ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам и композициям для получения гликозилированных белков (гликопротеинов) и, в частности, Fc-содержащих полипептидов, которые пригодны в качестве терапевтических средств для человека или животного.
ПРЕДШЕСТВУЮЩИЙ ИЗОБРЕТЕНИЮ УРОВЕНЬ ТЕХНИКИ
Моноклональные антитела часто осуществляют свою терапевтическую полезность за счет двух событий связывания. Сначала вариабельный домен антитела связывается со специфическим белком на клетке-мишени, например, с CD20 на поверхности злокачественных клеток. Затем следует рекрутинг эффекторных клеток, таких как клетки - естественные киллеры (NK), которые связываются с константной областью (Fc) антитела и разрушают клетки, с которыми связано антитело. Этот процесс, известный как антителозависимая клеточная цитотоксичность (ADCC), зависит от специфического N-гликозилирования по Asn297 в Fc-домене тяжелой цепи IgG1, Rothman et al., Mol. Immunol. 26: 1113-1123 (1989). Антитела, у которых отсутствует эта N-гликозилированная структура, по-прежнему связывают антиген, но не способны содействовать ADCC, очевидно, в результате сниженной аффинности в Fc-домене антитела к Fc-рецептору FcγRIIIa на поверхности NK-клеток.
Наличие N-гликозилирования играет роль не только в эффекторной функции антитела, конкретный состав N-связанного олигосахарида также является важным для конечного действия антитела. Отсутствие фукозы или наличие рассекающего N-ацетилглюкозамина положительно коррелирует с мощностью ADCC, Rothman (1989), Umana et al., Nat-Biotech. 17: 176-180 (1999), Shields et al., J. Biol Chem. 277: 26733-26740 (2002), и Shinkawa et al., J. Biol. Chem. 278: 3466-3473 (2003). Существуют также свидетельства, что сиалирование в Fc-области положительно коррелирует с противовоспалительными свойствами внутривенного иммуноглобулина (IVIG). См., например, Kaneko et al., Science, 313: 670-673, 2006; Nimmerjahn и Ravetch., J. Exp. Med., 204: 11-15, 2007.
Учитывая полезность специфического N-гликозилирования для функции и активности антител, был бы желателен способ для изменения композиции N-связанных олигосахаридов и изменения эффекторной функции антител.
Дрожжи и другие хозяева-грибы являются важными продуцирующими платформами для создания рекомбинантных белков. Дрожжи являются эукариотами и, таким образом, имеют общие эволюционные процессы с высшими эукариотами, включая множество посттрансляционных модификаций, которые происходят на секреторном пути. Современные достижения в гликоинженерии привели к созданию клеточных линий дрожжевого штамма Pichia pastoris с генетически модифицированными путями гликозилирования, которые позволяют им проводить последовательность ферментативных реакций, воспроизводящих процесс гликозилирования у людей. См., например, патенты США №№ 7029872, 7326681 и 7449308, которые описывают способы продуцирования рекомбинантного гликопротеина в клетке-хозяине низших эукариот, которые по существу идентичны их человеческим аналогам. Человекоподобные сиалированные биантенные N-связанные гликаны комплексного типа, аналогичные тем, что вырабатываются в Pichia pastoris вышеуказанными способами, продемонстрировали полезность для производства терапевтических гликопротеинов. Таким образом, был бы желателен способ для дальнейшей модификации или улучшения производства антител в дрожжах, таких как Pichia pastoris.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к Fc-содержащему полипептиду, который содержит мутации в аминокислотных положениях 243 и 264 Fc-области, где мутации в положении 243 выбраны из группы, состоящей из F243A, F243G, F243S, F243T, F243V, F243L, F243I, F243D, F243Y, F243E, F243R, F243W и F243K, и мутации в положении 264 выбраны из группы, состоящей из V264A, V264G, V264S, V264T, V264D, V264E, V264K, V264W, V264H, V264P, V264N, V264Q и V264L, где нумерация приведена согласно индексу EU по Kabat. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243Y и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243T и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264N. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243V и V264G. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой антитело или фрагмент антитела. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид по изобретению представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид по изобретению представляет собой фрагмент антитела, состоящий (или состоящий в основном) из SEQ ID NO:18 или SEQ ID NO:19.
В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело, включающее аминокислотную последовательность тяжелой цепи из SEQ ID NO:9 или ее вариант и аминокислотную последовательность легкой цепи из SEQ ID NO:2 или ее вариант. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело, включающее аминокислотную последовательность тяжелой цепи из SEQ ID NO:9 минус последний остаток лизина (K), записанный в SEQ ID NO:9.
В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело, включающее аминокислотную последовательность тяжелой цепи из SEQ ID NO:12 или ее вариант и аминокислотную последовательность легкой цепи из SEQ ID NO:11 или ее вариант.
В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело, включающее аминокислотную последовательность тяжелой цепи из SEQ ID NO:15 или ее вариант и аминокислотную последовательность легкой цепи из SEQ ID NO:14 или ее вариант.
В некоторых вариантах осуществления Fc-содержащие полипептиды по изобретению включают в себя N-гликаны, содержащие сиаловую кислоту (включая NANA, NGNA и их аналоги и производные). В одном из вариантов осуществления Fc-содержащие полипептиды по изобретению содержат смесь α-2,3- и α-2,6-связанных сиаловых кислот. В другом варианте осуществления Fc-содержащие полипептиды по изобретению содержат только α-2,6-связанную сиаловую кислоту. В одном из вариантов осуществления Fc-содержащие полипептиды по изобретению содержат α-2,6-связанную сиаловую кислоту и не содержат детектируемого количества α-2,3-связанной сиаловой кислоты. В одном из вариантов осуществления сиаловая кислота представляет собой N-ацетилнейраминовую кислоту (NANA) или N-гликолилнейраминовую кислоту (NGNA) или их смесь. В другом варианте осуществления сиаловая кислота представляет собой аналог или производное NANA или NGNA с ацетилированием в положении 9 на сиаловой кислоте. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах по изобретению содержат NANA и не содержат NGNA.
N-гликаны на Fc-содержащих полипептидах по изобретению могут необязательно содержать фукозу. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах содержат смесь фукозилированных и нефукозилированных N-гликанов. В другом варианте осуществления N-гликаны на Fc-содержащих полипептидах не содержат фукозу.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает одним или несколькими следующими свойствами по сравнению с родительским Fc-содержащим полипептидом: (i) сниженная эффекторная функция; (ii) повышенные противовоспалительные свойства; (iii) повышенное сиалирование; (iv) повышенная биодоступность (абсорбция или экспозиция), и (v) сниженное связывание с FcγRI, FcγRIIa, FcγRIIb и FcγRIIIa (FcγRIIIa-V158 или FcγRIIIa-F158), и FcγRIIIb. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает, по меньшей мере, 7, 10, 15, 30, 50, 100, 500 или 1000-кратным снижением эффекторной функции по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления эффекторная функция представляет собой ADCC. В другом варианте осуществления эффекторная функция представляет собой CDC.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает сниженной активностью ADCC по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид обладает, по меньшей мере, 7, 10, 15, 30, 50, 100, 500 или 1000-кратным снижением активности ADCC. В другом варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 100-кратным снижением активности ADCC. В другом варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 500-кратным снижением активности ADCC. В другом варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 1000-кратным снижением активности ADCC.
В другом варианте осуществления Fc-содержащий полипептид по изобретению обладает сниженной активностью CDC по сравнению с родительским Fc-содержащим полипептидом. В одном варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 100-кратным снижением активности CDC.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa (FcγRIIIa-V158 или FcγRIIIa-F158) и FcγRIIIb со сниженной аффинностью по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIa, по меньшей мере, с 50-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIb, по меньшей мере, с 20-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIIa LF, по меньшей мере, с 10-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIIa LV, по меньшей мере, с 1, 2 или 10-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIb, FcγRIIIa LF и FcγRIIIa LV со сниженной аффинностью по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает повышенными противовоспалительными свойствами по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает повышенной биодоступностью (абсорбцией или экспозицией) при введении парентерально по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает повышенной биодоступностью (абсорбцией или экспозицией) при введении подкожно по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию V264A.
Изобретение также относится к способу для получения Fc-содержащего полипептида в клетке-хозяине, включающему: (i) создание клетки-хозяина с помощью генетической инженерии для выработки Fc-содержащего полипептида, где клетка-хозяин содержит нуклеиновую кислоту, кодирующую мутации в аминокислотных положениях 243 и 264 Fc-области, где мутации в положении 243 выбраны из группы, состоящей из: F243A, F243G, F243S, F243T, F243V, F243L, F243I, F243D, F243Y, F243E, F243R, F243W и F243K и мутации в положении 264 выбраны из группы, состоящей из: V264A, V264G, V264S, V264T, V264D, V264E, V264K, V264W, V264H, V264P, V264N, V264Q и V264L, где нумерация приведена согласно индексу EU по Kabat; (ii) культивирование клетки-хозяина в условиях, приводящих к экспрессии Fc-содержащего полипептида; и (iii) выделение Fc-содержащего полипептида из клетки-хозяина. В одном из вариантов осуществления нуклеиновая кислота кодирует мутации F243A и V264A. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243Y и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации 243T и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243L и V264A. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243L и V264N. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243V и V264G. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой антитело или фрагмент антитела. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19.
В одном из вариантов осуществления способ получения Fc-содержащего полипептида осуществляют в клетке млекопитающего. В другом варианте осуществления способ получения Fc-содержащего полипептида осуществляют в растительной клетке. В другом варианте осуществления способ получения Fc-содержащего полипептида осуществляют в бактериях. В другом варианте осуществления способ получения Fc-содержащего полипептида осуществляют в клетке насекомого. В другом варианте осуществления способ получения Fc-содержащего полипептида осуществляют в клетке низших эукариот. В другом варианте осуществления способ получения Fc-содержащего полипептида осуществляют в дрожжевой клетке. В одном из вариантов осуществления способ получения Fc-содержащего полипептида осуществляют в Pichia pastoris.
В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, включает N-гликаны, содержащие сиаловую кислоту (включая NANA, NGNA, и их аналоги и производные). В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, содержит N-гликановую композицию, в которой, по меньшей мере, 40 моль%, 70 моль% или 90 моль% N-гликанов на Fc-содержащем полипептиде сиалированы (имеют структуру, выбранную из SA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2 или SAGalGlcNAcMan5GlcNAc2). В одном из вариантов осуществления, по меньшей мере, 47 моль% N-гликанов на антителах имеют структуру SA2Gal2GlcNAc2Man3GlcNAc2. В другом варианте осуществления, по меньшей мере, 47 моль% N-гликанов на антителах имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2. В другом варианте осуществления, по меньшей мере, 66 моль% N-гликанов на антителах имеют структуру SA2Gal2GlcNAc2Man3GlcNAc2. В другом варианте осуществления, по меньшей мере, 66 моль% N-гликанов на антителах имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2. В одном из вариантов осуществления Fc-содержащие полипептиды, полученные способом по изобретению, содержат смесь α-2,3- и α-2,6-связанных сиаловых кислот. В другом варианте осуществления Fc-содержащие полипептиды по изобретению содержат только α-2,6-связанную сиаловую кислоту. В одном из вариантов осуществления Fc-содержащие полипептиды по изобретению содержат α-2,6-связанную сиаловую кислоту и не содержат детектируемого количества α-2,3-связанной сиаловой кислоты. В одном из вариантов осуществления сиаловая кислота представляет собой N-ацетилнейраминовую кислоту (NANA) или N-гликолилнейраминовую кислоту (NGNA) или их смесь. В другом варианте осуществления сиаловая кислота представляет собой аналог или производное NANA или NGNA с ацетилированием в положении 9 на сиаловой кислоте. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах, полученных способом по изобретению, содержат NANA и не содержат NGNA.
N-гликаны на Fc-содержащих полипептидах, полученных способом по изобретению, могут необязательно содержать фукозу. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах, полученных способом по изобретению, содержат смесь фукозилированных и нефукозилированных N-гликанов. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах, полученных способом по изобретению, не содержат фукозу.
В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, содержит N-гликановую композицию, в которой общее количество и процентное содержание сиалированных N-гликанов повышено относительно родительского Fc-содержащего полипептида.
В некоторых вариантах осуществления Fc-содержащий полипептид, полученный способом по изобретению, обладает одним или несколькими следующими свойствами по сравнению с родительским Fc-содержащим полипептидом: (i) сниженная эффекторная функция; (ii) повышенные противовоспалительные свойства; (iii) повышенное сиалирование; (iv) повышенная биодоступность (абсорбция или экспозиция), и (v) сниженное связывание с FcγRI, FcγRIIa, FcγRIIb и FcγRIIIa. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению обладает, по меньшей мере, 7, 10, 15, 30, 50, 100, 500 или 1000-кратным снижением эффекторной функции по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления эффекторная функция представляет собой ADCC. В другом варианте осуществления эффекторная функция представляет собой CDC.
В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, обладает сниженной активностью ADCC по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид обладает, по меньшей мере, 7, 10, 15, 30, 50, 100, 500 или 1000-кратным снижением активности ADCC. В другом варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 100-кратным снижением активности ADCC. В другом варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 500-кратным снижением активности ADCC. В другом варианте осуществления Fc-содержащий полипептид обладает, по меньшей мере, 1000-кратным снижением активности ADCC.
В другом варианте осуществления Fc-содержащий полипептид, полученный способом по изобретению, обладает сниженной активностью CDC по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид обладает, по меньшей мере, 100-кратным снижением активности CDC.
В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, связывается с FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa (FcγRIIIa-V158 или FcγRIIIa-F158) и FcγRIIIb со сниженной аффинностью по сравнению родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIa, по меньшей мере, с 50-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIb, по меньшей мере, с 20-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIIa LF, по меньшей мере, с 10-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIIa LV по меньшей мере, с 1, 2 или 10-кратным снижением аффинности по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению связывается с FcγRIIb, FcγRIIIa LF и FcγRIIIa LV, со сниженной аффинностью по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, обладает повышенными противовоспалительными свойствами по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, обладает повышенной биодоступностью (абсорбцией или экспозицией) при введении парентерально по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид, полученный способом по изобретению, обладает повышенной биодоступностью (абсорбцией или экспозицией) при введении подкожно по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию V264A.
Изобретение также относится к способу снижения эффекторной функции Fc-содержащего полипептида, включающему введение мутаций в положениях 243 и 264 родительского Fc-содержащего полипептида, где указанный Fc-содержащий полипептид имеет сниженную эффекторную функцию по сравнению с родительским Fc-содержащим полипептидом, где нумерация приведена согласно индексу EU по Kabat. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243Y и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации 243T и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243L и V264A. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243L и V264N. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243V и V264G. В одном из вариантов осуществления эффекторная функция представляет собой ADCC. В другом варианте осуществления эффекторная функция представляет собой CDC. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой антитело или фрагмент антитела. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19. В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию V264A.
Изобретение также относится к способу усиления противовоспалительных свойств Fc-содержащего полипептида, включающему введение мутации в положении 243 и мутации в положении 264 родительского Fc-содержащего полипептида, где нумерация приведена согласно индексу EU по Kabat, где указанный Fc-содержащий полипептид обладает повышенной противовоспалительной активностью по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243Y и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации 243T и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264N. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243V и V264G. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой антитело или фрагмент антитела. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело или его антигенсвязывающий фрагмент, который связывается с антигеном, выбранным из группы, состоящей из TNF-α, IL-1, IL-2, IL-4, IL-5, IL-6, IL-8, IL-9, IL-10, IL-12, IL-15, IL-17, IL-18, IL-20, IL-21, IL-22, IL-23, IL-23R, IL-25, IL-27, IL-33, CD2, CD4, CD11A, CD14, CD18, CD19, CD23, CD25, CD40, CD40L, CD20, CD52, CD64, CD80, CD147, CD200, CD200R, TSLP, TSLPR, PD-1, PDL1, CTLA4, VLA-4, VEGF, PCSK9, α4β7-интегрина, E-селектина, фактора II, ICAM-3, бета2-интегрина, IFNγ, C5, CBL, LCAT, CR3, MDL-1, GITR, ADDL, CGRP, TRKA, IGF1R, RANKL, GTC или рецептора для любой из вышеперечисленных молекул. В одном из вариантов осуществления Fc-содержащий полипептид связывается с TNF-α. В другом варианте осуществления Fc-содержащий полипептид связывается с Her2. В другом варианте осуществления Fc-содержащий полипептид связывается с PCSK9. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид по изобретению представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид по изобретению представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19. В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию V264A.
Изобретение также относится к способу усиления противовоспалительных свойств Fc-содержащего полипептида, включающему выбор родительского Fc-содержащего полипептида, который подходит для лечения воспаления (например, антитела или иммуноадгезина, которые связываются с антигеном, вовлеченным в воспаление) и введение мутаций в положениях 243 или 264 в Fc-области, где нумерация приведена согласно индексу EU по Kabat, где Fc-содержащий полипептид обладает повышенной противовоспалительной активностью по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243Y и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации 243T и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264N. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243V и V264G. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению представляет собой антитело или фрагмент антитела. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело или его антигенсвязывающий фрагмент, который связывается с антигеном, выбранным из группы, состоящей из TNF-α, IL-1, IL-2, IL-4, IL-5, IL-6, IL-8, IL-9, IL-10, IL-12, IL-15, IL-17, IL-18, IL-20, IL-21, IL-22, IL-23, IL-23R, IL-25, IL-27, IL-33, CD2, CD4, CD11A, CD14, CD18, CD19, CD23, CD25, CD40, CD40L, CD20, CD52, CD64, CD80, CD147, CD200, CD200R, TSLP, TSLPR, PD-1, PDL1, CTLA4, VLA-4, VEGF, PCSK9, α4β7-интегрина, E-селектина, фактора II, ICAM-3, бета2-интегрина, IFNγ, C5, CBL, LCAT, CR3, MDL-1, GITR, ADDL, CGRP, TRKA, IGF1R, RANKL, GTC или рецептора для любой из вышеперечисленных молекул. В одном из вариантов осуществления Fc-содержащий полипептид связывается с TNF-α. В другом варианте осуществления Fc-содержащий полипептид связывается с Her2. В другом варианте осуществления Fc-содержащий полипептид связывается с PCSK9. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19. В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию V264A.
Изобретение также относится к способу лечения воспалительного состояния у нуждающегося в этом индивидуума, включающему введение индивидууму терапевтически эффективного количества Fc-содержащего полипептида, включающего мутации в положениях 243 и 264, где нумерация приведена согласно индексу EU по Kabat. В одном из вариантов осуществления Fc-содержащий полипептид снижает экспрессию гена, выбранного из группы, состоящей из IL-Ιβ, IL-6, RANKL, TRAP, ATP6v0d2, MDL-1, DAP12, CD11b, TIMP-1, MMP9, CTSK, PU-1, MCP1, MIP1α, Cxcl1-Groa, Cxcl2-Grob, CD18, TNF, FcγRI, FcγRIIb, FcγRIII и FcγRIV. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243Y и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации 243 и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243L и V264A. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243L и V264N. В другом варианте осуществления нуклеиновая кислота кодирует мутации F243V и V264G. В одном из вариантов осуществления Fc-содержащий полипептид вводят парентерально. В одном из вариантов осуществления Fc-содержащий полипептид вводят подкожно. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело или его антигенсвязывающий фрагмент. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело или его антигенсвязывающий фрагмент, которые подходят для лечения воспалительного состояния. В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, выбранным из группы, состоящей из TNF-α, IL-1, IL-2, IL-4, IL-5, IL-6, IL-8, IL-9, IL-10, IL-12, IL-15, IL-17, IL-18, IL-20, IL-21, IL-22, IL-23, IL-23R, IL-25, IL-27, IL-33, CD2, CD4, CD11A, CD14, CD18, CD19, CD23, CD25, CD40, CD40L, CD20, CD52, CD64, CD80, CD147, CD200, CD200R, TSLP, TSLPR, PD-1, PDL1, CTLA4, VLA-4, VEGF, PCSK9, α4β7-интегрина, E-селектина, фактора II, ICAM-3, бета2-интегрина, IFNγ, C5, CBL, LCAT, CR3, MDL-1, GITR, ADDL, CGRP, TRKA, IGF1R, RANKL, GTC или рецептора для любой из вышеперечисленных молекул. В одном из вариантов осуществления Fc-содержащий полипептид связывается с TNF-α. В другом варианте осуществления Fc-содержащий полипептид связывается с Her2. В другом варианте осуществления Fc-содержащий полипептид связывается с PCSK9. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19.
Другое изобретение, описываемое в настоящем документе, относится к фармацевтической композиции, которая включает Fc-содержащий полипептид, где, по меньшей мере, 70% N-гликанов на Fc-содержащем полипептиде включают олигосахаридную структуру, выбранную из группы, состоящей из SA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2 и SAGalGlcNAcMan5GlcNAc2, где Fc-содержащий полипептид содержит мутации в аминокислотных положениях 243 и 264 в Fc-области, где нумерация приведена согласно индексу EU по Kabat. В одном из вариантов осуществления мутации представляют собой F243A и V264A. В другом варианте осуществления мутации представляют собой F243Y и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации 243T и V264G. В другом варианте осуществления мутации представляют собой F243L и V264A. В другом варианте осуществления мутации представляют собой F243L и V264N. В другом варианте осуществления мутации представляют собой F243V и V264G. В одном из вариантов осуществления, по меньшей мере, 47 моль% N-гликанов имеют структуру SA2Gal2GlcNAc2Man3GlcNAc2. В другом варианте осуществления, по меньшей мере, 47 моль% N-гликанов имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2. В одном из вариантов осуществления сиалированные N-гликаны содержат смесь α-2,3- и α-2,6-связанных сиаловых кислот. В другом варианте осуществления сиалированные N-гликаны содержат только α-2,6-связанную сиаловую кислоту. В одном из вариантов осуществления сиалированные N-гликаны содержат α-2,6-связанную сиаловую кислоту и не содержат детектируемого количества α-2,3-связанной сиаловой кислоты. В одном из вариантов осуществления сиаловая кислота представляет собой N-ацетилнейраминовую кислоту (NANA) или N-гликолилнейраминовую кислоту (NGNA) или их смесь. В другом варианте осуществления сиаловая кислота представляет собой аналог или производное NANA или NGNA с ацетилированием в положении 9 на сиаловой кислоте. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах содержат NANA и не содержат NGNA. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19.
Другое изобретение, описываемое в настоящем документе, относится к фармацевтической композиции, которая включает Fc-содержащий полипептид, где, по меньшей мере, 70% N-гликанов на Fc-содержащем полипептиде включают олигосахаридную структуру, выбранную из группы, состоящей из SA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2 и SAGalGlcNAcMan5GlcNAc2, где остатки сиаловой кислоты присоединены посредством α-2,6-связи, где у N-гликанов отсутствует фукоза, и где Fc-содержащий полипептид содержит мутации в аминокислотных положениях 243 и 264 в Fc-области, где нумерация приведена согласно индексу EU по Kabat. В одном из вариантов осуществления мутации представляют собой F243A и V264A. В другом варианте осуществления мутации представляют собой F243Y и V264G. В другом варианте осуществления нуклеиновая кислота кодирует мутации 243T и V264G. В другом варианте осуществления мутации представляют собой F243L и V264A. В другом варианте осуществления мутации представляют собой F243L и V264N. В другом варианте осуществления мутации представляют собой F243V и V264G. В одном из вариантов осуществления, по меньшей мере, 47 моль% N-гликанов имеют структуру SA2Gal2GlcNAc2Man3GlcNAc2. В другом варианте осуществления, по меньшей мере, 47 моль% N-гликанов имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2. В одном из вариантов осуществления сиалированные N-гликаны содержат смесь α-2,3- и α-2,6-связанных сиаловых кислот. В другом варианте осуществления сиалированные N-гликаны содержат только α-2,6-связанную сиаловую кислоту. В одном из вариантов осуществления сиалированные N-гликаны содержат α-2,6-связанную сиаловую кислоту и не содержат детектируемого количества α-2,3-связанной сиаловой кислоты. В одном из вариантов осуществления сиаловая кислота представляет собой N-ацетилнейраминовую кислоту (NANA) или N-гликолилнейраминовую кислоту (NGNA) или их смесь. В другом варианте осуществления сиаловая кислота представляет собой аналог или производное NANA или NGNA с ацетилированием в положении 9 на сиаловой кислоте. В одном из вариантов осуществления N-гликаны на Fc-содержащих полипептидах содержат NANA и не содержат NGNA. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:18. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, включающий SEQ ID NO:19. В другом варианте осуществления Fc-содержащий полипептид представляет собой фрагмент антитела, состоящий (или состоящий по существу) из SEQ ID NO:18 или SEQ ID NO:19.
Изобретение также относится к Fc-содержащему полипептиду, включающему тяжелую цепь и легкую цепь, где тяжелая цепь содержит аминокислотную последовательность из SEQ ID NO:9 или ее вариант, и легкая цепь содержит аминокислотную последовательность из SEQ ID NO:2 или ее вариант, где вариант включает одно или несколько следующих свойств по сравнению с антителом, содержащим аминокислотную последовательность тяжелой цепи из SEQ ID NO:1 и аминокислотную последовательность легкой цепи из SEQ ID NO:2: сниженная эффекторная функция, повышенные противовоспалительные свойства; повышенное сиалирование; повышенная биодоступность (абсорбция или экспозиция) при парентеральном введении, и сниженное связывание с FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb. Изобретение также относится к Fc-содержащему полипептиду, включающему тяжелую цепь и легкую цепь, где тяжелая цепь содержит аминокислотную последовательность из SEQ ID NO:12 или ее вариант, и легкая цепь содержит аминокислотную последовательность из SEQ ID NO:11 или ее вариант, где вариант включает одно или несколько следующих свойств по сравнению с антителом, содержащим аминокислотную последовательность тяжелой цепи из SEQ ID NO:10 и аминокислотную последовательность легкой цепи из SEQ ID NO:11: сниженная эффекторная функция, повышенные противовоспалительные свойства; повышенное сиалирование; повышенная биодоступность (абсорбция или экспозиция) при парентеральном введении, и сниженное связывание с FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb. Изобретение также относится к Fc-содержащему полипептиду, включающему тяжелую цепь и легкую цепь, где тяжелая цепь содержит аминокислотную последовательность из SEQ ID NO:15 или ее вариант, и легкая цепь содержит аминокислотную последовательность из SEQ ID NO:14 или ее вариант, где вариант включает одно или несколько следующих свойств по сравнению с антителом, содержащим аминокислотную последовательность тяжелой цепи из SEQ ID NO:13 и аминокислотную последовательность легкой цепи из SEQ ID NO:14: сниженная эффекторная функция, повышенные противовоспалительные свойства; повышенное сиалирование; повышенная биодоступность (абсорбция или экспозиция) при парентеральном введении, и сниженное связывание с FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb. В одном из вариантов осуществления вариант содержит в пределах 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более консервативных или неконсервативных аминокислотных замен. В одном из вариантов осуществления вариант содержит, по меньшей мере, 75%, 80%, 85%, 90%, 95%, 98% или 99% последовательности, идентичной последовательности по изобретению.
Изобретение также относится к способу усиления противовоспалительных свойств Fc-содержащего полипептида, включающему введение мутации в положении 243 или мутации в положении 264 родительского Fc-содержащего полипептида, где нумерация приведена согласно индексу EU по Kabat, где указанный Fc-содержащий полипептид обладает повышенной противовоспалительной функцией по сравнению с родительским Fc-содержащим полипептидом. Изобретение также относится к способу усиления противовоспалительных свойств Fc-содержащего полипептида, включающему: выбор родительского Fc-содержащего полипептида, который подходит для лечения воспаления (например, антитела или иммуноадгезина, которые связываются с антигеном, вовлеченным в воспаление), и введение мутации в положении 243 или мутации в положении 264 родительского Fc-содержащего полипептида, где нумерация приведена согласно индексу EU по Kabat, где указанный Fc-содержащий полипептид обладает повышенной противовоспалительной функцией по сравнению с родительским Fc-содержащим полипептидом. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления Fc-содержащий полипептид содержит мутацию V264A. В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область.
Изобретение также относится к способу лечения воспалительного состояния у нуждающегося в этом индивидуума, включающему: введение индивидууму терапевтически эффективного количества Fc-содержащего полипептида с мутацией в положении 243 или мутацией в положении 264 родительского Fc-содержащего полипептида, где нумерация приведена согласно индексу EU по Kabat. В одном из вариантов осуществления Fc-содержащий полипептид вводят парентерально. В одном из вариантов осуществления Fc-содержащий полипептид вводят подкожно. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления Fc-содержащий полипептид содержит мутацию V264A. В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область.
В любом вышеуказанном варианте осуществления увеличение противовоспалительного действия может быть выявлено при помощи любого известного в данной области способа. В одном из вариантов осуществления увеличение противовоспалительного действия выявляется путем измерения снижения экспрессии гена, выбранного из группы, состоящей из IL-Ιβ, IL-6, RANKL, TRAP, ATP6v0d2, MDL-1, DAP12, CD11b, TIMP-1, MMP9, CTSK, PU-1, MCP1, MIP1α, Cxcl1-Groa, CXcl2-Grob, CD18, TNF, FcγRI, FcγRIIb, FcγRIII и FcγRIV.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фигура 1 представляет собой графическое изображение pGLY3483, экспрессионной плазмиды двойного мутеина F243A и V264A. Обе тяжелая и легкая цепи находились под контролем промотора, индуцируемого метанолом, AOX1. Ген PpTrp2 представлял собой локус, который использовали для интеграции плазмиды целиком. За исключением мутаций тяжелой цепи, структура экспрессионной плазмиды оставалась такой же, что и для дикого типа (родительского) и экспрессионных плазмид одиночного мутеина F243A, одиночного мутеина V264A и двойного мутеина.
Фигура 2 представляет собой изображение гелей из SDS-PAGE анализа, характеризующих невосстановленные (NR) и восстановленные (R) антитела, полученные при помощи материалов и способов по настоящему изобретению. Дорожка 1 содержит моноклональное антитело против Her2; дорожка 2 содержит одиночный Fc-мутеин, F243A; дорожка 3 содержит одиночный Fc-мутеин, V264A; и дорожка 4 содержит двойной Fc-мутеин, F243A/V264A.
Фигура 3 иллюстрирует аффинность к антигену для различных антител, полученных при помощи материалов и способов по настоящему изобретению, которую определяли путем анализа клеток с использованием клеточной линии SK-BR3, линии рака молочной железы человека со сверхэкспрессией Her2. ■ - Her2; ♦ - F243A/V264A GS6.0 гликозилирование; ▲ - F243A с GS6.0 гликозилированием; ▼ - V264A с GS6.0 гликозилированием; ◊ -контрольный IgG; x - антитело к Her2 из Pichia pastoris, произведенное с GFI5.0 гликозилированием.
Фигура 4 иллюстрирует MALDI-TOF MS анализ N-гликанов антитела к Her2, произведенного в Pichia pastoris GFI5.0, штамм YDX477. Пики соответствуют Man5GlcNAc2, 1261,24 (GS2.0), GlcNAc2Man3GlcNAc2, 1343,50 (G0) (преобладающий), GalGlcNAc2Man3GlcNAc2, 1505,97 (G1), и Gal2GlcNAc2Man3GlcNAc2, 1668,47 (G2).
Фигура 5 иллюстрирует MALDI-TOF MS анализ N-гликанов одиночного Fc-мутеина, F243A, антитела, произведенного в GFI5.0, штамм YDX551. Пики соответствуют Man5GlcNAc2, 1261,05, GlcNAc2Man3GlcNAc2, 1343,71, GalGlcNAc2Man3GlcNAc2, 1506,62, и Gal2GlcNAc2Man3GlcNAc2, 1668,97 (преобладающий).
Фигура 6 иллюстрирует MALDI-TOF MS анализ N-гликанов одиночного Fc-мутеина, V264A, антитела, произведенного в GFI5.0, штамм YDX551. Пики соответствуют Man5GlcNAc2, 1261,98, GalGlcNAc2Man3GlcNAc2, 1505,45, и Gal2GlcNAc2Man3GlcNAc2, 1668,85 (преобладающий).
Фигура 7 иллюстрирует MALDI-TOF MS анализ N-гликанов двойного Fc-мутеина, F243A/V264A, антитела, произведенного в GFI5.0, штамм YDX557. Основной пик соответствует Gal2GlcNAc2Man3GlcNAc2, 1668,39.
Фигура 8 иллюстрирует MALDI-TOF MS анализ N-гликанов двойного Fc-мутеина, F243A/V264A, антитела, произведенного в GFI6.0, штамм YGLY4563. Двойные пики на 2224,28 и 2245,83 (преобладающие) соответствуют NANAGal2GlcNAc2Man3GlcNAc2 и NANA2Gal2GlcNAc2Man3GlcNAc2, соответственно.
Фигуры 9A-9D представляют собой графические изображения связывания с FcγR для различных антител, полученных при помощи материалов и способов, описанных в примере 11: FcγRIIIaLF (фигура 9A); FcγRI (фигура 9B); FcγRIIb/c (фигура 9С); и FcγRIIIa LV (фигура 9D). Для фигур 9A-9D: ■ - антитело к Her2; ▲ - F243A, произведенное в GFI6.0; ▼ - V264A, произведенное в GFI6.0; ♦ - F243A/V264A, произведенное в GFI6.0; ● - F243A/V264A, произведенное в GFI6.0 и обработанное ПНГазой.
Фигура 10 представляет собой графическое изображение связывания с C1q для различных антител, полученных при помощи материалов и способов, описанных в примере 12: ◊ - антитело к CD20, положительный контроль; ■ - антитело к Her2; ▲ - F243A, произведенное с GS6.0 гликозилированием; ▼ - V264A, произведенное с GS6.0 гликозилированием; ♦ - F243A/V264A, произведенное с GS6.0 гликозилированием; ● - F243A/V264A с GS6.0 гликозилированием и ПНГазой.
Фигура 11 представляет собой графическое изображение ответа ADCC для различных антител, полученных при помощи материалов и способов, описанных в примере 13: ■ - антитело к Her2; ▲ - F243A произведенное с GS6.0 гликозилированием; ▼ - V264A, произведенное с GS6.0 гликозилированием; ♦ - F243A/V264A, произведенное с GS6.0 гликозилированием; ◊ - NK-клеточный лизис без антитела; X - антитело к Her2, произведенное в GFI2.0.
Фигура 12 представляет собой графическое изображение концентрации моноклонального антитела в сыворотке с течением времени у мышей, инъецированных: ■ - антитело к Her2; X - антитело к Her2, произведенное в GFI5.0; ♦ - F243A/V264A, произведенное в GFI6.0, как описано в примере 14.
Фигуры 13A-13E представляют собой графические изображения связывания с FcγR для различных антител, описанных в примере 15.
Фигура 14 представляет собой графическое изображение ответа ADCC для различных антител, полученных при помощи материалов и способов, описанных в примере 16. Результаты на фигурах 14A и 14B получены из экспериментов с использованием гетерозиготных F/V эффекторных клеток. Результаты на фигурах 14C и 14D получены из экспериментов с использованием F/F эффекторных клеток. Результаты на фигуре 14E получены из эксперимента с использованием V/V эффекторных клеток.
Фигура 15 представляет собой графическое изображение активности ADCC двойного Fc-мутанта антитела против Her2 по сравнению с предполагаемой добавочной нормативной кривой каждого из одиночных мутантов, как описано в примере 17.
Фигура 16 представляет собой графическое изображение ответа ADCC для различных антител, полученных при помощи материалов и способов, описанных в примере 18.
Фигура 17 представляет собой графическое изображение ответа CDC для различных антител, полученных при помощи материалов и способов, описанных в примере 19.
фигура 18 представляет собой графическое изображение действия Fc-мутеинов по изобретению на модели AIA, описанной в примере 20.
Фигура 19 представляет собой графическое изображение действия Fc-мутеинов по изобретению на модели AIA, описанной в примере 21.
Фигура 20 представляет собой графическое изображение связывания с FcγR для антител к TNFα, описанных в примере 22.
Фигура 21 представляет собой графическое изображение связывания с FcγR для антител к TNFα, описанных в примере 23.
Фигура 22 представляет собой графическое изображение действия Fc-мутеинов по изобретению на модели AIA, описанной в примере 24.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Термин "G0", применяемый в настоящем документе, относится к биантенному олигосахариду комплексного типа без галактозы или фукозы, GlcNAc2Man3GlcNAc2.
Термин "G1", применяемый в настоящем документе, относится к биантенному олигосахариду комплексного типа без фукозы и содержащему один остаток галактозила, GalGlcNAc2Man3GlcNAc2.
Термин "G2", применяемый в настоящем документе, относится к биантенному олигосахариду комплексного типа без фукозы и содержащему два остатка галактозила, Gal2GlcNAc2Man3GlcNAc2.
Термин "G0F", применяемый в настоящем документе, относится к биантенному олигосахариду комплексного типа, содержащему коровую фукозу, без галактозы, GlcNAc2Man3GlcNAc2F.
Термин "G1F", применяемый в настоящем документе, относится к биантенному олигосахариду комплексного типа, содержащему коровую фукозу и один остаток галактозила, GalGlcNAc2Man3GlcNAc2F.
Термин "G2F", применяемый в настоящем документе, относится к биантенному олигосахариду комплексного типа, содержащему коровую фукозу и два остатка галактозила, Gal2GlcNAc2Man3GlcNAc2F.
Термин "Man5", применяемый в настоящем документе, относится к олигосахаридной структуре, показанной как

Термин "GFI5.0", применяемый в настоящем документе, относится к гликоинжиниринговым штаммам Pichia pastoris, которые вырабатывают гликопротеины, преимущественно с Gal2GlcNAc2Man3GlcNAc2 N-гликанами.
Термин "GFI6.0", применяемый в настоящем документе, относится к гликоинжиниринговым штаммам Pichia pastoris, которые вырабатывают гликопротеины, преимущественно с NANA2Gal2GlcNAc2Man3GlcNAc2 N-гликанами.
Термин "GS5.0", применяемый в настоящем документе, относится к N-гликозилированной структуре Gal2GlcNAc2Man3GlcNAc2.
Термин "GS5.5", применяемый в настоящем документе, относится к N-гликозилированной структуре NANAGal2GlcNAc2Man3GlcNAc2, которая при выработке в штаммах Pichia pastoris, в которые гликоинженерными способами введена α2,6 сиалилтрансфераза, приводит к образованию α2,6-связанной сиаловой кислоты, и которая при выработке в штаммах Pichia pastoris, в которые гликоинженерными способами введена α2,3 сиалилтрансфераза, приводит к образованию α2,3-связанной сиаловой кислоты.
Термин "GS6.0", применяемый в настоящем документе, относится к N-гликозилированной структуре NANA2Gal2GlcNAc2Man3GlcNAc2, которая при выработке в штаммах Pichia pastoris, в которые гликоинженерными способами введена α2,6 сиалилтрансфераза, приводит к образованию α2,6-связанной сиаловой кислоты, и которая при выработке в штаммах Pichia pastoris, в которые гликоинженерными способами введена α2,3 сиалилтрансфераза, приводит к образованию α2,3-связанной сиаловой кислоты.
Термин "дикого типа" или "wt", применяемый в настоящем документе по отношению к штамму Pichia pastoris, относится к природному штамму Pichia pastoris, который не подвергали генетическим модификациям для контроля гликозилирования.
Термин "антитело", применяемый в настоящем документе, относится к молекуле иммуноглобулина, способной связывать конкретный антиген посредством, по меньшей мере, одного участка распознавания антигена, расположенного в вариабельной области молекулы иммуноглобулина. Применяемый в настоящем документе термин охватывает не только интактные поликлональные или моноклональные антитела, состоящие из четырех полипептидных цепей, т.е. двух идентичных пар полипептидных цепей, каждая пара имеет одну "легкую" цепь (LC) (приблизительно 25 кДа) и одну "тяжелую" цепь (HC) (приблизительно 50-70 кДа), но также их фрагменты, такие как Fab, Fab', F(ab')2, Fv, одна цепь (ScFv), их мутанты, слитые белки, включающие часть антитела, и любые другие модифицированные конфигурации молекулы иммуноглобулина, которая содержит участок распознавания антигена и, по меньшей мере, часть CH2-домена константной области тяжелой цепи иммуноглобулина, который включает N-связанный участок гликозилирования в CH2-домене, или их варианты. Применяемый в настоящем документе термин включает антитело любого класса, такое как IgG (например, IgGl, IgG2, IgG3 или IgG4), IgM, IgA, IgD и IgE, соответственно.
Термин "консенсусная последовательность CH2", применяемый в настоящем документе, относится к аминокислотной последовательности CH2-домена константной области тяжелой цепи, содержащей N-связанный участок гликозилирования, которая была получена из наиболее распространенных аминокислотных последовательностей, обнаруженных в CH2-доменах в ряде антител.
Термин "Fc-область" применяют для определения C-концевой области тяжелой цепи иммуноглобулина. "Fc-область" может быть нативной последовательностью Fc-области или вариантной Fc-областью. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи IgG человека определяется, как правило, протяженностью от аминокислотного остатка в положении Cys226, или от Pro230, до ее карбоксильного конца. Fc-область иммуноглобулина включает два константных домена, CH2 и CH3, и может необязательно включать шарнирную область. В одном из вариантов осуществления Fc-область включает аминокислотную последовательность SEQ ID NO:18. В одном из вариантов осуществления Fc-область включает аминокислотную последовательность SEQ ID NO:19. В другом варианте осуществления Fc-область включает аминокислотную последовательность SEQ ID NO:18 с добавлением остатка лизина (K) на 5'-конце. Fc-область содержит единичный N-связанный участок гликозилирования в CH2-домене, который соответствует участку Asn297 полноразмерной тяжелой цепи антитела.
Термин "Fc-содержащий полипептид" относится к полипептиду, такому как антитело или иммуноадгезин, который содержит Fc-область. Этот термин охватывает полипептиды, включающие или состоящие из (или состоящие по существу из) Fc-области. Полипептиды, включающие Fc-область, можно получать путем расщепления антител папаином или при помощи технологии рекомбинантных ДНК.
Термин "родительское антитело", "родительский иммуноглобулин" или "родительский Fc-содержащий полипептид", применяемый в настоящем документе, относится к антителу или Fc-содержащему полипептиду, в котором отсутствуют мутации в Fc-области, описываемые в настоящем документе. Родительский Fc-содержащий полипептид может включать нативную последовательность Fc-области или Fc-область с уже существующими модификациями аминокислотной последовательности. Нативная последовательность Fc-области содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, встречающейся в природе. Нативная последовательность Fc-области включает нативную последовательность Fc-области IgG1 человека, нативную последовательность Fc-области IgG2 человека, нативную последовательность Fc-области IgG3 человека и нативную последовательность Fc-области IgG4 человека, а также их природные варианты. Применяемые в качестве сравнения, родительское антитело или родительский Fc-содержащий полипептид можно экспрессировать в любой клетке. В одном из вариантов осуществления родительское антитело или родительский Fc-содержащий полипептид экпрессируется в той же самой клетке, что и Fc-содержащий полипептид по изобретению.
Применяемый в настоящем документе термин "иммуноадгезин" обозначает антителоподобные молекулы, которые сочетают в себе "связывающий домен" гетерологичного "адгезинового" белка (например, рецептора, лиганда или фермента) с константным доменом иммуноглобулина. Структурно, иммуноадгезины содержат слияние адгезиновой аминокислотной последовательности с желаемой специфичностью связывания, иной, чем участок распознавания антигена и участок связывания (антигенсвязывающий участок) антитела (т.е. "гетерологичной") и последовательность константного домена иммуноглобулина. Применяемый в настоящем документе термин "лигандсвязывающий домен" относится к любому поверхностному рецептору природной клетки или любой области или ее производному, сохраняющему, по меньшей мере, способность соответствующего природного рецептора к качественному связыванию лиганда. В конкретном варианте осуществления рецептор происходит из полипептида клеточной поверхности с внеклеточным доменом, который гомологичен члену генного суперсемейства иммуноглобулинов. Другие рецепторы, которые не являются членами суперсемейства иммуноглобулинов, но, тем не менее, подпадают под это определение, представляют собой рецепторы цитокинов, и, в частности, рецепторы с тирозинкиназной активностью (рецепторные тирозинкиназы), представители гематопоэтина и фактора роста нервов, которые предрасполагают млекопитающее к развитию обсуждаемого нарушения. В одном из вариантов осуществления нарушение представляет собой рак. Способы создания иммуноадгезинов хорошо известны в данной области. См., например, WO00/42072.
Термин "Her2" или "антитело к Her2", применяемый в настоящем документе, относится к антителу с аминокислотной последовательностью, аналогичной коммерчески доступному антителу к Her2, выработанному в клетках млекопитающих, т.е. клетках CHO, известному как трастузумаб.
Термин "Pichia антитело к Her2" или "антитело к Her2 из Pichia", применяемый в настоящем документе, относится к антителу с аминокислотной последовательностью, аналогичной коммерчески доступному антителу к Her2 (трастузумабу), выработанному в гликоинжиниринговых Pichia pastoris.
Термин "Fc-мутеин антитела", применяемый в настоящем документе, относится к антителу, включающему один из одиночных Fc-мутеинов или двойной Fc-мутеин, описываемые в настоящем документе.
Термин "Fc-мутеин", применяемый в настоящем документе, относится к Fc-содержащему полипептиду, в котором произведены одна или несколько точечных мутаций в Fc-области.
Термин "Fc-мутация", применяемый в настоящем документе, относится к мутации, сделанной в Fc-области Fc-содержащего полипептида. Примеры такой мутации включают мутации F243A или V264A, описываемые в настоящем документе.
Термин "одиночный Fc-мутеин", применяемый в настоящем документе, относится к Fc-содержащему полипептиду, включающему мутацию в положении 243 или 264 Fc-области. Термин "F243A" относится к мутации замены F (дикого типа) на A в положении 243 Fc-области Fc-содержащего полипептида. Термин "V264A" относится к мутации замены V (дикого типа) на A в положении 264 Fc-области Fc-содержащего полипептида. Положения 243 и 264 представляют собой положения аминокислот в CH2 домене Fc-области Fc-содержащего полипептида.
Термин "двойной Fc-мутеин", применяемый в настоящем документе, относится к Fc-содержащему полипептиду, включающему мутации в положениях 243 и 264 Fc-области. Термин "F243A/V264A" относится к двойному Fc-мутеину, включающему две точно указанные мутации.
На всем протяжении настоящего описания и формулы изобретения, нумерация остатков в тяжелой цепи иммуноглобулина или Fc-содержащего полипептида такая же, как в индексе EU по Kabat et al., Sequences of Proteins of Immunological Interest 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991), прямо включенной в настоящий документ в качестве ссылки. "Индекс EU по Kabat" относится к нумерации остатков человеческого антитела IgG1 EU.
Применяемый в настоящем документе термин "эффекторная функция" относится к биохимическому событию, которое является результатом взаимодействия Fc-области антитела с Fc-рецептором или лигандом. Типичные "эффекторные функции" включают связывание с C1q; обусловленную комплементом цитотоксичность; связывание с Fc-рецептором; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; негативную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора; BCR), и т.д. Такие эффекторные функции можно оценивать при помощи различных анализов, известных в данной области.
Термин "гликоинжиниринговый Pichia pastoris", применяемый в настоящем документе, относится к штамму Pichia pastoris, который был генетически изменен для экспрессии человекоподобных N-гликанов. Например, вышеописанные штаммы GFI5.0, GFI5.5 и GFI6.0.
Термины "N-гликан", "гликопротеин" и "гликоформа", применяемые в настоящем документе, относятся к N-связанному олигосахариду, например, тому, который присоединен посредством аспарагин-N-ацетилглюкозаминовой связи к аспарагиновому остатку полипептида. Преобладающими сахарами, обнаруженными на гликопротеинах, являются глюкоза, галактоза, манноза, фукоза, N-ацетилгалактозамин (GalNAc), N-ацетилглюкозамин (GlcNAc) и сиаловая кислота (SA, в том числе NANA, NGNA и их производные и аналоги, включая ацетилированную NANA или ацетилированную NGNA). В гликоинжинирингововом Pichia pastoris сиаловая кислота представляет собой исключительно N-ацетилнейраминовую кислоту (NANA) (Hamilton et al., Science 313 (5792): 1441-1443 (2006)). N-гликаны имеют общее пентасахаридное ядро Man3GlcNAc2, где "Man" относится к маннозе, "Glc" относится к глюкозе, "NAc" относится к N-ацетилу, и GlcNAc относится к N-ацетилглюкозамину. N-гликаны различаются в отношении числа ветвей (антенн), содержащих периферические сахара (например, GlcNAc, галактозу, фукозу и сиаловую кислоту), которые добавляют к коровой структуре Man3GlcNAc2 ("Man3"), которую также обозначают как "коровая триманноза", "коровый пентасахарид" или "олигоманнозное ядро". N-гликаны классифицируют в соответствии с составляющими их ветвей (например, с высоким содержанием маннозы, комплексного типа или гибридные).
Применяемый в настоящем документе термин "сиаловая кислота" или "SA" относится к любому члену семейства сиаловой кислоты, включая в качестве неограничивающих примеров: N-ацетилнейраминовую кислоту (Neu5Ac или NANA), N-гликолилнейраминовую кислоту (NGNA) и их любой аналог или производное (включая полученные путем ацетилирования по любой позиции в молекуле сиаловой кислоты). Сиаловая кислота является общим названием для группы приблизительно из 30-ти природных кислых углеводов, которые являются необходимыми компонентами огромного числа гликоконъюгатов. Schauer, Biochem. Society Transactions, 11, 270-271 (1983). Сиаловые кислоты, как правило, являются концевым остатком олигосахаридов. N-ацетилнейраминовая кислота (NANA) является наиболее распространенной формой сиаловой кислоты, и N-гликолилнейраминовая кислота (NGNA) является второй наиболее распространенной формой. Schauer, Glycobiologv, 1, 449-452 (1991). NGNA широко распространена на всем протяжении царства животных и, в зависимости от видов и ткани, часто составляет значительную долю сиаловой кислоты, связанной в гликоконъюгатах. Определенные виды, такие как курица и человек, являются исключением, поскольку у них в нормальных тканях отсутствует NGNA. Corfield, et al., Cell Biology Monographs, 10, 5-50 (1982). Сообщают, что в образцах сыворотки человека процентное содержание сиаловой кислоты в форме NGNA составляет 0,01% от общего количества сиаловой кислоты. Schauer, "Sialic Acids as Antigenic Determinants of Complex Carbohydrates", находящаяся в The Molecular Immunology of Complex Carbohydrates, (Plenum Press, New York, 1988).
Применяемый в настоящем документе термин "человекоподобный N-гликан" относится к N-связанным олигосахаридам, которые напоминают олигосахариды, полученные из клеток человека дикого типа, не подвергавшихся биоинженерии. Например, Pichia pastoris дикого типа и другие клетки низших эукариот, как правило, продуцируют белки с высоким содержанием маннозы в N-участках гликозилирования. Клетки-хозяева, описываемые в настоящем документе, производят гликопротеины (например, антитела), которые содержат человекоподобные N-гликаны, которые не являются гиперманнозилированными. В некоторых вариантах осуществления клетки-хозяева по настоящему изобретению способны к выработке человекоподобных N-гликанов с гибридным или комплексным типом N-гликанов. Конкретный тип "человекоподобных" гликанов, присутствующих на конкретном гликопротеине, произведенном клеткой-хозяином по изобретению, будет зависеть от специфических гликоинженерных шагов, которые проводят в клетке-хозяине.
Термин N-гликан "с высоким содержанием маннозы", применяемый в настоящем документе, относится к N-гликану с пятью или более маннозными остатками.
Термин N-гликан "комплексного" типа, применяемый в настоящем документе, относится к N-гликану, имеющему, по меньшей мере, один GlcNAc, присоединенный к 1,3 маннозному плечу, и, по меньшей мере, один GlcNAc, присоединенный к 1,6 маннозному плечу коровой триманнозы. N-гликаны комплексного типа могут также содержать остатки галактозы ("Gal") или N-ацетилгалактозамина ("GalNAc"), которые необязательно модифицированы сиаловой кислотой или ее производными (например, "NANA" или "NeuAc", где "Neu" относится к нейраминовой кислоте и "Ac" относится к ацетилу). N-гликаны комплексного типа могут также содержать внутрицепочечные замены, включающие "рассекающий" GlcNAc и коровую фукозу ("Fuc"). В качестве примера, когда N-гликан содержит рассекающий GlcNAc на коровой триманнозе, структура может быть представлена в виде Man3GlcNAc2(GlcNAc) или Man3GlcNAc3. Когда N-гликан содержит коровую фукозу, присоединенную к коровой триманнозе, структура может быть представлена в виде Man3GlcNAc2(Fuc). N-гликаны комплексного типа могут также иметь множественные «антенны» на "коровой триманнозе", часто обозначаемые как "гликаны с множественными антеннами".
Термин "гибридный" N-гликан, применяемый в настоящем документе, относится к N-гликану, имеющему, по меньшей мере, один GlcNAc на конце 1,3 маннозного плеча коровой триманнозы и ни одной или больше, чем одну маннозу на 1,6 маннозном плече коровой триманнозы.
Когда речь идет о "моль проценте" гликана, присутствующего в препарате гликопротеина, термин означает молярный процент конкретного гликана, присутствующего в совокупности N-связанных олигосахаридов, которые высвобождали, когда препарат белка обрабатывали ПНГазой, а затем оценивали количество способом, который не влияет на гликоформную композицию, (например, мечение совокупности гликанов, высвободившихся после обработки ПНГазой, флуоресцентной меткой, такой как 2-аминобензамид, а затем разделение при помощи высокоэффективной жидкостной хроматографии или капиллярного электрофореза, а затем определение количества гликанов по интенсивности флуоресценции). Например, 50 моль процент NANA2Gal2GlcNAc2Man3GlcNAc2 означает, что 50 процентов из высвободившихся гликанов представляют собой NANA2Gal2GlcNAc2Man3GlcNAc2 и оставшиеся 50 процентов состоят из других N-связанных олигосахаридов.
Применяемый в настоящем документе термин "противовоспалительное антитело" относится к антителу, предназначенному для применения при лечении воспаления. Противовоспалительные свойства Fc-содержащего полипептида можно измерять с использованием любого известного в данной области способа. В одном из вариантов осуществления противовоспалительные свойства Fc-содержащего полипептида измеряют с использованием моделей на животных, таких как модели, описанные в Kaneko et al., Science 313: 670-673 (2006), Anthony et al., Science 320: 373-376 (2008), и примерах 20-21 в настоящем документе. В другом варианте осуществления противовоспалительные свойства Fc-содержащего полипептида измеряют путем определения уровня биомаркера, связанного с воспалением (включая в качестве неограничивающих примеров CRP, провоспалительные цитокины, такие как фактор некроза опухолей (TNF-альфа), интерферон-гамма, интерлейкин 6 (IL-6), IL-8, IL-10, хемокины, маркер коагуляции D-димер, sCD14, кишечный пептид, связывающий жирные кислоты (IFABP) и гиалуроновую кислоту). В одном из вариантов осуществления противовоспалительные свойства Fc-содержащего полипептида измеряют путем определения уровня C-реактивного белка (CRP), с использованием известного в данной области способа. Снижение уровня C-реактивного белка указывает на то, что Fc-содержащий полипептид имеет противовоспалительные свойства.
"Консервативно модифицированные варианты" или "консервативная замена" относится к заменам аминокислот в белке другими аминокислотами со сходными характеристиками (например, заряд, размер боковой цепи, гидрофобность/гидрофильность, конформация остова и жесткость, и т.д.), таким образом, что замены часто могут быть произведены без изменения биологической активности белка. Специалисты в данной области признают, что в основном единичные замены аминокислот в несущественных областях полипептида по существу не изменяют биологическую активность (см., например, Watson et al. (1987) Molecular Biology of Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.)). Кроме того, замены структурно или функционально сходных аминокислот имеют меньше шансов нарушить биологическую активность. Типичные консервативные замены перечислены ниже:
Показано, что гликозилирование иммуноглобулина G (IgG) в Fc-области, по Asn297 (согласно системе нумерации EU), является необходимым для оптимального распознавания и активации эффекторных путей, в том числе антителозависимой клеточной цитотоксичности (ADCC) и обусловленной комплементом цитотоксичности (CDC), Wright & Morrison, Trends in Biotechnology. 15: 26-31 (1997), Tao & Morrison, J. Immunol, 143(8): 2595-2601 (1989). В связи с этим, гликоинжиниринг в константной области IgG стал областью активных исследований по разработке терапевтических моноклональных антител (МАТ). Установлено, что наличие N-связанного гликозилирования по Asn297 является критичным для активности МАТ в анализах иммунной эффекторной функции, включая ADCC, Rothman (1989), Lifely et al., Glycobiology, 5: 813-822 (1995), Umana (1999), Shields (2002), и Shinkawa (2003), и обусловленную комплементом цитотоксичность (CDC), Hodoniczky et al., Biotechnol Pros., 21(6): 1644-1652 (2005), и Jefferis et al., Chem. Immunol., 65: 111-128 (1997). Это влияние на функцию приписывают специфической конформации, которую принимает гликозилированный Fc-домен, и которая, по-видимому, пропадает, когда отсутствует гликозилирование. Более конкретно, IgG, в котором отсутствует гликозилирование в CH2-домене Fc-области, не связывается с FcγR, включая FcγRI, FcγRII и FcγRIII, Rothman (1989).
Похоже, что не только наличие гликозилирования играет роль в эффекторной функции антитела, но также важна конкретная композиция N-связанного олигосахарида. Например, наличие фукозы оказывает выраженное влияние на связывание с FcγRIIIa in vitro и ADCC in vitro, Rothman (1989), и Li et al., Nat. Biotechnol. 24(2): 2100-215 (2006). Рекомбинантные антитела, которые производятся клеточной культурой млекопитающих, такой как CHO или NS0, содержат N-связанные олигосахариды, которые в основном фукозилированы, Hossler et al., Biotechnology и Bioengineering, 95(5): 946-960 (2006), Umana (1999), и Jefferis et al., Biotechnol. Prog. 21: 11-16 (2005). Дополнительно, существуют доказательства, что сиалирование в Fc-области может придавать антителам противовоспалительные свойства. Внутривенный иммуноглобулин (IVIG), очищенный при помощи лектиновой колонки для обогащения сиалированной формой, демонстрировал четко выраженный противовоспалительный эффект, относящийся к сиалированному Fc-фрагменту, и связанный с повышением экспрессии ингибиторного рецептора FcγRIIb, Nimmerjahn и Ravetch., J. Exp. Med. 204: 11-15 (2007).
Гликозилирование в Fc-области антитела, полученного из клеточных линий млекопитающего, как правило, состоит из гетерогенной смеси гликоформ, с преобладающими формами, как правило, включающими сложные фукозилированные гликоформы: G0F, GIF и, в меньшей степени, G2F. Возможные условия, приводящие к неполному переносу галактозы на структуру G0F, в качестве неограничивающих примеров включают не оптимальный аппарат переноса галактозы, такой как β-1,4-галактозилтрансфераза, и плохой UDP-галактозный транспорт в аппарат Гольджи, субоптимальная клеточная культура и условия экспрессии белка, и стерическое препятствие из-за аминокислотных остатков, соседствующих с олигосахаридом. В то время как каждое из этих условий может модулировать окончательный уровень концевой галактозы, считают, что последующий перенос сиаловой кислоты на Fc-олигосахарид ингибируется закрытой конфигурацией желоба CH2-домена. См., например, Фиг. 1, Jefferis, R., Nature Biotech., 24 (10): 1230-1231, 2006. Без правильного концевого моносахарида, в частности, галактозы, или при недостаточно галактозилированных концевых формах, очень мала возможность получения сиалированной формы, способной действовать в качестве терапевтического белка, даже если она вырабатывается в присутствии сиалилтрансферазы. Белковая инженерия и структурный анализ Fc-гликоформ человеческих IgG показали, что профили гликозилирования находятся под влиянием конформации Fc, так как обнаружили, что можно достичь повышенных уровней галактозы и сиаловой кислоты на олигосахаридах из IgG3, выработанных CHO, когда конкретные аминокислоты из Fc-желоба мутированы в аланин, в том числе F241, F243, V264, D265 и R301. Lund et al., J. Immunol. 157(11): 4963-4969 (1996). Дополнительно показано, что определенные мутации оказывают некоторое действие на опосредованную клетками генерацию супероксида и опосредованный комплементом лизис красных клеток крови, которые используют в качестве суррогатных маркеров связывания с FcγRI и C1q, соответственно.
Опубликовано, что методами генетической инженерии могут быть созданы дрожжи для получения штаммов-хозяев, способных секретировать гликопротеины с высоко однородным гликозилированием. Choi et al., PNAS, USA 100(9): 5022-5027 (2003) описывают применение библиотек каталитических доменов α1,2-маннозидазы и каталитических доменов N-ацетилглюкозаминилтрансферазы I в комбинации с библиотекой лидерных последовательностей мембранного белка грибов типа II для определения местонахождения каталитических доменов в секреторных путях. Таким образом, были изолированы штаммы, которые вырабатывают in vivo гликопротеины с однородными Man5GlcNAc2 или GlcNAcMan5GlcNAc2 N-гликановыми структурами. Hamilton et al., Science 313 (5792): 1441-1443 (2006) описали выработку гликопротеина, эритропоэтина, в Pichia pastoris, как получение гликановой композиции, которая состоит преимущественно из бисиалированной гликановой структуры, GS6.0, NANA2Gal2GlcNAc2Man3GlcNAc2 (90,5%) и моносиалированной, GS5.5, NANAGal2GlcNAc2Man3GlcNAc2 (7,9%). Однако антитело, которое производится тем же штаммом, имеет значительно более низкое содержание сиалированного N-гликана из-за относительно низкого уровня концевого галактозного субстрата в антителе, как видно на фигуре 4. Кроме того, недавно было показано, что сиалирование Fc-олигосахарида наделяет противовоспалительными свойствами терапевтический внутривенный гамма-глобулин и его Fc-фрагменты, Kaneko et al., Science 313(5787): 670-673 (2006), и что противовоспалительное действие зависит от α2,6-, но не от α2,3-связанной формы сиаловой кислоты, Anthony et al., Science, 320: 373-376 (2008).
Организмы-хозяева и клеточные линии-хозяева
Fc-содержащие полипептиды по настоящему изобретению могут быть произведены в любом организме-хозяине или клеточной линии-хозяине. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению производят в клетке-хозяине, которая способна вырабатывать сиалированные N-гликаны.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению производят в клетке млекопитающего, где клетка по причине генетических или технологических воздействий или эндогенно вырабатывает гликопротеины, содержащие или смесь концевых α2,6- и α2,3-сиаловых кислот, или только концевую α2,6-сиаловую кислоту. Наращивание клеток млекопитающих в культуре (тканевая культура) стало рутинной процедурой. Примеры подходящих клеточных линий-хозяев от млекопитающих представляют собой линию клеток почки обезьяны CV1, трансформированную SV40 (COS-7, ATCC CRL 1651); линию клеток эмбриональной почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре); клетки почки детеныша хомяка (BHК, ATCC CCL 10); клетки яичника китайского хомячка/-DHFR (CHO); клетки Сертоли мыши (TM4,); клетки почки обезьяны (CV1, ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы buffalo (BRL3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (HepG2, HB 8065); опухоль молочной железы мыши (MMT060562, ATCC CCL51); клетки TRI; клетки MRC5; клетки FS4; гибридомные клеточные линии; NS0; SP2/0; и линия гепатомы человека (HepG2).
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению можно производить в растительной клетке, которая сконструирована для выработки сиалированных N-гликанов. См., например, Cox et al., Nature Biotechnology (2006) 24, 1591-1597 (2006), и Castilho et al., J. Biol. Chem. 285(21): 15923-15930 (2010).
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению можно производить в клетке насекомого, которая сконструирована для выработки сиалированных N-гликанов. См., например, Harrison и Jarvis, Adv. Вирус Res. 68: 159-91 (2006).
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению можно производить в бактериальной клетке, которая сконструирована для выработки сиалированных N-гликанов. См., например, Lizak et al., Bioconjugate Chem. 22: 488-496 (2011).
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению можно производить в клетке-хозяине или организме, относящемся к низшим эукариотам. Последние разработки позволяют производить полностью гуманизированные терапевтические средства в организмах-хозяевах, относящихся к низшим эукариотам, дрожжах и нитевидных грибах, таких как Pichia pastoris, Gerngross et al. патент США 7029872 и патент США № 7449308, содержание которых включено, таким образом, в качестве ссылки. См. также Jacobs et al., Nature Protocols 4(l): 58-70 (2009). Авторы заявки в настоящем документе дополнительно разработали модифицированные организмы-хозяева и клеточные линии-хозяева Pichia pastoris, способные экспрессировать антитела, содержащие две аминокислотные мутации в положениях 243 и 264 в Fc-области тяжелой цепи. Антитела с этими мутациями имеют повышенные уровни и более гомогенный состав α2,6-связанных сиалированных N-гликанов по сравнению с родительским антителом. Кроме того, авторы заявки неожиданно обнаружили, что аминокислотные мутации в положениях 243 и 264 в Fc-области тяжелой цепи приводят к образованию антитела, которое обладает пониженным связыванием со всеми рецепторами Fcγ и пониженным связыванием с C1q, последний из которых является заместителем для ADCC, которая независима от повышенных уровней α2,6-связанной сиаловой кислоты. Таким образом, основываясь на повышенном уровне и большей однородности N-гликана с концевой α2,6-связанной сиаловой кислотой, специалисты в данной области поймут и примут во внимание, что материалы и способы, описываемые в настоящем документе, можно использовать для производства рекомбинантных гликозилированных антител в клетках низших эукариот, таких как дрожжи и нитевидные грибы, и, в частности, Pichia pastoris, антител, которые имеют усиленные противовоспалительные свойства по сравнению с родительским антителом.
В связи со сниженным связыванием с FcγR и C1q, материалы и способы, описываемые в настоящем документе, можно использовать для производства рекомбинантных гликозилированных антител со сниженной эффекторной функцией по сравнению с родительским антителом. Антитела, производящиеся таким образом в Pichia pastoris при помощи способов по изобретению, вырабатываются в большом количестве со сниженной эффекторной функцией, и имеют преобладающие виды гликопротеина с концевым остатком α2,6-связанной сиаловой кислоты по сравнению с антителами, произведенными в гликоинжиниринговых клетках Pichia pastoris, в которых отсутствуют конкретные Fc-мутации, или в клетках-хозяевах Pichia pastoris, сохранивших свой эндогенный аппарат гликозилирования.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению производится в клетке-хозяине, более предпочтительно клетке-хозяине дрожжей или нитчатых грибов, которая сконструирована для выработки гликопротеинов с преобладающим N-гликаном, включающим концевую сиаловую кислоту. В одном из вариантов осуществления изобретения преобладающий N-гликан представляет собой α2,6-связанную форму SA2Gal2GlcNAc2Man3GlcNAc2, производящуюся в гликоинжиниринговых штаммах с α2,6-сиалилтрансферазой, которая не производит никакой α2,3-связанной сиаловой кислоты. В других вариантах осуществления штамм сконструирован для экспрессии α2,3-сиалилтрансферазы отдельно или в комбинации с α2,6-сиалилтрансферазой, что приводит к появлению α2,3-связанной или к комбинации α2,6- и α2,3-связанной сиаловой кислоты в качестве преобладающих N-гликанов.
Клеточные линии, которые используют для производства Fc-содержащих полипептидов по изобретению, могут быть любой клеточной линией, в частности, клеточными линиями со способностью вырабатывать один или несколько сиалированных гликопротеинов. Специалисты в данной области понимают, что материалы и способы, описываемые в настоящем документе, не ограничены конкретным штаммом Pichia pastoris, приводимом в качестве примера в настоящем документе, но могут включать любой штамм Pichia pastoris или штаммы других дрожжей или нитчатых грибов, которые производят N-гликаны с одной или несколькими концевыми галактозами, такими как Gal2GlcNAc2Man3. Концевая галактоза действует как субстрат для получения α2,6-связанной сиаловой кислоты, приводя к N-гликановой структуре NANA2Gal2GlcNAc2Man3GlcNAc2. Примеры подходящих штаммов описаны в патенте США № 7029872, US 2006-0286637 и Hamilton et al., Science 313 (5792): 1441-1443 (2006), содержание которых включено в настоящий документ, как если бы они были полностью здесь изложены.
В основном, низшие эукариоты, такие как дрожжи, применяют для экспрессии белков, в частности гликопротеинов, потому что их можно дешево культивировать, они дают высокий выход продукта, и при модификации соответствующим образом способны к подходящему гликозилированию. Дрожжи, в частности, обладают хорошо известной генетикой, позволяющей проводить быстрые трансформации, использовать стратегии по локализации исследуемого белка и не требующие усилий методы генетического нокаута. Подходящие векторы имеют последовательности контроля экспрессии, такие как промоторы, включая 3-фосфоглицераткиназный или другие ферменты гликолиза, и участок начала репликации, терминирующие последовательности и т.п. по желанию.
В то время как изобретение показано в настоящем документе с использованием метилотрофных дрожжей Pichia pastoris, подходящие клетки-хозяева низших эукариот включают Pichia pastoris, Pichia finlandica, Pichia trehalophila, Pichia koclamae, Pichia membranaefaciens, Pichia minuta (Ogataea minuta, Pichia lindneri), Pichia opuntiae, Pichia thermotolerans, Pichia salictaria, Pichia guercuum, Pichia pijperi, Pichia stiptis, Pichia methanolica, Pichia sp., Saccharomyces cerevisiae, Saccharomyces sp., Hansenula polymorpha, Kluyveromyces sp., Kluyveromyces lactis, Candida albicans, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Trichoderma reesei, Chrysosporiumi lucknowense, Fusarium sp., Fusarium gramineum, Fusarium venenatum и Neurospora crassa. Различные дрожжи, такие как K. lactis, Pichia pastoris, Pichia methanolica и Hansenula polymorpha, являются особенно подходящими для клеточной культуры, поскольку они способны расти с высокой плотностью клеток и секретировать большие количества рекомбинантного белка. Аналогично, для производства гликопротеинов по изобретению в промышленном масштабе можно использовать нитевидные грибы, такие как Aspergillus niger, Fusarium sp., Neurospora crassa и другие.
Низшие эукариоты, в частности, дрожжи и нитевидные грибы, могут быть генетически модифицированы таким образом, чтобы они экспрессировали гликопротеины, в которых паттерн гликозилирования является человекоподобным или гуманизированным. Как указано выше, применяемый в настоящем документе термин "человекоподобные N-гликаны" относится к N-связанным олигосахаридам, которые напоминают олигосахариды, полученные из клеток человека дикого типа, не подвергавшихся биоинженерии. В предпочтительных вариантах осуществления настоящего изобретения, клетки-хозяева по настоящему изобретению способны к выработке человекоподобных гликопротеинов с N-гликанами гибридного и/или комплексного типа; т.е. к "человекоподобному N-гликозилированию". Какие конкретные "человекоподобные" гликаны будут преимущественно присутствовать на гликопротеинах, произведенных в клетках-хозяевах по изобретению, зависит от конкретных проведенных биоинженерных шагов. Таким образом, можно получать гликопротеиновые композиции, в которых конкретная желаемая гликоформа является преобладающей в композиции. Это может быть достигнуто за счет удаления выбранных эндогенных ферментов гликозилирования и/или способами генетической инженерии в отношении клеток-хозяев, и/или добавлением экзогенных ферментов для имитации всего пути гликозилирования млекопитающих или его части, как описано в патенте США № 7449308. При желании можно проводить дополнительные шаги генетической инженерии в области гликозилирования, таким образом, чтобы гликопротеин можно было получать с наличием или отсутствием корового фукозилирования. Применение клеток-хозяев низших эукариот имеет дополнительное преимущество в том, что эти клетки способны производить высоко гомогенные композиции гликопротеинов, таким образом, что преобладающая гликоформа гликопротеина может присутствовать более чем в тридцати моль процентах гликопротеина в композиции. В конкретных аспектах преобладающая гликоформа гликопротеина может присутствовать более чем в сорока моль процентах, пятидесяти моль процентах, шестидесяти моль процентах, семидесяти моль процентах и, наиболее предпочтительно, более чем в восьмидесяти моль процентах гликопротеина, присутствующего в композиции.
Низшие эукариоты, в частности дрожжи, могут быть генетически модифицированы таким образом, чтобы они экспрессировали гликопротеины, в которых паттерн гликозилирования является человекоподобным или гуманизированным. Это может быть достигнуто за счет удаления выбранных эндогенных ферментов гликозилирования ферменты и/или добавления экзогенных ферментов, как описано у Gerngross et al., патент США № 7449308. Например, клетку-хозяина можно выбрать или сконструировать с уменьшенной активностью αl,6-маннозилтрансферазы, которая в противном случае будет добавлять остатки маннозы на N-гликан на гликопротеине.
В одном из вариантов осуществления клетка-хозяин дополнительно включает каталитический домен αl,2-маннозидазы, слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности αl,2-маннозидазы на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу Man5GlcNAc2, например, рекомбинантную гликопротеиновую композицию, включающую преимущественно гликоформу Man5GlcNAc2. Например, патенты США №№ 7029872 и 7449308 и опубликованная патентная заявка США № 2005/0170452 описывают клетки-хозяева низших эукариот, способные вырабатывать гликопротеин, включающий гликоформу Man5GlcNAc2.
В дополнительном варианте осуществления непосредственно предшествующая клетка-хозяин дополнительно включает каталитический домен GlcNAc-трансферазы I (GnT I), слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности GlcNAc трансферазы I на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу GlcNAcMan5GlcNAc2, например рекомбинантную гликопротеиновую композицию, включающую преимущественно гликоформу GlcNAcMan5GlcNAc2. Патенты США №№ 7029872 и 7449308 и опубликованная патентная заявка США № 2005/0170452 описывают клетки-хозяева низших эукариот, способные вырабатывать гликопротеин, включающий гликоформу GlcNAcMan5GlcNAc2. Гликопротеин, произведенный в вышеуказанных клетках, можно обрабатывать гексозаминидазой in vitro для получения рекомбинантного гликопротеина, включающего гликоформу Man5GlcNAc2.
В дополнительном варианте осуществления непосредственно предшествующая клетка-хозяин дополнительно включает каталитический домен маннозидазы II, слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности маннозидазы II на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу GlcNAcMan3GlcNAc2, например, рекомбинантная гликопротеиновая композиция, включающая преимущественно гликоформу GlcNAcMan3GlcNAc2. Патент США № 7029872 и опубликованная патентная заявка США № 2004/0230042 описывают клетки-хозяева низших эукариот, которые экспрессируют фермент маннозидаза II и способны к выработке гликопротеинов преимущественно с гликоформой GlcNAcMan3GlcNAc2. Гликопротеин, произведенный в вышеуказанных клетках можно обрабатывать гексозаминидазой in vitro для получения рекомбинантного гликопротеина, включающего гликоформу Man3GlcNAc2.
В дополнительном варианте осуществления непосредственно предшествующая клетка-хозяин дополнительно включает каталитический домен GlcNAc-трансферазы II (GnT II), слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности GlcNAc трансферазы II на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу GlcNAc2Man3GlcNAc2, например, рекомбинантную гликопротеиновую композицию, включающую преимущественно гликоформу GlcNAc2Man3GlcNAc2. Патенты США №№ 7029872 и 7449308 и опубликованная патентная заявка США № 2005/0170452 описывают клетки-хозяева низших эукариот, способные вырабатывать гликопротеин, включающий гликоформу GlcNAc2Man3GlcNAc2. Гликопротеин, произведенный в вышеуказанных клетках можно обрабатывать гексозаминидазой in vitro для получения рекомбинантного гликопротеина, включающего гликоформу Man3GlcNAc2.
В дополнительном варианте осуществления непосредственно предшествующая клетка-хозяин дополнительно включает каталитический домен галактозилтрансферазы, слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности галактозилтрансферазы на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу GalGlcNAc2Man3GlcNAc2 или Gal2GlcNAc2Man3GlcNAc2, или их смесь, например, рекомбинантную гликопротеиновую композицию, включающую преимущественно гликоформу GalGlcNAc2Man3GlcNAc2 или Gal2GlcNAc2Man3GlcNAc2 или их смесь. Патент США № 7029872 и опубликованная патентная заявка США № 2006/0040353 описывают клетки-хозяева низших эукариот, способные вырабатывать гликопротеин, включающий гликоформу Gal2GlcNAc2Man3GlcNAc2. Гликопротеин, произведенный в вышеуказанных клетках, можно обрабатывать галактозидазой in vitro для получения рекомбинантного гликопротеина, включающего гликоформу GlcNAc2Man3GlcNAc2, например, рекомбинантной гликопротеиновой композиции, включающей преимущественно гликоформу GlcNAc2Man3GlcNAc2.
В дополнительном варианте осуществления непосредственно предшествующая клетка-хозяин дополнительно включает каталитический домен сиалилтрансферазы, слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности сиалилтрансферазы на ЭР или аппарат Гольджи клетки-хозяина. В предпочтительном варианте осуществления сиалилтрансфераза представляет собой альфа 2,6-сиалилтрансферазу. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий преимущественно гликоформу NANA2Gal2GlcNAc2Man3GlcNAc2 или NANAGal2GlcNAc2Man3GlcNAc2 или их смесь. Клетки-хозяева низших эукариот, такие как дрожжи и нитевидные грибы, являются подходящими, если клетка-хозяин дополнительно включает способы получения CMP-сиаловой кислоты для переноса на N-гликан. Опубликованная патентная заявка США № 2005/0260729 раскрывает способ генетической инженерии низших эукариот для создания пути синтеза CMP-сиаловой кислоты, и опубликованная патентная заявка США № 2006/0286637 раскрывает способ генетической инженерии низших эукариот для выработки сиалированных гликопротеинов. Для увеличения сиалирования предпочтительным может являться создание клетки-хозяина, содержащей две или более копий пути синтеза CMP-сиаловой кислоты или две или более копий сиалилтрансферазы. Гликопротеин, произведенный в вышеуказанных клетках, можно обрабатывать нейраминидазой in vitro для получения рекомбинантного гликопротеина, включающего преимущественно гликоформу Gal2GlcNAc2Man3GlcNAc2 или гликоформу GalGlcNAc2Man3GlcNAc2 или их смесь.
Любая из предыдущих клеток-хозяев может дополнительно содержать одну или несколько GlcNAc-трансфераз, выбранных из группы, состоящей из GnT III, GnT IV, GnT V, GnT VI и GnT IX, для производства гликопротеинов с рассекающими (GnT III) и/или многоантенными (GnT IV, V, VI и IX) N-гликановыми структурами такими, как описано в опубликованных патентных заявках США №№ 2005/0208617 и 2007/0037248. Дополнительно, предшествующие клетки-хозяева могут производить рекомбинантные гликопротеины (например, антитела), включающие SA(l-4)Gal(l-4)GlcNAc(2-4)Man3GlcNAc2, в том числе антитела, содержащие NANA(l-4)Gal(l-4)GlcNAc(2-4)Man3GlcNAc2, NGNA(l-4)Gal(l-4)GlcNAc(2-4)Man3GlcNAc2 или комбинацию NANA(l-4)Gal(l-4)GlcNAc(2-4)Man3GlcNAc2 и NGNA(l-4)Gal(l-4)GlcNAc(2-4)Man3GlcNAc2. В одном из вариантов осуществления рекомбинантный гликопротеин содержит N-гликаны, включающие структуру, выбранную из группы, состоящей из SA(l-4)Gal(l-4)GlcNAc(2-4)Man3GlcNAc2, и лишенную любых α2,3-связанных сиаловых кислот.
В дополнительных вариантах осуществления клетка-хозяин, которая производит гликопротеины преимущественно с GlcNAcMan5GlcNAc2 N-гликанами, дополнительно включает каталитический домен галактозилтрансферазы, слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности галактозилтрансферазы на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу GalGlcNAcMan5GlcNAc2.
В дополнительном варианте осуществления непосредственно предшествующая клетка-хозяин, которая производит гликопротеины преимущественно с GalGlcNAcMan5GlcNAc2 N-гликанами дополнительно включает каталитический домен сиалилтрансферазы, слитый с сигнальным пептидом для клеточного таргетинга, который обычно не ассоциирован с каталитическим доменом и выбран для нацеливания активности сиалилтрансферазы на ЭР или аппарат Гольджи клетки-хозяина. Прохождение рекомбинантного гликопротеина через ЭР или аппарат Гольджи клетки-хозяина создает рекомбинантный гликопротеин, включающий гликоформу SAGalGlcNAcMan5GlcNAc2 (например, NANAGalGlcNAcMan5GlcNAc2 или NGNAGalGlcNAcMan5GlcNAc2 или их смесь).
Любая из предшествующих клеток-хозяев может дополнительно включать один или несколько переносчиков сахаров, таких как переносчики UDP-GlcNAc (например, переносчики UDP-GlcNAc Kluyveromyces lactis и Mus musculus), переносчики UDP-галактозы (например, переносчик UDP-галактозы Drosophila melanogaster) и переносчик CMP-сиаловой кислоты (например, человеческий переносчик сиаловой кислоты). Поскольку в клетках-хозяевах низших эукариот, таких как дрожжи и нитевидные грибы, отсутствуют вышеуказанные переносчики, предпочтительно, чтобы клетки-хозяева низших эукариот, такие как дрожжи и нитевидные грибы, были созданы методами генетической инженерии для включения вышеуказанных переносчиков.
Дополнительно, с любой из предшествующих клеток-хозяев можно проводить дополнительные манипуляции для повышения заполнения N-гликанами. См., например, Gaulitzek et al., Biotechnol. Bioengin. 103: 1164-1175 (2009); Jones et al., Biochim. Biosphys. Acta 1726: 121-137 (2005); WO2006/107990. В одном из вариантов осуществления любая из предшествующих клеток-хозяев может быть дополнительно сконструирована для включения, по меньшей мере, одной молекулы нуклеиновой кислоты, кодирующей гетерологичную олигосахарилтрансферазу с одной субъединицей (например, белок STT3A, белок STT3B, белок STT3C, белок STT3D Leishmania sp. или их сочетания), и молекулы нуклеиновой кислоты, кодирующей гетерологичный гликопротеин, и где клетка-хозяин экспрессирует эндогенные гены клетки-хозяина, кодирующие белки, которые составляют эндогенный комплекс олигосахарилтрансферазы. В одном из вариантов осуществления любая из предшествующих клеток-хозяев может быть дополнительно сконструирована для включения, по меньшей мере, одной молекулы нуклеиновой кислоты, кодирующей белок STT3D Leishmania sp., и молекулы нуклеиновой кислоты, кодирующей гетерологичный гликопротеин, и где клетка-хозяин экспрессирует эндогенные гены клетки-хозяина, кодирующие белки, которые составляют эндогенный комплекс олигосахарилтрансферазы.
Клетки-хозяева дополнительно включают клетки низших эукариот (например, дрожжей, таких как Pichia pastoris), которые созданы методами генетической инженерии для выработки гликопротеинов, не содержащих N-гликанов, устойчивых к α-маннозидазе. Этого можно достигнуть за счет делеции или разрушения одного или нескольких генов β-маннозилтрансферазы (например, ΒΜТ1, BMT2, BMT3 и BMT4) (см., опубликованную патентную заявку США № 2006/0211085) и гликопротеинов, имеющих фосфоманнозные остатки, из-за делеции или разрушения одного или обоих генов фосфоманнозилтрансферазы PNO1 и MNN4B (см. например, патенты США №№ 7198921 и 7259007), что в дополнительных аспектах может также включать удаление или разрушение гена MNN4A. Разрушение включает разрушение открытой рамки считывания, кодирующей конкретные ферменты, или нарушение экспрессии открытой рамки считывания или отмену трансляции РНК, кодирующей одну или несколько β-маннозилтрансфераз и/или фосфоманнозилтрансфераз, с использованием интерферирующей РНК, антисмысловой РНК или т.п. Дополнительно, клетки могут производить гликопротеины с N-гликанами, устойчивыми к α-маннозидазе, за счет добавления химических ингибиторов или за счет модификаций условий культивирования клеток. Эти клетки-хозяева могут быть модифицированы дополнительно, как описано выше, для выработки конкретных N-гликановых структур.
Клетки-хозяева дополнительно включают клетки низших эукариот (например, дрожжей, таких как Pichia pastoris), которые генетически модифицированы для контроля O-гликозилирования гликопротеина за счет делеции или разрушения одного или нескольких белков O-маннозилтрансферазы (Do1-P-Man: (Ser/Thr) гены белка манозилтрансферазы) (PMT) (см. патент США № 5714377) или за счет роста в присутствии ингибиторов Pmtp и/или α-маннозидазы, как описано в опубликованной международной заявке № WO 2007/061631, или за счет обоих факторов. Разрушение включает разрушение открытой рамки считывания, кодирующей Pmtp, или нарушение экспрессии открытой рамки считывания или отмену трансляции РНК, кодирующей один или несколько Pmtp с использованием интерферирующей РНК, антисмысловой РНК или т.п. Клетки-хозяева могут дополнительно включать любую из указанных выше клеток-хозяев, модифицированных для выработки конкретных N-гликановых структур.
Ингибиторы Pmtp в качестве неограничивающих примеров включают бензилиден тиазолидиндионы. Примерами бензилиден тиазолидиндионов, которые можно использовать, являются 5-[[3,4-бис(фенилметокси)фенил]метилен]-4-оксо-2-тиоксо-3-тиазолидинуксусная кислота, 5-[[3-(1-фенилэтокси)-4-(2-фенилэтокси)]фенил]метилен]-4-оксо-2-тиоксо-3-тиазолидинуксусная кислота и 5-[[3-(1-фенил-2-гидрокси)этокси)-4-(2-фенилэтокси)]фенил]метилен]-4-оксо-2-тиоксо-3-тиазолидинуксусная кислота.
В конкретных вариантах осуществления функция или экспрессия, по меньшей мере, одного эндогенного гена PMT снижена, нарушена или выключена. Например, в конкретных вариантах осуществления функция или экспрессия, по меньшей мере, одного эндогенного гена PMT, выбранного из группы, состоящей из генов PMT1, PMT2, PMT3 и ΡΜТ4, снижена, нарушена или выключена; или клетки-хозяева культивируются в присутствии одного или нескольких ингибиторов PMT. В дополнительных вариантах осуществления клетки-хозяева содержат одну или несколько делеций или разрушений генов PMT, и клетки-хозяева культивируются в присутствии одного или нескольких ингибиторов Pmtp. В частных аспектах этих вариантов осуществления, клетки-хозяева также экспрессируют секретируемую α-1,2-маннозидазу.
Делеции или разрушения PMT и/или ингибиторы Pmtp управляют O-гликозилированием посредством снижения присутствия O-гликозилирования, а именно, посредством снижения общего числа O-участков гликозилирования на гликопротеин, которые гликозилированы. Дополнительное добавление α-1,2-маннозидазы, которая секретируется клеткой, управляет O-гликозилированием за счет уменьшения длины цепи маннозы O-гликанов, которые находятся на гликопротеине. Таким образом, комбинирование делеций или разрушений PMT и/или ингибиторов Pmtp с экспрессией секретируемой α-1,2-маннозидазы управляет O-гликозилированием посредством уменьшения присутствия и длины цепи. В конкретных обстоятельствах конкретная комбинация делеций или разрушения PMT, ингибиторы Pmtp и α-1,2-маннозидазы эмпирически определяет, как конкретные гетерологичные гликопротеины (Fab и антитела, например) можно экспрессировать и транспортировать посредством аппарата Гольджи с различной степенью эффективности, и, таким образом, может требоваться конкретная комбинация делеций или разрушений PMT, ингибиторов Pmtp и α-1,2-маннозидазы. В другом аспекте гены, кодирующие один или несколько эндогенных ферментов маннозилтрансферазы, удалены. Эта делеция (делеции) может быть в комбинации с предлагаемой секретируемой α-1,2-маннозидазой и/или ингибиторами PMT или может быть вместо предлагаемой секретируемой α-1,2-маннозидазы и/или ингибиторов PMT.
Таким образом, управление О-гликозилированием может быть полезно для производства конкретных гликопротеинов в клетках-хозяевах, описываемых в настоящем документе, для улучшения общего выхода продукта или для выработки собранного надлежащим образом гликопротеина. Уменьшение или устранение O-гликозилирования, по-видимому, оказывает положительное воздействие на сборку и транспорт антител целиком и Fab-фрагментов, в то время как они проходят по секреторному пути и транспортируются на поверхность клетки. Таким образом, в клетках с управляемым O-гликозилированием, выработка антител, собранных надлежащим образом, или Fab-фрагментов, увеличена по сравнению с выработкой, получаемой в клетках, где O-гликозилирование не контролируется.
Для уменьшения или устранения вероятности образования N-гликанов и O-гликанов с β-связанными остатками маннозы, которые устойчивы к α-маннозидазам, способами генетической инженерии были созданы рекомбинантные гликоинжиниринговые клетки-хозяева Pichia pastoris для удаления гликопротеинов с N-гликанами, устойчивыми к α-маннозидазе, посредством делеции или разрушения одного или нескольких генов β-маннозилтрансферазы (например, BMT1, BMT2, BMT3 и BMT4) (см., патент США № 7465577 и патент США № 7713719). Делеция или разрушение BMT2 и одного или нескольких из BMT1, BMT3 и BMT4 также снижает или устраняет выявляемую перекрестную реактивность к антителам против белка клетки-хозяина.
Выработка гликопротеина в некоторых случаях может быть улучшена за счет сверхэкспрессии молекул нуклеиновой кислоты, кодирующей белки-шапероны человека или млекопитающих, или за счет замещения генов, кодирующих один или несколько эндогенных белков-шаперонов, молекулами нуклеиновой кислоты, кодирующей один или несколько белков-шаперонов человека или млекопитающих. Кроме того, экспрессия белков-шаперонов человека или млекопитающих в клетке-хозяине также, по-видимому, управляет O-гликозилированием в клетке. Таким образом, в настоящий документ дополнительно включены клетки-хозяева, где функция, по меньшей мере, одного эндогенного гена, кодирующего белок-шаперон, снижена или устранена, и в клетке-хозяине экспрессируется вектор, кодирующий, по меньшей мере, один гомолог белка-шаперона человека или млекопитающего. Также включены клетки-хозяева, в которых экспрессируются эндогенные шапероны клетки-хозяина и белки-шапероны человека или млекопитающего. В дополнительных аспектах клетка-хозяин из низших эукариот представляет собой клетку-хозяина из дрожжей или нитевидных грибов. Примеры использования шаперонов клеток-хозяев, в которые введены белки-шапероны человека для повышения выработки и снижения или контролирования O-гликозилирования рекомбинантных белков, описаны в опубликованных международных заявках №№ WO 2009105357 и WO2010019487 (описания которых включены в настоящий документ в качестве ссылки). Как и выше, дополнительно включены клетки-хозяева из низших эукариот, где, в дополнение к замещению генов, кодирующих один или несколько эндогенных белков-шаперонов, молекулами нуклеиновой кислоты, кодирующей один или несколько белков-шаперонов человека или млекопитающего, или сверхэкспрессии одного или нескольких белков-шаперонов человека или млекопитающего, как описано выше, снижена, нарушена или удалена функция или экспрессия, по меньшей мере, одного эндогенного гена, кодирующего белок O-маннозилтрансферазы (PMT). В конкретных вариантах осуществления снижена, нарушена или удалена функция, по меньшей мере, одного эндогенного гена PMT, выбранного из группы, состоящей из генов PMT1, ΡΜT2, PMT3 и PMT4.
Кроме того, O-гликозилирование может оказывать воздействие на аффинность антитела или Fab-фрагмента и/или авидность к антигену. Это может быть особенно значимым, когда конечная клетка-хозяин для выработки антитела или Fab не та же самая, что клетка-хозяин, которую применяли для селекции антитела. Например, O-гликозилирование может вмешиваться в аффинность антитела или Fab-фрагмента к антителу, таким образом, что антитело или Fab-фрагмент, которые в ином случае имеют высокую аффинность к антигену, могут не распознавать его, потому что O-гликозилирование может мешать способности антитела или Fab-фрагмента связывать антиген. В других случаях антитело или Fab-фрагмент, которые имеют высокую авидность к антигену, могут его не распознавать, потому что O-гликозилирование мешает авидности антитела или Fab-фрагмента к антигену. В двух предыдущих случаях антитело или Fab-фрагмент, которые могут быть особенно эффективны, когда вырабатываются в клеточной линии млекопитающего, могут не обнаруживаться, поскольку клетки-хозяева для выявления и селекции антитела или Fab-фрагмента относились к другому типу клеток, например, клеткам дрожжей или грибов (например, клетка-хозяин Pichia pastoris). Хорошо известно, что O-гликозилирование у дрожжей может значительно отличаться от O-гликозилирования в клетках млекопитающих. Это особенно актуально при сравнении O-гликозилирования дикого типа у дрожжей с муциновым типом или дистрогликановым типом O-гликозилирования у млекопитающих. В конкретных случаях O-гликозилирование может усиливать аффинность или авидность антитела или Fab-фрагментов к антигену, вместо того чтобы мешать связыванию с антигеном. Этот эффект является нежелательным, когда продуцирующая клетка-хозяин отличается от клетки-хозяина, использованной для выявления и селекции антитела или Fab-фрагмента (например, выявление и селекция происходят в дрожжах, а продуцирующий хозяин представляет собой клетку млекопитающего), поскольку в продуцирующем хозяине O-гликозилирование уже не того типа, который усиливает аффинность или авидность к антигену. Таким образом, регулирование O-гликозилирования делает возможным использование материалов и способов в настоящем документе для выявления и селекции антител или Fab-фрагментов со специфичностью к конкретному антигену в зависимости от аффинности или авидности антитела или Fab-фрагмента к антигену без выявления и селекции антитела или Fab-фрагмента, подверженного влиянию системы O-гликозилирования клетки-хозяина. Таким образом, регулирование O-гликозилирования дополнительно усиливает полезность дрожжевых или грибных клеток-хозяев для выявления и селекции антител или Fab-фрагментов, которые в конечном итоге будут производиться в клеточной линии млекопитающего.
Специалисты в данной области понимают, как использовать способы и материалы, описываемые в настоящем документе, в комбинации с другими линиями Pichia pastoris и линиями дрожжевых клеток, которые созданы методами генетической инженерии для производства конкретных N-гликанов или сиалированных гликопротеинов, такими как, но без ограничений, вышеописанные организмы-хозяева и клеточные линии, которые были созданы методами генетической инженерии для производства конкретных галактозилированных или сиалированных форм. См., например, US 2006-0286637, «Выработка сиалированных N-гликанов в низших эукариотах», в котором был генетически модифицирован путь захвата и утилизации галактозы, как источника углерода, описание которого включено в настоящий документ, как если бы он был полностью здесь изложен.
Дополнительно, способы в настоящем документе можно использовать для выработки рекомбинантных Fc-содержащих полипептидов в других клеточных линиях низших эукариот, которые сконструированы для выработки человекоподобных и человеческих гликопротеинов, и которые не обладают α2,6-сиалилтрансферазной активностью. Способы также можно использовать для выработки рекомбинантных Fc-содержащих полипептидов в эукариотических клеточных линиях, у которых выработка сиалированных N-гликанов является врожденной особенностью.
Уровни α2,3- и α2,6-связанной сиаловой кислоты на Fc-содержащих полипептидах можно измерять с использованием хорошо известных способов, в том числе ядерного магнитного резонанса (ЯМР), высокоэффективной нормально-фазовой жидкостной хроматографии (ВЭЖХ) и высокоэффективной анионообменной хроматографии с импульсным амперометрическим детектированием (HPAEC-PAD).
Биологические свойства Fc-мутеинов
Для многих Fc-содержащих полипептидов желательными характеристиками являются отсутствие или значительное снижение эффекторной функции, как показано за счет сниженного связывания с FcγR и C1q, Idusogie et al., J. Immunology, 164(8): 4178-84 (2000), и Shields et al., J. Biol. Chem., 276: 6591-6604 (2001), и повышенные противовоспалительные свойства. Авторы заявки в настоящем изобретении открыли, что конкретные модификации аминокислотных положений 243 и 264 в Fc-области IgG могут вызывать потерю или значительное снижение эффекторной функции независимо от наличия сиалирования по концевым положениям гликана. В частности, авторы заявки обнаружили, что модификация остатков F243 и V264 в Fc-области на аланин приводит к образованию антитела с пониженным связыванием с рецепторами Fcγ и C1q. Следует отметить, что антитела, произведенные с этими модификациями Fc-области, демонстрируют пониженное связывание с рецепторами Fcγ, независимо от того, присутствует ли у них в качестве концевого гликана α2,6-связанная форма сиаловой кислоты.
В связи с этим, авторы заявки разработали двойной Fc-мутеин, F243A/V264A, который будет производить Fc-содержащие полипептиды с вышеуказанными желаемыми характеристиками. Примеры в настоящем документе включают трансформацию клетки-хозяина полинуклеотидным вектором, кодирующим Fc-содержащий полипептид, который включает мутации в положениях 243 и 264 Fc-области, и культивирование трансформированной клетки-хозяина для выработки Fc-содержащего полипептида.
Выработка Fc-содержащих полипептидов
Fc-содержащие полипептиды по изобретению можно производить согласно любому известному в данной области способу, подходящему для создания полипептидов, содержащих Fc-область. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело или фрагмент антитела (в том числе, без ограничений, полипептид, состоящий из или состоящий по существу из Fc-области антитела). В другом варианте осуществления Fc-содержащий полипептид представляет собой иммуноадгезин. Способы получения антитела и фрагментов антител хорошо известны в данной области. Способы введения точечных мутаций в полипептид, например, сайт-направленный мутагенез, также хорошо известны в данной области.
В примерах, описываемых в настоящем документе, тяжелая и легкая цепи IgG1, содержащие консенсусную последовательность CH2, и двойные Fc-мутанты, описываемые в настоящем документе, экспрессировались в двух различных гликоинжиниринговых штаммах Pichia pastoris. Как описано в последующих примерах, последовательности генов тяжелой и легкой цепей находились под контролем индуцируемого метанолом промотора, AOX1, и имели встроенный селективный маркер устойчивости к блеомицину (Зеоцин). Эта стратегия позволяет интегрировать экспрессирующую кассету целиком в локус Trp2 путем гомологичной рекомбинации ДНК.
Секретируемые антитела захватывали из ферментационного бульона посредством аффинной хроматографии с белком A с последующим шагом очистки при помощи катионного обмена посредством Source 30S. Очищенные антитела были охарактеризованы при помощи SDS-PAGE (фигура 2) и гель-хроматографии (СЕК) и обратно-фазовой ВЭЖХ для оценки надлежащей сборки. Как видно на фигуре 2, антитела, произведенные посредством материалов и способов по настоящему документу, имели на SDS-PAGE профиль чистоты, который был таким же, что и для антитела к Her2, произведенного клеткой млекопитающего (CHO). Анализ IgG посредством СЕК и обратно-фазовой ВЭЖХ дополнительно продемонстрировал, что антитела, полученные из Fc-мутеинов гликоинжиниринговых штаммов, были собраны надлежащим образом и в отношении сборки сходны с антителом, произведенным клеткой млекопитающего (данные не показаны). Аффинность к антигену для различных антител, созданных посредством материалов и способов по настоящему документу, определяли путем анализа на клетках с использованием клеточной линии SK-BR3, которая в этом случае представляла собой линию рака молочной железы человека со сверхэкспрессией Her2. Как и ожидалось, все антитела, включая Fc-мутеины, одинаково хорошо связывались с клеточной линией SK-BR3 (фигура 3).
Анализ N-гликанов Fc-мутеинов
Для множества гликопротеинов, включая конкретные антитела, сиалирование концевого N-связанного гликана Fc-области IgG является обязательным для выработки гликопротеинов и антител, которые имеют правильную конформацию для проявления терапевтической активности. См., например, Anthony et al., Science, 320: 373-376 (2008), где концевое сиалирование было скоррелировано с противовоспалительной активностью для получения IVIG. Сиалирование требует присутствия галактозы в предпоследнем положении, на которой сиалилтрансфераза действует для образования сиалированного гликана. Таким образом, гликопротеины с отсутствием одной или нескольких концевых гликоформ галактозы не могут производить антитела, имеющие композицию α2,6-связанной сиаловой кислоты, ассоциированную с противовоспалительной активностью.
Как правило, антитела, произведенные в клеточной культуре млекопитающего, такой как клетки CHO, имеют гликоформную композицию, включающую: G0F (37%), G1F (43%), G2F (9%), G0 (4%), Gl (3%) и Man5 (3%), у которой концевая галактоза отсутствует или ее мало для того, чтобы действовать в качестве субстрата для переноса сиаловой кислоты. В случае выработки клеткой CHO, концевой гликан вырабатывается в виде α2,3-связанной формы; клетки CHO не экспрессируют α2,6-сиалилтрансферазу, необходимую для получения α2,6-связанной формы сиаловой кислоты, которая ассоциирована с противовоспалительной активностью. Однако сверхэкспрессия специфической α2,6-сиалилтрансферазы в CHO может приводить к образованию смеси α2,3-связанной и α2,6-связанной сиаловых кислот (Bragonzi et al., BBA 1474: 273-282 (2000); Biochem. Biophys. Res. Comm. 289: 243-249 (2001)). Гликоинжиниринговые штаммы Pichia pastoris GFI5.0, которые способны к выработке высоких уровней галактозилированных белков, не являющихся антителами, таких как эритропоэтин (Hamilton et al., Science, 313: 1441-1443 (2006)), производят антитела с относительно низкими количествами концевой галактозы, которая может быть использована для образования α2,6-связанной сиалированной формы (фигура 4). Антитела, произведенные в таких штаммах Pichia pastoris, как правило, имеют композицию, включающую гликоформы G0 (60%), G1 (17%), G2 (4%) и Man5 (8%). Даже антитела, произведенные в штаммах Pichia pastoris GFI6.0, которые имеют гликановую композицию, включающую G0 (43,5%), G1 (20,8%), G2 (2,7%), NANAGalGlcNAcMan5GlcNAc2 (5,5%) и NANAGal2GlcNAc2Man3GlcNAc2 (4,9%), имеют относительно низкий уровень α2,6-связанной сиалированной формы. Таким образом, антитела, произведенные в щтаммах GFI5.0 и 6.0, имеют значительно более низкие уровни галактозилирования и сиалирования по сравнению с белками, которые не являются антителами (такие как эритропоэтин), произведенными в тех же самых штаммах.
N-гликановая композиция антитела к Her2 и соответствующие Fc-мутеины антитела, произведенные по настоящему изобретению в гликоинжиниринговых штаммах Pichia pastoris GFI5.0 и GFI6.0, анализировали посредством лазерной десорбции/ионизации матрицы с помощью время-пролетной (MALDI-TOF) масс-спектрометрии после высвобождения из антитела при помощи пептид-N-гликозидазы F (фигуры 4-8). Количественный состав высвободившейся углеводной композиции антител к Her2 и Fc-мутеинов антитела определяли при помощи ВЭЖХ на колонках Allentech Prevail carbo (Alltech Associates, Deerfield IL).
Гликоформы антитела к Her2 из штамма Pichia pastoris GFI5.0 включали биантенные G0, G1 и G2 и другие нейтральные гликаны, с G0 в качестве преобладающей гликоформы (фигура 4). Эта гликановая композиция соответствует той, что производится на коммерческой основе в клетках CHO, за исключением того, что в антителе, полученном из Pichia pastoris, по природе отсутствует фукоза. Напротив, гликоформы из Fc-мутеинов антитела, произведенных в GFI5.0, имели разнородные гликановые композиции. Одиночные Fc-мутеины антитела, или F243A или V264A, демонстрировали повышенное содержание галактозилированных N-гликанов, которые могут служить субстратом для α2,6 сиалилтрансферазы, как это видно из наиболее высокого пика при MALDI-TOF масс-спектрометрии (фигуры 5 и 6), в то время как N-гликановая композиция для двойного Fc-мутеина антитела, F243A/V264A, демонстрировала еще большее увеличение галактозилированных N-гликанов с более чем 80% от общего числа N-гликанов (фигура 7). Уровень G2 (бигалактозилированного N-гликана), присутствующего на двойном Fc-мутеине антитела, представлял наибольшую долю G2, наблюдаемую из всех антител, которые оценивали авторы заявки. Когда эти же Fc-мутеины антитела экспрессировались в штамме GFI6.0, к G1 (моногалактозилированному) или G2 (бигалактозилированному) N-гликанам добавляли α2,6-связанную сиаловую кислоту. В случае одиночного Fc-мутеина антитела, почти все из G2 N-гликанов превратились в сиалированный гликан (данные не показаны). Хотя антитела, произведенные из одиночных Fc-мутеинов, демонстрировали высокий уровень α2,6-связанного сиалирования (40-51%, см. таблица 3), уровень был меньше, чем у антител, произведенных из Fc-мутеина (фигура 8 и таблица 3), для которого получили 74% сиалирования при помощи ферментации в биореакторе и 91% или больше сиалирования получили при ферментации в малом масштабе.
Не желая быть связанными какой-либо теорией, авторы заявки полагают, что перенос сиаловой кислоты на Fc-олигосахарид усиливается при более открытой конфигурации желоба CH2-домена у двойного Fc-мутеина по сравнению с одиночным Fc-мутеином. Следует также заметить, что α2,6-форма сиаловой кислоты, произведенная штаммом GFI6.0, представляет собой ту же самую форму, что и у людей, и отличается от α2,3-связанной формы сиаловой кислоты, присутствующей на антителах, произведенных в клеточных линиях CHO, Jassal et al., Biochem. Biophys. Res. Comm. 289: 243-249, 2001.
Связывание Fc-мутеинов с FcγR
С использованием анализа на основе ELISA, авторы заявки сравнили связывание с Fc-гамма рецептором (FcγR) для антитела к Her2 из Pichia pastoris, одниночных и двойных Fc-мутеинов антител и антитела к Her2. Как показано в экспериментах, описанных в примерах 11 и 15, двойной Fc-мутеин обладает сниженной аффинностью к FcγRI. Поскольку показано, что рецептор FcγRI стимулирует иммунный ответ после связывания антитела, эти данные позволяют предположить, что двойные мутеины антитела будут менее способны содействовать иммунному ответу.
Что касается FcγRIIb, рецептора, который имеет сниженную аффинность к антителу по сравнению с FcγRI и демонстрирует ингибирование иммунного ответа, с ним двойной Fc-мутеин связывается со сниженной аффинностью. Что касается FcγRIIa, похоже, что с ним двойной Fc-мутеин также связывается со сниженной аффинностью. Эти данные позволяют предположить, что конформационная структура двойных мутеинов антител изменяется таким образом, что способность ингибировать иммунный ответ через FcγRIIa и FcγRIIb/c значительно снижается или пропадает.
FcγRIIIa-F158 и FcγRIIIa-V158 представляют собой полиморфизмы рецептора, о которых известно, что они стимулируют иммунный ответ, но имеют низкую аффинность к антителу по сравнению с FcγRI. Двойной Fc-мутеин имеет незначительную аффинность к FcγRIIIa-F158, хотя все еще сохраняет некоторую аффинность к FcγRIIIa-V158. В совокупности эти данные позволяют предположить, что двойной Fc-мутеин менее склонен к активации и рекрутингу иммунных клеток, таких как макрофаги, моноциты и естественные клетки-киллеры, по сравнению с родительским антителом.
Связывание Fc-мутеинов с C1q
Связывание антитела с C1q является важным параметром для комплемент-зависимой цитотоксичности. Активность связывания (аффинность) молекулы иммуноглобулина (IgG) с C1q можно определять путем клеточного анализа, такого как приведенный в настоящем документе в примере 13. Специалисты в данной области понимают, что описанный анализ может быть легко адаптирован для применения с любой молекулой IgG.
Эффекты ADCC у Fc-мутеинов
Хорошо известно, что рецептор FcγRIIIa (CD16) отвечает за антителозависимую клеточную цитотоксичность (ADCC) (Daeron et al., Annu. Rev. Immunol. 15: 203-234, 1997). Авторы заявки обнаружили, что антитела, произведенные из двойного Fc-мутеина, описываемого в настоящем документе, обладают пониженным связыванием с FcγRIIIa (фигура 9C и D; таблицы 4 и 5) и, соответственно, существует вероятность потери эффекта FcγRIIIa-опосредованной ADCC.
Пример 13 представляет анализ in vitro для измерения элиминации B-клеток и ADCC по флуоресценции высвобождения.
Биодоступность Fc-мутеинов
Биодоступность относится к уровню и скорости, при которых активная субстанция, является ли она лекарственным средством или метаболитом, достигает циркуляции человека и тем самым получает доступ к месту действия. Биодоступность лекарственного средства определяется, в основном, свойствами лекарственной формы, в отличие от физико-химических свойств лекарственного средства, которые, в свою очередь, могут определять его способности к абсорбции. Химическая эквивалентность позволяет предположить, что лекарственные средства содержат одинаковый активный ингредиент(ы) и в одинаковом количестве, хотя другие неактивные компоненты могут различаться. Аналогично, биоэквивалентность позволяет предположить, что два лекарственных средства, при введении одному и тому же пациенту по одной и той же дозовой схеме лечения, будут создавать эквивалентные концентрации лекарственного средства в циркуляции и тканях. С другой стороны, два лекарственных средства, которые не являются идентичными, могут производить одинаковый терапевтический эффект (и неблагоприятные воздействия) при введении одному и тому же пациенту. Таким образом, биодоступность лекарственного средства по отношению к определению является любым эквивалентом, при котором возможно достичь терапевтической эквивалентности, даже когда различаются биодоступности. Для лекарственных средств с узким терапевтическим индексом, т.е. отношением минимальной токсической концентрации к средней эффективной концентрации, различия в биодоступности могут приводить к терапевтической неэквивалентности.
Авторы заявки неожиданно открыли, что двойные Fc-мутеины антител, произведенные в Pichia pastoris посредством материалов и способов по настоящему изобретению, при введении подкожно, имели лучшую биодоступность (абсорбцию или экспозицию), чем сравнимое с ними антитело, произведенное в клетках CHO известными коммерческими способами. В основном, подкожное введение предпочтительнее внутривенного введения из-за того, что подкожный состав пациент может вводить себе сам. Как показано на фигуре 12, сывороточные концентрации у мышей, которых лечили (как изложено в примере 14) двойным Fc-мутеином антитела, повысились приблизительно на 30% по сравнению с сывороточными концентрациями для аналога, произведенного в CHO, и были выше, чем для антитела, произведенного в Pichia pastoris, с отсутствием повышенного содержания α2,6-связанной сиалированной формы.
Выработка гликозилированных антител человека с повышенным уровнем сиалирования
В результате открытий по настоящему изобретению, авторы заявки разработали способ выработки антител, которые имеют повышенное α2,6-связанное сиалирование за счет модификации Fc-области антитела, способ, при котором создается штамм, который может производить in vivo антитела с повышенным галактозилированием и сиалированием по положению Asn297. Не желая быть связанными какой-либо теорией, авторы заявки предполагают, что отсутствие сиаловой кислоты и низкий уровень галактозы, присутствующей на N-гликане в положении Asn297 антитела, обусловлено не низкой эффективностью соответствующей гликозилтрансферазы, а скорее стерическим препятствием, присущим структуре Fc-области. Авторы заявки полагают, что структура Fc-области препятствует добавлению галактозы и сиаловой кислоты в течение короткого периода времени, когда антитело проходит по секреторному пути. В предыдущих работах предполагали, что высокие уровни галактозилирования МАТ к CD20 были возможны in vitro за счет использования более долгого времени инкубации, чем это обычно бывает in vivo, в сочетании с галактозилтрансферазой. Li et al., Nat. Biotechnol. 24(2): 210-215 (2006) считают, что перенос галактозы происходит, когда антитело находится в открытой конфигурации с минимумом стерических препятствий. Структурные или конформационные изменения, происходящие за счет двойной мутации, т.е. двойной Fc-мутеин, приводят к постоянной открытой конформации, позволяя значительно увеличить перенос галактозы и сиаловой кислоты.
Из исследований авторов заявки по настоящему изобретению, очевидно, что индивидуальная мутация по аминокислоте F243 или V264 имеет сходное влияние на повышение галактозилирования и сиалирования Fc-области молекулы IgG1. Lund et al., J. Immunology 157(11): 4963-4969 (1996) описывают умеренное увеличение сиалированного IgG3, произведенного в клеточной линии CHO-1, за счет изменения аминокислот в Fc-области, Phe241, Phe243, Val264, Asp265 или Tyr296, приводящих к уровням 12-42% для моносиалированной и 4-31% для бисиалированной форм. Как сообщалось в настоящем документе, антитела, произведенные посредством материалов и способов по настоящему изобретению, обладали значительным повышением сиалирования, когда конкретные сайты, F243 или V264, были замещены на аланин. Кроме того, были получены уровни более 91% для α2,6-сиалированных видов, если одновременно были мутированы оба сайта (таблица 3).
Биологические мишени
Следует отметить, что хотя в последующих примерах авторы заявки приводят примеры материалов и способов по изобретению с использованием антител IgG1 с последовательностями, аналогичными последовательностям для коммерчески доступных антител к Her2 и к TNF, изобретение не ограничено раскрытыми антителами. Специалисты в данной области поймут, что материалы и способы в настоящем документе можно использовать для производства любого Fc-содержащего полипептида, для которого желательны характеристики усиленной противовоспалительной активности или сниженной эффекторной функции. Дополнительно следует отметить, что не существует ограничений относительно типа Fc-содержащего полипептида или антитела, производимого таким образом по изобретению. Fc-область Fc-содержащего полипептида может быть из IgA, IgD, IgE, IgG или IgM. В одном из вариантов осуществления Fc-область Fc-содержащего полипептида происходит из IgG, включая IgG1, IgG2, IgG3 или IgG4. В одном из вариантов осуществления Fc-область Fc-содержащего полипептида происходит из IgG1. В конкретных вариантах осуществления антитела или фрагменты антител, произведенные посредством материалов и способов, описанных в настоящем документе, могут быть гуманизированными, химерными или антителами человека.
В некоторых вариантах осуществления Fc-содержащие полипептиды по изобретению связываются с биологической мишенью, вовлеченной в воспаление.
В некоторых вариантах осуществления Fc-содержащий полипептид по изобретению связывается с провоспалительным цитокином. В некоторых вариантах осуществления Fc-содержащий полипептид по изобретению связывается с молекулой, выбранной из группы, включающей: TNF-a, IL-1, IL-2, IL-4, IL-5, IL-6, IL-8, IL-9, IL-10, IL-12, IL-15, IL-17, IL-18, IL-20, IL-21, IL-22, IL-23, IL-23R, IL-25, IL-27, IL-33, CD2, CD4, CD11A, CD14, CD18, CD19, CD23, CD25, CD40, CD40L, CD20, CD52, CD64, CD80, CD147, CD200, CD200R, TSLP, TSLPR, PD-1, PDL1, CTLA4, VLA-4, VEGF, PCSK9, α4β7-интегрин, E-селектин, фактор II, ICAM-3, бета2-интегрин, IFNγ, C5, CBL, LCAT, CR3, MDL-1, GITR, ADDL, CGRP, TRKA, IGF1R, RANKL, GTC, αBLys или рецептор для любой из вышеперечисленных молекул. В одном из вариантов осуществления Fc-содержащий полипептид связывается с TNF-α. В другом варианте осуществления Fc-содержащий полипептид связывается с Her2. В другом варианте осуществления Fc-содержащий полипептид связывается с PCSK9. В другом варианте осуществления Fc-содержащий полипептид по изобретению связывается с TNFR. В другом варианте осуществления Fc-содержащий полипептид по изобретению связывается с LCAT. В другом варианте осуществления Fc-содержащий полипептид по изобретению связывается с TSLP. В другом варианте осуществления Fc-содержащий полипептид по изобретению связывается с PD-1. В другом варианте осуществления Fc-содержащий полипептид по изобретению связывается с IL-23.
В некоторых вариантах осуществления Fc-содержащие полипептиды по изобретению являются специфичными для антигена, выбранного из аутоиммунных антигенов, аллергенов, молекул MHC или антигена резус-фактора D. См., например, антигены, перечисленные в таблице 1 в WO2010/10910, включенном в настоящий документ в качестве ссылки.
Способы повышения противовоспалительных свойств или снижения эффекторной функции/цитотоксичности
Изобретение также относится к способу повышения противовоспалительных свойств Fc-содержащего полипептида, включающему: выбор родительского Fc-содержащего полипептида, который подходит для лечения воспаления (например, антитела или иммуноадгезина, которые связываются с антигеном, вовлеченным в воспаление) и введение мутаций в положениях 243 или 264 в Fc-содержащий полипептид, где нумерация приведена согласно индексу EU по Kabat, где Fc-содержащий полипептид имеет повышенные противовоспалительные свойства по сравнению с родительским Fc-содержащим полипептидом. В одном варианте осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243Y и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243T и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264N. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243V и V264G. В одном из вариантов осуществления родительский Fc-содержащий полипептид представляет собой антитело, фрагмент антитела или иммуноадгезин, который связывается с антигеном, вовлеченным в воспаление. В одном из вариантов осуществления родительский Fc-содержащий полипептид представляет собой антитело, фрагмент антитела или иммуноадгезин, который уже продается или разрабатывается для лечения воспалительных заболеваний. В другом варианте осуществления родительский Fc-содержащий полипептид представляет собой антитело, выбранное из группы, включающей: Мурономаб-CD3 (антитело против рецептора к CD3), Абциксимаб (анти-CD41 7E3 антитело), Ритуксимаб (антитело к CD20), Даклизумаб (антитело к CD25), Базиликсимаб (антитело к CD25), Паливизумаб (антитело против RSV (респираторно-синцитиального вируса)), Инфликсимаб (антитело к TNFα), Трастузумаб (антитело к Her2), Гемтузумаб озогамицин (антитело к CD33), Алемтузумаб (антитело к CD52), Ибритумомаб тиуксетен (антитело к CD20), Адалимумаб (антитело к TNFα), Омализумаб (антитело к IgE), Тозитумомаб-131I (йодированное производное антитела к CD20), Эфализумаб (антитело против CD11a), Цетуксимаб (антитело против рецептора к EGF), Голимумаб (антитело к TNFα), Бевацизумаб (антитело против VEGF-A), Натализумаб (против α4-интегрина), Эфализумаб (против CD11a), Цетолизумаб (антитело к TNFα), Тоцилизумаб (против IL-6R), Устенкинумаб (против IL-12/23), алемтузумаб (против CD52) и натализумаб (против интегрина α4), и их варианты. В другом варианте осуществления родительский Fc-содержащий полипептид представляет собой Fc-слитый белок, выбранный из группы, состоящей из: Аркалист/рилонацепт (слияние IL1R-Fc), Оренция/абатацепт (слияние CTLA-4-Fc), Амевив/алефацепт (слияние LFA-3-Fc), слияние Анакинра-Fc (IL-1Ra-Fc-слитый белок), этанерцепта (TNFR-Fc-слитый белок), FGF-21-Fc-слитого белка, GLP-1-Fc-слитого белка, RAGE-Fc-слитого белка, ActRIIA-Fc-слитого белка, ActRIIB-Fc-слитого белка, глюкагон-Fc-слитого белка, оксинтомодулин-Fc-слитого белка, GM-CSF-Fc-слитого белка, EPO-Fc-слитого белка, Инсулин-Fc-слитого белка, проинсулин-Fc-слитого белка и предшественник инсулина-Fc-слитого белка, и их аналогов и вариантов.
Изобретение также относится к способу уменьшения эффекторной функции Fc-содержащего полипептида, включающему введение мутаций в положениях 243 и 264 родительского Fc-содержащего полипептида, где указанный Fc-содержащий полипептид обладает уменьшенной эффекторной функцией по сравнению с родительским Fc-содержащим полипептидом, где нумерация произведена согласно индексу EU по Kabat. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В одном из вариантов осуществления Fc-содержащий полипептид представляет собой антитело или его антигенсвязывающий фрагмент. В одном из вариантов осуществления эффекторная функция представляет собой ADCC. В другом варианте осуществления эффекторная функция представляет собой CDC.
Изобретение также относится к способу снижения цитотоксичности Fc-содержащего полипептида, включающему: выбор родительского Fc-содержащего полипептида, подходящего для лечения воспалительных заболеваний (например, антитело или иммуноадгезин, которые связываются с антигеном, вовлеченным в воспаление), который связывается с антигеном, вовлеченным в воспаление, и введение мутаций в положениях 243 и 264 Fc-содержащего полипептида, где нумерация произведена согласно индексу EU по Kabat, где Fc-содержащий полипептид обладает сниженной цитотоксичностью по сравнению с родительским Fc-содержащим полипептидом.
В одном из вариантов осуществления родительский Fc-содержащий полипептид содержит нативную Fc-область. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию F243A. В другом варианте осуществления родительский Fc-содержащий полипептид содержит мутацию V264A.
Способы лечения
Изобретение также относится к способу лечения воспалительного заболевания у нуждающегося в этом индивидуума, включающему: введение индивидууму терапевтически эффективного количества Fc-содержащего полипептида, включающего мутации в положениях 243 и 264, где нумерация произведена согласно индексу EU по Kabat. В одном из вариантов осуществления Fc-содержащий полипептид содержит мутации F243A и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243Y и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243T и V264G. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264A. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243L и V264N. В другом варианте осуществления Fc-содержащий полипептид содержит мутации F243V и V264G. Fc-содержащий полипептид по изобретению можно вводить любым путем. В одном из вариантов осуществления Fc-содержащий полипептид вводят парентерально. В одном из вариантов осуществления Fc-содержащий полипептид вводят подкожно.
В одном из вариантов осуществления воспалительное заболевание представляет собой нежелательные воспалительные иммунные реакции.
В одном из вариантов осуществления воспалительное заболевание представляет собой аутоиммунное заболевание. В одном из вариантов осуществления воспалительное заболевание представляет собой рассеянный склероз. В одном из вариантов осуществления воспалительное заболевание представляет собой системную красную волчанку. В одном из вариантов осуществления воспалительное заболевание представляет собой диабет I типа.
В одном из вариантов осуществления воспалительное заболевание представляет собой синдром первичного иммунодефицита, в том числе врожденную агаммаглобулинемию и гипогаммаглобулинемию, вариабельный неклассифицируемый иммунодефицит, тяжелый комбинированный иммунодефицит или синдром Вискотта-Олдрича.
В одном из вариантов осуществления воспалительное заболевание представляет собой синдром вторичного иммунодефицита, в том числе B-клеточный лимфоцитарный лейкоз, инфекцию ВИЧ или трансплантацию аллогенного костного мозга.
В одном из вариантов осуществления воспалительное заболевание представляет собой идиопатическую тромбоцитопеническую пурпуру.
В одном из вариантов осуществления воспалительное заболевание представляет собой множественную миелому.
В одном из вариантов осуществления воспалительное заболевание представляет собой синдром Гийена-Барре.
В одном из вариантов осуществления воспалительное заболевание представляет собой заболевание Кавасаки.
В одном из вариантов осуществления воспалительное заболевание представляет собой хроническую воспалительную демиелинизирующую полинейропатию (CIDP).
В одном из вариантов осуществления воспалительное заболевание представляет собой аутоиммунную нейтропению.
В одном из вариантов осуществления воспалительное заболевание представляет собой гемолитическую анемию.
В одном из вариантов осуществления воспалительное заболевание представляет собой аутоиммунное заболевание, связанное с наличием антител против фактора VIII.
В одном из вариантов осуществления воспалительное заболевание представляет собой мультиочаговую нейропатию.
В одном из вариантов осуществления воспалительное заболевание представляет собой системный васкулит (позитивный по антителам к цитоплазме нейтрофилов - ANCA).
В одном из вариантов осуществления воспалительное заболевание представляет собой полимиозит.
В одном из вариантов осуществления воспалительное заболевание представляет собой дерматомиозит.
В одном из вариантов осуществления воспалительное заболевание представляет собой антифосфолипидный синдром.
В одном из вариантов осуществления воспалительное заболевание представляет собой септический синдром.
В одном из вариантов осуществления воспалительное заболевание представляет собой заболевание трансплантат против хозяина.
В одном из вариантов осуществления воспалительное заболевание представляет собой аллергию.
В одном из вариантов осуществления воспалительное заболевание представляет собой реакцию против резус-фактора D.
В одном из вариантов осуществления воспалительное заболевание представляет собой воспалительное заболевание сердечно-сосудистой системы. Fc-содержащие полипептиды по изобретению можно использовать для лечения атеросклероза, атеротромбоза, гипертензии коронарных артерий, острого коронарного синдрома и сердечной недостаточности, все эти заболевания ассоциированы с воспалением.
В одном из вариантов осуществления воспалительное заболевание представляет собой воспалительное заболевание центральной нервной системы. В другом варианте осуществления воспалительное заболевание представляет собой воспалительное заболевание периферической нервной системы. Например, Fc-содержащие полипептиды по изобретению можно использовать для лечения, например, болезни Альцгеймера, бокового амиотрофического склероза (также известного как ALS; заболевание Лу Герига), ишемического повреждения головного мозга, прионных болезней и ВИЧ-ассоциированной деменции.
В одном из вариантов осуществления воспалительное заболевание представляет собой воспалительное заболевание желудочно-кишечного тракта. Например, Fc-содержащие полипептиды по изобретению можно использовать для лечения воспалительных заболеваний кишечника, например, болезни Крона, язвенного колита, глютеновой болезни и синдрома раздраженного кишечника.
В одном из вариантов осуществления воспалительное заболевание представляет собой псориаз, атопический дерматит, артрит, в том числе ревматоидный артрит, остеоартрит и псориатический артрит.
В одном из вариантов осуществления воспалительное заболевание представляет собой стероид-зависимый атопический дерматит.
В одном из вариантов осуществления воспалительное заболевание представляет собой кахексию.
Примеры других воспалительных нарушений, которые можно лечить при помощи Fc-содержащих полипептидов по изобретению, также включают обыкновенные угри, астму, аутоиммунные заболевания, хронический простатит, гломерулонефрит, гиперчувствительности, воспалительные заболевания таза, реперфузионное повреждение, саркоидоз, отторжение трансплантата, васкулит, интерстициальный цистит и миопатии.
В одном из вариантов осуществления Fc-содержащий полипептид по изобретению вводят в дозе от 1 до 100 миллиграмм на килограмм массы тела. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению вводят в дозе от 0,001 до 10 миллиграмм на килограмм массы тела. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению вводят в дозе от 0,001 до 0,1 миллиграмма на килограмм массы тела. В одном из вариантов осуществления Fc-содержащий полипептид по изобретению вводят в дозе от 0,001 до 0,01 миллиграмма на килограмм массы тела.
Фармацевтические составы
Изобретение также относится к фармацевтическим составам, включающим Fc-содержащий полипептид по изобретению и фармацевтически приемлемый носитель.
В одном из вариантов осуществления изобретение относится к фармацевтической композиции, включающей Fc-содержащий полипептид, где, по меньшей мере, 70% N-гликанов на Fc-содержащем полипептиде содержат олигосахаридную структуру, выбранную из группы, состоящей из NANA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2), где Fc-содержащий полипептид содержит мутации в аминокислотных положениях 243 и 264 в Fc-области, где нумерация приведена согласно индексу EU по Kabat. В одном из вариантов осуществления мутации представляют собой F243A и V264A. В одном из вариантов осуществления, по меньшей мере, 47 моль% N-гликанов имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2. В одном из вариантов осуществления остатки сиаловой кислоты в сиалированных N-гликанах присоединены посредством α-2,6-связи. В одном из вариантов осуществления остатки сиаловой кислоты в сиалированных N-гликанах присоединены посредством α-2,6-связи и не содержат детектируемого количества α-2,3-связанной сиаловой кислоты. В одном из вариантов осуществления сиалированные N-гликаны не содержат N-гликолилнейраминовую кислоту (NGNA).
Применяемый в настоящем документе термин "фармацевтически приемлемый" означает нетоксический материал, который не влияет на эффективность биологической активности активного ингредиента(ов), одобрен федеральным регуляторным органом или регуляторным органом правительства штата, или указан в фармакопее США или в другой общепризнанной фармакопее для применения у животных и, более конкретно, у людей. Термин "носитель" относится к разбавителю, адъюванту, эксципиенту или носителю, с которым вводят терапевтическое средство, и в качестве неограничивающих примеров включает такие стерильные жидкости, как вода и масла. Характеристики носителя будут зависеть от пути введения.
Фармацевтические составы терапевтических и диагностических средств можно получать путем смешивания с приемлемыми носителями, эксципиентами или стабилизаторами в форме, например, лиофилизированных порошков, водных растворов или суспензий (см., например,

Способы введения могут быть различными. Подходящие пути введения включают пероральный, ректальный, трансмукозальный, кишечный, парентеральный, внутримышечный, подкожный, интрадермальный, интрамедуллярный, интратекальный, прямой внутрижелудочковый, внутривенный, интраперитонеальный, интраназальный, внутриглазной, ингаляцию, вдувание, местный, кожный, трансдермальный или внутриартериальный.
В определенных вариантах осуществления Fc-содержащие полипептиды по изобретению можно вводить инвазивным путем, таким как инъекция (см. выше). В некоторых вариантах осуществления изобретения Fc-содержащие полипептиды по изобретению, или их фармацевтическую композицию, вводят внутривенно, подкожно, внутримышечно, внутриартериально, внутрисуставно (например, в суставы с артритом), внутрь опухоли, или путем ингаляции, с помощью аэрозоля. Введение неинвазивными путями (например, перорально; например, в пилюле, капсуле или таблетке) также входит в объем настоящего изобретения.
В определенных вариантах осуществления Fc-содержащие полипептиды по изобретению можно вводить инвазивным путем, таким как инъекция (см. выше). В некоторых вариантах осуществления изобретения Fc-содержащие полипептиды по изобретению, или их фармацевтическую композицию, вводят внутривенно, подкожно, внутримышечно, внутриартериально, внутрисуставно (например, в суставы с артритом), внутрь опухоли, или путем ингаляции, с помощью аэрозоля.
Введение неинвазивными путями (например, перорально; например, в пилюле, капсуле или таблетке) также входит в объем настоящего изобретения.
Композиции можно вводить при помощи медицинских устройств, известных в данной области, Например, фармацевтическую композицию по изобретению можно вводить путем инъекции при помощи иглы для подкожного введения, в том числе, например, при помощи заранее заполненного шприца или автоинжектора.
Фармацевтические композиции по изобретению также можно вводить при помощи устройства для безыгольной подкожной инъекции, такого как устройства, раскрытые в патентах США №№ 6620135; 6096002; 5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556.
Фармацевтические композиции по изобретению также можно вводить путем инфузии. Примеры хорошо известных имплантатов и модулей, выполняющих введение фармацевтических композиций, включают: патент США № 4487603, который описывает имплантируемый инфузионный насос для дозирования лекарственного препарата с контролируемой скоростью; патент США № 4447233, который описывает инфузионный насос для лекарственного средства для доставки лекарственного средства с точной скоростью введения; патент США № 4447224, который описывает имплантируемый инфузионный аппарат с переменной скоростью введения для длительного введения лекарственного средства; патент США № 4439196, который описывает осмотическую систему доставки лекарственного средства с многокамерными отделениями. Множество других таких имплантататов, систем доставки и модулей хорошо известны специалистам в данной области.
С другой стороны, можно вводить антитело местно, а не системно, например, посредством инъекции антитела непосредственно в артритный сустав, часто в виде составов замедленного всасывания или пролонгированного высвобождения. Кроме того, можно вводить антитело в нацеленной системе доставки лекарственного средства, например, в липосоме, покрытой тканеспецифическими антителами, нацеливая, например, на артритный сустав или индуцированный патогеном очаг, который характеризуется иммунопатологией. Липосомы будут нацелены на поврежденную ткань и избирательно захвачены поврежденной тканью.
Режим введения зависит от нескольких факторов, включая скорость циркуляции терапевтического антитела в сыворотке или ткани, степени симптомов, иммуногенности терапевтического антитела и доступности клеток-мишеней в биологическом матриксе. Предпочтительно, режим введения обеспечивает доставку достаточного количества терапевтического антитела для того, чтобы улучшить состояние целевой болезни, наряду с тем одновременно минимизируя нежелательные побочные эффекты. Таким образом, количество доставляемого биопрепарата частично зависит от конкретного терапевтического антитела и тяжести состояния, которое лечат. Доступно руководство по выбору соответствующих доз терапевтических антител (см., например,

Определение соответствующей дозы производит врач, например, с учетом параметров или факторов, о которых известно, что они влияют или потенциально влияют на лечение. В основном, доза начинается с количества несколько меньшего, чем оптимальная доза, и в дальнейшем повышается небольшими шагами до тех пор, пока не будет достигнут желаемый или оптимальный эффект по отношению к любым негативным побочным эффектам. Важные диагностические параметры включают те из симптомов, которые вызывают, например, воспаление или уровень воспалительных цитокинов. Предпочтительно, биопрепарат, который будет использован, получают из тех же видов, к которым относится животное, предназначенное для лечения, тем самым минимизируя любой иммунный ответ на реагент. В случае пациентов-людей предпочтительными являются, например, химерные, гуманизированные и полностью человеческие Fc-содержащие полипептиды.
Fc-содержащие полипептиды могут быть обеспечены путем постоянного вливания или посредством доз, вводимых, например, ежедневно, 1-7 раз в неделю, раз в неделю, раз в две недели, раз в месяц, раз в два месяца, ежеквартально, раз в полугодие, ежегодно и т.д. Дозы могут быть обеспечены, например, внутривенно, подкожно, местно, перорально, назально, ректально, внутримышечно, интрацеребрально, интраспинально или путем ингаляции. Общая недельная доза составляет в основном, по меньшей мере, 0,05 мкг/кг массы тела, в более общем смысле, по меньшей мере, 0,2 мкг/кг, 0,5 мкг/кг, 1 мкг/кг, 10 мкг/кг, 100 мкг/кг, 0,25 мг/кг, 1,0 мг/кг, 2,0 мг/кг, 5 мг/мл, 10 мг/кг, 25 мг/кг, 50 мг/кг или больше (см., например, 
В других вариантах осуществления Fc-содержащий полипептид по настоящему изобретению вводят подкожно или внутривенно, раз в неделю, раз в две недели, "каждые 4 недели", раз в месяц, раз в два месяца или ежеквартально в дозе 10, 20, 50, 80, 100, 200, 500, 1000 или 2500 мг/на субъекта.
Применяемые в настоящем документе термины "терапевтически эффективное количество", "терапевтически эффективная доза" и "эффективное количество" относятся к количеству Fc-содержащего полипептида по изобретению, которое, при введении отдельно или в комбинации с дополнительным терапевтическим средством в клетку, ткань или субъекту, является эффективным для того, чтобы вызвать измеримое улучшение одного или нескольких симптомов заболевания или состояния или прогрессирования такого заболевания или состояния. «Терапевтически эффективная доза» дополнительно относится к такому количеству Fc-содержащего полипептида, которого достаточно для достижения, по меньшей мере, частичного улучшения симптомов, например, лечение, профилактика или улучшение соответствующего медицинского состояния, или повышения скорости лечения, исцеление, профилактика или ослабление таких состояний. При применении отдельного активного ингредиента, который вводят сам по себе, терапевтически эффективная доза относится отдельно к этому ингредиенту. При применении комбинации терапевтически эффективная доза относится к суммарному количеству активных ингредиентов, которые приводят к терапевтическому эффекту при введении в составе комбинации, последовательно или одновременно. Эффективное количество терапевтического средства приводит к улучшению диагностического показателя или параметра, по меньшей мере, на 10%; как правило, по меньшей мере, на 20%; предпочтительно, по меньшей мере, приблизительно на 30%; более предпочтительно, по меньшей мере, на 40%, и наиболее предпочтительно, по меньшей мере, на 50%. Эффективное количество может также приводить к улучшению субъективного показателя в случаях, когда для оценки тяжести заболевания применяют субъективные критерии.
ПРИМЕР 1
Штаммы и реагенты
Для работы с рекомбинантной ДНК использовали штаммы Escherichia coli TOP10 или DH5α (Invitrogen, CA). Рестрикционные эндонуклеазы, ферменты для модификации ДНК и ПНгазу F получали от New England Biolabs, Ipswich, MA. Олигонуклеотиды заказывали у Integrated DNA Technologies, Coralville, IA.
ПРИМЕР 2
Создание Fc-мутеинов IgG1 против Her2 и рекомбинантного экспрессирующего вектора Pichia pastoris
Получение одиночных и двойных Fc-мутеинов моноклонального антитела IgG1 к Her2 в Pichia pastoris проводили с использованием последовательностей и протоколов, перечисленных ниже.
A. Тяжелые и легкие цепи
Последовательности тяжелой и легкой цепей, SEQ ID NO:1 и 2, соответственно, использованные для получения моноклонального антитела IgG1 к Her2, являются такими, как изложено ниже. Аминокислотная последовательность тяжелой цепи двойного мутеина антитела против Her2 показана в SEQ ID NO:9. Тяжелые и легкие цепи подвергали оптимизации кодонов согласно частоте использования кодонов у Pichia pastoris, синтезировали у GeneArt AG (Josef-Engert-Str. 11, D-93053 Regensburg, Germany) и клонировали в pUC19.
Аланиновые мутации в Fc в аминокислотном положении F243 осуществляли при помощи набора для сайт-специфического мутагенеза QuikChange® (Strategene, CA) с использованием прямого и обратного праймеров, FcF243A-F (SEQ ID NO:3) и FcF243A-R (SEQ ID NO:4), соответственно. Аналогично, мутации в аминокислотном положении V264 осуществляли с использованием прямого и обратного праймеров, V254A-F (SEQ ID NO:5) и V264A-R (SEQ ID NO:6), соответственно. Двойную мутацию осуществляли с применением ферментативного расщепления между сайтами F243A и V264A в каждой из плазмид с одиночным мутеином с последующим лигированием.
B. Сигнальная последовательность
Сигнальную последовательность предомена α-спаривающего фактора (MF) сливали в рамке считывания с 5'-концом легкой или тяжелой цепи посредством сливающей ПЦР. Последовательность подвергали оптимизации кодонов, как описано выше. Последовательность Козака AAACG добавляли к 5'-концу метионина и перед последовательностью Козака добавляли сайт распознавания EcoR1 для клонирования. Последовательность ДНК (SEQ ID NO:7) и трансляция аминокислот (SEQ ID NO:8) показаны ниже.
C. Рекомбинантные плазмиды для экспрессии IgG1 и Fc-мутеинов IgGl
Тяжелые и легкие цепи IgG1 и его мутеинов, слитые с сигнальной последовательностью, клонировали под контролем промотора Pichia pastoris, AOX1, и перед терминатором Cyc S. cerevisiae, соответственно. Экспрессирующие кассеты с готовыми тяжелыми и легкими цепями помещали вместе в конечный экспрессирующий вектор. Геномную вставку в Pichia pastoris обеспечивали за счет линеаризации вектора при помощи Spe1 и нацеленной интеграции в сайт Trp2.
Краткие сведения о плазмидах, использованных в настоящем изобретении, представлены ниже в таблице 1. Графическое изображение конечной экспрессионной плазмиды для двойного Fc-мутеина антитела к Her2 показано на фигуре 1.
ПРИМЕР 3
Гликоинжиниринговые хозяева Pichia GFI5.0 и GFI6.0 для выработки антитела против Her2 и его Fc-мутеинов
Два различных гликоинжиниринговых хозяина Pichia были использованы в настоящем изобретении, GFI5.0 и GFI6.0. В соответствии с процедурами, описанными у Gerngross, US 7029872 и Gerngross, US 7449308, можно сконструировать векторы, которые пригодны для генетической инженерии клеток-хозяев низших эукариот, таких, которые способны к экспрессии желаемого полипептида с желаемой N-гликоформой в качестве преобладающего соединения. GFI5.0 и GFI6.0 штаммы были сконструированы из NRRL11430 (American Type Culture Collection (ATCC), P.O. Box 1549, Manassas, VA 20108, USA) согласно способам, описанным в Hamilton et al., Science, 313: 1441-1443 (2006) и Hamilton US 2006/0286637. Сконструированный штамм Pichia pastoris GFI5.0 способен вырабатывать белки с биантенной N-гликановой структурой с концевой галактозой. Генотип штамма GFI5.0, использованного в настоящем изобретении, RDP697, выглядит следующим образом: ura5∆::ScSUC2 ochl∆::lacZ bmt2∆::lacZ/KlMNN2-2 mnn4Ll∆::lacZ/MmSLC35A3 pnol∆::lacZ ADE1::lacZ/FB8/NA10/MmSLC35A3 his1::lacZ-URA5-lacZ/XB33/SpGALE/DmUGT arg1::HIS1/KD53/TC54. Генотип сконструированного штамма Pichia pastoris GFI6.0, YGLY3582, выглядит следующим образом: ura5∆::ScSUC2 ochl∆::lacZ bmt2∆::lacZ/KlMNN2-2 mnn4Ll∆::lacZ/MmSLC35A3 pnol∆mnn4∆::lacZ met16∆::lacZ his1∆::lacZ/ScGAL10/XB33/DmUGT arg1∆::HIS1/KD53/TC54 ADE1::lacZ/NA10/MmSLC35A3/FB8 PRO1::lacZ-URA5-lacZ/TrMDS1 TRP2:ARG1/MmCST/HsGNE/HsCSS/HsSPS/MmST6-33Y. Штамм GFI6.0 способен вырабатывать белки с биантенной N-гликановой структурой, в которой концевая 2,6-связанная сиаловая кислота присоединена к галактозе.
Сокращения, применяемые для описания генотипов, являются общеизвестными, понятны специалистам в данной области и включают следующие сокращения:
ScSUC2 инвертаза S. cerevisiae
OCH1 Альфа-1,6-маннозилтрансфераза
K1MN2-2 переносчик UDP-GlcNAc K. lactis
BMT1 перенос бета-маннозы (удаление бета-маннозы)
BMT2 перенос бета-маннозы (удаление бета-маннозы)
BMT3 перенос бета-маннозы (удаление бета-маннозы)
BMT4 перенос бета-маннозы (удаление бета-маннозы)
MNN4L1 MNN4-подобный 1 (удаление заряда)
MmSLC35A3 Мышиный гомолог переносчика UDP-GlcNAc
PNO1 фосфоманнозилирование N-гликанов (удаление заряда)
MNN4 маннозилтрансфераза (удаление заряда)
ScGAL10 UDP-глюкоза4-эпимераза
XB33 Усеченный HsGalT1, слитый с лидерной последовательностью ScKRE2
DmUGT переносчик UDP-галактозы
KD53 Усеченный DmMNSII, слитый с лидерной последовательностью ScMNN2
TC54 Усеченный RnGNTII, слитый с лидерной последовательностью ScMNN2
NA10 Усеченный HsGNTI, слитый с лидерной последовательностью PpSEC12
FB8 Усеченный MmMNS1A, слитый с лидерной последовательностью ScSEC12
TrMDS1 Секретируемый MNS1 T. reseei
ADE1 N-сукцинил-5-аминоимидазол-4-карбоксамид риботид (SAICAR) синтетаза
MmCST Мышиный переносчик CMP-сиаловой кислоты
HsGNE Человеческая UDP-GlcNAc 2-эпимераза/N-ацетилманнозаминкиназа
HsCSS Человеческая синтетаза CMP-сиаловой кислоты
HsSPS Человеческая N-ацетилнейраминат-9-фосфатсинтетаза
MmST6-33 Усеченная мышиная альфа-2,6-сиалилтрансфераза, слитая с лидерной последовательностью ScKRE2
LmSTT3d Каталитическая субъединица олигосахарилтрансферазы из Leishmania major
ПРИМЕР 4
Трансформация и скрининг дрожжей
Гликоинжиниринговые штаммы GFI5.0 и GS6.0 выращивали в обогащенной среде YPD (дрожжевой экстракт 1%, пептон 2% и 2% декстрозы), собирали в логарифмическую фазу путем центрифугирования и промывали три раза ледяным 1 M сорбитом. От 1 до 5 мкг плазмиды, расщепленной Spe1, смешивали с компетентными клетками дрожжей и проводили электропорацию при помощи Bio-Rad Gene Pulser Xcell™ (Bio-Rad, 2000 Alfred Nobel Drive, Hercules, CA 94547) со стандартной программой электропорации Pichia pastoris. Через час после восстановления в обогащенной среде при 24°C клетки высевали на планшет с минимальной средой с декстрозой (1,34% YNB, 0,0004% биотина, 2% декстрозы, 1,5% агара), содержащей 300 мкг/мл Зеоцина, и инкубировали при 24°C до появления трансформантов.
Для скрининга на штаммы с высоким титром, 96 трансформантов засевали в забуференную глицериновую комплексную среду (BMGY) и выращивали в течение 72 часов с последующей индукцией в забуференной комплексной среде с метанолом (BMMY) в течение 24 часов. Секрецию антитела оценивали путем анализа с белком А на частицах, как описано далее. 50 мкл супернатанта из культур из 96-луночного планшета разбавляли 1:1 50 мМ Трис pH 8,5 в 96-луночном планшете для несвязывающего анализа. Для каждого 96-луночного планшета, помещали 2 мл суспензии магнитных частиц BioMag с белком А (Qiagen, Valencia, CA) в пробирку, закрепленную на магнитном держателе. Через 2-3 минуты, когда частицы собирались по сторонам пробирки, удаляли буфер. Частицы отмывали три раза отмывочным буфером в объеме, равном начальному объему (100 мМ Трис, 150 мМ NaCl, pH 7,0), и ресуспендировали в таком же отмывочном буфере. 20 мкл частиц добавляли к каждой лунке планшета для анализа, содержащего разведенные образцы. Планшет накрывали, осторожно встряхивали, а затем инкубировали при комнатной температуре в течение 1 часа, встряхивая каждые 15 минут. После инкубации планшет с образцами помещали на магнитную подложку, заставляя частицы собираться с одной стороны каждой лунки. Удаляли супернатант из планшета в контейнер для отходов на Biomek NX Liquid Handler (Beckman Coulter, Fullerton, CA). Планшет с образцами снимали с магнита и отмывали частицы 100 мкл отмывочного буфера. Планшет снова помещали на магнит, перед тем как удалить буфер аспирацией. 20 мкл буфера для нанесения (Invitrogen, буфер для нанесения на гель E-PAGE, содержащий 25 мМ NEM (Pierce, Rockford, IL)) добавляли к каждой лунке, и планшет встряхивали. После центрифугирования при 500 об/мин на центрифуге Beckman Allegra 6, образцы инкубировали при 99°C в течение пять минут, а затем проводили форез на готовом высокопроизводительном геле E-PAGE (Invitrogen, Carlsbad, CA). Гели заливали раствором для окрашивания гелей (0,5 г Кумасси G250 Brilliant Blue, 40% MeOH, 7,5% уксусной кислоты), нагревали в микроволновой печи в течение 35 секунд, а затем инкубировали при комнатной температуре в течение 30 минут. Гели отмывали от красителя в дистиллированной воде в течение ночи. Колонии с высоким титром выбирали для дальнейшего ферментационного скрининга в Sixfors, подробно описанного в Примере 5. Краткое описание штаммов-продуцентов IgG1 дикого типа (родительского) и Fc-мутеинов приведено ниже в таблице 2.
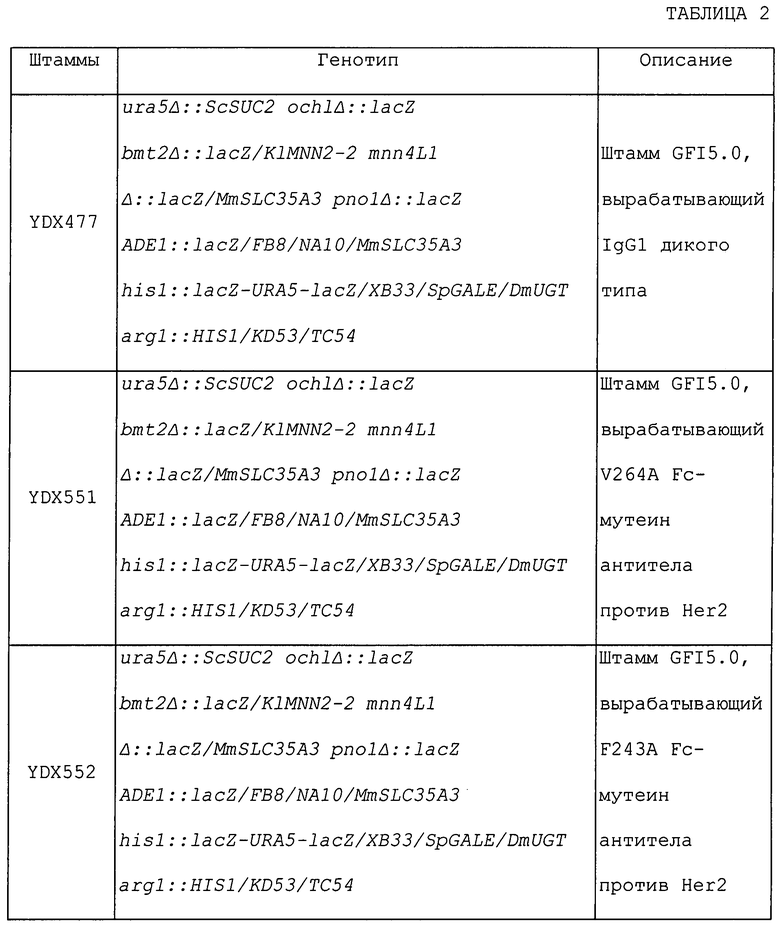


ПРИМЕР 5
Скрининг в биореакторе(Sixfors)
Ферментационный скрининг в биореакторе проводили, как описано далее. Периодическую ферментацию с добавлением субстрата гликоинжиниринговых Pichia pastoris проводили в 0,5-литровых биореакторах (мультиферментационная система Sixfors, ATR Biotech, Laurel, MD) при следующих условиях: pH 6,5, 24°C, 300 мл воздушного потока/мин, и начальная скорость мешалки 550 об/мин с начальным рабочим объемом 350 мл (330 мл среды BMGY [100 мМ фосфата калия, 10 г/л дрожжевого экстракта, 20 г/л пептона (BD, Franklin Lakes, NJ), 40 г/л глицерина, 18,2 г/л сорбита, 13,4 г/л YNB (BD, Franklin Lakes, NJ), 4 мг/л биотина] и 20 мл культуры). Использовали программное обеспечение IRIS для мультиферментеров (ATR Biotech, Laurel, MD) для увеличения скорости мешалки от 550 об/мин до 1200 об/мин линейно в интервале между 1 и 10 часами ферментации. Таким образом, концентрация растворенного кислорода могла колебаться в течение ферментации. Ферментацию проводили порционным способом до тех пор, пока не будет потреблено изначально внесенное количество глицерина (40 г/л) (как правило, 18-24 часа). Вторую фазу добавления субстрата начинали с добавления в биореактор 17 мл питающего глицеринового раствора (50% [масс./масс.] глицерина, 5 мг/л биотина и 12,5 мл/л PTM1 солей (65 г/л FeSO4×7H2O, 20 г/л ZnCl2, 9 г/л H2SO4, 6 г/л CuSO4×5H2O, 5 г/л H2SO4, 3 г/л MnSO4×7H2O, 500 мг/л CoCl2×6H2O, 200 мг/л NaMoO4×2H2O, 200 мг/л биотина, 80 мг/л NaI, 20 мг/л H3BO4). Снова проводили ферментацию порционным способом до потребления добавленного глицерина (как правило, 6-8 часов). Фазу индукции вызывали добавлением питающего раствора с метанолом (100% [масс./масс.] метанола, 5 мг/л биотина и 12,5 мл/л PTM1 солей) в количестве 0,6 г/час, как правило, в течение 36 часов перед сбором клеток. Весь объем содержимого удаляли из реактора и центрифугировали в центрифуге Sorvall Evolution RC, оснащенной ротором SLC-6000 (Thermo Scientific, Milford, MA), в течение 30 минут при 8500 об/мин Удаляли клеточную массу и сохраняли супернатант для очистки и анализа. Качественный состав гликанов оценивали посредством MALDI-время-пролетной (TOF) спектрометрии и мечения 2-аминобензидином (2-AB) согласно Li et al., Nat. Biotech. 24(2): 210-215 (2006). Гликаны высвобождали из антитела путем обработки PNGазой-F и анализировали MALDI-TOF для подтверждения структуры гликанов. Для оценки относительного количества присутствующих нейтральных и заряженных гликанов, гликаны, высвобожденные N-гликозидазой F, метили 2-AB и анализировали путем ВЭЖХ.
ПРИМЕР 6
Культивирование в биореакторе
Ферментации проводили в трехлитровых (Applikon, Foster City, CA) и 15-л (Applikon, Foster City, CA) стеклянных биореакторах и 40-литровых (Applikon, Foster City, CA) биореакторах из нержавеющей стали, стерилизуемых на месте паром. Посеянные культуры получали путем непосредственного введения среды BMGY в замороженные стоковые пробирки в объемном соотношении 1%. Засеянные флаконы инкубировали при 24°C в течение 48 часов для достижения оптической плотности (OD600) 20±5 для того, чтобы убедиться, что клетки перед переносом растут экспоненциально. Среда для культивирования содержала 40 г глицерина, 18,2 г сорбита, 2,3 г K2HPO4, 11,9 г H2PO4, 10 г дрожжевого экстракта (BD, Franklin Lakes, NJ), 20 г пептона (BD, Franklin Lakes, NJ), 4×10-3 г биотина и 13,4 г основы азотного агара для дрожжей (BD, Franklin Lakes, NJ) на литр. Биореактор засевали в объемном соотношении 10% клеток к начальной среде. Культивирования проводили способом периодических культур с подпиткой при следующих условиях: температура установлена на 24±0,5°C, pH контролировали на уровне 6,5±0,1 при помощи NH4OH, растворенный кислород поддерживали на уровне 1,7±0,1 мг/л посредством каскадной скорости перемешивания при добавлении О2. Скорость воздушного потока поддерживали на уровне 0,7 об/об в минуту. После истощения изначально внесенного глицерина (40 г/л), экспоненциально подкармливали 50% глицериновым раствором, содержащим 12,5 мл/л PTM1 солей, при 50% от максимальной скорости роста в течение восьми часов, до тех пор, пока влажная клеточная масса не достигала 250 г/л. Индукцию вызывали на тридцатой минуте фазы истощения, когда вводили экспоненциально метанол для поддержания специфической скорости роста 0,01 час-1. Когда интенсивность поглощения кислорода достигала 150 мМ/л/час, темп подачи метанола сохраняли постоянным, чтобы избежать недостатка кислорода.
ПРИМЕР 7
Очистка антител
Очистку секретируемого антитела может проводить специалист в данной области при помощи доступных опубликованных способов, например Li et al., Nat. Biotech. 24(2): 210-215 (2006), в котором антитела захватывают из ферментационного супернатанта посредством аффинной хроматографии с белком А и дополнительно очищают с использованием хроматографии на основе гидрофобных взаимодействий с фенил-сефарозной смолой с быстрой скоростью потока.
ПРИМЕР 8
MALDI-TOF анализ гликанов
N-гликаны анализировали, как описано в Choi et al., Proc. Natl. Acad. Sci. USA 100: 5022-5027 (2003), и Hamilton et al., Science 301: 1244-1246 (2003). После восстановления и карбоксиметилирования гликопротеинов, высвобождали N-гликаны с использованием обработки пептид-N-гликозидазой F. Свободные олигосахариды извлекали после осаждения белка этанолом. Молекулярные массы определяли при помощи Voyager PRO linear MALDI-TOF (Applied Biosystems) масс-спектрометра с отложенной экстракцией по инструкциям производителя. Результаты N-гликанового анализа антител к Her2, произведенных согласно вышеописанным примерам, показаны на фигурах 4-8.
ПРИМЕР 9
Анализ N-связанных гликанов посредством ВЭЖХ
Для оценки относительного количества каждой гликоформы, гликаны, высвобожденные N-гликозидазой F, метили 2-аминобензидином (2-AB) и анализировали при помощи ВЭЖХ, как описано в Choi et al., Proc. Natl. Acad. Sci. USA 100: 5022-5027 (2003), и Hamilton et al., Science 313: 1441-1443 (2006). Количества сиалированного антитела к Her2, произведенного в штамме GFI6.0 для одиночных и двойных мутеинов в 3-литровых биореакторах, и для дикого типа и двойного мутеина, произведенного в малых масштабах в 0,5-литровом биореакторе, показаны в таблице 3.
ПРИМЕР 10
Анализ аффинности к антигену
Клетки млекопитающих, экспрессирующие антиген, собирали путем трипсинизации, фильтровали через 40-мкм клеточный фильтр и суспендировали в 1% эмбриональной телячьей сыворотки (FBS) в фосфатно-солевом буфере (PBS) в 96-луночном планшете с глубокими лунками. Серийные разведения очищенного антитела добавляли к клеткам в конечной концентрации в диапазоне от 10 до 0,01 мкг/мл-1. Смесь антитело-клетки инкубировали в течение 45 минут на льду. Клетки отмывали в холодном PBS и окрашивали 2 мкг/мл-1 AlexaFluor488 против IgG человека (Invitrogen, Carlsbad, CA) в 1% FBS в PBS в течение 45 мин в темноте на льду. Клетки снова отмывали в холодном PBS, суспендировали в 1% FBS в PBS и переносили в 96-луночный планшет с U-образным дном (USA Scientific, Ocala, FL). Среднюю интенсивность флуоресценции (MFI) определяли на Guava ExpressPlus (Millipore, Billerica, MA), используя длину волны возбуждения 488 нм и длину волны испускания 525 нм. Результаты представлены на фигуре 3.
ПРИМЕР 11
Анализ связывания с FcγR
Анализ связывания с рецепторами Fcγ проводили, как описано в Shields et al., J. Biol. Chem. 276: 6591-6604 (2001) с незначительными модификациями. 96-луночные планшеты с высокой эффективностью связывания белка (Corning Costar, Lowell, MA) покрывали растворами рецепторов Fcγ в количестве 100 мкл на лунку в PBS в следующих концентрациях: 1 мкг/мл для FcγRI (R & D Systems) и FcγRIIa (произведенный Pichia pastoris), 2 мкг/мл для FcγRIIb/c (произведенный Pichia pastoris), 0,4 мкг мл для FcγRIIIa-V158 и 0,8 мкг/мл для FcγRIIIa-F158 (оба произведены Pichia pastoris). Рецепторы FcγRIIIa-V158 и FcγRIIIa-F158 экспрессировали с использованием P. pastoris, как описано в Li et al., Nat. Biotech. 24: 210-215(2006).
FcγRIIa также экспрессировали в гликоинжиниринговых Pichia с использованием такого же способа, который описан в Li et al. При помощи ПЦР проводили амплификацию внеклеточного домена FcγRIIa из кДНК человека и клонировали в вектор pCR2.1 topo. Рецепторы Fc гамма клонировали в экспрессирующий вектор Pichia с использованием пре-продомена альфа-спаривающего фактора S. cerevisiae и под контролем промотора AOX1. Конечный штамм, yGLY4665, получали трансформацией pGLY3249 в yGLY638 (хозяин GFI2.0).
FcγRIIb/c экспрессировали и производили в гликоинжиниринговом Pichia YGLY638 (хозяин GFI2.0). Последовательность ДНК внеклеточного домена человеческого Fc гамма рецептора IIb/c (NP_003992), несущая его С-концевую метку из девяти гистидинов, была подвергнута оптимизации кодонов для Pichia и обозначена pAS197 (GeneArt, Germany). Аминокислотную последовательность меченного гистидином внеклеточного домена FcγRIIb/c можно найти в SEQ ID NO:17. Для сборки плазмиды pGLY3246, кодон-оптимизированную hFcγRIIb/c (AfeI/KpnI) и αMFprepro Saccharomyces cerevisiae (EcoRI/тупой конец) клонировали в pGLY2219 по сайтам EcoR1 и Kpn1. Полученной плазмидой pGLY3246 трансформировали yGLY638 для создания yGLY4653. YGLY4653 ферментировали и очищали согласно Li et al.
Для FcγRI антитело в мономерной форме наносили в разбавителе для анализа (1% BSA, PBS, 0,05% Tween20). Для всех остальных рецепторов антитело после димеризации покрывали щелочной фосфатазой, конъюгированной с F(ab')2 к IgG человека, (Jackson ImmunoResearch, West Grove, PA) в течение одного часа при комнатной температуре. Связанное с FcγRI антитело также детектировали при помощи F(ab')2, и во всех планшетах определяли количество посредством измерения возбуждения при 340 нм и испускания при 465 нм после 18 часов инкубации с SuperPhos (Virolabs, Chantilly, VA).
Результаты представлены на фигуре 9. Как показано на фигуре 9A, одиночные Fc-мутеины (▲ и ▼) связывались с FcγRI (Fc рецептор гамма-цепь I, CD64), также как антитело к Her2 (■) (фигура 9 A) и антитело к Her2 из Pichia pastoris (данные не показаны), в то время как двойной Fc-мутеин (♦) показал приблизительно 14-кратное снижение аффинности к FcγRI (фигура 9A).
Для FcγRIIb/c одиночные Fc-мутеины (▲ и ▼) демонстрировали 10-кратное снижение связывания с рецептором по сравнению с антителом к Her2 (■) (фигура 9B), в то время как двойной Fc-мутеин, по-видимому, не связывается с FcγRIIb/c.
Для FcγRIIIa-F158 и FcγRIIIa-V158 оба одиночных Fc-мутеина (▲ и ▼) связывались с FcγRIIIa-F158 в 20 раз лучше, чем коммерческое антитело Герцептин (■) (фигура 9C), но связывание с FcγRIIIa-V158 было только незначительно лучше, чем у антитела Герцептин (фигура 9D). Двойной Fc-мутеин (♦) имеет незначительную аффинность (50-кратное снижение) к FcγRIIIa-F158 (фигура 9C), в то же время все еще сохраняя некоторую аффинность к FcγRIIIa-V158, хотя и в 30 раз слабее, чем у коммерческого антитела Герцептин (■) и антитела к Her2 из Pichia pastoris GS5.0 (данные не показаны) (фигура 9D).
Кроме того, аффинность двойного Fc-мутеина к обоим полиморфизмам FcγRIIIa, по-видимому, не меняется при высвобождении сиаловой кислоты нейраминидазой (данные не показаны). Таким образом, не желая быть связанными какой-либо теорией, авторы заявки связывают снижение аффинности к обоим полиморфизмам FcγRIIIa со структурными или конформационными изменениями, происходящими в результате двойной мутации, т.е. двойного Fc-мутеина, а не из-за повышенных уровней 2,6-связанных сиалированных N-гликанов.
ПРИМЕР 12
Анализ связывания с C1q для антител к Her2 и их Fc-мутеинов
Анализ связывания с C1q проводили при помощи способов, описанных у Idusogie et al., J. Immunology, 164: 4178-4184 (2000). Серийные разведения антитела наносили по 100 мкл на лунку в 50 мМ Na2HCO3 pH 9,0 на чистые планшеты для высокого связывания. C1q комплемента человека (US Biological, Swampscott, MA) наносили в концентрации 2 мкг/мл разбавителе для анализа (0,1% бычьего желатина, PBS, 0,05% Tween20) на два часа. C1q детектировали при помощи HRP, конъюгированного с овечьим поликлональным антителом к C1q человека (AbDSerotec), и количественно оценивали, измеряя OD450.
Результаты представлены на фигуре 10. Как показано на фигуре 10, одиночные и двойные Fc-мутеины антител, произведенные посредством материалов и способов, описанных в настоящем документе, обладают сниженным связыванием с C1q по сравнению с антителами, произведенными в клеточной культуре млекопитающего или в несиалированных штаммах Pichia pastoris. Связывание с C1q для антител, полученных из одиночных Fc-мутеинов (▲ и ▼), было снижено в 5-10 раз относительно антитела к Her2, в то время как связывание с C1q для двойного Fc-мутеина (♦) практически отсутствовало.
Как антитело к Her2, произведенное в штамме Pichia pastoris GFI5.0 (данные не показаны), так и антитело к Her2 показали сходную аффинность к C1q.
ПРИМЕР 13
Антителозависимая клеточная цитотоксичность для антитела против Her2 и его Fc-мутеинов
Антителозависимую клеточную цитотоксичность (ADCC) измеряли при помощи анализа инкорпорации европия. Линию аденокарциномы яичника человека SKOV3 культивировали в среде МакКоя 5A, дополненной 10% эмбриональной бычьей сывороткой (FBS). Мононуклеарные клетки периферической крови (PBMC) получали из Leucopaks. PBMC подвергали центрифугированию в градиенте плотности Фиколл-Гипак, отмывали в PBS, дополненном 2% FBS, и ресуспендировали в среде МакКоя 5A с 10% FBS. Генотип FcγRIIIA F158V определяли для каждого отдельного донора. Клетки SKOV3 метили EuDTPA. Клетки добавляли на 96-луночные планшеты для культивирования тканей в количестве 5000/лунку и эффекторные клетки (PBMC) добавляли в количестве 300000/лунку (E:T 60:1). Антитела добавляли в различных концентрациях и делали разведения вдоль по горизонтали планшета. Контроли включали базовые импульсы для фона, мишени и эффектора, и лунок со 100% лизисом. Смеси PBMC с антителом и без антитела инкубировали в течение четырех часов при 37°C. Высвобождение EuDTPA из лизированных PBMC определяли, смешивая 20 мкл супернатанта с DELPHIA Enhancement Solution (Perkin Elmer, Кат.№ 1244-105) для формирования высокофлуоресцентного хелата, который измеряли с использованием флуорометрии с временным разрешением. Для каждого разведения антитела делали три лунки и рассчитывали процент лизиса по формуле: ((высвобождение в эксперименте - фон)/(максимальное высвобождение - фон)) - ((естественная цитотоксичность - фон)/(максимальное высвобождение - фон)). Термины означают следующее: высвобождение в эксперименте представляет собой средний импульс для клеток-мишеней в присутствии эффекторных клеток и антитела, фон представляет собой среднее для супернатанта из последней отмывки после мечения клеток-мишеней, максимальное высвобождение представляет собой среднее для клеток-мишеней, инкубированных с лизирующим буфером DELPHIA (Perkin Elmer, Кат. № 4005-0010), и естественная цитотоксичность представляет собой средний импульс для клеток-мишеней в присутствии эффекторных клеток. Точки данных были подстроены к четырем параметрам модели логистической регрессии с использованием программного обеспечения Prism 5.0. Все кривые были ограничены одинаковым максимумом, минимумом и углом наклона.
Специалистам в данной области будет очевидно, что анализы в примерах 10-12 можно легко адаптировать под требования, относящиеся к любой молекуле иммуноглобулина. Более того, анализ ADCC in vivo на животной модели можно адаптировать для любого конкретного IgG, используя способы Borchmann et al., Blood, 102: 3737-3742 (2003); Niwa et al., Cancer Research, 64: 2127-2133 (2004).
Результаты представлены на фигуре 11. Одиночные Fc-мутеины продемонстрировали профиль активности ADCC, аналогичный профилю для антитела к Her2, и приблизительно в пять раз меньший, чем у антитела к Her2 из Pichia pastoris, произведенного в хозяине GFI5.0. Как показано на фигуре 11, активность ADCC практически отсутствовала у антитела, произведенного из двойного Fc-мутеина, что и ожидалось из данных по связыванию с FcγRIIIa.
ПРИМЕР 14
Подкожное исследование фармакокинетики (ФК) для антитела против Her2 и его Fc-мутеинов
Подкожное исследование фармакокинетики (ФК) проводили у мышей C57B6. Образцы антител вводили подкожно в дозе 1 мг/кг (n=3). 10 мкл крови собирали в капиллярную трубку в моменты времени для сбора - 1, 6, 24, 53, 77, 120, 192, 240, 288 и 360 часов, и переносили в микроцентрифужную пробирку, содержащую 90 мкл PBS, свободного от кальция и магния, и 1,8 мг/мл K2EDTA. Уровни IgG человека определяли на рабочей станции Gyros® Bioaffy (Uppsala,Sweden) с использованием иммунологического сэндвич-анализа. В качестве иммобилизованного антитела использовали 100 мкг/мл мышиного биотинилированного моноклонального антитела против каппа-цепи человека (BD Pharmingen, San Diego, CA) и в качестве детекторного антитела использовали 12,5 нМ ALEXA-647-меченого мышиного моноклонального антитела против Fc человека, Pan IgG (Southern Biotech, Birmingham, AL). Иммунологический анализ был полностью автоматизирован от иммобилизации антитела и добавления исследуемого вещества, до добавления детекторного антитела и отмывочных стадий. Стандарты и контроль качества (QC) готовили в 5% мышиной контрольной плазме (ЭДТА) в виде 20x стоков и разводили 1:20 в 5% мышиной плазме в буфере для анализа (PBS, 0,01% Tween) непосредственно перед анализом. Линейный диапазон для точного определения концентрации IgG был установлен с помощью известных количеств контроля качества и находился в пределах 5-5000 нг/мл. Точность и допустимые пределы точности составили +/-20% с нижним пределом количественной оценки (LLOQ) +/-25%. Стандарты и QC можно замораживать и оттаивать три раза без значительной потери сигнала. Стандарты, QC и образцы для исследования хранили при -70°C. Образцы для исследования оттаивали и разводили 1:20 в 5% мышиной плазме в буфере для анализа и дополнительно разводили 1:10, если уровни выходили за пределы линейного аналитического диапазона. Все стандарты, QC и образцы для исследования анализировали в двух повторениях и записывали средний результат. Концентрации определяли при помощи подбора логистической кривой с пятью параметрами. Фармакокинетические параметры рассчитывали для каждого животного при помощи WinNonlin с использованием некомпартментного анализа данных концентрация МАТ в сыворотке-время (WinNonlin Enterprise Version 5.01, Pharsight Corp, Mountain View, CA).
Как видно на фигуре 12, мыши, получавшие антитело против Her2 из Pichia pastoris с GFI5.0 гликозилированием и антитело против Her2, имеют сходные t1/2 и концентрации в сыворотке, в то время как мыши, получавшие двойной Fc-мутеин антитела против Her2, произведенный с GFI6.0 гликозилированием, демонстрировали более высокую Cmax (максимальная концентрация) и приблизительно 30% увеличение концентрации в сыворотке по сравнению с мышами, получавшими антитело против Her2, произведенное в клетках CHO. Таким образом, из трех образцов, инъецированных подкожно, двойной Fc-мутеин антитела продемонстрировал лучшую биодоступность (абсорбцию или экспозицию), основанную на более высокой Cmax и концентрациях в сыворотке.
ПРИМЕР 15
Другую серию анализов связывания с рецепторами Fcγ для антитела к Her2 по изобретению и его Fc-мутеинов проводили, как описано в примере 11, и результаты представлены на фигуре 13 и в таблицах 4 и 5.
Сравнение аффинности связывания с FcγR с антителом против Her2
V264A
Расчет соотношения: STD EC50/МАТ против антигена
Соотношение >1,0 большая аффинность, чем у АТ против Her2
Соотношение <1,0 меньшая аффинность, чем у АТ против Her2
Сравнение аффинности связывания с FcγR с антителом против Her2 из Pichia
Расчет соотношения: STD EC50/МАТ против антигена
Соотношение >1,0 большая аффинность, чем у АТ против Her2 из Pichia
Соотношение <1,0 меньшая аффинность, чем у АТ против Her2 из Pichia
ПРИМЕР 16
Оценка ADCC антител против HER2 и Fc-мутеинов
Анализ ADCC проводили с использованием клеток-мишеней SKOV3 (ATCC, Кат. № HTB-77). За день до анализа первичные эффекторные клетки NK (Biological Specialty, Кат. № 215-11-10) осаждали при 1000 об/мин в течение 15 минут и ресуспендировали до концентрации 1×106 клеток/мл в среде RPMI без фенолового красного (Invitrogen, кат.№ 11835-030), дополненной 10% FBS (Cellgro, кат.№ 35-016-CV). Ресуспендированные клетки NK инкубировали в течение ночи при 37°C при 5% CO2.
В день анализа, флакон с прикрепленными клетками-мишенями SKOV3 отмывали PBS, и отслаивали клетки с использованием 3 мл трипсина (Cellgro, кат.№ 25-053-CI) и инкубации в течение 2-5 минут при 37°C. Клетки собирали в 23 мл среды RPMI без фенолового красного с 10% FBS и пипетировали вверх-вниз, чтобы разбить сгустки. Собранные клетки центрифугировали при 1800 об/мин в течение 5 минут и ресуспендировали до концентрации 1×107 клеток/мл в среде RPMI без фенолового красного с 10% FBS. Клетки-мишени (1×107 клеток) метили 100 мкКи Chromium-51 (5 мКи хромата натрия в нормальном физиологическом растворе, Perkin Elmer, кат.№ NEZ03005MC). Клетки-мишени инкубировали при 37°C в течение одного часа со встряхиванием каждые 15 минут. Клетки центрифугировали при 1800 об/мин в течение 2 минут и ресуспендировали в 1 мл среды RPMI без фенолового красного с 10% FBS. Клетки отмывали два дополнительных раза одним мл среды RPMI без фенолового красного с 10% FBS, с центрифугированием при 1800 об/мин в течение 2 минут между каждой отмывкой. После финальной отмывки меченые клетки-мишени ресуспендировали в среде RPMI без фенолового красного с 10% FBS, до конечной концентрации 2,5×10 клетки/мл.
Исследуемые антитела, использованные в этих анализах представляли собой МАТ против HER2, произведенное в GFI5.0, МАТ против HER2 F243A, произведенное в GFI6.0 (GS6.0/F243A), МАТ против HER2 V264A, произведенное в GFI6.0 (GS6.0/V264A), и МАТ против HER2 F243A/V264A, произведенное в GFI6.0 (GS6.0/F243A/V264A). В то время как метили клетки-мишени SKOV3, исследуемые антитела разбавляли путем 3-кратной серийной титрации (начиная с 1 мкг/мл) в среде RPMI без фенолового красного с 10% FBS, в полистироловом 96-луночном планшете (Costar, кат. № 353077). В лунки отдельного 96-луночного планшета для анализа переносили клетки-мишени SKOV3, меченные 100 мкКи Cr-51 (=25,000 клеток). После приготовления планшета с разведениями антител, 10 мкл каждого разведения переносили в 96-луночный планшет для анализа, содержащий меченые клетки-мишени. Для контролей добавляли 10 мкл Triton-X100 (10% сток, Fluka Analytical, кат.№ 93443) или 10 мкл среды к контрольным лункам "максимальный лизис" или "спонтанное высвобождение", соответственно. Каждое разведение антитела тестировали в двух повторениях, в то же время тестировали 6 повторов каждого контроля.
Первичные NK-клетки осаждали при 1200 об/мин в течение 5 минут и осторожно ресуспендировали до концентрации 2,5×106 клеток/мл в среде RPMI без фенолового красного с 10% FBS. Ко всем лункам с образцами и контрольным лункам "без антитела" (т.е., исключая контроли "спонтанное высвобождение" и "максимальный лизис") добавляли 100 мкл NK-клеток (=250000 клеток) в соотношении эффектор:мишень 10:1. Планшет для анализа инкубировали при 37°C при 5% CO2 в течение четырех часов. После инкубации планшет для анализа центрифугировали при 300 об/мин в течение 5 минут. 30 мкл супернатантов добавляли к 250 мкл Microscint 20 (Perkin Elmer, Кат. № 6013621) в 96-луночном Picoplate (Perkin Elmer, Кат. № 6005185). Высвобождение Cr-51 измеряли в сцинтилляционном счетчике Packard Top Count.
Процент лизиса рассчитывали как: ((ADCC высвобождение в эксперименте - спонтанное высвобождение)/(максимальное высвобождение - спонтанное высвобождение))*100
Результаты этих анализов представлены на фигуре 14. Как показано на фигуре 14, одиночный мутеин антитела F243A и одиночный мутеин антитела V264A демонстрируют 5-10-кратное снижение активности ADCC по сравнению с родительским (дикого типа) антителом; в то время как двойной мутеин антитела F243A/V264A показывает более чем 100-кратное снижение активности ADCC по сравнению с родительским (дикого типа) антителом.
ПРИМЕР 17
Статистический анализ файла выходных данных ADCC для антитела к HER2 и его Fc-мутеинов
Задача исследования состояла в том, чтобы определить, является ли процент лизиса для варианта с двойной мутацией синергистичным по отношению к двум вариантам с единичной мутацией. Определение проводили путем сравнения ED50 (уровень антитела, соответствующий 50% лизису) между вариантами с двойной мутацией (группа 4) и предполагаемой добавочной нормативной кривой (показана на фигуре 15 и в таблице 6). Поскольку нижняя граница ED50 для кривой двойного мутанта находилась выше верхнего уровня анализируемых антител и значительно выше верхней границы ED50 для добавочной нормативной кривой, авторы сделали вывод, что эффект двойной мутации гораздо больше, чем аддитивный.
Статистические способы и результаты:
Задача состояла в том, чтобы определить, демонстрирует ли вариант антитела Герцептин с двойной мутацией (GS6.0/F243A/V264A) синергическое действие по сравнению с двумя вариантами с одиночной мутацией (GS6.0/F243A и GS6.0/V264A). Здесь было 4 группы антител: Группа 1 (GS5.0), Группа 2 (GS6.0/F243A), Группа 3 (GS6.0/V264A) и Группа 4 (GS6.0/F243A/V264A).
Показатели от трех доноров получали с отдельных 96-луночных планшетов. Наблюдения от каждого планшета были нормализованы путем превращения значений в процент лизиса следующим образом:
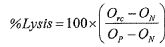 ,
,
где: Orc представляет собой наблюдаемый ответ в r-том ряду и c-той колонке каждого планшета;
ON представляет собой средний ответ отрицательных контролей для каждого планшета;
OP представляет собой средний ответ положительных контролей для каждого планшета.
Повторы значений процента лизиса затем усредняли для каждого уровня антитела и группы.
Данные от всех четырех групп и всех трех доноров моделировали совместно. Авторы предполагали, что связь «уровень антитела-ответ» подчиняется сигмоидному уравнению Хилла (Ур. 1). Авторы использовали (100% отсутствие лизиса) в качестве нашей зависимой переменной (Y) и логарифм уровня антитела в качестве независимой переменной (X). Для того чтобы провести сравнения относительно действия различных мутаций и построить модели, были сделаны некоторые допущения относительно параметров модели для каждой группы.
Форма применяемой модели допускает:
1. Хорда дуги (a) и плато (d) для всех четырех обработанных групп являются одними и теми же.
2. Один и тот же наклон кривой для всех обработанных групп, за исключением группы GS5.0, которой позволено иметь отличающийся наклон кривой.
3. Параметры EC50 (γ) различны для различных групп.
Уравнение Хилла:
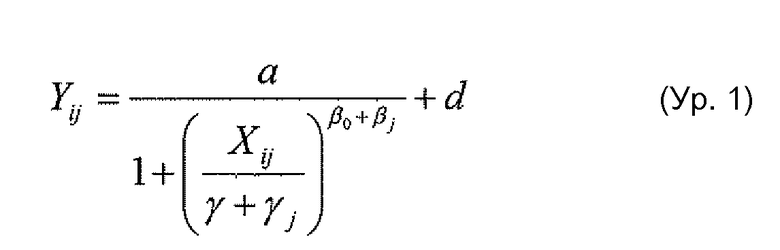
С ограничениями: γ1=0, β1=0 и β2=β3=β4=β
Здесь:
Yij представляет собой i-тый ответ (100 - процент лизиса) для j-того лечения
Xij представляет собой уровень антитела (на логарифмической шкале), соответствующий Yij
j=1 для GS5.0 j=2 для GS6.0/F243A
j=3 для GS60 V264A j=4 для GS6.0/F243A/V264A
Кроме того, дополнительно предполагали, что если два одиночных мутанта (GS6.0/F243A и GS6.0/V264A) являются аддитивными, то уравнение для комбинированных эффектов будет:
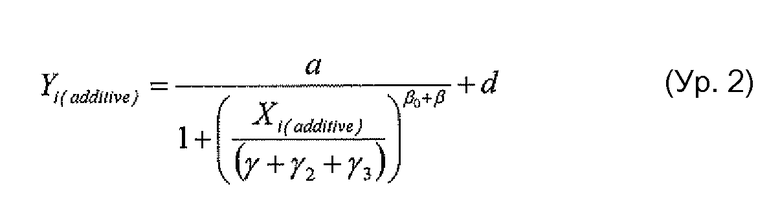 ,
,
с параметрами a, d, γ, γ2, γ3, β и β0, такими же, как в Ур. 1.
Модели (Ур. 1 и Ур. 2) построены с использованием PROC NLIN в SAS v9.2. Наблюдаемые значения данных и построенные по точкам кривые для четырех групп, а также предполагаемая добавочная нормативная кривая для комбинации групп с двумя одиночными мутациями представлены на фигуре 15. ED50 вместе с их доверительными интервалами (на оригинальной шкале) показаны в таблице 6.
Для того чтобы оценить эффекты GS6.0/F243A/V264A по сравнению с двумя единичными мутациями GS6.0/F243A и GS6.0/V264A, сравнивали ED50 группы 4 с ED50 добавочной нормативной кривой. Если доверительные интервалы не перекрывались, то делали вывод, что две величины являются статистически различными.
95% доверительный интервал для ED50 добавочной нормативной кривой составлял (0,3031, 0,5455), в то время как расчетные и доверительные интервалы для ED50 группы 4 были все больше 1 (рассчитанные значения находятся за пределами наблюдаемого диапазона уровней антитела). Нижняя граница 95% доверительного интервала для ED50 для двойной мутации >1, что гораздо больше, чем верхняя граница 95% доверительного интервала для ED50 добавочной нормативной кривой (комбинированная группа 2&3). Таким образом, можно сделать вывод, что две величины являются статистически различными.
Вышеописанное сравнение доверительных интервалов ED50 эквивалентно сравнению γ4 с суммой (γ2+γ3), т.е. проверка гипотезы H0: γ4=(γ2+γ3) против альтернативной Ha: γ4>(γ2+γ3).
Допустим, SSEreduced и SSEfull представляют собой остаточную сумму квадратов из усеченной модели и полной модели, соответственно. Полную модель получали путем аппроксимации Ур. 1 к данным, в то время как усеченную модель получали путем замещения γ4 в Ур. 1 на (γ2+γ3).
Гипотеза:
H0: γ4=(γ2+γ3) против Ha: γ4>(γ2+γ3)
Критерий значимости:
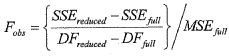
p-значение:
P(Fobs>F(DFreduced-DFfull)DFfull)<0,0001
Поскольку p-значение <0,0001, можно отклонить H0 и заключить, что γ4>(γ2+γ3).
Данные и сравнения, основанные на модели, указывают, что двойная мутация GS6.0/F243A/V264A имеет значительно более низкий процент лизиса, чем предполагаемые комбинированные эффекты двух одиночных мутаций. Эффект двойной мутации является гораздо большим, чем аддитивный эффект одиночных мутаций.
Ссылки, относящиеся к этому примеру:

ПРИМЕР 18
Создание Fc-мутеинов к TNFα и оценка ADCC
Получение двойных Fc-мутеинов (F243A/V264A) моноклонального антитела к TNF в Pichia pastoris проводили с использованием последовательностей и протоколов, перечисленных ниже. Последовательности тяжелой и легкой цепей родительского (дикого типа) антитела к TNFα изложены в SEQ ID NO:10 и 11. Последовательность тяжелой цепи двойного мутеина антитела к TNFα изложена в SEQ ID NO:12. Последовательности легкой цепи антитела дикого типа и двойного мутеина к TNFα являются идентичными.
Сигнальную последовательность предомена альфа-спаривающего фактора (SEQ ID NO:8) сливали в рамке считывания с концом легкой или тяжелой цепи посредством ПЦР слияния. Последовательность подвергали оптимизации кодонов и синтезировали у Genscript (GenScript USA Inc., 860 Centennial Ave. Piscataway, NJ 08854, USA). И тяжелую цепь, и легкую цепь клонировали в антитело-экспрессирующий вектор так же, как и при создании IgG1 против HER2 и его Fc-мутеинов.
Тяжелые и легкие цепи IgG1 и его мутеинов со слитой сигнальной последовательностью клонировали под контролем промотора AOX1 Pichia pastoris и перед терминатором Cyc S. cerevisiae, соответственно. Экспрессирующие кассеты с готовыми тяжелыми и легкими цепями помещали вместе в конечный экспрессирующий вектор. Геномную вставку в Pichia pastoris обеспечивали за счет линеаризации вектора при помощи Spe1 и нацеленной интеграции в сайт Trp2. Плазмида pGLY6964 кодирует антитело дикого типа IgG1 против TNFα. Плазмида pGLY7715 кодирует двойной мутеин F243A/V264A антитела IgG1 против TNFα.
Гликоинжиниринговые хозяева Pichia GFI5.0 YGLY8316 и GFI6.0 YGLY22834 для выработки антитела против TNFα и его Fc-мутеинов
Генотип GFI5.0 штамма YGLY16786, использованного для экспрессии антитела к TNFα и его Fc-мутеинов, выглядит следующим образом: ura5∆::ScSUC2 och1∆::lacZ bmt1∆::lacZ bmt2∆::lacZbmt3∆::lacZbmt4∆::lacZ/KlMNN2-2 mnn4L1∆::lacZ/MmSLC35A3 pno1∆::lacZ ADE1::lacZ/FB8/NA10/MmSLC35A3 his1::lacZ-URA5-lacZ/XB33/SpGALE/DmUGT arg1::HIS1/KD53/TC54 PRO1::ARG1/AOX1-ScMFpreTrMNS1PRO1::ARG1/AOX1-ScMFpreTrMNS1.
Генотип сконструированного штамма Pichia pastoris GFI6.0 YGLY23423, использованного для экспрессии двойного Fc-мутеина антитела против TNFα, выглядит следующим образом: ura5∆::ScSUC2 och1∆::lacZ bmt2∆::lacZ/KlMNN2-2 mnn4L1∆::lacZ/MmSLC35A3 pno1∆ mnn4∆::lacZ ADE1:lacZ/NA10/MmSLC35A3/FB8his1∆::lacZ/ScGAL10/XB33/DmUGT arg1∆::HIS1/KD53/TC54bmt4∆::lacZ bmt1∆::lacZ bmt3∆::lacZ TRP2:ARG1/MmCST/HsGNE/HsCSS/HsSPS/MmST6-33ste13∆::lacZ/TrMDS1 dap2∆::NatR TRP5:HygRMmCST/HsGNE/HsCSS/HsSPS/MmST6-33 Vps10-1∆::AOX1p_LmSTT3d.
Клетки трансформировали, подвергали скринингу и выделяли, как указано в примерах 4-7.
Оценка антителозависимой клеточной цитотоксичности (ADCC) Fc-мутеинов к TNFα
Анализ ADCC проводили с использованием клеток-мишеней Jurkat FlpIn, стабильно экспрессирующих не расщепляющийся вариант TNFα, у которого удалены 12 первых аминокислот - клетки-мишени Jurkat FlpIn TNFα (Δ1-12). Эти клетки получали путем трансфекции клеток Т-клеточного лейкоза человека Jurkat FlpIn (Invitrogen) не секретирующимся поверхностно-клеточным мутантом TNFα (Δ1-12 TNFα). Perez et al., Cell 63;251-258 (1990). ДНК Δ1-12 TNFα клонировали в вектор pcDNA5 (Invitrogen) для использования в трансфекции. Экспрессию TNFα на поверхности клетки подтверждали проточной цитометрией.
За день до анализа первичные эффекторные NK-клетки (Biological Specialty, Кат. № 215-11-10) осаждали при 1200 об/мин в течение 12 минут и осторожно ресуспендировали приблизительно до концентрации 1-1,5×106 клеток/мл в среде RPMI без фенолового красного, (Invitrogen, Кат. № 11835-030), дополненной 10% FBS, инактивированной нагреванием (Sigma, Кат. № F4135), 10 мМ Hepes (Gibco, Кат. № 15630), 2 мМ L-глутамином (Cellgro, Кат. № 25-005-CI) и 1X пенициллином/стрептомицином (Cellgro, Кат. № 30-002-CI). Ресуспендированные NK-клетки инкубировали в течение ночи при 37°C при 5% CO2.
В день анализа клетки-мишени Jurkat FlpIn TNFα (Δ1-12) центрифугировали при 1200 об/мин в течение 5 минут, и осторожно ресуспендировали клеточный осадок до концентрации 2×106 клеток/мл в среде RPMI без фенолового красного, дополненной 10% FBS, инактивированной нагреванием. Клетки метили 25 мкл метящего реагента DELFIA BATDA (из DELFIA EuTDA Cytotoxicity kit, Perkin Elmer, Кат. № AD0116). Клетки осторожно перемешивали и инкубировали при 37°C в течение 20 минут с легким перемешиванием каждые 10 минут. Клеточный объем доводили до 30 мл при помощи DPBS, содержащего 1,5 мМ пробеницид (Invitrogen, Кат. № P36400), и центрифугировали при 1200 об/мин в течение 5 минут. Клетки отмывали три раза при помощи 30 мл DPBS, содержащего 1,5 мМ пробеницида, с центрифугированием при 1200 об/мин в течение 5 минут между каждым отмыванием. После финального отмывания меченые клетки-мишени ресуспендировали в среде RPMI без фенолового красного, дополненной 5% FBS, инактивированной нагреванием и 1,5 мМ пробеницидом, до конечной концентрации 2,5×10 клеток/мл.
Исследуемые антитела, использованные в анализе, представляли собой: антитело дикого типа IgG1 против TNF (обозначенное "GFI774"), произведенное в GFI5.0, несиалированное антитело IgG1 против TNF (GFI774) F243A/V264A, произведенное в GFI5.0, и сиалированное антитело IgG1 против TNF (GFI774) F243A/V264A, произведенное в GFI6.0. Несиалированные антитела IgG1 против TNF - F243A/V264A - произведенные в GFI5.0, содержали следующие гликоформы: 84% G2, 2% G1 и 14% N-гликанов гибридного типа. Сиалированные антитела IgG1 против TNF - F243A/V264A - произведенные в GFI6.0, содержали следующие гликоформы: 27% Al, 50,6% A2 и 5,7% A1 гибридного типа (в целом, более чем 83% N-гликанов являются сиалированными).
В то время как метили клетки-мишени Jurkat FlpIn TNFα (Δ1-12), 2X концентрации антител разбавляли с использованием 3-кратной серийной титрации (начиная от 40 мкг/мл) в среде RPMI без фенолового красного, дополненной 5% FBS, инактивированной нагреванием и 1,5 мМ пробеницидом, в 96-луночном полипропиленовом круглодонном планшете (Costar, Кат. № 5699; крышки - Costar Кат. № 3931). После получения планшета с разведениями, 100 мкл каждого 2X разведения переносили в новый 96-луночный круглодонный полипропиленовый планшет (100 мкл только среды переносили для контролей "без антитела"). Среду RPMI без фенолового красного, дополненную 5% FBS, инактивированной нагреванием и 1,5 мМ пробеницидом, также переносили в 96-луночный планшет контролей "спонтанное высвобождение" и "максимальный лизис" (150 мкл на лунку) и контролей "фона" (200 мкл на лунку). Каждое разведение антитела тестировали в двух повторениях, в то время как каждый контроль тестировали в четырех повторениях.
Ко всем лункам, за исключением контролей "фона", добавляли 50 мкл меченных европием клеток Jurkat FlpIn TNFα (Δ1-12) (=12500 клеток) и осторожно перемешивали. Первичные NK-клетки осаждали при 1200 об/мин в течение 12 минут и осторожно ресуспендировали до концентрации 2,5×106 клеток/мл в среде RPMI без фенолового красного, дополненной 5% FBS, инактивированной нагреванием и 1,5 мМ пробеницидом. К каждой лунке с образцом (т.е., исключая контроли "спонтанное высвобождение", "максимальный лизис" и "фон") добавляли 50 мкл NK-клеток (=125000 клеток), для соотношения эффектор:мишень 10:1. Образцы осторожно перемешивали, и планшет для анализа инкубировали при 37°C в течение двух часов. Через 1 час и 15 минут добавляли 10 мкл 20% Triton-X100 (Pierce Surfact-Amps X-100, Кат. № 28314) или 10 мкл среды к контрольным лункам "максимальный лизис" или "спонтанное высвобождение", соответственно. Планшет инкубировали при 37°C в течение дополнительных 45 минут (общее время инкубации составило 2 часа). Пока планшеты инкубировались, раствор европия DELFIA (из DELFIA EuTDA Cytotoxicity kit) уравновешивали при комнатной температуре. Через два часа инкубации планшет для анализа центрифугировали при 1500 об/мин в течение 5 минут. Двадцать мкл супернатантов переносили в белый плоскодонный прозрачный планшет (из DELFIA EuTDA Cytotoxicity kit или Costar, Кат. № 3632), заботясь о том, чтобы не образовались пузыри. К каждой лунке добавляли 200 мкл раствора европия DELFIA, и герметично покрывали планшет алюминиевой фольгой. Планшет для анализа инкубировали при комнатной температуре в течение 15 минут с легким встряхиванием. Флуоресценцию измеряли с использованием установок прибора Perkin Elmer Envision для считывания европия.
Процент лизиса рассчитывали следующим образом:
1) среднее значение фонового контроля отнимали от всех значений в ряду,
2) % активности ADCC рассчитывали как: ((экспериментальное значение ADCC - спонтанное высвобождение)/(максимальный лизис - спонтанное высвобождение))*100,
и
3) окончательную величину процента лизиса записывали как процент активности ADCC из шага 2 минус контроль «без антитела».
Результаты этих анализов представлены на фигуре 16. Как видно на фигуре 16, несиалированный и сиалированный двойной мутеин F243A/V264A IgG1 против TNFα показал 1000-кратное снижение по ADCC по сравнению с родительским (дикого типа) полипептидом, произведенным в GS5.0.
ПРИМЕР 19
Оценка обусловленной комплементом цитотоксичности (CDC) двойных Fc-мутеинов против TNFα
Анализ CDC проводили с использованием клеток HEK293 FlpIn стабильно экспрессирующих нерасщепляемый вариант TNFα, у которого удалены первые 12 аминокислот. Клетки HEK293 FlpIn TNFα (Δ1-12) выращивали во флаконах для тканевых культур до 70% конфлюэнтности в минимальной питательной среде Дульбекко (DMEM) без фенолового красного (Gibco, Кат. № 21063), дополненной 10% FBS, инактивированной нагреванием (Sigma, Кат. № F4135), 100 мкг/мл гигромицина B (Cellgro, Кат. № 30-240-CR) и L-глутамина (Cellgro, Кат. № 25-005-CI). Клетки обрабатывали 2 мл трипсина (Cellgro, Кат. № MT25-053-CI), собирали в 8 мл среды DMEM без фенолового красного, с 10% инактивированной нагреванием FBS, и центрифугировали при 1200 об/мин в течение 10 минут. Супернатант удаляли, и ресуспендировали клеточный осадок в среде DMEM без фенолового красного, с 10% инактивированной нагреванием FBS, до концентрации 4×105 клеток/мл. Клетки высевали в 100 мкл (40000 клеток) на лунку в 96-луночный черный с прозрачным дном планшет (Costar, Кат. № 3603) и инкубировали в течение ночи при 37°C при 5% CO2.
Исследуемые антитела, использованные в анализе, представляли собой: антитело IgG1 против TNF (обозначенное "GFI774"), произведенное в GFI5.0, антитело IgG1 против TNF (GFI774) F243A/V264A, произведенное в GFI5.0, и антитело IgG1 против TNF (GFI774) F243A/V264A произведенное в GFI6.0 (эти антитела описаны в примере 18). В день анализа удаляли среду из лунок 96-луночного планшета мультиканальным дозатором и заменяли 50 мкл средой DMEM без фенолового красного, с 1X пенициллином/стрептомицином. 2-кратное серийное разведение исследуемого антитела (начиная с 30 мкг/мл) получали в среде DMEM без фенолового красного, содержащей 1X пенициллин/стрептомицин, 10 мкг/мл мышиного моноклонального антитела IgG1 против человеческого CD55 (IBGRL Research Products, Clone BRIC216, Кат. № 9404P) и 10 мкг/мл мышиного моноклонального антитела IgG2b против человеческого CD59 (IBGRL Research Products, Clone BRIC 229, Кат. № 9409P). В соответствующие лунки планшета для анализа добавляли 50 мкл разведенного антитела. Отрицательными контролями в анализе были отдельно среда для анализа и человеческий IgG (Jackson ImmunoResearch, Кат. № 009-000-003), разведенный в среде для анализа DMEM без фенолового красного (содержащей антитела к CD55 и CD59). Контролем лизиса в анализе был 0,25% Triton X100 (10% стоковый раствор, Fluka, Кат. № 93443), разведенный в среде для анализа DMEM без фенолового красного (содержащей антитела к CD55 и CD59). Концентрацию каждого исследуемого антитела тестировали в двух повторениях, в то время как контроли анализа тестировали в трех повторениях. Планшет для анализа, содержащий тестируемые образцы, перемешивали легким постукиванием, и инкубировали планшет приблизительно в течение 10 минут при 37°C, в то время пока готовили комплемент сыворотки человека. Три мл комплемента человека (QUIDEL, Кат. № A113) разбавляли 1:2 3 мл среды DMEM без фенолового красного, содержащей 1Х пенициллин/стрептомицин, и добавляли 50 мкл разведенного комплемента к лункам планшета для анализа. Планшет для анализа перемешивали легким постукиванием, и инкубировали планшет в течение 4 часов при 37°C при 5% CO2. Раствор 40% alamar blue (Biosource, Кат. № DAL1100) получали посредством разведения 100% стока средой DMEM без фенолового красного, содержащей 1X пенициллин/стрептомицин, и добавляли 50 мкл разведенного раствора alamar blue к лункам планшета для анализа (конечная концентрация 10%). Планшет для анализа перемешивали легким постукиванием, и инкубировали планшет в течение ночи (15-20 часов) при 37°C при 5% CO2. На следующий день планшет для анализа инкубировали в течение 10 минут на шейкере при комнатной температуре, и считывали флуоресценцию при волне возбуждения 544 нм, волне испускания 590 нм. Процент CDC рассчитывали как: (1 - (относительная единица флуоресценции (RFU) образца - RFU Triton)/(RFU среды - RFU Triton))*100.
Результаты этих анализов представлены на фигуре 17. Как показано на фигуре 17, несиалированный и сиалированный IgG1 против TNF - F243A/V264A двойной мутеин имеет приблизительно 10-кратное снижение активности CDC по сравнению с родительским антителом (дикого типа).
ПРИМЕР 20
Действие антитела к TNFα и его Fc-мутеинов на модели артрита, индуцированного антителами к коллагену (AIA)
ИНДУКЦИЯ МОДЕЛИ: AIA (Антитело-индуцированный артрит) индуцируют с использованием коммерческого артрогенного моноклонального антитела Arthrogen-CIA® (приобретенного у Chondrex), включающего в себя коктейль из 5 моноклональных антител, клон A2-10 (IgG2a), F10-21 (IgG2a), D8-6 (IgG2a), D1-2G (IgG2b) и D2-112 (IgG2b), которые распознают консервативные эпитопы на различных видах коллагена II типа.
ЖИВОТНЫЕ: Использовали 10-недельных самцов мышей B10.RIII, которые чувствительны к индукции артрита без дополнительных костимуляторных факторов. Этих животные приобретали у Jackson Laboratory.
КЛИНИЧЕСКАЯ ОЦЕНКА: Опухание лап измеряли ежедневно после индукции артрита. Тяжесть заболевания оценивали по шкале 0-3 на лапу следующим образом: 0, норма; 1, опухание одного пальца; 2, опухание двух или более пальцев; 3, отечность лапы целиком. Максимальная клиническая оценка для мыши составляла 12.
ДИЗАЙН ИССЛЕДОВАНИЯ: Артрит индуцировали пассивным переносом 3 мг патогенного коктейля МАТ против CII внутривенно в день 0.
Группы мышей лечили подкожно следующими реагентами:
Во всех группах n=5, за исключением групп A и H, в которых n=3.
Антитело с изотипом IgG1 использовали в качестве контроля. Антитело связано с мышиным антителом против гексона и имеет обозначение "27F11".
Образец, определенный как "несиалированное антитело против TNF", соответствует антителу к TNF, как описано в примере 18, произведенному в GFI5.0 и включающему мутации F243A/V264A. Образец, определенный как "α-2,6-сиалированное антитело против TNF", соответствует антителу к TNF, описанному в примере 18, произведенному в GFI6.0 и включающему мутации F243A/V264A. Несиалированные антитела IgG1 против TNF - F243A/V264A - произведенные в GFI5.0, содержали следующие гликоформы: 84% G2, 2% G1 и 14% N-гликанов гибридного типа. Сиалированные антитела IgG1 против TNF - F243A/V264A - произведенные в GFI6.0, содержали следующие гликоформы: 27% Al, 50,6% A2 и 5,7% A1 гибридного типа (в целом, более чем 83% N-гликанов являются сиалированными).
mTNFR-Ig представляет собой иммуноадгезин (Ig-слитый белок против рецептора к TNF), произведенный в клетках CHO, содержащий внеклеточный домен mTNFR2, соединенный с Fc mIgG1, начиная с шарнирной области и включая области CH2 и CH3, и содержащий аминокислотную последовательность SEQ ID NO:16.
Жидкость GAMMAGARD приобретали у Baxter Corp.
HUMIRA приобретали у Abbott Labs. HUMIRA содержит несиалированные N-гликаны с очень малым содержанием концевой галактозы.
Всем группам мышей вводили дозу в день -1 и день 7, за исключением mTNFR-Ig, который вводили в целом тремя дозами в дни -1, +3 и +7. Клиническую оценку проводили в течение 14 дней.
Результаты этих экспериментов представлены на фигурах 18A и 18B. В этом исследовании GAMMAGARD не продемонстрировал клинической эффективности. Mu-TNF-Ig продемострировал хорошую защиту от заболевания у четырех из пяти мышей. HUMIRA показала кинетику заболевания, весьма схожую с контрольным IgG1. Несиалированное антитело против TNF продемонстрировало некоторое ослабление заболевания. α-2,6-сиалированное антитело против TNF показало хорошую защиту от заболевания у пяти из пяти мышей, и оценки были сравнимы с терапией против TNF.
АНАЛИЗ ЭКСПРЕССИИ ГЕНОВ: Экспрессию воспалительных генов и генов ремоделирования костной ткани определяли путем анализа задних лап при помощи ПЦР с обратной транскрипцией у мышей, получавших изотип, α2,6-сиалированное МАТ или mTNFR-Ig (n=4 в группе). Для проведения количественной ПЦР выделяли тотальную РНК из задних лап при помощи РНК STAT-60 (Tel-Test, Friendswood, TX, USA). Тотальную РНК (5 мкг) подвергали воздействию ДНКазы (Roche). Проводили обратную транскрипцию обработанной ДНКазой тотальной РНК с использованием Superscript II (Gibco/BRL). Праймеры были разработаны с использованием Primer Express (PE Biosystems) или получены коммерческим путем от Applied Biosystems.
Количественную ПЦР в реальном времени на 10 нг кДНК от каждого образца проводили с использованием одного или двух способов. В первом способе использовали по 400 нМ двух геноспецифических немеченых праймеров в количественной ПЦР в реальном времени с SYBR green от Perkin Elmer, с использованием прибора ABI5700. Во втором способе применяли два немеченых праймера с концентрацей каждого 900 нМ вместе с 250 нМ FAM-меченного зонда (Applied Biosystems) в реакции TAQMAN™ для количественной ПЦР в реальном времени на системе для детекции последовательностей ABI7700. Отсутствие контаминации геномной ДНК подтверждали при помощи праймеров, которые распознают геномные области промотора CD4 - образцы с контаминацией ДНК, выявляемой при ПЦР в реальном времени исключали из исследования. Уровни убиквитина измеряли в отдельной реакции и использовали для нормализации данных способом Δ-Δ Ct, используя значение среднего порогового цикла (Ct) для убиквитина и интересующих генов для каждого образца; уравнение 1,8e (Ct убиквитина - Ct интересующего гена)×104 применяли для получения нормализованных значений. Результаты представлены в таблице 7. Показанные данные представляют собой кратное увеличение экспрессии воспалительных/связанных с ремоделингом костной ткани генов над экспрессией этих генов в задних лапах у контрольных наивных мышей.
ТАБЛИЦА 7
Кратность индукции по отношению к наивным мышам
ПРИМЕР 21
Действие антитела к TNFα или его Fc-мутеинов на модели артрита, индуцированного антителами к коллагену (AIA)
Эксперимент, описанный в примере 20, повторяли, как описано в этом примере, за исключением того, что вводили GAMMAGARD внутривенно (в то время как все остальные реагенты вводили подкожно).
ДИЗАЙН ИССЛЕДОВАНИЯ: Артрит индуцировали, как описано в примере 20.
Всем группам мышей вводили дозу в день -1, за исключением mTNFR-Ig, который вводили в целом двумя дозами в дни -1 и +3. Клиническую оценку проводили в течение 7 дней.
Группы мышей обрабатывали следующими реагентами:
Во всех группах n=5
Реагенты "Изотип IgG1", "несиалированное антитело против TNF", "α-2,6-сиалированное антитело против TNF”, mTNFR-Ig, GAMMAGARD и HUMIRA описаны в примере 20.
Реагент, определенный как "α-2,3-сиалированное антитело против TNF", соответствует антителу к TNF, описанному в примере 18, произведенному в GFI6.0 и включающему F243A/V264A, которое было in vitro обработано нейраминидазой для удаления α-2,6-связанной сиаловой кислоты, и дополнительно in vitro было обработано α-2,3-сиалилтрансферазой. В кратком изложении, очищенное антитело (4-5 мг/мл) находилось в сложном буфере, включавшем 6,16 мг хлорида натрия, 0,96 мг одноосновного двухводного фосфата натрия, 1,53 мг двухосновного двухводного фосфата натрия, 0,30 мг цитрата натрия, 1,30 мг моногидрата лимонной кислоты, 12 мг маннита, 1,0 мг полисорбата 80, на 1 мл с pH, скорректированным до 5,2. Нейраминидазу (l0 мЕд/мл) добавляли к смеси с антителом и инкубировали при 37°C в течение, по меньшей мере, пяти часов или до тех пор, пока десиалирование полностью не завершится. Десиалированный материал наносили на колонку для очистки CaptoMMC (GE Healthcare) для удаления нейраминидазы и заново собирали в буфер для сиалилтрансферазы (50 мМ Hepes pH 7,2, 150 мМ NaCl, 2,5 мМ CaCl2, 2,5 мМ MgC12, 2,5 мМ MnC12) в концентрации 4 мг/мл. Мышиный рекомбинантный фермент α-2,3 сиалилтрансфераза, экспрессированный в Pichia и очищенный при помощи гистидиновой метки, применяли для удлинения α-2,3 сиаловой кислотой. Ферментную смесь собирают в PBS в присутствии коктейля ингибиторов протеазы (Roche™, кат. № 11873580001) в концентрации 1,2 мг/мл. Перед реакцией сиалирования добавляли пепстатин (50 мкг/мл), химостатин (2 мг/мл) и 10 мМ CMP-сиаловой кислоты в ферментную смесь с последующей стерилизацией через 0,2-мкм фильтр. Один мл ферментной смеси добавляли к 10 мл десиалированного материала. Реакцию проводили при 37°C в течение 8 часов. Наличие сиалирования подтверждали определением массы посредством квадрупольной времяпролетной масс-спектрометрии с ионизацией электроспреем (ESI-Q-TOF). Конечный материал очищали при помощи MabSelect (GE Healthcare), собирали в вышеописанном буфере и стерильно фильтровали (0,2-мкм мембрана).
Реагент, определенный как "18 ADX PNG Humira", соответствует антителу к TNF, описанному в примере 18, произведенному в GFI6.0, содержащему F243A/V264A и обработанному in vitro ПНГазой F для удаления всех N-гликанов. Фермент ПНГазу F получали от Prozyme, Inc., и применяли в стехиометрии 2 мкл коммерческого фермента к 4 мг IgG1 при pH 7,4. Расщепленный материал очищали с использованием Mabselect (GE Healthcare). Конечный материал собирали и стерильно фильтровали (0,2-мкм мембрана).
Реагент, определенный как "20 ADX No Glyco", соответствует антителу дикого типа к TNF, описанному в примере 18, содержащему одиночную мутацию в положении 297, что приводит к отсутствию гликозилирования, и произведенное в GFI5.0 YGLY8316. Штамм, вырабатывающий это антитело, очищали посредством центрифугирования в течение 15 мин при 13000 g в Sorvall Evolution RC (kendo,Asheville, NC). Этап захвата проводили с использованием MabSelect (GE Healthcare), с последующим полирующим этапом с использованием Capto MMC (GE Healthcare). Конечный материал собирали и стерильно фильтровали (0,2-мкм мембрана).
Результаты этих экспериментов представлены на фигурах 19A и 19B.
АНАЛИЗ ЭКСПРЕССИИ ГЕНОВ: Экспрессию воспалительных генов и генов ремоделирования костной ткани определяли при помощи ПЦР с обратной транскрипцией. Для проведения количественной ПЦР выделяли тотальную РНК из задних лап при помощи РНК STAT-60 (Tel-Test, Friendswood, TX, USA). Тотальную РНК (5 мкг) подвергали воздействию ДНКазы (Roche). Проводили обратную транскрипцию обработанной ДНКазой тотальной РНК с использованием Superscript II (Gibco/BRL). Праймеры были разработаны с использованием Primer Express (PE Biosystems) или получены коммерческим путем от Applied Biosystems.
Количественную ПЦР в реальном времени на 10 нг кДНК от каждого образца проводили с использованием одного или двух способов. В первом способе использовали по 400 нМ двух геноспецифических немеченых праймеров в количественной ПЦР в реальном времени с SYBR green от Perkin Elmer, с использованием прибора ABI5700. Во втором способе применяли два немеченых праймера с концентрацей каждого 900 нМ вместе с 250 нМ FAM-меченного зонда (Applied Biosystems) в реакции TAQMAN™ для количественной ПЦР в реальном времени на системе для детекции последовательностей ABI7700. Отсутствие контаминации геномной ДНК подтверждали при помощи праймеров, которые распознают геномные области промотора CD4 - образцы с контаминацией ДНК, выявляемой при ПЦР в реальном времени исключали из исследования. Уровни убиквитина измеряли в отдельной реакции и использовали для нормализации данных способом Δ-Δ Ct, используя значение среднего порогового цикла (Ct) для убиквитина и интересующих генов для каждого образца; уравнение 1,8e (Ct убиквитина - Ct интересующего гена)×104 применяли для получения нормализованных значений. Результаты представлены в таблице 8. Показанные данные представляют собой кратное увеличение экспрессии воспалительных/связанных с ремоделингом костной ткани генов над экспрессией этих генов в задних лапах у контрольных наивных мышей.
Кратность индукции по отношению к наивным мышам
Humira
36ADV
Humira
19ADX
Guard
84ADU
82ABW
Humira
37ADV
ПРИМЕР 22
Анализы связывания антител к TNF и мутеинов с FcγR
Анализы связывания с рецепторами Fcγ проводили, как описано в примере 1, с использованием антител к TNF, описанных в примере 18. Результаты представлены на фигуре 20 и в таблицах 9-10.
Уменьшение связывания с рецепторами Fcγ по сравнению с антителом дикого типа (родительским), произведенным в GS 5.0
LV
↑ указывает на разы повышения аффинности
Уменьшение связывания с рецепторами Fcγ по сравнению с коммерческим антителом HUMIRA
LV
↑ указывает на разы повышения аффинности
ПРИМЕР 23
Создание дополнительных двойных Fc-мутеинов (по положениям 243/264) антитела к Her2 и их N-гликановая композиция
Дополнительный двойные Fc-мутеины антитела IgGl к Her2, описанного в примере 2, конструировали с использованием набора для сайт-специфического мутагенеза Stratagene QuikChange® кат. № 200518 (30 реакций), следуя протоколу, который предлагает производитель для способа насыщающего мутагенеза с вырожденными праймерами. Сигнальную последовательность предомена альфа-спаривающего фактора сливали в рамке считывания с 5'-концом легкой цепи и тяжелой цепи. Полученные тяжелые и легкие цепи IgG1 и его мутеинов со слитой сигнальной последовательностью клонировали под контролем промотора AOX1 Pichia pastoris и перед терминатором Cyc S. cerevisiae, соответственно. Экспрессирующие кассеты с готовыми тяжелыми и легкими цепями помещали вместе в конечный экспрессирующий вектор.
Векторы экспрессировали в гликоинжиниринговых клетках-хозяевах Pichia GFI6.0 YGLY3582 и YGLY22812.
Генотип штамма GFI6.0 YGLY3582 описан в примере 4 (такой же, как у YGLY4563).
Генотип сконструированного штамма Pichia pastoris GFI6.0 YGLY22812, использованный для экспрессии двойного Fc-мутеина антитела к TNFα, является следующим: ura5∆::ScSUC2 och1∆::lacZ bmt2∆::lacZ/KlMNN2-2 mnn4L1∆::lacZ/MmSLC35A3 pno1∆ mnn4∆::lacZ ADE1:lacZ/NA10/MmSLC35A3/FB8his1∆::lacZ/ScGAL10/XB33/DmUGT arg1∆::HIS1/KD53/TC54bmt4∆::lacZ bmt1∆::lacZ bmt3∆::lacZ TRP2:ARG1/MmCST/HsGNE/HsCSS/HsSPS/MmST6-33ste13∆::lacZ/TrMDS1 dap2∆::NatR TRP5:HygRMmCST/HsGNE/HsCSS/HsSPS/MmST6-33 Vps10-1∆::AOX1p_LmSTT3d. YGLY23294, YGLY23280, YGLY23301, YGLY25259 и YGLY23305 имеют одинаковый генотип, за исключением того, что они экспрессируют разные мутеины, перечисленные в таблице 11.
Штаммы культивировали во встряхиваемых колбах на 500 мл с 300 мл среды 2% BMGY и встряхивали при 24°C в течение 3 дней.
Протокол для индукции содержимого встряхиваемых колб: Собрать общий объем каждой культуры (300 мл) в пробирки фирмы Фалькон и центрифугировать при 2500 об/мин в течение 5 минут. Вылить супернатант и ресуспендировать клеточные осадки в конечном объеме 150 мл 2% BMMY и 360 мкл ΡΜTi4 (стоковая концентрация 0,65 мг/мл). Перенести в чистую встряхиваемую колбу на 500 мл и встряхивать при 24°C в течение 2 суток. Центрифугировать индуцированные культуры и собрать супернатант в чистые пробирки фирмы Фалькон.
Секретированные антитела очищали на колонках с белком A с использованием GE Healthcare, STREAMLINE rProtein A (Кат. № 17-1281-01) и хроматографических колонок BioRad poly-prep (10 мл) (Кат. № 731-1550). Использовали следующие буферы:
• Отмывочный буфер #1: 20 мМ Tris pH 7,0, 1M NaCl
• Отмывочный буфер #2: 20 мМ Tris pH 7,0
• Нейтрализующий буфер: 1M Tris pH 8,0-pH 9,0
• Элюирующий буфер: 100 мМ или 50 мМ цитрата натрия pH 3,0
• Очищающий раствор: 6 M мочевина в воде.
Протокол очистки проводят следующим образом:
• Добавить 500 мкл частиц STREAMLINE rProtein A к каждой колонке BioRad. Частицы должны находиться в 20% этаноле. Композиция взвеси частиц должна состоять из 50% частиц, 50% жидкости.
• Как только частицы белка A окажутся в колонке, их следует промыть 5 мл отмывочного буфера #2 (удалить элюат).
• Добавить l0 мл супернатанта на колонку BioRad. На этом этапе антитела связываются с частицами белка A (удалить элюат).
• Смыть нежелательный избыток белков, добавив 5 мл отмывочного буфера #1 на колонку (удалить элюат).
• Снова промыть колонку, добавив 5 мл отмывочного буфера #2 (удалить элюат).
• Добавить 1 мл нейтрализующего буфера в 15-мл пробирку для сбора белковых образцов.
• Поместить колонку BioRad в 15-мл пробирку для сбора образцов.
• Добавить 3 мл элюирующего буфера в колонку BioRad. Это удаляет желаемые антитела с частиц белка A.
• Собрать элюированный белок в 15-мл пробирку для сбора белковых образцов.
• Определить концентрацию элюированного белка путем анализа по Брэдфорду (использовать l0 мкл белка для анализа по Брэдфорду).
Для оценки относительного количества каждой гликоформы, гликаны, высвобожденные N-гликозидазой F, метили 2-аминобензидином (2-AB) и анализировали при помощи ВЭЖХ, как описано у Choi et al., Proc. Natl Acad. Sci. USA 100: 5022-5027 (2003), и Hamilton et al., Science 313: 1441-1443 (2006). Таблица 12 показывает гликановые профили произведенных антител (NQP=количественная оценка невозможна).
Связывание вышеописанных мутантов с различными рецепторами Fcγ определяли при помощи способа, описанного в примере 11. Результаты представлены на фигуре 21 и в таблицах 13 и 14.
Аффинность связывания по сравнению с двойным мутеином F243A/V264A
Аффинность связывания по сравнению с коммерчески доступным Герцептином
ПРИМЕР 23
Родительское антитело против PCKS9 и Fc-мутеины
Было также сконструировано антитело к PCSK9 с аминокислотной последовательностью тяжелой цепи из SEQ ID NO:13 и аминокислотной последовательностью легкой цепи из SEQ ID NO:14, и в тяжелую цепь были введены мутациии F243A и V264A, как описано в примере 2. Аминокислотная последовательность тяжелой цепи двойного мутеина против PCSK9 представлена в SEQ ID NO:15. Формы антитела дикого типа и двойного мутеина экспрессировали в гликоинжиниринговых Pichia GFI5.0 и GFI6.0, очищали согласно способам, описанным в примерах 3-7, и определяли их способность связываться с различными рецепторами Fcγ с использованием способов, описанных в примере 11. Двойной мутеин антитела против PCSK9 (произведенный в GFI6.0, включающее аминокислотную последовательность тяжелой цепи из SEQ ID NO:15 и аминокислотную последовательность легкой цепи из SEQ ID NO:14), по сравнению с родительским антителом (произведенным в GFI5.0) показал приблизительно 4-кратное снижение связывания с FcγRI, 8-60-кратное снижение связывания с FcγRIIIa LF, 2-6-кратное снижение связывания с FcγRIIIa LV и отсутствие выявляемого связывания с FcγRIIa или FcγRIIb/c.
ПРИМЕР 24
Действие антитела к Her2 и его Fc-мутеинов на модели артрита, индуцированного антителами к коллагену (AIA)
ИНДУКЦИЯ МОДЕЛИ: AIA (Антитело-индуцированный артрит) индуцируют с использованием коммерческого артрогенного моноклонального антитела Arthrogen-CIA® (приобретенного у Chondrex), включающего в себя коктейль из 5 моноклональных антител: клон A2-10 (IgG2a), F10-21 (IgG2a), D8-6 (IgG2a), D1-2G(IgG2b) и D2-112 (IgG2b), которые распознают консервативные эпитопы на различных видах коллагена II типа.
ЖИВОТНЫЕ: Использовали 10-недельных самцов мышей B10.RIII, которые чувствительны к индукции артрита без дополнительных костимуляторных факторов. Этих животных приобретали у Jackson Laboratory.
КЛИНИЧЕСКАЯ ОЦЕНКА: Опухание лап измеряли ежедневно после индукции артрита. Тяжесть заболевания оценивали по шкале 0-3 на лапу следующим образом: 0, норма; 1, опухание одного пальца; 2, опухание двух или более пальцев; 3, отечность лапы целиком. Максимальная клиническая оценка для мыши составляла 12.
ДИЗАЙН ИССЛЕДОВАНИЯ: Артрит индуцировали пассивным переносом 3 мг патогенного коктейля IV МАТ против CII внутривенно в день 0.
Всем группам мышей вводили дозу в день -1. Клиническую оценку проводили в течение 7 дней.
Группы мышей обрабатывали подкожно следующими реагентами:
Во всех группах n=5, за исключением группы E, в которой n=4
Антитело с изотипом IgG1 использовали в качестве контроля. Антитело связано с мышиным антителом против гексона и имеет обозначение "27F11".
Образец, определенный как "несиалированное антитело против Her2", соответствует антителу к Her2, содержащему мутации F243A/V264A и произведенному в GFI5.0, штамм YGLY19709. Штамм YGLY19709 получен из штамма YGLY13979, продуцирующего антитело к Her2, который был трансформирован плазмидой pGLY6301 для экспрессии LsSTT3d. Штамм YGLY13979 и плазмида pGLY6301 описаны в патентной заявке WO 2010/099186.
Образец, определенный как "α-2,6-сиалированное антитело против Her2", соответствует антителу к Her2, содержащему мутации F243A/V264A и произведенному в GFI6.0, штамм YGLY4563 (см. пример 4).
mTNFR-Ig представляет собой иммуноадгезин (Ig-слитый белок против рецептора TNF), произведенный в клетках CHO, содержащий внеклеточный домен mTNFR2, соединенный с Fc mIgG1, начиная с шарнирной области и включая области CH2 и CH3, и содержащий аминокислотную последовательность SEQ ID NO:16.
Результаты этих экспериментов показаны на фигуре 22.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ





Хотя настоящее изобретение описано в настоящем документе со ссылкой на проиллюстрированные варианты осуществления, следует понимать, что изобретение не ограничено ими. Специалисты в данной области, имеющие доступ к идеям в настоящем документе, понимают дополнительные модификации и варианты осуществления в объеме изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2711935C2 |
| ГИПЕРГЛИКОЗИЛИРОВАННЫЕ СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2708314C2 |
| ПРОДУКЦИЯ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ФУКОЗИЛИРОВАНИЕМ | 2008 |
|
RU2479629C2 |
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2831409C2 |
| ПОЛИПЕПТИДЫ ГЕМАГГЛЮТИНИНОВ, А ТАКЖЕ СВЯЗАННЫЕ С НИМИ РЕАГЕНТЫ И СПОСОБЫ | 2011 |
|
RU2663718C2 |
| АНТИТЕЛО ПРОТИВ MUC1 | 2019 |
|
RU2792347C2 |
| ВЕКТОР ДЛЯ ЭКСПРЕССИИ ПОЛИПЕПТИДОВ С СИАЛИДАЗНОЙ АКТИВНОСТЬЮ, СПОСОБ ОБЕСПЕЧЕНИЯ СИАЛИДАЗНОЙ АКТИВНОСТИ В КУЛЬТУРЕ КЛЕТОК И СПОСОБ РЕГУЛИРОВАНИЯ СВОЙСТВ Fc-СОДЕРЖАЩИХ МОЛЕКУЛ, ЭКСПРЕССИРУЕМЫХ В ЛИНИИ КЛЕТОК | 2007 |
|
RU2466189C2 |
| РЕКОМБИНАНТНЫЕ ГЛИКОПРОТЕИНЫ С СИАЛИРОВАННЫМИ О-ГЛИКАНАМИ И ЛИНИИ КЛЕТОК ДЛЯ ИХ ПРОДУКЦИИ | 2015 |
|
RU2714209C2 |
| СПОСОБЫ, ОТНОСЯЩИЕСЯ К МОДИФИЦИРОВАННЫМ ГЛИКАНАМ | 2009 |
|
RU2526250C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ГЛИКОЗИЛИРОВАНИЕМ | 2015 |
|
RU2704830C2 |






















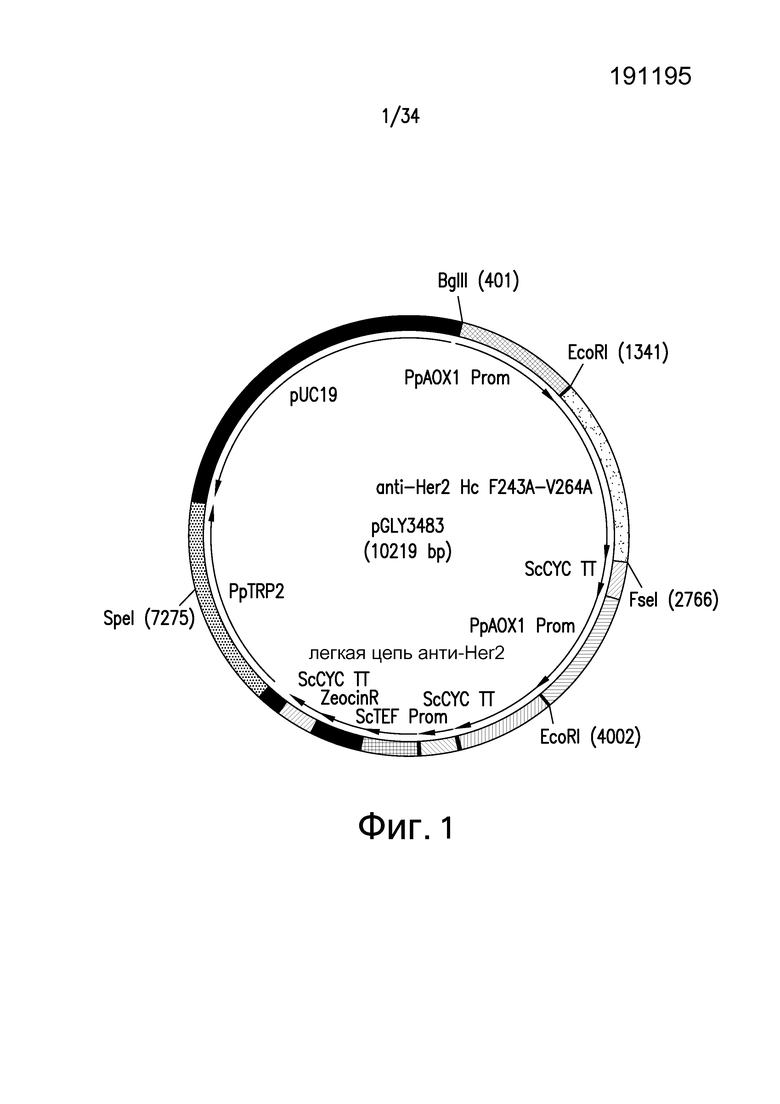
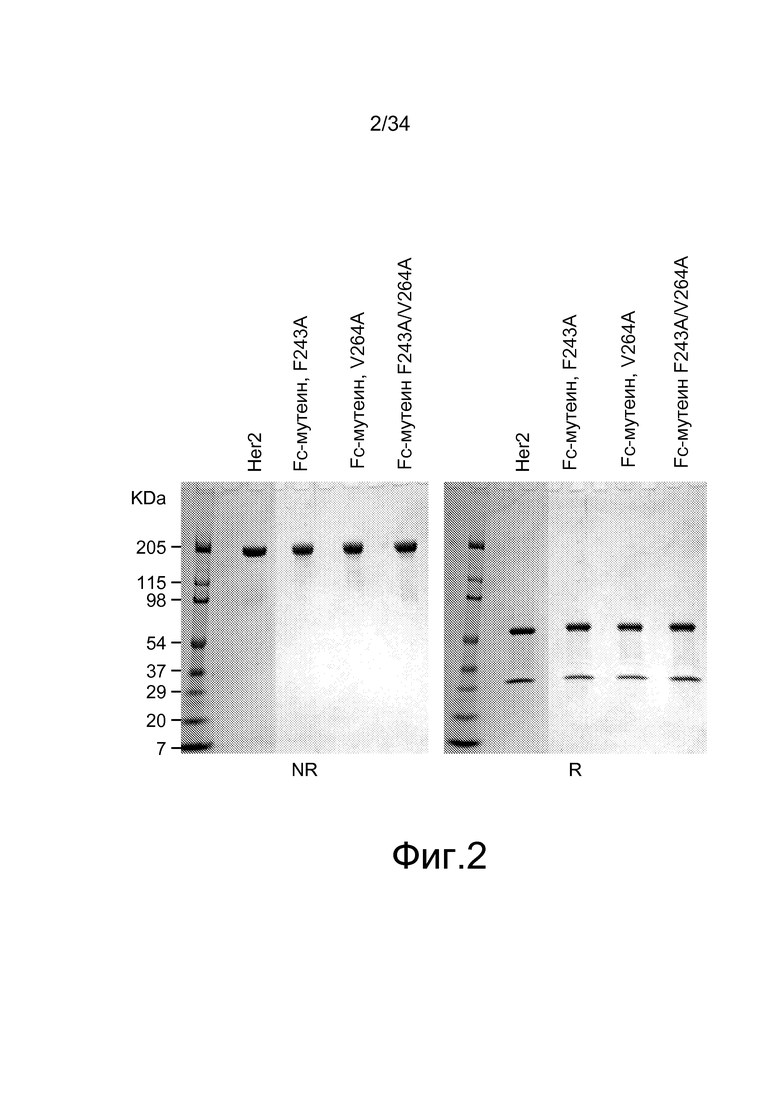
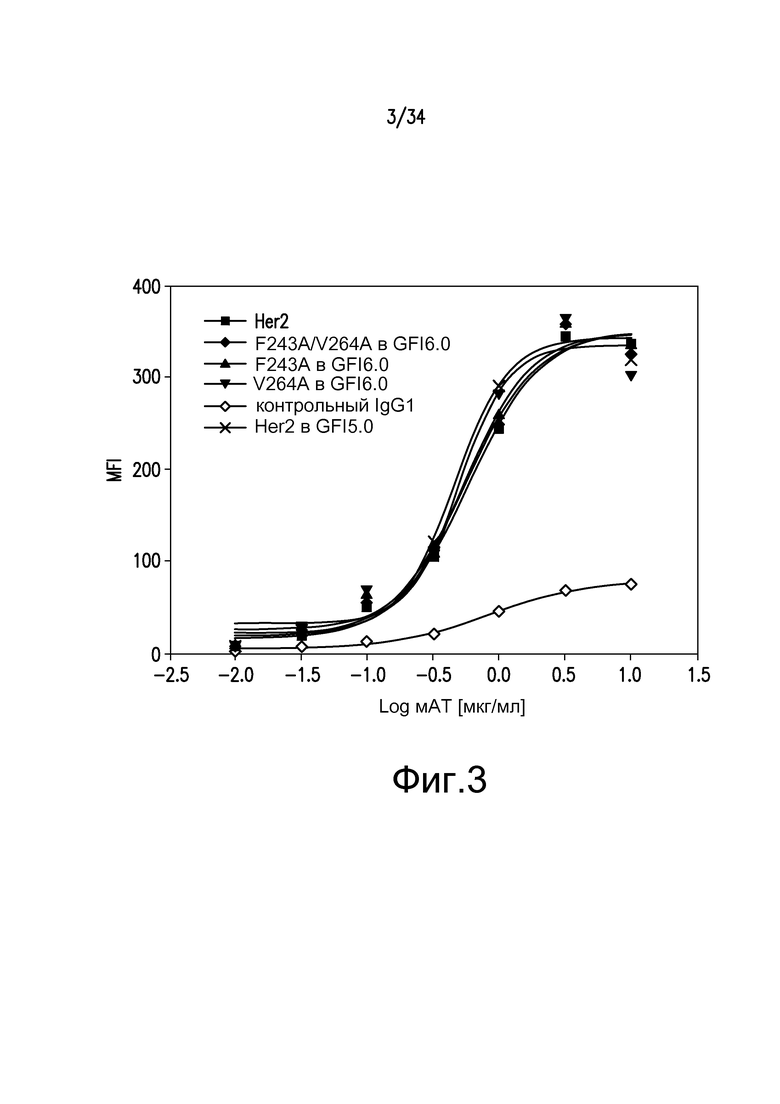
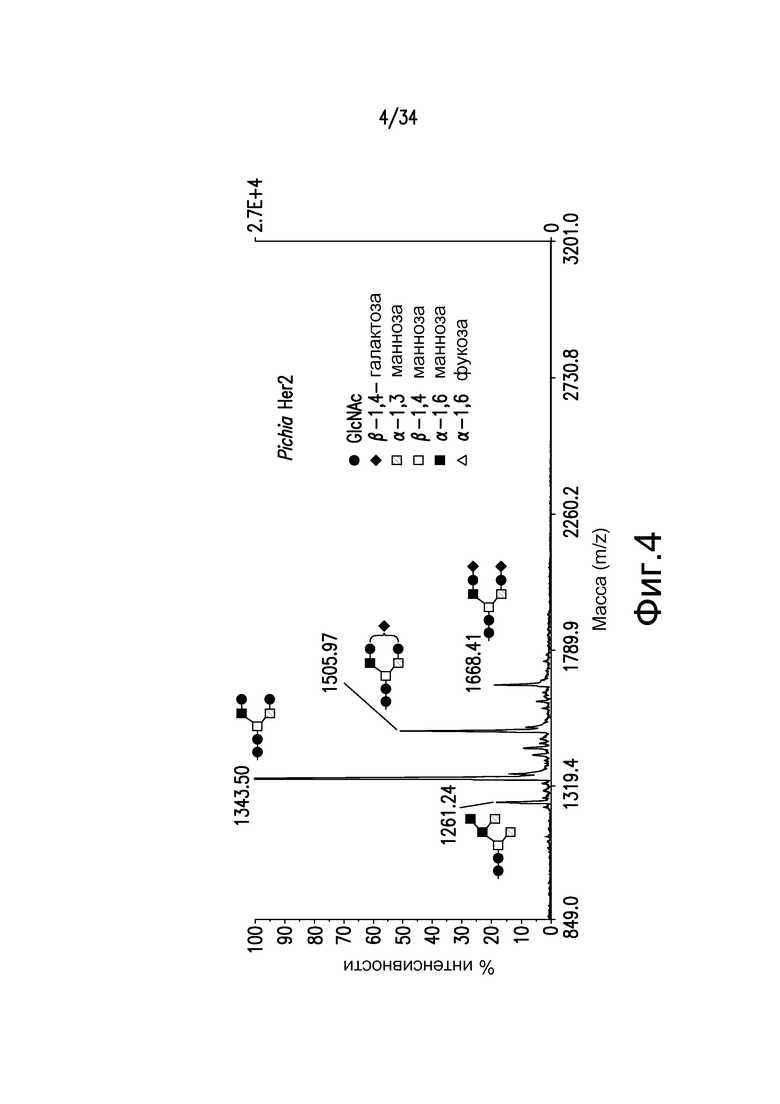
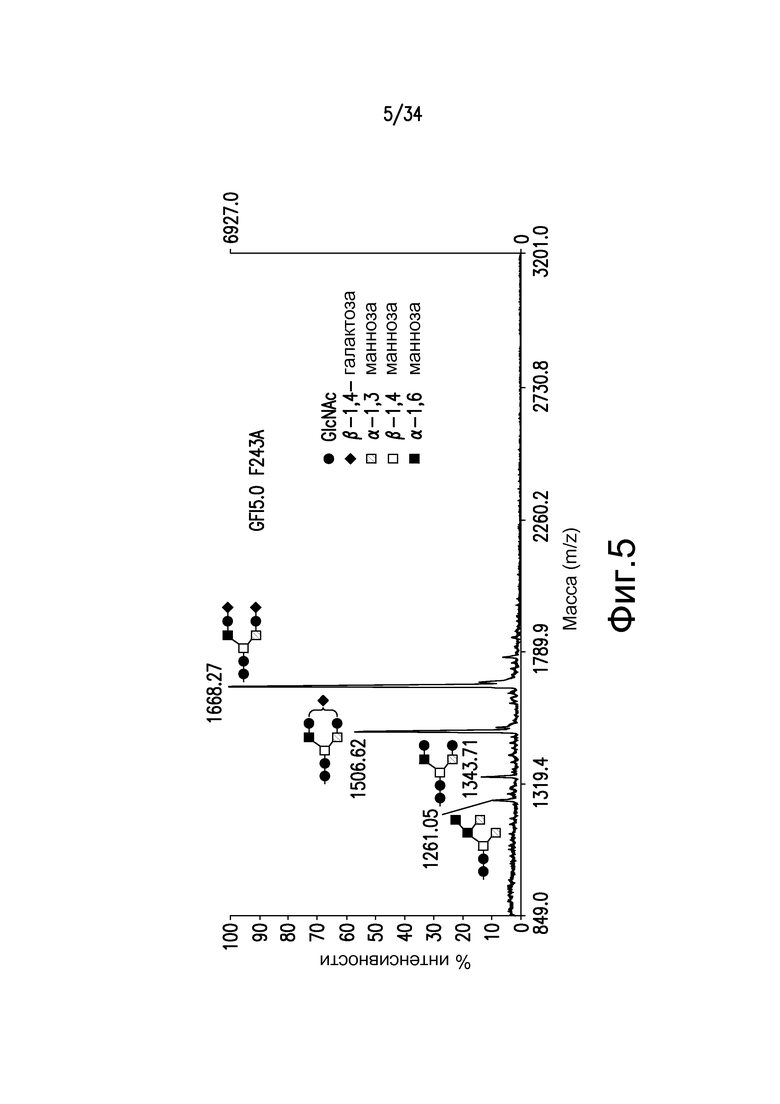
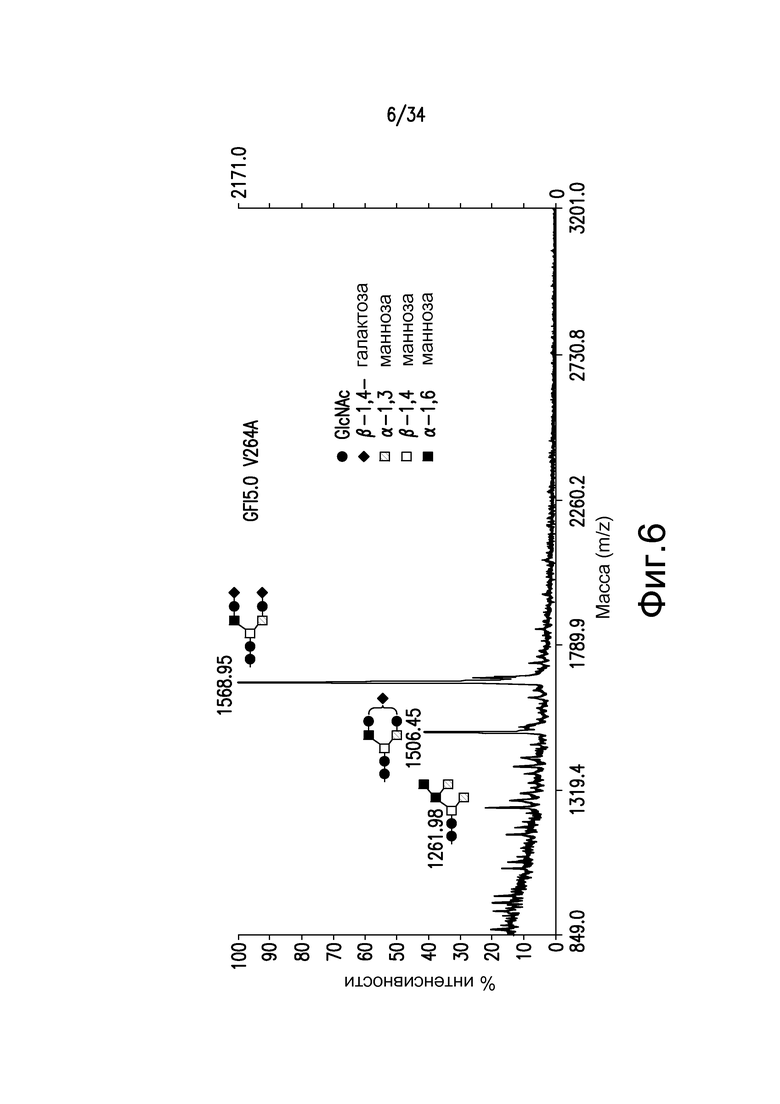
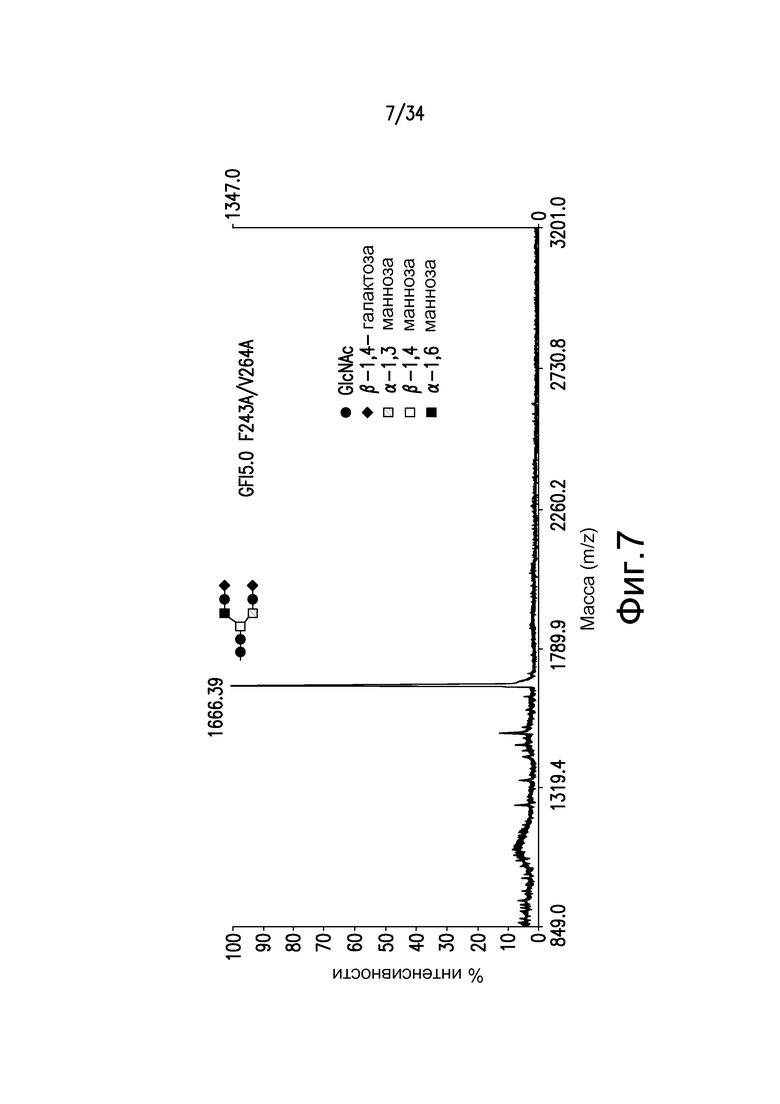
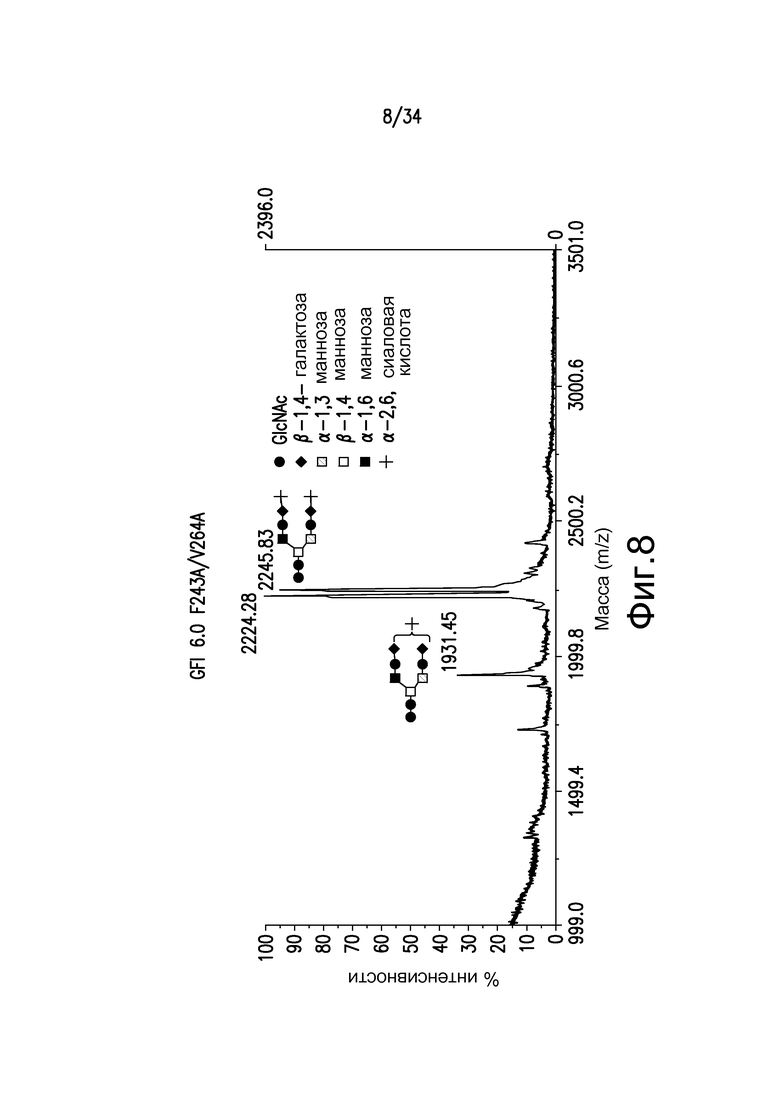
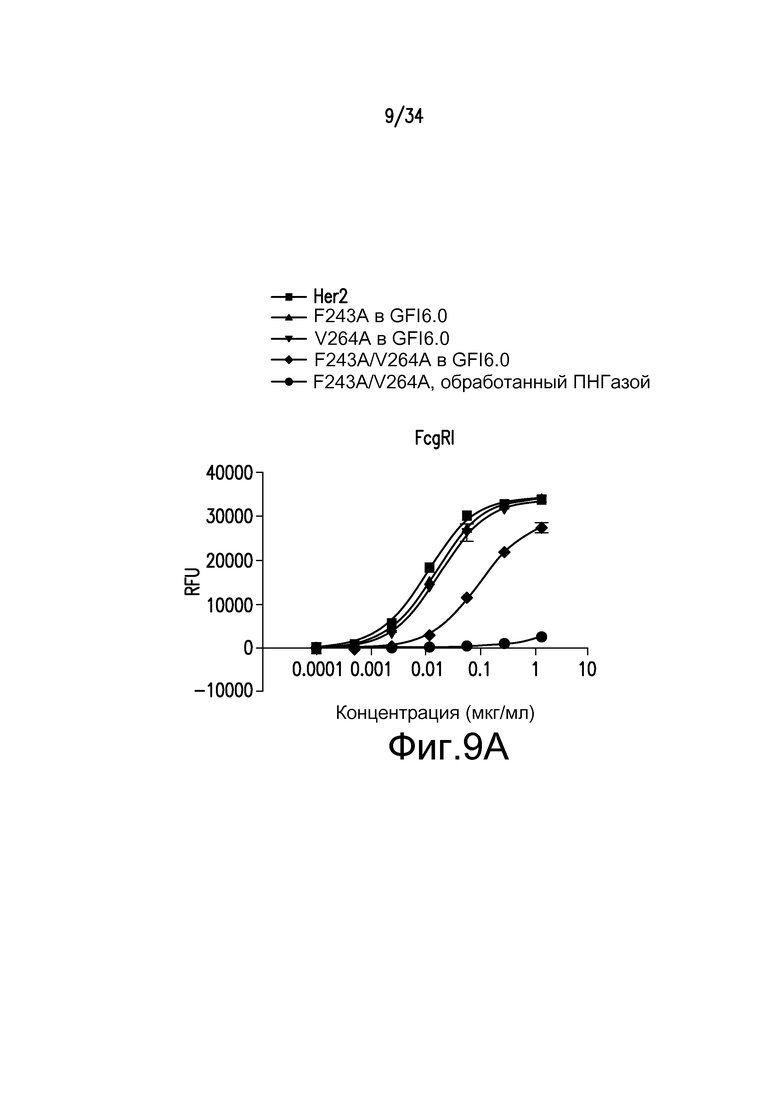
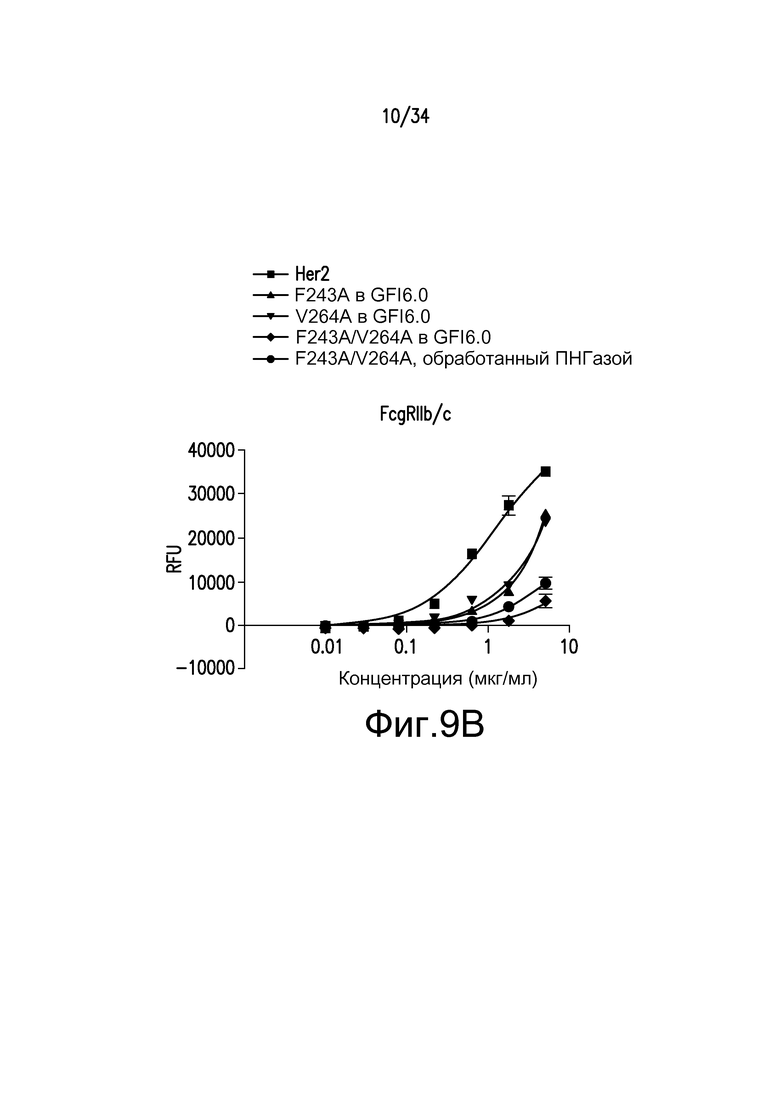
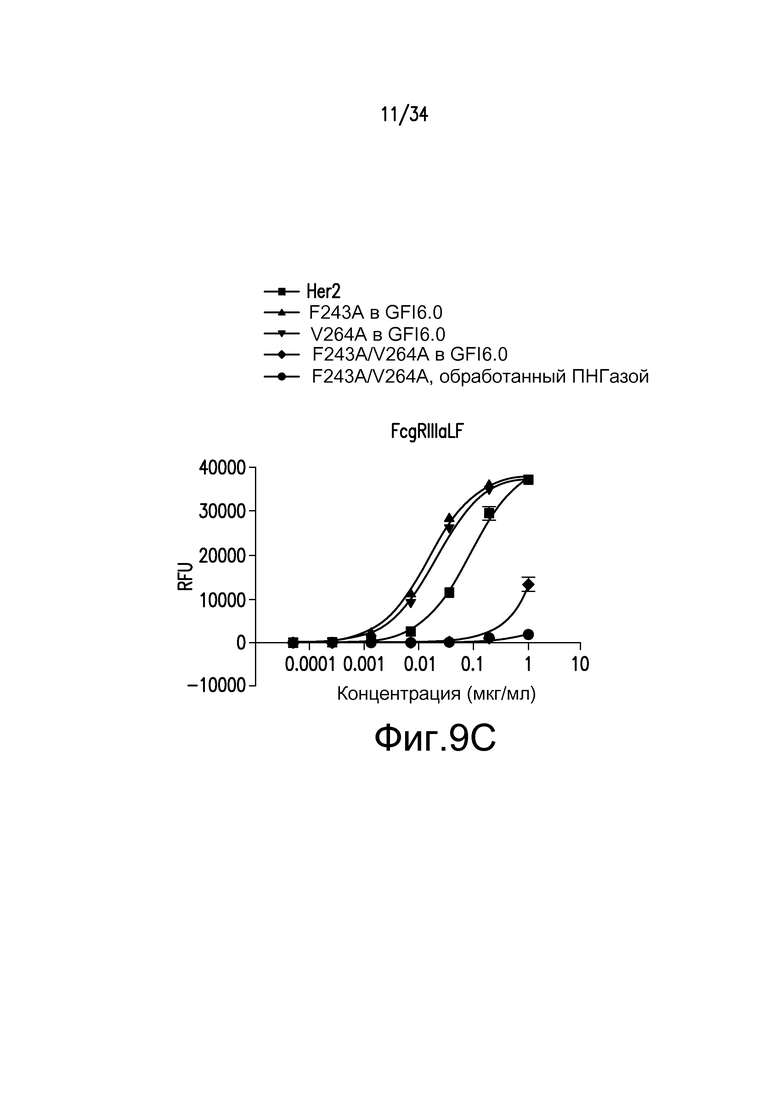
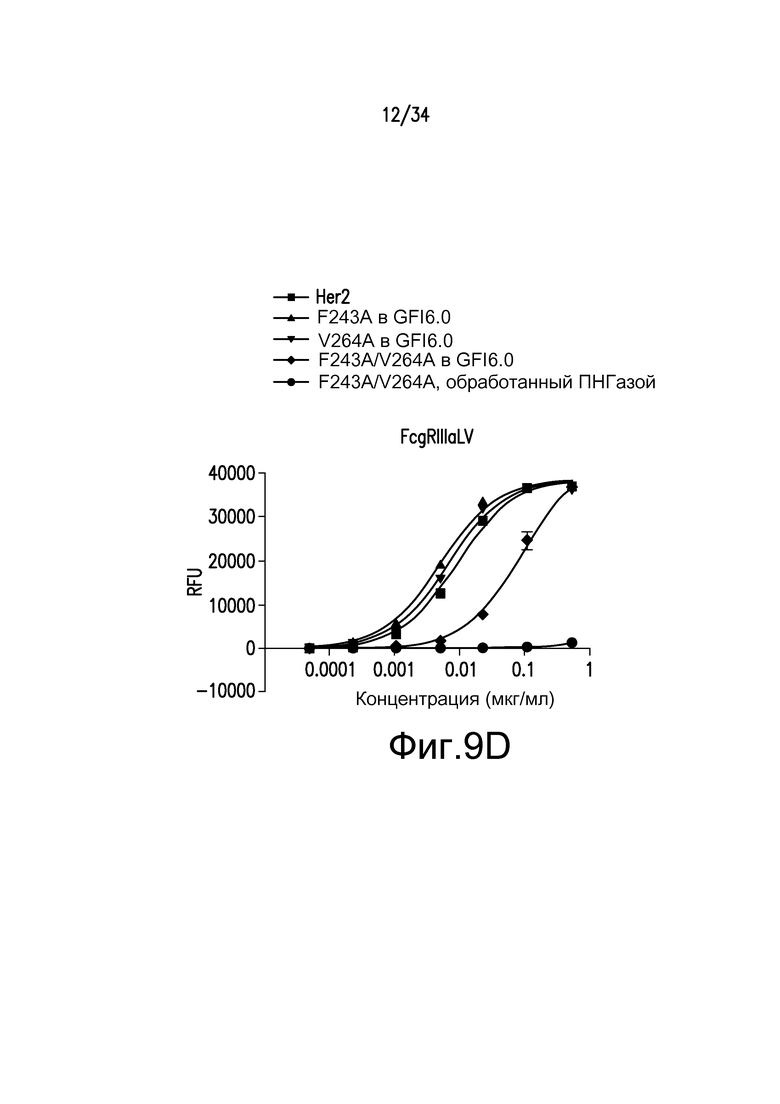
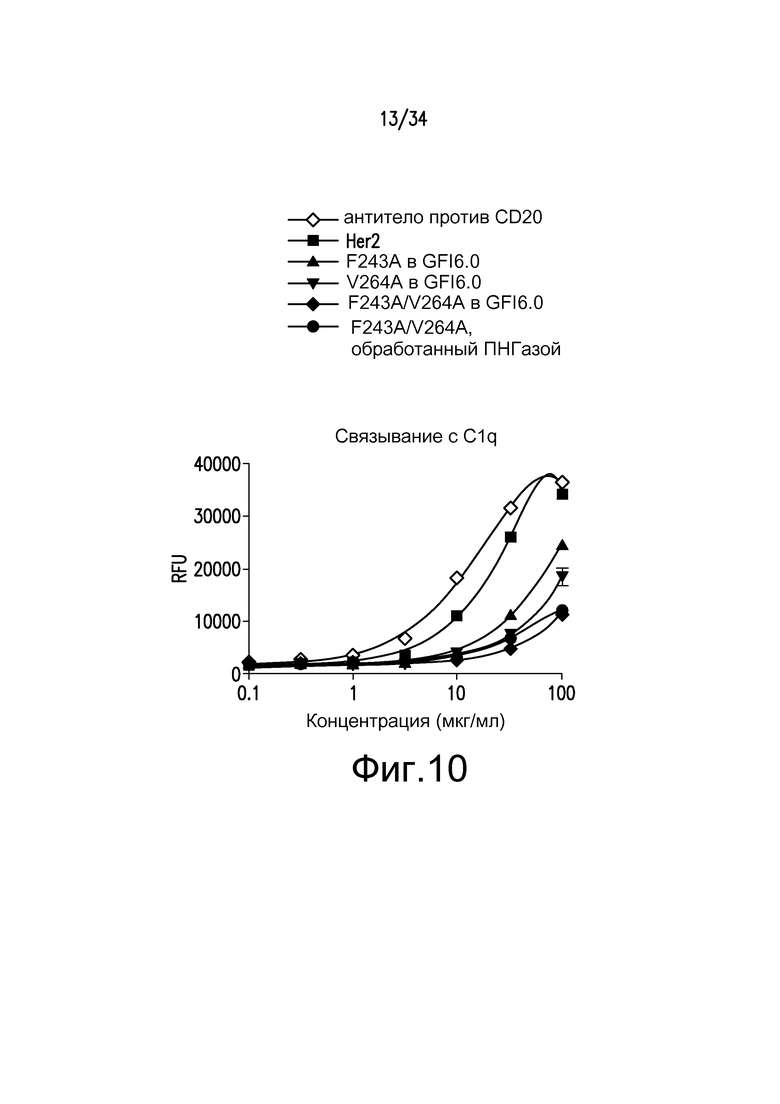
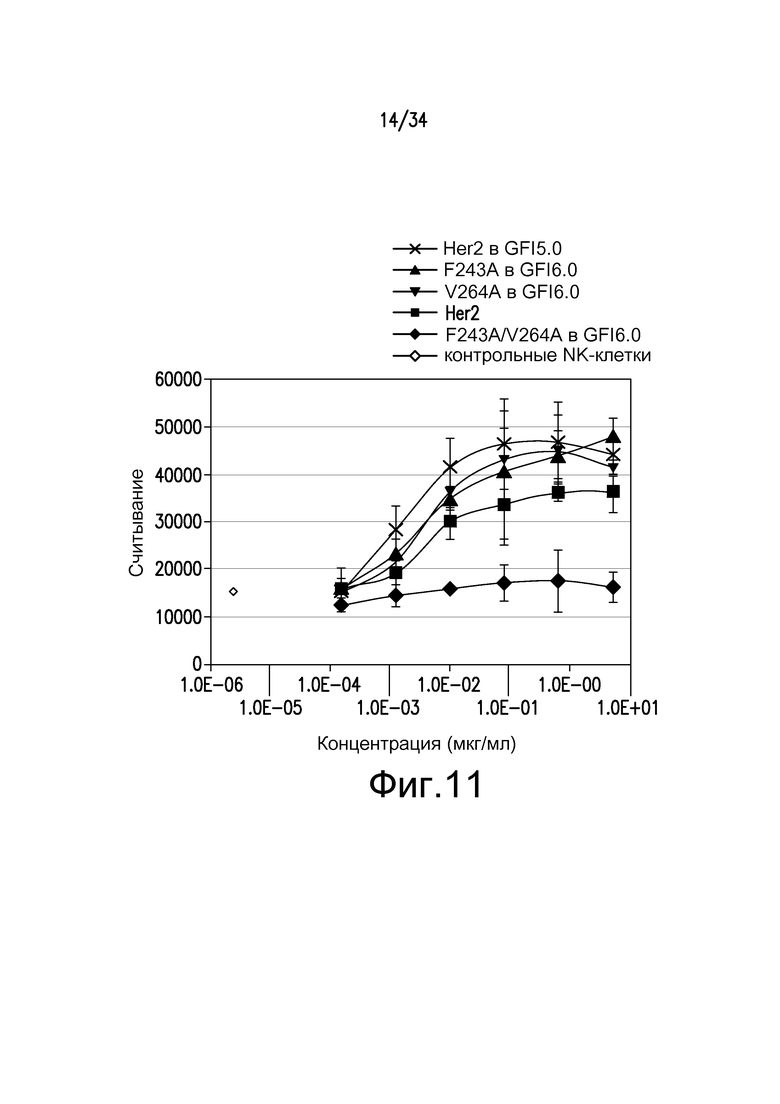
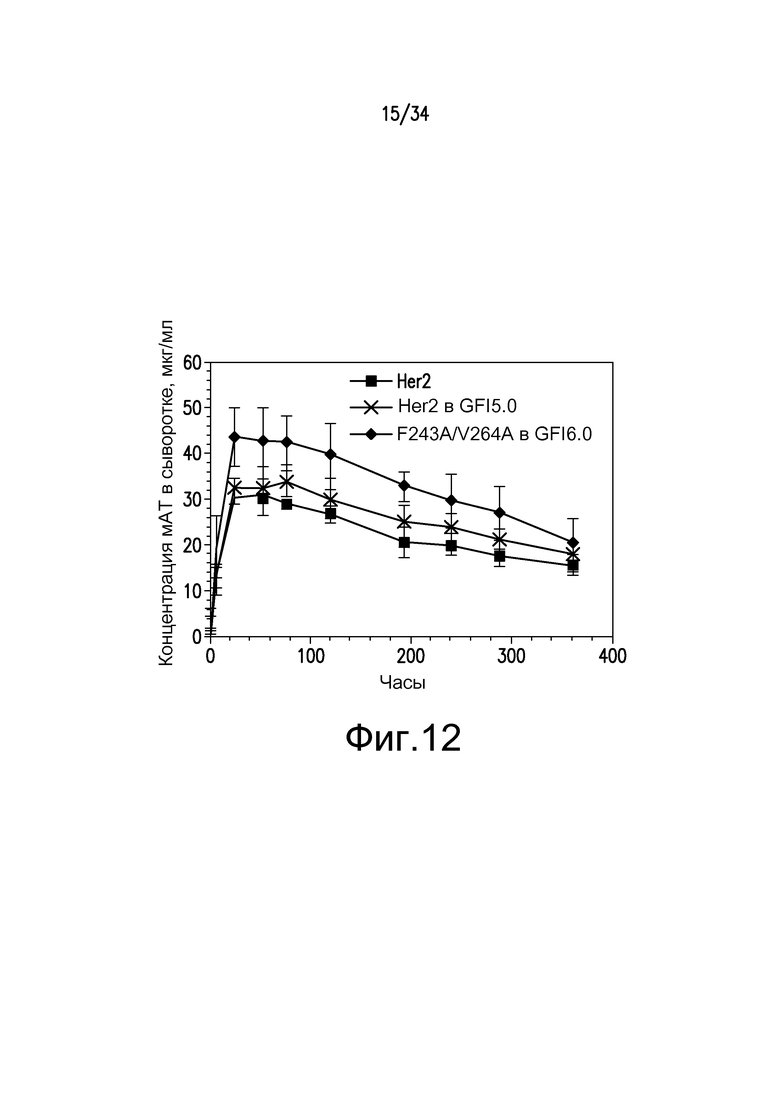
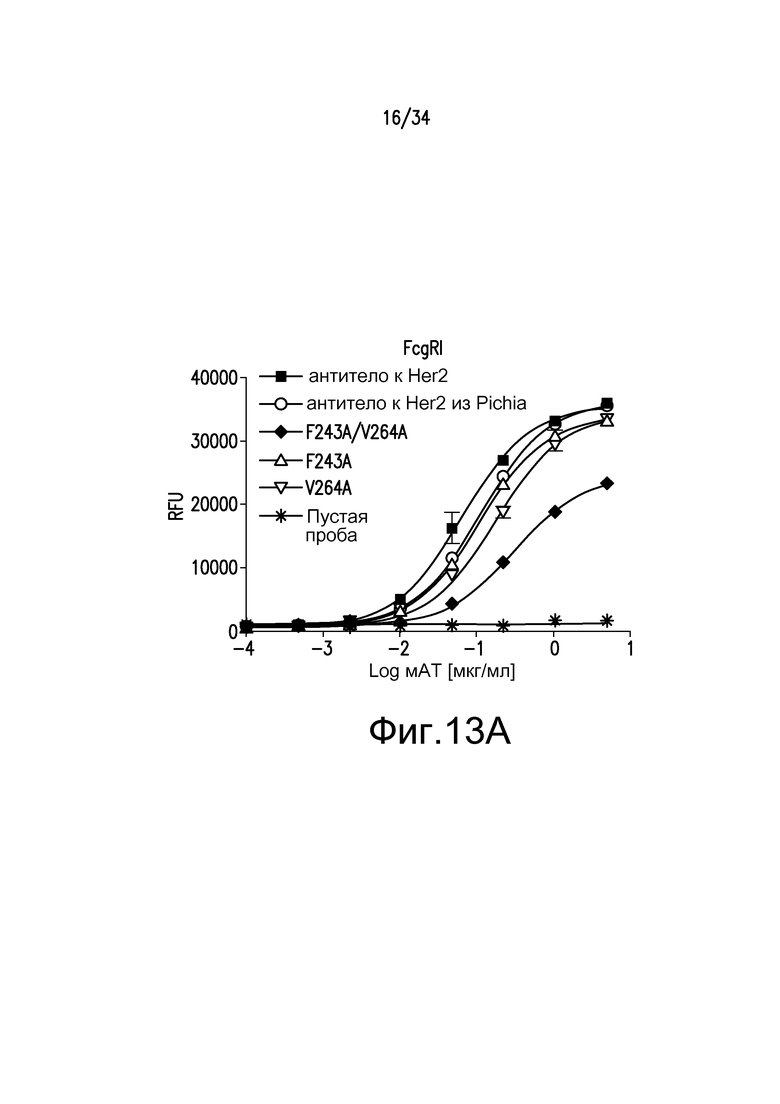
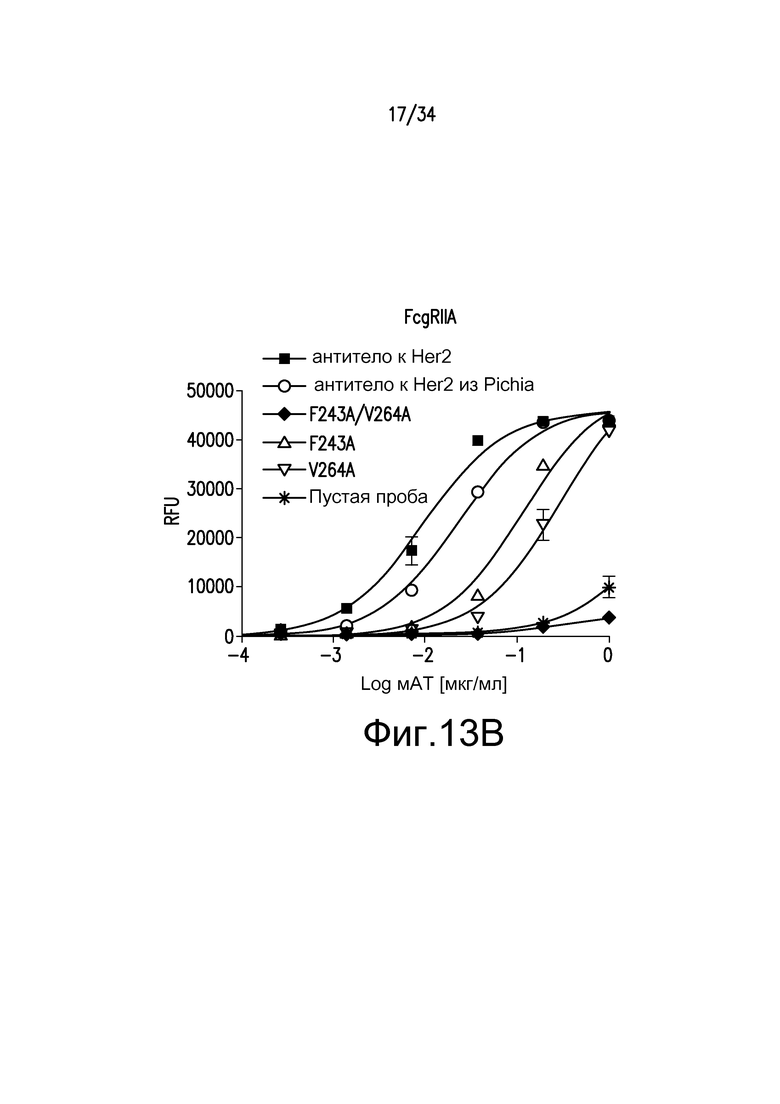
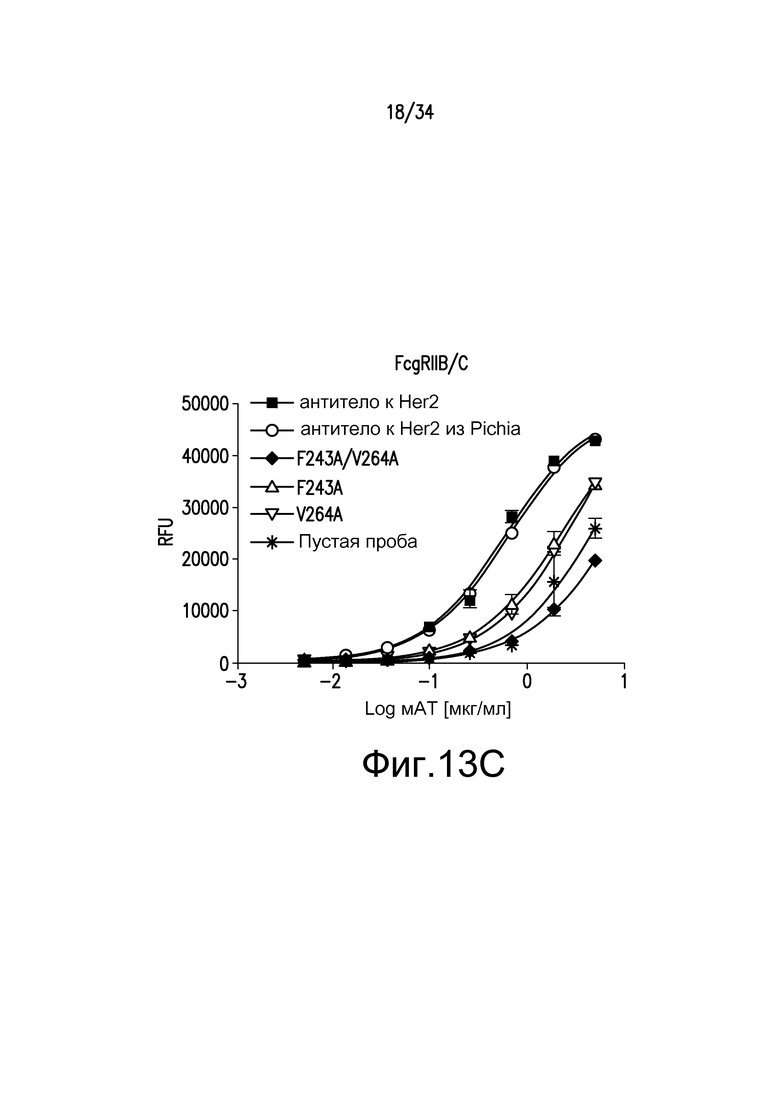
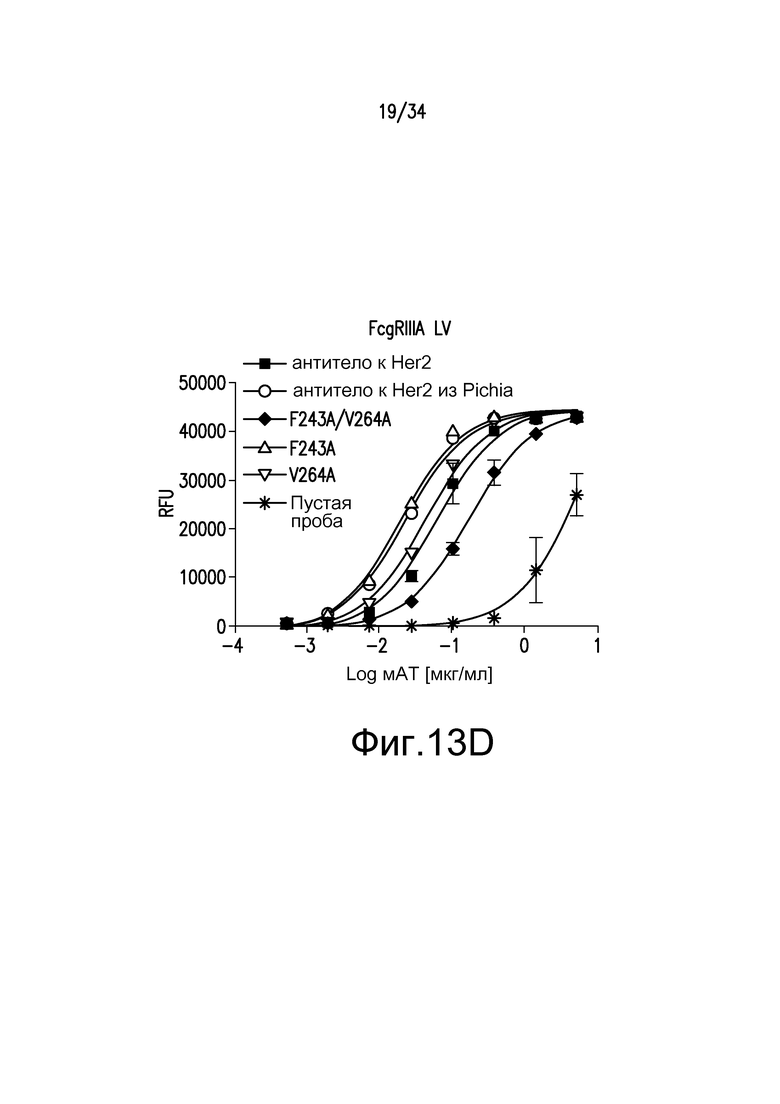
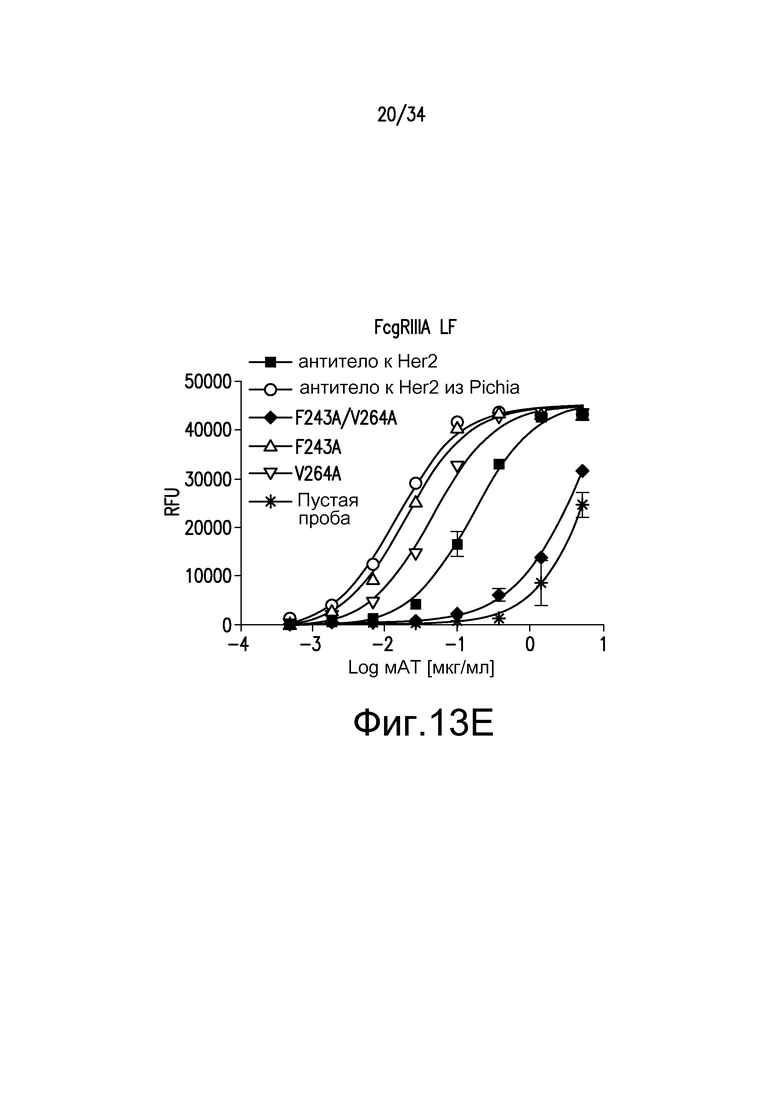
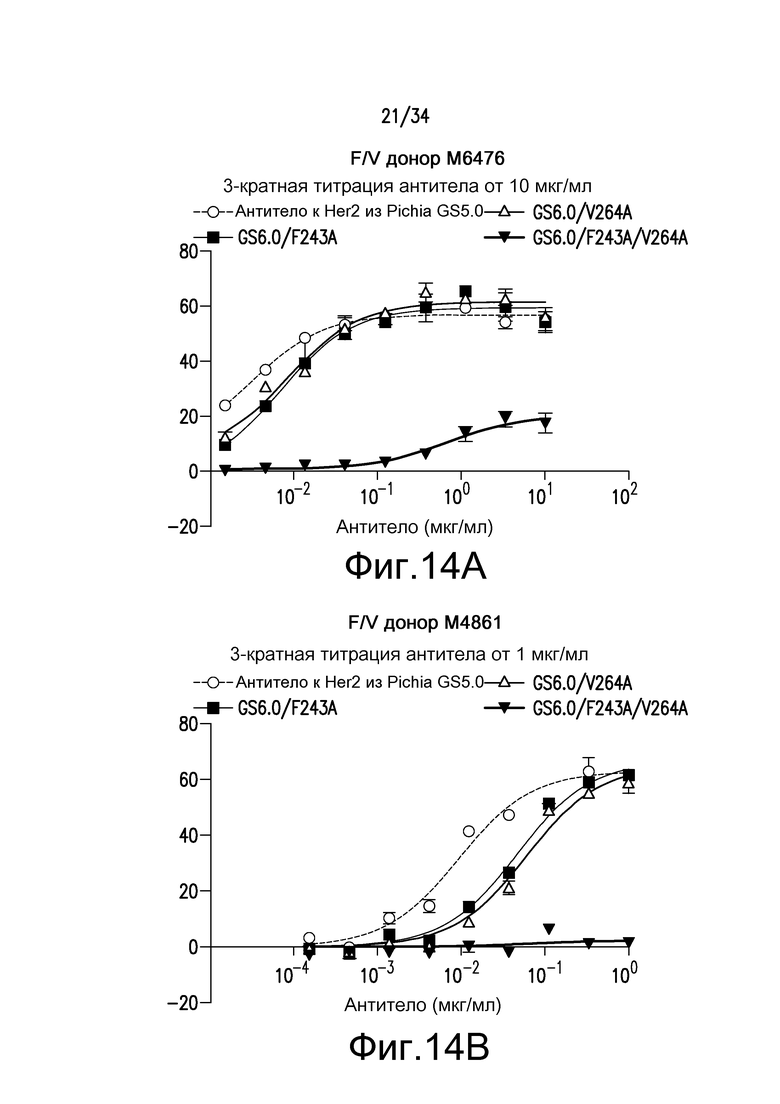
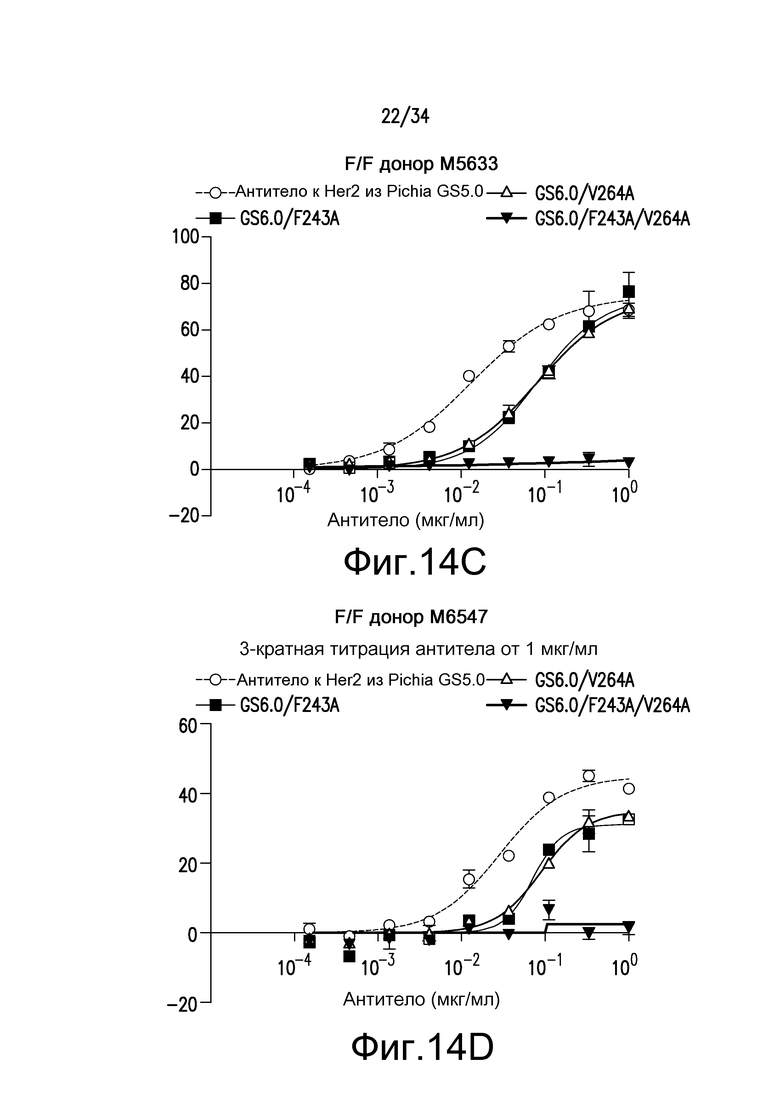
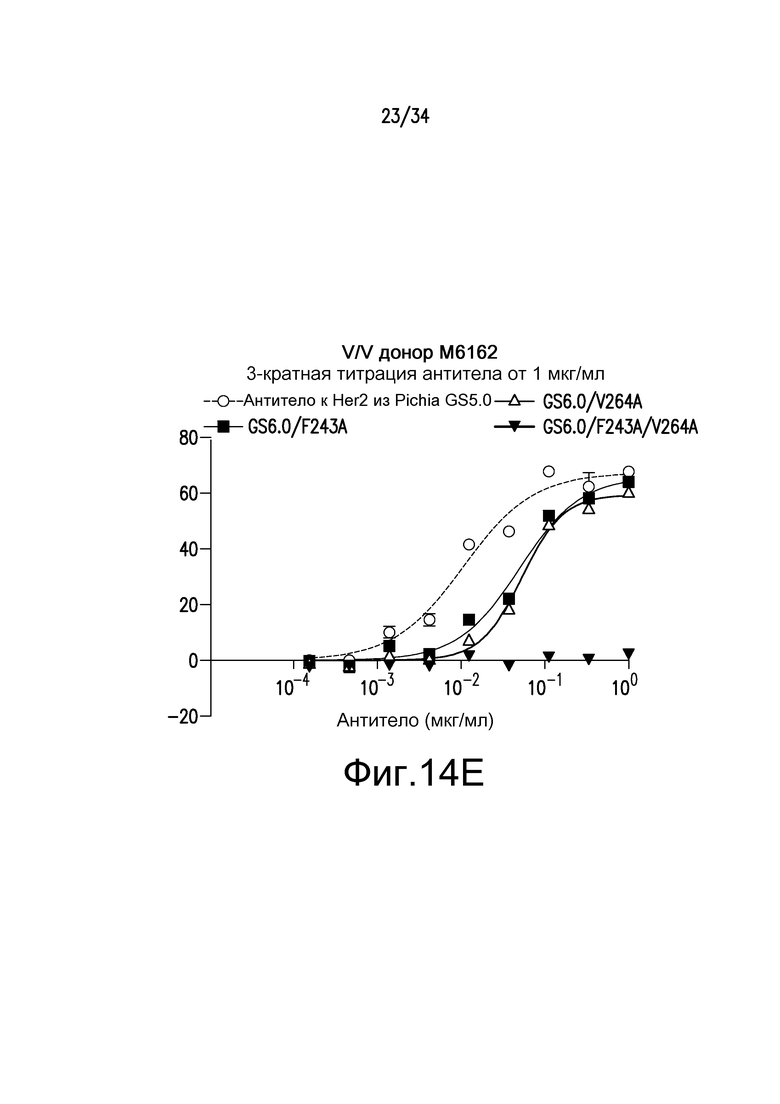
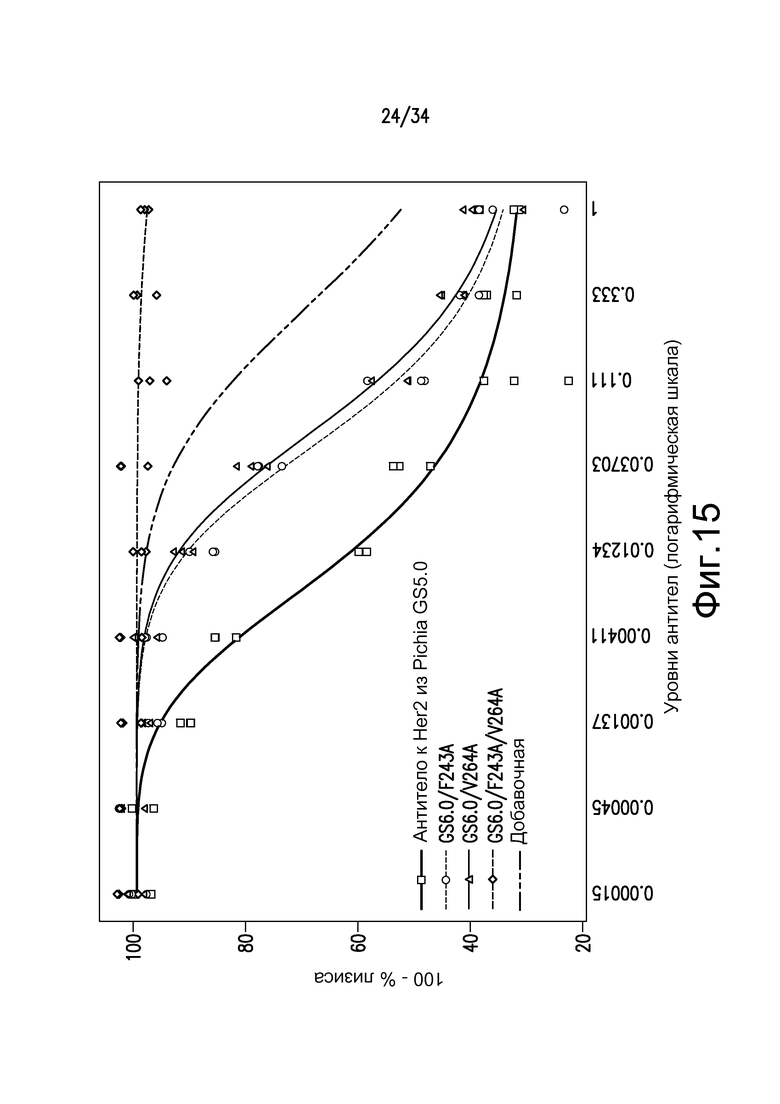
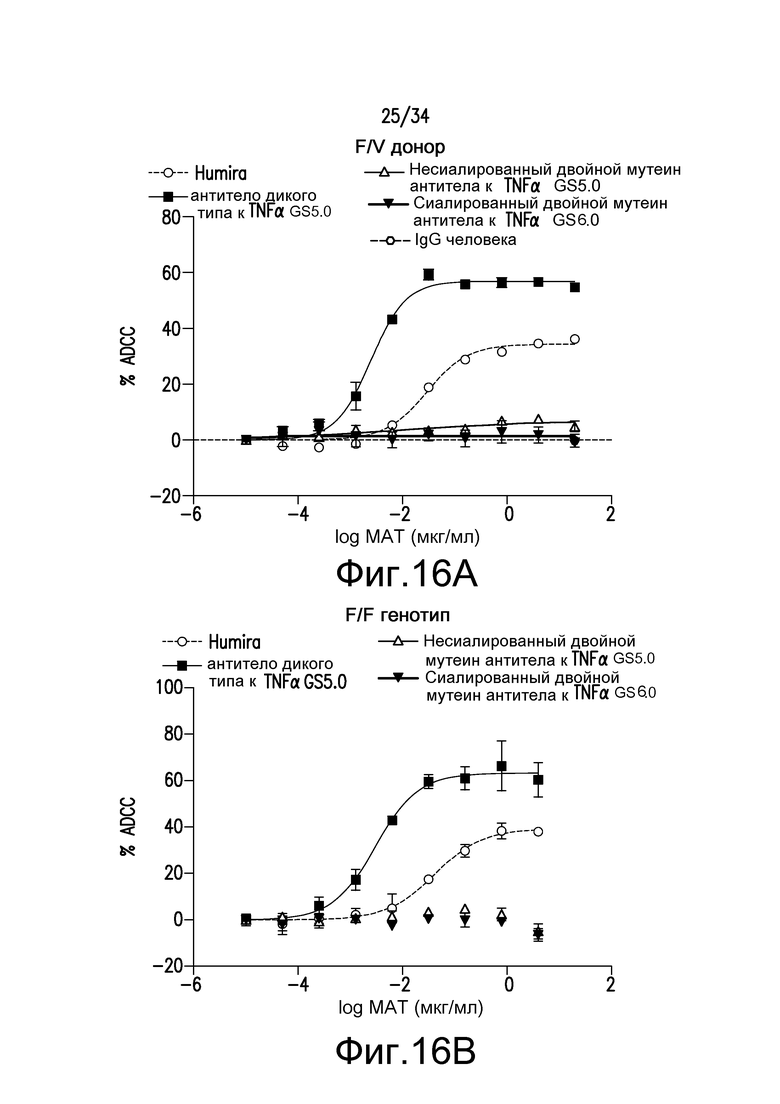
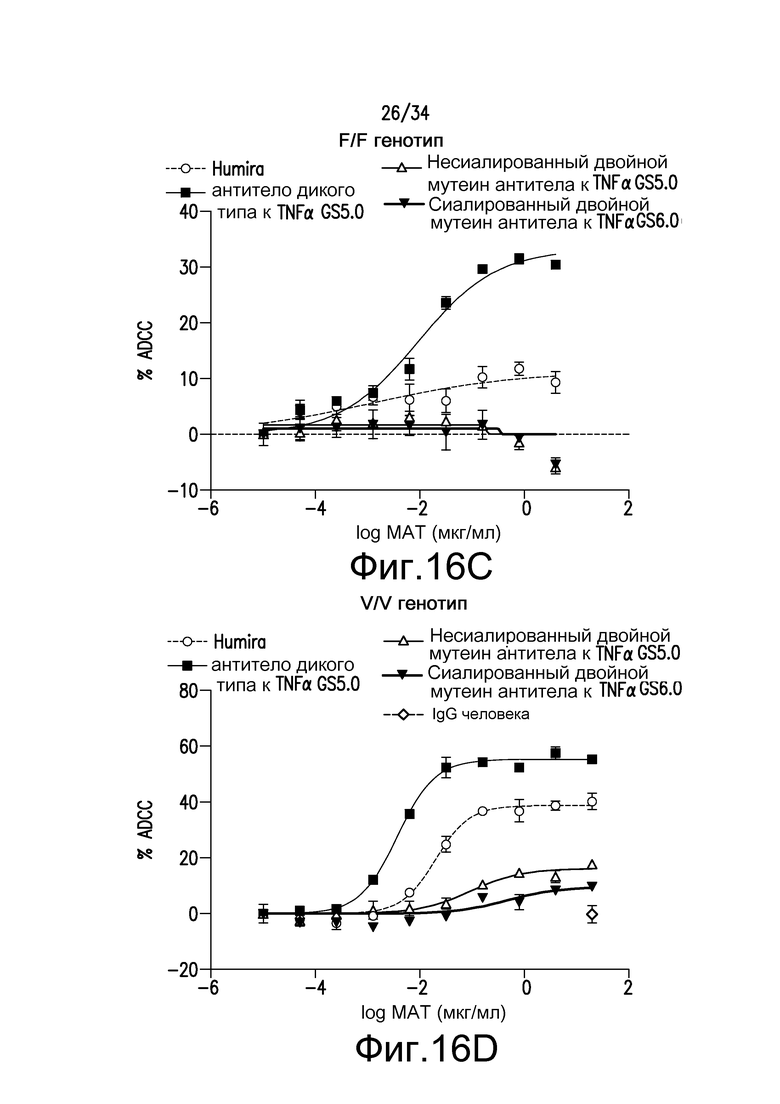
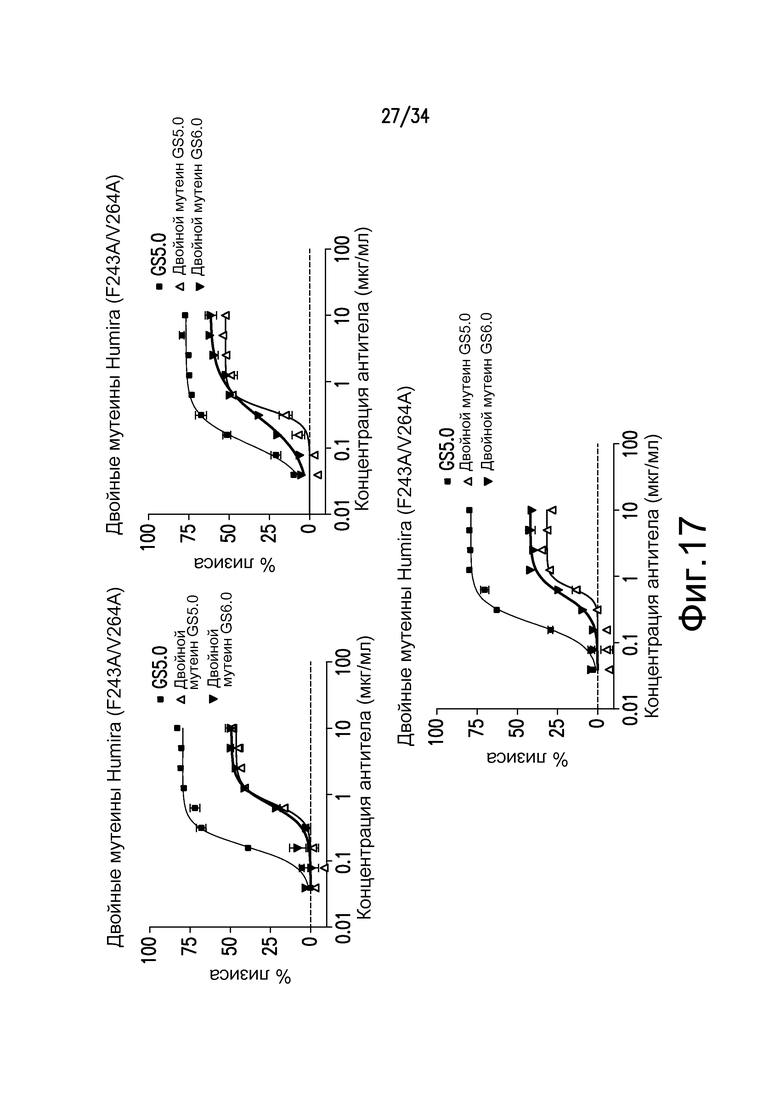
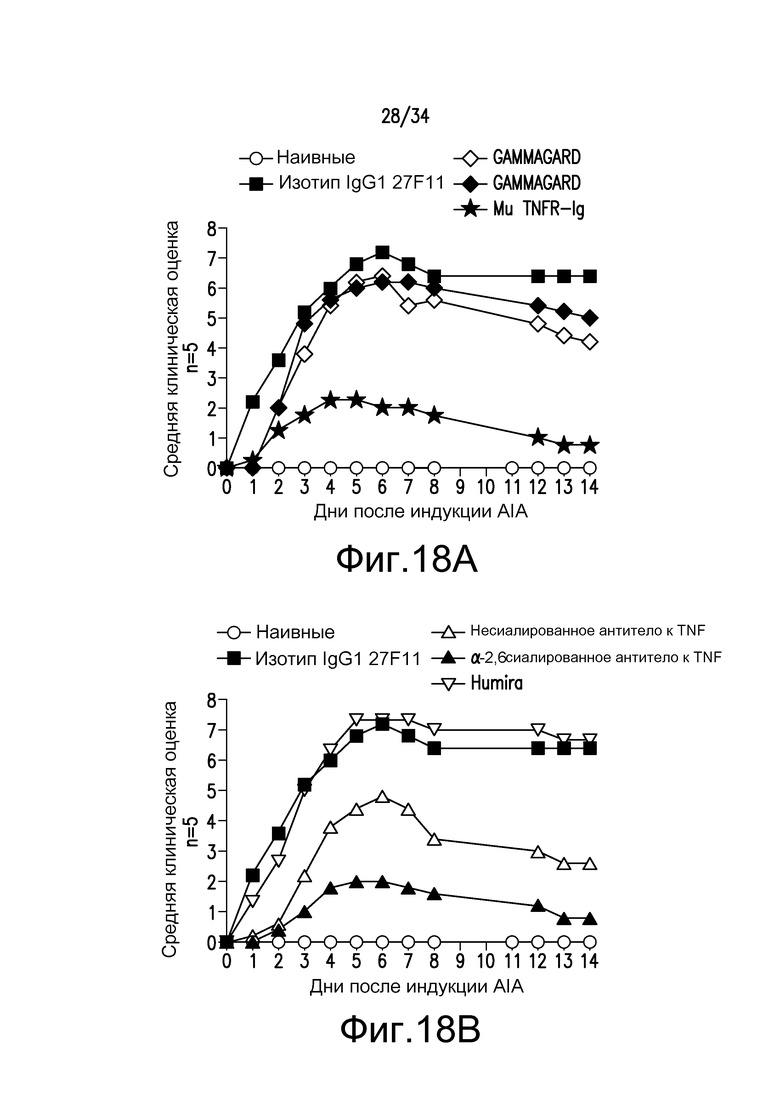
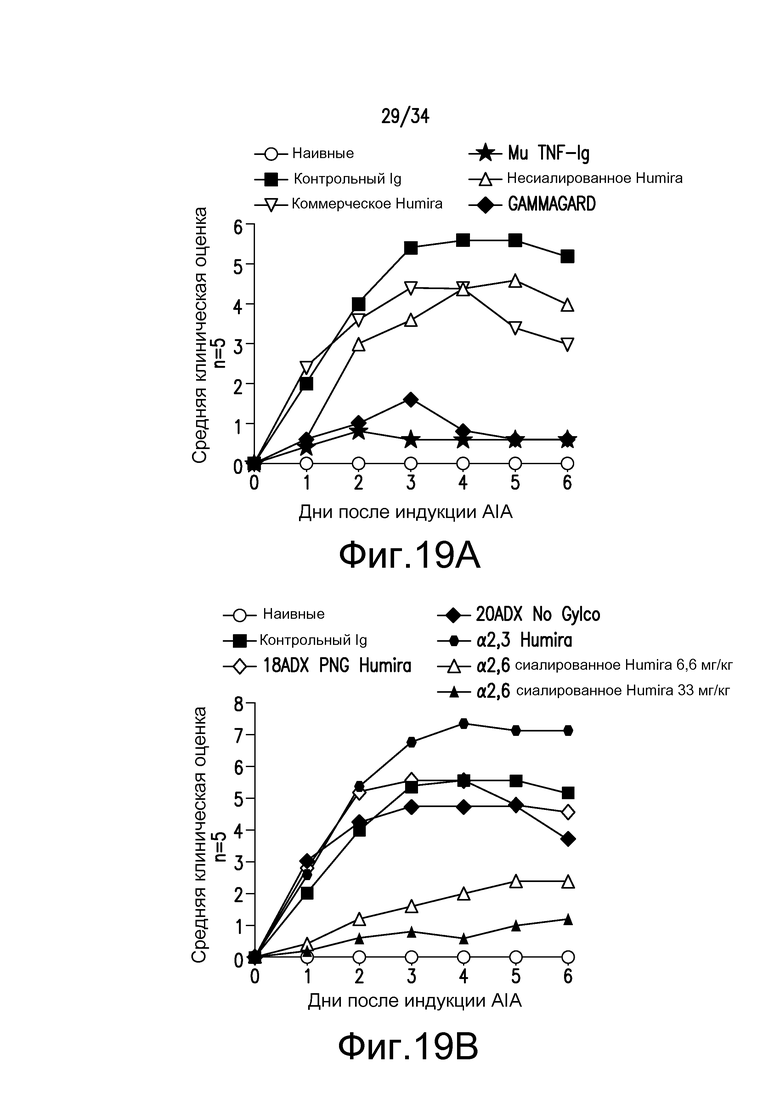
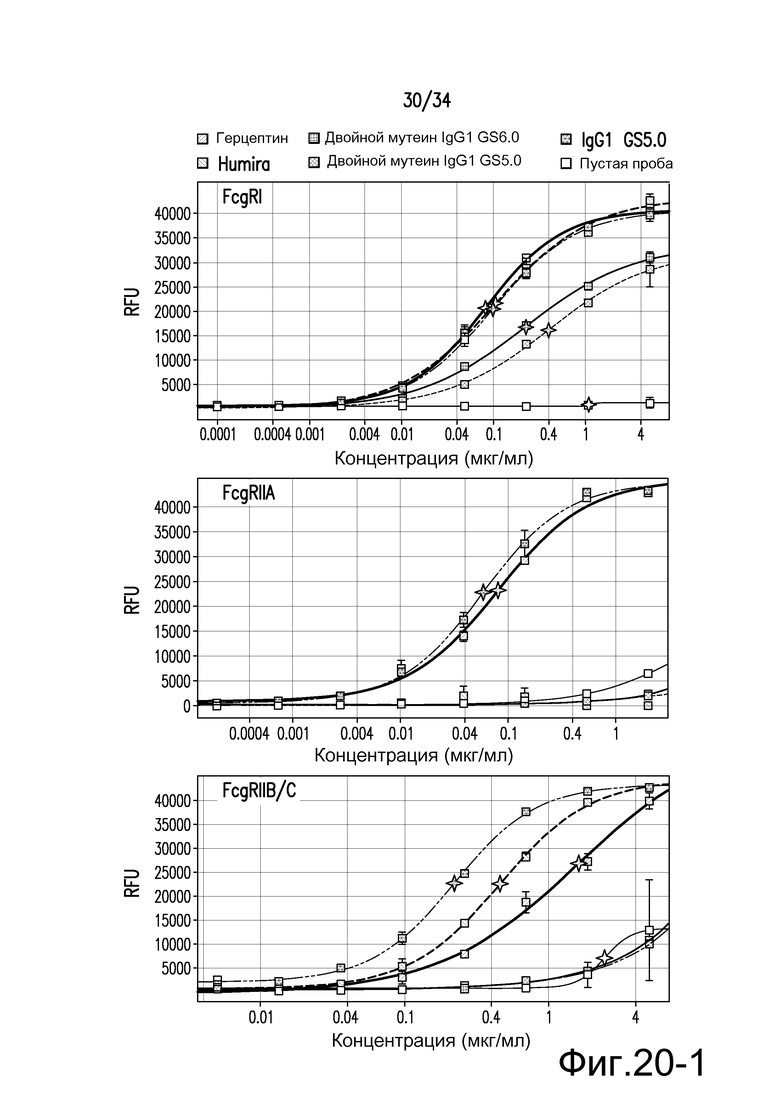
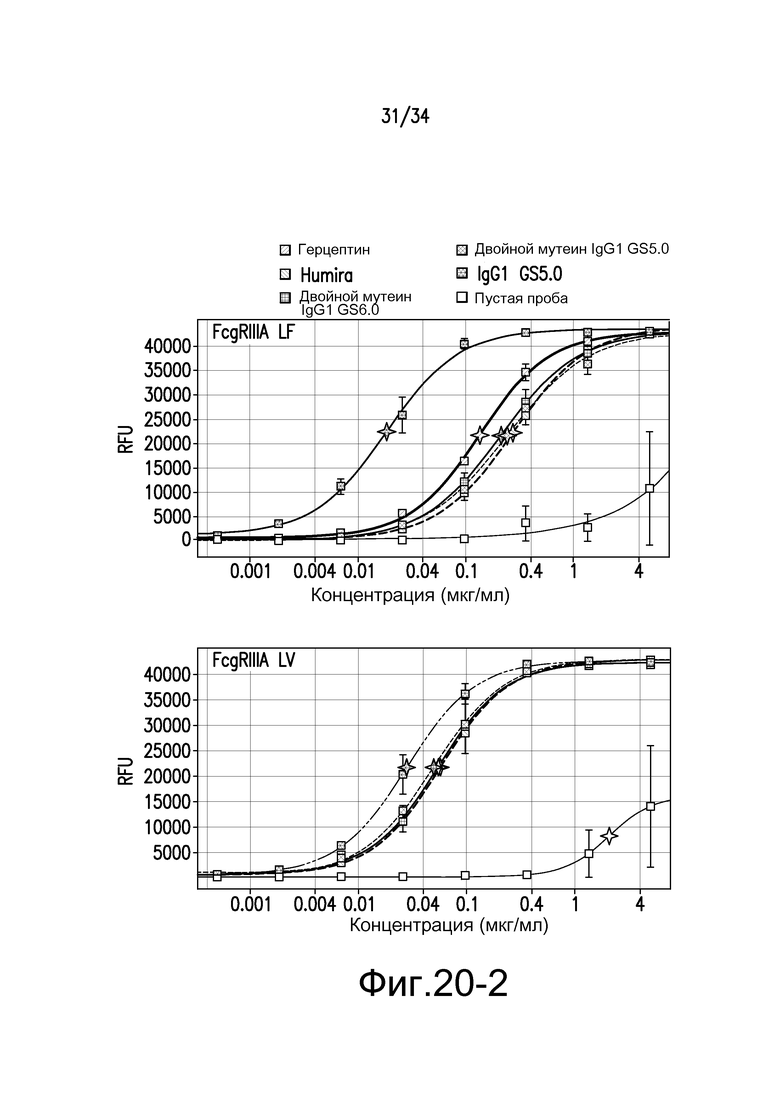
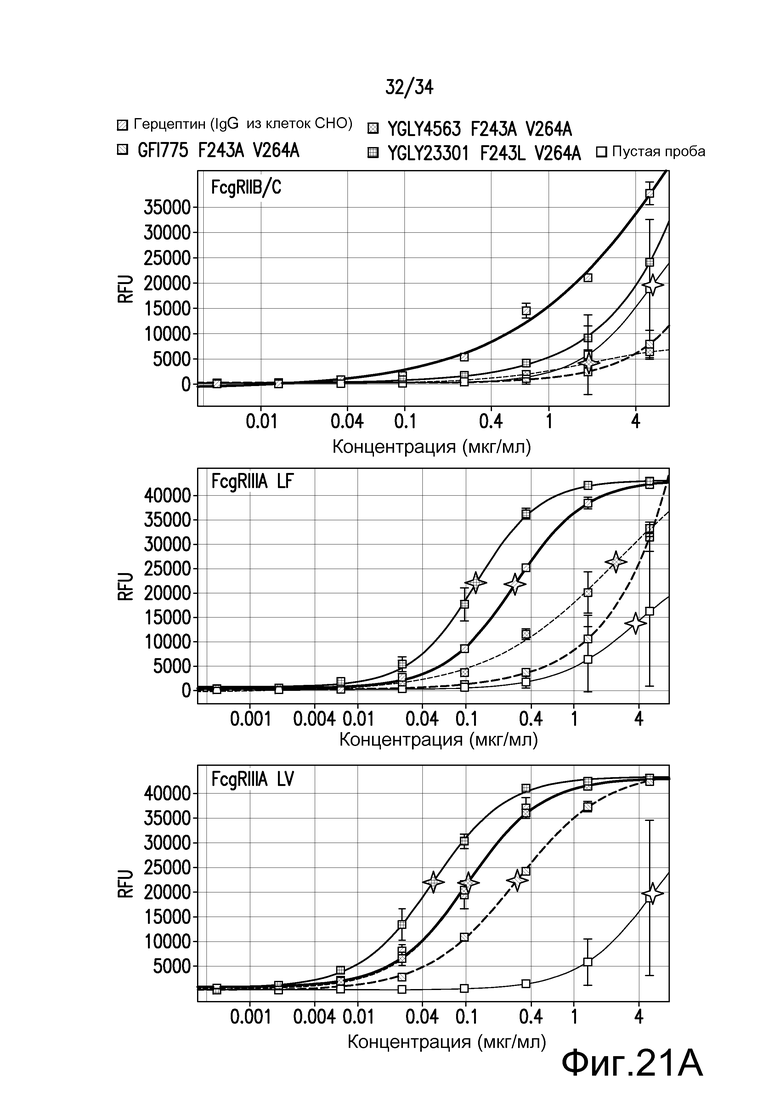
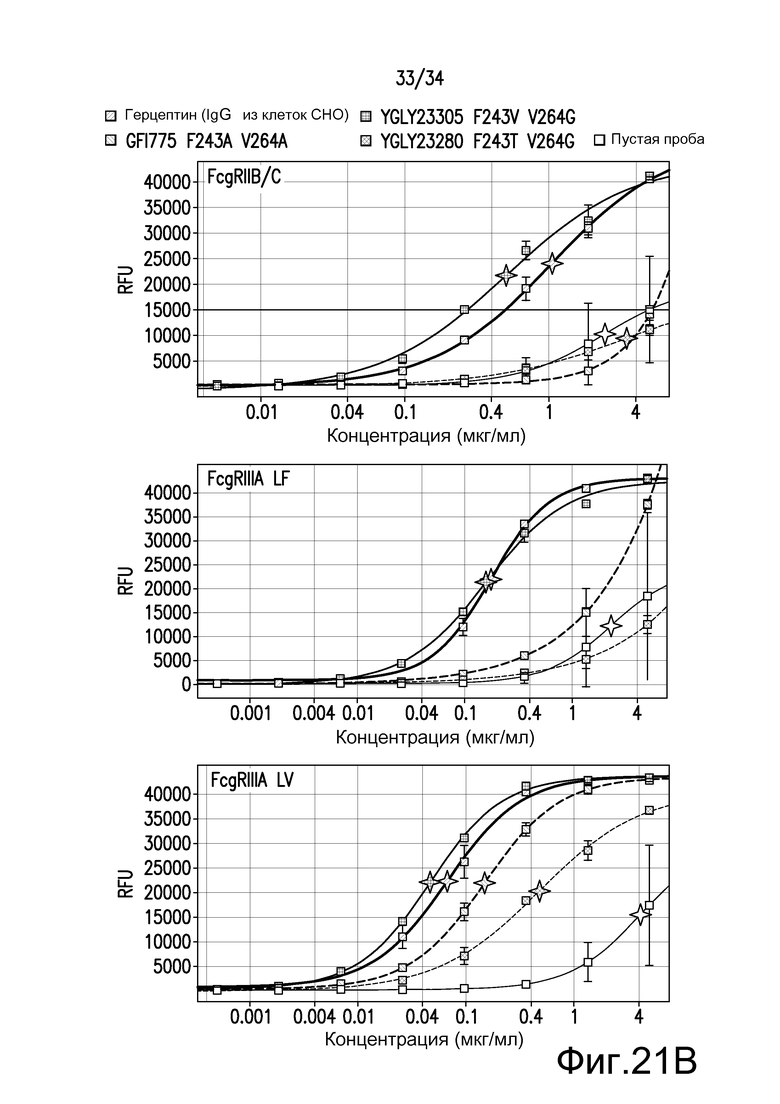
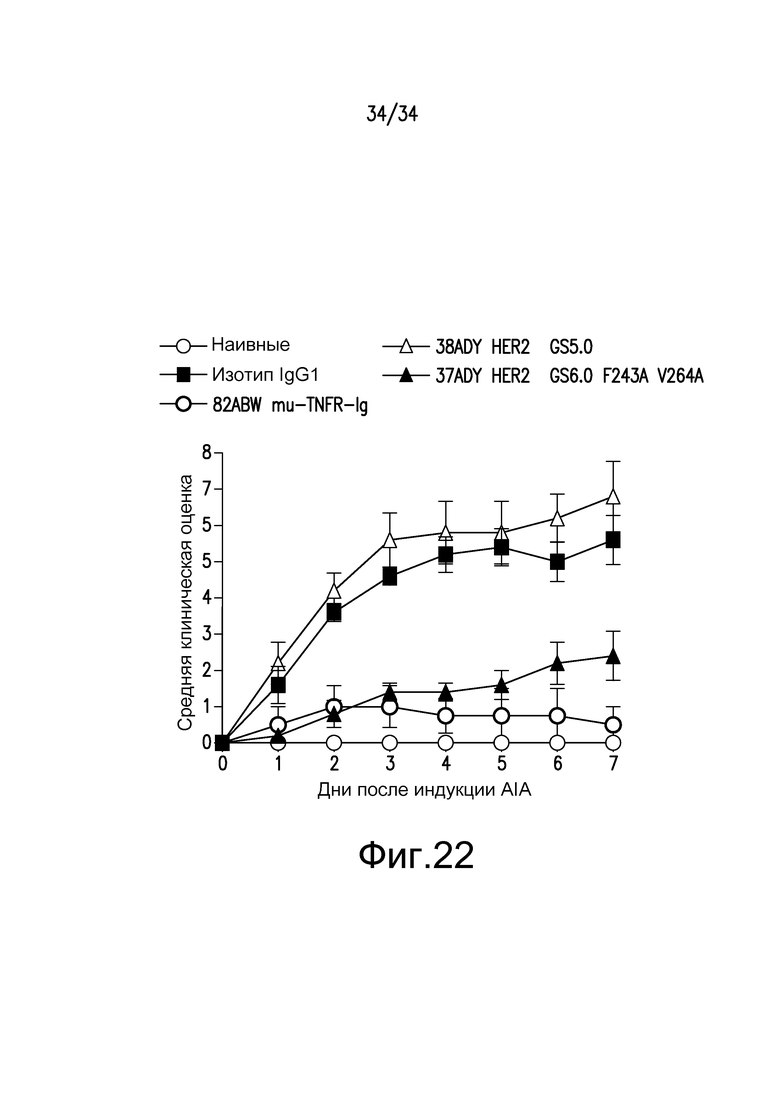
Изобретение относится к биотехнологии и представляет собой Fc-содержащий полипептид, содержащий аминокислотную последовательность SEQ ID NO: 18, где SEQ ID NO: 18 содержит мутации в аминокислотных положениях 243 и 264 Fc-области, где Fc-содержащий полипептид представляет собой антитело или фрагмент антитела, содержащий сиалированные N-гликаны, и где мутация в положении 243 представляет собой F243A и мутация в положении 264 представляет собой V264A, где нумерация произведена согласно индексу EU по Kabat, где Fc-содержащий полипептид обладает пониженным связыванием с FcγRI, FcγRIIa, FcγRIIb и/или FcγRIIIa по сравнению с родительским Fc-содержащим полипептидом. Изобретение относится также к способу получения указанного Fc-содержащего полипептида в клетке-хозяине, а также к способу лечения воспалительного состояния у нуждающегося в этом индивидуума, включающему введение индивидууму терапевтически эффективного количества указанного Fc-содержащего полипептида. Изобретение позволяет получать Fc-содержащий полипептид, обладающий пониженным связыванием с FcγRI, FcγRIIa, FcγRIIb и/или FcγRIIIa по сравнению с родительским Fc-содержащим полипептидом. 3 н. и 10 з.п. ф-лы, 22 ил., 14 табл., 24 пр.
1. Fc-содержащий полипептид, содержащий аминокислотную последовательность SEQ ID NO: 18, где SEQ ID NO: 18 содержит мутации в аминокислотных положениях 243 и 264 Fc-области, где Fc-содержащий полипептид представляет собой антитело или фрагмент антитела, содержащий сиалированные N-гликаны, и где мутация в положении 243 представляет собой F243A и мутация в положении 264 представляет собой V264A, где нумерация произведена согласно индексу EU по Kabat, где Fc-содержащий полипептид обладает пониженным связыванием с FcγRI, FcγRIIa, FcγRIIb и/или FcγRIIIa по сравнению с родительским Fc-содержащим полипептидом.
2. Fc-содержащий полипептид по п. 1, где остатки сиаловой кислоты в сиалированных N-гликанах присоединены посредством α-2,6-связей.
3. Fc-содержащий полипептид по п. 1 или 2, где по меньшей мере 70 мол.% N-гликанов в Fc-содержащем полипептиде имеют структуру, выбранную из группы, состоящей из SA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2.
4. Fc-содержащий полипептид по п. 1 или 2, где по меньшей мере 90 мол.% N-гликанов в Fc-содержащем полипептиде имеют структуру, выбранную из группы, состоящей из SA(1-4)Gal(1-4) GlcNAc(2-4)Man3GlcNAc2.
5. Fc-содержащий полипептид по п. 1 или 2, где по меньшей мере 66 мол.% N-гликанов в Fc-содержащем полипептиде имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2.
6. Способ получения Fc-содержащего полипептида по п. 1 в клетке-хозяине, включающий:
a) обеспечение генетически модифицированной клетки-хозяина с помощью генетической инженерии для выработки Fc-содержащего полипептида, где Fc-содержащий полипептид представляет собой антитело или фрагмент антитела, содержащий сиалированные N-гликаны, где клетка-хозяин содержит нуклеиновую кислоту, кодирующую мутации в аминокислотных положениях 243 и 264 Fc-области Fc-содержащего полипептида, где мутация в положении 243 представляет собой F243A и мутация в положении 264 представляет собой V264A и где нумерация произведена согласно индексу EU по Kabat;
b) культивирование клетки-хозяина в условиях, приводящих к экспрессии Fc-содержащего полипептида; и
c) выделение Fc-содержащего полипептида из клетки-хозяина.
7. Способ по п. 6, где остатки сиаловой кислоты в сиалированных N-гликанах присоединены посредством α-2,6-связей.
8. Способ по любому из пп. 6, 7, где Fc-содержащий полипептид содержит N-гликановую композицию, в которой количество и процентное содержание общего числа сиалированных N-гликанов повышено по сравнению с родительским Fc-содержащим полипептидом.
9. Способ по любому из пп. 6, 7, где по меньшей мере 70 мол.% N-гликанов в Fc-содержащем полипептиде имеют структуру, выбранную из группы, состоящей из SA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2.
10. Способ по любому из пп. 6, 7, где по меньшей мере 90 мол.% N-гликанов в Fc-содержащем полипептиде имеют структуру, выбранную из группы, состоящей из SA(1-4)Gal(1-4)GlcNAc(2-4)Man3GlcNAc2.
11. Способ по любому из пп. 6, 7, где по меньшей мере 66 мол.% N-гликанов в Fc-содержащем полипептиде имеют структуру NANA2Gal2GlcNAc2Man3GlcNAc2.
12. Способ по любому из пп. 6, 7, где Fc-содержащий полипептид обладает пониженным связыванием с FcγRI, FcγRIIa, FcγRIIb и/или FcγRIIIa по сравнению с родительским Fc-содержащим полипептидом.
13. Способ лечения воспалительного состояния у нуждающегося в этом индивидуума, включающий введение индивидууму терапевтически эффективного количества Fc-содержащего полипептида по любому из пп. 1-5.
| US20050054832 A1, 10.03.2010 | |||
| ОПТИМИЗИРОВАННЫЕ Fc-ВАРИАНТЫ, ИМЕЮЩИЕ ИЗМЕНЕННОЕ СВЯЗЫВАНИЕ С FcγR, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2004 |
|
RU2337107C2 |
| BAUDINO et al., "Crucial role of aspartic acid at position 265 in the CH2 domain for murine IgG2a and IgG2b Fc-associated effector functions", J.Immunol | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Водяные лыжи | 1919 |
|
SU181A1 |
| JASSAL et al., "Sialylation of human IgG-Fc carbohydrate by | |||

