Предпосылки создания изобретения
Область техники, к которой относится изобретение
Изобретение относится к способам получения лечебных белков, взаимодействующих с Fc-рецепторами, например антител, в которых состав олигосахаридных цепей оптимизирован для авидности антител в отношении своей мишени, а также аффиности связывания Fc-рецепторов, причем посредством этого оптимизируется эффекторная функциональная активность указанных антител по сравнению с неоптимизированными методами получения гликозилированных антител.
Описание родственных технических решений
Антитела представляют собой растворимые сывороточные гликопротеины, которые играют существенную роль в наследственном иммунитете. Углеводная структура всех продуцируемых в природе антител в консервативных позициях в константных областях тяжелой цепи изменяется с изотипом. Каждый изотип обладает определенным порядком N-соединенных олигосахаридных структур, который вариабельно влияет на сборку белка, секрецию или функциональную активность (Wright A. and Morrison S.L., Trends Biotech., 15: 26-32 (1997). При обращении к фиг.1 и 2 видно, что структура присоединенных N-соединенных олигосахаридов существенно изменяется в зависимости от степени процессинга и может включать много маннозных звеньев, а также комплекс биантеннальных олигосахаридов с или без рассекающего GlcNAc и коровых остатков фукозы (Wright A. and Morrison S.L., цит. выше). Типично происходит гетерогенный процессинг коровых олигосахаридных структур, связанный с определенным сайтом гликозилирования, так что даже моноклональные антитела существуют в виде многих гликоформ. Подобным образом, показано, что основные различия в гликозилировании антител имеют место между клеточными линиями, продуцирующими антитела, и даже видны небольшие различия для данной клеточной линии, растущей в различных условиях культивирования.
Известно, что сиаловая кислота в гликанах (статические группы) важна для пролонгирования времени полужизни в сыворотке гликопротеинов иных, чем антитела (Stockert R.J. (1995), Physiol. Rev., 75, 591-609). До сих пор роль сиаловой кислоты в моноклональных антителах (Mab) непонятна. Время полужизни Mab в сыворотке является особенно длительным, и конструирование Fc-слитых белков дает применимую стратегию при разработке лечебных белков, например белкового энтерацептика (protein enteracept).
Антитела и молекулы Т-клеточных рецепторов обладают участками, которые ответственны за специфическое связывание с клеточной поверхностью, которое модулирует клеточную реакцию. В иммунной системе такие функции классифицируют как гуморальные и клеточные. Антитела часто относят к адаптерным молекулам, связывающим гуморальный и клеточный иммунные механизмы: гуморальные реакции связывают, главным образом, с зрелыми секретированными циркулирующими антителами, способными к связыванию с высокой аффинностью с антигеном-мишенью. Клеточные реакции связывают с результатами клеточной активации за счет связывания комплексов Аг-Ат (ab-ag) и за счет последствий, вызванных высвобождением клеточных медиаторов в результате связывания комплекса Аг-Ат с эффекторными клетками. Такие клеточные реакции включают нейтрализацию мишени, опсонизацию и сенсибилизацию (если антиген отображается на поверхности клетки), сенсибилизацию тучных клеток и активацию комплемента. Для клеточных мишеней, т.е. антигенов клеточной поверхности, такие эффекторные функции ведут к тому, что обычно известно как антителозависимая клеточная цитотоксичность (ADCC) и комплементзависимая цитотоксичность (CDC).
Среди изотипов антител (например, IgE, IgD, IgA, IgM и IgG) IgG являются наиболее распространенными с подклассами IgG1, обнаруживающими наибольшие степень и ряд эффекторных функций. Антитела типа IgG1 являются наиболее используемыми антителами в иммунотерапии рака, где активность ADCC и CDC часто считается важной. Структурно-шарнирная область IgG и домены СН2 играют важную роль в эффекторных функциях антител. Присутствующие в Fc-области N-соединенные олигосахариды влияют на эффекторные функции. Ковалентно-связанные олигосахариды являются комплексными структурами биантенниального типа и являются высокогетерогенными (см. фиг.1 и 2). Консервативный N-связанный сайт гликозилирования в Asn297 размещен в каждом домене СН2. В зрелом антителе два комплексных биантенниальных олигосахарида скрываются между доменами СН2, образуя экстенсивные контакты с главной цепью полипептида. Обнаружено, что их присутствие существенно для антител, опосредующих эффекторные функции, такие как ADCC (Lifely M.R. et al., Glycobiology, 5: 813-822 (1995); Jefferis R. et al., Immunol. Rev., 163: 59-76 (1998); Wright A. and Morrison S.L., цит. выше).
Гетерогенные олигосахариды, декорирующие антитело Fc-части, или образованные антителами структуры, включая продуцированные различными клетками-хозяевами, содержат преимущественно остатки сиаловой кислоты, фукозы, галактозы и GlcNA как концевые сахара (Raju T.S. et al., Glycobiology, 2000, 10(5): 477-86). Показано, что некоторые такие концевые сахара, в частности, подвергнутые воздействию галактозы, остатки коровой фукозы и рассекающего GlcNA, влияют на структуру Fc-части молекулы и посредством этого изменяют эффекторные функции антител. Эффекторные функции, такие как активность ADCC и активность CDC, которые опираются на связывание с рецепторами клеточной поверхности, известными как Fc-рецепторы, а также на связывание с различными лигандами, включая комплементный белок C1q, могут быть изменены из-за состава присоединенного гликана (Presta L., 2003, Curr. Opin. Struct. Biol., 13(4): 519-25). Большинство N-соединенных гликанов, присоединенных к Fc, не являются сиалилированными в значительной степени (Idusogie E.E. et al., 2000, J. Immunol., 15: 164(8): 4178-84).
Основные структуры, обнаруженные в человеческом IgG и других полученных рекомбинантно IgG, представляют собой комплексные биантенниальные структуры с подвергнутыми воздействию остатками Gal или без них (фиг.1). Существует ряд клеток-хозяев млекопитающих, которые в настоящее время используются для экспрессии рекомбинантных антител для исследовательских целей, а также получения биофармацевтических препаратов. Виды клеток-хозяев, в зависимоти от происхождения, а также условий культивирования могут быть причиной изменения степени и структуры гликанов, присоединенных к рекомбинантно экспрессированным молекулам. Двумя обычно используемыми клеточными линиями для рекомбинантной экспрессии антител являются клетки яичников китайского хомячка (СНО) и клетки миеломы мыши (sp2/0, 653, Ns0). Хотя клетки СНО экспрессируют рекомбинантные антитела, которые фактически лишены гликана сиаловой кислоты, гликаны на 99% фукозилированы. Показано, что присутствие фукозы является основным фактором, вносящим вклад в восстановленный рецептор Fc-гамма-III и, следовательно, в ADCC. Клетки миеломы мыши экспрессируют рекомбинантные антитела с содержанием сиаловой кислоты до 50%, но, как правило, с меньшим содержанием фукозы. Как установлено выше, такие различия могут оказывать существенное влияние на активность антител in vivo.
Поэтому желательна возможность уменьшения сиалилирования гликанов, ассоциированных с лечебными антителами, способом, который устраняет необходимость процессинга после сбора и в то же время обеспечивает разумно гомогенную структуру в отношении содержания сиаловой кислоты.
Сущность изобретения
Настоящее изобретение относится к способам, линиям клеток-хозяев и экспрессирующим векторам и плазмидам, применимым для получения Fc-содержащих молекул, в частности лечебных средств на основе антител, с уменьшенным содержанием сиаловой кислоты. Конкретнее, изобретение относится к экспрессирующей плазмиде, кодирующей созданную инженерией сиалидазную кодирующую последовательность, и указанная плазмида, однажды включенная в секретирующую антитела клеточную линию, вызывает способность клетки-хозяина секретировать полипептид, обладающий сиалидазной активностью. В одном воплощении кодирующая последовательность в плазмиде кодирует каталитический домен сиалидазы Arthrobacter ureafaciences. В другом аспекте изобретения клетка-хозяин, включающая каталитический домен сиалидазы Arthrobacter ureafaciences, секретирует транслированный каталитический домен в культуральную среду.
Настоящее изобретение относится к способу регулирования свойств Fc-содержащей молекулы, включая минимизированное сиалилирование олигосахаридов, соединенных с Fc-участком, посредством чего оптимизируются авидность молекулы в отношении множества локализованных белков-мишеней и аффинность в отношении к одному или нескольким рецепторам Fc-гамма, например, FcγRI, FcγRIIA и FcγRIIIA; активность ADCC; макрофаговая или моноцитарная активация и время полужизни в сыворотке.
Изобретение также относится к получению высокогомогенных партий Fc-содержащих молекул, таких как антитела, содержащих максимально сиалилированные N-соединенные олигосахариды в Fc-домене. Изобретение также относится к очистке партий антител, обогащенных антителами, содержащими сиаловую кислоту в Fc-олигосахариде, а также антителами, не содержащими сиаловой кислоты в Fc-олигосахариде.
Краткое описание чертежей
Фиг.1 представляет собой схематическое изображение самой крупной структуры олигосахаридов, обнаруженной в человеческом IgG.
Фиг.2 отображает основные структуры олигосахаридов, обнаруженные в рекомбинантном IgG, продуцированном в клетках яичника китайского хомячка (СНО).
Фиг.3 показывает результаты анализов ВЭЖХ Fc-олигосахаридов. Сначала N-соединенные олигосахариды высвобождают из антител обработкой ферментом PNGase F. Высвобожденные олигосахариды метят антраниловой кислотой и меченые олигосахариды очищают гель-фильтрацией. Очищенные меченые олигосахариды анализируют ВЭЖХ, получая приведенную хроматограмму.
Фиг.4А и 4В представляют собой графики, показывающие связывание различных иммунных комплексов Ab1:TNF с человеческим FcγRII на клетках К562 двумя различными путями. (А) Конкурентное связывание, измеренное добавлением различных количеств немеченых комплексов Ab1 и TNF к клеткам в присутствии фиксированного количества Ab5 - меченого 125I человеческого IgG в комплексе с Ab6 - мышиными моноклональными антителами, специфичными для Ab5. (В) Прямое связывание, измеренное добавлением к клеткам К562 различных количеств Ab1 в комплексе с меченым 125I TNF.
Фиг.5, А-D, представляют собой графики исследований связывания FcγRIIIA с различными препаратами испытываемых Ab, используемых для конкурентных меченых радиоизотопами mAb против FcγRIIIA 3G8 при фиксированной концентрации, для связывания NK-клеточных FcγRIIIA: варианты природного гликозилирования Ab1 (A); варианты природного гликозилирования Ab5 (B); фракции Ab1 из колонки с лектином (С); и фракции Ab2 из колонки с лектином (D).
Фиг.6, А-D, представляют собой графики, показывающие результаты анализов ADCC in vitro, выполненных с использованием Ab1, различающихся содержанием сиаловой кислоты, клеток-мишеней К2, сверхэкспрессирующих TNF на своей клеточной поверхности, и человеческих эффекторных клеток РВМС, экспрессирующих FcγR. (А) Варианты природного гликозилирования Ab1, (В) варианты природного гликозилирования Ab2; (С) сравнение трех субпартий Ab1, различающихся содержанием сиаловой кислоты, после фракционирования на основе аффинности лектина к WGA, и ферментативно дегликозилированных (Gno) Ab1, (D) сравнение образца необработанных Ab1 и образца полностью сиалилированных Ab1 G2S2 или отрицательного контроля Ab, соответствующего изотипу Ab7. Образцы анализируют при трехкратном повторе (величина ошибки представлена ср.-кв. откл. (s.d.)), и приведенные результаты представляют три независимых эксперимента для каждой пары вариантов. Различие в активности между указанными испытываемыми образцами является значимым (P<0,0001 для графиков А, С и D; P=0,0016 для графика В), что определяют по F-критерию дополнительной суммы квадратов (extra sum of squares F-test).
Фиг.7, А и В, представляют собой графики, показывающие конкурентное связывание различных образцов антител IgG с человеческим рецептором FcγRI (CD64) на клетках U-937: (A) Ab1 G2 (полностью гликозилированные и несиалилированные) и Ab1 G2S2(hi) (полностью гликозилированные и полностью сиалилированные), различающихся только по отсутствию и присутствию сиаловой кислоты, (В) двух различных партий Ab3, различающихся по количеству загруженных видов олигосахаридов (видов, содержащих сиаловую кислоту), составляющих или 2% или 42% от всех олигосахаридов.
Фиг.8 представляет собой график, показывающий соотношение между временем после введения и сывороточной концентрацией Fc-части слитого белка (FcP1), который полностью сиалилирован (G2S2) или немодифицирован.
Фиг.9 представляет собой график, показывающий соотношение между временем после введения и сывороточной концентрацией Fc-части полностью сиалалированных Ab2 G2S2 или полностью асиалалированных Ab2 G2 описанными ферментативными способами.
Фиг.10, A-D, представляют собой графики, показывающие влияние сиаловой кислоты в препаратах Ab на аффинность в отношении лиганда-мишени на клеточной поверхности посредством конкурентного связывания с радиомечеными Ab: (A) природные варианты Ab1, (В) природные варианты Ab5, (С) варианты фракций Ab1 из колонки с лектином и (D) варианты фракций Ab2 из колонки с лектином. Образцы испытывают при двукратном повторе или четырехкратном повторе, и показанные результаты представляют 3 или 4 независимых эксперимента. Различие в связывании между указанными испытываемыми образцами является значимым (P<0,0001 для графиков А, С и D), что определяют по F-критерию дополнительной суммы квадратов.
Фиг.11, А-В, представляют собой графики, показывающие влияние сиаловой кислоты в препаратах Ab на аффинность в отношении лиганда-мишени, нанесенного на пластины EIA: (A) связывание природных вариантов Ab1 с TNF, (В) связывание Ab2 с анти-Id антителами.
Фиг.12, А-С, представляют собой графики, показывающие влияние сиаловой кислоты в препаратах Ab на аффинность в отношении лиганда-мишени, представленного в виде меченого радиоизотопом растворимого антигена к связанным с поверхностью Ab: (A) природные варианты Ab1, (В) варианты фракций Ab1 из колонки с лектином и (С) варианты фракций Ab2 из колонки с лектином. Осуществляют параллельные инкубации с меченым радиоизотопом Ag и 100-кратным избытком немеченого Ag для определения неспецифического связывания. Образцы испытывают при троехкратном повторе.
Фиг.13 является схематическим представлением экспрессирующей плазмиды р3629, сконструированной для экспрессии каталитического домена сиалидазы А из Arthrobacter ureafaciences, соединенного с сигнальной последовательностью hGH (человеческий фактор роста), с сайтами рестриктаз, используемыми для клонирования в исходный указанный вектор р2815.
Фиг.14 представляет собой график, представляющий антителозависимую клеточную цитотоксичность (ADCC) очищенных антител из клеточных линий, эксперессирующих секретированный каталитический домен сиалидазы; клоны 3, 5, 12, 13 и 17.
Подробное описание изобретения
Аббревиатуры
α1,3GT - α-1,3-галактозилтрансфераза; α2,3ST - α-2,3-сиалилтрансфераза; β1,4GT - β-1,4-галактозилтрансфераза; ADCC - антителозависимая клеточная цитотоксичность; АТСС - Американская коллекция типовых культур; BATDA - бис(ацетоксиметил)-2,2':6',2”-терпиридин-y,y”-дикарбоксилат; BSA - бычий сывороточный альбумин; CD-среда - химически определенная культуральная среда; CDC - комплементнаправленная цитотоксичность; CMP-Sia - цитидинмонофосфат-N-ацетилнейраминовая кислота; DMEM - модифицированная по Дульбекко среда Игла; Е:Т - отношение эффекторных клеток к клеткам-мишеням; FBS - сыворотка плода коровы; ESI-MS - масс-спектрометрия с ионизацией электронным распылением; NK-клетки - естественные клетки-киллеры; IgG - иммуноглобулин G; IMDM - среда Дульбекко, модифицированная по Искову; MALDI-TOF-MS - времяпролетная масс-спектрометрия с ионизацией с участием лазерной десорбции; МНХ - микофенольная кислота, гипоксантин, ксантин; NANA - N-ацетилнейраминовая кислота, изомерсиаловой кислоты; NGNA - N-гликолилнейраминовая кислота, изомерсиаловой кислоты; РВМС - одноядерные клетки периферической крови; РВМС - одноядерная клетка периферической крови; PBS - забуференный фосфатом физиологический раствор; PNGase F - пептид N-гликозидаза F; ОФ-ВЭЖХ (RP-HPLC) - высокоэффективная жидкостная хроматография с обращенной фазой; RT - комнатная температура; Sia - сиаловая кислота; UDP-Gal - уридиндифосфатгалактоза; UDP-GlcNAc - уридиндифосфат-N-ацетилглюкозамин.
Определения
Термин «активность ADCC» обозначает антителозависимую клеточную цитотоксичность и относится к явлению опосредуемого антителами разрушения клетки-мишени несенсибилизированными эффекторными клетками. Идентичность клетки-мишени изменяется, но она должна иметь связанный с поверхностью иммуноглобулин G, имеющий Fc-домен или часть Fc-домена, способную к активации Fc-рецептора. Это может быть, например, лимфоцит, лишенный обычных В- или Т-клеточных маркеров, или моноцит, макрофаг или многоядерный лейкоцит, в зависимости от идентичности клетки-мишени. Реакция является комплементнезависимой. Активность ADCC антител или другого Fc-содержащего белка по настоящему изобретению «усиливается», если его способность демонстрировать ADCC-опосредуемый киллинг клеток превышает способность антител или белка, по существу, с подобной последовательностью и Fc-доменом продуцироваться альтернативной клеткой-хозяином. Активность ADCC можно определить в стандартном анализе in vivo или in vitro киллинга клеток, таких как анализы, обсуждаемые в данном описании. Предпочтительно антитела с усиленной активностью добиваются такого же эффекта (предотвращения или ингибирования роста опухолевых клеток) при меньшей дозе и/или за более короткое время, чем ссылочные антитела, продуцируемые в альтернативной клетке-хозяине. Предпочтительно различие между возможностями антител в объеме настоящего изобретения и ссылочных антител составляет, по меньшей мере, примерно 1,5 раза, предпочтительнее, по меньшей мере, примерно 2 раза, даже предпочтительнее, по меньшей мере, примерно 3 раза, наиболее предпочтительно, по меньшей мере, примерно 5 раз, при определении, например, при сравнении рядом в выбранном стандартном анализе ADCC с высвобождением хрома.
Термин «аффинность», используемый в данном описании, предназначен для обозначения величины константы связывания простого одновалентного лиганда с его родственным партнером по связыванию, например, связывания Fab с антигеном или эпитопом. Аффинность можно измерить несколькими способами, включая измерение on- и off-rates (уровни связывания и уровни высвобождения kon и koff, соответственно), например, плазмонным резонансом (BiaCore), и выразить в виде константы общей ассоциации (Kass) или диссоциации (KD), где Kass представляет собой kon/koff, и KD представляет собой koff/kon. Kd также можно оценить эмпирически, например, измеряя концентрацию, при которой связывание лиганда с партнером по связыванию является полунасыщенным. Другим способом измерения KD является измерение конкурентным анализом, в котором один связующий агент или лиганд метят или превращают в мишень и выдерживают при постоянной концентрации, в то время как испытываемый связующий агент или лиганд добавляют в переменных концентрациях для устранения от конкуренции меченого вещества с его родственным партнером по связыванию и определения концентрации, при которой метка уменьшается вдвое.
Термин «авидность», используемый в данном описании, предназначен для оценки меры стремления лиганда оставаться связанным с партнером по связыванию, постольку поскольку как лиганд, так и партнер по связыванию может быть поливалентным, и стремление к нескольким событиям ассоциации и диссоциации для специфического лиганда может иметь место одновременно. Таким образом, авидность можно измерить по возрастанию кажущейся аффинности поливалентных конформаций партнера по связыванию с известной аффинностью.
Термин «Fc-содержащий белок» или «Fc-содержащая молекула», используемый в данном описании, относится к мономерному, димерному или гетеродимерному белку, имеющему лигандсвязывающий домен и, по меньшей мере, домен иммуноглобулина СН2 или СН3. Домены СН2 или СН3 могут образовывать, по меньшей мер, часть димерного участка белка/молекулы (например, антитела).
Предполагается, что термин «антитело» охватывает антитела, фрагменты расщепления, определенные белки и их варианты, в том числе, без ограничения, миметики антител или составляющие части антител, которые имитируют структуру и/или функцию антитела или определенных фрагментов или их частей, и сохраняют Fc-опосредуемые функции, в том числе, но без ограничения, связывание с Fc-рецепторами (например, FcγRI (CD64), FcγRIIA (CD32A), FcγRIIIA (CD16A) и FcRn), связывание комплемента (например, C1q), ADCC и CDC.
Термин «моноклональное антитело», используемый в данном описании, отражает специфическую форму Fc-содержащего слитого белка, в котором лигандсвязывающий домен остается, по существу, гомологичным, по меньшей мере, одному вариабельному домену тяжелой или легкой цепи антитела, по меньшей мере, одного вида антител животного.
Термин «эффекторные функции» антител или аналогов антител, используемый в данном описании, относится к процессам, посредством которых патогены или анормальные клетки, например, опухолевые клетки, разрушаются и удаляются из организма. Врожденные и адаптивные иммунные реакции используют большинство одних и тех же эффекторных механизмов для устранения патогенов, включая ADCC, CA (активацию комплемента), связывание C1q и опсинизацию.
Используемый в данном описании термин «клетка-хозяин» относится к любому виду клеточной системы, которую можно создать для получения белков, фрагментов белков или пептидов, представляющих интерес, в том числе, антител и фрагментов антител. Клетки-хозяева включают, без ограничения, культивированные клетки, например, культивированные клетки млекопитающего, такие как клетки СНО, клетки ВНК, клетки NSO, клетки SP/20, или гибридомные клетки, клетки дрожжей и клетки насекомых, но также клетки, являющиеся клетками трансгенного животного или культивированной ткани.
Термин «сиаловая кислота» относится к любому члену семейства содержащих девять атомов углерода карбоксилированных сахаров. Наиболее важным членом семейства сиаловой кислоты является N-ацетилнейраминовая кислота (2-кето-5-ацетамидо-3,5-дидезокси-D-глицеро-D-галактононулопираноз-1-оновая I кислота (часто сокращенно обозначаемая Neu5Ac, NeuAc или NANA). Вторым членом семейства является N-гликолилнейраминовая кислота (NGNA, Neu5Ge или NeuGe), в которой N-ацетильная группа NeuAc гидроксилирована. Такая форма является преимущественной в гликопротеинах от грызунов или микробных источников. Третьим членом семейства сиаловой кислоты является 2-кето-3-дезоксинонулозоновая кислота (KDN) (Nadano et al. (1986), J. Biol. Chem., 261: 11550-11557; Kanamori et al., J. Biol. Chem., 265: 21811-21819 (1990)). Также включаются 9-замещенные сиаловые кислоты, такие как 9-О-С-С6-ацил-Neu5Ac, подобные 9-О-лактил-Neu5Ac или 9-О-ацетил-Neu5Ac, 9-дезокси-9-фтор-Neu5Ac и 9-азидо-9-дезокси-Neu5Ac. Для представления о статическом семействе сиаловой кислоты см., например, Varki, Glycobiology, 2: 25-40 (1992); Sialic Acids: Chemistry, Metabolism and Function, R. Schauer, Ed. (Springer Verlag, New York (1992)).
Описание
Между тем авторы настоящего изобретения неожиданно обнаружили, что уровень сиалилирования Fc-олигосахаридов изменяет аффинность полученных рекомбинантно лечебных антител в отношении Fcγ-рецепторов, что приводит к модуляции различных аспектов биологических действий указанных антител. Конкретнее, обнаружилось, что Ab с высоким уровнем сиалилирования имеют существенно сниженную аффинность в отношении низкоаффинных рецепторов FcγRIIA (CD32A) и FcγRIIIA (CD16A) и имеют существенно пониженную активность в анализах ADCC in vitro, в которых, как полагают, FcγRIIIA является релевантным рецептором. Также обнаружилось, что Ab с высоким уровнем сиалилирования имеют повышенную аффинность в отношении высокоаффинного Fcγ-рецептора FcγRI (CD64) и что полностью сиалилированные Fc-содержащие белки имеют пониженное время полужизни в сыворотке по сравнению с асилилированными или частично сиалилированными Fc-содержащими белками. Также обнаружилось, что удаление (или отсутствие или пониженные уровни) сиаловой кислоты из Fc-олигосахаридов усиливает авидность полученных рекомбинантно лечебных антител в отношении их молекулы-мишени. Такие открытия и подтверждающие их сведения описаны в одновременно рассматриваемых предварительных заявках на патент США №60/695769, 60/809106, 60/841153.
Без желания привязываться к какой-либо теории, удаление заряженной статической группы из олигосахарида можно интерпретировать как придание большей жесткости всей структуре антитела, что расширяет сферу возможного взаимодействия для двух связывающих доменов друг относительно друга. Способность Ab связываться бивалентно с двумя эпитопами также будет зависеть от доступности, ориентации, плотности и подвижности эпитопа. Следует отметить, что антигенсвязывающее действие сиалилирования также может касаться Ab, узнающих вирусные или бактериальные поверхностные антигены и даже растворимые антигены, которые являются гомополимерами, так как жесткость Ab может определить, до какой степени отдельные молекулы Ab связываются бивалентно в растворимом иммунном комплексе, где не только некоторые из Ab могут связываться с более чем одним антигеном, но некоторые антигены могут связываться с более чем одним Ab.
Настоящее изобретение относится к способу регулирования свойств Fc-содержащей молекулы путем изменения сиалилирования Fc-олигосахаридов и измененных Fc-содержащих молекул. Сиаловая кислота имеет суммарный отрицательный заряд при физиологическом рН, и, таким образом, можно ожидать, что присутствие сиаловой кислоты в Fc-связанном углеводе может изменить трехмерную структуру и, следовательно, конформацию домена СН2 и посредством этого воздействовать на связывание Fc с различными лигандами или рецепторами. Измененная Fc-содержащая молекула влияет на аффинность в отношении одного или нескольких рецепторов FcγRI, FcγRIIA и FcγRIIIA, активность ADCC, активацию макрофагов или моноцитов и время полужизни в сыворотке.
Обогащение сиалилированных форм Fc-содержащих белков
Один подход к получению субпартий определенного Fc-содержащего белка, отличающихся содержанием сиаловой кислоты, должен состоять в получении Fc-содержащего белка с помощью гетерогенных Fc-олигосахаридов, включающих как сиалилированные, так и асилилированные молекулы, и пропускании его через колонку, содержащую иммобилизованный лектин, который имеет различную аффинность в отношении сиалилированных и асилилированных олигосахаридов. Несвязывающуюся прошедшую (Т, прошедшая) или несвязывающуюся в колонке фракцию можно отделить от связанной фракции (В, связанная), причем последнюю собирают в то время как через колонку пропускают буфер для элюирования. Также можно отдельно собрать слабосвязанную фракцию или фракцию, замедленную в колонке (R, замедленная), например, собирая Fc-содержащий белок, который элюируется во время продолжающегося промывания колонки исходным буфером для образца. В зависимости от используемого лектина несвязывающаяся фракция может иметь большее или меньшее содержание сиаловой кислоты, чем фракция, которая связывается.
Примерами лектинов, которые могут повысить концентрацию сиалилированных или асилилированных Fc-содержащих белков, является лектин из Maackia amurensis (MAA), который специфически связывает олигосахариды с концевой сиаловой кислотой, и лектин агглютинин из проростков пшеницы (WGA), который специфически связывает олигосахариды или с концевой сиаловой кислотой или с концевым N-ацетилглюкозамином (GlcNAc). Другим примером является лектин рицин I (RCA), который связывает олигосахариды с концевой галактозой. В последнем примере несвязывающаяся проходящая фракция может быть обогащена сиалилированными Fc-содержащими молекулами.
Ферментативная модификация Fc-содержащих белков
Альтернативным подходом к получению субпартий Fc-содержащего белка, различающихся по содержанию сиаловой кислоты, является обработка части препарата Fc-содержащего белка ферментом сиалидазой, причем посредством этого сиаловые кислоты удаляются. Полученный асиалилированный материал можно сравнить с исходным, частично сиалилированным материалом на различия в биологической активности. Чем выше содержание сиаловой кислоты в партии исходного Fc-содержащего белка, тем больше возможность обнаружения любых различий в биологической активности. Например, если только 10% Fc-олигосахаридов в исходном белковом препарате содержат сиаловую кислоту, может быть затруднительно обнаружить различия в биологической активности после обработки сиалидазой, когда сиаловую кислоту содержат 0-1% олигосахаридов. Сравнение биологической активности Fc-содержащего белка до и после обработки сиалидазой будет более трудным, если обработка сиалидазой приводит к различному разрушению фукозилированных и афукозилированных олигосахаридов, так как уровни фукозы оказывают глубокое влияние на некоторые виды биологической активности, такие как аффинность в отношении человеческого FcγRIIIA и активность ADCC. Например, если уменьшение содержания сиаловой кислоты от 30% олигосахаридов до 0% приводит к возрастанию доли афукозилированных олигосахаридов от 5% до 15%, тогда будет невозможно объяснить различия в активности ADCC только уменьшением содержания сиаловой кислоты. Такое влияние обработки сиалидазой на относительную долю фукозилированных и афукозилированных олигосахаридов возможно (и наблюдалось) из-за различия в сиалилировании фукозилированных и афукозилированных олигосахаридов до обработки сиалидазой для удаления остатков сиалиловых кислот.
Сиалилирования олигосахаридов, присутствующих в Fc-области, также можно достичь с использованием способов гликозилирования in vitro. С использованием таких способов возможно получение максимально сиалилированных гликоформ образцов антител. На основании открытия по настоящему изобретению максимально сиалилированные гликоформы антител или других Fc-содержащих конструкций будут иметь уменьшенное время полужизни в сыворотке по сравнению с асилилированными или малосилилированными антителами. Таким образом, способ по изобретению дает оптимальное средство регулирования как однородности гликоформ, включая антитела или другие рекомбинантные белковые конструкции, содержащие Fc-участок иммуноглобулина, так и функциональных аспектов in vivo указанных антител или конструкций.
Гликозилтрансферазы функционируют в природе, синтезируя олигосахариды. Они продуцируют специфические продукты с превосходной стереохимической и региохимической геометрией. Перенос гликозильных остатков приводит к наращиванию или синтезу олиго- или полисахаридов. Описано несколько типов гликозилтрансфераз, включая сиалилтрансферазы, фукозилтрансферазы, галактозилтрансферазы, N-ацетилгалактозилтрансферазы, N-ацетилглюкозаминилтрансферазы и т.п. Гликозилтрансферазы, применимые в настоящем изобретении, включают, например, α-сиалилтрансферазы, α-глюкозилтрансферазы, α-галактозилтрансферазы, α-фукозилтрансферазы, α-маннозилтрансферазы, α-ксилосилтрансферазы, α-N-ацетилгексозаминилтрансферазы, β-сиалилтрансферазы, β-глюкозилтрансферазы, β-галактозилтрансферазы, β-фукозилтрансферазы, β-маннозилтрансферазы, α-ксилосилтрансферазы и β-N-ацетилгексозаминилтрансферазы, такие как из Neisseria meningitidis или других бактериальных источников, и от крысы, мыши, кролика, коровы, свиньи, человека и насекомых как источников или вирусных источников. Предпочтительно гликозилтрансфераза представляет собой процессированный вариант фермента гликозилтрансферазы, в котором делитирован мембрансвязывающий домен.
Примеры галактозилтрансфераз включают α(1,3)-галактозилтрансферазу (E.C. No. 2.4.1.151, см., например, Dabkowski et al., Transplant. Proc., 25: 2921 (1993), и Yamamoto et al., Nature, 345: 229-233 (1990)) и α(1,4)-галактозилтрансферазу (E.C. No. 2.4.1.38). Можно использовать другие гликозилтрансферазы, такие как сиалилтрансфераза. Часто называемую сиалилтрансферазой α(2,3)-сиалилтрансферазу можно использовать при получении сиалиллактозы или других структур более высокого порядка. Данный фермент переносит сиаловую кислоту (NeuAc) из СМР-сиаловой кислоты в остаток Gal с образованием α-связи между двумя сахаридами. Связывание (соединение) между сахаридами находится между положением 2 NeuAc и положением 3 Gal. Являющаяся примером α(2,3)-сиалилтрансфераза, называемая α(2,3)-сиалилтрансферазой (ЕС 2.4.99.6), переносит сиаловую кислоту в невосстанавливающий концевой Gal дисахарида Galβ → 3Glc или гликозида. См. Van den Eijnden et al., J. Biol. Chem., 256: 3159 (1981), Weinstein et al., J. Biol. Chem., 257: 13845 (1982), и Wen et al., J. Biol. Chem., 267: 21011 (1992). Другой пример α-2,3-сиалилтрансферазы (ЕС 2.4.99.4) переносит сиаловую кислоту в невосстанавливающий концевой Gal дисахарида или гликозида. См. Rearick et al., J. Biol. Chem., 254: 4444 (1979), и Gillespie et al., J. Biol. Chem., 267: 21004 (1992). Другие примеры ферментов включают α-2,6-сиалилтрансферазу Galβ-1,4-GlcNAc (см. Kurosawa et al., Eur. J. Biochem., 219: 375-381 (1994)).
Другими глюкозилтрансферазами, особенно применимыми при получении олигосахаридов по изобретению, являются маннозилтрансферазы, в том числе, α(1,2)-маннозилтрансфераза, α(1,3)-маннозилтрансфераза, β(1,4)-маннозилтрансфераза, Dol-P-Man-синтаза, OCh1 и Pmt1.
Еще другие глюкозилтрансферазы включают N-ацетилгалактозаминилтрансферазы, в том числе, α(1,3)-N-ацетилгалактозаминилтрансферазу, β(1,4)-N-ацетилгалактозаминилтрансферазы (Nagata et al., J. Biol. Chem., 267: 12082-12089 (1992), и Smith et al., J. Biol. Chem., 269: 15162 (1994)), и полипептид N-ацетилгалактозаминилтрансферазу (Homa et al., J. Biol. Chem., 268: 12609 (1993)). Подходящие N-ацетилглюкозаминилтрансферазы включают GnTI (2.4.1.101, Hull et al., BBRC, 176: 608 (1991), GnTII и GnTIII (Ihara et al., J. Biochem., 113: 692 (1993)), GnTV (Shoreiban et al., J. Biol. Chem., 268: 15381 (1993)).
В случае тех воплощений, в которых способ должен осуществляться на практике в коммерческом масштабе, может быть выгодна иммобилизация гликозилтрансферазы на носителе. Такая иммобилизация облегчает удаление фермента из партии продукта и последующее повторное использование фермента. Иммобилизацию гликозилтрансфераз можно осуществить, например, удалением из трансферазы ее мембрансвязывающего домена и присоединение на его месте целлюлозсвязывающего домена. Специалистам в данной области техники будет понятно, что также можно использовать другие способы иммобилизации, и они описаны в доступной литературе.
Поскольку акцепторные субстраты могут, по существу, представлять собой любой моносахарид или олигосахарид с концевым сахаридным остатком, в отношении которого определенная гликозилтрансфераза показывает специфичность, субстрат может быть замещен в положении своего невосстанавливающего конца. Таким образом, гликозидный акцептор может представлять собой моносахарид, олигосахарид, сахарид с флуоресцентной меткой или производное сахарида, такое как аминогликозидный антибиотик, ганглиозид или гликопротеин, в том числе антитела и другие Fc-содержащие белки. В одной группе предпочтительных воплощений гликозидный акцептор представляет собой олигосахарид, предпочтительно, Galβ(1-3)GlcNAc, Galβ(1-4)GlcNAc, Galβ(1-3)GalNAc, Galβ(1-4)GalNAc, Man-α(1-3)Man, Man-α(1,6)Man или GalNAcβ(1-4)маннозу. В особенно предпочтительном воплощении олигосахаридный акцептор присоединяется к домену СН2 Fc-содержащего белка.
Применения субстрата активированного сахара, т.е. сахар-нуклеозидфосфата, можно избежать использованием любой реакции восстановления одновременно с реакцией гликотрансферазы (что также известно как рецикловая система). Например, как утверждается в, например, пат. США 6030815, в рецикловой системе с СМР-сиаловой кислотой используют синтазу СМР-сиаловой кислоты для восполнения СМР-сиаловой кислоты (СМР-NeuAc), так как она взаимодействует с акцептором сиалилтрансферазой в присутствии α(2,3)сиалилтрансферазы с образованием сиалилсахарида. Система, восстанавливающая СМР-сиаловую кислоту, применимая в изобретении, включает цитидинмонофосфат (СМР), нуклеозидтрифосфат (например, аденозинтрифосфат (АТФ), донор фосфата (например, фосфоенолпируват или ацетилфосфат), киназу (например, пируваткиназу или ацетаткиназу), способную переносить фосфат от донора фосфата к нуклеозиддифосфатам, и нуклеозидмонофосфаткиназу (например, миокиназу), способную переносить концевой фосфат от нуклеозидтрифосфата к СМР. α(2,3)Сиалилтрансферазу и синтазу СМР-сиаловой кислоты также можно рассматривать как часть системы регенерации СМР-сиаловой кислоты, так как удаление активированной сиаловой кислоты служит для поддержания лучшей скорости синтеза. Синтез и использование соединений сиаловой кислоты в процедуре сиалилирования с использованием фагмиды, включающей ген для модифицированного фермента синтетазы СМР-сиаловой кислоты, раскрываются в международной заявке WO 92/16640, опубликованной 1 октября 1992.
Другой способ получения олигосахаридов состоит в применении гликозилтрансферазы и активированных гликозилпроизводных в качестве сахаров-доноров, устраняющих потребность в сахарах-нуклеотидах как сахарах-донорах, как описано в пат. США 5952203. Активированные гликозилпроизводные действуют как альтернативы субстратам, встречающимся в природе, которыми являются дорогостоящие сахара-нуклеотиды, как правило, нуклеотиддифосфосахара или нуклеотидмонофосфосахара, в которых нуклеотидфосфат α-соединен с положением 1 сахара.
Активированные производные гликозидов, которые применимы, включают активированную удаляемую группу, такую как, например, фтор, хлор, бром, тозилатэфирная, мезилатэфирная, трифлатэфирная и т.п. Предпочтительные воплощения активированных производных гликозидов включают гликозилфториды и гликозилмезилаты, причем гликозилфториды являются особенно предпочтительными. Среди гликозилфторидов наиболее предпочтительными являются α-галактозилфторид, α-маннозилфторид, α-глюкозилфторид, α-фукозилфторид, α-ксилозилфторид, α-сиалилфторид, альфа-N-ацетилглюкозаминилфторид, α-N-ацетилгалактозаминилфторид, β-галактозилфторид, β-маннозилфторид, β-глюкозилфторид, β-фукозилфторид, β-ксилозилфторид, бета-сиалилфторид, β-N-ацетилглюкозаминилфторид и β-N-ацетилгалактозаминилфторид.
Гликозилфториды можно получить из свободного сахара, сначала ацетилируя сахар и затем обрабатывая его HF/пиридином. Из ацетилированных гликозилфторидов можно удалить защитные группы взаимодействием с умеренным (каталитическим) основанием в метаноле (например, с NaOMe/MeOH). Кроме того, многие гликозилфториды являются коммерчески доступными. Другие активированные гликозилпроизводные можно получить с использованием обычных способов, известных специалистам в данной области техники. Например, гликозилмезилаты можно получить обработкой полностью бензилированной гемиацетальной формы сахара мезилхлоридом с последующим каталитическим гидрированием для удаления бензильных групп.
Другим компонентом взаимодействия является каталитическое количество нуклеозидфосфата или его аналога. Нуклеозидмонофосфаты, подходящие для применения в настоящем изобретении, включают например, аденозинмонофосфат (АМФ), цитидинмонофосфат (СМР), уридинмонофосфат (UMP), гуанозинмонофосфат (GMP), инозинмонофосфат (IMP) и тимидинмонофосфат (ТМР). Нуклеозидтрифосфаты, подходящие для применения согласно настоящему изобретению, включают, аденозинтрифосфат (АТФ), цитидинтрифосфат (СТР), уридинтрифосфат (UTP), гуанозинтрифосфат (GTP), инозинтрифосфат (ITP) и тимидинтрифосфат (ТТР). Предпочтительным нуклеозидтрифосфатом является UTP. Предпочтительно нуклеозидфосфат представляет собой нуклеозиддифосфат, например, аденозиндифосфат (АДФ), цитидиндифосфат (CDP), уридиндифосфат (UDP), гуанозиндифосфат (GDP), инозиндифосфат (IDP) и тимидиндифосфат (TDP). Предпочтительным нуклеозиддифосфатом является UDP. Как отмечалось выше, настоящее изобретение также можно осуществить на практике с аналогом нуклеозидфосфатов. Подходящие аналоги включают, например, нуклеозидсульфаты и сульфонаты. Другие еще аналоги включают простые фосфаты, например пирофосфат.
Одной из процедур модификации рекомбинантных белков, полученных, например, в мышиных клетках, в которых преобладает гидроксилированная форма сиаловой кислоты (NGNA), является обработка белка сиалидазой для удаления сиаловой кислоты типа NGNA с последующим ферментативным галактозилированием с использованием реагента UDP-Gal и бета-1,4-Galтрансферазы для получения высокооднородных гликоформ G2. Затем препарт можно, необязательно, обработать реагентом CMP-NANA и альфа-2,3-сиалилтрансферазой и получить высокооднородные гликоформы G2S2.
Когда желательно удаление или элиминирование групп сиаловой кислоты из гликанов, присоединенных к Fc-области антител или Fc-содержащих молекул, можно использовать сиалидазу. Из литературы известен ряд сиалидаз различной специфичности. Идентифицирована растворимая сиалидаза клеток СНО (Ferrari et al., 1994, Glycobiology, 4: 367-373), и она, если проникает в культуральную среду, может быть ответственна за внеклеточное удаление сиаловых групп из гликанов рекомбинантных белков. Таким образом, возможно, что во время получения рекомбинантных белков может происходить добавление и удаление сиальных групп, что может объяснить различные и гетерогенные структуры гликанов в белках, продуцированных СНО-клеточными линиями.
Сиалидазы (нейраминидазы) с различной специфичностью в отношении субстратов, например, гликопротеинов, гликолипидов и ганглиозидов и связей, выделены и клонированы из различных видов от бактерий до человека. Ферменты с широкой специфичностью в отношении типа статической группы, например, гидроксилированных (NGNA) или негидроксилированных нейраминовых кислот и связей, которые могут представлять собой α2,3-, α2,6- или α2,8-, и разветвленных сиаловых кислот, соединенных с внутренним остатком, включают ферменты из Clostridium perfringens и сиалидазы из Arthrobacter ureafaciens (сиалидаза А, N-ацетилнейраминатгликобигидролаза; ЕС 3.2.1.18). Очищенные ферменты доступны коммерчески от, например, Prozyme, Inc, San Leonardo, CA. Нуклеотидная последовательность гена сиалидазы из А. ureafaciens клонирована (NCBI, инвентарный № AY934539) Lundbeck et al., 2005, Biotechnolo. Appl. Biochem., 41: 225-231.
С использованием способов, хорошо известных в технике, можно сконструировать клетки-хозяева, секретирующие ферменты, способные действовать на внеклеточные олигосахариды, как описано в, например, US7026152, в случае культур, способных продуцировать этанол за счет ферментации сахаров, высвобождаемых эндонуклеазами. Lundbeck et al. (цит. выше) синтезировали процессированную форму сиалидазы А, которая способна удалять остатки сиаловой кислоты из рекомбинантных мутеинов эритропоэтина. Создание клетки-хозяина млекопитающего, способной к экспрессии лечебных антител или другого Fc-содержащего белка, и одновременное десиалилирование такого экспрессированного белка во внеклеточной среде, не показано. Авторы настоящего изобретения показывают, что растворимая форма сиалидазы А может быть коэкспрессирована клетками, продуцирующими антитела, и полученные продукты антитела, извлеченные из культур таких клеток, имеют пониженное содержание сиаловой кислоты в Fc-части своих молекул. Антитела, оптимизированные в отношении сиаловой кислоты, полученные таким образом, имеют усиленную активность ADCC по сравнению с антителами, продуцированными обычными клеточными линиями.
Структурная характеризация вариантов сиаловой кислоты
Для структурной характеризации вариантов сиаловой кислоты, содержащих олигосахариды, препараты гликопротеинов, в том числе, препараты антител, обрабатывают пептид-N-гликозидазой F для высвобождения N-соединенных олигосахаридов. Фермент пептид-N-гликозидаза F (PNGase F) отщепляет аспаргинсвязанные олигосахариды. Высвобожденные олигосахариды метят флуоресцентной меткой антраниловой кислотой (2-аминобензойная кислота), очищают и анализируют ВЭЖХ так, как описано (см. Anumula K.R. and Dhume ST, Glycobiology, 1998, Jul, 8(7): 685-94). Как видно на фигуре 3, олигосахариды, выделенные на хроматограмме как G0, G1, G2, G2S1 и G2S2, можно обнаружить и определить количественно. Агликозилированные образцы, естественно лишенные гликанов или лишенные гликана химически или ферментативно, обозначены Gno.
Биологическая характеризация вариантов сиаловой кислоты
Сравнить Fc-содержащие белки на функциональность можно несколькими хорошо известными анализами in vitro. В частности, представляет интерес аффинность в отношении членов семейства Fcγ-рецепторов FcγRI, FcγRII и FcγRIII. Такие измерения можно осуществить с использованием рекомбинантных растворимых форм рецепторов или клеточноассоциированных форм рецепторов. Кроме того, аффинность в отношении рецептора FcRn, ответственного за длительное время полужизни в кровотоке IgG, можно измерить, например, BIAcore с использованием рекомбинантного растворимого FcRn. Функциональные анализы с использованием клеток, такие как анализы ADCC и анализы CDC, позволяют представить вероятные функциональные последствия определенных структур вариантов. В одном воплощении анализ ADCC строят при наличии NK-клеток, действующих как первичные эффекторные клетки, причем посредством этого отражаются функциональные действия на рецептор FcγRIII. Также можно использовать анализы фагоцитоза для сравнения иммунных эффекторных функций различных вариантов, как и анализы, которыми измеряют клеточные реакции, такие как образование супероксидов или высвобождение медиаторов воспаления.
Анализы аффинности и авидности
Антитела, которые по природе являются поливалентными, можно проанализировать для определения различных параметров связывания с белками-мишенями. Обычным форматом определения кажущегося Kd является ELISA (твердофазный иммуноферментный анализ) или РИА (RIA, радиоиммуноанализ). «ELISA» стал обычно используемым как анализ связывания, выполняемый на твердом носителе с использованием косвенных методов детекции. Как правило, в ELISA растворимые анализируемые вещества удаляют из раствора после специфического связывания с твердофазными реагентами. В данном способе твердофазные реагирующие вещества получают абсорбированием антигенов или антител на пластиковом титрационном микропланшете; в других способах твердофазные реагирующие вещества представляют собой клеточно-ассоциированные молекулы. Во всех протоколах твердофазные реагенты инкубируют с вторичными или третичными реагирующими веществами, ковалентно-связанными с ферментом. Несвязанные конъюгаты удаляют промывкой и добавляют хромогенный или флуорогенный субстрат. Так как субстрат гидролизуется связанным ферментным конъюгатом, образуется окрашенный или флуоресцентный продукт. Наконец, продукт детектируют визуально или с помощью спектрофотометра для прочтения микропланшетов. Интенсивность полученного сигнала пропорциональна количеству исходного анализируемого вещества в испытываемой смеси.
В варианте твердофазного анализа антиген может быть иммобилизован или захвачен косвенно, например, с использованием иммобилизованных антител, которые узнают несоответствующий домен на антигене, или с использованием антител или другого лиганда, который связывается с «меткой», созданной в белке-мишени, например, полигистидиновой последовательностью.
Другим способом измерения связывания антител против антигенов является использование целых клеток, которые экспрессируют (естественно или через генную инженерию) антиген на поверхности клеток. Клетки инкубируют с испытываемым раствором, содержащим первичные антитела. Несвязанные антитела вымывают и затем клетки инкубируют с ферментом, конъюгированным с антителами, специфичными для первичных антител. Несвязанный ферментный конъюгат вымывают и добавляют твердый субстрат. Уровень связанных первичных антител пропорционален количеству гидролизата субстрата. Это будет количественной мерой, если число клеток на единицу объема выдерживают постоянным. С другой стороны, детекцию осуществляют с использованием меченого изотопом лиганда через прямое связывание или конкуренцию, как описано выше. Протоколы анализов ELISA находятся в, например, Ausebel F.M. et al., Current Protocols in Molecular Biology, 2003, John Wiley & Sons, Inc.
Степени связывания, степени ассоциации и степени диссоциации также можно измерить с использованием технологии BIAcore, в которой используют твердофазный фиксатор или лиганд и подвижный жидкофазный фиксатор или лиганд, обнаруживаемые поверхностным плазмонным резонансом.
Способы оценки эффекторной функции
Роль гликозилирования антител в клиренсе и, следовательно, фармакокинетике лечебных Fc-содержащих белков кажется минимальной; оказывается, связывание неонатального Fc-рецептора (FcRn), которое представляется ответственным за удаление IgG из кровотока, не затрагивается удалением N-присоединенного олигосахарида на Fc-части антитела.
Fc-Рецепторы (FcR) IgG, которые связывают опосредуемые антителами IgG иммунные реакции с клеточными эффекторными функциями, включают Fc-гамма-рецепторы: FcRI (CD64), FcRII (CD32) (как FcRIIA, так и FcRIIB) и FcRIII (CD16). Все три найдены отраженными на моноцитах. Однако оказывается, что выработка таких рецепторов на различных клетках-мишенях происходит дифференцировано и в ответ на другие факторы. Поэтому измерение аффинности модифицированных гликозилированием Fc-содержащих биотерапевтических средств для Fc-гамма-рецепторов является одним из подходящих измерений для прогнозирования усиленных эффекторных функций.
Сообщается, что человеческие Ab IgG1 с низкими уровнями фукозы в своих Fc-гликанах имеют большую аффинность в отношении человеческого FcR CD16 и существенно усиливают активность in vitro в анализах ADCC с использованием человеческих эффекторных клеток РВМС (Shinkawa et al., J. Biol. Chem., 278(5): 3466-3473, 2003; Shields et al., J. Biol. Chem., 277(30): 26733-26740, 2002; Umana et al., Nat. Biotech., 17: 176-180, 1999).
Способ оценки эффекторных функций с использованием анализа ADCC in vitro можно осуществить количественно. Таким образом, анализ in vitro можно планировать для измерения способности связанных антител вызывать разрушение отображения своего родственного лиганда правильным отбором мишени и эффекторных клеточных линий и оценки «киллинга» клеток или по неспособности клеток продолжать деление или по высвобождению внутреннего содержимого, например, высвобождению 51Cr. Клетка-мишень может представлять собой клеточную линию, которая естественно экспрессирует мишень-лиганд для антитела, фрагмента антитела или слитого белка по изобретению, или может быть создана для экспрессии и сохранения белка-мишени на своей поверхности. Примером такой созданной клеточной линии является клетка К2 - клеточная линия миеломы мыши Sp2/0, которая устойчиво экспрессирует на своей поверхности рекомбинантный человеческий TNF, который остается в виде трансмембранной формы из-за введения делеции аминокислот 1-12 зрелого цитокина (Perez et al., Cell, 63: 251-258, 1990). Такая клеточная линия применима для оценки изменений активности ADCC антител против TNF, фрагментов антител или созданных слитых белков, направленных против TNF-альфа, имеющих Fc-домены или обладающих активностью Fc-доменов.
Эффекторными клетками для анализа активности ADCC in vitro могут являться РВМС (моноциты периферической крови) человека или другого млекопитающего. Эффекторные клетки РВМС могут быть свежеизвлеченными после забора крови у доноров одобренными способами. Другие моноциты или макрофаги, которые можно использовать, представляют собой клетки, полученные из истекающих жидкостей, например, перитонеальных эксудатов.
Модели in vivo для измерения клеточных иммунных функций также доступны. Например, антитела против CD3 можно использовать для измерения Т-клеточной активации у мышей, поскольку Т-клеточная активация зависит от способа, которым Fc-домен антитела занимает специфические Fcγ-рецепторы. In vitro сравнивали противоопухолевую активность версии с высоким содержанием и низким содержанием фукозы химерных человеческих Ab IgG1 против хемокинного рецептора СС 4, различия в их активности ADCC не наблюдали (с использованием мышиных эффекторных клеток), однако Ab с низким содержанием фукозы показывали более сильную активность in vivo. Человеческие эффекторные клетки не предоставлялись и сохранялись мышиные эндогенные NK-клетки (Niwa et al., Cancer Res., 64: 2127-2133, 2004). Так как рецепторы CD16 на человеческих NK-клетках показали повышенную чувствительность к уровням фукозы Ab IgG1, такие данные предполагают, что механизм, работающий в мышах, отличается от механизма, исследованного в человеческих эффекторных клетках. Одной из возможностей является позднее обнаруженный мышиный рецептор CD16-2 (Mechetina et al., Immunogen., 54: 463-468, 2002). Внеклеточный домен мышиного CD16-2 имеет существенно большую идентичность последовательности с человеческим CD16A (65%), чем известный лучше мышиный рецептор CD16, причем предполагается, что он может быть более чувствительным к уровням фукозы IgG, с которой он связывается, чем мышиный CD16. Его экспрессия в мышинах макрофагоподобных клетках J774, о которой имеются сообщения, согласуется с вероятностью того, что мышиные макрофаги, экспрессирующие CD16-2, могут быть ответственны за большую противоопухолевую активность за счет Ab с низким содержанием фукозы, описанных Niwa et al. (2004). Таким образом, исследование связывания Fc-рецепторов Fc-содержащими белками типа человеческого IgG1 к мышиным эффекторным клеткам не является предсказательным.
Способы получения белков
Различные способы, предполагающие получение Fc-содержащих белков, могут сжимать структуру Fc-олигосахаридов, включая сиаловую кислоту. В одном воплощении клетки-хозяева, секретирующие Fc-содержащие белки, культивируют в присутствии сыворотки, например, сыворотки плода коровы (FBS), которую предварительно не подвергают обработке при повышенной температуре (например, при 56°С в течение 30 минут). Это может привести к Fc-содержащему белку, который не содержит или содержит очень малое количество сиаловой кислоты из-за естественного присутствия в сыворотке активных ферментов сиалидаз, которые могут удалять сиаловую кислоту из Fc-содержащих белков, секретированных такими клетками-хозяевами. В другом воплощении клетки, секретирующие Fc-содержащие белки, культивируют или в присутствии сыворотки, которую подвергали обработке при повышенной температуре, причем посредством этого ферменты сиалидазы инактивируются, или в отсутствие сыворотки или других компонентов среды, которые могут содержать ферменты сиалидазы, так что Fc-содержащий белок имеет более высокие уровни сиаловой кислоты, для применений (например, терапевтических показаний), когда что может быть желательно.
В другом воплощении устанавливают условия, используемые для очистки и дальнейшей обработки Fc-содержащих белков, которые будут благоприятны для оптимального содержания сиаловой кислоты. Например, поскольку сиаловая кислота является лабильной, длительное воздействие в условиях низких рН, например, после элюирования из хроматографической колонки с белком А или во время процессов вирусной инактивации, может одновременно привести к уменьшению содержания сиаловой кислоты.
Создание клеток-хозяев
Как описано в данном описании, клетка-хозяин, выбранная для экспрессии рекомбинантного Fc-содержащего белка или моноклональных антител, является важным фактором, вносящим вклад в конечный состав, в том числе, без ограничения, изменение в состав олигосахаридных групп, оформляющих белок в домене СН2 иммуноглобулина. Таким образом, один аспект изобретения включает отбор соответствующих клеток-хозяев для применения и/или разработки продуцирующих клеток, экспрессирующих нужный лечебный белок.
В одном воплощении клетка-хозяин представляет собой клетку, в которой от природы не хватает сиалалтрансфераз или она их лишена. В другом воплощении клетка-хозяин представляет собой производную линию клеток-хозяев, отобранную для экспресии уменьшенных или недетектируемых уровней сиалилтрансфераз. В еще одном воплощении клетка-хозяин лишена по природе или генетически модифицирована или обработана таким образом, что лишена синтетазы СМР-сиаловой кислоты - фермента, который катализирует образование СМР-сиаловой кислоты, которая является источником сиаловой кислоты, используемой сиалилтрансферазой для переноса сиаловой кислоты к антителам. В родственном воплощении клетка-хозяин может быть лишена по природе или генетически модифицирована или обработана таким образом, что лишена синтетазы пировиноградной кислоты - фермента, который образует сиаловую кислоту из пировиноградной кислоты.
В другом воплощении клетка-хозяин может быть лишена по природе или генетически модифицирована или обработана таким образом, что лишена галактозилтрансфераз, так что антитела, экспрессированные в указанных клетках, лишены галактозы. В отсутствие галактозы сиаловая кислота не будет присоединяться. В отдельном воплощении клетка-хозяин может по природе сверхэкспрессировать или генетически модифицирована для сверхэкспрессии фермента сиалидазы, который удаляет сиаловую кислоту из антител во время получения. Такой фермент сиалидаза может действовать внутриклеточно на антитела перед секретированием антител или будущим секретированием в культуральную среду и действуют на антитела, которые уже секретированы в среду. Способы отбора клеточных линий с измененными гликозилазами и экспрессирующих гликопротеины с измененными составами углеводов описаны (Ripka and Stanley, 1986, Somatic Cell Mol. Gen., 12: 51-62; US 2004/0132140). Способы создания клеток-хозяев для получения антител с измененной картиной гликозилирования, приводящей к усиленной ADCC, описаны, например, в пат. США 6602864, где клетки-хозяева содержат нуклеиновую кислоту, кодирующую, по меньшей мере, одну модифицирующую гликопротеины гликозилтрансферазу, конкретно, β-(1,4)-N-ацетилглюкозаминилтрансферазу III (GnTIII).
Другие подходы к созданию методами генной инженерии свойств гликозилирования клетки-хозяина через манипуляцию с гликозилтрансферазой клетки-хозяина включают элиминирование или подавление активности, как описано в ЕР 1176195, конкретно, альфа-1,6-фукозилтрансферазы (генного продукта FUT8). Для специалистов в данной области техники может быть очевидно практическое осуществление способов создания клетки-хозяина методами генной инженерии в других конкретных примерах, цитированных выше. Кроме того, созданная клетка-хозяин может происходить от млекопитающего или может быть отобрана из клеток миеломы, лимфомы, дрожжей, насекомого или растения, или может представлять собой их дериватизированную, иммобилизованную или трасформированную клетку.
В другом воплощении способ подавления или элиминирования активности фермента, требуемого для присоединения сиаловой кислоты, можно выбрать из группы, состоящей из сайленсинга генов, такого как с использованием миРНК (siRNA), генетического подхода или добавления ингибитора фермента, например, путем коэкспрессии внутриклеточных Ab или пептида, специфического для фермента, который связывает и блокирует ферментативную активность, и других известных методов генной инженерии. В другом воплощении способ усиления экспрессии или активности фермента, блокирующего присоединение сиаловой кислоты, или фермента сиалидазы, который удаляет сиаловые кислоты, которые уже присоединены, можно выбрать из группы, состоящей из трансфекций рекомбинантными генами фермента, трансфекций факторов транскрипции, которые усиливают синтез ферментной РНК, или генетических модификаций, которые усиливают устойчивость ферментной РНК, причем все способы ведут к усиленной активности ферментов, таких как сиалидазы, что приводит к более низким уровням сиаловой кислоты в очищенном продукте. В другом воплощении в культуральную среду для клеток можно добавлять специфические ингибиторы ферментов.
Антитела
Антитела, описанные в данной заявке, могут находиться в организме или могут происходить от любого млекопитающего, такого как, но без ограничений, человек, мышь, кролик, крыса, грызун, примат, или могут представлять собой их любое сочетание и включают изолированные человеческие, антитела, антитела примата, грызуна, млекопитающего, химерные, гуманизированные и/или CDR-привитые антитела против интегрина, иммуноглобулины, продукты расщепления и другие их установленные части и варианты. Изобретение также относится к антителам, кодирующим нуклеиновые кислоты или комплементарным им, векторам, клеткам-хозяевам, композициям, препаратам, устройствам, трансгенным животным, трансгенным растениям и способам их получения и использования, описанным в данном описании, вместе с комбинациями с тем, что известно в технике.
Настоящее изобретение также относится к клеткам, клеточным линиям и клеточным культурам, которые экспрессируют иммуноглобулин или его фрагменты, способные к гликозилированию в домене СН2, которые связывают антиген, цитокин, интегрин, антитела, фактор роста, поверхностный антиген, который представляет собой маркер дифференцировки и направления дифференцировки клеток, гормон, рецептор или его слитый белок, белок крови, белок, вовлеченный в коагуляцию, их любые фрагменты и любой структурный или функциональный аналог перечисленных факторов. В предпочтительном воплощении иммуноглобулин, его фрагмент или производное связываются с антигеном на поверхности клетки-мишени. В особенно предпочтительном варианте клетка-мишень представляет собой опухолевую клетку, клетку сосудистой сети опухоли или иммуноцит. В определенном воплощении иммуноглобулин, его фрагмент или производное связываются с TNF, интегрином, В-клеточным антигеном или фактором ткани.
В еще одном воплощении клетки, клеточные линии и клеточные культуры по настоящему изобретению могут экспрессировать, что поддается обнаружению, слитый белок, включающий фактор роста или гормон. Примеры факторов роста, рассматриваемые в настоящем изобретении, включают, но не ограничиваются перечисленным, человеческий фатор роста, тромбоцитарный фактор роста, эпидермальный фактор роста, фактор роста фибробластов, нервный фактор роста, человеческий хорионический гонадотропин, эритропоэтин, тромбопоэтин, костный морфогенный белок, трансформирующий фактор роста, инсулиноподобный фактор роста или глюкагоноподобный пептид и их любые структурные или функциональные аналоги.
Изолированные антитела по изобретению включают антитела, имеющие изотипы с активностью ADCC, в особенности, человеческий IgG1 (например, IgG1-каппа и IgG1-лямбда), и менее предпочтительными являются IgG2 и IgG3 или гибридные изотипы, содержащие определенные измененные остатки в Fc-доменах, которые являются их эквивалентами из других видов. Антитела могут представлять собой полноразмерные антитела (например, IgG1) или могут включать только антигенсвязывающую часть и Fc-часть или домен, способные выявлять эффекторные функции, в том числе, ADCC, активацию комплемента и связывание C1q.
Кроме того, фрагменты иммуноглобулина, продуцируемые клетками, клеточными линиями и клеточными культурами по настоящему изобретению, могут включать, но без ограничения, Fc- или другие содержащие СН2-домен структуры, и их любые структурные или функциональные аналоги. В одном воплощении фрагмент иммуноглобулина представляет собой димерный слитый полипептид домена рецептора. В определенном воплощении димерный слитый полипептид домена рецептора представляет собой этанерцепт (etanercept). Этанерцепт представляет собой молекулу растворимого рецептора TNFα, которую вводят подкожно и которая связывается с TNFα в сыворотке пациента, переводя его в биологически неактивный. Этанерцепт представляет собой димерный слитый белок, состоящий из внеклеточной связывающей части человеческого рецептора фактора некроза опухоли (TNFR) в 75 килодальтон (р75), соединенной с Fc-частью человеческого IgG1. Fc-Компонент этанерцепта содержит домен СН2, домен СН3 и шарнирную область, но не домен СН1 IgG1.
Другие продукты, расположенные к получению с использованием клеточных линий по изобретению, включают лечебные или профилактические белки, получаемые в настоящее время с помощью других типов животных клеточных линий и содержащие СН2, способный к гликозилированию. Особенно предпочтительными являются такие лечебные гликозилированные, содержащие СН2-домен белки, которые связываются с антигенами-мишенями на клеточной поверхности клеток типа, который желательно вывести или элиминировать из организма. Создан ряд таких лечебных антител, которые содержат человеческий IgG1, в особенности IgG1, тяжелая цепь которого включает человеческий домен СН1, СН2 и СН3. Такие лечебные белки включают, но не ограничиваются, белки, описанные в данном описании ниже.
Инфликсимаб теперь продают как ремикад® (REMICADE®). Инфликсимаб представляет собой химерные моноклональные антитела IgG1κ с молекулярной массой приблизительно 149100 дальтон. Он состоит из человеческой константной и мышиной вариабельной областей. Инфликсимаб специфически связывается с человеческим фактором некроза опухоли альфа (TNF(альфа)) с константной ассоциации 1010 М-1. Инфликсимаб нейтрализует биологическую активность TNF(альфа) путем связывания с высокой аффинностью с растворимой и трансмембранной формами TNF(альфа) и ингибирует связывание TNF(альфа) с его рецепторами. Клетки, экспрессирующие трансмембранный TNF(альфа), связываемый инфликсимабом, можно лизировать in vitro или in vivo. Инфликсимаб показан для лечения ревматоидного артрита, болезни Крона и анкилозирующего спондилита. Инфликсимаб дают дозами 3-5 мг/кг внутривенной инфузией и затем последующими дополнительными подобными дозами через 2, 6 и/или 8 недель и с интервалами каждые 8 недель, в зависимости от заболевания, от которого лечат.
Даклизумаб (продаваемый как зенапакс® (ZENAPAX®)) представляет собой иммуносупрессорные гуманизированные антитела IgG1, получаемые технологией рекомбинантных ДНК, которые связываются специфически с субъединицей альфа (р55-альфа, CD25 или субъединица Тас) человеческого высокоаффинного рецептора интерлейкина II (IL-2), который экспрессируется на поверхности активированных лимфоцитов. Даклизумаб представляет собой мышиночеловеческие химерные антитела с привитыми определяющими комлементарность участками. Человеческие последовательности образованы от константных доменов человеческого IgG1 и вариабельных каркасных областей антител против миеломы Eu. Мышиные последовательности образованы от CDR мышиных анти-Тас антител. Даклизумаб показан для профилактики резкого отторжения органов у пациентов, получивших почечные трансплантаты, и как правило, используется как часть иммуносупрессивной схемы лечения, которая включает циклоспорин и кортикостероиды.
Базиликсимаб (продаваемый как симулект® (SIMULECT®)) представляет собой химерные (мышь/человек) антитела, получаемые технологией рекомбинантных ДНК, которые функционируют как иммуносупрессивное средство, специфически связываясь с и блокируя (альфа)-цепь рецептора интерлейкина-2 (IL-2R(альфа), также известного как антиген CD25) на поверхности активированных Т-лимфоцитов. Молекулярная масса белка, вычисленная на основании аминокислотной последовательности, составляет 144 килодальтона. Он представляет собой гликопротеин, полученный ферментацией определенной клеточной линии миеломы мыши, созданной генной инженерией для экспрессии плазмид, содержащих гены константной области тяжелой и легкой цепи человека (IgG1) и гены вариабельной области тяжелой и легкой цепи мыши, кодирующие антитело RFT5, которое селективно связывается с IL-2R(альфа). Базиликсимаб показан для профилактики резкого отторжения органов у пациентов, получивших трансплантацию почки, когда используется как часть иммуносупрессивной схемы лечения, которая включает циклоспорин и кортикостероиды.
Адалимумаб (продаваемый как гумира® (HUMIRA®)) представляет собой моноклональные антитела рекомбинантный человеческий IgG1, специфичные для фактора некроза опухоли (TNF) человека. Адалимумаб создавали с использованием технологии фазового дисплея, приводящей к антителам с вариабельными областями тяжелой и легкой цепи человеческого происхождения и константными областями IgG1-каппа человека. Гумира® показан для уменьшения признаков и симптомов и ингибирования развития структурного повреждения у взрослых пациентов с ревматоидным артритом от умеренного до тяжелого, которые имеют неадекватную реакцию на один или несколько DMARD. Гумиру® можно использовать один или в сочетании с МТХ или другими DMARD.
Ритуксимаб (продавемый как ритуксан® (RITUXAN®)) представляет собой созданные генной инженерией химерные мышиные/человеческие моноклональные антитела, направленные против антигена CD2, обнаруженного на поверхности нормальных и злокачественных В-лимфоцитов. Антитела представляют собой иммуноглобулин IgG1-каппа, содержащий последовательности вариабельных областей легкой и тяжелой цепи мыши и последовательности константной области человека. Ритуксимаб обладает аффиностью связывания в отношении антигена CD2 приблизительно 8,0 нМ. Ритуксимаб показан для лечения пациентов с рецидивной или рефракторной низкодифференцированной или фолликулярной, CD2-положительной В-клеточной неходжкинской лимфомы. Ритуксан® дают в количестве 375 мг/м 2 IV инфузиями один раз в неделю на протяжении 4-8 доз.
Трастузумаб (продаваемый как герцептин® (HERCEPTIN®)) представляет собой рекомбинантные образованные от ДНК гуманизированные моноклональные антитела, которые селективно связываются с высокой аффинностью в анализе на основе клеток (Kd=5 нМ) с внеклеточным доменом белка 2 рецептора эпидермального фактора роста человека HER2. Антитела представляют собой иммуноглобулин IgG1-каппа, который содержит каркасные области человека, с гипервариабельными участками мышиного антитела (4D5), которые связываются с HER2. Герцептин показан как средство монотерапии для лечения пациентов с метастазирующим раком молочной железы, опухоли которых сверхэкспрессируют белок HER2, и кто получал одно или несколько лечений химиотерапией против своего метастазирующего заболевания. Герцептин® в сочетании с паклитакселом показан для лечения пациентов с метастазирующим раком молочной железы, опухоли которых сверхэкспрессируют белок HER2, и кто не получал лечения химиотерапией против своего метастазирующего заболевания. Рекомендованной дозировкой является начальная ударная доза в 4 мг/кг трастузумаба, вводимая 90-минутной инфузией, и еженедельная поддерживающая доза в 2 мг/кг трастузумаба, которую можно вводить в виде 30-минутной инфузии, если начальная ударная доза хорошо переносится.
Алемтузумаб (продаваемый как кампат® (CAMPATH®)) представляет собой рекомбинантные образованные от ДНК гуманизированные моноклональные антитела (кампат-1Н), которые направлены против гликопротеина клеточной поверхности CD52 в 21-28 кД. Алемтузумаб связывается с CD52 - немодулирующим антигеном, который присутствует на поверхности, по существу, всех В- и Т-лимфоцитов, большинства моноцитов, макрофагов и NK-клеток, субпопуляции гранулоцитов и тканей мужской репродуктивной системы. Антитела кампат-1Н представляют собой IgG1-каппа с человеческими каркасными и константными областями и гипервариабельными участками из мышиных (крысиных) моноклональных антител (кампат-1G). Кампат показан для лечения В-клеточного хронического лимфолейкоза (B-CLL) у пациентов, которых лечили алкилирующими средствами и для которых лечение флударабилом оказалось недостаточным. Определение эффективности кампата основано на общих скоростях реакций. Кампат дают сначала в количестве 3 мг, которые вводят в виде 2-часовой IV-инфузии ежедневно; переносимую ежесуточную дозу следует повышать до 10 мг и продолжать до тех пор, пока она переносится. Когда такой уровень дозы является переносимым, можно начинать вводить поддерживающую дозу кампата в 30 мг и вводит ее три раза в течение недели и до 12 недель. Для большинства пациентов повышение дозы до 30 мг можно осуществлять в 3-7 дней.
Омализумаб (продаваемый как ксолаир® (XOLAIR®)) представляет собой рекомбинантные моноклональные антитела IgG1(каппа), которые селективно связываются с человеческим иммуноглобулином Е (IgE). Омализумаб ингибирует связывание IgE с высокоаффинным рецептором IgE (Fc(эпсилон)RI) на поверхности тучных клеток и базофилов. Уменьшение связанного с поверхностью IgE на клетках, несущих Fc(эпсилон)RI, ограничивает степень высвобождения медиаторов аллергической реакции. Лечение омализумабом также уменьшает число рецепторов Fc(эпсилон)RI на базофилах у аллергических пациентов. Омализумаб показан для взрослых и подростков (в возрасте 12 лет и выше) со стойкой астмой, у которых положительный кожный тест или реактивность in vitro на круглогодичный аэроаллерген и у кого симптомы неадекватно регулируются вдыхаемыми кортикостероидами. Омализумаб вводят SC каждые 2 или 4 недели в дозе 150-375 мг.
Эфализумаб (раптива® (RAPTIVA®)) представляет собой иммуносупрессивные рекомбинантные гуманизированные моноклональные антитела изотипа IgG1-каппа, которые связываются с человеческим CD11a. Эфализумаб связывается с CD11a - (альфа)субъединицей функционально-связанного антигена-1 лимфоцитов (LFA-1), которая экспрессируется на всех лейкоцитах, и снижает экспрессию CD11a на клеточной поверхности. Эфализумаб ингибирует адгезию лейкоцитов к клеткам других типов. Взаимодействие между LFA-1 и ICAM-1 вносит вклад в инициацию и поддержание многих процессов, в том числе активации Т-лимфоцитов, адгезии Т-лимфоцитов к эндотелиальным клеткам и миграции Т-лимфоцитов к местам воспаления, в том числе псориатической коже. Активация лимфоцитов и направленная миграция играет роль в патофизиологии хронического бляшечного псориаза. В псориатической коже экспрессия ICAM-1 на клеточной поверхности активируется на эндотелии и кератиноцитах. CD11a также экспрессируется на поверхности В-лимфоцитов, моноцитов, нейрофилов, природных киллерных клеток и других лейкоцитов. Следовательно, существует возможность воздействия эфализумаба на активацию, адгезию, миграцию и числа клеток иных, чем Т-лимфоциты. Рекомендованной дозой раптива® является одна общеукрепляющая доза SC в 0,7 мг/кг с последующими еженедельными дозами SC в 1 мг/кг (максимальная однократная доза не превышает в целом 200 мг).
В другом воплощении клеточная линия по изобретению устойчиво трансфицирована или создана иным способом для экспрессии полипептида, который образован не от иммуноглобулина, но который попадает в определение Fc-содержащего белка.
Нуклеиновые кислоты, кодирующие антитела и белки по данному изобретению, можно получить несколькими способами, хорошо известными в технике. В одном аспекте антитела обычно получают из гибридом, полученных иммунизацией мыши пептидами по изобретению. Антитела можно получить таким образом с использованием любых гибридомных методов, хорошо известных в технике, см., например, Ausubel et al., ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY, NY (1987-2001); Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor, NY (1989); Harlow and Lane, Antibodies, a Laboratory Manual, Cold Spring Harbor, NY (1989); Colligan et al., eds., Current Protocols in Immunology, John Wiley & Sons, Inc., NY, NY (1994-2001); Colligan et al., eds., Current Protocols in Protein Science, John Wiley & Sons, Inc., NY, NY (1997-2001); все работы включены в данное описание в качестве ссылок.
В другом обычном способе получения связывающей мишень части антитела, типично, вариабельных тяжелых и/или вариабельных легких областей антитела, указанные части выбирают из библиотеки таких связывающих областей, созданных в, например, фаговой библиотеке. Фаговую библиотеку можно создать с помощью встраивания библиотеки случайных олигонуклеотидов или библиотеки полинуклеотидов, содержащих последовательности, представляющие интерес, например, из В-клеток иммунизированного животного или человека (Smith G.P., 1985, Science, 228: 1315-1317). Фаговые библиотеки антител содержат пары вариабельных областей тяжелой (Н) и легкой (L) цепи в одном фаге, что допускает экспрессию одноцепных Fv-фрагментов или Fab-фрагментов (Hoogenboom et al., 2000, Immunol. Today, 21(8): 371-8). С многообразием фагемидной библиотеки можно манипулировать для усиления и/или изменения иммуноспецифичности моноклоналльных антител библиотеки для получения и последующей идентификации других нужных человеческих моноклональных антител. Например, молекулу иммуноглобулина с тяжелой (Н) цепью и легкой (L) цепью, кодирующую гены, можно произвольно смешать (перетасовать) для создания новых пар HL в собранной молекуле иммуноглобулина. Кроме того, любую Н или L цепь или обе, кодирующие гены, можно подвергнуть мутагенезу в гипервариабельной области (CDR) вариабельной области полипептида иммуноглобулина и затем скринировать на желательную аффинность и нейтрализующую способность. Библиотеки антител также можно создать синтетически, отбирая одну или несколько человеческих каркасных последовательностей и вводя коллекции кассет CDR, полученных из спектров человеческих антител или через созданную вариацию (Kretzschmar and von Ruden, 2000, Current Opinion in Biotechnology, 13: 598-602). Позиции многообразия не ограничиваются CDR, но также могут включать каркасные сегменты вариабельных областей или могут включать иные вариабельные области, чем области антител, например, пептидов.
Другими библиотеками связывающих компонентов-мишеней, которые могут включать иные вариабельные области, чем области антител, являются рибосомный дисплей, дрожжевой дисплей и бактериальные дисплеи. Рибосомный дисплей представляет собой способ трансляции мРНК в их родственные белки при сохранении белка, связанного с РНК. Нуклеотидную кодирующую последовательность извлекают RT-PCR (Mattheakis L.C. et al., 1994, Proc. Natl. Acad. Sci. USA, 91, 9022). Дрожжевой дисплей основан на конструировании слитых белков мембранассоциированного альфа-агглютининового дрожжевого рецептора адгезии aga1 и aga2 - части системы типа скрещивания (Broder et al., 1997, Nature Biotechnology, 15: 553-7). Бактериальный дисплей основан на слиянии мишени с экспортированными бактериальными белками, ассоциирующимися с клеточной мембраной или клеточной стенкой (Chen and Georgiou, 2002, Biotechnol. Bioeng., 79: 496-503).
При сравнении с технологией гибридом методы фагового дисплея и дисплеев других антител дают возможность выбора манипуляции против антигена-мишени in vitro и без ограничения возможности воздействия хозяина на антиген или наоборот.
Клетки-хозяева
Клетки-хозяева, описанные в данном описании, способны к продуцированию специфических антител с определенным содержанием сиаловой кислоты в олигосахаридной части указанных антител.
В отличие от большинства генов, которые транскрибируются из непрерывных геномных последовательностей ДНК, гены антител собираются из генных сегментов, которые могут быть совершенно разделенными в зародышевой линии. В частности, гены тяжелой цепи образуются путем рекомбинации трех геномных сегментов, кодирующих вариабельные (V), дополнительные (D) и соединяющие (J)/константные (С) области антитела. Функциональные гены легкой цепи образуются соединением двух генных сегментов: один кодирует V-область и другой кодирует J/C-область. Локусы как тяжелой цепи, так и легкой цепи каппа содержат множество генных сегментов V (оценки варьируются от сотен до тысяч), оцененные до интервала далеко за 1000 п.о. Локус лямбда является, напротив, намного меньшим и, как показано, у мыши на хромосоме 16 в интервале приблизительно 300 п.о. Он состоит из двух вариабельных генных сегментов и четырех генных сегментов связывающей/константной (J/C) области. Образование функционального гена требует рекомбинации между V и элементом J/C.
В В-клетке, в которой естественно продуцируется антитело, регуляция и транскрипция генов как реаранжированной тяжелой, так и легкой цепи каппа зависит как от активности тканеспецифического промотора выше V-области, так и от тканеспецифического энхансера, расположенного в J-C-интроне. Указанные элементы действуют синергично. Также в локусе легкой цепи каппа идентифицирован второй В-клеточный специфический энхансер. Этот дополнительный энхансер расположен на 9 п.о. ниже Скаппа. Таким образом, гибиридомный метод иммортализации антителоэкспрессирующих генов имеет в основании эндогенные промоторные и энхансерные последовательности пре-В-клеточной линии дифференцировки. С другой стороны, нуклеиновые кислоты по настоящему изобретению можно экспрессировать в клетке-хозяине путем запуска (путем манипуляции) в клетке-хозяине, содержащей эндогенную ДНК, кодирующую антитело по настоящему изобретению. Такие способы также хорошо известны в технике, например, описанные в патентах США №5580734, 5641670, 5733746 и 5733761, включенных в данное описание в качестве ссылок.
Клонирование геномной ДНК антитела в искусственный вектор является другим способом создания клеток-хозяев, способных экспрессировать антитела. Однако экспрессия моноклональных антител позади сильного промотора повышает шансы идентификации высокопродуцирующих линий и получения более высоких выходов моноклональных антител. Антитела по изобретению можно получить в клетке-хозяине трансфектоме с использованием, например, сочетания методов рекомбинантных ДНК и методов трансфекции генов, что хорошо известно в технике (см., например, Morrison S. (1985), Science, 229: 1202).
Системы клонирования и экспрессии полипептида во множестве различных клеток-хозяев хорошо известны. Подходящие клетки-хозяева включают бактерии, клетки млекопитающих, клетки растений, дрожжевые и бакуловирусные системы и трансгенные растения и трансгенных животных. Линии клеток млекопитающих, доступные в технике для экспрессии полноценных гликозилированных белков гетерологичных полипептидов, включают клетки яичника китайского хомячка (СНО), клетки HeLa, клетки почки детеныша хомяка (ВНК), клетки меланомы мыши NSO и производные клеточные линии, например, клетки миеломы крысы SP2/0, YB2/0 (ATC CRL-1662), эмбриональные почечные клетки человека (НЕК), эмбриональные ретинальные клетки человека, клетки PerC.6, клетки hep G2, клетки BSC-1 (например, ATCC CRL-26) и многие другие, доступные от, например, Американской коллекции типовых культур, Va (www.atcc.org). Обычно предпочтительным бактериальным хозяином является E. coli.
Клетки млекопитающих, такие как клетки СНО, клетки миеломы, клетки НЕК293, клетки ВНК (ВНК21, АТСС CRL-10), мышиные клетки Ltk- и клетки и NIH3T3, часто используют для устойчивой экспрессии гетерологичных генов. Клеточные линии, такие как Cos (COS-1, АТСС CRL 1650; COS-7, АТСС CRL-1651) и клетки НЕК293, обычно используют для временной экспрессии рекомбинантных белков.
Предпочтительные клетки-хозяева от млекопитающих для экспрессии рекомбинантных антител по изобретению включают клетки миеломы, такие как SP2/0, YB2/0 (ATC CRL-1662), NSO и Р3Х63.Ag8.653 (например, SP2/0-Ag14), из-за их высокой скорости экспрессии. В частности, для применения с клетками миеломы NSO другой предпочтительной экспрессирующей системой является система экспрессии гена GS, раскрытая в WO 87/04462, WO 89/01036 и ЕР 338841. Когда рекомбинантные экспрессирующие векторы, кодирующие гены антитела, вводят в клетки-хозяева от млекопитающих, антитела получают путем культивирования клеток-хозяев в течение периода времени, достаточного для экспрессии антител в клетках-хозяевах, предпочтительнее, секреции антител в культуральную среду, в которой росли клетки-хозяева. Антитела можно извлечь из культуральной среды с использованием стандартных способов очистки белков.
Клетки CHO СНО-К1 и DHFR-, DG44 и DUK-B11 (G. Urlaub, L.A. Chasin, 1980, Proc. Natl. Acad. Sci. U.S.A., 77, 4216-4220) используют для получения белков на высоком уровне, поскольку амплификация генов, представляющих интерес, становится возможной за счет внедрения селектируемого амплифицируемого маркера DHFR с использованием, например, лекарственного средства метотрексата (МТХ) (R.J. Kaufman, 1990. Methods Enzymol., 185: 537-566). Клетки СНО DHFR- можно успешно использовать для получения рекомбинантных Mab на высоком уровне. Клетки СНО DHFR- могут продуцировать антитела против МСР-1 со скоростью 80-110 мг на 106 клеток в сутки или более 200 мг на 106 клеток в сутки. Используют ряд промоторов для получения экспрессии Н- и L-цепей в таких клетках СНО, например, b-актиновый промотор, промотор человеческий CMV MIE, основной поздний промотор Ad-вируса (MLP), промотор RSV и вирус лейкоза мыши LTR. В литературе описан ряд векторов для экспрессии mAb, в которых две цепи Ig несут две различные плазмиды с независимым селектируемым/амплифицируемым маркером. Векторы, содержащие только цепь антитела, например, Н-цепь, связаны с маркером DHFR, а экспрессирующая кассета L-цепи с маркером Neor или наоборот, что можно использовать для получения до 180 мг гуманизированных mAb L-1 7 в сутки во вращающихся колбах. Способы, используемые для начального отбора и последующей амплификации, могут быть различными и хорошо известны специалистам в данной области техники. Вообще, высокий уровень экспрессии mAb можно получить с использованием следующих стадий: начальный отбор и последующая амплификация клонов-кандидатов, совместный отбор (например, в случаях, когда экспрессирующие векторы как Н-цепи, так и L-цепи содержат экспрессирующий элемент DHFR) и амплификация, соамплификация с использованием различных амплифицируемых маркеров, и начальный отбор и амплификация в культуре в массе, с последующим клонированием методом серийных разведений для идентификации отдельных высококэкспрессирующих клонов. Поскольку сайты интеграции могут влиять на эффективность экспрессии Н-цепи и L-цепи и общую экспрессию mAb, создают отдельный вектор, в котором два элемента, экспрессирующих цепь Ig, размещены в тандеме. Такие векторы также содержат доминантный селектируемый маркер, такой как Neor, и экспрессирующую кассету DHFR. Для общего представления см. Ganguly S. and A. Shartzman, Expression Systems, mammalian cells; в Encyclopedia of Bioprocess Technology: Fermentation, Biocatalysis, and Bioseparation. 1999, John Wiley & Sons, Inc.
Cockett et al. (1990, Bio/Technology, 8, 662-667) разработали систему GS для экспрессии на высоком уровне гетерологичных генов в клетках СНО. Трансфекцию экспрессирующего вектора, содержащего кДНК (под транскрипционным контролем промотора hCMV) и миниген GS (под контролем позднего промотора SV40) в клетки СНО-К1 (с последующим отбором с 20-500 мМ MSX) можно использовать для получения клонов, экспрессирующих антитела по изобретению с выходами, сравнимыми с выходом для систем DHFR- СНО. Система GS обсуждается в целом или частично в европейских патентах №0216846, 0256055 и 0323997 и заявке на европейский патент №89303964.4.
Хотя изобретение описано в общих терминах, воплощения изобретения будут дополнительно раскрыты в приведенных далее примерах.
Пример 1. Ферментативная модификация галактозилирования и сиалилирования антител
Для галактозилирования образцов очищенных антител ферментативным способом к образцам антител добавляют бычью β-1,4-галактозилтрансферазу (β1,4GT) и UDP-Gal, полученную от Sigma Chemical Co. (St. Louis, MO). Рекомбинантную α-2,3-сиалилтрансферазу (α2,3ST), рекомбинантную α-1,3-галактозилтрансферазу (α1,3GT) и CMP-Sia получают от Calbiochem (San Giego, CA). PNGase F получают от New England Biolabs (Beverly, MA) или от Prozyme (San Leandro, CA) или от Selectin BioSciences (Pleasant Hill, CA). β-Галактозидазу и β-глюкозаминидазу из Diplococcus pneumoniae получают или от ProZyme или от Selectin BioSciences. β-Галактозидаза из почки коровы и все другие ферменты или от ProZyme или от Selectin BioSciences. Колонки с белком А NAP-5 и HiTrap от Pharmacia Biotech (Piscataway, NJ). Все другие реагенты являются чистыми для анализа.
Получают ферментативно дегликозилированную форму (обозначаемую Gno) Ab1 для использования в качестве контрольных антител, которые утратили Fc-иммунную эффекторную функцию. Такой вариант получают, растворяя Ab1 (~10 мг в 1,0 мл буфера) в 100 мМ буфера MES (рН 7,0) и обрабатывая их 1000 Е PNGase F при 37оС в течение 24 часов. Добавляют другую аликвоту фермента и продолжают инкубацию еще в течение 24 часов. Дегликозилированные Ab1 очищают с использованием колонки с белком А HiTrap и вводят в PBS, рН 7,0. Гликоформу Gno характеризуют MALDI-TOF-MS для подтверждения дегликозилирования.
Кроме препаратов Ab, обрабатываемых в лаборатории, также сравнивают субпартии Ab, различающиеся по природе содержанием сиаловой кислоты, называемые в данном описании «природными вариантами». Немодифицированные антитела называют Ab1 PBS после того, как буфер для материала из исходной партии обменивали на PBS. Используют моноклональные Ab Ab1 и Ab3 человеческого IgG1, в которых члены пары различаются по степени Fc-сиалилирования, очевидно, из-за раличных способов, использованных для их получения (но продуцированных одним и тем же типом клеток-хозяев). Варианты Ab1 Ab1-20 и Ab1-29 содержат 20% и 29% сиалилированных гликанов, соответственно, и варианты Ab5 Ab5-20 и Ab5-26 содержат 0% и 26% сиалилированных гликанов, соответственно. Иными словами, члены каждой пары имеют одинаковые аминокислотные последовательности, одинаковые уровни Fc-фукозилирования и различающееся вполовину содержание GlcNAc (анализы масс-спектрометрией MALDI-TOF) и одинаковый низкий уровень агрегатов Ab (<1% по анализам SEC-ВЭЖХ).
Все препараты Ab и Fc-содержащих белков, используемые в различных биологических анализах, и способы, которыми их получали, сведены в таблице 1.
Общий перечень препаратов Fc-содержащих белков, используемых в данном примере
Все испытываемые образцы содержат шарнирную область, домены СН2 и СН3 человеческого IgG1. Ab1, Ab2, Ab3 и Ab5 являются моноклональными Ab IgG с человеческим IgG1 и константными областями каппа. Ab1 представляют собой полностью человеческие Ab, специфичные для человеческого TNF, и Ab2 представляют собой мышиные/человеческие химерные Ab, специфичные для человеческого TNF. Ab3 представляют собой полностью человеческие Ab, специфичные для одной из субъединиц гетеродимерного провоспалительного цитокина. Все четыре Ab экспрессируются в трансфицированных клетках миеломы мыши Sp2/0. Ab5 представляют собой полностью человеческие Ab, направленные на субъединицу гетеродимерного рецептора клеточной поверхности. FcP1 представляет собой димерный слитый белок, включающий шарнирную область, домены СН2 и СН3 человеческого IgG1.
Гликоформы G2 получают, подвергая образцы образцы IgG в буфере 100 мМ MES (рН 7,0) (~10 мг в 1,0 мл буфера) воздействию 50 миллиединиц β1,4GT, 5 мкмоль UDP-Gal и 5 мкмоль MnCl2 при 37°С в течение 24 часов. Добавляют еще одну аликвоту фермента и UDP-Gal и смесь инкубируют еще в течение 24 часов при 37оС. Образцы повторно галактозилированного IgG очищают с использованием колонки с белком А HiTrap. Олигосахариды высвобождают PNGase F и характеризуют MALDI-TOF-MS и ВЭЖХ, как описано ниже.
Гликоформу G2S2 получают, вводя образцы IgG в буфер 100 мМ MES (рН 7,0) (~10 мг в 1,0 мл буфера), с использованием колонки NAP-5 согласно предлагаемым протоколам изготовителя. В такой раствор добавляют по 50 миллиединиц каждого из β1,4GT и α2,3ST и по 5 мкмоль каждого из UDP-Gal, СМР-Sia (изомер NANA) и MnCl2. Смесь инкубируют при 37°С. Через 24 часа добавляют вторую аликвоту ферментов вместе с нуклеотидсодержащими сахарами и смесь инкубируют еще в течение 24 часов при 37°С. Гликоформу G2S2 образцов IgG очищают так, как описано выше. В случае одной определенной партии Ab1 G2S2 Ab1 G2S2(lo) сиаловая кислота, которая первоначально была присоединена, затем утрачена при хранении, возможно, из-за загрязнения сиалидазой. Анализы показывают, что только 30% Fc-олигосахаридов в Ab1 G2S2(lo) содержат сиаловую кислоту, в то время как в Ab1 G2S2(hi) ~95% содержат сиаловую кислоту.
Гликановые структуры препаратов Ab анализируют различными способами. Для того, чтобы выполнить анализ MALDI-TOF-MS исходных Ab IgG, образцы IgG вносят в буфер 10 мМ трис-HCl, рН 7,0 и доводят концентрацию до ~1 мг/мл буфера. Примерно 2 мкл раствора IgG смешивают с 2 мкл матричного раствора (матричный раствор получают, растворяя 10 мг синапиновой кислоты в 1,0 мл 50% ацетонитрила в воде, содержащей 0,1% трифторуксусной кислоты) и 2 мл полученного раствора загружают на мишень и сушат на воздухе. MALDI-TOF-MS получают с использованием прибора Voyager DE от Applied BioSystems (Foster City, CA).
Для того, чтобы осуществить анализ MALDI-TOF-MS высвобожденных Fc-глканов, образцы IgG (~50 мкг) до и после реакции гликозилирования in vitro гидролизуют PNGase F в 10 мМ буфере трис-HCl (50 мкл), рН 7,0, в течение 4 часов при 37°С. Гидролиз останавливают, подкисляя реакционную смесь 50% уксусной кислотой (~5 мкл), и затем смесь пропускают через колонку с катионообменной смолой, как описано ранее (Papac et al., 1996; Papac et al., 1998; Raju et al., 2000). Такие образцы, содержащие смесь кислых и нейтральных олигосахаридов, анализируют MALDI-TOF-MS по типу с положительными и отрицательными ионами, как описано в тех же источниках (Papac et al., 1996; Papac et al., 1998; Raju et al., 2000), с использованием прибора Voyager DE от Applied BioSystems (Foster City, CA).
Анализ гликанов ВЭЖХ осуществляют, гидролизуя образцы IgG (~50 мкг) в 10 мМ буфере трис-HCl (50 мкл), рН 7,0, при 37°С в течение 4-8 часов. Дериватизацию высвобожденных олигосахаридов антраниловой кислотой (2-аминобензойная кислота) осуществляют так, как описано (см. Anumula K.R., Anal. Biochem., 2000, Jul, 15, 283(1): 17-26). Коротко, сначала получают раствор 4% тригидрата ацетата натрия (мас./об.) и 2% борной кислоты (мас./об.) в метаноле. Затем получают свежий раствор реагента дериватизации, растворяя ~30 мг антраниловой кислоты (Aldrich) и ~20 мг цианоборогидрида натрия (Aldrich) в 1,0 мл раствора в метаноле ацетата-бората натрия. Полученные из IgG олигосахариды (<3 нмоль в 20-50 мкл воды) смешивают с 0,1 мл раствора реагента с антраниловой кислотой (АА) в 1,6 мл флаконах для замораживания из полипропилена с навинчивающейся крышкой с уплотнительными кольцами (Sigma) и плотно закрывают. Флаконы греют при 80°С в печи или нагревательном блоке (Reacti-Therm, Pierce) в течение 1-2 часов. После охлаждения флаконов до комнатной температуры образцы разбавляют водой для доведения объема до ~0,5 мл. Дериватизированные олигосахариды очищают с использованием колонок с NAP-5.
Пример 2. Связывание с клеточными Fc-рецепторами с низкой аффинностью
Из нескольких типов Fc-рецепторов на эффекторных клетках рецепторы Fcгамма типов II и III считаются рецепторами с низкой или промежуточной аффинностью. Как правило, мономерное связывание может быть при слишком низким для детектируемой аффинности или на очень низких уровнях. Например, связывание мономерного IgG с Fcгамма типа IIA трудно измерить. Такие рецепторы функционируют как связывающие иммунные комплексы, которые из-за их мультивалентной природы связываются сильнее, вероятно, из-за низкой скорости высвобождения комплекса.
Человеческие клетки К562, экспрессирующие FcγRIIA как единственный Fcγ-рецептор, используют в двух типах анализа связывания для того, чтобы проверить, влияют ли изменения содержания сиаловой кислоты в Fc-гликане на связывание с таким человеческим Fcγ-рецептором с низкой аффинностью. Для того, чтобы получить достаточную авидность связывания с FcγRIIA, который имеет низкую аффинность в отношении мономерного IgG, получают иммунные комплексы смешиванием испытываемых Ab против TNF с гомотримерным TNF в молярном отношении 2:1, отношении, которое, как показано, приводит только к следовым количествам свободных Ab или свободного TNF. Зависимость иммунных комплексов поясняется, когда связывание одних меченых радиоизотопами Ab2 с клетками К562 не поддается обнаружению в концентрациях до 1 мкг/мл, но комплексы Ab2:TNF показывают существенное связывание при 0,02 мкг/мл (данные не приводятся).
Формат конкурентного связывания. Получают два набора иммунных комплексов IgG, меченый комплекс, содержащий человеческие антитела IgG1 с иррелеавантной специфичностью, в комплексе с нечеловеческими Ab, специфичными к анти-V участку, и Ab5. Для того, чтобы создать меченый комплекс, химерные моноклональные Ab с V-участками хомяка с человеческим IgG1 и константными областями легкой цепи каппа иодируют с использованием реагента IODO-GEN, как описано ранее (Knight et al., 1993). Затем подмешивают крысиные моноклональные Ab IgG2a, специфичные для идиотипа V-участка химеры хомяк-человек, в молярном отношении 1:1 в PBS в течение 30 мин для образования меченых радиоизотопами иммунных комплексов. Показано, что крысиные анти-Id не вносят вклад в связывание FcγRIIA непосредственно, как в случае, когда получают комплексы с деглигликозилированными химерами хомяк-человек, и происходит слабое связывание; в то время как комплексы с немодифицированными химерными Ab показывают высокие уровни связывания (данные не приводятся). Кроме того, отсутствует детектируемая перекрестная реактивность между агентами, используемыми для получения отдельных иммунных комплексов, которая может показать, что один иммунный комплекс может связываться с другим иммунным комплексом (данные не приводятся).
Для испытываемых комплексов варианты Ab1 с сиаловой кислотой смешивают с гомотримером человеческим TNF в молярном отношении 2:1 (как показано анализом методом светорассеяния, приводящем к очень небольшому количеству несвязанных Ab плюс несвязанный TNF) в PBS при комнатной температуре в течение 30 минут. В одном наборе экспериментов сравнивают друг с другом комплексы природных вариантов Ab1 с 20 и 29% сиаловой кислоты. В другом наборе экспериментов комплекс Ab1-29:TNF сравнивают с препаратом, усиленным колонкой с лектином, Ab1-43:TNF. В обоих случаях контрольным комплексом является Ab1-Gno:TNF, в котором антитело освобождено от гликана ферментативно.
Человеческие клетки К562 высевают в количестве 3х105 клеток/лунка в 96-луночные планшеты в IMDM, 5% FBS. Добавляют фиксированное количество меченого радиоизотопами комплекса антител к различным количествам комплекса испытываемых антител и объединенную смесь добавляют к клеткам К652 таким образом, чтобы в каждой лунке содержался иодированный комплекс антител в конечной концентрации 0,1 мкг/мл. Планшеты инкубируют в течение 16-18 часов при 4°С, после чего несвязанные Ab удаляют промывкой 3 раза IMDM, 5% FBS, и определяют число импульсов, связанных с клетками, с использованием счетчика гамма-квантов.
Результаты. Возрастающие количества немеченых иммунных комплексов-конкурентов ингибируют по возрастающей связывание мечеными радиоизотопами иммунными комплексами. Варианты с сиаловой кислотой немодифицированные Ab1 (сиалилированные на 29%) и Ab1 МААВ (сиалилированные на 43%) показывают, что в случае комплекса с более сиалалированными Ab требуются концентрации в 5-10 раз более высокие, чем в случае менее сиалилированных Ab, для того, чтобы получить одинаковую степень связывания с FcγRII (фиг.4А). В случае природных вариантов Ab1, различающихся содержанием сиаловой кислоты на 9% (20 и 29), различие составляет примерно в более чем 4-кратную большую авидность в случае менее сиалилированного препарата (не показано). Таким образом, наличие изомерной формы NGNA сиаловой кислоты как результата рекомбинантной экспрессии в клетке-хозяине миеломы мыши на данном человеческом IgG1 уменьшает авидность иммунных комплексов в отношении человеческого FcγRII.
Связывание иммунных комплексов с клетками К562. Испытываемые образцы Ab1 смешивают с меченым 125I человеческим TNF в фиксированном молярном отношении 2:1, и затем различные количества полученного иммунного комплекса добавляют к 3×105 клеток К562 в 96-луночном культуральном планшете. Сравнение комплексов Ab1 G2:TNF (несиалилированные Ab) с комплексами Ab1 G2S2(hi):TNF (полностью сиалилированные Ab) показывает, что полностью сиалилированные Ab связываются со значительно меньшей авидностью, причем в случае высоко сиалалированных вариантов требуются в 10 раз более высокие концентрации, чем в случае асиалилированных вариантов для достижения одной и той же степени связывания (фиг.4В). Такие результаты показывают, что наличие изомера NANA сиаловой кислоты, введенного ферментатиными модификациями in vitro, снижает авидность антител в отношении человеческого FcγRII, что можно связать с уменьшением аффинности связывания с мишенью (TNF), причем посредством этого комплексы Ab:TNF становятся менее устойчивыми или снижается аффинность константной области в отношении Fc-рецептора, или происходит и то и другое.
Связывание Ab с клеточным FcγRIIIa. Для того, чтобы проанализировать связывание FcγRIIIa на природных киллерных клетках (NK), человеческие РВМС выделяют так, как описано выше, и из РВМС выделяют клетки NK магнитным клеточным сортингом с использованием набора NK Cell Isolation Kit (Milteyi Biotec). Клетки NK культивируют в течение ночи в 96-луночном планшете в количестве 1×105 клеток на лунку в среде DMEM с 10% FBS при 37оС с 5% СО2. Метят mAb против FcγRIIIa 3G822 (BD Biosciences Pharmingen) 125I с использованием пробирок с иодом (Iodogen tubes, Pierce) до специфической активности 11 мкКи/мкг. Иодированные mAb 3G8 предварительно смешивают с различными количествами немеченых конкурентных Ab в DMEM, 10% FBS, и смесь Ab добавляют к клеткам NK до конечной концентрации 0,3 мкг/мл иодированных 3G8. Клетки инкубируют при 4°С в течение 16 час и затем несвязанный IgG удаляют, промывая 4 раза PBS. Число СРМ (импульсов), связанных с клетками, определяют с использованием счетчика гамма-квантов.
Клетки U-937 (без предварительной обработки для усиления экспрессии FcγR), культивированные в среде RPMI 1640 с добавлением 2 мМ L-глутамина, 1 мМ пирувата натрия и 10% FBS (среда для U-937), высевают в 96-луночный планшет с получением 3×105 клеток на лунку в 50 мкл среды для U-937. Ab2 (человеческий IgG1) метят 125I до специфической активности 17 мкКи/мкг. Иодированные Ab Ab2 предварительно смешивают с различными количествами образцов немеченых конкурентных Ab2 в среде для U-937. Затем 50 мкл смеси Ab добавляют к 50 мкл клеток U-937 для получения конечной концентрации 0,2 мкг/мл иодированных Ab3 во всех лунках. Клетки инкубируют при 4°С в течение 16 час и немеченые Ab удаляют, промывая три раза средой для U-937. Число СРМ, связанных с клетками, определяют с использованием счетчика гамма-квантов.
Для того, чтобы проверить, показывают ли варианты Ab различную аффинность в отношении FcγRIIIa, свежевыделенные клетки NK берут у здоровых доноров и используют в опытах по конкурентному связыванию с участием меченых радиоизотопами mAb3 3G8 - Ab против FcγRIIIa, конкурирующих за связывание с Fc, и немеченых Ab как конкурентов. Используют свободные не включенные в комплексы Ab вместо иммунных комплексов (которые, как правило, показывают значительно большее связывание с FcγRIIIa), так что результаты не будут искажаться различиями в устойчивости самих растворимых комплексов, на которые может влиять содержание Fc-сиаловой кислоты (неопубликованные данные авторов изобретения). Результаты показывают, что более сиалилированный природный вариант Ab1 Ab1-29 имеет пониженную аффинность в отношении FcγRIIIa на клетках NK, причем требуется его концентрация в 4 раза большая, чем Ab1-20, для достижения одной и той же степени связывания (фиг.5А). Существует подобное различие с природными вариантами Ab5, когда требуется концентрация Ab5-26 в 5 раз большая, чем Ab5-0, для конкуренции против mAb3 3G8 одной и той же степени (фиг.5В). Подобные результаты получают в каждом эксперименте с использованием клеток NK от, по меньшей мере, двух других доноров крови (данные не приводятся; аллотип FcγRIIIa не определен). Такие результаты показывают, что более высокие уровни сиалилирования могут снизить аффинность IgG в отношении FcγRIIIa и, следовательно, почти определенно вносят вклад в наблюдаемое снижение активности ADCC.
Однако, когда один и тот же эксперимент осуществляют с парами вариантов, полученных фракционированием на лектине, видно, что более сиалилированные варианты связываются с FcγRIIIa так же хорошо, и вероятно, чуть лучше, чем менее сиалилированные варианты (фиг.5С и 5D). Причинами различных результатов с двумя парами природных вариантов и двумя парами вариантов, полученных на лектине, неизвестны, но весьма возможно, что существуют различия в местоположении имеющихся остатков сиаловой кислоты.
Пример 3. Анализы ADCC in vitro
Клетки-мишени для Ab против TNF включают клеточную линию Sp2/0 миеломы мыши, которая устойчиво экспрессирует на своей клеточной поверхности рекомбинантный человеческий TNF, который остается в трансмембранной форме из-за введения делеции аминокислот 1-12 зрелого TNF (Perez et al., 1990). Клетки К2 культивируют в среде Исков, содержащей инактивированную нагреванием FBS, 2 мМ L-глутамина, 1 мМ пирувата натрия, 0,1 мМ заменимых аминокислот и МНХ. Культуральные среды и добавки закупают у Gibco (Invitrogen). Клетки пересевают 1:5 каждые 2-3 дня. В день анализа клетки К2 центрифугируют и промывают один раз PBS. Клетки доводят до примерно 1×106 клеток/мл культуральной средой и добавляют 15 мкл флуоресцентного метящего реагента BATDA (из набора Delfia EuTDA Cytotoxicity Reagent Kit, Perkin-Elmer Life Sciences) к 5 мл клеток (Blomberg et al., 1996). Клетки инкубируют в течение 30 минут при 37оС, затем дважды промывают PBS при 1000 об/мин, 5 мин. Непосредственно перед смешиванием с эффекторными клетками РВМС клетки-мишени центрифугируют и ресуспендируют в концентрации 2×105 клеток/мл в среде Исков, содержащей 1% BSA.
Эффекторные клетки РВМС выделяют после забора крови у здоровых доноров в гепаринизированные емкости (vacutainers) и двукратно разбавляют PBS. Тридцать мл разбавленной крови наносят поверх 15 мл фиколл-пака (Amersham, Uppsala, Швеция) в 50-мл конической пробирке и центрифугируют при 1500 об/мин 30 мин при комнатной температуре (RT). Поверхностный слой (желтый слой), содержащий РВМС, собирают и дважды промывают PBS и центрифугируют при 1300 об/мин, 10 мин, RT. Клетки ресуспендируют в среде Исков, содержащей 5% инактивированной нагреванием FBS, 2 мМ L-глутамина, 1 мМ пирувата натрия и 0,1 мМ заменимых аминокислот. РВМС активируют в течение приблизительно 4 часов при 37°С, 5% СО2, инкубацией на 100-мм чашках Петри (Corning), сенсибилизированных ОКТ3 (10 мкг/мл в PBS, Ortho Pharmaceutical), в течение ночи при 4°С и ополаскивают PBS. РВМС собирают, промывают один раз средой Исков, содержащей 1% BSA, считают и ресуспендируют до приблизительно 1×107 клеток/мл.
Испытываемые образцы Ab, включая отрицательный контроль вариант Ab1 Gno, серийно разводят в среде Исков - 1% BSA. Добавляют пятьдесят микролитров клеток-мишеней (~10000) и 100 мкл антител в круглодонные лунки 96-луночного планшета (Corning). К смеси добавляют пятьдесят (50) микролитров эффекторных клеток (~500000) и планшет центрифугируют при 1000 об/мин в течение 5 мин, RT. Отношение Е:Т, как правило, составляет 50:1, однако иногда используют 35:1. Для фоновой флуоресценции лунки инкубируют с эффекторными клетками, клетками-мишенями и средой. Для максимальной флуоресценции в фоновые лунки добавляют по 10 мкл раствора для лизиса (из набора Delfia EuTDA Cytotoxicity Reagent Kit). Для анализа ADCC клетки инкубируют при 37°С, 5% СО2, в течение приблизительно 2 часов. Переносят по 20 мкл супернатанта в лунки 96-луночного планшета с плоскодонными лунками (Corning). Добавляют 200 мкл раствора европия (набор Delfia EuTDA Cytotoxicity Reagent Kit) и планшет помещают на планшетный шейкер на 10 минут при RT. Флуоресценцию измеряют флуориметром с разрешением во времени, EnVision Instrument (Perkin-Elmer Life Sciences). Процент специфического лизиса в каждом образце вычисляют по следующей формуле: % специфического высвобождения = ([экспериментальное высвобождение - спонтанное высвобождение] - [максимальное высвобождение - спонтанное высвобождение])×100.
Начальные оценки действия сиаловой кислоты фокусируются на активности ADCC in vitro двух пар природных вариантов. Ab1-29 и Ab1-20 инкубируют при различных концентрациях с мечеными европием экспрессирующими Ag1 клетками-мишенями. Как видно на фигуре 6А, существует четкое различие в цитотоксической активности, когда в случае Ab1-29 с более высокими уровнями Fc-сиалилирования требуется приблизительно в 7 раз большие концентрации, чем концентрации Ab1-20, для того, чтобы запустить лизис клеток в одной и той же степени. Результаты показывают, что обогащенные сиалилированными гликоформами Ab1 МААВ менее эффективны, чем немодифицированные Ab1 PBS. Приблизительно в 3 раза больше требуется материала Ab1 МААВ-43% для достижения той же степени лизиса, что с образцом Ab1 PBS-29%. Эксперименты с экспрессирующими Ag5 клетками-мишенями показывают такую же картину, как для пары природных вариантов Ab2. Для того, чтобы достичь такой же степени лизиса клеток, как для варианта Ab2-0 с недетектируемой сиаловой кислотой, концентрация Ab2-26 требуется приблизительно в 6 раз большая (как видно на фиг.6В). Таким образом, влияние изменения природного гликозилирования на такое измерение ADCC не является Ab- или мишенеспецифическим.
В характерном эксперименте по сравнению активности ADCC субпартий Ab1, отличающихся содержанием в них сиаловой кислоты после фракционирования на лектине, Ab1 МААВ (сиалилированые на 43%) сравнивают с партией немодифицированных Ab1, из которой они получены (Ab1 PBS). Во втором эксперименте по сравнению субпартий Ab1, отличающихся содержанием в них сиаловой кислоты, Ab1 WGAT (сиалилированые на 29%), Ab1 WGAR (сиалилированые на 40%) и Ab1 WGAB (сиалилированые на 32%) сравнивают друг с другом.
Результаты анализа также показывают обратное соотношение между содержанием сиаловой кислоты и эффективностью в анализе ADCC независимо от способа, которым получают Ab (фиг.6С). Иными словами, Ab1 WGAT, которые содержат примерно такое же количество сиаловой кислоты, как и немодифицированные Ab1 показывают такую же активность, как немодифицированные Ab1. Однако полученные фракции WGA теряют эффективность с возрастанием содержания сиаловой кислоты (фиг.6С).
В эксперименте сравнивают два образца с более заметными различиями по содержанию сиаловой кислоты - ферментативно модифицированные Ab1 G2 (сиалилированые на 0%) и Ab1 G2S2(hi) (сиалилированые на ~95%). Свежие РВМС извлекают центрифугированием в градиенте плотности в фиколл-паке. Предварительно инкубируют 5×105 РВМС в объеме 100 мл в течение приблизительно 10 минут с различными количествами необработанных Ab1, Ab1 G2S2(hi) (полностью галактозилированные и сиалилированные) или Ab7, родственными по изотипу, Ab отрицательного контроля. Клетки К2, экспрессирующие связанный с поверхностью рекомбинантный человеческий TNF, используют в качестве мишеней, метя их 200 мКи 51Cr. Меченые клетки добавляют к смеси РВМС/Ab, центрифугируют при 1000 об/мин в течение 1 мин и инкубируют при 37°С в течение 4 часов. Известно, что такое время инкубации (4 часа) обнаруживает первичный лизис клеток, вызванный клетками NK (в пределах популяции клеток РВМС), которые экспрессируют FcγRIIIA, а не макрофагами, которые обычно экспрессируют FcγRI (CD64), FcγRIIA (CD32A) и FcγRIIIA (CD16A). Затем определяют количество радиоактивности в клеточных супернатантах с использованием счетчика Topcount. Приведенные результаты (фиг.6D) представляют два независимых эксперимента, осуществленных с использованием РВМС от разных доноров, и показывают более чем 10-кратное изменение в эффективности лизиса клеток между Ab, которые полностью сиалилированы, и антителами, которые почти десиалилированы.
Также сравнивают другие пары препаратов Ab в анализе ADCC. Фракции, полученные на лектине WGA из галактозилированных Ab2, оценивают в анализах ADCC с использованием клеток-мишеней, экспрессирующих Ag2. Снова более сиалилированный матерал является менее активным, хотя имеется только 4-кратное различие в величинах их ЕС50, несмотря на существенное различие в их содержании сиаловой кислоты (5% против 67%). Для сравнения, фракции, полученные на лектине WGA из Ab1, показывают, что для достижения одной и той же степени лизиса клеток концентрации сиалилированного на 41% варианта требуются приблизительно в 6 раз большие, чем варианта, сиалилированного на 29%.
Такие результаты для всех трех испытываемых Ab последовательно показывают, что более высокие уровни Fc-сиаловой кислоты ассоциируются с пониженной активностью ADCC. Хотя и не количественно, различия между изменением величины активности ADCC и содержанием сиаловой кислоты в препаратах Ab являются согласующимся соотношением в пределах панели четырех вариантов Ab1, где величины ЕС50 типично составляют 0,3 нг/мл, 2 нг/мл, 2 нг/мл и 10 нг/мл для Ab1-20, Ab1-29, Ab1-WGA-29 и Ab1-WGA-41, соответственно. Результаты с лектиновыми фракциями также подтверждают, что сиалилированные препараты Ab содержат молекулярные группы с различными уровнями активности ADCC. Примечательно, что, за исключением Ab3-0 и Ab3-26, варианты, анализируемые в данном случае, не склонны показывать различия в максимальном уровне достигаемого лизиса.
Так как в данном способе измерения активности ADCC, главным образом, посредниками являются FcγRIIIA-положительные клетки NK, результаты подразумевают, что в то время как наличие сиаловой кислоты в Fc-олигосахаридах усиливает связывание с FcγRI, ее присутствие существенно ослабляет связывание с FcγRIIIA.
Пример 4. Связывание с высокоаффинным Fc-рецептором
Связывание с испытываемыми Ab, различающимися содержанием сиаловой кислоты, с высокоаффинным человеческим Fc-рецептором FcγRI (CD64) измеряют с использованием формата конкурентного связывания на клетках U-937 - клеточной линии моноцитов человека. Клетки U-937 культивируют в среде RPMI 1640 с добавлением 2 мМ L-глутамина, 1 мМ пирувата натрия и 10% FBS в Т-колбах и сохраняют в инкубаторе с 5% СО2 при 37°С. Химерные Ab мышиный/человеческий IgG1 Ab2 иодируют с использованием пробирок для иодирования, предварительно сенсибилизированных IODO-Gen, до специфической активности 17,2 мКи/мг. Клетки U-937 ресуспендируют при 6×106 клеток/мл в свежей культуральной среде и затем высевают в 96-луночные культуральные планшеты с фильтрами, Millipore, при плотности 3×105 клеток на лунку. Клетки предварительно не обрабатывают для того, чтобы вызвать более высокую экспрессию FcγR. Иодированные Ab2 предварительно смешивают с различными количествами немеченых Mab-конкурентов (испытываемые образцы) с использованием культуральной среды как разбавителя, в объеме 50 мкл. Затем смеси добавляют к 50 мкл культуры клеток U-937 и получают конечную концентрацию иодированных Ab2 0,2 нг/мл. Затем клетки инкубируют при 4°С в течение 16 часов. Несвязанный IgG удаляют трехкратной промывкой средой с отсасыванием с использованием системы вакуумирования планшетов. Число импульсов, связанных с клетками, определяют с использованием счетчика гамма-квантов.
Фиг.7А показывает, что по сравнению с Ab1 G2 (без сиаловой кислоты) Ab1 G2S2(hi) (сиалилированные на ~95%) связываются с высокоаффинным FcR (CD64) на клетках U-937 с 5-10-кратной более высокой аффинностью, т.е. Ab1 G2S2(hi) требуется только в одной пятой-одной десятой концентрации для получения той же самой степени ингибирования связывания иодированных Ab2. Ab1 G2 не показывают заметного различия с необработанными Ab1 (данные не приводятся), причем последние представляют собой гетерогенную смесь различных гликоформ, большинство которых содержит меньше галактозы (т.е. гликоформы G0 и G1), чем образец Ab1 G2.
Фиг.7В показывает, что две разные партии Ab3, различающиеся по количеству заряженных олигосахаридных групп (групп, содержащих сиаловую кислоту), составляющих или 2% или 42% от общего количества олигосахаридов, подобным образом показывают, что партия, характеризующаяся наличием большего содержания сиаловой кислоты, имеет более высокую аффинность в отношении FcγRI.
После регистрации уменьшенного связывания с FcγRIIIa клеток NK препаратами антител с более высоким содержанием сиаловой кислоты для двух пар природных вариантов гликозилирования Ab1 и Ab5 (пример 3, фиг.5, А и В) рассматривается возможность того, что эффект имеет место из-за простого электростатического отталкивания между отрицательно заряженной сиаловой кислотой и отрицательно заряженной клеточной поверхностью. Однако обратное действие содержания сиаловой кислоты на аффинность связывания для рецептора FcγRI на человеческих клетках U-937 не дает такую же картину для Ab5 или других Ab (данные не приводятся).
Следует отметить, что в то время как два образца Ab1 различаются по отсутствию/присутствию формы NANA сиаловой кислоты, два образца Ab3, как полагают, различаются по количеству формы NGNA сиаловой кислоты (полученной в мышиных клетках-хозявах).
Пример 5. Измерение времени полужизни в сыворотке
В настоящем примере Fc-содержащий слитый белок, включающий N-концевой пептид, слитый с последовательностью вариабельной области антитела, и домены шарнирный, СН2 и СН3 человеческого IgG1, экспрессированный в клетках миеломы мыши, обрабатывают для образования полностью сиалилированной (G2S2) формы. Здоровым самкам крысы CD1 (4 на группу обработки) дают внутривенную инъекцию или немодифицированной формы FcP1, содержащей Fc-олигосахариды, сиалилированные на 5%, или группам самок крысы CD1 инъецируют отдельно внутривенно полностью сиалилированную версию (сиалилированные на ~95%). Кровь собирают из заглазничных сосудов через 1 час, 5 часов, 24 часа, 72 часа, 7 дней, 14 дней и 21 день, и затем последний отбор крови у животных проводят сердечной пункцией под наркозом СО2 на 28 день. Получают сыворотку из образцов крови и измеряют концентрацию человеческого Fc в сыворотке с использованием колориметрического ELISA. Коротко, сначала 96-луночные планшеты для EIA сенсибилизируют поликлональными козьими антителами против человеческого Fc. Различные разведения образцов сыворотки инкубируют в лунках в течение 1 часа при комнатной температуре. Несвязанный белок удаляют промывкой, и связанный человеческий Fc детектируют с использованием конъюгированных с ферментом козьих антител против человеческого IgG и затем с помощью соответствующих цветных субстратов.
Результаты исследования показаны на фиг.8. Вычисленная площадь под кривой (AUC) составляет 95±1,6 день·нг/мл×10-3 для немодифицированных антител и, соответственно, 48±1,9 день·нг/мл×10-3. Это показывает, что более высокая степень сиалилирования Fc-олигосахарида ассоциируется с более высокой скоростью клиренса у здоровых крыс.
Во втором эксперименте здоровым мышам инъецируют однократной дозой в 3 мг/кг ферментативно модифицированные Ab2, которые являются или полностью асиалилированными (G2) или полностью сиалилированными (G2S2). Человеческий Fc в сыворотке контролируют и измеряют с использованием колориметрического ELISA, как описано выше. Результаты данного эксперимента приводятся на фиг.9. Спустя приблизительно одну неделю Ab2 G2S2 начинают быстрее выводиться из сыворотки мышей и через 20 дней Ab2 G2S2, оставшихся в сыворотке, приблизительно в 1000 раз меньше, чем концентрация Ab2-G2.
Клиренс вариантов Ab1 с сиаловой кислотой из большого круга кровообращения у мышей. Другое прямое измерение влияния содержания сиаловой кислоты осуществляют путем количественной оценки скорости выведения отдельных гликозилированных видов из сыворотки после инъекции образца, содержащего смесь гликанов, присоединенных к Ab1.
Один и тот же препарат гетерогенно-гликозилированных Ab1 инъецируют i.p. 18 здоровым 8-10-недельным мышам Balb/c в дозе 20 мг/кг. Берут кровь у 6 мышей в день 3, у других 6 мышей в день 14 и у последних 6 мышей в день 28. Получают сыворотку из каждого образца крови и Ab1 очищают повторно от сыворотки с использованием колонки против аффинности к Id, специфичной для V-областей Ab1. Затем анализируют структуры Fc-гликанов образцов повторно очищенных Ab1 анализами ВЭЖХ и определяют относительную долю различных гликанов так, как описано в данном описании ранее.
Обнаруживают, что галактозилированная гликоформа, утратившая сиаловую кислоту (G2S0), остается у мышей в своем относительном количестве в течение 4 недель, в то время как гликоформы Ab, содержащие гликаны с 1 остатком сиаловой кислоты (G2S1), и гликоформы с 2 остатками сиаловой кислоты (G2S2) выводятся с большей скоростью. Таким образом, полностью сиалалированные Fc-содержащие белки имеют более короткое время полужизни, чем асиалилированные или частично сиалилированные композиции.
Пример 6. Содержание сиаловой кислоты и авидность антител
Результаты, описанные в данном примере, подтверждают теорию, что изменение содержания сиаловой кислоты Fc-гликана Fc-домена (димеризованный шарнир-СН2-СН3) будет влиять на весь белок. Что касается двухвалентности антител и слитых белков, включающих гликозилированный Fc, такое влияние может проявляться в авидности белка в отношении специфической мишени. Эксперименты в данном примере осуществляют для проверки такой теории и также демонстрации специфического влияния содержания сиаловой кислоты на аффинность связывания мишени.
Связывание с антигеном клеточной поверхности. Те же самые Ag-экспрессирующие клеточные линии, используемые в описанных выше анализах активности ADCC, используют в анализах связывания для проверки различий между вариантами с сиаловой кислотой в их авидности связывания антигенов. Анализы осуществляют в конкурентном формате, в котором одни из меченых радиоизотопами Ab (любые из Ab1, Ab2 и Ab5), поддерживаемые в фиксированной концентрации, инкубируют с Ag-экспрессирующими клетками в присутствии различных количеств немеченых испытываемых Ab. Иодированные Ab, полученные по способу Iodogen, обычно имеют специфическую активность 10 мкКи/мкг.
Клетки, экспрессирующие на поверхности TNF, высевают в 96-луночные культуральные планшеты в количестве 50000 клеток на лунку, а клетки, экспрессирующие Ag2, в количестве 180000 клеток на лунку, в среде IMDM с 5% FBS. Соответствующие меченые 125I Ab предварительно смешивают с определяющими титр количествами испытываемых Ab и смесь добавляют к соответствующим Ag-экспрессирующим клеткам. Планшеты инкубируют при RT в течение 2 часов для связывания Ab с клетками. Затем клетки три раза промывают IMDM, 5% FBS, для удаления несвязанных Ab и определяют число импульсов, связанных с клетками, с использованием счетчика гамма-квантов.
В случае вариантов Ab5 Ab5-экспрессирующие клетки высевают в 96-луночные культуральные планшеты в количестве 186000 клеток на лунку в 50 мкл DMEM, 10% FBS. Меченые 125I Ab2 предварительно смешивают с определяющими титр количествами испытываемых Ab и 50 мкл смеси добавляют к Ag-экспрессирующим клеткам. Планшеты инкубируют при 4оС в течение 16 часов для связывания Ab с антигеном на клетках. Затем клетки три раза промывают DMEM, 10% FBS, для удаления несвязанных Ab и определяют число импульсов, связанных с клетками, с использованием счетчика гамма-квантов. Образцы испытывают при двукратном или четырехкратном повторении и приведенные результаты представляют 3 или 4 независимых эксперимента. Различие в связывании между указанными испытываемыми образцами является значимым (P<0,0001 для графиков а, с и d) по определению по F-критерию дополнительной суммы квадратов.
Результаты приводятся на фиг.10, A-D: связывание мечеными радиоизотопом Ab1 с Ag1-экспрессирующими клетками в присутствии немеченых природных вариантов Ab1 в качестве конкурентов (фиг.10А); связывание мечеными радиоизотопом Ab5 с Ag5-экспрессирующими клетками в присутствии немеченых природных вариантов Ab5 в качестве конкурентов (фиг.10В); связывание мечеными радиоизотопом Ab1 с Ag1-экспрессирующими клетками в присутствии немеченых полученных на лектине вариантов Ab1 в качестве конкурентов (фиг.10С); связывание мечеными радиоизотопом Ab3 с Ag3-экспрессирующими клетками в присутствии немеченых полученных на лектине вариантов Ab3 в качестве конкурентов (фиг.10D).
Связывание Ab с твердофазным лигандом. Рекомбинантный растворимый TNF или анти-Id2 наносят на планшеты для EIA, добавляя 50 мкл Ag или Ab анти-Id в количестве 1 мкг/мл в PBS в каждую лунку и инкубируя планшеты при 4°С в течение ночи. Лунки промывают и затем предварительно обрабатывают 50 мкл 1% BSA, 0,125% желатина в PBS в течение 1 часа при RT для минимизации неспецифического связывания. Меченые 125I Ab1 или меченые 125I Ab3 предварительно смешивают с определяющими титр количествами препаратов соответствующих испытываемых Ab в IMDM, 5% FBS и 50 мкл смеси добавляют в лунки с покрытием-мишенью. Конечная концентрация меченых радиоизотопом Ab составляет 100 нг/мл во всех лунках. Планшеты инкубируют при RT в течение 2 часов для связывания Ab с мишенями-покрытиями. Клетки промывают для удаления несвязанных Ab и определяют число связанных импульсов с использованием счетчика гамма-квантов.
Связывание Ab, нанесенных на планшеты, с растворимым антигеном. Наносят на 96-луночные планшеты варианты с сиаловой кислотой Ab1 или Ab3 и затем инкубируют с различными количествами меченого радиоизотопом растворимого антигена для определения (а) связывания меченого радиоизотопом растворимого Ag1 с нанесенными на планшет природными вариантами Ab1, (b) связывания меченого радиоизотопом растворимого Ag1 с нанесенными на планшет фракционированными на лектине вариантами Ab1 и (с) связывания меченого радиоизотопом растворимого Ag3 с нанесенными на планшет фракционированными на лектине вариантами Ab3. Параллельно осуществляют инкубацию с меченым радиоизотопом Ag и 100-кратным избытком немеченого Ag для определения неспецифического связывания. Образцы испытывают при трехкратном повторении. Варианты Ab2 не анализируют из-за недоступности растворимого Ag2.
Статистический анализ. Различие в эффективности между вариантами антител анализируют путем сравнения кривых с использованием совместных 4-параметрических логистических регрессий с общим минимумом, максимумом и наклоном после предварительного критерия для наклона и интервала, данного общим плато для нулевой концентрации (т.е. всегда предполагая, без испытания, общее «дно» для возрастающих кривых и общий верх для убывающих кривых). Проверку значимости осуществляют по F-критерию дополнительной суммы квадратов в GraphPad Prism v4. Величину Р<0,05 рассматривают как значимую. Анализы на СРМ взвешивают обратно пропорционально СРМ2, поскольку среднеквадратическое отклонение СРМ возрастает пропорционально его среднему (т.е., коэффициент вариации СРМ CV не связан со средним).
Результаты
Эксперименты по связыванию антигена, выполненные в конкурентном формате с одними и теми же Ag-экспрессирующими клетками-мишенями, которые использовали в анализах ADCC, неожиданно показали, что Ab1-29 сообразно связывают антиген клеточной поверхности с примерно в 3 раза меньшей аффинностью, чем Ab1-20 (фиг.10А). Напротив, Ab5-26 показывают аффинность, которая неотличима от аффинности Ab5-0 (фиг.10В). Такие же анализы, осуществленные с двумя парами вариантов, полученных на лектине, показывают схожие результаты для природных вариантов Ab1, т.е., в случае высоко сиалилированных Ab1-WGA-41 требуются концентрации в 4-6 раз большие, чем в случае менее сиалилированных Ab1-WGA-29, для достижения одной и той же степени конкурентного связывания (фиг.10С), и в случае более сиалилированных Ab2-GT-WGA-67 требуются концентрации в 4-6 раз большие, чем в случае менее сиалилированных Ab2-GT-WGA-5 (фиг.10D).
Представляет интерес, что такую же картину ослабленного связывания вариантами Ab1 и Ab2 с большей степенью сиалилирования также наблюдают в экспериментах, анализирующих связывание с мишенями (растворимым рекомбинантным антигеном или Ab анти-Id), иммобилизованными на 96-луночных планшетах для EIA (фиг.11, А и В). Такие результаты показывают, что различия в степени сиалилирования Fc могут влиять на связывание антигена так же, как FcγRIIIA, но что степень сиалалирования не влияет на связывание антигена всех Ab.
Из результатов по связыванию иммобилизованных мишеней полагают, что повышенное сиалалирование Fc может служить для снижения жесткости шарнирной области Ab. В случае связывания Ab1 и Ab2 с антигеном клеточной поверхности пониженная жесткость шарнира может привести больше к моновалентному связыванию и меньше бивалентному (высокая авидность) связыванию с антигеном, в зависимости от размещения антигенных детерминант на твердом носителе или клеточной поврехности. Шарнирная область Ab5 также может иметь уменьшенную жесткость, но жесткость может не потребоваться для таких Ab для достижения максимального связывания с Ag5.
Для того, чтобы разграничить, влияния ли жесткости Ab или изменения действительной аффинности связывания вызваны сиалилированием Fc, проверяют связывание Ab с растворимым лигандом как меру чисто аффинности моновалентного связывания. Результаты действительно показывают, что для трех пар вариантов с сиаловой кислотой, которые показывают различия в связывании с антигеном клеточной поверхности, не существует детектируемых различий, наблюдаемых между парами вариантов при их связывании с растворимыми мишенями (фиг.12, А-С). Все вместе такие результаты показывают, что различия в связывании с иммобилизованными мишенями (на клеточной поверхности или на сенсибилизованном планшете) имеют место не из-за различий в действительной аффинности между каждым плечом Fab и мишени. Следовательно, различия между вариантами с сиаловой кислотой Ab1 и Ab2 в их связывании с иммобилизованными мишениям имеют место из-за различий в степени бивалентного связывания с клетками.
Пример 7. Получение вектора для секреции сиалидазы
Сборка плазмиды, экспрессирующей сиалидазу. Нуклеотидную последовательность, кодирующую каталитический домен сиалидазы из Arthrobacter ureafaciens, остатки 40-535, инвентарный № AY934539, GenBank, синтезируют на основе такой последовательности. Гибрид синтезированный ген (SEQ ID NO: 2), кодирующий сигнальную последовательность человеческого гормона роста - каталитический домен Arthrobacter ureafaciens (SEQ ID NO: 1 с сигнальной последовательностью в виде первых 26 аминокислот, и каталитический домен оставшиеся 494 аминокислоты) клонируют в плазмиду р2815 с использованием уникальных сайтов рестрикции BamH I и Not I. Такая плазмида обладает промотором CMV, кодирующей последовательностью для сигнальной последовательности человеческого гормона роста для влияния на секрецию и геном устойчивости к неомицину для устойчивой селекции. Кодирующую последовательность для каталитического домена фермента, лигированную с сигнальной кодирующей последовательностью hGH (SEQ ID NO: 1), подтверждают расщеплением рестриктазами и секвенированием.
Временная и устойчивая трансфекция. Для временной трансфекции клетки НЕК293 трансфицируют 15 мкг очищенной плазмиды р3629 или контрольной плазмиды (полный вектор) с использованием липофектамина 2000. Плазмидную ДНК и 90 мкл липофектамина 2000 разводят в Optimem, объединяют и затем инкубируют в течение 20 минут при комнатной температуре. Затем добавляют коктейль для трансфекции к слитым на 70% клеткам НЕК293 в среде для выращивания в течение ночи. На следующий день среду для выращивания заменяют средой 293SFM и клетки инкубируют в течение 5 суток для сбора среды и анализа. Для устойчивой трансфекции клетки С168М подвергают электропорации 10 мкг р3629, линеаризированной рестриционным расщеплением Bgl II. Трансфицированные клетки сохраняют в среде для выращивания с 700 мкг/мл антибиотика генетицина для отбора стабильных трансфектантов. Клоны, устойчивые к антибиотику, размножают и анализируют на сиалидазу.
Анализ на сиалидазную активность. Сиалидазную активность анализируют с использованием 2'-(4-метилумбеллиферил)-α-D-N-ацетилнейраминовой кислоты. Флуорометрический анализ супернатанта жизнеспособоных клеточных культур выполняют, смешивая его с 200 мкл 150 мкМ 2'-(4-метилумбеллиферил)-α-D-N-ацетилнейраминовой кислоты в 100 мМ цитратфосфатном буфере, рН 6,5, при 37°С с последующим добавлением 2 мл 0,5 М раствора Na2CO3 для прекращения реакции. Возбуждение осуществляют при 366 нм, а испускание при 446 нм. Флуорометрические единицы нормализуют против числа жизнеспособных клеток. С другой стороны, сиалидазную активность в культуральной среде определяют инкубацией в течение ночи сиалилированного ремикада (Remicade) со средами от трансфицированных клеток и анализом на сиаловую кислоту, как описано ниже.
Определение сиаловой кислоты. Сиалидазную активность определяют, анализируя удаление сиаловой кислоты из очищенных антител после инкубации с супернатантами клеточных культур. Высвобождают N-соединенные олигосахариды обработкой образцов IgG (0,05-0,5 мг в 0,1 мл) PNGase F в 10 мМ трис-HCl буфере, рН 7,0, при 37°С в течение 4-6 час. Аликвоту такого раствора (~0,01 мл) пропускают через колонку, содержащую катионообменную смолу, и анализируют MALDI-TOF-MS как описано ранее. Оставшуюся часть образца подвергают восстановительному аминированию антраниловой кислотой и последующему анализу ВЭЖХ, как описано Anumula. Коротко, сначала получают раствор 4% тригидрата ацетата натрия (мас./об.) и 2% борной кислоты (мас./об.) в метаноле. Получают свежий реагент дериватизации, растворяя ~30 мг антраниловой кислоты (Aldrich) и ~20 мг цианоборогидрида натрия (Aldrich) в 1,0 мл раствора в метаноле ацетата-бората натрия. Полученные из IgG олигосахариды (<3 нмоль в 20-50 мкл воды) смешивают с 0,1 мл раствора реагента с антраниловой кислотой (АА) в 1,6 мл флаконах для замораживания из полипропилена с навинчивающейся крышкой с уплотнительными кольцами (Sigma) и плотно закрывают. Флаконы греют при 80°С в нагревательном блоке (Reacti-Therm, Pierce) в течение ~1 часа. После охлаждения флаконов до комнатной температуры образцы разбавляют водой для доведения объема до ~0,5 мл. Дериватизированные олигосахариды очищают так, как описано ранее.
Очистка антител. Экспрессированные рекомбинантные антитела из устойчиво трансфицированных клеток очищают аффинной хроматографией на белке А. Клеточные супернатанты разводят 10× буфером для белка А (0,2 М трис, 1,4 М NaCl, 10 мМ ЭДТК, рН 8,5) до 1× и очищают на 1-мл колонке с белком А. Элюированные антитела перед дальнейшим анализом диализуют в PBS, pH 7,2.
Анализ антителозависимой клеточной цитотоксичности (ADCC). Клетки-мишени для анализов, включающие Ab1 и Ab3, получают по Centocor трансфекций клеток миеломы мыши Sp2/0 трансмембранной формой Ag1 и Ag3, соответственно. Как экспрессирующие Ag1, так и экспрессирующие Ag3 клетки выращивают в IMDM, содержащей инактивированную нагреванием FBS, 2 мМ L-глутамина, 1 мМ пирувата натрия, 0,1 мМ заменимых аминокислот. Прилипающие клетки, экспрессирующие Ag2, получают от АТСС и выращивают в среде DMEM, содержащей 10% FBS, 2 мМ L-глутамина, 1 мМ пирувата натрия и 0,1 мМ заменимых аминокислот. Все клетки пересевают дважды в неделю и сохраняют в логарифмической фазе роста. Культуральные среды и добавки закупают у Gibco (Invitrogen).
В день анализа экспрессирующие Ag миеломные клетки-мишени центрифугируют и промывают один раз PBS. Прилипающие экспрессирующие Ag2 клетки-мишени извлекают трипсином и дважды промывают. Клетки доводят до 1×106 клеток/мл культуральной средой и к 5 мл клеток добавляют 15 мкл BATDA (реагент для флуоресцентного мечения (бисацетоксиметил)-2,2':6'2”-терпиридин-6,6”-дикарбоксилат) (из набора Delfia EuTDA Cytotoxicity Kit, Perkin-Elmer Life Sciences; Blomber K. et al.). Клетки инкубируют в течение 30 минут при 37°С, встряхивая время от времени, и затем дважды промывают средой. Непосредственно перед смешиванием с эффекторными клетками клетки-мишени центрифугируют и ресуспендируют в 2×105 клеток/мл в культуральных средах.
Эффекторные одноядерные клетки периферической крови (РВМС) извлекают из гепаринизированной крови здоровых доноров. Образцы крови разбавляют забуференным фосфатом физиологическим раствором (PBS) и выделяют РВМС центрифугированием в градиенте плотности фиколл-гипака (Amersham). После центрифугирования РВМС собирают, дважды промывают и держат в течение ночи в культуральной среде при 37°С с 5% СО2. На следующий день РВМС собирают, промывают и ресуспендируют в среде в 1×107 клеток/мл.
Для анализов цитотоксичности антитела, разведенные в 100 мкл культуральной среды, добавляют в 96-луночные планшеты с круглодонными лунками. К разведениям Ab добавляют пятьдесят мкл эффекторных клеток и 50 мкл клеток-мишеней, меченых BATDA, при соотношении эффекторных клеток к клеткам-мишеням 50:1. Планшет недолго центрифугируют для приведения эффекторов и мишеней в контакт друг с другом и затем инкубируют в течение 2 час при 37°С в атмосфере с 5% СО2. После инкубации трансфицируют 20 мкл супернатанта в лунки 96-луночного планшета с плоскодонными лунками и в каждую лунку добавляют 200 мкл усилителя раствора европия (из набора Delfia Cytotoxicity Kit). После встряхивания планшета в течение 10 мин измеряют флуоресценцию флуорометром с разрешением во времени (EnVision Instrument, Perkin-Elmer). Процент специфической цитотоксичности вычисляют как (экспериментальное высвобождение - спонтанное высвобождение)/максимальное высвобождение - спонтанное высвобождение)×100. Спонтанное высвобождение определяют, инкубируя мишени со средой вместо эффекторных клеток, и максимальное высвобождение определяют, инкубируя мишени с 10 мкл раствора для лизиса, содержащего диджитонин (из набора Delfia EuTDA Cytotoxicity Kit).
Результаты и обсуждение
Сконструирован экспрессирующий вектор р3629 (фиг.13), позволяющий экспрессировать каталитический домен сиалидазы А из Arthrobacter ureafaciens в клетках млекопитающих. Кодирующая последовательность для лидерной последовательности гормона роста человека оперативно связана с каталитическим доменом фермента для того, чтобы усилить внеклеточную секрецию сиалидазы. Клетки НЕК293 временно трансфицированы экспрессирующей плазмидой и собран супернатант для активности сиалидазы на очищенных антителах против TNFα. Антитела инкубируют в течение ночи с кондиционной средой от клеток, трансфицированных р3629, или контрольных трансфицированных плазмидой клеток. Анализ ВЭЖХ олигосахаридов, высвобожденных из антител после инкубации с кондиционным супернатантом, не обнаруживает их вовсе, за исключением гликанов, высвобожденных из контрольной трансфицированной исходной клеточной линии. Следовательно, сиалидазная активность присутствует только в супернатанте клеток, трансфицированных р3629, но не в супернатанте, трансфицированном контролем.
Пример 8. Коэкспрессия антител и сиалидазы
Целью данных исследований является получение линии клеток-хозяев, способных секретировать фермент сиалидазу в культуральные среды, которые больше можно не трансфицировать последовательностью, кодирующей антитело, причем посредством этого продуцируются гликозилированные антитела, которые могут быть десиалилированы. Линию клеток миеломы мыши С168М, которая экспрессирует антитела, трансфицируют вектором р3629, полученным в примере 7, и стабильные клоны отбирают и скринируют на сиалидазную активность в супернатанте. Как показано ниже в таблице 2, из 17 клонов в данном эксперименте, анализированных флуорометрией (MFU), шесть являются положительными на сиалидазную активность, и экспрессия сиалидазы продолжается в течение 6 недель, что указывает на стабильные клоны.
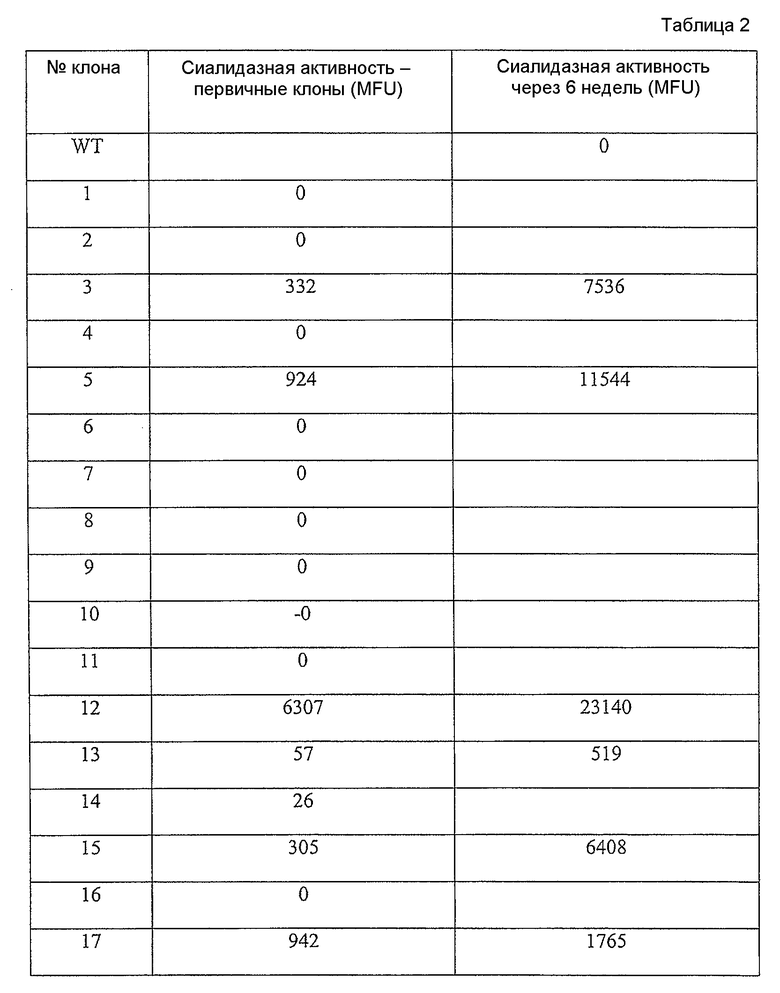
Дополнительный анализ очищенных антител из положительных на сиалидазу А клонов методом SDS-PAGE показывает, что они еще экспрессируют исходные антитела, и экспрессия сиалидазы не влияет на уровни экспрессии. Анализ углеводов антител показывает, что все клоны содержат менее 2% сиаловой кислоты, в отличие от антител из нетрансфицированных сиалидазой клеток-хозяев, которые содержат почти 15% сиаловой кислоты (таблица 3).
EC40 (нг/мл)
Антитела с низким содержанием сиаловой кислоты из клонов, сверхэкспрессирующих сиалидазу, анализируют в отношении усиленной активности ADCC в сравнении с WT C168M (фиг.14). Как видно из таблицы 3, некоторые образцы показывают меньшие ЕС40 и больший максимальный лизис, чем анти-TNFα антитела дикого типа, что указывает, что меньшее содержание сиаловой кислоты оказывает влияние на биологические свойства таких антител.
Создана экспрессирующая плазмида, которая будет секретировать каталитический домен фермента сиалидазы А из Arthrobacter ureafaciens в культуральные среды, для клеток млекопитающих. Такой секретированный фермент активен в клеточных супенатантах и способен удалять остаток сиаловой кислоты из N-присоединенных олигосахаридов, присутствующих в Fc-области антител. Устойчивая трансфекция экспрессирующих антитела клеточных линий такой экспрессирующей конструкцией приводит к клонам, которые секретируют активность сиалидазы А в супернатанты культур вместе с антителами. Антитела, извлеченные из таких клеточных культур, содержат меньше сиаловой кислоты, что преобразуется в функциональные улучшения в активности ADCC. Такие результаты предполагают, что способ можно использовать для получения клеток-хозяев, которые экспрессируют рекомбинантные гликопротеины, и использовать для создания культур для минимально сиалилированных гликоконъюгатов.
Пример 9. Экспрессия сиалидазы в СНО
Клетки СНО являются важными клетками-хозяевами для получения биофармацевтических препаратов. Плазмиду р3629 (фиг.13) используют для устойчивой трансфекции клеточной линии СНО, способной к дополнительной трансфекции вектором для экспрессии лечебного белка, такого как антитела.
Клетки СНО (С1835А) трансфицируют 30 мкг р3629, линеаризированной рестрикционным расщеплением Bgl II с использованием FuGene 6 (Roche, Inc.). Трансфицированные клетки выдерживают в среде для выращивания с 700 мкг/мл антибиотика генетицина (Invitrogen, Inc.) для отбора стабилиьных трансфектантов. Устойчивые к антибиотику клоны объединяют, размножают и анализируют на сиалидазную активность.
Сиалидазную активность анализируют в клеточных супернатантах от жизнеспособных клеточных культур с использованием флуорофора. К 200 мкл клеточного супернатанта добавляют 150 мкМ 2'-(4-метилумбеллиферил)-альфа-D-N-ацетилнейраминовой кислоты в 100 мМ цитратфосфатном буфере, рН 6,5, при 37°С с последующим добавлением 2 мл 0,5 М раствора Na2CO3 для прекращения реакции. Возбуждение при 366 нм, а испускание при 446 нм. Флуорометрические единицы (FU) нормализуют путем деления отдельных флуорометрических показателей для каждой клеточной линии на общее число жизнеспособных клеток. Сиалидазная активность в культуральной среде, измеренная в FU, приводится в таблице 4.
Так как сиалидазная активность через 3 недели и 8 недель после отбора остается постоянной или возрастает со временем, устойчивая трансфекция и секреция фермента сиалидазы в культуральный супернатант подтверждается.
Будет понятно, что изобретение можно осуществить на практике иначе, чем конкретно описано в приведенном выше описании и примерах. Многие модификации и изменения в воплощении настоящего изобретения возможны в свете описанных выше положений и, следовательно, входят в объем прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2011 |
|
RU2604811C2 |
| СПОСОБЫ РАЗРУШЕНИЯ КЛЕТОК С ИСПОЛЬЗОВАНИЕМ ЭФФЕКТОРНЫХ ФУНКЦИЙ АНТИ-EphA4 АНТИТЕЛ | 2007 |
|
RU2429246C2 |
| FC ВАРИАНТЫ АНТИТЕЛА | 2012 |
|
RU2607014C2 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ ИЛИ БИСПЕЦИФИЧЕСКАЯ МОЛЕКУЛА (ВАРИАНТЫ), СПОСОБ УНИЧТОЖЕНИЯ ИЛИ УМЕНЬШЕНИЯ КОЛИЧЕСТВА КЛЕТОК-МИШЕНЕЙ В ОРГАНИЗМЕ СУБЪЕКТА, СПОСОБ ЛЕЧЕНИЯ СУБЪЕКТА, ИНФИЦИРОВАННОГО ПАТОГЕНОМ И СПОСОБ ВАКЦИНАЦИИ СУБЪЕКТА | 1997 |
|
RU2201766C2 |
| АНТИТЕЛА ПРОТИВ А2 ТЕНАСЦИНА С И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2584597C2 |
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ДВА АНТИТЕЛА, СКОНСТРУИРОВАННЫХ ТАК, ЧТОБЫ ОНИ ОБЛАДАЛИ ПОНИЖЕННОЙ И ПОВЫШЕННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2013 |
|
RU2650788C2 |
| ГУМАНИЗИРОВАННЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К СЕА С СОЗРЕВШЕЙ АФФИННОСТЬЮ | 2010 |
|
RU2570554C2 |
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2815683C2 |
| УСТОЙЧИВЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2018 |
|
RU2779602C2 |
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2727836C2 |



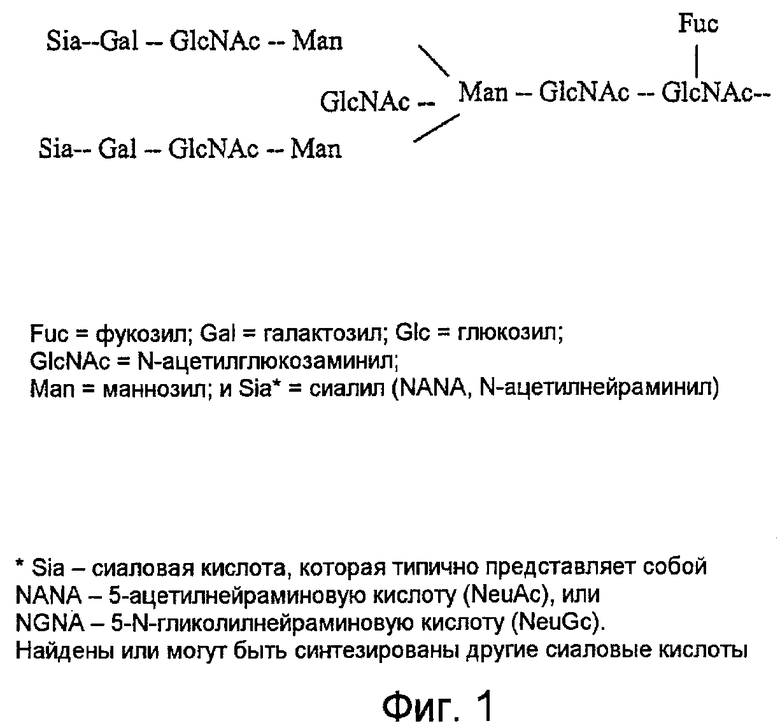
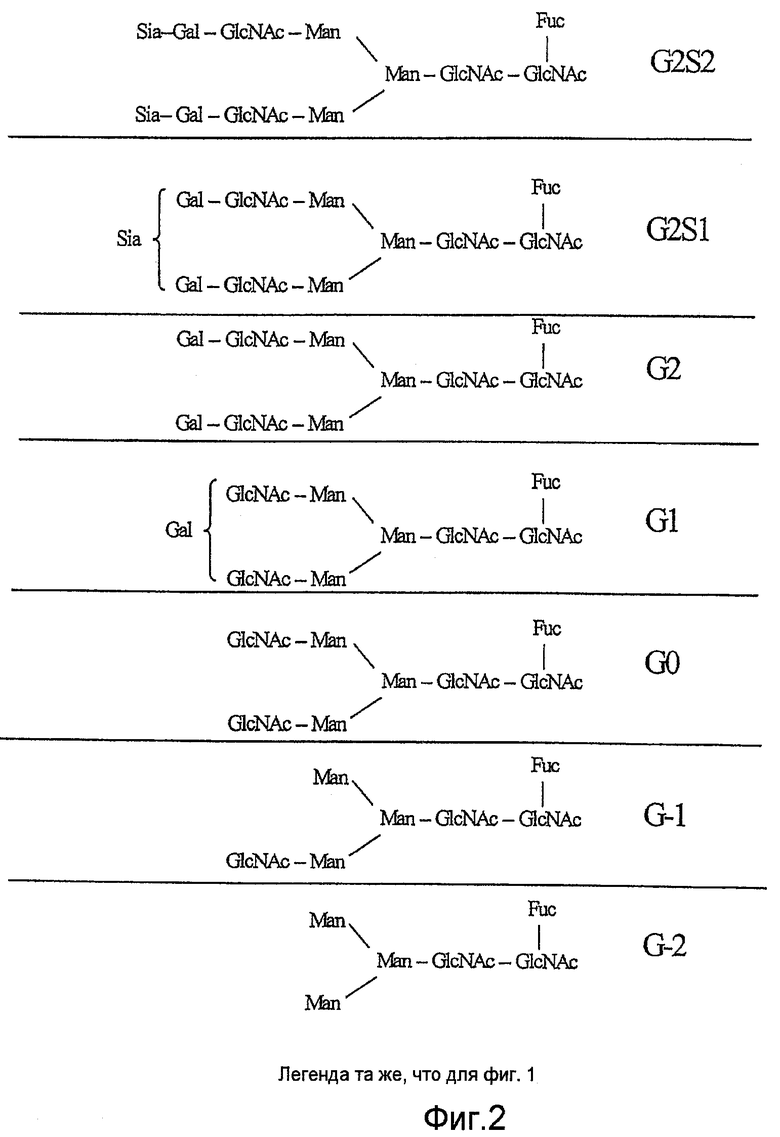
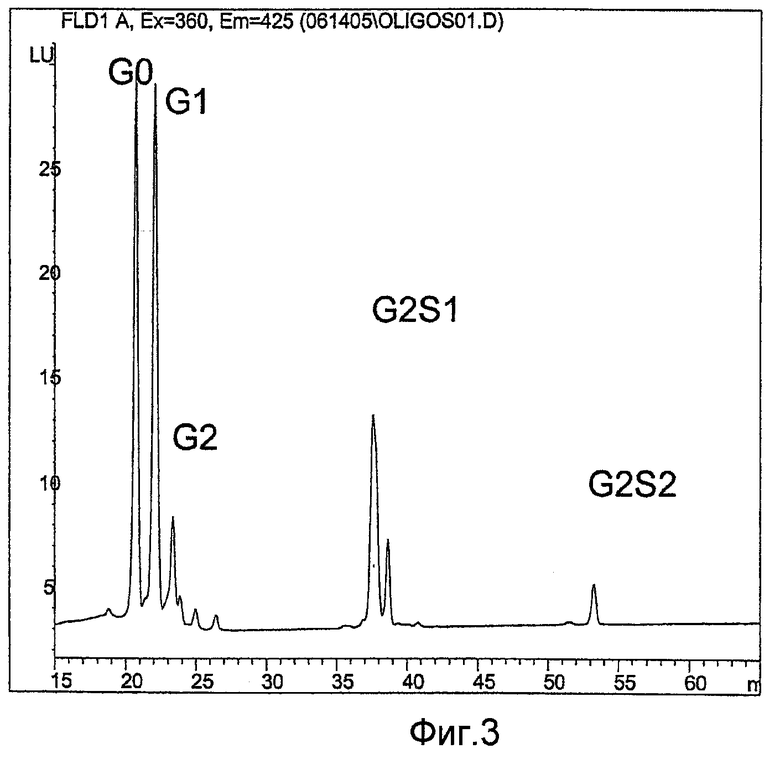
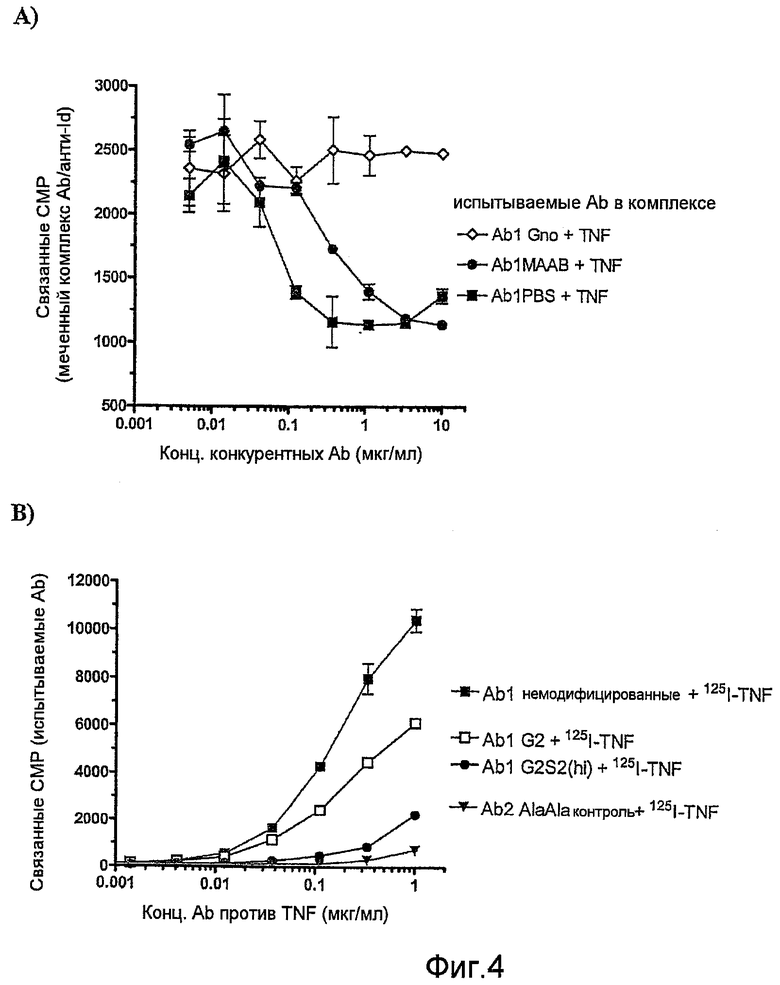

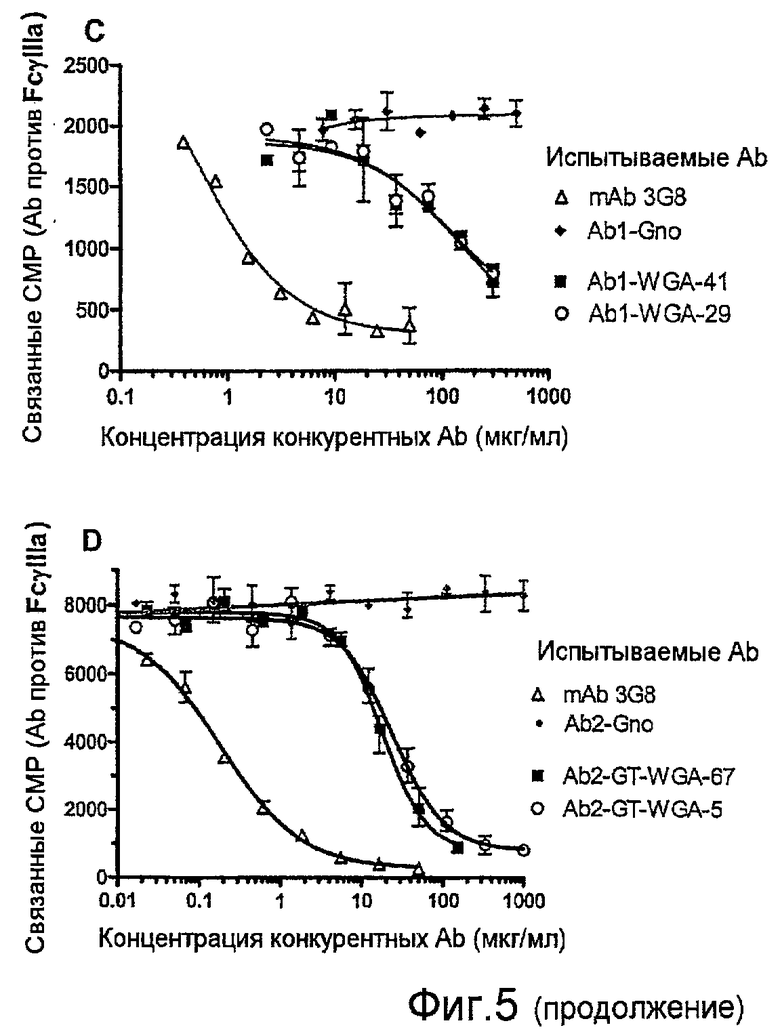
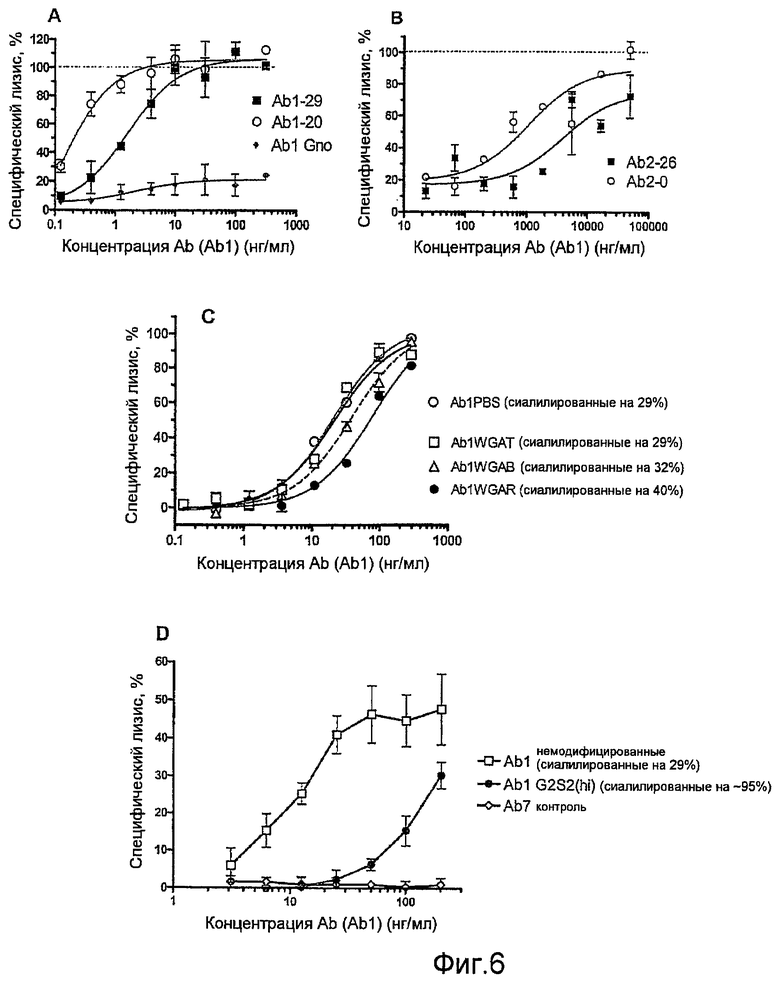
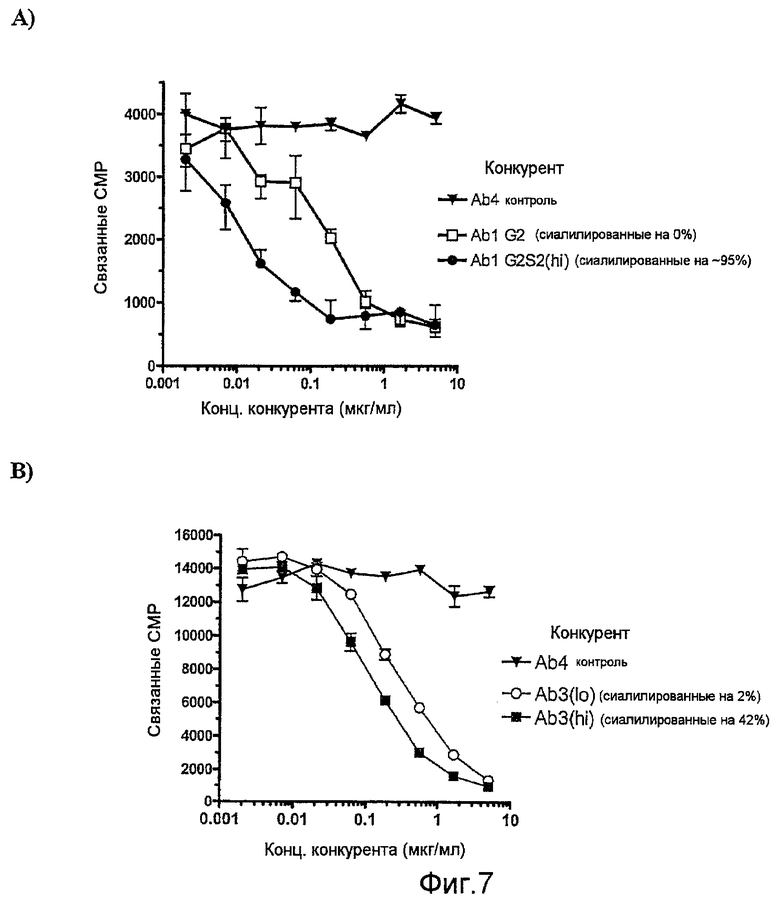

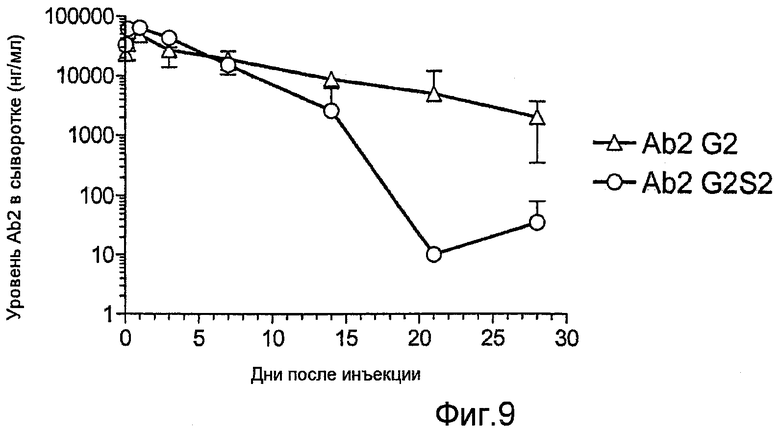
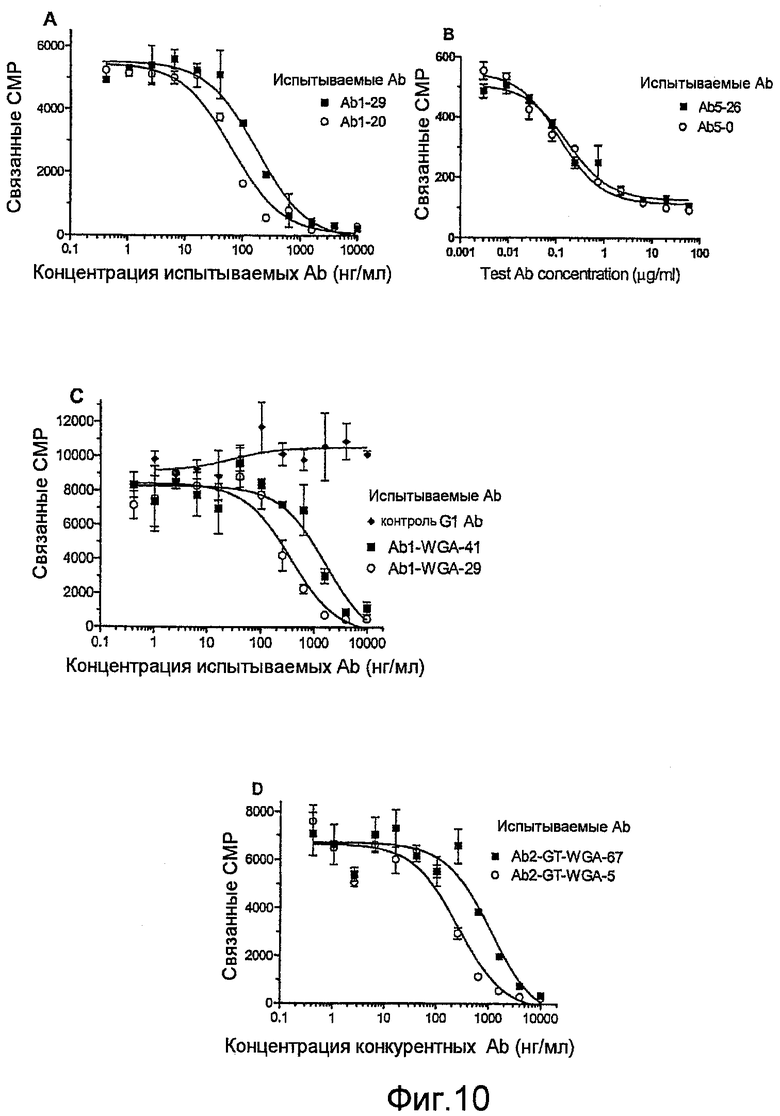
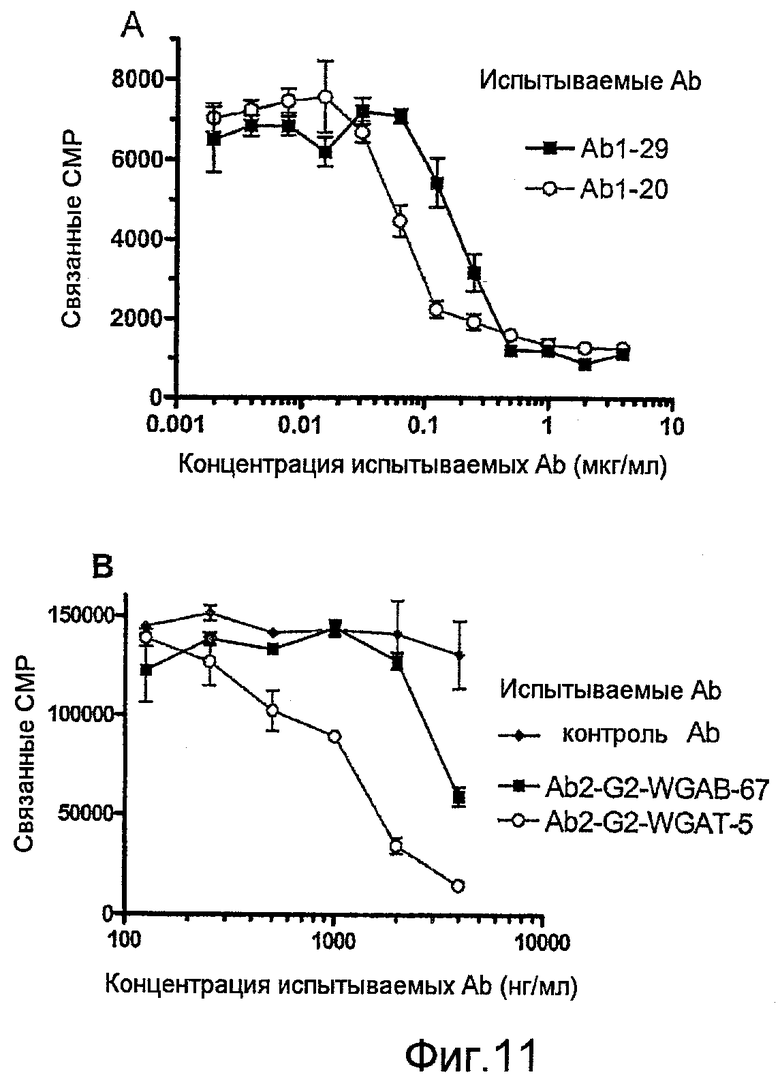
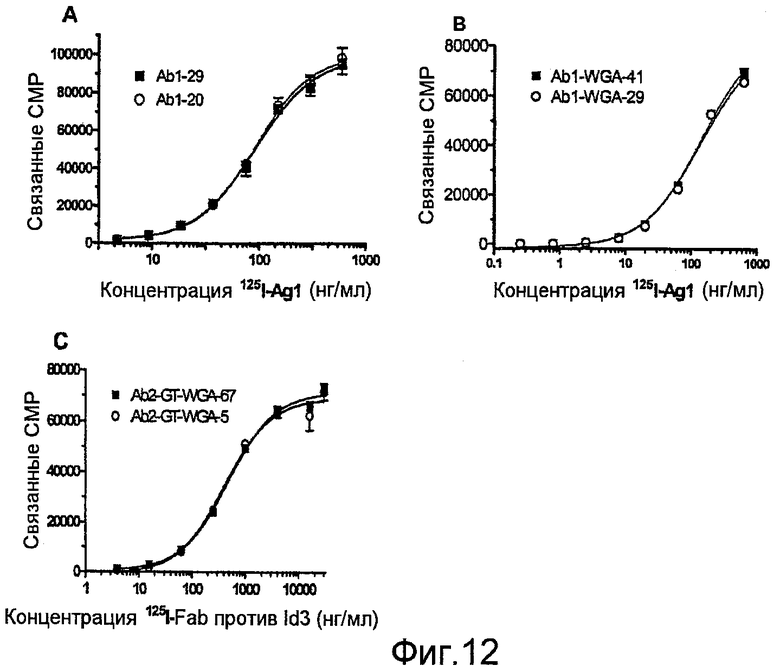
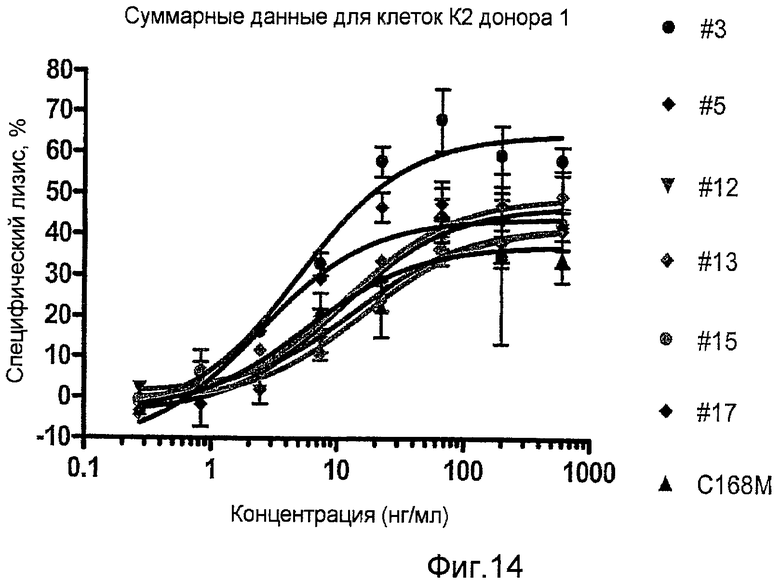
Изобретение относится к области биоинженерии и молекулярной биологии. Предложен способ обеспечения сиалидазной ферментативной активности в культуре клеточной линии на основе трансфекции клетки вектором для экспрессии полипептида, имеющего сиалидазную ферментативную активность и включающего последовательность, кодирующую каталитический домен фермента сиалидазы с пониженным количеством остатков сиаловой кислоты, где полипептид секретируется в среду вместе с Fc-содержащим белком. Так же предложен соответствующий вектор. Изобретение применимо при лечении заболеваний или состояний, при которых желательно регулировать аффинность в отношении одного или нескольких рецепторов FcγRI, FcγRIIA и FcγRIIIA, активность ADCC, активацию макрофагов или моноцитов, время полужизни в сыворотке и авидность. 3 н. и 11 з.п. ф-лы, 14 ил., 4 табл., 9 пр.
1. Способ обеспечения сиалидазной ферментативной активности в культуре клеточной линии, включающий создание трансфицированной клетки млекопитающего для экспрессии Fc-содержащей молекулы путем трансфекции клетки млекопитающего вектором для экспрессии полипептида, имеющего сиалидазную ферментативную активность, включающим нуклеотидную последовательность, кодирующую каталитический домен фермента сиалидазы с пониженным количеством остатков сиаловой кислоты, причем полипептид секретируется в культуру вместе с Fc-содержащей молекулой, в результате чего на гликаны Fc-содержащей молекулы может воздействовать сиалидазная ферментативная активность полипептида.
2. Способ по п.1, где вектор кодирует каталитический домен фермента сиалидазы А из Arthrobacter ureafaciencs.
3. Способ по п.1, где Fc-содержащая молекула представляет собой антитело, и линию клеток млекопитающего выбирают из группы, состоящей из HEK293, СНО, SP2/0, клеток миеломы или их любого производного, иммортализованных или трансформированных клеток.
4. Способ по п.3, где антитело представляет собой антитело против TNFα.
5. Экспрессирующий вектор, предназначенный для экспрессии полипептида, имеющего сиалидазную ферментативную активность, в культуре линии клеток млекопитающего, экспрессирующей Fc-содержащую молекулу, включающий нуклеотидную последовательность, кодирующую каталитический фрагмент фермента сиалидазы с пониженным количеством остатков сиаловой кислоты, а также сигнальную последовательность, промоторную последовательность и последовательность гена устойчивости к антибиотику.
6. Экспрессирующий вектор по п.5, где фермент сиалидаза представляет собой фермент сиалидазу А из Arthrobacter ureafaciencs, и линию клеток млекопитающего выбирают из группы, состоящей из HEK293, СНО, SP2/0, клеток миеломы или лимфомы или их любого производного, иммортализованных или трансформированных клеток.
7. Экспрессирующий вектор по п.5, где сигнальная последовательность представляет собой сигнальную последовательность гормона роста человека, промоторная последовательность представляет собой промоторную последовательность CMV, и ген устойчивости к антибиотику является геном устойчивости к неомицину.
8. Способ регулирования свойств Fc-содержащей молекулы, экспрессированной в клеточной линии, включающий снижение сиалилирования олигосахаридов Fc-области посредством трансфекции клеточной линии вектором, кодирующим каталитический домен фермента сиалидазы, где экспрессированная Fc-содержащая молекула включает пониженное количество остатков сиаловой кислоты.
9. Способ по п.8, где регулируемыми свойствами Fc-содержащей молекулы являются авидность молекулы в отношении множества локализованных белков-мишеней; аффинность в отношении одного или нескольких Fc-гамма-рецепторов FcγRI, FcγRIIA и FcγR-IIIA; активность ADCC; активация макрофагов или моноцитов и время полужизни в сыворотке.
10. Способ по п.8, где вектор кодирует каталитический домен фермента сиалидазы А из Arthrobacter ureafaciencs.
11. Способ по п.8, где Fc-содержащая молекула представляет собой антитело, и линию клеток млекопитающего выбирают из группы, состоящей из HEK293, СНО, SP2/0, клеток миеломы или лимфомы или их любого производного, иммортализованных или трансформированных клеток.
12. Способ по п.11, где антитело представляет собой антитело против TNFα.
13. Способ по п.8, где Fc-содержащая молекула имеет связывающий домен, специфичный для мишени, причем указанная мишень представляет собой иммобилизованную мишень.
14. Способ по п.8, где Fc-содержащая молекула имеет связывающий домен, специфичный для мишени, причем указанная мишень экспрессируется на поверхности клетки.
| DODON MD, QUASH GA | |||
| The antigenicity of asialylated IgG: its relationship to rheumatoid factor | |||
| Immunology | |||
| Приспособление для изготовления в грунте бетонных свай с употреблением обсадных труб | 1915 |
|
SU1981A1 |
| GALLOWAY G, LEUNG AY, HUNNEYBALL IM, Stanworth DR | |||
| The successful use of asialylated IgG as an immunogen and arthritogen in the rabbit | |||
| Immunology | |||
| Гребенчатая передача | 1916 |
|
SU1983A1 |

