[001] Данная заявка испрашивает приоритет по предварительной заявке на выдачу патента США с серийным номером 62/417845, поданной 4 ноября 2016 года, и по предварительной заявке на выдачу патента США с серийным номером 62/567932, поданной 10 октября 2017 года, каждая из которых таким образом включена в данный документ посредством ссылки в полном объеме.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[002] Человеческие антитела представляют собой наиболее быстро растущий класс терапевтических средств. Среди технологий, применяемых в настоящее время для их получения, разработка трансгенных животных (например, грызуны), модифицированных с помощью генетического материала, кодирующего полные или неполные человеческие антитела, совершила переворот в области терапевтических моноклональных человеческих антител для лечения различных заболеваний. До сих пор существует необходимость в разработке улучшенных систем in vivo для получения моноклональных человеческих антител, которые увеличивают до максимума репертуары человеческих антител у трансгенных животных-хозяев.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[003] В определенных аспектах в данном документе предусмотрены улучшенные системы in vivo для идентификации и разработки новых терапевтических средств, представляющих собой антитела, и терапевтических средств на основе антител, которые могут применяться для лечения ряда заболеваний, поражающих людей. Как раскрыто в данном документе, в определенных вариантах осуществления отличные от человека животные (например, грызуны), предусмотренные в данном документе, которые имеют сконструированные локусы иммуноглобулинов, в частности сконструированные локусы легких λ-цепей иммуноглобулинов (Ig), и/или которые иным образом экспрессируют, продуцируют или содержат репертуары антител, характеризующиеся легкими цепями с человеческими Vλ-областями, являются пригодными, например, при разработке разнообразия человеческих Vλ-последовательностей для целей идентификации и разработки новых терапевтических средств на основе антител. В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, обеспечивают улучшенные системы in vivo для разработки антител и/или терапевтических средств на основе антител для введения людям. В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, обеспечивают улучшенные системы in vivo для разработки антител и/или терапевтических средств на основе антител, которые содержат человеческие Vλ-домены, характеризующиеся улучшенными характеристиками по сравнению с антителами и/или терапевтическими средствами на основе антител, полученными из существующих систем in vivo, которые содержат последовательности человеческих Vλ-областей.
[004] В определенных аспектах в данном документе предусматривается отличное от человека животное с локусом легкой λ-цепи Ig, который содержит сконструированные вариабельную и константную области иммуноглобулина; в некоторых определенных вариантах осуществления дополнительно содержит сконструированную регуляторную область (или последовательность). Как описано в данном документе, в определенных вариантах осуществления предусмотренные отличные от человека животные содержат в своем геноме зародышевого типа локус легкой λ-цепи Ig, содержащий сконструированную вариабельную область легкой λ-цепи Ig, характеризующуюся наличием одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ, одного или более человеческих генов Cλ-области и гена Cλ-области грызуна, при этом сегменты человеческих генов Vλ, Jλ и Cλ функционально связаны друг с другом и функционально связаны с указанным геном Cλ-области грызуна.
[005] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или по меньшей мере 25 сегментов человеческого гена Vλ.
[006] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 5-25, 5-24, 5-23, 5-22, 5-21, 5-20, 5-19, 5-18, 5-17, 5-16, 5-15, 5-14, 5-13, 5-12, 5-11, 5-10, 5-9, 5-8, 5-7 или 5-6 сегментов человеческого гена Vλ. В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 10-70, 10-69, 10-68, 10-67, 10-66, 10-65, 10-64, 10-63, 10-62, 10-61, 10-60, 10-59, 10-58, 10-57, 10-56, 10-55, 10-54, 10-53, 10-52, 10-51, 10-50, 10-49, 10-48, 10-47, 10-46, 10-45, 10-44, 10-43, 10-42, 10-41, 10-40, 10-39, 10-38, 10-37, 10-36, 10-35, 10-34, 10-33, 10-32, 10-31, 10-32, 10-31, 10-30, 10-29, 10-28, 10-27, 10-26, 10-25, 10-24, 10-23, 10-22, 10-21, 10-20, 10-19, 10-18, 10-17, 10-16, 10-15, 10-14, 10-13, 10-12 или 10-11 сегментов человеческого гена Vλ.
[007] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 6-25, 7-25, 8-25, 9-25, 10-25, 11-25, 12-25, 13-25, 14-25, 15-25, 16-25, 17-25, 18-25, 19-25, 20-25, 21-25, 22-25, 23-25 или 24-25 сегментов человеческого гена Vλ. В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 11-70, 12-70, 13-70, 14-70, 15-70, 16-70, 17-70, 18-70, 19-70, 20-70, 21-70, 22-70, 23-70, 24-70, 25-70, 26-70, 27-70, 28-70, 29-70, 30-70, 31-70, 32-70, 33-70, 34-70, 35-70, 36-70, 37-70, 38-70, 39-70, 40-70, 41-70, 42-70, 43-70, 44-70, 45-70, 46-70, 47-70, 48-70, 49-70, 50-70, 51-70, 52-70, 53-70, 54-70, 55-70, 56-70, 57-70, 58-70, 59-70, 60-70, 61-70, 62-70, 63-70, 64-70, 65-70, 66-70, 67-70, 68-70 или 69-70 сегментов человеческого гена Vλ.
[008] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 6-24, 7-23, 8-22, 9-21, 10-20, 11-19, 12-18, 13-17, 14-16 или 15-16 сегментов человеческого гена Vλ. В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 11-69, 12-68, 13-67, 14-66, 15-65, 16-64, 17-63, 18-62, 19-61, 20-60, 21-59, 22-58, 23-57, 24-56, 25-55, 26-54, 27-53, 28-52, 29-51, 30-50, 31-49, 32-48, 33-47, 34-48, 35-47, 36-46, 37-45, 38-44, 39-43, 40-42 или 41-42 сегмента человеческого гена Vλ.
[009] В определенных вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 5, 16 или 25 функциональных сегментов человеческого гена Vλ. В определенных вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит 10, 27 или 40 сегментов человеческого гена Vλ. В определенных вариантах осуществления сегменты человеческого гена Vλ предусматривают последовательные сегменты человеческого гена Vλ таким образом, как указанные сегменты человеческого гена Vλ находятся в человеческом локусе легкой λ-цепи Ig в клетке человека.
[0010] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит по меньшей мере 5 сегментов человеческого гена Jλ (например, без ограничения 5 сегментов человеческого гена Jλ, 6 сегментов человеческого гена Jλ, 7 сегментов человеческого гена Jλ, 8 сегментов человеческого гена Jλ и т.д.). В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит по меньшей мере 4 человеческих гена Cλ-области (например, без ограничения 4 человеческих гена Cλ-области, 5 человеческих генов Cλ-области, 6 человеческих генов Cλ-области, 7 человеческих генов Cλ-области, 8 человеческих генов Cλ-области и т.д.). В определенных вариантах осуществления предусмотренные отличные от человека животные содержат локус легкой λ-цепи Ig, который содержит по меньшей мере 25 сегментов человеческого гена Vλ, по меньшей мере 5 сегментов человеческого гена Jλ и по меньшей мере 4 человеческих гена Cλ-области в эндогенном аллеле легкой λ-цепи Ig. В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат только один мышиный (например, мышь или крыса) ген Cλ-области (например, ген Cλ1-области мыши или сегмент гена Cλ1 мыши) в эндогенном отличном от человеческого локусе легкой λ-цепи Ig. В некоторых вариантах осуществления указанный локус легкой λ-цепи Ig дополнительно содержит человеческую Eλ-область (или последовательность), которая характеризуется тремя элементами последовательности.
[0011] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат сегменты человеческих генов Vλ, Jλ и Cλ в эндогенном отличном от человеческого локусе легкой λ-цепи Ig в природной конфигурации или конфигурации зародышевого типа. В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат сегменты человеческих генов Vλ, Jλи Cλ в эндогенном отличном от человеческого локусе легкой λ-цепи Ig в конфигурации, которая в природе не присутствует в человеческом локусе легкой λ-цепи иммуноглобулина генома зародышевого типа клетки человека.
[0012] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат последовательность ДНК в эндогенном отличном от человеческого локусе легкой λ-цепи Ig, который включает множество кодирующих человеческие Vλ, Jλ и Cλ последовательностей, чередующихся (или расположенных рядом, связанных и т.д.) с некодирующей последовательностью человеческой легкой λ-цепи иммуноглобулина. В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат последовательность ДНК в эндогенном отличном от человеческого локусе легкой λ-цепи Ig, который включает множество кодирующих человеческие Vλ, Jλ и Cλ последовательностей, чередующихся с некодирующей отличной от человеческой (например, мышиная) последовательностью легкой λ-цепи иммуноглобулина.
[0013] В некоторых вариантах осуществления предусмотренные отличные от человека животные характеризуются экспрессией антител из эндогенных отличных от человеческих локусов легких λ-цепей Ig в геноме зародышевого типа указанных отличных от человека животных, при этом антитела содержат человеческие Vλ- домены и человеческие или отличные от человеческих Cλ-домены. В некоторых вариантах осуществления предусмотренные отличные от человека животные характеризуются повышенным использованием человеческих Vλ-областей из сконструированных локусов легких λ-цепей иммуноглобулина (например, при соотношении 60:40 κ:λ) по сравнению с одним или более отличными от человека эталонными модифицированными животными или животными дикого типа (например, без ограничения при соотношении 95:5 κ:λ).
[0014] В некоторых вариантах осуществления предусмотрены отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, геном которых содержит эндогенный локус легкой λ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ, Jλ и Cλ функционально связаны с сегментом отличного от человеческого гена Cλ, и данный эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более отличных от человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина и один или более человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина.
[0015] В некоторых вариантах осуществления предусмотрены отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, геном зародышевого типа которых содержит эндогенный локус легкой λ-цепи иммуноглобулина, содержащий: (a) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, где (a) и (b) функционально связаны с (c) и сегментом отличного от человеческого гена Cλ, и где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит: один или более отличных от человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина и один или более человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина.
[0016] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, дополнительно содержит три человеческих Eλ. В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один человеческий Eλ, характеризующийся наличием трех элементов последовательности. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один человеческий Eλ, характеризующийся наличием трех элементов последовательности, которые действуют (или функционируют) модульным образом.
[0017] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, содержит два отличных от человеческих Eλ. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит два Eλ грызуна. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, содержит два Eλ мыши. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит Eλ мыши и Eλ3-1 мыши. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, не содержит (или в нем отсутствует) Eλ2-4 мыши. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит два Eλ крысы.
[0018] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, представленный в данном документе, предусматривает делецию сегментов эндогенных генов Vλи Jλ, полностью или частично. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, представленный в данном документе, предусматривает делецию генных сегментов Vλ2-Vλ3-Jλ2-Cλ2 и генных сегментов Vλ1-Jλ3-Cλ3-Jλ1. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P и генных сегментов Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1. В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, представленный в данном документе, предусматривает делецию отличных от человеческих Eλ2-4. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, представленный в данном документе, предусматривает делецию Vλ2, Vλ3, Jλ2, Cλ2, Jλ4P, Cλ4P, Eλ2-4, Vλ1, Jλ3, Jλ3P, Cλ3 и Jλ1. В некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе содержит Cλ1, Eλ и Eλ3-1, если присутствуют только отличные от человеческих сегменты генов или элементы последовательности.
[0019] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, содержит вставку сегментов человеческого гена Vλ от Vλ4-69 до Vλ3-1, по меньшей мере пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, сегмента человеческого гена Jλ, Jλ7, и сегмента гена Cλ1 грызуна. В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, содержит вставку сегментов человеческого гена Vλ от Vλ5-52 до Vλ3-1, по меньшей мере пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, сегмента человеческого гена Jλ, Jλ7, и сегмента гена Cλ1 грызуна. В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина, предусмотренный в данном документе, содержит вставку сегментов человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, по меньшей мере пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, сегмента человеческого гена Jλ, Jλ7, и сегмента гена Cλ1 грызуна. В некоторых определенных вариантах осуществления вставка предусматривает некодирующую ДНК человека, которая встречается в природе между человеческими Vλ5-52 и Vλ1-40 и Vλ3-27 и Vλ3-1, некодирующую ДНК человека, которая в природе встречается между парами сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, и некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно сегмента человеческого гена Jλ, Jλ7.
[0020] В некоторых вариантах осуществления сегмент отличного от человеческого гена Cλ представляет собой сегмент гена Cλ грызуна или содержит его. В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент мышиного (например, мышь или крыса) гена Cλ или содержит его. В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ крысы или содержит его. В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ мыши или содержит его. В некоторых определенных вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1мыши.
[0021] В некоторых вариантах осуществления ген (или сегмент гена) Cλ мыши содержит последовательность, которая на по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% идентична или на 100% идентична гену Cλ мыши, выбранному из группы, состоящей из Cλ1 мыши, Cλ2 мыши и Cλ3 мыши. В некоторых вариантах осуществления ген Cλ мыши содержит последовательность, которая практически идентична или идентична гену Cλ мыши, выбранному из группы, состоящей из Cλ1 мыши, Cλ2 мыши и Cλ3 мыши. В некоторых определенных вариантах осуществления ген Cλ1 мыши представляет собой SEQ ID NO: 1 или содержит ее. В некоторых определенных вариантах осуществления ген Cλ2 мыши представляет собой SEQ ID NO: 3 или содержит ее. В некоторых определенных вариантах осуществления ген Cλ3 мыши представляет собой SEQ ID NO: 5 или содержит ее. В некоторых определенных вариантах осуществления ген Cλ мыши содержит последовательность, которая идентична гену Cλ1мыши.
[0022] В некоторых вариантах осуществления ген (или сегмент гена) Cλ мыши содержит последовательность, которая на 50%-100%, 55%-100%, 60%-100%, 65%-100%, 70%-100%, 75%-100%, 80%-100%, 85%-100%, 90%-100%, 95%-100% или 98%-100% идентична гену Cλ мыши, выбранному из группы, состоящей из Cλ1 мыши, Cλ2 мыши и Cλ3 мыши.
[0023] В некоторых вариантах осуществления ген (или сегмент гена) Cλ мыши содержит последовательность, которая на 50%-98%, 50%-95%, 50%-90%, 50%-85%, 50%-80%, 50%-75%, 50%-70%, 50%-65%, 50%-60% или 50%-55% идентична гену Cλ мыши, выбранному из группы, состоящей из Cλ1 мыши, Cλ2 мыши и Cλ3 мыши.
[0024] В некоторых вариантах осуществления ген (или сегмент гена) Cλ мыши содержит последовательность, которая на 55%-98%, 60%-95%, 65%-90%, 70%-85% или 75%-80% идентична гену Cλ мыши, выбранному из группы, состоящей из Cλ1 мыши, Cλ2 мыши и Cλ3 мыши.
[0025] В некоторых вариантах осуществления ген (или сегмент гена) Cλ крысы содержит последовательность, которая на по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% идентична или на 100% идентична гену Cλ крысы, выбранному из группы, состоящей из гена Cλ1 крысы, Cλ2 крысы, Cλ3 крысы и Cλ4 крысы. В некоторых вариантах осуществления ген Cλ крысы содержит последовательность, которая практически идентична или идентична гену Cλ крысы, выбранному из группы, состоящей из Cλ1 крысы, Cλ2 крысы, Cλ3 крысы и Cλ4 крысы. В некоторых определенных вариантах осуществления ген Cλ1 крысы представляет собой SEQ ID NO: 7 или содержит ее. В некоторых определенных вариантах осуществления ген Cλ2 крысы представляет собой SEQ ID NO: 9 или содержит ее. В некоторых определенных вариантах осуществления ген Cλ3 крысы представляет собой SEQ ID NO: 11 или содержит ее. В некоторых определенных вариантах осуществления ген Cλ4 крысы представляет собой SEQ ID NO: 13 или содержит ее.
[0026] В некоторых вариантах осуществления ген (или сегмент гена) Cλ крысы содержит последовательность, которая на 50%-100%, 55%-100%, 60%-100%, 65%-100%, 70%-100%, 75%-100%, 80%-100%, 85%-100%, 90%-100%, 95%-100% или 98%-100% идентична гену Cλ крысы, выбранному из группы, состоящей из гена Cλ1 крысы, Cλ2 крысы, Cλ3 крысы и Cλ4 крысы.
[0027] В некоторых вариантах осуществления ген (или сегмент гена) Cλ крысы содержит последовательность, которая на 50%-98%, 50%-95%, 50%-90%, 50%-85%, 50%-80%, 50%-75%, 50%-70%, 50%-65%, 50%-60% или 50%-55% идентична гену Cλ крысы, выбранному из группы, состоящей из гена Cλ1 крысы, Cλ2 крысы, Cλ3 крысы и Cλ4 крысы.
[0028] В некоторых вариантах осуществления ген (или сегмент гена) Cλ крысы содержит последовательность, которая на 55%-98%, 60%-95%, 65%-90%, 70%-85% или 75%-80% идентична гену Cλ крысы, выбранному из группы, состоящей из гена Cλ1 крысы, Cλ2 крысы, Cλ3 крысы и Cλ4 крысы.
[0029] В некоторых вариантах осуществления предусмотренных отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани, геном зародышевого типа или геном указанных отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани дополнительно содержит (i) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с отличной от человеческой константной областью тяжелой цепи иммуноглобулина; или (ii) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с отличной от человеческой константной областью тяжелой цепи иммуноглобулина, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с отличной от человеческой Cκ-областью иммуноглобулина.
[0030] В некоторых вариантах осуществления вставленные один или более сегментов человеческого гена VH, один или более сегментов человеческого гена DH и один или более сегментов человеческого гена JH заменяют сегменты отличных от человеческих генов VH, DH. В определенных вариантах осуществления вставка предусматривает некодирующую ДНК человека, которая встречается в природе между сегментами человеческих VH, DH и JH и их комбинациями. В некоторых вариантах осуществления отличная от человеческой константная область тяжелой цепи иммуноглобулина представляет собой эндогенную отличную от человеческой константную область тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина содержит вставку сегментов человеческого гена VH от VH3-74 до VH6-1, сегментов человеческого гена DH от DH1-1 до DH7-27 и сегментов человеческого гена JH, JH1-JH6. В определенных вариантах осуществления вставка предусматривает некодирующую ДНК человека, которая встречается (находится) в природе между человеческими VH3-74 и VH6-1, некодирующую ДНК человека, которая встречается (находится) в природе между человеческими DH1-1 и DH7-27, и некодирующую ДНК человека, которая встречается (находится) в природе между человеческими JH1 и JH6. В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина содержит вставку всех функциональных сегментов человеческого гена VH, всех функциональных сегментов человеческого гена DH и всех функциональных сегментов человеческого гена JH.
[0031] В некоторых вариантах осуществления в локусе тяжелой цепи иммуноглобулина отсутствует эндогенный отличный от человеческого ген Adam6. В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более отличных от человеческих полипептидов Adam6. В некоторых вариантах осуществления одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены между первым и вторым сегментами человеческого гена VH. В некоторых вариантах осуществления первый сегмент человеческого гена VH представляет собой человеческий VH1-2, и второй сегмент человеческого гена VH представляет собой человеческий VH6-1. В некоторых вариантах осуществления одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены между сегментом человеческого гена VH и сегментом человеческого гена DH. В некоторых вариантах осуществления одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены вместо человеческого псевдогена Adam6.
[0032] В некоторых вариантах осуществления вставленные один или более сегментов человеческого гена Vκ и один или более сегментов человеческого гена Jκ заменяют сегменты отличных от человеческих генов Vκ и Jκ. В некоторых определенных вариантах осуществления вставка предусматривает некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов Vκ и Jκ и их комбинациями. В некоторых вариантах осуществления отличная от человеческой Cκ-область иммуноглобулина представляет собой эндогенную отличную от человеческой Cκ-область. В некоторых вариантах осуществления локус легкой κ-цепи иммуноглобулина содержит вставку проксимальной дупликации Vκ, полностью или частично, человеческого локуса легкой κ-цепи иммуноглобулина. В некоторых вариантах осуществления локус легкой κ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vκ от Vκ 2-40 до Vκ4-1 и сегментов человеческого гена Jκ от Jκ1 до Jκ5. В некоторых определенных вариантах осуществления вставка предусматривает некодирующую ДНК человека, которая встречается в природе между человеческими Vκ2-40 и Vκ4-1, и некодирующую ДНК человека, которая встречается в природе между человеческими Jκ1 и Jκ5.
[0033] В некоторых вариантах осуществления отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани, предусмотренных в данном документе, отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань являются гетерозиготными или гомозиготными в отношении локуса тяжелой цепи иммуноглобулина, описанного в данном документе (например, эндогенного локуса тяжелой цепи иммуноглобулина, описанного в данном документе).
[0034] В некоторых вариантах осуществления отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани, предусмотренных в данном документе, отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань являются гетерозиготными или гомозиготными в отношении локуса легкой κ-цепи иммуноглобулина, описанного в данном документе (например, эндогенного локуса легкой κ-цепи иммуноглобулина, описанного в данном документе).
[0035] В некоторых вариантах осуществления отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани, предусмотренных в данном документе, отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань являются гетерозиготными или гомозиготными в отношении локуса легкой λ-цепи иммуноглобулина, описанного в данном документе (например, эндогенного локуса легкой λ-цепи иммуноглобулина, описанного в данном документе).
[0036] В некоторых вариантах осуществления отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани, предусмотренных в данном документе, геном зародышевого типа указанных отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более отличных от человеческих полипептидов Adam6, и при этом животное является гетерозиготным или гомозиготным в отношении указанной вставки.
[0037] В некоторых вариантах осуществления отличная от человеческой клетка представляет собой отличный от человеческого лимфоцит. В некоторых вариантах осуществления отличная от человеческой клетка выбрана из B-клетки, дендритной клетки, макрофага, моноцита и T-клетки.
[0038] В некоторых вариантах осуществления отличная от человеческой клетка представляет собой отличную от человеческой эмбриональную стволовую (ES) клетку. В некоторых вариантах осуществления отличная от человеческой ES-клетка представляет ES-клетку грызуна. В определенных вариантах осуществления ES- клетка грызуна представляет собой ES-клетку мыши (например, из линии 129, линии C57BL, BALB/c или их помеси). В некоторых определенных вариантах осуществления эмбриональная стволовая клетка грызуна представляет собой эмбриональную стволовую клетку мыши и является помесью линий 129 и C57BL. В некоторых определенных вариантах осуществления эмбриональная стволовая клетка грызуна представляет собой эмбриональную стволовую клетку мыши и является помесью линий 129, C57BL и BALB/c.
[0039] В некоторых вариантах осуществления предусмотрено применение отличной от человеческой ES-клетки, описанной в данном документе, для получения отличного от человека животного. В определенных вариантах осуществления отличная от человеческой ES-клетка представляет собой ES-клетку мыши и применяется для получения мыши, содержащей сконструированный локус легкой λ-цепи иммуноглобулина, описанный в данном документе. В определенных вариантах осуществления отличная от человеческой ES-клетка представляет собой ES-клетку крысы и применяется для получения крысы, содержащей сконструированный локус легкой λ-цепи иммуноглобулина, описанный в данном документе.
[0040] В некоторых вариантах осуществления отличная от человеческой ткань выбрана из адипозной ткани, ткани мочевого пузыря, головного мозга, молочной железы, костного мозга, глаза, сердца, кишечника, почки, печени, легкого, лимфатического узла, мышцы, поджелудочной железы, плазмы крови, сыворотки крови, кожи, ткани селезенки, желудка, вилочковой железы, яичка, яйцеклетки и их комбинации.
[0041] В некоторых вариантах осуществления предусмотрена иммортализованная клетка, созданная, образованная, произведенная или полученная из выделенной отличной от человеческой клетки или ткани, описанной в данном документе.
[0042] В некоторых вариантах осуществления предусмотрен отличный от человеческого эмбрион, образованный, созданный, произведенный или полученный из отличной от человеческой ES-клетки, описанной в данном документе. В некоторых определенных вариантах осуществления отличный от человеческого эмбрион представляет собой эмбрион грызуна; в некоторых вариантах осуществления - эмбрион мыши; в некоторых вариантах осуществления - эмбрион крысы.
[0043] В некоторых вариантах осуществления представлен набор, предусматривающий отличное от человека животное, отличную от человеческой клетку или отличную от человеческой ткань, иммортализованную клетку, отличную от человеческой ES-клетку, отличный от человеческого эмбрион, описанные в данном документе.
[0044] В некоторых вариантах осуществления предусмотрен набор, описанный в данном документе, для применения в производстве и/или разработке лекарственного средства (например, антитело или его фрагмент) для терапии или диагностики.
[0045] В некоторых вариантах осуществления предусмотрен набор, описанный в данном документе, для применения в производстве и/или разработке лекарственного средства (например, антитело или его фрагмент) для лечения, предупреждения или уменьшения интенсивности проявления заболевания, нарушения или состояния.
[0046] В некоторых вариантах осуществления предусмотрен способ получения отличного от человека животного, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи иммуноглобулина, при этом способ включает: (a) введение фрагмента ДНК в отличную от человеческой эмбриональную стволовую клетку, при этом указанный фрагмент ДНК содержит нуклеотидную последовательность, которая включает (i) один или более сегментов человеческого гена Vλ, (ii) один или более сегментов человеческого гена Jλ и (iii) один или более сегментов человеческого гена Cλ, где (i)-(iii) функционально связаны с сегментом отличного от человеческого гена Cλ, и где нуклеотидная последовательность дополнительно содержит один или более человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина; (b) получение отличной от человеческой эмбриональной стволовой клетки, созданной на (a); и (c) получение отличного от человека животного с применением отличной от человеческой эмбриональной стволовой клетки с (b).
[0047] В некоторых вариантах осуществления предусмотрен способ получения отличного от человека животного, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи иммуноглобулина, при этом сконструированный эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом отличного от человеческого и/или человеческого гена Cλ, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более отличных от человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина и один или более человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина; при этом способ включает модификацию генома зародышевого типа отличного от человека животного таким образом, что оно содержит сконструированный локус легкой λ-цепи иммуноглобулина, который содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом отличного от человеческого и/или человеческого гена Cλ, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более человеческих энхансеров легкой λ-цепи (Eλ) иммуноглобулина, с получением, тем самым, указанного отличного от человека животного.
[0048] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, один или более сегментов человеческого гена Vλ предусматривают от Vλ4-69 до Vλ3-1, от Vλ5-52 до Vλ3-1 или от Vλ3-27 до Vλ3-1. В некоторых вариантах осуществления способа получения отличного от человека животного один или более сегментов человеческого гена Vλ предусматривают от Vλ5-52 до Vλ1-40 и/или от Vλ3-27 до Vλ3-1. В некоторых определенных вариантах осуществления способа получения отличного от человека животного один или более сегментов человеческого гена Vλ предусматривают некодирующую ДНК человека, которая встречается в природе между человеческими Vλ5-52 и Vλ1-40 и/или Vλ3-27 и Vλ3-1. В некоторых вариантах осуществления способа получения отличного от человека животного один или более сегментов человеческого гена Jλ и один или более сегментов человеческого гена Cλ предусматривают пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3- Cλ3, Jλ6-Cλ6, и человеческий генный сегмент Jλ7. В некоторых определенных вариантах осуществления способа получения отличного от человека животного пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержат некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλ и Cλ, и человеческий генный сегмент Jλ7 содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[0049] В некоторых определенных вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, вставка сегментов человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1 содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческого гена Vλ, вставка пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержит некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλ-Cλ, и вставка сегмента человеческого гена Jλ7 содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[0050] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, сегмент отличного от человеческого гена Cλ представляет собой сегмент гена Cλ грызуна; в некоторых определенных вариантах осуществления сегмент гена Cλ1 мыши.
[0051] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, фрагмент ДНК дополнительно содержит один или более маркеров отбора. В некоторых вариантах осуществления способа получения отличного от человека животного фрагмент ДНК дополнительно содержит один или более участков сайт-специфической рекомбинации. В некоторых определенных вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, фрагмент ДНК дополнительно содержит одну или более совокупностей участков сайт-специфической рекомбинации, которые рекомбинируют с помощью одной и той же рекомбиназы. В некоторых определенных вариантах осуществления способа получения отличного от человека животного фрагмент ДНК дополнительно содержит одну или более совокупностей участков сайт-специфической рекомбинации, которые рекомбинируют с помощью разных рекомбиназ.
[0052] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, фрагмент ДНК вводят в отличную от человеческой эмбриональную стволовую клетку, геном зародышевого типа которой содержит (i) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с отличной от человеческой константной областью тяжелой цепи иммуноглобулина; или (ii) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с отличной от человеческой константной областью тяжелой цепи иммуноглобулина, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с отличной от человеческой Cκ-областью иммуноглобулина.
[0053] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, фрагмент ДНК вводят в отличную от человеческой эмбриональную стволовую клетку, геном зародышевого типа которой содержит (i) эндогенный локус тяжелой цепи иммуноглобулина дикого типа или (ii) эндогенный локус тяжелой цепи иммуноглобулина дикого типа и эндогенный локус легкой κ-цепи иммуноглобулина дикого типа; и где способ дополнительно включает стадию скрещивания мыши, созданной, образованной, произведенной или полученной из указанной отличной от человеческой эмбриональной стволовой клетки, со второй мышью.
[0054] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, модификацию генома зародышевого типа отличного от человека животного, с тем чтобы оно содержало сконструированный локус легкой λ-цепи иммуноглобулина, проводят в отличной от человеческой эмбриональной стволовой клетке, геном зародышевого типа которой содержит (i) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с отличной от человеческой константной областью тяжелой цепи иммуноглобулина; или (ii) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с отличной от человеческой константной областью тяжелой цепи иммуноглобулина, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с отличной от человеческой Cκ-областью иммуноглобулина.
[0055] В некоторых определенных вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, вставка одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH предусматривает некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена VH, некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена DH, и некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена JH. В некоторых определенных вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, вставка одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ предусматривает некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена Vκ, и некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена Jκ.
[0056] В некоторых вариантах осуществления способа получения отличного от человека животного, предусмотренного в данном документе, модификацию генома зародышевого типа отличного от человека животного, с тем чтобы оно содержало сконструированный локус легкой λ-цепи иммуноглобулина, проводят в отличной от человеческой эмбриональной стволовой клетке, геном зародышевого типа которой содержит (i) эндогенный локус тяжелой цепи иммуноглобулина дикого типа или (ii) эндогенный локус тяжелой цепи иммуноглобулина дикого типа и эндогенный локус легкой κ-цепи иммуноглобулина дикого типа; и где способ дополнительно включает стадию скрещивания мыши, созданной, образованной, произведенной или полученной из указанной отличной от человеческой эмбриональной стволовой клетки, со второй мышью.
[0057] В некоторых вариантах осуществления мышь, описанная в данном документе, имеет геном зародышевого типа, содержащий локусы дикого типа H-цепи Ig и κ-цепи Ig, гомозиготные или гетерозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный или гетерозиготный гуманизированный локус H-цепи Ig содержит вставленную кодирующую Adam6 последовательность грызуна, или гомозиготный или гетерозиготный гуманизированный локус H-цепи Ig (со вставкой кодирующей Adam6 последовательности или без нее) и гомозиготный или гетерозиготный инактивированный локус κ-цепи Ig.
[0058] В некоторых вариантах осуществления предусмотрено отличное от человека животное, созданное, образованное, произведенное, полученное или получаемое по способу, описанному в данном документе.
[0059] В некоторых вариантах осуществления предусмотрен способ получения антитела в отличном от человека животном, при этом способ включает стадии (a) иммунизации отличного от человека животного, описанного в данном документе, с помощью представляющего интерес антигена, (b) содержания отличного от человека животного в условиях, достаточных для того, чтобы у грызуна выработался иммунный ответ на представляющий интерес антиген, и (c) выделения антитела из отличного от человека животного или отличной от человеческой клетки, которое связывает представляющий интерес антиген. В некоторых вариантах осуществления антитело содержит человеческий вариабельный домен легкой лямбда-цепи.
[0060] В некоторых вариантах осуществления предусмотрен способ получения нуклеиновой кислоты, кодирующей человеческий вариабельный домен легкой лямбда-цепи, в отличном от человека животном, при этом способ включает стадии: (a) иммунизации отличного от человека животного, описанного в данном документе, с помощью представляющего интерес антигена, (b) содержания отличного от человека животного в условиях, достаточных для того, чтобы у грызуна выработался иммунный ответ на представляющий интерес антиген, и (c) выделения нуклеиновой кислоты, кодирующей человеческий вариабельный домен легкой лямбда-цепи, из отличного от человека животного или отличной от человеческой клетки. В некоторых вариантах осуществления способ дополнительно включает выделение нуклеиновой кислоты, кодирующей человеческий вариабельный домен тяжелой цепи, из отличного от человека животного или отличной от человеческой клетки.
[0061] В некоторых вариантах осуществления способа получения антитела или нуклеиновой кислоты в отличном от человека животном отличная от человеческой клетка представляет собой B-клетку. В некоторых вариантах осуществления способа получения антитела или нуклеиновой кислоты в отличном от человека животном отличная от человеческой клетка представляет собой гибридому.
[0062] В некоторых вариантах осуществления способа получения антитела в отличном от человека животном антитело, выделенное из грызуна или клетки грызуна, которое связывает представляющий интерес антиген, содержит человеческий вариабельный домен тяжелой цепи и человеческий вариабельный домен легкой лямбда-цепи.
[0063] В некоторых вариантах осуществления способа получения антитела или нуклеиновой кислоты в отличном от человека животном человеческий вариабельный домен тяжелой цепи содержит перегруппированный сегмент человеческого гена VH, выбранный из группы, состоящей из VH3-74, VH3-73, VH3-72, VH2-70, VH1-69, VH3-66, VH3-64, VH4-61, VH4-59, VH1-58, VH3-53, VH5-51, VH3-49, VH3-48, VH1-46, VH1-45, VH3-43, VH4-39, VH4-34, VH3-33, VH4-31, VH3-30, VH4-28, VH2-26, VH1-24, VH3-23, VH3-21, VH3-20, VH1-18, VH3-15, VH3-13, VH3-11, VH3-9, VH1-8, VH3-7, VH2-5, VH7-4-1, VH4-4, VH1-3, VH1-2 и VH6-1.
[0064] В некоторых вариантах осуществления способа получения антитела или нуклеиновой кислоты в отличном от человека животном человеческий вариабельный домен легкой лямбда-цепи содержит перегруппированный сегмент человеческого гена Vλ, выбранный из группы, состоящей из Vλ4-69, Vλ8-61, Vλ4-60, Vλ6-57, Vλ10-54, Vλ5-52, Vλ1-51, Vλ9-49, Vλ1-47, Vλ7-46, Vλ5-45, Vλ1-44, Vλ7-43, Vλ1-40, Vλ5-39, Vλ5-37, Vλ1-36, Vλ3-27, Vλ3-25, Vλ2-23, Vλ3-22, Vλ3-21, Vλ3-19, Vλ2-18, Vλ3-16, Vλ2-14, Vλ3-12, Vλ2-11, Vλ3-10, Vλ3-9, Vλ2-8, Vλ4-3 и Vλ3-1.
[0065] В некоторых вариантах осуществления предусмотрен способ индукции антиген-специфического иммунного ответа у отличного от человека животного, при этом способ включает стадии (a) иммунизации отличного от человека животного, описанного в данном документе, с помощью представляющего интерес антигена, (b) содержания отличного от человека животного в условиях, достаточных для того, чтобы у грызуна выработался иммунный ответ на представляющий интерес антиген.
[0066] В некоторых вариантах осуществления предусмотрено отличное от человека животное, геном зародышевого типа которого содержит гомозиготный эндогенный локус легкой λ-цепи иммуноглобулина, содержащий вставку (i) сегментов человеческого гена Vλ от Vλ4-69 до Vλ3-1, от Vλ5-52 до Vλ3-1, от Vλ3-27 до Vλ3-1 или от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, (ii) пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, (iii) сегмента человеческого гена Jλ, Jλ7, и (iv) трех человеческих энхансеров легкой λ-цепи иммуноглобулина (или человеческого энхансера легкой λ-цепи иммуноглобулина с тремя элементами последовательности); где (i)-(iv) функционально связаны друг с другом, и вставка находится против хода транскрипции относительно сегмента отличного от человеческого гена Cλ, и где в эндогенном локусе легкой λ-цепи иммуноглобулина отсутствует эндогенный отличный от человеческого Eλ2-4 иммуноглобулина.
[0067] В некоторых вариантах осуществления предусмотрено отличное от человека животное, геном зародышевого типа которого содержит гомозиготный эндогенный локус легкой λ-цепи иммуноглобулина, содержащий: (i) сегменты человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, (ii) пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, (iii) сегмент человеческого гена Jλ, Jλ7, и (iv) три энхансера человеческой легкой λ-цепи иммуноглобулина (или энхансер человеческой легкой λ-цепи иммуноглобулина с тремя элементами последовательности); где (i)-(iv) функционально связаны друг с другом, и (i)-(iii) находится против хода транскрипции (или в 5'-направлении) относительно сегмента отличного от человеческого гена Cλ, и где в эндогенном локусе легкой λ-цепи иммуноглобулина отсутствует эндогенный отличный от человеческого Eλ2-4 иммуноглобулина, сегменты человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1 содержат некодирующую ДНК человека, которая встречается в природе между сегментами человеческого гена Vλ, пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержат некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλ-Cλ, и сегмент человеческого гена Jλ, Jλ7, содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[0068] В некоторых определенных вариантах осуществления предусмотренного отличного от человека животного отличный от человеческого ген (или сегмент отличного от человеческого гена) Cλ представляет собой ген (или сегмент гена) Cλ1 мыши. В некоторых определенных вариантах осуществления предусмотренного отличного от человека животного эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит эндогенные отличные от человеческих энхансеры легкой λ-цепи иммуноглобулина, Eλ и Eλ3-1. В некоторых определенных вариантах осуществления предусмотренного отличного от человека животного эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию эндогенных отличных от человеческих генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P и эндогенных отличных от человеческих генных сегментов Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1.
[0069] В некоторых вариантах осуществления отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, предусмотрены для применения в изготовлении и/или разработке лекарственного средства (например, антитело или его фрагмент) для терапии или диагностики.
[0070] В некоторых вариантах осуществления отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, предусмотрены для применения в изготовлении лекарственного препарата для лечения, предупреждения или уменьшения интенсивности проявления заболевания, нарушения или состояния.
[0071] В некоторых вариантах осуществления предусмотрено применение отличного от человека животного, отличной от человеческой клетки или отличной от человеческой ткани, описанных в данном документе, в изготовлении и/или разработке лекарственного средства или вакцины для применения в медицине, как, например, для применения в качестве лекарственного препарата.
[0072] В некоторых вариантах осуществления предусмотрено применение отличного от человека животного или отличной от человеческой клетки, описанных в данном документе, в изготовлении и/или разработке антитела или его фрагмента.
[0073] В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, представляют собой грызуна, клетку грызуна или ткань грызуна; в некоторых вариантах осуществления мышь, клетку мыши или ткань мыши; в некоторых вариантах осуществления крысу, клетку крысы или ткань крысы. В некоторых вариантах осуществления мышь, клетка мыши или ткань мыши, описанные в данном документе, предусматривают генетический фон, который включает линию 129, линию BALB/c, линию C57BL/6, смешанную линию 129xC57BL/6 или их комбинации.
[0074] Используемые в данной заявке термины «приблизительно» и «примерно» применяются как эквиваленты. Предполагается, что все числа, используемые в настоящей заявке, вместе с терминами «приблизительно» или «примерно» или без них, охватывают любые нормальные отклонения, понятные специалисту в соответствующей области техники.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0075] Графические материалы, включенные в данный документ, которые состоят из последующих фигур, приведены только в иллюстративных целях, а не для ограничения.
[0076] На фигуре 1 показано схематическое изображение, без соблюдения масштаба, иллюстративной стратегии построения сконструированного эндогенного локуса легкой λ-цепи Ig у грызуна, характеризующегося наличием множества кодирующих человеческие Vλ, Jλ и Cλ последовательностей, которые функционально связаны друг с другом и функционально связаны с Cλ-областью грызуна (или гена Cλ грызуна). Как изображено, показано пять отдельных нацеливающих векторов (6286, 6571, 6596, 6597 и 6680) с различными количествами генетического материала из человеческого локуса легкой λ-цепи Ig и их последовательно вставляют в эндогенный локус легкой λ-цепи Ig грызуна (например, мышь) (показано вверху). Первый нацеливающий вектор (6286) вставлен по ходу транскрипции относительно Cλ1-области грызуна и построен так, чтобы содержать модульную энхансерную область (или последовательность) человеческой λ-цепи (Eλ) Ig, характеризующуюся тремя элементами последовательности. Второй нацеливающий вектор (6571) вставлен против хода транскрипции относительно Cλ1-области грызуна и сконструирован так, чтобы содержать пять функциональных сегментов человеческого гена Vλ, четыре пары функциональных сегментов человеческих генов Jλ-Cλ и человеческий генный сегмент Jλ7 (человеческие Jλ1-Cλ1-Jλ2-Cλ2-Jλ3-Cλ3-Jλ4-Cλ4-Jλ5-Cλ5-Jλ6-Cλ6-Jλ7). Третий (6596) и четвертый (6597) нацеливающие векторы содержали добавочные совокупности дополнительных сегментов человеческого гена Vλ (одиннадцать и девять соответственно), которые последовательно добавляются к общему содержанию сегментов человеческого гена Vλ эндогенного локуса легкой λ-цепи Ig мыши после успешного нацеливания первого нацеливающего вектора. Оба нацеливающих вектора содержали области перекрывания (заштрихованные прямоугольники) на 3'-концах для облегчения гомологичной рекомбинации с 5'-концом предыдущего нацеливающего вектора, уже интегрированного в эндогенный локус легкой λ-цепи Ig мыши. Также показан альтернативный пятый нацеливающий вектор (6680), который содержит тот же генетический материал, что и нацеливающий вектор 6597, за исключением того, что этот альтернативный нацеливающий вектор содержал 5'-плечо гомологии с последовательностью, идентичной последовательности в 5'-направлении (или против хода транскрипции) относительно генного сегмента Vλ2 грызуна, тем самым облегчая делецию эндогенных генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P-Eλ2-4-Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1 при гомологичной рекомбинации с нацеливающим вектором. Если не указано иное, закрашенные символы обозначают сегменты генов и/или последовательности грызуна, тогда как незакрашенные символы обозначают сегменты человеческих генов и/или человеческие последовательности. Также показаны участки распознавания сайт-специфической рекомбинации (например, loxP, Frt), фланкирующие кассеты для отбора (HYG: ген устойчивости к гигромицину [HYGR] под транскрипционным контролем промотора убиквитина; NEO: ген устойчивости к неомицину [NEOR] под транскрипционным контролем промотора убиквитина). Выбранные месторасположения участков сочленения нуклеотидов отмечены линией под каждым участком сочленения, и каждый обозначен с помощью SEQ ID NO.
[0077] На фигуре 2 показано схематическое изображение, без соблюдения масштаба, иллюстративных аллелей легкой λ-цепи Ig грызуна после последовательной вставки нацеливающих векторов, описанных в примере 1. Аллель 6597: аллель легкой λ-цепи Ig, который содержит 25 функциональных сегментов человеческого гена Vλ, четыре пары функциональных сегментов человеческих генов Jλ-Cλ и человеческий генный сегмент Jλ7, функционально связанные с Cλ-областью грызуна (например, Cλ1-область мыши), и при этом локус легкой λ-цепи Ig дополнительно содержит эндогенные сегменты генов Vλ-Jλ-Cλ, три (т.е. E2.4, E и E3.1) эндогенных энхансерных области (или последовательности) λ-цепи Ig и модульную человеческую энхансерную область (или последовательность) λ-цепи Ig, характеризующуюся тремя элементами последовательности. Аллель 6680: аллель легкой λ-цепи Ig после сайт-специфической делеции эндогенных сегментов генов Vλ-Jλ-Cλ и энхансера λ-цепи Ig, Eλ2-4, при этом аллель легкой λ-цепи Ig содержит 25 функциональных сегментов человеческого гена Vλ, четыре пары функциональных сегментов человеческих генов Jλ-Cλ и человеческий генный сегмент Jλ7, функционально связанные с Cλ-областью грызуна (например, Cλ1-область мыши), при этом локус легкой λ-цепи Ig дополнительно содержит две (т.е. E и E3.1) эндогенные энхансерные области (или последовательности) λ-цепи Ig и модульную человеческую энхансерную область (или последовательность, см. выше) λ-цепи Ig. Если не указано иное, закрашенные символы обозначают сегменты генов и/или последовательности грызуна, тогда как незакрашенные символы обозначают сегменты человеческих генов и/или человеческие последовательности. Показаны участки распознавания сайт-специфической рекомбинации (например, Frt), фланкирующие кассеты для отбора (HYG: ген устойчивости к гигромицину [HYGR] под транскрипционным контролем промотора убиквитина). Пунктирными линиями отмечена удаленная область между двумя проиллюстрированными аллелями λ-цепи Ig. Выбранные месторасположения участков сочленения нуклеотидов отмечены линией под каждым участком сочленения, и каждый обозначен с помощью SEQ ID NO.
[0078] На фигуре 3 показано схематическое изображение, без соблюдения масштаба, альтернативной иллюстративной стратегии построения сконструированного эндогенного локуса легкой λ-цепи Ig у грызуна, характеризующегося наличием множества кодирующих человеческие Vλ, Jλ и Cλ последовательностей, которые функционально связаны друг с другом и функционально связаны с Cλ-областью грызуна. Как изображено, показаны два различных нацеливающих вектора с разными количествами генетического материала из человеческого локуса легкой λ-цепи Ig и они одновременно вставлены в сконструированный локус легкой λ-цепи Ig грызуна (например, мышь) (показано вверху), который содержит пять сегментов человеческого гена Vλ, кластер человеческих Jλ-Cλ и ген Cλ1 мыши. Нацеливающий вектор 6596 модифицирован с удалением кассеты для отбора по устойчивости к неомицину и введением перекрывающихся последовательностей (заштрихованные прямоугольники) на 5'- и 3'-концах с получением областей гомологии для облегчения рекомбинации с соответствующей человеческой последовательностью. Второй нацеливающий вектор сконструирован так, чтобы содержать перекрывающуюся область на 3'-конце конструкции (заштрихованные прямоугольники), которая обладает общей гомологией последовательности с модифицированным нацеливающим вектором 6596 (усеченным нацеливающим вектором 6596), что облегчает гомологичную рекомбинацию с 5'-концом усеченного нацеливающего вектора 6596. Эти два нацеливающих вектора содержат добавочные совокупности дополнительных сегментов человеческого гена Vλ (одиннадцать и девять соответственно), которые последовательно добавляются к общему содержанию сегментов человеческого гена Vλ эндогенного локуса легкой λ-цепи Ig мыши после успешного нацеливания первого нацеливающего вектора. Второй нацеливающий вектор содержал 5'-плечо гомологии с последовательностью, идентичной последовательности в 5'-направлении (или против хода транскрипции) относительно генного сегмента Vλ2 грызуна, тем самым облегчая делецию эндогенных генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P-Eλ2-4-Vλ1-Jλ3-Jλ3P-Cλ3- Jλ1 при гомологичной рекомбинации с нацеливающим вектором. Два нацеливающих вектора вводили совместно электропорацией с помощью направляющих РНК (gRNA) для облегчения интеграции в сконструированном локусе легкой λ-цепи Ig, которые отмечены стрелками возле месторасположения каждой последовательность, и каждая обозначена с помощью SEQ ID NO. Если не указано иное, закрашенные символы обозначают сегменты генов и/или последовательности грызуна, тогда как незакрашенные символы обозначают сегменты человеческих генов и/или человеческие последовательности. Также показаны участки распознавания сайт- специфической рекомбинации (например, loxP, Frt), фланкирующие кассеты для отбора (HYG: ген устойчивости к гигромицину [HYGR] под транскрипционным контролем промотора убиквитина; NEO: ген устойчивости к неомицину [NEOR] под транскрипционным контролем промотора убиквитина). Выбранные месторасположения участков сочленения нуклеотидов отмечены линией под каждым участком сочленения, и каждый обозначен с помощью SEQ ID NO.
[0079] На фигуре 4 показано схематическое изображение, без соблюдения масштаба, аллеля дикого типа и иллюстративного сконструированного аллеля легкой λ-цепи Ig грызуна у грызунов, применяемых в экспериментах, описанных в примере 3. Аллель дикого типа: локус дикого типа легкой λ-цепи Ig мыши (см. также, например, фигуру 2 из патента США №9006511); аллель 6571: аллель легкой λ-цепи Ig, который содержит 5 функциональных сегментов человеческого гена Vλ, четыре пары функциональных сегментов человеческих генов Jλ-Cλ и человеческий генный сегмент Jλ7, функционально связанные с Cλ-областью грызуна (например, Cλ1-область мыши), и при этом локус легкой λ-цепи Ig дополнительно содержит эндогенные сегменты генов Vλ-Jλ-Cλ, три эндогенных энхансерных области (или последовательности) λ-цепи Ig и модульную человеческую энхансерную область (или последовательность, см. выше) λ-цепи Ig. Аллель 6597: см. выше; аллель 6680: см. выше. Выбранные месторасположения участков сочленения нуклеотидов отмечены линией под каждым участком сочленения, и каждый обозначен с помощью SEQ ID NO.
[0080] На фигурах 5A и 5B показаны репрезентативные контурные графики, на которых обозначены гейтированные по одиночным клеткам спленоциты (A), демонстрирующие экспрессию CD19 (ось y) и CD3 (ось x), и абсолютное число клеток на селезенку (B), собранное у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и у мышей дикого типа (WT) из одного помета.
[0081] На фигурах 6A и 6B показаны репрезентативные контурные графики, на которых обозначены зрелые и переходные В-клетки в спленоцитах, гейтированных по CD19+ (A), демонстрирующих экспрессию IgD (ось y) и IgM (ось x), и абсолютное число клеток на селезенку (B), собранное у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и у мышей дикого типа (WT) из одного помета. Конкретные субпопуляции B-клеток указаны на каждом точечном графике (например, зрелые, переходные).
[0082] На фигурах 7A и 7B показаны репрезентативные контурные графики, на которых обозначена экспрессия λ-цепи Ig мыши (mIgλ, ось y), κ-цепи Ig мыши (mIgκ, ось x) или человеческой λ-цепи Ig (hIgλ, ось y) в гейтированных по CD19+ спленоцитах, собранных у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета.
[0083] На фигурах 8A и 8B показаны репрезентативные контурные графики, на которых обозначены гейтированный по одиночным клеткам костный мозг (A), демонстрирующий экспрессию CD19 (ось y) и CD3 (ось x), и абсолютное число клеток на бедренную кость (B), собранное у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета.
[0084] На фигурах 9A и 9B показаны репрезентативные контурные графики, на которых обозначены гейтированный по CD19+IgMlowB220int костный мозг (A), демонстрирующий экспрессию c-набора (ось y) и CD43 (ось x), и абсолютное число клеток на бедренную кость (B), собранное у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета. Конкретные субпопуляции B-клеток указаны на каждом точечном графике (например, про-B-клетки, пре-B-клетки).
[0085] На фигурах 10A и 10B показаны репрезентативные контурные графики, на которых обозначены гейтированный по CD19+ костный мозг (A), демонстрирующий экспрессию IgM (ось y) и B220 (ось x), и абсолютное число клеток на бедренную кость (B), собранное у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета. Конкретные субпопуляции B-клеток указаны на каждом точечном графике (например, незрелые, зрелые, пре- и про-B-клетки).
[0086] На фигурах 11A и 11B показаны репрезентативные контурные графики, на которых обозначен незрелый костный мозг (гейтированный по CD19+IgM+B220int), демонстрирующий экспрессию λ-цепи Ig мыши (mIgλ, ось y), κ-цепи Ig мыши (mIgκ, ось x) или человеческой λ-цепи Ig (hIgλ, ось y) у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета.
[0087] На фигурах 12A и 12B показаны репрезентативные контурные графики, на которых обозначен зрелый костный мозг (гейтированный по CD19+IgM+B220+), демонстрирующий экспрессию λ-цепи Ig мыши (mIgλ, ось y), κ-цепи Ig мыши (mIgκ, ось x) или человеческой λ-цепи Ig (hIgλ, ось y) у мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета.
[0088] На фигуре 13 показан репрезентативный средний процент экспрессирующих κ-цепь Ig (% κ C) и экспрессирующих человеческую λ-цепь Ig (% hum λ C) B-клеток в селезенке, незрелом костном мозге (незрелом BM) и зрелом костном мозге (зрелом BM) у выбранных сконструированных линий мышей, описанных в данном документе. Данные представлены в виде средних значений с также указанным стандартным отклонением. 6680HO/VI HO/Adam6 HO: сконструированная линия мышей, содержащих гомозиготный сконструированный локус легкой λ-цепи Ig, сконструированный так, чтобы содержать 25 функциональных сегментов человеческого гена Vλ, четыре пары функциональных сегментов человеческих генов Jλ-Cλ и человеческий генный сегмент Jλ7, функционально связанные с Cλ-областью грызуна (например, Cλ1-область мыши), при этом локус легкой λ-цепи Ig дополнительно содержит две эндогенные энхансерные области (или последовательности) λ-цепи Ig и модульную человеческую энхансерную область (или последовательность, см. выше) λ-цепи Ig; и гомозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный гуманизированный локус H-цепи Ig содержит вставленную кодирующую Adam6 последовательность грызуна (см., например, патенты США №№8642835 и 8697940; включенные таким образом в данный документ посредством ссылки в полном объеме); 6889HO/VI HO/Adam6 HO: сконструированная линия мышей, содержащих гомозиготный сконструированный локус легкой λ-цепи Ig, содержащий 25 функциональных сегментов человеческого гена Vλ, четыре пары функциональных сегментов человеческих генов Jλ-Cλ и человеческий генный сегмент Jλ7, функционально связанные с Cλ-областью грызуна (например, Cλ1-область мыши), при этом локус легкой λ-цепи Ig дополнительно содержит две эндогенные энхансерные области (или последовательности) λ-цепи Ig и модульную человеческую энхансерную область (или последовательность, см. выше) λ-цепи Ig; и гомозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный гуманизированный локус H-цепи Ig содержит вставленную кодирующую Adam6 последовательность грызуна (см., например, патенты США №№8642835 и 8697940; включенные таким образом в данный документ посредством ссылки в полном объеме). Число мышей для каждой показанной когорты по генотипу составляло по меньшей мере три и не более восьми животных на группу.
[0089] На фигурах 14A и 14B показаны репрезентативные иммуноблоты (вестерн-блоты) SDS-PAGE в невосстанавливающих условиях с применением сыворотки крови, выделенной из сконструированных мышей, гомозиготных в отношении вставки нацеливающего вектора 6680 (6680HO), и мышей дикого типа (WT) из одного помета, на которых обозначена экспрессия легких λ-цепей мыши (B, правое изображение) или человека (A, левое изображение); каждый образец загружали на дорожки при объеме, составляющем 1,5 мкл сыворотки крови. PHS: объединенная сыворотка крови человека при объеме, составляющем 0,25 мкл (Labquip Ltd, номер по кат.9101A). Значения молекулярной массы в кДа указаны в правой части каждого изображения геля.
[0090] На фигуре 15A показано репрезентативное использование сегментов человеческого гена Vλ (вверху) и человеческого гена Jλ (внизу) в примированных человеческим Cλ последовательностях, амплифицированных из РНК, выделенной из спленоцитов, собранных у мышей 6889HET (n=5).
[0091] На фигуре 15B показано репрезентативное использование сегментов человеческого гена Vλ в примированных Cλ мыши последовательностях, амплифицированных из РНК, выделенной из спленоцитов, собранных у мышей 6889HET (n=5).
[0092] На фигуре 15C показано репрезентативное использование сегментов человеческого гена Vλ (вверху) и человеческого гена Jλ (внизу) в примированных человеческим Cλ последовательностях, амплифицированных из РНК, выделенной из спленоцитов, собранных у мышей 6889HO/VI HO/Adam6 HO (n=6).
[0093] На фигуре 15D показано репрезентативное использование сегментов человеческого гена Vλ в примированных Cλ мыши последовательностях, амплифицированных из РНК, выделенной из спленоцитов, собранных у мышей 6889HO/VI HO/Adam6 HO (n=6).
[0094] На фигурах 16A и 16B показаны репрезентативные титры общего IgG (A) и антигенспецифического IgG (B) в сыворотке крови на 0 и 22 сутки, собранной у иммунизированных мышей, гетерозиготных в отношении вставки нацеливающего вектора 6597 (6597HET, n=6) или 6680 (6680HET, n=6), и иммунизированных контролей дикого типа (WT, n=6).
[0095] На фигурах 17A-C показаны репрезентативные титры человеческой легкой λ-цепи (hIgλ, слева), легкой λ-цепи мыши (mIgλ, посередине) и легкой κ-цепи мыши (mIgκ, справа) в антигенспецифическом IgG в сыворотке крови на 0 и 22 сутки, собранной у иммунизированных мышей, гетерозиготных в отношении вставки нацеливающего вектора 6597 (6597HET, n=6) или 6680 (6680HET, n=6), и иммунизированных контролей дикого типа (WT, n=6).
[0096] На фигурах 18A и 18B показаны репрезентативные контурные графики, на которых обозначены гейтированные по одиночным клеткам спленоциты (слева), демонстрирующие экспрессию CD19 (ось y) и CD3 (ось x), и общее число B-клеток на селезенку (справа), собранных у мышей, гомозиготных в отношении вставки нацеливающего вектора 6889 (6889HO VI HO Adam6 HO), и эталонных сконструированных мышей (VI). 6889HO/VI HO/Adam6 HO: см. выше; VI: сконструированная линия мышей, содержащих гомозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный гуманизированный локус H-цепи Ig содержит вставленную кодирующую Adam6 последовательность грызуна (см., например, патенты США №№8642835 и 8697940; включенные таким образом в данный документ посредством ссылки в полном объеме). Живые спленоциты по одной клетке определяли с помощью окрашивания на жизнеспособность (Thermo Fisher).
[0097] На фигуре 19 показаны репрезентативные контурные графики, на которых обозначена экспрессия человеческой λ-цепи Ig (hIgλ, ось y) и κ-цепи Ig мыши (mIgκ, ось x) в гейтированных по CD19+ спленоцитах, собранных у мышей, гомозиготных в отношении вставки нацеливающего вектора 6889 (6889HO VI HO Adam6 HO), и у эталонных сконструированных мышей (VI). 6889HO/VI HO/Adam6 HO: см. выше; VI: см. выше.
[0098] На фигуре 20 показаны репрезентативные контурные графики, на которых обозначены гейтированные по одиночным клеткам лимфоциты из костного мозга, демонстрирующие экспрессию IgM (ось y) и B220 (ось x), собранные из бедренных костей мышей, гомозиготных в отношении вставки нацеливающего вектора 6889 (6889HO VI HO Adam6 HO), и эталонных сконструированных мышей (VI). 6889HO/VI HO/Adam6 HO: см. выше; VI: см. выше. Субпопуляции незрелых и зрелых B-клеток указаны на каждом контурном графике.
[0099] На фигуре 21 показаны репрезентативные контурные графики, на которых обозначен незрелый (гейтированный по CD19+IgM+B220int, левая колонка) и зрелый (гейтированный по CD19+IgM+B220+, правая колонка) костный мозг, демонстрирующий экспрессию человеческой λ-цепи Ig (hIgλ, ось y) и κ-цепи Ig мыши (mIgκ, ось x) от мышей, гомозиготных в отношении вставки нацеливающего вектора 6889 (6889HO VI HO Adam6 HO), и эталонных сконструированных мышей (VI). 6889HO/VI HO/Adam6 HO: см. выше; VI: см. выше.
КРАТКОЕ ОПИСАНИЕ ВЫБРАННЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ПЕРЕЧНЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[00100] ДНК Cλ1 мыши (SEQ ID NO: 1):
GCCAGCCCAAGTCTTCGCCATCAGTCACCCTGTTTCCACCTTCCTCTGAAG AGCTCGAGACTAACAAGGCCACACTGGTGTGTACGATCACTGATTTCTACCCAG GTGTGGTGACAGTGGACTGGAAGGTAGATGGTACCCCTGTCACTCAGGGTATGG AGACAACCCAGCCTTCCAAACAGAGCAACAACAAGTACATGGCTAGCAGCTAC CTGACCCTGACAGCAAGAGCATGGGAAAGGCATAGCAGTTACAGCTGCCAGGT CACTCATGAAGGTCACACTGTGGAGAAGAGTTTGTCCCGTGCTGACTGTTCC
[00101] Аминокислотная последовательность Cλ1 мыши (SEQ ID NO: 2):
GQPKSSPSVTLFPPSSEELETNKATLVCTITDFYPGVVTVDWKVDGTPVTQG METTQPSKQSNNKYMASSYLTLTARAWERHSSYSCQVTHEGHTVEKSLSRADCS
[00102] ДНК Cλ2 мыши (SEQ ID NO: 3):
GTCAGCCCAAGTCCACTCCCACTCTCACCGTGTTTCCACCTTCCTCTGAGG AGCTCAAGGAAAACAAAGCCACACTGGTGTGTCTGATTTCCAACTTTTCCCCGA GTGGTGTGACAGTGGCCTGGAAGGCAAATGGTACACCTATCACCCAGGGTGTG GACACTTCAAATCCCACCAAAGAGGGCAACAAGTTCATGGCCAGCAGCTTCCTA CATTTGACATCGGACCAGTGGAGATCTCACAACAGTTTTACCTGTCAAGTTACA CATGAAGGGGACACTGTGGAGAAGAGTCTGTCTCCTGCAGAATGTCTC
[00103] Аминокислотная последовательность Cλ2 мыши (SEQ ID NO: 4):
GQPKSTPTLTVFPPSSEELKENKATLVCLISNFSPSGVTVAWKANGTPITQGV DTSNPTKEGNKFMASSFLHLTSDQWRSHNSFTCQVTHEGDTVEKSLSPAECL
[00104] ДНК Cλ3 мыши (SEQ ID NO: 5):
GTCAGCCCAAGTCCACTCCCACACTCACCATGTTTCCACCTTCCCCTGAGG AGCTCCAGGAAAACAAAGCCACACTCGTGTGTCTGATTTCCAATTTTTCCCCAA GTGGTGTGACAGTGGCCTGGAAGGCAAATGGTACACCTATCACCCAGGGTGTG GACACTTCAAATCCCACCAAAGAGGACAACAAGTACATGGCCAGCAGCTTCTT ACATTTGACATCGGACCAGTGGAGATCTCACAACAGTTTTACCTGCCAAGTTAC ACATGAAGGGGACACTGTGGAGAAGAGTCTGTCTCCTGCAGAATGTCTC
[00105] Аминокислотная последовательность Cλ3 мыши (SEQ ID NO: 6):
GQPKSTPTLTMFPPSPEELQENKATLVCLISNFSPSGVTVAWKANGTPITQGV DTSNPTKEDNKYMASSFLHLTSDQWRSHNSFTCQVTHEGDTVEKSLSPAECL
[00106] ДНК Cλ1 крысы (SEQ ID NO: 7):
GTCAGCCCAAGTCCACTCCCACACTCACAGTATTTCCACCTTCAACTGAG GAGCTCCAGGGAAACAAAGCCACACTGGTGTGTCTGATTTCTGATTTCTACCCG AGTGATGTGGAAGTGGCCTGGAAGGCAAATGGTGCACCTATCTCCCAGGGTGT GGACACTGCAAATCCCACCAAACAGGGCAACAAATACATCGCCAGCAGCTTCT TACGTTTGACAGCAGAACAGTGGAGATCTCGCAACAGTTTTACCTGCCAAGTTA CACATGAAGGGAACACTGTGGAGAAGAGTCTGTCTCCTGCAGAATGTGTC
[00107] Аминокислотная последовательность Cλ1 крысы (SEQ ID NO: 8):
GQPKSTPTLTVFPPSTEELQGNKATLVCLISDFYPSDVEVAWKANGAPISQGV DTANPTKQGNKYIASSFLRLTAEQWRSRNSFTCQVTHEGNTVEKSLSPAECV
[00108] ДНК Cλ2 крысы (SEQ ID NO: 9):
ACCAACCCAAGGCTACGCCCTCAGTCACCCTGTTCCCACCTTCCTCTGAAG AGCTCAAGACTGACAAGGCTACACTGGTGTGTATGGTGACAGATTTCTACCCTG GTGTTATGACAGTGGTCTGGAAGGCAGATGGTACCCCTATCACTCAGGGTGTGG AGACTACCCAGCCTTTCAAACAGAACAACAAGTACATGGCTACCAGCTACCTGCTTTTGACAGCAAAAGCATGGGAGACTCATAGCAATTACAGCTGCCAGGTCACTC ACGAAGAGAACACTGTGGAGAAGAGTTTGTCCCGTGCTGAGTGTTCC
[00109] Аминокислотная последовательность Cλ2 крысы (SEQ ID NO: 10):
DQPKATPSVTLFPPSSEELKTDKATLVCMVTDFYPGVMTVVWKADGTPITQG VETTQPFKQNNKYMATSYLLLTAKAWETHSNYSCQVTHEENTVEKSLSRAECS
[00110] ДНК Cλ3 крысы (SEQ ID NO: 11):
GTCAGCCCAAGTCCACTCCCACACTCACAGTATTTCCACCTTCAACTGAG GAGCTCCAGGGAAACAAAGCCACACTGGTGTGTCTGATTTCTGATTTCTACCCG AGTGATGTGGAAGTGGCCTGGAAGGCAAATGGTGCACCTATCTCCCAGGGTGT GGACACTGCAAATCCCACCAAACAGGGCAACAAATACATCGCCAGCAGCTTCT TACGTTTGACAGCAGAACAGTGGAGATCTCGCAACAGTTTTACCTGCCAAGTTA CACATGAAGGGAACACTGTGGAAAAGAGTCTGTCTCCTGCAGAGTGTGTC
[00111] Аминокислотная последовательность Cλ3 крысы (SEQ ID NO: 12):
GQPKSTPTLTVFPPSTEELQGNKATLVCLISDFYPSDVEVAWKANGAPISQGV DTANPTKQGNKYIASSFLRLTAEQWRSRNSFTCQVTHEGNTVEKSLSPAECV
[00112] ДНК Cλ4 крысы (SEQ ID NO: 13):
ACCAACCCAAGGCTACGCCCTCAGTCACCCTGTTCCCACCTTCCTCTGAAG AGCTCAAGACTGACAAGGCTACACTGGTGTGTATGGTGACAGATTTCTACCCTG GTGTTATGACAGTGGTCTGGAAGGCAGATGGTACCCCTATCACTCAGGGTGTGG AGACTACCCAGCCTTTCAAACAGAACAACAAGTACATGGCTACCAGCTACCTGC TTTTGACAGCAAAAGCATGGGAGACTCATAGCAATTACAGCTGCCAGGTCACTC ACGAAGAGAACACTGTGGAGAAGAGTTTGTCCCGTGCTGAGTGTTCC
[00113] Аминокислотная последовательность Cλ4 крысы (SEQ ID NO: 14):
DQPKATPSVTLFPPSSEELKTDKATLVCMVTDFYPGVMTVVWKADGTPITQG VETTQPFKQNNKYMATSYLLLTAKAWETHSNYSCQVTHEENTVEKSLSRAECS
ОПРЕДЕЛЕНИЯ
[00114] Объем настоящего изобретения определяется формулой изобретения, прилагаемой к настоящему документу, и не ограничивается определенными вариантами осуществления, описанными в данном документе; специалистам в данной области в ходе ознакомления с настоящим описанием будет известно о различных модификациях, которые могут быть эквивалентны таким описанным вариантам осуществления или иным образом находящимся в пределах объема формулы изобретения.
[00115] В целом, используемые в данном документе термины соответствуют их значениям, понятным в данной области, если явно не указано иное. Точные определения некоторых терминов приведены ниже; значения этих и других терминов в конкретных случаях в рамках этого описания будут понятны специалистам в данной области из контекста. Дополнительные определения для следующих и других терминов изложены по всему описанию. Патенты и непатентная литература, указанная в ссылках, приведенных в данном описании или его соответствующих частях, включены в данный документ с помощью ссылки в полном объеме.
[00116] Введение, используемое в данном документе, предусматривает введение композиции субъекту или в систему (например, в клетку, орган, ткань, организм или их соответствующий компонент или набор компонентов). Специалист в данной области поймет, что путь введения может варьироваться, например, в зависимости от субъекта или системы, которым будет вводиться композиция, природы композиции, цели введения и т.д. Например, в определенных вариантах осуществления введение субъекту-животному (например, человек или грызун) может быть бронхиальным (в том числе посредством бронхиальной инстилляции), трансбуккальным, энтеральным, внутрикожным, внутриартериальным, интрадермальным, внутрижелудочным, интрамедуллярным, внутримышечным, интраназальным, внутрибрюшинным, интратекальным, внутривенным, интравентрикулярным, чресслизистым, назальным, пероральным, ректальным, подкожным, подъязычным, местным, трахеальным (в том числе посредством интратрахеальной инстилляции), трансдермальным, вагинальным и/или витреальным. В некоторых вариантах осуществления введение может подразумевать прерывистое введение доз. В некоторых вариантах осуществления введение может предусматривать непрерывное введение доз (например, перфузия) в течение по меньшей мере выбранного периода времени.
[00117] Уменьшение интенсивности проявления, используемое в данном документе, предусматривает предупреждение, ослабление или временное облегчение состояния или улучшение состояния субъекта. Уменьшение интенсивности проявления включает полное излечение или полное предупреждение заболевания, нарушения или состояния, но это не являются необходимым.
[00118] Примерно, применяемое в отношении одного или более представляющих интерес значений, предусматривает значение, сходное с указанным эталонным значением. В некоторых вариантах осуществления термин «примерно» или «приблизительно» относится к диапазону значений, находящихся в пределах 25%, 20%, 19%, 18%, 17%, 16%, 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1% или меньше в любую сторону (большую или меньшую) от указанного эталонного значения, если иное не указано или иное не очевидно из контекста (за исключением случаев, в которых такое число будет превышать 100% относительно возможного значения).
[00119] Биологически активный, используемый в данном документе, относится к характеристике любого средства, обладающего активностью в биологической системе in vitro или in vivo (например, в организме). Например, средство, которое в случае присутствия в организме оказывает биологическое воздействие в пределах этого организма, считается биологически активным. В конкретных вариантах осуществления, в которых белок или полипептид являются биологически активными, часть этого белка или полипептида, обладающая по меньшей мере одним общим с данным белком или полипептидом видом биологической активности, обычно называется «биологически активной» частью.
[00120] Сравнимый, используемый в данном документе, относится к двум или более средствам, объектам, сопутствующим условиям, совокупностям условий и т.д., которые могут не быть идентичными друг другу, но которые являются достаточно сходными для обеспечения возможности сравнения между ними, так что на основании наблюдаемых различий или сходств можно сделать разумные выводы. Специалистам в данной области будет понятно из контекста, какая степень идентичности необходима в любых заданных обстоятельствах для двух или более таких средств, объектов, сопутствующих условий, наборов условий и т.д., чтобы считать их сравнимыми.
[00121] Консервативный, используемый в данном документе, относится к случаям, описывающим консервативную аминокислотную замену, в том числе замену аминокислотного остатка другим аминокислотным остатком, имеющим R-группу боковой цепи со сходными химическими свойствами (например, зарядом или гидрофобностью). Как правило, консервативная аминокислотная замена практически не будет изменять представляющие интерес функциональные свойства белка, например способность рецептора связываться с лигандом. Примеры групп аминокислот, которые имеют боковые цепи со сходными химическими свойствами, включают алифатические боковые цепи, такие как у глицина (Gly, G), аланина (Ala, A), валина (Val, V), лейцина (Leu, L) и изолейцина (Ile, I); алифатические боковые цепи с гидроксильными группами, такие как у серина (Ser, S) и треонина (Thr, T); амидосодержащие боковые цепи, такие как у аспарагина (Asn, N) и глутамина (Gln, Q); ароматические боковые цепи, такие как у фенилаланина (Phe, F), тирозина (Tyr, Y) и триптофана (Trp, W); основные боковые цепи, такие как у лизина (Lys, K), аргинина (Arg, R) и гистидина (His, H); кислые боковые цепи, такие как у аспарагиновой кислоты (Asp, D) и глутаминовой кислоты (Glu, E); и серосодержащие боковые цепи, такие как у цистеина (Cys, C) и метионина (Met, M). Группы консервативных аминокислотных замен включают, например, валин/лейцин/изолейцин (Val/Leu/Ile, V/L/I), фенилаланин/тирозин (Phe/Tyr, F/Y), лизин/аргинин (Lys/Arg, K/R), аланин/валин (Ala/Val, A/V), глутамат/аспартат (Glu/Asp, E/D) и аспарагин/глутамин (Asn/Gln, N/Q). В некоторых вариантах осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке на аланин, используемую, например, в аланин-сканирующем мутагенезе. В некоторых вариантах осуществления проводят консервативную замену, которая имеет положительное значение в матрице логарифмической функции правдоподобия PAM250, раскрытой в публикации Gonnet, G.H. et al., 1992, Science 256:1443-1445. В некоторых вариантах осуществления замена представляет собой умеренно консервативную замену, где замена имеет неотрицательное значение в матрице логарифмической функции правдоподобия PAM250.
[00122] Контроль, используемый в данном документе, относится к понятному в данной области значению «контроля», являющемуся стандартом, с которым сравнивают результаты. Как правило, контроли применяют для расширения полноты в экспериментах путем разделения переменных для того, чтобы сделать вывод об этих переменных. В некоторых вариантах осуществления контроль представляет собой реакцию или анализ, проводимые одновременно с тестовыми реакцией или анализом для обеспечения объекта сравнения. «Контроль» также предусматривает «контрольное животное». «Контрольное животное» может иметь модификацию, описанную в данном документе, модификацию, отличную от описанной в данном документе, или не иметь модификации (т.е. быть животным дикого типа). В одном эксперименте используют «тест» (т.е. тестируемую переменную). Во втором эксперименте, «контроле», тестируемую переменную не используют. В некоторых вариантах осуществления контроль представляет собой исторический контроль (т.е. тест или анализ, проведенный ранее, или количество или результат, известные заранее). В некоторых вариантах осуществления контроль представляет собой или включает напечатанную или иным образом сохраненную запись. Контроль может представлять собой положительный контроль или отрицательный контроль.
[00123]
[00124] Полученный из, при использовании в отношении перегруппированного гена вариабельной области или вариабельного домена, «полученного из» неперегруппированной вариабельной области и/или сегментов неперегрупированного гена вариабельной области, относится к способности восходить последовательностью перегрупированного гена вариабельной области или вариабельного домена к группе сегментов неперегрупированного гена вариабельной области, которые были перегрупированы с образованием перегрупированного гена вариабельной области, который экспрессирует вариабельный домен (учитывая, когда это применимо, отличия при сплайсинге и соматические мутации). Например, то, что перегрупированный ген вариабельной области претерпел соматическую мутацию, не изменяет тот факт, что он получен из сегментов неперегрупированного гена вариабельной области.
[00125] Разрушение, используемое в данном документе, относится к результату события гомологичной рекомбинации с участием молекулы ДНК (например, с участием эндогенной гомологичной последовательности, такой как ген или локус гена). В некоторых вариантах осуществления разрушение может приводить к вставке, делеции, замене, замещению, миссенс-мутации или сдвигу рамки считывания в последовательности(-ях) ДНК или представлять собой таковые или быть любой их комбинацией. Вставки могут предусматривать вставку полных генов или фрагментов генов, например экзонов, которые могут иметь происхождение, отличное от такового для эндогенной последовательности (например, гетерологичная последовательность). В некоторых вариантах осуществления разрушение может приводить к повышению уровня экспрессии и/или активности гена или продукта гена (например, полипептида, кодируемого геном). В некоторых вариантах осуществления разрушение может приводить к снижению уровня экспрессии и/или активности гена или продукта гена. В некоторых вариантах осуществления разрушение может приводить к изменению последовательности гена или кодируемого продукта гена (например, кодируемого полипептида). В некоторых вариантах осуществления разрушение может приводить к усечению или фрагментации гена или кодируемого продукта гена (например, кодируемого полипептида). В некоторых вариантах осуществления разрушение может приводить к удлинению гена или кодируемого продукта гена. В некоторых таких вариантах осуществления разрушение может приводить к сборке слитого полипептида. В некоторых вариантах осуществления разрушение может влиять на уровень экспрессии, но не на активность гена или продукта гена. В некоторых вариантах осуществления разрушение может влиять на активность, но не на уровень экспрессии гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного воздействия на уровень экспрессии гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного воздействия на активность гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного воздействия ни на уровень экспрессии, ни на активность гена или продукта гена.
[00126] Определение, измерение, установление, оценивание, анализ и анализирование используются в данном документе взаимозаменяемо для обозначения любой формы измерения и предусматривают определение присутствия или отсутствия элемента. Эти термины предусматривают количественные и/или также качественные определения. Анализ может быть относительным или абсолютным. «Анализ на наличие» может представлять собой определение количества чего-либо присутствующего и/или определение того, присутствует ли или отсутствует это.
[00127] Эндогенный локус или эндогенный ген, используемый в данном документе, относится к генетическому локусу, находящемуся в исходном или эталонном организме до внесения разрушения, делеции, замещения, изменения или модификации, описанных в данном документе. В некоторых вариантах осуществления эндогенный локус имеет последовательность, обнаруживаемую в природе. В некоторых вариантах осуществления эндогенный локус представляет собой локус дикого типа. В некоторых вариантах осуществления эндогенный локус представляет собой сконструированный локус. В некоторых вариантах осуществления эталонный организм представляет собой организм дикого типа. В некоторых вариантах осуществления эталонный организм представляет собой сконструированный организм. В некоторых вариантах осуществления эталонный организм представляет собой организм, разводимый в лаборатории (или дикого типа, или сконструированный).
[00128] Эндогенный промотор, используемый в данном документе, относится к промотору, связанному в естественных условиях, например в организме дикого типа, с эндогенным геном.
[00129] Сконструированный, используемый в данном документе, относится в целом к аспекту проведения манипуляций человеком. Например, в некоторых вариантах осуществления полинуклеотид может считаться «сконструированным» в том случае, если две или более последовательности, которые не соединены друг с другом в этом порядке в природе, подвергнуты манипуляции человеком для непосредственного соединения друг с другом в сконструированном полинуклеотиде. В некоторых таких конкретных вариантах осуществления сконструированный полинуклеотид может содержать регуляторную последовательность, которая обнаруживается в природе во функциональной связи с первой кодирующей последовательностью, но не во функциональной связи со второй кодирующей последовательностью, соединенную человеком таким образом, чтобы она была функционально связанной со второй кодирующей последовательностью. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления первая и вторая последовательности нуклеиновых кислот, каждая из которых кодирует полипептидные элементы или домены, которые в природе не связаны друг с другом, могут быть связаны друг с другом в единый сконструированный полинуклеотид. Аналогично, в некоторых вариантах осуществления клетка или организм могут считаться «сконструированными» в том случае, если они были подвергнуты манипуляции таким образом, чтобы их генетическая информация была изменена (например, был введен новый генетический материал, который ранее отсутствовал, или ранее присутствовавший генетический материал был изменен или удален). Что является общепринятой практикой и как понятно специалистам в данной области, потомство сконструированных полинуклеотида или клетки, как правило, по-прежнему называют «сконструированным», даже если фактическую манипуляцию проводили на предшествующем объекте. Кроме того, как будет понятно специалистам в данной области, доступно множество методик, посредством которых можно осуществлять «конструирование», описанное в данном документе. Например, в некоторых вариантах осуществления «конструирование» может предусматривать отбор или разработку (например, последовательностей нуклеиновых кислот, полипептидных последовательностей, клеток, тканей и/или организмов) путем использования компьютерных систем, запрограммированных для проведения анализа или сравнения, или в иных случаях для обеспечения анализа, рекомендации в отношении и/или отбора, последовательностей, изменений и т.д.). В качестве альтернативы или дополнительно, в некоторых вариантах осуществления «конструирование» может предусматривать применение методик химического синтеза in vitro, и/или технологий с применением рекомбинантных нуклеиновых кислот, таких как, например, амплификация (например, посредством полимеразной цепной реакции), гибридизация, мутация, трансформация, трансфекция нуклеиновых кислот и т.д., и/или любой из множества методик контролируемого спаривания. Как будет понятно специалистам в данной области, множество таких общепринятых методик (например, для рекомбинантной ДНК, синтеза олигонуклеотидов и культивирования и трансформации тканей (например, электропорация, липофекция и т.д.)) хорошо известно из уровня техники и описано в различных общих и более специфических литературных источниках, которые цитируются и/или рассматриваются по всему настоящему описанию. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989 и Principles of Gene Manipulation: An Introduction to Genetic Manipulation, 5th Ed., ed. By Old, R.W. and S.B. Primrose, Blackwell Science, Inc., 1994.
[00130] Функциональный, используемый в данном документе, относится к форме или фрагменту объекта (например, ген или сегмент гена), которые проявляют конкретные свойство (например, образуют часть кодирующей последовательности) и/или активность. Например, в случае иммуноглобулинов вариабельные домены кодируются сегментами уникальных генов (т.е. V, D и/или J), которые организованы (или рекомбинированы) с образованием функциональных кодирующих последовательностей. При наличии в геноме сегменты генов организованы в кластеры, хотя в действительности имеют место вариации. «Функциональный» сегмент гена представляет собой сегмент гена, представленный в экспрессируемой последовательности (т.е. вариабельном домене), в отношении которого была выделена соответствующая геномная ДНК (т.е. клонирована) и идентифицирована по последовательности. Некоторые последовательности сегментов генов иммуноглобулинов содержат открытые рамки считывания и считаются функциональными, хотя не представлены в экспрессируемом репертуаре, в то время как другие последовательности сегментов генов иммуноглобулинов содержат мутации (например, точечные мутации, вставки, делеции и т.д.), приводящие к образованию стоп-кодона и/или усеченной последовательности, что впоследствии делает такие последовательности сегментов генов неспособными обеспечивать свойство/свойства и/или активность/виды активности, связанные с немутированной(-ыми) последовательностью(-ями). Такие последовательности не представлены среди экспрессируемых последовательностей и, следовательно, классифицируются как псевдогены.
[00131] Ген, используемый в данном документе, относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, продукт, представляющий собой РНК, и/или продукт, представляющий собой полипептид). В некоторых вариантах осуществления ген содержит кодирующую последовательность (т.е. последовательность, которая кодирует конкретный продукт). В некоторых вариантах осуществления ген содержит некодирующую последовательность. В некоторых конкретных вариантах осуществления ген может содержать как кодирующую (например, экзонная), так и некодирующую (например, интронная) последовательность. В некоторых вариантах осуществления ген может содержать одну или более регуляторных последовательностей (например, промоторы, энхансеры и т.д.) и/или интронных последовательностей, которые, например, могут контролировать или влиять на один или более аспектов экспрессии гена (например, специфическая для типа клеток экспрессия, индуцируемая экспрессия и т.д.). С целью внесения ясности отметим, что используемый в настоящем изобретении термин «ген» обычно обозначает часть нуклеиновой кислоты, которая кодирует полипептид или его фрагмент; при этом термин может необязательно охватывать регуляторные последовательности, что будет понятно из контекста специалистам в данной области. Это определение не подразумевает исключение применения термина «ген» к не кодирующим белок единицам экспрессии, а скорее поясняет то, что в большинстве случаев термин, используемый в данном документе, относится к нуклеиновой кислоте, кодирующей полипептид.
[00132] Гетерологичный, используемый в данном документе, относится к средству или объекту из другого источника. Например, при использовании по отношению к полипептиду, гену или продукту гена, присутствующему в конкретных клетке или организме, термин поясняет, что соответствующий полипептид, ген или продукт гена 1) был сконструирован человеком; 2) был введен в клетку или организм (или их предшественника) человеком (например, посредством генной инженерии) и/или 3) в естественных условиях не вырабатывается соответствующими клеткой или организмом (например, соответствующий тип клетки или тип организма) или отсутствует в них. «Гетерологичный» также предусматривает полипептид, ген или продукт гена, которые обычно присутствуют в конкретных нативной клетке или организме, но были изменены или модифицированы, например, за счет мутации или помещения под контроль не встречающихся в природе ассоциированных и в некоторых вариантах осуществления не являющихся эндогенными регуляторных элементов (например, промотор).
[00133] Клетка-хозяин, используемая в данном документе, относится к клетке, в которую были введены нуклеиновая кислота или белок. Специалистам в данной области при прочтении данного раскрытия будет понятно, что такие термины относятся не только к конкретной рассматриваемой клетке, но также используются для обозначения потомства такой клетки. Поскольку в последующих поколениях вследствие влияния либо мутаций, либо окружающей среды могут происходить определенные модификации, такое потомство в действительности может не быть идентичным исходной клетке, но оно по-прежнему включено в объем фразы «клетка-хозяин». В некоторых вариантах осуществления клетка-хозяин представляет собой или предусматривает прокариотическую или эукариотическую клетку. Как правило, клетка-хозяин представляет собой любую клетку, подходящую для приема и/или продуцирования гетерологичных нуклеиновой кислоты или белка, независимо от того, к какому царству живой природы относится клетка. Иллюстративные клетки предусматривают клетки прокариот и эукариот (одноклеточных или многоклеточных), бактериальные клетки (например, штаммы Escherichia coli, Bacillus spp., Streptomyces spp.и т.д.), клетки микобактерий, клетки грибов, клетки дрожжей (например, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris, Pichia methanolica и т.д.), клетки растений, клетки насекомых (например, SF-9, SF-21, клетки насекомых, зараженные бакуловирусами, Trichoplusia ni и т.д.), клетки отличных от человека животных, клетки человека или продукты слияния клеток, такие как, например, гибридомы или квадромы. В некоторых вариантах осуществления клетка представляет собой клетку человека, обезьяны, человекообразной обезьяны, хомячка, крысы или мыши. В некоторых вариантах осуществления клетка является эукариотической и выбрана из следующих клеток: CHO (например, CHO K1, DXB-11 CHO, Veggie-CHO), COS (например, COS-7), клетки сетчатки, Vero, CV1, клетки почки (например, HEK293, 293 EBNA, MSR 293, MDCK, HaK, BHK), HeLa, HepG2, WI38, MRC 5, Colo205, HB 8065, HL-60 (например, BHK21), Jurkat, Daudi, A431 (эпидермальной), CV-1, U937, 3T3, L-клетки, клетки C127, SP2/0, NS-0, MMT 060562, клетки Сертоли, клетки BRL 3A, клетки HT1080, клетки миеломы, опухолевой клетки и линии клеток, полученной из вышеупомянутой клетки. В некоторых вариантах осуществления клетка содержит один или более вирусных генов, например клетка сетчатки, которая экспрессирует вирусный ген (например, клетка PER.C6®). В некоторых вариантах осуществления клетка-хозяин представляет собой или предусматривает выделенную клетку. В некоторых вариантах осуществления клетка-хозяин является частью ткани. В некоторых вариантах осуществления клетка-хозяин является частью организма.
[00134] Идентичность, используемая в данном документе применительно к сравнению последовательностей, относится к идентичности, определяемой с помощью ряда различных алгоритмов, известных из уровня техники, которые можно применять для измерения идентичности нуклеотидных и/или аминокислотных последовательностей. В некоторых вариантах осуществления значения идентичности, описываемые в данном документе, определяют с применением выравнивания ClustalW версии 1.83 (медленного), в котором используются штраф за открытие гэпа 10,0, штраф за продление гэпа 0,1, а также с применением матрицы сходства Gonnet (MACVECTOR™10.0.2, MacVector Inc., 2008).
[00135] In vitro, используемый в данном документе, относится к событиям, которые происходят в искусственной окружающей среде, например в тестовой пробирке или реакционном сосуде, в культуре клеток и т.д., а не в многоклеточном организме.
[00136] In vivo, используемый в данном документе, относится к событиям, которые происходят в многоклеточном организме, таком как человек и/или отличное от человека животное. В случае систем на основе клеток термин можно использовать для обозначения событий, которые происходят в пределах живой клетки (в противоположность, например, системам in vitro).
[00137] Выделенный, используемый в данном документе, относится к веществу и/или объекту, которые были (1) отделены от по меньшей мере некоторых компонентов, с которыми они были связаны при первоначальной выработке (будь то в природе и/или в условиях проведения эксперимента), и/или (2) разработаны, выработаны, получены и/или произведены человеком. Выделенные вещества и/или объекты могут быть отделены от приблизительно 10%, приблизительно 20%, приблизительно 30%, приблизительно 40%, приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или больше чем приблизительно 99% других компонентов, с которыми они были первоначально связаны. В некоторых вариантах осуществления выделенные средства отделены от 10%-100%, 15%-100%, 20%-100%, 25%-100%, 30%-100%, 35%-100%, 40%-100%, 45%-100%, 50%-100%, 55%-100%, 60%-100%, 65%-100%, 70%-100%, 75%-100%, 80%-100%, 85%-100%, 90%-100%, 95%-100%, 96%-100%, 97%-100%, 98%-100% или 99%-100% других компонентов, с которыми они были первоначально связаны. В некоторых вариантах осуществления выделенные средства отделены от 10%-100%, 10%-99%, 10%-98%, 10%-97%, 10%-96%, 10%-95%, 10%-90%, 10%-85%, 10%-80%, 10%-75%, 10%-70%, 10%-65%, 10%-60%, 10%-55%, 10%-50%, 10%-45%, 10%-40%, 10%-35%, 10%-30%, 10%-25%, 10%-20% или 10%-15% других компонентов, с которыми они были первоначально связаны. В некоторых вариантах осуществления выделенные средства отделены от 11%-99%, 12%-98%, 13%-97%, 14%-96%, 15%-95%, 20%-90%, 25%-85%, 30%-80%, 35%-75%, 40%-70%, 45%-65%, 50%-60% или 55%-60% других компонентов, с которыми они были первоначально связаны. В некоторых вариантах осуществления выделенные средства являются чистыми на приблизительно 80%, на приблизительно 85%, на приблизительно 90%, на приблизительно 91%, на приблизительно 92%, на приблизительно 93%, на приблизительно 94%, на приблизительно 95%, на приблизительно 96%, на приблизительно 97%, на приблизительно 98%, на приблизительно 99% или больше чем на приблизительно 99%. В некоторых вариантах осуществления выделенные средства характеризуются чистотой 80%-99%, 85%-99%, 90%-99%, 95%-99%, 96%-99%, 97%-99% или 98%-99%. В некоторых вариантах осуществления выделенные средства характеризуются чистотой 80%-99%, 80%-98%, 80%-97%, 80%-96%, 80%-95%, 80%-90% или 80%-85%. В некоторых вариантах осуществления выделенные средства характеризуются чистотой 85%-98%, 90%-97% или 95%-96%. В некоторых вариантах осуществления вещество является «чистым», если оно практически не содержит других компонентов. В некоторых вариантах осуществления, что будет понятно специалистам в данной области, вещество может по-прежнему считаться «выделенным» или даже «чистым» после объединения с определенными другими компонентами, такими как, например, один или более носителей или наполнителей (например, буфер, растворитель, вода и т.д.); в таких вариантах осуществления процент выделения или чистоты вещества рассчитывают без учета таких носителей или наполнителей. В качестве лишь одного примера, в некоторых вариантах осуществления биологический полимер, такой как полипептид или полинуклеотид, встречающийся в природе, считается «выделенным», если: a) он ввиду своего происхождения или источника получения не связан с некоторыми или всеми компонентами, сопровождающими его в его нативном состоянии в природе; b) он практически не содержит других полипептидов или нуклеиновых кислот того же вида из вида, который вырабатывает их в природе; или c) он экспрессируется или иным образом связан с компонентами в клетке или другой системе экспрессии, не принадлежащей к виду, который вырабатывает его в природе. Таким образом, например, в некоторых вариантах осуществления полипептид, синтезируемый химическим путем или синтезируемый в клеточной системе, отличной от той, которая продуцирует его в природе, считается «выделенным» полипептидом. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления полипептид, который был подвергнут одной или более методикам очистки, может считаться «выделенным» полипептидом в той степени, в которой он был отделен от других компонентов: a) с которыми он связан в природе и/или b) с которыми он был связан при исходном продуцировании.
[00138] Локус или локусы, используемые в данном документе, относятся к конкретному(-ым) месторасположению(-иям) гена (или значимой последовательности), последовательности ДНК, кодирующей полипептид последовательности или положению в хромосоме генома организма. Например, «локус иммуноглобулина» может относиться к конкретному месторасположению сегмента гена иммуноглобулина (например, V, D, J или C), последовательности ДНК сегмента гена иммуноглобулина, кодирующей сегмент гена иммуноглобулина последовательности или положению сегмента гена иммуноглобулина в хромосоме генома организма, которое было идентифицировано как то, в котором находится такая последовательность. «Локус иммуноглобулина» может содержать регуляторный элемент сегмента гена иммуноглобулина, в том числе без ограничения энхансер, промотор, регуляторную 5'- и/или 3'-последовательность или область или их комбинацию. «Локус иммуноглобулина» может содержать ДНК, которая обычно находится между сегментами генов в локусе дикого типа, но в самой ДНК отсутствует сегмент гена иммуноглобулина (например, последовательность ДНК иммуноглобулина, которая в природе находится между группой сегментов гена V и группой сегментов гена J, последовательность ДНК иммуноглобулина, которая в природе находится между группой сегментов гена J и геном константной области, или последовательность ДНК иммуноглобулина, которая в природе находится на 3'-конце гена константной области). Специалистам в данной области будет понятно, что в некоторых вариантах осуществления хромосомы могут содержать несколько сотен или даже несколько тысяч генов и демонстрировать физическую совместную локализацию сходных генетических локусов при сравнении разных видов. Такие генетические локусы могут быть описаны как имеющие общую синтению.
[00139] Отличное от человека животное, используемое в данном документе, относится к любому позвоночному организму, который не является человеком. В некоторых вариантах осуществления отличное от человека животное, является круглоротым, костной рыбой, хрящевой рыбой (например, акула или скат), земноводным, пресмыкающимся, млекопитающим и птицей. В некоторых вариантах осуществления отличное от человека животное, представляет собой млекопитающее. В некоторых вариантах осуществления отличное от человека млекопитающее является приматом, козой, овцой, свиньей, собакой, коровой или грызуном. В некоторых вариантах осуществления отличное от человека животное, представляет собой грызуна, такого как крыса или мышь.
[00140] Нуклеиновая кислота, используемая в данном документе, относится к любому соединению и/или веществу, которое включено или может быть включено в состав олигонуклеотидной цепи. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой соединение и/или вещество, которое включено или может быть включено в состав олигонуклеотидной цепи посредством фосфодиэфирной связи. Как будет очевидно из контекста, в некоторых вариантах осуществления «нуклеиновая кислота» относится к отдельным остаткам нуклеиновой кислоты (например, нуклеотиды и/или нуклеозиды); в некоторых вариантах осуществления «нуклеиновая кислота» относится к олигонуклеотидной цепи, содержащей отдельные остатки нуклеиновой кислоты. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой РНК или содержит ее; в некоторых вариантах осуществления «нуклеиновая кислота» представляет собой ДНК или содержит ее. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более остатков природной нуклеиновой кислоты, содержит их или состоит из них. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более аналогов нуклеиновой кислоты, содержит их или состоит из них. В некоторых вариантах осуществления аналог нуклеиновой кислоты отличается от «нуклеиновой кислоты» тем, что в нем не используется фосфодиэфирный остов. Например, в некоторых вариантах осуществления «нуклеиновая кислота» представляет собой одну или более «пептидных нуклеиновых кислот», которые известны из уровня техники и имеют в остове пептидные связи вместо фосфодиэфирных связей, содержит их или состоит из них. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления «нуклеиновая кислота» имеет одну или более фосфоротиоатных и/или 5'-N-фосфорамидитных связей вместо фосфодиэфирных связей. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более природных нуклеозидов (например, аденозин, тимидин, гуанозин, цитидин, уридин, дезоксиаденозин, дезокситимидин, дезоксигуанозин и дезоксицитидин), содержит их или состоит из них. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более аналогов нуклеозидов (например, 2- аминоаденозин, 2-тиотимидин, инозин, пирролопиримидин, 3-метиладенозин, 5-метилцитидин, C5-пропинилцитидин, C5-пропинилуридин, 2-аминоаденозин, C5-бромуридин, C5-фторуридин, C5-йодуридин, C5-пропинилуридин, C5-пропинилцитидин, C5-метилцитидин, 2-аминоаденозин, 7-дезазааденозин, 7-дезазагуанозин, 8-оксоаденозин, 8-оксогуанозин, O(6)-метилгуанин, 2-тиоцитидин, метилированные основания, интеркалированные основания и их комбинации), содержит их или состоит из них. В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более модифицированных сахарных остатков (например, 2'-фторрибоза, рибоза, 2'-дезоксирибоза, арабиноза и гексоза) по сравнению с таковыми в природных нуклеиновых кислотах. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, которая кодирует функциональный продукт гена, такой как РНК или полипептид. В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более интронов. В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более экзонов. В некоторых вариантах осуществления «нуклеиновую кислоту» получают посредством одного или более из выделения из природного источника, ферментативного синтеза путем полимеризации на основе комплементарной матрицы (in vivo или in vivo), воспроизведения в рекомбинантной клетке или системе и химического синтеза. В некоторых вариантах осуществления длина «нуклеиновой кислоты» составляет по меньшей мере, например, без ограничения 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 20, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000 или более остатков. В некоторых вариантах осуществления «нуклеиновая кислота» является однонитевой; в некоторых вариантах осуществления «нуклеиновая кислота» является двухнитевой. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, содержащую по меньшей мере один элемент, который кодирует полипептид или является комплементарным по отношению к последовательности, которая кодирует полипептид. В некоторых вариантах осуществления «нуклеиновая кислота» обладает ферментативной активностью.
[00141] Функционально связанный, используемый в данном документе, относится к расположению рядом, при котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать надлежащим образом. Например, сегменты неперегруппированного гена вариабельной области «функционально связаны» со смежным геном константной области, если сегменты неперегруппированного гена вариабельной области способны к перегруппировке с образованием перегрупированного гена вариабельной области, который экспрессируется в сочетании с геном константной области в виде полипептидной цепи антигенсвязывающего белка. Контролирующая последовательность, «функционально связанная» с кодирующей последовательностью, лигирована с ней таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с функционированием контролирующих последовательностей. «Функционально связанные» последовательности предусматривают как последовательности, контролирующие экспрессию, смежные с представляющим интерес геном, так и последовательности, контролирующие экспрессию, действующие в транс-положении или на некотором расстоянии, осуществляя контроль над представляющим интерес геном (или представляющей интерес последовательностью). Термин «последовательность, контролирующая экспрессию» предусматривает полинуклеотидные последовательности, необходимые для осуществления воздействия на экспрессию и процессинг кодирующих последовательностей, с которыми они лигированы. «Последовательности, контролирующие экспрессию» включают соответствующие последовательности для инициации, терминации транскрипции, промоторные и энхансерные последовательности; сигналы для эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую mRNA; последовательности, повышающие эффективность трансляции (т.е. консенсусную последовательность Kozak); последовательности, повышающие стабильность полипептида; и при необходимости последовательности, усиливающие секрецию полипептида. Природа таких контролирующих последовательностей различается в зависимости от организма-хозяина. Например, у прокариот такие контролирующие последовательности обычно предусматривают промотор, участок связывания рибосом и последовательность для терминации транскрипции, тогда как у эукариот обычно такие контролирующие последовательности предусматривают промоторы и последовательность для терминации транскрипции. Подразумевается, что термин «контролирующие последовательности» включает компоненты, присутствие которых является необходимым для экспрессии и процессинга, а также может включать дополнительные компоненты, присутствие которых является благоприятным, например лидерные последовательности и последовательности партнера по слиянию.
[00142] Физиологические условия, используемые в данном документе, относятся к понятным из уровня техники стандартным условиям, при которых клетки или организмы живут и/или репродуцируются. В некоторых вариантах осуществления термин предусматривает условия внешней или внутренней окружающих сред, которые могут существовать в природе относительно организма или системы клеток. В некоторых вариантах осуществления физиологические условия представляют собой условия, присутствующие в пределах организма человека или отличного от человека животного, в частности те условия, присутствующие в месте и/или в пределах области хирургического вмешательства. Физиологические условия, как правило, предусматривают, например, температурный диапазон, составляющий 20-40°C, атмосферное давление, составляющее 1, показатель pH, составляющий 6-8, концентрацию глюкозы, составляющую 1-20 мМ, концентрацию кислорода при уровнях, равным атмосферным, и силу тяжести, которая встречается на планете Земля. В некоторых вариантах осуществления условия в лаборатории регулируют и/или поддерживают при физиологических условиях. В некоторых вариантах осуществления физиологические условия встречаются в организме.
[00143] Полипептид, используемый в данном документе, относится к любой полимерной цепи из аминокислот. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, встречающуюся в природе. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, не встречающуюся в природе. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, которая содержит части, встречающиеся в природе отдельно друг от друга (т.е. из двух или более различных организмов, например части, принадлежащие организму человека и отличному от человека организму). В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, которая является сконструированной в том смысле, что она разработана и/или получена в результате действий человека. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, кодируемую последовательностью, не встречающейся в природе (например, последовательность, которая сконструирована в том смысле, что она разработана и/или получена в результате действий человека для кодирования указанного полипептида).
[00144] Рекомбинантный, используемый в данном документе, подразумевает обозначение полипептидов, разрабатываемых, конструируемых, получаемых, экспрессируемых, создаваемых или выделяемых с помощью рекомбинантных способов, таких как полипептиды, экспрессируемые с помощью рекомбинантного вектора экспрессии, вводимого путем трансфекции в клетку-хозяина, полипептиды, выделяемые из комбинаторной библиотеки рекомбинантных человеческих полипептидов (Hoogenboom, H. R., 1997, TIB Tech. 15:62-70; Azzazy, H. and W.E. Highsmith, 2002, Clin. Biochem. 35:425-45; Gavilondo, J. V. and J.W. Larrick, 2002, BioTechniques 29:128-45; Hoogenboom H., and P. Chames, 2000, Immunol. Today 21:371-8), антитела, выделяемые из животного (например, мышь), трансгенного по человеческим генам иммуноглобулинов (см., например, Taylor, L. D. et al., 1992, Nucl. Acids Res. 20:6287-95; Kellermann, S-A. and L.L. Green, 2002, Curr. Opin. Biotechnol. 13:593-7; Little, M. et al., 2000, Immunol. Today 21:364-70; Osborn, M.J. et al., 2013, J. Immunol. 190:1481-90; Lee, E-C. et al., 2014, Nat. Biotech. 32(4):356-63; Macdonald, L.E. et al., 2014, Proc. Natl. Acad. Sci. U.S.A. 111(14):5147-52; Murphy, A.J. et al., 2014, Proc. Natl. Acad. Sci. U.S.A. 111(14):5153-8), или полипептиды, получаемые, экспрессируемые, создаваемые или выделяемые любыми другими способами, предусматривающими сшивку выбранных элементов последовательностей друг с другом. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательностей обнаруживаются в природе. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательностей разрабатывают in silico. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательностей получают в результате мутагенеза (например, in vivo или in vitro) элемента известной последовательности, например, из природного или синтетического источника. Например, в некоторых вариантах осуществления рекомбинантный полипептид состоит из последовательностей, обнаруживаемых в геноме представляющего интерес организма-источника (например, человек, мышь и т.д.). В некоторых вариантах осуществления рекомбинантный полипептид имеет аминокислотную последовательность, полученную в результате мутагенеза (например, in vitro или in vivo, например в отличном от человека животном), так что аминокислотные последовательности рекомбинантных полипептидов представляют собой последовательности, которые хоть и происходят из полипептидных последовательностей и являются родственными им, но в естественных условиях могут не существовать in vivo в пределах генома отличного от человека животного.
[00145] Эталон, используемый в данном документе, относится к стандартным или контрольным средству, животному, когорте, индивидууму, популяции, образцу, последовательности или значению, с которыми сравнивают представляющие интерес средство, животное, когорту, индивидуума, популяцию, образец, последовательность или значение. В некоторых вариантах осуществления эталонные средство, животное, когорту, индивидуума, популяцию, образец, последовательность или значение тестируют и/или определяют практически одновременно с тестированием или определением представляющих интерес средства, животного, когорты, индивидуума, популяции, образца, последовательности или значения. В некоторых вариантах осуществления эталонные средство, животное, когорта, индивидуум, популяция, образец, последовательность или значение представляют собой исторический эталон, необязательно существующий в материальной форме на материальном носителе. В некоторых вариантах осуществления эталон может относиться к контролю. «Эталонный» также предусматривает «эталонное животное». «Эталонное животное» может иметь модификацию, описанную в данном документе, модификацию, отличную от описанной в данном документе, или не иметь модификации (т.е. быть животным дикого типа). Как будет понятно специалистам в данной области, эталонные средство, животное, когорту, индивидуума, популяцию, образец, последовательность или значение обычно определяют или характеризуют в условиях, сравнимых с условиями, используемыми для определения или получения характеристик представляющих интерес средства, животного (например, млекопитающее), когорты, индивидуума, популяции, образца, последовательности или значения.
[00146] Замещение, используемое в данном документе, относится к процессу, посредством которого «замещаемую» последовательность нуклеиновой кислоты (например, ген), обнаруживаемую в локусе хозяина (например, в геноме), удаляют из этого локуса, а другую, «замещающую» нуклеиновую кислоту, располагают на ее месте. В некоторых вариантах осуществления замещаемая последовательность нуклеиновой кислоты и замещающие последовательности нуклеиновых кислот сравнимы друг с другом в том, что, например, они являются гомологичными друг другу и/или содержат соответствующие элементы (например, элементы, кодирующие белок, регуляторные элементы и т.д.). В некоторых вариантах осуществления замещаемая последовательность нуклеиновой кислоты содержит одно или более из промотора, энхансера, донорного участка сплайсинга, акцепторного участка сплайсинга, интрона, экзона, нетранслируемой области (UTR); в некоторых вариантах осуществления замещающая последовательность нуклеиновой кислоты содержит одну или более кодирующих последовательностей. В некоторых вариантах осуществления замещающая последовательность нуклеиновой кислоты представляет собой гомолог или вариант (например, мутант) замещаемой последовательности нуклеиновой кислоты. В некоторых вариантах осуществления замещающая последовательность нуклеиновой кислоты является ортологом или гомологом замещаемой последовательности. В некоторых вариантах осуществления замещающая последовательность нуклеиновой кислоты представляет собой или содержит человеческую последовательность нуклеиновой кислоты. В некоторых вариантах осуществления, в том числе в тех, в которых замещающая последовательность нуклеиновой кислоты представляет собой или содержит человеческую последовательность нуклеиновой кислоты, замещаемая последовательность нуклеиновой кислоты представляет собой или содержит последовательность грызуна (например, последовательность мыши или крысы). В некоторых вариантах осуществления, в том числе в тех, в которых замещающая последовательность нуклеиновой кислоты представляет собой или содержит человеческую последовательность нуклеиновой кислоты, замещаемая последовательность нуклеиновой кислоты представляет собой или содержит человеческую последовательность. В некоторых вариантах осуществления замещающая последовательность нуклеиновой кислоты представляет собой вариант или мутант (т.е. последовательность, содержащую одно или более различий в последовательности, например замен, по сравнению с замещаемой последовательностью) замещаемой последовательности. Последовательность нуклеиновой кислоты, размещенная таким образом, может содержать одну или более регуляторных последовательностей, которые являются частью первичной последовательности нуклеиновой кислоты, используемой для получения последовательности, размещенной таким образом (например, промоторы, энхансеры, 5'- или 3'-нетранслируемые области и т.д.). Например, в различных вариантах осуществления замещение представляет собой замену эндогенной последовательности гетерологичной последовательностью, что в результате приводит к выработке продукта гена с последовательностью нуклеиновой кислоты, размещенной таким образом (содержащей гетерологичную последовательность), но не к экспрессии эндогенной последовательности; при этом эндогенная геномная последовательность замещается последовательностью нуклеиновой кислоты, которая кодирует полипептид, имеющий функцию, сходную с таковой полипептида, кодируемого эндогенной последовательностью (например, эндогенная геномная последовательность кодирует полный или неполный отличный от человеческого полипептид вариабельного домена, и фрагмент ДНК кодирует один или более человеческих полных или неполных полипептидов вариабельных доменов). В различных вариантах осуществления эндогенный сегмент отличного от человеческого гена иммуноглобулина или его фрагмент заменен сегментом человеческого гена иммуноглобулина или его фрагментом.
[00147] Практически, используемое в данном документе относится к качественному фактору проявления полной или практически полной меры или степени выраженности представляющих интерес характеристики или свойства. Специалисту в области биологии будет понятно, что биологические и химические явления редко, если это вообще происходит, продолжаются до окончания и/или протекают до завершения или достигают или избегают абсолютного результата. Таким образом, термин «практически» используется в данном документе для охвата потенциального отсутствия завершенности, присущего многим биологическим и химическим явлениям.
[00148] Практически гомологичный, используемый в данном документе, относится к сравнению между аминокислотными последовательностями или последовательностями нуклеиновых кислот. Как будет понятно специалистам в данной области, две последовательности обычно считаются «практически гомологичными», если они содержат гомологичные остатки в соответствующих положениях. Гомологичные остатки могут быть идентичными остатками. В качестве альтернативы, гомологичные остатки могут представлять собой неидентичные остатки с соответственно сходными структурными и/или функциональными характеристиками. Например, как хорошо известно специалистам в данной области, определенные аминокислоты, как правило, классифицируют как «гидрофобные» или «гидрофильные» аминокислоты и/или как имеющие «полярные» или «неполярные» боковые цепи. Замена одной аминокислоты другой того же типа часто может считаться «гомологичной» заменой. Типичное распределение аминокислот по категориям обобщено в таблице ниже.

[00149] Как хорошо известно из уровня техники, аминокислотные последовательности или последовательности нуклеиновых кислот можно сравнивать с помощью любого из множества алгоритмов, в том числе таковых, доступных в коммерческих компьютерных программах, таких как BLASTN для нуклеотидных последовательностей и BLASTP, BLAST с гэпами и PSI-BLAST для аминокислотных последовательностей. Такие иллюстративные программы описаны в Altschul, S. F. et al., 1990, J. Mol. Biol., 215(3): 403-10; Altschul, S.F. et al., 1996, Meth. Enzymol. 266:460-80; Altschul, S.F. et al., 1997, Nucleic Acids Res., 25:3389-402; Baxevanis, A.D. and B.F.F. Ouellette (eds.) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins, Wiley, 1998; и Misener et al. (eds.) Bioinformatics Methods and Protocols, Methods in Molecular Biology, Vol.132, Humana Press, 1998. В дополнение к идентификации гомологичных последовательностей, программы, упомянутые выше, как правило, дают представление о степени гомологии. В некоторых вариантах осуществления две последовательности считаются практически гомологичными, если по меньшей мере, например, без ограничения 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше их соответствующих остатков являются гомологичными на протяжении рассматриваемого фрагмента из остатков. В некоторых вариантах осуществления рассматриваемый фрагмент представляет собой полную последовательность. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 9, 10, 11, 12, 13, 14, 15, 16, 17 или больше остатков. В некоторых вариантах осуществления рассматриваемый фрагмент содержит смежные остатки вдоль полной последовательности. В некоторых вариантах осуществления рассматриваемый фрагмент содержит расположенные с промежутками остатки вдоль полной последовательности, например расположенные с промежутками остатки, сводимые вместе за счет складчатой конформации полипептида или его части. В некоторых вариантах осуществления рассматриваемый фрагмент составляет по меньшей мере, например, без ограничения 10, 15, 20, 25, 30, 35, 40, 45, 50 или больше остатков.
[00150] Практически идентичный, используемый в данном документе, относится к сравнению между аминокислотными последовательностями или последовательностями нуклеиновых кислот. Как будет понятно специалистам в данной области, две последовательности обычно считаются «практически идентичными», если они содержат идентичные остатки в соответствующих положениях. Как хорошо известно из уровня техники, аминокислотные последовательности или последовательности нуклеиновых кислот можно сравнивать с помощью любого из множества алгоритмов, в том числе таковых, доступных в коммерческих компьютерных программах, таких как BLASTN для нуклеотидных последовательностей и BLASTP, BLAST с гэпами и PSI-BLAST для аминокислотных последовательностей. Такие иллюстративные программы описаны в Altschul, S. F. et al., 1990, J. Mol. Biol., 215(3): 403-10; Altschul, S.F. et al., 1996, Meth. Enzymol. 266:460-80; Altschul, S.F. et al., 1997, Nucleic Acids Res., 25:3389-402; Baxevanis, A.D. and B.F.F. Ouellette (eds.) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins, Wiley, 1998; и Misener et al. (eds.) Bioinformatics Methods and Protocols, Methods in Molecular Biology, Vol.132, Humana Press, 1998. В дополнение к идентификации идентичных последовательностей, программы, упомянутые выше, как правило, дают представление о степени идентичности. В некоторых вариантах осуществления две последовательности считаются практически идентичными, если по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше их соответствующих остатков являются идентичными на протяжении рассматриваемого фрагмента из остатков. В некоторых вариантах осуществления рассматриваемый фрагмент из остатков представляет собой полную последовательность. В некоторых вариантах осуществления рассматриваемый фрагмент из остатков составляет, например, без ограничения по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50 или больше остатков.
[00151] Нацеливающая конструкция или нацеливающий вектор, используемые в данном документе, относятся к молекуле полинуклеотида, содержащей нацеливающую область. Нацеливающая область содержит последовательность, идентичную или практически идентичную последовательности в целевых клетке, ткани или животном, и обеспечивает интеграцию нацеливающей конструкции в некоторое положение в геноме клетки, ткани или животного посредством гомологичной рекомбинации. Также включены и описаны в данном документе нацеливающие области, которые нацеливаются с помощью участков распознавания для сайт-специфической рекомбиназы (например, участки loxP и/или Frt). В некоторых вариантах осуществления нацеливающая конструкция, описанная в данном документе, дополнительно содержит представляющие особенный интерес последовательность нуклеиновой кислоты или ген, селектируемый маркер, контролирующие и/или регуляторные последовательности, а также другие последовательности нуклеиновой кислоты, обеспечивающие возможность рекомбинации, опосредованной добавлением экзогенных белков, которые способствуют или содействуют рекомбинации с вовлечением таких последовательностей. В некоторых вариантах осуществления нацеливающая конструкция, описанная в данном документе, дополнительно содержит представляющий интерес полный или неполный ген, где представляющий интерес ген является гетерологичным геном, который кодирует полный или неполный полипептид, имеющий функцию, сходную с таковой у белка, кодируемого эндогенной последовательностью. В некоторых вариантах осуществления нацеливающая конструкция, описанная в данном документе, дополнительно содержит представляющий интерес гуманизированный полный или неполный ген, где представляющий интерес гуманизированный ген кодирует полный или неполный полипептид, имеющий функцию, сходную с таковой у полипептида, кодируемого эндогенной последовательностью. В некоторых вариантах осуществления нацеливающая конструкция (или нацеливающий вектор) могут содержать последовательность нуклеиновой кислоты, в отношении которой проведена манипуляция человеком. Например, в некоторых вариантах осуществления нацеливающая конструкция (или нацеливающий вектор) могут быть построены так, чтобы содержать сконструированный или рекомбинантный полинуклеотид, который содержит две или более последовательности, которые не связаны друг с другом в таком порядке в природе, пока не подвергаются манипуляции человеком для связывания напрямую друг с другом в сконструированном или рекомбинантном полинуклеотиде.
[00152] Трансген или трансгенная конструкция, используемые в данном документе обозначают последовательность нуклеиновой кислоты (кодирующую, например, представляющий интерес полный или неполный полипептид), которая была введена в клетку человеком с помощью таких способов, которые описаны в данном документе. Трансген может быть частично или полностью гетерологичным, т.е. чужеродным, для трансгенных животного или клетки, в которые он введен. Трансген может содержать одну или более последовательностей регуляции транскрипции и любую другую нуклеиновую кислоту, такую как интроны или промоторы, которые могут быть необходимы для экспрессии выбранной последовательности нуклеиновой кислоты.
[00153] Трансгенное животное, трансгенное отличное от человека животное или Tg+ используется взаимозаменяемо в данном документе и обозначает любое не встречающееся в природе отличное от человека животное, у которого одна или более клеток отличного от человека животного содержат гетерологичные нуклеиновую кислоту и/или ген, кодирующие представляющий интерес полный или неполный полипептид. Например, в некоторых вариантах осуществления «трансгенное животное» или «трансгенное отличное от человека животное» относится к животному или отличному от человека животному, которое содержит трансген или трансгенную конструкцию, описанные в данном документе. В некоторых вариантах осуществления гетерологичные нуклеиновую кислоту и/или ген вводят в клетку напрямую или опосредованно, путем введения в клетку-предшественника, путем преднамеренной генетической манипуляции, например с помощью микроинъекции или путем инфицирования рекомбинантным вирусом. Термин «генетическая манипуляция» не включает классические методики скрещивания, а скорее относится к введению молекулы (молекул) рекомбинантной ДНК. Данная молекула может быть интегрирована в хромосому, или она может представлять собой внехромосомно реплицирующуюся ДНК. Термин «Tg+» предусматривает животных, которые являются гетерозиготными или гомозиготными по гетерологичным нуклеиновой кислоте и/или гену, и/или животных, которые имеют одну или множество копий гетерологичных нуклеиновой кислоты и/или гена.
[00154] Вариант, используемый в данном документе, относится к объекту, демонстрирующему значительную структурную идентичность с эталонным объектом, но структурно отличающемуся от эталонного объекта по наличию или уровню одного или более химических фрагментов по сравнению с эталонным объектом. Во многих вариантах осуществления «вариант» также отличается функционально от своего эталонного объекта. Как правило, то, уместно ли считать конкретный объект «вариантом» эталонного объекта, зависит от степени его структурной идентичности с эталонным объектом. Как будет понятно специалистам в данной области, любой биологический или химический эталонный объект имеет определенные характерные структурные элементы. «Вариант» по определению представляет собой отличающийся химический объект, имеющий один или более общих таких характерных структурных элементов. В качестве лишь пары примеров полипептид может иметь характерный элемент последовательности, состоящий из множества аминокислот, характеризующихся определенными положениями друг относительно друга в линейном или трехмерном пространстве и/или обеспечивающих конкретную биологическую функцию, или нуклеиновая кислота может иметь характерный элемент последовательности, состоящий из множества нуклеотидных остатков, характеризующихся определенными положениями друг относительно друга в линейном или трехмерном пространстве. В другом примере «вариантный полипептид» может отличаться от эталонного полипептида вследствие одного или более различий в аминокислотной последовательности и/или одного или более различий в химических фрагментах (например, углеводы, липиды и т.д.), присоединенных ковалентной связью к полипептидному остову. В некоторых вариантах осуществления «вариантный полипептид» демонстрирует общую идентичность последовательности с эталонным полипептидом, составляющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97% или 99%. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления «вариантный полипептид» не имеет по меньшей мере одного характерного элемента последовательности, общего с эталонным полипептидом. В некоторых вариантах осуществления эталонный полипептид обладает одним или более видами биологической активности. В некоторых вариантах осуществления «вариантный полипептид» обладает одним или более видами биологической активности, общими с эталонным полипептидом. В некоторых вариантах осуществления у «вариантного полипептида» отсутствует один или более видов биологической активности эталонного полипептида. В некоторых вариантах осуществления «вариантный полипептид» демонстрирует пониженный уровень одного или более видов биологической активности по сравнению с эталонным полипептидом. Во многих вариантах осуществления представляющий интерес полипептид считается «вариантом» исходного или эталонного полипептида, если представляющий интерес полипептид имеет аминокислотную последовательность, идентичную таковой исходной молекулы, за исключением небольшого количества изменений в конкретных положениях в последовательности. Как правило, в варианте заменены менее 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% остатков по сравнению с исходной молекулой. В некоторых вариантах осуществления «вариант» имеет, например, без ограничения 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 замену остатка(-ов) по сравнению с исходной молекулой. Зачастую «вариант» имеет очень небольшое количество (например, менее 5, 4, 3, 2 или 1) количество замененных функциональных остатков (т.е. остатков, принимающих участие в конкретном виде биологической активности). Кроме того, «вариант» обычно имеет не более, например, без ограничения 5, 4, 3, 2 или 1 добавления или делеции, а зачастую не имеет добавлений или делеций по сравнению с исходной молекулой. Более того, любые добавления или делеции обычно охватывают меньше чем приблизительно 25, приблизительно 20, приблизительно 19, приблизительно 18, приблизительно 17, приблизительно 16, приблизительно 15, приблизительно 14, приблизительно 13, приблизительно 10, приблизительно 9, приблизительно 8, приблизительно 7, приблизительно 6 и зачастую охватывают меньше чем приблизительно 5, приблизительно 4, приблизительно 3 или приблизительно 2 остатка. В некоторых вариантах осуществления исходный или эталонный полипептид обнаруживается в природе. Как будет понятно специалистам в данной области, множество вариантов конкретного представляющего интерес полипептида можно зачастую обнаружить в природе, особенно если представляющий интерес полипептид представляет собой полипептид возбудителя инфекции.
[00155] Вектор, используемый в данном документе, относится к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она связана. В некоторых вариантах осуществления векторы способны к внехромосомной репликации и/или экспрессии нуклеиновых кислот, с которыми они соединены, в клетке-хозяине, такой как эукариотическая и/или прокариотическая клетка. Векторы, способные управлять экспрессией функционально связанных генов, называются в данном документе «векторами экспрессии».
[00156] Дикий тип, используемый в данном документе, относится к объекту, обладающему структурой и/или активностью, обнаруживаемой в природе в «нормальном» (в противоположность мутантному, болезненному, измененному и т.д.) состоянии или окружении. Специалистам в данной области будет понятно, что гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллелях).
ПОДРОБНОЕ ОПИСАНИЕ ОПРЕДЕЛЕННЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[00157] В определенных аспектах в данном документе, среди прочего, предусмотрены сконструированные отличные от человека животные, имеющие гетерологичный генетический материал, кодирующий человеческие вариабельные домены и, в некоторых вариантах осуществления, человеческие константные домены, при этом гетерологичный генетический материал содержит последовательности человеческих генов (т.е. сегменты человеческих генов) Vλ, Jλ и Cλ и другие человеческие последовательности, которые обеспечивают подходящую перегруппировку и экспрессию антител, имеющих часть из организма человека и часть из отличного от человеческого организма, или антител, имеющих последовательность, которая является, практически или практически полностью, человеческой. В различных вариантах осуществления предусмотренные сконструированные отличные от человека животные содержат гетерологичный генетический материал, который вставлен таким образом, что антитела, содержащие легкие цепи, которые имеют человеческий Vλ-домен и человеческий или отличный от человеческого Cλ-домен, экспрессируются в репертуаре антител отличного от человека животного. Кроме того, предусмотренные сконструированные отличные от человека животные содержат гетерологичный генетический материал, который вставлен таким образом, что антитела, содержащие легкие цепи, которые имеют человеческий Vλ-домен и человеческий или отличный от человеческого Cλ-домен, экспрессируются из сконструированных локусов легких λ-цепей Ig, которые содержат человеческие и отличные от человеческих энхансерные области (или последовательности) λ-цепи Ig в геноме зародышевого типа отличного от человека животного.
[00158] Не желая связывать себя какой-либо конкретной теорией, предполагают, что в вариантах осуществления отличных от человека животных, описанных в данном документе, обеспечивается улучшенная система in vivo, в которой используется экспрессия антител, содержащих человеческие Vλ-домены, для получения терапевтических антител. Также предполагается, что в вариантах осуществления отличных от человека животных, описанных в данном документе, в некоторых вариантах осуществления предусмотрены альтернативные сконструированные формы локусов легких λ-цепей Ig, которые содержат гетерологичный генетический материал, для разработки терапевтических средств на основе человеческих антител (например, моноклональные человеческие антитела, полиспецифические связывающие средства, scFv, гибридные полипептиды и т.д.), направленных на вызывающие заболевания мишени, связанные со смещенными ответами антител (например, ответы антител, характеризующиеся преобладающей долей легких либо κ-, либо λ-цепей). Таким образом, варианты осуществления отличных от человека животных, описанных в данном документе, в частности, применимы для разработки человеческих антител, направленных на мишени, связанные с низкой иммуногенностью (например, вирусы) по причине, отчасти, отклоненных репертуаров и/или ответов антител.
[00159] В частности, в определенных аспектах в настоящем изобретении описано получение отличного от человека животного (например, грызун, такой как крыса или мышь), имеющего геном зародышевого типа, который содержит сконструированный локус легкой λ-цепи Ig, который, в некоторых вариантах осуществления, характеризуется введением множества последовательностей человеческих генов Vλ, Jλ и Cλ, функционально связанных с отличной от человеческой Cλ-областью, что приводит к экспрессии антител, которые содержат легкие цепи, содержащие человеческий Vλ-домен и человеческий или отличный от человеческого Cλ-домен. Как описано в данном документе, получение такого сконструированного локуса легкой λ-цепи Ig приводит к экспрессии антител, которые содержат легкие цепи, содержащие человеческий Vλ-домен и человеческий или отличный от человеческого Cλ-домен, из указанного сконструированного локуса легкой λ-цепи Ig в геноме зародышевого типа отличного от человека животного. В некоторых вариантах осуществления геном зародышевого типа предусмотренных отличных от человека животных дополнительно содержит (1) гуманизированные локусы H-цепи Ig и κ-цепи Ig или (2) гуманизированный локус H-цепи Ig и функционально подавленные или иным образом лишенные функциональности локусы легких κ-цепей Ig. Предусмотренные отличные от человека животные, описанные в данном документе, экспрессируют репертуары антител, которые содержат легкие λ-цепи Ig, содержащие человеческие Vλ-домены.
[00160] В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, содержат последовательности человеческой и отличной от человеческой легких λ-цепей Ig в одном локусе легкой λ-цепи Ig. В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, содержат последовательности человеческих легких λ-цепей Ig и последовательности мышиных (например, мышь или крыса) легких λ-цепей Ig в локусе легкой λ-цепи Ig. Во многих вариантах осуществления отличных от человека животных, описанных в данном документе, последовательности отличных от человеческих легких λ-цепей Ig представляют собой или содержат мышиные последовательности (например, мышь или крыса).
[00161] В некоторых вариантах осуществления последовательности легких λ-цепей Ig содержат межгенную ДНК, которая происходит от человека и/или мышиных (например, мышь или крыса). В некоторых вариантах осуществления последовательности легких λ-цепей Ig содержат межгенную ДНК, которая является синтетической и основана на первичной последовательности, которая происходит от человека и/или мышиных (например, мышь или крыса). В некоторых вариантах осуществления указанная межгенная ДНК происходит из того же локуса иммуноглобулина, в который межгенная ДНК таким образом помещена, вставлена, расположена или встроена (например, межгенная ДНК λ-цепи Ig в локусе легкой λ-цепи Ig). В некоторых определенных вариантах осуществления отличные от человека животные, описанные в данном документе, содержат сконструированный локус легкой λ-цепи Ig, который содержит межгенную ДНК, содержащую последовательность(-ти) легких λ-цепей Ig отличного от человеческого происхождения (например, последовательность легкой λ-цепи Ig мыши или крысы).
[00162] В различных вариантах осуществления гуманизированный локус H-цепи Ig содержит множество сегментов человеческих генов VH, DH и JH, функционально связанных с отличной от человеческой константной областью H-цепи Ig (например, эндогенная отличная от человеческой константная область H-цепи Ig, которая содержит один или более генов константных областей H-цепи Ig, как, например, IgM, IgG и т.д.). В различных вариантах осуществления гуманизированный локус легкой κ-цепи Ig содержит множество сегментов человеческих генов Vκ и Jκ, функционально связанных с отличной от человеческой константной областью κ-цепи Ig. В некоторых вариантах осуществления предусмотренные отличные от человека животные имеют геном зародышевого типа, который содержит локусы (или аллели) иммуноглобулинов, изображенные в графических материалах, предусмотренных в данном документе (например, см. фигуры 1, 2, 3 и/или 4). Такие сконструированные отличные от человека животные обеспечивают источник человеческих антител и фрагментов человеческих антител и/или нуклеиновых кислот, кодирующих такие человеческие антитела и фрагменты человеческих антител, и обеспечивают улучшенную систему in vivo, подходящую для использования человеческих Vλ-последовательностей, предназначенных для получения человеческих терапевтических антител.
[00163] Как описано в данном документе, в определенных вариантах осуществления предусмотрены отличные от человека животные, имеющие геном, в котором содержится множество сегментов человеческих генов легких λ-цепей (например, Vλ, Jλ и Cλ) вместо сегментов отличных от человеческих генов легких λ-цепей иммуноглобулина в эндогенных локусах легких λ-цепей иммуноглобулина и содержится некодирующая межгенная ДНК человека между сегментами человеческих генов вариабельной области. В некоторых вариантах осуществления отличные от человека животные, предусмотренные в данном документе, имеют геном, в котором дополнительно содержатся сегменты человеческих генов вариабельной области тяжелых (т.е. VH, DH и JH) и легких κ-цепей (например, Vκ и Jκ) вместо сегментов отличных от человеческих генов вариабельной области тяжелых (т.е. VH, DH и JH) и легких κ-цепей (например, Vκ и Jκ) в эндогенных локусах тяжелых и легких κ-цепей иммуноглобулина соответственно. Во многих вариантах осуществления сегменты человеческих генов иммуноглобулина (тяжелых и/или легких цепей) сконструированы с помощью межгенной ДНК человека (т.е. некодирующей межгенной ДНК иммуноглобулина человека), которая в природе связана с указанными сегментами генов (т.е. некодирующей геномной ДНК, связанной с указанными сегментами генов, которая встречается в природе в человеческом локусе иммуноглобулина клетки человека). Такая межгенная ДНК включает, например, промоторы, лидерные последовательности и последовательности сигнала рекомбинации, которые обеспечивают подходящую рекомбинацию и экспрессию сегментов человеческих генов в случае вариабельных доменов антител. Специалисты в данной области поймут, что отличные от человеческих локусы иммуноглобулинов также содержат такую некодирующую межгенную ДНК, и что, после ознакомления с данным изобретением, можно применять другую человеческую или отличную от человеческой межгенную ДНК для создания таких сконструированных локусов иммуноглобулинов, что приводит к тому же уровню экспрессии человеческих вариабельных доменов в случае антител у отличного от человека животного. Такие подобные сконструированные локусы иммуноглобулинов должны лишь содержать кодирующие человеческие последовательности (т.е. экзоны) из требуемых сегментов человеческих генов или комбинации сегментов человеческих генов для достижения экспрессии антител, которые содержат человеческие вариабельные домены.
[00164] Различные аспекты некоторых вариантов осуществления подробно описаны в следующих разделах, каждый из которых можно применять в отношении любого аспекта или варианта осуществления, описанного в данном документе. Использование разделов не предназначено для ограничения, и использование «или» означает «и/или», если не указано иное.
Репертуары антител у отличных от человека животных
[00165] Иммуноглобулины (также называемые антителами) представляют собой крупные (~150 кДа) гликопротеины Y-образной формы, которые продуцируются B-клетками иммунной системы хозяина для нейтрализации чужеродных антигенов (например, вирусов, бактерий и т.д.). Каждый иммуноглобулин (Ig) состоит из двух идентичных тяжелых цепей и двух идентичных легких цепей, каждая из которых имеет два структурных компонента: вариабельный домен и константный домен. Вариабельные домены тяжелых и легких цепей различаются у антител, продуцируемых различными B-клетками, но являются идентичными у всех антител, продуцируемых одной B-клеткой или клоном B-клетки. Вариабельные домены тяжелых и легких цепей каждого антитела совместно составляют антигенсвязывающую область (или антигенсвязывающий участок). Иммуноглобулины могут существовать в виде различных разновидностей, которые называются изотипами или классами, на основании константных областей (или доменов) тяжелых цепей, которые они содержат. Константный домен тяжелой цепи является идентичным во всех антителах одного и того же изотипа, но отличается в антителах различных изотипов. В таблице ниже кратко описаны девять изотипов антител мыши и человека (человеческих).

[00166] Дополнительные изотипы были идентифицированы у других видов. Изотипы придают специализированные биологические свойства антителу благодаря различным структурным характеристикам между различными изотипами, и они обнаруживаются в различных месторасположениях (клетках, тканях и т.д.) в организме животного. Изначально B-клетки продуцируют IgM и IgD с идентичными антигенсвязывающими областями. После активации B-клетки переключаются на различные изотипы с помощью процесса, называемого переключением классов, который предусматривает изменение константного домена антитела, продуцированного B-клеткой, при этом вариабельные домены остаются теми же, тем самым сохраняя антигенную специфичность исходного антитела (B-клетка).
[00167] Два отдельных локуса (κ-цепи Ig и λ-цепи Ig) содержат сегменты генов, которые кодируют легкие цепи антител, и проявляют как аллельное, так и изотипическое исключение. Соотношения экспрессии κ+и λ+ B-клеток варьируются среди видов. Например, у людей показано соотношение, составляющее приблизительно 60:40 (κ:λ). У мышей и крыс наблюдается соотношение, составляющее 95:5 (κ:λ). Интересно отметить, соотношение κ:λ, наблюдаемое у кошек (5:95), является противоположным таковому у мышей и крыс. Было проведено несколько исследований с целью выяснить возможные причины, лежащие в основе таких наблюдаемых соотношений, и были предложены в качестве обоснования как сложность локуса (т.е., число сегментов генов, в частности сегментов гена V), так и эффективность перегруппировки сегментов генов. Человеческий локус легкой λ-цепи иммуноглобулина имеет протяженность свыше 1000 т. н. и содержит примерно 70 сегментов гена Vλ (от 29 до 33 функциональных) и семь пар сегментов генов Jλ-Cλ (от четырех до пяти функциональных), организованных в три кластера (см., например, фиг.1 из патента США №9006511). Большинство наблюдаемых областей Vλ в репертуаре экспрессируемых антител закодировано сегментами генов, содержащимися в наиболее проксимальном кластере (т.е. кластере A). Локус легкой λ-цепи иммуноглобулина мыши поразительно отличается от человеческого локуса и, в зависимости от линии, содержит лишь несколько сегментов генов Vλ и Jλ , организованных в два различных генных кластера (см., например, фиг.2 из патента США №9006511).
[00168] Разработка терапевтических антител для лечения различных заболеваний человека главным образом была сконцентрирована на создании сконструированных линий отличных от человека животных, в частности сконструированных линий грызунов, несущих различные количества генетического материала в своих геномах, соответствующего человеческим генам иммуноглобулинов (краткий обзор изложен, например, в  , M. et al., 2015, Arch. Immunol. Ther. Exp.63:101-8). Первоначальные попытки создания таких линий трансгенных грызунов фокусировались на интеграции частей человеческих локусов иммуноглобулинов, которые могли, сами по себе, обеспечивать рекомбинацию сегментов генов и получение тяжелых и/или легких цепей, которые являлись полностью человеческими, при этом имели инактивированные эндогенные локусы иммуноглобулинов (см., например, , M. et al., 1989, Proc. Nat. Acad. Sci. U.S.A. 86:67-09-13; , M. et al., 1991, Eur. J. Immunol. 21:1323-6; Taylor, L.D. et al., 1992, Nucl. Acids Res. 20:6287-6295; Davies, N.P. et al., 1993, Biotechnol. 11:911-4; Green, L.L. et al., 1994, Nat. Genet. 7:13-21; Lonberg, N. et al., 1994, Nature 368:856-9; Taylor, L.D. et al., 1994, Int. Immunol. 6:579-91; Wagner, S.D. et al., 1994, Eur. J. Immunol. 24:2672-81; Fishwild, D.M. et al., 1996, Nat. Biotechnol. 14:845-51; Wagner, S.D. et al., 1996, Genomics 35:405-14; Mendez, M.J. et al., 1997, Nat. Genet. 15:146-56; Green, L.L. et al., 1998, J. Exp.Med. 188:483-95; Xian, J. et al., 1998, Transgenics 2:333-43; Little, M. et al., 2000, Immunol. Today 21:364-70; Kellermann, S.A. and L.L. Green, 2002, Cur. Opin. Biotechnol. 13:593-7). В частности, некоторые попытки включали интеграцию последовательностей человеческих легких λ-цепей Ig (см., например, публикации заявок на выдачу патентов США №№2002/0088016 A1, 2003/0217373 A1 и 2011/0236378 A1; патенты США №№6998514 и 7435871; Nicholson, I.C. et al., 1999, J. Immunol. 163:6898-906; Popov, A.V et al., 1999, J. Exp.Med. 189(10):1611-19). Такие попытки были сфокусированы на случайной интеграции дрожжевых искусственных хромосом, содержащих последовательности человеческих Vλ, Jλ и Cλ, с созданием тем самым линий мышей, которые экспрессируют полностью человеческие легкие λ-цепи (т.е. человеческий вариабельный и человеческий константный домены). Более поздние попытки предусматривали сходные стратегии с применением конструкций, которые также содержат последовательности человеческих Vλ, Jλ и Cλ (Osborn, M.J. et al., 2013, J. Immunol. 190:1481-90; Lee, E-C. et al., 2014, Nat. Biotech. 32(4):356-63).
, M. et al., 2015, Arch. Immunol. Ther. Exp.63:101-8). Первоначальные попытки создания таких линий трансгенных грызунов фокусировались на интеграции частей человеческих локусов иммуноглобулинов, которые могли, сами по себе, обеспечивать рекомбинацию сегментов генов и получение тяжелых и/или легких цепей, которые являлись полностью человеческими, при этом имели инактивированные эндогенные локусы иммуноглобулинов (см., например, , M. et al., 1989, Proc. Nat. Acad. Sci. U.S.A. 86:67-09-13; , M. et al., 1991, Eur. J. Immunol. 21:1323-6; Taylor, L.D. et al., 1992, Nucl. Acids Res. 20:6287-6295; Davies, N.P. et al., 1993, Biotechnol. 11:911-4; Green, L.L. et al., 1994, Nat. Genet. 7:13-21; Lonberg, N. et al., 1994, Nature 368:856-9; Taylor, L.D. et al., 1994, Int. Immunol. 6:579-91; Wagner, S.D. et al., 1994, Eur. J. Immunol. 24:2672-81; Fishwild, D.M. et al., 1996, Nat. Biotechnol. 14:845-51; Wagner, S.D. et al., 1996, Genomics 35:405-14; Mendez, M.J. et al., 1997, Nat. Genet. 15:146-56; Green, L.L. et al., 1998, J. Exp.Med. 188:483-95; Xian, J. et al., 1998, Transgenics 2:333-43; Little, M. et al., 2000, Immunol. Today 21:364-70; Kellermann, S.A. and L.L. Green, 2002, Cur. Opin. Biotechnol. 13:593-7). В частности, некоторые попытки включали интеграцию последовательностей человеческих легких λ-цепей Ig (см., например, публикации заявок на выдачу патентов США №№2002/0088016 A1, 2003/0217373 A1 и 2011/0236378 A1; патенты США №№6998514 и 7435871; Nicholson, I.C. et al., 1999, J. Immunol. 163:6898-906; Popov, A.V et al., 1999, J. Exp.Med. 189(10):1611-19). Такие попытки были сфокусированы на случайной интеграции дрожжевых искусственных хромосом, содержащих последовательности человеческих Vλ, Jλ и Cλ, с созданием тем самым линий мышей, которые экспрессируют полностью человеческие легкие λ-цепи (т.е. человеческий вариабельный и человеческий константный домены). Более поздние попытки предусматривали сходные стратегии с применением конструкций, которые также содержат последовательности человеческих Vλ, Jλ и Cλ (Osborn, M.J. et al., 2013, J. Immunol. 190:1481-90; Lee, E-C. et al., 2014, Nat. Biotech. 32(4):356-63).
[00169] Еще одни попытки включали специфическую вставку сегментов человеческих генов Vλ и Jλ в эндогенные локусы легких цепей Ig грызуна (κ-цепь и λ-цепь) таким образом, что указанные сегменты человеческих генов Vλ и Jλ функционально связаны с эндогенными константными областями легких цепей Ig (см., например, патенты США №№9006511, 9012717, 9029628, 9035128, 9066502, 9150662 и 9163092; все из которых включены в данный документ с помощью ссылки в полном объеме). У таких животных все из сегментов человеческого гена Vλ из кластеров A и B и либо один, либо четыре сегмента человеческого гена Jλ вставлены в эндогенные локусы легких κ-цепей Ig и λ-цепей Ig. В результате несколько различных сегментов человеческих генов Vλ и Jλ продемонстрировали надлежащую перегруппировку в обоих сконструированных локусах легких цепей Ig грызуна с образованием функциональных человеческих Vλ-доменов, которые экспрессировались в тесной связи как с Cκ-областями, так и с Cλ-областями в легких цепях репертуара антител грызуна (см., например, таблицу 7 и фигуры 11-13 из патента США №9006511). В частности, мыши, имеющие сконструированные локусы легких κ-цепей Ig, несущие сегменты человеческих генов Vλ и Jλ, демонстрировали соотношение κ:λ, составляющее приблизительно 1:1 в отделе, представляющем собой селезенку (см., например, таблицу 4 из патента США №9006511). Действительно, обе сконструированные линии мышей (т.е. сконструированные локусы легких κ-цепей Ig или λ-цепей Ig) продемонстрировали, что человеческие Vλ-домены можно экспрессировать из эндогенных локусов легких цепей Ig у грызунов, которые обычно проявляют большое смещение в экспрессии легких цепей (см. выше). Настоящее изобретение основано на признании того, что можно получать другие структуры сконструированного локуса легкой цепи Ig с целью повышения до максимума использования сегментов человеческих генов Vλ и Jλ в репертуарах антител в отношении терапевтических мишеней у отличных от человека животных, в частности, по сравнению с отличными от человека животными, которые содержат локус легкой λ-цепи Ig, который характеризуется отсутствием сложности и высокого качества (например, мыши и крысы), обычно связанных с человеческим локусом легкой λ-цепи Ig (т.е. который находится в клетке человека). Такие альтернативные структуры сконструированного локуса легкой цепи Ig обеспечивают способность к получению уникальных репертуаров антител, обусловленного их строением.
[00170] В настоящем изобретении описано, среди прочего, успешное получение отличного от человека животного, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи Ig, содержащий множество сегментов человеческих генов Vλ, Jλ и Cλ, функционально связанных с отличной от человеческой константной областью легкой λ-цепи Ig. В частности, в настоящем изобретении конкретно продемонстрировано успешное получение сконструированного отличного от человека животного, которое экспрессирует антитела, содержащие человеческие вариабельные домены и отличные от человеческих константные домены, при этом антитела включают легкие цепи, которые содержат человеческий Vλ-домен. Как описано в данном документе, экспрессии таких легких цепей достигают путем вставки указанного множества сегментов человеческих генов Vλ, Jλ и Cλ в эндогенный локус (или аллель) легкой λ-цепи Ig. Так же, как описано в данном документе, предусмотренные отличные от человека животные в некоторых вариантах осуществления сконструированы таким образом, что экспрессия легких цепей, которые содержат эндогенные Vλ-домены, является инактивированной (например, с помощью делеции генов). Таким образом, в настоящем изобретении, в по меньшей мере некоторых вариантах осуществления, охвачена разработка улучшенной системы in vivo для получения человеческих антител посредством обеспечения сконструированного отличного от человека животного, содержащего альтернативно сконструированный локус легкой λ-цепи Ig, что приводит к экспрессируемому репертуару антител, содержащему человеческие Vλ-домены.
ДНК-вставки
[00171] Как правило, молекула полинуклеотида, содержащая последовательности человеческих легких λ-цепей Ig (например, Vλ, Jλ, Cλ и энхансеры λ-цепи Ig) или их часть(части) вставлена в вектор, предпочтительно ДНК- вектор, с целью репликации молекулы полинуклеотида в клетке-хозяине.
[00172] Последовательности человеческих легких λ-цепей Ig можно клонировать непосредственно из известных последовательностей или источников (например, библиотеки) или синтезировать из последовательностей зародышевых типов, спроектированных in silico на основе опубликованных последовательностей, доступных из GenBank или других находящихся в открытом доступе баз данных (например, IMGT). В качестве альтернативы, библиотеки бактериальных искусственных хромосом (BAC) могут предоставлять представляющие интерес последовательности ДНК иммуноглобулинов (например, сегменты человеческого гена Vλ, пары сегментов человеческих генов Jλ-Cλ, человеческие Eλ-области или -последовательности и их комбинации). Библиотеки BAC могут содержать вставки размером 100-150 т. н., и они способны нести вставки размером до 300 т. н. (Shizuya, et al., 1992, Proc. Natl. Acad. Sci., USA 89:8794-8797; Swiatek, et al., 1993, Genes and Development 7:2071-2084; Kim, et al., 1996, Genomics 34 213-218; включенные в данный документ посредством ссылки в полном объеме). Например, была описана библиотека BAC человека, несущая вставки со средним размером 164-196 т. н. (Osoegawa, K. et al., 2001, Genome Res. 11(3):483-96; Osoegawa, K. et al., 1998, Genomics 52:1-8, статья с номером GE985423). Геномные библиотеки BAC человека и мыши были сконструированы и являются коммерчески доступными (например, ThermoFisher). Геномные библиотеки BAC также могут служить источником последовательностей ДНК иммуноглобулинов, а также областей транскрипционного контроля.
[00173] В качестве альтернативы, последовательности ДНК иммуноглобулинов можно выделять, клонировать и/или переносить из искусственных хромосом дрожжей (YAC). Например, была определена нуклеотидная последовательность человеческого локуса легкой λ-цепи Ig (см., например, Dunham, I. et al., 1999, Nature 402:489-95). Кроме того, ранее YAC были применены для сборки трансгена человеческого локуса легкой λ-цепи Ig (см., например, Popov, A.V. et al., 1996, Gene 177:195-201; Popov, A.V. et al., 1999, J. Exp.Med. 189(10):1611-19). Полный локус легкой λ-цепи Ig (человека или грызуна) можно клонировать, и он может содержаться в нескольких YAC. Если используются несколько YAC, и они содержат перекрывающиеся гомологичные области, их можно рекомбинировать в штаммах дрожжей-хозяев с получением единой конструкции, представляющей полный локус или необходимые части локуса (например, область, на которую нацеливаются с помощью нацеливающего вектора). Плечи YAC можно дополнительно модифицировать с помощью кассет для отбора для клеток млекопитающих путем их усовершенствования таким образом, чтобы содействовать введению конструкций в эмбриональные стволовые клетки или эмбрионы с помощью способов, известных из уровня техники и/или описанных в данном документе.
[00174] ДНК и аминокислотные последовательности сегментов человеческих генов легких λ-цепей Ig для применения в построении сконструированного локуса легкой λ-цепи Ig, описанного в данном документе, могут быть получены из опубликованных баз данных (например, GenBank, IMGT и т.д.) и/или опубликованных последовательностей антител. ДНК-вставки, содержащие сегменты человеческих генов легких λ-цепей Ig, в некоторых вариантах осуществления содержат одну или более энхансерных последовательностей (или областей) человеческих легких λ-цепей Ig. В некоторых вариантах осуществления ДНК-вставки содержат энхансерную последовательность (или область) человеческой легкой λ-цепи Ig, которая содержит один или более элементов последовательности, например один, два, три и т.д. В некоторых определенных вариантах осуществления ДНК-вставки содержат энхансерную последовательность (или область) человеческой легкой λ-цепи Ig, называемую человеческим Eλ, которая имеет три различных элемента последовательности. Таким образом, в некоторых вариантах осуществления человеческий Eλ, описанный в данном документе, является модульным, и один или более элементов последовательности функционируют вместе как энхансерная последовательность (или область). В некоторых определенных вариантах осуществления ДНК-вставки, содержащие энхансерные последовательности человеческих легких λ-цепей Ig, содержат энхансерные последовательности человеческих легких λ-цепей Ig, функционально связанные с последовательностью отличной от человеческой легкой λ-цепи Ig (например, последовательность отличной от человеческой константной области легкой λ-цепи Ig). В некоторых определенных вариантах осуществления ДНК-вставки, содержащие энхансерные последовательности человеческих легких λ-цепей Ig, содержат энхансерные последовательности человеческих легких λ-цепей Ig, функционально связанные с последовательностью отличной от человеческой легкой λ-цепи Ig (например, последовательность отличной от человеческой константной области легкой λ-цепи Ig) и функционально связанные с одним или более сегментами человеческого гена Vλ, одной или более парами сегментов человеческих генов Jλ-Cλ и/или одним или более сегментами человеческого гена Jλ. В некоторых определенных вариантах осуществления ДНК-вставки, содержащие энхансерные последовательности человеческих легких λ-цепей Ig, содержат энхансерные последовательности человеческих легких λ-цепей Ig, функционально связанные с последовательностью отличной от человеческой легкой λ-цепи Ig (например, последовательность отличной от человеческой константной области легкой λ-цепи Ig), одним или более сегментами человеческого гена Vλ, одним или более сегментами человеческого гена Jλ и одним или более сегментами человеческого гена Cλ.
[00175] ДНК-вставки можно получать с помощью способов, известных из уровня техники. Например, ДНК-вставку можно получить как часть большей плазмиды. Такое получение позволяет эффективно клонировать и отбирать правильные конструкции, как это известно из уровня техники. ДНК-вставки, содержащие полные или неполные последовательности человеческих легких λ-цепей Ig, описанные в данном документе, можно располагать между подходящими участками рестрикции на плазмиде таким образом, что их можно легко выделять из оставшихся последовательностей плазмиды для включения в требуемое отличное от человека животное.
[00176] Различные способы, используемые в получении плазмид и трансформации организмов-хозяев, известны из уровня техники. В отношении других систем экспрессии, подходящих как для прокариотических, так и для эукариотических клеток, а также общих процедур рекомбинации см. Principles of Gene Manipulation: An Introduction to Genetic Manipulation, 5th Ed., ed. By Old, R.W. and S.B. Primrose, Blackwell Science, Inc., 1994, и Molecular Cloning: A Laboratory Manual, 2nd Ed., ed. by Sambrook, J. et al., Cold Spring Harbor Laboratory Press: 1989.
Нацеливающие векторы
[00177] Нацеливающие векторы можно применять для введения ДНК-вставки в геномный целевой локус, и они содержат ДНК-вставку и плечи гомологии, которые фланкируют указанную ДНК-вставку. Нацеливающие векторы могут быть в линейной форме или в кольцевой форме, и они могу быть однонитевыми или двухнитевыми. Нацеливающие векторы могут представлять собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). Для удобства пользования плечи гомологии называются в данном документе как 5'- и 3'- (т.е. против хода и по ходу транскрипции) плечи гомологии. Эта терминология относится к относительному положению плеч гомологии относительно ДНК-вставки в нацеливающем векторе. 5'- и 3'-плечи гомологии соответствуют областям в локусе, подлежащем нацеливанию, или области в другом нацеливающем векторе, которые соответственно называются в данном документе как «целевая 5'-последовательность» и «целевая 3'-последовательность». В некоторых вариантах осуществления плечи гомологии также могут функционировать как целевая 5'- или 3'-последовательность.
[00178] В некоторых вариантах осуществления способы, описанные в данном документе, предусматривают два, три или более нацеливающих векторов, которые способны к рекомбинации друг с другом. В различных вариантах осуществления нацеливающие векторы представляют собой крупные нацеливающие векторы (LTVEC), описанные где-либо еще в данном документе. В таких вариантах осуществления каждый из первого, второго и третьего нацеливающих векторов содержит 5'- и 3'-плечо гомологии. 3'-Плечо гомологии первого нацеливающего вектора содержит последовательность, которая перекрывается с 5'-плечом гомологии второго нацеливающего вектора (т.е. перекрывающиеся последовательности), которые обеспечивают гомологичную рекомбинацию между первым и вторым LTVEC.
[00179] В случае способов двойного нацеливания 5'-плечо гомологии первого нацеливающего вектора и 3'-плечо гомологии второго нацеливающего вектора гомологичны соответствующим сегментам в целевом геномном локусе (т.е. целевой последовательности), что способствует гомологичной рекомбинации первого и второго нацеливающих векторов с соответствующими геномными сегментами и обеспечивает модификацию целевого геномного локуса.
[00180] В случае способов тройного нацеливания 3'-плечо гомологии второго нацеливающего вектора содержит последовательность, которая перекрывается с 5'-плечом гомологии третьего нацеливающего вектора (т.е. перекрывающиеся последовательности), которые обеспечивают гомологичную рекомбинацию между вторым и третьим LTVEC. 5'-Плечо гомологии первого нацеливающего вектора и 3'-плечо гомологии третьего нацеливающего вектора гомологичны соответствующим сегментам в целевом геномном локусе (т.е. целевой последовательности), что способствует гомологичной рекомбинации первого и третьего нацеливающих векторов с соответствующими геномными сегментами и обеспечивает модификацию целевого геномного локуса.
[00181] Плечо гомологии и целевая последовательность или два плеча гомологии «соответствуют» или «являются соответствующими» друг другу, если две области имеют достаточный уровень идентичности последовательностей друг с другом, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации. Термин «гомология» предусматривает последовательности ДНК, которые либо идентичны, либо обладают идентичностью последовательности с соответствующей последовательностью. Идентичность последовательности между указанной целевой последовательностью и соответствующим плечом гомологии, выявленным в нацеливающем векторе (т.е. перекрывающаяся последовательность), или между двумя плечами гомологии может представлять любую степень идентичности последовательностей, которая обеспечивает возникновение гомологичной рекомбинации. В качестве одного примера степень идентичности последовательности между плечом гомологии нацеливающего вектора (или его фрагмента) и целевой последовательностью другого нацеливающего вектора или целевой последовательностью целевого геномного локуса (или его фрагмента) может составлять, например, без ограничения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности, вследствие чего последовательности подвергаются гомологичной рекомбинации.
[00182] Кроме того, соответствующая область гомологии между плечом гомологии и соответствующей целевой последовательностью может иметь любую длину, достаточную для активации гомологичной рекомбинации в целевом геномном локусе. Например, данное плечо гомологии и/или соответствующая целевая последовательность могут содержать соответствующие области гомологии, которые имеют длину, например, без ограничения по меньшей мере приблизительно 5-10 т. н., 5-15 т. н., 5-20 т. н., 5-25 т. н., 5-30 т. н., 5-35 т. н., 5-40 т. н., 5-45 т. н., 5-50 т. н., 5-55 т. н., 5-60 т. н., 5-65 т. н., 5-70 т. н., 5-75 т. н., 5-80 т. н., 5-85 т. н., 5-90 т. н., 5-95 т. н., 5-100 т. н., 100-200 т. н. или 200-300 т. н. или больше (как описано где-либо еще в данном документе), вследствие чего плечо гомологии характеризуется достаточной гомологией, чтобы подвергнуться гомологичной рекомбинации с соответствующей(-ими) целевой(-ыми) последовательностью(-ями) в целевом геномном локусе клетки или в другом нацеливающем векторе. В некоторых вариантах осуществления данное плечо гомологии и/или соответствующая целевая последовательность содержат соответствующие области гомологии, которые имеют длину, например, без ограничения по меньшей мере приблизительно 10-100 т. н., 15-100 т. н., 20-100 т. н., 25-100 т. н., 30-100 т. н., 35-100 т. н., 40-100 т. н., 45-100 т. н., 50-100 т. н., 55-100 т. н., 60-100 т. н., 65-100 т. н., 70-100 т. н., 75-100 т. н., 80-100 т. н., 85-100 т. н., 90-100 т. н. или 95-100 т. н. или больше (как описано где-либо еще в данном документе), вследствие чего плечо гомологии характеризуется достаточной гомологией, чтобы подвергнуться гомологичной рекомбинации с соответствующей(-ими) целевой(-ыми) последовательностью(-ями) в целевом геномном локусе клетки или в другом нацеливающем векторе.
[00183] Перекрывающиеся последовательности 3'-плеча гомологии первого нацеливающего вектора и 5'-плеча гомологии второго нацеливающего вектора или 3'-плеча гомологии второго нацеливающего вектора и 5'-плеча гомологии третьего нацеливающего вектора могут быть любой длины, которая является достаточной для обеспечения гомологичной рекомбинации между указанными нацеливающими векторами. Например, данная перекрывающаяся последовательность плеча гомологии может содержать соответствующие перекрывающиеся области, которые имеют длину по меньшей мере приблизительно 1-5 т. н., 5-10 т. н., 5-15 т. н., 5-20 т. н., 5-25 т. н., 5-30 т. н., 5-35 т. н., 5-40 т. н., 5-45 т. н., 5-50 т. н., 5-55 т. н., 5-60 т. н., 5-65 т. н., 5-70 т. н., 5-75 т. н., 5-80 т. н., 5-85 т. н., 5-90 т. н., 5-95 т. н., 5-100 т. н., 100-200 т. н. или 200-300 т. н. или больше, вследствие чего перекрывающаяся последовательность плеча гомологии характеризуется достаточной гомологией, чтобы подвергнуться гомологичной рекомбинации с соответствующей перекрывающейся последовательностью в другом нацеливающем векторе. В некоторых вариантах осуществления данная перекрывающаяся последовательность плеча гомологии содержит перекрывающуюся область, которая имеет длину по меньшей мере приблизительно 1-100 т. н., 5-100 т. н., 10-100 т. н., 15-100 т. н., 20-100 т. н., 25-100 т. н., 30-100 т. н., 35-100 т. н., 40-100 т. н., 45-100 т. н., 50-100 т. н., 55-100 т. н., 60-100 т. н., 65-100 т. н., 70-100 т. н., 75-100 т. н., 80-100 т. н., 85-100 т. н., 90-100 т. н. или 95-100 т. н. или больше, вследствие чего перекрывающаяся последовательность плеча гомологии характеризуется достаточной гомологией, чтобы подвергнуться гомологичной рекомбинации с соответствующей перекрывающейся последовательностью в другом нацеливающем векторе. В некоторых вариантах осуществления перекрывающаяся последовательность составляет 1-5 т. н. включительно. В некоторых вариантах осуществления перекрывающаяся последовательность составляет от приблизительно 1 т. н. до приблизительно 70 т. н. включительно. В некоторых вариантах осуществления перекрывающаяся последовательность составляет от приблизительно 10 т. н. до приблизительно 70 т. н. включительно. В некоторых вариантах осуществления перекрывающаяся последовательность составляет от приблизительно 10 т. н. до приблизительно 50 т. н. включительно. В некоторых вариантах осуществления перекрывающаяся последовательность составляет по меньшей мере 10 т. н. В некоторых вариантах осуществления перекрывающаяся последовательность составляет по меньшей мере 20 т. н. Например, перекрывающаяся последовательность может составлять от приблизительно 1 т. н. до приблизительно 5 т. н. включительно, от приблизительно 5 т. н. до приблизительно 10 т. н. включительно, от приблизительно 10 т. н. до приблизительно 15 т. н. включительно, от приблизительно 15 т. н. до приблизительно 20 т. н. включительно, от приблизительно 20 т. н. до приблизительно 25 т. н. включительно, от приблизительно 25 т. н. до приблизительно 30 т. н. включительно, от приблизительно 30 т. н. до приблизительно 35 т. н. включительно, от приблизительно 35 т. н. до приблизительно 40 т. н. включительно, от приблизительно 40 т. н. до приблизительно 45 т. н. включительно, от приблизительно 45 т. н. до приблизительно 50 т. н. включительно, от приблизительно 50 т. н. до приблизительно 60 т. н. включительно, от приблизительно 60 т. н. до приблизительно 70 т. н. включительно, от приблизительно 70 т. н. до приблизительно 80 т. н. включительно, от приблизительно 80 т. н. до приблизительно 90 т. н. включительно, от приблизительно 90 т. н. до приблизительно 100 т. н. включительно, от приблизительно 100 т. н. до приблизительно 120 т. н. включительно, от приблизительно 120 т. н. до приблизительно 140 т. н. включительно, от приблизительно 140 т. н. до приблизительно 160 т. н. включительно, от приблизительно 160 т. н. до приблизительно 180 т. н. включительно, от приблизительно 180 т. н. до приблизительно 200 т. н. включительно, от приблизительно 200 т. н. до приблизительно 220 т. н. включительно, от приблизительно 220 т. н. до приблизительно 240 т. н. включительно, от приблизительно 240 т. н. до приблизительно 260 т. н. включительно, от приблизительно 260 т. н. до приблизительно 280 т. н. включительно или от приблизительно 280 т. н. до приблизительно 300 т. н. включительно. В качестве одного примера перекрывающаяся последовательность может составлять от приблизительно 20 т. н. до приблизительно 60 т. н. включительно. В качестве альтернативы, перекрывающаяся последовательность может составлять по меньшей мере 1 т. н., по меньшей мере 5 т. н., по меньшей мере 10 т. н., по меньшей мере 15 т. н., по меньшей мере 20 т. н., по меньшей мере 25 т. н., по меньшей мере 30 т. н., по меньшей мере 35 т. н., по меньшей мере 40 т. н., по меньшей мере 45 т. н., по меньшей мере 50 т. н., по меньшей мере 60 т. н., по меньшей мере 70 т. н., по меньшей мере 80 т. н., по меньшей мере 90 т. н., по меньшей мере 100 т. н., по меньшей мере 120 т. н., по меньшей мере 140 т. н., по меньшей мере 160 т. н., по меньшей мере 180 т. н., по меньшей мере 200 т. н., по меньшей мере 220 т. н., по меньшей мере 240 т. н., по меньшей мере 260 т. н., по меньшей мере 280 т. н. или по меньшей мере 300 т. н.
[00184] Плечи гомологии, в некоторых вариантах осуществления, могут соответствовать локусу, который является нативным по отношению к клетке (например, локус, подлежащий нацеливанию), или, в качестве альтернативы, они могут соответствовать области гетерологичного или экзогенного сегмента ДНК, который был интегрирован в геном клетки, в том числе, например, трансгенам, кассетам экспрессии или гетерологичным или экзогенным областям ДНК. В качестве альтернативы, плечи гомологии, в некоторых вариантах осуществления, могут соответствовать области нацеливающего вектора в клетке. В некоторых вариантах осуществления плечи гомологии нацеливающего вектора могут соответствовать области искусственной хромосомы дрожжей (YAC), бактериальной искусственной хромосомы (BAC), искусственной хромосомы человека или любой другой сконструированной области, содержащейся в соответствующей клетке-хозяине. Кроме того, плечи гомологии нацеливающего вектора могут соответствовать или быть получены из области библиотеки BAC, космидной библиотеки или библиотеки фагов P1. В некоторых определенных вариантах осуществления плечи гомологии нацеливающего вектора соответствуют локусу, который является нативным, гетерологичным или экзогенным по отношении к прокариоту, клетке дрожжей, птице (например, курица), отличному от человека млекопитающему, грызуну, человеку, крысе, мыши, хомяку кролику, свинье, крупному рогатому скоту, оленю, овце, козе, кошке, собаке, хорьку, примату (например, игрунка, макак-резус), одомашненному млекопитающему, сельскохозяйственному млекопитающему или любому другому представляющему интерес организму. В некоторых вариантах осуществления плечи гомологии соответствуют локусу клетки, на который нельзя целенаправленно воздействовать с применением традиционного способа или на который можно целенаправленно воздействовать разве лишь некорректно или лишь со в значительной степени низкой эффективностью в отсутствие однонитевого разрыва или двухнитевого разрыва, индуцированного средством на основе нуклеаз (например, белок Cas). В некоторых вариантах осуществления плечи гомологии получены из синтетической ДНК.
[00185] В некоторых вариантах осуществления одно из 5'- или 3'-плеч гомологии нацеливающего(-их) вектора(-ов) соответствует геномному локусу, подлежащему нацеливанию, при этом другое из 5'- или 3'-плеч гомологии соответствует области другого нацеливающего вектора.
[00186] В некоторых вариантах осуществления 5'- и 3'-плечи гомологии нацеливающего(-их) вектора(-ов) соответствуют геному, подлежащему нацеливанию. В качестве альтернативы, плечи гомологии могут быть из родственного генома. Например, геном, подлежащий нацеливанию, представляет собой геном мыши из первой линии, а нацеливающие плечи происходят из генома мыши из второй линии, где первая линия и вторая линия являются различными. В определенных вариантах осуществления плечи гомологии происходят из генома одного и того же животного или из генома одной и той же линии, например, геном, подлежащий нацеливанию, представляет собой геном мыши из первой линии, а нацеливающие плечи происходят из генома мыши одной и той же мыши или из одной и той же линии.
[00187] Плечо гомологии нацеливающего вектора может иметь любую длину, которая является достаточной для обеспечения события гомологичной рекомбинации с соответствующей целевой последовательностью, в том числе, например, длину, составляющую по меньшей мере 1-5 т. н. включительно, 5-10 т. н. включительно, 5-15 т. н. включительно, 5-20 т. н. включительно, 5-25 т. н. включительно, 5-30 т. н. включительно, 5-35 т. н. включительно, 5-40 т. н. включительно, 5-45 т. н. включительно, 5-50 т. н. включительно, 5-55 т. н. включительно, 5-60 т. н. включительно, 5-65 т. н. включительно, 5-70 т. н. включительно, 5-75 т. н. включительно, 5-80 т. н. включительно, 5-85 т. н. включительно, 5-90 т. н. включительно, 5-95 т. н. включительно, 5-100 т. н. включительно, 100-200 т. н. включительно или 200-300 т. н. включительно или больше. В некоторых вариантах осуществления плечо гомологии нацеливающего вектора имеет длину, которая является достаточной для обеспечения события гомологичной рекомбинации с соответствующей целевой последовательностью, которая составляет длину по меньшей мере 1-100 т. н. включительно, 5-100 т. н. включительно, 10-100 т. н. включительно, 15-100 т. н. включительно, 20-100 т. н. включительно, 25-100 т. н. включительно, 30-100 т. н. включительно, 35-100 т. н. включительно, 40-100 т. н. включительно, 45-100 т. н. включительно, 50-100 т. н. включительно, 55-100 т. н. включительно, 60-100 т. н. включительно, 65-100 т. н. включительно, 70-100 т. н. включительно, 75-100 т. н. включительно, 80-100 т. н. включительно, 85-100 т. н. включительно, 90-100 т. н. включительно или 95-100 т. н. включительно или больше. Как описано в данном документе, в крупных нацеливающих векторах можно применять нацеливающие плечи большей длины.
[00188] Можно использовать средства на основе нуклеаз (например, системы CRISPR/Cas) в комбинации с нацеливающими векторами для облегчения модификации целевого локуса (например, локуса легкой λ-цепи Ig). Такие средства на основе нуклеаз могут способствовать гомологичной рекомбинации между нацеливающим вектором и целевым локусом. Если средства на основе нуклеаз применяют в комбинации с нацеливающим вектором, нацеливающий вектор может содержать 5'- и 3'-плечи гомологии, соответствующие целевым 5'- и 3'-последовательностям, расположенным достаточно близко к участку расщепления нуклеазами, чтобы способствовать точности события гомологичной рекомбинации между целевыми последовательностями и плечами гомологии после однонитевого разрыва или двухнитевого разрыва в участке расщепления нуклеазами. Термин «участок расщепления нуклеазами» предусматривает последовательность ДНК, в которой однонитевой или двухнитевой разрыв осуществляется с помощью средства на основе нуклеаз (например, участок расщепления для Cas9). Целевые последовательности в локусе, подлежащем нацеливанию, которые соответствуют 5'-и 3'-плечам гомологии нацеливающего вектора «расположены достаточно близко» к участку расщепления нуклеазами, если расстояние является таким, чтобы способствовать точности события гомологичной рекомбинации между целевыми 5'- и 3'-последовательностями и плечами гомологии после однонитевого или двухнитевого разрыва в участке распознавания. Таким образом, в определенных вариантах осуществления целевые последовательности, соответствующие 5'- и/или 3'-плечам гомологии нацеливающего вектора, находятся на расстоянии одного нуклеотида от данного участка распознавания или на расстоянии от по меньшей мере 10 нуклеотидов до приблизительно 14 т. н. от данного участка распознавания. В некоторых вариантах осуществления участок расщепления нуклеазами непосредственно примыкает к по меньшей мере одной или обеим из целевых последовательностей.
[00189] Пространственная взаимосвязь целевых последовательностей, которые соответствуют плечам гомологии нацеливающего вектора, и участка расщепления нуклеазами может варьироваться. Например, целевые последовательности могут быть расположены в 5'-направлении по отношению к участку расщепления нуклеазами, целевые последовательности могут быть расположены в 3'-направлении по отношению к участку распознавания, или целевые последовательности могут фланкировать участок расщепления нуклеазами.
[00190] Комбинированное применение нацеливающего вектора (в том числе, например, крупного нацеливающего вектора) со средством на основе нуклеаз может приводить к повышению эффективности нацеливания по сравнению с применением нацеливающего вектора отдельно. Например, если нацеливающий вектор применяют в сочетании со средством на основе нуклеаз, то эффективность нацеливания нацеливающего вектора может повышаться в по меньшей мере два раза, по меньшей мере три раза, по меньшей мере четыре раза, по меньшей мере пять раз, по меньшей мере шесть раз, по меньшей мере семь раз, по меньшей мере восемь раз, по меньшей мере девять раз, по меньшей мере десять раз или в диапазоне, образованном такими целыми числами, таком как 2-10 раз, по сравнению с применением нацеливающего вектора отдельно.
[00191] Некоторые нацеливающие векторы представляют собой «крупные нацеливающие векторы» или «LTVEC», которые включают нацеливающие векторы, содержащие плечи гомологии, которые соответствуют и получены из последовательностей нуклеиновых кислот, более крупных, чем таковые обычно используемые в других подходах, предназначенных для осуществления гомологичной рекомбинации в клетках. LTVEC может иметь длину, например, по меньшей мере 10 т. н., или общая сумма длин 5'-плеча гомологии и 3'-плеча гомологии может составлять, например, по меньшей мере 10 т. н. LTVEC также предусматривают нацеливающие векторы, содержащие ДНК-вставки, более крупные по размеру, чем таковые обычно используемые в других подходах, предназначенных для осуществления гомологичной рекомбинации в клетках. Например, LTVEC делают возможным модификацию крупных локусов, в отношении которых не могут быть применены традиционные нацеливающие векторы на основе плазмид из-за их ограничений по размеру. Например, локус, подлежащий нацеливанию, может представлять собой (т.е. 5'- и 3'-плечи гомологии могут соответствовать локусу) локус клетки, на который нельзя целенаправленно воздействовать с применением традиционных способов или на который можно целенаправленно воздействовать разве лишь некорректно или лишь со в значительной степени низкой эффективностью в отсутствие однонитевого разрыва или двухнитевого разрыва, индуцированного средством на основе нуклеаз (например, белок Cas).
[00192] В некоторых вариантах осуществления в способах, описанных в данном документе, применяют два или три LTVEC, которые способны к рекомбинации друг с другом и с целевым геномным локусом в трехнаправленном или четырехнаправленном событии рекомбинации. Такие способы делают возможной модификацию крупных локусов, чего невозможно достичь с применением единственного LTVEC.
[00193] Примеры LTVEC включают векторы, полученные из бактериальной искусственной хромосомы (BAC), искусственной хромосомы человека или искусственной хромосомы дрожжей (YAC). Примеры LTVEC и способы их получения описаны, например, в патентах США №№6586251, 6596541 и №7105348 и в публикации международной патентной заявки с номером WO 2002/036789, каждое из которых включено в данный документ посредством ссылки в полном объеме. LTVEC могут быть в линейной форме или кольцевой форме.
[00194] LTVEC могут иметь любую длину, в том числе, например, от приблизительно 20 т. н. до приблизительно 300 т. н. включительно, от приблизительно 20 т. н. до приблизительно 50 т. н. включительно, от приблизительно 50 т. н. до приблизительно 75 т. н. включительно, от приблизительно 75 т. н. до приблизительно 100 т. н. включительно, от приблизительно 100 т. н. до 125 т. н. включительно, от приблизительно 125 т. н. до приблизительно 150 т. н. включительно, от приблизительно 150 т. н. до приблизительно 175 т. н. включительно, от приблизительно 175 т. н. до приблизительно 200 т. н. включительно, от приблизительно 200 т. н. до приблизительно 225 т. н. включительно, от приблизительно 225 т. н. до приблизительно 250 т. н. включительно, от приблизительно 250 т. н. до приблизительно 275 т. н. включительно или от приблизительно 275 т. н. до приблизительно 300 т. н. включительно. В качестве альтернативы, LTVEC может составлять по меньшей мере 10 т. н., по меньшей мере 15 т. н., по меньшей мере 20 т. н., по меньшей мере 30 т. н., по меньшей мере 40 т. н., по меньшей мере 50 т. н., по меньшей мере 60 т. н., по меньшей мере 70 т. н., по меньшей мере 80 т. н., по меньшей мере 90 т. н., по меньшей мере 100 т. н., по меньшей мере 150 т. н., по меньшей мере 200 т. н., по меньшей мере 250 т. н., по меньшей мере 300 т. н., по меньшей мере 350 т. н., по меньшей мере 400 т. н., по меньшей мере 450 т. н. или по меньшей мере 500 т. н. или больше. В некоторых вариантах осуществления размер LTVEC может быть слишком большим, чтобы обеспечить возможность скрининга событий нацеливания с помощью традиционных анализов, например саузерн-блоттинга и ПЦР длинных фрагментов (например, от 1 т. н. до 5 т. н.).
[00195] В некоторых вариантах осуществления LTVEC содержит ДНК-вставку, имеющую длину в диапазоне от приблизительно 5 т. н. до приблизительно 200 т. н. включительно, от приблизительно 5 т. н. до приблизительно 10 т. н. включительно, от приблизительно 10 т. н. до приблизительно 20 т. н. включительно, от приблизительно 20 т. н. до приблизительно 30 т. н. включительно, от приблизительно 30 т. н. до приблизительно 40 т. н. включительно, от приблизительно 40 т. н. до приблизительно 50 т. н. включительно, от приблизительно 60 т. н. до приблизительно 70 т. н. включительно, от приблизительно 80 т. н. до приблизительно 90 т. н. включительно, от приблизительно 90 т. н. до приблизительно 100 т. н. включительно, от приблизительно 100 т. н. до приблизительно 110 т. н. включительно, от приблизительно 120 т. н. до приблизительно 130 т. н. включительно, от приблизительно 130 т. н. до приблизительно 140 т. н. включительно, от приблизительно 140 т. н. до приблизительно 150 т. н. включительно, от приблизительно 150 т. н. до приблизительно 160 т. н. включительно, от приблизительно 160 т. н. до приблизительно 170 т. н. включительно, от приблизительно 170 т. н. до приблизительно 180 т. н. включительно, от приблизительно 180 т. н. до приблизительно 190 т. н. включительно или от приблизительно 190 т. н. до приблизительно 200 т. н. включительно. В некоторых вариантах осуществления длина ДНК-вставки может находиться в диапазоне от приблизительно 5 т. н. до приблизительно 10 т. н. включительно, от приблизительно 10 т. н. до приблизительно 20 т. н. включительно, от приблизительно 20 т. н. до приблизительно 40 т. н. включительно, от приблизительно 40 т. н. до приблизительно 60 т. н. включительно, от приблизительно 60 т. н. до приблизительно 80 т. н. включительно, от приблизительно 80 т. н. до приблизительно 100 т. н. включительно, от приблизительно 100 т. н. до приблизительно 150 т. н. включительно, от приблизительно 150 т. н. до приблизительно 200 т. н. включительно, от приблизительно 200 т. н. до приблизительно 250 т. н. включительно, от приблизительно 250 т. н. до приблизительно 300 т. н. включительно, от приблизительно 300 т. н. до приблизительно 350 т. н. включительно или от приблизительно 350 т. н. до приблизительно 400 т. н. включительно. В некоторых вариантах осуществления LTVEC содержит ДНК-вставку, имеющую длину в диапазоне от приблизительно 400 т. н. до приблизительно 450 т. н. включительно, от приблизительно 450 т. н. до приблизительно 500 т. н. включительно, от приблизительно 500 т. н. до приблизительно 550 т. н. включительно, от приблизительно 550 т. н. до приблизительно 600 т. н. включительно, от приблизительно 600 т. н. до приблизительно 650 т. н. включительно, от приблизительно 650 т. н. до приблизительно 700 т. н. включительно, от приблизительно 700 т. н. до приблизительно 750 т. н. включительно или от приблизительно 750 т. н. до приблизительно 800 т. н. включительно.
[00196] В некоторых вариантах осуществления общая сумма длин 5'-плеча гомологии и 3-плеча гомологии LTVEC составляет по меньшей мере 10 т. н. В некоторых вариантах осуществления длина 5'-плеча гомологии LTVEC находится в диапазоне от приблизительно 1 т. н. до приблизительно 100 т. н. включительно, и/или длина 3'-плеча гомологии LTVEC находится в диапазоне от приблизительно 1 т. н. до приблизительно 100 т. н. включительно. Общая сумма длин 5'- и 3'-плеч гомологии может составлять, например, от приблизительно 1 т. н. до приблизительно 5 т. н. включительно, от приблизительно 5 т. н. до приблизительно 10 т. н. включительно, от приблизительно 10 т. н. до приблизительно 20 т. н. включительно, от приблизительно 20 т. н. до приблизительно 30 т. н. включительно, от приблизительно 30 т. н. до приблизительно 40 т. н. включительно, от приблизительно 40 т. н. до приблизительно 50 т. н. включительно, от приблизительно 50 т. н. до приблизительно 60 т. н. включительно, от приблизительно 60 т. н. до приблизительно 70 т. н. включительно, от приблизительно 70 т. н. до приблизительно 80 т. н. включительно, от приблизительно 80 т. н. до приблизительно 90 т. н. включительно, от приблизительно 90 т. н. до приблизительно 100 т. н. включительно, от приблизительно 100 т. н. до приблизительно 110 т. н. включительно, от приблизительно 110 т. н. до приблизительно 120 т. н. включительно, от приблизительно 120 т. н. до приблизительно 130 т. н. включительно, от приблизительно 130 т. н. до приблизительно 140 т. н. включительно, от приблизительно 140 т. н. до приблизительно 150 т. н. включительно, от приблизительно 150 т. н. до приблизительно 160 т. н. включительно, от приблизительно 160 т. н. до приблизительно 170 т. н. включительно, от приблизительно 170 т. н. до приблизительно 180 т. н. включительно, от приблизительно 180 т. н. до приблизительно 190 т. н. включительно или от приблизительно 190 т. н. до приблизительно 200 т. н. включительно. В качестве альтернативы, длина каждого плеча гомологии, в некоторых вариантах осуществления, может составлять по меньшей мере 5 т. н., по меньшей мере 10 т. н., по меньшей мере 15 т. н., по меньшей мере 20 т. н., по меньшей мере 30 т. н., по меньшей мере 40 т. н., по меньшей мере 50 т. н., по меньшей мере 60 т. н., по меньшей мере 70 т. н., по меньшей мере 80 т. н., по меньшей мере 90 т. н., по меньшей мере 100 т. н., по меньшей мере 110 т. н., по меньшей мере 120 т. н., по меньшей мере 130 т. н., по меньшей мере 140 т. н., по меньшей мере 150 т. н., по меньшей мере 160 т. н., по меньшей мере 170 т. н., по меньшей мере 180 т. н., по меньшей мере 190 т. н. или по меньшей мере 200 т. н. Аналогично, общая сумма длин 5'- и 3'-плеч гомологии, в некоторых вариантах осуществления, может составлять по меньшей мере 5 т. н., по меньшей мере 10 т. н., по меньшей мере 15 т. н., по меньшей мере 20 т. н., по меньшей мере 30 т. н., по меньшей мере 40 т. н., по меньшей мере 50 т. н., по меньшей мере 60 т. н., по меньшей мере 70 т. н., по меньшей мере 80 т. н., по меньшей мере 90 т. н., по меньшей мере 100 т. н., по меньшей мере 110 т. н., по меньшей мере 120 т. н., по меньшей мере 130 т. н., по меньшей мере 140 т. н., по меньшей мере 150 т. н., по меньшей мере 160 т. н., по меньшей мере 170 т. н., по меньшей мере 180 т. н., по меньшей мере 190 т. н. или по меньшей мере 200 т. н.
[00197] В некоторых вариантах осуществления LTVEC и ДНК-вставка спроектированы с обеспечением возможности делеции эндогенной последовательности в целевом локусе размером от приблизительно 5 т. н. до приблизительно 10 т. н. включительно, от приблизительно 10 т. н. до приблизительно 20 т. н. включительно, от приблизительно 20 т. н. до приблизительно 40 т. н. включительно, от приблизительно 40 т. н. до приблизительно 60 т. н. включительно, от приблизительно 60 т. н. до приблизительно 80 т. н. включительно, от приблизительно 80 т. н. до приблизительно 100 т. н. включительно или от приблизительно 100 т. н. до приблизительно 150 т. н. включительно, от приблизительно 150 т. н. до приблизительно 200 т. н. включительно, от приблизительно 200 т. н. до приблизительно 300 т. н. включительно, от приблизительно 300 т. н. до приблизительно 400 т. н. включительно, от приблизительно 400 т. н. до приблизительно 500 т. н. включительно, от приблизительно 500 т. н. до приблизительно 600 т. н. включительно, от приблизительно 600 т. н. до приблизительно 700 т. н. включительно, от приблизительно 700 т. н. до приблизительно 800 т. н. включительно или от приблизительно 500 т. н. до приблизительно 1 млн. н. включительно, от приблизительно 1 млн. н. до приблизительно 1,5 млн. н. включительно, от приблизительно 1,5 млн. н. до приблизительно 2 млн. н. включительно, от приблизительно 2 млн. н. до приблизительно 2,5 млн. н. включительно или от приблизительно 2,5 млн. н. до приблизительно 3 млн. н. включительно. В качестве альтернативы размер делеции может составлять от приблизительно 3 млн. н. до приблизительно 4 млн. н. включительно, от приблизительно 4 млн. н. до приблизительно 5 млн. н. включительно, от приблизительно 5 млн. н. до приблизительно 10 млн. н. включительно, от приблизительно 10 млн. н. до приблизительно 20 млн. н. включительно, от приблизительно 20 млн. н. до приблизительно 30 млн. н. включительно, от приблизительно 30 млн. н. до приблизительно 40 млн. н. включительно, от приблизительно 40 млн. н. до приблизительно 50 млн. н. включительно, от приблизительно 50 млн. н. до приблизительно 60 млн. н. включительно, от приблизительно 60 млн. н. до приблизительно 70 млн. н. включительно, от приблизительно 70 млн. н. до приблизительно 80 млн. н. включительно, от приблизительно 80 млн. н. до приблизительно 90 млн. н. включительно или от приблизительно 90 млн. н. до приблизительно 100 млн. н. включительно. В качестве альтернативы, размер делеции может составлять по меньшей мере 10 т. н., по меньшей мере 20 т. н., по меньшей мере 30 т. н., по меньшей мере 40 т. н., по меньшей мере 50 т. н., по меньшей мере 60 т. н., по меньшей мере 70 т. н., по меньшей мере 80 т. н., по меньшей мере 90 т. н., по меньшей мере 100 т. н., по меньшей мере 150 т. н., по меньшей мере 200 т. н., по меньшей мере 250 т. н., по меньшей мере 300 т. н., по меньшей мере 350 т. н., по меньшей мере 400 т. н., по меньшей мере 450 т. н. или по меньшей мере 500 т. н. или больше.
[00198] В некоторых вариантах осуществления LTVEC и ДНК-вставка спроектированы с обеспечением возможности вставки в целевой локус экзогенной последовательности нуклеиновой кислоты, имеющей размер в диапазоне от приблизительно 5 т. н. до приблизительно 10 т. н. включительно, от приблизительно 10 т. н. до приблизительно 20 т. н. включительно, от приблизительно 20 т. н. до приблизительно 40 т. н. включительно, от приблизительно 40 т. н. до приблизительно 60 т. н. включительно, от приблизительно 60 т. н. до приблизительно 80 т. н. включительно, от приблизительно 80 т. н. до приблизительно 100 т. н. включительно, от приблизительно 100 т. н. до приблизительно 150 т. н. включительно, от приблизительно 150 т. н. до приблизительно 200 т. н. включительно, от приблизительно 200 т. н. до приблизительно 250 т. н. включительно, от приблизительно 250 т. н. до приблизительно 300 т. н. включительно, от приблизительно 300 т. н. до приблизительно 350 т. н. включительно или от приблизительно 350 т. н. до приблизительно 400 т. н. включительно. В качестве альтернативы, размер вставки, в некоторых вариантах осуществления, может составлять от приблизительно 400 т. н. до приблизительно 450 т. н. включительно, от приблизительно 450 т. н. до приблизительно 500 т. н. включительно, от приблизительно 500 т. н. до приблизительно 550 т. н. включительно, от приблизительно 550 т. н. до приблизительно 600 т. н. включительно, от приблизительно 600 т. н. до приблизительно 650 т. н. включительно, от приблизительно 650 т. н. до приблизительно 700 т. н. включительно, от приблизительно 700 т. н. до приблизительно 750 т. н. включительно или от приблизительно 750 т. н. до приблизительно 800 т. н. включительно. В качестве альтернативы размер вставки может составлять, в некоторых вариантах осуществления, по меньшей мере 10 т. н., по меньшей мере 20 т. н., по меньшей мере 30 т. н., по меньшей мере 40 т. н., по меньшей мере 50 т. н., по меньшей мере 60 т. н., по меньшей мере 70 т. н., по меньшей мере 80 т. н., по меньшей мере 90 т. н., по меньшей мере 100 т. н., по меньшей мере 150 т. н., по меньшей мере 200 т. н., по меньшей мере 250 т. н., по меньшей мере 300 т. н., по меньшей мере 350 т. н., по меньшей мере 400 т. н., по меньшей мере 450 т. н. или по меньшей мере 500 т. н. или больше.
[00199] В еще одних случаях размер ДНК-вставки и/или области эндогенного локуса, подвергаемого изменению, делеции, нацеливанию, модифицированию, конструированию и т.д., составляет по меньшей мере 100, 200, 300, 400, 500, 600, 700, 800 или 900 нуклеотидов, или по меньшей мере 1 т. н., 2 т. н., 3 т. н., 4 т. н., 5 т. н., 6 т. н., 7 т. н., 8 т. н., 9 т. н., 10 т. н., 11 т. н., 12 т. н., 13 т. н., 14 т. н., 15 т. н., 16 т. н., 17 т. н., 18 т. н., 19 т. н., 20 т. н. или больше. В некоторых вариантах осуществления размер ДНК-вставки и/или области эндогенного локуса, подвергаемого изменению, делеции, нацеливанию, модифицированию, конструированию и т.д., составляет от нуклеотидов до 20 т. н., от 200 нуклеотидов до 20 т. н., от 300 нуклеотидов до 20 т. н., от 400 нуклеотидов до 20 т. н., от 500 нуклеотидов до 20 т. н., от 600 нуклеотидов до 20 т. н., от 700 нуклеотидов до 20 т. н., от 800 нуклеотидов до 20 т. н., от 900 нуклеотидов до 20 т. н., от 1 т. н. до 20 т. н., от 2 т. н. до 20 т. н., от 3 т. н. до 20 т. н., от 4 т. н. до 20 т. н., от 5 т. н. до 20 т. н., от 6 т. н. до 20 т. н., от 7 т. н. до 20 т. н., от 8 т. н. до 20 т. н., от 9 т. н. до 20 т. н., от 10 т. н. до 20 т. н., от 11 т. н. до 20 т. н., от 12 т. н. до 20 т. н., от 13 т. н. до 20 т. н., от 14 т. н. до 20 т. н., от 15 т. н. до 20 т. н., от 16 т. н. до 20 т. н., от 17 т. н. до 20 т. н., от 18 т. н. до 20 т. н. или от 19 т. н. до 20 т. н. В некоторых вариантах осуществления размер ДНК-вставки и/или области эндогенного локуса, подвергаемого изменению, делеции, нацеливанию, модифицированию, конструированию и т.д., составляет от 100 нуклеотидов до 19 т. н., от 100 нуклеотидов до 18 т. н., от 100 нуклеотидов до 17 т. н., от 100 нуклеотидов до 16 т. н., от 100 нуклеотидов до 15 т. н., от 100 нуклеотидов до 14 т. н., от 100 нуклеотидов до 13 т. н., от 100 нуклеотидов до 12 т. н., от 100 нуклеотидов до 11 т. н., от 100 нуклеотидов до 10 т. н., от 100 нуклеотидов до 9 т. н., от 100 нуклеотидов до 8 т. н., от 100 нуклеотидов до 7 т. н., от 100 нуклеотидов до 6 т. н., от 100 нуклеотидов до 5 т. н., от 100 нуклеотидов до 4 т. н., от 100 нуклеотидов до 3 т. н., от 100 нуклеотидов до 2 т. н., от 100 нуклеотидов до 1 т. н., от 100 нуклеотидов до 900 нуклеотидов, от 100 нуклеотидов до 800 нуклеотидов, от 100 нуклеотидов до 700 нуклеотидов, от 100 нуклеотидов до 600 нуклеотидов, от 100 нуклеотидов до 500 нуклеотидов, от 100 нуклеотидов до 400 нуклеотидов, от 100 нуклеотидов до 300 нуклеотидов или от 100 нуклеотидов до 200 нуклеотидов. В некоторых вариантах осуществления размер ДНК-вставки и/или области эндогенного локуса, подвергаемого изменению, делеции, нацеливанию, модифицированию, конструированию и т.д., составляет от 200 нуклеотидов до 19 т. н., от 300 нуклеотидов до 18 т. н., от 400 нуклеотидов до 17 т. н., от 500 нуклеотидов до 16 т. н., от 600 нуклеотидов до 15 т. н., от 700 нуклеотидов до 14 т. н., от 800 нуклеотидов до 13 т. н., от 900 нуклеотидов до 12 т. н., от 1 т. н. до 11 т. н., от 2 т. н. до 10 т. н., от 3 т. н. до 9 т. н., от 4 т. н. до 8 т. н., от 5 т. н. до 7 т. н. или от 5 т. н. до 6 т. н.
Предусмотренные отличные от человека животные
[00200] В определенных аспектах предусмотрены отличные от человека животные, которые экспрессируют антитела, содержащие легкие цепи, которые содержат полную или неполную последовательность человеческой легкой λ-цепи Ig, в результате интеграции генетического материала, который соответствует по меньшей мере части человеческого локуса легкой λ-цепи Ig и который кодирует по меньшей мере человеческий Vλ-домен (т.е. перегруппированную последовательность человеческих Vλ-Jλ), вместо соответствующих последовательностей отличных от человеческих легких λ-цепей Ig в геноме зародышевого типа отличного от человека животного. Подходящие примеры, описанные в данном документе, включают без ограничения грызунов, в частности крыс или мышей.
[00201] Последовательность человеческой легкой λ-цепи Ig, в некоторых вариантах осуществления, содержит генетический материал из человеческого локуса легкой λ-цепи Ig, где последовательность человеческой легкой λ-цепи Ig кодирует легкую цепь иммуноглобулина, которая содержит кодируемую часть генетического материала из человеческого локуса легкой λ-цепи Ig. В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig, описанная в данном документе, содержит по меньшей мере один сегмент человеческого гена Vλ, и по меньшей мере один человеческий генный сегмент Jλ, и одну или более последовательностей, необходимых для обеспечения перегруппировки (например, последовательность[-и] сигнала рекомбинации) по меньшей мере одного указанного сегмента человеческого гена Vλ с по меньшей мере одним указанным сегментом человеческого гена Jλ с образованием функциональной перегруппированной последовательности человеческих Vλ-Jλ, которая кодирует человеческий Vλ-домен. Во многих вариантах осуществления последовательность человеческой легкой λ-цепи Ig содержит множество сегментов человеческого гена Vλ и одну или более последовательностей, необходимых для обеспечения перегруппировки указанных сегментов человеческого гена Vλ с по меньшей мере одним сегментом человеческого гена Jλ. Во многих вариантах осуществления последовательность человеческой легкой λ-цепи Ig, описанная в данном документе, представляет собой геномную последовательность человеческого локуса легкой λ-цепи Ig (например, выделенная и/или клонированная из бактериальной искусственной хромосомы) и содержит множество сегментов человеческого гена Vλ в конфигурации зародышевого типа. В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig содержит последовательности человеческих Vλ, Jλи Cλ в конфигурации зародышевого типа (т.е., как указано, последовательности человеческих Vλ, Jλи Cλ находятся в локусе легкой λ-цепи Ig в клетке человека). В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig представляет собой или содержит человеческую последовательность, которая изображена в графических материалах (например, см. фигуры 1-4). В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig кодирует полный или неполный полипептид легкой λ-цепи Ig, при этом полипептид легкой λ-цепи Ig находится в иммуноглобулине, в частности иммуноглобулине, который экспрессируется B-клеткой человека. Также предусмотрены отличные от человека животные, отличные от человеческих эмбрионы, клетки и нацеливающие конструкции для получения отличных от человека животных, отличных от человеческих эмбрионов и клеток, содержащих указанную последовательность человеческой легкой λ-цепи Ig вместо соответствующей последовательности отличной от человеческой легкой λ-цепи Ig (например, эндогенный локус легкой λ-цепи Ig грызуна).
[00202] В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig вставлена вместо соответствующей последовательности отличной от человеческой легкой λ-цепи Ig в геном зародышевого типа отличного от человека животного. В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig вставлена против хода транскрипции относительно последовательности отличной от человеческой легкой λ-цепи Ig (например, последовательность отличной от человеческой константной области легкой λ-цепи Ig). В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig вставлена среди одной или более последовательностей отличных от человеческих легких λ-цепей Ig, таким образом, что указанная последовательность человеческой легкой λ-цепи Ig расположена рядом с последовательностями отличных от человеческих легких λ-цепей Ig, (например, см. фигуры 1, 2, 3 и/или 4).
[00203] В некоторых вариантах осуществления одна или более последовательностей (или их часть) отличной от человеческой легкой λ-цепи Ig не удалены из отличного от человеческого локуса легкой λ-цепи Ig. В некоторых вариантах осуществления одна или более последовательностей отличных от человеческих легких λ-цепей Ig (например, Vλ, Jλ и/или Cλ) из отличного от человеческого локуса легкой λ-цепи Ig подвержены изменению, замещению, разрушению, делеции или замене с помощью, среди прочего, последовательности человеческой легкой λ-цепи Ig, описанной в данном документе (например, последовательность, которая содержит один или более сегментов человеческого гена Vλ, один или более сегментов человеческого гена Jλ , один или более сегментов человеческого гена Cλ или их комбинации), функционально связанной с отличной от человеческой константной областью легкой λ-цепи Ig и одним или более энхансерами и/или регуляторными элементами отличного от человеческого локуса легкой λ-цепи Ig. В некоторых вариантах осуществления весь или практически весь отличный от человеческого локус легкой λ-цепи Ig заменен одной или более последовательностями человеческой легкой λ-цепи Ig (которые описаны в данном документе), которые функционально связаны с отличной от человеческой константной областью легкой λ-цепи Ig и одним или более энхансерами отличной от человеческой легкой λ-цепи Ig и/или регуляторными элементами отличного от человеческого локуса легкой λ-цепи Ig. В некоторых определенных вариантах осуществления один или более отличных от человеческих генов константных областей легких λ-цепей Ig не подвержены делеции или замене у отличного от человека животного, которое содержит последовательность человеческой легкой λ-цепи Ig, описанную в данном документе. В качестве лишь одного неограничивающего примера, в случае вставки последовательности человеческой легкой λ-цепи Ig, которая вставлена в отличный от человеческого локус легкой λ-цепи Ig, указанная вставка осуществлена таким образом, чтобы сохранять целостность последовательностей отличных от человеческих легких λ-цепей Ig около места вставки (например, отличная от человеческой константная область легкой λ-цепи Ig и/или энхансерная область или последовательность отличной от человеческой легкой λ-цепи Ig). Таким образом, такие отличные от человека животные имеют константную область легкой λ-цепи Ig дикого типа. В некоторых вариантах осуществления отличный от человеческого локус легкой λ-цепи Ig, который подвержен изменению, замещению, разрушению, делеции, замене или конструированию с помощью одной или более последовательностей человеческих легких λ-цепей Ig, описанных в данном документе, представляет собой мышиный (например, мышь или крыса) локус легкой λ-цепи Ig. В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig вставлена в одну копию (т.е. аллель) отличного от человеческого локуса легкой λ-цепи Ig из двух копий указанного отличного от человеческого локуса легкой λ-цепи Ig с получением отличного от человека животного, которое является гетерозиготным по отношению к последовательности человеческой легкой λ-цепи Ig. В некоторых вариантах осуществления предусмотрено отличное от человека животное, которое является гомозиготным в отношении локуса легкой λ-цепи Ig, который содержит последовательность человеческой легкой λ-цепи Ig, описанную в данном документе.
[00204] В некоторых вариантах осуществления сконструированный отличный от человеческого локус легкой λ-цепи Ig, описанный в данном документе, содержит сегменты человеческих генов Vλ, Jλ и Cλ, функционально связанные с отличной от человеческой константной областью легкой λ-цепи Ig и одним или более энхансерами и/или регуляторными элементами отличной от человеческой легкой λ-цепи Ig. В некоторых вариантах осуществления сконструированный отличный от человеческого локус легкой λ-цепи Ig, описанный в данном документе, содержит сегменты человеческих генов Vλ, Jλ и Cλ, функционально связанные с отличной от человеческой константной областью легкой λ-цепи Ig, одним или более энхансерами и/или регуляторными элементами отличной от человеческой легкой λ-цепи Ig и одним или более энхансерами и/или регуляторными элементами человеческой легкой λ-цепи Ig.
[00205] В некоторых вариантах осуществления отличное от человека животное содержит сконструированный локус легкой λ-цепи Ig, описанный в данном документе, который произвольным образом интегрирован в его геном (например, в виде части произвольным образом интегрированной последовательности человеческой легкой λ-цепи Ig). Таким образом, такие отличные от человека животные могут быть описаны как имеющие трансген человеческой легкой λ-цепи Ig, содержащий множество сегментов человеческих генов Vλ, Jλ и/или Cλ, сконфигурированных таким образом, чтобы указанные сегменты человеческих генов Vλ, Jλ и/или Cλ были способны к перегруппировке и кодированию полной или неполной легкой λ-цепи Ig из антитела в экспрессируемом репертуаре отличного от человека животного. Сконструированный локус или трансген легкой λ-цепи Ig, описанный в данном документе, можно выявить с применением разнообразных способов, в том числе, например, ПЦР, вестерн-блоттинга, саузерн-блоттинга, определения полиморфизма длин рестрикционных фрагментов (RFLP) или анализа приобретения или потери аллеля. В некоторых вариантах осуществления описанное в данном документе отличное от человека животное является гетерозиготным в отношении сконструированного локуса легкой λ-цепи Ig, описанного в данном документе. В некоторых вариантах осуществления описанное в данном документе отличное от человека животное является гемизиготным в отношении сконструированного локуса легкой λ-цепи Ig, описанного в данном документе. В некоторых вариантах осуществления описанное в данном документе отличное от человека животное содержит одну или более копий сконструированного локуса или трансгена легкой λ-цепи Ig, описанного в данном документе. В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, содержит локус легкой λ-цепи Ig, изображенный в графических материалах (например, см. фигуры 1, 2, 3 и/или 4).
[00206] В некоторых вариантах осуществления предусмотрены композиции и способы для получения отличных от человека животных, геном зародышевого типа которых содержит сконструированный локус легкой λ-цепи Ig, который содержит одну или более последовательностей человеческих легких λ-цепей Ig (например, сегменты человеческих генов Vλ, Jλ и/или Cλ) вместо последовательностей отличных от человеческих легких λ-цепей Ig, в том числе кодирующие человеческие легкие λ-цепи Ig последовательности, которые включают конкретные полиморфные формы сегментов человеческих Vλ, Jλ и/или Cλ (например, конкретные аллели или варианты V и/или J), в том числе композиции и способы для получения отличных от человека животных, которые экспрессируют антитела, содержащие легкие λ-цепи Ig, которые содержат человеческие вариабельные домены и человеческие или отличные от человеческих константные домены, собранные из локуса легкой λ-цепи Ig, который содержит сегменты человеческих Vλ, Jλ и Cλ, функционально связанные с отличной от человеческой константной областью легкой λ-цепи Ig. В некоторых вариантах осуществления также представлены композиции и способы для получения отличных от человека животных, которые экспрессируют такие антитела под контролем эндогенного(-ых) энхансера(ов) и/или эндогенной(-ых) регуляторной(-ых) последовательности(ей). В некоторых вариантах осуществления также представлены композиции и способы для получения отличных от человека животных, которые экспрессируют такие антитела под контролем гетерологичного(-ых) энхансера(ов) и/или гетерологичной(-ых) регуляторной(-ых) последовательности(ей).
[00207] В определенных вариантах осуществления способы, описанные в данном документе, включают осуществление вставки последовательности, кодирующей полную или неполную человеческую легкую λ-цепь Ig, против хода транскрипции относительно отличной от человеческой константной области легкой λ-цепи Ig (например, мышиная Cλ-область), таким образом, что экспрессируется антитело, при этом антитело характеризуется наличием легкой цепи, которая содержит по меньшей мере человеческий Vλ-домен и, в некоторых вариантах осуществления, человеческие Vλ- и Cλ-домены, и экспрессируется как на поверхности B-клеток, так и в сыворотке крови отличного от человека животного.
[00208] В некоторых вариантах осуществления способы включают осуществление серии вставок генетического материала, соответствующего человеческому локусу легкой λ-цепи Ig. В некоторых вариантах осуществления генетический материал, соответствующий человеческому локусу легкой λ-цепи Ig, может быть синтетическим или геномным (например, клонированным из бактериальной искусственной хромосомы). В некоторых вариантах осуществления генетический материал, соответствующий человеческому локусу легкой λ-цепи Ig, может быть сконструирован на основе опубликованных источников и/или бактериальных искусственных хромосом таким образом, что указанный генетический материал содержит сегменты человеческих Vλ, Jλ и/или Cλ в ориентации, отличной от той, в которой они находятся в человеческом локусе легкой λ-цепи Ig, тем не менее указанный генетический материал все еще содержит последовательности для обеспечения перегруппировки указанных сегментов человеческих Vλ, Jλ и/или Cλ с кодированием функциональной легкой λ-цепи Ig. В качестве лишь одного примера, генетический материал, соответствующий человеческому локусу легкой λ-цепи Ig, может быть спроектирован с применением руководства, предусмотренного в данном документе для построения последовательности человеческой легкой λ-цепи Ig, которая содержит сегменты человеческих Vλ, Jλ и/или Cλ в порядке и/или группировке, которые отличаются от таковых, имеющих место в человеческом локусе легкой λ-цепи Ig клетки человека. В таком примере содержание сегментов человеческих Vλ, Jλ и/или Cλ будет эквивалентным соответствующим сегментам в клетке человека, однако, порядок и группировка будут отличными. При построении человеческого локуса легкой λ-цепи Ig для создания отличного от человека животного, описанного в данном документе, необходимые последовательности сигнала рекомбинации могут быть сконфигурированы таким образом, что сегменты, принадлежащие организму человека, могут правильно перегруппироваться и образовывать функциональную легкую λ-цепь Ig. Руководство для конфигурации зародышевого типа сегментов и последовательностей человеческих легких λ-цепей Ig, необходимых для надлежащей рекомбинации, можно найти в Molecular Biology of B Cells, London: Elsevier Academic Press, 2004, Ed. Honjo, T., Alt, F.W., Neuberger, M. Chapters 4 (pp.37-59) and 5 (61-82); включенных в данный документ с помощью ссылки в полном объеме.
[00209] В некоторых вариантах осуществления осуществление серии вставок предусматривает осуществление множества вставок частей гетерологичного генетического материала в один клон ES-клетки. В некоторых вариантах осуществления осуществление серии вставок предусматривает осуществление последовательных вставок частей гетерологичного генетического материала в успешные клоны ES-клетки.
[00210] В некоторых вариантах осуществления способы включают осуществление вставки приблизительно 11822 п. о. ДНК по ходу транскрипции относительно мышиной (например, мышь или крыса) Cλ1-области таким образом, что указанная ДНК функционально связана с указанной мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК включает одну или более энхансерных областей (или последовательностей) человеческой легкой λ-цепи Ig. В некоторых определенных вариантах осуществления способы включают осуществление вставки приблизительно 11822 п. о. ДНК, которая содержит три энхансерные области (или последовательности) человеческой легкой λ-цепи Ig, при этом указанные три энхансерные области (или последовательности) человеческой легкой λ-цепи Ig вставлены по ходу транскрипции относительно указанной мышиной (например, мышь или крыса) Cλ1-области.
[00211] В некоторых вариантах осуществления способы включают осуществление вставки приблизительно 125473 п. о. ДНК против хода транскрипции относительно мышиной (например, мышь или крыса) Cλ1-области таким образом, что указанная ДНК функционально связана с указанной мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ3-10, Vλ3-9, Vλ2-8, Vλ4-3, Vλ3-1, пары сегментов человеческих Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, и сегмент человеческого гена Jλ, Jλ7. В некоторых определенных вариантах осуществления способы включают осуществление вставки приблизительно 11822 п. о. ДНК, которая содержит одну или более энхансерных областей (или последовательностей) человеческой легкой λ-цепи Ig, при этом одна или более энхансерных областей (или последовательностей) человеческой легкой λ-цепи Ig вставлены по ходу транскрипции относительно указанной мышиной (например, мышь или крыса) Cλ1-области.
[00212] В некоторых вариантах осуществления способы включают осуществление вставки приблизительно 171458 п. о. ДНК против хода транскрипции относительно мышиной (например, мышь или крыса) Cλ1-области таким образом, что указанная ДНК функционально связана с указанной мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ2-11, Vλ3-12, Vλ2-14, Vλ3-16, Vλ3-19, Vλ3-21, Vλ3-22, Vλ2-23, Vλ3-25 и Vλ3-27. В некоторых определенных вариантах осуществления способы включают осуществление вставки приблизительно 171458 п. о. ДНК против хода транскрипции относительно человеческого генного сегмента Vλ3-10, который функционально связан с мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ , Vλ2-11, Vλ3-12, Vλ2-14, Vλ3-16, Vλ3-19, Vλ3-21, Vλ3-22, Vλ2-23, Vλ3-25 и Vλ3-27.
[00213] В некоторых вариантах осуществления способы включают осуществление вставки приблизительно 121188 п. о. ДНК против хода транскрипции относительно мышиной (например, мышь или крыса) Cλ1-области таким образом, что указанная ДНК функционально связана с указанной мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ3-27, Vλ1-36, Vλ5-37, Vλ5-39, Vλ1-40, Vλ7-43, Vλ1-44, Vλ5-45, Vλ7-46, Vλ1-47, Vλ9-49, Vλ1-51 и Vλ5-52. В некоторых определенных вариантах осуществления способы включают осуществление вставки приблизительно 121188 п. о. ДНК против хода транскрипции относительно человеческого генного сегмента Vλ3-27, который функционально связан с мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ3-27, Vλ1-36, Vλ5-37, Vλ5-39, Vλ1-40, Vλ7-43, Vλ1-44, Vλ5-45, Vλ7-46, Vλ1-47, Vλ9-49, Vλ1-51 и Vλ5-52.
[00214] В некоторых вариантах осуществления способы включают осуществление вставки приблизительно 121188 п. о. ДНК против хода транскрипции относительно мышиной (например, мышь или крыса) Cλ1-области таким образом, что указанная ДНК функционально связана с указанной мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ3-27, Vλ1-36, Vλ5-37, Vλ5-39, Vλ1-40, Vλ7-43, Vλ1-44, Vλ5-45, Vλ7-46, Vλ1-47, Vλ9-49, Vλ1-51 и Vλ5-52, и при этом ДНК содержит плечо гомологии, которое содержит последовательность, расположенную в 5'-направлении относительно генного сегмента Vλ2 мыши. В некоторых определенных вариантах осуществления способы включают осуществление вставки приблизительно 121188 п. о. ДНК против хода транскрипции относительно человеческого генного сегмента Vλ3-27, который функционально связан с мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК включает сегменты человеческого гена Vλ, Vλ3-27, Vλ1-36, Vλ5-37, Vλ5-39, Vλ1-40, Vλ7-43, Vλ1-44, Vλ5-45, Vλ7-46, Vλ1-47, Vλ9-49, Vλ1-51 и Vλ5-52, и при этом ДНК содержит плечо гомологии, которое содержит последовательность мыши, расположенную в 5'-направлении относительно генного сегмента Vλ2 мыши, для управления делецией геномной последовательности λ-цепи Ig мыши (например, локуса легкой λ-цепи Ig) после гомологичной рекомбинации с указанным фрагментом ДНК.
[00215] Вставки дополнительных сегментов человеческих генов Vλ, Jλ и/или Cλ можно достигать с применением способов, описанных в данном документе, для дополнительного расширения расхождения сконструированного локуса легкой λ-цепи Ig. Например, в некоторых вариантах осуществления способы могут включать осуществление вставки приблизительно 300 т. н. ДНК против хода транскрипции относительно мышиной (например, мышь или крыса) Cλ1-области таким образом, что указанная ДНК функционально связана с указанной мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ10-54, Vλ6-57, Vλ4-60, Vλ8-61 и Vλ4-69. В таких вариантах осуществления указанная ДНК вставлена против хода транскрипции относительно человеческого генного сегмента Vλ5-52, который функционально связан с мышиной (например, мышь или крыса) Cλ1-областью, при этом ДНК содержит сегменты человеческого гена Vλ, Vλ10-54, Vλ6-57, Vλ4-60, Vλ8-61 и Vλ4-69. В некоторых определенных вариантах осуществления указанная ДНК содержит человеческий ген VpreB. Дополнительные сегменты человеческого Vλ, описанные выше, можно клонировать непосредственно из коммерчески доступных клонов BAC и группировать в меньшем фрагменте ДНК с применением рекомбинантных методик, описанных в данном документе или иным образом известных из уровня техники. В качестве альтернативы, дополнительные сегменты человеческого гена Vλ, описанные выше, можно синтезировать во фрагмент ДНК и добавлять в сконструированный локус легкой λ-цепи Ig, описанный выше. Сходным образом дополнительные сегменты человеческих генов Jλ и/или Cλ можно получать из коммерчески доступных клонов BAC или синтезировать непосредственно из опубликованных последовательностей. Также эндогенные энхансерные области (или последовательности) легкой λ-цепи Ig могут быть удалены из сконструированного локуса легкой λ-цепи Ig, описанного в данном документе. Иллюстративное изображение, на котором показан сконструированный локус легкой λ-цепи Ig у отличных от человека животных, описанных в данном документе, представлено на любой из фигур 1, 2, 3 и 4.
[00216] При необходимости последовательность человеческой легкой λ-цепи Ig (т.е. последовательность, содержащая сегменты человеческих генов Vλ, Jλ и/или Cλ), кодирующая полную или неполную легкую λ-цепь Ig, может быть отдельно модифицирована для включения кодонов, оптимизированных для экспрессии у отличного от человека животного (например, см. патенты США №№5670356 и 5874304). Кодон-оптимизированные последовательности представляют собой синтетические последовательности и предпочтительно кодируют полипептид (или биологически активный фрагмент полноразмерного полипептида, обладающий фактически такой же активностью, как и полноразмерный полипептид), идентичный кодируемому исходным полинуклеотидом, не являющимся кодон- оптимизированным. В некоторых вариантах осуществления последовательность человеческой легкой λ-цепи Ig, кодирующая полную или неполную легкую λ-цепь Ig, может отдельно включать измененную последовательность для оптимизации использования кодонов в отношении конкретного клеточного типа (например, клетки грызуна). Например, кодоны каждой нуклеотидной последовательности, которая должна быть встроена в геном отличного от человека животного (например, грызуна), можно оптимизировать для экспрессии в клетке отличного от человека животного. Такая последовательность может быть описана как кодон-оптимизированная последовательность.
[00217] В некоторых вариантах осуществления осуществление вставки нуклеотидной последовательности, кодирующей полную или неполную человеческую легкую λ-цепь Ig, предусматривает минимальную модификацию генома зародышевого типа отличного от человека животного, описанного в данном документе, и приводит к экспрессии антител, содержащих легкие цепи, которые являются полностью или частично человеческими. Способы создания сконструированных отличных от человека животных, в том числе с помощью нокаутов и нокинов, известны из уровня техники (см., например, Gene Targeting: A Practical Approach, Joyner, ed., Oxford University Press, Inc. 2000). Например, создание трансгенных грызунов необязательно может подразумевать разрушение генетических локусов одного или более эндогенных генов (или генных сегментов) грызуна и введение одного или более гетерологичных генов (или генных сегментов, или нуклеотидных последовательностей) в геном грызуна, в некоторых вариантах осуществления, в то же местоположение, что и эндогенный ген (или генные сегменты) грызуна. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая полную или неполную человеческую легкую λ-цепь Ig, введена против хода транскрипции относительно мышиного (например, мышь или крыса) гена константной области легкой λ-цепи Ig произвольным образом вставленного трансгена легкой λ-цепи Ig в геном зародышевого типа грызуна. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая полную или неполную человеческую легкую λ-цепь Ig, введена против хода транскрипции относительно мышиного (например, мышь или крыса) гена константной области легкой λ-цепи Ig эндогенного локуса легкой λ-цепи Ig в геном зародышевого типа грызуна; в некоторых определенных вариантах осуществления эндогенный локус легкой λ-цепи Ig изменен, модифицирован или сконструирован для содержания сегментов человеческих генов λ-цепи Ig (например, Vλ, Jλ и/или Cλ), функционально связанных с Cλ1-областью грызуна.
[00218] Схематическое изображение (без соблюдения масштаба) иллюстративных сконструированных локусов легких λ-цепей Ig представлено на фигурах 1-4. В частности, на фигурах 1 и 3 представлены иллюстративные стратегии для построения сконструированных локусов легких λ-цепей Ig, характеризующихся вставкой нуклеотидных последовательностей, содержащих множество сегментов человеческих Vλ, Jλ и Cλ. Как проиллюстрировано на фигуре 1, фрагмент ДНК, содержащий человеческую Eλ-последовательность (или -область), вставлен по ходу транскрипции относительно Cλ-области грызуна с помощью гомологичной рекомбинации. Данный фрагмент ДНК содержит кассету для отбора по устойчивости к неомицину (например, ген устойчивости к неомицину [NEOR] фланкированный участками распознавания рекомбинации loxP), расположенную в 3'-направлении относительно человеческой Eλ-последовательности, которая содержит три элемента человеческого Eλ, сконструированные по ходу транскрипции (или в 3'-направлении) относительно Cλ1-области грызуна. Также на фигуре 1 проиллюстрирован фрагмент ДНК, содержащий первую часть сегментов человеческого Vλ, совокупность пар сегментов человеческих Jλ-Cλ (например, человеческие Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cl3, Jλ6-Cλ6) и сегмент Jλ7, принадлежащий организму человека, вставленный против хода транскрипции относительно Cλ1-области грызуна с помощью гомологичной рекомбинации. Как проиллюстрировано, кассета для отбора по устойчивости к гигромицину (например, ген устойчивости к гигромицину [HYGR] фланкированный участками распознавания рекомбинации Frt) расположена на 5'-конце нацеливающего вектора и против хода транскрипции относительно последовательности человеческой легкой λ-цепи Ig, содержащейся в нацеливающем векторе. Кассету для отбора по устойчивости к гигромицину удаляют с помощью гомологичной рекомбинации с последующими нацеливающими векторами, описанными в разделе примеров ниже. Затем нацеливающий вектор вводят с помощью электропорации в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном зародышевого типа которого содержит сконструированный локус легкой λ-цепи Ig. Как только положительность клона ES- клетки грызуна подтверждена, вводят с помощью электропорации последовательным образом другие изображенные нацеливающие векторы и осуществляют подтверждение на каждой стадии для завершения построения сконструированного локуса легкой λ-цепи Ig (см. фигуру 2). Конечный нацеливающий вектор может быть спроектирован с плечом гомологии (нацеливающий вектор 6680) или без него (нацеливающий вектор 6597), которое обеспечивает направление делеции сегментов эндогенной легкой λ-цепи Ig с помощью гомологичной рекомбинации с получением в результате двух потенциальных сконструированных аллелей легких λ-цепей Ig (фигура 2). Кроме того, любую оставшуюся кассету для отбора можно удалить при необходимости с помощью опосредованной рекомбиназами делеции. Альтернативная стратегия для осуществления вставки дополнительных сегментов человеческого гена Vλ в сконструированный локус легкой λ-цепи Ig с применением направляющих РНК (gRNA) представлена на фигуре 3.
[00219] Как только последовательность человеческой легкой λ-цепи Ig вставлена против хода транскрипции относительно отличной от человеческой константной области легкой λ-цепи Ig из клона BAC, получают нацеливающий вектор для интеграции в локус легкой λ-цепи Ig. Клон BAC, подлежащий нацеливанию с помощью последовательности человеческой легкой λ-цепи Ig для получения нацеливающего вектора, может содержать 5'- и/или 3'-фланкирующую геномную ДНК мышиного происхождения (например, мышь или крыса). В качестве альтернативы или дополнительно, клон BAC, подлежащий нацеливанию с помощью последовательности человеческой легкой λ-цепи Ig для получения нацеливающего вектора, может содержать 5'- и/или 3'-фланкирующую геномную ДНК человеческого происхождения, таким образом, что образуется область перекрывания последовательности человеческой легкой λ-цепи Ig. Таким образом, обеспечивается возможность успешного нацеливания нескольких сконструированных клонов BAC (например, см. фигуру 1). Конечные нацеливающие векторы вводят в локус легкой λ-цепи Ig в геном отличной от человеческой клетки (например, эмбриональная стволовая клетка грызуна). В некоторых вариантах осуществления нацеливающие векторы, описанные в данном документе, вводят в локус легкой λ-цепи Ig в геном зародышевого типа отличной от человеческой клетки, которая дополнительно содержит геномную ДНК человеческих VH, DH и JH (например, содержащая множество сегментов человеческих генов VH, DH и JH), функционально связанную с одним или более генов константной области H-цепи Ig, и/или геномную ДНК человеческих Vκ и Jκ (например, содержащая множество сегментов человеческих генов Vκи Jκ), функционально связанную с геном константной области κ-цепи Ig (например, см. патенты США №№8502018, 8642835, 8697940 и 8791323, включенные в данный документ с помощью ссылки в полном объеме).
[00220] Нацеливающий вектор вводят в эмбриональные стволовые клетки грызуна (например, мышь) с помощью электропорации таким образом, что последовательность, содержащаяся в нацеливающем векторе, вставляется в геном эмбриональных стволовых клеток грызуна, и что приводит к получению способности отличной от человеческой клетки или отличного от человека животного (например, мышь), обеспечивающей экспрессию антител с полными или неполными человеческими легкими λ-цепями Ig. Как описано в данном документе, создают трансгенного грызуна, в котором в зародышевого типа генома грызуна был получен сконструированный локус легкой λ-цепи Ig (например, эндогенный локус легкой λ-цепи Ig, содержащий последовательность человеческой легкой λ-цепи Ig, функционально связанную с эндогенной Cλ-областью грызуна, описанной в данном документе). Антитела экспрессируются на поверхности B-клеток грызуна и в сыворотке крови указанного грызуна, при этом антитела характеризуются легкими цепями с человеческими Vλ-доменами и, в некоторых вариантах осуществления, человеческими Vλ- и Cλ-доменами. Если эндогенный локус легкой λ-цепи Ig в зародышевого типа генома грызуна не подлежал нацеливанию с помощью нацеливающего вектора, то сконструированный локус легкой λ-цепи Ig предпочтительно вставлен в месторасположение, отличное от такового эндогенного локуса легкой λ-цепи Ig грызуна (например, произвольным образом вставленный трансген).
[00221] Получение сконструированного локуса легкой λ-цепи Ig в отличном от человека животном, описанном выше, обеспечивает получение сконструированной линии грызунов, которая продуцируют антитела, которые содержат легкие λ-цепи Ig, экспрессированные на основе такого сконструированного локуса легкой λ-цепи Ig с человеческим Vλ-доменом, и в некоторых вариантах осуществления человеческими Vλ- и Cλ-доменами. При эффективном использовании присутствия сконструированного локуса H-цепи Ig, который содержит множество сегментов человеческих генов VH, DH и JH, функционально связанных с генами константной области H-цепи Ig, получена сконструированная линия грызунов, которая продуцируют антитела и компоненты антител для разработки терапевтических средств на основе человеческих антител. Таким образом, на практике получена единственная сконструированная линия грызунов, обладающая возможностью обеспечить альтернативную систему in vivo для использования человеческих Vλ-доменов для разработки новых лекарственных препаратов на основе антител для лечения заболевания человека.
[00222] В некоторых вариантах осуществления в геноме отличного от человека животного, описанного в данном документе, дополнительно содержатся (например, с помощью стратегий кроссбридинга или множественного генного нацеливания) одна или более человеческих вариабельных областей тяжелой и/или легкой цепей иммуноглобулинов, как описано в патентах США №№8502018, 8642835, 8697940 и 8791323; все из которых включены в данный документ с помощью ссылки в полном объеме. В качестве альтернативы, сконструированный локус легкой λ-цепи Ig, описанный в данном документе, может быть встроен в эмбриональную стволовую клетку, содержащую гуманизированные локусы H-цепи Ig и/или κ-цепи Ig, или отличное от человека животное, содержащее сконструированный локус легкой λ-цепи Ig, описанный в данном документе, можно скрещивать с другим отличным от человека животным, содержащим гуманизированные локусы H-цепи Ig и/или κ-цепи Ig. Известны различные такие животные, содержащие гуманизированные локусы H- цепи Ig и/или κ-цепи Ig, например линия VELOCIMMUNE® (см., например, патенты США №№8502018 и/или 8642835; включенные в данный документ с помощью ссылки в полном объеме), линия XENOMOUSE™ (см., например, Mendez, M.J. et al., 1997, Nat. Genetics 15(2):146-56, и Jakobovits, A. et al., 1995, Ann. NY Acad. Sci. 764:525-35). Гомозиготность сконструированного локуса легкой λ-цепи Ig, описанного в данном документе, может быть впоследствии достигнута путем скрещивания. В качестве альтернативы, в случае произвольным образом вставленного сконструированного трансгена легкой λ-цепи Ig (описанного выше), линии грызунов могут быть выбраны на основании, среди прочего, уровня экспрессии человеческих Vλ-доменов из трансгена.
[00223] В качестве альтернативы и/или дополнительно, в некоторых вариантах осуществления в геноме зародышевого типа отличного от человека животного, описанного в данном документе, дополнительно содержится удаленный, инактивированный, функционально подавленный или иным образом нефункциональный эндогенный локус легкой κ-цепи Ig. Генетические модификации для удаления или лишения функциональности гена или генетического локуса могут быть достигнуты с применением способов, описанных в данном документе, и/или способов, известных в уровне техники.
[00224] Трансгенное отличное от человека животное-основоположника можно идентифицировать на основании присутствия сконструированного локуса легкой λ-цепи Ig в его геноме зародышевого типа и/или экспрессии антител с полной или неполной последовательностью человеческой легкой λ-цепи Ig в тканях или клетках отличного от человека животного. Затем трансгенное отличное от человека животное- основоположника можно использовать для разведения дополнительных отличных от человека животных, несущих сконструированный локус легкой λ-цепи Ig, получая тем самым когорту отличных от человека животных, каждое из которых несет одну или более копий сконструированного локуса легкой λ-цепи Ig. Более того, трансгенных отличных от человека животных, несущих сконструированный локус легкой λ-цепи Ig, описанный в данном документе, при необходимости можно дополнительно скрещивать с другими трансгенными отличными от человека животными, несущими другие трансгены (например, человеческие гены иммуноглобулинов).
[00225] В некоторых вариантах осуществления также можно получать трансгенных отличных от человека животных так, чтобы они содержали выбранные системы, которые обеспечивают контролируемую, управляемую, индуцируемую и/или специфическую для типа клеток экспрессию трансгена или интегрированной(-ых) последовательности(-ей). Например, отличные от человека животные описанные в данном документе, могут быть сконструированы с содержанием последовательности, кодирующей полную или неполную человеческую легкую λ-цепь Ig из антитела, которое экспрессируется при определенных условиях (например, кратко описано в Rajewski, K. et al., 1996, J. Clin. Invest. 98(3):600-3). Иллюстративные системы предусматривают систему для рекомбиназ Cre/loxP из бактериофага P1 (см., например, Lakso, M. et al., 1992, Proc. Natl. Acad. Sci. U.S.A. 89:6232-6) и систему для рекомбиназ FLP/Frt из S. cerevisiae (O’Gorman, S. et al, 1991, Science 251:1351-5). Таких животных можно получать путем создания «дважды» трансгенных животных, например, путем спаривания двух трансгенных животных, одного, содержащего трансген, содержащий выбранную модификацию (например, сконструированный локус легкой λ-цепи Ig, описанный в данном документе), и другого, содержащего трансген, кодирующий рекомбиназу (например, рекомбиназа Cre).
[00226] Отличных от человека животных, описанных в данном документе, можно получать, как описано выше, или с использованием способов, известных из уровня техники, таким образом, чтобы они содержали дополнительные человеческие, гуманизированные или иным образом сконструированные гены, зачастую в зависимости от предполагаемого применения отличного от человека животного. При необходимости, генетический материал таких человеческих, гуманизированных или иным образом сконструированных генов можно вводить посредством дальнейшего изменения генома клеток (например, эмбриональные стволовые клетки), имеющих генетические модификации или изменения, описанные выше, или с помощью методик скрещивания, известных из уровня техники, с другими генетически модифицированными или сконструированными линиями. В некоторых вариантах осуществления описанных в данном документе отличных от человека животных получают таким образом, что они дополнительно содержат трансгенные человеческие гены или сегменты генов H-цепи Ig и/или легкой κ-цепи Ig (см., например, Murphy, A.J. et al., (2014) Proc. Natl. Acad. Sci. U.S.A. 111(14):5153-5158; патент США №8502018; патент США №8642835; патент США №8697940; патент США №8791323 и публикацию заявки на выдачу патента США №2013/0096287 A1; включенные в данный документ посредством ссылки в полном объеме).
[00227] В некоторых вариантах осуществления отличных от человека животных, описанных в данном документе, можно получать путем введения нацеливающего вектора, описанного в данном документе, в клетку из модифицированной линии. В виде лишь одного примера, нацеливающий вектор, описанный выше, может быть введен в мышь VELOCIMMUNE®. Мыши VELOCIMMUNE® экспрессируют антитела, которые имеют полностью человеческие вариабельные домены и константные домены мыши. В некоторых вариантах осуществления отличных от человека животных, описанных в данном документе, получают таким образом, чтобы они дополнительно содержали человеческие гены иммуноглобулина (гены вариабельной и/или константной области). В некоторых вариантах осуществления описанные в данном документе отличные от человека животные содержат сконструированный локус легкой λ-цепи Ig, описанный в данном документе, и генетический материал от гетерологичного вида (например, люди), где генетический материал кодирует полные или неполные один или более человеческих вариабельных доменов тяжелой цепи и/или легкой κ-цепи Ig.
[00228] Например, описанные в данном документе отличные от человека животные, содержащие сконструированный локус легкой λ-цепи Ig, описанный в данном документе, могут дополнительно содержать (например, с помощью стратегий кроссбридинга или множественного генного нацеливания) одну или более модификаций, описанных в Murphy, A.J. et al., (2014) Proc. Natl. Acad. Sci. U.S.A. 111(14):5153-8; Macdonald, L.E. et al., 2014, Proc. Natl. Acad. Sci. U.S.A. 111(14):5147-52; патенты США №№8502018, 8642835, 8697940 и 8791323; все из которых включены в данный документ посредством ссылки в полном объеме. В некоторых вариантах осуществления грызуна, содержащего сконструированный локус легкой λ-цепи Ig, описанный в данном документе, скрещивают с грызуном, содержащим гуманизированный локус вариабельной области H-цепи Ig и/или легкой κ-цепи Ig (см., например, патенты США №№8502018, 8642835, 8697940 и/или 8791323; включенные в данный документ с помощью ссылки в полном объеме). В некоторых вариантах осуществления грызуна, содержащего сконструированный локус легкой λ-цепи Ig, описанный в данном документе, скрещивают с грызуном, содержащим гуманизированный локус вариабельной области H-цепи Ig (см., например, патенты США №№8502018, 8642835, 8697940 и/или 8791323; включенные в данный документ с помощью ссылки в полном объеме) и инактивированный эндогенный локус легкой κ-цепи Ig (см., например, патенты США №№9006511, 9012717, 9029628, 9035128, 9066502, 9150662 и 9163092, включенные в данный документ с помощью ссылки в полном объеме).
[00229] Хотя в данном документе активно рассматриваются варианты осуществления, в которых описано построение сконструированного локуса легкой λ-цепи Ig у мыши (т.е. мышь со сконструированным локусом легкой λ-цепи Ig, характеризующимся наличием множества сегментов человеческих генов Vλ, Jλ и Cλ, функционально связанных с Cλ-областью мыши таким образом, что экспрессируются антитела, содержащие полные или неполные человеческие легкие λ-цепи Ig), также предусмотрены другие отличные от человека животные, которые содержат сконструированный локус легкой λ-цепи Ig. Такие отличные от человека животные предусматривают любых из тех, которых можно подвергнуть генетической модификации для экспрессии антител, описанных в данном документе, в том числе, например, млекопитающих, например мышь, крысу, кролика, свинью, бычьих (например, корова, бык, буйвол), оленя, овцу, козу, курицу, кошку, собаку, хорька, примата (например, игрунка, макак-резус) и т.д. Например, для тех отличных от человека животных, для которых подходящие генетически модифицируемые ES-клетки являются труднодоступными, используют другие способы для получения отличного от человека животного, содержащего генетическую модификацию. Такие способы предусматривают, например, модификацию генома клеток, отличных от ES-клеток (например, фибробласт или индуцированная плюрипотентная клетка), и использование переноса ядер соматических клеток (SCNT) для переноса генетически модифицированного генома в подходящую клетку, например в безъядерный ооцит, и вынашивание модифицированной клетки (например, модифицированный ооцит) отличным от человека животным в подходящих условиях с образованием эмбриона.
[00230] Способы модификации генома зародышевого типа отличного от человека животного (например, геном свиньи, коровы, грызуна, курицы и т.д.) предусматривают, например, использование нуклеазы с «цинковыми пальцами» (ZFN), эффекторной нуклеазы, подобной активатору транскрипции (TALEN), или белка Cas (т.e. системы CRISPR/Cas) для включения сконструированного локуса легкой λ-цепи Ig, описанного в данном документе. Руководство для осуществления способов модификации генома зародышевого типа отличного от человека животного, можно найти, например, в заявках на выдачу патентов США №№14/747461 (поданной 23 июня 2015 года), 14/948221 (поданной 20 ноября 2015 года) и 14/974623 (поданной 18 декабря 2015 года); из которых все три заявки таким образом включены в данный документ с помощью ссылки в полном объеме.
[00231] В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, является млекопитающим. В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, является мелким млекопитающим, например из надсемейства Dipodoidea или Muroidea. В некоторых вариантах осуществления генетически модифицированное животное, описанное в данном документе, является грызуном. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из мыши, крысы и хомяка. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из надсемейства Muroidea. В некоторых вариантах осуществления генетически модифицированное животное, описанное в данном документе, относится к семейству, выбранному из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяки, крысы и мыши Нового света, полевки), Muridae (настоящие мыши и крысы, карликовые песчанки, иглистые мыши, косматые хомяки), Nesomyidae (лазающие мыши, скальные мыши, белохвостые крысы, малагасийские крысы и мыши), Platacanthomyidae (например, колючие сони) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В некоторых определенных вариантах осуществления генетически модифицированный грызун, описанный в данном документе, выбран из настоящей мыши или крысы (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. В некоторых определенных вариантах осуществления генетически модифицированная мышь, описанная в данном документе, является представителем семейства Muridae. В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, является грызуном. В некоторых определенных вариантах осуществления грызун, описанный в данном документе, выбран из мыши и крысы. В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, является мышью.
[00232] В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, является грызуном, который является мышью линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. В некоторых определенных вариантах осуществления описанная в данном документе мышь представляет собой линию 129, выбранную из группы, состоящей из линий 129P1, 129P2, 129P3, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129/SvJae, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2 (см., например, Festing et al., 1999, Mammalian Genome 10:836; Auerbach, W. et al., 2000, Biotechniques 29(5):1024-1028, 1030, 1032). В некоторых определенных вариантах осуществления генетически модифицированная мышь, описанная в данном документе, является помесью вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. В некоторых определенных вариантах осуществления мышь, описанная в данном документе, является помесью вышеупомянутых линий 129 или помесью вышеупомянутых линий BL/6. В некоторых определенных вариантах осуществления линия 129 в помеси, описанной в данном документе, представляет собой линию 129S6 (129/SvEvTac). В некоторых вариантах осуществления мышь, описанная в данном документе, принадлежит к линии BALB, например линии BALB/c. В некоторых вариантах осуществления мышь, описанная в данном документе, является помесью линии BALB и другой вышеупомянутой линии.
[00233] В некоторых вариантах осуществления отличное от человека животное, описанное в данном документе, является крысой. В некоторых определенных вариантах осуществления крыса, описанная в данном документе, выбрана из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. В некоторых определенных вариантах осуществления линия крыс, описанная в данном документе, представляет собой помесь двух или более линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
[00234] Плюрипотентная и/или тотипотентная клетка крысы может быть получена от любой линии крыс, включающей, например, линию крыс ACI, линию крыс Dark Agouti (DA), линию крыс Wistar, линию крыс LEA, линию крыс Sprague Dawley (SD) или линию крыс Fischer, такую как Fisher F344 или Fisher F6. Плюрипотентные и/или тотипотентные клетки крысы также могут быть получены из линии, происходящей из помеси двух или более линий, перечисленных выше. Например, плюрипотентная и/или тотипотентная клетка крысы может быть получена из линии DA или линии ACI. Линия крыс ACI характеризуется наличием окраски «черный агути» с белыми брюхом и лапами и гаплотипа RT1av1. Такие линии доступны из множества источников, в том числе Harlan Laboratories. Примером линии ES-клеток крысы от крысы ACI является ES-клетка крысы ACI.G1. Линия крыс Dark Agouti (DA) характеризуется наличием окраски «агути» и гаплотипа RT1av1. Такие крысы доступны из множества источников, в том числе Charles River и Harlan Laboratories. Примерами линии ES-клеток крысы от крысы DA являются линия ES- клеток крысы DA.2B и линия ES-клеток крысы DA.2C. В некоторых вариантах осуществления плюрипотентные и/или тотипотентные клетки крысы получены из инбредной линии крыс (см., например, публикацию заявки на выдачу патента США №2014-0235933 A1, опубликованную 21 августа 2014 года, включенную в данный документ посредством ссылки в полном объеме).
Конкретные иллюстративные варианты осуществления - сконструированные локусы H-цепи Ig
[00235] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат сконструированный локус легкой λ-цепи Ig, описанный в данном документе, и дополнительно содержат сконструированные локусы (или аллели) H-цепи Ig, характеризующиеся наличием множества сегментов человеческих генов VH, DH и JH, группированных в конфигурации зародышевого типа и функционально связанных с отличными от человеческих константными областями H-цепи Ig, энхансерами и регуляторными областями человеческой H-цепи Ig. В некоторых вариантах осуществления сконструированный локус (или аллель) H-цепи Ig, описанный в данном документе, содержит один или более сегментов человеческого гена VH, один или более сегментов человеческого гена DH и один или более сегментов человеческого гена JH, функционально связанных с отличной от человеческой константной областью H-цепи Ig.
[00236] В некоторых вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит 5, 10, 15, 20, 25, 30, 35, 40 или больше (например, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52 и т.д.) сегментов человеческого гена VH. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит все или практически все функциональные сегменты человеческого гена VH, которые находятся между человеческими генными сегментами VH3-74 и VH6-1 включительно, из человеческого локуса H-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит по меньшей мере сегменты человеческого гена VH, VH3-74, VH3-73, VH3-72, VH2-70, VH1-69, VH3-66, VH3-64, VH4-61, VH4-59, VH1-58, VH3-53, VH5-51, VH3-49, VH3-48, VH1-46, VH1-45, VH3-43, VH4-39, VH4-34, VH3-33, VH4-31, VH3-30, VH4-28, VH2-26, VH1-24, VH3-23, VH3-21, VH3-20, VH1-18, VH3-15, VH3-13, VH3-11, VH3-9, VH1-8, VH3-7, VH2-5, VH7-4-1, VH4-4, VH1-3, VH1-2 и VH6-1.
[00237] В некоторых вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит 5, 10, 15, 20, 25 или больше (например, 26, 27 и т.д.) сегментов человеческого гена DH. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит все или практически все функциональные сегменты человеческого гена DH, которые находятся между человеческими генными сегментами DH1-1 и DH7-27 включительно, из человеческого локуса H-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит по меньшей мере сегменты человеческого гена DH, DH1-1, DH2-2, DH3-3, DH4-4, DH5-5, DH6-6, DH1-7, DH2-8, DH3-9, DH3-10, DH5-12, DH6-13, DH2-15, DH3-16, DH4-17, DH6-19, DH1-20, DH2-21, DH3-22, DH6-25, DH1-26 и DH7-27.
[00238] В некоторых вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит 1, 2, 3, 4, 5, 6 или больше функциональных сегментов человеческого гена JH. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит все или практически все функциональные сегменты человеческого гена JH, которые находятся между человеческими генными сегментами JH1 и JH6 включительно, из человеческого локуса H-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) H-цепи Ig содержит по меньшей мере сегменты человеческого гена JH, JH1, JH2, JH3, JH4, JH5 и JH6.
[00239] В некоторых вариантах осуществления отличная от человеческой константная область H-цепи Ig содержит один или более отличных от человеческих генов константной области H-цепи Ig, таких как, например, ген иммуноглобулина M (IgM), иммуноглобулина D (IgD), иммуноглобулина G (IgG), иммуноглобулина E (IgE) и иммуноглобулина A (IgA). В некоторых определенных вариантах осуществления отличная от человеческой константная область H-цепи Ig включает гены константных областей IgM грызуна, IgD грызуна, IgG3 грызуна, IgG1 грызуна, IgG2b грызуна, IgG2a грызуна, IgE грызуна и IgA грызуна. В некоторых вариантах осуществления указанные сегменты человеческих генов VH, DH и JH функционально связаны с одним или более энхансерами (т.е. энхансерными последовательностями или энхансерными областями) отличной от человеческой H-цепи Ig. В некоторых вариантах осуществления указанные сегменты человеческих генов VH, DH и JH функционально связаны с одной или более регуляторными областями (или регуляторными последовательностями) отличной от человеческой H-цепи Ig. В некоторых вариантах осуществления указанные сегменты человеческих генов VH, DH и JH функционально связаны с одним или более энхансерами (или энхансерными последовательностями) отличной от человеческой H-цепи Ig и с одной или более регуляторными областями (или регуляторными последовательностями) отличной от человеческой H-цепи Ig.
[00240] В некоторых вариантах осуществления сконструированный локус H- цепи Ig, описанный в данном документе, не содержит эндогенный ген Adam6. В некоторых вариантах осуществления сконструированный локус H-цепи Ig, описанный в данном документе, не содержит эндогенный ген Adam6 (или кодирующую Adam6 последовательность) в том же положении генома зародышевого типа, в каком он находится в геноме зародышевого типа отличного от человека животного дикого типа того же вида. В некоторых вариантах осуществления сконструированный локус H-цепи Ig, описанный в данном документе, не содержит человеческий псевдоген Adam6. В некоторых вариантах осуществления сконструированный локус H-цепи Ig, описанный в данном документе, содержит вставку по меньшей мере одной нуклеотидной последовательности, которая кодирует один или более отличных от человеческих (например, грызуна) полипептидов Adam6. Указанная вставка может находиться вне сконструированного локуса тяжелой цепи иммуноглобулина, описанного в данном документе (например, против хода транскрипции относительно самого близко расположенного к 5' сегмента гена VH), в пределах сконструированного локуса H-цепи Ig или где-либо еще в геноме зародышевого типа отличного от человека животного (например, произвольным образом введенная кодирующая отличный от человеческого Adam6 последовательность), отличной от человеческой клетки или ткани.
[00241] В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, не экспрессируют на выявляемом уровне полную или неполную эндогенную отличную от человеческой VH-область в молекуле антитела. В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, не содержат (или характеризуются отсутствием или делецией) одной или более нуклеотидных последовательностей, которые кодируют полную или неполную эндогенную отличную от человеческой VH-область (например, VH, DH и/или JH) в молекуле антитела. В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, характеризуются геномом зародышевого типа, в котором предусматривается делеция сегментов эндогенных отличных от человеческих генов VH, DH и JH, полностью или частично. В различных вариантах осуществления предусмотренное отличное от человека животное является фертильным.
[00242] Руководство для получения нацеливающих векторов, отличных от человеческих клеток и отличных от человека животных, несущих такие сконструированные локусы (или аллели) H-цепи Ig, можно найти, например, в патентах США №№8642835 и 8697940, которые включены с помощью ссылки в полном объеме. Специалисты в данной области знают ряд технологий, известных в уровне техники, для осуществления таких генетических конструирования и/или манипуляции в отношении отличных от человеческих (например, млекопитающего) геномов или для получения, обеспечения или изготовления иным образом таких последовательностей, предназначенных для введения в геном зародышевого типа отличных от человека животных.
Конкретные иллюстративные варианты осуществления - сконструированные локусы легких κ-цепей Ig
[00243] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат сконструированный локус легкой λ-цепи Ig, описанный в данном документе, и дополнительно содержат сконструированные локусы (или аллели) легких κ-цепей Ig, характеризующиеся наличием множества сегментов человеческих генов Vκ и Jκ, группированных в конфигурации зародышевого типа и функционально связанных с отличной от человеческой константной областью легкой κ-цепи Ig, энхансерами и регуляторными областями κ-цепи Ig. В некоторых вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит один или более сегментов человеческого гена Vκ и один или более сегментов человеческого гена Jκ, функционально связанных с отличной от человеческой константной областью κ-цепи (Cκ) Ig.
[00244] В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит по меньшей мере сегменты человеческого гена Vκ, которые находятся в дистальном вариабельном кластере (или дистальном плече, или дистальной дупликации) человеческого локуса легкой κ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит по меньшей мере сегменты человеческого гена Vκ, которые находятся в проксимальном вариабельном кластере (или проксимальном плече, или проксимальной дупликации) человеческого локуса легкой κ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит сегменты человеческого гена Vκ, которые находятся в дистальном и проксимальном вариабельных кластерах человеческого локуса легкой κ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Vκ, которые находятся между человеческими генными сегментами Vκ2-40 (или Vκ3D-7) и Vκ4-1 включительно, из человеческого локуса легкой κ-цепи Ig, который встречается в природе.
[00245] В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит 5, 10, 15, 20, 25, 30, 35 или больше (например, 36, 37, 38, 39, 40 и т.д.) сегментов человеческого гена Vκ. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит сегменты человеческого гена Vκ, Vκ3D-7, Vκ1D-8, Vκ1D-43, Vκ3D-11, Vκ1D-12, Vκ1D-13, Vκ3D-15, Vκ1D-16, Vκ1D-17, Vκ3D-20, Vκ6D-21, Vκ2D-26, Vκ2D-28, Vκ2D-29, Vκ2D-30, Vκ1D-33, Vκ1D-39, Vκ2D-40, Vκ2-40, Vκ1-39, Vκ1-33, Vκ2-30, Vκ2-28, Vκ1-27, Vκ2-24, Vκ6-21, Vκ3-20, Vκ1-17, Vκ1-16, Vκ3-15, Vκ1-12, Vκ3-11, Vκ1-9, Vκ1-8, Vκ1-6, Vκ1-5, Vκ5-2 и Vκ4-1. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит по меньшей мере сегменты человеческого гена Vκ, Vκ3D-7, Vκ1D-8, Vκ1D-43, Vκ3D-11, Vκ1D-12, Vκ1D-13, Vκ3D-15, Vκ1D-16, Vκ1D-17, Vκ3D-20, Vκ6D-21, Vκ2D-26, Vκ2D-28, Vκ2D-29, Vκ2D-30, Vκ1D-33, Vκ1D-39 и Vκ2D-40. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит по меньшей мере сегменты человеческого гена Vκ, Vκ2-40, Vκ1-39, Vκ1-33, Vκ2-30, Vκ2-28, Vκ1-27, Vκ2-24, Vκ6-21, Vκ3-20, Vκ1-17, Vκ1-16, Vκ3-15, Vκ1-12, Vκ3-11, Vκ1-9, Vκ1-8, Vκ1-6, Vκ1-5, Vκ5-2 и Vκ4-1.
[00246] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит 1, 2, 3, 4, 5 или больше функциональных сегментов человеческого гена Jκ. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Jκ, которые находятся между человеческими генными сегментами Jκ1 и Jκ5 включительно, из человеческого локуса легкой κ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой κ-цепи Ig содержит по меньшей мере сегменты человеческого гена Jκ, Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5.
[00247] В некоторых вариантах осуществления указанные сегменты человеческих генов Vκ и Jκ функционально связаны с одним или более энхансерами (т.е. энхансерными последовательностями или энхансерными областями) отличной от человеческой легкой κ-цепи Ig. В некоторых вариантах осуществления указанные сегменты человеческих генов Vκ и Jκ функционально связаны с одной или более регуляторными областями (или регуляторными последовательностями) отличной от человеческой легкой κ-цепи Ig. В некоторых вариантах осуществления указанные сегменты человеческих генов Vκ и Jκ функционально связаны с одним или более энхансерами (или энхансерными последовательностями, или энхансерными областями) отличной от человеческой легкой κ-цепи Ig и одной или более регуляторными областями (или регуляторными последовательностями) отличной от человеческой легкой κ-цепи Ig.
[00248] В некоторых вариантах осуществления отличная от человеческой Cκ-область сконструированного локуса (или аллеля) легкой κ-цепи Ig содержит Cκ-область грызуна, такую как, например, Cκ-область мыши или Cκ-область крысы. В некоторых определенных вариантах осуществления отличная от человеческой Cκ-область сконструированного локуса (или аллеля) легкой κ-цепи Ig представляет собой или содержит Cκ-область мыши из генетического фона, который предусматривает линию 129, линию BALB/c, линию C57BL/6, помесную линию 129xC57BL/6 или их комбинации.
[00249] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат сконструированный локус легкой λ-цепи Ig, описанный в данном документе, и дополнительно содержат инактивированные локусы (или аллели) легких κ-цепей Ig.
[00250] В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, не экспрессируют на выявляемом уровне полную или неполную эндогенную отличную от человеческой Vκ-область в молекуле антитела. В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, не содержат (или характеризуются отсутствием или делецией) одной или более нуклеотидных последовательностей, которые кодируют полную или неполную эндогенную отличную от человеческой Vκ-область в молекуле антитела. В различных вариантах осуществления предусмотренные отличное от человека животное, отличная от человеческой клетка или отличная от человеческой ткань, описанные в данном документе, характеризуются геномом зародышевого типа, в котором предусматривается делеция сегментов эндогенных отличных от человеческих генов Vκ и Jκ, полностью или частично.
[00251] Руководство для получения нацеливающих векторов, отличных от человеческих клеток и отличных от человека животных, несущих такие сконструированные локусы (или аллели) легких κ-цепей Ig, можно найти, например, в патентах США №№8642835 и 8697940, которые таким образом включены с помощью ссылки в полном объеме. Специалисты в данной области знают ряд технологий, известных в уровне техники, для осуществления таких генетических конструирования и/или манипуляции в отношении отличных от человеческих (например, млекопитающего) геномов или для получения, обеспечения или изготовления иным образом таких последовательностей, предназначенных для введения в геном зародышевого типа отличных от человека животных.
Конкретные иллюстративные варианты осуществления - сконструированные локусы легких λ-цепей Ig
[00252] В некоторых вариантах осуществления предусмотренные отличные от человека животные содержат сконструированный локус легкой λ-цепи Ig, характеризующийся наличием множества сегментов человеческих генов Vλ, Jλи Cλ, группированных в конфигурации зародышевого типа и вставленных против хода транскрипции относительно сегмента отличного от человеческого гена Cλ (или гена Cλ-области) и функционально связанных с ним. Как описано в данном документе, такой сконструированный локус легкой λ-цепи Ig дополнительно содержит одну или более энхансерных областей (или энхансерных последовательностей) человеческой легкой λ-цепи Ig. В некоторых вариантах осуществления сконструированный локус легкой λ-цепи Ig содержит один или более сегментов человеческого гена Vλ и один или более сегментов человеческого гена Jλ, функционально связанных с отличной от человеческой константной областью легкой λ-цепи (Cλ) Ig. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит сегменты человеческих генов Vλ, которые находятся в по меньшей мере кластере A человеческого локуса легкой λ-цепи Ig; в некоторых вариантах осуществления, кластере A и кластере B человеческого локуса легкой λ-цепи Ig; в некоторых определенных вариантах осуществления, кластере A, кластере B и кластере C человеческого локуса легкой λ-цепи Ig.
[00253] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит 5, 10, 15, 20, 25, 30 или больше (например, 31, 32, 33, 34, 35 и т.д.) сегментов человеческого гена Vλ. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Vλ , которые находятся между человеческими генными сегментами Vλ4-69 и Vλ3-1 включительно, из человеческого локуса легкой λ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Vλ , которые находятся между человеческими генными сегментами Vλ5-52 и Vλ3-1 включительно, из человеческого локуса легкой λ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Vλ , которые находятся между человеческими генными сегментами Vλ3-27 и Vλ3-1 включительно, из человеческого локуса легкой λ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит сегменты человеческого гена Vλ, Vλ4-69, Vλ8-61, Vλ4-60, Vλ6-57, Vλ10-54, Vλ5-52, Vλ1-51, Vλ9-49, Vλ1-47, Vλ7-46, Vλ5-45, Vλ1-44, Vλ7-43, Vλ1-40, Vλ5-39, Vλ5-37, Vλ1-36, Vλ3-27, Vλ3-25, Vλ2-23, Vλ3-22, Vλ3-21, Vλ3-19, Vλ2-18, Vλ3-16, Vλ2-14, Vλ3-12, Vλ2-11, Vλ3-10, Vλ3-9, Vλ2-8, Vλ4-3 и Vλ3-1. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит по меньшей мере функциональные сегменты человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1.
[00254] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит 1, 2, 3, 4, 5, 6, 7 или больше функциональных сегментов человеческого гена Jλ. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Jλ, которые находятся между человеческими генными сегментами Jλ1 и Jλ7 включительно, из человеческого локуса легкой λ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит по меньшей мере сегменты человеческого гена Jλ, Jλ1, Jλ2, Jλ3, Jλ6 и Jλ7.
[00255] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит 1, 2, 3, 4, 5, 6, 7 или больше функциональных сегментов человеческого гена Cλ. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит все или практически все функциональные сегменты человеческого гена Cλ, которые находятся между человеческими генными сегментами Cλ1 и Cλ7 включительно, из человеческого локуса легкой λ-цепи Ig, который встречается в природе. В некоторых определенных вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig содержит по меньшей мере сегменты человеческого гена Cλ, Cλ1, Cλ2, Cλ3 и Cλ6.
[00256] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig не содержит те же энхансерные области (или энхансерные последовательности) отличной от человеческой легкой λ-цепи Ig, что встречаются в локусе (или аллеле) легкой λ-цепи Ig дикого типа. В некоторых вариантах осуществления в сконструированном локусе (или аллеле) легкой λ-цепи Ig полностью или частично отсутствует по меньшей мере одна энхансерная область (или энхансерная последовательность) отличной от человеческой легкой λ-цепи Ig (например, энхансер 2-4 λ-цепи Ig или Eλ2-4).
[00257] В некоторых вариантах осуществления указанные сегменты человеческих генов Vλ и Jλ функционально связаны с одним или более энхансерами (т.е. энхансерными последовательностями или энхансерными областями) отличной от человеческой легкой λ-цепи Ig и одним или более энхансерами (т.е. энхансерными последовательностями или энхансерными областями) человеческой легкой λ-цепи Ig. В некоторых вариантах осуществления указанные сегменты человеческих генов Vλ и Jλ функционально связаны с одной или более регуляторными областями (или регуляторными последовательностями) отличной от человеческой легкой λ-цепи Ig. В некоторых вариантах осуществления, указанные сегменты человеческих генов Vλ и Jλ функционально связаны с одним или более энхансерами (или энхансерными последовательностями, или энхансерными областями) отличной от человеческой легкой λ-цепи Ig, одним или более энхансерами (т.е. энхансерными последовательностями или энхансерными областями) человеческой легкой λ-цепи Ig и одной или более регуляторными областями (или регуляторными последовательностями) отличной от человеческой легкой λ-цепи Ig.
[00258] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, не содержит человеческий ген VpreB (или последовательность, кодирующую человеческий ген VpreB).
[00259] В некоторых вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig содержит Cλ-область грызуна, такую как, например, Cλ-область мыши или Cλ-область крысы. В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig представляет собой или содержит Cλ-область мыши из генетического фона, который предусматривает линию 129, линию BALB/c, линию C57BL/6, помесную линию 129xC57BL/6 или их комбинации.
[00260] В некоторых вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig, описанного в данном документе, содержит последовательность, которая на по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% идентична SEQ ID NO: 1 (Cλ1 мыши), SEQ ID NO: 3 (Cλ2 мыши) или SEQ ID NO: 5 (Cλ3 мыши). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig, описанного в данном документе, содержит последовательность, которая практически идентична или идентична SEQ ID NO: 1 (Cλ1 мыши), SEQ ID NO: 3 (Cλ2 мыши) или SEQ ID NO: 5 (Cλ3 мыши). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig, описанного в данном документе, представляет собой или содержит последовательность Cλ1-области мыши.
[00261] В некоторых вариантах осуществления отличная от человеческой Cλ-область, которая кодируется последовательностью, расположенной в сконструированном локусе (или аллеле) легкой λ-цепи Ig, описанном в данном документе, содержит последовательность, которая на по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% идентична SEQ ID NO: 2 (Cλ1 мыши), SEQ ID NO: 4 (Cλ2 мыши) или SEQ ID NO: 6 (Cλ3 мыши). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область, которая кодируется последовательностью, расположенной в сконструированном локусе (или аллеле) легкой λ-цепи Ig, описанного в данном документе, содержит последовательность, которая практически идентична или идентична SEQ ID NO: 2 (Cλ1 мыши), SEQ ID NO: 4 (Cλ2 мыши) или SEQ ID NO: 6 (Cλ3 мыши). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область, которая кодируется последовательностью, расположенной в сконструированном локусе (или аллеле) легкой λ-цепи Ig, описанном в данном документе, представляет собой или содержит полипептид Cλ1-области мыши.
[00262] В некоторых вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig, описанного в данном документе, содержит последовательность, которая на по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% идентична SEQ ID NO: 7 (Cλ1 крысы), SEQ ID NO: 9 (Cλ2 крысы), SEQ ID NO: 11 (Cλ3 крысы) или SEQ ID NO: 13 (Cλ4 крысы). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig, описанного в данном документе, содержит последовательность, которая практически идентична или идентична SEQ ID NO: 7 (Cλ1 крысы), SEQ ID NO: 9 (Cλ2 крысы), SEQ ID NO: 11 (Cλ3 крысы) или SEQ ID NO: 13 (Cλ4 крысы). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область сконструированного локуса (или аллеля) легкой λ-цепи Ig, описанного в данном документе, представляет собой или содержит последовательность Cλ1-области крысы.
[00263] В некоторых вариантах осуществления отличная от человеческой Cλ-область, которая кодируется последовательностью, расположенной в сконструированном локусе (или аллеле) легкой λ-цепи Ig, описанном в данном документе, содержит последовательность, которая на по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% идентична SEQ ID NO: 8 (Cλ1 крысы), SEQ ID NO: 10 (Cλ2 крысы), SEQ ID NO: 12 (Cλ3 крысы) или SEQ ID NO: 14 (Cλ4 крысы). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область, которая кодируется последовательностью, расположенной в сконструированном локусе (или аллеле) легкой λ-цепи Ig, описанном в данном документе, содержит последовательность, которая практически идентична или идентична SEQ ID NO: 8 (Cλ1 крысы), SEQ ID NO: 10 (Cλ2 крысы), SEQ ID NO: 12 (Cλ3 крысы) или SEQ ID NO: 14 (Cλ4 крысы). В некоторых определенных вариантах осуществления отличная от человеческой Cλ-область, которая кодируется последовательностью, расположенной в сконструированном локусе (или аллеле) легкой λ-цепи Ig, описанном в данном документе, представляет собой или содержит полипептид Cλ1-области крысы.
[00264] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, характеризуется наличием одного или более уникальных соединений нуклеотидных последовательностей (или комбинаций уникальных соединений последовательностей), образующихся в результате осуществления вставки генетического материала человека, соответствующего (геномной или синтетической) последовательности человеческой легкой λ-цепи Ig, вместо последовательности отличной от человеческой легкой λ-цепи Ig или в ее пределах в эндогенном локусе. Иллюстративные соединения нуклеотидных последовательностей представлены под SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128 и SEQ ID NO: 129.
[00265] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, содержит одну или более из SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128 и SEQ ID NO: 129.
[00266] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, содержит SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122 и SEQ ID NO: 123.
[00267] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, содержит SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122 и SEQ ID NO: 123.
[00268] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, содержит SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 128 и SEQ ID NO: 129.
[00269] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, содержит SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 126 и SEQ ID NO: 127.
[00270] В некоторых вариантах осуществления сконструированный локус (или аллель) легкой λ-цепи Ig, описанный в данном документе, содержит SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124 и SEQ ID NO: 125.
[00271] Методологию в отношении сегментов человеческих генов Vλ, Jλи Cλ можно найти, например, в Lefranc, M.P., 2000, Nomenclature of the human immunoglobulin lambda (IGL) genes, Current Protocols in Immunology, No. Supplement, 40:A.1p.1-A.1p.37. Среди прочего, в настоящем изобретении продемонстрировано, что наличие сегментов человеческих генов Vλ и Jλ в локусах (или аллелях) легких λ-цепей Ig обеспечивает увеличение разнообразия репертуара легких цепей у предусмотренного отличного от человека животного, по сравнению с разнообразием легких цепей в экспрессируемом репертуаре антител у отличного от человека животного, которое не содержит таких сконструированных аллелей легких λ-цепей Ig.
Способы
[00272] В определенных аспектах отличных от человека животных, описанных в данном документе, можно применять для получения человеческого антитела и/или последовательностей нуклеиновой кислоты, кодирующих человеческие антитела, при этом человеческое антитело содержит вариабельные домены, полученные из последовательностей нуклеиновой кислоты, кодируемых генетическим материалом клетки отличного от человека животного, описанного в данном документе. Например, отличное от человека животное, описанное в данном документе, иммунизируют с помощью представляющего интерес антигена, при условиях и в течение времени, достаточных для того, чтобы у отличного от человека животного выработался иммунный ответ на указанный представляющий интерес антиген. Антитела выделяют из отличного от человека животного (или одной или более клеток, например одной или более B-клеток), иммунизированного таким образом, и определяют их характеристики с использованием различных анализов, посредством которых, например, измеряют аффинность, специфичность, проводят картирование эпитопов, определяют способность блокирования взаимодействия лиганд-рецептор, измеряют уровень подавления активации рецепторов и т.д. В различных вариантах осуществления антитела, продуцируемые отличными от человека животными, описанными в данном документе, содержат один или более человеческих вариабельных доменов, которые получены из одной или более нуклеотидных последовательностей человеческой вариабельной области, выделенных из отличного от человека животного. В некоторых вариантах осуществления в отличных от человека животных, описанных в данном документе, может быть индуцирована выработка антител к лекарственным средствам (например, антиидиотипического антитела).
[00273] В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, представляют улучшенную систему in vivo и источник биологических материалов (например, клетки) для продуцирования человеческих антител, которые применимы во множестве анализов. В различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для разработки терапевтических средств, которые направлены на представляющий интерес полипептид (например, трансмембранный или секретируемый полипептид), и/или модулируют один или более видов активности, связанных с указанным представляющим интерес полипептидом, и/или модулируют взаимодействия указанного представляющего интерес полипептида с другими партнерами по связыванию (например, лигандом или рецепторным полипептидом). Например, в различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для разработки терапевтических средств, которые направлены на один или более рецепторных полипептидов, модулируют активность рецепторного полипептида и/или модулируют взаимодействия рецепторного полипептида с другими партнерами по связыванию. В различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для идентификации, скрининга и/или разработки кандидатных терапевтических средств (например, антител, siRNA и т.д.), которые связывают один или более представляющих интерес полипептидов. В различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для скрининга и разработки кандидатных терапевтических средств (например, антител, siRNA и т.д.), которые блокируют активность одного или более представляющих интерес полипептидов или которые блокируют активность одного или более представляющих интерес рецепторных полипептидов. В различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для определения профиля связывания антагонистов и/или агонистов одного или более представляющих интерес полипептидов. В некоторых вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для определения эпитопа или эпитопов одного или более кандидатных терапевтических антител, которые связывают один или более представляющих интерес полипептидов.
[00274] В различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для определения фармакокинетических профилей одного или более человеческих антител-кандидатов. В различных вариантах осуществления каждое из одного или более отличных от человека животных, описанных в данном документе, и одного или более контрольных или эталонных отличных от человека животных подвергают воздействию одного или более человеческих антител-кандидатов в различных дозах (например, 0,1 мг/кг, 0,2 мг/кг, 0,3 мг/кг, 0,4 мг/кг, 0,5 мг/кг, 1 мг/кг, 2 мг/кг, 3 мг/кг, 4 мг/кг, 5 мг/кг, 7,5 мг/кг, 10 мг/кг, 15 мг/кг, 20 мг/кг, 25 мг/кг, 30 мг/кг, 40 мг/кг или 50 мг/кг или больше). Кандидатные терапевтические антитела можно вводить дозами посредством любого желаемого пути введения, в том числе парентерального и отличного от парентерального путей введения. Парентеральные пути предусматривают, например, внутривенный, внутриартериальный, интрапортальный, внутримышечный, подкожный, внутрибрюшинный, интраспинальный, интратекальный, интрацеребровентрикулярный, внутричерепной, интраплевральный или другие пути инъекции. Непарентеральные пути включают, например, пероральный, интраназальный, трансдермальный, внутрилегочный, ректальный, трансбуккальный, вагинальный, внутриглазной. Введение также можно осуществлять посредством непрерывной инфузии, местного введения, замедленного высвобождения из имплантатов (гелей, мембран и т.п.) и/или внутривенной инъекции. У отличных от человека животных (гуманизированных и контрольных), отбирают кровь в различные моменты времени (например, 0 ч., 6 ч., 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней или до 30 или больше дней). Можно проводить различные анализы для определения фармакокинетических профилей вводимых кандидатных терапевтических антител с использованием образцов, полученных от отличных от человека животных, описанных в данном документе, в том числе без ограничения анализы уровня общего IgG, гуморального ответа на терапевтические средства, агглютинации и т.д.
[00275] В различных вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для измерения терапевтического эффекта, заключающегося в блокировании или модулировании активности представляющего интерес полипептида, и эффекта в отношении экспрессии генов в результате клеточных изменений или, в случае рецепторного полипептида, численности молекул рецепторного полипептида на поверхности клеток отличных от человека животных. В различных вариантах осуществления отличное от человека животное, описанное в данном документе, или выделенные из него клетки подвергают воздействию кандидатного терапевтического средства, которое связывает представляющий интерес полипептид и, через дополнительный период времени, анализируют в отношении эффектов на конкретные клеточные процессы, которые связаны с указанным представляющим интерес полипептидом, например взаимодействий лиганд-рецептор или сигнальной трансдукции.
[00276] В определенных аспектах отличные от человека, животные описанные в данном документе, экспрессируют человеческие вариабельные домены антител, таким образом, можно получить клетки, клеточные линии и культуры клеток, которые служат в качестве источника человеческих вариабельных доменов антител, предназначенных для применения в анализах связывания и функциональных анализах, например для анализа в отношении связывания или функции антагониста или агониста, в частности, где антагонист или агонист является специфическим по отношению к человеческому антигену, представляющему интерес, или специфическим по отношению к эпитопу, который функционирует при взаимодействии (связывании) лиганд-рецептор. В различных вариантах осуществления эпитопы, с которыми связываются кандидатные терапевтические антитела или siRNA, можно определять с применением клеток, выделенных из отличных от человека животных, описанных в данном документе.
[00277] Клетки предусмотренных отличных от человека животных можно выделять и использовать по ситуации или их можно поддерживать в культуре в течение многих поколений. В различных вариантах осуществления клетки предусмотренного отличного от человека животного подвергают иммортализации (например, посредством использования вируса) и поддерживают в культуре в течение неопределенного периода времени (например, в последовательных культурах).
[00278] В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, обеспечивают систему in vivo для получения вариантов человеческих вариабельных доменов антител, которые связывают представляющий интерес полипептид (например, варианты человеческих Vλ-доменов). Такие варианты предусматривают человеческие вариабельные домены антител с необходимой функциональностью, специфичностью, низкой способностью к перекрестной реакции с общим эпитопом, который есть для двух или более вариантов представляющего интерес полипептида. В некоторых вариантах осуществления отличных от человека животных, описанных в данном документе, применяют для получения панелей человеческих вариабельных доменов антител, содержащих серии вариантных вариабельных доменов, которые подвергают скринингу в отношении необходимой или улучшенной функциональности.
[00279] В определенных аспектах отличные от человека животные, описанные в данном документе, обеспечивают систему in vivo для получения библиотек человеческих вариабельных областей антител (например, библиотека человеческих Vλ-доменов). Такие библиотеки обеспечивают источник последовательностей вариабельных областей тяжелых и/или легких цепей, которые могут быть привиты на различные Fc-области на основе необходимой эффекторной функции, могут применяться в качестве источника для «созревания аффинности» последовательности вариабельной области с применением методик, известных из уровня техники (например, сайт-направленный мутагенез, ПЦР с внесением ошибок и т.д.), и/или могут применяться в качестве источника компонентов антител для получения терапевтических молекул на основе антител, таких как, например, химерные рецепторы антигенов (т.е., молекула, сконструированная с применением компонентов антител, например, scFv), полиспецифические связывающие средства (например, биспецифические связывающие средства) и слитые белки (например, однодоменные антитела, scFv и т.д.).
[00280] В некоторых аспектах отличные от человека животные, описанные в данном документе, обеспечивают систему in vivo для анализа и тестирования лекарственного средства или вакцины. В различных вариантах осуществления кандидатные лекарственное средство или вакцину можно доставлять одному или более отличным от человека животным, описанным в данном документе, с последующим наблюдением за отличными от человека животными для определения одного или более иммунных ответов на лекарственное средство или вакцину, профиля безопасности лекарственного средства или вакцины или эффекта в отношении заболевания или состояния и/или одного или более симптомов заболевания или состояния. Иллюстративные способы, используемые для определения профиля безопасности, включают измерения токсичности, оптимальной концентрации дозы, ответа с участием антитела (т.е. антитела к лекарственному средству), эффективности лекарственного средства или вакцины и возможных факторов риска. Такие лекарственные средства или вакцины можно улучшать и/или разрабатывать с участием таких отличных от человека животных.
[00281] Эффективность вакцины можно определять с помощью ряда способов. Вкратце, отличных от человека животных, описанных в данном документе, вакцинируют с помощью способов, известных из уровня техники, и затем заражают вакциной, или вакцину вводят уже инфицированному отличному от человека животному. Ответ отличного(-ых) от человека животного(-ых) на вакцину можно измерять путем мониторинга отличного(-ых) от человека животного(-ых) (или выделенных из него(них) клеток) и/или проведения одного или более анализов в их отношении для определения эффективности вакцины. Ответ отличного(-ых) от человека животного(-ых) на вакцину затем сравнивают с ответом контрольных животных, используя один или более показателей, известных из уровня техники и/или описанных в данном документе.
[00282] Эффективность вакцины можно дополнительно определять с помощью анализов нейтрализации вируса. Вкратце, отличных от человека животных, описанных в данном документе, иммунизируют и сыворотку крови собирают в разные дни после иммунизации. Серийные разведения сыворотки крови предварительно инкубируют с вирусом, и в ходе этого периода времени антитела в сыворотке крови, специфичные к вирусу, связываются с ним. Смесь вирус/сыворотка крови затем добавляют к пермиссивным клеткам для определения инфицирующей способности с помощью анализа бляшкообразования или анализа микронейтрализации. Если антитела в сыворотке крови нейтрализуют вирус, то наблюдается меньшее количество бляшек или более низкое количество относительных люциферазных единиц по сравнению с контрольной группой.
[00283] В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, продуцируют человеческие вариабельные домены антител и, следовательно, обеспечивают систему in vivo для получения человеческих антител, предназначенных для применения в путях диагностического применения (например, иммунология, серология, микробиология, клеточная патология и т.д.). В различных вариантах осуществления отличных от человека животных, описанных в данном документе, можно применять для получения человеческих вариабельных доменов антител, которые связывают соответствующие антигенные участки, для идентификации клеточных изменений, таких как, например, экспрессия специфических маркеров клеточной поверхности, характеризующих патологические изменения. Такие антитела можно конъюгировать с различными химическими частицами (например, радиоактивной меткой) и применять при необходимости в различных анализах in vivo и/или in vitro.
[00284] В некоторых вариантах осуществления отличные от человека животные, описанные в данном документе, обеспечивают улучшенную систему in vivo для разработки и отбора человеческих антител, предназначенных для применения при онкологических и/или инфекционных заболеваниях. В различных вариантах осуществления отличным от человека животным, описанным в данном документе, и контрольным отличным от человека животным (например, характеризующееся генетической модификацией, которая отличается от таковой, описанной в данном документе, или без генетической модификации, т.е. дикого типа) можно имплантировать опухоль (или опухолевые клетки) или их можно инфицировать вирусом (например, вирус гриппа, HIV, HCV, HPV и т.д.). После имплантации или инфицирования отличным от человека животным можно вводить кандидатное терапевтическое средство. Перед введением кандидатного терапевтического средства опухоли или вирусу может быть предоставлено достаточное количество времени для развития в одном или более местоположениях в пределах организма отличных от человека животных. В качестве альтернативы и/или дополнительно, иммунный ответ у таких отличных от человека животных можно мониторить таким образом, чтобы определять характеристики и отбирать потенциальные человеческие антитела, которые можно разрабатывать в качестве терапевтического средства.
Наборы
[00285] В некоторых аспектах в настоящем изобретении дополнительно предусмотрены упаковка или набор, содержащие один или более контейнеров, заполненных по меньшей мере одним отличным от человека животным, отличной от человеческой клеткой, фрагментом ДНК, нацеливающим вектором или любой их комбинацией, описанными в данном документе. Наборы можно применять согласно любому применимому способу (например, способу исследования). Необязательно связанным с таким(-и) контейнером(-ами) может быть уведомление в форме, предписанной правительственным органом, регулирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, при этом уведомление отражает (а) одобрение органом, регулирующим производство, применение или продажу, для введения человеку, (b) указания по применению и/или (c) договор, который регулирует передачу материалов и/или биологических продуктов (например, отличное от человека животное или отличная от человеческой клетка, описанные в данном документе) между двумя или более организациями, и их комбинации.
[00286] Другие признаки определенных вариантов осуществления станут очевидными при прочтении следующих описаний иллюстративных вариантов осуществления, которые приведены в иллюстративных целях и не подразумеваются как ограничивающие их.
Дополнительные иллюстративные варианты осуществления
[00287] В иллюстративном варианте осуществления 1 в данном документе предусмотрен грызун, геном зародышевого типа которого содержит эндогенный локус легкой λ-цепи иммуноглобулина, содержащий (a) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, где (a) и (b) функционально связаны с (c) и сегментом гена Cλ грызуна, и где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина.
[00288] В иллюстративном варианте осуществления 2 в данном документе предусмотрен грызун согласно варианту осуществления 1, где эндогенный локус легкой λ-цепи иммуноглобулина содержит два Eλ грызуна.
[00289] В иллюстративном варианте осуществления 3 в данном документе предусмотрен грызун согласно варианту осуществления 2, где два Eλ грызуна представляют собой Eλ мыши и Eλ3-1 мыши.
[00290] В иллюстрированном варианте осуществления 4 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-3, где эндогенный локус легкой λ-цепи иммуноглобулина содержит три человеческих Eλ.
[00291] В иллюстративном варианте осуществления 5 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-4, где геном зародышевого типа дополнительно содержит (i) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или (ii) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00292] В иллюстративном варианте осуществления 6 в данном документе предусмотрен грызун согласно варианту осуществления 5, где вставка одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH обеспечивает замену сегментов генов VH, DH грызуна.
[00293] В иллюстративном варианте осуществления 7 в данном документе предусмотрен грызун согласно варианту осуществления 6, где вставка содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов VH, DH и JH и их комбинациями.
[00294] В иллюстративном варианте осуществления 8 в данном документе предусмотрен грызун согласно варианту осуществления 5 или 6, где вставка одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ обеспечивает замену сегментов генов Vκ и Jκ грызуна.
[00295] В иллюстративном варианте осуществления 9 в данном документе предусмотрен грызун согласно варианту осуществления 8, где вставка содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов Vκ и Jκ и их комбинациями.
[00296] В иллюстративном варианте осуществления 10 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-8, где константная область тяжелой цепи иммуноглобулина грызуна представляет собой эндогенную константную область тяжелой цепи иммуноглобулина грызуна.
[00297] В иллюстративном варианте осуществления 11 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-10, где Cκ-область грызуна представляет собой эндогенную Cκ-область грызуна.
[00298] В иллюстративном варианте осуществления 12 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-9, где эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию эндогенных сегментов генов Vλ и Jλ, полностью или частично.
[00299] В иллюстративном варианте осуществления 13 в данном документе предусмотрен грызун согласно варианту осуществления 12, где эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию генных сегментов Vλ2-Vλ3-Jλ2-Cλ2 и генных сегментов Vλ1-Jλ3-Cλ3-Jλ1.
[00300] В иллюстративном варианте осуществления 14 в данном документе предусмотрен грызун согласно варианту осуществления 12, где эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P и генных сегментов Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1.
[00301] В иллюстративном варианте осуществления 15 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-14, где сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00302] В иллюстративном варианте осуществления 16 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-13, где эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию Eλ2-4 грызуна.
[00303] В иллюстративном варианте осуществления 17 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-16, где грызун не экспрессирует на выявляемом уровне эндогенные легкие λ-цепи иммуноглобулина.
[00304] В иллюстративном варианте осуществления 18 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-17, где локус тяжелой цепи иммуноглобулина содержит вставку сегментов человеческого гена VH от VH3-74 до VH6-1, сегментов человеческого гена DH от DH1-1 до DH7-27 и сегментов человеческого гена JH, JH1-JH6.
[00305] В иллюстративном варианте осуществления 19 в данном документе предусмотрен грызун согласно варианту осуществления 18, где вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими VH3-74 и VH6-1, некодирующую ДНК человека, которая встречается в природе между человеческими DH1-1 и DH7-27, и некодирующую ДНК человека, которая встречается в природе между человеческими JH1 и JH6.
[00306] В иллюстративном варианте осуществления 20 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-19, где локус легкой κ-цепи иммуноглобулина содержит вставку проксимальной дупликации Vκ, полностью или частично, человеческого локуса легкой κ-цепи иммуноглобулина.
[00307] В иллюстративном варианте осуществления 21 в данном документе предусмотрен грызун согласно варианту осуществления 20, где локус легкой κ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vκ от Vκ2-40 до Vκ4-1 и сегментов человеческого гена Jκ, от Jκ1 до Jκ5.
[00308] В иллюстративном варианте осуществления 22 в данном документе предусмотрен грызун согласно варианту осуществления 21, где вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими Vκ2-40 и Vκ4-1, и некодирующую ДНК человека, которая встречается в природе между человеческими Jκ1 и Jκ5.
[00309] В иллюстративном варианте осуществления 23 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-22, где эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, по меньшей мере пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, сегмента человеческого гена Jλ, Jλ7, и сегмента гена Cλ1 грызуна.
[00310] В иллюстративном варианте осуществления 24 в данном документе предусмотрен грызун согласно вариантам осуществления 23, где вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими Vλ5-52 и Vλ1-40 и Vλ3-27 и Vλ3-1, некодирующую ДНК человека, которая в природе встречается между парами сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, и некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно сегмента человеческого гена Jλ, Jλ7.
[00311] В иллюстративном варианте осуществления 25 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-24, где в локусе тяжелой цепи иммуноглобулина отсутствует эндогенный ген Adam6 грызуна.
[00312] В иллюстративном варианте осуществления 26 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-25, где локус тяжелой цепи иммуноглобулина дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна.
[00313] В иллюстративном варианте осуществления 27 в данном документе предусмотрен грызун согласно варианту осуществления 26, где одна или более нуклеотидных последовательностей вставлены между первым и вторым сегментами человеческого гена VH.
[00314] В иллюстративном варианте осуществления 28 в данном документе предусмотрен грызун согласно варианту осуществления 26, где одна или более нуклеотидных последовательностей вставлены вместо человеческого псевдогена Adam6.
[00315] В иллюстративном варианте осуществления 29 в данном документе предусмотрен грызун согласно варианту осуществления 27, где первый сегмент человеческого гена VH представляет собой человеческий VH1-2, и второй сегмент человеческого гена VH представляет собой человеческий VH6-1.
[00316] В иллюстративном варианте осуществления 30 в данном документе предусмотрен грызун согласно варианту осуществления 26, где одна или более нуклеотидных последовательностей вставлены между сегментом человеческого гена VH и сегментом человеческого гена DH.
[00317] В иллюстративном варианте осущесвления 31 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-30, где грызун является гетерозиготным или гомозиготным в отношении эндогенного локуса тяжелой цепи иммуноглобулина.
[00318] В иллюстративном варианте осуществления 32 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 5-31, где грызун является гетерозиготным или гомозиготным в отношении эндогенного локуса легкой κ-цепи иммуноглобулина.
[00319] В иллюстративном варианте осуществления 33 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-32, где грызун является гетерозиготным или гомозиготным в отношении эндогенного локуса легкой λ-цепи иммуноглобулина.
[00320] В иллюстративном варианте осуществления 34 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 1-33, где грызун представляет собой крысу или мышь.
[00321] В иллюстративном варианте осуществления 35 в данном документе предусмотрена выделенная клетка грызуна, геном зародышевого типа которой содержит эндогенный локус легкой λ-цепи иммуноглобулина, содержащий (a) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, (i) где (a) и (b) функционально связаны с (c) и с сегментом гена Cλ грызуна, и (ii) где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина.
[00322] В иллюстративном варианте осуществления 36 в данном документе предусмотрена иммортализованная клетка, полученная с помощью клетки грызуна согласно варианту осуществления 35.
[00323] В иллюстративном варианте осуществления 37 в данном документе предусмотрена выделенная клетка грызуна согласно варианту осуществления 35, где клетка грызуна представляет собой эмбриональную стволовую клетку грызуна.
[00324] В иллюстративном варианте осуществления 38 в данном документе предусмотрен эмбрион грызуна, полученный с помощью эмбриональной стволовой клетки грызуна согласно варианту осуществления 35.
[00325] В иллюстративном варианте осуществления 39 в данном документе предусмотрен способ получения грызуна, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи иммуноглобулина, при этом способ включает: (a) введение фрагмента ДНК в эмбриональную стволовую клетку грызуна, при этом указанный фрагмент ДНК содержит нуклеотидную последовательность, которая содержит (i) один или более сегментов человеческого гена Vλ, (ii) один или более сегментов человеческого гена Jλ и (iii) один или более сегментов человеческого гена Cλ, где (i)-(iii) функционально связаны с сегментом гена Cλ грызуна, и где нуклеотидная последовательность дополнительно содержит один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина; (b) получение эмбриональной стволовой клетки грызуна, созданной на (a); и (c) получение грызуна с применением эмбриональной стволовой клетки грызуна с (b).
[00326] В иллюстративном варианте осуществления 40 в данном документе предусмотрен способ согласно варианту осуществления 39, где нуклеотидная последовательность дополнительно содержит и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина.
[00327] В иллюстративном варианте осуществления 41 в данном документе предусмотрен способ получения грызуна, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи иммуноглобулина, при этом сконструированный эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом гена Cλ грызуна или человека, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина; при этом способ включает модификацию генома зародышевого типа грызуна, таким образом, что в нем содержится сконструированный локус легкой λ-цепи иммуноглобулина, который содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом гена Cλ грызуна или человека, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина, с получением, тем самым, указанного грызуна.
[00328] В иллюстративном варианте осуществления 42 в данном документе предусмотрен способ согласно варианту осуществления 39 или 41, где один или более сегментов человеческого гена Vλ включают от Vλ5-52 до Vλ1-40 и/или от Vλ3-27 до Vλ3-1.
[00329] В иллюстративном варианте осуществления 43 в данном документе предусмотрен способ согласно варианту осуществления 42, где один или более сегментов человеческого гена Vλ содержат некодирующую ДНК человека, которая встречается в природе между человеческими Vλ5-52 и Vλ1-40 и/или Vλ3-27 и Vλ3-1.
[00330] В иллюстративном варианте осуществления 44 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39-43, где один или более сегментов человеческого гена Jλ и один или более сегментов человеческого гена Cλ содержат пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3- Cλ3, Jλ6-Cλ6, и человеческий генный сегмент Jλ7.
[00331] В иллюстративном варианте осуществления 45 в данном документе предусмотрен способ согласно варианту осуществления 44, где пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержат некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλи Cλ, и человеческий генный сегмент Jλ7содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[00332] В иллюстративном варианте осуществления 46 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39-45, где сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00333] В иллюстративном варианте осуществления 47 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39-46, где эндогенный локус легкой λ-цепи иммуноглобулина содержит три человеческих Eλ.
[00334] В иллюстративном варианте осуществления 48 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39-46, где эндогенный локус легкой λ-цепи иммуноглобулина содержит два Eλ грызуна.
[00335] В иллюстративном варианте осуществления 49 в данном документе предусмотрен способ согласно варианту осуществления 48, где два Eλ грызуна представляют собой Eλ мыши и Eλ3-1 мыши.
[00336] В иллюстративном варианте осуществления 50 в данном документе предусмотрен способ согласно любому из вариантов осуществления 38 и 42-49, где фрагмент ДНК дополнительно содержит один или более маркеров отбора.
[00337] В иллюстративном варианте осуществления 51 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39 и 42-50, где фрагмент ДНК дополнительно содержит один или более участков сайт- специфической рекомбинации.
[00338] В иллюстративном варианте осуществления 52 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39 и 42-51, где фрагмент ДНК с (a) вводят в эмбриональную стволовую клетку грызуна, геном зародышевого типа которой содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00339] В иллюстративном варианте осуществления 53 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39 и 42-51, где фрагмент ДНК с (a) вводят в эмбриональную стволовую клетку грызуна, геном зародышевого типа которой содержит эндогенный локус тяжелой цепи иммуноглобулина дикого типа или эндогенный локус тяжелой цепи иммуноглобулина дикого типа и эндогенный локус легкой κ-цепи иммуноглобулина дикого типа; и где способ дополнительно включает стадию скрещивания мыши, полученной с помощью указанной отличной от человеческой эмбриональной стволовой клетки, со второй мышью.
[00340] В иллюстративном варианте осуществления 54 в данном документе предусмотрен способ согласно любому из вариантов осуществления 47-49, где модификацию генома зародышевого типа грызуна, с тем чтобы он содержал сконструированный локус легкой λ-цепи иммуноглобулина, осуществляют в эмбриональной стволовой клетке грызуна, геном зародышевого типа которой дополнительно содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00341] В иллюстративном варианте осуществления 55 в данном документе предусмотрен способ согласно варианту осуществления 52 или 54, где вставка одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH содержит некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена VH, некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена DH, и некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена JH.
[00342] В иллюстративном варианте осуществления 56 в данном документе предусмотрен способ согласно варианту осуществления 52 или 54, где вставка одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ содержит некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена Vκ, и некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена Jκ.
[00343] В иллюстративном варианте осуществления 57 в данном документе предусмотрен способ согласно любому из вариантов осуществления 41-49, где модификацию генома зародышевого типа отличного от человека животного, с тем чтобы он содержал сконструированный локус легкой λ-цепи иммуноглобулина, осуществляют в отличной от человеческой эмбриональной стволовой клетке, геном зародышевого типа которой содержит эндогенный локус тяжелой цепи иммуноглобулина дикого типа или эндогенный локус тяжелой цепи иммуноглобулина дикого типа и эндогенный локус легкой κ-цепи иммуноглобулина дикого типа; и где способ дополнительно включает стадию скрещивания мыши, полученной с помощью указанной отличной от человеческой эмбриональной стволовой клетки, со второй мышью.
[00344] В иллюстративном варианте осуществления 58 в данном документе предусмотрен способ согласно варианту осуществления 53 или 57, где вторая мышь характеризуется геномом зародышевого типа, содержащим локусы H-цепи Ig и κ-цепи Ig дикого типа.
[00345] В иллюстративном варианте осуществления 59 в данном документе предусмотрен способ согласно варианту осуществления 53 или 57, где вторая мышь характеризуется геномом зародышевого типа, содержащим гомозиготные или гетерозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный или гетерозиготный гуманизированный локус H-цепи Ig содержит вставленную последовательность, кодирующую Adam6 грызуна.
[00346] В иллюстративном варианте осуществления 60 в данном документе предусмотрен способ согласно варианту осуществления 53 или 57, где вторая мышь характеризуется геномом зародышевого типа, содержащим гомозиготный или гетерозиготный гуманизированный локус H-цепи Ig и гомозиготный или гетерозиготный инактивированный локус κ-цепи Ig.
[00347] В иллюстративном варианте осуществления 61 в данном документе предусмотрен способ получения антитела у грызуна, при этом способ включает стадии (1) иммунизации грызуна с помощью представляющего интерес антигена, при этом грызун характеризуется геномом зародышевого типа, содержащим эндогенный локус легкой λ-цепи иммуноглобулина, который содержит (ai) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, где (a) и (b) функционально связаны с (c) и с сегментом гена Cλ грызуна, и где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина; (2) содержания грызуна в условиях, достаточных для того, чтобы у грызуна выработался иммунный ответ на представляющий интерес антиген; и (3) выделения антитела из грызуна или клетки грызуна, которое связывает представляющий интерес антиген.
[00348] В иллюстративном варианте осуществления 62 в данном документе предусмотрен способ согласно варианту осуществления 61, где грызун характеризуется геномом зародышевого типа, в котором дополнительно содержатся эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00349] В иллюстративном варианте осуществления 63 в данном документе предусмотрен способ согласно варианту осуществления 61 или 62, где клетка грызуна представляет собой B-клетку.
[00350] В иллюстративном варианте осуществления 64 в данном документе предусмотрен способ согласно варианту осуществления 61 или 62, где клетка грызуна представляет собой гибридому.
[00351] В иллюстративном варианте осуществления 65 в данном документе предусмотрен способ согласно любому из вариантов осуществления 61-64, где эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, сегмента человеческого гена Jλ, Jλ7.
[00352] В иллюстративном варианте осуществления 66 в данном документе предусмотрен способ согласно варианту осуществления 65, где вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческим Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, некодирующую ДНК человека, которая в природе встречается между парами сегментов человеческих генов Jλ-Cλ Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, и некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно сегмента человеческого гена Jλ, Jλ7.
[00353] В иллюстративном варианте осуществления 67 в данном документе предусмотрен способ согласно любому из вариантов осуществления 61-66, где сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00354] В иллюстративном варианте осуществления 68 в данном документе предусмотрен способ согласно любому из вариантов осуществления 62-67, где локус тяжелой цепи иммуноглобулина содержит вставку сегментов человеческого гена VH от VH3-74 до VH6-1, сегментов человеческого гена DH от DH1-1 до DH7-27 и сегментов человеческого гена JH, JH1-JH6, и при этом сегменты человеческих генов VH, DH и JH функционально связаны с эндогенной константной областью тяжелой цепи иммуноглобулина грызуна.
[00355] В иллюстративном варианте осуществления 69 в данном документе предусмотрен способ согласно варианту осуществления 68, где вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими VH3-74 и VH6-1, некодирующую ДНК человека, которая встречается в природе между человеческими DH1-1 и DH7-27, и некодирующую ДНК человека, которая встречается в природе между человеческими JH1 и JH6.
[00356] В иллюстративном варианте осуществления 70 в данном документе предусмотрен способ согласно варианту осуществления 68, где сегменты человеческих генов VH, DH и JH обеспечивают замену сегментов генов VH, DH и JH грызуна.
[00357] В иллюстративном варианте осуществления 71 в данном документе предусмотрен способ согласно любому из вариантов осуществления 62-70, где локус легкой κ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vκ от Vκ2-40 до Vκ4-1 и сегментов человеческого гена Jκ, Jκ1-Jκ5, и при этом сегменты человеческих генов Vκ и Jκ функционально связаны с эндогенной Cκ-областью иммуноглобулина грызуна.
[00358] В иллюстративном варианте осуществления 72 в данном документе предусмотрен способ согласно варианту осуществления 71, где вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими Vκ2-40 и Vκ4-1, и некодирующую ДНК человека, которая встречается в природе между человеческими Jκ1 и Jκ5.
[00359] В иллюстративном варианте осуществления 73 в данном документе предусмотрен способ согласно варианту осуществления 71, где сегменты человеческих генов Vκ и Jκ обеспечивают замену сегментов генов Vκ и Jκ грызуна.
[00360] В иллюстративном варианте осуществления 74 в данном документе предусмотрен способ согласно любому из вариантов осуществления 61-73, где в геноме зародышевого типа грызуна дополнительно содержится вставка одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна.
[00361] В иллюстративном варианте осуществления 75 в данном документе предусмотрен способ согласно любому из вариантов осуществления 62-74, где в локусе тяжелой цепи иммуноглобулина отсутствует эндогенный ген Adam6 грызуна.
[00362] В иллюстративном варианте осуществления 76 в данном документе предусмотрен способ согласно варианту осуществления 75, где локус тяжелой цепи иммуноглобулина дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна.
[00363] В иллюстративном варианте осуществления 77 в данном документе предусмотрен способ согласно варианту осуществления 76, где одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены между первым и вторым сегментами человеческого гена VH.
[00364] В иллюстративном варианте осуществления 78 в данном документе предусмотрен способ согласно варианту осуществления 77, где первый сегмент человеческого гена VH представляет собой человеческий VH1-2, и второй сегмент человеческого гена VH представляет собой человеческий VH6-1.
[00365] В иллюстративном варианте осуществления 79 в данном документе предусмотрен способ согласно варианту осуществления 76, где одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены вместо человеческого псевдогена Adam6.
[00366] В иллюстративном варианте осуществления 80 в данном документе предусмотрен способ согласно варианту осуществления 76, где одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены между сегментом человеческого гена VH и сегментом человеческого гена DH.
[00367] В иллюстративном варианте осуществления 81 в данном документе предусмотрен способ согласно любому из вариантов осуществления 61-80, где антитело, полученное из грызуна или клетки грызуна, которое связывает представляющий интерес антиген, содержит человеческий вариабельный домен тяжелой цепи и человеческий вариабельный домен легкой лямбда-цепи.
[00368] В иллюстративном варианте осуществления 82 в данном документе предусмотрен способ согласно варианту осуществления 81, где человеческий вариабельный домен тяжелой цепи содержит перегруппированный сегмент человеческого гена VH, выбранный из группы, состоящей из VH3-74, VH3-73, VH3-72, VH2-70, VH1-69, VH3-66, VH3-64, VH4-61, VH4-59, VH1-58, VH3-53, VH5-51, VH3-49, VH3-48, VH1-46, VH1-45, VH3-43, VH4-39, VH4-34, VH3-33, VH4-31, VH3-30, VH4-28, VH2-26, VH1-24, VH3-23, VH3-21, VH3-20, VH1-18, VH3-15, VH3-13, VH3-11, VH3-9, VH1-8, VH3-7, VH2-5, VH7-4-1, VH4-4, VH1-3, VH1-2 и VH6-1.
[00369] В иллюстративном варианте осуществления 83 в данном документе предусмотрен способ согласно варианту осуществления 81 или 82, где человеческий вариабельный домен легкой лямбда-цепи содержит перегруппированный сегмент человеческого гена Vλ, выбранный из группы, состоящей из Vλ4-69, Vλ8-61, Vλ4-60, Vλ6-57, Vλ10-54, Vλ5-52, Vλ1-51, Vλ9-49, Vλ1-47, Vλ7-46, Vλ5-45, Vλ1-44, Vλ7-43, Vλ1-40, Vλ5-39, Vλ5-37, Vλ1-36, Vλ3-27, Vλ3-25, Vλ2-23, Vλ3-22, Vλ3-21, Vλ3-19, Vλ2-18, Vλ3-16, Vλ2-14, Vλ3-12, Vλ2-11, Vλ3-10, Vλ3-9, Vλ2-8, Vλ4-3 и Vλ3-1.
[00370] В иллюстративном варианте осуществления 84 в данном документе предусмотрен способ согласно любому из вариантов осуществления 39-83, где грызун представляет собой мышь или крысу.
[00371] В иллюстративном варианте осуществления 85 в данном документе предусмотрен грызун, геном зародышевого типа которого содержит гомозиготный эндогенный локус легкой λ-цепи иммуноглобулина, который содержит (i) сегменты человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, (ii) пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, (iii) сегмент человеческого гена Jλ, Jλ7, и (iv) три энхансера человеческой легкой λ-цепи иммуноглобулина; где (i)-(iv) функционально связаны друг с другом и (i)-(iii) расположены против хода транскрипции относительно сегмента гена Cλ грызуна, и где в эндогенном локусе легкой λ-цепи иммуноглобулина отсутствует эндогенный Eλ2-4 иммуноглобулина грызуна, сегменты человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1 содержат некодирующую ДНК человека, которая встречается в природе между сегментами человеческого гена Vλ, пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержат некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλ-Cλ, и сегмент человеческого гена Jλ, Jλ7, содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[00372] В иллюстративном варианте осуществления 86 в данном документе предусмотрен грызун согласно варианту осуществления 85, где сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00373] В иллюстративном варианте осуществления 87 в данном документе предусмотрен грызун согласно варианту осуществления 85 или 86, где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит эндогенные энхансеры легкой λ-цепи иммуноглобулина грызуна, Eλ и Eλ3-1.
[00374] В иллюстративном варианте осуществления 88 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 85-87, где эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию эндогенных генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P и генных сегментов Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1 грызуна.
[00375] В иллюстративном варианте осуществления 89 в данном документе предусмотрен грызун согласно любому из вариантов осуществления 85-88, где грызун представляет собой крысу или мышь.
[00376] В некоторых вариантах осуществления в данном документе предусмотрен грызун, геном зародышевого типа которого содержит эндогенный локус легкой λ-цепи иммуноглобулина, содержащий (a) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, где (a) и (b) функционально связаны с (c) и с сегментом гена Cλ грызуна, и где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина.
[00377] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит два Eλ грызуна.
[00378] В некоторых вариантах осуществления два Eλ грызуна представляют собой Eλ мыши и Eλ3-1 мыши.
[00379] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит три человеческих Eλ.
[00380] В некоторых вариантах осуществления геном зародышевого типа дополнительно содержит (i) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или (ii) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00381] В некоторых вариантах осуществления вставка одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH обеспечивает замену сегментов гена VH, DH грызуна.
[00382] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов VH, DH и JH и их комбинациями.
[00383] В некоторых вариантах осуществления вставка одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ обеспечивает замену сегментов генов Vκ и Jκ грызуна.
[00384] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов Vκ и Jκ и их комбинациями.
[00385] В некоторых вариантах осуществления константная область тяжелой цепи иммуноглобулина грызуна представляет собой эндогенную константную область тяжелой цепи иммуноглобулина грызуна.
[00386] В некоторых вариантах осуществления, где Cκ-область грызуна представляет собой эндогенную Cκ-область грызуна.
[00387] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию сегментов эндогенных генов Vλ и Jλ, полностью или частично.
[00388] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию генных сегментов Vλ2-Vλ3-Jλ2-Cλ2 и генных сегментов Vλ1-Jλ3-Cλ3-Jλ1.
[00389] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P и генных сегментов Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1.
[00390] В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00391] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию Eλ2-4 грызуна.
[00392] В некоторых вариантах осуществления у грызуна не экспрессируются на выявляемом уровне эндогенные легкие λ-цепи иммуноглобулина.
[00393] В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина содержит вставку сегментов человеческого гена VH от VH3-74 до VH6-1, сегментов человеческого гена DH от DH1-1 до DH7-27 и сегментов человеческого гена JH, JH1-JH6.
[00394] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими VH3-74 и VH6-1, некодирующую ДНК человека, которая встречается в природе между человеческими DH1-1 и DH7-27, и некодирующую ДНК человека, которая встречается в природе между человеческими JH1-JH6.
[00395] В некоторых вариантах осуществления локус легкой κ-цепи иммуноглобулина содержит вставку проксимальной дупликации Vκ, полностью или частично, человеческого локуса легкой κ-цепи иммуноглобулина.
[00396] В некоторых вариантах осуществления локус легкой κ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vκ от Vκ2-40 до Vκ4-1 и сегментов человеческого гена Jκ от Jκ1 до Jκ5.
[00397] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими Vκ2-40 и Vκ4-1, и некодирующую ДНК человека, которая встречается в природе между человеческими Jκ1-Jκ5.
[00398] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, по меньшей мере пар сегментов человеческих генов Jλ- Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, сегмента человеческого гена Jλ, Jλ7, и сегмента гена Cλ1 грызуна.
[00399] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими Vλ5-52 и Vλ1-40 и Vλ3-27 и Vλ3-1, некодирующую ДНК человека, которая в природе встречается между парами сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, и некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно сегмента человеческого гена Jλ, Jλ7.
[00400] В некоторых вариантах осуществления в локусе тяжелой цепи иммуноглобулина отсутствует эндогенный ген Adam6 грызуна.
[00401] В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна.
[00402] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей вставлены между первым и вторым сегментами человеческого гена VH.
[00403] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей вставлены вместо человеческого псевдогена Adam6.
[00404] В некоторых вариантах осуществления первый сегмент человеческого гена VH представляет собой человеческий VH1-2, и второй сегмент человеческого гена VH представляет собой человеческий VH6-1.
[00405] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей вставлены между сегментом человеческого гена VH и сегментом человеческого гена DH.
[00406] В некоторых вариантах осуществления грызун является гетерозиготным или гомозиготным в отношении эндогенного локуса тяжелой цепи иммуноглобулина.
[00407] В некоторых вариантах осуществления грызун является гетерозиготным или гомозиготным в отношении эндогенного локуса легкой κ-цепи иммуноглобулина.
[00408] В некоторых вариантах осуществления грызун является гетерозиготным или гомозиготным в отношении эндогенного локуса легкой λ-цепи иммуноглобулина.
[00409] В некоторых вариантах осуществления грызун представляет собой крысу или мышь.
[00410] В некоторых вариантах осуществления в данном документе предусмотрена выделенная клетка грызуна, геном зародышевого типа которой содержит эндогенный локус легкой λ-цепи иммуноглобулина, содержащий (a) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, (i) где (a) и (b) функционально связаны с (c) и с сегментом гена Cλ грызуна, и (ii) где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина.
[00411] В некоторых вариантах осуществления в данном документе предусмотрена иммортализованная клетка, полученная с помощью клетки грызуна, предусмотренной в данном документе.
[00412] В некоторых вариантах осуществления клетка грызуна представляет собой эмбриональную стволовую клетку грызуна.
[00413] В некоторых вариантах осуществления в данном документе предусмотрен эмбрион грызуна, полученный с помощью эмбриональной стволовой клетки грызуна, предусмотренной в данном документе.
[00414] В некоторых вариантах осуществления в данном документе предусмотрен способ получения грызуна, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи иммуноглобулина, при этом способ включает (a) введение фрагмента ДНК в эмбриональную стволовую клетку грызуна, при этом указанный фрагмент ДНК содержит нуклеотидную последовательность, которая включает (i) один или более сегментов человеческого гена Vλ, (ii) один или более сегментов человеческого гена Jλ и (iii) один или более сегментов человеческого гена Cλ, где (i)-(iii) функционально связаны с сегментом гена Cλ грызуна, и где нуклеотидная последовательность дополнительно содержит один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина; (b) получение эмбриональной стволовой клетки грызуна, созданной на (a); и (c) получение грызуна с применением эмбриональной стволовой клетки грызуна с (b).
[00415] В некоторых вариантах осуществления нуклеотидная последовательность дополнительно содержит и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина.
[00416] В некоторых вариантах осуществления в данном документе предусмотрен способ получения грызуна, геном зародышевого типа которого содержит сконструированный эндогенный локус легкой λ-цепи иммуноглобулина, при этом сконструированный эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом гена Cλ грызуна или человека, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина; при этом способ включает модификацию генома зародышевого типа грызуна, таким образом, что он содержит сконструированный локус легкой λ-цепи иммуноглобулина, который содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом гена Cλ грызуна или человека, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина, с получением, тем самым, указанного грызуна.
[00417] В некоторых вариантах осуществления один или более сегментов человеческого гена Vλ содержат от Vλ5-52 до Vλ1-40 и/или от Vλ3-27 до Vλ3-1.
[00418] В некоторых вариантах осуществления один или более сегментов человеческого гена Vλ содержат некодирующую ДНК человека, которая встречается в природе между человеческими Vλ5-52 и Vλ1-40 и/или Vλ3-27 и Vλ3-1.
[00419] В некоторых вариантах осуществления один или более сегментов человеческого гена Jλ и один или более сегментов человеческого гена Cλ содержат пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, и человеческий генный сегмент Jλ7.
[00420] В некоторых вариантах осуществления пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержат некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλ и Cλ, и человеческий генный сегмент Jλ7 содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[00421] В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00422] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит три человеческих Eλ.
[00423] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит два Eλ грызуна.
[00424] В некоторых вариантах осуществления два Eλ грызуна представляют собой Eλ мыши и Eλ3-1 мыши.
[00425] В некоторых вариантах осуществления фрагмент ДНК дополнительно содержит один или более маркеров отбора.
[00426] В некоторых вариантах осуществления фрагмент ДНК дополнительно содержит один или более участков сайт-специфической рекомбинации.
[00427] В некоторых вариантах осуществления фрагмент ДНК с (a) вводят в эмбриональную стволовую клетку грызуна, геном зародышевого типа которой содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00428] В некоторых вариантах осуществления фрагмент ДНК с (a) вводят в эмбриональную стволовую клетку грызуна, геном зародышевого типа которой содержит эндогенный локус тяжелой цепи иммуноглобулина дикого типа или эндогенный локус тяжелой цепи иммуноглобулина дикого типа и эндогенный локус легкой κ-цепи иммуноглобулина дикого типа; и где способ дополнительно включает стадию скрещивания мыши, полученной с помощью указанной отличной от человеческой эмбриональной стволовой клетки, со второй мышью.
[00429] В некоторых вариантах осуществления модификацию генома зародышевого типа грызуна, с тем чтобы он содержал сконструированный локус легкой λ-цепи иммуноглобулина, осуществляют в эмбриональной стволовой клетке грызуна, геном зародышевого типа которой дополнительно содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκи Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00430] В некоторых вариантах осуществления вставка одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH содержит некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена VH, некодирующую ДНК человека которая встречается в природе между одним или более сегментами человеческого гена DH, и некодирующую ДНК человека, которая встречается в природе между одним или более сегментами человеческого гена JH.
[00431] В некоторых вариантах осуществления вставка одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ содержит некодирующую ДНК человека которая встречается в природе между одним или более сегментами человеческого гена Vκ, и некодирующую ДНК человека которая встречается в природе между одним или более сегментами человеческого гена Jκ.
[00432] В некоторых вариантах осуществления модификацию генома зародышевого типа отличного от человека животного, с тем чтобы он содержал сконструированный локус легкой λ-цепи иммуноглобулина, осуществляют в отличной от человеческой эмбриональной стволовой клетке, геном зародышевого типа которой содержит эндогенный локус тяжелой цепи иммуноглобулина дикого типа или эндогенный локус тяжелой цепи иммуноглобулина дикого типа и эндогенный локус легкой κ-цепи иммуноглобулина дикого типа; и где способ дополнительно включает стадию скрещивания мыши, полученной с помощью указанной отличной от человеческой эмбриональной стволовой клетки, со второй мышью.
[00433] В некоторых вариантах осуществления вторая мышь характеризуется геномом зародышевого типа, содержащим локусы H-цепи Ig и κ-цепи Ig дикого типа.
[00434] В некоторых вариантах осуществления вторая мышь характеризуется геномом зародышевого типа, содержащим гомозиготные или гетерозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный или гетерозиготный гуманизированный локус H-цепи Ig содержит вставленную последовательность, кодирующую Adam6 грызуна.
[00435] В некоторых вариантах осуществления вторая мышь характеризуется геномом зародышевого типа, содержащим гомозиготный или гетерозиготный гуманизированный локус H-цепи Ig и гомозиготный или гетерозиготный инактивированный локус κ-цепи Ig.
[00436] В некоторых вариантах осуществления предусмотрен способ получения антитела у грызуна, при этом способ включает стадии (1) иммунизации грызуна с помощью представляющего интерес антигена, при этом грызун характеризуется геномом зародышевого типа, содержащим эндогенный локус легкой λ-цепи иммуноглобулина, который содержит (ai) один или более сегментов человеческого гена Vλ, (b) один или более сегментов человеческого гена Jλ и (c) один или более сегментов человеческого гена Cλ, где (a) и (b) функционально связаны с (c) и с сегментом гена Cλ грызуна, и где эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) иммуноглобулина грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) иммуноглобулина; (2) содержания грызуна в условиях, достаточных для того, чтобы у грызуна выработался иммунный ответ на представляющий интерес антиген; и (3) выделения антитела из грызуна или клетки грызуна, которое связывает представляющий интерес антиген.
[00437] В некоторых вариантах осуществления грызун характеризуется геномом зародышевого типа, который дополнительно содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна; или эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с константной областью тяжелой цепи иммуноглобулина грызуна, и эндогенный локус легкой κ-цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с Cκ-областью иммуноглобулина грызуна.
[00438] В некоторых вариантах осуществления клетка грызуна представляет собой B-клетку.
[00439] В некоторых вариантах осуществления клетка грызуна представляет собой гибридому.
[00440] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, пар сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2- Cλ2, Jλ3-Cλ3, Jλ6-Cλ6, и сегмента человеческого гена Jλ, Jλ7.
[00441] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческим Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, некодирующую ДНК человека, которая в природе встречается между парами сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, и некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно сегмента человеческого гена Jλ, Jλ7..
[00442] В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00443] В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина содержит вставку сегментов человеческого гена VH от VH3-74 до VH6-1, сегментов человеческого гена DH от DH1-1 до DH7-27 и сегментов человеческого гена JH, JH1-JH6, и при этом сегменты человеческих генов VH, DH и JH функционально связаны с эндогенной константной областью тяжелой цепи иммуноглобулина грызуна.
[00444] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими VH3-74 и VH6-1, некодирующую ДНК человека, которая встречается в природе между человеческими DH1-1 и DH7-27, и некодирующую ДНК человека, которая встречается в природе между человеческими JH1-JH6.
[00445] В некоторых вариантах осуществления сегменты человеческих генов VH, DH и JH обеспечивают замену сегментов генов VH, DH и JH грызуна.
[00446] В некоторых вариантах осуществления локус легкой κ-цепи иммуноглобулина содержит вставку сегментов человеческого гена Vκ от Vκ2-40 до Vκ4-1 и сегментов человеческого гена Jκ, Jκ1-Jκ5, и при этом сегменты человеческих генов Vκ и Jκ функционально связаны с эндогенной Cκ-областью иммуноглобулина грызуна.
[00447] В некоторых вариантах осуществления вставка содержит некодирующую ДНК человека, которая встречается в природе между человеческими Vκ2-40 и Vκ4-1, и некодирующую ДНК человека, которая встречается в природе между человеческими Jκ1-Jκ5.
[00448] В некоторых вариантах осуществления сегменты человеческих генов Vκ и Jκ обеспечивают замену сегментов гена Vκ и Jκ грызуна.
[00449] В некоторых вариантах осуществления в геноме зародышевого типа грызуна дополнительно содержится вставка одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна.
[00450] В некоторых вариантах осуществления в локусе тяжелой цепи иммуноглобулина отсутствует эндогенный ген Adam6 грызуна.
[00451] В некоторых вариантах осуществления локус тяжелой цепи иммуноглобулина дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна.
[00452] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены между первым и вторым сегментами человеческого гена VH.
[00453] В некоторых вариантах осуществления первый сегмент человеческого гена VH представляет собой человеческий VH1-2, и второй сегмент человеческого гена VH представляет собой человеческий VH6-1.
[00454] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены вместо человеческого псевдогена Adam6.
[00455] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна, вставлены между сегментом человеческого гена VH и сегментом человеческого гена DH.
[00456] В некоторых вариантах осуществления антитело, выделенное из грызуна или клетки грызуна, которое связывает представляющий интерес антиген, содержит человеческий вариабельный домен тяжелой цепи и человеческий вариабельный домен легкой лямбда-цепи.
[00457] В некоторых вариантах осуществления человеческий вариабельный домен тяжелой цепи содержит перегруппированный сегмент человеческого гена VH, выбранный из группы, состоящей из VH3-74, VH3-73, VH3-72, VH2-70, VH1-69, VH3-66, VH3-64, VH4-61, VH4-59, VH1-58, VH3-53, VH5-51, VH3-49, VH3-48, VH1-46, VH1-45, VH3-43, VH4-39, VH4-34, VH3-33, VH4-31, VH3-30, VH4-28, VH2-26, VH1-24, VH3-23, VH3-21, VH3-20, VH1-18, VH3-15, VH3-13, VH3-11, VH3-9, VH1-8, VH3-7, VH2-5, VH7-4-1, VH4-4, VH1-3, VH1-2 и VH6-1.
[00458] В некоторых вариантах осуществления человеческий вариабельный домен легкой лямбда-цепи содержит перегруппированный сегмент человеческого гена Vλ, выбранный из группы, состоящей из Vλ4-69, Vλ8-61, Vλ4-60, Vλ6-57, Vλ10-54, Vλ5-52, Vλ1-51, Vλ9-49, Vλ1-47, Vλ7-46, Vλ5-45, Vλ1-44, Vλ7-43, Vλ1-40, Vλ5-39, Vλ5-37, Vλ1-36, Vλ3-27, Vλ3-25, Vλ2-23, Vλ3-22, Vλ3-21, Vλ3-19, Vλ2-18, Vλ3-16, Vλ2-14, Vλ3-12, Vλ2-11, Vλ3-10, Vλ3-9, Vλ2-8, Vλ4-3 и Vλ3-1.
[00459] В некоторых вариантах осуществления грызун представляет собой мышь или крысу.
[00460] В некоторых вариантах осуществления в данном документе предусмотрен грызун, геном зародышевого типа которого содержит гомозиготный эндогенный локус легкой λ-цепи иммуноглобулина, который содержит (i) сегменты человеческого гена Vλот Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1, (ii) пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, (iii) сегмент человеческого гена Jλ, Jλ7, и (iv) три энхансера человеческой легкой λ-цепи иммуноглобулина; где (i)-(iv) функционально связаны друг с другом и (i)-(iii) расположены против хода транскрипции относительно сегмента гена Cλ грызуна, и где в эндогенном локусе легкой λ-цепи иммуноглобулина отсутствует эндогенный Eλ2-4 иммуноглобулина грызуна, сегменты человеческого гена Vλ от Vλ5-52 до Vλ1-40 и от Vλ3-27 до Vλ3-1 содержат некодирующую ДНК человека, которая встречается в природе между сегментами человеческого гена Vλ, пары сегментов человеческих генов Jλ-Cλ, Jλ1-Cλ1, Jλ2-Cλ2, Jλ3-Cλ3 и Jλ6-Cλ6, содержат некодирующую ДНК человека, которая встречается в природе между парами сегментов человеческих генов Jλ-Cλ, и сегмент человеческого гена Jλ, Jλ7, содержит некодирующую ДНК человека, которая встречается в природе против хода транскрипции (или в 5'-направлении) относительно человеческого Jλ7.
[00461] В некоторых вариантах осуществления сегмент гена Cλ грызуна представляет собой сегмент гена Cλ1 мыши.
[00462] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит эндогенные энхансеры легкой λ-цепи иммуноглобулина грызуна, Eλ и Eλ3-1.
[00463] В некоторых вариантах осуществления эндогенный локус легкой λ-цепи иммуноглобулина предусматривает делецию эндогенных генных сегментов Vλ2-Vλ3-Jλ2-Cλ2-Jλ4P-Cλ4P и Vλ1-Jλ3-Jλ3P-Cλ3-Jλ1 грызуна.
[00464] В некоторых вариантах осуществления грызун представляет собой крысу или мышь.
ПРИМЕРЫ
[00465] Следующие примеры приведены для того, чтобы обеспечить специалистов в данной области описанием того, как получать и применять способы и композиции, описанные в данном документе, и они не предназначены для ограничения объема того, что авторы настоящего изобретения рассматривают в качестве своего изобретения. Если не указано иное, температура указана в градусах Цельсия, а давление является атмосферным или близким к нему.
Пример 1. Построение нацеливающего вектора для конструирования локуса легкой λ-цепи Ig грызуна
[00466] В данном примере показаны иллюстративные способы построения нацеливающего вектора для вставки в геном отличного от человека животного, такого как грызун (например, мышь). В способах, описанных в данном примере, продемонстрировано получение отличного от человека животного, геном зародышевого типа которого содержит сконструированный локус легкой λ-цепи Ig. В частности, в данном примере продемонстрировано построение серий нацеливающих векторов для конструирования эндогенного локуса легкой λ-цепи Ig у отличного от человека животного, таким образом, что отличное от человека животное экспрессирует и/или продуцирует антитела, которые содержат легкие λ-цепи Ig с человеческими и отличными от человеческих вариабельными доменами или, в некоторых вариантах осуществления, человеческими константными доменами из указанного эндогенного локуса легкой λ-цепи Ig в геноме зародышевого типа отличного от человека животного. Как описано ниже, серии нацеливающих векторов, содержащих различные количества генетического материала, соответствующего человеческому локусу легкой λ-цепи Ig (т.е. человеческие Vλ, Jλ, Cλ и энхансерные последовательности человеческой λ-цепи Ig), подвергают вставке в эндогенный локус легкой λ-цепи Ig грызуна. В частности, указанный генетический материал подвергают вставке против хода транскрипции относительно гена (или сегмента гена) Cλ грызуна таким образом, что сегменты человеческих генов Vλ, Jλ и Cλ функционально связаны с указанным геном Cλ грызуна. Способы, описанные в данном примере, обеспечивают сохранение и/или делецию эндогенных сегментов генов (или последовательностей) λ-цепи Ig грызуна. Иллюстративное схематическое изображение серий нацеливающих векторов для построения сконструированного эндогенного локуса легкой λ-цепи Ig представлено на фигурах 1-4.
[00467] Серии нацеливающих векторов, содержащих различные количества человеческих Vλ, Jλ, Cλ и энхансерных последовательностей (или областей) человеческой λ-цепи Ig, для осуществления вставки в локус легкой λ-цепи Ig грызуна создавали с применением технологии VELOCIGENE® (см., например, патент США №6586251 и Valenzuela et al., 2003, Nature Biotech. 21(6):652-9; включенные в данный документ с помощью ссылки в полном объеме) и методик молекулярной биологии, известных из уровня техники. Способы, описанные в данном примере, можно применять при необходимости с использованием любых человеческих Vλ, Jλ, Cλ и энхансерных последовательностей или комбинации последовательностей (или фрагментов последовательностей) человеческой λ-цепи Ig. В таблице 1 изложены краткие описания каждого нацеливающего вектора, проиллюстрированного на фигуре 1.
[00468] Вкратце, приблизительно 12 т. н. (11822 п. о.) геномной последовательности человеческой λ-цепи Ig из клона бактериальной искусственной хромосомы (BAC) для человека CTD-2502m16 лигировали в клон BAC для мыши RP23-60e14. Данный клон BAC для мыши конструировали с целью укоротить клон BAC на приблизительно 90 т. н., вставить уникальные участки распознавания рестрикционными ферментами AsiSI и PI-SceI по ходу транскрипции относительно гена Cλ1 мыши и заменить исходный ген устойчивости к хлорамфениколу (CMR) на ген устойчивости к спектиномицину (SpecR) и уникальный участок рестрикции I-CeuI посредством двух последовательных стадий бактериальной гомологичной рекомбинации (BHR) перед лигированием с геномной последовательностью человеческой λ-цепи Ig. Клон BAC для человека CTD-2502m16 также модифицировали с помощью двух последовательных стадий BHR для замещения приблизительно 53 т. н. человеческой последовательности на 3'-конце с помощью кассеты для отбора по устойчивости к неомицину и уникального участка рестрикции PI-SceI и замещения гена CMR и приблизительно 101,5 т. н. человеческой последовательности на 5'-конце с помощью кассеты для отбора по устойчивости к гигромицину и уникального участка рестрикции AsiSI с размещением, тем самым, участка AsiSI и кассеты для отбора по устойчивости к неомицину размером приблизительно 2885 п. о. против хода транскрипции и приблизительно 1418 п. о. по ходу транскрипции соответственно относительно модульной человеческой энхансерной области (см., например, Asenbauer, H. and H.G. Klobeck, 1996, Eur. J. Immunol. 26(1):142-50, которая таким образом включена посредством ссылки в полном объеме). Геномная последовательность человеческой λ-цепи Ig содержала приблизительно 7,5 т. н., соответствующих энхансерной области (или последовательности) человеческой λ-цепи (Eλ) Ig, которая является модульной и содержит три элемента последовательности (фигура 1; Asenbauer, H. and H.G. Klobeck, 1996, Eur. J. Immunol. 26(1):142-50), и соответственно 2,9 т. н. и 1,4 т. н. 5'- и 3'-фланкирующей последовательности, а также кассету для отбора по устойчивости к неомицину (т.е. ген устойчивости к неомицину [NEOR] под транскрипционным контролем промотора убиквитина и фланкированный участками loxP). Модифицированные клоны BAC для человека и мыши расщепляли с помощью участков AsiSI и PI-SceI и лигировали вместе. После лигирования со сконструированным клоном BAC для мыши полученный в результате нацеливающий вектор содержал приблизительно 39166 п. о. последовательности мыши в виде 5'-плеча гомологии и включал генные сегменты Vλ1, Jλ3, Jλ3P, Cλ3, Jλ1 и Cλ1 λ-цепи Ig мыши (нацеливающий вектор 6286, фигура 1). 3'-Плечо гомологии (приблизительно 30395 п. о.) содержало энхансер λ-цепи (mEλ) Ig мыши. С целью упрощения на изображении нацеливающего вектора 6286 на фигуре 1 плечи гомологии мыши не показаны. Гомологичная рекомбинация с данным нацеливающим вектором в результате приводила к осуществлению вставки трех энхансерных последовательностей человеческой λ-цепи Ig, а также 5'- и 3'-фланкирующих последовательностей без какой-либо делеции последовательности мыши. Опосредованной рекомбиназами делеции кассеты для отбора по устойчивости к неомицину достигали в ES-клетках с помощью транзиентной экспрессии рекомбиназы Cre (см., например, Lakso, M. et al., 1992, Proc. Natl. Acad. Sci. USA 89: 6232-6; Orban, P.C. et al., 1992, Proc. Natl. Acad. Sci. U.S.A. 89:6861-5; Gu, H. et al., 1993, Cell 73(6):1155-64; Araki, K. et al., 1995, Proc. Natl. Acad. Sci. U.S.A. 92:160-4; Dymecki, S.M., 1996, Proc. Natl. Acad. Sci. U.S.A. 93(12):6191-6; все из которых включены в данный документ с помощью ссылки в полном объеме).
[00469] Вторую конструкцию (нацеливающий вектор 6571) конструировали так, чтобы она содержала группу из пяти функциональных сегментов человеческого гена Vλи практически весь кластер человеческих Jλ-Cλ (т.е. человеческие Jλ1-Cλ1-Jλ2-Cλ2-Jλ3-Cλ3-Jλ4-Cλ4-Jλ5-Cλ5-Jλ6-Cλ6-hJλ7), охватывающий приблизительно 125473 п. о., которые получали с помощью клона BAC для человека CTD-2079i4. Для построения нацеливающего вектора человеческий ген Cλ7 в клоне BAC для человека CTD-2079i4 сперва заменяли с помощью BHR геном Cλ1 мыши и приблизительно 1588 п. о. фланкирующей последовательности, который амплифицировали с помощью ПЦР с применением клона BAC для мыши RP23-60e14 в качестве матрицы. 5'-Плечо гомологии, содержащее приблизительно 37161 п. о. последовательности мыши, соответствующей последовательности 5' гена Cλ1 мыши в клоне BAC для мыши RP23-60e14, затем лигировали с модифицированным клоном BAC для человека CTD-2079i4, содержащим синтетический ген Cλ1 мыши, с применением уникальных участков распознавания рестрикционными ферментами I-CeuI и PI-SceI, по отдельности введенных в клоны BAC как для мыши, так и для человека с помощью BHR. Данное 5'-плечо гомологии содержало генные сегменты Vλ1, Jλ3, Jλ3P, Cλ3 и Jλ1 мыши (фигура 1). 3'-Плечо гомологии содержало приблизительно 9189 п. о. человеческой последовательности, соответствующей двум человеческим Eλ из нацеливающего вектора 6286 (фигура 1).
[00470] Третью конструкцию (нацеливающий вектор 6596) конструировали так, чтобы она содержала дополнительные одиннадцать функциональных сегментов человеческого гена Vλ. Данный нацеливающий вектор содержал приблизительно 171458 п. о. человеческой последовательности из клона BAC RP11-761L13. Согласно схеме включали три сегмента человеческого гена Vλ с обеспечением гомологичной 3'-последовательности, перекрывающейся (приблизительно 33469 п. о.) с нацеливающим вектором 6571. Как описано выше, 5'-плечо гомологии, содержащее приблизительно 37161 п. о. последовательности мыши, лигировали с 5'-концом фрагмента ДНК, содержащего сегменты человеческого гена Vλ, с применением уникальных участков распознавания рестрикционными ферментами I-CeuI и AscI, по отдельности введенных в клоны BAC как для мыши, так и для человека с помощью BHR.
[00471] Четвертую конструкцию (нацеливающий вектор 6597) конструировали так, чтобы она содержала дополнительные девять функциональных сегментов человеческого гена Vλ. Данный нацеливающий вектор содержал приблизительно 121188 п. о. человеческой последовательности из двух клонов BAC, RP11-22L18 и RP11-761L13. Как описано выше, 3'-конец данной человеческой последовательности содержал дополнительные сегменты человеческого гена Vλ, которые обеспечивали гомологичную 3'-последовательность, перекрывающуюся с нацеливающим вектором 6596 (приблизительно 27468 п. о.). Как описано выше в отношении нацеливающих векторов 6571 и 6596, приблизительно 37161 п. о. 5'-плеча гомологии, содержащего последовательность мыши, из клона BAC RP23-60e14, лигировали с 5'-концом человеческой последовательности с применением уникальных участков распознавания рестрикционными ферментами I-CeuI и AscI, по отдельности введенных в клоны BAC как для мыши, так и для человека с помощью BHR.
[00472] Сходным образом конструировали пятую конструкцию (нацеливающий вектор 6680) так, чтобы она содержала те же дополнительные девять функциональных сегментов человеческого гена Vλ, что и нацеливающий вектор 6597, за исключением того, что 5'-плечо гомологии изменяли с обеспечением возможности делеции локуса легкой λ-цепи Ig мыши посредством гомологичной рекомбинации. Данное 5'-плечо гомологии содержало приблизительно 22298 п. о. из клона BAC для мыши RP23-15m16, и его лигировали с 5'-концом человеческой последовательности (фрагмент ~121188 п. о., выше) с применением уникальных участков распознавания рестрикционными ферментами I-CeuI и AscI, по отдельности введенных в клоны BAC как для мыши, так и для человека с помощью BHR. Данное 5'-плечо гомологии содержало 5'-последовательность мыши из генного сегмента Vλ2 мыши, который при гомологичной рекомбинации обеспечивает эффективное удаление локуса легкой λ-цепи Ig мыши. Данный нацеливающий вектор содержал ту же гомологичную перекрывающуюся 3'-последовательность, что и нацеливающий вектор 6597 (описанный выше). На фигуре 2 проиллюстрированы различные аллели, которые образованы в результате осуществления вставки нацеливающих векторов 6597 или 6680.
[00473] Сходным образом создавали дополнительную сконструированную линию мышей посредством совместной электропорации двух различных нацеливающих векторов в ES-клетки с помощью применения направляющих РНК (gRNA) с использованием системы CRISPR/Cas9 (фигура 3), см., например, патент США №9228208 (выданный 5 января 2016 года) и публикации заявок на выдачу патентов США №№U.S. 2015-0159174 A1 (поданной 15 октября 2014 года), U.S. 2015-0376650 A1 (поданной 5 июня 2015 года), U.S. 2015-0376628 A1 (поданной 23 июня 2015 года), U.S. 2016-0060657 A1 (поданной 30 октября 2015 года), U.S. 2016-0145646 A1 (поданной 20 ноября 2015 года) и U.S. 2016-0177339 A1 (поданной 18 декабря 2015 года); все из которых включены в данный документ с помощью ссылки в полном объеме. ES-клетки характеризовались геномом, гетерозиготным в отношении вставки конструкции нацеливающего вектора 6571.
[00474] Вкратце, как показано на фигуре 3, усеченный нацеливающий вектор 6596 (т.е. без 5'-плеча гомологии и кассеты, описанных выше) проектировали так, чтобы он содержал 3'-плечо гомологии размером приблизительно 33 т. н., которое содержит перекрывающуюся последовательность, соответствующую трем сегментам человеческого гена Vλ в нацеливающем векторе 6571, последовательность размером приблизительно 111 т. н., которая содержит 11 дополнительных сегментов человеческого гена Vλ, и последовательность размером приблизительно 27 т. н., которая содержит один сегмент человеческого гена Vλ и служит в качестве области перекрывания со вторым нацеливающим вектором. Второй нацеливающий вектор (нацеливающий вектор 6680) содержит ту же последовательность области перекрывания размером приблизительно 27 т. н., расположенную на 3'-конце нацеливающего вектора, последовательность размером приблизительно 94 т. н., содержащую дополнительные девять сегментов человеческого гена Vλ, кассету для отбора по устойчивости к неомицину (например, ген устойчивости к неомицину [NEOR] под транскрипционным контролем промотора убиквитина, фланкированный участками распознавания рекомбинации Frt) и 5'-плечо гомологии λ-цепи мыши размером приблизительно 22 т. н. ES-клетки, используемые при электропорации этих двух нацеливающих векторов, характеризовались геномом, гетерозиготным в отношении вставки нацеливающего вектора 6571 (фигура 3). В такие ES-клетки два нацеливающих вектора, описанных выше, вводят совместно с помощью электропорации вместе с направляющей РНК (gRNA), которая нацеливается на ген устойчивости к гигромицину из нацеливающего вектора 6571 в отношении нуклеотидной последовательности CGACCTGATG CAGCTCTCGG (SEQ ID NO: 130), и двумя gRNA, которые нацеливаются на область, расположенную против хода транскрипции относительно генного сегмента Vλ2 мыши (т.е. 3'-конца генного сегмента Vλ2 мыши на минус-нити; gRNA1: GTACATCTTG TCTTCAACGT, SEQ ID NO: 139, приблизительно 1000 п. о. против хода транскрипции относительно Vλ2 мыши; gRNA2: GTCCATAATT AATGTAGTTA C, SEQ ID NO: 140, приблизительно 380 п. о. против хода транскрипции относительно Vλ2 мыши) и обеспечивают образование двухнитевых разрывов в таких последовательностях. Два введенных совместно с помощью электропорации нацеливающих вектора вставляли с применением гомологичной рекомбинации в геном ES-клеток в последовательность устойчивости к гигромицину с заменой области, содержащий и окружающей кассету для отбора по устойчивости к гигромицину. Полученные в результате ES-клетки содержали сконструированный эндогенный локус λ-цепи Ig, который содержал человеческую вариабельную область иммуноглобулина, содержащую 25 функциональных сегментов человеческого гена Vλ, функционально связанных с кластером человеческих Jλ-Cλ, человеческим генным сегментом Jλ7 и функционально связанных с геном Cλ1 мыши (фигура 3).
[00475] Нацеливающие векторы, описанные выше, вводили в эмбриональные стволовые (ES) клетки мыши для создания сконструированного локуса легкой λ-цепи Ig. Положительные клоны ES-клеток подтверждали после вставки каждого нацеливающего вектора в геном ES-клеток (см. ниже) перед вставкой следующего нацеливающего вектора. В некоторых случаях создавали промежуточные линии для фенотипического анализа.

[00476] Нуклеотидную последовательность в месте выбранных точек соединения после осуществления вставки нацеливающих векторов, описанных выше, подтверждали с помощью секвенирования. Выбранные точки соединения, отмеченные на фигурах 1-4, представлены ниже.


[00477] При построении сконструированных локусов, в частности, сконструированных локусов иммуноглобулинов, авторы настоящего изобретения признали, что некоторые сегменты человеческого гена Vλ из некоторых гаплотипов могут отсутствовать и, следовательно, не представлены в выбранных клонах BAC, охватывающих человеческий локус легкой λ-цепи Ig. В качестве лишь одного примера, в одном докладе был приведен факт, что открытые позже аллели содержат вставку/делецию одного или более сегментов человеческого гена Vλ в кластере B человеческого локуса легкой λ-цепи Ig по сравнению с ранее описанными аллелями (например, человеческие Vλ1-50, Vλ5-48, Vλ5-45 и Vλ5-39; см. Moraes, J.C. and G.A. Passos, 2003, Immunogenetics 55(1):10-5). Таким образом, авторы настоящего изобретения разработали стратегию включения сегментов человеческого гена Vλ, которые отсутствуют в конкретном клоне BAC, применяемом для проектирования и построения нацеливающего вектора.
[00478] Вкратце, клоны BAC для человека картировали относительно человеческого локуса легкой λ-цепи Ig с помощью секвенирования концов. В частности, с применением сборки GRCh37/hg19 (UCSC Genome Browser, человек, фев. 2009 года) были идентифицированы клоны BAC для человека RP11-346I4, CTD-2523F21 и CTD-2523E22, охватывающие область кластера B человеческого локуса легкой λ-цепи Ig, которая содержит от человеческого Vλ7-46 до человеческого Vλ1-36. Например, один или более отсутствующих сегментов человеческого гена Vλ (например, Vλ5-39, Vλ5-37 и/или Vλ1-36) можно вставить в нацеливающий вектор, описанный выше (6680 или 6889), с применением любого из таких клонов BAC, которые, как было идентифицировано, содержат один или более сегментов человеческого гена Vλ, которые являются необходимыми для вставки. Например, клон BAC CTD-2523F21 модифицируют путем замены 3'-области кассетой для отбора (например, геном устойчивости к гигромицину [HYGR] под транскрипционным контролем промотора убиквитина), фланкированной участками распознавания рекомбиназами (например, lox2372), и 3'-плечом гомологии размером ~27 т. н. с последовательностью, перекрывающейся с нацеливающим вектором 6597 (см. выше). 5'-Конец клона ВАС для человека служит в качестве 5'-плеча гомологии с последовательностью, перекрывающейся с нацеливающим вектором 6680 или 6889, тем самым облегчая гомологичную рекомбинацию и осуществление вставки каких- либо отсутствующих сегментов человеческого гена Vλ наряду с кассетой для отбора. Можно применять необязательную последнюю стадию транзиентной экспрессии рекомбиназы (например, Cre) для удаления кассеты для отбора.
Пример 2. Получение грызунов со сконструированным локусом легкой λ-цепи Ig
[00479] В данном примере продемонстрировано получение отличного от человека животного (например, грызуна), геном зародышевого типа которого содержит эндогенный локус легкой λ-цепи Ig, содержащий вставку множества последовательностей человеческих Vλ, Jλ и Cλ, при этом последовательности человеческих Vλ, Jλи Cλ функционально связаны с геном (или сегментом гена) Cλ грызуна, и при этом эндогенный локус легкой λ-цепи Ig дополнительно содержит один или более энхансеров человеческой λ-цепи (Eλ) Ig. В некоторых вариантах осуществления указанный эндогенный локус легкой λ-цепи Ig предусматривает делецию одной или более эндогенных энхансерных областей (или последовательностей) легкой λ-цепи Ig. Такие отличные от человека животные характеризуются, в некоторых вариантах осуществления, экспрессией легких λ-цепей Ig, которые являются полностью человеческими (т.е. человеческими вариабельными и константными доменами).
[00480] Подлежащую нацеливанию вставку нацеливающих векторов, описанных в примере 1, подтверждали с помощью полимеразной цепной реакции. Подвергнутую нацеливанию ДНК BAC, подтвержденную полимеразной цепной реакцией, затем вводили в эмбриональные стволовые (ES) клетки мыши, представляющие собой гибрид F1 (129S6SvEvTac/C57BL6NTac), посредством электропорации с последующим культивированием в среде для отбора. В некоторых вариантах осуществления ES-клетки, применяемые для электропорации серий нацеливающих векторов, могут характеризоваться геномом зародышевого типа, который содержит локусы H-цепи Ig и κ-цепи Ig дикого типа, гомозиготные гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гомозиготный гуманизированный локус H-цепи Ig содержал вставленную последовательность, кодирующую Adam6 грызуна (см., например, патенты США №№8642835 и 8697940; таким образом включенные посредством ссылки в полном объеме), или гомозиготный гуманизированный локус H-цепи Ig (см., например, патенты США №№8642835 и 8697940, выше) и гомозиготный инактивированный локус κ-цепи Ig. В других вариантах осуществления после того, как подвергнутые целенаправленному воздействию ES-клетки, описанные в данном документе, применяют для получения мышей (см. ниже), полученных в результате мышей, содержащих сконструированный человеческий локус λ-цепи Ig, описанный в данном документе, применяют для скрещивания с мышами, содержащими гуманизированные локусы H-цепи Ig и κ-цепи Ig, при этом гуманизированный локус H-цепи Ig содержит вставленную последовательность, кодирующую Adam6 грызуна (см., например, патенты США №№8642835 и 8697940, выше), или гуманизированный локус H-цепи Ig (см., например, патенты США №№8642835 и 8697940, выше) и инактивированный локус κ-цепи Ig. Устойчивые к лекарственным средствам колонии отбирали через 10 дней после электропорации, и скрининга с помощью TAQMAN™, и кариотипирования на предмет правильного нацеливания, описанного ранее (Valenzuela et al., выше; Frendewey, D. et al., 2010, Methods Enzymol. 476:295-307). В таблице 2 изложены иллюстративные совокупности праймеров/зондов, применяемых для скрининга положительных клонов ES-клеток (F: прямой праймер; R: обратный праймер; P: зонд).
[00481] Использовали способ VELOCIMOUSE® (DeChiara, T.M. et al., 2010, Methods Enzymol. 476:285-294; DeChiara, T.M., 2009, Methods Mol. Biol. 530:311-324; Poueymirou et al., 2007, Nat. Biotechnol. 25:91-99), при котором подвергнутые целенаправленному воздействию ES-клетки вводили в эмбрионы мышей Swiss Webster на стадии неуплотненных 8 бластомеров для получения здоровых мышей поколения F0, произошедших полностью из ES-клеток, гетерозиготных в отношении аллеля сконструированного локуса легкой λ-цепи Ig. Гетерозиготную мышь поколения F0 скрещивали с мышами C57Bl6/NTac для получения гетерозигот F1, которых скрещивали между собой с получением животных поколения F2 для фенотипических анализов.
[00482] В целом, в данном примере проиллюстрировано получение грызуна (например, мыши), геном зародышевого типа которого содержит сконструированный локус легкой λ-цепи Ig, характеризующийся наличием множества последовательностей человеческих Vλ, Jλ и Cλ, функционально связанных с геном Cλ грызуна, при этом сконструированный локус легкой λ-цепи Ig грызуна содержит эндогенные энхансерные последовательности (или области) легкой λ-цепи Ig грызуна и энхансерные последовательности (или области) легкой λ-цепи Ig человека. Стратегия, описанная в данном документе, для осуществления вставки последовательностей человеческих Vλ, Jλ и Cλ в эндогенный локус легкой λ-цепи Ig грызуна делает возможным создание грызуна, который экспрессирует антитела, содержащие человеческие Vλ-домены, слитые с Cλ-доменом либо человека, либо грызуна. Как описано в данном документе, такие человеческие Vλ-домены экспрессируются с помощью эндогенных локусов легких λ-цепей Ig в геноме зародышевого типа грызуна.




Пример 3. Фенотипическая оценка грызунов со сконструированным локусом легкой λ-цепи Ig
[00483] В данном пример продемонстрирована характеристика различных популяций иммунных клеток у грызунов (например, мыши), сконструированных так, чтобы они содержали эндогенный локус легкой λ-цепи Ig, описанный выше. В частности, в данном примере конкретно продемонстрировано, что грызуны со сконструированным эндогенным локусом легкой λ-цепи Ig, описанным в данном документе, проявляют сходное развитие B-клеток при сравнении с мышами дикого типа из одного помета. В частности, каждое из нескольких сконструированных грызунов, несущих различные количества генетического материала, соответствующего человеческому локусу легкой λ-цепи Ig, на выявляемом уровне экспрессирует легкие λ-цепи Ig с человеческими вариабельными доменами и константными доменами человека или грызуна на поверхности B-клеток грызуна.
[00484] Вкратце, собирали селезенки и бедренные кости у выбранных сконструированных линий мышей, гомозиготных или гетерозиготных в отношении аллелей легких λ-цепей Ig, изображенных на фигуре 4, и у мышей дикого типа из одного помета. Костный мозг собирали из бедренных костей с помощью промывания 1x фосфатно-буферным солевым раствором (PBS, Gibco) с 2,0% фетальной бычьей сыворотки (FBS). Эритроциты из препаратов селезенки и костного мозга лизировали с помощью буфера для лизиса ACK (Gibco) с последующей промывкой с помощью 1xPBS с 2,0% FBS. Выделенные клетки (1x106) инкубировали с выбранными смесями антител в течение 30 мин. при +4°C (см. Таблицу 3).

[00485] После окрашивания клетки промывали и фиксировали в 2% формальдегиде. Получение данных осуществляли на проточном цитометре BD FORTESSA™ и анализировали с помощью программного обеспечения FLOWJO™. Репрезентативные результаты представлены на фигурах 5-13 и 18-21. Сходные данные получали для других линий, изображенных на фигуре 4, но лишь выбранные указанные линии показаны.
[00486] Из результатов видно, что каждая линия, несущая различные количества генетического материала, соответствующего человеческому локусу легкой λ-цепи Ig, демонстрировала сходные профили популяций иммунных клеток в отделах, представляющих собой селезенку и костный мозг. В частности, как очевидно из данных, показанных на фигурах 5-7, сконструированные мыши демонстрировали сходное количество CD19+ B-клеток селезенки, сходные популяции зрелых и переходных B-клеток в селезенке, сходный уровень использования каппа-цепи в селезенке и сходные популяции B-клеток краевой зоны и фолликулярных B-клеток по сравнению с их контролями дикого типа из одного помета. Кроме того, мыши, содержащие сконструированный локус легкой λ-цепи Ig, описанный в данном документе, в присутствии дополнительных гуманизированных локусов H-цепи Ig и κ-цепи Ig, демонстрируют отсутствие больших различий в развитии B-клеток по сравнению с мышами, содержащими отдельно гуманизированные локусы H-цепи Ig и κ-цепи Ig (например, см. фигуры 18A и 18B). Также, как показано у мышей, представленных на фигурах 8-12, сконструированные мыши имели сходное количество CD19+, про-, пре-, незрелых и зрелых B-клеток и сходный уровень использования каппа-цепи в костном мозгу по сравнению с их контролями дикого типа из одного помета. Краткое описание экспрессии легких цепей в выбранных сконструированных линиях (гомозиготная - HO; гетерозиготная - HET) по сравнению с их контролями дикого типа из одного помета представлено на фигуре 13. Мыши, гомозиготные в отношении сконструированного гуманизированного локуса λ-цепи Ig, описанного в данном документе, а также гомозиготные в отношении гуманизированных локусов H-цепи Ig и κ-цепи Ig и гомозиготные в отношении последовательности, кодирующей Adam6 грызуна, демонстрировали повышенное использование локуса лямбда-цепи (приблизительно 40%) по сравнению с типичным периферическим использованием (например, 5% в селезенка) лямбда-цепи, известным для мышей дикого типа (см. колонки касательно мышей 6680HO/VI HO/Adam6 HO и 6689HO/VI HO/Adam6 HO, фигура 13). Также была выявлена небольшая доля положительных в отношении λ-цепи Ig мыши B-клеток (~3-5%) у таких мышей, что подтверждает то, ген Cλ мыши в сконструированном локусе легких λ-цепей также экспрессируется в контексте функциональны легких λ-цепей у таких мышей.
Пример 4. Экспрессия антител у грызунов со сконструированным локусом легкой λ-цепи Ig
[00487] В данном примере продемонстрирована экспрессия антител у отличных от человека животных, при этом антитела содержат легкие цепи, характеризующиеся наличием человеческих Vλ-областей и Cλ-областей человека или грызуна, и при этом легкие цепи экспрессируются из сконструированного эндогенного локуса легкой λ-цепи Ig грызуна. В частности, в данном примере конкретно продемонстрирована экспрессия антител в сыворотке крови отличных от человека животных (например, грызуны), геном зародышевого типа которых содержит эндогенный локус легкой λ-цепи Ig, содержащий вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Cλ, при этом сегменты человеческих генов Vλ, Jλ и Cλ функционально связаны с геном Cλ грызуна, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи (Eλ) Ig грызуна и один или более энхансеров человеческой легкой λ-цепи (Eλ) Ig.
[00488] Вкратце, брали кровь у выбранных сконструированных линий мышей и у мышей дикого типа из одного помета (см. Пример 3). Сыворотку отделяли от крови с применением пробирок Eppendorf, центрифугированных при 9000 rcf в течение пяти минут при 4°C. Собранную сыворотку применяли для иммуноблоттинга для идентификации экспрессии легких цепей Ig антител. Сыворотку крови мыши разбавляли при 1,5:10 в PBS (без Ca2+ и Mg2+) и прогоняли на Трис-глициновых гелях Novex 4-20% в восстанавливающих и невосстанавливающих условиях. Гели переносили на поливинилидендифторидные (PVDF) мембраны в соответствии с инструкциями производителя. Блоты блокировали в течение ночи с помощью 5% обезжиренного молока в Трис-буферном солевом растворе с 0,05% Tween-20 (TBST, Sigma). PVDF-мембраны подвергали воздействию различных первичных антител (антитело мыши к hIgλ, конъюгированное с HRP (Southern Biotech); антитело козы к mIgλ, конъюгированное с HRP, Southern Biotech), разбавленных при 1:1000 в 0,1% обезжиренного молока в TBST в течение одного часа при комнатной температуре. Блоты промывали четыре раза по десять минут на промывку и проявляли в течение одной минуты с помощью реагента для выявления для вестерн-блоттинга Amersham ECL (GE Healthcare Life Sciences) в соответствии с инструкциями производителя. Затем блоты визуализировали с применением документирующей системы для гелей с камерой ImageQuant LAS-4000 Cooled CCD от GE Healthcare. Изображения получали с 15-секундными интервалами, пока не получали 20 изображений или изображения не были получены с помощью полного воздействия, в зависимости от того, что получалось первым. Репрезентативные результаты представлены на фигурах 14A и 14B. Из результатов видно, что все сконструированные линии экспрессировали на выявляемых уровнях антитела, содержащие человеческие легкие λ-цепи Ig, в их сыворотке крови (фигура 14 и данные не показаны).
Пример 5. Использование сегментов человеческих генов у грызунов со сконструированным локусом легкой λ-цепи Ig
[00489] В данном примере продемонстрировано использование человеческих генов в легких цепях антител, экспрессируемых у грызунов (например, мыши), сконструированных так, чтобы они содержали эндогенный локус легкой λ-цепи Ig, описанный выше. Использование сегментов человеческих генов λ-цепей Ig в выбранных сконструированных линиях грызунов, описанных выше, определяли с помощью анализа репертуара антител путем секвенирования нового поколения.
[00490] Вкратце, собирали спленоциты у мышей, гетерозиготных в отношении вставки 25 функциональных сегментов человеческого гена Vλ, 4 функциональных кластеров человеческих Jλ-Cλ и человеческого генного сегмента Jλ7 против хода транскрипции относительно гена Cλ1 мыши и энхансера человеческой λ-цепи Ig, вставленного между геном Cλ1 мыши и эндогенным энхансером 3.1 λ-цепи Ig мыши (гетерозиготные в отношении 6889 мыши, представленные на фигуре 4). B-клетки подвергали положительному обогащению относительно общего количества спленоцитов с помощью магнитных гранул с антителом к CD19 мыши и колонок MACS (Miltenyi Biotech). Выделяли общую РНК из B-клеток селезенки с применением набора RNeasy Plus (Qiagen).
[00491] Проводили обратную транскрипцию с помощью праймера олиго-dT с последующей геноспецифической ПЦР для получения cDNA, содержащей последовательности человеческих константных областей λ-цепи Ig (Cλ1, Cλ2, Cλ3 и Cλ6), а также cDNA, содержащей последовательность Cλ1 мыши, с применением набора для амплификации cDNA SMARTer(RACE (Clontech). Во время обратной транскрипции последовательность специфической ДНК (PIIA: 5'-CCCATGTACT CTGCGTTGAT ACCACTGCTT-3', SEQ ID NO: 133) присоединяли к 3'-концу новосинтезированных cDNA. Очищали cDNA с помощью геля NucleoSpin и набора PCR Clean-Up (Clontech), затем дополнительно амплифицировали с применением обратного праймера, комплементарного PIIA (5'-AAGCAGTGGT ATCAACGCAG AGTACAT-3'), спаренной со специфическим к человеческому Cλ праймером (5'-CACYAGTGTG GCCTTGTTGG CTTG-3', SEQ ID NO: 131) и специфическим к Cλ1 мыши праймером (5'-CACCAGTGTG GCCTTGTTAG TCTC-3', SEQ ID NO: 132).
[00492] Очищенные ампликоны затем амплифицировали с помощью ПЦР с применением специфического к PIIA праймера (5'-GTGACTGGAG TTCAGACGTG TGCTCTTCCG ATCTAAGCAG TGGTATCAAC GCAGAGT-3', SEQ ID NO: 134) и вложенного специфического к человеческому Cλ праймера (5'-ACACTCTTTC CCTACACGAC GCTCTTCCGA TCTCAGAGGA GGGCGGGAAC AGAGTG-3', SEQ ID NO: 135) или вложенного специфического к Cλ1 мыши праймера (5'-ACACTCTTTC CCTACACGAC GCTCTTCCGA TCTAAGGTGG AAACAGGGTG ACTGATG-3', SEQ ID NO: 136). Продукты ПЦР размером в диапазоне 450-690 п. о. выделяли и собирали с помощью Pippin Prep (SAGE Science). Эти фрагменты дополнительно амплифицировали с помощью ПЦР с применением следующих праймеров: 5'-AATGATACGG CGACCACCGA GATCTACACX XXXXXACACT CTTTCCCTAC ACGACGCTCT TCCGATC-3', SEQ ID NO: 137, и 5'-CAAGCAGAAG ACGGCATACG AGATXXXXXG TGACTGGAGT TCAGACGTGT GCTCTTCCGA TCT-3', SEQ ID NO: 138 («XXXXXX» представляет собой последовательности с шагом 6 п. о. для обеспечения мультиплексирования образцов для секвенирования). Продукты ПЦР размером в диапазоне 490 п. о.-710 п. о. выделяли и собирали с помощью Pippin Prep, затем количественно определяли с помощью qPCR с применением набора для количественного определения KAPA Library (KAPA Biosystems) перед загрузкой в секвенатор Miseq (Illumina) для секвенирования (v3, 600 циклов).
[00493] Для биоинформатического анализа полученные в результате последовательности с Illumina демультиплексировали и выравнивали в отношении качества. Перекрывающиеся риды парных концов затем группировали и присваивали им характеристику с применением локальной установки igblast (NCBI, v2.2.25+). Риды выравнивали относительно базы данных сегментов Vλ и Jλ зародышевого типа как человека, так и мыши и сортировали по наилучшему совпадению. Последовательность помечали как неопределенную и удаляли из анализа, если выявляли множество наилучших совпадений с одинаковыми баллами. Получали совокупность скриптов Perl для анализирования результатов и хранения данных в базе данных mysql. Репрезентативные результаты представлены на фигурах 15A (примированные человеческим Cλ) и 15B (примированные Cλ мыши).
[00494] В другом эксперименте B-клетки селезенки, собранные у мышей (n=3), гомозиготных в отношении вставки нацеливающего вектора 6889 (6889HO/VI HO/Adam6 HO, см. выше), анализировали в отношении использования сегментов человеческих генов λ-цепи Ig с помощью анализа репертуара антител путем секвенирования нового поколения (описанный выше). Выделенную из B-клеток селезенки mRNA амплифицировали с помощью 5'RACE с применением праймеров к mCλ мыши (n=3) и человеческим константным областям hCλ (n=3) и секвенировали с помощью MiSeq. Репрезентативные результаты представлены на фигурах 15C (примированные человеческим Cλ) и 15D (примированные Cλ мыши).
[00495] У мышей, полученных с применением нацеливающего вектора 6889 (т.е. совместная электропорация двух нацеливающих векторов и gRNA), показана экспрессия всех 25 функциональных сегментов человеческого гена Vλ. Кроме того, продемонстрирована экспрессия сегментов человеческого гена Vλ из B-клеток таких мышей в выделенных B-клетках со значениями частоты, сходными по сравнению с таковыми сегментов человеческого гена Vλ, наблюдаемыми в пуповинной крови человека. В целом, в данном примере продемонстрировано, что грызуны, содержащие в своих геномах зародышевого типа сконструированные локусы легких λ-цепей Ig, описанные в данном документе, экспрессируют легкие λ-цепи Ig, содержащие последовательности человеческих λ-цепей Ig, в B-клетках. Кроме того, такие последовательности человеческой λ-цепи Ig можно легко отличать в легких цепях, содержащих Cλ-домен мыши или человека.
Пример 6. Продуцирование антител в сконструированных грызунах
[00496] В данном примере продемонстрировано продуцирование антител в грызуне, который содержит сконструированный эндогенный локус легкой λ-цепи Ig, описанный выше, с применением представляющего интерес антигена (например, мембранный белок, проходящий через мембрану однократно или многократно и т.д.). При необходимости, для иммунизации грызунов, содержащих сконструированный эндогенный локус легкой λ-цепи Ig, описанный в данном документе, можно применять способы, описанные в данном примере, или способы иммунизации, хорошо известные из уровня техники, с использованием полипептидов или их фрагментов (например, пептиды, полученные из требуемого эпитопа) или комбинации полипептидов или их фрагментов.
[00497] Когорты мышей со сконструированным локусом легкой λ-цепи Ig, описанным в данном документе, и гуманизированным локусом H-цепи Ig (описанным выше) или сконструированным локусом легкой λ-цепи Ig, описанным в данном документе, и гуманизированными локусами H-цепи Ig и легкой κ-цепи Ig (описанными выше), заражали представляющим интерес антигеном с применением способов иммунизации, известных в уровне техники. Гуморальный иммунный ответ мониторили с использованием иммунологического анализа ELISA (т.e. определения титра в сыворотке). При достижении требуемого иммунного ответа собирали спленоциты (и/или другую лимфатическую ткань) и подвергали их слиянию с клетками миеломы мыши для сохранения их жизнеспособности и образования иммортализированных линий клеток гибридомы. Проводили скрининг (например, с помощью анализа ELISA) и отбор линий клеток гибридомы для идентификации линий клеток гибридомы, которые продуцировали антиген-специфические антитела. При необходимости, гибридомы можно дополнительно характеризовать в отношении относительной аффинности связывания и изотипа. С использованием этой методики и иммуногена описанного выше, получено несколько антиген-специфических химерных антител (т.e. антител, имеющих человеческие вариабельные домены и константные домены грызуна).
[00498] ДНК, кодирующую вариабельные домены тяжелой цепи и легких цепей, можно выделять и связать с требуемыми изотипами (константными доменами) тяжелой цепи и легкой цепи для получения полностью человеческих антител. Такой белок антитела может продуцироваться в клетке, такой как клетка CHO. Полностью человеческие антитела затем характеризуют в отношении относительной аффинности связывания и/или активности нейтрализации представляющего интерес антигена.
[00499] ДНК, кодирующую антиген-специфические химерные антитела или вариабельные домены легкой и тяжелой цепей, можно выделять напрямую из антиген-специфических лимфоцитов. Изначально выделяли химерные антитела с высокой аффинностью с человеческим вариабельным доменом и константным доменом грызуна и их характеризовали и подвергали отбору в отношение требуемых характеристик, включая аффинность, селективность, эпитоп и т.д. Константные домены грызуна заменяли на требуемые человеческие константные домены для получения полностью человеческих антител. Хотя выбранный константный домен может изменяться в зависимости от конкретного применения, у вариабельного домена сохраняются характеристики высокой аффинности связывания с антигеном или специфичности к мишеням. Также антиген-специфические антитела выделяли напрямую из антиген-положительных B-клеток (из иммунизированных мышей) без слияния с клетками миеломы, как описано, например, в патенте США №7582298, таким образом включенном посредством ссылки в полном объеме. С помощью данного способа получали несколько полностью человеческих антиген-специфических антител (т.e. антител, обладающих человеческими вариабельными доменами и человеческими константными доменами).
[00500] В одном эксперименте мышей 6597het (n=6) и 6680het (n=6) иммунизировали посредством введения в подушечку стопы внеклеточного домена (ECD) рецепторного полипептида с целью определения иммунного ответа у сконструированных мышей.
[00501] Вкратце, мышей примировали с помощью 2,35 мкг антигена (ECD рецепторного полипептида) плюс 10 мкг адъюванта CpG (Invivogen ODN1826). Мышам вводили семь раз бустерную инъекцию 2,35 мкг антигена (ECD рецепторного полипептида), 10 мкг адъюванта CpG и 25 мкг Adju-Phos (Brenntag). Через два дня после конечной инъекции брали кровь у выбранных сконструированных линий мышей и контролей. Сыворотку крови отделяли от крови с применением коллекторных пробирок для капиллярной крови Mictrotainer capillary (BD, номер по кат.365967) с помощью центрифугирования при 9000 rcf в течение пяти минут при 4°C. Анализы ELISA сыворотки крови проводили с определением титров общего IgG (фигура 16A), антиген-специфического IgG (фигура 16B), mIgκ (фигура 17C) mIgλ (фигура 17B) и hIgλ (фигура 17A).
[00502] В случае анализа ELISA общего IgG планшеты Maxisorp (Nunc) покрывали 1 мкг/мл антитела козы к IgG+IgM+IgA мыши H&L (Abcam) в DPBS (с Ca и Mg) на лунку и инкубировали в течение ночи при 4°C. На следующий день планшеты промывали четыре раза в PBS-T (PBS без Ca или Mg плюс 0,1%Tween-20) и блокировали в PBS-T с помощью 1% BSA в течение одного часа при комнатной температуре. Сыворотку крови в десять раз разбавляли (начиная при 1:100 и заканчивая при 1:109) в PBS-T с помощью 0,1% BSA, а стандарт IgG мыши (Sigma) разбавляли в три раза (начиная при 1 мкг/мл и заканчивая при 0,05 нг/мл) в PBS-T с помощью 0,1% BSA. Добавляли 100 мкл каждого разбавления стандарта и образца в планшет и инкубировали при комнатной температуре в течение одного часа с последующим промыванием четыре раза в PBS-T. Добавляли в каждую лунку 100 мкл антитела козы к IgG мыши, перекрестно реагирующего с человеческими иммуноглобулинами, конъюгированного с HRP (Southern Biotech), разбавленного при 1:2500, и планшеты инкубировали в течение часа. Планшеты промывали четыре раза в PBS-T и добавляли в каждую лунку 100 мкл субстратного реагента TMB (BD Biosciences) в течение десяти минут. Реакцию останавливали с помощью 100 мкл 1 н. серной кислоты на лунку и измеряли поглощение при 450 нм. Данные анализировали в GraphPad Prism и аппроксимировали с помощью четырехпараметрической кривой.
[00503] В случае антигенспецифического анализа ELISA планшеты Maxisorp (Nunc) покрывали 1 мкг/мл антигена в DPBS (с Ca и Mg) на лунку и инкубировали в течение ночи при 4°C. На следующий день планшеты промывали четыре раза в PBS-T и блокировали в Sea Block (ThermoFisher), разбавленном при 1:2 в PBS-T в течение одного часа при комнатной температуре. Сыворотку крови разбавляли (как описано выше) в Sea Block, разбавленном при 1:5 в PBS-T. Каждое разбавление образца добавляли в каждый планшет и инкубировали при комнатной температуре в течение одного часа. Затем планшеты промывали четыре раза в PBS-T. Добавляли в каждую лунку 100 мкл либо антитела козы к IgG мыши, перекрестно реагирующего с человеческим иммуноглобулином, конъюгированного с HRP (Southern Biotech), разбавленного при 1:2500, либо антитела козы к mIgκ, конъюгированного с HRP (Southern Biotech), разбавленного при 1:4000, либо антитела козы к mIgλ, конъюгированного с HRP (Southern Biotech), разбавленного при 1:4000, либо антитела козы к hIgλ, перекрестно реагирующего с иммуноглобулином мыши, конъюгированного с HRP (Southern Biotech), разбавленного при 1:4000, и планшеты инкубировали в течение одного часа. Планшеты промывали четыре раза в PBS-T и проявляли, как описано выше. Репрезентативные результаты представлены на фигурах 16 и 17.
[00504] В целом, в данном примере конкретно продемонстрировано, что у грызунов, сконструированных так, чтобы они содержали локусы легких λ-цепей Ig, описанные в данном документе, вырабатывались сильные гуморальные ответы на иммунизацию с помощью представляющего интерес антигена. Кроме того, у таких сконструированных грызунов продемонстрированы уровни общего и антиген-специфического IgG, сравнимые с контролями дикого типа, что подтверждает способность к сильному иммунному ответу у таких сконструированных животных. Действительно, титры hIgλ были выше, чем титры mIgλ после иммунизации (фигуры 17A и 17B). Таким образом, сконструированные грызуны, описанные в данном документе, обеспечивают улучшенную систему in vivo для получения антител, предназначенных для разработки терапевтических средств на основе человеческих антител, в частности терапевтических средств на основе человеческих антител, в которых используются последовательности человеческих легких λ-цепей Ig.
ЭКВИВАЛЕНТЫ
[00505] Специалистам в данной области будет понятно, что различные изменения, модификации и улучшения в отношении настоящего изобретения будут легко осуществлены специалистами в данной области. Подразумевается, что такие изменения, модификации и улучшения являются частью настоящего изобретения, и подразумевается, что они находятся в пределах сути и объема настоящего изобретения. Соответственно, вышеприведенные описание и графические материалы представлены исключительно в качестве примера, и любое открытие, описанное в настоящем изобретении, подробно описано в нижеследующей формуле изобретения.
[00506] Порядковые термины, такие как «первый», «второй», «третий» и т.д., используемые в формуле изобретения для модификации элемента формулы изобретения, сами по себе не означают какой-либо приоритет, предшествование или порядок расположения одного элемента формулы изобретения перед другим или временной порядок, в котором выполняются действия способа, но используются исключительно в качестве отметок для проведения различий между одним элементом формулы изобретения, имеющим определенное название, и другим элементом, имеющим такое же название (за исключением используемого порядкового термина), для проведения различий между элементами формулы изобретения.
[00507] Форму единственного числа в описании и формуле изобретения, если четко не указано противоположное, следует понимать как охватывающие определяемые объекты в форме множественного числа. Признаки, раскрытые в пунктах формулы изобретения или частях описания, в которых содержится «или» между одним или более членами группы, считаются охваченными, если один, более одного или все члены группы присутствуют в указанном продукте или способе, используются в них или иным образом имеют отношение к ним, если не указано противоположное или иное не очевидно из контекста. Настоящее изобретение охватывает варианты осуществления, в которых только один член группы присутствует в указанном продукте или способе, используется в них или иным образом имеет отношение к ним. Настоящее изобретение также охватывает варианты осуществления, в которых более одного или все члены группы присутствуют в указанном продукте или способе, используются в них или иным образом имеют отношение к ним. Кроме того, следует понимать, что настоящее изобретение охватывает все варианты, комбинации и перестановки, в которых одно или более ограничений, элементов, условий, описательных терминов и т.д. из одного или более перечисленных пунктов формулы изобретения входят в другой пункт формулы изобретения, зависимый от того же основного пункта формулы изобретения (или, в зависимости от того, что применимо, любого другого пункта формулы изобретения), если не указано иное или если специалисту в данной области не будет очевидным возникновение противоречия или несоответствия. В тех случаях, когда элементы представлены в виде перечней (например, в виде группы Маркуша или в аналогичном формате), следует понимать, что также раскрыта каждая подгруппа элементов, и любой(-ые) элемент(-ы) можно исключить из группы. Следует понимать, что, как правило, в тех случаях, когда настоящее изобретение или аспекты настоящего изобретения рассматриваются как включающие конкретные элементы, признаки и т.д., определенные варианты осуществления настоящего изобретения или аспектов настоящего изобретения состоят из таких элементов, признаков и т.д. или состоят главным образом из них. В целях простоты эти варианты осуществления не были в каждом случае специально недвусмысленно изложены в данном документе. Также следует понимать, что любой вариант осуществления или аспект настоящего изобретения может быть явным образом исключен из формулы изобретения независимо от упоминания конкретного исключения в описании.
[00508] Специалистам в данной области будут понятны типичные среднеквадратические отклонения или ошибки, присущие значениям, получаемым в анализах или других способах, описанных в данном документе. Публикации, веб-сайты и другие справочные материалы, упоминаемые в данном документе для описания предпосылок настоящего изобретения и для предоставления дополнительных подробностей, относящихся к его практическому осуществлению, настоящим включены в данный документ посредством ссылки в полном объеме
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> REGENERON PHARMACEUTICALS, INC.
<120> ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ СО СКОНСТРУИРОВАННЫМ ЛОКУСОМ
ЛЕГКОЙ ЛЯМБДА-ЦЕПИ ИММУНОГЛОБУЛИНА
<130> RPB-01825
<140> PCT/US2017/060006
<141> 2017-11-03
<150> 62/567,932
<151> 2017-10-04
<150> 62/417,845
<151> 2016-11-04
<160> 141
<170> PatentIn версия 3.5
<210> 1
<211> 317
<212> ДНК
<213> Mus musculus
<400> 1
gccagcccaa gtcttcgcca tcagtcaccc tgtttccacc ttcctctgaa gagctcgaga 60
ctaacaaggc cacactggtg tgtacgatca ctgatttcta cccaggtgtg gtgacagtgg 120
actggaaggt agatggtacc cctgtcactc agggtatgga gacaacccag ccttccaaac 180
agagcaacaa caagtacatg gctagcagct acctgaccct gacagcaaga gcatgggaaa 240
ggcatagcag ttacagctgc caggtcactc atgaaggtca cactgtggag aagagtttgt 300
cccgtgctga ctgttcc 317
<210> 2
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 2
Gly Gln Pro Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp
20 25 30
Phe Tyr Pro Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro
35 40 45
Val Thr Gln Gly Met Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Met Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu
65 70 75 80
Arg His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val
85 90 95
Glu Lys Ser Leu Ser Arg Ala Asp Cys Ser
100 105
<210> 3
<211> 314
<212> ДНК
<213> Mus musculus
<400> 3
gtcagcccaa gtccactccc actctcaccg tgtttccacc ttcctctgag gagctcaagg 60
aaaacaaagc cacactggtg tgtctgattt ccaacttttc cccgagtggt gtgacagtgg 120
cctggaaggc aaatggtaca cctatcaccc agggtgtgga cacttcaaat cccaccaaag 180
agggcaacaa gttcatggcc agcagcttcc tacatttgac atcggaccag tggagatctc 240
acaacagttt tacctgtcaa gttacacatg aaggggacac tgtggagaag agtctgtctc 300
ctgcagaatg tctc 314
<210> 4
<211> 105
<212> Белок
<213> Mus musculus
<400> 4
Gly Gln Pro Lys Ser Thr Pro Thr Leu Thr Val Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Lys Glu Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asn
20 25 30
Phe Ser Pro Ser Gly Val Thr Val Ala Trp Lys Ala Asn Gly Thr Pro
35 40 45
Ile Thr Gln Gly Val Asp Thr Ser Asn Pro Thr Lys Glu Gly Asn Lys
50 55 60
Phe Met Ala Ser Ser Phe Leu His Leu Thr Ser Asp Gln Trp Arg Ser
65 70 75 80
His Asn Ser Phe Thr Cys Gln Val Thr His Glu Gly Asp Thr Val Glu
85 90 95
Lys Ser Leu Ser Pro Ala Glu Cys Leu
100 105
<210> 5
<211> 314
<212> ДНК
<213> Mus musculus
<400> 5
gtcagcccaa gtccactccc acactcacca tgtttccacc ttcccctgag gagctccagg 60
aaaacaaagc cacactcgtg tgtctgattt ccaatttttc cccaagtggt gtgacagtgg 120
cctggaaggc aaatggtaca cctatcaccc agggtgtgga cacttcaaat cccaccaaag 180
aggacaacaa gtacatggcc agcagcttct tacatttgac atcggaccag tggagatctc 240
acaacagttt tacctgccaa gttacacatg aaggggacac tgtggagaag agtctgtctc 300
ctgcagaatg tctc 314
<210> 6
<211> 105
<212> Белок
<213> Mus musculus
<400> 6
Gly Gln Pro Lys Ser Thr Pro Thr Leu Thr Met Phe Pro Pro Ser Pro
1 5 10 15
Glu Glu Leu Gln Glu Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asn
20 25 30
Phe Ser Pro Ser Gly Val Thr Val Ala Trp Lys Ala Asn Gly Thr Pro
35 40 45
Ile Thr Gln Gly Val Asp Thr Ser Asn Pro Thr Lys Glu Asp Asn Lys
50 55 60
Tyr Met Ala Ser Ser Phe Leu His Leu Thr Ser Asp Gln Trp Arg Ser
65 70 75 80
His Asn Ser Phe Thr Cys Gln Val Thr His Glu Gly Asp Thr Val Glu
85 90 95
Lys Ser Leu Ser Pro Ala Glu Cys Leu
100 105
<210> 7
<211> 314
<212> ДНК
<213> Rattus norvegicus
<400> 7
gtcagcccaa gtccactccc acactcacag tatttccacc ttcaactgag gagctccagg 60
gaaacaaagc cacactggtg tgtctgattt ctgatttcta cccgagtgat gtggaagtgg 120
cctggaaggc aaatggtgca cctatctccc agggtgtgga cactgcaaat cccaccaaac 180
agggcaacaa atacatcgcc agcagcttct tacgtttgac agcagaacag tggagatctc 240
gcaacagttt tacctgccaa gttacacatg aagggaacac tgtggagaag agtctgtctc 300
ctgcagaatg tgtc 314
<210> 8
<211> 105
<212> Белок
<213> Rattus norvegicus
<400> 8
Gly Gln Pro Lys Ser Thr Pro Thr Leu Thr Val Phe Pro Pro Ser Thr
1 5 10 15
Glu Glu Leu Gln Gly Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Ser Asp Val Glu Val Ala Trp Lys Ala Asn Gly Ala Pro
35 40 45
Ile Ser Gln Gly Val Asp Thr Ala Asn Pro Thr Lys Gln Gly Asn Lys
50 55 60
Tyr Ile Ala Ser Ser Phe Leu Arg Leu Thr Ala Glu Gln Trp Arg Ser
65 70 75 80
Arg Asn Ser Phe Thr Cys Gln Val Thr His Glu Gly Asn Thr Val Glu
85 90 95
Lys Ser Leu Ser Pro Ala Glu Cys Val
100 105
<210> 9
<211> 314
<212> ДНК
<213> Rattus norvegicus
<400> 9
accaacccaa ggctacgccc tcagtcaccc tgttcccacc ttcctctgaa gagctcaaga 60
ctgacaaggc tacactggtg tgtatggtga cagatttcta ccctggtgtt atgacagtgg 120
tctggaaggc agatggtacc cctatcactc agggtgtgga gactacccag cctttcaaac 180
agaacaacaa gtacatggct accagctacc tgcttttgac agcaaaagca tgggagactc 240
atagcaatta cagctgccag gtcactcacg aagagaacac tgtggagaag agtttgtccc 300
gtgctgagtg ttcc 314
<210> 10
<211> 105
<212> Белок
<213> Rattus norvegicus
<400> 10
Asp Gln Pro Lys Ala Thr Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Lys Thr Asp Lys Ala Thr Leu Val Cys Met Val Thr Asp
20 25 30
Phe Tyr Pro Gly Val Met Thr Val Val Trp Lys Ala Asp Gly Thr Pro
35 40 45
Ile Thr Gln Gly Val Glu Thr Thr Gln Pro Phe Lys Gln Asn Asn Lys
50 55 60
Tyr Met Ala Thr Ser Tyr Leu Leu Leu Thr Ala Lys Ala Trp Glu Thr
65 70 75 80
His Ser Asn Tyr Ser Cys Gln Val Thr His Glu Glu Asn Thr Val Glu
85 90 95
Lys Ser Leu Ser Arg Ala Glu Cys Ser
100 105
<210> 11
<211> 314
<212> ДНК
<213> Rattus norvegicus
<400> 11
gtcagcccaa gtccactccc acactcacag tatttccacc ttcaactgag gagctccagg 60
gaaacaaagc cacactggtg tgtctgattt ctgatttcta cccgagtgat gtggaagtgg 120
cctggaaggc aaatggtgca cctatctccc agggtgtgga cactgcaaat cccaccaaac 180
agggcaacaa atacatcgcc agcagcttct tacgtttgac agcagaacag tggagatctc 240
gcaacagttt tacctgccaa gttacacatg aagggaacac tgtggaaaag agtctgtctc 300
ctgcagagtg tgtc 314
<210> 12
<211> 105
<212> Белок
<213> Rattus norvegicus
<400> 12
Gly Gln Pro Lys Ser Thr Pro Thr Leu Thr Val Phe Pro Pro Ser Thr
1 5 10 15
Glu Glu Leu Gln Gly Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Ser Asp Val Glu Val Ala Trp Lys Ala Asn Gly Ala Pro
35 40 45
Ile Ser Gln Gly Val Asp Thr Ala Asn Pro Thr Lys Gln Gly Asn Lys
50 55 60
Tyr Ile Ala Ser Ser Phe Leu Arg Leu Thr Ala Glu Gln Trp Arg Ser
65 70 75 80
Arg Asn Ser Phe Thr Cys Gln Val Thr His Glu Gly Asn Thr Val Glu
85 90 95
Lys Ser Leu Ser Pro Ala Glu Cys Val
100 105
<210> 13
<211> 314
<212> ДНК
<213> Rattus norvegicus
<400> 13
accaacccaa ggctacgccc tcagtcaccc tgttcccacc ttcctctgaa gagctcaaga 60
ctgacaaggc tacactggtg tgtatggtga cagatttcta ccctggtgtt atgacagtgg 120
tctggaaggc agatggtacc cctatcactc agggtgtgga gactacccag cctttcaaac 180
agaacaacaa gtacatggct accagctacc tgcttttgac agcaaaagca tgggagactc 240
atagcaatta cagctgccag gtcactcacg aagagaacac tgtggagaag agtttgtccc 300
gtgctgagtg ttcc 314
<210> 14
<211> 105
<212> Белок
<213> Rattus norvegicus
<400> 14
Asp Gln Pro Lys Ala Thr Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Lys Thr Asp Lys Ala Thr Leu Val Cys Met Val Thr Asp
20 25 30
Phe Tyr Pro Gly Val Met Thr Val Val Trp Lys Ala Asp Gly Thr Pro
35 40 45
Ile Thr Gln Gly Val Glu Thr Thr Gln Pro Phe Lys Gln Asn Asn Lys
50 55 60
Tyr Met Ala Thr Ser Tyr Leu Leu Leu Thr Ala Lys Ala Trp Glu Thr
65 70 75 80
His Ser Asn Tyr Ser Cys Gln Val Thr His Glu Glu Asn Thr Val Glu
85 90 95
Lys Ser Leu Ser Arg Ala Glu Cys Ser
100 105
<210> 15
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 15
gcatggccta gagataacaa gac 23
<210> 16
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 16
ggccttggat aacctcagga tac 23
<210> 17
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 17
tccatcccaa tagatctcat tccttccc 28
<210> 18
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 18
ccctgtcaag tctccaaggt tg 22
<210> 19
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 19
cactgtggcc caaggatcac 20
<210> 20
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 20
cactctgccc agggagtgtc tgg 23
<210> 21
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 21
gcatggccta gagataacaa gactg 25
<210> 22
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 22
gtgctcttcc cttgggaga 19
<210> 23
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 23
tccatcccaa tagagcgatc gca 23
<210> 24
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 24
ggtggagagg ctattcggc 19
<210> 25
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 25
gaacacggcg gcatcag 17
<210> 26
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 26
tgggcacaac agacaatcgg ctg 23
<210> 27
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 27
agctgaatgg aaacaaggca a 21
<210> 28
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 28
ggagacaatg ccccagtga 19
<210> 29
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 29
tgacatgaac catctgtttc tctctcgaca a 31
<210> 30
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 30
ccaccgccaa gttgacctc 19
<210> 31
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 31
tgaaggacta aggcccagga tag 23
<210> 32
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 32
agtacagcaa gggcccagcc t 21
<210> 33
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 33
tggctcagtg acaagagtc 19
<210> 34
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 34
ccagggacac agcctttgc 19
<210> 35
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 35
tgcattgcag agaccaggga cc 22
<210> 36
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 36
tgcggccgat cttagcc 17
<210> 37
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 37
acgagcgggt tcggcccatt c 21
<210> 38
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 38
ttgaccgatt ccttgcgg 18
<210> 39
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 39
tgtcgggcgt acacaaatcg 20
<210> 40
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 40
gggcgtcggt ttccactatc 20
<210> 41
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 41
ccgtctggac cgatggctgt gt 22
<210> 42
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 42
cgacgtctgt cgagaagttt ctg 23
<210> 43
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 43
cacgccctcc tacatcgaa 19
<210> 44
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 44
agttcgacag cgtgtccgac ctga 24
<210> 45
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 45
aacaaccgag ctccaggtgt 20
<210> 46
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 46
agggcagcct tgtctccaa 19
<210> 47
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 47
cctgccagat tctcaggctc cctg 24
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 48
ggaggtcagg aatgagggac 20
<210> 49
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 49
cacttgctca ctgcaaaagc a 21
<210> 50
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 50
tgtgggattt tggaattcta tctcactgat aggaaag 37
<210> 51
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 51
gcagagagga ttcaagagct gg 22
<210> 52
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 52
tttttgcaat gcttcacctg a 21
<210> 53
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 53
caggtgtctg tattggaggt caatggca 28
<210> 54
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 54
gatttgctga gggcagggt 19
<210> 55
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 55
ccccaagtct gatccttcct t 21
<210> 56
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 56
ccttcatact cttgcatcct cccttctcca 30
<210> 57
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 57
gctgaccaac gatcgcctaa 20
<210> 58
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 58
taagcgccac actgcacct 19
<210> 59
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 59
ttccttctct tctgtgactc aattatttgt ggaca 35
<210> 60
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 60
aactgctgat gcactgggc 19
<210> 61
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 61
tgaatgcatg gagttggcc 19
<210> 62
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 62
tctcctttgc agtggcttaa ttagctgagt ca 32
<210> 63
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 63
ccctggtgaa gcatgtttgc 20
<210> 64
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 64
tgtggcctgt ctgccttacg 20
<210> 65
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 65
ccaagcagga ggtgctcagt tcccaa 26
<210> 66
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 66
gggacaggtg aagggcctat c 21
<210> 67
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 67
tggtccacag gatgcagttg 20
<210> 68
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 68
cgcacctgta tctaaccagt cccagcatc 29
<210> 69
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 69
cacacctaga ccccggaagt c 21
<210> 70
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 70
tcgctttgcc agttgattct c 21
<210> 71
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 71
tccacactgt cggctgggag ctca 24
<210> 72
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 72
cgcttcaatg acccaacca 19
<210> 73
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 73
tgttgaaacg taatccccaa tg 22
<210> 74
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 74
ctcccaccag gtgccacatg ca 22
<210> 75
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 75
gggctacttg aggaccttgc t 21
<210> 76
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 76
gacagccctt acagagtttg gaa 23
<210> 77
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 77
cagggcctcc atcccaggca 20
<210> 78
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 78
agtgcaaaca gcaagatgag atct 24
<210> 79
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 79
ggcgctgagc agaaaacaa 19
<210> 80
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 80
agaccaccaa gaaggcccag agtgacc 27
<210> 81
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 81
aagaccagga gctctgccta agt 23
<210> 82
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 82
cccatcacga actgaagttg ag 22
<210> 83
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 83
ccccagtgtg tgaatcactc taccctcc 28
<210> 84
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 84
cccttcatga tgctttgtca tc 22
<210> 85
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 85
gtagtggcaa aggcagattc ct 22
<210> 86
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 86
ccttcactcc ccgaatgccc tcc 23
<210> 87
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 87
gccctgctcc agtcttattc c 21
<210> 88
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 88
ctgcgtctgg gctttgct 18
<210> 89
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 89
ccacagatcc caagttgagc ctgc 24
<210> 90
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 90
gtgagcggta ccctggaatc 20
<210> 91
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 91
agcctcgtct tcggtcagga c 21
<210> 92
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 92
tgaacgattc tctgggtcca cc 22
<210> 93
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 93
cctgagccag gatggaatga ag 22
<210> 94
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 94
ggccgtgatt taagaggttg ttag 24
<210> 95
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 95
actgtggacc ccagataatt cccctg 26
<210> 96
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 96
gagtgcagtg gcagaatctt g 21
<210> 97
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 97
ggcagggagc attggtaga 19
<210> 98
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 98
tactgaaatc tcagcctccc aggc 24
<210> 99
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 99
tggctccagc tcaggaaav 19
<210> 100
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 100
cccgggagtt acagtaatag tca 23
<210> 101
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 101
cacagcttcc ttgaccatca ctggg 25
<210> 102
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 102
ccagcccacc caattatgct a 21
<210> 103
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 103
gcgtttaggg ccaggtacaa at 22
<210> 104
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 104
tggatctgtc aaacactttc agagca 26
<210> 105
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 105
gaggctgcag ggatgtaac 19
<210> 106
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 106
cccattccag gtccaattct ca 22
<210> 107
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 107
tttgtaaagt gcataacaca gaccctga 28
<210> 108
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 108
gggtacaatg agacaagaat caga 24
<210> 109
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 109
gaaaggcaaa cacaagtcac agatg 25
<210> 110
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 110
tcagccctct ggaatgtaag gatca 25
<210> 111
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 111
gctgcatctt ctcaagtctt taagt 25
<210> 112
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 112
gggaaccagt caggaactca tac 23
<210> 113
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 113
taagcagacc tatgcatcgc tca 23
<210> 114
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 114
gtgctccttg ttcccttcac ag 22
<210> 115
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 115
ctgaagcatc tgcaccatca aatc 24
<210> 116
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
зонд"
<400> 116
ccacccacat gtgcccgtgt g 21
<210> 117
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 117
ccctattcac tgagttctgg aagctctgct atttccatga tcgttcacac tgacccctgt 60
tgatcttacc ggtaccgaag ttcctattcc gaagttccta 100
<210> 118
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 118
ttctctagaa agtataggaa cttcctaggg tttcaccggt ggcgcgccga tgtacatcag 60
ttcagtctgg aaaggtggaa cagctccagg tgaaggcagg 100
<210> 119
<211> 150
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 119
ctctacgggt gatgttcatc taaggtgaca ggagtcagtg agggcttctc aagctttatc 60
tatgtcgggt gcggagaaag aggtaatgaa atggcactcg agccctgctg gtgccttctg 120
ttgtatccac gccttcagta gatttgatga 150
<210> 120
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 120
gagtttttcc ctttcctgtc tgtcgaaggc taaggtctaa gcctgtctgg tcacactagg 60
taaagaattt ctttcttctc tagatgcttt gtctcatttc 100
<210> 121
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 121
tatgtcactg gaatttagag tagtgtgtgg aatgtcttgg caacctggac acgcgtcctg 60
gcacccagtg agaaagtggc cctgagggag aggctcatag 100
<210> 122
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 122
agcagccgac atttagcaaa gaggattgga aaatgaaccc ccccttaaaa tacagttaaa 60
cacagaggag ggagcaaacc ggtataactt cgtataatgt 100
<210> 123
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 123
atgctatacg aagttatgtc gacctcgagg gggggcccgg taccatctat gtcgggtgcg 60
gagaaagagg taatgaaatg gtctcattcc ttccctgtct caaggcataa tggttcaata 120
tgcacctgta 130
<210> 124
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 124
ttctctccaa gacttgaggt gctttttgtt gtatactttc cctttctgta ttctgcttca 60
tacctatact ggtaccgaag ttcctattcc gaagttccta 100
<210> 125
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 125
ttctctagaa agtataggaa cttcctaggg tttcaccggt ggcgcgcctg ccatttcatt 60
acctctttct ccgcacccga catagataag ctttggattg gattcagtga gcaagaattc 120
acaaacacaa tggacttatc 140
<210> 126
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 126
ttctctccaa gacttgaggt gctttttgtt gtatactttc cctttctgta ttctgcttca 60
tacctatact ggtaccgaag ttcctattcc gaagttccta 100
<210> 127
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 127
ttctctagaa agtataggaa cttcctaggg tttcaccggt ggcgcgcccc cctgctggtg 60
ccttttgttg tatccacgcc ttcagtagat ttgatgatgc 100
<210> 128
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 128
ttctctccaa gacttgaggt gctttttgtt gtatactttc cctttctgta ttctgcttca 60
tacctatact ggtaccgaag ttcctattcc gaagttccta 100
<210> 129
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 129
ttctctagaa agtataggaa cttcctaggg tttcaccggt ggcgcgccga tgtacatcag 60
ttcagtctgg aaaggtggaa cagctccagg tgaaggcagg 100
<210> 130
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 130
cgacctgatg cagctctcgg 20
<210> 131
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 131
cacyagtgtg gccttgttgg cttg 24
<210> 132
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 132
caccagtgtg gccttgttag tctc 24
<210> 133
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 133
cccatgtact ctgcgttgat accactgctt 30
<210> 134
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 134
gtgactggag ttcagacgtg tgctcttccg atctaagcag tggtatcaac gcagagt 57
<210> 135
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 135
acactctttc cctacacgac gctcttccga tctcagagga gggcgggaac agagtg 56
<210> 136
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 136
acactctttc cctacacgac gctcttccga tctaaggtgg aaacagggtg actgatg 57
<210> 137
<211> 67
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<220>
<221> модифицированное основание
<222> (30)..(35)
<223> a, c, t, g, неизвестное или другое
<400> 137
aatgatacgg cgaccaccga gatctacacn nnnnnacact ctttccctac acgacgctct 60
tccgatc 67
<210> 138
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<220>
<221> модифицированное основание
<222> (25)..(29)
<223> a, c, t, g, неизвестное или другое
<400> 138
caagcagaag acggcatacg agatnnnnng tgactggagt tcagacgtgt gctcttccga 60
tct 63
<210> 139
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 139
gtacatcttg tcttcaacgt 20
<210> 140
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 140
gtccataatt aatgtagtta c 21
<210> 141
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 141
aagcagtggt atcaacgcag agtacat 27
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ λ ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2011 |
|
RU2724736C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ГИБРИДНУЮ ЛЕГКУЮ ЦЕПЬ ИММУНОГЛОБУЛИНА | 2011 |
|
RU2601297C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ АЛЬФА ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2011 |
|
RU2590594C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ ДЛЯ ТЕСТИРОВАНИЯ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ | 2019 |
|
RU2805212C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ λ ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2020 |
|
RU2833566C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2664185C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2770016C2 |
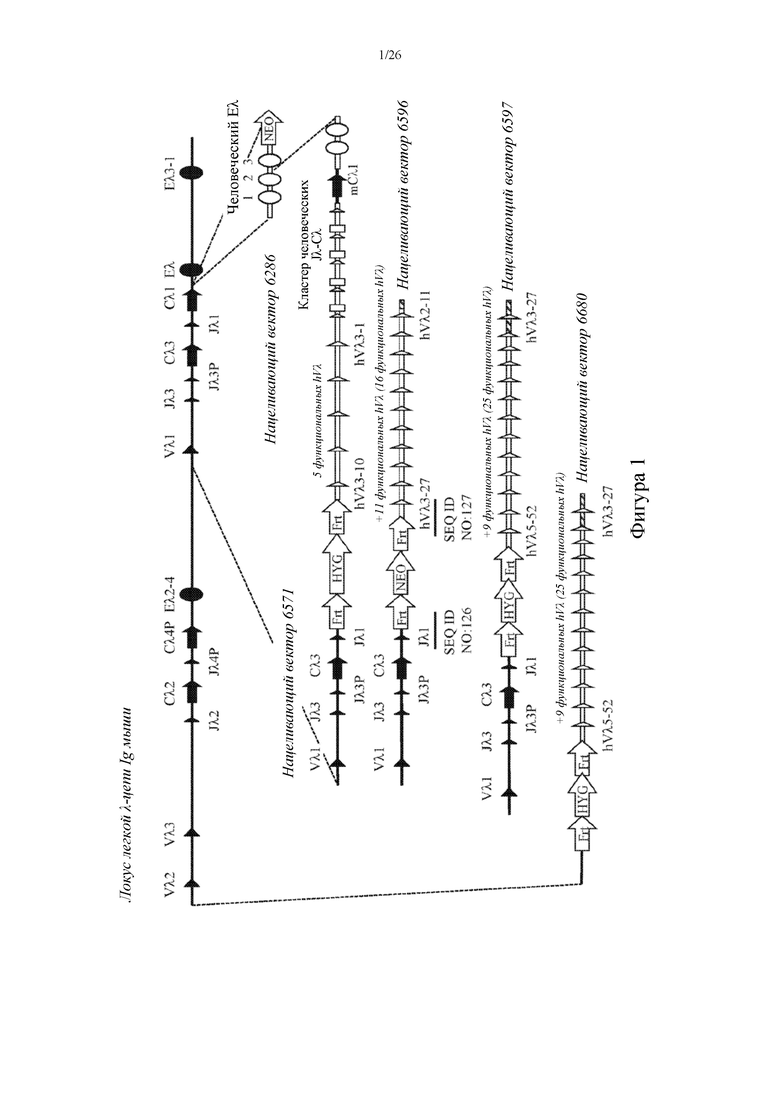

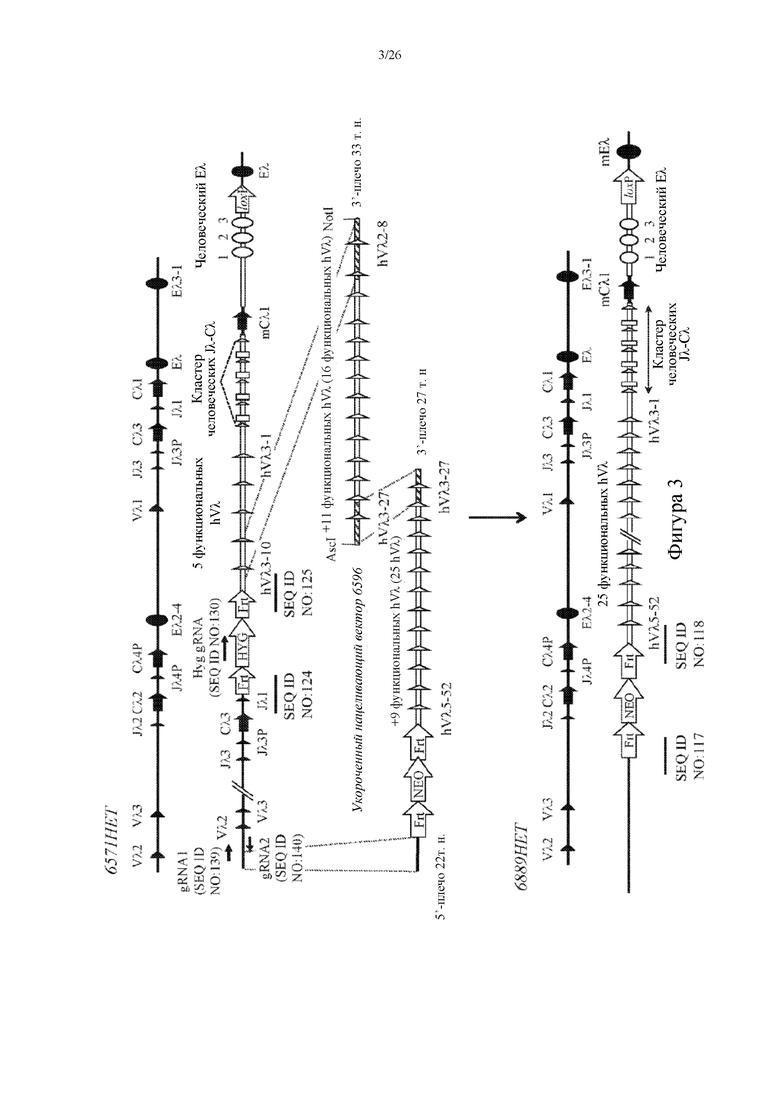


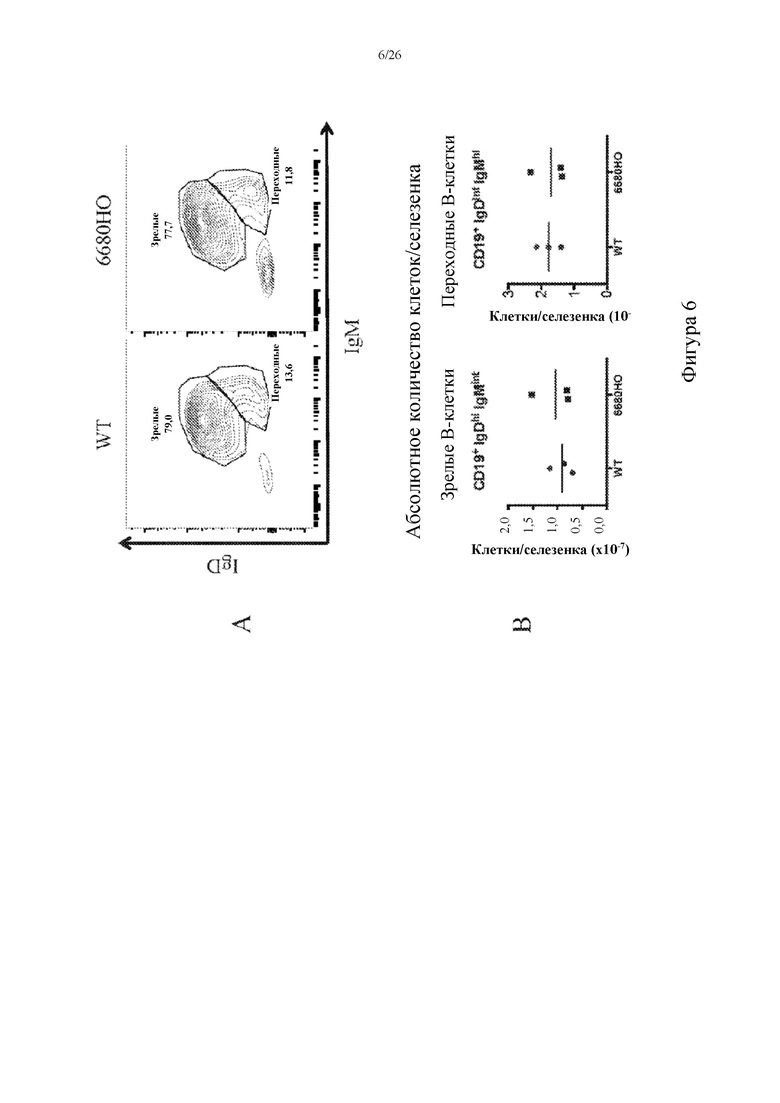



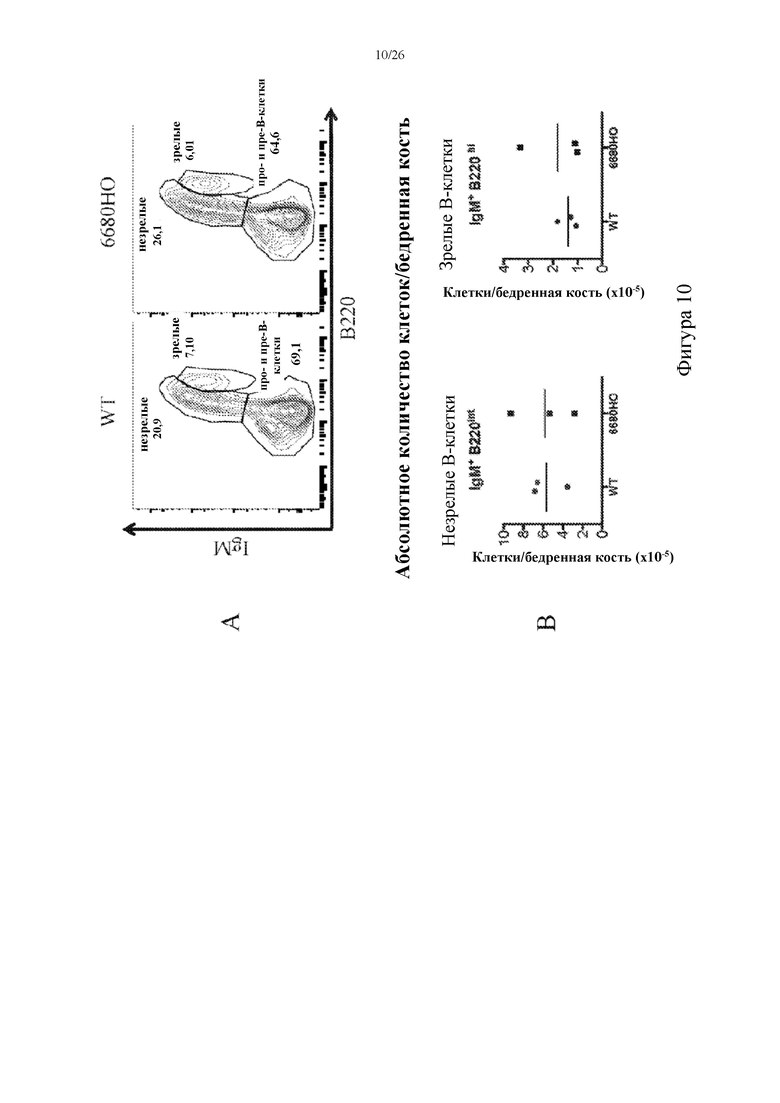


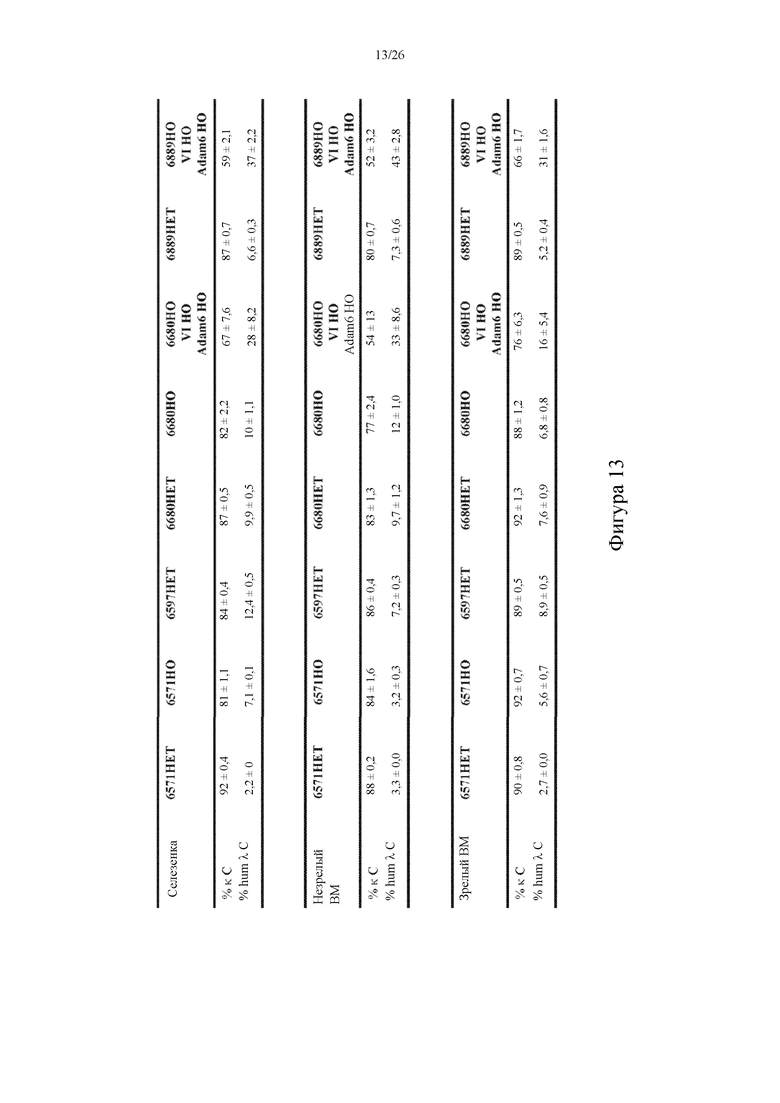


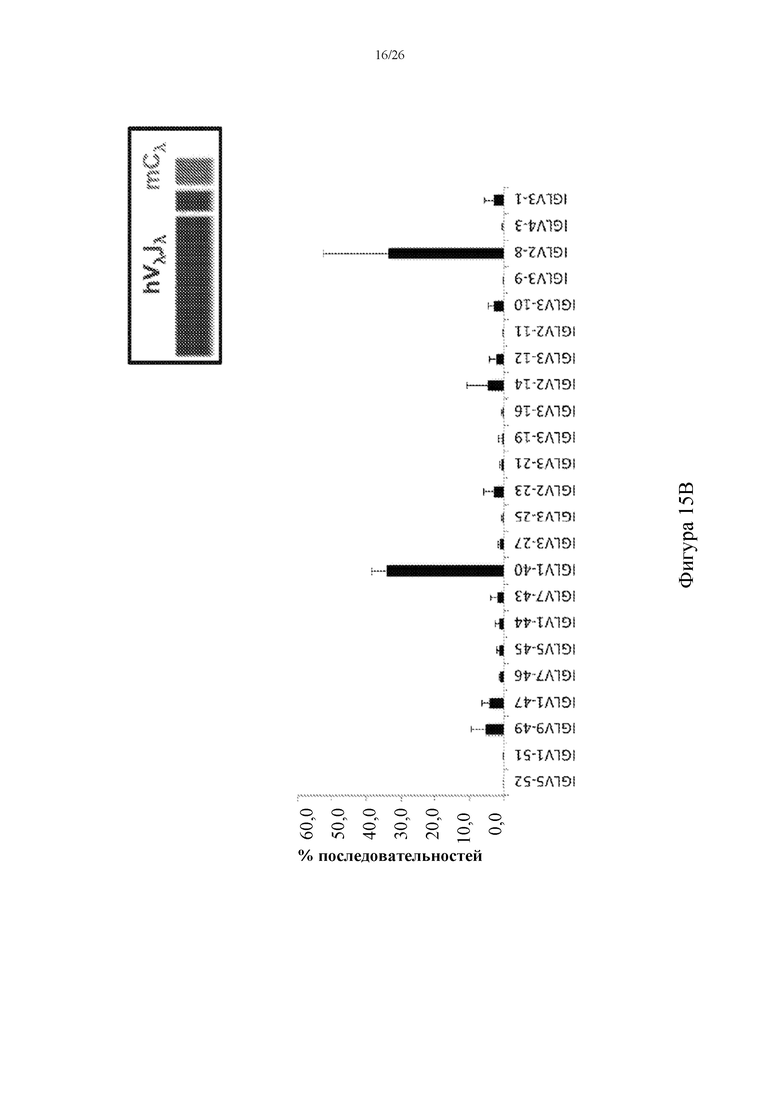


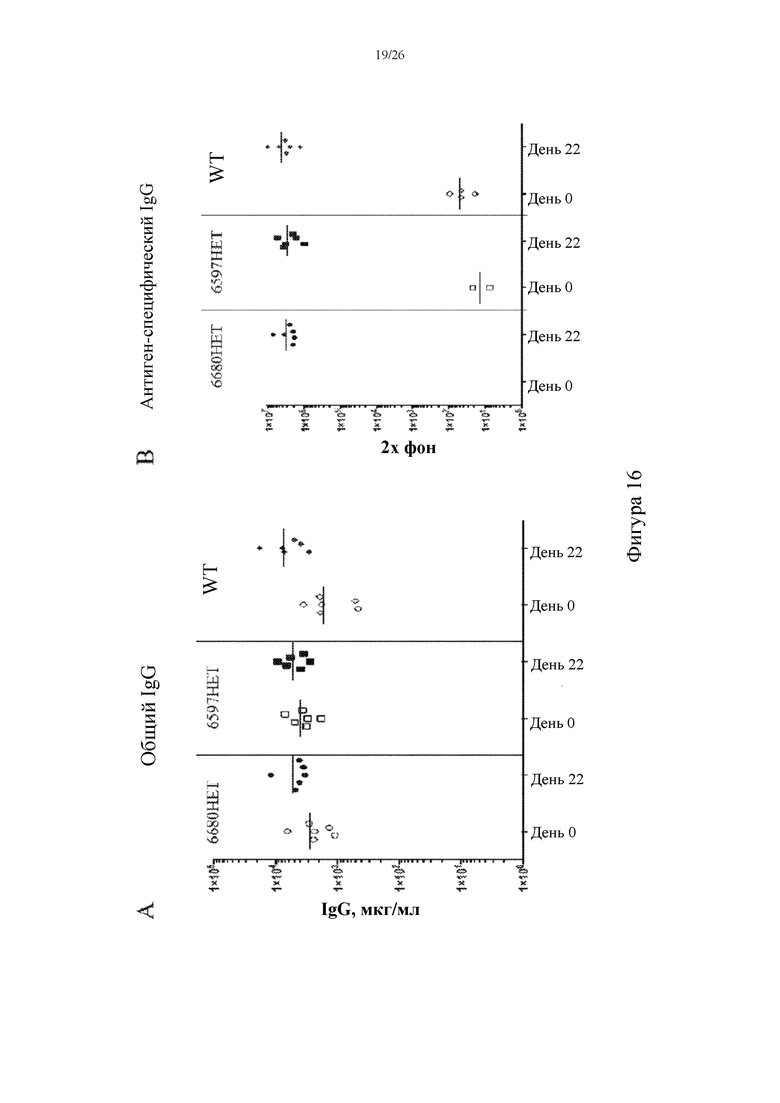
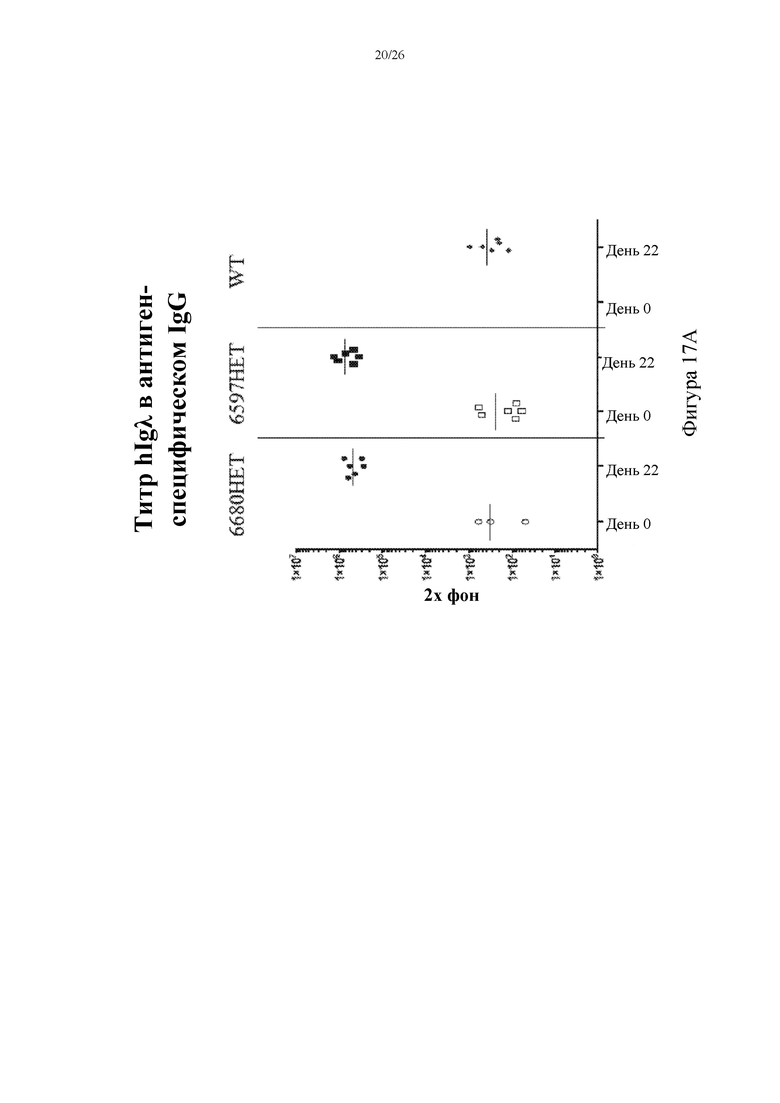

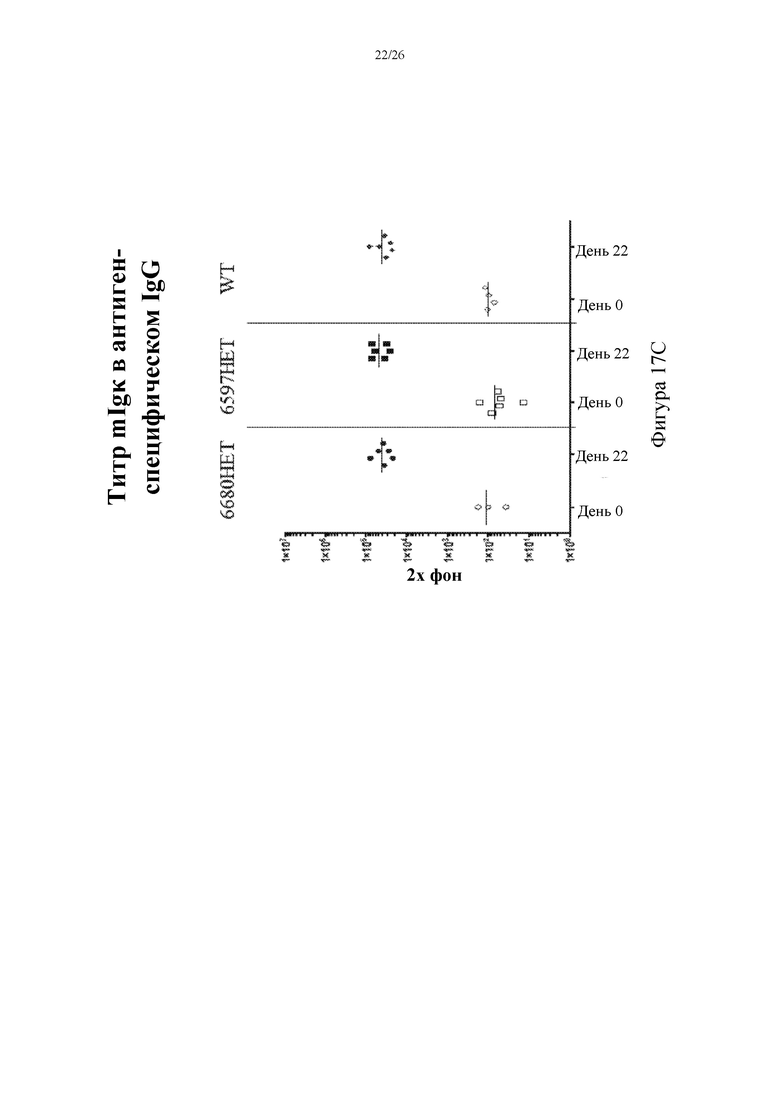




Изобретение относится к области биотехнологии, в частности, к мыши, геном зародышевого типа которой содержит эндогенный локус легкой λ цепи иммуноглобулина, содержащий: один или более сегментов человеческого гена Vλ, один или более сегментов человеческого гена Jλ и один или более сегментов человеческого гена Сλ, а также к способу ее получения. Изобретение эффективно для получения антитела. 3 н. и 18 з.п. ф-лы, 21 ил., 4 табл., 6 пр.
1. Мышь для получения антитела, геном зародышевого типа которой содержит эндогенный локус легкой λ цепи иммуноглобулина, содержащий:
(a) один или более сегментов человеческого гена Vλ,
(b) один или более сегментов человеческого гена Jλ и
(c) один или более сегментов человеческого гена Сλ,
где (а) и (b) функционально связаны с (с) и с сегментом гена Сλ мыши, и
где эндогенный локус легкой λ цепи иммуноглобулина дополнительно содержит один или более энхансеров мышиной легкой λ-цепи (Еλ) иммуноглобулина и один или более энхансеров человеческой легкой λ-цепи (Еλ) иммуноглобулина, и причем мышь экспрессирует легкую λ-цепь иммуноглобулина, содержащую человеческий вариабельный λ-домен и либо человеческий константный λ-домен, либо мышиный константный λ-домен.
2. Мышь по п. 1, где эндогенный локус легкой λ цепи иммуноглобулина содержит два мышиных Eλ.
3. Мышь по п. 2, где два мышиных Eλ представляют собой Еλ мыши и Еλ3-1 мыши.
4. Мышь по п. 1, где эндогенный локус легкой λ цепи иммуноглобулина содержит три человеческих Eλ.
5. Мышь по п. 1, где геном зародышевого типа дополнительно содержит
(i) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с мышиной константной областью тяжелой цепи иммуноглобулина; или
(ii) эндогенный локус тяжелой цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH, при этом сегменты человеческих генов VH, DH и JH функционально связаны с мышиной константной областью тяжелой цепи иммуноглобулина, и эндогенный локус легкой к цепи иммуноглобулина, содержащий вставку одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ, при этом сегменты человеческих генов Vκ и Jκ функционально связаны с мышиной Сκ-областью иммуноглобулина.
6. Мышь по п. 5, где вставка одного или более сегментов человеческого гена VH, одного или более сегментов человеческого гена DH и одного или более сегментов человеческого гена JH обеспечивает замену сегментов мышиных генов VH, DH.
7. Мышь по п. 6, где вставка содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов VH, DH и JH и их комбинациями.
8. Мышь по п. 5, где вставка одного или более сегментов человеческого гена Vκ и одного или более сегментов человеческого гена Jκ обеспечивает замену сегментов мышиных генов Vκ и Jκ.
9. Мышь по п. 8, где вставка содержит некодирующую ДНК человека, которая встречается в природе между сегментами человеческих генов Vκ и Jκ и их комбинациями.
10. Мышь по п. 5, где мышиная константная область тяжелой цепи иммуноглобулина представляет собой эндогенную мышиную константную область тяжелой цепи иммуноглобулина.
11. Мышь по п. 5, где мышиная Сκ-область представляет собой эндогенную мышиную Сκ-область.
12. Мышь по п. 1, где эндогенный локус легкой λ-цепи иммуноглобулина содержит делецию эндогенных сегментов генов Vλ и Jλ, полностью или частично.
13. Мышь по п. 1, где сегмент мышиного гена Сλ представляет собой сегмент мышиного гена Сλ1.
14. Мышь по п. 5, где локус легкой κ-цепи иммуноглобулина содержит вставку проксимальной дупликации Vκ полностью или частично, человеческого локуса легкой κ-цепи иммуноглобулина.
15. Мышь по п. 5, где в локусе тяжелой цепи иммуноглобулина отсутствует эндогенный мышиный ген Adam6.
16. Мышь по п. 15, где локус тяжелой цепи иммуноглобулина дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более мышиных полипептидов Adam6.
17. Мышь по п. 5, где мышь является гомозиготной в отношении эндогенного локуса тяжелой цепи иммуноглобулина.
18. Мышь по п. 5, где мышь является гомозиготной в отношении эндогенного локуса легкой κ-цепи иммуноглобулина.
19. Мышь по п. 1, где мышь является гомозиготной в отношении эндогенного локуса легкой λ-цепи иммуноглобулина.
20. Способ получения мыши, геном зародышевого типа которой содержит сконструированный эндогенный локус легкой λ цепи иммуноглобулина, при этом способ включает
(a) введение фрагмента ДНК в мышиную эмбриональную стволовую клетку, при этом указанный фрагмент ДНК содержит нуклеотидную последовательность, которая содержит
(i) один или более сегментов человеческого гена Vλ,
(ii) один или более сегментов человеческого гена Jλ и
(iii) один или более сегментов человеческого гена Сλ,
где (i)-(iii) функционально связаны с сегментом мышиного гена Сλ, и
где нуклеотидная последовательность дополнительно содержит один или более энхансеров человеческой легкой λ-цепи (Еλ) иммуноглобулина;
(b) получение мышиной эмбриональной стволовой клетки, образованной на (а); и
(c) создание мыши с применением мышиной эмбриональной стволовой клетки из (b).
21. Способ получения мыши, геном зародышевого типа которой содержит сконструированный эндогенный локус легкой λ цепи иммуноглобулина, при этом сконструированный эндогенный локус легкой λ цепи иммуноглобулина содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Сλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом мышиного или человеческого гена Сλ, и при этом эндогенный локус легкой λ цепи иммуноглобулина дополнительно содержит один или более энхансеров мышиной легкой λ-цепи (Еλ) иммуноглобулина и один или более энхансеров человеческой легкой λ-цепи (Еλ) иммуноглобулина, при этом способ включает
модификацию генома зародышевого типа мыши для содержания сконструированного локуса легкой λ-цепи иммуноглобулина, который содержит вставку одного или более сегментов человеческого гена Vλ, одного или более сегментов человеческого гена Jλ и одного или более сегментов человеческого гена Сλ, при этом сегменты человеческих генов Vλ и Jλ функционально связаны с сегментом мышиного или человеческого гена Сλ, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров мышиной легкой λ-цепи (Еλ) иммуноглобулина и один или более энхансеров человеческой легкой λ-цепи (Еλ) иммуноглобулина, с получением тем самым указанной мыши.
| US2014283150 A1, 18.09.2014 | |||
| WO2011163311 A1, 29.12.2011 | |||
| WO2013096142 A1, 27.06.2013 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЫШЕЙ | 2009 |
|
RU2425880C2 |

