты (2′-дезокси-2′-фтор-β-D-арабинонуклеиновой кислоты, FANA), их аналога, производного и комбинаций.
12. Способ по п. 1, отличающийся тем, что указанный по меньшей мере один олигонуклеотид, представляющий собой фосфотиоатный аналог, содержит по меньшей мере одну из олигонуклеотидных последовательностей, соответствующих SEQ ID NOS: 29-94.
13. Способ увеличения экспрессии гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) в клетках или тканях млекопитающих in vivo или in vitro, включающий: осуществление контакта указанных клеток или тканей с по меньшей мере одним олигонуклеотидом, представляющим собой фосфотиоатный аналог, длиной приблизительно от 11 до 30 нуклеотидов, специфичным к некодирующим и/или кодирующим последовательностям природной антисмысловой цепи полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA), при этом указанный по меньшей мере один антисмысловой олигонуклеотид, представляющий собой фосфотиоатный аналог, имеет по меньшей мере 90% идентичность последовательности с 11 по 30 последовательных нуклеотидов в пределах по меньшей мере одной нуклеотидной последовательности, соответствующей SEQ ID NOS: 1-11; и увеличение экспрессии гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) в клетках или тканях млекопитающих in vivo или in vitro.
14. Олигонуклеотид, представляющий собой фосфотиоатный аналог, длиной от 11 до 30 нуклеотидов, содержащий по меньшей мере одну модификацию, при этом указанная по меньшей мере одна модификация выбрана из: по меньшей мере одного модифицированного остатка сахара; по меньшей мере одной модифицированной межнуклеотидной связи; по меньшей мере одного модифицированного нуклеотида и их комбинаций; при этом указанный олигонуклеотид представляет собой соединение, которое специфически гибридизуется с комплементарной областью природного антисмыслового олигонуклеотида гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и увеличивает экспрессию указанного гена альфа-субъединицы потенциалзависимого натриевого канала (SCNA) in vivo или in vitro по сравнению с нормальным контролем.
15. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 14, отличающийся тем, что указанный олигонуклеотид имеет длину от 11 до 30 нуклеотидов и указанный природный антисмысловой олигонуклеотид выбран из группы, состоящей из нуклеотидов природного антисмыслового транскрипта: 1-1123 последовательности SEQ ID NO: 12 и 1-2352 последовательности SEQ ID NO: 13, 1-267 последовательности SEQ ID NO: 14, 1-1080 последовательности SEQ ID NO: 15, 1-173 последовательности SEQ ID NO: 16, 1-618 последовательности SEQ ID NO: 17, 1-871 последовательности SEQ ID NO: 18, 1-304 последовательности SEQ ID NO: 19, 1-293 последовательности SEQ ID NO: 20, 1-892 последовательности SEQ ID NO: 21, 1-260 последовательности SEQ ID NO: 22, 1-982 последовательности SEQ ID NO: 23, 1-906 последовательности SEQ ID NO: 24, 1-476 последовательности SEQ ID NO: 25, 1-285 последовательности SEQ ID NO: 26, 1-162 последовательности SEQ ID NO: 27 и 1-94 последовательности SEQ ID NO: 28.
16. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанная по меньшей мере одна модификация включает межнуклеотидную связь, выбранную из группы, состоящей из: фосфотиоата, алкилфосфоната, фосфодитиоата, алкилфосфонотиоата, фосфорамидата, карбамата, карбоната, триэфирфосфата, ацетамидата, карбоксиметилового эфира и их комбинаций.
17. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит по меньшей мере одну фосфотиоатную межнуклеотидную связь.
18. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит остов из фосфотиоатных межнуклеотидных связей.
19. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит по меньшей мере один модифицированный нуклеотид, при этом указанный модифицированный нуклеотид выбран из: пептидо-нуклеиновой кислоты, закрытой нуклеиновой кислоты, их аналога, производного и комбинаций.
20. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит множество модификаций, при этом указанные модификации включают модифицированные нуклеотиды, выбранные из: фосфотиоата, алкилфосфоната, фосфодитиоата, алкилфосфонотиоата, фосфорамидата, карбамата, карбоната, триэфирфосфата, ацетамидата, карбоксиметилового эфира и их комбинаций.
21. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит множество модификаций, при этом указанные модификации включают модифицированные нуклеотиды, выбранные из: пептидо-нуклеиновых кислот, закрытых нуклеиновых кислот, их аналогов, производных и их комбинаций.
22. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит по меньшей мере один модифицированный остаток сахара, выбранный из: 2′-O-метоксиэтил-модифицированного остатка сахара, 2′-метокси-модифицированного остатка сахара, 2′-O-алкил-модифицированного остатка сахара, бициклического остатка сахара и их комбинаций.
23. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит множество модификаций, при этом указанные модификации включают модифицированные остатки сахаров, выбранные из: 2′-O-метоксиэтил-модифицированного остатка сахара, 2′-метокси-модифицированного остатка сахара, 2′-O-алкил-модифицированного остатка сахара, бициклического остатка сахара и их комбинаций.
24. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид имеет длину по меньшей мере примерно от 11 до 30 нуклеотидов и гибридизуется с природной антисмысловой цепью полинуклеотида, кодирующего альфа-субъединицу потенциалзависимого натриевого канала типа I (SCN1A), при этом указанный олигонуклеотид увеличивает экспрессию полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала типа I (SCN1A).
25. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид гибридизуется с и увеличивает экспрессию по меньшей мере одного полинуклеотида, кодирующего альфа-субъединицу потенциалзависимого натриевого канала типа I (SCN1A), in vivo или in vitro по сравнению с нормальным контролем.
26. Олигонуклеотид, представляющий собой фосфотиоатный аналог, по п. 15, отличающийся тем, что указанный олигонуклеотид содержит последовательности, соответствующие SEQ ID NOS: 29-94.
27. Фармацевтическая композиция для предотвращения и/или лечения заболевания или состояния, ассоциированного с по меньшей мере одним полинуклеотидом, кодирующим альфа-субъединицу потенциалзависимого натриевого канала (SCNA), и/или по меньшей мере одним продуктом, который он кодирует, при этом указанная композиция содержит эффективное количество одного или более олигонуклеотидов, представляющих собой фосфотиоатный аналог, длиной от 11 до 30 нуклеотидов, специфичных к одному или более полинуклеотидам альфа-субъединицы потенциалзависимого натриевого канала (SCNA), по п. 14 и фармацевтически приемлемое вспомогательное вещество.
28. Фармацевтическая композиция по п. 27, отличающаяся тем, что указанные олигонуклеотиды, представляющие собой фосфотиоатные аналоги, имеют по меньшей мере примерно 40% идентичность последовательности по сравнению с любой из нуклеотидных последовательностей, соответствующих SEQ ID NOS: 29-94.
29. Фармацевтическая композиция по п. 27, отличающаяся тем, что указанные олигонуклеотиды, представляющие собой фосфотиоатные аналоги, содержат нуклеотидные последовательности, соответствующие SEQ ID NOS: 29-94.
30. Фармацевтическая композиция по п. 29, отличающаяся тем, что олигонуклеотиды, соответствующие SEQ ID NOS: 29-94, содержат одну или более модификаций или замен.
31. Фармацевтическая композиция по п. 30, отличающаяся тем, что указанные одна или более модификаций выбраны из: фосфотиоата, метилфосфоната, пептидо-нуклеиновой кислоты, закрытой нуклеиновой кислоты и их комбинаций.
32. Способ предотвращения или лечения заболевания, ассоциированного с по меньшей мере одним полинуклеотидом, кодирующим альфа-субъединицу потенциалзависимого натриевого канала (SCNA), и/или по меньшей мере одним продуктом, который он кодирует, включающий: введение пациенту терапевтически эффективной дозы по меньшей мере одного олигонуклеотида, представляющего собой фосфотиоатный аналог и имеющего длину от 11 до 30 нуклеотидов, который связывается с природной антисмысловой последовательностью указанного по меньшей мере одного полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и увеличивает экспрессию указанного по меньшей мере одного полинуклеотида альфа-субъединицы потенциалзависимого натриевого канала (SCNA), что приводит к предотвращению или лечению заболевания, ассоциированного с указанным по меньшей мере одним полинуклеотидом альфа-субъединицы потенциалзависимого натриевого канала (SCNA) и/или по меньшей мере одним кодируемым им продуктом.
33. Способ по п. 32, отличающийся тем, что заболевание, ассоциированное с указанным по меньшей мере одним полинуклеотидом альфа-субъединицы потенциалзависимого натриевого канала (SCNA), выбрано из: заболевания или расстройства, ассоциированного с аномальной функцией и/или экспрессией гена SCNA; неврологического заболевания или расстройства, конвульсий, боли (включая хроническую боль), нарушенной электрической возбудимости, сопряженной с дисфункцией натриевого канала; заболевания или расстройства, ассоциированного с дисфункцией натриевого канала; заболевания или расстройства, ассоциированного с неправильной регуляцией активности альфа-субъединицы потенциалзависимого натриевого канала (например, паралича, гиперкалиемического периодического паралича, врожденной парамиотонии, обостряемой действием калия миотонии, синдрома 3 удлиненного интервала QT, заболевания двигательных замыкательных пластинок, атаксии и т.д.); заболевания желудочно-кишечного тракта, возникшего вследствие дисфункции энтеральной нервной системы (например, колита, илеита, синдрома воспаленного кишечника и т.д.); сердечно-сосудистого заболевания или расстройства (например, гипертензии, застойной сердечной недостаточности и т.д.); заболевания или расстройства урогенитального тракта с вовлечением симпатической и парасимпатической иннервации (например, доброкачественной гиперплазии предстательной железы, импотенции); заболевания или расстройства, ассоциированного с нервно-мышечной системой (например, мышечной дистрофии, рассеянного склероза, эпилепсии, аутизма, мигрени (например, спорадических и семейных гемиплегических мигреней и т.д.), тяжелой миоклонической эпилепсии младенчества (SMEI), генерализованной эпилепсии с фебрильными припадками плюс (ГЭФП+; GEFS+) и т.д.), и ассоциированной с геном SCNA эпилепсии.
| ВИРУСНЫЕ ВЕКТОРЫ И ИХ ПРИМЕНЕНИЕ В ГЕННОЙ ТЕРАПИИ | 1994 |
|
RU2233333C2 |
| RU 2008145510 A, 27.05.2010 | |||
| US 20040203024 A1, 14.10.2004. | |||

