Область применения изобретения
Генетически модифицированные мыши, которые содержат вариабельную последовательность лямбда (Vλ) легкой цепи мыши или человека, функционально связанную с константной областью легкой цепи мыши или человека (λ или каппа (κ)). Генетически модифицированные мыши, которые экспрессируют эпитоп-связывающие белки, которые содержат легкую цепь иммуноглобулина, содержащую вариабельный домен, полученный из вариабельного лямбда (hVλ) генного сегмента человека, соединительного лямбда (hJλ) генного сегмента человека, и константный домен легкой цепи (CL) мыши. Генетически модифицированные мыши, содержащие вариабельную последовательность нуклеиновой кислоты неперегруппированной легкой цепи лямбда (λ) иммуноглобулина в эндогенном локусе легкой цепи мыши. Мыши, допускающие перегруппировку и экспрессию химерной легкой цепи λ человека/ CL мыши из эндогенного локуса легкой цепи, которые содержат замену всех генных сегментов вариабельной области эндогенной легкой цепи мыши на один или несколько генных сегментов hVλ и один или несколько генных сегментов hJλ. Соматически видоизмененные антитела, содержащие домены hVλ и домены CL мыши.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
Мыши, которые экспрессируют антитела, являющиеся полностью человеческими, или частично человеческими и частично мышиными, известны в данной области. Например, описаны трансгенные мыши, которые экспрессируют полностью человеческие антитела из трансгенов, содержащих гены вариабельной области легкой и тяжелой цепей иммуноглобулина человека. Также известны генетически модифицированные мыши, которые содержат замену эндогенных генных сегментов вариабельной области тяжелой цепи (HCVR) и генных сегментов вариабельной области легкой цепи (LCVR) каппа (κ) мыши на генные сегменты HCVR и LCVR человека, и которые продуцируют химерные антитела с химерной каппа-цепью человек/мышь.
Легкие цепи антител кодируются одним из двух раздельных локусов: каппа (κ) и лямбда (λ). Легкие цепи антител мыши главным образом являются κ-типа. Соотношение использования κ и λ легкой цепи у человека составляет приблизительно 60:40, в то время как у мышей это соотношение составляет приблизительно 95:5. Несимметричное использование легких цепей κ у мыши, по имеющимся данным, поддерживается у генетически модифицированных мышей, способных экспрессировать полностью или частично человеческие антитела. Таким образом, мыши, которые продуцируют полностью или частично человеческие антитела, по-видимому, ограничены в использовании вариабельных лямбда.
В данной области существует необходимость в получении лямбда вариабельных областей, либо мыши, либо человека, для использования при получении эпитоп-связывающих белков. В данной области существует необходимость в мышах, которые продуцируют полностью или частично человеческие антитела, где мыши проявляют повышенное использование вариабельных областей лямбда (Vλ).
В данной области существует необходимость в мышах, которые продуцируют полностью или частично человеческие антитела, где мыши проявляют повышенное использование вариабельных областей лямбда (Vλ).
Краткое описание изобретения
Изобретение относится к генетически модифицированным мышам, эмбрионам, клеткам, тканям, а также конструкциям нуклеиновых кислот для видоизменения мышей, и к способам и композициям для получения и использования их. Изобретение относится к мышам и клеткам, которые продуцируют вариабельные области лямбда (λ) (человека или не относящиеся к человеческому роду) в связи с легкой цепью каппа (κ). Также изобретение относится к мышам и клеткам, которые продуцируют вариабельные области λ человека в связи с легкой цепью κ или λ, например, из эндогенного локуса легкой цепи мыши. Также изобретение относится к способам получения антител, которые содержат вариабельные области лямбда. Также изобретение относится к способам отбора тяжелых цепей, которые экспрессируются с родственными вариабельными областями лямбда.
Химерные и человеческие антигенсвязывающие белки (например, антитела), и нуклеиновые кислоты, кодирующие их, обеспечены при условии, если содержат соматически видоизмененные вариабельные области, в том числе антитела, которые имеют легкие цепи, содержащие вариабельные домены, полученные из генного сегмента Vλ и Jλ человека, слитого с константным доменом легкой цепи мыши.
В одном из аспектов, изобретение относится к мыши, которая экспрессирует последовательность вариабельной области λ человека на легкой цепи, которая включает константную область мыши. В одном из аспектов изобретение относится к мыши, которая экспрессирует последовательность вариабельной области λ человека на легкой цепи, которая содержит константную область κ. В одном из аспектов изобретение относится к мыши, которая экспрессирует из эндогенного локуса легкой цепи мыши легкую цепь, которая содержит последовательность вариабельной области λ человека. В одном из аспектов изобретение относится к мыши, которая содержит перегруппированный ген легкой цепи, который содержит последовательность вариабельной области λ человека, связанную с последовательностью константной области мыши; в одном из вариантов осуществления последовательность константной области мыши представляет собой константную последовательность λ; в одном из вариантов осуществления последовательность константной области мыши представляет собой константную последовательность κ.
В одном из аспектов изобретение относится к генетически модифицированной мыши, где мышь содержит неперегруппированный генный сегмент легкой цепи λ человека (hVλ) и J-сегмент λ человека (hJλ). В одном из вариантов осуществления неперегруппированный hVλ и hJλ находятся в локусе легкой цепи мыши. В одном из вариантов осуществления неперегруппированный hVλ и неперегруппированный hJλ находятся в трансгене и функционально связаны с последовательностью константной области человека или мыши. В одном из вариантов осуществления неперегруппированный hVλ и неперегруппированный hJλ находятся на эписоме. В одном из вариантов осуществления мышь способна продуцировать иммуноглобулин, который содержит легкую цепь, полученную из последовательности неперегруппированного hVλ и последовательности hJλ, и последовательности нуклеиновой кислоты константной области (CL) легкой цепи мыши. Таким образом изобретение относится к способам и композициям для получения и использования генетически модифицированных мышей. Изобретение относится к антителам, которые содержат (а) вариабельный домен тяжелой цепи человека (hVH), слитый с константной областью тяжелой цепи мыши, и (b) Vλ человека, слитый с доменом CL мыши; в том числе, к антителам, в которых один или несколько вариабельных доменов соматически видоизменены, например, во время селекции антитела или иммунных клеток в мыши по данному изобретению. В одном из вариантов осуществления данные неперегруппированный hVλ и неперегруппированный hJλ функционально связаны с константной областью κ (Сκ) человека или мыши. В одном из вариантов осуществления данные неперегруппированный hVλ и неперегруппированный hJλ функционально связаны с константной областью λ (Сλ) человека или мыши.
В одном из аспектов изобретение относится к мыши, содержащей в своей зародышевой линии, в эндогенном локусе легкой цепи мыши, последовательность вариабельной области легкой цепи λ человека, где последовательность вариабельной области лямбда человека экспрессируется в легкой цепи, которая содержит последовательность гена константной области иммуноглобулина мыши.
В одном из вариантов осуществления эндогенный локус легкой цепи мыши представляет собой локус λ. В одном из вариантов осуществления эндогенный локус легкой цепи мыши представляет собой локус κ.
В одном из вариантов осуществления мышь лишена эндогенной вариабельной последовательности легкой цепи в эндогенном локусе легкой цепи мыши.
В одном из вариантов осуществления все, или практически все, эндогенные генные сегменты вариабельной области легкой цепи мыши заменены одним или несколькими генными сегментами вариабельной области λ человека.
В одном из вариантов осуществления последовательность вариабельной области легкой цепи λ человека содержит последовательность Jλ человека. В одном из вариантов осуществления, последовательность Jλ человека выбрана из группы, состоящей из Jλ1, Jλ2, Jλ3, Jλ7, и их комбинации.
В одном из вариантов осуществления последовательность вариабельной области легкой цепи λ человека содержит фрагмент кластера А локуса легкой цепи человека. В конкретном варианте осуществления, фрагмент кластера А локуса легкой цепи λ человека простирается от hVλ3-27 через hVλ3-1 включительно.
В одном из вариантов осуществления, последовательность вариабельной области легкой цепи λ человека содержит фрагмент кластера В локуса легкой цепи человека. В конкретном варианте осуществления, фрагмент кластера В локуса легкой цепи λ человека простирается от hVλ5-52 через hVλ1-40 включительно.
В одном из вариантов осуществления последовательность вариабельной области легкой цепи λ человека содержит геномный фрагмент кластера А и геномный фрагмент кластера В. В одном из вариантов осуществления последовательность вариабельной области легкой цепи λ человека содержит по меньшей мере один генный сегмент кластера А и по меньшей мере один генный сегмент кластера В.
В одном из вариантов осуществления более чем 10% интактного набора легкой цепи мыши получено из по меньшей мере двух генных сегментов hVλ, отобранных из 2-8, 2-23, 1-40, 5-45, и 9-49. В одном из вариантов осуществления более чем 20% интактного набора легкой цепи мыши получено из по меньшей мере трех генных сегментов hVλ, выбранных из 2-8, 2-23, 1-40, 5-45, и 9-49. В одном из вариантов осуществления более чем 30% интактного набора легкой цепи мыши получено из по меньшей мере, трех генных сегментов hVλ, выбранных из 2-8, 2-23, 1-40, 5-45, и 9-49.
В одном из аспектов изобретение относится к мыши, которая экспрессирует легкую цепь иммуноглобулина, которая содержит вариабельную последовательность λ человека, слитую с константной областью мыши, где мышь демонстрирует соотношение использования κ к использованию λ приблизительно 1:1.
В одном из вариантов осуществления легкая цепь иммуноглобулина экспрессируется из эндогенного локуса легкой цепи мыши.
В одном из аспектов изобретение относится к мыши, которая содержит последовательность вариабельной области легкой цепи λ (Vλ) и по меньшей мере одну J-последовательность (J), прилегающую к последовательности константной области легкой цепи κ мыши.
В одном из вариантов осуществления у мыши отсутствует функциональный генный сегмент Vκ мыши и/или генный сегмент Jκ мыши.
В одном из вариантов осуществления Vλ представляет собой Vλ человека (hVλ), и J представляет собой Jλ человека (hJλ). В одном из вариантов осуществления данные hVλ и hJλ представляют собой неперегруппированные генные сегменты.
В одном из вариантов осуществления мышь содержит совокупность неперегруппированных генных сегментов hVλ и по меньшей мере один генный сегмент hJλ. В конкретном варианте осуществления совокупность неперегруппированных генных сегментов hVλ составляют по меньшей мере 12 генных сегментов, по меньшей мере 28 генных сегментов или по меньшей мере 40 генных сегментов.
В одном из вариантов осуществления по меньшей мере один генный сегмент hJλ выбран из группы, имеющей в своем составе Jλ1, Jλ2, Jλ3, Jλ7, и их комбинации.
В одном из вариантов осуществления эндогенный локус легкой цепи λ мыши делетирован полностью или частично.
В одном из вариантов осуществления последовательность константной области легкой цепи κ мыши находится в эндогенном локусе легкой цепи κ мыши.
В одном из вариантов осуществления, приблизительно от 10% приблизительно до 45% В-клеток мыши экспрессируют антитело, которое содержит легкую цепь, содержащую вариабельный домен легкой цепи λ (Vλ) человека и константный домен легкой цепи κ (Сκ) мыши.
В одном из вариантов осуществления вариабельный домен λ человека получен из перегруппированной последовательности hVλ/hJλ, выбранной из группы, имеющей в своем составе 3-1/1, 3-1/7, 4-3/1, 4-3/7, 2-8/1, 3-9/1, 3-10/1, 3-10/3, 3-10/7, 2-14/1, 3-19/1, 2-23/1, 3-25/1, 1-40/1, 1-40/2, 1-40/3, 1-40/7, 7-43/1, 7-43/3, 1-44/1, 1-44/7, 5-45/1, 5-45/2, 5-45/7, 7-46/1, 7-46/2, 7-46/7, 9-49/1, 9-49/2, 9-49/7 и 1-51/1.
В одном из вариантов осуществления мышь дополнительно содержит межгенную область Vκ-Jκ человека из локуса легкой цепи κ человека, где межгенная область Vκ-Jκ человека сопряжена с последовательностью Vλ и последовательностью J. В конкретном варианте осуществления межгенная область Vκ-Jκ человека расположена между последовательностью Vλ и J-последовательностью.
В одном из аспектов изобретение относится к мыши, которая содержит (а) по меньшей мере от 12 до по меньшей мере 40 неперегруппированных генных сегментов вариабельной области легкой цепи λ человека и, по меньшей мере, один генный сегмент Jλ человека в эндогенном локусе легкой цепи мыши; (b) межгенную область Vκ-Jκ человека, расположенную между по меньшей мере от 12 до, по меньшей мере 40 генными сегментами вариабельной области легкой цепи человека, и по меньшей мере одной последовательностью Jλ человека; где мышь продуцирует антитело, содержащее легкую цепь, содержащую домен Vλ человека и домен Cκ мыши.
В одном из аспектов изобретение относится к мыши, которая экспрессирует антитело, содержащее легкую цепь, которая содержит вариабельную последовательность λ и константную последовательность κ.
В одном из вариантов осуществления мышь демонстрирует использование κ и использование λ в соотношении приблизительно 1:1.
В одном из вариантов осуществления популяция незрелых В-клеток, полученная из костного мозга мыши, демонстрирует использование κ и использование λ в соотношении приблизительно 1:1.
В одном из аспектов изобретение относится к генетически модифицированной мыши, где мышь содержит неперегруппированные генные сегменты Vλ и Jλ иммуноглобулина, функционально связанные с локусом легкой цепи мыши, который содержит ген CL мыши.
В одном из вариантов осуществления генные сегменты Vλ и/или Jλ представляют собой генные сегменты человека. В одном из вариантов генные сегменты Vλ и/или Jλ представляют собой генные сегменты мыши, и CL представляет собой Cκ мыши.
В одном из вариантов осуществления эндогенный локус легкой цепи мыши представляет собой локус легкой цепи κ. В одном из вариантов осуществления эндогенный локус легкой цепи мыши представляет собой локус легкой цепи λ.
В одном из вариантов осуществления неперегруппированные генные сегменты Vλ и Jλ находятся в эндогенном локусе легкой цепи мыши.
В одном из вариантов осуществления неперегруппированные генные сегменты Vλ и Jλ иммуноглобулина находятся в трансгене.
В одном из вариантов осуществления мышь дополнительно содержит замену одного или нескольких генных сегментов V, D, и/или J тяжелой цепи на один или более сегментов генов V, D и/или J человека в эндогенном локусе тяжелой цепи иммуноглобулина мыши.
В одном из вариантов осуществления мышь содержит неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина в эндогенном локусе легкой цепи κ мыши, который содержит ген Сκ мыши.
В одном из вариантов осуществления мышь содержит неперегруппированный генный сегмент вариабельной области легкой цепи λ (Vλ) и J-генный сегмент λ (Jλ) иммуноглобулина человека в эндогенном локусе легкой цепи λ мыши, который содержит ген Cλ мыши.
В одном из вариантов осуществления локус вариабельного гена легкой цепи («локус VL») содержит по меньшей мере один генный сегмент Vλ (hVλ) человека. В одном из вариантов осуществления локус VL содержит по меньшей мере один генный сегмент Jλ (hJλ) человека. В другом варианте осуществления локус VL содержит до четырех генных сегментов hJλ. В одном из вариантов осуществления локус VL содержит сопредельную последовательность, содержащую геномную последовательность λ человека и κ человека.
В одном из вариантов осуществления локус вариабельного гена легкой цепи κ («локус κ») содержит по меньшей мере один генный сегмент Vλ человека (hVλ). В одном из вариантов осуществления локус κ содержит по меньшей мере один генный сегмент Jλ человека (hJλ). В одном из вариантов осуществления локус κ содержит до четырех генных сегментов hJλ. В одном из вариантов осуществления локус κ содержит по меньшей мере один hVλ и по меньшей мере один hJλ, и лишен, или практически лишен, функционального генного сегмента области Vκ, и лишен, или практически лишен, функционального генного сегмента области Jκ. В одном из вариантов осуществления мышь не содержит функционального генного сегмента области Vκ. В одном из вариантов осуществления мышь не содержит функционального генного сегмента области Jκ.
В одном из вариантов осуществления локус вариабельных генов легкой цепи λ («локус λ») содержит по меньшей мере один генный сегмент hVλ. В одном из вариантов осуществления локус λ содержит по меньшей мере один генный сегмент Jλ человека (hJλ). В другом варианте осуществления локус λ содержит до четырех генных сегментов hJλ.
В одном из вариантов осуществления локус VL содержит некоторое количество hVλ. В одном из вариантов осуществления совокупность hVλ выбрана таким образом, чтобы приводить к экспрессии спектра вариабельной области легкой цепи λ, который воспроизводит приблизительно 10%, приблизительно 20%, приблизительно 30%, приблизительно 40%, приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80% или приблизительно 90%, или более от использования Vλ, наблюдаемого у человека. В одном из вариантов осуществления локус VL содержит генные сегменты hVλ 1-40, 1-44, 2-8, 2-14, 3-21, и их комбинацию.
В одном из вариантов осуществления hVλ включают 3-1, 4-3, 2-8, 3-9, 3-10, 2-11 и 3-12. В конкретном варианте осуществления локус VL содержит смежную последовательность локуса легкой цепи λ человека, которая простирается от Vλ3-12 до Vλ3-1. В одном из вариантов осуществления локус VL содержит по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 hVλ. В конкретном варианте осуществления hVλ включают 3-1, 4-3, 2-8, 3-9, 3-10, 2-11 и 3-12. В конкретном варианте осуществления локус VL содержит смежную последовательность локуса λ человека, которая простирается от Vλ3-12 до Vλ3-1. В одном из вариантов осуществления локус VL находится в эндогенном локусе κ. В конкретном варианте осуществления локус VL находится в эндогенном локусе κ и эндогенный локус легкой цепи λ делетирован частично или полностью. В одном из вариантов осуществления локус VL находится в эндогенном локусе λ. В конкретном варианте осуществления локус VL находится в эндогенном локусе λ и эндогенный локус κ делетирован частично или полностью.
В одном из вариантов осуществления локус VL содержит 13-28 или более hVλ. В конкретном варианте осуществления hVλ включают 2-14, 3-16, 2-18, 3-19, 3-21, 3-22, 2-23, 3-25, и 3-27. В конкретном варианте осуществления локус κ содержит смежную последовательность локуса λ человека, которая простирается от Vλ3-27 до Vλ3-1. В одном из вариантов осуществления локус VL находится в эндогенном локусе κ. В конкретном варианте осуществления локус VL находится в эндогенном локусе κ и эндогенный локус легкой цепи λ делетирован частично или полностью. В другом варианте осуществления локус VL находится в эндогенном локусе λ. В конкретном варианте осуществления локус VL находится в эндогенном локусе λ и эндогенный локус κ делетирован частично или полностью.
В одном из вариантов осуществления локус VL содержит 29-40 hVλ. В конкретном варианте осуществления локус κ содержит прилегающую последовательность локуса λ человека, которая простирается от Vλ3-29 до Vλ3-1, и прилегающую последовательность локуса λ человека, которая простирается от Vλ5-52 до Vλ1-40. В конкретном варианте осуществления вся, или практически вся, последовательность между hVλ1-40 и hVλ3-29 в генетически модифицированной мыши состоит по существу из последовательности λ человека длиной приблизительно 959 п.о., встречающейся в природе (например, в человеческой популяции) в 5'-3' направлении генного сегмента hVλ1-40 (в прямом направлении 3'-нетранслируемой области), участок рестрикционного фермента (например, PI-SceI), за которым следует последовательность λ человека в 3'-5' направлении от генного сегмента hVλ3-29, длинной приблизительно 3431 п.о. встречающийся в природе. В одном из вариантов осуществления локус VL находится в эндогенном локусе κ мыши. В конкретном варианте осуществления локус VL находится в эндогенном локусе κ мыши и эндогенный локус легкой цепи λ мыши делетирован частично или полностью. В другом варианте осуществления локус VL находится в эндогенном локусе λ мыши. В конкретном варианте осуществления локус VL находится в эндогенном локусе λ мыши и эндогенный локус κ мыши делетирован частично или полностью.
В одном из вариантов осуществления локус VL содержит по меньшей мере один hJλ. В одном из вариантов осуществления локус VL содержит совокупность hJλ. В одном из вариантов осуществления локус VL содержит по меньшей мере 2, 3, 4, 5, 6, или 7 hJλ. В определенном варианте осуществления локус VL содержит четыре hJλ. В определенном варианте осуществления данные четыре hJλ представляют собой hJλ1, hJλ2, hJλ3, и hJλ7. В одном из вариантов осуществления локус VL представляет собой локус κ. В определенном варианте осуществления локус VL находится в эндогенном локусе κ и эндогенный локус легкой цепи λ делетирован частично или полностью. В одном из вариантов осуществления локус VL содержит один hJλ. В определенном варианте осуществления один hJλ представляет собой hJλ1. В одном из вариантов осуществления локус VL находится в эндогенном локусе κ. В определенном варианте локус VL находится в эндогенном локусе κ и эндогенный локус легкой цепи λ делетирован частично или полностью. В другом варианте осуществления локус VL находится в эндогенном локусе λ. В определенном варианте локус VL находится в эндогенном локусе λ и эндогенный локус легкой цепи κ делетирован частично или полностью.
В одном из вариантов осуществления локус VL содержит по меньшей мере один hVλ, по меньшей мере один hJλ, и ген Сκ мыши. В одном из вариантов осуществления локус VL содержит по меньшей мере один hVλ, по меньшей мере один hJλ, и ген Сλ мыши. В определенном варианте осуществления ген Сλ мыши представляет собой Сλ2. В определенном варианте осуществления ген Сλ мыши по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, 96%, 97%, 98%, или, по меньшей мере на 99% идентичен Сλ2 мыши.
В одном из вариантов осуществления мышь содержит замену в эндогенном локусе κ мыши эндогенных генных сегментов Vκ мыши на один или несколько генных сегментов hVλ, где генные сегменты hVλ функционально связаны с эндогенной областью гена Сκ мыши, так что у мыши перегруппированы генные сегменты Vλ человека, и продуцируется обратная химерная легкая цепь иммуноглобулина, которая содержит домен Vλ человека и Сκ мыши. В одном из вариантов осуществления 90-100% неперегруппированных генных сегментов Vκ мыши заменены на, по меньшей мере, один неперегруппированный генный сегмент hVλ. В определенном варианте осуществления все, или практически все, эндогенные генные сегменты Vκ мыши заменены на, по меньшей мере, один неперегруппированный генный сегмент hVλ. В одном из вариантов осуществления замена составляет по меньшей мере 12, по меньшей мере 28 или по меньшей мере 40 неперегруппированных генных сегментов hVλ. В одном из вариантов осуществления замена происходит, по меньшей мере на 7 функциональных неперегруппированных генных сегментов hVλ, по меньшей мере, на 16 функциональных неперегруппированных генных сегментов hVλ, или, по меньшей мере, на 27 функциональных неперегруппированных генных сегментов hVλ. В одном из вариантов осуществления мышь содержит замену всех генных сегментов Jκ мыши на, по меньшей мере, один неперегруппированный генный сегмент hJλ. В одном из вариантов осуществления по меньшей мере один неперегруппированный генный сегмент hJλ выбран из Jλ1, Jλ2, Jλ3, Jλ4, Jλ5, Jλ6, Jλ7, и их комбинации. В определенном варианте осуществления один или несколько генных сегментов hVλ выбран из генных сегментов hVλ 3-1, 4-3, 2-8, 3-9, 3-10, 2-11, 3-12, 2-14, 3-16, 2-18, 3-19, 3-21, 3-22, 2-23, 3-25, 3-27, 1-40, 7-43, 1-44, 5-45, 7-46, 1-47, 5-48, 9-49, 1-50, 1-51, 5-52 и их комбинации. В определенном варианте осуществления по меньшей мере один неперегруппированный генный сегмент hJλ выбран из Jλ1, Jλ2, Jλ3, Jλ7, и их комбинации.
В одном из вариантов осуществления мышь содержит замену эндогенных генных сегментов Vλ мыши в эндогенном локусе λ мыши на один или несколько генных сегментов Vλ человека в эндогенном локусе λ мыши, где данные генные сегменты hVλ функционально связаны с областью гена Сλ мыши, так что у мыши перегруппированы генные сегменты hVλ, и продуцируется обратная химерная легкая цепь иммуноглобулина, которая содержит домен hVλ и Сλ мыши. В определенном варианте осуществления ген Cλ мыши представляет собой Cλ2. В определенном варианте осуществления ген Сλ мыши, по меньшей мере, на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или, по меньшей мере на 98% идентичен Сλ2 мыши. В одном из вариантов осуществления 90-100% неперегруппированных генных сегментов Vλ мыши заменены на, по меньшей мере, один неперегруппированный генный сегмент hVλ. В определенном варианте осуществления все, или практически все, эндогенные генные сегменты Vλ мыши заменены на, по меньшей мере, один неперегруппированный генный сегмент hVλ. В одном из вариантов осуществления замена составляет по меньшей мере 12, по меньшей мере 28 или по меньшей мере 40 неперегруппированных генных сегментов hVλ. В одном из вариантов осуществления замена происходит по меньшей мере на 7 функциональных неперегруппированных генных сегментов hVλ, по меньшей мере на 16 функциональных неперегруппированных генных сегментов hVλ или по меньшей мере на 27 функциональных неперегруппированных генных сегментов hVλ. В одном из вариантов осуществления мышь содержит замену всех генных сегментов Jλ мыши на по меньшей мере один неперегруппированный генный сегмент hJλ. В одном из вариантов осуществления по меньшей мере один неперегруппированный генный сегмент hJλ выбран из Jλ1, Jλ2, Jλ3, Jλ4, Jλ5, Jλ6, Jλ7, и их комбинации. В определенном варианте осуществления один или несколько генных сегментов hVλ выбран из генных сегментов hVλ 3-1, 4-3, 2-8, 3-9, 3-10, 2-11, 3-12, 2-14, 3-16, 2-18, 3-19, 3-21, 3-22, 2-23, 3-25, 3-27, 1-40, 7-43, 1-44, 5-45, 7-46, 1-47, 5-48, 9-49, 1-50, 1-51, 5-52, и их комбинации. В определенном варианте осуществления по меньшей мере один неперегруппированный генный сегмент hJλ выбран из Jλ1, Jλ2, Jλ3, Jλ7, и их комбинации.
В одном из аспектов изобретение относится к генетически модифицированной мыши, которая содержит последовательность межгенной области Vκ-Jκ человека, расположенную в эндогенном локусе легкой цепи κ мыши.
В одном из вариантов осуществления последовательность межгенной области Vκ-Jκ человека расположена в эндогенном локусе легкой цепи κ мыши, которая содержит генный сегмент hVλ и hJλ, и последовательность межгенной области Vκ-Jκ человека расположена между генными сегментами hVλ и hJλ. В определенном варианте осуществления данные генные сегменты hVλ и hJλ способны к рекомбинации для формирования функционального вариабельного домена легкой цепи λ человека у мыши.
В одном из вариантов осуществления изобретение относится к мыши, которая содержит совокупность генных сегментов hVλ и один или несколько hJλ, и последовательность межгенной области Vκ-Jκ человека расположена, относительно транскрипции, в 5'-3' направлении от самой проксимальной или 3' последовательности hVλ, и в 3'-5' направлении или 5' от первой последовательности hJλ.
В одном из вариантов осуществления межгенная область Vκ-Jκ человека представляет собой область, расположенную приблизительно в 130 п.о. в 5'-3' направлении или 3' от генного сегмента Vκ4-1 человека, приблизительно в 130 п.о. в 5'-3' направлении от 3'-нетранслируемой области генного сегмента Vκ4-1 человека, и перекрывает приблизительно 600 п.о. в 3'-5' направлении или 5' от генного сегмента Jκ1 человека. В определенном варианте осуществления размер межгенной области Vκ-Jκ человека составляет приблизительно 22,8 т.п.о. В одном из вариантов осуществления межгенная область Vκ-Jκ приблизительно на 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, или приблизительно на 95% или более, идентична межгенной области Vκ-Jκ человека, простирающейся от конца 3'-нетранслируемой области генного сегмента Vκ4-1 человека до приблизительно 600 п.о. в 3'-5' направлении генного сегмента Jκ1 человека. В одном из вариантов осуществления межгенная область Vκ-Jκ содержит последовательность SEQ ID NO:100. В определенном варианте осуществления межгенная область Vκ-Jκ содержит функциональный фрагмент последовательности SEQ ID NO:100. В определенном варианте осуществления межгенная область Vκ-Jκ представляет собой последовательность SEQ ID NO:100.
В одном из аспектов изобретение относится к мыши, клеткам мыши (например, эмбриональная стволовая клетка мыши), эмбриону мыши и тканям мыши, которые содержат описанную последовательность межгенной области Vκ-Jκ человека, где последовательность межгенной области является эктопической. В определенном варианте осуществления эктопическая последовательность расположена в гуманизированном эндогенном локусе иммуноглобулина мыши.
В одном из аспектов изобретение относится к выделенной конструкции нуклеиновой кислоты, которая содержит описанную последовательность межгенной области Vκ-Jκ человека. В одном из вариантов осуществления конструкция нуклеиновой кислоты содержит направляющие плечи для нацеливания последовательности межгенной области Vκ-Jκ человека на локус легкой цепи мыши. В определенном варианте осуществления локус легкой цепи мыши представляет собой локус κ. В определенном варианте осуществления данные направляющие плечи нацеливают данную последовательность межгенной области Vκ-Jκ человека на модифицированный эндогенный локус κ мыши, где нацеливание относится к положению между последовательностью hVλ и последовательностью hJλ.
В одном из аспектов изобретение относится к генетически модифицированной мыши, где мышь содержит не более, чем две аллели легкой цепи, где данные аллели легкой цепи содержат (a) неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина человека в эндогенном локусе легкой цепи мыши, который содержит ген CL мыши; и, (b) неперегруппированный генный сегмент VL и JL иммуноглобулина в эндогенном локусе легкой цепи мыши, который содержит ген CL мыши.
В одном из вариантов осуществления эндогенный локус легкой цепи мыши представляет собой локус κ. В другом варианте осуществления эндогенный локус легкой цепи мыши представляет собой локус λ.
В одном из вариантов осуществления не более двух аллелей легкой цепи выбрано из аллеля κ и аллеля λ, двух аллелей κ, и двух аллелей λ. В определенном варианте осуществления один из двух аллелей легкой цепи представляет собой аллель λ, который содержит ген Сλ2.
В одном из вариантов осуществления мышь содержит один функциональный локус легкой цепи иммуноглобулина и один нефункциональный локус легкой цепи иммуноглобулина, где функциональный локус легкой цепи содержит неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина человека в эндогенном локусе легкой цепи κ мыши, который содержит ген Сκ мыши.
В одном из вариантов осуществления мышь содержит один функциональный локус легкой цепи иммуноглобулина и один нефункциональный локус легкой цепи иммуноглобулина, где функциональный локус легкой цепи содержит неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина человека в эндогенном локусе легкой цепи λ мыши, который содержит ген Сκ мыши. В одном из вариантов осуществления ген Сλ представляет собой Сλ2. В определенном варианте осуществления ген Сλ мыши по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентичен Сλ2 мыши.
В одном из вариантов осуществления мышь дополнительно содержит по меньшей мере один аллель тяжелой цепи иммуноглобулина. В одном из вариантов осуществления по меньшей мере один аллель тяжелой цепи иммуноглобулина содержит генный сегмент VH человека, и генный сегмент DH человека, и генный сегмент JH в эндогенном локусе тяжелой цепи мыши, который содержит ген тяжелой цепи человека, который экспрессирует тяжелую цепь человек/мышь. В определенном варианте осуществления мышь содержит два аллеля тяжелой цепи иммуноглобулина, и мышь экспрессирует тяжелую цепь человек/мышь.
В одном из вариантов осуществления мышь содержит первый аллель легкой цепи, который содержит неперегруппированный hVκ и неперегруппированный hJκ в эндогенном локусе κ мыши, содержащем эндогенный ген Сκ; и второй аллель легкой цепи, который содержит неперегруппированный hVλ и неперегруппированный hJλ в эндогенном локусе κ мыши, содержащем эндогенный ген Сκ. В определенном варианте осуществления первый и второй аллели легкой цепи являются единственными функциональными аллелями легкой цепи генетически модифицированной мыши. В определенном варианте осуществления мышь содержит нефункциональный локус λ. В одном из вариантов осуществления генетически модифицированная мышь не экспрессирует легкую цепь, которая содержит константную область λ.
В одном из вариантов осуществления мышь содержит первый аллель легкой цепи, который содержит неперегруппированный hVκ и неперегруппированный hJκ в эндогенном локусе κ мыши, содержащем эндогенный ген Сκ; и второй аллель легкой цепи, который содержит неперегруппированный hVλ и неперегруппированный hJλ в эндогенном локусе λ мыши, содержащем эндогенный ген Сλ. В определенном варианте осуществления первый и второй аллели легкой цепи являются единственными функциональными аллелями легкой цепи генетически модифицированной мыши. В одном из вариантов осуществления ген Сλ представляет собой Сλ2. В определенном варианте осуществления ген Сλ мыши, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или по меньшей мере на 98% идентичен Сλ2 мыши.
В одном из вариантов осуществления мышь содержит шесть аллелей иммуноглобулина, где первый аллель содержит неперегруппированный генный сегмент Vλ и Jλ в эндогенном локусе легкой цепи κ мыши, который содержит ген Сκ мыши, второй аллель содержит неперегруппированный генный сегмент Vκ и Jκ иммуноглобулина в эндогенном локусе легкой цепи κ мыши, который содержит ген Сκ мыши, третий аллель содержит неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина в эндогенном локусе легкой цепи λ мыши, который содержит ген Сλ мыши, четвертый и пятый аллели, каждый независимо друг от друга, содержат неперегруппированный генный сегмент VH и DH и JH в эндогенном локусе тяжелой цепи мыши, который содержит ген тяжелой цепи мыши, и шестой аллель содержит либо (а) неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина в эндогенном локусе легкой цепи λ мыши, содержащем ген Сλ мыши, либо (b) локус λ, который является нефункциональным, либо (с) делецию всего или части локуса λ.
В одном из вариантов осуществления первый аллель содержит неперегруппированные hVλ и hJλ. В одном из вариантов осуществления второй аллель содержит неперегруппированные hVκ и hJκ. В одном из вариантов осуществления третий аллель содержит неперегруппированные hVλ и hJλ. В одном из вариантов осуществления четвертый и пятый аллели, каждый независимо друг от друга, содержат неперегруппированный hVH и hDH, и hJH. В одном из вариантов осуществления шестой аллель содержит эндогенный локус λ мыши, который делетирован полностью или частично.
В одном из вариантов осуществления мышь содержит шесть аллелей иммуноглобулина, где первый аллель содержит неперегруппированный генный сегмент Vλ и Jλ в эндогенном локусе легкой цепи λ мыши, который содержит ген Сλ мыши, второй аллель содержит неперегруппированный генный сегмент Vλ и Jλ иммуноглобулина в эндогенном локусе легкой цепи λ мыши, который содержит ген Сλ мыши, третий аллель содержит неперегруппированный генный сегмент Vκ и Jκ иммуноглобулина в эндогенном локусе легкой цепи κ мыши, который содержит ген Сκ мыши, четвертый и пятый аллели, каждый независимо друг от друга, содержат неперегруппированный генный сегмент VH и DH, и JH в эндогенном локусе тяжелой цепи мыши, который содержит ген тяжелой цепи мыши, и шестой аллель содержит либо (а) неперегруппированный генный сегмент Vκ и Jκ иммуноглобулина в эндогенном локусе легкой цепи κ мыши, содержащем ген Сκ мыши, либо (b) локус κ, который является нефункциональным, либо (с) делецию одного или нескольких элементов локуса κ.
В одном из вариантов осуществления первый аллель содержит неперегруппированные генные сегменты hVλ и hJλ. В одном из вариантов осуществления второй аллель содержит неперегруппированные генные сегменты hVλ и hJλ. В одном из вариантов осуществления третий аллель содержит неперегруппированные генные сегменты hVκ и hJκ. В одном из вариантов осуществления четвертый и пятый аллели, каждый независимо друг от друга, содержит неперегруппированный генный сегмент hVH и hDH, и hJH. В одном из вариантов осуществления шестой аллель содержит эндогенный локус κ мыши, который функционально отключен.
В одном из вариантов осуществления генетически модифицированная мышь содержит В-клетку, которая содержит перегруппированный ген антитела, содержащий перегруппированный домен hVλ, функционально связанный с доменом CL мыши. В одном из вариантов осуществления домен CL мыши выбран из домена Сλ мыши и Сκ мыши. В определенном варианте осуществления домен Сλ мыши получен из гена Сλ2. В определенном варианте осуществления домен Сλ мыши получен из домена Сλ, который, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере, на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентичен Cλ2 мыши.
В одном из аспектов изобретение относится к генетически модифицированной мыши, которая экспрессирует область Vλ в CL, который представляет собой Cκ. В одном из аспектов изобретение относится к генетически модифицированной мыши, которая экспрессирует область hVλ в CL, выбранном из Сκ человека, Сλ человека или Сκ мыши. В одном из аспектов изобретение относится к генетически модифицированной мыши, которая экспрессирует область hVλ в Сκ мыши.
В одном из вариантов осуществления приблизительно 10-50% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 9-28% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В определенном варианте осуществления приблизительно 23-34% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 9-11% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В определенном варианте осуществления приблизительно 19-31% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 9-17% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В определенном варианте осуществления приблизительно 21-38% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 24-27% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В определенном варианте осуществления приблизительно 10-14% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 9-13% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В определенном варианте осуществления приблизительно 31-48% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 15-21% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши. В определенном варианте осуществления приблизительно 30-38% спленоцитов мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 33-48% экпрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В одном из вариантов осуществления приблизительно 52-70% костного мозга мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 31-47% незрелые В-клетки (а именно, CD19-положительные/В220-промежуточно положительные/IgM-положительные) экспрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В одном из вариантов осуществления приблизительно 60% костного мозга мыши представляют собой В-клетки (а именно, CD19-положительные), или которые приблизительно 38,3% незрелые В-клетки (а именно, CD19-положительные/В220-промежуточно положительные/IgM-положительные) экспрессируют легкую цепь иммуноглобулина, содержащую домен hVλ, слитый с доменом Сκ мыши.
В одном из вариантов осуществления мышь экспрессирует антитело, содержащее легкую цепь, которая включает вариабельный домен, полученный из генного сегмента V человека и J человека, и константный домен, полученный из гена костантной области мыши. В одном из вариантов осуществления ген константной области мыши представляет собой ген Сκ. В другом варианте осуществления ген константной области мыши представляет собой ген Сλ. В определенном варианте осуществления область Сλ представляет собой Сλ2. В определенном варианте осуществления ген Сλ мыши получен из гена Сλ, который, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или, по меньшей мере на 98% идентичен Сλ2 мыши. В определенном варианте осуществления антитело дополнительно содержит тяжелую цепь, содержащую вариабельный домен, полученный из генного сегмента V человека, генного сегмента D человека и генного сегмента J человека, и константный домен тяжелой цепи, полученный из гена константной области тяжелой цепи мыши. В одном из вариантов осуществления ген константной области тяжелой цепи мыши содержит последовательность шарнир-СН2-СН3 константного домена тяжелой цепи. В другом варианте осуществления ген константной области тяжелой цепи мыши содержит последовательность СН1-шарнир-СН2-СН3 константного домена тяжелой цепи. В другом варианте осуществления ген константной области тяжелой цепи мыши содержит последовательность СН1-СН2-СН3-СН4 константного домена тяжелой цепи. В другом варианте осуществления ген константной области тяжелой цепи мыши содержит последовательность СН2-СН3-СН4 константного домена тяжелой цепи.
В одном из вариантов осуществления мышь экспрессирует антитело, содержащее легкую цепь, которая содержит перегруппированную последовательность Vλ-Jλ человека и последовательность Сκ мыши. В одном из вариантов осуществления перегруппированная последовательность Vλ-Jλ человека получена в результате перегруппировки генных сегментов hVλ, выбранных из генных сегментов 3-1, 4-3, 2-8, 3-9, 3-10, 2-14, 3-19, 2-23, 3-25, 1-40, 7-43, 1-44, 5-45, 7-46, 1-47, 9-49 и 1-51. В одном из вариантов осуществления перегруппированная последовательность Vλ-Jλ человека получена в результате перегруппировки генных сегментов hJλ, выбранных из генных сегментов Jλ1, Jλ2, Jλ3 и Jλ7.
В одном из вариантов осуществления мышь экспрессирует антитело, содержащее легкую цепь, которая содержит перегруппированную вариабельную область легкой цепи λ иммуноглобулина, содержащий последовательность Vλ/Jλ человека, выбранную из 3-1/1, 3-1/7, 4-3/1, 4-3/7, 2-8/1, 3-9/1, 3-10/1, 3-10/3, 3-10/7, 2-14/1, 3-19/1, 2-23/1, 3-25/1, 1-40/1, 1-40/2, 1-40/3, 1-40/7, 7-43/1, 7-43/3, 1-44/1, 1-44/7, 5-45/1, 5-45/2, 5-45/7, 7-46/1, 7-46/2, 7-46/7, 9-49/1, 9-49/2, 9-49/7 и 1- 51/1. В определенном варианте осуществления В-клетка экспрессирует антитело, содержащее вариабельный домен тяжелой цепи иммуноглобулина человека, слитый с константным доменом тяжелой цепи мыши, и вариабельный домен легкой цепи λ иммуноглобулина человека, слитый с константным доменом легкой цепи κ мыши.
В одном из аспектов изобретение относится к мыши, которая экспрессирует антитело, содержащее (a) тяжелую цепь, содержащую вариабельный домен тяжелой цепи, полученный из неперегруппированного генного сегмента вариабельной области тяжелой цепи человека, где вариабельный домен тяжелой цепи слит с константной областью тяжелой цепи (СН) мыши; и (b) легкую цепь, содержащую вариабельный домен легкой цепи, полученный из неперегруппированных hVλ и hJλ, где вариабельный домен легкой цепи слит с областью CL мыши.
В одном из вариантов осуществления мышь содержит (i) локус тяжелой цепи, который содержит замену всех или практически всех эндогенных функциональных генных сегментов V, D и J мыши на все, или практически все функциональные генные сегменты V, D, и J человека, и ген СН мыши, (ii) первый локус легкой цепи κ, содержащий замену всех или практически всех функциональных эндогенных генных сегментов Vκ и Jκ на все, или практически все, или совокупность, функциональных генных сегментов hVλ и hJλ, и ген Сκ мыши, (iii) второй локус легкой цепи κ, содержащий замену всех, или практически всех, функциональных эндогенных генных сегментов Vκ и Jκ мыши на все, или практически все, или совокупность, функциональных генных сегментов hVκ и hJκ, и ген Сκ мыши. В одном из вариантов осуществления мышь не экспрессирует антитело, которое содержит область Сλ. В одном из вариантов осуществления мышь содержит делецию гена Сλ и/или генного сегмента Vλ и/или генного сегмента Jλ. В одном из вариантов осуществления мышь содержит нефункциональный локус легкой цепи λ. В определенном варианте осуществления локус легкой цепи λ делетирован полностью или частично.
В одном из вариантов осуществления мышь содержит (i) локус тяжелой цепи, который содержит замену всех или практически всех эндогенных функциональных генных сегментов V, D и J мыши на все, или практически все функциональные генные сегменты V, D, и J человека, и ген СН мыши, (ii) первый локус легкой цепи λ, включающий замену всех или практически всех функциональных эндогенных генных сегментов Vλ и Jλ на все, или практически все, или совокупность, функциональных генных сегментов hVλ и hJλ, и ген Сλ мыши, (iii) второй локус легкой цепи λ, содержащий замену всех, или практически всех, функциональных эндогенных генных сегментов Vλ и Jλ мыши на все, или практически все, или совокупность, функциональных генных сегментов hVλ и hJλ, и ген Сλ мыши. В определенном варианте осуществления ген Сλ мыши представляет собой Сλ2. В определенном варианте осуществления ген Сλ мыши получен из гена Сλ, который, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или, по меньшей мере на 98% идентичен Сλ2 мыши.
В одном из вариантов осуществления мышь содержит делецию гена Сκ и/или генного сегмента Vκ, и/или генного сегмента Jκ. В одном из вариантов осуществления мышь содержит нефункциональный локус легкой цепи κ.
В одном из аспектов изобретение относится к генетически модифицированной мыши, которая экспрессирует антитело, где более, чем 10%, более, чем 15%, более, чем 20%, более, чем 25%, более, чем 30%, более, чем 35%, более, чем 40%, более, чем 60%, более, чем 70%, более, чем 80%, или более, чем 90% общего количества антитела IgG, продуцируемого мышью, содержит вариабельный домен λ-происхождения, и где мышь экспрессирует антитела, содержащие вариабельный домен κ-происхождения, слитый с областью Сκ мыши. В определенном варианте осуществления приблизительно 15-40%, 20-40%, 25-40%, 30-40%, или 35-40% общего антитела, продуцируемого мышью, содержит вариабельный домен λ-происхождения.
В одном из вариантов осуществления вариабельный домен λ-происхождения получен из hVλ и hJλ. В одном из вариантов осуществления вариабельный домен λ-происхождения находится на легкой цепи, которая включает область Сκ мыши. В определенном варианте осуществления вариабельная область λ-происхождения находится на легкой цепи, которая содержит область Сλ мыши. В другом специфическом варианте осуществления область Сλ представляет собой область Сλ2. В одном из вариантов осуществления вариабельный домен κ-происхождения получен из hVκ и hJκ, и в определенном варианте осуществления находится на легкой цепи, которая содержит область Сκ мыши.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, которая содержит гомологичное плечо в 3'-5' направлении и гомологичное плечо в 5'-3' направлении, где данные гомологичные плечи в 3'-5' направлении и 5'-3' направлении нацеливают данную конструкцию на локус κ мыши, и конструкция содержит функциональный неперегруппированный сегмент hVλ и функциональный неперегруппированный сегмент hJλ, и селекционную или маркерную последовательность.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания последовательности λ мыши в направлении 3'-5' Vλ2 мыши, селекционную кассету, фланкированную 5' и 3' участками распознавания рекомбиназ, и направляющее плечо для нацеливания последовательности λ мыши 3' Jλ2 мыши. В одном из вариантов осуществления селекционная кассета представляет собой кассету Frt Hyg-TK. В одном из вариантов осуществления 3' направляющее плечо содержит Cλ2, Jλ4, Cλ4 и энхансер 2.4 мыши.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса λ мыши в 5'-направлении относительно Vλ1, селекционную кассету, фланкированную 5' и 3' участками распознавания рекомбиназ, и направляющее плечо 3' для нацеливания последовательности λ мыши 3' относительно Сλ1 мыши. В одном из вариантов осуществления селекционная кассета представляет собой неомициновую кассету, фланкированную lox-сайтами. В одном из вариантов осуществления 3' направляющее плечо содержит 3' энхансер λ мыши и 3' энхансер 3.1 λ мыши.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса λ мыши в 5'-направлении относительно Vλ2, селекционную кассету, фланкированную 5' и 3' участками распознавания рекомбиназ, и направляющее плечо 3' для нацеливания последовательности λ мыши 3' относительно Jλ2 мыши и 5' относительно Сλ2. В одном из вариантов осуществления селекционная кассета представляет собой кассету Frt гигромицин-TK. В одном из вариантов осуществления 3' направляющее плечо содержит генные сегменты Cλ2-Jλ4-Cλ4 и энхансер 2.4 λ мыши.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса λ мыши в 5'-направлении относительно Vλ2, селекционную кассету, фланкированную 5' и 3' участками распознавания рекомбиназ, геномный фрагмент человека, содержащий смежную область локуса легкой цепи λ человека от hVλ3-12 в направлении 5'-3' до конца hJλ1, и направляющее плечо 3' для нацеливания последовательности λ мыши 3' относительно Jλ2. В одном из вариантов осуществления селекционная кассета представляет собой Frt неомициновую кассету. В одном из вариантов осуществления 3'-направляющее плечо содержит генные сегменты Cλ2-Jλ4-Cλ4 мыши и энхансер 2.4 λ мыши.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, включающей смежную область локуса легкой цепи λ человека от hVλ3-12 в направлении 5'-3' до конца hJλ1.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса λ мыши в 5'-направлении относительно Vλ2, селекционную кассету, фланкированную 5' и 3' участками распознавания рекомбиназ, и геномный фрагмент человека, содержащий смежную область локуса легкой цепи λ человека от hVλ3-27 в направлении 5'-3' до конца hVλ2-8. В одном из вариантов осуществления селекционная кассета представляет собой Frt гигромициновую кассету. В одном из вариантов осуществления геномный фрагмент человека содержит 3' направляющее плечо. В определенном варианте осуществления 3' направляющее плечо содержит приблизительно 53 т.п.о. локуса легкой цепи λ человека от hVλ3-12 в направлении 5'-3' до конца hVλ2-8.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей смежную область локуса легкой цепи λ человека от hVλ3-27 в направлении 5'-3' до конца hVλ3-12.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса λ мыши в 5'-направлении относительно Vλ2, селекционную кассету, фланкированную 5' и 3' участками распознавания рекомбиназ, первый геномный фрагмент человека, содержащий смежную область локуса легкой цепи λ человека от hVλ5-52 в направлении 5'-3' до конца hVλ1-40, участок рестрикционного фермента и второй геномный фрагмент человека, содержащий смежную область локуса легкой цепи λ человека от hVλ3-29 в направлении 5'-3' до конца hVλ82Κ. В одном из вариантов осуществления селекционная кассета представляет собой Frt неомициновую кассету. В одном из вариантов осуществления участок фермента рестрикции представляет собой участок для хоминг-эндонуклеазы. В определенном варианте осуществления хоминг-эндонуклеазой является PI-SceI. В одном из вариантов осуществления второй геномный фрагмент человека представляет собой 3' направляющее плечо. В определенном варианте осуществления 3' направляющее плечо содержит приблизительно 27 т.п.о. локуса легкой цепи λ человека от hVλ3-29 в 5'-3' направлении до конца hVλ82K.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей смежную область локуса легкой цепи λ человека от hVλ5-52 в направлении 5'-3' до конца hVλ1-40.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса κ мыши в 5'-направлении относительно эндогенных генных сегментов Vκ, два смежных участка распознавания рекомбиназ, и селекционную кассету 3' к смежным участкам распознавания рекомбиназ, и 3' направляющее плечо для нацеливания последовательности κ мыши 5' относительно вариабельных генных сегментов легкой цепи κ. В одном из вариантов осуществления данные смежные участки распознавания рекомбиназ находятся в противоположной ориентации относительно друг друга. В определенном варианте осуществления данные участки распознавания рекомбиназ являются разными. В другом определенном варианте осуществления и участками распознавания рекомбиназ являются участок loxP и участок lox511. В одном из вариантов осуществления селекционной кассетой является неомициновая кассета.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, направляющее плечо для нацеливания локуса κ мыши в 5'-направлении относительно генных сегментов Jκ мыши, селекционную кассету, участки распознавания рекомбиназ 3' к селекционной кассете, и 3' направляющее плечо для нацеливания последовательности κ мыши 5' относительно генных сегментов Jκ мыши, и 5' к интронному энхансеру κ мыши. В одном из вариантов осуществления селекционной кассетой является кассета гигромицин-ТК. В одном из вариантов осуществления участок распознавания рекомбиназы находится в той же ориентации относительно транскрипции, что и селекционная кассета. В определенном варианте осуществления участком распознавания рекомбиназы является участок loxP.
В одном из аспектов изобретение относится к выделенной конструкции ДНК, содержащей в 5'-3' направлении относительно направления транскрипции, первый геномный фрагмент мыши, содержащий 5'-последовательность эндогенных генных сегментов Vκ мыши, первый участок распознавания рекомбиназы, второй участок распознавания рекомбиназы и второй геномный фрагмент мыши, содержащий 3' последовательность эндогенных генных сегментов Jκ мыши, и 5' интронного энхансера κ мыши.
В одном из аспектов изобретение относится к генетически модифицированной мыши, где генетическая модификация содержит модификацию с одной или несколькими конструкциями ДНК, описанными ранее или в настоящем описании.
В одном из аспектов изобретение относится к применению выделенной конструкции ДНК для создания мыши, как описано в настоящем описании. В одном из аспектов изобретение относится к применению выделенной конструкции ДНК, как описано в настоящем описании, в способе получения антигенсвязывающего белка.
В одном из аспектов изобретение относится к стволовой клетке не-человеческого происхождения, которая содержит направляющий вектор, содержащий конструкцию ДНК, как описано ранее и в настоящем описании. В одном из аспектов изобретение относится к стволовой клетке не-человеческого происхождения, где стволовая клетка не-человеческого происхождения получена от мыши, описанной в настоящем описании.
В одном из вариантов осуществления стволовая клетка не-человеческого происхождения представляет собой эмбриональную стволовую (ES) клетку. В определенном варианте осуществления ES-клетка представляет собой ES-клетку мыши.
В одном из аспектов изобретение относится к применению не-человеческой стволовой клетки, как описано в настоящем описании, для получения мыши, как описано в настоящем описании. В одном из аспектов изобретение относится к применению не-человеческой стволовой клетки, как описано в настоящем описании, для получения антигенсвязывающего белка.
В одном из аспектов изобретение относится к эмбриону мыши, где эмбрион мыши содержит предусмотренную в настоящем описании генетическую модификацию. В одном из вариантов осуществления изобретение относится к эмбриону-хозяину мыши, который содержит донорскую ES-клетку, где донорская ES-клетка включает описанную в настоящем описании генетическую модификацию. В одном из вариантов осуществления эмбрион мыши представляет собой эмбрион на стадии преморулы. В определенном варианте осуществления эмбрион на стадии преморулы представляет собой 4-клеточный эмбрион или 8-клеточный эмбрион. В другом специфическом варианте осуществления эмбрион мыши представляет собой бластоцисту.
В одном из аспектов изобретение относится к применению мышиного эмбриона, как описано в настоящем описании, для получения мыши, как описано в настоящем описании. В одном из аспектов изобретение относится к применению эмбриона мыши, как описано в настоящем описании, для получения антигенсвязывающего белка.
В одном из аспектов изобретение относится к клетке, которая не является клеткой человека, где клетка, которая не является клеткой человека, содержит перегруппированную последовательность гена легкой цепи иммуноглобулина, полученной из генетически модифицированной мыши, как описано в настоящем описании. В одном из вариантов осуществления клетка представляет собой В-клетку. В одном из вариантов осуществления клетка представляет собой гибридому. В одном из вариантов осуществления клетка кодирует вариабельный домен легкой цепи иммуноглобулина и/или вариабельный домен тяжелой цепи иммуноглобулина, который соматически видоизменен.
В одном из аспектов изобретение относится к не-человеческая клетке, где не-человеческая клетка содержит перегруппированную последовательность гена легкой цепи иммуноглобулина, полученной из генетически модифицированной мыши, как описано в настоящем описании. В одном из вариантов осуществления клетка представляет собой В-клетку. В одном из вариантов осуществления клетка представляет собой гибридому. В одном из вариантов осуществления клетка кодирует вариабельный домен легкой цепи иммуноглобулина и/или вариабельный домен тяжелой цепи иммуноглобулина, который соматически видоизменен.
В одном из аспектов изобретение относится к применению не-человеческой клетки, как описано в настоящем описании, для получения мыши, как описано в настоящем описании. В одном из аспектов изобретение относится к использованию не-человеческой клетки, как описано в настоящем описании, для получения антигенсвязывающего белка.
В одном из аспектов изобретение относится к В-клетке мыши, которая экспрессирует легкую цепь иммуноглобулина, которая содержит (a) вариабельную область, полученную из генного сегмента hVλ и генного сегмента hJλ; и, (b) ген CL мыши. В одном из вариантов осуществления ген CL мыши выбран из гена Сκ и Сλ. В определенном варианте осуществления ген Сλ представляет собой Сλ2. В определенном варианте осуществления ген Сλ мыши получен из гена Сλ, который, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или по меньшей мере на 98% идентичен Сλ2 мыши. В одном из вариантов осуществления В-клетка мыши дополнительно экспрессирует родственную тяжелую цепь, которая содержит (c) вариабельную область, полученную из сегмента hVH, hDH, и (d) hJH. В одном из вариантов осуществления В-клетка не содержит перегруппированный ген λ. В другом варианте осуществления В-клетка не содержит перегруппированный ген κ.
В одном из аспектов изобретение относится к способу получения антитела в генетически модифицированной мыши, включающему: (а) воздействие на генетически модифицированную мышь антигеном, где в геноме мыши содержится по меньшей мере один hVλ и по меньшей мере один hJλ в эндогенном локусе легкой цепи, содержащем ген CL мыши; (b) развитие у генетически модифицированной мыши иммунного ответа на антиген; и (c) выделение из организма мыши (b) антитела, которое специфически распознает антиген, или выделение из организма мыши (b) клетки, содержащей домен иммуноглобулина, который специфически распознает антиген, где антитело содержит легкую цепь, происходящую из hVλ, hJλ, и гена мыши CL. В определенном варианте осуществления ген CL мыши представляет собой ген Сκ мыши.
В одном из вариантов осуществления изобретение относится к способу получения антитела в генетически модифицированной мыши, включающему: (а) воздействие на генетически модифицированную мышь антигеном, где в геноме мыши содержится по меньшей мере один hVλ в эндогенном локусе κ и, по меньшей мере один hJλ в эндогенном локусе κ, где локус κ содержит ген Cκ мыши; (b) развитие у генетически модифицированной мыши иммунного ответа на антиген; и, (c) выделение из организма мыши (b) антитела, которое специфически распознает антиген, или выделение из организма мыши (b) клетки, содержащей домен иммуноглобулина, который специфически распознает антиген, где антитело содержит легкую цепь, происходящую из hVλ, hJλ, и гена мыши CL.
В одном из вариантов осуществления константный ген легкой цепи κ выбран из гена Сκ человека и гена Сκ мыши.
В одном из вариантов осуществления изобретение относится к способу получения антитела в генетически модифицированной мыши, включающему: (а) воздействие на генетически модифицированную мышь антигеном, где в геноме мыши содержится по меньшей мере один hVλ в локусе легкой цепи λ и по меньшей мере один Jλ в локусе легкой цепи λ, где локус легкой цепи λ содержит ген Cλ мыши; (b) развитие у генетически модифицированной мыши иммунного ответа на антиген; и, (c) выделение из организма мыши (b) антитела, которое специфически распознает антиген, или выделение из организма мыши (b) клетки, содержащей домен иммуноглобулина, который специфически распознает антиген, или выявление в мыши (B) последовательности нуклеиновой кислоты, кодирующей вариабельный домен тяжелой и/или легкой цепи, который связывает антиген, где антитело содержит легкую цепь, происходящую из hVλ, hJλ и гена Cλ мыши.
В одном из вариантов осуществления константный ген легкой цепи λ выбран из гена Сλ человека и гена Сλ мыши. В одном из вариантов осуществления константный ген легкой цепи λ представляет собой ген Сλ человека. В определенном варианте осуществления ген Сλ человека выбран из Cλ1, Cλ2, Cλ3 и Cλ7. В одном из вариантов осуществления константный ген легкой цепи λ представляет собой ген Сλ мыши. В определенном варианте осуществления ген Сλ мыши выбран из Cλ1, Cλ2 и Cλ3. В более специфическом варианте осуществления ген Сλ мыши представляет собой Сλ2. В другом специфическом варианте осуществления ген Сλ происходит из гена Сλ, который, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или, по меньшей мере на 98% идентичен Сλ2 мыши.
В одном из аспектов изобретение относится к способу получения перегруппированного гена антитела в генетически модифицированной мыши, включающему: (a) воздействие на генетически модифицированную мышь антигеном, где генетическая модификация содержит hVλ и hJλ в эндогенном локусе легкой цепи, где эндогенный локус легкой цепи содержит ген CL мыши или его функциональный фрагмент; и, (b) выявление перегруппированного гена иммуноглобулина у указанной мыши, где перегруппированный ген иммуноглобулина содержит генный сегмент вариабельной области легкой цепи λ и ген CL, или его функциональный фрагмент.
В одном из вариантов осуществления способ дополнительно включает клонирование последовательности нуклеиновой кислоты, кодирующей вариабельную область тяжелой и/или легкой цепи мыши, где вариабельная область тяжелой и/или легкой цепи происходит из антитела, которое содержит Vλ человека и CL мыши.
В одном из вариантов осуществления ген CL мыши, или его функциональный фрагмент, выбран из гена CL человека и гена CL мыши, или их функционального фрагмента.
В одном из вариантов осуществления изобретение относится к способу получения перегруппированного гена антитела в генетически модифицированной мыши, включающему: (а) воздействие на генетически модифицированную мышь антигеном, где генетическая модификация содержит hVλ и hJλ в локусе легкой цепи κ, где локус легкой цепи κ содержит ген Cκ мыши или его функциональный фрагмент; и, (b) выявление перегруппированного гена иммуноглобулина у указанной мыши, где перегруппированный ген иммуноглобулина содержит генный сегмент вариабельной области легкой цепи λ и ген Cκ, или его функциональный фрагмент.
В одном из вариантов осуществления константный ген легкой цепи κ, или его функциональный фрагмент, выбран из гена Сκ человека и гена Сκ мыши, или их функционального фрагмента.
В одном из вариантов осуществления способ дополнительно включает клонирование последовательности нуклеиновой кислоты, кодирующей вариабельную область тяжелой и/или легкой цепи мыши, где вариабельная область тяжелой и/или легкой цепи происходит из антитела, которое содержит Vλ человека и Cκ мыши.
В одном из вариантов осуществления изобретение относится к способу получения перегруппированного гена иммуноглобулина в генетически модифицированной мыши, включающему: (а) воздействие на генетически модифицированную мышь антигеном, где генетическая модификация включает hVλ и hJλ в локусе легкой цепи λ мыши, где локус легкой цепи λ содержит ген Cλ мыши или его функциональный фрагмент; и, (b) выявление перегруппированного гена иммуноглобулина у указанной мыши, где перегруппированный ген иммуноглобулина содержит генный сегмент вариабельной области легкой цепи λ и ген Cλ, или его функциональный фрагмент.
В одном из вариантов осуществления константный ген легкой цепи λ, или его функциональный фрагмент, выбран из гена Сλ человека и гена Сλ мыши, или их функционального фрагмента. В определенном варианте осуществления константный ген легкой цепи λ представляет собой ген Сλ мыши, или его функциональный фрагмент.
В одном из вариантов осуществления способ дополнительно включает клонирование последовательности нуклеиновой кислоты, кодирующей вариабельную область тяжелой и/или легкой цепи мыши, где вариабельная область тяжелой и/или легкой цепи происходит из антитела, которое содержит Vλ человека и Сλ мыши.
В одном из аспектов изобретение относится к способу получения антитела, включающему воздействие на мышь, как описано в настоящем описании, антигеном, обеспечение повышения иммунного ответа у мыши, который включает получение антитела, специфически связывающего антиген, выявление у мыши перегруппированной последовательности нуклеиновой кислоты, которая кодирует тяжелую цепь, и выявление у мыши перегруппированной последовательности нуклеиновой кислоты, которая кодирует последовательность вариабельного домена родственной легкой цепи антитела, где антитело специфически связывается с антигеном, и использование последовательностей нуклеиновых кислот вариабельных доменов тяжелой и легкой цепей, слитых с константными доменами человека, для получения необходимого антитела, где необходимое антитело содержит легкую цепь, содержащую домен Vλ, слитый с доменом CL. В одном из вариантов осуществления домен Vλ является доменом человека, и домен CL представляет собой домен Сλ человека или мыши. В одном из вариантов осуществления домен Vλ представляет собой домен мыши, и домен CL представляет собой домен Сκ человека или мыши.
В одном из вариантов осуществления изобретение относится к способу получения антитела, включающему воздействие на мышь, как описано в настоящем описании, антигеном, обеспечение у мыши повышения иммунного ответа, который включает получение антитела, специфически связывающего антиген, выявление у мыши перегруппированной последовательности нуклеиновой кислоты, которая кодирует тяжелую цепь, и выявление у мыши перегруппированной последовательности нуклеиновой кислоты, которая кодирует последовательность вариабельного домена родственной легкой цепи антитела, где антитело специфически связывается с антигеном, и использование последовательностей нуклеиновых кислот вариабельных доменов тяжелой и легкой цепей, слитых с последовательностями нуклеиновых кислот константных доменов человека, для получения необходимого антитела, где необходимое антитело содержит легкую цепь, содержащую домен Vλ, слитый с доменом Сκ.
В одном из вариантов осуществления изобретение относится к способу получения антитела, включающему воздействие на мышь, как описано в настоящем описании, антигеном, обеспечение повышения иммунного ответа у мыши, который включает получение антитела, специфически связывающего антиген, выявление у мыши перегруппированной последовательности нуклеиновой кислоты, которая кодирует вариабельный домен тяжелой цепи, и выявление у мыши перегруппированной последовательности нуклеиновой кислоты, которая кодирует последовательность вариабельного домена родственной легкой цепи антитела, где антитело специфически связывается с антигеном, и использование последовательностей нуклеиновых кислот, слитых с последовательностями нуклеиновых кислот, кодирующих константный домен тяжелой цепи человека и константный домен легкой цепи человека, для получения необходимого антитела, где антитело, которое специфически связывает антиген, содержит легкую цепь, которая содержит домен Vλ человека, слитый с областью Сλ мыши.
В одном из вариантов осуществления Сλ мыши выбрана из Cλ1, Cλ2 и Cλ3. В определенном варианте осуществления область Сλ мыши представляет собой Сλ2.
В одном из аспектов изобретение относится к способу получения последовательности перегруппированного гена вариабельной области легкой цепи антитела, включающему (a) воздействие на мышь, как описано в настоящем описании, антигеном; (b) обеспечение повышения у мыши иммунного ответа; (c) выявление у мыши клетки, которая содержит последовательность нуклеиновой кислоты, которая кодирует последовательность перегруппированного домена Vλ человека, слитого с доменом CL мыши, где клетка также кодирует родственную тяжелую цепь, содержащую домен VH человека и домен CH мыши, и где клетка экспрессирует антитело, которое связывает антиген; (d) клонирование из клетки последовательности нуклеиновой кислоты, кодирующей домен Vλ человека и последовательности нуклеиновой кислоты, кодирующей родственный домен VH человека; и, (e) применение клонированной последовательности нуклеиновой кислоты, кодирующей домен Vλ человека, и клонированной последовательности нуклеиновой кислоты, кодирующей родственный домен VH человека, для получения полностью человеческого антитела.
В одном из вариантов осуществления изобретение относится к способу получения последовательности перегруппированного гена вариабельной области легкой цепи антитела, включающему (a) воздействие на мышь, как описано в настоящем описании, антигеном; (b) обеспечение повышения у мыши иммунного ответа; (c) выявление у мыши клетки, которая содержит последовательность нуклеиновой кислоты, которая кодирует последовательность перегруппированного домена Vλ человека, смежного в этой же молекуле нуклеиновой кислоты с последовательностью нуклеиновой кислоты, кодирующей домен Cκ мыши, где клетка также кодирует родственную тяжелую цепь, содержащую домен VH человека и домен CH мыши, и где клетка экспрессирует антитело, которое связывает антиген; (d) клонирование из клетки последовательности нуклеиновой кислоты, кодирующей домен Vλ человека и последовательности нуклеиновой кислоты, кодирующей родственный домен VH человека; и, (e) применение клонированной последовательности нуклеиновой кислоты, кодирующей домен Vλ человека, и клонированной последовательности нуклеиновой кислоты, кодирующей родственный домен VH человека, для получения полностью человеческого антитела.
В одном из вариантов осуществления изобретение относится к способу получения последовательности перегруппированного гена вариабельной области легкой цепи антитела, включающему (a) воздействие на мышь, как описано в настоящем описании, антигеном; (b) обеспечение повышения у мыши иммунного ответа; (c) выявление у мыши клетки, которая содержит ДНК, кодирующую последовательность перегруппированного домена Vλ человека, слитую с доменом Cλ мыши, где клетка также кодирует родственную тяжелую цепь, содержащую домен VH человека и домен CH мыши, и где клетка экспрессирует антитело, которое связывает антиген; (d) клонирование из клетки последовательности нуклеиновой кислоты, кодирующей перегруппированный домен Vλ человека, и последовательности нуклеиновой кислоты, кодирующей родственный домен VH человека; и, (e) применение клонированной последовательности нуклеиновой кислоты, кодирующей домен Vλ человека, и клонированной последовательности нуклеиновой кислоты, кодирующей родственный домен VH человека, для получения полностью человеческого антитела. В одном из вариантов осуществления домен Сλ мыши представляет собой домен Сλ2 мыши. В определенном варианте осуществления домен Сλ мыши получен из гена Сλ, который, по меньшей мере, на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, или по меньшей мере на 98% идентичен Сλ2 мыши.
В одном из аспектов изобретение относится к генетически модифицированной мыши, которая экспрессирует легкую цепь, происходящую из λ человека, слитую с эндогенной константной областью (CL) легкой цепи, где мышь, после иммунизации антигеном, продуцирует антитело, содержащее домен Vλ человека, слитый с доменом CL мыши. В одном из вариантов осуществления домен CL мыши выбран из домена Сκ и домена Сλ. В одном из вариантов осуществления домен CL мыши представляет собой домен Сκ. В одном из вариантов осуществления домен CL мыши представляет собой домен Сλ. В определенном варианте осуществления домен Сλ представляет собой Сλ2. В определенном варианте осуществления домен Сλ мыши происходит из гена Сλ, который по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентичен Сλ2 мыши.
В одном из аспектов изобретение относится к генетически модифицированной мыши, содержащей модифицированный эндогенный локус легкой цепи κ или λ, как описано в настоящем описании, которая экспрессирует совокупность легких цепей λ иммуноглобулина, связанную с совокупностью тяжелых цепей иммуноглобулина. В одном из вариантов осуществления тяжелая цепь содержит последовательность человека. В различных вариантах осуществления последовательность человека выбрана из вариабельной последовательности, СН1, шарнира, СН2, СН3 и их комбинации. В одном из вариантов осуществления совокупность легких цепей λ иммуноглобулина содержит последовательность человека. В различных вариантах осуществления последовательность человека выбрана из вариабельной последовательности, константной последовательности и их комбинации. В одном из вариантов осуществления мышь содержит “молчащий” эндогенный локус иммуноглобулина и экспрессирует тяжелую цепь и/или легкую цепь λ из трансгена или внехромосомной эписомы. В одном из вариантов осуществления мышь содержит замену в эндогенном локусе мыши некоторых или всех эндогенных генных сегментов тяжелой цепи мыши (а именно, V, D, J), и/или некоторых, или всех эндогенных константных последовательностей тяжелой цепи мыши (например, CH1, шарнир, CH2, CH3, или их комбинации), и/или некоторых или всех эндогенных последовательностей легкой цепи мыши (например, V, J, константная, или их комбинации), на одну или несколько последовательностей иммуноглобулина человека.
В одном из аспектов изобретение относится к мыши, подходящей для получения антител, которые имеют легкую цепь, происходящую из λ человека, где все, или практически все антитела, полученные в мыши, экспрессируются с легкой цепью, происходящей из λ человека. В одном из вариантов осуществления легкая цепь, происходящая из λ человека, экспрессируется из эндогенного локуса легкой цепи. В одном из вариантов осуществления эндогенный локус легкой цепи представляет собой локус легкой цепи κ. В определенном варианте осуществления локус легкой цепи κ представляет собой локус легкой цепи κ мыши.
В одном из аспектов изобретение относится к способу получения легкой цепи λ-происхождения для антитела человека, включающему получение от мыши, как описано в настоящем описании, последовательности легкой цепи и последовательности тяжелой цепи, и применение последовательности легкой цепи и последовательности тяжелой цепи при получении антитела человека.
В одном из аспектов изобретение относится к способу получения антигенсвязывающего белка, включающему воздействие на мышь, как описано в настоящем описании, антигеном; обеспечение наращивания иммунного ответа у мыши; и получение от мыши антигенсвязывающего белка, который связывает антиген, или получение от мыши последовательности, которую используют для получения антигенсвязывающего белка, который связывает антиген.
В одном из аспектов изобретение относится к клетке, полученной от мыши, как описано в настоящем описании. В одном из вариантов осуществления клетка выбрана из эмбриональной стволовой клетки, плюрипотентной клетки, индуцированной плюрипотентной клетки, В-клетки и гибридомы.
В одном из аспектов изобретение относится к клетке, которая содержит генетическую модификацию, как описано в настоящем описании. В одном из вариантов осуществления клетка представляет собой клетку мыши. В одном из вариантов осуществления клетка выбрана из гибридомы и квадрогибридомы. В одном из вариантов осуществления клетка экспрессирует легкую цепь иммуноглобулина, которая содержит вариабельную последовательность λ человека, слитую с константной последовательностью мыши. В определенном варианте осуществления константная последовательность мыши представляет собой константную последовательность κ мыши.
В одном из аспектов изобретение относится к ткани, полученной от мыши, как описано в настоящем описании.
В одном из аспектов изобретение относится к применению мыши или клетки, как описано в настоящем описании, для получения антигенсвязывающего белка. В одном из вариантов осуществления антигенсвязывающий белок представляет собой белок человека. В одном из вариантов осуществления белок человека представляет собой антитело человека.
В одном из аспектов изобретение относится к антигенсвязывающему белку, продуцированному мышью, клеткой, тканью, или способом, как описано в настоящем описании. В одном из вариантов осуществления антигенсвязывающий белок представляет собой белок человека. В одном из вариантов осуществления белок человека представляет собой антитело человека.
Любые из вариантов осуществления и аспектов, описанных в настоящем описании, могут быть использованы в сочетании друг с другом, если не указано иное или очевидно из контекста. Другие варианты осуществления будут очевидны специалисту в данной области из обзора последующего описания.
Краткое описание чертежей
На фиг. 1 представлена подробная иллюстрация, без соблюдения масштаба, локуса легкой цепи λ человека, включающего кластеры генных сегментов Vλ (А, В и С) и пары областей Jλ и Cλ (пары J-C).
На фиг. 2 представлена общая иллюстрация, без соблюдения масштаба, стратегии выбора мишени, используемой для инактивации эндогенного локуса легкой цепи λ мыши.
На фиг. 3 представлена общая иллюстрация, без соблюдения масштаба, стратегии выбора мишени, используемой для инактивации эндогенного локуса легкой цепи κ мыши.
На фиг. 4А представлена общая иллюстрация, без соблюдения масштаба, первичного направленного вектора для нацеливания эндогенного локуса легкой цепи λ мыши с помощью последовательностей легкой цепи λ человека, включающих 12 генных сегментов hVλ и генный сегмент hJλ1 (Направленный вектор 12/1-λ).
На фиг. 4В представлена общая иллюстрация, без соблюдения масштаба, четырех первичных направленных векторов для нацеливания эндогенного локуса легкой цепи κ мыши с помощью последовательностей легкой цепи λ человека, включающих 12 генных сегментов hVλ и генный сегмент hJλ1 (Направленный вектор 12/1-κ), 12 генных сегментов hVλ и генные сегменты hJλ1, 2, 3 и 7 (Направленный вектор 12/4-κ), 12 генных сегментов hVλ, геномную последовательность Vκ-Jκ человека и генный сегмент hJλ1 (Направленный вектор 12(κ)1-κ) и 12 генных сегментов hVλ, геномную последовательность Vκ-Jκ и генные сегменты hJλ1, 2, 3 и 7 (Направленный вектор 12(κ)4-κ).
На фиг. 5А представлена общая иллюстрация, без соблюдения масштаба, стратегии выбора мишени для прогрессивной инсерции 40 генных сегментов hVλ и единственного генного сегмента hJλ в локус легкой цепи λ мыши.
На фиг. 5В представлена общая иллюстрация, без соблюдения масштаба, стратегии выбора мишени для прогрессивной инсерции 40 генных сегментов hVλ и единственного генного сегмента hJλ в локус κ мыши.
На фиг. 6 представлена общая иллюстрация, без соблюдения масштаба, этапов выбора мишени и молекулярной инженерии, применяемых при создании уникальных человеческих λ-κ гибридных направленных векторов для конструирования гибридного локуса легкой цепи, содержащего межгенную последовательность κ человека, совокупность генных сегментов hJλ, или и то, и другое.
На фиг. 7А представлена общая иллюстрация, без соблюдения масштаба, структуры локуса для модифицированного локуса легкой цепи λ мыши, содержащего 40 генных сегментов hVλ и единственный генный сегмент hJλ, функционально связанных с эндогенным геном Cλ2.
На фиг. 7В представлена общая иллюстрация, без соблюдения масштаба, структуры локуса для четырех независимых, модифицированных локусов легкой цепи κ мыши, содержащих 40 генных сегментов hVλ и либо один, либо четыре генных сегмента hJλ с, или без, смежной геномной последовательностью Vκ-Jκ человека, функционально связанных с эндогенным геном Сκ.
На фиг. 8А представлены контурные изображения спленоцитов Igλ+ и Igκ+, селектированных на CD19+ от мыши дикого типа (WT), мыши, гомозиготной по 12 генным сегментам hVλ и четырем генным сегментам hJλ, в том числе геномной последовательности Vκ-Jκ человека (12hVλ-Vκ-Jκ-4hJλ) и мыши, гомозиготной по 40 генным сегментам hVλ и одному генному сегменту hJλ (40hVλ-1hJλ).
На фиг. 8В представлено общее количество В-клеток CD19+ в селезенках, полученных от мышей дикого типа (WT), мышей, гомозиготных по 12 генным сегментам hVλ и четырем генным сегментам hJλ, в том числе геномной последовательности Vκ-Jκ человека (12hVλ-Vκ-Jκ-4hJλ) и мышей, гомозиготных по 40 генным сегментам hVλ и одному генному сегменту hJλ(40hVλ-1hJλ).
На фиг. 9А, на верхней панели, представлены контурные изображения спленоцитов, селектированных в синглетах, и окрашенных на В- и Т-клетки (CD19+ и CD3+, соответственно) от мыши дикого типа (WT) и мыши, гомозиготной по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ). На нижней панели представлены контурные изображения спленоцитов, селектированных на CD19+ и окрашенных на экспрессию Igλ+ и Igκ+ от мыши дикого типа (WT) и мыши, гомозиготной по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ).
На фиг. 9В представлено общее количество В-клеток CD19+, CD19+IgK + и CD19+Igλ+ в селезенках, полученных от мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ).
На фиг. 9С представлены контурные изображения спленоцитов, селектированных на CD19+, и окрашенных на иммуноглобулин D (IgD) и иммуноглобулин М (IgM) от мыши дикого типа (WT) и мыши, гомозиготной по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ). Зрелые (72 для WT, 51 для 40hVλ-Vκ-Jκ-4hJλ) и промежуточные (13 для WT, 22 для 40hVλ-Vκ-Jκ-4hJλ) B-клетки показаны в каждом из контурных изображений.
На фиг. 9D представлено общее количество В-клеток CD19+, промежуточных В-клеток (CD19+IgMhiIgDlo) и зрелых В-клеток (CD19+IgMloIgDhi) в селезенках, полученных от мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ).
На фиг. 10А, на верхней панели, представлены контурные изображения костного мозга, окрашенного на В- и Т-клетки (CD19+ и CD3+, соответственно), полученного от мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ). На нижней панели представлены контурные изображения костного мозга, селектированного на CD19+ и окрашенного на ckit+ и CD43+, полученного от мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ). Про- и пре- В-клетки замечены в контурных изображениях на нижней панели.
На фиг. 10В представлено количество про- (CD19+CD43+ckit+) и пре- (CD19+CD43-ckit-) В-клеток в костном мозге, собранном из бедренной кости мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ).
На фиг. 10С представлены контурные изображения костного мозга, селектированного в синглетах и окрашенного на иммуноглобулин M (IgM) и B220, полученного от мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ). Незрелые, зрелые, пре/про В-клетки замечены в каждом из контурных изображений.
На фиг. 10D представлено общее количество незрелых (B220intIgM+) и зрелых (B220hiIgM+) B-клеток в костном мозге, выделенном из бедренной кости мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ).
На фиг. 10Е представлены контурные изображения костного мозга, селектированного на незрелых (B220intIgM+) и зрелых (B220hiIgM+) В-клетках, и окрашенного на экспрессию Igλ и Igκ, выделенного из бедренной кости мышей дикого типа (WT) и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам Jλ, в том числе геномной последовательности Vκ-Jκ человека (40hVλ-Vκ-Jκ-4hJλ).
На фиг. 11 представлено выравнивание нуклеотидных последовательностей соединения Vλ-Jλ-Cκ из восемнадцати независимых клонов ОТ-ПЦР, амплифицированных из РНК спленоцитов мышей, несущих последовательности генов легкой цепи λ человека в эндогенном локусе легкой цепи κ мыши. A6 = SEQ ID NO:57; B6 = SEQ ID NO:58; F6 = SEQ ID NO:59; B7 = SEQ ID NO:60; E7 = SEQ ID NO:61; F7 = SEQ ID NO:62; C8 = SEQ ID NO:63; E12 = SEQ ID NO:64; 1-4 = SEQ ID NO:65; 1-20 = SEQ ID NO:66; 3B43 = SEQ ID NO:67; 5-8 = SEQ ID NO:68; 5-19 = SEQ ID NO:69; 1010 = SEQ ID NO:70; 11A1 = SEQ ID NO:71; 7A8 = SEQ ID NO:72; 3A3 = SEQ ID NO:73; 2-7 = SEQ ID NO:74. Основания, обозначенные строчными буквами, означают незародышевые основания, полученные либо в результате мутации, и/или N-дополнения во время рекомбинации. Консенсусные аминокислоты в пределах области рамки считывания 4 (FWR4), кодируемые нуклеотидной последовательностью hJλ1 и Сκ мыши, указаны внизу данного выравнивания последовательностей.
На фиг. 12 представлено выравнивание нуклеотидной последовательностей соединения Vλ-Jλ-Cκ из двенадцати независимых клонов ОТ-ПЦР, амплифицированных из РНК спленоцитов мышей, несущих последовательности генов легкой цепи λ человека, в том числе смежную геномную последовательность Vκ-Jκ человека в эндогенном локусе легкой цепи κ мыши. 5-2 = SEQ ID NO:87; 2-5 = SEQ ID NO:88; 1-3 = SEQ ID NO:89; 4B-1 = SEQ ID NO:90; 3B-5 = SEQ ID NO:91; 7A-1 = SEQ ID NO:92; 5-1 = SEQ ID NO:93; 4A-1 = SEQ ID NO:94; 11A-1 = SEQ ID NO:95; 5-7 = SEQ ID NO:96; 5-4 = SEQ ID NO:97; 2-3 = SEQ ID NO:98. Основания, обозначенные строчными буквами, означают незародышевые основания, полученные либо в результате мутации, и/или N-дополнения во время рекомбинации. Консенсусные аминокислоты в пределах области рамки считывания 4 (FWR4), кодируемые нуклеотидной последовательностью каждого Jλ1 человека и Сκ мыши, указаны внизу данного выравнивания последовательностей.
На фиг. 13 представлено выравнивание нуклеотидной последовательностей соединения Vλ-Jλ-Cκ из трех независимых клонов ОТ-ПЦР, амплифицированных из РНК спленоцитов мышей, несущих последовательности легкой цепи λ человека в эндогенном локусе легкой цепи λ мыши. 2D1 = SEQ ID NO:101; 2D9 = SEQ ID NO:102; 3E15 = SEQ ID NO:103. Основания, обозначенные строчными буквами, означают незародышевые основания, полученные либо в результате мутации, и/или N-дополнения во время рекомбинации. Консенсусные аминокислоты в пределах области рамки считывания 4 (FWR4), кодируемые нуклеотидной последовательностью hJλ1 и Сλ2 мыши, указаны внизу данного выравнивания последовательностей.
Подробное описание изобретения
Несмотря на то, что характерные черты различных вариантов осуществления подробно обсуждены, описания специфических аспектов, вариантов осуществления и примеров не ограничивают сущность формулы изобретения; это является формулой изобретения, которая описывает объем настоящего изобретения. Все термины и фразы, используемые в описании, включают значения, обыкновенно приписываемые им в данной области.
Термин «смежный» включает ссылку на нахождение в той же самой молекуле нуклеиновой кислоты, например, две последовательности нуклеиновых кислот являются «смежными», если они находятся в одной и той же молекуле нуклеиновой кислоты, но разделены другой последовательностью нуклеиновой кислоты. Например, перегруппированная последовательность V(D)J является «смежной» с последовательностью гена константной области, хотя за конечным кодоном последовательности V(D)J не следует сразу же первый кодон последовательности константной области. В другом примере две последовательности генных сегментов V являются «смежными», если они находятся в одном и том же геномном фрагменте, несмотря на то что они могут быть разделены последовательностью, которая не кодирует кодон области V, например, они могут быть разделены регуляторной последовательностью, например, промотором или другой некодирующей последовательностью. В одном из вариантов осуществления смежная последовательность включает геномный фрагмент, который содержит геномные последовательности, распределенные как в геноме дикого типа.
Фраза «полученный из», когда используется в отношении вариабельной области, «полученной из» указанного гена, или генного сегмента, включает возможность установить последовательность до специфического неперегруппированного генного сегмента или генных сегментов, которые были перегруппированы для формирования гена, который экспрессирует вариабельный домен (принимая во внимание, где применимо, различия сплайсинга и соматические мутации).
Используемый в настоящем описании термин «функциональный» в отношении генного сегмента вариабельной области, или J-генного сегмента, относится к использованию в экспрессируемом спектре антител; например, среди генных сегментов человека являются функциональными Vλ 3-1, 4-3, 2-8, и тому подобное, в то время как генные сегменты Vλ 3-2, 3-4, 2-5, и тому подобное, являются нефункциональными.
«Локус тяжелой цепи» включает расположение на хромосоме, например, хромосоме мыши, где у мыши дикого типа обнаружены вариабельная ДНК-последовательность тяжелой цепи (VH), ДНК-последовательность реанжировки тяжелой цепи (DH), соединительная ДНК-последовательность тяжелой цепи (JH), и ДНК-последовательность константной области (СН) тяжелой цепи.
«Локус κ» включает положение на хромосоме, например, хромосоме мыши, в котором у мыши дикого типа обнаружены вариабельная ДНК-последовательность κ (Vκ), соединительная ДНК-последовательность κ (Jκ) и константная ДНК-последовательность κ (Cκ).
«Локус λ» включает положение на хромосоме, например, хромосоме мыши, в котором у мыши дикого типа обнаружены вариабельная ДНК-последовательность λ (Vλ), соединительная ДНК-последовательность λ (Jλ) и константная ДНК-последовательность λ (Cλ).
Термин «неперегруппированный» включает состояние локуса иммуноглобулина, когда генные сегменты V и генные сегменты J (для тяжелых цепей так же генные сегменты D) сохраняются в разрозненном состоянии, но их возможно соединить для образования перегруппированного гена V(D)J, который содержит отдельные V,(D),J из спектра V(D)J.
Мыши, экспрессирующие вариабельные домены λ человека
Ранее сообщалось о мышах, которые экспрессируют антитела, являющиеся полностью человеческими, или частично человеческими и частично мышиными. Полученные методами генной инженерии мыши VELOCIMMUNE® содержат замену неперегруппированных генных сегментов V(D)J в эндогенных локусах мыши на генные сегменты V(D)J человека. Мыши VELOCIMMUNE® экспрессируют химерные антитела, имеющие вариабельные домены человека и константные домены мыши (см., например, патент США № 7605237). Большинство других публикаций относятся к мышам, которые экспрессируют полностью человеческие антитела из полностью человеческих трансгенов в мышах, которые имеют “молчащие” эндогенные локусы иммуноглобулина.
Легкие цепи антител кодируются одним из двух раздельных локусов: каппа (κ) и лямбда (λ). Легкие цепи антител мыши главным образом являются κ-типа. Мыши, которые продуцируют антитела мыши, и модифицированные мыши, которые продуцируют полностью человеческие, или химерные человек-мышь, антитела, демонстрируют смещение в использовании легкой цепи. У человека также наблюдается смещение в использовании легкой цепи, но не так четко выраженное, как у мышей; соотношение между использованием легких цепей κ и легких цепей λ у мышей составляет приблизительно 95:5, тогда как у человека это соотношение составляет приблизительно 60:40. Считается, что более выраженное смещение у мышей не оказывает значительного влияния на разнообразие антител, потому что у мышей вариабельный локус λ, прежде всего, не является настолько разнообразным. У человека дело обстоит иначе. У человека локус легкой цепи λ отличается большим разнообразием.
У человека локус легкой цепи λ имеет протяженность более 1000 т.п.о. и содержит свыше 80 генов, которые кодируют вариабельные (V) или соединительные (J) сегменты (фиг. 1). В пределах локуса легкой цепи λ человека более половины всех обнаруженных доменом Vλ кодируются генными сегментами 1-40, 1-44, 2-8, 2-14, и 3-21. В целом, считают, что приблизительно 30, или около того, генных сегментов Vλ человека являются функциональными. Существуют семь генных сегментов Jλ, и только четыре из них рассматриваются как в целом функциональные генные сегменты Jλ - Jλ1, Jλ2, Jλ3, и Jλ7.
У человека локус легкой цепи λ по структуре похож на локус κ как мыши, так и человека, тем, что локус легкой цепи λ человека имеет несколько генных сегментов вариабельной области, которые способны рекомбинироваться для формирования функционального белка легкой цепи. Локус легкой цепи λ человека содержит приблизительно 70 генных сегментов V и 7 пар генных сегментов Jλ-Cλ. По-видимому, только четыре из этих пар генных сегментов Jλ-Cλ являются функциональными. В некоторых аллелях пятая пара генных сегментов Jλ-Cλ, по некоторым сведениям, является псевдогеном (Cλ6). По-видимому, данные 70 генных сегментов Vλ содержат 38 функциональных генных сегментов. Данные 70 последовательностей Vλ расположены в трех кластерах, каждый из которых содержит различные члены определенных групп семейства генов V (кластеры А, В и С; фиг.1). Это является потенциально богатым источником относительно неиспользованного разнообразия для получения антител с V-областями человека животными, не являющимися человеком.
В существенном контрасте, локус легкой цепи λ мыши содержит только два или три (в зависимости от штамма) генных сегмента области Vλ мыши (фиг. 2). По меньшей мере по этой причине выраженное κ-смещение у мышей, как считается, не является чрезвычайно вредным для общего разнообразия антител.
Согласно опубликованным картам локуса легкой цепи λ мыши, локус состоит, по существу, из двух кластеров генных сегментов в пределах участка приблизительно 200 т.п.о. (фиг. 2). Данные два кластера включают два набора генов V, J, и C, которые способны к независимой перегруппировке: Vλ2-Jλ2-Cλ2-Jλ4-Cλ4 и Vλ1-Jλ3-Cλ3-Jλ1-Cλ1. Хотя было установлено, что Vλ2 рекомбинируется со всеми генными сегментами Jλ, Vλ1, по-видимому, рекомбинируется исключительно с Сλ1. Считается, что Cλ4 представляет собой псевдоген с дефектами участков сплайсинга.
Локус легкой цепи κ мыши сильно отличается. Структура и количество генных сегментов, которые принимают участие в рекомбинационных событиях, приводящих к формированию функционального белка легкой цепи из локуса κ мыши, являются более сложными (фиг. 3). Таким образом, легкие цепи λ мыши не вносят большого вклада в разнообразие популяции антител у обычной мыши.
Использование богатого разнообразия локуса легкой цепи λ человека у мыши вероятно приведет к образованию, среди прочего, источника более полного спектра доменов V легкой цепи человека. В предыдущих попытках использовать это разнообразие использовали трансгены человека, содержащие фрагменты локуса легкой цепи λ человека, случайным образом введенные в геном мыши (См., например, патент США № 6998514 и патент США № 7435871). Мыши, содержащие эти случайным образом интегрированные трансгены, по имеющимся данным, экспрессируют полностью легкие цепи λ человека, однако, в некоторых случаях, один или оба эндогенных локуса легкой цепи остаются интактными. Эта ситуация является нежелательной, поскольку последовательности легкой цепи λ человека конкурируют с легкой цепью мыши (κ или λ) в экспрессируемом спектре антител мыши.
В отличие от этого, авторы настоящего изобретения описывают генетически модифицированных мышей, которые способны экспрессировать одну или несколько последовательностей нуклеиновых кислот легкой цепи λ непосредственно из локуса легкой цепи мыши, в том числе, посредством замены в эндогенном локусе легкой цепи мыши. Генетически модифицированные мыши, способные экспрессировать последовательности легкой цепи λ человека из эндогенного локуса, далее могут быть скрещены с мышами, которые содержат локус тяжелой цепи человека, и таким образом могут быть использованы для экспрессии антител, содержащих области V (тяжелые и легкие), которые являются полностью человеческими. В различных вариантах осуществления области V экспрессируются с константными областями мыши. В различных вариантах осуществления не присутствуют эндогенные генные сегменты иммуноглобулина мыши, и области V экспрессируются с константными областями человека. Эти антитела могут оказаться полезными в многообразных практических применениях, и в диагностических, и в терапевтических.
Многочисленные преимущества могут быть реализованы для различных вариантов осуществления экспрессии связывающих белков, происходящих из генных сегментов Vλ и Jλ человека в мыши. Преимущества могут быть реализованы в результате помещения последовательностей λ человека в эндогенный локус легкой цепи, например, локус κ или λ мыши. Антитела, продуцированные такими мышами, могут иметь легкие цепи, которые содержат домены Vλ человека, слитые с областью CL мыши, особенно, областью Сκ или Сλ мыши. Мыши также будут экспрессировать домены Vλ человека, которые подходят для идентификации и клонирования для использования с областями CL человека, особенно, областями Сκ и/или Сλ. Поскольку развитие В-клеток у таких мышей, в остальном, нормальное, возможно образование совместимых доменов Vλ (в том числе, соматически видоизмененных доменов Vλ) в рамках либо Сλ, либо Сκ области.
Описаны генетически модифицированные мыши, которые содержат неперегруппированный генный сегмент Vλ в локусе легкой цепи κ или λ иммуноглобулина. Описаны мыши, которые экспрессируют антитела, содержащие легкую цепь, имеющую домен Vλ человека, слитый с областью Сκ и/или Сλ.
Стерильные транскрипты локуса легкой цепи κ иммуноглобулина
Вариации характерных черт экспрессии последовательностей λ иммуноглобулина человека нашли свое отражение в различных вариантах генетически модифицированных мышей, способных к такой экспрессии. Таким образом, в некоторых вариантах осуществления, данные генетически модифицированные мыши содержат определенную некодирующую последовательность (последовательности) из локуса человека. В одном из вариантов осуществления генетически модифицированная мышь содержит генные сегменты Vλ и Jλ человека в эндогенном локусе легкой цепи κ, и дополнительно содержит геномный фрагмент легкой цепи κ человека. В определенном варианте осуществления геномный фрагмент легкой цепи κ человека представляет собой некодирующую последовательность, встречающуюся в природе между генным сегментом Vκ человека и генным сегментом Jκ человека.
Локусы легкой цепи κ человека и мыши содержат последовательности, которые кодируют стерильные транскрипты, в которых отсутствуют либо стартовый кодон, либо открытая рамка считывания, и которые рассматриваются в качестве элементов, которые регулируют транскрипцию локусов легкой цепи κ. Эти стерильные транскрипты происходят из межгенной последовательности, расположенной в 5'-3' направлении, или 3', от наиболее проксимального генного сегмента Vκ и в направлении 3'-5', или 5', от интронного энхансера легкой цепи κ (Eκi), который находится в направлении 3'-5' гена константной области (Сκ) легкой цепи κ. Стерильные транскрипты возникают в результате перегруппировки межгенной последовательности для формирования сегмента VκJκ1, слитого с Сκ.
Замена локуса легкой цепи κ в направлении 3'-5' от гена Сκ удалит межгенную область, кодирующую стерильные транскрипты. Следовательно, в различных вариантах осуществления, замена последовательности легкой цепи κ мыши в направлении 3'-5' от гена Сκ мыши на генные сегменты легкой цепи λ человека будет иметь своим результатом гуманизированный локус легкой цепи κ мыши, который содержит генные сегменты Vλ и Jλ человека, но не межгенную область легкой цепи κ, которая кодирует стерильные транскрипты.
Как описано в настоящем описании, гуманизация эндогенного локуса легкой цепи κ мыши с помощью генных сегментов легкой цепи λ человека, где посредством гуманизации удаляется межгенная область, приводит в результате к существенному снижению использования локуса легкой цепи κ, в сочетании со значительным повышением использования легкой цепи λ. Следовательно, несмотря на то, что гуманизированная мышь, которая лишена межгенной области, является полезной тем, что может продуцировать антитела с вариабельными доменами легкой цепи человека (например, домены λ или κ человека), применимость локуса снижена.
Кроме того, описано гуманизация эндогенного локуса легкой цепи κ мыши с помощью генных сегментов Vλ и Jλ человека в сочетании с инсерцией межгенной области κ человека для получения локуса Vλ, который содержит в отношении транскрипции, между конечным генным сегментом Vλ человека и первым генным сегментом Jλ человека, межгенную область κ; который демонстрирует популяцию В-клеток с более высокой экспрессией, чем локус, в котором отсутствует межгенная область κ. Это наблюдение находится в согласии с гипотезой о том, что межгенная область - непосредственно через стерильный транскрипт, или опосредованно, - подавляет использование из эндогенного локуса легкой цепи λ. В рамках такой гипотезы, включение межгенной области в результате приведет к снижению использования эндогенного локуса легкой цепи λ, оставляя мыши ограниченный выбор, но используя модифицированнй локус (λ в κ) для получения антител.
В различных вариантах осуществления замена последовательности легкой цепи κ мыши в направлении 3'-5' от гена Сκ мыши на последовательность легкой цепи λ человека дополнительно содержит межгенную область легкой цепи κ человека, расположенную, в отношении транскрипции, между 3'-нетранслируемой областью наиболее 3' генного сегмента Vλ и 5' по отношению к первому генному сегменту Jλ человека. Альтернативно, такая межгенная область может быть исключена из замены эндогенного локуса легкой цепи κ (в направлении 3'-5' от гена Сκ мыши) в результате создания делеции в эндогенном локусе легкой цепи λ. Таким же образом, в рамках этого варианта осуществления, мышь продуцирует антитела из эндогенного локуса легкой цепи κ, содержащего последовательности легкой цепи λ человека.
Подходы к конструированию мышей для экспрессии доменов Vλ человека
Описаны различные подходы к получения генетически модифицированных мышей, которые продуцируют антитела, содержащие легкую цепь, которая включает домен Vλ человека, слитый с эндогенной областью CL (например, Сκ или Сλ). Описаны генетические модификации, которые, в различных вариантах осуществления, включают делецию одного или обоих локусов эндогенной легкой цепи. Например, для удаления легких цепей λ мыши из эндогенного спектра антител можно осуществить делецию первого кластера гена Vλ-Jλ-Cλ и замену, полностью или частично, генных сегментов Vλ-Jλ второго кластера гена на генные сегменты Vλ-Jλ человека. Также предусмотрены эмбрионы генетически модифицированных мышей, клетки, и направленные конструкции для получения таких мышей, эмбрионов мышей и клеток.
Делеция одного эндогенного кластера гена Vλ-Jλ-Cλ и замена генных сегментов Vλ-Jλ другого эндогенного кластера гена Vλ-Jλ-Cλ задействуют относительно минимальное нарушение в ассоциации природной константной области антитела и функции у данного животного, в различных вариантах осуществления, поскольку эндогенные гены Сλ остаются интактными и, следовательно, сохраняют нормальную функциональность и способность к ассоциации с константной областью эндогенной тяжелой цепи. Таким образом, в таких вариантах осуществления модификация не затрагивает другие константные области эндогенной тяжелой цепи, находящиеся под воздействием функциональных константных областей легкой цепи для сборки функциональной молекулы антитела, содержащей две тяжелые цепи и две легкие цепи. Более того, в различных вариантах осуществления модификация не затрагивает сборку функциональной мембранно-связанной молекулы антитела, с участием эндогенной тяжелой цепи и легкой цепи, например, домен hVλ, связанный с областью Сλ мыши. Поскольку, по меньшей мере, один функциональный ген Сλ сохраняется в эндогенном локусе, животные, содержащие замену генных сегментов Vλ-Jλ эндогенного кластера гена Vλ-Jλ-Cλ на генные сегменты Vλ-Jλ человека, должны быть способны продуцировать нормальные легкие цепи λ, которые способны связывать антиген во время иммунного ответа через генные сегменты Vλ-Jλ, присутствующие в экспрессируемом спектре антител данного животного.
Схематическое изображение (без соблюдения масштаба) делетированного эндогенного кластера гена Vλ-Jλ-Cλ мыши представлено на фиг. 2. Как показано на данном изображении, локус легкой цепи λ мыши упорядочен в два кластера гена, оба содержат функциональные генные сегменты, способные к рекомбинации для создания функциональной легкой цепи λ мыши. Эндогенный кластер генов Vλ1-Jλ3-Cλ3-Jλ1-Cλ1 мыши делетирован с помощью направленной конструкции (Направленный вектор 1) с неомициновой кассетой, фланкированной центрами рекомбинации. Другой эндогенный кластер генов (Vλ2-Vλ3-Jλ2-Cλ2-Jλ4-Cλ4) делетирован частично с помощью направленной конструкции (Направленный вектор 2) с гигромицин-тимидин киназной кассетой, фланкированной центрами рекомбинации. В этом втором случае нацеливания эндогенные генные сегменты Cλ2-Jλ4-Cλ4 сохраняются. Вторая направленная конструкция (направленный вектор 2) сконструирована с использованием участков рекомбинации, отличающихся от тех, которые используются в первой направленной конструкции (направленный вектор 1), там самым обеспечивается селективная делеция селекционной кассеты после успешного осуществления нацеливания. Полученный двусторонне-целевой локус является функционально выключенным, в котором не может продуцироваться эндогенная легкая цепь λ. Данный модифицированный локус можно использовать для инсерции генных сегментов Vλ и Jλ человека для получения эндогенного локуса λ мыши, содержащего генные сегменты Vλ и Jλ человека, посредством чего, после рекомбинации в модифицированном локусе, животное продуцирует легкие цепи λ, содержащие перегруппированные генные сегменты Vλ и Jλ человека, связанные с эндогенным генным сегментом Сλ мыши.
Генетическое модифицирование мыши для получения эндогенных генных сегментов λ нефункциональными, в различных вариантах осуществления, приводит к получению мыши, которая продуцирует исключительно легкие цепи κ в своем спектре антител, что делает такую мышь подходящей для оценки роли легких цепей λ в иммунном ответе, и которые подходят для получения спектра антител, содержащих домены Vκ, но не домены Vλ.
Генетически модифицированную мышь, которая экспрессирует hVλ, связанный с геном Сλ мыши, рекомбинированные в эндогеном локусе легкой цепи λ мыши, можно получить, используя любой способ, известный в данной области. Схематическое изображение (без соблюдения масштаба) замены эндогенных генных сегментов Vλ2-Vλ3-Jλ2 мыши на генные сегменты Vλ и Jλ человека представлено на фиг. 4А. Как показано на данном изображении, эндогенный локус легкой цепи λ мыши, который был воссоздан нефункциональным, заменен с помощью направленной конструкции (направленный вектор 12/1-λ), которая включает неомициновую кассету, фланкированную участками рекомбинации. Генные сегменты Vλ2-Vλ3-Jλ2 заменены на геномный фрагмент, содержащий последовательность λ человека, которая включает 12 генных сегментов hVλ и единственный генный сегмент hJλ.
Таким образом, этот первый подход располагает один или несколько генных сегментов hVλ в эндогенном локусе легкой цепи λ, смежно с единственным генным сегментом hJλ (фиг. 4А).
Дополнительные модификации в модифицированном эндогенном локусе легкой цепи λ могут быть обеспечены с использованием аналогичных технологий для инсерции большего числа генных сегментов hVλ. Например, на Фиг. 5А представлено схематическое изображение двух дополнительных направленных конструкций (направленные векторы +16-λ и +12-λ), использованных для прогрессивной вставки дополнительных генных сегментов hVλ человека. Как представлено на данном изображении, дополнительные геномные фрагменты, содержащие определенные генные сегменты hVλ человека, вставлены в модифицированный эндогенный локус легкой цепи λ на последовательных этапах, с использованием гомологии, обеспеченной предыдущей вставкой последовательностей легкой цепи λ человека. После рекомбинации с каждой проиллюстрированной направленной конструкцией, в последовательной манере, 28 дополнительных генных сегментов hVλ были встроены в модифицированный эндогенный локус легкой цепи λ. В результате этого создан химерный локус, который продуцирует белок легкой цепи λ, который содержит генные сегменты Vλ-Jλ человека, соединенные с геном Сλ мыши.
Описанные выше способы вставки генных сегментов легкой цепи λ человека в локус λ мыши сохраняют энхансеры, расположенные в 5'-3' направлении от генных сегментов Cλ2-Jλ4-Cλ4 (обозначенные Enh2.4, Enh и Enh3.1 на фиг. 4А и фиг. 5А). Такой подход в результате приводит к единственному модифицированному аллелю в эндогенном локусе легкой цепи λ мыши (фиг. 7А).
Изобретение также относится к композициям и способам получения мыши, которая экспрессирует легкую цепь, содержащую генные сегменты hVλ и Jλ, функционально связанные с генным сегментом Сλ мыши, в том числе к композициям и способам получения мыши, которая экспрессирует такие гены из эндогенного локуса легкой цепи λ мыши. Способы включают выборочное приведение в нефункциональное состояние одного эндогенного кластера гена Vλ-Jλ-Cλ мыши (например, посредством направленной делеции), и применение генных сегментов hVλ и Jλ в эндогенном локусе легкой цепи λ мыши для экспрессии домена hVλ у мыши.
Альтернативно, во втором способе, генные сегменты легкой цепи λ человека могут быть расположены в эндогенном локусе легкой цепи κ. Генетическая модификация, в различных вариантах осуществления, включает делецию эндогенного локуса легкой цепи κ. Например, для того, чтобы удалить легкие цепи κ мыши из эндогенного спектра антител, может быть осуществлена делеция генных сегментов Vκ и Jκ мыши. Также предусмотрены генетически модифицированные эмбрионы мыши, клетки и направленные конструкции для получения мышей, эмбрионов мышей и клеток.
По изложенным выше причинам, делеция генных сегментов Vκ и Jκ мыши осуществляется с относительно минимальным ущербом. Схематическое изображение (без соблюдения масштаба) делетированных генных сегментов Vκ и Jκ мыши представлено на фиг. 3. Эндогенные генные сегменты Vκ и Jκ мыши удалены посредством опосредованной рекомбиназами делеции последовательностей мыши, расположенных между двумя точно расположенными направленными векторами, каждый из которых содержит сайт-специфические участки рекомбинации. Первый направленный вектор (направленный вектор Jκ) используется в первом случае нацеливания для удаления генных сегментов Jκ мыши. Второй направленный вектор (Направленный вектор Vκ) используется во втором случае нацеливания для удаления последовательности, расположенной в 5'-направлении от наиболее удаленного генного сегмента Vκ мыши. Оба направленных вектора содержат сайт-специфические участки рекомбинации, что делает возможным выборочное удаление обеих селекционных кассет и всех промежуточных последовательностей легкой цепи κ мыши после успешного осуществления нацеливания на мишень. Полученный в результате делетированный локус является функционально “молчащим” в том смысле, что эндогенная легкая цепь κ не может быть получена. Этот модифицированный локус можно использовать для инсерции генных сегментов hVλ и Jλ для создания эндогенного локуса κ мыши, содержащего генные сегменты hVλ и Jλ, посредством чего, после рекомбинации в модифицированном локусе, животное продуцирует легкие цепи λ, содержащие перегруппированные генные сегменты hVλ и Jλ, функционально связанные с эндогенным генным сегментом Сκ мыши. Различные направленные векторы, содержащие последовательноси легкой цепи λ человека, можно использовать в сочетании с этим делетированным локусом κ мыши, для создания гибридного локуса легкой цепи, содержащего генные сегменты λ человека, функционально связанные с областью Сκ мыши.
Таким образом, второй подход фиксирует один или несколько генных сегментов Vλ человека, расположенных в локусе легкой цепи κ мыши сопредельно с единственным генным сегментом Jλ человека (направленный вектор 12/1-κ, фиг. 4B).
В различных вариантах осуществления модификации данного подхода могут быть проведены для добавления генных сегментов и/или регуляторных последовательностей для оптимизации использования последовательностей легкой цепи λ человека из локуса легкой цепи κ мыши в пределах спектра антител мыши.
В третьем подходе один или несколько генных сегментов hVλ расположены в локусе легкой цепи κ мыши, смежном с четырьмя генными последовательностями hJλ (направленный вектор 12/4-κ, фиг. 4B).
В третьем подходе один или несколько генных сегментов hVλ расположены в локусе легкой цепи κ мыши, смежном с межгенной последовательностью κ человека и единственной генной последовательностью hJλ (направленный вектор 12(κ)1-κ, фиг. 4B).
В четвертом способе один или несколько генных сегментов hVλ помещены в локус легкой цепи κ мыши, смежный с межгенной последовательностью κ человека и четырьмя генными последовательностями hJλ (направленный вектор 12(κ)4-κ, фиг. 4B).
Все вышеописанные подходы вставки генных сегментов легкой цепи λ человека в локус κ мыши сохраняют элемент интронного энхансера κ в направлении 3'-5' от гена Сκ (обозначенный Eκi, Фиг. 4В и Фиг. 5В) и 3' энхансер κ в направлении 5'-3' от гена Сκ (обозначен Еκ3', Фиг. 4В и Фиг. 5В). В результате данного подхода получают четыре отдельных модифицированных аллеля в эндогенном локусе легкой цепи κ мыши (фиг. 7B).
В различных вариантах осуществления генетически модифицированная мышь содержит “нокаут” локус эндогенной легкой цепи λ мыши. В одном из вариантов осуществления локус легкой цепи λ выключен с помощью метода, в котором удаляется область, простирающаяся от Vλ2 до Jλ2, и область, простирающаяся от Vλ1 до Cλ1 (фиг. 2). Любые стратегии, в результате которых снижается или исключается способность эндогенного локуса легкой цепи λ экспрессировать эндогенные домены λ, подходят для использования с вариантами осуществления настоящего изобретения.
Антитела, содержащие домен лямбда, полученные от генетически модифицированных мышей
Мыши, содержащие последовательности λ человека либо в локусе легкой цепи κ, либо в локусе легкой цепи λ, будут экспрессировать легкую цепь, которая содержит область hVλ, слитую с областью CL (Сκ или Сλ) мыши. Эти мыши успешно скрещиваются с мышами, которые (a) содержат функционально “молчащий” локус легкой цепи (например, нокаут эндогенный локус легкой цепи κ или λ мыши); (b) содержат эндогенный локус легкой цепи λ мыши, который содержит генные сегменты hV и hJ, функционально связанные с эндогенным геном Сλ мыши; (с) содержат эндогенный локус легкой цепи κ мыши, который включает генные сегменты hVκ и hJκ, функционально связанные с эндогенным геном Сκ мыши; и, (d) мышь, у которой один аллель κ содержит hVκ и hJκ; другой аллель κ содержит hVλ и hJλ; один аллель λ содержит hVλ и hJλ, и другой аллель λ является молчащим или выключенным, или оба аллеля λ содержат hVλ и hJλ; и два аллеля тяжелой цепи, каждый из которых содержат hVH, hDH и hJH.
Антитела, которые содержат домены hVλ, экспрессируемые либо в области Сκ, либо в области Сλ, используют для получения полностью человеческих антител с помощью клонирования нуклеиновых кислот, кодирующих домены hVλ в экспрессионные конструкции, которые несут гены, кодирующие Сλ человека. Полученные экспрессионные конструкции трансфицируют в подходящие клетки-хозяева для экспрессии антител, которые демонстрируют полностью человеческий домен hVλ, слитый с hCλ.
ПРИМЕРЫ
Следующие примеры предоставлены для того, чтобы описать, как получать и применять способы и композиции по настоящему изобретению, и они не предназначены для ограничения объема настоящего изобретения. Если не оговорено иначе, температура указана в градусах по Цельсию и давление является близким к атмосферному.
Пример I
Делеция локусов легкой цепи иммуноглобулина мыши
Различные конструкции направленного действия были созданы с помощью технологии VELOCIGENE® (см., например, патент США № 6586251 и Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6):652-659) для модификации геномных библиотек Бактериальной Искусственной Хромосомы (BAC) мыши для того, чтобы инактивировать локусы легкой цепи κ и λ мыши.
Делеция локуса легкой цепи λ мыши. ДНК клона RP23-135k15 ВАС мыши (Invitrogen) была модифицирована посредством гомологичной рекомбинации для инактивации эндогенного локуса легкой цепи λ мыши в результате направленной делеции кластера гена Vλ-Jλ-Cλ (фиг. 2).
Кратко, весь проксимальный кластер, содержащий генные сегменты Vλ1-Jλ3-Cλ3-Jλ1-Cλ1, был делетирован в ходе единственного случая нацеливания с использованием направленного вектора, содержащего неомициновую кассету, фланкированную сайтами loxP, с 5' на гомологичное плечо мыши, содержащее 5'-последовательность генного сегмента Vλ1, и на 3' гомологичное плечо мыши, содержащим 3'-последовательность генного сегмента Сλ1 (фиг. 2, Направленный вектор 1).
Вторая нацеливающая конструкция была подготовлена для точной делеции дистального эндогенного кластера гена λ мыши, содержащего Vλ2-Jλ2-Cλ2-Jλ4-Cλ4, за исключением того, что нацеливающая конструкция содержала 5' гомологичное плечо мыши, содержащее последовательность 5' генного сегмента Vλ2, и 3' гомологичное плечо мыши, которое содержало последовательность 5' к эндогенному генному сегменту Сλ2 (фиг. 2, направленный вектор 2). Таким образом, вторая нацеливающая конструкция точно удаляет Vλ2-Jλ2, оставляя Cλ2-Jλ4-Cλ4 интактным в эндогенном локусе λ мыши. Присутствие клеток ES, содержащих инактивированный эндогенный локус λ (как описано выше), было подтверждено с использованием способов кариотипирования и скрининга (например, TAQMAN®), известных в уровне техники. Затем, ДНК была выделена из модифицированных клеток ES и подвержена обработке рекомбиназой CRE, тем самым опосредуя делецию проксимальной направленной кассеты, содержащей маркерный ген неомицина, оставляя только единственный участок loxP в точке делеции (фиг. 2, внизу).
Делеция локуса легкой цепи κ мыши. Несколько нацеливающих конструкций были созданы с использованием методов, аналогичных описанным выше, для модификации ДНК клонов ВАС мыши RP23-302g12 и RP23-254m04 (Invitrogen) посредством гомологичной рекомбинации для инактивации локуса легкой цепи κ мыши в двухстадийном процессе (Фиг. 3).
Кратко, генные сегменты Jκ (1-5) эндогенного локуса легкой цепи κ мыши были удалены в ходе единственного случая нацеливания с использованием направленного вектора, содержащего гигромицин-тимидинкиназную кассету (hyg-TK), содержащую единственный сайт loxP 3' к кассете hyg-TK (фиг. 3, направленный вектор Jκ). Гомологичные плечи, использованные для создания этого направленного вектора, содержат геномную последовательность мыши 5' и 3' эндогенных генных сегментов Jκ мыши. Во втором случае нацеливания второй направленный вектор был подготовлен для удаления части геномной последовательности мыши в направлении 3'-5' (5') к наиболее дистальному эндогенному генному сегменту Vκ мыши (фиг. 3, направленный вектор Vκ). Этот направленный вектор содержал инвертированный сайт lox511, сайт loxP и неомициновую кассету. Гомологичные плечи, использованные для создания этого направленного вектора, содержали геномную последовательность мыши в направлении 3'-5' от наиболее дистального генного сегмента Vκ мыши. Направленные векторы использовали в последовательной манере (а именно, сначала Jκ, затем Vκ) в ДНК-мишени в клетках ES. Наличие ES, несущих двусторонне-нацеленную хромосому (а именно, единственный эндогенный локус κ мыши, нацеленный обоими направленными векторами), было подтверждено с помощью методов кариотипирования и скрининга (например, Taqman™), известных в данной области. Затем ДНК выделяли из модифицированных клеток ES и подвергали обработке рекомбиназой Cre, тем самым опосредуя делецию эндогенных генных сегментов Vκ мыши и обеих селективных кассет, оставляя два объединенных lox-сайта в противоположной ориентации относительно друг друга (фиг. 3, внизу; SEQ ID NO:1).
Таким образом, два модифицированных эндогенных локуса легкой цепи (κ и λ), содержащие интактный энхансер и константные области, были созданы для прогрессивной инсерции неперегруппированных генных сегментов эмбрионального типа λ человека в точно определенной манере с использованием направленных векторов, описанных ниже.
Пример II
Замена локусов легкой цепи мыши на мини-локус легкой цепи λ человека
Множественные направленные векторы были сконструированы для прогрессивной инсерции генных сегментов λ человека в эндогенные локусы легкой цепи κ и λ мыши с использованием методов, аналогичных описанным выше. Множественные независимые первичные модификации были проделаны с эндогенными локусами легкой цепи, каждая приводит к образованию химерного локуса легкой цепи, содержащего генные сегменты hVλ и Jλ, оперативно связанные с константными генами и энхансерами легкой цепи мыши.
Мини-локус λ человека, содержащий 12 генных сегментов Vλ человека и один генный сегмент Jλ человека. Серии первичных направленных векторов были сконструированы таким образом, чтобы содержать первые 12 последовательных генных сегментов Vλ человека из кластера А и генный сегмент hJλ1, или четыре генных сегмента hJλ, используя клон ВАС человека, называемый RP11-729g4 (Invitrogen). На фиг. 4А и 4В представлены направленные векторы, которые были сконструированы для осуществления начальной инсерции генных сегментов легкой цепи λ человека в локусах легкой цепи λ и κ мыши, соответственно.
Для первого комплекта первичных направленных векторов фрагмент ДНК, размером 124125 п.о., из клона ВАС 729g4, содержащий 12 генных сегментов hVλ и генный сегмент hJλ1, был сконструирован таким образом, чтобы содержать участок PI-SceI, 996 п.о. в направлении 5'-3' (3') от генного сегмента hJλ1, для лигирования 3' гомологичного плеча мыши. Два различных комплекта гомологичных плеч использовали для лигирования с этим фрагментом человека; один комплект гомологичных плеч содержал эндогенные последовательности λ мыши из клона ВАС 135k15 (фиг. 4А), и другой комплект содержал эндогенную последовательность κ 5' и 3' генных сегментов Vκ и Jκ мыши из клонов ВАС мыши RP23-302g12 и RP23-254m04, соответственно (фиг. 4В).
Для направленного вектора 12/1-λ (Фиг. 4А) был сконструирован сайт PI-SceI на 5'-конце ДНК-фрагмента размером 27847 п.о., содержащем Cλ2-Jλ4-Cλ4 мыши и энхансер 2.4 модифицированного локуса λ мыши, описанного в Примере 1. Фрагмент мыши, размером ~28 т.п.о., использовали в качестве 3' гомологичного плеча посредством лигирования с фрагментом λ человека размером ~124 т.п.о., что создавало 3'-соединение, содержащее от 5' до 3', генный сегмент hJλ1, 996 п.о. последовательности λ человека 3' от генного сегмента hJλ1, 1229 п.о. из последовательности λ мыши 5' к гену Сλ2 мыши, ген мыши Сλ2 и оставшуюся часть мышиного фрагмента ~28 т.п.о. В направлении 3'-5' (5') от генного сегмента Vλ3-12 человека была дополнительная последовательность λ человека, размером 1456 п.о., до начала гомологичного плеча мыши 5', которая содержала 23792 п.о. геномной ДНК мыши, соответствующей последовательности 5' эндогенного локуса λ мыши. Между гомологичным плечом 5' и началом последовательности λ человека находилась неомициновая кассета, фланкированная сайтами Frt.
Таким образом, направленный вектор 12/1-λ включал от 5' до 3', 5'-гомологичное плечо, содержащее ~24 т.п.о. геномной последовательности λ мыши 5' от эндогенного локуса λ, 5' сайт Frt, неомициновую кассету, 3' сайт Frt, ~123 т.п.о. геномной последовательности λ человека, содержащий первые 12 последовательных генных сегмента hVλ и генный сегмент hJλ1, сайт PI-SceI, и 3' гомологичное плечо, содержащее ~28 т.п.о. геномной последовательности мыши, в том числе эндогенные генные сегменты Cλ2-Jλ4-Cλ4, последовательность энхансера 2.4 мыши и дополнительную геномную последовательность мыши в направлении 5'-3' (3') от энхансера 2.4 (фиг. 4А).
Аналогично, в направленном векторе 12/1-κ (Фиг. 4В) использовался тот же самый фрагмент λ человека размером ~124, за исключением того, что гомологичные плечи мыши, содержащие последовательность κ мыши, были использованы так, чтобы нацеливание на эндогенный локус κ могло быть достигнуто посредством гомологичной рекомбинации. Таким образом, направленный вектор 12/1-κ включал от 5' до 3', 5' гомологичное плечо, содержащее геномную последовательность мыши, размером ~23 т.п.о., 5' от эндогенного локуса κ, сайт I-CeuI, 5' сайт Frt, неомициновую кассету, 3' сайт Frt, ~124 т.п.о. геномной последовательности λ человека, содержащей первые 12 последовательных генных сегмента hVλ и генный сегмент hJλ1, сайт PI-SceI, и 3' гомологичное плечо, содержащее ~28 т.п.о. геномной последовательности мыши, в том числе, эндогенный ген Сκ мыши, Eκi и Eκ3' и дополнительную геномную последовательность мыши в направлении 5'-3' (3') от Еκ3' (фиг. 4В, направленный вектор 12/1-κ).
Гомологичная рекомбинация с любым из этих двух первичных направленных векторов приводит к образованию модифицированного локуса легкой цепи мыши (κ или λ), содержащего 12 генных сегментов hVλ и генный сегмент hJλ1, функционально связанных с эндогенным константным геном легкой цепи мыши и энхансерами (Cκ или Cλ2 и Eκi/Eκ3', или Enh2.4/Enh3.1), что после рекомбинации, приводит к формированию химерной легкой цепи λ.
Мини-локус λ человека с 12 генными сегментами Vλ человека и четырьмя генными сегментами Jλ человека. В другом способе, для того, чтобы добавить разнообразия химерному локусу легкой цепи λ, был сконструирован третий направленный вектор для инсерции первых 12 последовательных генных сегментов Vλ человека из кластера А, и генных сегментов hJλ1, 2, 3 и 7, в локус легкой цепи κ мыши (Фиг. 4В, Направленный вектор 12/4-κ). Сегмент ДНК, содержащий генные сегменты hJλ1, Jλ2, Jλ3 и Jλ7, был создан посредством синтеза ДНК de novo (Integrated DNA Technologies), включая каждый генный сегмент Jλ и геномную последовательность человека ~100 п.о. из обеих ближайших 5' и 3' областей каждого генного сегмента Jλ. Участок PI-SceI был сконструирован на 3'-конце этого фрагмента ДНК размером ~1 т.п.о., и лигирован с кассетой устойчивости к хлорамфениколу. Гомологичные плечи с помощью ПЦР были амплифицированы из последовательности λ человека в положениях 5' и 3' относительно генного сегмента hJλ1 клона ВАС 729g4 человека. Гомологичную рекомбинацию с этим промежуточным направленным вектором осуществляли на модифицированном клоне ВАС 729g4, который ранее был намечен мишенью в направлении 3'-5' (5') от генного сегмента Vλ3-12 человека с помощью неомициновой кассеты, фланкированной участками Frt, который также содержал участок I-CeuI в 5'-положении относительно 5'-участка Frt. двухнацеленный клон ВАС 729g4 включал от 5' до 3', участок I-CeuI, 5' участок Frt, неомициновую кассету, 3' участок Frt, фрагмент размером ~123 т.п.о., содержащий первые 12 генных сегментов hVλ, фрагмент размером ~1 т.п.о., содержащий генные сегменты Jλ1, 2, 3 и 7 человека, участок PI-SceI, и кассету устойчивости к хлорамфениколу. Этот промежуточный направленный вектор был расщеплен совместно с помощью I-CeuI и PI-SceI, и последовательно лигирован в модифицированный клон ВАС мыши (описан выше) для создания третьего направленного вектора.
В результате этого лигирования получают третий направленный вектор для инсерции последовательностей λ человека в эндогенный локус легкой цепи κ мыши, что включает от 5' до 3', 5' гомологичное плечо мыши, содержащее геномную последовательность размером ~23 т.п.о. 5' эндогенного локуса κ мыши, участок I-CeuI, 5' участок Frt, неомициновую кассету, 3' участок Frt, фрагмент размером ~123 т.п.о., содержащий первые 12 генных сегментов hVλ , фрагмент размером ~1 т.п.о, содержащий генные сегменты hJλ1, 2, 3 и 7, участок PI-SceI и 3' гомологичное плечо, содержащее геномную последовательность мыши размером ~28 т.п.о., в том числе эндогенный ген Сκ мыши, Eκi и Eκ3', и дополнительную эндогенную последовательность мыши в направлении 5'-3' (3') от Eκ3' (Фиг. 4В, направленный вектор 12/4-κ). В результате гомологичной рекомбинации с этим третьим направленным вектором получается модифицированный локус легкой цепи κ мыши, содержащий 12 генных сегментов hVλ и четыре генных сегмента hJλ, функционально связанных с эндогенным геном Сκ мыши, что, после рекомбинации, приводит к формированию химерной легкой цепи λ человека/κ мыши.
Мини-локус λ человека с интегрированной последовательностью легкой цепи κ человека. Аналогичным образом, два дополнительных направленных вектора, похожих на сконструированные для осуществления первичной инсерции генных сегментов λ человека в эндогенный локус легкой цепи κ (фиг. 4В, Направленный вектор 12/1-κ и 12/4-κ), были созданы для прогрессивной инсерции генных сегментов легкой цепи λ человека, с использованием сконструированных единственным образом направленных векторов, содержащих смежные геномные последовательности λ и κ человека. Эти направленные векторы были сконструированы таким образом, чтобы включали геномную последовательность κ человека размером ~23 т.п.о., по природе расположенную между генными сегментами Vκ4-1 и Jκ1. Эта геномная последовательность κ человека была специфически помещена в этих двух дополнительных направленных векторах между генными сегментами Vλ и Jλ человека (фиг. 4В, Направленные векторы 12(κ)1-κ и 12(κ)4-κ).
Оба направленных вектора, содержащих геномную последовательность κ человека, были созданы с использованием модифицированного клона ВАС RP11-729g4, описанного выше (фиг. 6). Этот модифицированный клон ВАС был нацелен с помощью спектиномициновой селекционной кассеты, фланкированной сайтами рестрикции NotI и AsiSI (фиг. 6, вверху слева). Гомологичная рекомбинация с использованием спектиномициновой кассеты приводит в результате к двусторонне нацеленному клону ВАС 729g4, который включает от 5' до 3', участок I-CeuI, 5' участок Frt, неомициновую кассету, 3' участок Frt, фрагмент размером ~123 т.п.о., содержащий первые 12 генных сегментов hVλ, сайт NotI приблизительно 200 п.о. в направлении 5'-3' (3') к девятичленной последовательности генного сегмента hVλ3-1, спектиномициновую кассету и AsiSI. Отдельный клон ВАС человека, содержащий последовательность κ человека (CTD-2366j12) был нацелен два раза, независимо друг от друга, для конструкции сайтов рестрикции в участках между генными сегментами hVκ4-1 и hJκ1, для обеспечения последующего клонирования фрагмента размером ~23 т.п.о. для лигирования с генными сегментами hVλ, содержащимися в двусторонне нацеленном модифицированном клоне ВАС 729g4 (фиг. 6, вверху справа).
Кратко, размер клона ВАС 2366j12 составляет около 132 т.п.о., и он содержит генные сегменты hVκ 1-6, 1-5, 2-4, 7-3, 5-2, 4-1, геномную последовательность κ человека в направлении 5'-3' от генных сегментов Vκ, генные сегменты hJκ 1-5, hCκ и приблизительно 20 т.п.о. дополнительной геномной последовательности локуса κ человека. Этот клон сначала был нацелен с помощью направленного вектора, содержащего гигромициновую кассету, фланкированную участками Frt и участком NotI в направлении 5'-3' (3') от 3'-участка Frt. Гомологичные плечи для этого направленного вектора содержали геномную последовательность 5' и 3' генных сегментов Vκ человека в пределах клона ВАС, так что после гомологичной рекомбинации с этим направленным вектором, генные сегменты Vκ были удалены и участок NotI был сконструирован ~133 п.о. в направлении 5'-3' от генного сегмента hVκ4-1 (фиг. 6, вверху справа). Этот модифицированный клон ВАС 2366j12 был нацелен независимо двумя направленными векторами на 3'-конце для удаления генных сегментов hJκ с кассетой хлорамфеникола, которые также содержали либо генный сегмент hJλ1, участок PI-SceI и участок AsiSI, или геномный фрагмент λ человека, содержащий четыре генных сегмента hJλ (выше), участок PI-SceI и участок AsiSI (фиг. 6, вверху справа). Гомологичные плечи для этих двух похожих направленных векторов содержали последовательность 5' и 3' генных сегментов hJκ. В результате гомологичной рекомбинации с этими вторыми направленными векторами и модифицированным клоном ВАС 2366j12 образуется двусторонне нацеленный клон 2366j12, который включает от 5' до 3', 5'-участок Frt, гигромициновую кассету, 3'-участок Frt, участок NotI, 22800 п.о. геномного фрагмента локуса κ человека, содержащего межгенную область между генными сегментами Vκ4-1 и Jκ1, либо генный сегмент hJλ1, либо геномный фрагмент λ человека, содержащий hJλ1, Jλ2, Jλ3 и Jλ7, участок PI-SceI и кассету хлорамфеникола (фиг. 6, вверху справа). Два последних направленных вектора для осуществления двух дополнительных модификаций, были получены в двух этапах лигирования с использованием двусторонне нацеленных клонов 729g4 и 2366j12.
Двусторонне нацеленные клоны 729g4 и 2366j12 были расщеплены с помощью NotI и AsiSI, с образованием одного фрагмента, содержащего неомициновую кассету и генные сегменты hVλ, и другого фрагмента, содержащего ~23 т.п.о. геномного фрагмента локуса κ человека, содержащего межгенную область между генными сегментами Vκ4-1 и Jκ1, либо генный сегмент hJλ1, либо геномный фрагмент, содержащий генные сегменты hJλ1, Jλ2, Jλ3 и Jλ7, участок PI-SceI и кассету хлорамфеникола, соответственно. В результате лигирования этих фрагментов получали два уникальных клона ВАС, содержащих от 5' до 3', генные сегменты hVλ, геномную последовательность κ человека между генными сегментами Vκ4-1 и Jκ1, либо генный сегмент hJλ1, либо геномный фрагмент, содержащий генные сегменты hJλ1, Jλ2, Jλ3 и Jλ7, участок PI-SceI и кассету хлорамфеникола (фиг. 6, внизу). Эти новые клоны ВАС затем были расщеплены с помощью I-CeuI и PI-SceI, для высвобождения уникальных фрагментов, содержащих неомициновую кассету в направлении 3'-5' и смежные последовательности λ и κ человека, и лигированы в модифицированный клон ВАС 302g12 мыши, который содержал от 5'-3', геномную последовательность 5' эндогенного локуса κ мыши, участок I-CeuI, 5' участок Frt, неомициновую кассету, 3' участок Frt, генные сегменты hVλ (от 3-12 до 3-1), участок NotI ~200 п.о. в направлении 5'-3' от Vλ3-1, ~23 т.п.о. последовательности κ человека, в природе обнаруженную между генными сегментами Vκ4-1 и Jκ1 человека, либо генный сегмент hJλ1 или геномный фрагмент, содержащий генные сегменты hJλ1, Jλ2, Jλ3 и Jλ7, Eκi мыши, ген Cκ мыши и Eκ3' (фиг. 4, Направленные векторы 12hVλ-VκJκ-hJλ1 и 12hVλ-VκJκ4hJλ). Гомологичная рекомбинация с обоими этими направленными векторами создает в результате два отдельных модифицированных локуса легкой цепи κ мыши, содержащий 12 генных сегментов hVλ, геномную последовательность κ человека, и либо один, либо четыре генных сегмента hJλ, функционально связанных с эндогенным геном Сκ мыши, что, после рекомбинации, приводит к формированию химерной легкой цепи λ человека/κ мыши.
Пример III
Конструирование дополнительных генных сегментов Vλ человека в мини-локусе легкой цепи λ человека
Дополнительные генные сегменты hVλ были добавлены независимо к каждой из первоначальных модификаций, описанных в Примере 2, с использованием аналогичных направленных векторов и способов (фиг. 5A, Направленный вектор +16-λ, и фиг. 5B, Направленный вектор +16-κ).
Введение 16 дополнительных генных сегментов Vλ человека. Расположенные в 3'-5' направлении (5') гомологичные плечи использовали при конструировании направленных векторов для добавления 16 дополнительных генных сегментов hVλ к модифицированным локусам легкой цепи, описанным в Примере 2, содержащим 5' геномную последовательность мыши либо эндогенных локусов легкой цепи κ, либо эндогенных локусов легкой цепи λ. 3' гомологичные плечи были одинаковыми для всех направленных векторов и содержали геномные последовательности человека, перекрывающиеся с 5'-концом последовательности λ человека модификаций, как описано в Примере 2.
Коротко, два направленных вектора были сконструированы для введения 16 дополнительных генных сегментов hVλ в модифицированные локусы легкой цепи мыши, описанные в Примере 2 (фиг. 5A и 5B, Направленный вектор +16-λ или +16-κ). Фрагмент ДНК размером ~172 т.п.о. из клона ВАС RP11-761I13 человека (Invitrogen), содержащий 21 последовательных генных сегментов hVλ из кластера А, был сконструирован с 5' гомологичным плечом, содержащим 5' геномную последовательность мыши либо эндогенных локусов легкой цепи κ или λ мыши и 3' гомологичное плечо, содержащее геномную последовательность λ человека. 5' гомологичные плечи κ или λ мыши, использованные в этих нацеливающих конструкциях, были теми же 5' гомологичными плечами, что и описанные в Примере 2 (фиг. 5A и 5B). 3' гомологичное плечо включало перекрывающийся участок 53057 п.о. геномной последовательности λ человека, соответствующей эквивалентному 5'-концу фрагмента размером ~123 т.п.о. геномной последовательности λ человека, описанной в Примере 2. Эти два направленных вектора содержали от 5' до 3', 5' гомологичное плечо мыши, содержащее либо ~23 т.п.о. геномной последовательности 5' эндогенного локуса легкой цепи κ мыши, либо ~24 т.п.о. геномной последовательности мыши 5' эндогенного локуса легкой цепи λ, 5' участок Frt, гигромициновую кассету, 3' участок Frt и 171457 п.о. геномной последовательности λ человека, включающей 21 последовательных генных сегментов hVλ, ~53 т.п.о. которых перекрываются с 5'-концом последовательности λ человека, описанной в Примере 3 и выполняют функцию 3' гомологичного плеча для этой нацеливающей конструкции (фиг. 5А и 5В, Направленные векторы +16-λ или +16-κ). В результате гомологичной рекомбинации с этими направленными векторами получают независимо модифицированные локусы легкой цепи κ и λ мыши, каждый содержащий 28 генных сегментов hVλ и генный сегмент hJλ1, функционально связанные с эндогенными константными генами мыши (Сκ или Сλ2), что после рекомбинации, приводит к формированию химерной легкой цепи.
Аналогичным образом, направленный вектор +16-κ также использовали для введения 16 дополнительных генных сегментов hVλ в другие первичные модификации, описанные в Примере 2, которые объединяют множественные генные сегменты hJλ с, или без, интегрированной последовательностью κ человека (Фиг. 4B). Гомологичная рекомбинация с этим направленным вектором в эндогенном локусе κ мыши, содержащем другие первичные модификации, создает локусы легкой цепи κ мыши, содержащие 28 генных сегментов hVλ и генные сегменты hJλ1, 2, 3 и 7 с, или без, геномной последовательностью Vκ-Jκ человека, функционально связанные с эндогенным геном Сκ мыши, что после рекомбинации, приводит к формированию химерной легкой цепи λ-κ.
Введение 12 дополнительных генных сегментов Vλ человека. Дополнительные генные сегменты hVλ были добавлены независимо в каждую из модификаций, описанных выше, с использованием аналогичных направленных векторов и методов. Конечные структуры локусов, полученные в результате гомологичной рекомбинации с направленными векторами, содержащие дополнительные генные сегменты hVλ, представлены на Фиг. 7А и 7В.
Кратко, направленный вектор был сконструирован для введения 12 дополнительных генных сегментов hVλ в модифицированные локусы легкой цепи κ и λ мыши, описанные выше (фиг. 5А и 5В, Направленные векторы +12-λ или +12-κ). Фрагмент ДНК размером 93674 п.о. из клона ВАС RP11-22I18 человека (Invitrogen), содержащий 12 последовательных генных сегментов hVλ из кластера В, был сконструирован с 5' гомологичным плечом, содержащим геномную последовательность мыши 5' либо к эндогенным локусам легкой цепи κ, либо легкой цепи λ мыши, и 3' гомологичным плечом, содержащим геномную последовательность λ человека. 5' гомологичные плечи, использованные в этой нацеливающей конструкции, были такими же, как и 5' гомологичные плечи, использованные для добавления 16 генных сегментов hVλ, описанные выше (фиг. 5A и 5B). 3' гомологичное плечо было создано в результате конструирования участка PI-SceI ~3431 п.о. 5' к генному сегменту Vλ3-29Р человека, содержащему 27468 п.о. геномного фрагмента последовательности λ человека из клона ВАС RP11-76I13 человека. Этот участок PI-SceI выполняет функцию точки лигирования для соединения фрагмента размером ~94 т.п.о. дополнительной последовательности λ человека с фрагментом размером ~27 т.п.о. последовательности λ человека, который перекрывается с 5'-концом последовательности λ человека в предыдущей модификации с использованием направленных векторов +16-λ или +16-κ (фиг. 5А и 5В). Эти два направленных вектора включали от 5' до 3', 5' гомологичное плечо, содержащее либо ~23 т.п.о. геномной последовательности мыши 5' от эндогенного локуса легкой цепи κ, или ~24 т.п.о. геномной последовательности мыши 5' от эндогенного локуса легкой цепи λ, 5' участок Frt, неомициновую кассету, 3' участок Frt и 121188 п.о. геномной последовательности λ человека, содержащей 16 генных сегментов hVλ и участок PI-SceI, ~27 т.п.о. которой перекрываются с 5'-концом последовательности λ человека из вставки 16 дополнительных генных сегментов hVλ, и выполняют функцию 3' гомологичного плеча для этой нацеливающей конструкции (фиг. 5А и 5В, Направленные векторы +12-λ или +12-κ). Гомологичная рекомбинация с этими направленными векторами независимо создает модифицированные локусы легкой цепи κ и λ мыши, содержащие 40 генных сегментов hVλ и генный сегмент Jλ1 человека, функционально связанных с эндогенными константными генами мыши (Сκ или Сλ2), что после рекомбинации приводит к формированию химерной легкой цепи (внизу фиг. 5А и 5В).
Аналогичным образом, направленный вектор +12-κ также использовали для введения 12 дополнительных генных сегментов hVλ в другие первичные модификации, которые объединяют множественные генные сегменты hJλ с, или без, интегрированной последовательностью κ человека (Фиг. 4B). Гомологичная рекомбинация с этим направленным вектором в эндогенном локусе κ мыши, содержащем другие модификации, создает локус легкой цепи κ мыши, содержащий 40 генных сегментов hVλ и генные сегменты hJλ1, 2, 3 и 7 с, или без, геномной последовательностью Vκ-Jκ, функционально связанные с эндогенным геном Сκ мыши, что, после рекомбинации, приводит к формированию химерной легкой цепи λ-κ.
Пример IV
Выявление клеток-мишеней ES, несущих генные сегменты легкой цепи λ человека
ДНК целевого ВАС, созданного в соответствии с вышеприведенными Примерами, использовали для электропорации клеток ES мыши, для получения модифицированных клеток ES для создания химерных мышей, которые экспрессируют генные сегменты легкой цепи λ человека. Клетки ES, содержащие вставку неперегруппированных генных сегментов легкой цепи λ человека, были идентифицированы с помощью количественного анализа TAQMAN®. Были разработаны специфические наборы праймеров и зондов для вставки последовательностей λ человека и соответствующих селекционных кассет (появление аллеля, GOA), утраты эндогенных последовательностей мыши и любых селекционных кассет (утрата аллеля, LOA) и сохранения фланкирующих последовательностей мыши (сохранение аллеля, AR). Для каждой дополнительной вставки последовательностей λ человека использовали дополнительные наборы праймеров и зондов, для подтверждения присутствия дополнительных последовательностей λ человека, а также предыдущие наборы праймеров и зондов, используемых для подтверждения сохранения ранее нацеленных последовательностей человека. В Таблице 1 представлены праймеры и соответствующие зонды, использованные в количественном ПЦР-анализе. В Таблице 2 представлены комбинации, использованные для подтверждения вставки каждого участка генных сегментов легкой цепи λ человека в клонах клеток ES.
Клетки ES, несущие генные сегменты легкой цепи λ человека, при желании трансфицированы с использованием конструкции, которая экспрессирует FLP с целью удаления Frt неомициновой кассеты, введенной посредством инсерции нацеливающей конструкции, содержащей генные сегменты Vλ5-52 - Vλ1-40 (фиг. 5А и 5В). Неомициновую кассету при желании можно удалить посредством скрещивания с мышами, которые экспрессируют FLP рекомбиназу (например, патент США № 6774279). При желании, неомициновую кассету можно оставить в мышах.
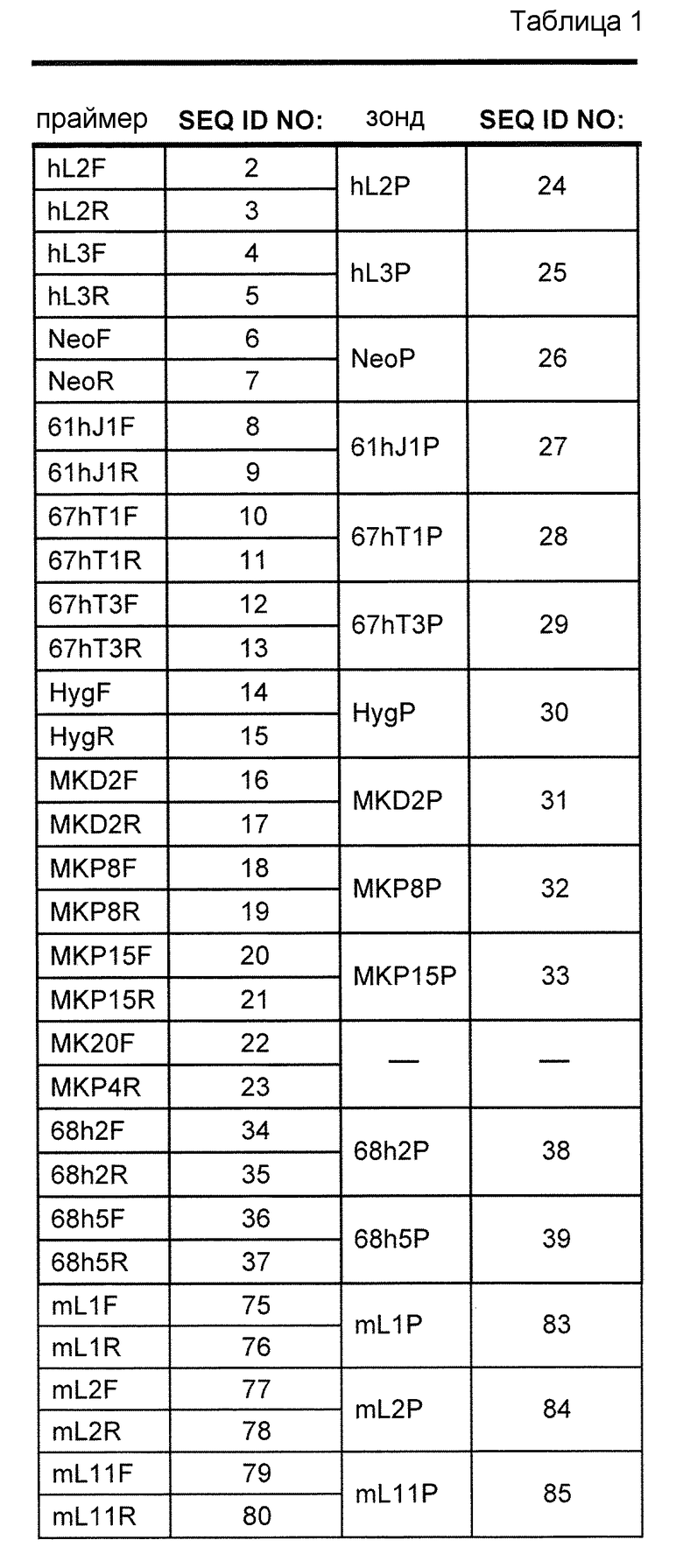
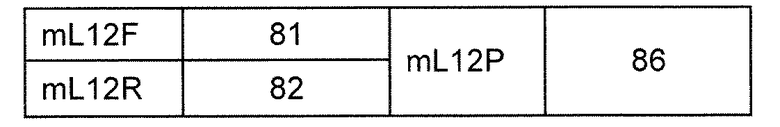
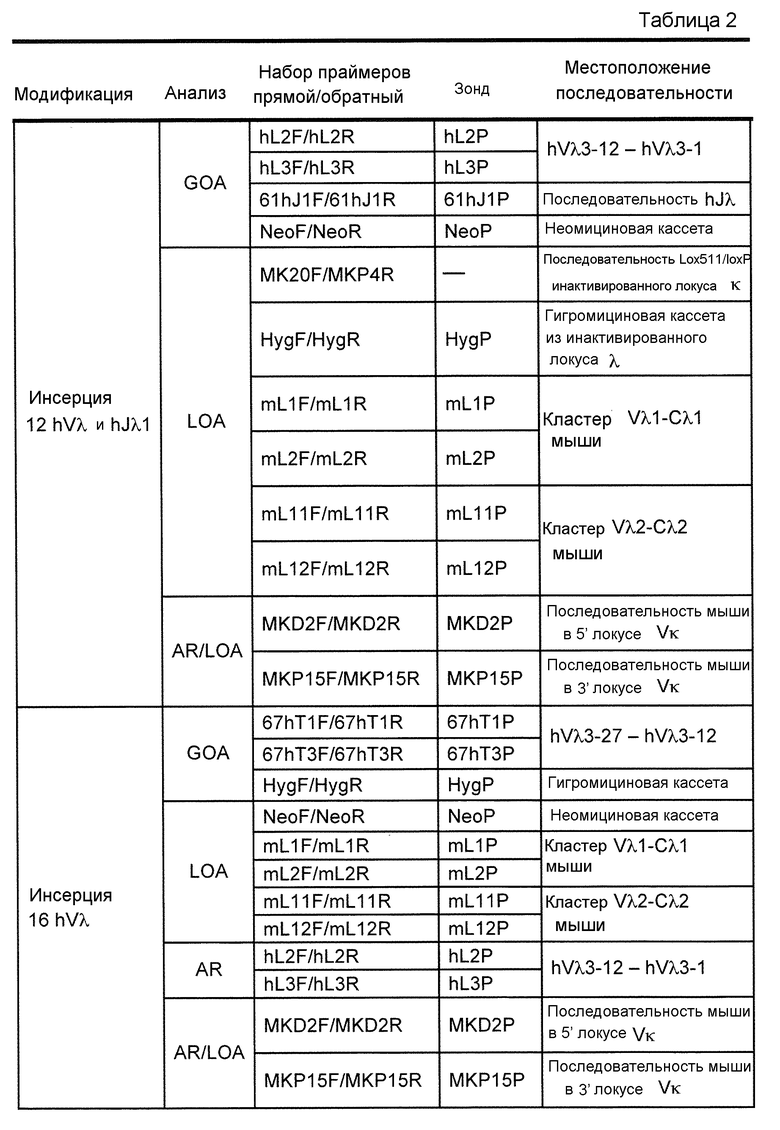
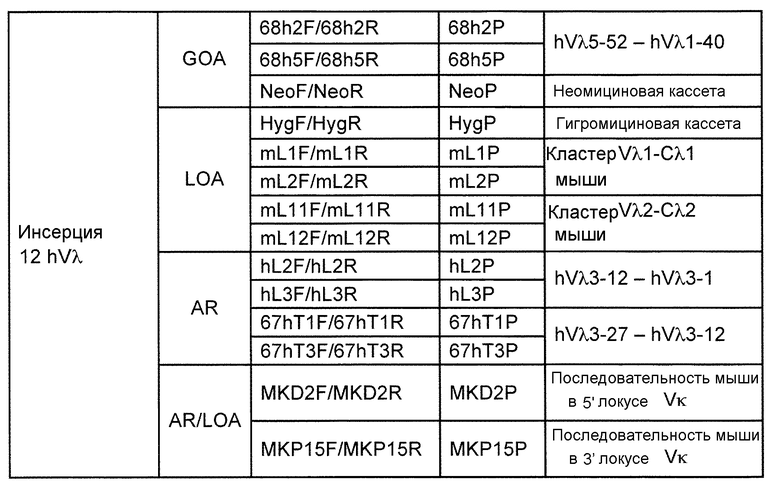
Пример V
Создание мышей, экспрессирующих легкие цепи λ человека из эндогенного локуса легкой цепи
Нацеленные клетки ES, описанные выше, использовали в качестве донорских клеток ES и вводили в эмбрион мыши на 8-клеточной стадии развития, используя метод VELOCIMOUSE® (см., например, патент США № 7294754 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1): 91-99.) VELOCIMICE® (Мыши поколения F0, полностью происходящие из клеток ES донора), независимо несущие генные сегменты λ человека, были выявлены в результате генотипирования с использованием модификации анализа аллелей (Valenzuela et al., выше), который выявляет присутствие уникальных генных сегментов λ человека (выше).
Соотношение использования легких цепей κ/λ мышами, несущими генные сегменты легкой цепи λ человека. Мышей, гомозиготных по каждой из трех последовательных инсерций генных сегментов hVλ с единственным генным сегментом hJλ (фиг. 5В) и мышей, гомозиготных по первой инсерции генных сегментов hVλ либо с единственным генным сегментом hJλ, либо четырьмя генными сегментами Jλ человека, включая геномную последовательность Vκ-Jκ человека (фиг. 4В), анализировали на экспрессию легкой цепи κ и λ в спленоцитах, используя способ проточной цитометрии.
Кратко, у мышей из групп (диапазон от трех до семи животных в группе) удаляли селезенки и измельчали их, используя стеклянные пластинки. После лизиса эритроцитов (RBC) с помощью лизирующего буфера ACK (Lonza Walkersville), спленоциты окрашивали флуоресцентным красителем конъюгированных антител, специфических для CD19 мыши (клон 1D3; BD Biosciences), CD3 мыши (17A2; Biolegend), Igκ мыши (187.1; BD Biosciences) и Igλ мыши (RML-42; Biolegend). Результат был получен с помощью проточного цитометра BD™ LSR II (BD Biosciences) и проведен анализ с использованием программного обеспечения FLOWJO™ (Tree Star, Inc.). В Таблице 3 представлена средняя величина в процентах для экспрессии В-клеток (CD19+), легкой цепи κ (CD19+Igκ+Igλ-) и легкой цепи λ (CD19+Igκ-Igλ+), наблюдаемой в спленоцитах, полученных от групп животных, несущих каждую генетическую модификацию.
В аналогичном эксперименте, В-клеточный состав в селезеночном компартменте, полученном от мышей, гомозиготных по первой инсерции 12 генных сегментов hVλ и четырех генных сегментов hJλ, включающих геномную последовательность Vκ-Jκ человека, функционально связанную с геном Сκ мыши (фиг. 4В, внизу), и от мышей, гомозиготных по 40 генным сегментам hVλ и одному генному сегменту hJλ (фиг. 5В, внизу или фиг. 7В, вверху), был проанализирован на экспрессию Igκ и Igλ с использованием проточной цитометрии (как описано выше). На Фиг. 8A продемонстрирована экспрессия Igλ и Igκ в CD19+ В-клетках для репрезентативной мыши из каждой группы. Количество CD19+ B-клеток в селезенке также было зарегистрировано для каждой мыши (фиг. 8В).
В другом эксперименте, В-клеточные составы из селезеночного и костно-мозгового компартментов от мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам hJλ, включающим геномную последовательность Vκ-Jκ человека, функционально связанную с геном Сκ мыши (Фиг. 7В, внизу) были проанализированы на предмет прогрессии на протяжении развития В-клеток с использованием проточной цитометрии различных маркеров клеточной поверхности.
Кратко, две группы (N=3 в каждой, возраст 9-12 недель, самцы и самки) мышей дикого типа и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам hJλ, включающим геномную последовательность Vκ-Jκ человека, функционально связанную с геном Сκ мыши, были безболезненно умерщвлены, и селезенки и костный мозг были собраны. Костный мозг был отобран из бедренных костей посредством промывания сильным напором струи полной среды RPMI (среда RPMI, дополненная фетальной телячьей сывороткой, пируватом натрия, Hepes, 2-меркаптоэтанолом, заменимыми аминокислотами и гентамицином). Эритроциты в препаратах селезенок и костного мозга были лизированы с помощью лизирующего буфера АСК (Lonza Walkersville), с последующим отмыванием с помощью полной среды RPMI. Клетки 1×106 инкубировали с анти-мышиными CD16/CD32 (2.4G2, BD Biosciences) на льду в течение 10 минут, с последующим мечением с помощью отобранной панели антител в течение 30 мин на льду.
Панель для костного мозга: анти-мышиное FITC-CD43 (1B11, BioLegend), PE-ckit (2B8, BioLegend), PeCy7-IgM (II/41, eBioscience), PerCP-Cy5.5-IgD (11-26c.2a, BioLegend), APC-B220 (RA3-6B2, eBioscience), APC-H7-CD19 (ID3, BD) и Pacific Blue-CD3 (17A2, BioLegend).
Панель для костного мозга и селезенки: анти-мышиное FITC-Igκ (187.1, BD), PE-Igλ (RML-42, BioLegend), PeCy7-IgM (II/41, ebioscience), PerCP-Cy5.5-IgD (11-26c.2a, BioLegend), Pacific Blue-CD3 (17A2, BioLegend), APC-B220 (RA3-6B2, eBioscience), APC-H7-CD19 (ID3, BD).
После окрашивания клетки отмывали и фиксировали в 2% формальдегиде. Получение было осуществлено с помощью проточного цитометра FACSCANTOII™ (BD Biosciences) и анализ был проведен с использованием программного обеспечения FLOWJO™ (Tree Star, Inc.). На фиг. 9А-9D представлены результаты для селезеночного компартмента одной репрезентативной мыши из каждой группы. На фиг. 10A-10E представлены результаты для костно-мозгового компартмента одной репрезентативной мыши из каждой группы. В Таблице 4 представлены средние величины в процентах для экспрессии В-клеток (CD19+), легкой цепи κ (CD19+Igκ+Igλ-) и легкой цепи λ (CD19+Igκ-Igλ+), наблюдаемой в спленоцитах, полученных от групп животных, несущих различные генетические модификации. В Таблице 5 представлены средние величины в процентах для экспрессии В-клеток (CD19+), зрелых В-клеток (B220hiIgM+), незрелых В-клеток (B220intIgM+), незрелых В-клеток, экспрессирующих легкую цепь κ (B220intIgM+Igκ+)и незрелых В-клеток, экспрессирующих легкую цепь λ (B220intIgM+Igλ+), наблюдаемую в костном мозге мышей дикого типа и мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам hJλ, включающим геномную последовательность Vκ-Jκ человека, функционально связанную с геном Сκ мыши. Этот эксперимент был повторен с дополнительной группой мышей, описанных выше, и были получены аналогичные результаты (данные не представлены).
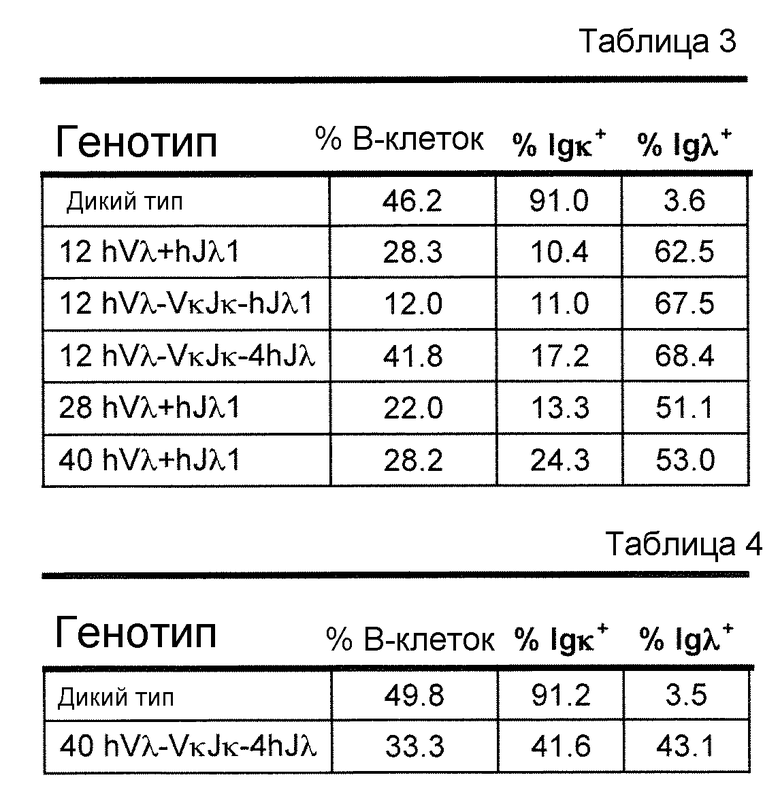
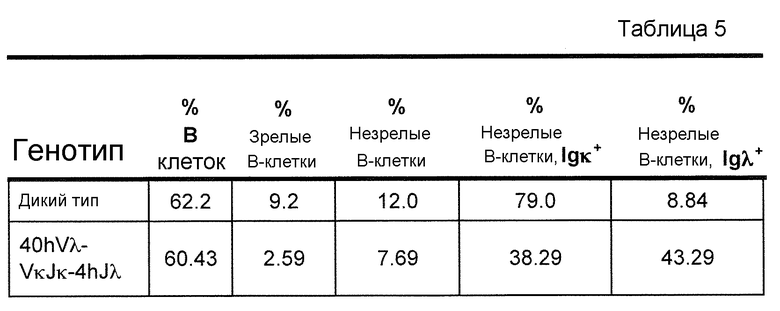
Использование гена Vλ человека у мышей, несущих генные сегменты легкой цепи λ человека. Проводили анализ мышей, гомозиготных по первой инсерции последовательностей λ человека (hVλ3-12-hVλ3-1 и hJλ1, фиг. 5B) и гомозиготных по третьей инсерции последовательностей λ человека (hVλ5-52-hVλ3-1 и hJλ1, Фиг. 5B), на использование генов легкой цепи λ человека, посредством полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР), с использованием РНК, выделенной из спленоцитов.
Кратко, собирали селезенки и заливали 10 мл RPMI-1640 (Sigma) с 5% HI-FBS в стерильных пакетах одноразового использования. Каждый пакет, содержащий одну селезенку, затем помещали в STOMACHER™ (Seward) и гомогенизировали в течение 30 секунд при средних установках. Гомогенизированные селезенки фильтровали с использованием 0,7 мкм клеточного фильтра и затем осаждали с помощью центрифуги (1000 об/мин в течение 10 минут) и эритроциты лизировали в течение трех минут в BD PHARM LYSE™ (BD Biosciences). Спленоциты разбавляли с помощью RPMI-1640 и снова центрифугировали, с последующим ресуспендированием в 1 мл PBS (Irvine Scientific). РНК выделяли из спленоцитов, используя стандартные методы, известные в данной области.
ОТ-ПЦР осуществляли на РНК спленоцитов с использованием праймеров, специфичных для генных сегментов hVλ человека и гена Сκ мыши (Таблица 6). ПЦР-продукты очищали на геле и клонировали в вектор pCR2.1-TOPO TA (Invitrogen) и проводили секвенирование с использованием праймеров M13 Прямой (GTAAAACGACGGCCAG; SEQ ID NO:55) и M13 Обратный (CAGGAAACAGCTATGAC; SEQ ID NO:56), расположенных в пределах данного вектора в позициях, фланкирующих сайт клонирования. Восемьдесят четыре общих клона, полученные из первой и третьей инсерции последовательностей λ человека, были секвенированы для определения использования гена hVλ (Таблица 7). Нуклеотидная последовательность соединения hVλ-hJλ1-mCκ для отобранных клонов ОТ-ПЦР представлена на фиг. 11.
Аналогичным образом проводили анализ мышей, гомозиготных по третьей инсерции генных последовательностей легкой цепи λ человека (а именно, 40 генных сегментов hVλ и четырех генных сегментов hJλ, включая геномную последовательность Vκ-Jκ человека, Фиг. 7В, внизу), функционально связанных с эндогенным геном Сκ мыши, на использование гена легкой цепи λ человека, посредством ОТ-ПЦР, с использованием РНК, выделенной из спленоцитов (как описано выше). Использование генных сегментов легкой цепи λ человека для 26 отобранных ОТ-ПЦР клонов представлено в Таблице 8. Нуклеотидная последовательность соединения hVλ-hJλ-mCκ для отобранных клонов ОТ-ПЦР представлена на фиг. 12.
Аналогичным образом проводили анализ мышей, гомозиготных по первой инсерции генных сегментов легкой цепи λ человека (12 генных сегментов hVλ и hJλ1, фиг. 4А и фиг. 5А), функционально связанных с эндогенным геном Сλ2 мыши, на использование гена легкой цепи λ человека, посредством ОТ-ПЦР, с использованием РНК, выделенной из спленоцитов (как описано выше). Праймеры, специфичные для генных сегментов hVλ (Таблица 6) были спарены с одним из двух праймеров, специфичных для гена Сλ2 мыши; Сλ2-1 (SEQ ID NO:104) или Сλ2-2 (SEQ ID NO:105).
Множественные генные сегменты hVλ перегруппированные в hλ1 наблюдали в клонах ОТ-ПЦР мышей, несущих генные сегменты легкой цепи λ человека в эндогенном локусе легкой цепи λ мыши. Нуклеотидная последовательность соединения hVλ-hJλ-mCλ2 для отобранных клонов ОТ-ПЦР представлена на Фиг. 13.
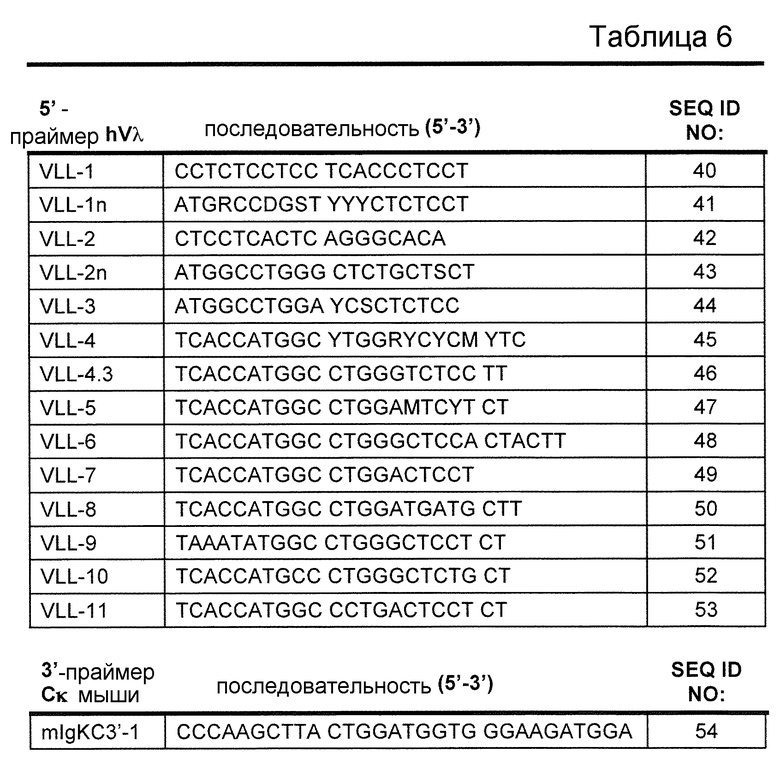
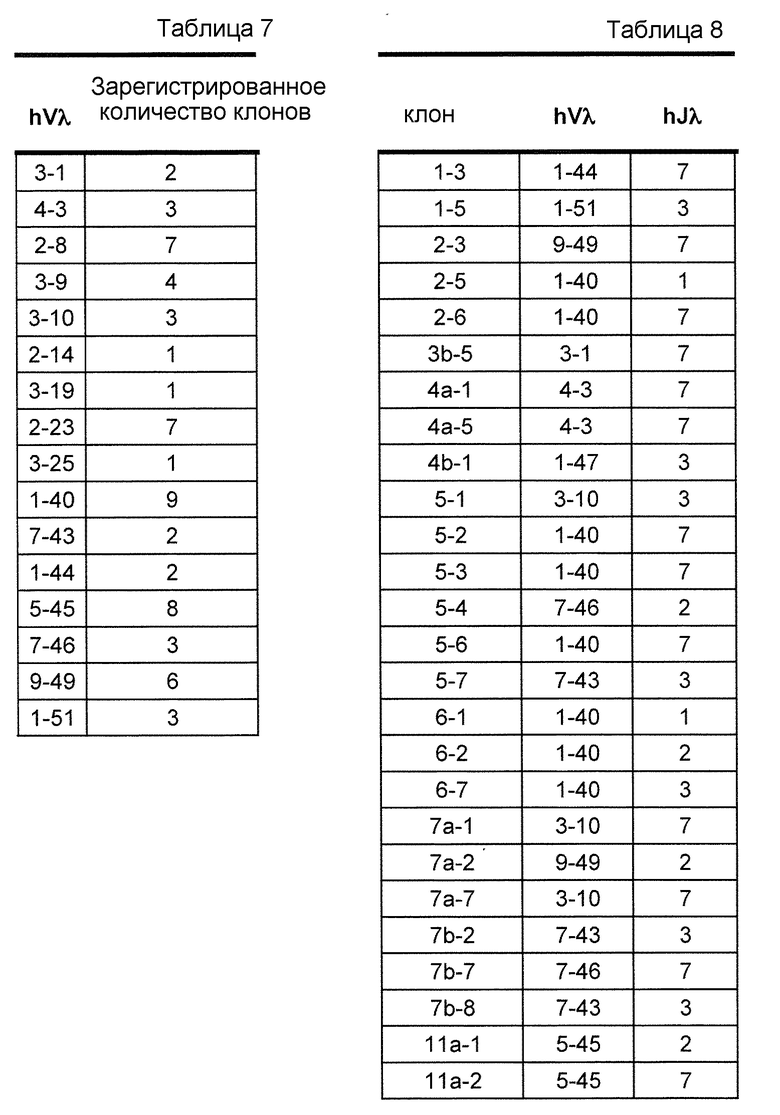
На фиг. 11 представлена последовательность соединения hVλ-hJλ1-mCκ для ОТ-ПЦР клонов, полученных от мышей, несущих первую и вторую инсерцию генных сегментов hVλ с единственным генным сегментом hJλ. Последовательности, представленные на Фиг. 11, демонстрируют уникальные перегруппировки, вовлекающие различные генные сегменты hVλ с hJλ1, рекомбинированные с геном Сκ мыши. И гетерозиготные мыши, несущие единственный модифицированный эндогенный локус κ, содержащий 12 генных сегментов hVλ и hJλ1, и гомозиготные мыши, несущие два модифицированных эндогенных локуса κ, содержащие 40 генных сегментов hVλ и hJλ1, были в состоянии продуцировать генные сегменты λ человека, функционально связанные с геном Сκ мыши, и продуцировать В-клетки, которые экспрессируют легкие цепи λ человека. Эти перегруппировки демонстрируют, что химерные локусы могут независимо перегруппировывать генные сегменты λ человека в нескольких независимых В-клетках в этих мышах. Кроме того, эти модификации локуса эндогенной легкой цепи κ не воспроизводили никакие из генных сегментов hVλ нефункционально, или предохраняли химерный локус от многократной рекомбинации генного сегмента hVλ и hJλ (Jλ1) во время развития В-клетки, о чем свидетельствуют 16 различных генных сегментов hVλ, которые наблюдали перегруппированными с hJλ1 (Таблица 7). Кроме того, эти мыши продуцировали функциональные антитела, содержащие перегруппированные генные сегменты Vλ-Jλ человека, функционально связанные с генами Сκ мыши, как часть спектра эндогенной легкой цепи иммуноглобулина.
На фиг. 12 представлена последовательность соединения hVλ-hJλ-mCκ для отобранных клонов ОТ-ПЦР, полученных от мышей, гомозиготных по 40 генным сегментам hVλ и четырем генным сегментам hJλ, содержащим геномную последовательность Vκ-Jκ человека. Последовательности, представленные на Фиг. 12, демонстрируют дополнительные уникальные перегруппировки, затрагивающие множественные различные генные сегменты hVλ, охватывающие полностью химерный локус, с несколькими различными генными сегментами hJλ, перегруппированными и функционально связанными с геном Сκ мыши. Гомозиготные мыши, несущие модифицированные эндогенные локусы κ, содержащие 40 генных сегментов hVλ и четыре генных сегмента hJλ, также были способны продуцировать генные сегменты λ человека, функционально связанные с геном Сκ мыши, и продуцировать В-клетки, которые экспрессируют легкие цепи λ человека. Эти перегруппировки дополнительно демонстрируют, что химерные локусы на всех стадиях были способны независимо перегруппировывать генные сегменты λ человека в множественных независимых В-клетках у этих мышей. Кроме того, эти дополнительные модификации в эндогенном локусе легкой цепи κ демонстрируют, что каждая инсерция генных сегментов λ человека не приводила к образованию никаких нефункциональных генных сегментов hVλ и/или Jλ, или предохраняла химерный локус от рекомбинации генных сегментов hVλ и Jλ во время развития В-клетки, о чем свидетельствуют 12 различных генных сегментов hVλ, которые были обнаружены перегруппированными со всеми четырьмя генными сегментами hJλ (Таблица 8) из 26 отобранных клонов ОТ-ПЦР. Кроме того, эти мыши также продуцировали функциональные антитела, содержащие перегруппированные генные сегменты Vλ-Jλ человека, функционально связанные с областями Сκ мыши, как часть спектра эндогенной легкой цепи иммуноглобулина.
На фиг. 13 представлена последовательность соединения hVλ-hJλ-mCλ2 для трех отдельных клонов ОТ-ПЦР, полученных от мышей, гомозиготных по 12 генным сегментам hVλ и генному сегменту hJλ1. Последовательности, представленные на фиг. 13, демонстрируют дополнительные уникальные перегруппировки, вовлекающие различные генные сегменты hVλ, охватывающие всю длину первой инсерции, с hJλ1, перегруппированным и функционально связанным с геном Сλ2 мыши (2D1 = Vλ2-8Jλ1; 2D9 = Vλ3-10Jλ1; 3E15 = Vλ3-1Jλ1). Один клон демонстрировал непродуктивную перегруппировку вследствие добавлений N к соединению hVλ-hJλ (2D1, Фиг. 13). Это не редкость при рекомбинации V(D)J, поскольку показано, что соединение генных сегментов во время рекомбинации является неточным. Хотя этот клон представляет собой непродуктивный рекомбинант, присутствующий в спектре легких цепей этих мышей, это является демонстрацией того, что генетический механизм, способствующий множественности J-сегментов среди генов антител, работает нормально у этих мышей и приводит в результате к образованию спектра антител, включающего легкие цепи с более значительным разнообразием.
Гомозиготные мыши, несущие модифицированные эндогенные локусы λ, содержащие 12 генных сегментов hVλ и генный сегмент hJλ1, также были способны продуцировать генные сегменты λ человека, функционально связанные с эндогенным геном Сλ мыши, и продуцировать В-клетки, которые экспрессируют обратные химерные легкие цепи λ, содержащие области hVλ, связанные с областями Сλ мыши. Эти перегруппировки дополнительно демонстрируют, что генные сегменты легкой цепи λ человека, помещенные в другой локус легкой цепи (а именно, локус λ) были способны независимо перегруппировывать генные сегменты λ человека в нескольких независимых В-клетках у этих мышей. Кроме того, модификации в эндогенном локусе легкой цепи λ демонстрируют, что инсерция генных сегментов λ человека не воспроизводит никаких нефункциональных генных сегментов hVλ и/или hJλ1, или предохраняет химерный локус от рекомбинации генных сегментов hVλ и hJλ1 во время развития В-клеток. Кроме того, эти мыши также продуцировали функциональные антитела, содержащие генные сегменты Vλ-Jλ человека, функционально связанные с областями Сλ мыши, как часть спектра эндогенной легкой цепи иммуноглобулина.
Как показано в этом Примере, мыши, несущие генные сегменты легкой цепи λ человека в эндогенных локусах легкой цепи κ и λ, способны к перегруппировке генных сегментов легкой цепи λ человека и экспрессии их в связи с областью Сκ и/или Сλ мыши, в качестве части нормального спектра антител мыши, поскольку функциональная легкая цепь требуется на различных контрольных точках в ходе развития В-клеток, как в селезенке, так и в костном мозге. Кроме того, ранние субпопуляции В-клеток (например, пре-, про- и промежуточные В-клетки) демонстрируют нормальный фенотип у этих мышей, в сравнении с однопометными животными дикого типа (Фиг. 9D, 10A и 10B). Наблюдали небольшой дефицит в костном мозге и периферических популяций В-клеток, что можно объяснить делецией субпопуляции самореактивных незрелых В-клеток и/или недостаточной ассоциацией легкой цепи λ человека с тяжелой цепью мыши. Тем не менее, использование Igκ/Igλ, наблюдаемое у этих мышей, демонстрирует ситуацию, которая больше похожа на экспрессию легкой цепи человека, чем таковую, наблюдаемую у мышей.
Пример VI
Разведение мышей, экспрессирующих легкие цепи λ человека из эндогенного локуса легкой цепи
Для оптимизации использования генных сегментов λ человека в эндогенном локусе легкой цепи мыши, мышей, несущих неперегруппированные генные сегменты λ человека, скрещивали с другими мышами, несущими делецию противоположного эндогенного локуса легкой цепи (либо κ, либо λ). Например, генные сегменты λ человека, помещенные в эндогенный локус κ, будут единственно функциональными генными сегментами легкой цепи, присутствующими у мыши, которая также несет делецию в эндогенном локусе легкой цепи λ. Таким образом, полученное потомство будет экспрессировать только легкие цепи λ человека, как описано в вышеприведенных примерах. Размножение осуществляется с использованием стандартных технологий, принятых в данной области, и, альтернативно, коммерческими компаниями, например, The Jackson Laboratory. Проводится скрининг пород мышей, несущих генные сегменты легких цепей λ человека в эндогенном локусе κ и делецию эндогенного локуса легкой цепи λ, на наличие уникальных обратно-химерных (человека-мышь) легких цепей λ и отсутствие эндогенных легких цепей λ мыши.
Мышей, несущих неперегруппированный локус легкой цепи λ человека, также скрещивают с мышами, которые содержат замену эндогенного локуса вариабельного гена тяжелой цепи мыши на локус вариабельного гена тяжелой цепи человека (см. патент США № 6596541, Regeneron Pharmaceuticals, созданная методами генной инженерии мышь VELOCIMMUNE®). Мышь VELOCIMMUNE® включает, отчасти, геном, содержащий вариабельные области тяжелой цепи человека, функционально связанные с эндогенными локусами константной области мыши, так что мышь продуцирует антитела, содержащие вариабельную область тяжелой цепи человека и константную область тяжелой цепи мыши, в ответ на антигенную стимуляцию. ДНК, кодирующая вариабельные области тяжелых цепей антител, может быть выделена и функционально связана с ДНК, кодирующей константные области тяжелой цепи человека. ДНК затем может быть экспрессирована в клетках, способных к экспрессии полностью человеческой тяжелой цепи антитела. На основании соответствующего графика размножения, получены мыши, несущие замену эндогенного локуса тяжелой цепи мыши на локус тяжелой цепи человека, и неперегруппированный локус легкой цепи λ человека в эндогенном локусе легкой цепи κ. Антитела, содержащие соматические видоизмененные вариабельные области тяжелой цепи человека и вариабельные области легкой цепи λ человека, могут быть выделены после иммунизации представляющим интерес антигеном.
Пример VII
Продуцирование антител мышами, экспрессирующими тяжелые цепи человека и легкие цепи λ человека
После скрещивания мышей, которые содержат неперегруппированный локус легкой цепи λ человека, с различными породами, содержащими модификации и делеции других эндогенных локусов Ig (как описано выше), отобранных мышей иммунизировали представляющим интерес антигеном.
В целом, мышь VELOCIMMUNE®, содержащую один из однократно перегруппированных областей легкой цепи зародышевой линии человека, сенсибилизировали антигеном, и лимфатические клетки (такие, как В-клетки) восстанавливали из сыворотки крови животных. Лимфатические клетки могут быть слиты с клеточной линией миеломы для получения бессмертных гибридомных клеточных линий, и проводят скрининг и отбор таких гибридомных клеточных линий для выявления гибридомных клеточных линий, которые продуцируют антитела, содержащие тяжелую цепь человека и легкую цепь λ человека, которые являются специфическими для антигена, использованного для иммунизации. ДНК, кодирующая вариабельные области тяжелых цепей и легких цепей λ, может быть выделена и связана с подходящими изотипическими константными областями тяжелой цепи и легкой цепи. Вследствие наличия дополнительных генных сегментов hVλ, по сравнению с эндогенным локусом λ мыши, разнообразие спектра легких цепей резко возрастает и придает более высокое разнообразие антигенспецифическому спектру после иммунизации. Полученные клонированные последовательности антител могут быть в дальнейшем продуцированы в клетке, такой как клетка СНО. Альтернативно, ДНК, кодирующая антигенспецифические химерные антитела, или вариабельные домены легкой и тяжелой цепей, может быть выделена непосредственно из антигенспецифических лимфоцитов (например, В-клеток).
Первоначально, высокоаффинные химерные антитела изолированы имеющими вариабельную область человека и константную область мыши. Как описано выше, антитела характеризуются и отбираются по требуемым характерным особенностям, включающим аффинность, селективность, эпитоп, и тому подобное. Константные области мыши заменяют на желаемую константную область человека для создания полностью человеческого антитела, содержащего соматически видоизмененную тяжелую цепь человека и легкую цепь человека λ, полученную из неперегруппированного локуса легкой цепи λ человека по настоящему изобретению. Подходящие константные области человека включают, например, IgG1, IgG2, IgG3 или IgG4, дикого типа или модифицированные.
| название | год | авторы | номер документа |
|---|---|---|---|
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ λ ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2020 |
|
RU2833566C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ГИБРИДНУЮ ЛЕГКУЮ ЦЕПЬ ИММУНОГЛОБУЛИНА | 2011 |
|
RU2601297C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ λ ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2011 |
|
RU2724736C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| МЫШИ, У КОТОРЫХ ВЫРАБАТЫВАЮТСЯ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ V-ДОМЕНЫ | 2011 |
|
RU2750219C2 |
| МЫШИ, У КОТОРЫХ ВЫРАБАТЫВАЮТСЯ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ VL-ДОМЕНЫ | 2011 |
|
RU2612903C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ СО СКОНСТРУИРОВАННЫМ ЛОКУСОМ ЛЕГКОЙ ЛЯМБДА-ЦЕПИ ИММУНОГЛОБУЛИНА | 2017 |
|
RU2757665C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2664185C2 |
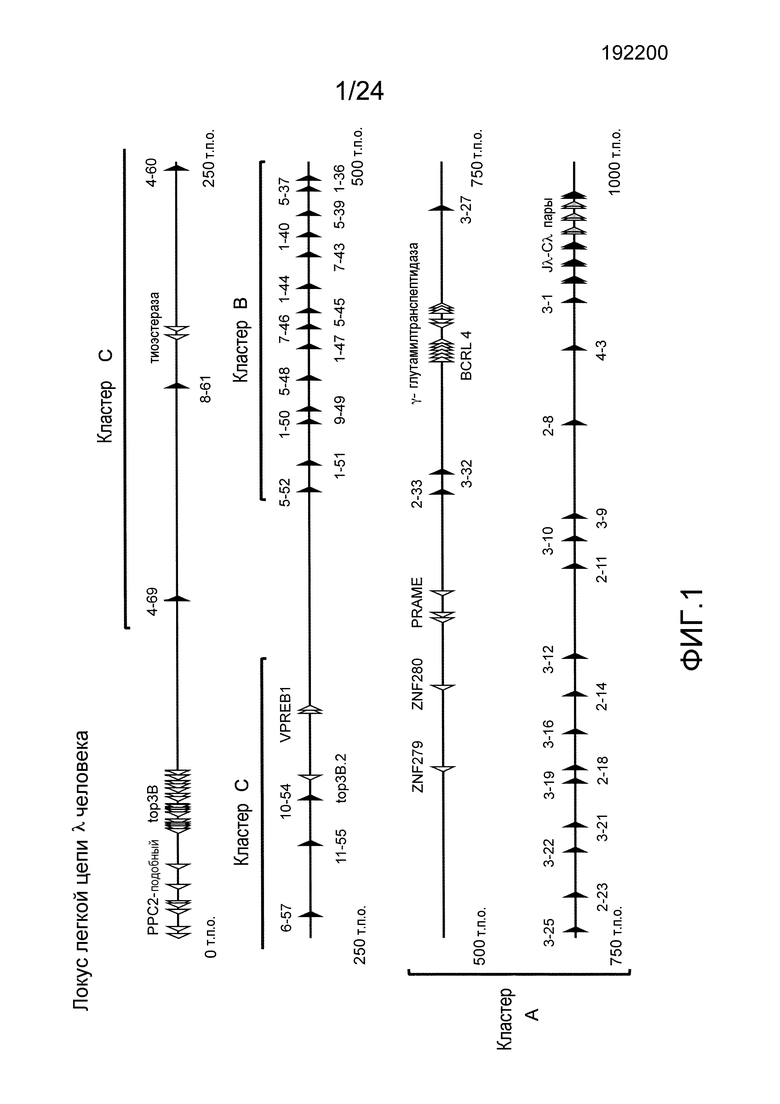
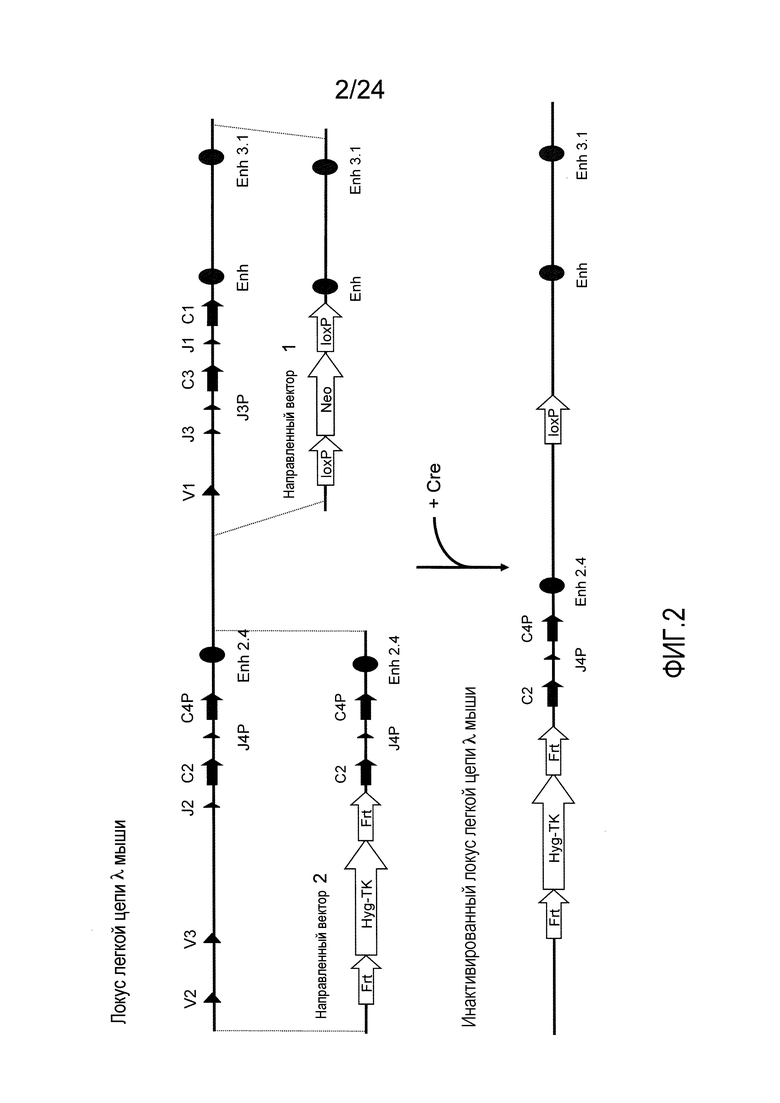
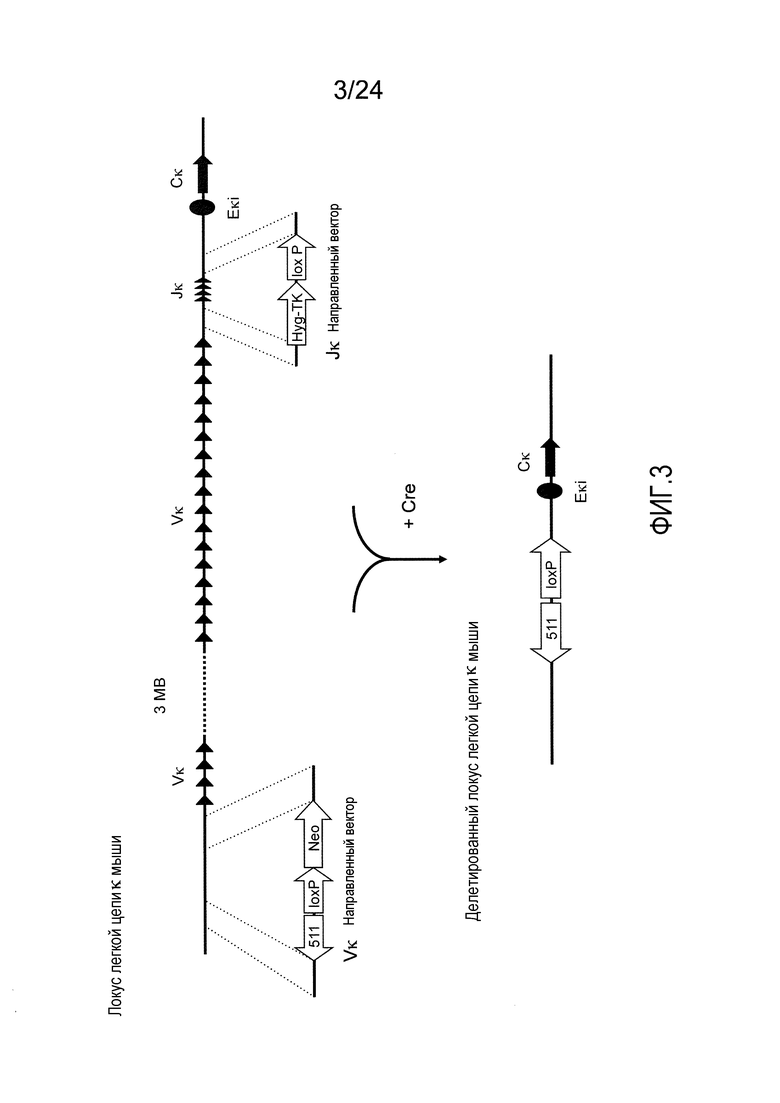
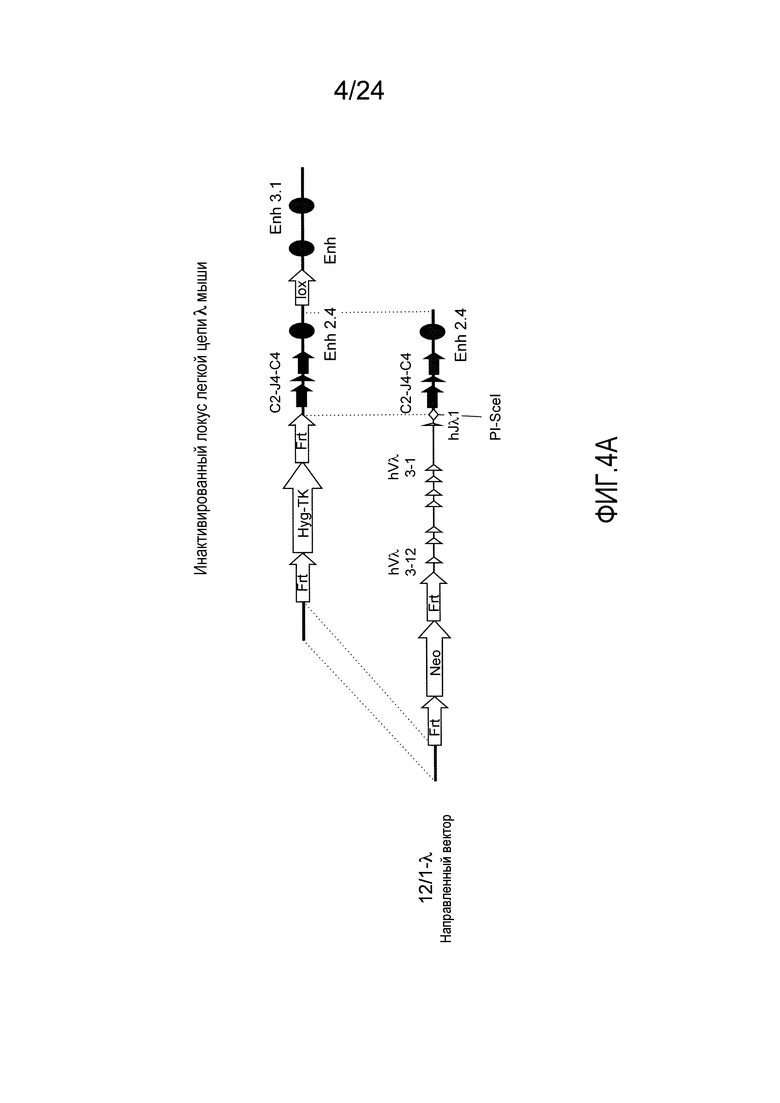
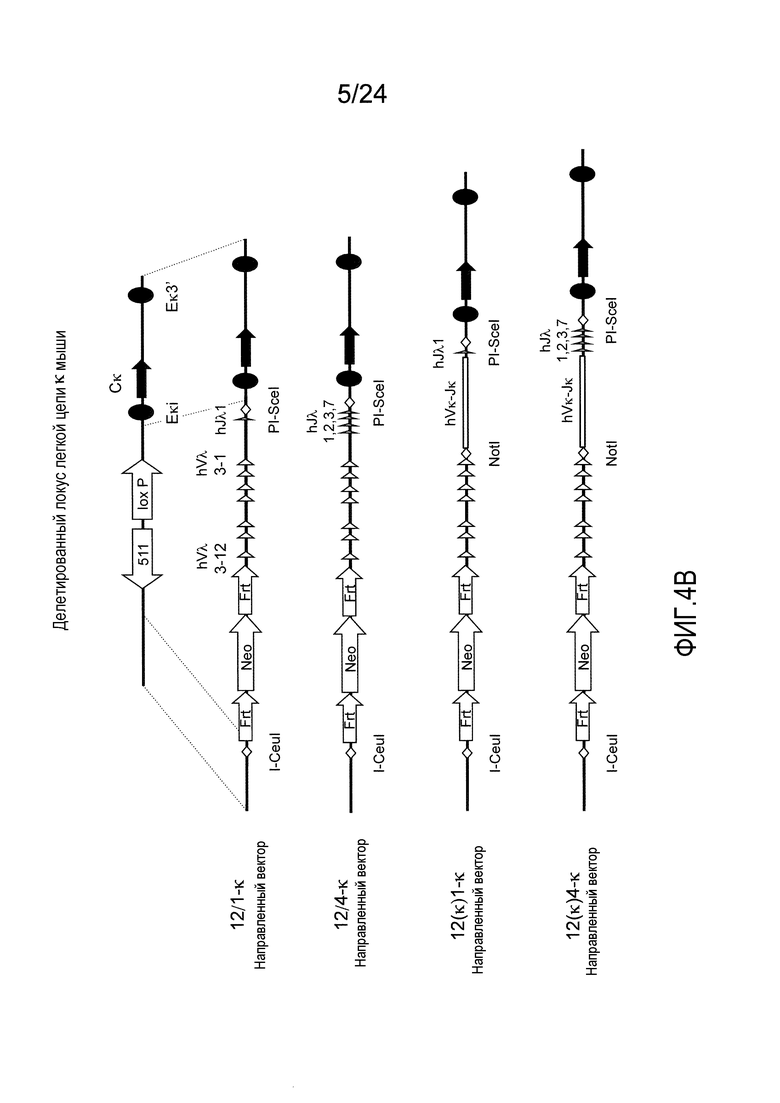
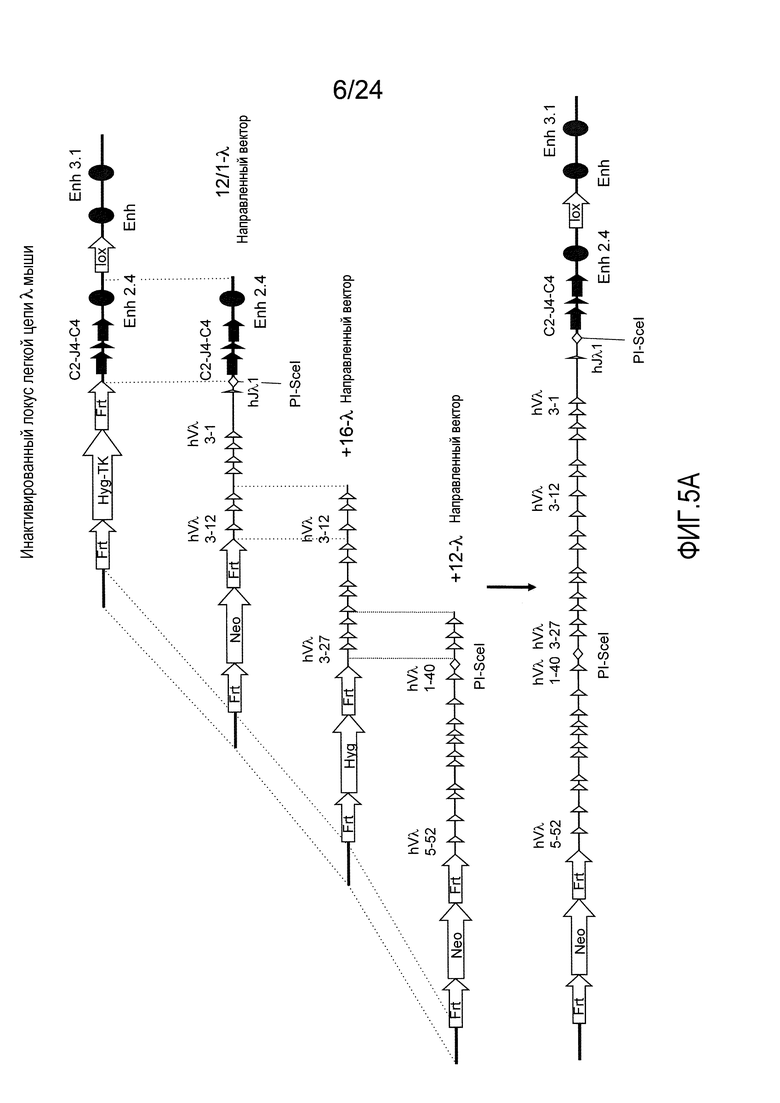
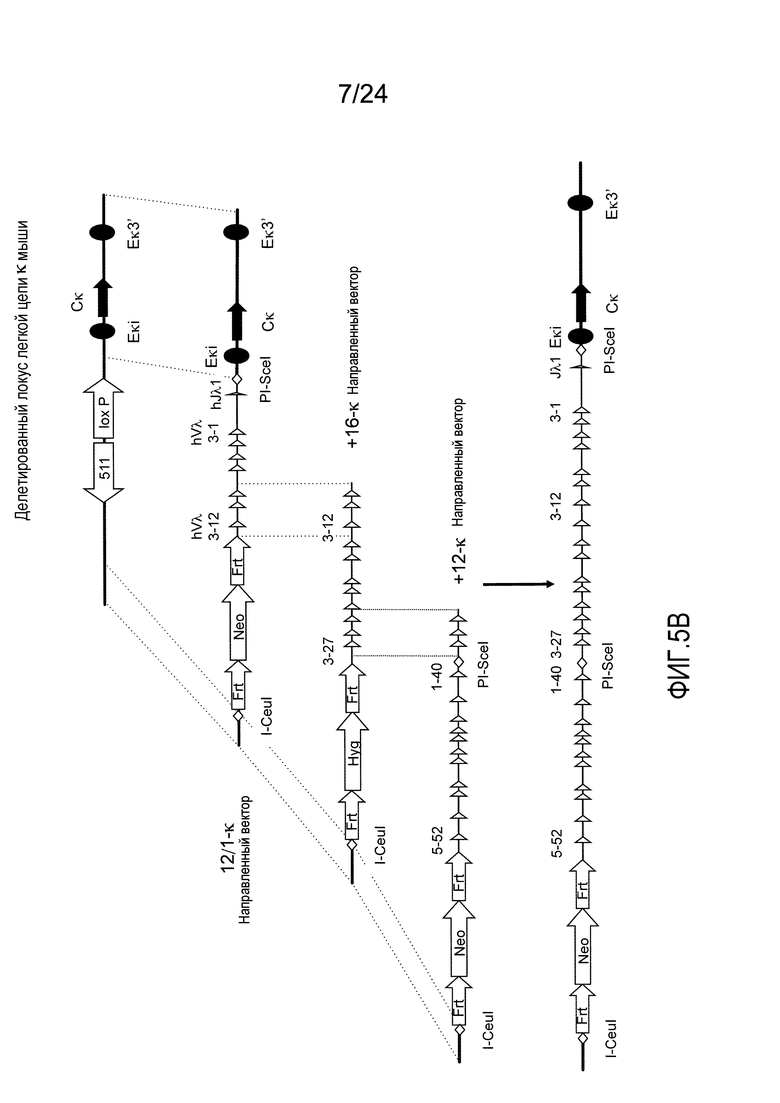
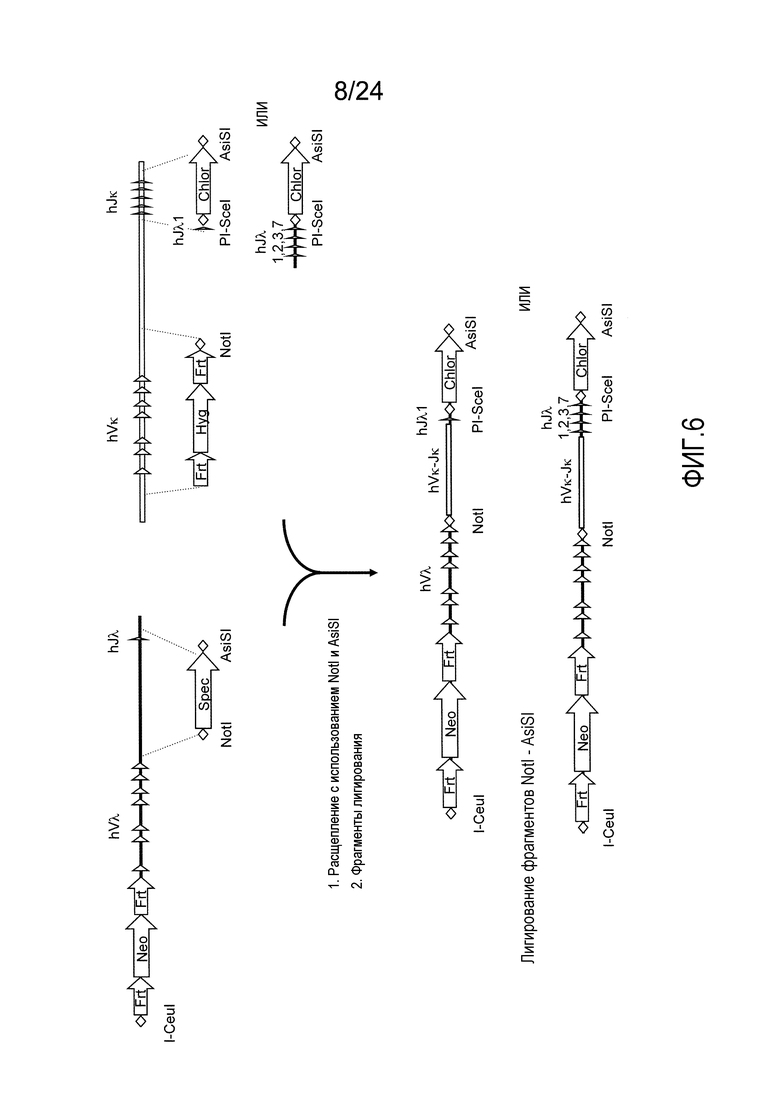

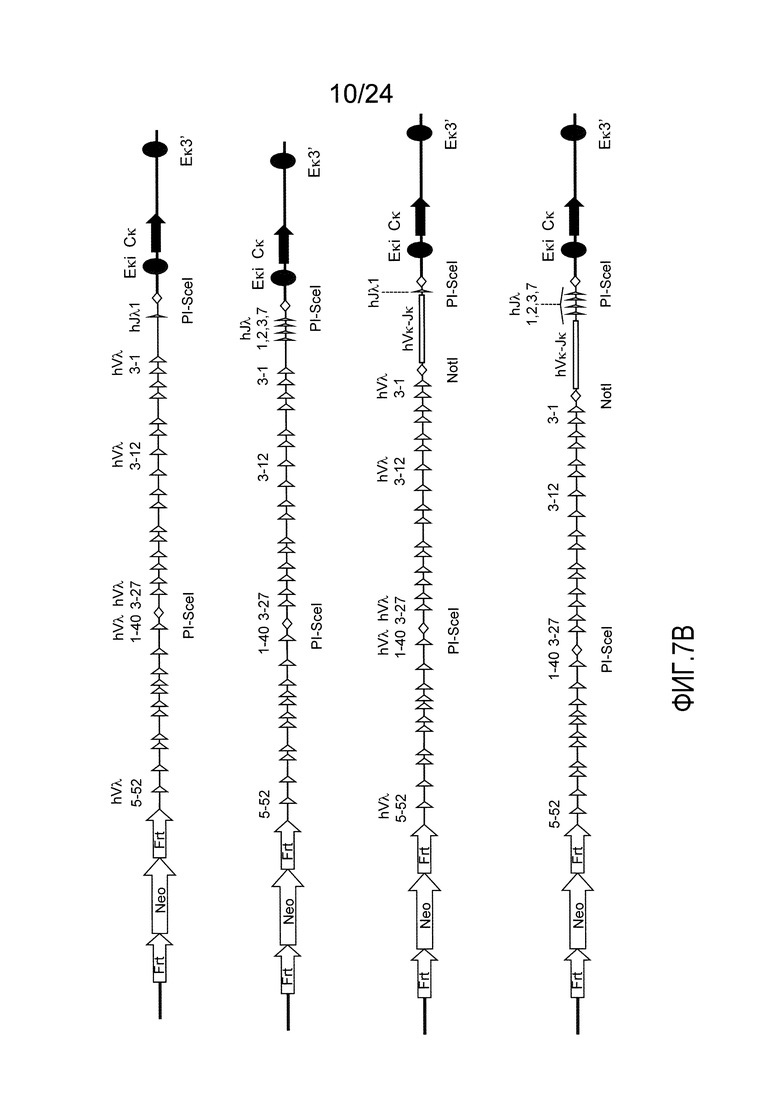
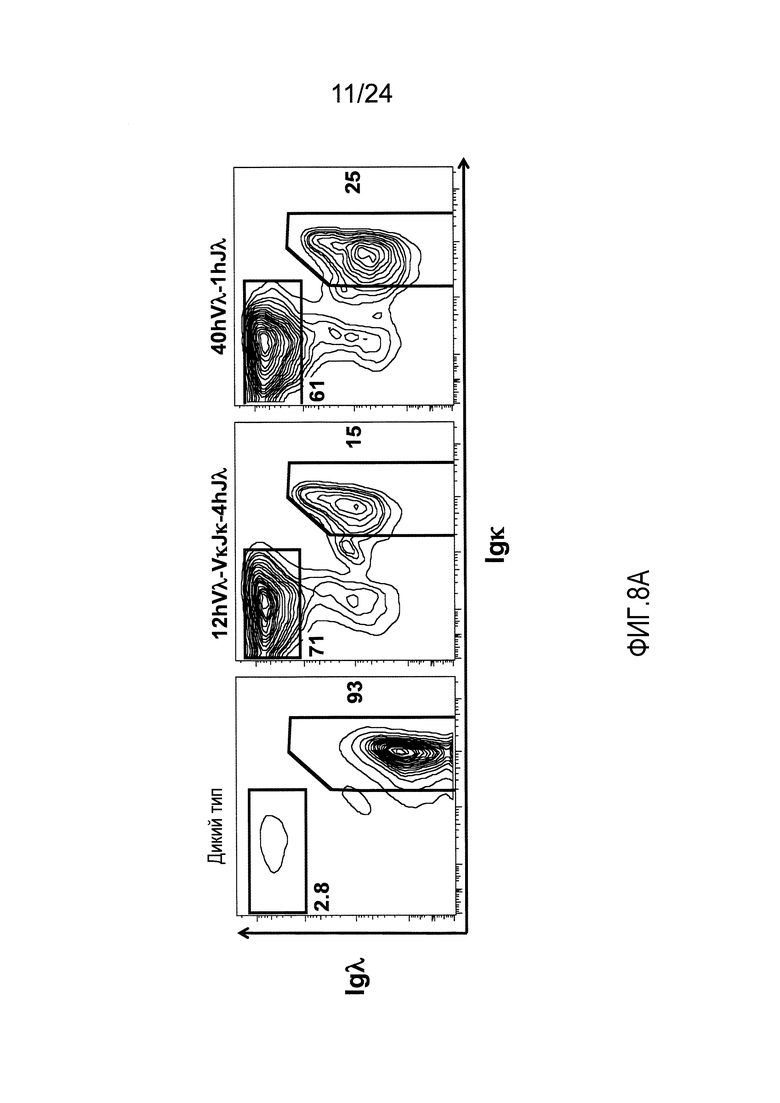
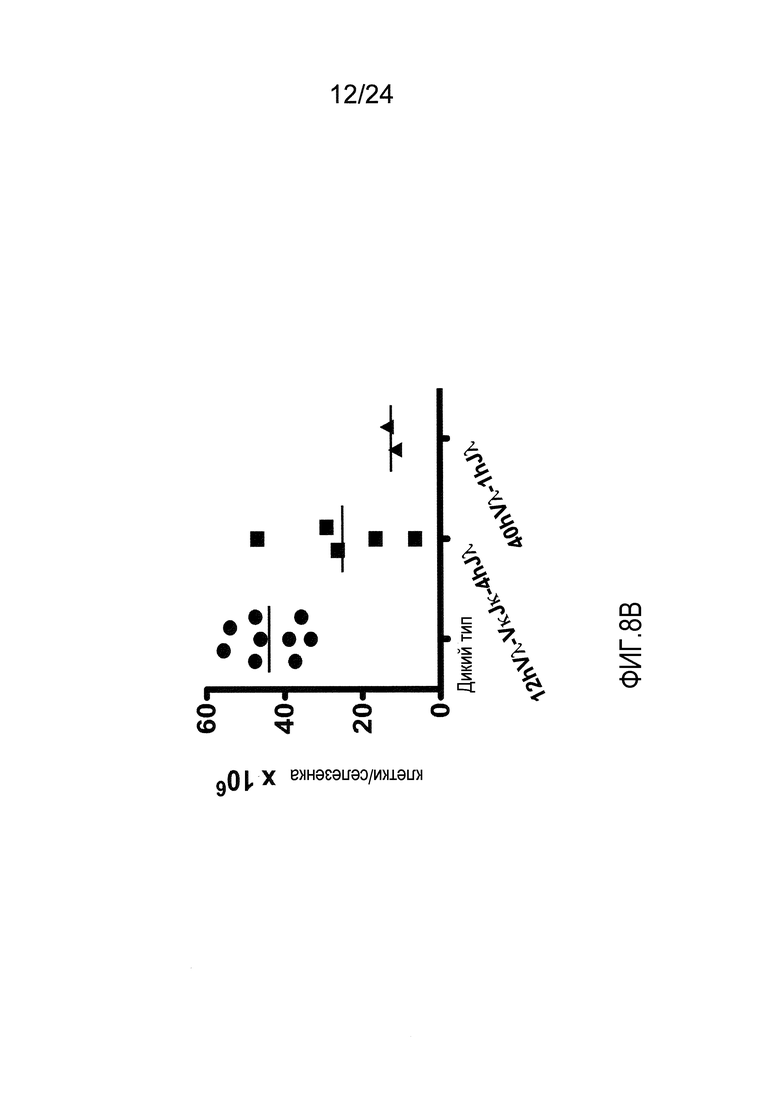
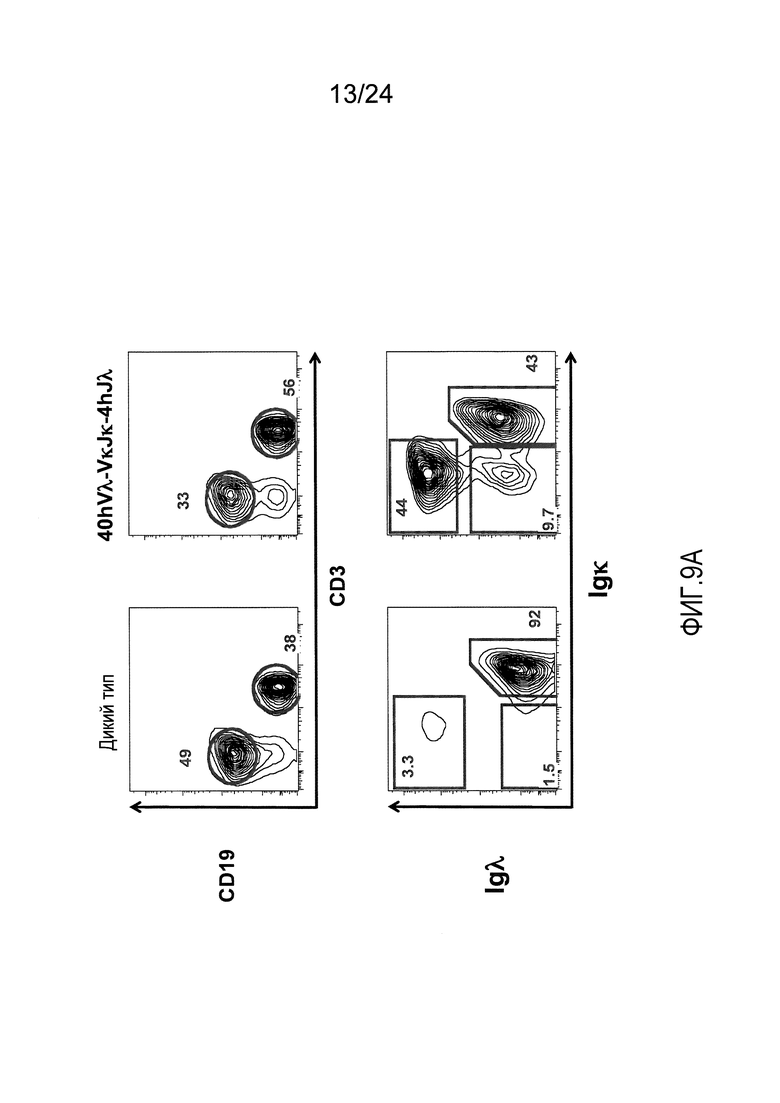
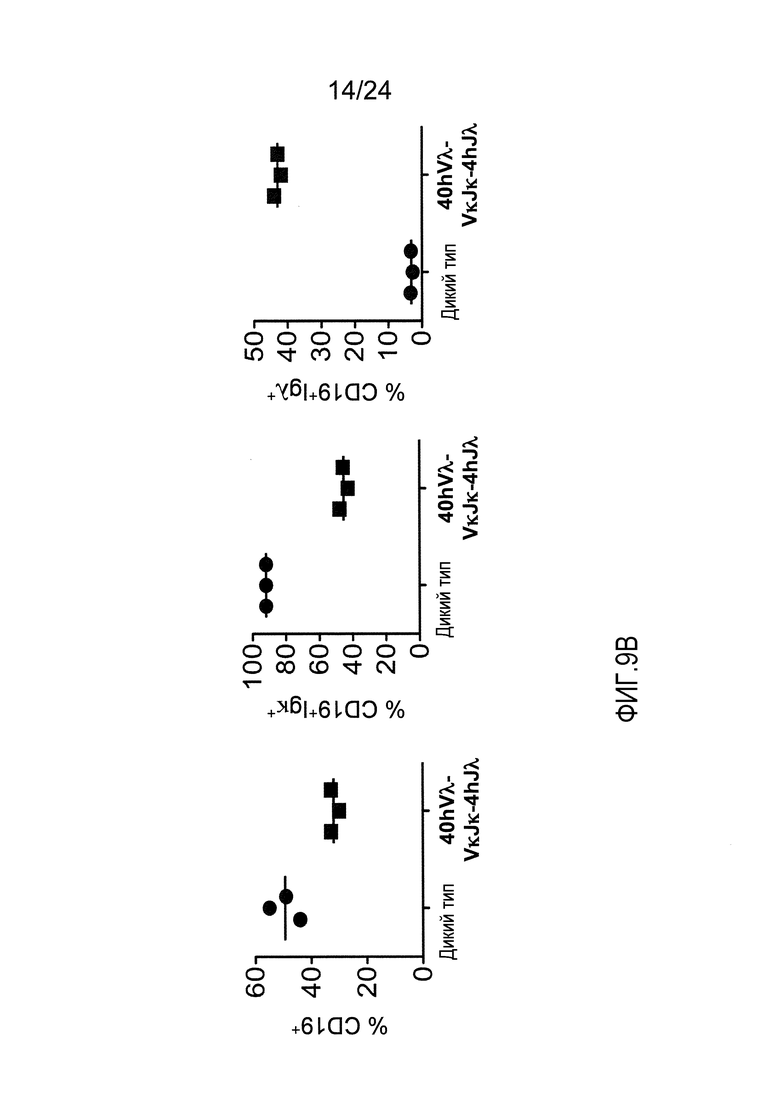
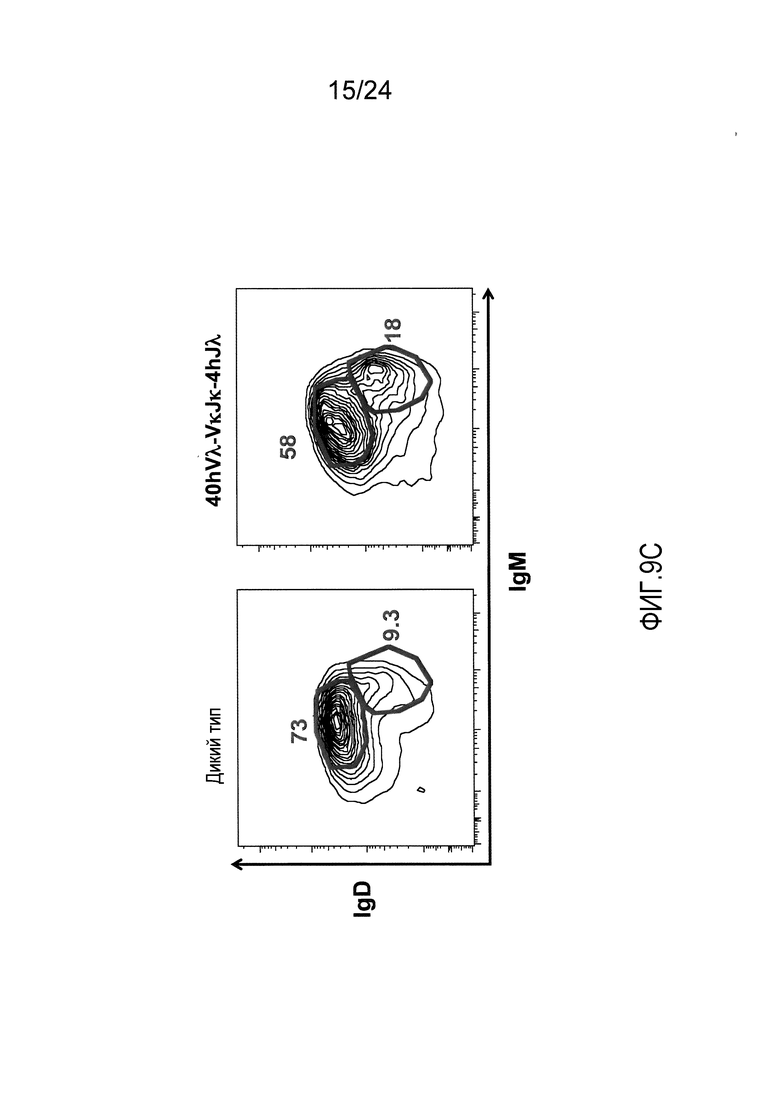
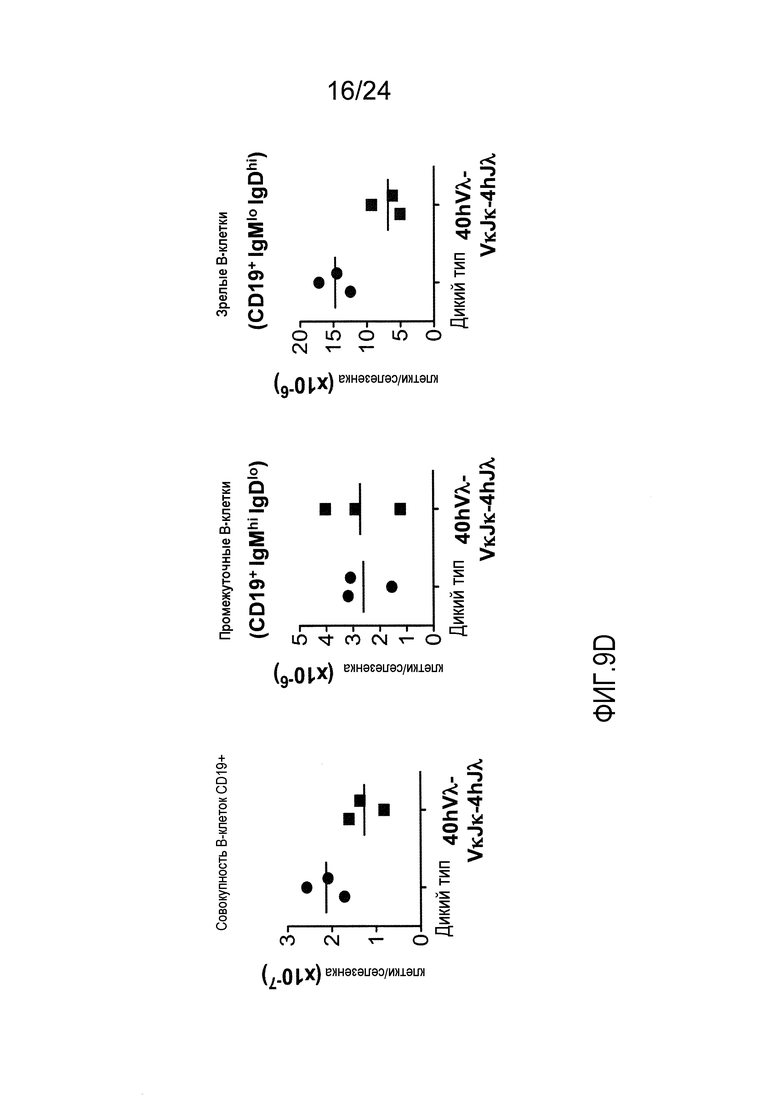
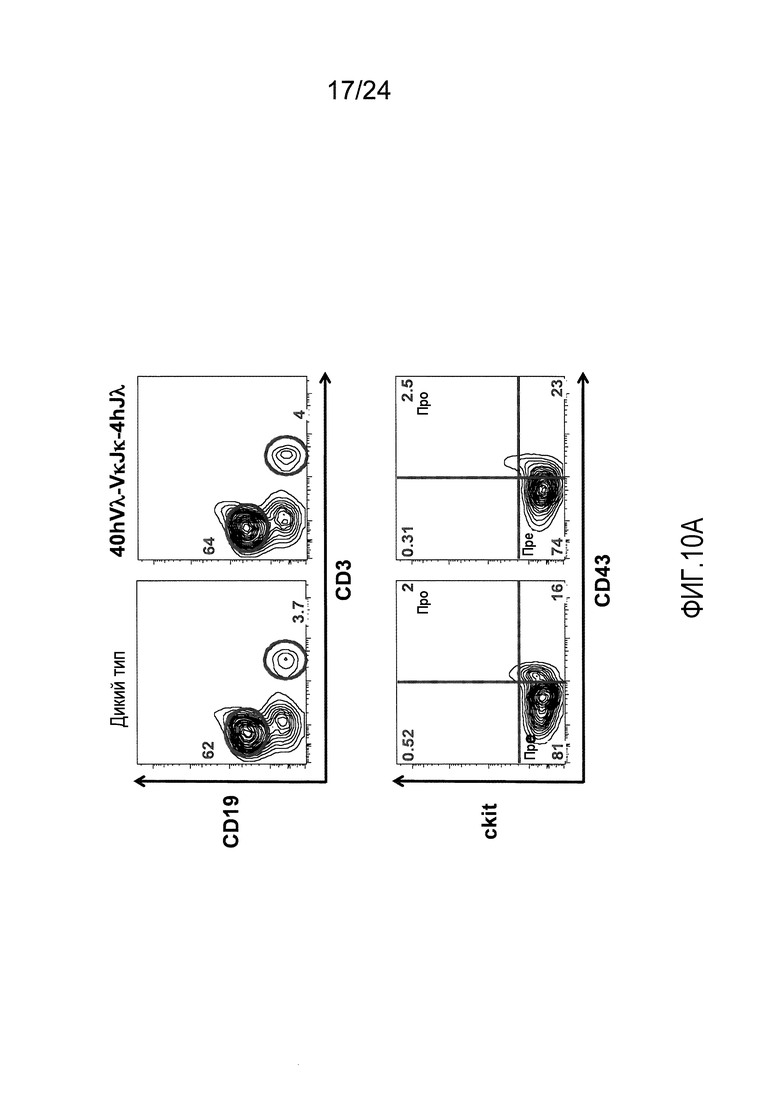
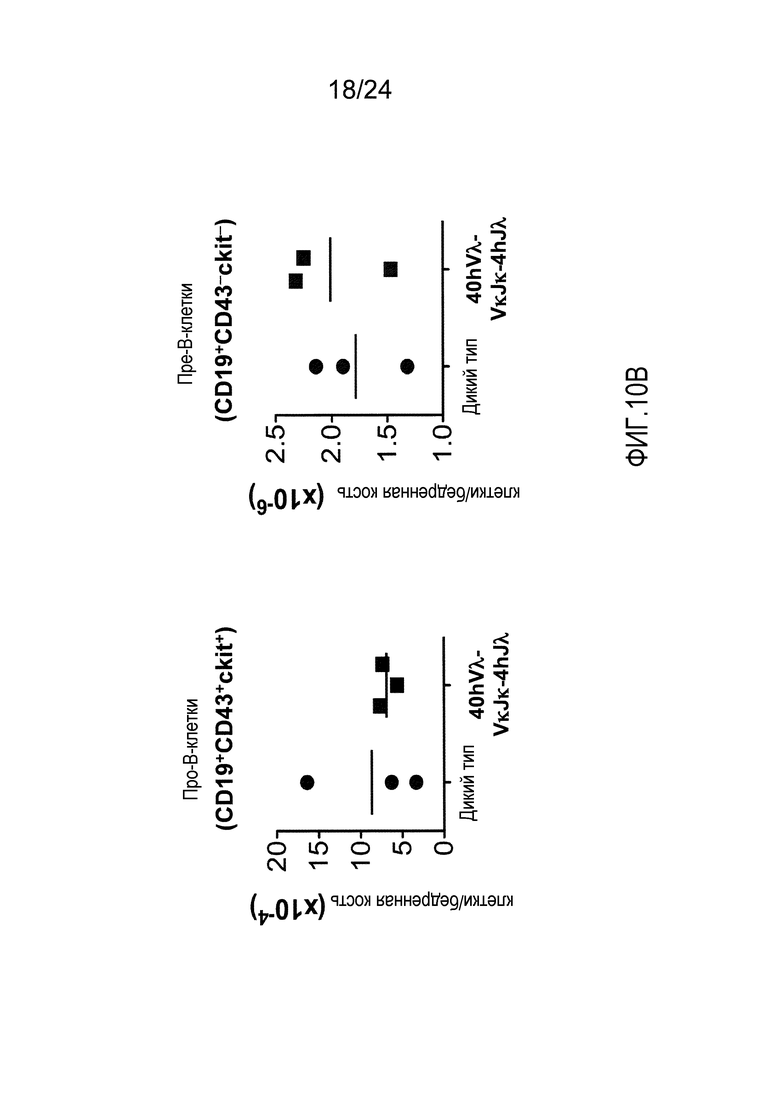
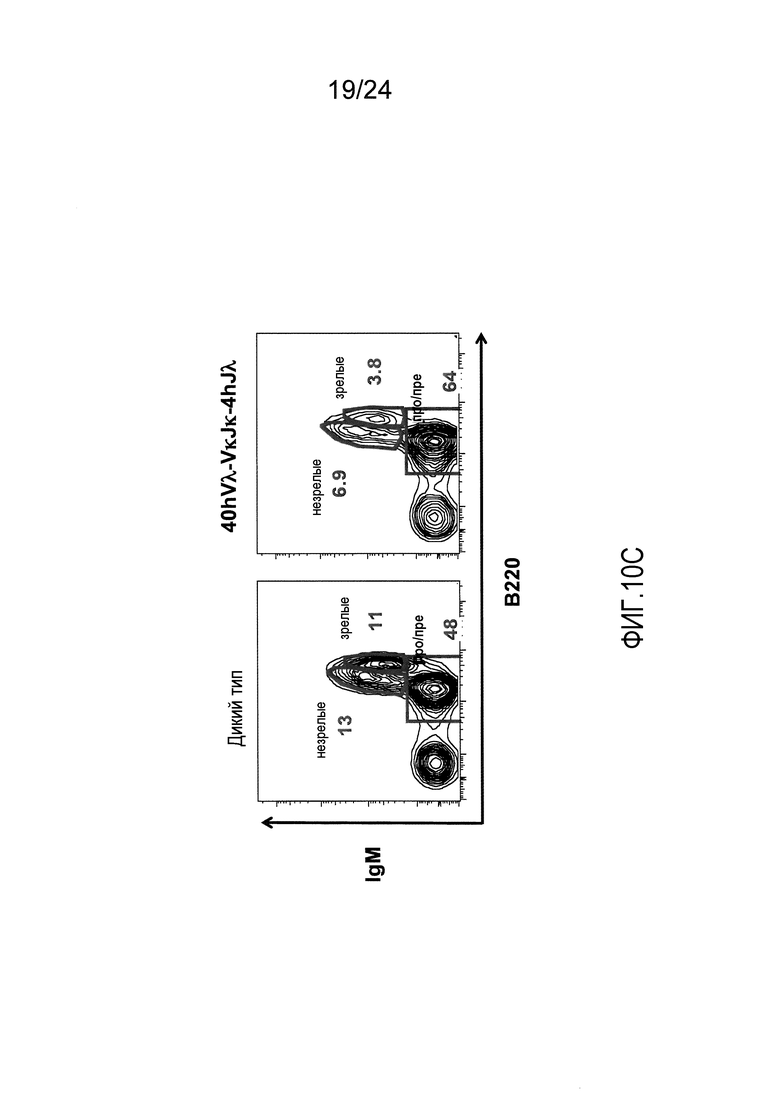
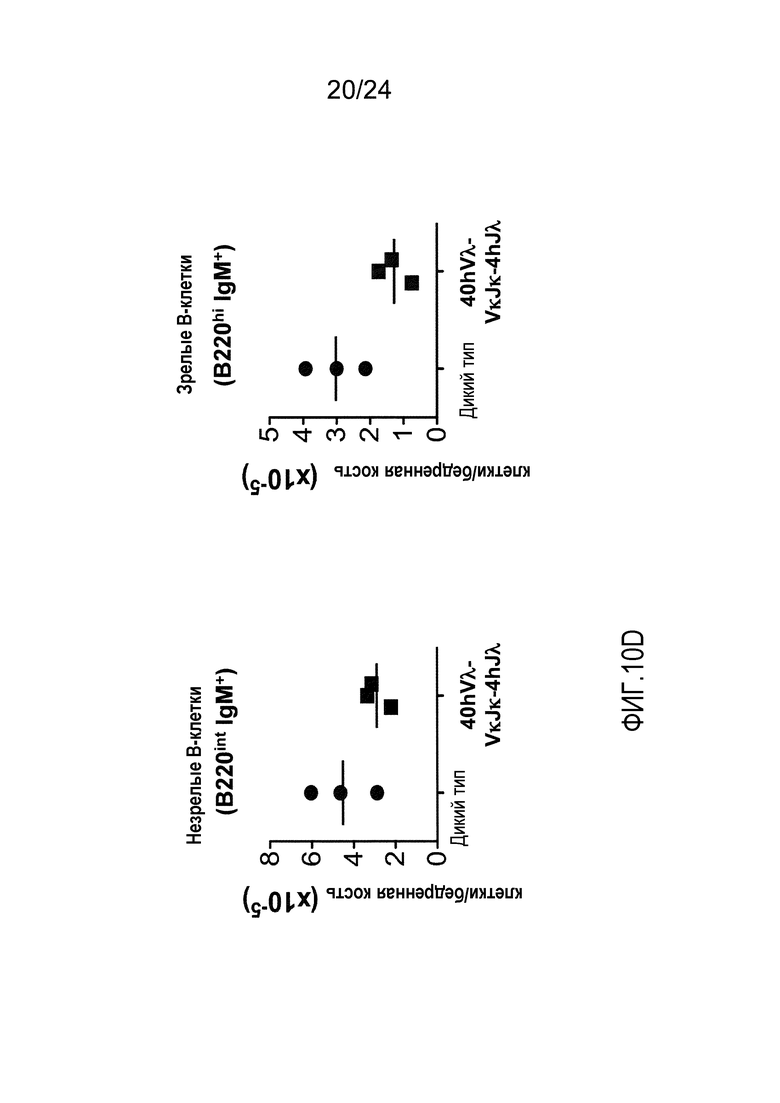
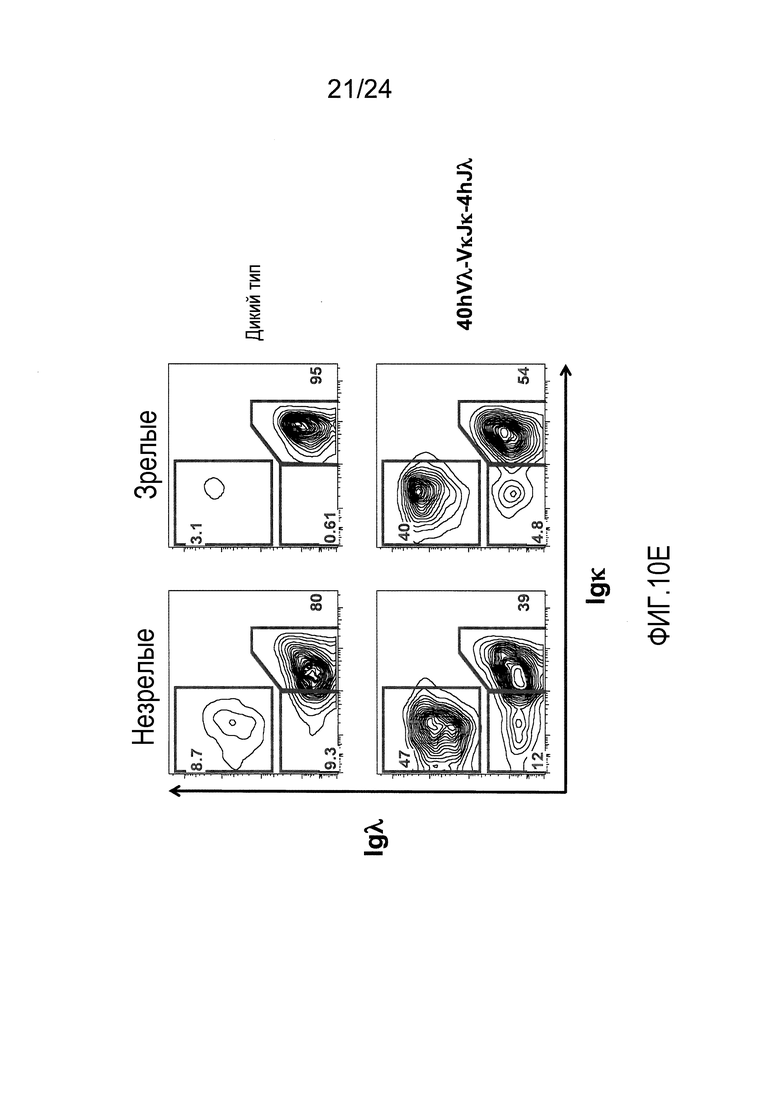


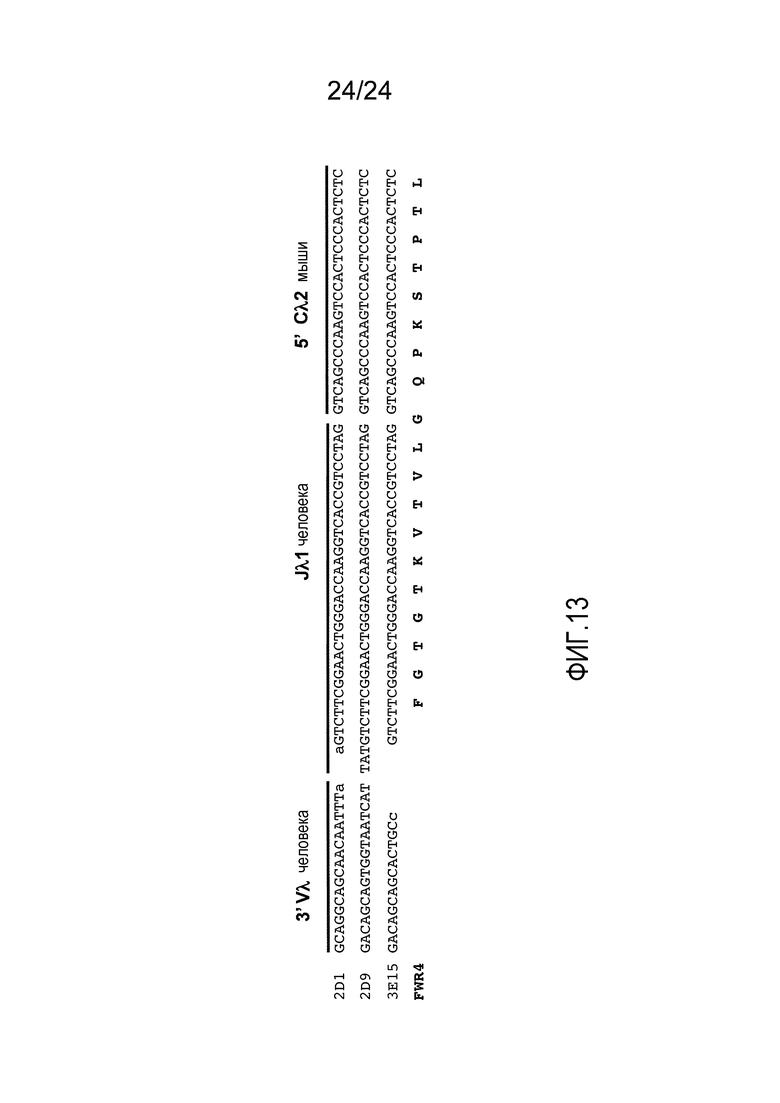





























Изобретение относится к генетически модифицированным мышам, которые экспрессируют вариабельные последовательности λ человека (hVλ), в том числе к мышам, которые экспрессируют последовательности hVλ из эндогенного локуса легкой цепи λ мыши, мышам, которые экспрессируют последовательности hVλ из эндогенного локуса легкой цепи κ мыши, и к мышам, которые экспрессируют последовательности hVλ из трансгена или эписомы, где последовательность hVλ связана с константной последовательностью мыши. Изобретение относится к мышам, которые являются источником соматически видоизмененных вариабельных последовательностей λ человека, подходящих для получения антигенсвязывающих белков. Изобретение относится к композициям и способам получения антигенсвязывающих белков, которые содержат вариабельные последовательности λ человека, в том числе, антител человека. 4 н. и 10 з.п. ф-лы, 24 ил., 8 табл., 7 пр.
1. Мышь, содержащая эндогенный аллель λ (лямбда) легкой цепи, содержащий неперегруппированный генный сегмент Vλ человека и неперегруппированный генный сегмент Jλ человека, причем мышь содержит делецию в первом генном кластере Vλ-Jλ-Cλ эндогенного аллеля λ легкой цепи мыши и замену, полную или частичную, генных сегментов Vλ-Jλ второго генного кластера Vλ-Jλ-Cλ аллеля генными сегментами Vλ и Jλ человека, для получения антител, содержащих вариабельный домен λ человека.
2. Мышь по п. 1, у которой эндогенный аллель λ (лямбда) легкой цепи содержит последовательность константной области, которая представляет собой последовательность константной области Сλ мыши.
3. Мышь по п. 2, которая содержит совокупность генных сегментов Vλ человека.
4. Мышь по любому из предыдущих пунктов, у которой эндогенный аллель λ (лямбда) легкой цепи содержит:
(a) по меньшей мере 12 неперегруппированных генных сегментов Vλ человека; или
(b) 13-28 неперегруппированных генных сегментов Vλ человека.
5. Мышь по п. 4, у которой генные сегменты Vλ человека включают:
(a) hVλ3-1, hVλ4-3, hVλ2-8, hVλ3-9, hVλ3-10, hVλ2-11 и hVλ3-12; или
(b) hVλ2-14, hVλ3-16, hVλ2-18, hVλ3-19, hVλ3-21, hVλ3-22, hVλ2-23, hVλ3-25 и hVλ3-27.
6. Мышь по п. 1, у которой эндогенный аллель λ (лямбда) легкой цепи содержит один неперегруппированный генный сегмент Jλ человека, который представляет собой hJλ1.
7. Мышь по п. 1, у которой эндогенный аллель λ (лямбда) легкой цепи содержит четыре неперегруппированных генных сегмента Jλ человека.
8. Мышь по п. 7, у которой четыре неперегруппированных генных сегмента Jλ человека представляют собой Jλ1, Jλ2, Jλ3 и Jλ7.
9. Мышь по п. 1, у которой эндогенный аллель λ (лямбда) легкой цепи лишен эндогенного генного сегмента Vλ.
10. Мышь по любому из пп 1-3, 6 и 9, у которой эндогенный аллель κ (каппа) легкой цепи делетирован.
11. Мышь по п. 1, у которой эндогенный аллель λ (лямбда) легкой цепи мыши содержит:
(a) непрерывную последовательность локуса λ легкой цепи человека, которая простирается от Vλ3-12 до Vλ3-1;
(b) непрерывную последовательность локуса λ легкой цепи человека, которая простирается от Vλ3-27 до Vλ3-1; или
(c) непрерывную последовательность локуса λ легкой цепи человека, которая простирается от Vλ3-29 до Vλ3-1, и непрерывную последовательность локуса λ легкой цепи человека, которая простирается от Vλ5-52 до Vλ1-40.
12. Выделенная эмбриональная стволовая (ES) клетка мыши, содержащая эндогенный аллель λ (лямбда) легкой цепи, содержащий неперегруппированный генный сегмент Vλ человека и неперегруппированный генный сегмент Jλ человека, где клетка содержит делецию в первом генном кластере Vλ-Jλ-Cλ эндогенного аллеля λ легкой цепи мыши и замену, полную или частичную, генных сегментов Vλ-Jλ второго генного кластера Vλ-Jλ-Сλ аллеля генными сегментами Vλ и Jλ человека, для получения мыши, которая экспрессирует антитела, содержащие вариабельный домен λ легкой цепи человека.
13. Эмбрион мыши, содержащий ES-клетку мыши по п. 12, для получения мыши, которая экспрессирует антитела, содержащие вариабельный домен λ легкой цепи человека.
14. Способ получения антитела в организме мыши, включающий:
(a) воздействие на мышь по п. 1 антигеном;
(b) развитие у мыши иммунного ответа на антиген;
(c) выделение из организма мыши из (b) антитела, которое специфически распознает антиген, или выделение из организма мыши из (b) клетки, содержащей домен иммуноглобулина, который специфически распознает антиген, или выявление в организме мыши из (b) последовательности нуклеиновой кислоты, кодирующей вариабельный домен тяжелой и/или легкой цепи, который связывает антиген, где антитело содержит легкую цепь, происходящую из hVλ, hJλ и гена Сλ мыши.
| Дисковая электростатическая машина | 1930 |
|
SU26373A1 |
| WO 2009143472 A2, 26.11.2009 | |||
| WO 2008054606 A2, 08.05.2008 | |||
| RU 94046095 A1, 27.03.1997. | |||

