ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По данной заявке испрашивается приоритет по предварительной заявке США № 61/656546, зарегистрированной 7 июня 2012 года, описание которой, таким образом, явно включено в качестве ссылки в полном объеме.
ВКЛЮЧЕНИЕ МАТЕРИАЛА, ПОДАННОГО В ЭЛЕКТРОННОМ ВИДЕ, В КАЧЕСТВЕ ССЫЛКИ
Включенным в полном объеме в качестве ссылки является машинно-читаемый список последовательностей, поданный одновременно с заявкой и определенный следующим образом: один файл размером 11342 байт ASCII (текстовый) под названием "72747_ST25.txt", созданный 13 мая 2013 года.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Сорняки могут быстро истощать содержание в почве полезных питательных веществ, необходимых для сельскохозяйственных культур и других желаемых растений. Существует множество типов гербицидов, используемых в настоящее время для контроля сорняков. Одним из крайне популярных гербицидов является глифосат.
Разработаны сельскохозяйственные культуры, такие как кукуруза, соя, канола, хлопок, сахарная свекла, пшеница, газонная трава и рис, резистентные к глифосату. Таким образом, поля с активно растущей кукурузой, резистентной к глифосату, например, можно опрыскивать для контроля сорняков без значительного повреждения растений кукурузы.
С внедрением генетически сконструированных, устойчивых к глифосату сельскохозяйственных культур (GTC) в середине 1990-х годов сельхозпроизводители получили простой, удобный, гибкий и недорогой инструмент для контроля широкого спектра широколиственных и травянистых сорняков, не имеющий аналогов в сельском хозяйстве. Таким образом, производители быстро приняли GTC и во многих случаях отказались от многих из лучших общепринятых агрономических приемов, таких как севооборот, севооборот в комбинации с действием гербицидов, использование баковой смеси, внедрение механического контроля вместе с химическим и культуральным контролем сорняков. В настоящее время толерантные к глифосату соя, хлопок, кукуруза и канола являются коммерчески доступными в США и в других областях Западного полушария. Все больше GTC (например, пшеница, рис, сахарная свекла, газонная трава, и т.д.) готовы для внедрения, ожидая принятия на глобальном рынке. Многие другие резистентные к глифосату виды находятся в стадии от эксперимента до разработки (например, люцерна, сахарный тростник, подсолнечник, свекла, горох, морковь, огурец, латук, лук, клубника, помидор и табак; лесохозяйственные виды, такие как тополь и ликвидамбар; и садоводческие виды, такие как бархатцы, петуния и бегония; см. веб-сайт "isb.vt.edu/cfdocs/fieldtestsl.cfm, 2005"). Кроме того, стоимость глифосата за последние годы значительно снизилась до такой точки, что несколько программ общепринятого контроля сорняков могут эффективно конкурировать по цене и действию с системами глифосат-GTC.
Глифосат успешно используют для уничтожения сорняков и в других несельскохозяйственных областях для тотальной борьбы с сорняками в течение более 15 лет. Во многих случаях, как и с GTC, глифосат используют 1-3 раз в год в течение 3, 5, 10, до 15 лет подряд. Эти обстоятельства привели к избыточному доверию к глифосату и технологии GTC и тяжелому давлению отбора по отношению к местным видам сорняков в пользу растений, являющихся от природы более устойчивыми к глифосату или у которых развился механизм резистентности к гербицидной активности глифосата.
Обширное использование программ контроля сорняков только с помощью глифосата приводит к селекции резистентных к глифосату сорняков и является селективным для размножения видов сорняков, от природы более устойчивых к глифосату, чем большинство видов-мишеней (т.е. сдвиги в популяциях сорняков). (Ng et al., 2003; Simarmata et al, 2003; Lorraine-Colwill et al, 2003; Sfiligoj, 2004; Miller et al, 2003; Heap, 2005; Murphy et al., 2002; Martin et al., 2002). Хотя глифосат широко используют по всему миру в течение более 15 лет, сообщают, что только у небольшого количества сорняков развилась резистентность к глифосату (Heap, 2005); однако, большинство из них идентифицировали в последние 3-5 лет. Резистентные сорняки включают травянистые и широколиственные виды - Lolium rigidum, Lolium multiflorum, Eleusine indica, Ambrosia artemisiifolia, Conyza canadensis, Conyza bonariensis и Plantago lanceolata. Кроме того, сорняки, ранее не представлявшие собой агрономическую проблему до широкого использования GTC, теперь становятся все более распространенными и трудноконтролируемыми в случае GTC, занимающих >80% акров хлопка и сои в США и >20% акров кукурузы в США (Gianessi, 2005). Эти сдвиги в популяциях сорняков преимущественно (но не исключительно) происходят у трудноконтролируемых широколиственных сорняков. Некоторые примеры включают виды Ipomoea, Amaranthus, Chenopodium, Taraxacum и Commelina.
В областях, где сельхозпроизводители сталкиваются с резистентными к глифосату сорняками или сдвигами в сторону более трудноконтролируемых видов сорняков, сельхозпроизводители могут компенсировать недостаточность действия глифосата использованием его в баковой смеси или заменой другими гербицидами, с помощью которых будут контролировать упущенные сорняки. Одним из популярных и эффективных средств для использования в баковой смеси для контроля широколиственных диких растений во многих случаях является 2,4-дихлорфеноксиуксусная кислота (2,4-D). 2,4-D используют в агрономии и в несельскохозяйственных условиях широкого спектра для контроля широколиственных сорняков в течение более 60 лет. Сообщают об отдельных случаях устойчивых видов, но 2,4-D остается одним из наиболее широко используемых гербицидов по всему миру. Ограничением для дальнейшего использования 2,4-D является то, что его селективность в случае двудольных сельскохозяйственных культур, таких как соя или хлопок, очень плоха, и, таким образом, 2,4-D, как правило, не используют на (и, как правило, не вблизи) чувствительных двудольных сельскохозяйственных культур. Кроме того, использование 2,4-D в случае травянистых сельскохозяйственных культур в некоторой степени ограничено природой повреждения сельскохозяйственных культур, которое может происходить. 2,4-D в комбинации с глифосатом используют для обеспечения более надежной burndown обработки перед посадкой сои и хлопка с нулевой обработкой почвы; однако, по причине чувствительности этих двудольных видов к 2,4-D, эту burndown обработку необходимо проводить по меньшей мере за 14-30 дней до посадки (Agriliance, 2003).
2,4-D принадлежит к феноксикислотному классу гербицидов, как и MCPA. 2,4-D используют в случае многих однодольных сельскохозяйственных культур (таких как кукуруза, пшеница и рис) для селективного контроля широколиственных сорняков без существенного повреждения желаемых сельскохозяйственных культур. 2,4-D является синтетическим производным ауксина, действующим, нарушая регуляцию нормального клеточно-гормонального гомеостаза и препятствуя сбалансированному, контролируемому росту; однако, точный механизм действия все еще неизвестен. Триклопир и флуроксипир являются гербицидами на основе пиридилоксиуксусной кислоты, механизм действия которых также является таким, как у синтетического ауксина.
Эти гербициды имеют различные уровни селективности в отношении конкретных растений (например, двудольные растения являются более чувствительными, чем травянистые растения). Различия метаболизма у различных растений являются одним из объяснений разных уровней селективности. В основном, растения медленно метаболизируют 2,4-D, таким образом, разный ответ растений на 2,4-D, более вероятно, можно объяснить различной активностью в сайтах-мишенях (WSSA, 2002). Метаболизм 2,4-D у растений, как правило, происходит посредством двухфазного механизма, как правило, гидроксилирования с последующей конъюгацией с аминокислотами или глюкозой (WSSA, 2002).
С течением времени в популяциях микроорганизмов появился альтернативный и эффективный путь деградации этого конкретного ксенобиотика, приводящий к полной минерализации 2,4-D. Последующие нанесения гербицида приводят к селекции микроорганизмов, которые могут использовать гербицид в качестве источника углерода для роста, обеспечивая им конкурентное преимущество в почве. По этой причине составляемый в настоящее время 2,4-D имеет относительно короткое время полужизни в почве, и не обнаруживали значительных переходящих эффектов в отношении последующих сельскохозяйственных культур. Это повышает применимость 2,4-D в качестве гербицида.
Одним из организмов, подвергаемых обширным исследованиям на предмет его способности деградировать 2,4-D, является Ralstonia eutropha (Streber et al., 1987). Кодирующим геном на первом ферментативном этапе цикла минерализации, является tfdA. См. патент США № 6153401 и GENBANK рег. № M16730. TfdA катализирует преобразование кислоты 2,4-D в дихлорфенол (DCP) через реакцию α-кетоглутарат-зависимой диоксигеназы (Smejkal et al., 2001). DCP обладает невысокой гербицидной активностью по сравнению с 2,4-D. TfdA используют в трансгенных растениях для придания резистентности к 2,4-D двудольным растениям (например, хлопку и табаку), в норме чувствительным к 2,4-D (Streber et al. (1989), Lyon et al. (1989), Lyon (1993) и патент США № 5608147).
Большое количество генов tfdA-типа, кодирующих белки, способные деградировать 2,4-D, идентифицированы в окружающей среде и депонированы в базе данных Genbank. Многие гомологи схожи с tfdA (>85% идентичности аминокислот) и обладают схожими с tfdA ферментативными свойствами. Однако, существует ряд гомологов, имеющих значительно меньшую идентичность по отношению к tfdA (25-50%), но имеющие характерные остатки, ассоциированные с α-кетоглутарат-зависимыми Fe+2-диоксигеназами. Таким образом, не ясны субстратные специфичности этих дивергентных диоксигеназ.
Одним уникальным примером низкой гомологии по отношению к tfdA (31% идентичности аминокислот) является sdpA из Delftia acidovorans (Kohler et al., 1999, Westendorf et al., 2002, Westendorf et al., 2003). Показано, что этот фермент катализирует первый этап в минерализации (S)-дихлорпропа (и других (S)-феноксипропионовых кислот), а также 2,4-D (феноксиуксусной кислоты) (Westendorf et al., 2003). До настоящего времени не сообщали об использовании этого гена в трансформации растений.
Разработка новых технологий устойчивых к гербицидам сельскохозяйственных культур (HTC) имеет ограниченный успех, главным образом, по причине эффективности, низкой стоимости и удобства GTC. Таким образом, среди производителей имеет место очень высокий уровень внедрения GTC. Это приводит к низкой мотивации для разработки новых технологий HTC.
Арилоксиалканоатные химические субструктуры являются распространенными веществами во многих коммерческих гербицидах, включая феноксиацетатауксины (такие как 2,4-D и дихлорпроп), пиридилоксиацетатауксины (такие как флуроксипир и триклопир), арилоксифеноксипропионатные (AOPP) ингибиторы ацетил-кофермент A карбоксилазы (ACCase) (такие как галоксифоп, квизалофоп, и диклофоп) и 5-замещенные феноксиацетатные ингибиторы протопорфириногеноксидазы IX (такие как пирафлуфен и флумиклорак). Однако эти классы гербицидов являются совершенно разными, и в современной литературе нет доказательств общих путей деградации среди этих классов химических веществ. Недавно описан мультифункциональный фермент для деградации гербицидов, охватывающих множество типов действия (PCT US/2005/014737; зарегистрированной 2 мая 2005 года).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам повышения высоты растений и/или урожайности сельскохозяйственных культур, резистентных к гербициду 2,4-D, посредством обработки растений 2,4-D при нормах внесения, не являющихся вредными для растений. В частности, изобретение относится к способу с использованием обработки 2,4-D для повышения урожайности сельскохозяйственных культур, экспрессирующих ген AAD-12 для резистентности к 2,4-D. Это изобретение дополнительно относится к применению 2,4-D для улучшения урожайности сельскохозяйственных культур, являющихся резистентными к 2,4-D. Предоставляемый способ представляет конкретный интерес для обработки сельскохозяйственных растений, включая кукурузу, сою, озимый и яровой рапс, (канолу), сахарную свеклу, пшеницу, подсолнечник, ячмень и рис.
В некоторых вариантах осуществления резистентные к 2,4-D сельскохозяйственные культуры являются трансгенными сельскохозяйственными культурами, трансформированными арилоксиалканоатдиоксигеназой (AAD). В дополнительном варианте осуществления арилоксиалканоатдиоксигеназа (AAD) является AAD-1 или AAD-12. AAD-1 ранее описана в патенте США 2009/0093366, а AAD-12 ранее описана в WO 2007/053482, полное содержание которых включено в настоящее описание в качестве ссылок.
Повышающий урожайность эффект обработки 2,4-D можно наблюдать при нормах внесения от 25 г кэ/га до 5000 г кэ/га, или от 100 г кэ/га до 2500 г кэ/га, или, в частности, от 1000 г кэ/га до 2000 г кэ/га. В одном из вариантов осуществления используют от 1000 г кэ/га до 1500 г кэ/га 2,4-D. В другом варианте осуществления используют от 2000 г кэ/га до 2500 г кэ/га. Кроме того, этот повышающий урожайность эффект обработки 2,4-D является особенно выраженным при нанесении 2,4-D в фазу от 2 до 8 листьев сельскохозяйственных культур до цветения. Однако необходимая норма внесения и/или фаза роста сельскохозяйственной культуры варьируется в зависимости от растений, их роста и климатических условий.
Термин "повышение" урожайности означает, что урожайность растения повышается на 50% или более. В одном из вариантов осуществления повышение урожайности составляет по меньшей мере 10%. В другом варианте осуществления повышение урожайности составляет по меньшей мере 20%. В другом варианте осуществления повышение урожайности составляет от 10% до 60%. В другом варианте осуществления повышение урожайности составляет от 20% до 50%. В другом варианте осуществления повышение урожайности является статистически значимым. Активность 2,4-D, усиливающую рост, в отношении резистентных к 2,4-D сельскохозяйственных культур можно измерять в полевых испытаниях или вегетационных опытах. О гербициде, имеющем различный механизм действия, как правило, известно, оказывает ли он неблагоприятное воздействие на урожайность, или не влияет на урожайность.
Один из аспектов относится к способу повышения урожайности резистентных к 2,4-D сельскохозяйственных культур, включающий обработку растений стимулирующим количеством гербицида, содержащего остаток арилоксиалканоата.
В одном из вариантов осуществления резистентные к 2,4-D сельскохозяйственные культуры являются трансгенными растениями, трансформированными с использованием арилоксиалканоатдиоксигеназы (AAD). В дополнительном варианте осуществления арилоксиалканоатдиоксигеназа (AAD) является AAD-1 или AAD-12. В другом варианте осуществления гербицид, содержащий остаток арилоксиалканоата, является фенокси-гербицидом или феноксиуксусным гербицидом. В дополнительном варианте осуществления гербицид, содержащий остаток арилоксиалканоата, является 2,4-D. В дополнительном варианте осуществления 2,4-D содержит 2,4-D-холин или 2,4-D-диметиламин (DMA).
В одном из вариантов осуществления трансгенные растения, трансформированные с использованием арилоксиалканоатдиоксигеназы, (AAD) выбраны из хлопка, сои и канолы. В другом варианте осуществления обработку осуществляют по меньшей мере один раз при норме внесения 2,4-D, используемой также для контроля сорняков. В другом варианте осуществления обработку осуществляют два раза при норме внесения 2,4-D, используемой также для контроля сорняков. В дополнительном варианте осуществления 2,4-D наносят в фазы роста V3 и R2 сои с устойчивостью к 2,4-D. В другом варианте осуществления обработку осуществляют по меньшей мере три раза при норме внесения 2,4-D, используемой также для контроля сорняков. В другом варианте осуществления гербицид, содержащий остаток арилоксиалканоата, достигает резистентных к 2,4-D сельскохозяйственных культур посредством поглощения корнями.
В другом варианте осуществления для контроля сорняков резистентные к 2,4-D сельскохозяйственные культуры также обрабатывают гербицидом, иным, чем 2,4-D. В дополнительном варианте осуществления гербицид, иной, чем 2,4-D, является фосфорным гербицидом или арилоксифеноксипропионовым гербицидом. В дополнительном варианте осуществления фосфорный гербицид включает глифосат, глуфосинат, их производные или их комбинации. В дополнительном варианте осуществления фосфорный гербицид находится в форме соли аммония, соли изопропиламмония, соли изопропиламина или соли калия. В другом варианте осуществления фосфорный гербицид достигает резистентных к 2,4-D сельскохозяйственных культур посредством поглощения корнями. В другом варианте осуществления арилоксифеноксипропионовый гербицид включает хлоразифоп, феноксапроп, флуазифоп, галоксифоп, квизалофоп, их производные или их комбинации. В дополнительном варианте осуществления арилоксифеноксипропионовый гербицид достигает резистентных к 2,4-D сельскохозяйственных культур посредством поглощения корнями.
В одном из вариантов осуществления резистентные к 2,4-D сельскохозяйственные культуры обрабатывают по меньшей мере один раз при дозе от 25 г кэ/га до 5000 г кэ/га 2,4-D. В другом варианте осуществления резистентные к 2,4-D сельскохозяйственные культуры обрабатывают по меньшей мере один раз при дозе от 100 г кэ/га до 2000 г кэ/га 2,4-D. В другом варианте осуществления резистентные к 2,4-D сельскохозяйственные культуры обрабатывают по меньшей мере один раз при дозе от 100 г кэ/га до 2500 г кэ/га 2,4-D. В другом варианте осуществления резистентные к 2,4-D сельскохозяйственные культуры обрабатывают по меньшей мере один раз при дозе от 1000 г кэ/га до 2000 г кэ/га 2,4-D. В дополнительном варианте осуществления 2,4-D включает 2,4-D-холин или 2,4-D-диметиламин (DMA).
Один из вариантов осуществления относится к способу повышения урожайности резистентных к 2,4-D сельскохозяйственных культур. Способ включает
(a) трансформацию растительных клеток с использованием молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую арилоксиалканоатдиоксигеназу (AAD);
(b) селекцию трансформированных клеток;
(c) регенерацию растений из трансформированных клеток; и
(d) обработку растений стимулирующим количеством гербицида, содержащего остаток арилоксиалканоата.
В одном из вариантов осуществления арилоксиалканоатдиоксигеназа (AAD) является AAD-1 или AAD-12. В другом варианте осуществления молекула нуклеиновой кислоты содержит селективный маркер, не являющийся арилоксиалканоатдиоксигеназой (AAD). В дополнительном варианте осуществления или альтернативном варианте осуществления селективный маркер является геном фосфинотрицинацетилтрансферазы (pat) или геном резистентности к биалафосу (bar). В другом варианте осуществления молекула нуклеиновой кислоты оптимизирована для растений.
Другой аспект относится к применению гербицида, содержащего остаток арилоксиалканоата, в производстве трансгенных растений с резистентностью к 2,4-D с повышенной урожайностью по сравнению с их нетрансгенными родительскими растениями. В одном из вариантов осуществления гербицид, содержащий остаток арилоксиалканоата, является 2,4-D. В дополнительном варианте осуществления 2,4-D наносят по меньшей мере один раз при дозе от 25 г кэ/га до 5000 г кэ/га 2,4-D. В другом варианте осуществления 2,4-D наносят по меньшей мере один раз при дозе от 100 г кэ/га до 2000 г кэ/га 2,4-D. В другом варианте осуществления 2,4-D наносят по меньшей мере один раз при дозе от 100 г кэ/га до 2500 г кэ/га 2,4-D. В другом варианте осуществления 2,4-D наносят по меньшей мере один раз при дозе от 1000 г кэ/га до 2000 г кэ/га 2,4-D. В дополнительном варианте осуществления 2,4-D включает 2,4-D-холин или 2,4-D-диметиламин (DMA). В дополнительном варианте осуществления резистентные к 2,4-D сельскохозяйственные культуры обрабатывают 2,4-D по меньшей мере два раза перед цветением. В другом варианте осуществления резистентные к 2,4-D сельскохозяйственные культуры являются трансгенными растениями, трансформированными с использованием арилоксиалканоатдиоксигеназы (AAD). В дополнительном варианте осуществления арилоксиалканоатдиоксигеназа (AAD) является AAD-1 или AAD-12.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ И ПОСЛЕДОВАТЕЛЬНОСТЕЙ
На фиг. 1 представлена общая химическая реакция, катализируемая ферментами AAD-12 по настоящему изобретению. На фиг. 2 представлена типичная карта плазмиды pDAB4468. На фиг. 3 представлена типичная карта плазмиды pDAS1740.
SEQ ID NO: 1 представляет собой нуклеотидную последовательность AAD-12 из Delftia acidovorans.
SEQ ID NO: 2 представляет собой транслированную белковую последовательность, кодируемую SEQ ID NO: 1.
SEQ ID NO: 3 представляет собой оптимизированную для растений нуклеотидную последовательность AAD-12 (v1).
SEQ ID NO: 4 представляет собой транслированную белковую последовательность, кодируемую SEQ ID NO: 3.
SEQ ID NO: 5 представляет собой оптимизированную для E. coli нуклеотидную последовательность AAD-12 (v2).
SEQ ID NO: 6 представляет собой последовательность прямого праймера M13.
SEQ ID NO: 7 представляет собой последовательность обратного праймера M13.
SEQ ID NO: 8 представляет собой последовательность прямого праймера PTU AAD-12 (v1).
SEQ ID NO: 9 представляет собой последовательность обратного праймера PTU AAD-12 (v1).
SEQ ID NO: 10 представляет собой последовательность прямого праймера для ПЦР кодирующей области AAD-12 (v1).
SEQ ID NO: 11 представляет собой последовательность обратного праймера для ПЦР кодирующей области AAD-12 (v1).
SEQ ID NO: 12 представляет собой последовательность праймера "sdpacodF" AAD-12 (v1).
SEQ ID NO: 13 представляет собой последовательность праймера "sdpacodR" AAD-12 (v1).
SEQ ID NO: 14 представляет собой последовательность праймера "Nco1 of Brady".
SEQ ID NO: 15 представляет собой последовательность праймера "Sac1 of Brady".
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Как применяют в настоящем описании, фраза "трансформированный" или "трансформация" относится к встраиванию ДНК в клетку. Фразы "трансформант" или "трансгенный" относятся к растительным клеткам, растениям и т.п., трансформированным или подвергнутым трансформации. Встраиваемая ДНК, как правило, находится в форме вектора, содержащего вставку ДНК.
Как применяют в настоящем описании, фраза "селективный маркер" или "ген селективного маркера" относится к гену, необязательно, используемому в трансформации растений, например, для защиты растительных клеток от селективного средства или обеспечения резистентности/устойчивости к селективному средству. Только те клетки или растения, которые получали функциональный селективный маркер, способны делиться или расти в условиях наличия селективного средства. Примеры селективных средств могут включать, например, антибиотики, включая спектиномицин, неомицин, канамицин, паромомицин, гентамицин и гигромицин. Эти селективные маркеры включают ген неомицинфосфотрансферазы (npt II), экспрессирующий фермент, придающий резистентность к антибиотику канамицину, и гены для родственных антибиотиков неомицина, паромомицина, гентамицина и G418, или ген гигромицинфосфотрансфераза (hpt), экспрессирующий фермент, придающий резистентность к гигромицину. Другие гены селективных маркеров могут включать гены, кодирующие резистентность к гербициду, включая Bar (резистентность к BASTA® (глуфосинат аммония) или фосфинотрицину (PPT)), ацетолактатсинтазу (ALS, резистентность к ингибиторам, таким как сульфонилкарбамиды (SU), имидазолиноны (IMI), триазолопиримидины (TP), пиримидинилоксибензоаты (POB) и сульфониламинокарбонилтриазолиноны, предотвращающие первый этап синтеза аминокислот с разветвленной цепью, глифосату, 2,4-D и резистентность или чувствительность к металлам. Фраза "маркер-позитивный" относится к растениям, трансформированным с включением гена селективного маркера.
В выбранный экспрессирующий вектор можно встраивать различные селективные или детектируемые маркеры, делая возможной идентификацию и селекцию трансформированных растений, или трансформантов. Доступно множество способов подтверждения экспрессии селективных маркеров в трансформированных растениях, включая, например, секвенирование ДНК и ПЦР (полимеразную цепную реакцию), Саузерн-блоттинг, РНК-блоттинг, иммунологические способы определения белка, экспрессирующегося с вектора, например, преципитированного белка, белков репортерных генов - к фосфинотрицину, или других белков, таких как β-глюкуронидаза (GUS), люцифераза, зеленый флуоресцентный белок (GFP), DsRed, β-галактозидаза, хлорамфениколацетилтрансфераза (CAT), щелочная фосфатаза и т.п. (См. Sambrook, et al., Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Press, N.Y., 2001, содержание которого включено в настоящее описание в качестве ссылки в полном объеме).
Гены селективных маркеров используют для селекции трансформированных клеток или тканей. Гены селективных маркеров включают гены, кодирующие резистентность к антибиотикам, такие как гены, кодирующие неомицинфосфотрансферазу II (NEO) и гигромицинфосфотрансферазу (HPT), а также гены, придающие резистентность к гербицидным соединениям. Гены резистентности к гербицидам, как правило, кодируют модифицированный белок-мишень, нечувствительный к гербициду, или фермент, деградирующий или детоксифицирующий гербицид в растении до того, как он может подействовать. См. DeBlock et al. (1987) EMBO J., 6:2513-2518; DeBlock et al. (1989) Plant Physiol., 91:691-704; Fromm et al. (1990) 8:833-839; Gordon-Kamm et al. (1990) 2:603-618). Например, резистентности к глифосату или сульфонилкарбамидным гербицидам достигали с использованием генов, кодирующих мутантные ферменты-мишени 5-енолпирувилшикимат-3-фосфатсинтетазу (EPSPS) и ацетолактатсинтазу (ALS). Резистентности к глуфосинату аммония, бромоксинилу и 2,4-дихлорфеноксиацетату (2,4-D) достигали с использованием бактериальных генов, кодирующих фосфинотрицинацетилтрансферазу, нитрилазу или 2,4-дихлорфеноксиацетатмонооксигеназу, детоксифицирующие соответствующие гербициды. Ферменты/гены резистентности к 2,4-D ранее описаны в патенте США № 2009/0093366 и WO 2007/053482, содержание которых, таким образом, включено в настоящее описание в качестве ссылок в полном объеме.
Другие гербициды могут ингибировать конус нарастания или меристему, включая имидазолинон или сульфонилкарбамид. Примеры генов этой категории кодируют мутантные ферменты ALS и AHAS, как описано, например, в Lee et al., EMBO J. 7: 1241 (1988); и Miki et al., Theon. Appl. Genet. 80:449 (1990), соответственно.
Гены резистентности к глифосату включают мутантные гены 5-енолпирувилшикимат-3-фосфатсинтетазы (EPSP) (посредством встраивания рекомбинантных нуклеиновых кислот и/или различных форм мутагенеза in vivo нативных генов EPSP), гены aroA и гены глифосатацетилтрансферазы (GAT), соответственно. Гены резистентности к другим фосфоновым соединениям включают гены глуфосинат (гены фосфинотрицинацетилтрансфераы (PAT) из видов Streptomyces, включая Streptomyces hygroscopicus и Streptomyces viridichromogenes) и пиридинокси- или феноксипроионовые кислоты и циклогексоны (гены, кодирующие ингибитор ACCase). См., например, патент США № 4940835 Shah, et al. и патент США № 6248876 Barry et al., в которых описывают нуклеотидные последовательности форм EPSP, которые могут придавать растению резистентность к глифосату. Молекулу ДНК, кодирующую мутантный ген aroA, можно получать в ATCC под регистрационным номером 39256, и нуклеотидная последовательность мутантного гена описана в патенте США № 4769061 Comai, европейской патентной заявке № 0 333 033 Kumada et al. и патенте США № 4975374 Goodman et al., в которых описывают нуклеотидные последовательности генов глутаминсинтетазы, придающих резистентность к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена PAT представлена в европейской патентной заявке № 0 242 246 Leemans et al. Кроме того, в DeGreef et al., Bio/Technology 7:61 (1989) описывают получение трансгенных растений, экспрессирующих химерные гены bar, кодирующие активность PAT. Примерами генов, придающих резистентность к феноксипропионовым кислотам и циклогексонам, включая сетоксидим и галоксифоп, являются гены Acc1-S1, Acc1-S2 и Acc1-S3, описываемые в Marshall et al., Theon. Appl. Genet. 83:435 (1992). Гены GAT, способные придавать резистентность к глифосату, описывают в WO 2005012515 Castle et al. Гены, придающие резистентность к гербицидам 2,4-D, фоп и пиридилоксиауксину, описывают в WO 2005107437 и патентной заявке США с серийным № 11/587893.
Другие гербициды могут ингибировать фотосинтез, включая триазин (гены psbA и 1s+) или бензонитрил (ген нитрилазы). В Przibila et al., Plant Cell 3: 169 (1991) описывают трансформацию Chlamydomonas с использованием плазмид, кодирующих мутантные гены psbA. Нуклеотидные последовательности генов нитрилазы описывают в патенте США № 4810648 Stalker, и молекулы ДНК, содержащие эти гены, доступны в ATCC под регистрационными №№ 53435, 67441 и 67442. Клонирование и экспрессию ДНК, кодирующей глутатион-S-трансферазу, описывают в Hayes et al., Biochem. J. 285: 173 (1992).
В целях по настоящему изобретению, гены селективных маркеров включают, в качестве неограничивающих примеров, гены, кодирующие: неомицинфосфотрансферазу II (Fraley et al. (1986) CRC Critical Reviews in Plant Science, 4: 1-25); цианамидгидратазу (Maier-Greiner et al. (1991) Proc. Natl. Acad. Sci. USA, 88:4250-4264); аспартаткиназу; дигидродипиколинатсинтетазу (Perl et al. (1993) Bio/Technology, 11:715-718); триптофандекарбоксилазу (Goddijn et al. (1993) Plant Mol. Bio., 22:907-912); дигидродипиколинатсинтетазу и десенсибилизированную аспартаткиназу (Perl et al. (1993) Bio/Technology, 11:715-718); ген bar (Toki et al. (1992) Plant Physiol., 100: 1503-1507 и Meagher et al. (1996) and Crop Sci., 36: 1367); триптофандекарбоксилазу (Goddijn et al. (1993) Plant Mol. Biol., 22:907-912); неомицинфосфотрансферазу (NEO) (Southern et al. (1982) J. Mol. Appl. Gen., 1:327; гигромицинфосфотрансферазу (HPT или HYG) (Shimizu et al. (1986) Mol. Cell Biol., 6: 1074); дигидрофолатредуктазу (DHFR) (Kwok et al. (1986) PNAS USA 4552); фосфинотрицинацетилтрансферазу (DeBlock et al. (1987) EMBO J., 6:2513); дегалогеназу 2,2-дихлорпропионовой кислоты (Buchanan-Wollatron et al. (1989) J. Cell. Biochem. 13D:330); синтетазу ацетогидроксикислот (Anderson et al., патент США № 4761373; Haughn et al. (1988) Mol. Gen. Genet. 221:266); 5-енолпирувилшикиматфосфатсинтетазу (aroA) (Comai et al. (1985) Nature 317:741); галоарилнитрилазу (Stalker et al., опубликованная заявка PCT WO87/04181); ацетил-кофермент A карбоксилазу (Parker et al. (1990) Plant Physiol. 92: 1220); дигидроптероатсинтетазу (sul I) (Guerineau et al. (1990) Plant Mol. Biol. 15: 127); и полипептид фотосистемы II массой 32 кДа (psbA) (Hirschberg et al. (1983) Science, 222: 1346).
Также включены гены, кодирующие резистентность к: хлорамфениколу (Herrera-Estrella et al. (1983) EMBO J., 2:987-992); метотрексату (Herrera-Estrella et al. (1983) Nature, 303:209-213; Meijer et al. (1991) Plant Mol Bio., 16:807-820 (1991); гигромицину (Waldron et al. (1985) Plant Mol. Biol., 5: 103-108; Zhijian et al. (1995) Plant Science, 108:219-227 и Meijer et al. (1991) Plant Mol. Bio. 16:807-820); стрептомицину (Jones et al. (1987) Mol. Gen. Genet., 210:86-91); спектиномицину (Bretagne-Sagnard et al. (1996) Transgenic Res., 5: 131-137); блеомицину (Hille et al. (1986) Plant Mol. Biol., 7: 171-176); сульфонамиду (Guerineau et al. (1990) Plant Mol. Bio., 15: 127-136); бромоксинилу (Stalker et al. (1988) Science, 242:419-423); 2,4-D (Streber et al. (1989) Bio/Technology, 7:811-816); глифосату (Shaw et al. (1986) Science, 233:478-481); и фосфинотрицину (DeBlock et al. (1987) EMBO J., 6:2513-2518). Все ссылки, процитированные в описании, таким образом, включены в настоящее описание в качестве ссылок в полном объеме, если не указано иначе.
Представленный выше список генов селективных маркеров и репортерных генов не является ограничивающим. Любой репортерный или ген селективного маркера находятся в объеме настоящего изобретения. При необходимости, такие гены можно секвенировать известными в этой области способами.
Репортерные гены и гены селективных маркеров синтезируют для оптимальной экспрессии в растениях. Т.е. кодирующую последовательность гена модифицируют для повышения экспрессии в растениях. Синтетический маркерный ген конструируют для экспрессии в растениях на высоком уровне, приводящей к более высокой эффективности трансформации. В этой области доступны способы синтетической оптимизации генов. Фактически, несколько генов оптимизируют для повышения экспрессии продукта гена в растениях.
Последовательность маркерного гена можно оптимизировать для экспрессии в конкретном виде растений или, альтернативно, ее можно модифицировать для оптимальной экспрессии в семействах растений. Предпочтительные для растения кодоны можно определять из кодонов, встречающихся с наибольшей частотой в белках, экспрессирующихся в наибольших количествах в конкретном интересующем виде растений. См., например, EPA 0359472; EPA 0385962; WO 91/16432; Perlak et al. (1991) Proc. Natl. Acad. Sci. USA, 88:3324-3328; и Murray et al. (1989) Nucleic Acids Research, 17: 477-498; патент США № 5380831; и патент США № 5436391, включенные в настоящее описание в качестве ссылок. Таким образом, нуклеотидные последовательности можно оптимизировать для экспрессии в любом растении. Установлено, что можно оптимизировать или синтезировать всю последовательность гена или любую ее часть. То есть, также можно использовать полностью оптимизированные или частично оптимизированные последовательности.
Кроме того, разработано несколько стратегий трансформации с использованием системы опосредованной Agrobacterium трансформации. Например, бинарная векторная стратегия основана на двухплазмидной системе, где Т-ДНК находится в иной плазмиде, чем плазмида Ti. В стратегии коинтеграции небольшую часть Т-ДНК помещают в тот же вектор, что и чужеродный ген, который затем рекомбинирует с плазмидой Ti.
Как применяют в настоящем описании, фраза "растение" включает двудольные растения и однодольные растения. Примеры двудольных растений включают табак, арабидопсис, сою, помидор, папайю, канолу, подсолнечник, хлопок, люцерну, картофель, виноград, голубиный горох, горох, декоративную капусту, нут, сахарную свеклу, рапс, арбуз, дыню, перец, арахис, тыкву, редис, шпинат, гигантскую тыкву, брокколи, капусту, морковь, цветную капусту, сельдерей, китайскую капусту, огурец, баклажан и латук. Примеры однодольных растений включают кукурузу, рис, пшеницу, сахарный тростник, ячмень, рожь, сорго, орхидею, бамбук, банан, рогоз, лилию, овес, лук, просо и тритикале.
Предметная разработка гена резистентности к 2,4-D и последующих резистентных сельскохозяйственных культур обеспечивает исключительные средства контроля широколиственных, резистентных к глифосату (или высокоустойчивых и подвергнутых сдвигу) видов сорняков для использования в сельскохозяйственных культурах. 2,4-D является относительно недорогим и надежным гербицидом широкого спектра действия против широколиственных растений, который будет приносить исключительную пользу сельхозпроизводителям, если можно обеспечивать боле высокую устойчивость сельскохозяйственных культур в равной степени в случае двудольных и однодольных сельскохозяйственных культур. Устойчивые к 2,4-D трансгенные двудольные сельскохозяйственные культуры также будут иметь большую гибкость по срокам и норме внесения. Дополнительной пользой указанного признака устойчивости к гербициду 2,4-D является предотвращение повреждения в норме восприимчивых сельскохозяйственных культур при дрейфе 2,4-D, испарении, преобразовании (или другом явлении перемещения за пределы территории), неправильном применении, вандализме и т.п. Дополнительным преимуществом гена AAD-12 является то, что, в отличие от всех гомологов tfdA, охарактеризованных к настоящему времени, AAD-12 способна деградировать пиридилоксиацетатные ауксины (например, триклопир, флуроксипир) в дополнение к ахиральным феноксиауксинам (например, 2,4-D, MCPA, 4-хлорфеноксиуксусной кислоте). См. таблицу 1. Общая картина химических реакций, катализируемых указанным ферментом AAD-12, представлена на фиг. 1. (Добавление O.sub.2 является стереоспецифичным; распад промежуточного соединения на фенол и глиоксилат является спонтанным.) Следует понимать, что с помощью химических структур на фиг. 1 иллюстрируют молекулярные остовы, и что различные R-группы и т.п. (такие, как представленные в таблице 1) включены, но не обязательно конкретно проиллюстрированы на фиг. 1. Для воздействия на конкретные спектры сорняков и условия окружающей среды в различных областях во всем мире используют множество смесей различных комбинаций феноксиауксинов. Использование гена AAD-12 в растениях обеспечивает защиту против гораздо более широкого спектра ауксиновых гербицидов, таким образом, увеличивая гибкость и спектры сорняков, которые можно контролировать.
К настоящему времени идентифицирован единственный ген (AAD-12), который при генетическом конструировании для экспрессии в растениях обладает свойствами, позволяющими использовать феноксиауксиновые гербициды на растениях, у которых никогда не было наследственной устойчивости, или она была недостаточно высокой, чтобы иметь возможность использовать эти гербициды. Кроме того, AAD-12 может обеспечивать защиту от пиридилоксиацетатных гербицидов в полевых условиях, повышая потенциальную пользу от этих гербицидов, природной устойчивости к которым также недостаточно, чтобы сделать возможной селективность. Растения, содержащие AAD-12 в отдельности, теперь можно обрабатывать последовательно или с использованием баковой смеси с одним, двумя или комбинацией нескольких феноксиауксиновых гербицидов. Норма внесения каждого феноксиауксинового гербицида может находиться в диапазоне от 25 до 4000 г кэ/га, и чаще - от 100 до 2000 г кэ/га для контроля широкого спектра двудольных сорняков. Аналогично, один, два или смесь нескольких пиридилоксиацетатауксиновых соединений можно наносить на растения, экспрессирующие AAD-12, со сниженным риском повреждения от указанных гербицидов. Норма внесения для пиридилоксиацетатного гербицида может находиться в диапазоне от 25 до 2000 г кэ/га, и чаще - 35-840 г кэ/га для дополнительного контроля двудольных сорняков.
Глифосат широко используют, т.к. он помогает контролировать очень широкий спектр широколиственных и травянистых видов сорняков. Однако повторное использование глифосата в GTC и при несельскохозяйственном применении приводит и будет продолжать приводить к селекции сорняков со сдвигами в сторону природно более устойчивых видов или резистентных к глифосату биотипов. Использование баковых смесей гербицидов, используемых в эффективных нормах внесения, обеспечивающих контроль одних и тех же видов, но имеющих различные механизмы действия, предписывают в большинстве стратегий борьбы с резистентностью к гербицидам, как способ задержки появления резистентных сорняков. Стэкинг AAD-12 с признаком устойчивости к глифосату (и/или с признаками устойчивости к другим гербицидам) может представлять механизм, позволяющий контролировать резистентные к глифосату виды двудольных сорняков в GTC, делая возможным использование глифосата, феноксиауксинов (например, 2,4-D) и пиридилоксиацетатауксиновых гербицидов (например, триклопира) избирательно в одной сельскохозяйственной культуре. Нанесение этих гербицидов может являться одновременным нанесением в баковой смеси, содержащей два или более гербицидов с различными механизмами действия; отдельным нанесением одной композиции гербицидов с последовательными нанесениями в предпосевном, предвсходном или послевсходовом и раздельном режиме нанесений в диапазоне от приблизительно 2 часов до приблизительно 3 месяцев; или, альтернативно, любую комбинацию любого количества гербицидов, представляющих каждый химический класс, можно наносить в любом режиме в пределах от приблизительно 7 месяцев от посадки сельскохозяйственной культуры до сбора урожая сельскохозяйственной культуры (или предуборочного интервала для отдельного гербицида, в зависимости от того, какой из них является наиболее коротким).
Важно иметь гибкость в контроле широкого спектра травянистых и широколиственных сорняков в отношении режима нанесения, нормы внесения отдельных гербицидов и способности контролировать представляющие затруднение или резистентные сорняки. Норма внесения глифосата на сельскохозяйственную культуру со стэком гена резистентности к глифосату/AAD-12 может находиться в диапазоне приблизительно 250-2500 г кэ/га; феноксиауксиновые гербициды (один или несколько) можно наносить при норме внесения приблизительно 25-4000 г кэ/га; и пиридилоксиацетатауксиновые гербициды (один или несколько) можно наносить при норме внесения 25-2000 г кэ/га. Оптимальные комбинации и режим этих нанесений будет зависеть от конкретной ситуации, вида и окружающей среды, и лучше всего их будет определять специалист в области контроля сорняков, получающий пользу от настоящего изобретения.
Проростки, как правило, резистентны на всем протяжении цикла роста. Трансформированные растения, как правило, будут резистентны к нанесению нового гербицида в любое время экспрессии гена. В настоящем описании представлена устойчивость к 2,4-D на всем протяжении жизненного цикла при использовании конститутивных промоторов, протестированных к настоящему времени (главным образом, CsVMV и AtUbi10). Это, как правило, является ожидаемым, но представляет собой усовершенствование по сравнению с другими неметаболизируемыми активными средствами, где на устойчивость может значительно влиять, например, сниженная экспрессия места приложения действия механизма резистентности. Одним из примеров является Roundup Ready cotton, где растения устойчивы при раннем опрыскивании, но если опрыскать их слишком поздно, глифосат концентрируется в меристеме (т.к. он не метаболизируется и перемещается); используемые вирусные промоторы Monsanto плохо экспрессируются в цветах. В этом отношении заявленное изобретение представляет собой усовершенствование.
Составы гербицидов (например, состав сложных эфиров, кислот или солей; или растворимый концентрат, эмульгируемый концентрат или растворимая жидкость) и добавки для баковых смесей (например, вспомогательные средства, поверхностно-активные вещества, замедлители дрейфа или средства совместимости) могут значительно влиять на контроль сорняков с помощью указанного гербицида или комбинации одного или нескольких гербицидов. Любая их комбинация с любым из указанных выше гербицидных химических веществ входит в объем настоящего изобретения.
Специалист в этой области также увидит пользу комбинирования двух или более механизмов действия для увеличения спектра контролируемых сорняков и/или для контроля природно более устойчивых или резистентных видов сорняков. Это также может распространяться на химические вещества, в случае которых устойчивость к гербицидам в сельскохозяйственных культурах стала возможной под влиянием человека (трансгенно или нетрансгенно) помимо GTC. В действительности, признаки резистентности к глифосату (например, EPSPS резистентного растения или бактерии, глифосатоксидоредуктазу (GOX), GAT), резистентности к глуфосинату (например, Pat, bar), ацетолактатсинтаза (ALS)-ингибиторной резистентности к гербицидам (например, имидазолинону, сульфонилкарбамиду, триазолопиримидин сульфонанилиду, пиримидинилтиобензоатам и другим химическим веществам - AHAS, Csrl, SurA и др.), резистентности к бромоксинилу (например, Bxn), резистентности к ингибиторам фермента HPPD (4-гидроксифенилпируватдиоксигеназы), резистентности к ингибиторам фитоендесатуразы (PDS), резистентности к гербицидам, ингибирующим фотосистему II (например, psbA), резистентности к гербицидам, ингибирующим фотосистему I, резистентности к гербицидам, ингибирующим протопорфириногеноксидазу IX (PPO) (например, PPO-1), резистентности гербицидам на основе фенилмочевины (например, CYP76B1), дикамба-деградирующие ферменты (см, например, US 20030135879) и другие могут быть обособлены или быть во множестве комбинаций для обеспечения способности эффективно контролировать или предотвращать сдвиги сорняков и/или резистентность к любому гербициду из указанных выше классов. В некоторых предпочтительных вариантах осуществления можно использовать модифицированную in vivo EPSPS, а также гены резистентности к глифосату класса I, класса II и класса III.
Что касается дополнительных гербицидов, некоторые дополнительные предпочтительные ингибиторы ALS включают, в качестве неограничивающих примеров сульфонилкарбамиды (такие как хлорсульфурон, галосульфурон, никосульфурон, сульфометурон, сульфосульфурон, трифлоксисульфурон), имидазолиноны (такие как имазамокс, имазетапир, имазахин), триазолопиримидин сульфонанилиды (такие как клорансулам-метил, диклосулам, флорасулам, флуметсулам, метосулам и пеноксулам), пиримидинилтиобензоаты (такие как биспирибак и пиритиобак) и флукарбазон. Некоторые предпочтительные ингибиторы HPPD включают, в качестве неограничивающих примеров, мезотрион, изоксафлутол и сулкотрион. Некоторые предпочтительные ингибиторы PPO включают, в качестве неограничивающих примеров, флумиклорак, флумиоксазин, флуфенпир, пирафлуфен флутиацет, бутафенацил, карфентразон, сульфентразон и дифениловые простые эфиры (такие как ацифлуорфен, фомезафен, лактофен и оксифлуорфен).
Кроме того, AAD-12 в отдельности или подвергнутую стэкингу с одним или несколькими дополнительными признаками HTC можно подвергать стэкингу с одним или несколькими дополнительными входными (например, резистентность к насекомым, резистентность к грибам или стрессоустойчивость и др.) или выходными (например, повышенная урожайность урожай, улучшенный профиль масел, улучшенное качество волокон и др.) признаками. Таким образом, настоящее изобретение можно использовать для обеспечения полного агрономического пакета улучшенного качества сельскохозяйственной культуры со способностью гибко и экономически эффективно контролировать любое количество сельскохозяйственных вредителей.
Настоящее изобретение частично относится к идентификации фермента, который не только способен деградировать 2,4-D, но также, неожиданно, обладает новыми свойствами, что отличает фермент по настоящему изобретению, например, от ранее известных белков tfdA. Несмотря на то, что этот фермент имеет очень низкую гомологию с tfdA, гены по настоящему изобретению все равно, как правило, можно классифицировать в то же общее семейство α-кетоглутарат-зависимых диоксигеназ. Это семейство белков отличается тремя консервативными остатками гистидина в мотиве "HX(D/E)X23-26(T/S)X114-183HX10-13R", содержащем активный центр. Гистидины координируют в активном центре ион Fe2+, который важен для каталитической активности (Hogan et al., 2000). Предварительные эксперименты по экспрессии in vitro, обсуждаемые в настоящем описании, адаптировали для облегчения селекции новых свойств. Эти эксперименты также показывают, что фермент AAD-12 уникален по отношению к другому, в корне отличающемуся ферменту того же класса, описываемому в зарегистрированной ранее патентной заявке (PCT US/2005/014737; зарегистрированной 2 мая 2005 года). Фермент AAD-1 по этой заявке имеет всего лишь приблизительно 25% идентичности последовательности по отношению к белку AAD-12 по настоящему изобретению.
Более конкретно, настоящее изобретение частично относится к применению фермента, способного деградировать не только 2,4-D, но также и пиридилоксиацетатные гербициды. Ранее не сообщали об α-кетоглутарат-зависимом ферменте диоксигеназе, имеющем способность деградировать гербициды различных химических классов и механизмов действия. Предпочтительные ферменты и гены для применения по настоящему изобретению обозначают в настоящем описании как гены и белки AAD-12 (арилоксиалканоатдиоксигеназы).
Белки по настоящему изобретению при анализе являлись тест-положительными на превращение 2,4-D в 2,4-дихлорфенол ("DCP"; гербицидно неактивный). Частично очищенные белки по настоящему изобретению могут быстро преобразовывать 2,4-D в DCP in vitro. Дополнительным преимуществом предоставляемых AAD-12-трансформированных растений является то, что родительские гербициды метаболизируются до неактивных форм, таким образом, снижая потенциал сбора гербицидных остатков вместе с зерном или соломой.
Настоящее изобретение также включает способы контроля сорняков, где указанные способы включают нанесение пиридилоксиацетатного и/или феноксиауксинового гербицида на растения, содержащие ген AAD-12.
В свете этих результатов, теперь предоставляют новые растения, содержащие полинуклеотид, кодирующий этот тип фермента. До этого момента не существовало мотивации получать такие растения, и не ожидали, что такие растения смогут эффективно продуцировать этот фермент, делающий растения резистентными не только к гербицидам на основе феноксикислот (таким как 2,4-D), но также и к пиридилоксиацетатным гербицидам. Таким образом, настоящее изобретение обеспечивает множество преимуществ, которые до настоящего момента в этой области не считали возможными.
Общедоступные штаммы (депонируемые в коллекциях культур, таких как ATCC или DSMZ) можно приобретать и подвергать скринингу на новые гены с использованием способов, представленных в настоящем описании. Последовательности, представленные в настоящем описании, можно использовать для амплификации и клонирования гомологичных генов в систему рекомбинантной экспрессии для дополнительного скрининга и тестирования по настоящему изобретению.
Как указано выше в разделе "Уровень техники", одним из организмов, интенсивно исследуемых на способность деградировать 2,4-D, являлась Ralstonia eutropha (Streber et al., 1987). Геном, кодирующим первый фермент в пути деградации, является tfdA. См. патент США № 6153401 и GENBANK рег. № M16730. TfdA катализирует превращение кислоты 2,4-D в гербицидно неактивный DCP посредством реакции α-кетоглутарат-зависимой диоксигеназы (Smejkal et al., 2001). TfdA используют в трансгенных растениях для придания резистентности к 2,4-D двудольным растениям (например, хлопку и табаку), в норме восприимчивым к 2,4-D (Streber et al., 1989; Lyon et al., 1989; Lyon et al., 1993). Большое количество генов tfdA-типа, кодирующих белки, способные деградировать 2,4-D, идентифицировали из окружающей среды и депонировали в базе данных Genbank. Многие гомологи очень похожи на tfdA (>85% идентичности аминокислот) и имеют аналогичные tfdA ферментативные свойства. Однако, к настоящему времени идентифицировали небольшую коллекцию гомологов α-кетоглутарат-зависимой диоксигеназы, имеющих низкий уровень гомологии с tfdA.
Настоящее изобретение частично относится к неожиданным данным о новом использовании и функциях отдаленно родственного фермента sdpA из Delftia acidivorans (Westendorf et al., 2002, 2003) с низкой гомологией по отношению к tfdA (31% идентичности аминокислот). Ранее показано, что этот фермент α-кетоглутарат-зависимая диоксигеназа, очищенная в своей нативной форме, деградирует 2,4-D и S-дихлорпроп (Westendorf et al., 2002 и 2003). Однако ранее не сообщали о ферменте α-кетоглутарат-зависимой диоксигеназе, имеющей способность деградировать гербициды химического класса пиридилоксиацетатов. SdpA никогда не экспрессировали в растениях, не было и мотивации для этого, частично потому, что разработка новых технологий HTC ограничена, главным образом, эффективностью, низкой стоимостью и удобством GTC (Devine, 2005).
С учетом новой активности белки и гены по настоящему изобретению обозначают в настоящем описании как белки и гены AAD-12. К настоящему времени подтверждено, что AAD-12 деградирует разнообразные феноксиацетатауксиновые гербициды in vitro. Однако, как впервые сообщено в настоящем описании, неожиданно обнаруживали, что этот фермент также способен деградировать дополнительные субстраты класса арилоксиалканоатных молекул. Субстраты значительной агрономической важности включают пиридилоксиацетатауксиновые гербициды. Это новое открытие является основой для значительных возможностей для устойчивых к гербицидам сельскохозяйственных культур (HTC) и признаков селективных маркеров. Этот фермент уникален по своей способности проявлять свою гербицид-деструктивную активность по отношению к гербицидам широколиственных растений широкого спектра (феноксиацетат- и пиридилоксиацетатауксинам).
Таким образом, настоящее изобретение частично относится к деградации 2,4-дихлорфеноксиуксусной кислоты, других феноксиацетатауксиновых гербицидов и пиридилоксиацетатных гербицидов с помощью рекомбинантно экспрессирующегося фермента арилоксиалканоатдиоксигеназы (AAD-12). Настоящее изобретение также частично относится к идентификации и применению генов, кодирующих деградирующий фермент арилоксиалканоатдиоксигеназу (AAD-12), способную деградировать фенокси- и/или пиридилоксиауксиновые гербициды.
Фермент по настоящему изобретению делает возможной трансгенную экспрессию, приводящую к устойчивости к комбинациям гербицидов, с помощью которых будут контролировать почти все широколиственные сорняки. AAD-12 может служить в качестве исключительного признака устойчивой к гербицидам сельскохозяйственной культуры (HTC) для стэкинга с другими признаками HTC [например, резистентностью к глифосату, резистентностью к глуфосинату, резистентностью к ALS-ингибиторам (например, имидазолинону, сульфонилкарбамиду, триазолопиримидин сульфонанилиду), резистентностью к бромоксинилу, резистентностью к HPPD-ингибиторам, резистентностью к PPO-ингибиторам и др.] и, например, признаками резистентности к насекомым (Cry1F, Cry1Ab, Cry 34/45, другие белки Bt. или инсектицидные белки, не происходящие из Bacillis, и др.). Дополнительно, AAD-12 может служить в качестве селективного маркера для облегчения селекции первичных трансформантов растений, генетически сконструированных со вторым геном или группой генов.
Кроме того, ген микроорганизма по настоящему изобретению переконструировали таким образом, что белок кодируется кодонами, смещенными в сторону их использования и однодольными, и двудольными растениями (гемикот). Арабидопсис, кукурузу, табак, хлопок, сою, канолу и рис трансформировали с использованием конструкций, содержащих AAD-12, и они демонстрировали высокие уровни резистентности к фенокси- и пиридилоксиауксиновым гербицидам. Таким образом, настоящее изобретение также относится к генам, "оптимизированным для растений", кодирующим белки по настоящему изобретению.
Оксиалканоатные группы применимы для включения в гербициды стабильной кислотной функциональной группы. Кислотная группа посредством "кислотной ловушки" может придавать желаемую характерную черту действия гербицида - подвижность во флоэме, и, таким образом, ее можно включать в новые гербициды в целях повышения подвижности. Аспекты настоящего изобретения также относятся к механизму получения HTC. Существует множество потенциальных коммерческих и экспериментальных гербицидов, которые могут служить в качестве субстратов для AAD-12. Таким образом, применение генов по настоящему изобретению также может приводить к устойчивости к гербициду также в отношении других гербицидов.
Признаки HTC по настоящему изобретению можно использовать в новых комбинациях с другими признаками HTC (включая, в качестве неограничивающих примеров, устойчивость к глифосату). Эти комбинации признаков дают начало новым способам контроля видов сорняков (и т.п.) по причине новоприобретенной резистентности или наследственной устойчивости к гербицидам (например, глифосату). Таким образом, в дополнение к признакам HTC в объем изобретения входят новые способы контроля сорняков с использованием гербицидов, устойчивости к которым в трансгенных сельскохозяйственных культурах достигали с помощью указанного фермента.
Настоящее изобретение можно применять в отношении извлечения коммерческой выгоды из признака резистентности к 2,4-D, подвергнутого стэкингу, например, с существующими в настоящее время признаками резистентности к глифосату у сои. Таким образом, настоящее изобретение относится к инструменту для борьбы со сдвигами в популяциях видов широколиственных сорняков и/или селекции резистентных к гербицидам широколиственных сорняков, достигших широкого распространения в результате крайне высокого доверия сельхозпроизводителей в отношении глифосата для контроля сорняков в различных сельскохозяйственных культурах.
Трансгенная экспрессия генов AAD-12 по настоящему изобретению представлена в качестве примера в арабидопсисе, табаке, сое, хлопке, рисе, кукурузе и каноле. Соя является предпочтительной сельскохозяйственной культурой для трансформации по настоящему изобретению. Однако настоящее изобретение можно использовать во множестве других однодольных (таких как пастбищные злаки или газонная трава) и двудольных сельскохозяйственных культур, таких люцерна, клевер, виды деревьев и др. Аналогично, 2,4-D (или другие субстраты AAD-12) можно более успешно использовать в случае травянистых сельскохозяйственных культур, в которых устойчивость является умеренной, и устойчивость, повышенная с помощью этого признака, будет предоставлять сельхозпроизводителям возможность использования этих гербицидов в более эффективных нормах внесения и с более широким режимом нанесения без риска повреждения сельскохозяйственных культур.
Кроме того, настоящее изобретение относится к одному гену, который может обеспечивать резистентность к гербицидам, с помощью которых можно контролировать широколиственные сорняки. Этот ген можно использовать во множестве сельскохозяйственных культур, чтобы сделать возможным использование комбинации гербицидов широкого спектра. С помощью настоящего изобретения также можно контролировать сорняки, резистентные к существующим в настоящее время химическим веществам, и оно может помогать контролировать сдвиги популяций сорняков, возникающие в результате существующих в настоящее время агрономических технологий. AAD-12 по настоящему изобретению также можно использовать в попытках эффективно детоксифицировать дополнительные гербицидные субстраты в негербицидные формы. Таким образом, настоящее изобретение относится к разработке дополнительных признаков HTC и/или технологии селективных маркеров.
Отдельно или в дополнение к использованию генов по настоящему изобретению для получения HTC, гены по настоящему изобретению также можно использовать в качестве селективных маркеров для успешной селекции трансформантов в культурах клеток, теплицах и в полевых условиях. Гены по настоящему изобретению обладают крайне важным значением просто в качестве селективного маркера для биотехнологических проектов. Недифференцированное действие AAD-12 в отношении других арилоксиалканоатауксиновых гербицидов предоставляет множество возможностей для использования этого гена в целях получения HTC и/или селективного маркера.
Нельзя просто описать термин "резистентность" и не использовать глагол "иметь устойчивость" или прилагательное "устойчивый". В этой области потрачено бесчисленное количество часов в дебатах о терминах "устойчивые к гербицидам сельскохозяйственные культуры" (HTC) и "резистентные к гербицидам сельскохозяйственные культуры" (HRC). HTC является предпочтительным термином в этой области. Однако, официальным определением "резистентности" согласно Weed Science Society of America является "врожденная способность растения выживать и размножаться после воздействия дозы гербицида, в норме летальной для растения дикого типа. У растений резистентность может являться природной или индуцируемой с помощью таких технологий, как генная инженерия или селекция вариантов, осуществляемая с помощью культуры тканей или мутагенеза". Как применяют в настоящем описании, если не указано иначе, "резистентность" к гербициду является наследственной и позволяет растению расти и размножаться в присутствие типичной гербицидно-эффективной обработки с помощью гербицида для указанного растения, как предложено в последнем, на момент подачи настоящей заявки, издании Herbicide Handbook. Как считают специалисты в этой области, растение все равно можно считать "резистентным", даже если очевидна некоторая степень повреждения растения в результате обработки гербицидом. Как применяют в настоящем описании, термин "устойчивость" является более широким, чем термин "резистентность", и включает "резистентность", как определено в настоящем описании, а также улучшенную способность конкретного растения сопротивляться различным степеням индуцируемого гербицидами повреждения, которое, как правило, происходит у растений дикого типа того же генотипа при той же дозе гербицида.
Перенос функциональной активности в растительные или бактериальные системы может включать последовательность нуклеиновой кислоты, кодирующую аминокислотную последовательность белка по настоящему изобретению, интегрированную в вектор, экспрессирующий белок, подходящий для хозяина, в котором вектор будет находиться. Одним из способов получения последовательности нуклеиновой кислоты, кодирующей белок с функциональной активностью, является выделение нативного генетического материала из видов бактерий, продуцирующих интересующий белок, с использованием информации, прогнозируемой из аминокислотной последовательности белка, как представлено в настоящем описании. Нативные последовательности можно оптимизировать для экспрессии в растениях, например, как более подробно описано ниже. Оптимизированный полинуклеотид также можно конструировать на основе последовательности белка.
Существует ряд способов получения белков для применения по настоящему изобретению. Например, можно использовать антитела против белков, представленных в настоящем описании, для идентификации и выделения таких белков из смеси белков. В частности, можно индуцировать антитела против частей белков, являющихся наиболее консервативными или наиболее различающимися по сравнению с другими родственными белками. Затем эти антитела можно использовать для конкретной идентификации эквивалентных белков с характерной активностью посредством иммунопреципитации, твердофазного иммуноферментного анализа (ELISA) или иммуноблоттинга. Антитела против белков, представленных в настоящем описании, или против эквивалентных белков, или против фрагментов этих белков можно легко получать с использованием стандартных способов. Такие антитела являются аспектом настоящего изобретения. Антитела по настоящему изобретению включают моноклональные и поликлональные антитела, предпочтительно, продуцируемые в ответ на являющийся примером или предполагаемый белок.
Специалисту в этой области будет понятно, что белки (и гены) по настоящему изобретению можно получать из разнообразных источников. Так как известно, что полные опероны деградации гербицидов кодируются мобильными генетическими элементами, такими как плазмиды, а также элементами, интегрируемыми в геном, белки по настоящему изобретению можно получать из широкого спектра микроорганизмов, например, включая рекомбинантные бактерии и/или бактерии дикого типа.
Мутантные формы изолятов бактерий можно получать способами, хорошо известными в этой области. Например, аспорогенные мутанты можно получать, подвергая изолят мутагенезу с использованием этилметансульфоната (EMS). Мутантные штаммы также можно получать с использованием ультрафиолетового излучения и нитрозогуанидина хорошо известными в этой области способами.
Белок "из" или "получаемый из" любого из изолятов по настоящему изобретению, упомянутый или предложенный в настоящем описании, означает, что белок (или аналогичный белок) можно получать из изолята или некоторого другого источника, такого как другой штамм бактерий или растение. "Полученный из" также имеет это значение и включает белки, получаемые из указанного типа бактерий, например, модифицированные для экспрессии в растении. Специалисту в этой области будет понятно, что, принимая во внимание описание бактериального гена и белка, можно конструировать растение для получения белка. Препараты антител, зонды нуклеиновых кислот (например, ДНК, РНК или ПНК) и т.п. можно получать с использованием полинуклеотидных и/или аминокислотных последовательностей, представленных в настоящем описании, и использовать их для скрининга и выделения других родственных генов из других (природных) источников.
Для клонирования и секвенирования белков и генов, представленных в настоящем описании, можно использовать стандартные способы молекулярной биологии. Дополнительную информацию можно найти в Sambrook et al., 1989, включенном в настоящее описание в качестве ссылки.
Полинуклеотиды и зонды: Настоящее изобретение дополнительно относится к последовательностям нуклеиновой кислоты, кодирующим белки для применения по настоящему изобретению. Настоящее изобретение дополнительно относится к способам идентификации и определения свойств генов, кодирующих белки, имеющих желаемую гербицидную активность. В одном из вариантов осуществления настоящее изобретение относится к уникальным нуклеотидным последовательностям, применимым в качестве зондов для гибридизации и/или праймеров для способов ПЦР. С помощью праймеров получают характерные фрагменты генов, которые можно использовать в идентификации, определении свойств и/или выделении конкретных интересующих генов. Нуклеотидные последовательности по настоящему изобретению кодируют белки, отличающиеся от ранее описанных белков.
Полинуклеотиды по настоящему изобретению можно использовать для получения полных "генов", кодирующих белки или пептиды в желаемой клетке-хозяине. Например, как будет понятно специалистам в этой области, полинуклеотиды по настоящему изобретению можно соответствующим образом помещать под контроль промотора в интересующем хозяине, как известно в этой области. Уровень экспрессии гена и временная/тканеспецифическая экспрессия могут значительно влиять на применимость изобретения. Как правило, более высокие уровни экспрессии белка деструктивного гена будут приводить к более быстрой и более полной деградации субстрата (в этом случае - гербицида-мишени). Желательно, чтобы промоторы контролировали экспрессию гена-мишени на высоких уровнях, если высокая экспрессия не имеет последующего негативного влияния на здоровье растения. Как правило, желательно иметь ген AAD-12, конститутивно экспрессирующимся во всех тканях для полной защиты растения на во все фазы роста. Однако, альтернативно, можно использовать ген резистентности, экспрессирующийся в зависимости от фазы вегетативного цикла; это позволит использовать гербицид-мишень в фазу плодоношения для контроля сорняков и затем контролировать половое размножение целевой сельскохозяйственной культуры посредством нанесения в течение фазы цветения. Кроме того, желаемые уровни и периоды экспрессии также могут зависеть от типа растения и желаемого уровня устойчивости. В некоторых предпочтительных вариантах осуществления используют сильные конститутивные промоторы в комбинации с энхансерами транскрипции и т.п. для повышения уровней экспрессия и устойчивости до желаемых уровней. Некоторые такие варианты осуществления более подробно описаны ниже перед разделом "Примеры".
Как известно специалистам в этой области, ДНК, как правило, существует в двухцепочечной форме. В такой структуре одна цепь комплементарна другой цепи, и наоборот. Т.к. ДНК реплицируется (например) в растении, продуцируются дополнительные комплементарные цепи ДНК. Термин "кодирующая цепь" часто используют в этой области для обозначения цепи, связывающейся с антисмысловой цепью. мРНК транскрибируется с "антисмысловой" цепи ДНК. "Смысловая" или "кодирующая" цепь имеет серию кодонов (кодон представляет собой три нуклеотида, которые можно читать как единицу из трех остатков для обозначения конкретной аминокислоты), которая может быть считана как открытая рамка считывания (ORF) для образования интересующего белка или пептида. В случае продукции белка in vivo цепь ДНК, как правило, транскрибируется в комплементарную цепь мРНК, используемую в качестве матрицы для белка. Таким образом, настоящее изобретение включает применение примеров полинуклеотидов, представленных в приложенном списке последовательностей, и/или эквивалентов, включая комплементарные цепи. Настоящее изобретение включает РНК и ПНК (пептид-нуклеиновые кислоты), являющиеся функционально эквивалентными примерам молекул ДНК.
Белки и гены для применения по настоящему изобретению можно идентифицировать и получать, например, с использованием олигонуклеотидных зондов. Эти зонды являются детектируемыми нуклеотидными последовательностями, которые можно определять с помощью соответствующей метки, или которые можно получать, по существу, флуоресцентными, как описано в международной патентной заявке № WO 93/16094. Зонды (и полинуклеотиды по настоящему изобретению) могут представлять собой ДНК, РНК или ПНК. В дополнение к аденину (A), цитозину (C), гуанину (G), тимину (T) и урацилу (U; в случае молекул РНК), синтетические зонды (и полинуклеотиды) по настоящему изобретению также могут содержать инозин (нейтральное основание, способное спариваться со всеми четырьмя основаниями; иногда используемое вместо смеси всех четырех оснований в синтетических зондах) и/или другие синтетические (неприродные) основания. Таким образом, когда в настоящем описании упоминают синтетический, вырожденный олигонуклеотид и, в общем, используют "N" или "n", "N" или "n" может представлять собой G, A, T, C или инозин. Как применяют в настоящем описании, неопределенные коды соответствуют стандартным соглашениям о номенклатуре IUPAC на момент подачи настоящей заявки (например, R означает A или G, Y означает C или T и т.д.).
Как хорошо известно в этой области, если молекулу зонда гибридизуют с образцом нуклеиновой кислоты, можно разумно предположить, что зонд и образец обладают существенной гомологией/сходством/идентичностью. Предпочтительно, сначала осуществляют гибридизацию полинуклеотида с последующими промываниями в условиях пониженной строгости, умеренной строгости или в строгих условиях способами, хорошо известными в этой области, как описано, например, в Keller, G. H., M. M. Manak (1987) DNA Probes, Stockton Press, New York, N.Y., pp. 169-170. Например, как там указано, условий пониженной строгости можно достигать сначала промыванием 2-кратным SSC (стандартным цитратно-солевым буфером)/0,1% SDS (додецилсульфатом натрия) в течение 15 минут при комнатной температуре. Как правило, осуществляют два промывания. Затем можно достигать большей строгости, понижая концентрацию соли и/или повышая температуру. Например, после указанного выше промывания можно осуществлять два промывания 0,1-кратным SSC/0,1% SDS в течение 15 минут, каждое при комнатной температуре, с последующими промываниями 0,1-кратным SSC/0,1% SDS в течение 30 минут, каждое при 55°C. Эти температуры можно использовать с другими протоколами гибридизации и промывания, приведенными в настоящем описании и известными специалисту в этой области (в качестве соли вместо SSC можно использовать, например, SSPE). 2-кратный SSC/0,1% SDS можно получать, добавляя 50 мл 20-кратного SSC и 5 мл 10% SDS в 445 мл воды. 20-кратный SSC можно получать, комбинируя NaCl (175,3 г/0,150 M), цитрат натрия (88,2 г/0,015 M) и воду, доводя pH до 7,0 с помощью 10 Н NaOH, а затем доводя объем до 1 литра. 10% SDS можно получать, растворяя 10 г SDS в 50 мл автоклавированной воды, а затем разбавляя до 100 мл.
Детекция зонда обеспечивает средства для определения известным способом того, поддерживается ли гибридизация. Такой анализ зонда представляет быстрый способ идентификации генов по настоящему изобретению. Сегменты нуклеотидов, используемые в качестве зондов по изобретению, можно синтезировать с использованием ДНК-синтезатора и стандартных способов. Эти нуклеотидные последовательности также можно использовать в качестве праймеров для ПЦР для амплификации генов по настоящему изобретению.
Гибридизационные характеристики молекулы можно использовать для определения полинуклеотидов по настоящему изобретению. Таким образом, настоящее изобретение включает полинуклеотиды (и/или комплементарные им полинуклеотиды, предпочтительно, полностью комплементарные им полинуклеотиды), гибридизующиеся с полинуклеотидом, приведенным в качестве примера в настоящем описании. То есть, одним из способов определения гена (и кодируемого им белка), например, является определение его способности гибридизоваться (в любых условиях, конкретно представленных в настоящем описании) с известным или конкретно приведенным в качестве примера геном.
Как применяют в настоящем описании, термин "строгие" условия гибридизации относится к условиям, с помощью которых достигают той же или приблизительно той же степени специфичности гибридизации, что и в условиях, используемых авторами настоящего изобретения. В частности, гибридизацию иммобилизованной ДНК при Саузерн-блоттинге с мечеными 32P специфичными для гена зондами можно осуществлять стандартными способами (см., например, Maniatis et al. 1982). В основном, гибридизацию и последующие промывания можно осуществлять в условиях, позволяющих определять последовательности-мишени. В случае двухцепочечных ДНК-зондов к гену гибридизацию можно осуществлять в течение ночи при температуре на 20-25°C ниже температуры плавления (Tm) гибрида ДНК в 6-кратном SSPE, 5-кратном растворе Денхардта, 0,1% SDS, 0,1 мг/мл денатурированной ДНК.
Как правило, промывания можно осуществлять следующим образом: (1) дважды при комнатной температуре в течение 15 минут в 1-кратном SSPE, 0,1% SDS (промывание в условиях пониженной строгости); и (2) один раз при Tm-20°C в течение 15 минут в 0,2-кратном SSPE, 0,1% SDS (промывания в условиях умеренной строгости).
В случае олигонуклеотидных зондов гибридизацию можно осуществлять в течение ночи при температуре на 10-20°C ниже температуры плавления (Tm) гибрида в 6-кратном SSPE, 5-кратном раствор Денхардта, 0,1% SDS, 0,1 мг/мл денатурированной ДНК.
Как правило, промывания можно осуществлять следующим образом: (1) дважды при комнатной температуре в течение 15 минут в 1-кратном SSPE, 0,1% SDS (промывание в условиях пониженной строгости); и (2) один раз при температуре гибридизации в течение 15 минут в 1-кратном SSPE, 0,1% SDS (промывание в условиях умеренной строгости).
В основном, для изменения строгости можно изменять соль и/или температуру. В случае меченого фрагмента ДНК длиной >70 оснований или около того, можно использовать следующие условия: (1) пониженной строгости: 1- или 2-кратный SSPE, комнатная температура; (2) пониженной строгости: 1- или 2-кратный SSPE, 42°C; (3) умеренной строгости: 0,2- или 1-кратный SSPE, 65°C, или (4) строгие условия: 0,1-кратный SSPE, 65°C
Образование и стабильность дуплекса зависят от фактической комплементарности между двумя цепями гибрида и, как указано выше, можно допускать некоторую степень несовпадения. Таким образом, последовательности зондов по настоящему изобретению включают мутации (одиночные и множественные), делеции, инсерции в описываемых последовательностях и их комбинации, где указанные мутации, инсерции и делеции делают возможным образование стабильных гибридов с интересующим полинуклеотидом-мишенью. Мутации, инсерции и делеции можно получать в указанной полинуклеотидной последовательности множеством способов, и эти способы, как правило, известны специалистам в этой области. Другие способы могут стать известными в будущем.
Технология ПЦР: Полимеразная цепная реакция (ПЦР) представляет собой повторяющийся, ферментативный, примированный синтез последовательности нуклеиновой кислоты. Этот способ хорошо известен и общеупотребителен среди специалистов в этой области (см. Mullis, патенты США №№ 4683195, 4683202, и 4800159; Saiki et al., 1985). ПЦР основана на ферментативной амплификации интересующего фрагмента ДНК, фланкированного двумя олигонуклеотидными праймерами, гибридизующимися на противоположных цепях последовательности-мишени. Предпочтительно, праймеры ориентированы по 3'-концам, направленным друг против друга. Повторяющиеся циклы термической денатурации матрицы, отжига праймеров на комплементарных им последовательностях и элонгации отожженных праймеров с помощью ДНК-полимеразы приводят к амплификации сегмента, определяемого по 5'-концам праймеров для ПЦР. Продукт элонгации каждого праймера может служить в качестве матрицы для других праймеров, таким образом, в каждом цикле удваивается количество фрагмента ДНК, образовавшееся в предыдущем цикле. Это приводит к экспоненциальному накоплению конкретного фрагмента-мишени в несколько миллионов раз за несколько часов. Используя термостабильную ДНК-полимеразу, такую как Taq-полимераза, выделенная из термофильной бактерии Thermus aquaticus, можно полностью автоматизировать амплификацию. Другие ферменты, которые можно использовать, известны специалистам в этой области.
Примеры последовательности ДНК или их сегменты можно использовать в качестве праймеров для ПЦР-амплификации. При осуществлении ПЦР-амплификации можно допускать определенную степень несовпадения между праймерами и матрицей. Таким образом, мутации, делеции, и инсерции (особенно добавления нуклеотидов на 5'-конец) примеров праймеров входят в объем настоящего изобретения. В указанном праймере можно осуществлять мутации, инсерции и делеции способами, как правило, известными специалистам в этой области.
Модификация генов и белков: Гены и белки по настоящему изобретению можно подвергать слиянию с другими генами и белками для получения химерных или слитных белков. Гены и белки, применимые по настоящему изобретению, включают не только конкретно приведенные в качестве примеров полноразмерные последовательности, но также части, сегменты и/или фрагменты (включая смежные фрагменты и варианты с внутренними и/или концевыми делециями по сравнению с полноразмерными молекулами) этих последовательностей, их варианты, мутантные, химерные и слитные формы. Белки по настоящему изобретению могут содержать замены аминокислот при условии, что они сохраняют желаемую функциональную активность. "Варианты" генов имею нуклеотидные последовательности, кодирующие те же белки или эквивалентные белки, имеющие активность, эквивалентную или схожую с белком, приведенным в качестве примера.
Два лучших результата поисков с помощью BLAST с использованием нативной нуклеотидной последовательностью aad-12 демонстрируют достаточный уровень гомологии (приблизительно 85%) по 120 парам оснований в последовательности. Можно ожидать, что гибридизация в конкретных условиях будет включать эти две последовательности. См. GENBANK рег. №№ DQ406818.1 (89329742; Rhodoferax) и AJ6288601.1 (44903451; Sphingomonas). Rhodoferax очень схожа с Delftia, но Sphingomonas филогенетически представляет совершенно другой класс.
Термины "варианты белков" и "эквивалентные белки" относятся к белкам, имеющим ту же или, по существу, ту же биологическую/функциональную активность против субстратов-мишеней и эквивалентные последовательности по отношению к белкам, приведенным в качестве примеров. Как применяют в настоящем описании, ссылка на "эквивалентную" последовательность относится к последовательностям, содержащим замены аминокислот, делеции, добавления или инсерции, улучшающие или не влияющие на активность в значительной степени неблагоприятно. Фрагменты, сохраняющие активность также включены в это определение. Фрагменты и другие эквиваленты, сохраняющие ту же или аналогичную функцию или активность, что и соответствующий фрагмент белка, приведенного в качестве примера, входят в объем настоящего изобретения. Изменения, такие как замены аминокислот или добавления, можно осуществлять в различных целях, таких как повышение (или снижение) протеазной стабильности белка (без значительного/существенного снижения функциональной активности белка), удаление или добавление участка рестрикции и т.п. Варианты генов можно легко конструировать, например, с использованием стандартных способов получения точечных мутаций.
Кроме того, например, в патенте США № 5605793 описывают способы получения дополнительного молекулярного разнообразия с использованием реассемблирования ДНК после случайной или направленной фрагментации. Это можно обозначать как "перетасовку" генов, как правило, включающую смешивание фрагментов (желаемого размера) двух или более различных молекул ДНК с последующими повторяющимися раундами ренатурации. Это может улучшать активность белка, кодируемого исходным геном. Результатом является химерным белок, имеющий улучшенную активность, измененную субстратную специфичность, повышенную ферментативную стабильность, измененную стереоспецифичность или другие характеристики.
"Перетасовку" можно планировать и направлять после получения и исследования атомных 3D (трехмерных) координат и кристаллической структуры интересующего белка. Таким образом, "направленную перетасовку" можно направлять на конкретные сегменты белка, идеальные для модификации, такие как сегменты, экспонируемые на поверхности, и, предпочтительно, не внутренние сегменты, участвующие в фолдинге белка и важные для 3D структурной целостности.
Конкретные изменения в "активном центре" фермента можно осуществлять для воздействия на присущую ему функциональность в отношении активности или стереоспецифичности. Muller et. al. (2006). Известную кристаллическую структуру tauD использовали в качестве модели диоксигеназы для определения остатков активного центра, в то время как он был связан со своим характерным субстратом таурином. Elkins et al. (2002) “X-ray crystal structure of Escerichia coli taurine/alpha-ketoglutarate dioxygenase complexed to ferrous iron and substrates,” Biochemistry 41(16):5185-5192. Что касается оптимизации последовательности и конструирования активных центров фермента, см. Chakrabarti et al., PNAS, (Aug. 23, 2005), 102(34): 12035-12040.
Фрагменты полноразмерных генов можно получать стандартными способами с использованием коммерчески доступных экзонуклеаз или эндонуклеаз. Например, для систематического отщепления нуклеотидов с концов этих генов можно использовать ферменты, такие как Bal31, или сайт-специфический мутагенез. Кроме того, гены, кодирующие активные фрагменты, можно получать с использованием различных ферментов рестрикции. Чтобы напрямую получать активные фрагменты этих белков, можно использовать протеазы.
Как представлено в настоящем описании, в объем изобретения входит то, что белки могут являться укороченными и все равно сохранять функциональную активность. Термин "укороченный белок" означает, что можно отщеплять часть белка, в то время как остальной укороченный белок после расщепления сохраняет и проявляет желаемую активность. Расщепление можно осуществлять с использованием различных протеаз. Кроме того, эффективно расщепленные белки можно получать способами молекулярной биологии, где основания ДНК, кодирующей указанный белок, удаляют посредством обработки рестрикционными эндонуклеазами, или другими способами, доступными специалистам в этой области. После укорачивания указанные белки можно экспрессировать в гетерологичных системах, таких как E. coli, бакуловирусы, системы на основе вирусов растений, дрожжи и т.п., а затем помещать в системы анализа на основе клеток насекомых, как представлено в настоящем описании, для определения активности. В этой области хорошо известно, что можно успешно получать укороченные белки таким образом, что они сохраняют функциональную активность, имея меньшую последовательность, чем полноразмерная последовательность. Например, белки B.t. можно использовать в укороченной форме (коровый белок) (см., например, Hofte et al. (1989), и Adang et al. (1985)). Как применяют в настоящем описании, термин "белок" может включать функционально активные укороченные формы.
Как применяют в настоящем описании, если не указано иначе, процент идентичности и/или схожести последовательности двух нуклеиновых кислот определяют с использованием алгоритма Karlin and Altschul, 1990, модифицированного, как указано в Karlin and Altschul, 1993. Такой алгоритм включен в программы NBLAST и XBLAST Altschul et al., 1990. Нуклеотидные поиски в BLAST осуществляют с использованием программы NBLAST, вес совпадений = 100, длина "слова" = 12. Можно использовать Gapped BLAST, как описано в Altschul et al., 1997. При использовании программ BLAST и Gapped BLAST используют параметры соответствующих программ по умолчанию (NBLAST и XBLAST). См. веб-сайт NCBI/NIH. В целях сравнения для получения выравнивания с пропусками использовали функцию AlignX Vector NTI Suite 8 (InforMax, Inc., North Bethesda, Md., U.S.A.) с параметрами по умолчанию. Они являлись следующими: штраф за открытие пропуска - 15, штраф за удлинение пропуска - 6,66, и штраф за разделение пропуска - 8.
Также можно изменять различные свойства и трехмерные черты белка без неблагоприятного воздействия на его активность/функциональность. Консервативные аминокислотные замены можно допускать/включать таким образом, чтобы не влиять неблагоприятно на активность и/или трехмерную конфигурацию молекулы. Аминокислоты можно разделять на следующие классы: неполярные, незаряженные полярные, основные и кислые. Консервативные замены, при которых аминокислоту одного класса заменяют другой аминокислотой того же типа, попадают в объем настоящего изобретения при условии, что замена не влияет неблагоприятно на биологическую активность соединения. В таблице 1 представлен список примеров аминокислот, принадлежащих к каждому классу. В некоторых случаях также можно осуществлять неконсервативные замены. Однако предпочтительные замены не изменяют значительно функциональную/биологическую активность белка.
Как применяют в настоящем описании, ссылка на "выделенные полинуклеотиды и/или "очищенные" белки относится к этим молекулам, когда они не связаны с другими молекулами, с которыми их обнаруживают в природе. Таким образом, ссылка на "выделенные" и/или "очищенные" означает участие "рук человека", как представлено в настоящем описании. Например, бактериальный "ген" по настоящему изобретению, помещенный в растение для экспрессии, является "выделенным полинуклеотидом". Аналогично, белок, полученный из бактериального белка и продуцируемый растением, является "выделенным белком".
По причине вырожденности/избыточности генетического кода аминокислотные последовательности, представленные в настоящем описании, могут кодироваться множеством различных последовательностей ДНК. Специалист в этой области может получать альтернативные последовательности ДНК, кодирующие одинаковые или, по существу, одинаковые белки. Эти варианты последовательностей ДНК входят в объем настоящего изобретения. Это более подробно описано ниже в разделе под названием "Оптимизация последовательности для экспрессии в растениях".
Оптимизация последовательности для экспрессии в растениях: Для достижения высокого уровня экспрессии гетерологичных генов в растениях, как правило, предпочтительно реконструировать гены таким образом, чтобы они более эффективно экспрессировались в растительных клетках (цитоплазме). Кукуруза является одним из таких растений, в которых предпочтительным может являться реконструирование гетерологичных генов перед трансформацией для повышения уровня их экспрессии в указанном растении. Таким образом, дополнительным этапом в конструировании генов, кодирующих бактериальный белок, является реконструирование гетерологичного гена для оптимальной экспрессии с использованием отклонения частоты использования кодонов, более соответствующего последовательности-мишени растения, принадлежащего к двудольным или однодольным видам. Последовательности также можно оптимизировать для экспрессии в любом из более конкретных типов растений, обсуждаемых где-либо в настоящем описании.
Трансгенные хозяева: Гены по настоящему изобретению, кодирующие белки, можно встраивать в широкий спектр микроорганизмов- или растений-хозяев. Настоящее изобретение включает трансгенные растительные клетки и трансгенные растения. Предпочтительными растениями (и растительными клетками) являются кукуруза, арабидопсис, табак, соя, хлопок, канола, рис, пшеница, газонная трава, бобово-злаковые растения (например, люцерна и клевер), пастбищные злаки и т.п. По настоящему изобретению также можно получать другие типы трансгенных растений, такие как плодовые растения, овощные культуры, декоративные растения и деревья. В общем, в различных аспектах по настоящему изобретению можно использовать двудольные и/или однодольные растения.
В предпочтительных вариантах осуществления экспрессия гена, прямо или косвенно, приводит к внутриклеточной продукции (и сохранению) интересующих белков. Таким образом, растения можно делать резистентными к гербицидам. Таких хозяев можно обозначать как трансгенные, рекомбинантные, трансформированные и/или трансфицированные хозяева и/или клетки. В некоторых аспектах по настоящему изобретению (например, при клонировании и получении интересующего гена) клетки микроорганизмов (предпочтительно, бактерий) можно получать и использовать стандартными способами с выгодой раскрытия предмета изобретения.
Растительные клетки, трансфицированные с использованием полинуклеотида по настоящему изобретению, можно подвергать регенерации в целые растения. Настоящее изобретение включает культуры клеток, включая культуры клеток тканей, жидкие культуры и культуры на твердой среде. Семена, продуцируемые и/или используемые для получения растений по настоящему изобретению, также входят в объем настоящего изобретения. Другие растительные ткани и части также включены в настоящее изобретение. Настоящее изобретение аналогично включает способы получения растений или клеток, содержащих полинуклеотид по настоящему изобретению. Одним из предпочтительных способов получения таких растений является посадка семян по настоящему изобретению.
Хотя растения могут являться предпочтительными, настоящее изобретение также включает продукцию высокоактивной рекомбинантной AAD-12, например, в штамме-акцепторе Pseudomonas fluorescens (Pf). Настоящее изобретение включает предпочтительные температуры роста для поддержания растворимой активной AAD-12 в этом хозяине; условия ферментации, в которых AAD-12 продуцируется как более 40% общего белка клетки или по меньшей мере 10 г/л; способ очистки, приводящий к эффективному выделению активной рекомбинантной AAD-12 из Pf-хозяина; схему очистки, выход которой составляет по меньшей мере 10 г активной AAD-12 на кг клеток; схему очистки, выход которой может составлять 20 г активной AAD-12 на кг клеток; способ составления, при котором можно хранить AAD-12 и восстанавливать ее активность в растворе; и способ лиофилизации, при котором можно сохранять активность AAD-12 для длительного хранения и срока годности.
Встраивание генов для получения трансгенных хозяев: Одним из аспектов настоящего изобретения является трансформация/трансфекция растений, растительных клеток и других клеток-хозяев с использованием полинуклеотидов по настоящему изобретению, с которых экспрессируются белки по настоящему изобретению. Растения, трансформированные таким образом, можно делать резистентными к различным гербицидам с различными механизмами действия.
Доступен широкий спектр способов встраивания гена, кодирующего желаемый белок, в хозяина-мишень в условиях, делающих возможным стабильное сохранение и экспрессию гена. Эти способы хорошо известны специалистам в этой области и описаны, например, в патенте США № 5135867.
Векторы, содержащие полинуклеотид AAD-12, включены в объем настоящего изобретения. Например, для подготовки к встраиванию чужеродных генов в высшие растения доступно большое количество клонирующих векторов, содержащих систему репликации в E. coli и маркер, делающий возможной селекцию трансформированных клеток. Векторы включают, например, pBR322, серию pUC, серию M13 mp, pACYC184,и т.д. Таким образом, последовательность, кодирующую белок, можно встраивать в вектор в подходящий участок рестрикции. Полученную плазмиду используют для трансформации E. coli. Клетки E. coli культивируют в подходящей питательной среде, затем собирают и лизируют. Плазмиду выделяют посредством очистки от геномной ДНК. В качестве способов анализа, как правило, осуществляют, анализ последовательности, рестрикционный анализ, электрофорез и другие способы биохимии и молекулярной биологии. После каждой манипуляции используемую последовательность ДНК можно рестрицировать и соединять со следующей последовательностью ДНК. Последовательность каждой плазмиды можно клонировать в одинаковые или разные плазмиды. В зависимости от способа встраивания желаемых генов в растение, необходимыми могут являться другие последовательности ДНК. Например, если для трансформации растительной клетки используют плазмиду Ti или Ri, то, по меньшей мере, правый край, но часто и правый край, и левый край Т-ДНК плазмиды Ti или Ri необходимо соединять в виде фланкирующих областей генов, подлежащих встраиванию. Использование Т-ДНК для трансформации растительных клеток активно исследуют и описывают в EP 120 516; Hoekema (1985); Fraley et al. (1986); и An et al. (1985).
Для встраивания ДНК в растительную клетку хозяина доступно большое количество способов. Эти способы включают трансформацию с помощью Т-ДНК с использованием Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве объектов трансформации, слияние, инъекцию, биолистику (бомбардировку микрочастицами), использование вискеров карбида кремния, обработку аэрозольным путем, PEG или электропорацию, а также другие возможные способы. При использовании Agrobacteria для трансформации, ДНК, подлежащую встраиванию, необходимо клонировать в специальные плазмиды, а именно в промежуточный вектор или бинарный вектор. Промежуточные векторы можно встраивать в плазмиду Ti или Ri посредством гомологичной рекомбинации благодаря последовательностям, гомологичным последовательностям в Т-ДНК. Плазмида Ti или Ri также содержит vir-область, необходимую для переноса Т-ДНК. Промежуточные векторы не могут самостоятельно реплицироваться в Agrobacteria. Промежуточный вектор можно переносить в Agrobacterium tumefaciens посредством плазмиды-помощника (конъюгации). Бинарные векторы могут самостоятельно реплицироваться и в E. coli, и в Agrobacteria. Они содержат ген селективного маркера и линкер или полилинкер, фланкированные правой и левой краевыми областями Т-ДНК. С помощью них можно напрямую трансформировать Agrobacteria (Holsters, 1978). Agrobacterium, используемая в качестве клетки-хозяина, должна содержать плазмиду, несущую vir-область. Vir-область необходима для переноса Т-ДНК в растительную клетку. Можно включать дополнительную Т-ДНК. Трансформированную таким образом бактерию используют для трансформации растительных клеток. Для переноса ДНК в растительную клетку растительные эксплантаты можно культивировать преимущественно с Agrobacterium tumefaciens или Agrobacterium rhizogenes. Затем в подходящей среде, которая может содержать антибиотики или биоциды для селекции, из инфицированного растительного материала (например, кусочков листьев, сегментов стебля, корней, а также протопластов или клеток, культивируемых в суспензии) можно осуществлять регенерацию целых растений. Затем получаемые таким образом растения можно тестировать на наличие встроенной ДНК. В случае инъекции и электропорации для плазмид нет специальных требований. Можно использовать общепринятые плазмиды, такие как, например, производные pUC.
Трансформированные клетки растут внутри растений обычным образом. Они могут образовывать половые клетки и передавать трансформированные признаки потомству растений. Такие растения можно выращивать общепринятым образом и скрещивать с растениями, имеющими те же трансформированные наследственные факторы или другие наследственные факторы. Полученные гибридные особи имеют соответствующие фенотипические свойства. В некоторых предпочтительных вариантах осуществления изобретения гены, кодирующие бактериальный белок, экспрессируются из транскрипционных единиц, встроенных в геном растения. Предпочтительно, указанными транскрипционными единицами являются рекомбинантные векторы, способные к стабильной интеграции в геном растения и делающие возможной селекцию линий трансформированных растений, экспрессирующих мРНК, кодирующую белки.
При встраивании ДНК интегрируется в геном и является в нем относительно стабильной (и не выходит обратно). В норме она содержит селективный маркер, придающий трансформированным растительным клеткам резистентность к биоциду или антибиотику, такому как, помимо прочего, канамицин, G418, блеомицин, гигромицин или хлорамфеникол. Селективные маркеры растений, как правило, также могут придавать резистентность к различным гербицидам, таким как глуфосинат (например, PAT/bar), глифосат (EPSPS), ингибиторы ALS (например, имидазолинон, сульфонилкарбамид, триазолопиримидин сульфонанилид и др.), бромоксинил, резистентность к ингибиторам HPPD, ингибиторам PPO, ингибиторам ACC-ase и многим другим. Таким образом, используемый в отдельности маркер должен делать возможной селекцию трансформированных клеток, а не клеток, не содержащих встроенную ДНК. Предпочтительно, интересующие гены экспрессируются в растительной клетке с помощью конститутивных или индуцибельных промоторов. При экспрессии мРНК транслируется в белки, таким образом, интересующие аминокислоты встраиваются в белок. Гены, кодирующие белок, экспрессирующийся в растительных клетках, могут находиться под контролем конститутивного промотора, тканеспецифического промотора или индуцибельного промотора.
Существует несколько способов встраивания чужеродных рекомбинантных векторов в растительные клетки и получения растений, стабильно сохраняющих и экспрессирующих встроенный ген. Такие способы включают встраивание генетического материала, которым покрывают микрочастицы, непосредственно в клетки (патенты США №№ 4945050 Cornell и 5141131 DowElanco, в настоящее время Dow AgroSciences, LLC). Кроме того, растения можно трансформировать с помощью технологии с использованием Agrobacterium, см. патенты США №№ 5177010 University of Toledo; 5104310 Texas A&M; европейскую патентную заявку № 0131624B1; европейские патентные заявки №№ 120516, 159418B1 и 176112 Schilperoot; патенты США №№ 5149645, 5469976, 5464763, 4940838 и 4693976 Schilperoot; европейские патентные заявки №№ 116718, 290799, 320500 Max Planck; европейские патентные заявки №№ 604662 и 627752 и патент США № 5591616 Japan Tobacco; европейские патентные заявки №№ 0267159 и 0292435 и патент США № 5231019 Ciba Geigy, в настоящее время Syngenta; патенты США №№ 5463174 и 4762785 Calgene; и патенты США №№ 5004863 и 5159135 Agracetus. Другая технология трансформации включает технологию с использованием вискеров. См. патенты США №№ 5302523 и 5464765 Zeneca, в настоящее время Syngenta. Другая технология трансформации с прямой доставкой ДНК включает технологию aerosol beam. См. патент США № 6809232. Для трансформации растений также используют технологию электропорации. См. WO 87/06614 Boyce Thompson Institute; патенты США №№ 5472869 и 5384253 Dekalb; и WO 92/09696 и WO 93/21335 Plant Genetic Systems. Кроме того, для получения трансгенных растений, экспрессирующих интересующий белок, также можно использовать вирусные векторы. Например, однодольные растения можно трансформировать с помощью вирусного вектора с использованием способов, описываемых в патенте США № 5569597 Mycogen Plant Science и Ciba-Geigy (в настоящее время Syngenta), а также патентах США №№ 5589367 и 5316931 Biosource, в настоящее время Large Scale Biology.
Как указано выше, способ, которым конструкцию ДНК встраивают в растение-хозяина, не является критичным для настоящего изобретения. Можно использовать любой способ, обеспечивающий эффективную трансформацию. Например, различные способы трансформации растительных клеток представлены в настоящем описании и включают использование плазмид Ti или Ri и т.п. для осуществления опосредованной Agrobacterium трансформации. Во многих случаях желательно иметь конструкцию, используемую для трансформации, фланкируемую с одной или обеих сторон краевыми областями Т-ДНК, более конкретно - правой краевой областью. Особенно полезно, когда для конструкции в качестве способа трансформации используют трансформацию, опосредованную Agrobacterium tumefaciens или Agrobacterium rhizogenes, хотя краевые области Т-ДНК могут найти применение в других способах трансформации. При использовании Agrobacterium для трансформации растительных клеток можно использовать вектор, который можно встраивать в хозяина для гомологичной рекомбинации с Т-ДНК или плазмидой Ti или Ri, присутствующих в хозяине. Встраивание вектора можно осуществлять посредством электропорации, трехродительского скрещивания и других способов трансформации грамотрицательных бактерий, известных специалистам в этой области. Способ трансформации Agrobacterium-хозяина с использованием вектора не является критичным для настоящего изобретения. Плазмида Ti или Ri, содержащая Т-ДНК для рекомбинации, может быть способна или неспособна вызывать образование галлов и не является критичной для указанного изобретения при условии, что в указанном хозяине присутствуют vir-гены.
В некоторых случаях при использовании Agrobacterium для трансформации экспрессирующую конструкцию внутри краевых областей Т-ДНК будут встраивать в широкий спектр векторов, таких как pRK2 или его производные, как описано в Ditta et al. (1980) и EPO 0 120 515. В экспрессирующую конструкцию и Т-ДНК будут включать один или несколько маркеров, как представлено в настоящем описании, делающих возможной селекцию трансформированных Agrobacterium и трансформированных растительных клеток. Конкретный используемый маркер не является существенным для настоящего изобретения, при этом предпочтительный маркер зависит от используемого хозяина и конструкции.
Для трансформации растительных клеток с использованием Agrobacterium эксплантаты можно комбинировать и инкубировать с трансформированными Agrobacterium в течение периода времени, достаточного, чтобы сделать возможной их трансформацию. После трансформации Agrobacteria уничтожают посредством селекции с использованием соответствующего антибиотика и культивируют растительные клетки с соответствующей селективной средой. После образования каллюсов можно стимулировать образование побега, используя соответствующие растительные гормоны, способами, хорошо известными в области культивирования растительных тканей и регенерации растений. Однако промежуточная стадия каллюса не всегда является обязательной. После образования побега указанные растительные клетки можно переносить в среду, стимулирующую образование корня, таким образом, завершая регенерацию растения. Затем растения можно выращивать для получения семян и указанные семена можно использовать для получения будущих поколений. Независимо от способа трансформации, ген, кодирующий бактериальный белок, предпочтительно, встраивают в вектор для переноса гена, адаптированный для экспрессии указанного гена в растительной клетке посредством включения в вектор промоторного регуляторного элемента растений, а также 3'-нетранслируемых областей терминации транскрипции, таких как Nos и т.п.
В дополнение к многочисленным технологиям трансформации растений также можно варьировать тип ткани, приводимой в контакт с чужеродными генами. Такая ткань будет включать, в качестве неограничивающих примеров, эмбриогенную ткань, ткань каллюса типов I, II и III, гипокотиль, меристему, ткань корня, ткани для экспрессии во флоэме и т.п. Почти все ткани растений можно трансформировать при дедифференцировке с использованием подходящих способов, представленных в настоящем описании.
Как указано выше, при желании можно использовать множество селективных маркеров. Предпочтение конкретного маркера остается на усмотрение специалиста в этой области, но можно использовать любой из следующих селективных маркеров вместе с любым другим геном, не указанным в настоящем описании, который может функционировать как селективный маркер. Такие селективные маркеры включают, в качестве неограничивающих примеров, ген аминогликозидфосфотрансферазы транспозона Tn5 (Aph II), кодирующий резистентность к антибиотикам канамицину, неомицину и G41; резистентность к гигромицину; резистентность к метотрексату, а также гены, кодирующие резистентность или устойчивость к глифосату; фосфинотрицину (биалафосу или глуфосинату); ALS-ингибирующим гербицидам (имидазолинонам, сульфонилкарбамидам и триазолопиримидиновым гербицидам), ингибиторам ACC-ase (например, арилоксипропионатам или циклогександионам) и др., таким как бромоксинил, и ингибиторам HPPD (например, мезотриону) и т.п.
В дополнение к селективному маркеру желательным может являться использование репортерного гена. В некоторых случаях репортерный ген можно использовать с селективным маркером или без него. Репортерными генами являются гены, как правило, не присутствующие в организме- или ткани-реципиенте и, как правило, кодирующие белки, приводящие к некоторым фенотипическим изменениям или ферментативному свойству. Примеры таких генов представлены в Weising et al., 1988. Предпочтительные репортерные гены включают ген бета-глюкуронидазы (GUS) локуса uidA E. coli, ген хлорамфениколацетилтрансферазы из Tn9 E. coli, ген зеленого флуоресцентного белка биолюминесцентной медузы Aequorea victoria и гены люциферазы светлячка Photinus pyralis. Затем можно осуществлять анализ для определения экспрессии репортерного гена в подходящее время после встраивания указанного гена в реципиентные клетки. Такой предпочтительный анализ включает использование гена, кодирующего бета-глюкуронидазу (GUS), локуса uidA E. coli, как описано в Jefferson et al., (1987), для идентификации трансформированных клеток.
В дополнение к промоторным регуляторным элементам растений в растительных клетках для экспрессии чужеродных генов можно эффективно использовать промоторные регуляторные элементы из множества источников. Например, можно использовать промоторные регуляторные элементы бактериального происхождения, такие как промотор октопинсинтазы, промотор нопалинсинтазы, промотор маннопинсинтазы; промоторы вирусного происхождения, такие как промотор вируса мозаики цветной капусты (35S и 19S), 35T (являющийся реконструированным промотором 35S, см. патент США № 6166302, особенно пример 7E) и т.п. Промоторные регуляторные элементы растений включают, в качестве неограничивающих примеров, промотор малой субъединицы рибулозо-1,6-бифосфат (RUBP)-карбоксилазы (ssu), промотор бета-конглицинина, промотор бета-фазеолина, промотор ADH, промоторы белков теплового шока и тканеспецифические промоторы. Могут присутствовать другие элементы, такие как последовательности прикрепления к матриксу, последовательности связывания с ядерным матриксом, интроны, энхансеры, последовательности полиаденилирования и т.п., и, таким образом, могут улучшать эффективность транскрипции или интеграцию ДНК. Такие элементы могут являться или не являться необходимыми для функционирования ДНК, хотя они могут обеспечивать лучшую экспрессию или функционирование ДНК, влияя на транскрипцию, стабильность мРНК и т.п. Такие элементы, при желании, можно включать в ДНК для достижения оптимального функционирования трансформированной ДНК в растении. Типичные элементы включают, в качестве неограничивающих примеров, Adh-интрон 1, Adh-интрон 6, лидерную последовательность белка оболочки вируса мозаики люцерны, последовательности UTR осмотина, лидерную последовательность белка оболочки вируса полосчатости кукурузы, а также другие, доступные специалистам в этой области. Также можно использовать конститутивные промоторные регуляторные элементы, таким образом, регулируя непрерывную экспрессию гена во всех типах клеток и в любой момент времени (например, актин, убиквитин, CaMV 35S и т.п.). Тканеспецифические промоторные регуляторные элементы отвечают за экспрессию гена в конкретных типах клеток или тканей, таких как листья или семена (например, зеин, олеозин, напин, ACP, глобулин и т.п.), и их также можно использовать.
Промоторные регуляторные элементы также могут являться активными (или неактивными) в течение конкретной фазы роста растения, а также активными в тканях и органах растения. Их примеры включают, в качестве неограничивающих примеров, специфические для пыльцы, эмбриоспецифические, специфические для кукурузных рылец, специфические для хлопкового волокна, специфические для корней, специфические для эндосперма семян и т.п. В конкретных обстоятельствах желательным может являться использование индуцибельного промоторного регуляторного элемента, отвечащего за экспрессию генов в ответ на конкретный сигнал, такой как: физический стимул (гены белков теплового шока), свет (RUBP-карбоксилаза), гормон (Em), метаболиты, химические вещества (чувствительные к тетрациклину) и стресс. Можно использовать другие желательные транскрипционные и трансляционные элементы, функционирующие в растениях. В этой области известно множество специфичных для растений векторов для переноса генов.
Для экспрессии бактериального белка также можно использовать системы на основе РНК-вирусов растений. При этом ген, кодирующий белок, можно встраивать в промоторную область гена белка оболочки подходящего вируса растений, которым инфицируют интересующее растение-хозяина. Затем белок может экспрессироваться, таким образом, обеспечивая защиту растения от повреждения гербицидов. Системы на основе РНК-вирусов растений описывают в патенте США № 5500360 Mycogen Plant Sciences, Inc. и патентах США №№ 5316931 и 5589367 Biosource, в настоящее время Large Scale Biology.
Средства дополнительного повышения уровней устойчивости или резистентности. В настоящем описании показано, что растениям по настоящему изобретению можно придавать новые признаки резистентности к гербицидам без наблюдаемых неблагоприятных воздействий на фенотип, включая урожайность. Такие растения входят в объем настоящего изобретения. Растения, приведенные в качестве примеров и предложенные в настоящем описании, могут выдерживать 2-, 3-, 4- и 5-кратные типичные нормы внесения, например, по меньшей мере одного гербицида по настоящему изобретению. Улучшения этих уровней устойчивости входят в объем настоящего изобретения. Например, в этой области известны различные способы, и их предсказуемо можно оптимизировать и разрабатывать далее для повышения экспрессии указанного гена.
Один из таких способов включает повышение копийности генов AAD-12 по настоящему изобретению (в экспрессирующих кассетах и т.п.). Трансформированные объекты также можно подвергать селекции в пользу имеющих множество копий генов.
Можно использовать сильные промоторы и энхансеры для "форсирования" экспрессии. Примеры таких промоторов включают предпочтительный промотор 35T, в котором используют энхансеры 35S. Для таких целей включены промоторы 35S, убиквитина кукурузы, убиквитина арабидопсиса, актина A.t. и CSMV. Также предпочтительными являются другие сильные вирусные промоторы. Энхансеры включают двойной энхансер 4 OCS и 35S. Для повышения эффективности трансформации и экспрессии трансгена также можно использовать последовательности прикрепления к матриксу (MAR).
Для вариантов осуществления настоящего изобретения также можно использовать перетасовку (направленное развитие) и факторы транскрипции.
Также можно конструировать варианты белков, отличающиеся на уровне последовательности, но сохраняющие ту же или схожую общую необходимую трехмерную структуру, поверхностное распределение заряда и т.п. См., например, патент США № 7058515; Larson et al., Protein Sci. 2002 11: 2804-2813, “Thoroughly sampling sequence space: Large-scale protein design of structural ensembles”; Crameri et al., Nature Biotechnology 15, 436-438 (1997), “Molecular evolution of an arsenate detoxification pathway by DNA shuffling”; Stemmer, W. P. C. 1994. “DNA shuffling by random fragmentation and reassembly: in vitro recombination for molecular evolution” Proc. Natl. Acad. Sci. USA 91: 10747-10751; Stemmer, W. P. C. 1994. “Rapid evolution of a protein in vitro by DNA shuffling” Nature 370: 389-391; Stemmer, W. P. C. 1995. Searching sequence space. Bio/Technology 13: 549-553; Crameri, A., et al. 1996. “Construction and evolution of antibody-phage libraries by DNA shuffling” Nature Medicine 2: 100-103; and Crameri, A., et al. 1996. “Improved green fluorescent protein by molecular evolution using DNA shuffling” Nature Biotechnology 14: 315-319.
Активность рекомбинантных полинуклеотидов, встроенных в растительные клетки, может зависеть от влияния эндогенной ДНК растений, смежной со вставкой. Таким образом, другим вариантом является использование преимущества объектов, о которых известно, что они являются прекрасными участками для инсерций в геноме растений. См. например WO 2005/103266 A1, относящийся к объектам хлопка crylF и crylAc; геном AAD-12 по настоящему изобретению можно замещать в локусах генома вставки crylF и/или crylAc. Таким образом, по настоящему изобретению можно использовать, например, направленную гомологичную рекомбинацию. Этот тип технологии является объектом, например, WO 03/080809 A2 и соответствующей опубликованной патентной заявки США № 20030232410, относящейся к использованию цинковых пальцев для направленной рекомбинации. Также известно использование рекомбиназы (например, cre-10 x и flp-frt).
Считают, что детоксификация AAD-12 происходит в цитоплазме. Таким образом, аспекты настоящего изобретения включают средства для дополнительной стабилизации этого белка и мРНК (включая блокирование деградации мРНК), и, таким образом, можно использовать известные в этой области способы. Можно конструировать белки, резистентные к деградации протеазами и т.п. (участки расщепления протеазами можно эффективно удалять, реконструируя аминокислотную последовательность белка). Такие варианты осуществления включают использование 5'- и 3'-структуры типа "стебель-петля", таких как UTR из осмотина, и per5 (AU-богатые 5'-нетранслируемые последовательности). Также можно использовать 5'-кэпы, такие как 7-метил- или 2'-O-метильные группы, например, остаток 7-метилгуаниловой кислоты. См., например: Proc. Natl. Acad. Sci. USA Vol. 74, No. 7, pp. 2734-2738 (July 1977) Importance of 5'-terminal blocking structure to stabilize mRNA in eucaryotic protein synthesis. Также можно использовать белковые комплексы или лиганд-блокирующие группы.
В объеме настоящего изобретения также можно осуществлять компьютерный дизайн 5'- или 3'-UTR, наиболее подходящих для AAD-12 (синтетические шпильки). Компьютерное моделирование в целом, а также перетасовка генов и направленное развитие описывают где-либо в настоящем описании. Более конкретно, что касается компьютерного моделирования и UTR, способы компьютерного моделирования для использования в прогнозировании/оценке производных 5'- и 3'-UTR по настоящему изобретению включают, в качестве неограничивающих примеров: MFold версии 3.1, доступный в Genetics Corporation Group, Madison, Wis. (см. Zucker et al., Algorithms and Thermodynamics for RNA Secondary Structure Prediction: A Practical Guide. In RNA Biochemistry and Biotechnology, 11-43, J. Barciszewski & B. F. C. Clark, eds., NATO ASI Series, Kluwer Academic Publishers, Dordrecht, NL, (1999); Zucker et al., Expanded Sequence Dependence of Thermodynamic Parameters Improves Prediction of RNA Secondary Structure. J. Mol. Biol. 288, 911-940 (1999); Zucker et al., RNA Secondary Structure Prediction. In Current Protocols in Nucleic Acid Chemistry S. Beaucage, D. E. Bergstrom, G. D. Glick, and R. A. Jones eds., John Wiley & Sons, New York, 11.2.1-11.2.10, (2000)), COVE (анализ структуры РНК с использованием ковариационных моделей (стохастических бесконтекстных грамматических способов)) v. 2.4.2 (Eddy & Durbin, Nucl. Acids Res. 1994, 22: 2079-2088) свободно распространяемый в качестве исходной программы и который можно загружать посредством доступа к веб-сайту genetics.wustl.edu/eddy/software/, и FOLDALIGN, также свободно распространяемый и доступный для загрузки на веб-сайте bioinf.au.dk.FOLDALIGN/ (см. Finding the most significant common sequence and structure motifs in a set of RNA sequences. J. Gorodkin, L. J. Heyer and G. D. Stormo. Nucleic Acids Research, Vol. 25, no. 18 pp 3724-3732, 1997; Finding Common Sequence and Structure Motifs in a set of RNA Sequences. J. Gorodkin, L. J. Heyer, and G. D. Stormo. ISMB 5; 120-123, 1997).
Варианты осуществления по настоящему изобретению можно использовать в комбинации с естественно развившимися или химически индуцированными мутантами (мутантов можно подвергать селекции с помощью способов скрининга, а затем трансформировать с использованием AAD-12 и, возможно, других генов). Можно комбинировать растения по настоящему изобретению с резистентностью к ALS и/или развившейся резистентностью к глифосату. Резистентность к аминопиралиду, например, также можно комбинировать или "подвергать стэкингу" с геном AAD-12.
Общепринятые способы скрещивания также можно комбинировать с настоящим изобретением для получения сильной комбинации, интрогрессии и улучшения желаемых признаков.
Дополнительные улучшения также включают использование вместе с соответствующими антидотами для дополнительной защиты растений и/или для приданий перекрестной резистентности к большему количеству гербицидов. Антидоты, как правило, действуют, усиливая иммунную систему растений посредством активации/экспрессии cP450. Антидоты являются химическими средствами, снижающими фитотоксичность гербицидов в отношении сельскохозяйственных культур с помощью физиологического или молекулярного механизма, не нарушая эффективность контроля сорняков.
Антидоты гербицидов включают беноксакор, клоквинтоцет, циометринил, дихлормид, дициклонон, диэтолят, фенхлоразол, фенклорим, флуразол, флуксофеним, фурилазол, изоксадифен, мефенпир, мефенат, нафтойный ангидрид и оксабетринил. В вариантах осуществления настоящего изобретения также можно использовать растительные активаторы (новый класс соединений, защищающих растения посредством активации их механизмов защиты). Они включают ацибензолар и пробеназол.
Можно использовать коммерчески доступные антидоты для защиты крупносеменных травянистых сельскохозяйственных культур, таких как кукуруза, сорго обыкновенное и культура высеваемого в воду риса, при нанесении предпосевных или предвсходовых гербицидов из семейств тиокарбамата и хлорацетанилида. Также разрабатывают антидоты для защиты озимых зерновых сельскохозяйственных культур, таких как пшеница, при послевсходовом нанесении арилоксифеноксипропионатных и сульфонилкарбамидных гербицидов. Также хорошо известно использование антидотов для защиты кукурузы и риса от гербицидов сульфонилкарбамида, имидазолинона, циклогександиона, изоксазола и трикетона. Индуцированное антидотом повышение детоксификации гербицида в защищенных растениях широко признают в качестве основного механизма действия антидота. Антидоты индуцируют кофакторы, такие как глутатион и гербицид-детоксифицирующие ферменты, такие как глутатион-S-трансферазы, цитохром-P450-монооксигеназы и глюкозилтрансферазы. Hatzios K K, Burgos N (2004) "Metabolism-based herbicide resistance: regulation by safeners," Weed Science: Vol. 52, No. 3 pp. 454-467.
Использование гена цитохром-p450-монооксигеназы, подвергнутого стэкингу с AAD-12, является одним из предпочтительных вариантов осуществления. Существуют P450, участвующие в метаболизме гербицидов; cP450 может иметь, например, происхождение из млекопитающего или растительное происхождение. Известно, что у высших растений цитохром-P450-монооксигеназа (P450) осуществляет вторичный метаболизм. Она также играет важную роль в окислительном метаболизме ксенобиотиков во взаимодействии с NADPH-цитохром-P450-оксидоредуктазой (редуктазой). Сообщают, что резистентность к некоторым гербицидам является результатом метаболизма с помощью P450, а также глутатион-S-трансферазы. С помощью молекулярного клонирования охарактеризовали ряд микросомальных видов P450, участвующих в метаболизме ксенобиотиков у млекопитающих. Сообщают, что некоторые из них эффективно метаболизируют несколько гербицидов. Таким образом, трансгенные растения с P450 растений или млекопитающих могут демонстрировать резистентность к нескольким гербицидам.
Одним из предпочтительных вариантов осуществления изложенного выше является применение cP450 для резистентности к ацетохлору (продукты на основе ацетохлора включают гербициды Surpass®, Keystone®, Keystone LA, FulTime® и TopNotch®) и/или трифлуралину (такому как Treflan®). Некоторые предпочтительные варианты осуществления включают такую резистентность у сои и/или кукурузы. Дополнительные инструкции, касающиеся таких вариантов осуществления, см., например, в Inui et al., "A selectable marker using cytochrome P450 monooxygenases for Arabidopsis transformation," Plant Biotechnology 22, 281-286 (2005) (относящейся к системе селекции для трансформации Arabidopsis thaliana посредством Agrobacterium tumefaciens, в которой используют цитохром-P450-монооксигеназы человека, метаболизирующие гербициды; устойчивые к гербицидам саженцы трансформировали и подвергали селекции с использованием гербицидов ацетохлор, амипрофос-метил, хлорпрофам, хлорсульфурон, норфлуразон и пендиметалин); Siminszky et al., "Expression of a soybean cytochrome P450 monooxygenase cDNA in yeast and tobacco enhances the metabolism of phenylurea herbicides," PNAS Vol. 96, Issue 4, 1750-1755, Feb. 16, 1999; Sheldon et al, Weed Science: Vol. 48, No. 3, pp. 291-295, "A cytochrome P450 monooxygenase cDNA (CYP71A10) confers resistance to linuron in transgenic Nicotiana tabacum"; and "Phytoremediation of the herbicides atrazine and metolachlor by transgenic rice plants expressing human CYP1A1, CYP2B6, and CYP2C19," J Agric Food Chem. 2006 Apr. 19; 54(8):2985-91 (относящиеся к тестированию цитохром-p450-монооксигеназы человека в рисе, где, как сообщают, растения риса демонстрировали высокую устойчивость к хлорацетамидам (ацетохлору, алахлору, метоахлору, претилахлору и тенилхлору), оксиацетамидам (мефенацету), пиридазинонам (норфлуразону), 2,6-динитроаналинам (трифлуралину и пендиметалину), фосфамидатам (амипрофос-метилу, тиокарбаматам (пирибутикарбу) и мочевине (хлортолурону)).
Также можно изменять или использовать различные химические составы 2,4-D, чтобы сделать гены AAD-12 по настоящему изобретению более эффективными. Такие возможные изменения включают получение лучших субстратов и лучших уходящих групп (большей электроотрицательности). Для повышения гербицидной активности вместе с 2,4-D также можно использовать ингибиторы транспорта ауксинов (например, дифлуфензопир).
Если конкретно не указано или очевидно иное, термины в единственном числе означают "по меньшей мере один", как применяют в настоящем описании. Все патенты, патентные заявки, предварительные заявки и публикации, указанные или процитированные в настоящем описании, включены в настоящее описание в качестве ссылок в полном объеме до тех пор, пока они не противоречат представленному в настоящем описании.
Далее приведены примеры, в которых иллюстрируют способы практического осуществления изобретения.
Эти примеры нельзя истолковывать как ограничивающие. Все процентные доли представляют собой доли по массе, и все пропорции в смесях растворителей являются пропорциями по объему, если не указано иначе.
ПРИМЕРЫ
ПРИМЕР 1
Способ идентификации генов, придающих резистентность к 2,4-D в полевых условиях
Как способ идентификации генов, обладающих свойствами деградации гербицидов в полевых условиях, можно осуществлять поиск в существующих на настоящий момент общедоступных базах данных, таких как NCBI (National Center for Biotechnology Information). Для начала необходимо иметь уже идентифицированную последовательность функционального гена, кодирующего белок с желаемыми свойствами (т.е. активностью α-кетоглутаратдиоксигеназы). Затем эту белковую последовательность используют в качестве вводной информации для алгоритма BLAST (Basic Local Alignment Search Tool) (Altschul et al., 1997) для сравнения с доступными белковыми последовательностями, хранящимися в NCBI. Используя параметры по умолчанию, с помощью этого поиска получают приблизительно 100 гомологичных белковых последовательностей на различных уровнях. Они находятся в диапазоне от высокоидентичных (85-98%) до малоидентичных (23-32%) на уровне аминокислот. Как правило, ожидают, что только последовательности с высокой гомологией будут сохранять свойства, схожие со свойствами вводной последовательности. В этом случае будут выбирать только последовательности с ≥50% гомологии. Как приведено в настоящем описании в качестве примера, можно использовать клонирование и рекомбинантно экспрессирующихся гомологов всего с 31% консервативных аминокислот (относительно tfdA из Ralstonia eutropha) для придания коммерческих уровней резистентности не только к предполагаемому гербициду, но также и к субстратам, ранее никогда не тестированными с этими ферментами.
С помощью базы данных NCBI (см. веб-сайт ncbi.nlm.nih.gov; учетный № AF516752) в качестве гомолога всего с 31% идентичности аминокислот относительно tfdA идентифицировали единственный ген (sdpA). Процент идентичности определяли, сначала переводя последовательности ДНК sdpA и tfdA, хранящиеся в базе данных, в белки, затем используя ClustalW в пакете программ VectorNTI для осуществления множественного выравнивания последовательностей.
ПРИМЕР 2
Оптимизация последовательности для экспрессии в растениях и бактериях
Для достижения более высоких уровней экспрессии гетерологичных генов в растениях предпочтительным может являться реконструирование кодирующей белок последовательности генов таким образом, чтобы они более эффективно экспрессировались в растительных клетках. Кукуруза является одним из таких растений, где предпочтительным может являться реконструирование гетерологичной области, кодирующей белок, перед трансформацией для повышения уровня экспрессии гена и уровня кодируемого белка в растении. Таким образом, дополнительным этапом в конструировании генов, кодирующих бактериальный белок, является реконструирование гетерологичного гена для оптимальной экспрессии.
Обобщение данных о содержании G+C в областях генов кукурузы, кодирующих белок
Одной из причин реконструирования бактериального белка для экспрессии в кукурузе является неоптимальное содержание G+C в нативном гене. Например, очень низкое содержание G+C во многих нативных бактериальных генах (и последующее смещение в сторону высокого содержания A+T) приводит к образованию последовательностей, имитирующих или дублирующих последовательности контроля гена растения, которые, как известно, являются, в значительной степени богатыми A+T. Наличие некоторых A+T-богатых последовательностей в ДНК генов, встраиваемых в растения, (например, областей TATA-бокс, в норме обнаруживаемых в промоторах генов) может приводить к аномальной транскрипции генов. С другой стороны, наличие других регуляторных последовательностей в транскрибированной мРНК (например, последовательностей сигнала полиаденилирования (AAUAAA) или последовательностей, комплементарных малой ядерной РНК, участвующей в сплайсинге пре-мРНК) может приводить к нестабильности РНК. Таким образом, одной из целей при конструировании генов, кодирующих бактериальный белок, для экспрессии в кукурузе, более предпочтительно, обозначаемых как гены, оптимизированные для растений, является получение последовательности ДНК, имеющей более высокое содержание G+C, и, предпочтительно, близкое к таковому в генах кукурузы, кодирующих метаболические ферменты. Другой целью при конструировании генов, оптимизированных для растений и кодирующих бактериальный белок, является получение последовательности ДНК, в которой модификации последовательности не препятствуют трансляции.
В таблице 2 показано, насколько высоким является содержание G+C в кукурузе. Что касается данных в таблице 2, кодирующие области генов получали из записей в GenBank (выпуск 71), и композиции оснований вычисляли с использованием программы MacVector™ (Accelerys, San Diego, Calif.). При вычислениях игнорировали последовательности интронов.
По причине пластичности, обеспечиваемой избыточностью/вырожденностью генетического кода (т.е. некоторые аминокислоты определяются несколькими кодонами), эволюция геномов в различных организмах или классах организмов привела к различному использованию избыточных кодонов. Это "отклонение частоты использования кодонов" отражено в средней композиции оснований в областях, кодирующих белок. Например, организмы с относительно низким содержанием G+C используют кодоны, содержащие A или T в третьем положении избыточных кодонов, в то время как организмы, имеющие высокое содержание G+C, используют кодоны, содержащие G или C в третьем положении. Считают, что наличие "минорных" кодонов в мРНК может снижать абсолютную скорость трансляции этой мРНК, особенно когда относительный избыток нагруженной тРНК, соответствующей минорному кодону, является низким. Продолжением этого является то, что снижение скорости трансляции посредством отдельных минорных кодонов будет, по меньшей мере, аддитивным для многочисленных минорных кодонов. Таким образом, мРНК, имеющая высокое относительное содержание минорных кодонов, будет иметь, соответственно, низкую скорость трансляции. Эту скорость будут отражаться в последующих низких уровнях кодируемого белка.
Предпочтительные кодоны для аминокислот в белках, экспрессирующихся в кукурузе
При конструировании генов, кодирующих бактериальный белок, для экспрессии в кукурузе (или другом растении, таком как хлопок или соя) определяют отклонение частоты использования кодонов в растении. Отклонение частоты использования кодонов в кукурузе является статистическим распределением кодонов, которые растение использует для кодирования своих белков, и предпочтительное использование кодонов представлено в таблице 3. После определения отклонения определяют процентную частоту кодонов в интересующих генах. Необходимо определять предпочтительные первичные кодоны в растении, а также предпочтительные кодоны второго, третьего и четвертого выбора, если существует возможность множественного выбора. Затем можно конструировать новую последовательность ДНК, кодирующую аминокислотную последовательность в бактериальном белке, но новая последовательность ДНК отличается от нативной бактериальной последовательности ДНК (кодирующей белок) заменами на кодоны растения (первые предпочтительные, вторые предпочтительные, третьи предпочтительные или четвертые предпочтительные) для определения аминокислот в каждом положении в аминокислотной последовательности белка. Затем новую последовательность анализируют на предмет участков распознавания рестрикционных ферментов, которые можно получать посредством модификации. Идентифицированные участки дополнительно модифицируют, заменяя кодоны предпочтительными кодонами первого, второго, третьего или четвертого выбора. Другими участками в последовательности, которые могут влиять на транскрипцию или трансляцию интересующего гена, являются экзон-интронные сочленения (5' или 3'), сигналы полиаденилирования или сигналы терминации для РНК-полимеразы. Последовательность дополнительно анализируют и модифицируют для снижения частоты TA- или GC-дуплетов. В дополнение к дуплетам, блоки G или C в последовательности, содержащие приблизительно более четырех одинаковых остатков, могут влиять на транскрипцию последовательности. Таким образом, эти блоки также модифицируют, заменяя кодоны первого или второго выбора и т.д. предпочтительными кодонами следующего выбора.
Предпочтительно, гены, оптимизированные для растений и кодирующие бактериальный белок, содержат приблизительно 63% кодонов первого выбора, от приблизительно 22% до приблизительно 37% кодонов второго выбора и от приблизительно 15% до приблизительно 0% кодонов третьего или четвертого выбора, где общая процентная доля составляет 100%. Наиболее предпочтительно, гены, оптимизированные для растений, содержат приблизительно 63% кодонов первого выбора, по меньшей мере приблизительно 22% кодонов второго выбора, приблизительно 7,5% кодонов третьего выбора и приблизительно 7,5% кодонов четвертого выбора, где общая процентная доля составляет 100%. Описываемый выше способ позволяет специалисту в этой области модифицировать гены, являющиеся чужеродными для конкретного растения, таким образом, что гены оптимально экспрессируются в растениях. Способ дополнительно проиллюстрирован в заявке PCT WO 97/13402.
Таким образом, для конструирования генов, оптимизированных для растений и кодирующих бактериальный белок, последовательность ДНК конструируют кодирующей аминокислотную последовательность указанного белка с использованием избыточного генетического кода, установленного по таблице отклонений частот использования кодонов, составленной из последовательностей генов конкретного растения или растений. Получаемая последовательность ДНК имеет более высокую степень разнообразия кодонов, желаемую композицию оснований, может содержать стратегически расположенные участки распознавания рестрикционных ферментов и не содержит последовательности, которые могут препятствовать транскрипции гена или трансляции продукта мРНК. Таким образом, для трансформации хозяев, включая растения, можно использовать синтетические гены, функционально эквивалентные белкам/генам по настоящему изобретению. Дополнительные инструкции, касающиеся получения синтетических генов можно найти, например, в патенте США № 5380831.
Анализ перестроек в AAD-12 растений: При обширном анализе 876 пар оснований (п.н.) последовательности ДНК, кодирующей области нативного AAD-12 (SEQ ID NO: 1), выявляли наличие нескольких мотивов последовательности, которые, как считают, являются неблагоприятными для оптимальной экспрессии в растении, а также неоптимальную композицию кодонов. Белок, кодируемый SEQ ID NO: 1 (AAD-12), представлен как SEQ ID NO: 2. Для улучшения продукции рекомбинантного белка в однодольных, а также двудольных растениях, разрабатывали "оптимизированную для растений" последовательность ДНК AAD-12 (v1) (SEQ ID NO: 3), кодирующую белок (SEQ ID NO: 4), являющийся тем же, что и нативный SEQ ID NO: 2, за исключением добавления остатка аланина во втором положении (подчеркнуто в SEQ ID NO: 4). Дополнительный кодон для аланина (GCT; подчеркнутый в SEQ ID NO: 3) кодирует часть участка распознавания рестрикционного фермента NcoI (CCATGG), перекрывающегося с инициирующим трансляцию кодоном ATG. Таким образом, он служит двойной цели облегчения последующих действий по клонированию одновременно с улучшением контекста последовательности, окружающего инициаторный кодон ATG, для оптимизации инициации трансляции. Белки, кодируемые нативной и оптимизированной для растений (v1) кодирующими областями, являются на 99,3% идентичными, отличаясь только по аминокислоте в положении 2. В отличие от этого, нативные и оптимизированные для растений (v1) последовательности ДНК кодирующих областей являются только на 79,7% идентичными.
В таблице 4 показаны различия композиций кодонов нативной (колонки A и D) и оптимизированной для растений (колонки B и E) последовательностей, и приведено сравнение с теоретической оптимизированной для растений последовательностью (колонки C и F).
При рассмотрении таблицы 4 видно, что нативная и оптимизированная для растений кодирующие области, хотя и кодируют приблизительно идентичные белки, существенно отличаются друг от друга. Версия, оптимизированная для растений, (v1) точно имитирует композицию кодонов в теоретической оптимизированной для растений кодирующей области, кодирующей белок AAD-12.
"Amino acid" - "Аминокислота", "Native #" - "Количество в нативной последовательности", "Plant Opt v1 #" - "Количество в оптимизированной для растений версии (v1)", "Theor. Plant Opt. #" - "Количество в теоретической оптимизированной для растений версии", "Totals" - "Всего"]
Сравнение композиции кодонов кодирующих областей нативной AAD-12, оптимизированной для растений версии (v1) и теоретической оптимизированной для растений версии
Перестройка для экспрессии в E. coli: Специально сконструированные штаммы Escherichia coli и соответствующие векторные системы часто используют для получения относительно больших количеств белков для биохимических и аналитических исследований. Иногда обнаруживают, что нативный ген, кодирующий желаемый белок, плохо подходит для высокоуровневой экспрессии в E. coli, даже если организмом-источником гена может являться другой род бактерий. В таких случаях можно и желательно реконструировать кодирующую белок область гена, чтобы сделать его более подходящим для экспрессии в E. coli. Гены E. coli класса II определяют как те, которые на высоком уровне и непрерывно экспрессируются в течение экспоненциальной фазы роста клеток E. coli. (Henaut, A. and Danchin, A. (1996) in Escherichia coli and Salmonella typhimurium cellular and molecular biology, vol. 2, pp. 2047-2066. Neidhardt, F., Curtiss III, R., Ingraham, J., Lin, E., Low, B., Magasanik, B., Reznikoff, W., Riley, M., Schaechter, M. and Umbarger, H. (eds.) American Society for Microbiology, Washington, D.C.). Исследуя композиции кодонов кодирующих областей генов E. coli класса II, можно разрабатывать среднюю композицию кодонов для этих кодирующих областей генов E. coli класса II.
Считают, что кодирующая белок область, имеющая среднюю композицию кодонов, имитирующую таковую в генах класса II, будет подходить для экспрессии в течение экспоненциальной фазы роста E. coli. Используя эти руководства, новую последовательность ДНК, кодирующую белок AAD-12 (SEQ ID NO: 4), включающую дополнительный аланин во втором положении, как указано выше, конструировали в соответствии со средней композицией кодонов кодирующих областей генов класса II E. coli. Исходную последовательность, конструирование которой основывалось только на композиции кодонов, дополнительно реконструировали для включения конкретных последовательностей участков распознавания рестрикционных ферментов, подходящих для клонирования в векторы, экспрессирующиеся в E. coli. Избегали неблагоприятных черт последовательности, таких как структуры "стебель-петля" с высокой стабильностью, а также внутригенные последовательности, гомологичные 3'-концу рибосомальной РНК 16S (т.е. последовательности Шайна-Дальгарно). E. coli-оптимизированную последовательность (v2) описывают как SEQ ID NO: 5, и она кодирует белок, описываемый как SEQ ID NO: 4.
Нативная и E. coli-оптимизированная (v2) последовательности ДНК являются на 84,0% идентичными, в то время как оптимизированная для растений (v1) и E. coli-оптимизированная (v2) последовательности ДНК являются на 76,0% идентичными. В таблице 5 представлены композиции кодонов нативной кодирующей области AAD-12 (колонки A и D), кодирующей области AAD-12, оптимизированной для экспрессии в E. coli (v2; колонки B и E), и композиция кодонов теоретической кодирующей области для белка AAD-12, имеющей оптимальную композицию кодонов генов класса II E. coli (колонки C и F).
При рассмотрении таблицы 6 видно, что нативная и E. coli-оптимизированная кодирующие области, хотя и кодируют приблизительно идентичные белки, существенно отличаются друг от друга. E. coli-оптимизированная версия (v2) точно имитирует композицию кодонов в теоретической E. coli-оптимизированной кодирующей области, кодирующей белок AAD-12.
Сравнение композиции кодонов кодирующих областей нативной AAD-12, E. coli-оптимизированной версии (v2) и теоретической версии, оптимизированной для генов класса II E. coli
Конструирование последовательности ДНК с отклонением в использовании кодонов для сои, кодирующей EPSPS сои, имеющую мутации, придающие устойчивость к глифосату. В этом примере описывают конструирование новой последовательности ДНК, кодирующей мутантную 5-енолпирувилшикимат-3-фосфатсинтетазу (EPSPS) сои, но оптимизированной для экспрессии в клетках сои. Аминокислотную последовательность EPSPS сои с тремя мутациями описывают как SEQ ID NO: 5 в WO 2004/009761. Мутантные аминокислоты в описываемой таким образом последовательности находятся в положении 183 (треонин нативного белка, замененный изолейцином), положении 186 (аргинин нативного белка, замененный лизином) и положении 187 (пролин нативного белка, замененный серином). Таким образом, можно получать аминокислотную последовательность нативного белка EPSPS сои, заменяя замещенные аминокислоты SEQ ID NO:5 в WO 2004/009761 нативными аминокислотами в соответствующих положениях. Такую нативную белковую последовательность описывают как SEQ ID NO: 20 в PCT/US2005/014737 (зарегистрированной 2 мая 2005 года). Последовательность белка EPSPS сои с двумя мутациями, содержащую мутацию в положении 183 (треонин нативного белка, замененный изолейцином) и положении 187 (пролин нативного белка, замененный серином), описывают как SEQ ID NO: 21 в PCT/US2005/014737.
Таблицу использования кодонов в кодирующих белок последовательностях сои (с максимальным содержанием глицина), вычисленного с помощью 362096 кодонов (приблизительно 870 кодирующих последовательностей) получали на веб-сайте "kazusa.или.jp/codon". Эти данные преобразовывали, как показано в таблице 6. В колонках D и H таблицы 6 представлены распределения (в % использованиях всех кодонов для этой аминокислоты) синонимичных кодонов для каждой аминокислоты, как обнаруживают в кодирующих белок областях генов сои. Очевидно, что некоторые синонимичные кодоны для некоторых аминокислот (аминокислоту могут определять 1, 2, 3, 4 или 6 кодонов) относительно редко представлены в кодирующих белок областях сои (например, сравнительно использование кодонов GCG и GCT, определяющих аланин).
Таблицу отклонений использования кодонов в сое вычисляли с помощью данных из таблицы 6.
Игнорировали кодоны, обнаруживаемые в генах сои менее чем приблизительно в 10% случаев от общей частоты наличия конкретной аминокислоты. Для уравновешивания распределения оставшихся кодонов для аминокислот вычисляли средневзвешенную частоту для каждого кодона, используя формулу:
Средневзвешенный % C1=1/(%C1+%C2+%C3+и т.д.)×% C1×100
где C1 является рассматриваемым кодоном, C2, C3 и т.д. представляют оставшиеся синонимичные кодоны, и значения % для соответствующих кодонов взяты из колонок D и H таблицы 6 (игнорируя значения для редких кодонов, выделенные полужирным шрифтом).
Средневзвешенный % значения для каждого кодона представлен в колонках C и G таблицы 6. TGA условно выбирали в качестве терминатора трансляции. Затем отклонения частоты использования кодонов включали в специализированную таблицу генетического кода для использования в программе для конструирования генов OptGene™ (Ocimum Biosolutions LLC, Indianapolis, Ind.).
Частота синонимичных кодонов в кодирующих белок последовательностях сои и вычисление отклонения частоты использования кодонов для конструирования синтетического гена, оптимизированного для сои
Для получения оптимизированной для сои последовательности ДНК, кодирующей белок EPSPS с двумя мутациями, белковую последовательность SEQ ID NO: 21 из PCT/US2005/014737 подвергали обратной трансляции с помощью программы OptGene™ с использованием представленного выше генетического кода с отклонением частот использования кодонов для сои. Затем полученную таким образом исходную последовательность ДНК модифицировали, компенсируя изменения кодонов (с сохранением общих средневзвешенных частот кодонов) для снижения количества CG- и TA-дуплетов между смежными кодонами, повышения количества CT- и TG-дуплетов между смежными кодонами, удаления внутрицепочечных вторичных структур с высокой стабильностью, удаления или добавления участков распознавания рестрикционных ферментов и удаления других последовательностей, которые могут являться неблагоприятными для экспрессии или действий по клонированию сконструированного гена. Осуществляли дополнительные улучшения последовательности для устранения потенциальных растительных участков сплайсинга интронов, длинных участков остатков A/T или C/G и других мотивов, которые могут препятствовать стабильности РНК, транскрипции или трансляции кодирующей области в растительных клетках. Осуществляли другие изменения для устранения длинных внутренних открытых рамок считывания (рамок, иных, чем +1). Все эти изменения осуществляли в пределах ограничений для сохранения композиции кодонов с отклонением частот использования кодонов для сои, как описано выше, и одновременно сохраняя аминокислотную последовательность, описываемую как SEQ ID NO: 21 в PCT/US2005/014737.
Последовательность ДНК с отклонением частот использования кодонов для сои, кодирующую белок EPSPS SEQ ID NO: 21, описывают как основания 1-1575 SEQ ID NO: 22 в PCT/US2005/014737. Синтез фрагмента ДНК, содержащего SEQ ID NO: 22 из PCT/US2005/014737, осуществлял коммерческий поставщик (PicoScript, Houston Tex.).
ПРИМЕР 3
Клонирование экспрессирующих векторов и векторов для трансформации
Конструирование E. coli, экспрессирующий вектор pET: Используя рестрикционные ферменты, соответствующие участкам, добавленным вместе с дополнительными линкерами для клонирования, (Xba 1, Xho 1) AAD-12 (v2) вырезали из вектора Picoscript и лигировали в вектор pET280 с резистентностью к стрептомицину/спектиномицину. Затем с помощью лигированных продуктов трансформировали E. coli TOP10F' и помещали их на чашки с агаром с бульоном Луриа + 50 мкг/мл стрептомицина и спектиномицина (LB S/S).
Чтобы различать продукты лигирования AAD-12 (v2):pET280 и pCR2.1:pET280 приблизительно 20 выделенных колоний помещали в 6 мл LB-S/S и выращивали при 37°C в течение 4 часов со встряхиванием. Затем каждую культуру наносили на чашки LB + 50 мкг/мл канамицина, которые инкубировали при 37°C в течение ночи. Колонии, выращенные на LB-K, считали содержащими лигированный вектор pCR2.1 и отбраковывали. Из оставшихся культур выделяли плазмиды, как указано выше, и проверяли на соответствие посредством расщепления XbaI/XhoI. Конечной экспрессирующей конструкции присваивали обозначение pDAB3222.
Конструирование экспрессирующего вектора для Pseudomonas: Открытую рамку считывания AAD-12 (v2) сначала клонировали в модифицированный экспрессирующий вектор pET (Novagen), "pET280 S/S", как фрагмент XbaI-XhoI. Полученную плазмиду pDAB725 подтверждали расщеплением рестрикционными ферментами и реакциями секвенирования. Открытую рамку считывания AAD-12 (v2) из pDAB725 переносили в экспрессирующий вектор для Pseudomonas, pMYC1803, как фрагмент XbaI-XhoI. Позитивные колонии подтверждали посредством расщепления рестрикционными ферментами. С помощью завершенной конструкции pDAB739 трансформировали экспрессирующие штаммы Pseudomonas MB217 и MB324.
Завершение конструирования бинарных векторов: Оптимизированный для растений ген AAD-12 (v1) получали из Picoscript (завершали реконструирование гена (см. выше) и привлекали Picoscript для конструирования) и внутренне подтверждали последовательность (SEQ ID NO: 3), чтобы убедиться в отсутствии изменений в ожидаемой последовательности. Осуществляли реакции секвенирования с прямым (SEQ ID NO: 6) и обратным (SEQ ID NO: 7) праймерами M13 с использованием реагентов "Dye Terminator Cycle Sequencing with Quick Start Kit" Beckman Coulter, как указано выше. Анализировали данные о последовательности, и результаты свидетельствовали об отсутствии аномалий в оптимизированной для растений последовательности ДНК AAD-12 (v1). Ген AAD-12 (v1) клонировали в pDAB726 как фрагмент NcoI-SacI. Полученную конструкцию обозначали как pDAB723, содержащую: [промотор AtUbi10: Nt OSM 5'UTR: AAD-12 (v1): Nt OSM3' UTR: ORF1 polyA 3'UTR] (проверенную посредством рестрикции PvuII и NotI). Затем фрагмент NotI-NotI, содержащий описываемую кассету, клонировали по участку NotI бинарного вектора pDAB3038. Полученный бинарный вектор pDAB724, содержащий следующую кассету [промотор AtUbi10: Nt OSM 5'UTR: AAD-12 (v1): Nt OSM 3'UTR: ORF1 polyA 3'UTR: промотор CsVMV: PAT: ORF25/26 3'UTR] подвергали рестрикции (с использованием BamHI, NcoI, NotI, SacI и XmnI) для проверки правильной ориентации. Проверенную завершенную конструкцию (pDAB724) использовали для трансформации Agrobacterium.
Клонирование дополнительных конструкций для трансформации: Все другие конструкции, полученные для трансформации соответствующих видов растений, конструировали с использованием аналогичных способов, как представленные выше в настоящем описании, и других стандартных способов молекулярного клонирования (Maniatis et al., 1982).
ПРИМЕР 4
Трансформация арабидопсиса и селекция
Условия выращивания Arabidopsis thaliana: Семена арабидопсиса дикого типа суспендировали в 0,1% растворе агарозу (Sigma Chemical Co., St. Louis, Mo.). Суспендированные семена хранили при 4°C в течение 2 дней для выполнения требований покоя и обеспечения синхронного прорастания семян (стратификации).
Sunshine Mix LP5 (Sun Gro Horticulture, Bellevue, Wash.) покрывали мелкозернистым вермикулитом и осуществляли подпочвенное орошение раствором Хогланда до увлажнения. Почвенной смеси позволяли дренироваться в течение 24 часов. Стратифицированные семена высевали на вермикулит и закрывали куполами-увлажнителями (KORD Products, Bramalea, Ontario, Canada) на 7 дней.
Семена прорастали и растения выращивали в Conviron (модели CMP4030 и CMP3244, Controlled Environments Limited, Winnipeg, Manitoba, Canada) в условиях длинного дня (16 часов освещения/8 часов темноты) при интенсивности света 120-150 мкмоль/м2⋅с при постоянной температуре (22°C) и влажности (40-50%). Растения орошали сначала раствором Хогланда, а затем деионизированной водой, чтобы сохранять почву влажной, но не мокрой.
Трансформация Agrobacterium: Для получения колонии для инокуляции 4 мл препарационной культуры (жидкая LB + эритромицин) использовали чашку с LB + агар с эритромицином (Sigma Chemical Co., St. Louis, Mo.) (200 мг/л) или спектиномицином (100 мг/л), содержащую штриховую колонию DH5α. Культуры инкубировали в течение ночи при 37°C с постоянным встряхиванием. Для очистки плазмидной ДНК использовали Qiagen (Valencia, Calif.) Spin Mini Preps, осуществляя очистку по инструкциям производителя.
Электрокомпетентные клетки Agrobacterium tumefaciens (штаммов Z707s, EHA101s и LBA4404s) получали с использованием протокола из Weigel and Glazebrook (2002). Компетентные клетки Agrobacterium трансформировали с использованием способа электропорации, адаптированного из Weigel and Glazebrook (2002). 50 мкл компетентных клеток Agrobacterium размораживали на льду и добавляли к клеткам 10-25 нг желаемой плазмиды. Смесь ДНК и клеток добавляли в предварительно охлажденные кюветы для электропорации (2 мм). Для трансформации использовали Eppendorf Electroporator 2510 со следующими параметрами: напряжение: 2,4 кВ, длительность импульса: 5 мс.
После электропорации в кювету добавляли 1 мл бульона YEP (на литр: 10 г дрожжевого экстракта, 10 г бакто-пептона, 5 г NaCl), и суспензию клеток-YEP переносили в культуральную пробирку емкостью 15 мл. Клетки инкубировали при 28°C на водяной бане с постоянным встряхиванием в течение 4 часов. После инкубации культуру помещали в чашки с YEP + агар с эритромицином (200 мг/л) или спектиномицином (100 мг/л) и стрептомицином (Sigma Chemical Co., St. Louis, Mo.) (250 мг/л). Чашки инкубировали в течение 2-4 дней при 28°C.
Выбирали колонии, наносили штрихами на свежие чашки с YEP + агар с эритромицином (200 мг/л) или спектиномицином (100 мг/л) и стрептомицином (250 мг/л) и инкубировали при 28°C в течение 1-3 дней. Для проверки наличия встроенного гена выбирали колонии для ПЦР-анализа с использованием праймеров, специфичных для вектора. Для выделения плазмидной ДНК из выбранных колоний Agrobacterium использовали Qiagen Spin Mini Preps, осуществляя выделение по инструкциям производителя со следующим исключением: для очистки ДНК использовали аликвоты по 4 мл из 15 мл ночной препарационной культуры (жидкий YEP + эритромицин (200 мг/л) или спектиномицин (100 мг/л)) и стрептомицин (250 мг/л)). Альтернативой использованию Qiagen Spin Mini Prep DNA являлся лизис трансформированных клеток Agrobacterium, суспендированных в 10 мкл воды, при 100°C в течение 5 минут. Плазмидную ДНК из бинарного вектора, используемого при трансформации Agrobacterium, включали в качестве контроля. Реакцию ПЦР проводили с использованием Taq ДНК-полимеразы от Takara Minis Bio Inc. (Madison, Wis.) по инструкциям производителя в 0,5-кратных концентрациях. Реакции ПЦР осуществляли в термоциклере MJ Research Peltier, запрограммированном на следующие условия; 1) 94°C в течение 3 минут, 2) 94°C в течение 45 секунд, 3) 55°C в течение 30 секунд, 4) 72°C в течение 1 минуты, в течение 29 циклов, затем 1 цикл при 72°C в течение 10 минут. После завершения циклов реакционную смесь хранили при 4°C. Продукты амплификации анализировали с помощью электрофореза в 1%-ном агарозном геле и визуализировали с помощью окрашивания бромистым этидием. Выбирали колонии, продукт ПЦР которых являлся идентичным контрольной плазмиде.
Трансформация арабидопсиса: Арабидопсис трансформировали с использованием способа погружения цветочных почек. Выбранную колонию использовали для инокуляции одной или нескольких 15-30 мл прекультур в бульоне YEP, содержащем эритромицин (200 мг/л) или спектиномицин (100 мг/л) и стрептомицин (250 мг/л). Культуры инкубировали в течение ночи при 28°C с постоянным встряхиванием при 220 об./мин. Каждую прекультуру использовали для инокуляции двух 500 мл культур в бульоне YEP, содержащем эритромицин (200 мг/л) или спектиномицин (100 мг/л) и стрептомицин (250 мг/л), и инкубировали культуры в течение ночи при 28°C с постоянным встряхиванием. Затем клетки осаждали при приблизительно 8700×g в течение 10 минут при комнатной температуре и выбрасывали полученный супернатант. Клеточный осадок осторожно ресуспендировали в 500 мл инфильтрационных сред, содержащих: ½ × солей Мурашиге и Скуга/витаминов B5 по Гамборгу, 10% (масс./об.) сахарозы, 0,044 мкМ бензиламинопурина (10 мкл/литр стокового раствора 1 мг/мл в DMSO) и 300 мкл/литр Silwet L-77. Растения возрастом приблизительно 1 месяц погружали в среды на 15 секунд, будучи уверенными в погружении самых новых соцветий. Затем растения клали набок и закрывали (прозрачным или непрозрачным материалом) на 24 часа, затем промывали водой и помещали вертикально. Растения выращивали при 22°C с фотопериодом 16 часов освещения/8 часов темноты. Приблизительно через 4 недели после погружения собирали семена.
Селекция трансформированных растений: Свежесобранным семенам T1 [ген AAD-12 (v1)] позволяли высушиваться в течение 7 дней при комнатной температуре. Семена T1 высевали в лотки для проращивания размером 26,5×51 см (T.O. Plastics Inc., Clearwater, Minn.), в каждый из которых вносили аликвоты по 200 мг стратифицированных семян T1 (приблизительно 10000 семян), предварительно суспендированных в 40 мл 0,1% раствора агарозы и хранившихся при 4°C в течение 2 дней для выполнения требований покоя и обеспечения синхронного прорастания семян.
Sunshine Mix LP5 (Sun Gro Horticulture Inc., Bellevue, Wash.) покрывали мелкозернистым вермикулитом и осуществляли подпочвенное орошение раствором Хогланда до увлажнения, затем позволяя ему дренироваться под действием силы тяжести. Каждую аликвоту по 40 мл стратифицированных семян равномерно высевали на вермикулит с помощью пипетки и закрывали куполами-увлажнителями (KORD Products, Bramalea, Ontario, Canada) на 4-5 дней. Купола снимали за 1 день до начальной селекции трансформантов с использованием послевсходового опрыскивания глуфосинатом (селекция на котрансформированный ген PAT).
Через семь дней после посадки (DAP) и вновь через 11 DAP растения T1 (фазы семядоли и 2-4-листьев, соответственно) опрыскивали 0,2% раствора гербицида Liberty (200 г активного ингредиента глуфосината/л, Bayer Crop Sciences, Kansas City, Mo.) при объеме раствора для опрыскивания 10 мл/лоток (703 л/га) с использованием пневматического распылителя DeVilbiss для доставки эффективного количества 280 г активного ингредиента глуфосината/га на нанесение. Выжившие растения (активно растущие растения) определяли через 4-7 дней после последнего распыления и переносили по-отдельности в 3-дюймовые горшки с подготовленными горшечными средами (Metro Mix 360). Перенесенные растения покрывали куполами-увлажнителями на 3-4 дня и помещали в вегетационную камеру с температурой 22°C до помещения в теплицу или непосредственно в теплице. Затем купола снимали и растения выращивали в теплице (22+5°C, 50+30% RH, 14 часов освещения:10 часов темноты, минимум 500 мЭ/м2⋅с1 естественного + дополнительного освещения) по меньшей мере 1 день до тестирования на способность AAD-12 (v1) (гена, оптимизированного для растений) обеспечивать резистентность к феноксиауксиновым гербицидам.
Затем растениям T1 случайным образом распределяли различные количества 2,4-D. В случае арабидопсиса 50 г кэ/га 2,4-D является эффективной дозой для различения восприимчивых растений и растений со значимыми уровнями резистентности. Также для определения относительных уровней резистентности наносили повышающиеся количества (50, 200, 800 или 3200 г кэ/га).
Все нанесения ауксинового гербицида осуществляли с использованием распылителя DeVilbiss, как описано выше, для нанесения объема раствора для опрыскивания 703 л/га (0,4 мл раствор/3-дюймовый горшок) или наносили с помощью автоматического устройства для опрыскивания при объеме раствора для опрыскивания 187 л/га. Используемый 2,4-D являлся 2,4-D технической категории (Sigma, St. Louis, Mo.), растворенным в DMSO и разведенным в воде (конечная концентрация DMSO <1%), или коммерческим составом диметиламиновой соли (456 г кэ/л, NuFarm, St Joseph, Mo.). Используемый дихлорпроп являлся дихлорпропом коммерческой категории, составленным в виде калиевой соли R-дихлорпропа (600 г активного ингредиента/л, AH Marks). При повышении количеств гербицидов выше 800 г кэ/га pH распыляемого раствора становится крайне кислым, приводя к ожогу листьев молодых уязвимых растений арабидопсиса и затруднению оценки первичных эффектов гербицидов. Общепринятой практикой стало нанесение этих больших количеств гербицидов в 200 мМ буфере HEPES, pH 7,5.
Некоторые отдельные растения T1 подвергали воздействию альтернативных коммерческих гербицидов вместо феноксиауксина. Одним из интересующих пунктов являлось определение того, могут ли пиридилоксиацетатауксиновын гербициды, триклопир и флуроксипир, эффективно подвергаться деградации в полевых условиях. Гербициды наносили на растения T1 с использованием автоматического устройства для опрыскивания при объеме раствора для опрыскивания 187 л/га. Растения T1, проявляющие устойчивость к 2,4-D DMA, дополнительно оценивали в поколении T2.
Результаты селекции трансформированных растений: Первые трансформации арабидопсиса осуществляли с использованием AAD-12 (v1) (гена, оптимизированного для растений). Сначала трансформантов T1 выбирали по данным о нетрансформированных семенах с использованием схемы селекции с использованием глуфосината. Более 300000 семян T1 подвергали скринингу и идентифицировали 316 резистентных к глуфосинату растений (ген PAT), приравниваемых к частоте трансформации/селекции 0,10%, попадающей в нормальный диапазон частоты селекции конструкций, где для селекции использовали PAT + Liberty. Затем растения T1, подвергнутые селекции, как указано выше, переносили в отдельные горшки и опрыскивали различными количествами коммерческих арилоксиалканоатных гербицидов.
Ответ растений Arabidopsis T1, трансформированных с использованием AAD-12 v1 (оптимизированного для растений), на диапазон количеств 2,4-D, наносимого в послевсходовый переиод, по сравнению с популяцией резистентных растений (T4), гомозиготных по AAD-1 v3, растениями, трансформированными с использованием Pat-Cry1F, ауксин-восприимчивым контролем
Средние
Средние
Средние
В таблице 7 сравнивают ответ AAD-12 (v1) и контрольных генов по приданию резистентности к 2,4-D трансформантам T1 Arabidopsis. Ответ представлен в терминах % видимых повреждений через 2 WAT. Данные представлены в виде гистограммы отдельных растений, проявляющих небольшие повреждения или их отсутствие (<20%), умеренные повреждения (20-40%) или тяжелые повреждения (>40%). Так как каждое растение T1 является независимым трансформантом, можно ожидать значительной вариации индивидуальных ответов растений T1 в пределах указанного количества. Для каждой обработки представлены арифметическое среднее и стандартное отклонение. В последней колонке также указан диапазон индивидуального ответа для каждого количества и трансформации. PAT /Cry1F-трансформированные Arabidopsis служили в качестве ауксин-восприимчивого трансформированного контроля. Ген AAD-12 (v1) придавал резистентность к гербицидам отдельным растениям T1 Arabidopsis. В пределах указанной обработки уровень ответа растений значительно варьировался, и это можно приписать тому, что каждое растение представляет собой независимый трансформант.
Ответ растений T1 Arabidopsis на диапазон количеств R-дихлорпропа, наносимого в послевсходовый период
Средние
Средние
повреждений
Важно отметить, что при каждом тестируемом количестве 2,4-D отдельные растения оставались неповрежденными, в то время как некоторые растения имели тяжелые повреждения. Среднее значение повреждений во всей популяции на норму внесения представлено в таблице 7 исключительно для демонстрации значимого различия между растениями, трансформированными с использованием AAD-12 (v1), и растений дикого типа или PAT/Cry1F-трансформированных контролей. Уровни повреждения имели тенденцию к повышению, и доля неповрежденных растений являлась более низкой при повышении норм внесения до 3200 г кэ/га (или ~6-кратной нормой внесения в полевых условиях). Также при этих высоких нормах внесения раствор для опрыскивания становился сильнокислым, если его не забуферивали. Arabidopsis, выращенный, главным образом, в вегетационной камере, имеет очень тонкую кутикулу, и тяжелые ожоговые эффекты могут затруднять тестирование при этих повышенных нормах внесения. Тем не менее, многие отдельные растения выживали при 3200 г кэ/га 2,4-D с небольшими повреждениями или без них.
В таблице 8 представлен аналогично проведенный анализ дозозависимого ответа растений T1 Arabidopsis на феноксипропионовую кислоту, дихлорпроп. Данные свидетельствуют о том, что гербицидно активный (R-) изомер дихлорпропа не является подходящим субстратом для AAD-12 (v1). То, что AAD-1 достаточно хорошо будет метаболизировать R-дихлорпроп для придания коммерчески приемлемой устойчивости, является одной из отличительных характеристик, разделяющих два гена (таблица 8). AAD-1 и AAD-12 считают R- и S-специфичными α-кетоглутаратдиоксигеназами, соответственно.
AAD-12 (v1) в качестве селективного маркера: Исходно возможность использования AAD-12 (v1) в качестве селективного маркера при использовании 2,4-D в качестве селективного средства анализировали с помощью арабидопсиса, трансформированного, как описано выше. Приблизительно 50 семян Arabidopsis поколения T4 (гомозиготных по AAD-12 (v1)) добавляли к приблизительно 5000 семян дикого типа (восприимчивым). Сравнивали несколько обработок, для каждого из лотков с растениями использовали один или два времени нанесения 2,4-D по одной из следующих схем обработок: через 7 DAP, через 11 DAP или через 7, затем 11 DAP. Т.к. все отдельные растения также содержали ген PAT в том же векторе для трансформации, AAD-12, подвергаемый селекции с помощью 2,4-D, можно напрямую сравнивать с PAT, подвергаемым селекции с помощью глуфосината.
Обработки проводили с использованием распылителя DeVilbiss, как описано выше. Растения идентифицировали как резистентные или восприимчивые через 17 DAP. Оптимальная обработка представляла собой 75 г кэ/га 2,4-D, наносимого через 7 и 11 дней после посадки (DAP), являлась в равной степени эффективной по частоте селекции и приводила к меньшему повреждению гербицидами отдельных трансформированных растений, чем при схеме селекции с использованием Liberty. Эти результаты свидетельствуют о том, что AAD-12 (v1) можно эффективно использовать в качестве альтернативного селективного маркера для популяции трансформированного Arabidopsis.
Наследование: Множество объектов T1 самоопылялись с образованием семян T2. Эти семена являлись потомством, тестируемым посредством нанесения 2,4-D (200 г кэ/га) на 100 случайных сибсов T2. Каждое отдельное растение T2 переносили в 7,5-см квадратные горшки перед опрыскиванием (автоматическое устройство для опрыскивания с объемом раствора для опрыскивания 187 л/га). Семьдесят пять процентов семейств T1 (растений T2) расщеплялись в соответствии с предполагаемой моделью 3 резистентных: 1 восприимчивое растение для доминантно наследуемого единого локуса с менделевским наследованием, как определяли с помощью анализа с использованием критерия хи-квадрата (P>0,05).
Собирали семена от 12 до 20 отдельных растений T2 (семена T3). Двадцать пять сибсов T3 из каждого из восьми случайно выбранных семейств T2 являлись потомством, тестируемым, как описано выше. В каждой линии идентифицировали приблизительно треть семейств T2, как предполагают, гомозигот (нерасщепляющиеся популяции). Эти данные свидетельствуют о том, что AAD-12 (v1) стабильно интегрируется и наследуется по менделевскому типу по меньшей мере в трех поколениях.
Сравнение ответа растений T2 с AAD-12 (v1) и трансформированных контрольных растений Arabidopsis на некорневое нанесение различных ауксиновых гербицидов
Дополнительный анализ резистентности к гербицидам у AAD-12 Arabidopsis с помощью некорневого нанесения: Способность AAD-12 (v1) обеспечивать резистентность к другим арилоксиалканоатауксиновым гербицидам у трансгенного арабидопсиса определяли с помощью некорневого нанесения различных субстратов. Семена Arabidopsis поколения T2 стратифицировали и высевали в лотки для селекции так же, как и у Arabidopsis. Аналогично сажали трансформированную контрольную линию, содержащую PAT и ген резистентности к насекомым Cry1F. Рассаду переносили в отдельные 3-дюймовые горшки в теплицу. Все растения опрыскивали с использованием автоматического устройства для опрыскивания, установленного на 187 л/га. Растения опрыскивали с использованием диапазона пиридилоксиацетатных гербицидов: 280-2240 г кэ/га триклопира (Garlon 3A, Dow AgroSciences) и 280-2240 г кэ/га флуроксипира (Starane, Dow AgroSciences); и метаболит 2,4-D, полученный в результате активности AAD-12, 2,4-дихлорфенол (DCP, Sigma) (в молярном эквиваленте 280-2240 г кэ/га 2,4-D, использовали DCP технической категории). Все растворы для нанесения составляли в воде. Каждую обработку повторяли 3-4 раза. Растения оценивали через 3 и 14 дней после обработки.
Не наблюдали эффекта метаболита 2,4-D, 2,4-дихлорфенола (DCP), в отношении трансгенного контрольного Arabidopsis (Pat/Cry1F) без AAD-12. AAD-12-трансформированные растения также являлись определенно защищенными от повреждения гербицидами триклопиром и флуроксипиром, наблюдаемого у трансформированных нерезистентных контролей (см. таблицу 9). Эти результаты подтверждают, что AAD-12 (v1) у Arabidopsis обеспечивает резистентность к тестируемым пиридилоксиуксусным ауксинам. Это первое сообщение о ферменте со значимой активностью в отношении гербицидов на основе пиридилоксиуксусной кислоты. Не сообщали о другом деградирующем 2,4-D ферменте с аналогичной активностью.
Молекулярный анализ Arabidopsis с AAD-12 (v1): Анализ Invader (способы по процедурам набора Third Wave Agbio) для анализа копийности гена PAT осуществляли с использованием тотальной ДНК, полученной с помощью набора Qiagen DNeasy, на многочисленных линиях, гомозиготных по AAD-12 (v1), для определения стабильной интеграции единицы трансформации растения, содержащей PAT и AAD-12 (v1). При анализе оценивали прямое физическое сцепление этих генов, т.к. они содержатся в одной плазмиде.
Результаты показали, что все анализируемые резистентные к 2,4-D растения содержали PAT (и, таким образом, как подразумевают, AAD-12 (v1)). Анализ копийности показал, что общее количество вставок находится в диапазоне от 1 до 5 копий. Это также коррелирует с данными об экспрессии белка AAD-12 (v1), свидетельствующими о том, что наличие фермента приводит к значимо высоким уровням резистентности ко всем коммерчески доступным феноксиуксусным и пиридилоксиуксусным кислотам.
Arabidopsis, трансформированный с использованием молекулярного стэкинга AAD-12 (v1) и гена резистентности к глифосату: Получали семена Arabidopsis T1, как описано выше, содержащие плазмиду pDAB3759 (AAD-12 (v1) + EPSPS), кодирующую предполагаемый признак резистентности к глифосату. Трансформанты T1 подвергали селекции с использованием AAD-12 (v1) в качестве селективного маркера, как описано выше. Растения T1 (объекты, трансформированные по-отдельности) получали в первом опыте по селекции и переносили в трехдюймовые горшки в теплицу, как описано выше. Также тестировали три различные контрольные линии Arabidopsis: Columbia-0 дикого типа, линии, гомозиготные по AAD-12 (v1)+PAT T4 (трансформированные с использованием pDAB724), и линию, гомозиготную по PAT+Cry1F (трансформированный контроль). Растения, трансформированные с использованием pDAB3759 и pDAB724, подвергали предварительной селекции по устойчивости к 2,4-D на стадии всходов. Через четыре дня после пересаживания растения равномерно разделяли для некорневой обработки с помощью автоматического устройства для опрыскивания, как описано выше, с 0, 26,25, 105, 420 или 1680 г кэ/га глифосата (Glyphomax Plus, Dow AgroSciences) в воде. Все обработки повторяли от 5 до 20 раз. Растения оценивали через 7 и 14 дней после обработки.
"AAD-12 v1 gene" - "ген AAD-12 v1", "Averages" - "Средние", "% Injury" - "% повреждения", "Ave" - "Среднее", "Std Dev" - "Стандартное отклонение", "Untreated control" - "Необработанный контроль, "g ae/ha" - "г кэ/га глифосата", "Wild type" - "Дикий тип"]
Ответ Arabidopsis на диапазон количеств глифосата, наносимого в послевсходовый период (через 14 DAT)
(Средние)
повреждения
%
Средние
%
Средние
%
Растения, определенные при исходной оценке резистентности как устойчивые к 2,4-D, впоследствии определяли как устойчивые к глифосату при сравнении с ответом трех контрольных линий. Эти результаты свидетельствуют о том, что растениям можно придавать резистентность к двум гербицидам с различными механизмами действия, включая устойчивость к 2,4-D и глифосату, что позволяет использовать оба гербицида в послевсходовый период. Кроме того, AAD-12+2,4-D эффективно использовали в качестве селективного маркера для точной селекции по резистентности.
Arabidopsis с AAD-12, подвергнутый генетическому стэкингу с AAD-1 для достижения более широкого спектра устойчивости к гербицидам: Растения с AAD-12 (v1) (pDAB724) и AAD-1 (v3) (pDAB721) подвергали реципрокному скрещиванию и собирали семена F1. Восемь семян F1 сажали и позволяли вырасти для получения семян. Из восьми растений F1 получали образцы тканей и подвергали их анализу вестерн-блоттинга для подтверждения наличия обоих генов. Делали вывод о том, что все 8 тестируемых растений экспрессировали белки AAD-1 и AAD-12. Семена собирали и позволяли им высушиваться в течение недели перед посадкой.
Высевали одну сотню семян F2 и наносили 280 г активного ингредиента/га глуфосината. Девяносто шесть растений F2 выживали после селекции с использованием глуфосината, что соответствует ожидаемому соотношению расщепления для двух независимо подобранных локусов резистентности к глуфосинату (15 R:1 S). Затем резистентные к глуфосинату растения обрабатывали 560 г кэ/га R-дихлорпропа +560 г кэ/га триклопира, наносимых на растения то той же схеме опрыскивания, которую использовали для других тестов. Растения классифицировали в 3 и 14 DAT. Шестьдесят три из 96 растений, выживших после селекции с использованием глуфосината, также выживали после нанесения гербицидов. Эти данные соответствуют ожидаемому профилю расщепления (9R: 6S) двух независимо подобранных доминантных признаков, где каждый придает резистентность только к одному из ауксиновых гербицидов (R-дихлорпропу или триклопиру). Результаты свидетельствуют о том, что AAD-12 (pDAB724) можно успешно подвергать стэкингу с AAD-1 (pDAB721), таким образом, увеличивая спектр гербицидов, которые можно наносить на интересующую сельскохозяйственную культуру [(2,4-D + R-дихлорпроп) и (2,4-D + флуроксипир + триклопир), соответственно]. Это можно применять для придания крайне восприимчивым видам устойчивости к 2,4-D посредством общепринятого стэкинга двух отдельных генов резистентности к 2,4-D. Дополнительно, если ген используют в качестве селективного маркера для третьего и четвертого интересующих генов посредством независимых трансформаций, то каждую пару генов можно объединять посредством общепринятых способов скрещивания, а затем подвергать селекции в поколении F1 посредством парного опрыскивания гербицидами, являющимися исключающими между ферментами AAD-1 и AAD-12 (как показано с использованием R-дихлорпропа и триклопира для AAD-1 и AAD-12, соответственно).
Другие стэки с AAD также входят в объем настоящего изобретения. Белок TfdA, приведенный где-либо в настоящем описании (Streber et al.), например, можно использовать вместе с генами AAD-12 по настоящему изобретению для придания трансгенным растениям по настоящему изобретению спектров резистентности к гербицидам.
ПРИМЕР 5
Вискер-опосредованная трансформация кукурузы с использованием селекции имазетапиром
Клонирование AAD-12 (v1): Ген AAD-12 (v1) вырезали из промежуточного вектора pDAB3283 в виде фрагмента NcoI/SacI. Его лигировали непосредственно в аналогично рестрицированный вектор pDAB3403, содержащий промотор ZmUbi1 для однодольных растений. Два фрагмента лигировали с использованием ДНК-лигазы T4 и с их использованием трансформировали клетки DH5α. Получали минипроросты полученных колоний с использованием набора Qiagen QIA Spin mini prep, и колонии подвергали расщеплению ферментами для проверки ориентации. Эта первая промежуточная конструкция (pDAB4100) содержит кассету ZmUbi1:AAD-12 (v1). Эту конструкцию расщепляли NotI и PvuI для выделения генетической кассеты и расщепления нежелательного остова. Ее лигировали в рестрицированный NotI pDAB2212, содержащий AHAS качестве селективного маркера, запускаемый промотором актина риса OsAct1. Конечную конструкцию обозначали как pDAB4101 или pDAS1863, и она содержала ZmUbi1/AAD-12 (v1)/ZmPer5::OsAct1/AHAS/LZmLip.
Инициация суспензионной культуры каллюса: Для получения незрелых эмбрионов для инициации культуры каллюса осуществляли скрещивания F1 между выращенными в теплице Hi-II родителями A и B (Armstrong et al. 1991). При достижении эмбрионами размера 1,0-1,2 мм (приблизительно 9-10 дней после опыления) собирали початки и стерилизовали их поверхность посредством чистки мылом Liqui-Nox®, погружали в 70% этанол на 2-3 минуты, затем погружали в 20% коммерческий отбеливатель (0,1% гипохлорит натрия) на 30 минут.
Початки промывали в стерильной дистиллированной воде, асептически отделяли незрелые зиготные эмбрионы и культивировали их в среде 15Ag10 (среда N6 (Chu et al., 1975), 1,0 мг/л 2,4-D, 20 г/л сахарозы, 100 мг/л гидролизата казеина (ферментативно расщепленного), 25 мМ L-пролин, 10 мг/л AgNO3, 2,5 г/л Gelrite, pH 5,8) в течение 2-3 недель с удалением щитков из среды. Ткань, демонстрирующую правильную морфологию (Welter et al., 1995), избирательно переносили через двухнедельные интервалы в свежую среду 15Ag10 приблизительно на 6 недель, затем через двухнедельные интервалы ее переносили в среду 4 (среда N6, 1,0 мг/л 2,4-D, 20 г/л сахарозы, 100 мг/л гидролизата казеина (ферментативно расщепленного), 6 мМ L-пролин, 2,5 г/л Gelrite, pH 5,8) приблизительно на 2 месяца.
Для инициации эмбриогенной суспензионной культуры приблизительно 3 мл объема клеток (PCV) ткани каллюса, полученной из одного эмбриона, добавляли в приблизительно 30 мл жидкой среды H9CP+ (базовая смесь солей MS (Murashige and Skoog, 1962), модифицированные витамины MS, содержащие в 10 раз меньше никотиновой кислоты и в 5 раз больше хлорида тиамина, 2,0 мг/л 2,4-D, 2,0 мг/л α-нафтилуксусной кислоты (NAA), 30 г/л сахарозы, 200 мг/л гидролизат казеина (кислотная вытяжка), 100 мг/л миоинозитола, 6 мМ L-пролин, 5% об./об. кокосового молока (добавленного непосредственно перед субкультивированием), pH 6,0). Суспензионные культуры поддерживали в условиях темноты в конических колбах Эрленмейера емкостью 125 мл в термошейкере, установленном на 125 об./мин. при 28°C. Клеточные линии, как правило, становятся стабильными в течение от 2 до 3 месяцев после инициации. В течение периода стабилизации суспензии субкультивировали каждые 3,5 дня, добавляя 3 мл PCV клеток и 7 мл кондиционированной среды в 20 мл свежей жидкой среды H9CP+ с использованием пипетки с широким отверстием. Когда начиналось удвоение роста ткани, суспензии укрупняли и поддерживали во флаконах емкостью 500 мл, при этом 12 мл PCV клеток и 28 мл кондиционированной среды переносили в 80 мл среды H9CP+. Когда суспензии полностью стабилизировались, их подвергали криоконсервации для дальнейшего использования.
Криоконсервация и размораживание суспензий: Через два дня после субкультивирования 4 мл PCV клеток суспензии и 4 мл кондиционированной среды добавляли в 8 мл криопротектора (растворенного в среде H9CP+ без кокосового молока, с 1 M глицерином, 1 M DMSO, 2 M сахарозой, стерилизованной фильтрацией) и позволяли встряхиваться при 125 об./мин. и 4°C в течение 1 часа во флаконе емкостью 125 мл. Через 1 час 4,5 мл добавляли в охлажденный сосуд для криоконсервации Corning емкостью 5,0 мл. После наполнения отдельные сосуды хранили в течение 15 минут при 4°C в программируемом криозамораживателе, затем позволяли им замораживаться со скоростью -0,5°C/минуту до достижения конечной температуры -40°C. После достижения конечной температуры сосуды переносили в коробки в штативах внутри криохранилища Cryoplus 4 (Form a Scientific), наполненного парами жидкого азота.
Для размораживания сосуды извлекали из криохранилища и помещали в закрытый контейнер с сухим льдом, затем погружали в водяную баню, поддерживаемую при 40-45°C, до снижения "кипения". При размораживании содержимое выливали на стопку из ~8 стерильных фрагментов фильтровальной бумаги Whatman толщиной 70 мм (№ 4) в закрытые чашки Петри 100×25 мм. В течение нескольких минут жидкости позволяли впитываться в фильтры, затем верхний фильтр, содержащий клетки, переносили на среду GN6 (среда N6, 2,0 мг/л 2,4-D, 30 г/л сахарозы, 2,5 г/л Gelrite, pH 5,8) на 1 неделю. Через 1 неделю только ткань с представляющей интерес морфологией переносили с фильтровальной бумаги непосредственно на свежую среду GN6. Эту ткань субкультивировали каждые 7-14 дней до получения от 1 до 3 граммов для инициации суспензионной культуры в приблизительно 30 мл среды H9CP+ в конических колбах Эрленмейера емкостью 125 мл. Три миллилитра PCV субкультивировали в свежей среде H9CP+ каждые 3,5 дня до получения в общей сложности 12 мл PCV, после чего субкультивирование осуществляли, как описано выше.
Стабильная трансформация: Приблизительно за 24 часа до трансформации 12 мл PCV ранее подвергнутой криоконсервации суспензии эмбриогенных клеток кукурузы и 28 мл кондиционированной среды субкультивировали в 80 мл жидкой среды GN6 (среда GN6 без Gelrite) в конической колбе Эрленмейера емкостью 500 мл и помещали на шейкер при 125 об./мин. и 28°C. Это повторяли 2 раза с использованием одной и той же клеточной линии таким образом, что всего 36 мл PCV распределяли по 3 флаконам. Через 24 часа жидкие среды GN6 удаляли и заменяли их 72 мл осмотической среды GN6 S/M (среда N6, 2,0 мг/л 2,4-D, 30 г/л сахарозы, 45,5 г/л сорбита, 45,5 г/л маннита, 100 мг/л миоинозитола, pH 6,0) на флакон для осуществления плазмолиза клеток. Флаконы помещали на шейкер при 125 об./мин. в темноту на 30-35 минут при 28°C, и в течение этого времени получали суспензию 50 мг/мл вискеров из карбида кремния, добавляя соответствующий объем 8,1 мл жидкой среды GN6 S/M в ~405 мг предварительно автоклавированных стерильных вискеров из карбида кремния (Advanced Composite Materials, Inc.).
После инкубации в GN6 S/M содержимое каждого флакона объединяли в колбе для центрифугирования емкостью 250 мл. Когда все клетки осаждались на дно, отбирали всю жидкую среду GN6 S/M, кроме ~44 мл, собирали в стерильный флакон емкостью 1 л для дальнейшего использования. Предварительно увлажненную суспензию вискеров центрифугировали на вортексе в течение 60 секунд при максимальной скорости, а затем 8,1 мл добавляли в бутыль, в которую в качестве последнего этапа добавляли 170 мкг ДНК. Бутыль незамедлительно помещали в модифицированный коммерческий смеситель для краски Red Devil 5400 и перемешивали в течение 10 секунд. После перемешивания смесь клеток, сред, вискеров и ДНК добавляли к содержимому флакона емкостью 1 л вместе с 125 мл свежей жидкой среды GN6 для восстановления раствора, устанавливающего осмотический градиент. Клеткам позволяли восстанавливаться на шейкере при 125 об./мин. в течение 2 часов при 28°C перед фильтрацией через фильтровальную бумагу Whatman №4 (5,5 см) с использованием стеклянного коллектора для клеток, соединенного с вакуумным трубопроводом.
Приблизительно 2 мл диспергированной суспензии пипетировали на поверхности фильтра одновременно с воздействием вакуума. Фильтры помещали на чашки размером 60×20 мм со среды GN6. Чашки культивировали в течение 1 недели при 28°C в закрытой коробке.
Через 1 неделю фильтровальную бумагу переносили на чашки размером 60×20 мм со средой GN6 (3P) (среда N6, 2,0 мг/л 2,4-D, 30 г/л сахарозы, 100 мг/л миоинозитола, 3 мкМ имазетапир от Pursuit® DG, 2,5 г/л Gelrite, pH 5,8). Чашки помещали в коробки и культивировали в течение дополнительной недели.
Через две недели после трансформации ткань заливали, соскабливая все клетки на чашке в 3,0 мл расплавленной среды GN6 с агарозой (среда N6, 2,0 мг/л 2,4-D, 30 г/л сахарозы, 100 мг/л миоинозитола, 7 г/л легкоплавкой агарозы, pH 5,8, автоклавированная в течение всего 10 минут при 121°C), содержащей 3 мкМ имазетапира от Pursuit® DG. Ткань разрушали и 3 мл агарозы и ткани равномерно выливали на поверхность планшета размером 100×15 мм с GN6 (3P). Это повторяли для всех остальных чашек. После заливки чашки по-отдельности запечатывали с использованием Nescofilm® или Parafilm M®, а затем культивировали до появления предполагаемых изолятов.
Способ выделения изолятов и регенерации: Предположительно трансформированные объекты выделяли из содержащих Pursuit® запечатанных чашек приблизительно через 9 недель после трансформации посредством переноса на свежую селекционную среду той же концентрации в чашки 60×20 мм. Если приблизительно через 2-3 недели устойчивый рост являлся очевидным, объект считали резистентным и отбирали его для молекулярного анализа.
"Event" - "Объект", "Spray treatment" - "Опрыскивание", "% Injury" - "% повреждения", "AAD-12 ELISA" - "ELISA на AAD-12", "AAD-12 PCR" - "ПЦР на AAD-12", "cloning region" - "область клонирования", "AHAS Copy #" - "Копийность AHAS", "g ae/ha" - "г кэ/га имазетапира", "Band smaller than expected" - "Полоса меньше ожидаемой"]
Характеристика растений кукурузы T0, трансформированных с использованием AAD-12
(ppm TSP)
Инициировали регенерацию, перенося ткань каллюса в индукционную среду с цитокининами 28 (3P), содержащую 3 мкМ имазетапир от Pursuit.RTM. DG, соли и витамины MS, 30,0 г/л сахарозы, 5 мг/л BAP, 0,25 мг/л 2,4-D, 2,5 г/л Gelrite; pH 5,7. Клеткам позволяли расти при низкой освещенности (13 μΕm-2 s-1) в течение одной недели, затем при большей освещенности (40 μΕm-2 s-1) в течение еще одной недели перед переносом в регенерационную среду 36 (3P), идентичную 28 (3P), за исключением отсутствия регуляторов роста растений. Удаляли небольшие (3-5 см) проростки и помещали их в культуральные пробирки размером 150×25 мм, содержащие неселекционную среду SHGA (базовые соли и витамины Шенка и Хильдебрандта, 1972; 1 г/л миоинозитола, 10 г/л сахарозы, 2,0 г/л Gelrite, pH 5,8). При развитии у проростков достаточной корневой системы и системы побегов, их пересаживали в почву в теплице.
В 4 экспериментах полные проростки, состоящие из побега и корня, образовывались in vitro на залитых селекционных чашках в условиях темноты без прохождения общепринятой фазы каллюса. Ткани листьев девяти из этих "ранних регенераторов" подвергали ПЦР кодирующей области и ПЦР транскрипционной единицы растений (PTU) для определения гена AAD-12 и генетической кассеты, соответственно. Все содержали интактную кодирующую область AAD-12, в то время как 3 не имели полноразмерную PTU (таблица 11). Эти "ранние регенераторы" идентифицировали как объекты 4101, чтобы отличать их от полученных общепринятым способом объектов, идентифицированных как объекты "1283". Растения от 19 дополнительных объектов, полученных стандартной селекцией и регенерацией, отправляли в теплицу, выращивали до созревания и перекрестно опыляли с патентованной инбредной линией для получения семян T1. Некоторые из объектов, по-видимому, являются клонами друг друга в связи со схожим характером образования полос при Саузерн-блоттинге, таким образом, получали только 14 уникальных объектов. Растения T0 от объектов являлись устойчивыми к 70 г/га имазетапира. При анализе Invader (ген AHAS) выявляли копийность вставки в диапазоне от 1 до >10 копий. Тринадцать объектов содержали полную кодирующую область AAD-12; однако, при последующем анализе выявляли, что в случае девяти объектов полная транскрипционная единица растений не встроилась. Ни один из аномальных объектов 1863 не пропускали дальше стадии T1 и для дальнейшего определения характеристики использовали объекты 4101.
Молекулярный анализ - Кукуруза, материалы и способы: Сбор ткани, выделение ДНК и количественный анализ. Свежую ткань помещали в пробирки и лиофилизировали при 4°C в течение 2 дней. После полного высушивания ткани в пробирку помещали гранулу вольфрама (Valenite) и образцы подвергали сухому измельчению в течение 1 минуты с использованием шаровой мельницы Kelco. Затем осуществляли стандартный способ выделения ДНК с использованием DNeasy (Qiagen, DNeasy 69109). Затем аликвоту выделенной ДНК окрашивали Pico Green (Molecular Probes P7589) и анализировали с помощью флуориметра (BioTek) с известными стандартами для получения концентрации в нг/мкл.
Анализ Invader: Образцы ДНК разбавляли до 20 нг/мкл, затем подвергали денатурации посредством инкубации в термоциклере при 95°C в течение 10 минут. Затем получали смесь Signal Probe mix с использованием предоставленной смеси олигонуклеотидов и MgCl2 (Third Wave Technologies). Аликвоту 7,5 мкл помещали в каждую лунку планшета для анализа Invader, затем аликвоту 7,5 мкл контролей, стандартов и 20 нг/мкл разбавленных неизвестных образцов. Каждую лунку покрывали 15 мкл минерального масла (Sigma). Затем планшеты инкубировали при 63°C в течение 1 часа и анализировали с помощью флуориметра (Biotek).
С помощью вычисления % сигнала выше фона для целевого зонда, разделенного на % сигнала выше фона для зонда внутреннего контроля, получали соотношение. Соотношение для стандартов с известной копийностью, полученное и валидированное с помощью анализа Саузерн-блоттинга, использовали для определения приблизительной копийности для неизвестных объектов.
Полимеразная цепная реакция: В качестве матрицы использовали всего 100 нг тотальной ДНК. Использовали 20 мМ каждого праймера с набором Takara Ex Taq PCR Polimerase kit (Minis TAKRR001A). Праймерами для PTU AAD-12 (v1) являлись прямой GAACAGTTAGACATGGTCTAAAGG (SEQ ID NO: 8) и обратный GCTGCAACACTGATAAATGCCAACTGG (SEQ ID NO: 9). Реакцию ПЦР осуществляли в термоциклере 9700 Geneamp (Applied Biosystems), подвергая образцы воздействию 94°C в течение 3 минут и 35 циклам воздействия 94°C в течение 30 секунд, 63°C в течение 30 секунд и 72°C в течение 1 минуты и 45 секунд, затем 72°C в течение 10 минут.
Праймерами для ПЦР кодирующей области AAD-12 (v1) являлись прямой ATGGCTCAGACCACTCTCCAAA (SEQ ID NO: 10) и обратный AGCTGCATCCATGCCAGGGA (SEQ ID NO: 11). Реакцию ПЦР осуществляли в термоциклере 9700 Geneamp (Applied Biosystems), подвергая образцы воздействию 94°C в течение 3 минут и 35 циклам воздействия 94°C в течение 30 секунд, 65°C в течение 30 секунд, и 72°C в течение 1 минуты и 45 секунд, затем 72°C в течение 10 минут. Продукты ПЦР анализировали с помощью электрофореза в 1% агарозном геле, окрашиваемом EtBr.
Анализ Саузерн-блоттинга: Анализ Саузерн-блоттинга осуществляли с использованием геномной ДНК, полученной с помощью набора Qiagen DNeasy. Всего 2 мкг геномной ДНК из листьев или 10 мкг геномной ДНК из каллюса в течение ночи подвергали расщеплению рестрикционными ферментами BsmI и SwaI для получения данных о PTU.
После расщепления в течение ночи аликвоту ~100 нг анализировали с использованием 1%-ного геля, чтобы убедиться в полном расщеплении. После этой проверки образцы анализировали с использованием большого 0,85%-ного агарозного геля в течение ночи при 40 Вольтах. Затем гель подвергали денатурации в 0,2 M NaOH, 0,6 M NaCl в течение 30 минут. Затем гель подвергали нейтрализации в 0,5 M Трис-HCl, 1,5 M NaCl, pH 7,5, в течение 30 минут. Затем использовали устройство для блоттинга, содержащее 20-кратный SSC, для осуществления переноса геля на нейлоновую мембрану (Millipore INYC00010) под действием силы тяжести в течение ночи. После переноса в течение ночи мембрану подвергали воздействию УФ-излучения с помощью кросслинкера (Stratagene UV stratalinker 1800) при 1200×100 микроджоулях. Затем мембрану промывали в 0,1% SDS, 0,1 SSC в течение 45 минут. После промывания в течение 45 минут мембрану сушили в течение 3 часов при 80°C, а затем хранили при 4°C до гибридизации. Фрагмент матрицы для гибридизации получали с помощью указанной выше ПЦР кодирующей области с использованием плазмидной ДНК. Продукт подвергали электрофорезу в 1%-ном агарозном геле и вырезали, а затем экстрагировали из геля с использованием способа экстракции из геля Qiagen (28706). Затем мембрану подвергали прегибридизации на этапе 60°C в течение 1 часа в буфере Perfect Hyb (Sigma H7033). Способ Prime it RmT dCTP-labeling rxn (Stratagene 300392) использовали для получения зонда на основе p32 (Perkin Elmer). Зонд очищали с использованием колонок Probe Quant. G50 (Amersham 27-5335-01). Два миллиона CPM использовали для гибридизации Саузерн-блотов в течение ночи. После гибридизации в течение ночи блоты подвергали двум промываниям по 20 минут при 65°C в 0,1% SDS, 0,1% SSC. Затем блоты экспонировали с пленкой в течение ночи, инкубируя при -80°C.
Послевсходовая устойчивость к гербицидам у кукурузы T0, трансформированной с использованием AAD-12: Четырем объектам T0 позволяли акклиматизироваться в теплице и выращивали до появления 2-4 новых, выглядящих нормальными листьев из мутовки (т.е. растения перешли от культуры тканей в условия выращивания в теплице). Растения выращивали при 27°C в теплице в условиях 16 часов освещения: 8 часов темноты. Затем растения обрабатывали коммерческими составами Pursuit® (имазетапир) или 2,4-D-амин 4. Pursuit® распыляли для демонстрации функционирования гена селективного маркера, присутствующего в тестируемых объектах. Нанесение гербицидов осуществляли с использованием автоматического устройства для опрыскивания при объеме раствора для опрыскивания 187 л/га, высота распыления 50 см. Растения опрыскивали летальной дозой имазетапира (70 г кэ/га) или количеством соли 2,4-D DMA, способным значительно повреждать нетрансформированные линии кукурузы (2240 г кэ/га). Летальную дозу определяли как количество, вызывающее >95% повреждения у инбредных Hi-II. Hi-II представляет собой генетический фон для трансформантов по настоящему изобретению.
Несколько отдельных растений являлись невредимыми после обработки гербицидами, резистентность к которым им придали соответствующие гены. Однако, отдельный клон '001' объекта ''001'' (так называемый 4101(0)-001-001) получил незначительные повреждения, но восстановился к 14 DAT. Три из четырех объектов использовали в дальнейшем, отдельные растения скрещивали с 5XH751 и получали следующее поколение. Каждое устойчивое к гербициду растение являлось положительным на наличие кодирующей области AAD-12 (анализ ПЦР) или наличие гена AHAS (анализ Invader) в случае растений, устойчивых к 2,4-D и имазетапиру, соответственно. Белок AAD-12 определяли у всех устойчивых к 2,4-D растений T0, содержащих интактную кодирующую область. Копийность трансгенов (AHAS и, как подразумевают, AAD-12) значительно варьировалась от 1 до 15 копий. Отдельные растения T0 выращивали до созревания и перекрестно опыляли с патентованной инбредной линией для получения семян T1.
Верификация высокой устойчивости к 2,4-D у кукурузы T1: Семена T1 с AAD-12 (v1) высевали в 3-дюймовые горшки, содержащие среды Metro Mix, и в фазу 2 листьев растения опрыскивали 70 г кэ/га имазетапира для устранения растений, не содержащих вставку. Выжившие растения пересаживали в 1-галлонные горшки, содержащие среды Metro Mix, и помещали в те же условия выращивания, которые описаны выше. В фазу V3-V4 растения опрыскивали в автоматическом устройстве для опрыскивания, установленном на 187 л/га при 560 или 2240 г кэ/га 2,4-D DMA. Растения классифицировали в 3 и 14 DAT и сравнивали с контрольными растениями 5XH751 x Hi-II. Шкалу оценок 0-10 (от отсутствия повреждений до критических повреждений ауксинами) разрабатывали для различения повреждений опорного корня. Оценку опорного корня осуществляли в 14 DAT для демонстрации устойчивости к 2,4-D. 2,4-D вызывает деформацию опорного корня, что является соответствующим индикатором повреждения ауксиновыми гербицидами у кукурузы. Данные оценки опорного корня (как видно из таблицы ниже) свидетельствуют о том, что 2 из 3 тестируемых объектов являлись стабильно устойчивыми к 2240 г кэ/га 2,4-D DMA. Объект "pDAB4101(0)001.001" являлся очевидно нестабильным; однако, два других объекта являлись стабильно устойчивыми к 2,4-D и 2,4-D + имазетапир или 2,4-D + глифосат (см. таблицу 12).
"Herbicide" - "Гербицид", "Untransformed control" - "Нетрансформированный контроль", "g ae/ha" - "г кэ/га", "A scale of 0-10, 10 being the highest, was used for grading the 2,4-D DMA injury. Results are a visual average of four replications per tratment" - "Шкалу от 0 до 10, где 10 является наивысшим баллом, использовали для классификации повреждений от 2,4-D DMA. Результаты представляют собой среднее при визуальной оценке для четырех повторений на обработку"]
Повреждение опорного корня трансформированных с использованием AAD-12 (v1) растений T1 и нетрансформированных контрольных растений кукурузы: среднее повреждение опорного корня
(шкала 0-10)
Наследование AAD-12 (v1) у кукурузы: Также проводили тестирование потомства семи семей T1 с AAD-12 (v1), скрещенных с 5XH751. Семена сажали в трехдюймовые горшки, как описано выше. В фазу трех листьев все растения опрыскивали 70 г кэ/га имазетапира в автоматическом устройстве для опрыскивания, как описано выше. Через 14 DAT подсчитывали резистентные и восприимчивые растения. Четыре из шести тестируемых линий расщеплялись в соответствии с единым локусом, доминантным признаком с менделевским наследованием (1R:1S), как определяли при анализе с использованием хи-квадрата. Затем выжившие растения опрыскивали 2,4-D и все растения считали устойчивыми к 2,4-D (количества >560 г кэ/га). AAD-12 наследуется как устойчивый ген резистентности к арилоксиалканоатауксинам во многих видах при реципрокном скрещивании с коммерческим гибридом.
Стэкинг AAD-12 (v1) для повышения спектра гербицидов: AAD-12 (v1) (pDAB4101) и элитную инбредную линию Roundup Ready (BE1146RR) реципрокно скрещивали и собирали семена F1. Сажали семена от двух линий F1 и обрабатывали 70 г кэ/га имазетапира в фазу V2 для устранения, растений, не содержащих вставку. Из выживших растений повторяющиеся разделяли и обрабатывали 1120 г кэ/га 2,4-D DMA + 70 г кэ/га имазетапира (для подтверждения наличия гена AHAS) или 1120 г кэ/га 2,4-D DMA + 1680 г кэ/га глифосата (для подтверждения наличия гена Round Up Ready) в автоматическом устройстве для опрыскивания, откалиброванном на 187 л/га. Растения классифицировали в 3 и 16 DAT. Данные опрыскивания показали, что AAD-12 (v1) можно общепринятым образом подвергать стэкингу с геном устойчивости к глифосату (такому как ген Roundup CP4-EPSPS) или другими генами устойчивости к гербицидам для получения увеличенного спектра гербицидов, которые можно безопасно наносить на кукурузу. Аналогично, у растений F1 наблюдали устойчивость к имидазолинону + 2,4-D + глифосату и не наблюдали негативного фенотипа при молекулярных или бридинговых стэковых комбинациях этих многочисленных трансгенов.
Данные, демонстрирующие увеличение спектра устойчивости к гербицидам в результате AAD-12 (v1) и BE1146RR (элитной устойчивой инбредной линии, сокращенной обозначаемой как AF) у F1: средний % повреждения через 16 DAT
Устойчивость трансформированных растений кукурузы pDAB4101 к гербицидам 2,4-D, триклопиру и флуроксипиру в полевых условиях: Полевые испытания уровня устойчивости проводили на двух объектах с AAD-12 (v1) pDAB4101 (4101(0)003.R.003.AF и 4101(0)005.R001.AF) и одном контрольном гибриде Roundup Ready (RR) (2P782) в Fowler, Ind. и Wayside, Miss. Семена сажали с использованием конической сеялки с междурядным расстоянием 40 дюймов в Wayside и 30 дюймов в Fowler. Дизайн эксперимента представлял собой рандомизированный полноблочный план с 3 повторениями. Варианты эксперимента представляли собой 2,4-D (диметиламиновая соль) при 1120, 2240 и 4480 г кэ/га, триклопир при 840 г кэ/га, флуроксипир при 280 г кэ/га и необработанный контроль. Объекты с AAD-12 (v1) содержали ген AHAS в качестве селективного маркера. Объекты кукурузы F2 расщеплялись, поэтому растения с AAD-12 (v1) обрабатывали имазетапиром при 70 г кэ/га для удаления растений, не содержащих вставку. Когда кукуруза достигала фазы V6, осуществляли обработки гербицидами с использованием пневматического ранцевого опрыскивателя, выпускающего объем раствора для опрыскивания 187 л/га под давлением 130-200 кПа. Визуальную оценку повреждений осуществляли в 7, 14 и 21 дни после обработки. Оценку повреждения опорного корня осуществляли на 28 DAT по шкале 0-10, где 0-1 представляет сращение опорного корня, 1-3 - умеренное утолщение опорного корня/отклонение и пролиферация корня, 3-5 - умеренное сращение опорного корня, 5-9 - тяжелое сращение опорного корня и деформация, и 10 представляет полное ингибирование опорных корней.
Ответ объекта с AAD-12 (v1) на 2,4-D, триклопир и флуроксипир через 14 дней после обработки представлен в таблице 14. Повреждение сельскохозяйственной культуры являлось наиболее тяжелым через 14 DAT. Контрольная кукуруза RR (2P782) являлась тяжело поврежденной (44% через 14 DAT) 2,4-D при 4480 г кэ/га, что представляет собой 8-кратную (8 x) норму внесения в полевых условиях. Все объекты с AAD-12 (v1) демонстрировали исключительную устойчивость к 2,4-D через 14 DAT при 0% повреждений при 1-, 2- и 4-кратных нормах внесения, соответственно. Контрольная кукуруза (2P782) являлась тяжело поврежденной (31% через 14 DAT) при 2-кратной норме внесения триклопира (840 г кэ/га). Объекты с AAD-12 (v1) демонстрировали устойчивость при 2-кратной норме внесения триклопира в среднем с 3% повреждения через 14 DAT для двух объектов. Флуроксипир при 280 г кэ/га вызывал 11% видимых повреждений кукурузы дикого типа через 14 DAT. Объекты с AAD-12 (v1) демонстрировали повышенную устойчивость в среднем с 8% повреждения через 5 DAT.
Видимые повреждения объектов с AAD-12 и кукурузы дикого типа после некорневых обработок 2,4-D, триклопиром и флуоксипиром в полевых условиях: % видимых повреждений через 14 DAT
(г кэ/га)
Обработка ауксиновыми гербицидами кукурузы в фазе роста V6 может вызывать деформацию опорных корней. В таблице 15 представлена тяжесть повреждения опорного корня, вызываемого 2,4-D, триклопиром и флуроксипиром. Триклопир при 840 г кэ/га вызывал наиболее тяжелое срастание опорного корня и деформацию, приводящие к средней оценке повреждения опорного корня у контрольной кукурузы дикого типа 2P782 в 7 баллов.
"Treatment" - "Обработка", "Rate (g ae/ha)" - "Норма внесения (г кэ/га)", "control" - "контроль", "Untreated" - "Необработанные", "Fluoxypyr" - "Флуоксипир", "Triclopyr" - "Триклопир", "Dicamba" - "Дикамба"]
Оценка повреждения опорного корня у растений кукурузы с AAD-12 и дикого типа в ответ на 2,4-D, триклопир и флуоксипир в полевых условиях: Оценка повреждения опорного корня (по шкале 0-10) через 28 DAT
Оба объекта кукурузы с AAD-12 (v1) демонстрировали отсутствие повреждения опорного корня после обработки триклопиром. Повреждение опорного корня у кукурузы 2P782 повышалось с повышением норм внесения 2,4-D. При 4480 г кэ/га 2,4-D объекты с AAD-12 демонстрировали отсутствие повреждения опорного корня; в то время как у гибрида 2P782 наблюдали тяжелое срастание опорного корня и деформацию. Флуроксипир вызывал только умеренное утолщение и отклонение опорного корня у кукурузы дикого типа, при этом у объектов с AAD-12 (v1) не наблюдали повреждения опорного корня.
Эти данные четко свидетельствуют о том, что AAD-12 (v1) придает кукурузе высокий уровень устойчивости к 2,4-D, триклопиру и флуроксипиру при нормах внесения, сильно превышающих коммерчески используемые и вызывающих у кукурузы без AAD-12 (v1) тяжелые видимые повреждения и повреждения опорного корня.
ПРИМЕР 6
Трансформация табака
Трансформацию табака с использованием Agrobacterium tumefaciens осуществляли способом аналогичным, но идентичным, опубликованным способам (Horsch et al., 1988). Для получения источника ткани для трансформации семена табака (Nicotiana tabacum cv. KY160) поверхностно стерилизовали и высевали на поверхность среды TOB, представляющей собой среду Мурашиге-Скуга без гормонов (Murashige and Skoog, 1962), отвержденную агаром. Растения выращивали в течение 6-8 недель в освещенной инкубаторной комнате при 28-30°C и стерильно собирали листья для использования в трансформации. Из этих листьев стерильно вырезали фрагменты размером приблизительно один квадратный сантиметр, исключая среднюю жилку. Культуры штаммов Agrobacterium (EHA101S, содержащего pDAB3278, также обозначаемый как pDAS1580, AAD-12 (v1)+PAT), выращенные в течение ночи во флаконе на шейкере, установленном на 250 об./мин., при 28°C, осаждали центрифугированием, ресуспендировали в стерильных солях Мурашиге-Скуга и доводили до конечной оптической плотности 0,5 при 600 нм. Фрагменты листьев погружали в эту бактериальную суспензию приблизительно на 30 секунд, затем высушивали на стерильных бумажных салфетках, помещали правой стороной вверх на среду TOB+ (среда Мурашиге-Скуга, содержащая 1 мг/л индолил-3-уксусной кислоты и 2,5 мг/л бензиладенина) и инкубировали в темноте при 28°C. Через два дня фрагменты листьев перемещали на среду TOB+, содержащую 250 мг/л цефотаксима (Agri-Bio, North Miami, Fla.) и 5 мг/л глуфосината аммония (активный ингредиент в Basta, Bayer Crop Sciences), и инкубировали при 28-30°C при освещении. Фрагменты листьев перемещали на свежую среду TOB+ с цефотаксимом и Basta дважды в неделю в течение первых двух недель и раз в неделю впоследствии. Через 4-6 недель после обработки фрагментов листьев с использованием бактерий небольшие растения, развившиеся из трансформированных очагов, удаляли из этого препарата ткани и высаживали в среду TOB, содержащую 250 мг/л цефотаксима и 10 мг/л Basta, в сосуды Phytatray™ II (Sigma). Эти проростки выращивали в освещенной инкубаторной комнате. Через 3 недели получали фрагменты стеблей и переукореняли в тех же средах. Растения были готовы к отправке в теплицу еще через 2-3 недели.
Растения перемещали в теплицу, смывая агар с корней, пересаживая в почву в квадратные горшки 13,75 см, помещая горшок в мешок Ziploc® (SC Johnson & Son, Inc.), помещая водопроводную воду на дно мешка, и помещая его под непрямое освещение в теплицу при 30°C на одну неделю. Через 3-7 открывали мешки; растения удобряли и позволяли им расти в открытом мешке до акклиматизации растений в теплице, когда мешок удаляли. Растения выращивали в общепринятых теплых условиях в теплице (30°C, 16 часов освещения, 8 часов темноты, минимальный естественный + дополнительный свет = 500 мЭ/м2 с1).
До размножения из растений T0 брали образцы на анализ ДНК для определения копийности вставки. Для удобства анализировали ген PAT, молекулярно связанный с AAD-12 (v1). Свежую ткань помещали в пробирки и лиофилизировали при 4°C в течение 2 дней. После полного высушивания ткани в пробирку помещали гранулу вольфрама (Valenite) и образцы подвергали сухому измельчению в течение 1 минуты с использованием шаровой мельницы Kelco. Затем осуществляли стандартное выделение ДНК с использованием DNeasy (Qiagen, DNeasy 69109). Затем аликвоту выделенной ДНК окрашивали Pico Green (Molecular Probes P7589) и анализировали с помощью флуориметра (BioTek) с известными стандартами для получения концентрации в нг/мкл.
Образцы ДНК разбавляли до 9 нг/мкл, а затем подвергали денатурации посредством инкубации в термоциклере при 95°C в течение 10 минут. Затем получали смесь Signal Probe mix с использованием предоставленной смеси олигонуклеотидов и MgCl2 (Third Wave Technologies). Аликвоту 7,5 мкл помещали в каждую лунку планшета для анализа Invader, затем аликвоту 7,5 мкл контролей, стандартов и 20 нг/мкл разбавленных неизвестных образцов. Каждую лунку покрывали 15 мкл минерального масла (Sigma). Затем планшеты инкубировали при 63°C в течение 1,5 часов и анализировали с помощью флуориметра (Biotek).
При вычислении % сигнала выше фона для целевого зонда, разделенного на % сигнала выше фона для зонда внутреннего контроля, получали соотношение. Соотношение для стандартов с известной копийностью, полученных и валидированных с помощью анализа Саузерн-блоттинга, использовали для идентификации приблизительной копийности для неизвестных объектов.
Все объекты также анализировали на наличие гена AAD-12 (v1) посредством ПЦР с использованием тех же выделенных образцов ДНК. В качестве матрицы всего использовали 100 нг тотальной ДНК. Использовали 20 мМ каждого праймера с набором Takara Ex Taq PCR Polimerase. Праймерами для ПЦР транскрипционной единицы растений (PTU) AAD-12 являлись (SdpacodF: ATGGCTCATGCTGCCCTCAGCC) (SEQ ID NO: 12) и (SdpacodR: CGGGCAGGCCTAACTCCACCAA) (SEQ ID NO: 13). Реакцию ПЦР осуществляли в термоциклере 9700 Geneamp (Applied Biosystems), подвергая образцы воздействию 94°C в течение 3 минут и 35 циклам воздействия 94°C в течение 30 секунд, 64°C в течение 30 секунд и 72°C в течение 1 минуты и 45 секунд, затем 72°C в течение 10 минут. Продукты ПЦР анализировали с помощью электрофореза в 1%-ном агарозном геле, окрашиваемом EtBr. 4-12 клональных линий от каждого из 18 ПЦР-позитивных объектов с 1-3 копиями гена PAT (и, очевидно, AAD-12 (v1), т.к. эти гены физически связаны) подвергали регенерации и переносили в теплицу.
Объекты табака T0, трансформированные с использованием pDAS1580 (AAD-12 (v1)+PAT)
Послевсходовая устойчивость к гербицидам в трансформированных растениях табака T0 с AAD-12 (v1): Растения T0 от каждого из 19 объектов проверяли с использованием широкого диапазона 2,4-D, триклопира или флуроксипира, распыляемых на растения высотой
3-4 дюйма. Опрыскивания осуществляли, как описано выше, с использованием автоматического устройства для опрыскивания при объеме раствора для опрыскивания 187 л/га. На соответствующие клоны от каждого объекта наносили диметиламиновую соль 2,4-D (Riverside Corp), смешанную с деионизированной водой, в количестве 0, 140, 560 или 2240 г кэ/га. Флуроксипир наносили аналогично в количестве 35, 140 или 560 г кэ/га. Триклопир наносили в количестве 70, 280 или 1120 г кэ/га. Каждую обработку повторяли 1-3 раза. Оценки повреждений проводили в 3 и 14 DAT. Каждый тестируемый объект являлся более устойчивым к 2,4-D, чем нетрансформированная контрольная линия KY160. У нескольких объектов возникала некоторая начальная эпинастия, связанная с ауксиновыми гербицидами, при дозах 560 г кэ/га 2,4-D или менее. Некоторые объекты являлись неповрежденными, когда 2,4-D наносили в количестве 2240 г кэ/га (эквивалентно 4-кратной норме внесения). В целом, объекты с AAD-12 (v1) являлись более восприимчивыми к флуроксипиру, затем - триклопиру, и меньше всего повреждались 2,4-D. Качество объектов в отношении величины резистентности определяли с использованием ответов растений T0 на 560 г кэ/га флуроксипира. Резистентность объектов классифицировали как "низкую" (>40% повреждений через 14 DAT), "среднюю" (20-40% повреждений), "высокую" (<20% повреждений). Некоторые объекты имели непостоянный ответ среди дублирующих растений, и их считали "переменными".
Подтверждение высокой устойчивости к 2,4-D у табака T1: От каждого объекта сохраняли от двух до четырех отдельных растений T0, выживших после воздействия высоких доз 2,4-D и флуроксипира, и позволяли им самоопыляться в теплице для получения семян T1. Семена T1 стратифицировали и высевали в лотки для селекции, так же, как в случае Arabidopsis, с последующим избирательным удалением нетрансформированных растений без вставок в этой расщепленной популяции с использованием 560 г активного ингредиента/га глуфосината (селекция по гену PAT). Выжившие растения переносили в отдельные 3-дюймовые горшки в теплицу. Эти линии обеспечивали высокие уровни резистентности к 2,4-D в поколении T0. У растений T1, полученных не непосредственно из культуры ткани, предполагали улучшенное постоянство ответа. Эти растения сравнивали с табаком дикого типа KY160. Все растения опрыскивали с использованием автоматического устройства для опрыскивания, установленного на 187 л/га. Растения опрыскивали количествами диметиламиновой соли (DMA) 2,4-D в диапазоне 140-2240 г кэ/га, триклопира - в диапазоне 70-1120 г кэ/га, или флуроксипира - в диапазоне 35-560 г кэ/га. Все растворы для опрыскивания составляли в воде. Каждую обработку повторяли 2-4 раза. Растения оценивали через 3 и 14 дней после обработки. Растениям присваивали степень повреждения в отношении замедленного роста, хлороза и некроза. Поколение T1 расщеплялось, поэтому ожидали некоторый переменный ответ по причине различий в зиготности.
Ответ расщепляющихся растений табака T1 с AAD-12 на фенокси- и пиридилоксиауксиновые гербициды
В случае 2,4-D не наблюдали повреждений при 4-кратной норме внесения (2240 г кэ/га) или ниже. Некоторые повреждения наблюдали при обработках триклопиром в одной линии объектов, но наибольшие повреждения наблюдали при обработке флуроксипиром. Повреждения флуроксипиром являлись кратковременными, и прирост на одном объекте являлся практически неотличимым от необработанного контроля на 14 DAT (таблица 17). Важно отметить, что нетрансформированный табак чрезвычайно восприимчив к флуроксипиру. Эти результаты свидетельствуют о том, что с помощью AAD-12 (v1) можно получать коммерческий уровень устойчивости к 2,4-D, даже у крайне восприимчивой к ауксинам двудольной сельскохозяйственной культуры, такой как табак. Эти результаты также показали, что можно придавать резистентность к гербицидам на основе пиридилоксиуксусной кислоты, триклопиру и флуроксипиру. Наличие возможности назначать обработку устойчивой к гербицидам сельскохозяйственной культуры, защищенной AAD-12, с использованием различных активных ингредиентов, имеющих различные спектры контроля сорняков, является крайне применимым для сельхозпроизводителей.
Наследование AAD-12 (v1) у табака: Также осуществляли тестирование 100 растений-потомков в семи линиях T1 из линий с AAD-12 (v1). Семена стратифицировали, высевали и пересаживали в соответствии со способом, описываемым выше, за исключением того, что растения, не содержащие вставку, не удаляли посредством селекции с использованием Liberty. Затем все растения опрыскивали 560 г кэ/га 2,4-D DMA, как описано выше. Через 14 DAT подсчитывали резистентные и восприимчивые растения. Пять из семи тестируемых линий расщеплялись в соответствии с одним локусом, доминантным признаком с менделевским наследованием (3R: 1S), как определяли с помощью анализа с использованием хи-квадрата. AAD-12 наследуется как устойчивый ген резистентности к арилоксиалканоатауксинам во многих видах.
Устойчивость растений табака с pDAS1580 к гербицидам 2,4-D, дихлорпропу, триклопиру и флуроксипиру в полевых условиях: Полевые испытания уровня устойчивости осуществляли на трех линиях с AAD-12 (v1) (объекты pDAS1580-[1]-018.001, pDAS1580-[1]-004.001 и pDAS1580-[1]-020.016) и одной линии дикого типа (KY160) на полевых станциях в Indiana и Miss. Саженцы табака выращивали в теплице посредством посадки семян T1 в 72-луночные поддоны для рассады (Hummert International), содержащие среды Metro 360, согласно указанным выше условиям выращивания. Растения без вставки избирательно удаляли посредством селекции с использованием Liberty, как описано выше. Саженцы переносили на полевые станции и сажали с интервалом 14 или 24 дюймов с использованием промышленных рассадопосадочных машин. Для поддержания интенсивного роста растений на станции в Миссисипи использовали капельное орошение, а на станции в Индиане - орошение дождеванием.
Дизайн эксперимента представлял собой план с расщепленными делянками с 4 повторениями. Основной делянкой являлась обработка гербицидом, и подделянкой являлась линия табака. Варианты эксперимента представляли собой 2,4-D (диметиламиновая соль) в количестве 280, 560, 1120, 2240 и 4480 г кэ/га, триклопир в количестве 840 г кэ/га, флуроксипир в количестве 280 г кэ/га и необработанный контроль. Опытные участки содержали один ряд по 25-30 футов. Обработки гербицидами осуществляли через 3-4 недели после пересаживания с использованием пневматического ранцевого опрыскивателя, выпускающего объем раствора для опрыскивания 187 л/га под давлением 130-200 кПа. Визуальную оценку повреждений, ингибирования роста и эпинастии проводили на 7, 14 и 21 день после обработки.
Ответ растений табака с AAD-12 на 2,4-D, триклопир и флуроксипир в полевых условиях
Ответ объектов с AAD-12 (v1) на 2,4-D, триклопир и флуроксипир представлен в таблице 18. Нетрансформированная линия табака являлась тяжело поврежденной (63% через 14 DAT) 2,4-D при 560 г кэ/га, рассматриваемой как 1-кратная норма внесения. Все линии AAD-12 (v1) демонстрировали исключительную устойчивость к 2,4-D через 14 DAT со средними повреждениями 1, 4 и 4% повреждения, наблюдаемыми при 2-, 4- и 8-кратных нормах внесения, соответственно. Нетрансформированная линия табака являлась тяжело поврежденной (53% через 14 DAT) 2-кратной нормой внесения триклопира (840 г кэ/га); в то время как линии с AAD-12 (v1) демонстрировали устойчивость в среднем с 5% повреждения через 14 DAT среди трех линий. Флуроксипир при 280 г кэ/га вызывал тяжелые повреждения (99%) у нетрансформированной линии через 14 DAT. Линии с AAD-12 (v1) демонстрировали повышенную устойчивость в среднем с 11% повреждения через 14 DAT.
Эти результаты свидетельствуют о том, что линии трансформированных объектов с AAD-12 (v1) проявляют высокий уровень устойчивости к 2,4-D, триклопиру и флуроксипиру при кратном повышении коммерчески используемых норм внесения, являющихся летальными или вызывающими тяжелые эпинастические деформации у нетрансформированного табака в типичных полевых условиях.
Защита с помощью AAD-12 (v1) от повышенных количеств 2,4-D: Результаты, демонстрирующие защиту с помощью AAD-12 (v1) от повышенных количеств 2,4-D DMA в теплице, представлены в таблице 19. Растения T1 с AAD-12 (v1) от объекта расщеплялись как 3R:1S при селекции с использованием 560 г активного ингредиента/га Liberty с использованием того же способа, как описано выше. Семена T1 с AAD-1 (v3) также сажали в качестве трансформированных контролей табака (см. PCT/US2005/014737). Нетрансформированные KY160 служили в качестве восприимчивого контроля. Растения опрыскивали с использованием автоматического устройства для опрыскивания, установленного на 187 л/га при 140, 560, 2240, 8960 и 35840 г кэ/га 2,4-D DMA, и оценивали на 3 и 14 DAT.
AAD-12 (v1) и AAD-1 (v3) эффективно защищали табак от повреждения 2,4-D при дозах до 4-кратной коммерчески используемой нормы внесения. Однако, растения с AAD-12 (v1) четко демонстрировали значительное преимущество по сравнению с AAD-1 (v3), защищая от 64-кратных стандартных норм внесения.
"Treatment" - "Обработка", "control" - "контроль", "Average % Injury of Replicates 14 DAT" - "Средний % повреждений у дублирующих растений через 14 DAT", "g ae/ha" - "г кэ/га"]
Результаты, демонстрирующие защиту от повышенных количеств 2,4-D, обеспечиваемую AAD-12 (v1) и AAD-1 (v3)
Стэкинг AAD-12 для повышения гербицидного спектра: Реципрокно скрещивали растения, гомозиготные по AAD-12 (v1) (pDAS1580) и AAD-1 (v3) (pDAB721) (см. PCT/US2005/014737 в случае последнего), и собирали семена F1. Семена F1 от двух реципрокно скрещенных растений с каждым геном стратифицировали и обрабатывали 4 дублирующих растения от каждого скрещивания по той же схеме опрыскивания, которую использовали для других тестов с одной из следующих обработок: 70, 140, 280 г кэ/га флуроксипира (селекция по гену AAD-12 (v1)); 280, 560, 1120 г кэ/га R-дихлорпропа (селекция по гену AAD-1 (v3)); или 560, 1120, 2240 г кэ/га 2,4-D DMA (для подтверждения устойчивости к 2,4-D). Также сажали гомозиготные растения T2 с каждым геном для использования в качестве контролей. Растения классифицировали через 3 и 14 DAT. Результаты опрыскивания представлены в таблице 20.
Результаты подтверждают, что AAD-12 (v1) можно успешно подвергать стэкингу с AAD-1 (v3), таким образом, повышая спектр гербицидов, которые можно наносить на интересующую сельскохозяйственную культуру (феноксиуксусные кислоты + феноксипропионовые кислоты vs. феноксиуксусных кислот + пиридилоксиуксусных кислот для AAD-1 и AAD-12, соответственно). Дополняющая природа паттернов перекрестной резистентности к гербицидам делает возможным удобное использование двух этих генов в качестве дополняющих и способных к стэкингу селективных маркеров в полевых условиях. В случае сельскохозяйственных культур, у которых устойчивость при использовании одного гена может быть незначительной, специалисту в этой области будет понятно, что можно повышать устойчивость посредством стэкинга со вторым геном устойчивости к тому же гербициду. Это можно осуществлять с использованием того же гена с теми же или другими промоторами; однако, как наблюдали в этом случае, стэкинг и трекинг двух дополняющих признаков можно облегчать посредством различающейся перекрестной защиты от феноксипропионовых кислот [с помощью AAD-1 (v3)] или пиридилоксиксусных кислот [с помощью AAD-12 (v1)].
Сравнение перекрестной устойчивости к ауксиновым гербицидам у растений T2 с AAD-12 (v1) (pDAS1580) и AAD-1 (v3) (pDAB721) с гибридом F1 AAD-12 x AAD-1 и диким типом
ПРИМЕР 7
Трансформация сои
Улучшение сои осуществляют с помощью способов переноса генов для таких признаков как устойчивость к гербицидам (Padgette et al., 1995), модификация аминокислот (Falco et al., 1995) и резистентность к насекомым (Parrott et al., 1994). Для введения чужеродных признаков в сельскохозяйственную культуру необходимы способы, которые сделают возможным рутинное получение трансгенных линий с использованием последовательностей селективных маркеров, содержащих простые вставки. Для упрощения разведения трансгены должны наследоваться как единый функциональный локус. Сообщают о доставке чужеродных генов в культурную сою посредством бомбардировки микрочастицами рубчика зиготного эмбриона (McCabe et al., 1988) или соматических эмбриогенных культур (Finer and McMullen, 1991) и опосредованной Agrobacterium трансформации эксплантатов семядоли (Hinchee et al., 1988) или зиготных эмбрионов (Chee et al., 1989).
Трансформанты, полученные посредством опосредованной Agrobacterium трансформации, как правило, содержат простые вставки с низкой копийностью (Birch, 1991). Существуют преимущества и недостатки, связанные с каждой из трех тканей-мишеней, исследуемых на предмет переноса генов в сою, рубчиком зиготных эмбрионов (Chee et al., 1989; McCabe et al., 1988), семядолей (Hinchee et al., 1988) и соматическими эмбриогенными культурами (Finer и McMullen, 1991). Последние из них интенсивно исследуют как ткань-мишень для прямого переноса генов. Эмбриогенные культуры, как правило, хорошо пролиферируют, и их можно поддерживать в течение длительного периода. Однако срок хранения эмбриогенных суспензий связан со стерильностью и хромосомными аберрациями первичных трансформантов (Singh et al., 1998), и, таким образом, для систем трансформации сои с использованием этой ткани, по-видимому, необходима непрерывная инициация новых культур. Для этой системы необходим высокий уровень 2,4-D, концентрация 40 мг/л, для инициации эмбриогенного каллюса, и это представляет фундаментальную проблему в использовании гена AAD-12 (v1), т.к. трансформированный локус не может развиваться в дальнейшем с 2,4-D в среде. Таким образом, трансформация с использованием меристемы является идеальной для разработки резистентных к 2,4-D растений с использованием AAD-12 (v1).
Gateway-клонирование бинарных конструкций: Кодирующую последовательность AAD-12 (v1) клонировали в пять различных донорных векторов Gateway, содержащих различные промоторы растений. Затем полученные экспрессирующие кассеты AAD-12 (v1) для растений клонировали в конечный бинарный вектор Gateway с помощью реакции клоназы LR (Invitrogen Corporation, Carlsbad Calif., кат. №11791-019).
Фрагмент NcoI-SacI, содержащий кодирующую последовательность AAD-12 (v1), вырезали из DASPICO12 и лигировали по соответствующим участкам рестрикции NcoI-SacI в следующие донорные векторы Gateway: pDAB3912 (attL1//промотор CsVMV//AtuORF23 3'UTR//attL2); pDAB3916 (attL1//промотор AtUbi10//AtuORF23 3'UTR//attL2); pDAB4458 (attL1//промотор AtUbi3//AtuORF23 3'UTR//attL2); pDAB4459 (attL1//промотор ZmUbi1//AtuORF23 3'UTR//attL2) и pDAB4460 (attL1//промотор AtAct2//AtuORF23 3'UTR//attL2). Полученные конструкции, содержащие следующие экспрессирующие кассеты для растений, обозначали как: pDAB4463 (attL1//промотор CsVMV//AAD-12 (v1)//AtuORF23 3'UTR//attL2); pDAB4467 (attL1//промотор AtUbi10//AAD-12 (v1)//AtuORF23 3'UTR//attL2); pDAB4471 (attL1//промотор AtUbi3//AAD-12 (v1)//AtuORF23 3'UTR//attL2); pDAB4475 (attL1//промотор ZmUbi1//AAD-12 (v1)//AtuORF23 3'UTR//attL2) и pDAB4479 (attL1//промотор AtAct2//AAD-12 (v1)//AtuORF23 3'UTR//attL2). Эти конструкции проверяли посредством расщепления рестрикционными ферментами и секвенирования.
Экспрессирующие кассеты для растений подвергали рекомбинации в конечный бинарный вектор Gateway pDAB4484 (RB7 MARv3//attR1-ccdB-резистентность к хлорамфениколу-attR2//промотор CsVMV//PATv6//AtuORF1 3'UTR) с помощью реакции клоназы LR Gateway. В технологии Gateway используют сайт-специфическую рекомбинацию на основе фага лямбда вместо использования рестрикционной эндонуклеазы и лигазы для встраивания интересующего гена в экспрессирующий вектор. Invitrogen Corporation, Gateway Technology: A Universal Technology to Clone DNA Sequences for Functional Analysis and Expression in multiple Systems, Technical Manual, Catalog #'s 12535-019 and 12535-027, Gateway Technology Version E, Sep. 22, 2003, #25-022. Рекомбинационные последовательности ДНК (attL, и attR,) и смесь ферментов клоназы LR позволяют переносить любой фрагмент ДНК, фланкированный участком рекомбинации, в любой вектор, содержащий соответствующий участок. Участок attL1 донорного вектора соответствует attR1 бинарного вектора. Аналогично, участок attL2 донорного вектора соответствует attR2 бинарного вектора. Используя технологии Gateway экспрессирующую кассету для растений (из донорного вектора), фланкированную участками attL, можно рекомбинировать по участкам attR бинарного вектора. Полученные конструкции, содержащие следующие экспрессирующие кассеты для растений, помечали как: pDAB4464 (RB7 MARv3//промотор CsVMV//AAD-12 (v1)//AtuORF23 3'UTR//промотор CsVMV//PATv6 AtuORF1 3'UTR); pDAB4468 (RB7 MARv3//промотор AtUbi10//AAD-12 (v1)//AtuORF23 3'UTR//промотор CsVMV//PATv6//AtuORF1 3'UTR); pDAB4472 (RB7 MARv3//промотор AtUbi3//AAD-12 (v1)//AtuORF23 3'UTR//промотор CsVMV//PATv6//AtuORF1 3'UTR); pDAB4476 (RB7 MARv3//промотор ZmUbi1//AAD-12 (v1)//AtuORF23 3'UTR//промотор CsVMV//PATv6 AtuORF1 3'UTR); и pDAB4480 (RB7 MARv3//промотор AtAct2//AAD-12 (v1)//AtuORF23 3'UTR//промотор CsVMV//PATv6//AtuORF1 3'UTR). Эти конструкции проверяли посредством расщепления рестрикционными ферментами и секвенирования.
Способ трансформации 1 - Трансформация, опосредованная Agrobacterium: В первых сообщениях о трансформации сои использовали меристемные клетки в области семядольного узла (Hinchee et al., 1988) и размножение побегов из апикальной меристемы (McCabe et al., 1988). В способе с использованием A. tumefciciens и семядольного узла получали эксплантат и с помощью композиции сред для культивирования стимулировали пролиферацию ауксилярной меристемы в узле (Hinchee et al., 1988). Остается неясным, инициируется ли на самом деле культура дифференцированного, но тотипотентного, каллюса с помощью этих обработок. Выделение многочисленных клонов объекта трансформации из одного эксплантата и редкое выделение химерных растений (Clemente et al., 2000; Olhoft et al., 2003) свидетельствуют о происхождении из одной клетки с последующим размножением трансгенных клеток для получения культуры пролиферирующей трансгенной меристемы или равномерно трансформированного побега, в дальнейшем подвергаемого размножению побегов. Исходно способ размножения побегов сои основан на бомбардировке микрочастицами (McCabe et al., 1988), но недавно его адаптировали для опосредованной Agrobacterium трансформации (Martinell et al., 2002), очевидно не достигающего того же уровня или типа дифференцировки, как при способе с использованием семядольного узла, т.к. система основана на успешной идентификации химер зародышевой линии. Кроме того, этот способ не основан на использовании 2,4-D, что будет идеальным для системы селекции с 2,4-D. Таким образом, способ с использованием семядольного узла может являться способом выбора для разработки резистентных к 2,4-D культурных сортов сои.
Получение устойчивых фенотипов с AAD-12 (v1) с помощью трансформации растений. Для получения трансгенных растений с AAD-12 (v1) использовали полученные из семян эксплантаты "Maverick" и способ опосредованной Agrobacterium трансформации семядольного узла.
Получение Agrobacterium и инокуляция: Для инициации трансформации использовали штамм Agrobacterium EHA101 (Hood et al. 1986), несущий каждый из пяти бинарных векторов pDAB (таблица 8). Каждый бинарный вектор содержит кассету гена AAD-12 (v1) и гена для селекции растений (PAT) в области Т-ДНК. Плазмиды вносили в штамм Agrobacterium EHA101 посредством электропорации. Затем выбранные колонии анализировали на предмет встраивания генов перед обработкой Agrobacterium эксплантатов сои. Семена Maverick использовали во всех экспериментах по трансформации, и их получали из University of Missouri, Columbia, Mo.
Осуществляли опосредованную Agrobacterium трансформацию сои (Glycine max) с использованием гена PAT в качестве селективного маркера вместе с гербицидом глуфосинатом в качестве селективного средства. Семена проращивали на минимальной среде B5 (Gamborg et al. 1968), отвержденной 3 г/л фитагеля (Sigma-Aldrich, St. Louis, Mo.). Затем выбранные побеги переносили на среду для укоренения. Оптимальной схемой селекции являлось использование глуфосината в концентрации 8 мг/л в среде в фазы появления первого и второго побегов и в концентрации 3-4 мг/л в среде в течение роста побега в длину.
Перед переносом выросших в длину побегов (3-5 см) на среду для укоренения отрезанный конец междоузлий погружали в 1 мг/л индолил-3-масляной кислоты на 1-3 мин для стимуляции укоренения (Khan et al. 1994). У побегов проращивали корни в стеклянных культуральных пробирках 25×100 мм, содержащих среду для укоренения, а затем их переносили в почвенную смесь для акклиматизации проростков в Metro-mix 200 (Hummert International, Earth City, Mo.) в открытых коробках Magenta в Conviron. При появлении и росте побега в длину для селекции использовали глуфосинат, активный ингредиент гербицида Liberty (Bayer Crop Science). Укоренившиеся проростки подвергали акклиматизации в открытых коробках Magenta в течение нескольких недель перед скринингом и переносом в теплицу для дальнейшей акклиматизации и оценки.
Анализ предполагаемо трансформированных проростков и анализы для оценки растений T0 в теплице: Для скрининга на предмет предполагаемых трансформантов на концевые листочки из выбранных листьев этих проростков наносили 50 мг/л глуфосината дважды с недельным интервалом для наблюдения результатов. Затем подвергнутые скринингу проростки переносили в теплицу и после акклиматизации на листья снова наносили глуфосинат для подтверждения устойчивости этих проростков в GH и считали их предполагаемыми трансформантами.
В дальнейшем растения, перенесенные в теплицу, можно анализировать на наличие активного гена PAT недеструктивным способом посредством нанесения на часть листа первичного трансформанта T0 или его потомка раствора глуфосината [0,05-2% об./об. гербицида Liberty, предпочтительно - 0,25-1,0% (об./об.) = 500-2000 м.д. глуфосината, Bayer Crop Science]. В зависимости от используемой концентрации оценку повреждения глуфосинатом можно осуществлять через 1-7 дней после обработки. Растения также можно тестировать на устойчивость к 2,4-D недеструктивным способом посредством избирательного нанесения раствора 2,4-D в воде (0,25-1% об./об. коммерческого состава диметиламиновой соли 2,4-D, предпочтительно - 0,5% об./об.=2280 м.д. 2,4-D кэ) на концевой листочек новых распустившихся трехлисточковых узлов, одного или двух, предпочтительно - двух, ниже последнего появившегося трехлисточкового узла. Этот анализ позволяет оценивать восприимчивые к 2,4-D растения через период от 6 часов до нескольких дней после нанесения посредством оценки переориентации или вращения листа на >90 градусов относительно плоскости смежных листочков. Растения, устойчивые к 2,4-D, не будут отвечать на 2,4-D. Растениям T0 позволяли самоопыляться в теплице для получения семян T1. Растения T1 (и в той мере, достаточной для получения клонов растений T0) будут опрыскивать с использованием диапазона доз гербицидов для определения уровня защиты от гербицидов, обеспечиваемой генами AAD-12 (v1) и PAT в трансгенной сое. Нормы внесения 2,4-D, используемые для растений T0, как правило, будут содержать одну или две избирательные нормы внесения в диапазоне 100-1120 г кэ/га, наносимые с использованием автоматического устройства для опрыскивания, как описано выше. Растения T1 будут обрабатывать гербицидом с более широким диапазоном доз 50-3200 г кэ/га 2,4-D. Аналогично, растения T0 и T1 можно подвергать скринингу на резистентность к глуфосинату посредством послевсходовой обработки 200-800 и 50-3200 г кэ/га глуфосината, соответственно. Ген резистентности к глифосату (у растений, трансформированных с использованием конструкций, содержащих EPSPS) или другой устойчивости к глифосату можно оценивать в поколении T1 посредством послевсходовых нанесений глифосата в диапазоне доз 280-2240 г кэ/га глифосат. Отдельные растения T0 оценивали на наличие кодирующей области интересующего гена (AAD-12 (v1) или PAT v6) и копийность. Определение наследования AAD-12 (v1) будут осуществлять с использованием расщепления потомства T1 и T2 в отношении устойчивости к гербицидам, как описано в представленных выше примерах.
Субпопуляцию исходных трансформантов оценивали в поколении T0 указанными выше способами. Любое растение, для которого подтверждали наличие кодирующей области AAD-12 (v1), независимо от промотора, запускающего транскрипцию гена, не отвечало на нанесение 2,4-D на листья, в отличие от сои дикого типа Maverick. Растения, трансформированные исключительно с использованием PAT, отвечали на нанесение 2,4-D на листья так же, как и растения дикого типа.
Субпопуляцию растений, имеющих размер, схожий с размером контрольных растений дикого типа, обрабатывали 2,4-D с использованием 560 или 1120 г кэ 2,4-D. Все растения, содержащие AAD-12 (v1), являлись очевидно резистентными к обработке гербицидом по сравнению с соей дикого типа Maverick. У двух растений с AAD-12 (v1) наблюдали незначительный уровень повреждений (на 2 DAT), однако, повреждения являлись временными, и на 7 DAT повреждений не наблюдали. Контрольные растения дикого типа являлись тяжело поврежденными через 7-14 DAT при 560 г кэ/га 2,4-D и погибали при 1120 г кэ/га. Эти данные соответствуют тому факту, что AAD-12 (v1) может придавать высокую устойчивость (>2-кратной нормы внесения) восприимчивой сельскохозяйственной культуре, такой как соя. Затем отбирали образцы подвергнутых скринингу растений для молекулярных и биохимических анализов для подтверждения встраивания генов AAD-12 (v1), копийности и уровней экспрессии генов.
Молекулярные анализы - Соя: Сбор ткани, выделение ДНК и количественный анализ. Свежую ткань помещали в пробирки и лиофилизировали при 4°C в течение 2 дней. После полного высушивания ткани в пробирку помещали гранулу вольфрама (Valenite) и образцы подвергали сухому измельчению с использованием шаровой мельницы Kelco в течение 1 минуты. Затем осуществляли стандартное выделение ДНК с использованием DNeasy (Qiagen, DNeasy 69109). Затем аликвоту выделенной ДНК окрашивали Pico Green (Molecular Probes P7589) и анализировали с помощью флуориметра (BioTek) с известными стандартами для получения концентрация в нг/мкл.
Полимеразная цепная реакция: в качестве матрицы использовали всего 100 нг тотальной ДНК. Использовали 20 мМ каждого праймера с набором Takara Ex Taq PCR Polimerase (Minis TAKRR001A). Праймерами для PTU AAD-12 (v1) являлись (прямой ATAATGCCAGCCTGTTAAACGCC) (SEQ ID NO: 8) и (обратный CTCAAGCATATGAATGACCTCGA) (SEQ ID NO: 9). Реакцию ПЦР осуществляли в термоциклере 9700 Geneamp (Applied Biosystems), подвергая образцы воздействию 94°C в течение 3 минут и 35 циклам воздействия 94°C в течение 30 секунд, 63°C в течение 30 секунд, и 72°C в течение 1 минуты и 45 секунд, затем 72°C в течение 10 минут. Праймерами для ПЦР кодирующей области AAD-12 (v1) являлись (прямой ATGGCTCATGCTGCCCTCAGCC) (SEQ ID NO: 10) и (обратный CGGGCAGGCCTAACTCCACCAA) (SEQ ID NO: 11). Реакцию ПЦР осуществляли в термоциклере 9700 Geneamp (Applied Biosystems), подвергая образцы воздействию 94°C в течение 3 минут и 35 циклам воздействия 94°C в течение 30 секунд, 65°C в течение 30 секунд, и 72°C в течение 1 минуты и 45 секунд, затем 72°C в течение 10 минут. Продукты ПЦР анализировали с помощью электрофореза в 1% агарозном геле, окрашиваемом EtBr.
Анализ Саузерн-блоттинга: Осуществляли анализ Саузерн-блоттинга с использованием тотальной ДНК, полученной с помощью набора Qiagen DNeasy. Всего 10 мкг геномной ДНК подвергали расщеплению в течение ночи для получения данных о встраивании. После расщепления в течение ночи аликвоту ~100 нг анализировали с помощью 1%-геля, чтобы убедиться в полном расщеплении. После этой проверки образцы анализировали с помощью большого 0,85%-ного агарозного геля в течение ночи при 40 Вольтах. Затем гель подвергали денатурации в 0,2 M NaOH, 0,6 M NaCl в течение 30 минут. Затем гель подвергали нейтрализации в 0,5 M Трис-HCl, 1,5 M NaCl, pH 7,5, в течение 30 минут. Затем использовали устройство для блоттинга, содержащее 20-кратный SSC, для осуществления переноса геля на нейлоновую мембрану (Millipore INYC00010) под действием силы тяжести в течение ночи. После переноса в течение ночи мембрану подвергали воздействию УФ-излучения с помощью кросслинкера (Stratagene UV stratalinker 1800) при 1200×100 микроджоулях. Затем мембрану промывали в 0,1% SDS, 0,1 SSC в течение 45 минут. После промывания в течение 45 минут мембрану сушили в течение 3 часов при 80°C, а затем хранили при 4°C до гибридизации. Фрагмент матрицы для гибридизации получали с помощью указанной выше ПЦР кодирующей области с использованием плазмидной ДНК. Продукт подвергали электрофорезу в 1%-ном агарозном геле и вырезали, а затем экстрагировали из геля с использованием способа экстракции из геля Qiagen (28706). Затем мембрану подвергали прегибридизации на этапе 60°C в течение 1 часа в буфере Perfect Hyb (Sigma H7033). Способ Prime it RmT dCTP-labeling rxn (Stratagene 300392) использовали для получения зонда на основе p32 (Perkin Elmer). Зонд очищали с использованием колонок Probe Quant. G50 (Amersham 27-5335-01). Два миллиона CPM использовали для гибридизации Саузерн-блотов в течение ночи. После гибридизации в течение ночи блоты подвергали двум промываниям по 20 минут при 65°C в 0,1% SDS, 0,1% SSC. Затем блоты экспонировали с пленкой в течение ночи, инкубируя при -80°C.
Биохимические анализы - Соя: Сбор образцов ткани и выделение белка AAD-12 (v1) из листьев сои. Приблизительно 50 до 100 мг ткани листьев собирали из листьев N-2, подвергнутых нанесению 2,4-D, но после 1 DAT. Концевой листочек N-2 удаляли, разрезали на небольшие фрагменты или 2 диска из листьев из отдельного локуса (~0,5 см в диаметре) и незамедлительно замораживали на сухом льду. Таким образом, осуществляли анализ белка (ELISA и анализ вестерн-блоттинга).
Оценка потомства T1: Растениям T0 позволяли самоопыляться для получения семейств T1. Тестирование потомства (анализ расщепления) осуществляли с использованием глуфосината в концентрации 560 г активного ингредиента/га в качестве селективного средства, наносимого в фазу роста V1-V2. Затем выжившие растения анализировали на предмет устойчивости к 2,4-D в одну или несколько фаз роста V2-V6. Семена получали посредством самоопыления, чтобы сделать возможным более широкое тестирование гербицидов на трансгенной сое.
Трансгенные по AAD-12 (v1) растения сои Maverick получали с помощью системы опосредованной Agrobacterium трансформации. Полученные растения T0 являлись устойчивыми к нанесению 2-кратной нормы внесения 2,4-D и давали фертильные семена. Частота фертильных трансгенных растений сои составляла до 5,9%. Встраивание гена AAD1-12 (v1) в геном сои подтверждали анализом Саузерн-блоттинга. Этот анализ показал, что большинство трансгенных растений содержало низкое количество копий. Растения, подвергнутые скринингу с использованием антител против AAD-12 (v1), демонстрировали положительный результат при ELISA и соответствующую полосу при анализе Вестерн блоттинга.
Способ трансформации 2 - Aerosol-Beam опосредованная трансформация эмбриогенной ткани каллюса сои: Культивирование эмбриогенной ткани каллюса соя и последующее beaming можно осуществлять, как описано в патенте США № 6809232 (Held et al.), для получения трансформантов с использованием конструкций, представленных в настоящем описании.
Способ трансформации 3 - Биолистическая бомбардировка сои: Этот способ можно осуществлять с использованием зрелой меристемы рубчика, полученной из семян (McCabe et al. (1988)). После осуществления способов биолистической бомбардировки можно ожидать восстановления трансформированных растений сои.
Способ трансформации 4 - Вискер-опосредованная трансформация: Получение вискеров и трансформацию с использованием вискеров можно осуществлять способами, описываемыми ранее Terakawa et al. (2005)). После осуществления способов биолистической бомбардировки можно ожидать восстановления трансформированных растений сои.
Семена Maverick поверхностно стерилизовали в 70% этаноле в течение 1 мин с последующим погружением в 1% гипохлорит натрия на 20 минут, а затем промыванием три раза в стерильной дистиллированной воде. Семена замачивали в дистиллированной воде на 18-20 часов. Из семян вырезали эмбриональные рубчики и обнажали апикальную меристему, удаляя первичные листья. Эмбриональные рубчики апикальной областью вверх помещали в среду для бомбардировки [BM: MS (Murashige and Skoog 1962) базовая солевая среда, 3% сахарозы и 0,8% фитагеля Sigma, pH 5,7] в 5-см чашки для культивирования, содержащие 12 мл среды для культивирования.
Способ трансформации 5 - Трансформацию эмбриогенной ткани каллюса, опосредованную бомбардировкой микрочастицами, можно оптимизировать описанными ранее способами (Khalafalla et al., 2005; El-Shemy et al., 2004, 2006).
ПРИМЕР 8
AAD-12 (v1) в хлопке
Способ трансформации хлопка: Семена хлопка (генотип Co310) поверхностно стерилизовали в 95% этаноле в течение 1 минуты, промывали, стерилизовали 50% коммерческим отбеливателем в течение двадцати минут, а затем промывали 3 раза стерильной дистиллированной водой перед проращиванием на среде G-media (таблица 21) в сосудах Magenta GA-7 и хранением при высокой интенсивности освещения 40-60 μΕ/m2 с фотопериодом, установленным на 16 часов освещения и 8 часов темноты, при 28°C.
Квадратные сегменты семядоли (~5 мм) получали из проростков возрастом 7-10 дней и помещали в жидкие среды M (таблица 21) в чашках Петри (Nunc, кат. №0875728). Вырезанные сегменты обрабатывали раствором, содержащим Agrobacterium, (в течение 30 минут), затем переносили на полутвердую среду M (таблица 21) и подвергали совместному культивированию в течение 2-3 дней. После совместного культивирования сегменты переносили на среду MG (таблица 21). Карбенициллин является антибиотиком, используемым для уничтожения Agrobacterium, а глуфосинат-аммоний является селективным средством, позволяющим расти только клеткам, содержащим перенесенный ген.
Получение Agrobacterium: 35 мл среды Y (таблица 21) (содержащей стрептомицин (стоковый раствор 100 мг/мл) и эритромицин (стоковый раствор 100 мг/мл)) инокулировали с использованием одной петли бактерий для выращивания в течение ночи в темноте при 28°C при встряхивании при 150 об./мин. На следующий день содержащий Agrobacterium раствор выливали в стерильную пробирку Oak Ridge (Nalge-Nunc, 3139-0050) и центрифугировали в Beckman J2-21 при 8000 об./мин. в течение 5 минут. Выливали супернатант, ресуспендировали осадок в 25 мл жидкой среды M (таблица 21) и центрифугировали на вортексе. Помещали аликвоту в стеклянную культуральную пробирку (Fisher, 14-961-27) для считывания с использованием Klett (Klett-Summerson, модель 800-3). Разбавляли новую суспензию с использованием жидких сред M для считывания с помощью колориметра Klett при 10 колониеобразующих единицах на мл с общим объемом 40 мл.
Через три недели из сегментов семядоли выделяли каллюс и переносили на свежую среду MG. Каллюс переносили на среду MG еще на 3 недели. При непосредственном сравнении среду MG можно дополнять дихлорпропом (добавляемым в среды в концентрациях 0,01 и 0,05 мг/л) для дополнения деградации 2,4-D, т.к. дихлорпроп не является субстратом фермента AAD-12, однако дихлорпроп более активен в отношении хлопка, чем 2,4-D. При раздельном сравнении сегменты помещали на среду MG, не содержащую регуляторов роста по сравнению со стандартной средой MG, они демонстрировали сниженное каллюсообразование, но каллюс все равно рос. Затем каллюс переносили на среду CG (таблица 21) и через три недели снова переносили на свежую селекционную среду. Еще через три недели ткань каллюс переносили на среду D (таблица 21) без регуляторов роста растений для индукции эмбриогенного каллюса. Через 4-8 недель на этой среде образовывался эмбриогенный каллюс, и его можно отличать от неэмбриогенного каллюса по его желтовато-белому цвету и гранулярным клеткам. Вскоре после этого начиналась регенерация эмбрионов, и они становились различимо зелеными по цвету. Регенерация и образование эмбрионов хлопка может занять время, одним из способов ускорения этого процесса является подвергание ткани стрессу. Общепринятым способом для осуществления этого является дессикация посредством изменений в микроокружении ткани и чашки с использованием меньшего количества сред для культивирования и/или различных способов заключения чашки (заматывание лентой по сравнению с парафильмом).
Среды для трансформации хлопка
(1 мМ)
(1 мМ)
Выделяли хорошо развившиеся эмбрионы большего размера и переносили их на среды DK (таблица 21) для развития эмбрионов. Через 3 недели (или когда эмбрионы разовьются) проросшие эмбрионы переносили на свежие среды для проращивания побега и корня. Через 4-8 недель любые хорошо развившиеся растения переносили в почву и выращивали до созревания. Через несколько месяцев растения вырастали до такой степени, когда их можно опрыскивать для определения того, имеют ли они резистентность к 2,4-D.
Трансформация клеток: Начинали несколько экспериментов, в которых сегменты семядоли обрабатывали Agrobacterium, содержащими pDAB724. Более 2000 полученных сегментов обрабатывали с использованием различных вариантов ауксинов для пролиферации каллюса хлопка с pDAB724: 0,1 или 0,5 мг/л R-дихлорпропа, 2,4-D в стандартной концентрации и без обработки ауксинами. Каллюс подвергали селекции с глуфосинатом аммония по причине включения гена PAT в конструкцию. Анализ линии каллюса в форме ПЦР и Invader будут использовать для определения и проверки наличия гена на стадии каллюса; затем линии каллюса, являющиеся эмбриогенными, будут отправлять на анализ вестерн-блоттинга. Эмбриогенный каллюс хлопка подвергали стрессу с использованием способов дессикации для улучшения качества и количества выделенной ткани. Почти 200 объектов каллюса подвергали скринингу на интактную PTU и экспрессию с использованием анализа вестерн-блоттинга на ген AAD-12 (v1).
Регенерация растений: Линии хлопка с AAD-12 (v1), с помощью которых получали растения согласно указанному выше способу, будут отправлять в теплицу. Для демонстрации того, что ген AAD-12 (v1) обеспечивает резистентность хлопка к 2,4-D, растения хлопка с AAD-12 (v1) и растения хлопка дикого типа опрыскивали с использованием автоматического устройства для опрыскивания, выпускающего 560 г кэ/га 2,4-D при объеме раствора для опрыскивания 187 л/га. Растения будут оценивать через 3 и 14 дней после обработки. Растения, выжившие при селективной норме внесения 2,4-D, будут подвергать самоопылению для получения семян T1 или ауткроссингу с элитной линией хлопка для получения семян F1. Полученные затем семена будут выращивать и оценивать на резистентность к гербицидам, как описано выше. Объекты с AAD-12 (v1) можно комбинировать с другими желаемыми признаками HT или IR.
ПРИМЕР 9
Трансформация других сельскохозяйственных культур с использованием Agrobacterium
В свете настоящего описания по настоящему изобретению можно трансформировать дополнительные сельскохозяйственные культуры с использованием известных в этой области способов. Опосредованную Agrobacterium трансформацию ржи см., например, в Popelka and Altpeter (2003). Опосредованную Agrobacterium трансформацию сои см., например, в Hinchee et al., 1988. Опосредованную Agrobacterium трансформацию сорго см., например, в Zhao et al., 2000. Опосредованную Agrobacterium трансформацию ячменя см., например, в Tingay et al., 1997. Опосредованную Agrobacterium трансформацию пшеницы см., например, в Cheng et al., 1997. Опосредованную Agrobacterium трансформацию риса см., например, в Hiei et al., 1997.
Латинские названия этих и других растений представлены ниже. Следует понимать, что эти и другие (без Agrobacterium) способы трансформации можно использовать для трансформации с использованием AAD-12 (v1), например, этих и других растений, включая, в качестве неограничивающих примеров, кукурузу (Zea mays), пшеницу (Triticum spp.), рис (Oryza spp. и Zizania spp.), ячмень (Hordeum spp.), хлопок (Abroma augusta и Gossypium spp.), сою (Glycine max), сахарную и столовую свеклу (Beta spp.), сахарный тростник (Arenga pinnata), помидор (Lycopersicon esculentum и другие spp., Physalis ixocarpa, Solanum incanum и другие spp., и Cyphomandra betacea), картофель (Solanum tubersoum), батат (Ipomoea betatas), рожь (Secale spp.), перец (Capsicum annuum, sinense и frutescens), латук (Lactuca sativa, perennis и pulchella), капусту (Brassica spp), сельдерей (Apium graveolens), баклажан (Solanum melongena), арахис (Arachis hypogea), сорго (все виды сорго), люцерну (Medicago sativua), морковь (Daucus carota), фасоль (Phaseolus spp. и другие роды), овес (Avena sativa и strigosa), горох (Pisum, Vigna и Tetragonolobus spp.), подсолнечник (Helianthus annuus), тыкву гигантскую (Cucurbita spp.), огурец (Cucumis sativa), табак (Nicotiana spp.), арабидопсис (Arabidopsis thaliana), газонную траву (Lolium, Agrostis, Poa, Cynadon и другие роды), клевер (Tifolium), горошек (Vicia). Такие растения, например, с генами AAD-12 (v1), включены в настоящее изобретение.
AAD-12 (v1) обладает потенциалом для повышения применимости ключевых ауксиновых гербицидов для сезонного использования во многих лиственных и хвойных системах лесоводства. Виды деревьев, резистентные к триклопиру, 2,4-D и/или флуроксипиру будут повышать гибкость чрезмерного использования этих гербицидов без опасений, связанных с повреждениями. Эти виды будут включать, в качестве неограничивающих примеров: ольху (Alnus spp.), ясень (Fraxinus spp.), виды осины и тополя (Populus spp.), бук (Fagus spp.), березу (Betula spp.), вишню (Prunus spp.), эвкалипт (Eucalyptus spp.), гикори (Carya spp.), клен (Acer spp.), дуб (Quercus spp) и сосну (Pinus spp). Использование резистентности к ауксинам для селективного контроля сорняков у декоративных и плодоносящих видов также входит в объем настоящего изобретения. Примеры могут включать, в качестве неограничивающих примеров, розу (Rosa spp.), бересклет (Euonymus spp.), петунию (Petunia spp), бегонию (Begonia spp.), рододендрон (Rhododendron spp), яблоню лесную или яблоню (Malus spp.), грушу (Pyrus spp.), персик (Prunus spp) и бархатцы (Tagetes spp.).
ПРИМЕР 10
Дополнительное доказательство неожиданных результатов: AAD-12 по сравнению с AAD-2
Исходное клонирование AAD-2 (v1): Другой ген из базы данных NCBI (см. веб-сайт ncbi.nlm.nih.gov; рег. №AP005940) идентифицировали в качестве гомолога с идентичностью по отношению к tfdA всего 44% аминокислот. Для удобства этот ген в настоящем описании обозначают как AAD-2 (v1). Процент идентичности определяли, сначала преобразуя последовательности ДНК AAD-2 и tfdA (SEQ ID NO: 12 из PCT/US2005/014737 и регистрационный номер GenBank M16730, соответственно) в белки (SEQ ID NO: 13 из PCT/US2005/014737 и регистрационный номер GenBank M16730, соответственно), затем используя ClustalW в пакете программ VectorNTI для осуществления множественного выравнивания последовательностей.
Из Northern Regional Research Laboratory получали штамм Bradyrhizobium japonicum, содержащий ген AAD-2 (v1) (NRRL, штамм №B4450). Лиофилизированный штамм восстанавливали согласно протоколу NRRL и хранили при -80°C в 20% глицерине для внутреннего использования в качестве бактериального штамма Dow DB 663. Затем чашку с триптическим соевым агаром засевали петлей клеток из этого замороженного стока для выделения и инкубировали при 28°C в течение 3 дней. Отдельную колонию использовали для инокуляции 100 мл триптического соевого бульона в колбе с отбойниками емкостью 500 мл, который инкубировали в течение ночи при 28°C в напольной качалке при 150 об./мин. Из него выделяли тотальную ДНК с использованием грамотрицательного протокола для набора Qiagen's DNeasy (Qiagen, кат. №69504). Для амплификации гена-мишени из геномной ДНК конструировали следующие праймеры, прямой: 5'-ACTAGTAACAAAGAAGGAGATATACCATGACGAT-3' [(brjap 5'(speI) SEQ ID NO: 14 из PCT/US2005/014737 (с добавленным участком рестрикции Spe I и участком связывания рибосомы (RBS))], и обратный: 5'-TTCTCGAGCTATCACTCCGCCGCCTGCTGCTGC-3' [(brjap 3' (xhoI) SEQ ID NO: 15 из PCT/US2005/014737 (с добавленным участком рестрикции XhoI)].
Реакционные смеси объемом пятьдесят микролитров составляли следующим образом: буфер Fail Safe Buffer - 25 мкл, ea. праймер - 1 мкл (50 нг/мкл), геномная ДНК - 1 мкл (200 нг/мкл), H.sub.2O - 21 мкл, Taq-полимераза - 1 мкл (2,5 Ед./мкл). Три буфера Fail Safe Buffer-A, B и C использовали в трех отдельных реакциях. Затем осуществляли ПЦР в следующих условиях: цикл термической денатурации при 95°C в течение 3,0 минут; 95°C - 1,0 минута, 50°C - 1,0 минута, 72°C - 1,5 минуты в течение 30 циклов; с последующим конечным циклом при 72°C в течение 5 минут, с использованием системы FailSafe PCR System (Epicenter, кат. №F599100). Для подтверждения нуклеотидной последовательности полученный продукт ПЦР размером ~1 т.п.н. клонировали в pCR 2.1 (Invitrogen, кат. №K4550-40), следуя включенному протоколу, с использованием химически-компетентных TOP10F' E. coli в качестве штамма-акцептора.
Десятью из полученных белых колоний инокулировали 3 мкл бульона Луриа + 1000 мкг/мл ампициллина (LB Amp) и выращивали в течение ночи при 37°C со встряхиванием. Из каждой культуры выделяли плазмиды, используя набор Nucleospin Plus Plasmide Miniprep Kit (BD Biosciences, кат. №K3063-2) и следуя включенному протоколу. Осуществляли рестрикцию выделенной ДНК для подтверждения наличия продукта ПЦР в векторе pCR2.1. Плазмидную ДНК расщепляли с использованием рестрикционного фермента EcoRI (New England Biolabs, кат. №R0101S). Секвенирование осуществляли с помощью набора Beckman CEQ Quick Start Kit (Beckman Coulter, кат. №608120) с использованием праймеров M13, прямого [5'-GTAAAACGACGGCCAG-3'] (SEQ ID NO: 6) и обратного [5'-CAGGAAACAGCTATGAC-3'] (SEQ ID NO: 7), по инструкциям производителя. Этой последовательности гена и соответствующему ей белку давали новое общее обозначение AAD-2 (v1) для внутренней согласованности.
Получение бинарного вектора AAD-2 (v1): Ген AAD-2 (v1) амплифицировали с помощью ПЦР из pDAB3202. При реакции ПЦР осуществляли изменения в праймерах для встраивания участков рестрикции AflIII и SacI в 5'-праймер и 3'-праймер, соответственно. См. PCT/US2005/014737. Праймеры "NcoI of Brady" [5'-TATACCACATGTCGATCGCCATCCGGCAGCTT-3'] (SEQ ID NO: 14) и "SacI of Brady" [5'-GAGCTCCTATCACTCCGCCGCCTGCTGCTGCAC-3'] (SEQ ID NO: 15) использовали для амплификации фрагментов ДНК с использованием системы Fail Safe PCR System (Epicentre). Продукт ПЦР клонировали в клонирующий вектор pCR2.1 TOPO TA (Invitrogen) и подтверждали последовательность с помощью прямого и обратного праймеров М13 с использованием реагентов для секвенирования Beckman Coulter "Dye Terminator Cycle Sequencing with Quick Start Kit". С помощью данных о последовательности идентифицировали клон с правильной последовательностью (pDAB716). Затем фрагмент гена AflIII/SacI AAD-2 (v1) клонировали в вектор NcoI/SacI pDAB726. Полученную конструкцию (pDAB717); промотор AtUbi10: Nt OSM 5'UTR: AAD-2 (v1): Nt OSM 3'UTR: ORF1 polyA 3'UTR подтверждали с помощью рестрикции (с использованием NcoI/SacI). Эту конструкцию клонировали в бинарный вектор pDAB3038 как фрагмент ДНК NotI-NotI. Полученную конструкцию (pDAB767); промотор AtUbi10: Nt OSM5'UTR: AAD-2 (v1): Nt OSM 3'UTR: ORF1 polyA 3'UTR: промотор CsVMV: PAT: ORF25/26 3'UTR подвергали рестрикции (с использованием Nod, EcoRI, HinDIII, NcoI, PvuII и SalI) для подтверждения правильной ориентации. Затем завершенную конструкцию (pDAB767) использовали для трансформации Agrobacterium.
Оценка трансформированного арабидопсиса: Свежесобранные семена T1, трансформированные с использованием оптимизированного для растений гена AAD-12 (v1) или нативного гена AAD-2 (v1), выращивали и подвергали селекции на резистентность к глуфосинату, как описано выше. Затем растениям случайным образом присваивали различные нормы внесения 2,4-D (50-3200 г кэ/га). Нанесение гербицида осуществляли с помощью автоматического устройства для опрыскивания при объеме раствора для опрыскивания187 л/га. Используемым 2,4-D являлся коммерческий состав диметиламиновой соли (456 г кэ/л, NuFarm, St Joseph, Mo.), смешанный в 200 мМ буфере Трис (pH 9,0) или 200 мМ буфере HEPES (pH7,5).
AAD-12 (v1) и AAD-2 (v1) обеспечивали детектируемую резистентность к 2,4-D по сравнению с трансформированными и нетрансформированными контрольными линиями; однако, отдельные конструкции являлись широко вариабельными по своей способности придавать резистентность к 2,4-D отдельным растениям арабидопсиса T1. Неожиданно, трансформанты с AAD-2 (v1) и AAD-2 (v2) являлись намного менее резистентными к 2,4-D, чем с геном AAD-12 (v1), по доле высоко устойчивых растений, а также общим средним повреждениям. Растения, трансформированные с использованием AAD-2 (v1), не выживали при 200 г кэ/га 2,4-D относительно неповрежденными (<20% видимых повреждений), и общие повреждения в популяции составляли приблизительно 83% (см. PCT/US2005/014737). В отличие от этого, AAD-12 (v1) имели средние повреждения в популяции приблизительно 6% при обработке 3200 г кэ/га 2,4-D. Устойчивость немного улучшалась в случае оптимизированного для растений AAD-2 (v2) по сравнению с нативным геном; однако сравнение обоих генов AAD-12 и AAD-2, оптимизированных для растений, указывает на значительное преимущество AAD-12 (v1) в полевых условиях.
Эти результаты являются неожиданными с учетом того, что при сравнении AAD-2 (v1) (см. PCT/US2005/014737) и AAD-12 (v2) in vitro выявили, что оба являлись высокоэффективными в деградации 2,4-D и оба имели специфичность S-типа в отношении хиральных арилоксиалканоатных субстратов. AAD-2 (v1) экспрессируется в отдельных растениях T1 на различных уровнях; однако этот экспрессирующийся белок придавал небольшую защиту от повреждений 2,4-D. Не наблюдали значимых различий в уровне экспрессии белка (в полевых условиях) для нативных и оптимизированных для растений генов AAD-2 (см. PCT/US2005/014737). Эти данные подтверждают полученные ранее результаты, которые делают неожиданной функциональную экспрессию AAD-12 (v1) в полевых условиях и являющуюся ее результатом резистентность к гербицидам 2,4-D и пиридилоксиацетатным гербицидам.
ПРИМЕР 11
Использование феноксиауксиновых гербицидов для посевов сои, хлопка и других двудольных сельскохозяйственных культур, трансформированных только с использованием AAD-12 (v1)
AAD-12 (v1) может сделать возможным использование феноксиауксиновых гербицидов (например, 2,4-D и MCPA) и пиридилоксиауксинов (триклопира и флуроксипира) для контроля широкого спектра широколиственных сорняков непосредственно в случае сельскохозяйственных культур, в норме восприимчивых к 2,4-D. С помощью нанесения 2,4-D в количестве от 280 до 2240 г кэ/га можно контролировать большинство видов широколиственных сорняков, присутствующих в агрономических условиях. Чаще используют 560-1120 г кэ/га. В случае триклопира нормы внесения, как правило, будут находиться в диапазоне 70-1120 г кэ/га, чаще - 140-420 г кэ/га. В случае флуроксипира нормы внесения, как правило, будут находиться в диапазоне 35-560 г кэ/га, чаще - 70-280 кэ/га.
Преимуществом этого дополнительного инструмента является крайне низкая стоимость гербицидного компонента для широколиственных растений и потенциальный кратковременный последействующий контроль сорняков, обеспечиваемый более высокими количествами 2,4-D, триклопира и флуроксипира при их использовании, в то время как непоследействующий гербицид, такой как глифосат, не будет обеспечивать контроль проросших в дальнейшем сорняков. Этот инструмент также обеспечивает механизм комбинирования механизмов действия гербицида с удобством HTC в качестве интегрированной стратегии резистентности к гербицидам и регуляции сдвигов в популяциях сорняков.
Дополнительным преимуществом, обеспечиваемым этим инструментом, является возможность получения баковой смеси гербицидов для контроля широкого спектра широколиственных сорняков (например, 2,4-D, триклопира и флуроксипира) с общеупотребительными последействующими гербицидами для контроля сорняков. Эти гербициды, как правило, наносят до посадки или во время нее, но часто они менее эффективны в отношении проросших, укоренившихся сорняков, которые могут существовать на поле до посадки. Повышая применимость этих арилоксиауксиновых гербицидов до включения посадочных, предвсходных или предпосевных обработок, увеличивают гибкость программ последействующего контроля сорняков. Специалисту в этой области будет понятно, что программы с использованием последействующих гербицидов будут отличаться с учетом интересующей сельскохозяйственной культуры, но типичные программы будут включать гербициды из групп хлорацетамидных и динитроанилиновых гербицидов, а также гербициды, такие как кломазон, сульфентразон и множество ALS-ингибирующих, PPO-ингибирующих и HPPD-ингибирующих гербицидов.
Дополнительные преимущества могут включать устойчивость к 2,4-D, триклопиру или флуроксипиру, необходимую до посадки после нанесения ауксиновых гербицидов на основе арилоксиуксусной кислоты (см. пример выше); и меньшее количество проблем с контаминационными повреждениями двудольных сельскохозяйственных культур, являющихся результатом неполной очистки наливных цистерн, содержавших 2,4-D, триклопир или флуроксипир. Дикамбу (и многие другие гербициды) все равно можно использовать для последующего контроля AAD-12 (v1)-трансформированных двудольных самосевных сельскохозяйственных культур.
Специалистам в этой области также будет понятно, что указанный выше пример можно применить к любой восприимчивой к 2,4-D (или другому арилоксиауксиновому гербициду) сельскохозяйственной культуре, которая будет защищена с помощью гена AAD-12 (v1) в случае стабильной трансформации. Поэтому специалисту в области контроля сорняков будет понятно, что использование различных коммерческих фенокси- или пиридилоксиауксиновых гербицидов в отдельности или в комбинации с гербицидом становится возможным благодаря трансформации с использованием AAD-12 (v1). Конкретные количества других гербицидов, характерные для этих химических веществ, можно определять с помощью маркировок гербицидов, собранных в книге CPR (Crop Protection Reference), или аналогичном сборнике или любых коммерческих или академических источниках по защите сельскохозяйственных культур, таких как Crop Protection Guide от Agriliance (2005). Каждый альтернативный гербицид, который можно использовать в HTC благодаря AAD-12 (v1), используемый в отдельности, в баковой смеси или последовательно, считают входящим в объем настоящего изобретения.
ПРИМЕР 12
Использование феноксиауксиновых и пиридилоксиауксиновых гербицидов для посевов кукурузы, риса и других однодольных видов, трансформированных только с использованием AAD-12 (v1)
Аналогично, трансформация видов трав (в качестве неограничивающих примеров, таких как, кукуруза, рис, пшеница, ячмень или газонная трава и пастбищные злаки) с использованием AAD-12 (v1) будет делать возможным использование высокоэффективных фенокси- и пиридилоксиауксинов в случае сельскохозяйственных культур, где в норме селективность является неопределенной. Большинство видов трав имеют природную устойчивость к ауксиновым гербицидам, таким как феноксиауксины (т.е. 2,4-D). Однако относительно низкий уровень селективности сельскохозяйственных культур приводит к сниженной применимости в случае этих сельскохозяйственных культур по причине укороченного временного окна нанесения или неприемлемого риска повреждения. Таким образом, однодольные сельскохозяйственные культуры, трансформированные с использованием AAD-12 (v1), будут делать возможным использование аналогичной комбинации обработок, описываемой для двудольных сельскохозяйственных культур, таких как нанесение 2,4-D при норме внесения от 280 до 2240 г кэ/га для контроля большинства видов широколиственных сорняков. Чаще используют 560-1120 г кэ/га. В случае триклопира нормы внесения, как правило, будут находиться в диапазоне 70-1120 г кэ/га, чаще - 140-420 г кэ/га. В случае флуроксипира нормы внесения, как правило, будут находиться в диапазоне 35-560 г кэ/га, чаще - 70-280 кэ/га.
Преимуществом этого дополнительного инструмента является крайне низкая стоимость гербицидного компонента для широколиственных растений и потенциальный кратковременный последействующий контроль сорняков, обеспечиваемый более высокими количествами 2,4-D, триклопира или флуроксипира. В отличие от этого, непоследействующий гербицид, такой как глифосат, не будет обеспечивать контроль проросших в дальнейшем сорняков. Этот инструмент также будет обеспечивать механизм чередования механизмов действия гербицидов с удобством HTC в качестве интегрированной стратегии резистентности к гербицидам и регуляции сдвигов в популяциях сорняков в стратегии комбинирования устойчивых к глифосату сельскохозяйственных культур/HTC AAD-12 (v1) при чередовании видов сельскохозяйственных культур или его отсутствии.
Дополнительным преимуществом, обеспечиваемым этим инструментом, является возможность получения баковой смеси гербицидов для контроля широкого спектра широколиственных сорняков (например, 2,4-D, триклопира и флуроксипира) с общеупотребительными последействующими гербицидами для контроля сорняков. Эти гербициды, как правило, наносят до посадки или во время нее, но часто они менее эффективны в отношении проросших, укоренившихся сорняков, которые могут существовать на поле до посадки. Повышая применимость этих арилоксиауксиновых гербицидов до включения посадочных, предвсходных или предпосевных обработок, увеличивают гибкость программ последействующего контроля сорняков. Специалисту в этой области будет понятно, что программы с использованием последействующих гербицидов будут отличаться с учетом интересующей сельскохозяйственной культуры, но типичные программы будут включать гербициды из групп хлорацетамидных и динитроанилиновых гербицидов, а также гербициды, такие как кломазон, сульфентразон и множество ALS-ингибирующих, PPO-ингибирующих и HPPD-ингибирующих гербицидов.
Повышенная устойчивость кукурузы, риса и других однодольных растений к фенокси- или пиридилоксиауксинам должна позволить использовать эти гербициды для посевов без ограничений, связанных с фазами роста или потенциальным наклонением сельскохозяйственных культур, развития явлений, таких как "крысиный хвост", наклон сельскохозяйственных культур, индуцируемая регуляторами роста ломкость стебля у кукурузы или деформация опорных корней. Каждый альтернативный гербицид, который можно использовать в HTC благодаря AAD-12 (v1), используемый в отдельности, в баковой смеси или последовательно, считают входящим в объем настоящего изобретения.
ПРИМЕР 13
AAD-12 (v1) в рисе
Описание сред: Используемые среды для культивирования доводили до pH 5,8 с помощью 1 M KOH и отверждали с помощью 2,5 г/л фитагеля (Sigma). Эмбриогенные каллюсы культивировали в чашках Петри размером 100×20 мм, содержащих 40 мл полутвердой среды. Проростки риса выращивали на 50 мл среды в коробках Magenta. Клеточные суспензии поддерживали в конических колбах емкостью 125 мл, содержащих 35 мл жидкой среды, и вращали при 125 об./мин. Индукцию и поддержание эмбриогенных культур осуществляли в темноте при 25-26°C, и регенерацию растений и культивирование целых растений осуществляли при фотопериоде 16 часов (Zhang et al. 1996).
Индукцию и поддержание эмбриогенного каллюса осуществляли на минимальной среде NB, описанной ранее (Li et al. 1993), но адаптированной для содержания 500 мг/л глутамина. Суспензионные культуры инициировали и поддерживали в жидкой среде SZ (Zhang et al. 1998) с включением 30 г/л сахарозы вместо мальтозы. Осмотическая среда (NBO) состояла из среды NB с добавлением 0,256 M маннита и сорбита. Каллюс, резистентный к гигромицину B, подвергали селекции на среде NB, дополненной 50 мг/л гигромицина B, в течение 3-4 недель. Предварительную регенерацию осуществляли на среде (PRH50), состоящей из среды NB без 2,4-дихлорфеноксиуксусной кислоты (2,4-D), но с добавлением 2 мг/л 6-бензиламинопурина (BAP), 1 мг/л α-нафтилуксусной кислоты (NAA), 5 мг/л абсцизовой кислоты (ABA) и 50 мг/л гигромицина B, в течение 1 недели. Затем осуществляли регенерацию проростков посредством культивирования на среде для регенерации (RNH50), содержащей среду NB без 2,4-D, и дополненную 3 мг/л BAP, 0,5 мг/л NAA и 50 мг/л гигромицина B, до регенерации побегов. Побеги переносили на среду для укоренения с половинной концентрацией базовых солей Мурашиге и Скуга и витаминов B5 по Гамборгу, дополненную 1% сахарозой и 50 мг/л гигромицина B (1/2MSH50).
Получение культуры ткани: Зрелые обезвоженные семена Oryza sativa L. japonica cv. Taipei 309 стерилизовали, как описано в Zhang et al. 1996. Индуцировали эмбриогенные ткани, культивируя стерильные зрелые семена риса на среде NB в темноте. Первичный каллюс диаметром приблизительно 1 мм отделяли от щитка и использовали для инициации клеточной суспензии в жидкой среде SZ. Затем суспензии поддерживали, как описано в Zhang 1995. Полученные с помощью суспензии эмбриогенные ткани удаляли из жидкой культуры через 3-5 дней после предшествующего субкультивирования, помещали на осмотическую среду NBO для получения круга диаметром приблизительно 2,5 см на чашке Петри и культивировали в течение 4 часов перед бомбардировкой. Через 16-20 часов после бомбардировки ткани переносили со среды NBO на селективную среду NBH50 с гигромицином B, проверяя, чтобы поверхность, подвергнутая бомбардировке, находилась сверху, и инкубировали в темноте в течение 14-17 дней. Затем вновь образовавшийся каллюс отделяли от исходных подвергнутых бомбардировке эксплантатов и помещали вблизи на ту же среду. Еще через 8-12 дней визуально определяли относительно плотный, непрозрачный каллюс и переносили его на среду для предварительной регенерации PRH50 на 7 дней в темноте. Затем растущий каллюс, ставший более плотным и непрозрачным, субкультивировали на среде для регенерации RNH50 в течение 14-21 дня с фотопериодом 16 часов. Регенерировавшие побеги переносили в коробки Magenta, содержащие среду 1/2 MSH50. Многочисленные растения, регенерировавшие из одного эксплантата, считали сибсами и рассматривали как одну независимую линию растений. Растение оценивали как положительное по гену hph, если оно давало толстые белые корни и активно росло на среде 1/2 MSH50. Когда проростки достигали верха коробок Magenta, их переносили в почву в 6-см горшок при 100% влажности на неделю, затем перемещали в вегетационную камеру с периодом освещения 14 часов при 30°C и периодом темноты при 21°C на 2-3 недели до пересаживания в 13-см горшки в теплицы. Семена собирали и высушивали при 37°C в течение одной недели перед хранением.
Бомбардировка микрочастицами: Все бомбардировки осуществляли с использованием системы Biolistic PDS-1000/He™ (Bio-Rad, Laboratories, Inc.). Три миллиграмма частиц золота диаметром 1,0 микрометр промывали однократно 100% этанолом, дважды - стерильной дистиллированной водой и ресуспендировали в 50 мкл воды в силиконизированной пробирке Eppendorf. Пять микрограммов плазмидной ДНК, представляющей pDOW3303 (Hpt-содержащий вектор) и pDAB4101 (AAD-12 (v1)+AHAS) в молярном соотношении 1:6, 20 мкл спермидина (0,1 M) и 50 мкл хлорида кальция (2,5 M) добавляли к суспензии золота. Смесь инкубировали при комнатной температуре в течение 10 мин, центрифугировали при 10000 об./мин. в течение 10 сек, ресуспендировали в 60 мкл холодного 100% этанола и распределяли по 8-9 мкл на каждый макроноситель. Образцы ткани бомбардировали при 1100 фунт/дюйм2 и вакууме 27 дюймов ртутного столба, как описано в Zhang et al. (1996).
Послевсходовая устойчивость к гербицидам у риса T0, трансформированного с использованием AAD-12 (v1): Проростки риса в фазе 3-5 листьев опрыскивали летальной дозой 0,16% (об./об.) раствора Pursuit (для подтверждения наличия гена AHAS), содержащего 1% Sunit II (об./об.) и 1,25% UAN (об./об.), с использованием автоматического устройства для опрыскивания, откалиброванного на 187 л/га. Оценку восприимчивости или резистентности осуществляли через 36 дней после обработки (DAT). Десять из 33 объектов, отправленных в теплицу, являлись стабильно устойчивыми к Pursuit; другие демонстрировали различные уровни повреждения гербицидами. Отбирали растения и осуществляли молекулярный анализ, при котором определяли семь из этих 10 объектов как содержащие и PTU AAD-12 (v1), и полную кодирующую область AHAS.
Наследование AAD-12 (v1) у риса T1: Тестирование 100 растений-потомков осуществляли на пяти линиях T1 из линий с AAD-12 (v1), содержащих и PTU AAD-12 (v1), и кодирующую область AHAS. Семена высаживали с учетом указанного выше способа и опрыскивали 140 г кэ/га имазетапира с использованием автоматического устройства для опрыскивания, как описано выше. Через 14 DAT подсчитывали резистентные и восприимчивые растения. Две из пяти тестируемых линий расщеплялись в соответствии с единым локусом, доминантным признаком с менделевским наследованием (3R: 1S), как определяли с помощью анализа с использованием хи-квадрата. AAD-12 наследовался совместно с селективным маркером AHAS, как определяли с помощью указанного ниже тестирования на устойчивость к 2,4-D.
Подтверждение высокой устойчивости к 2,4-D у риса T1: Следующие линии T1, расщепляющиеся по AAD-12 (v1), как единому локусу, высаживали в 3-дюймовые горшки, содержащие среды Metro Mix: pDAB4101(20)003 и pDAB4101(27)002. В фазе 2-3 листьев их опрыскивали 140 г кэ/га имазетапира. Удаляли растения, не содержащие вставку, и отдельные растения опрыскивали в фазе V3-V4 в автоматическом устройстве для опрыскивания, установленном на 187 л/га при 1120, 2240 или 4480 г кэ/га 2,4-D DMA (2-кратная, 4-кратная и 8-кратная типичные нормы внесения для коммерческого использования, соответственно). Растения классифицировали через 7 и 14 DAT и сравнивали с нетрансформированным коммерческим культиваром риса "Lamont" в качестве растений отрицательного контроля.
Ответ растений T1 с AAD-12 (v1) и нетрансформированного контроля на различные уровни 2,4-D DMA: средний % повреждения через 14 DAT
(20)003
(27)002
Данные о повреждениях (таблица 22) свидетельствуют о том, что линии, трансформированные с использованием AAD-12 (v1), являются более устойчивыми к большим количествам 2,4-D DMA, чем нетрансформированные контроли. Линия pDAB4101(20)003 являлась более устойчивой к высоким уровням 2,4-D, чем линия pDAB4101(27)002. Данные также свидетельствуют о том, что устойчивость к 2,4-D является стабильной в течение по меньшей мере двух поколений.
Сбор ткани, выделение ДНК и количественный анализ: Свежую ткань помещали в пробирки и лиофилизировали при 4°C в течение 2 дней. После полного высушивания ткани в пробирку помещали гранулу вольфрама (Valenite) и образцы подвергали сухому измельчению с использованием шаровой мельницы Kelco в течение 1 минуты. Затем осуществляли стандартное выделение ДНК с использованием DNeasy (Qiagen, Dneasy 69109). Затем аликвоту выделенной ДНК окрашивали Pico Green (Molecular Probes P7589) и сканировали на флуориметре (BioTek) с известными стандартами для получения концентрации в нг/мкл.
Экспрессия AAD-12 (v1): Все 33 линии трансгенного риса T0 и 1 нетрансгенный контроль анализировали на экспрессию AAD-12 с использованием ELISA-блот. AAD-12 определяли в клонах 20 линий, но не в линии контрольных растений Taipei 309. Двенадцать из 20 линий, содержащих некоторое количество клонов, устойчивых к имазетапиру, экспрессировали белок AAD-12, являлись положительным при ПЦР на PTU AAD-12 и положительными на кодирующую область AHAS. Уровни экспрессии находились в диапазоне от 2,3 до 1092,4 м.д. общего растворимого белка.
Устойчивость растений риса с pDAB4101 к гербицидам 2,4-D и триклопиру в полевых условиях: Полевое испытание уровня устойчивости осуществляли с использованием объекта с AAD-12 (v1) pDAB4101[20] и одного риса дикого типа (Clearfield 131) (нетрансгенного резистентного к имидазолинону сорта) в Wayside, Miss. Дизайн эксперимента представлял собой рандомизированный полноблочный план с одним повторением. Варианты эксперимента представляли собой 2-кратные нормы внесения 2,4-D (диметиламиновой соли) при 2240 г кэ/га и триклопира при 560 г кэ/га вместе с необработанным контролем. В рамках каждого варианта эксперимента два ряда pDAB4101[20] поколения T1 и два ряда риса Clearfield высаживали с использованием небольшой сеялки с расстоянием между рядами 8 дюймов. Рис pDAB4101[20] содержал ген AHAS в качестве селективного маркера для гена AAD-12 (v1). Имазетапир наносили в фазу одного листа в качестве селективного средства для удаления с опытных участков каких-либо растений без AAD-12 (v1). Когда рис достигал фазы 2 листьев, производили обработки гербицидами с использованием пневматического ранцевого опрыскивателя, выпускающего объем раствора для опрыскивания 187 л/га под давлением 130-200 кПа. Визуальную оценку повреждений проводили через 7, 14 и 21 дней после обработки.
Ответы объекта с AAD-12 (v1) на 2,4-D и триклопир представлены в таблице 23. Нетрансформированная линия риса (Clearfield) являлась тяжело поврежденной (30% через 7 DAT и 35% через 15 DAT) 2,4-D при 2240 г кэ/га, что считают 4-кратной нормой внесения для коммерческого использования. Объект с AAD-12 (v1) демонстрировал исключительную устойчивость к 2,4-D без видимых повреждений через 7 или 15 DAT. Нетрансформированный рис являлся тяжело поврежденным (15% через 7 DAT и 25% через 15 DAT) 2-кратной нормой внесения триклопира (560 г кэ/га). Объект с AAD-12 (v1) демонстрировал исключительную устойчивость к 2-кратным нормам внесения триклопира без наблюдаемых повреждений через 7 или 15 DAT.
Эти результаты свидетельствуют о том, что трансформированный рис с AAD-12 (v1) демонстрировал высокий уровень резистентности к 2,4-D и триклопиру при нормах внесения, вызывающих тяжелые видимые повреждения у риса Clearfield. Они также свидетельствуют о возможности стэкинга многочисленных генов устойчивости к гербицидам с AAD-12 I у множества видов для обеспечения резистентности к более широкому спектру эффективных химических веществ.
Ответ растений риса с AAD-12 поколения T1 на 2,4-D и триклопир в полевых условиях
гербицидами
ПРИМЕР 14
AAD-12 (v1) в каноле
Трансформация канолы: Ген AAD-12 (v1), придающий резистентность к 2,4-D, использовали для трансформации Brassica napus var. Nexera*710 с помощью опосредованной Agrobacterium трансформации и плазмиды pDAB3759. Конструкция содержала ген AAD-12 (v1), запускаемый промотором CsVMV, и ген Pat, запускаемый промотором AtUbi10, и признак резистентности к глифосату EPSPS, запускаемый промотором AtUbi10.
Семена поверхностно стерилизовали 10% коммерческим отбеливателем в течение 10 минут и промывали 3 раза стерильной дистиллированной водой. Затем семена помещали на минимальную среду MS с половинной концентрацией (Murashige and Skoog, 1962) и поддерживали в режиме роста при 25°C и фотопериоде 16 часов освещения /8 часов темноты.
Сегменты гипокотиля (3-5 мм) вырезали из 5-7-дневных проростков и помещали на среду для индукции каллюса K1D1 (среда MS с 1 мг/л кинетина и 1 мг/л 2,4-D) на 3 дня в качестве предварительной обработки. Затем сегменты переносили на чашку Петри, обрабатывали штаммом Agrobacterium Z707S или LBA4404, содержащим pDAB3759. Agrobacterium выращивали в течение ночи при 28°C в темноте на шейкере при 150 об./мин., затем ресуспендировали в среде для культивирования.
Через 30 мин обработки сегментов гипокотиля Agrobacterium, их помещали обратно на среду для индукции каллюса на 3 дня. После совместного культивирования сегменты помещали на K1D1TC (среду для индукции каллюса, содержащую 250 мг/л карбенициллина и 300 мг/л тиментина) на одну или две недели восстановления. Альтернативно, сегменты помещали непосредственно на селективную среду K1D1H1 (указанную выше среду с 1 мг/л Herbiace). Карбенициллин и тиментин являлись антибиотиками, используемыми для уничтожения Agrobacterium. Селективное средство Herbiace позволяет расти трансформированным клеткам.
Затем привитые сегменты гипокотиля помещали на среду для регенерации побегов B3Z1H1 (среда MS, 3 мг/л бензиламинопурина, 1 мг/л зеатина, 0,5 г/л MES [2-(N-морфолино)этансульфоновой кислоты], 5 мг/л нитрата серебра, 1 мг/л Herbiace, карбенициллина и тиментина). Через 2-3 недели побеги начинали регенерировать. Сегменты гипокотиля вместе с побегами переносили на среду B3Z1H3 (среда MS, 3 мг/л бензиламинопурина, 1 мг/л зеатина, 0,5 г/л MES [2-(N-морфолино)этансульфоновой кислоты], 5 мг/л нитрата серебра, 3 мг/л Herbiace, карбенициллина и тиментина) еще на 2-3 недели.
Побеги отрезали от сегментов гипокотиля и переносили на среду для удлинения побегов MESH5 или MES 10 (MS, 0,5 г/л MES, 5 или 10 мг/л Herbiace, карбенициллина, тиментина) на 2-4 недели. Удлинившиеся побеги культивировали для индукции корней на MSI.1 (MS с 0,1 мг/л индолилмасляной кислоты). Когда у растений появлялась хорошо развитая корневая система, их переносили в почву. Растения подвергали акклиматизации в контролируемых условиях окружающей среды в Conviron в течение 1-2 недели перед переносом в теплицу.
Молекулярный анализ - Канола, материалы и способы: Сбор ткани, выделение ДНК и количественный анализ. Свежую ткань помещали в пробирки и лиофилизировали при 4°C в течение 2 дней. После полного высушивания ткани в пробирку помещали гранулу вольфрама (Valenite) и образцы подвергали сухому измельчению с использованием шаровой мельницы Kelco в течение 1 минуты. Затем осуществляли стандартное выделение ДНК с использованием DNeasy (Qiagen, DNeasy 69109). Затем аликвоту выделенной ДНК окрашивали Pico Green (Molecular Probes P7589) и анализировали с помощью флуориметра (BioTek) с известными стандартами для получения концентрации в нг/мкл.
Полимеразная цепная реакция: В качестве матрицы использовали всего 100 нг тотальной ДНК. Использовали 20 мМ каждого праймера с набором Takara Ex Taq PCR Polimerase kit (Minis TAKRR001A). Праймерами для ПЦР кодирующей области AAD-12 (v1) являлись (SEQ ID NO: 10) (прямой) и (SEQ ID NO: 11) (обратный). Реакцию ПЦР осуществляли в термоциклере 9700 Geneamp (Applied Biosystems), подвергая образцы воздействию 94°C в течение 3 минут и 35 циклам 94°C в течение 30 секунд, 65°C в течение 30 секунд и 72°C в течение 2 минут, затем 72°C в течение 10 минут. Продукты ПЦР анализировали с помощью электрофореза в 1% агарозном геле, окрашиваемом EtBr. 35 образцов от 35 растений с AAD-12 (v1) при тестировании являлись положительными. Три образца отрицательного контроля при тестировании являлись отрицательными.
ELISA: Используя общепринятый ELISA, описываемый в предыдущем разделе, определяли белок AAD-12 в 5 различных трансформированных объектах канолы. Уровни экспрессии находились в диапазоне от 14 до более 700 м.д. общего растворимого белка (TSP). Образцы трех различных нетрансформированных растений тестировали параллельно без детектируемого сигнала, что свидетельствует о том, что антитела, используемые в анализе, имеют минимальную перекрестную реактивность с клеточным матриксом канолы. Эти образцы также подтверждали как положительные при анализе вестерн-блоттинга. Краткое изложение результатов представлено в таблице 24.
Уровни экспрессии AAD-12 (v1) в растениях канолы
(нг/мл)
(ELISA)
Послевсходовая устойчивость к гербицидам у канолы T0, трансформированной с использованием AAD-12(v1): Сорок пять объектов T0 из трансформированных с использованием pDAB3759 отправляли в теплицу на период времени и позволяли им акклиматизироваться. Растения выращивали до появления 2-4 новых, выглядящих нормальными листьев (т.е. растения переносили из условий культивирования ткани в условия роста в теплице). Затем растения обрабатывали летальной дозой коммерческих составов 2,4-D-Амин 4 при норме внесения 560 г кэ/га. Нанесение гербицида осуществляли с использованием автоматического устройства для опрыскивания при объеме раствора для опрыскивания 187 л/га, высоте опрыскивания 50 см. Летальную дозу определяют как количество, вызывающее >95% повреждений у нетрансформированных контролей.
Двадцать четыре из объектов являлись устойчивыми к нанесению гербицида 2,4-D DMA. Некоторые объекты имели незначительные повреждения, но восстанавливались через 14 DAT. Объекты совершенствовали до T1 (и поколения T2) посредством самоопыления в контролируемых условиях в мешках.
Наследование AAD-12 (v1) у канолы: Также проводили тестирование 100 растений потомков на 11 линиях T1 с AAD-12 (v1). Семена высевали и пересаживали в 3-дюймовые горшки, наполненные средами Metro Mix. Затем все растения опрыскивали 560 г кэ/га 2,4-D DMA, как описано выше. Через 14 DAT подсчитывали резистентные и восприимчивые растения. Семь из 11 тестируемых линий расщеплялись в соответствии с единым локусом, доминантным признаком с менделевским наследование (3R:1S), как определяли с помощью анализа с использованием хи-квадрата. AAD-12 наследуется как стабильный ген резистентности к арилоксиалканоатауксинам у многих видов, и его можно подвергать стэкингу с одним или несколькими дополнительными генами резистентности к гербицидам.
Наследование AAD-12 (v1) у канолы: Также проводили тестирование 100 растений потомков на 11 линиях T1 с AAD-12 (v1). Семена высевали и пересаживали в 3-дюймовые горшки, наполненные средами Metro Mix. Затем все растения опрыскивали 560 г кэ/га 2,4-D DMA, как описано выше. Через 14 DAT подсчитывали резистентные и восприимчивые растения. Семь из 11 тестируемых линий расщеплялись в соответствии с единым локусом, доминантным признаком с менделевским наследование (3R:1S), как определяли с помощью анализа с использованием хи-квадрата. AAD-12 наследуется как стабильный ген резистентности к арилоксиалканоатауксинам у многих видов, и его можно подвергать стэкингу с одним или несколькими дополнительными генами резистентности к гербицидам.
Подтверждение высокой устойчивости к 2,4-D у канолы T1: В случае T1 с AAD-12 (v1) 5-6 мг семян стратифицировали, высевали и добавляли тонкий слой сред Sunshine Mix #5 в качестве верхнего слоя почвы. Проросшие растения подвергали селекции с использованием 560 г кэ/га 2,4-D через 7 и 13 дней после посадки.
Ответ T1 с AAD-12 (v1) и нетрансформированного контроля на послевсходовую обработку различными количествами 2,4-D DMA: Средний % повреждений через 14 DAT
2,4-D DMA
Выжившие растения пересаживали в 3-дюймовые горшки, содержащие среды Metro Mix. Выжившие растения из потомков T1, подвергнутых селекции с использованием 560 г кэ/га 2,4-D, также пересаживали в 3-дюймовые горшки, наполненные почвой Metro Mix. В фазу 2-4 листьев растения опрыскивали 280, 560, 1120 или 2240 г кэ/га 2,4-D DMA. Растения классифицировали через 3 и 14 DAT и сравнивали с нетрансформированными контрольными растениями. Выборка данных о повреждениях объектов T1 через 14 DAT представлена в таблице 25. Данные позволяют предполагать, что многочисленные объекты являются стабильно резистентными к 2240 г кэ/га 2,4-D, в то время как другие объекты демонстрировали менее стабильную устойчивость к нормам внесения до 1120 г кэ/га 2,4-D. Выжившие растения пересаживали в горшки 51/4'', содержащие среды Metro Mix, помещали в те же условия выращивания, как описано выше, и подвергали самоопылению для получения только гомозиготных семян.
Устойчивость растений канолы pDAB3759 к гербицидам 2,4-D, дихлорпропу, триклопиру и флуроксипиру в полевых условиях: Полевое испытание уровня устойчивости проводили на двух объектах с AAD-12 (v1) 3759(20)018.001 и 3759(18)030.001 и каноле дикого типа (Nex710) в Fowler, Ind. Дизайн эксперимента представлял собой рандомизированный полноблоковый план с 3 повторениями. Варианты эксперимента представляли собой 2,4-D (диметиламиновая соль) при 280, 560, 1120, 2240 и 4480 г кэ/га, триклопир при 840 г кэ/га, флуроксипир при 280 г кэ/га и необработанный контроль. В рамках каждого варианта эксперимента высаживали один 20-футовый ряд/объект для объекта 3759(18)030.0011, 3759(18)018.001 и линии дикого типа (Nex710) с использованием 4-рядной сеялки с расстоянием между рядами 8 дюймов. Когда канола достигала фазы 4-6 листьев, проводили обработки гербицидами с использованием пневматического ранцевого опрыскивателя, выпускающего объем раствора для опрыскивания 187 л/га под давлением 130-200 кПа. Визуальную оценку повреждений проводили через 7, 14 и 21 день после нанесения.
Ответ растений канолы с AAD-12 (pDAB3759) на 2,4-D, триклопир и флуроксипир
Ответ канолы на 2,4-D, триклопир и флуроксипир представлен в таблице 26. Канола дикого типа (Nex710) являлась тяжело поврежденной (72% через 14 DAT) 2,4-D при 2240 г кэ/га, что считают 4-кратной нормой внесения. Все объекты с AAD-12 (v1) демонстрировали исключительную устойчивость к 2,4-D через 14 DAT со средними повреждениями 2, 3 и 2%, наблюдаемыми при 1-, 2- и 4-кратных нормах внесения, соответственно. Канола дикого типа являлась тяжело поврежденной (25% через 14 DAT) 2-кратной нормой внесения триклопира (840 г кэ/га). Объекты с AAD-12 (v1) демонстрировали устойчивость при 2-кратных нормах внесения триклопира со средними повреждениями 6% через 14 DAT для двух объектов. Флуроксипир при 280 г кэ/га вызывал у нетрансформированной линии тяжелые повреждения (37%) через 14 DAT. Объекты с AAD-12 (v1) демонстрировали повышенную устойчивость со средними повреждениями 8% через 5 DAT.
Эти результаты свидетельствуют о том, что объекты, трансформированные с использованием AAD-12 (v1), демонстрировали высокий уровень резистентности к 2,4-D, триклопиру и флуроксипиру при нормах внесения, являвшихся летальными или вызывающими тяжелые эпинастические деформации у нетрансформированной канолы. Показано, что AAD-12 имеет относительную эффективность в отношении 2,4-D > триклопира > флуроксипира.
ПРИМЕР 15
Трансформация и селекция объекта сои DAS-68416-4 с AAD-12
Трансгенный объект сои (Glycine max) DAS-68416-4 получали с помощью опосредованной Agrobacterium трансформации эксплантатов семядольного узла сои. Для инициации трансформации использовали инактивированный штамм Agrobacterium EHA101 (Hood et al., 2006), несущий бинарный вектор pDAB4468 (фиг. 2) с селективным маркером (pat) и интересующим геном (AAD-12) внутри T-области цепи ДНК.
Осуществляли опосредованную Agrobacterium трансформацию. В кратком изложении, семена сои (cv Maverick) проращивали на базовых средах, выделяли семядольные узлы и инфицировали Agrobacterium. Среды для инициации прорастания побегов, удлинения побегов и укоренения дополняли цефотаксимом, тиментином и ванкомицином для удаления Agrobacterium. Использовали селекцию глуфосинатом для ингибирования роста нетрансформированных побегов. Подвергнутые селекции побеги переносили на среду для укоренения для развития корней, а затем переносили на почвенную смесь для акклиматизации проростков.
На концевые листочки подвергнутых селекции проростков наносили глуфосинат для скрининга на предмет предполагаемых трансформантов. Подвергнутые скринингу проростки переносили в теплицу, позволяли им акклиматизироваться, а затем на листья наносили глуфосинат для подтверждения устойчивости и считали их предполагаемыми трансформантами. Отбирали подвергнутые скринингу растения и осуществляли молекулярные анализы для подтверждения наличия гена селективного маркера и/или интересующего гена. Для получения семян T1 растениям T0 позволяли самоопыляться в теплице.
Растения T1 подвергали обратному скрещиванию и подвергали интрогрессии в элитную идиоплазму (Maverick). Этот объект сои DAS-68416-4 получали из независимого трансформированного изолята. Объект подвергали скринингу с учетом его уникальных характеристик, таких как единый участок инсерции, нормальное менделевское расщепление и стабильная экспрессия, и превосходная комбинация эффективности, включая устойчивость к гербицидам, и агрономических характеристик при различном генетическом фоне и во многих местностях с различным климатом. Дополнительное описание объекта сои DAS-68416-4 представлено в WO 2011/066384, включенном в качестве ссылки в настоящее описание в полном объеме.
ПРИМЕР 16
Получение агрономических данных за 2008 год
Агрономическое исследование с использованием объекта сои DAS-68416-4 и нетрансгенного контроля (var. Maverick) проводили в 2008 году в шести местах, находящихся в Айове, Иллинойсе, Индиане, Небраске и Онтарио, Канада (2 места). Оценивали агрономические показатели, включая подсчет густоты стояния/популяции, мощность проростка/растения, высоту растения, полегание, частоту заболеваний, повреждение насекомыми и дни до цветения, для исследования эквивалентности объекта сои DAS-68416-4 (с обработками гербицидами и без них) по сравнению с контрольной линией Maverick. Это исследование обозначают как Агрономический эксперимент S1.
Агрономические показатели, оцениваемые в агрономическом эксперименте S1
10=Эквивалентность росту нетрансформированных растений
9=Жизнеспособность растений составляет 90% по сравнению с нетрансформированными, и т.д.
повреждения
10=Эквивалентность росту нетрансформированных растений
9=Жизнеспособность растений составляет 90% по сравнению с нетрансформированными, и т.д.
Семена тестируемой и контрольной сои высевали при норме высева приблизительно 112 семян на 25-футовый ряд с расстоянием между рядами приблизительно 30 дюймов (75 см). В каждой местности создавали три повторяющихся опытных участка для каждого варианта эксперимента, при этом каждый опытный участок состоял из 2-25-футовых рядов. Опытные участки распределяли согласно рандомизированному полноблоковому плану (RCB) с уникальной рандомизацией в каждой местности. Каждый опытный участок сои ограничивали двумя рядами нетрансгенной сои схожей зрелости. Все место проведения испытаний окружали минимум 10 футами нетрансгенной сои схожей относительной зрелости.
Обработки гербицидами осуществляли при объеме раствора для опрыскивания приблизительно 20 галлонов на акр (187 л/га). Эти обработки планировали для воспроизведения максимальной отмеченной на маркировке нормы внесения, используемой в коммерческой практике. 2,4-D наносили в виде трех избыточных обработок распылением всего 3 фунта кэ/акр за сезон. Отдельные нанесения 1,0 фунта кэ/акр (1,120 г/га) осуществляли предвсходно и приблизительно в фазы роста V4 и R2. Глуфосинат наносили в виде двух избыточных обработок распылением всего 0,74 фунта активного ингредиента/акр (828 г активного ингредиента/га) за сезон. Отдельные нанесения 0,33 фунта активного ингредиента/акр и 0,41 фунта активного ингредиента/акр (374 и 454 г активного ингредиента/га) осуществляли приблизительно в фазы роста V6 и R1.
Для агрономических данных проводили дисперсионный анализ среди мест полевых испытаний с использованием смешанной модели (SAS версии 8; SAS Institute 1999). Входные элементы данных считали фиксированными эффектами, а местность, блок внутри местности, местность по входным элементам данных, и входные элементы данных по блоку внутри местности обозначали как случайные эффекты. Значимость общего эффекта условий оценивали с использованием критерия Фишера. С помощью критерия Стьюдента осуществляли попарное сопоставление между входными элементами данных для контроля и неопрысканного трансгенного объекта сои DAS-68416-4 (неопрысканный), объекта сои DAS-68416-4, опрысканного глуфосинатом (объект сои DAS-68416-4 + глуфосинат), объекта сои DAS-68416-4, опрысканного 2,4-D (объект сои DAS-68416-4 + 2,4-D), и объекта сои DAS-68416-4, опрысканного глуфосинатом и 2,4-D (объект сои DAS-68416-4 + оба). Также вычисляли скорректированные значения P с использованием средней доли ложных отклонений гипотезы (FDR) для контроля эффекта множественных сравнений (Benjamini and Hochberg, 1995).
Проводили анализ агрономических данных, собранных для контроля, неопрысканного объекта сои DAS-68416-4, объекта сои DAS-68416-4+2,4-D, объекта сои DAS-68416-4+глуфосинат, и объекта сои DAS-68416-4+оба гербицида. Не наблюдали статистически значимых различий для подсчета густоты стояния, ранней популяции, мощности проростков, повреждений после обработки, полегания, конечного подсчета густоты стояния или дней до цветения (таблица 28). В случае высоты растений вычисляли значимый двусторонний критерий Стьюдента между контролем и объектом сои DAS-68416-4 + опрыскивание 2,4-D. Однако, не наблюдали значимый общий эффект условий, различия являлись очень незначительными между обработанным объектом сои DAS-68416-4 и контролем, и не наблюдали различий между различными вариантами эксперимента с объектом сои DAS-68416-4. Учитывая эти результаты, объект сои DAS-68416-4 являлся агрономически эквивалентным почти изогенному нетрансгенному контролю.
Анализ агрономических характеристик из агрономического эксперимента S1
(Pr>F)a
(Значение Pb Скоррект. P)c
(Значение P Скоррект. P)
(Значение P Скоррект. P)
ПРИМЕР 17
Получение агрономических данных за 2009 год
Агрономическое исследование с использованием объекта сои DAS-68416-4 и нетрансгенного контроля (var. Maverick) проводили в 2009 году в 8 местах, находящихся в Арканзасе, Айове, Иллинойсе, Индиане, Миссури и Небраске. Оценивали агрономические показатели, включая подсчет густоты стояния/популяции, мощность проростка/растения, высоту растения частоту заболеваний, повреждение насекомыми и дни до цветения, для исследования эквивалентности объекта сои DAS-68416-4 (с обработками гербицидами и без них) контролю (таблица 29).
Данные, собранные в агрономических исследованиях и исследованиях урожайности в 2009 году.
S - тестирования только опрысканных растений
Для испытаний использовали рандомизированный полноблочный план. Входными элементами данных являлись объект сои DAS-68416-4, контрольная линия Maverick и коммерчески доступные нетрансгенные линии сои. Высевали семена тестируемой, контрольной и референсной сои при норме высева приблизительно 112 семян на ряд с расстоянием между рядами приблизительно 30 дюймов (75 см). В каждой местности создавали 4 повторяющихся опытных участка для каждого варианта эксперимента, при этом каждый опытный участок состоял из 2-25-футовых рядов. Каждый опытный участок сои ограничивали 2 рядами нетрансгенной сои (Maverick). Все место испытаний окружали минимум 4 рядами (или 10 футами) нетрансгенной сои (Maverick). Для получения агрономически приемлемой сельскохозяйственной культуры осуществляли соответствующие практики по контролю насекомых, сорняков и заболеваний.
Осуществляли обработки гербицидами для воспроизведения максимальной отмеченной на маркировке нормы внесения, используемой в коммерческой практике. Варианты эксперимента состояли из неопрысканного контроля и обработок гербицидами 2,4-D, глуфосинатом, 2,4-D/глуфосинатом, наносимых в конкретные фазы роста. Для обработки 2,4-D гербицид наносили при норме внесения 1,0 фунт кэ/акр (1,120 г кэ/га) в фазы роста V4 и R2. Для обработок глуфосинатом нанесения осуществляли на растения в фазы роста V4 и V6-R2. Для обработок обоими гербицидами глуфосинат наносили при норме внесения 0,33 фунта активного ингредиента/акр (374 г активного ингредиента/га) и 0,41 фунта активного ингредиента/акр (454 г активного ингредиента/га) для нанесений в фазы V4 и V6-R2, соответственно. Входными элементами данных для обработок обоими гербицидами являлись объект сои DAS-68416-4 и контроли, включая нетрансгенную Maverick. Ожидали, что опытные участки с Maverick погибнут после нанесения гербицидов.
Для агрономических данных проводили дисперсионный анализ среди мест полевых испытаний с использованием смешанной модели (SAS версии 8; SAS Institute 1999). Входные элементы данных считали фиксированными эффектами, а местность, блок внутри местности, местность по входным элементам данных, и входные элементы данных по блоку внутри местности обозначали как случайные эффекты. Аналогично осуществляли анализ в отдельных местах проведения испытаний с входными элементами данных в качестве фиксированных эффектов и блоком и входными элементами данных по блоку в качестве случайных эффектов. Для статистического анализа данные округляли. Описывали значимые различия с 95% уровнем значимости, и значимость общего эффекта условий оценивали с использованием критерия Фишера. С помощью критерия Стьюдента осуществляли попарное сопоставление между неопрысканным трансгенным объектом с AAD-12 (неопрысканный), объектом с AAD-12, опрысканным глуфосинатом (AAD-12+глуфосинат), объектом с AAD-12, опрысканным 2,4-D (AAD-12+2,4-D), и объектом с AAD-12, опрысканным глуфосинатом и 2,4-D (AAD-12+2,4-D+глуфосинат), и контролем.
По причине большого количества сопоставлений, осуществляемых в этом исследовании, проблемой являлся эффект множественных сравнений. Эффект множественных сравнений является проблемой, когда в одном исследовании осуществляют большое количество сравнений для обнаружения неожиданных эффектов. В этих условиях вероятность ложно определенных различий на основе связанных со сравнениями значений p является очень высокой (1-0,95количество сравнений). В этом исследовании осуществляли четыре сравнения на анализируемый параметр (16 анализируемых типов наблюдений для агрономических данных), приводящих к 64 сравнениям для агрономических данных. Таким образом, вероятность определения одного или нескольких ложных различий на основе нескорректированных значений p составляла 99% для агрономических данных (1-0,9564).
Проводили анализ агрономических данных, собранных для контроля, неопрысканного объекта с AAD-12, AAD-12+глуфосинат, AAD-12+2,4-D, и AAD-12+2,4-D+глуфосинат. В случае анализа по всей местности не наблюдали статистически значимых различий для мощности проростков, конечной популяции, мощности растений/повреждений (V4, R1), полегания, частоты заболеваний, повреждения насекомыми, дней до цветения, дней до созревания, количества бобов, количества семян, урожайности и высоты растения. В случае подсчета густоты стояния и ранней популяции вычисляли значимый двусторонний критерий Стьюдента между контролем и AAD-12 + глуфосинат, но это не сопровождалось значимым общим эффектом условий или FDR-скорректированным значением p. В случае мощности растений/повреждения (R2) вычисляли значимый двусторонний критерий Стьюдента и значимый общий эффект условий между контролем и AAD-12 + глуфосинат и AAD-12+2,4-D+глуфосинат, но это не сопровождалось значимым FDR-скорректированным значением p. Средние результаты для всех этих переменных также находились в пределах диапазона, обнаруживаемого для референсных линий, тестируемых в этом исследовании.
ПРИМЕР 18
Трансформация и селекция объекта pDAS 1740-278 с AAD-1
Объект pDAS 1740-278 с AAD-1 получали с помощью вискер-опосредованной трансформации линии кукурузы Hi-II. Используемый способ трансформации описывают в патентной заявке США № 20090093366. Линию кукурузы трансформировали с использованием FspI-фрагмента плазмиды pDAS 1740 (фиг. 3), также обозначаемого как pDAB3812. Эта плазмидная конструкция содержит экспрессирующую кассету растений, содержащую RB7 MARv3:: промотор убиквитина 1 Zea mays v2//AAD1 v3//PER5 3'UTR Zea mays:: RB7 MARv4 транскрипционную единицу растений (PTU).
Получали множество объектов. Объектам, выжившим и продуцирующим здоровую, резистентную к галоксифопу ткань каллюса, присваивали уникальные коды идентификации, представляющие предполагаемые трансформированные объекты, и их непрерывно переносили на свежую селективную среду. Растения подвергали регенерации из ткани, полученной от каждого уникального объекта, и переносили в теплицу.
Брали образцы листьев для молекулярного анализа для подтверждения наличия трансгена AAD-I с помощью Саузерн-блоттинга, подтверждения границ ДНК и подтверждения сопутствующего геномного маркера. Положительные растения T0 опыляли с использованием инбредных линий для получения семян T1. Растения T1 объекта pDAS 1470-278-9 (DAS-40278-9) подвергали селекции, самоопылению и охарактеризовывали в течение пяти поколений. В это же время растения T1 подвергали обратному скрещиванию и интрогрессии в элитную идиоплазму (XHH 13) посредством селекции с помощью маркера в течение нескольких поколений. Этот объект получали из независимого трансформированного изолята. Объект подвергали селекции с учетом его уникальных характеристик, таких как единый участок инсерции, нормальное менделевское расщепление и стабильная экспрессия, лучшая комбинация эффективности, включая устойчивость к гербицидам, и агрономических характеристик при различном генетическом фоне и во многих местностях с различным климатом. Дополнительное описание, касающееся объекта кукурузы pDAS-1740-278-9, приведено в WO 2011/022469, включенном в настоящее описание в качестве ссылки в полном объеме.
ПРИМЕР 19
Обработка гербицидами и агрономические данные
Обработки гербицидами осуществляли с использованием объема раствора для опрыскивания приблизительно 20 галлонов на акре (187 л/га).
Эти обработки осуществляли для воспроизведения максимальной отмеченной на маркировке нормы внесения, используемой в коммерческой практике. Использовали Weedar 64 (026491-0006) в концентрации 39%, 3,76 фунта кэ/галлон, 451 г кэ/л и Assure II (106155) в концентрации 10,2%, 0,87 фунта активного ингредиента/галлон, 104 г активного ингредиента/га.
2,4-D (Weedar 64) наносили в виде 3 избыточных нанесений распылением на тестируемые входные элементы данных 4 и 5 (всего 3 фунта кэ/акр за сезон). Отдельные нанесения осуществляли в предвсходный период и приблизительно в фазы V4 и V8-V8,5. Для Weedar 64 отдельные плановые нормы внесения составляли 1,0 фунт кэ/акр или 1120 г кэ/га. Точные нормы внесения находились в диапазоне 1096-1231 г кэ/акр.
Квизалофоп (Assure II)наносили в виде одного избыточного нанесения распылением на тестируемые входные элементы данных 3 и 5. Время нанесения приходилось приблизительно на фазу роста V6. Для Assure II плановая норма внесения составляла 0,0825 фунта активного ингредиента/акр или 92 г активного ингредиента/га. Точные нормы внесения находились в диапазоне 90,8-103 г активного ингредиента/га. Агрономические характеристики регистрировали для всех тестируемых элементов в блоках 2, 3 и 4 в каждой местности. В таблице 30 представлены измеряемые характеристики.
Агрономические данные для объекта кукурузы pDAS- 1740-278-9
повреждения
Проводили анализ агрономических данных, собранных для контроля, неопрысканного объекта с aad-1, aad-1 + 2,4-D, aad-1 + квизалофоп, и aad-1 + оба гербицида. В случае анализа по всей местности не наблюдали статистически значимых различий для значений ранней популяции (V1 и V4), мощности, конечной популяции, повреждений сельскохозяйственной культуры, времени до выметывания пестичных столбиков, времени до сбрасывания пыльцы, полегания стеблей, полегания корней, частоты заболеваний, повреждения насекомыми, дней до созревания, высоты растения и жизнеспособности пыльцы (формы и цвета) в сводном анализе по местности. В случае ремонтантности и высоты початка вычисляли значимый двусторонний критерий Стьюдента между контролем и объектом с aad-1 + квизалофоп, но это не сопровождалось значимым общим эффектом условий или значениями p, скорректированными по средней доле ложных отклонений гипотезы (FDR) (таблица 31).
Результаты сводного анализа агрономических характеристик по местностям для объекта кукурузы DAS-40278-9 с aad-1 (опрысканного и неопрысканного) и контроля
(0,303, 0,819)
(0,443, 0,819)
(0,556, 0,819)
(0,393, 0,819)
(0,883, 0,984)
(0,687, 0,863)
(0,423, 0,819)
(0,263, 0,819)
(0,197, 0,819)
(0,161, 0,819)
(0,633, 0,819)
(0,729, 0,889)
(0,747, 0,889)
(0,802, 0,924)
(0,943, 1,00)
(0,521, 0,819)
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
(0,130, 0,819)
(единицы количества тепла)f
(0,996, 1,00)
(0,781, 0,917)
(0,088, 0,819)
(0,224, 0,819)
(единицы количества тепла)f
(0,564, 0,819)
(0,480, 0,819)
(0,245, 0,819)
(0,245, 0,819)
0 минут (%)g
(0,931, 1,00)
(0,546, 0,819)
(0,439, 0,819)
(0,605, 0,819)
30 минут (%)
(0,618, 0,819)
(0,409, 0,819)
(0,739, 0,889)
(0,409, 0,819)
60 минут (%)
(0,809, 0,924)
(0,470, 0,819)
(0,629, 0,819)
(0,629, 0,819)
120 минут (%)
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
(0,337, 0,819)
30 минут (%)h
(0,850, 0,960)
(0,306, 0,819)
(0,573, 0,819)
(0,573, 0,819)
60 минут (%)
(0,612, 0,819)
(0,315, 0,819)
(0,315, 0,819)
(0,612, 0,819)
120 минут (%)
(0,356, 0,819)
(0,356, 0,819)
(0,356, 0,819)
(0,356, 0,819)
(0,457, 0,819)
(0,457, 0,819)
(0,457, 0,819)
(0,373, 0,819)
(0,250, 0,819)
(0,034m, 0,819)
(0,144, 0,819)
(0,106, 0,819)
(0,383, 0,819)
(0,265, 0,819)
(0,265, 0,819)
(0,265, 0,819)
(0,500, 0,819)
(0,500, 0,819)
(0,736, 0,889)
(0,500, 0,819)
(0,558, 0,819)
(0,302, 0,819)
(0,185, 0,819)
(0,104, 0,819)
(0,206, 0,819)
(0,109, 0,819)
(0,350, 0,819)
(0,286, 0,819)
(0,089, 0,819)
(0,018m, 0,786)
(0,214, 0,819)
(0,016m, 0,186)
ПРИМЕР 20
Дополнительные агрономические испытания
Агрономические характеристики линии кукурузы 40278 по сравнению с почти изогенной линией кукурузы оценивали в различных условиях окружающей среды. Варианты экспериментов включали 4 генетически различающихся гибрида и соответствующие им почти изогенные контрольные гибриды, тестируемые всего в 21 местности.
Четыре тестируемых гибрида являлись гибридами с созреванием от среднего до позднего в диапазоне от 99 до 113 дней относительного созревания. В эксперименте A тестировали объект DAS-40278-9 с генетическим фоном инбредной линии C × конверсии BC3S1. Этот гибрид имеет относительное созревание 109 дней, и его тестировали в 16 местностях (Таблица 32). В эксперименте B тестировали гибрид с фоном инбредной линии E × конверсии BC3S1, гибрид с относительным созреванием 113 дней. Этот гибрид тестировали в 14 местностях с использованием немного отличающегося набора местностей, чем в эксперименте A. В экспериментах C и D тестировали гибриды с фоном конверсии BC2S1 × инбредной линии D и конверсии BC2S1 × инбредной линии F, соответственно. Оба эти гибрида имеют относительное созревание 99 дней, и их тестировали в 10 одинаковых местностях.
Для каждого испытания использовали рандомизированный полноблочный план с двумя повторениями на местность и двухрядными опытными участками. Длина ряда составляла 20 футов, и каждый ряд высевали при 34 семенах на ряд. При проведении испытаний использовали стандартную региональную агротехнику.
Данные собирали и анализировали по восьми агрономическим характеристикам; высоте растения, высоте початка, полеганию побегов, полеганию корней, конечной популяции, влажности зерна, тестовой массе и урожайности. Параметры высота растения и высота початка представляют информацию о внешнем виде гибридов. Агрономические характеристики процент полегания стеблей и полегания корней определяют собираемость урожая гибрида. С помощью подсчета конечной популяции измеряют качество семян и сезонные условия выращивания, влияющие на урожай. С помощью процента влажности зерна при сборе определяют созревание гибрида, и с помощью урожайности (бушели/акр, скорректированные по влажности) и тестовой массы (массы в фунтах на бушель кукурузы, скорректированный по 15,5% влажности) описывают репродуктивную способность гибрида.
Проводили дисперсионный анализ по местам полевых испытаний с использованием линейной модели. Входной элемент данных и местность включали в модель в качестве фиксированных эффектов. Изучали смешанные модели, включающие местность и местность по входному элементу данных в качестве случайных эффектов, но местность по входному элементу данных объясняла лишь небольшую часть дисперсии и ее компонента дисперсии часто не отличалась значимо от нуля. В случае полегания стеблей и корней использовали логарифмическое преобразование для стабилизации дисперсии, однако средние значения и диапазоны записывали по исходной шкале. Описывали значимые различия с 95% уровнем значимости. Значимость общего эффекта условий оценивали с использованием критерия Стьюдента.
Результаты этих исследований агрономических характеристик можно найти в таблице 32. Не наблюдали статистически значимых различий для любого из четырех гибридов 40278 по сравнению с изогенными контрольными линиями (при p<0,05) для параметров высоты початка, полегания стеблей, полегания корней, влажности зерна, тестовой массы и урожайности. Подсчет конечной популяции и высота растения являлись статистически значимыми в экспериментах A и B, соответственно, но не обнаруживали аналогичных различий при сравнениях с другими тестируемыми гибридами 40278. Некоторые из наблюдаемых колебаний могут являться результатом низкого уровня генетической изменчивости, оставшихся после обратного скрещивания объекта DAS-40278-9 в элитные инбредные линии. Общий диапазон значений всех измеряемых параметров находится в пределах диапазона значений, получаемого для общепринятых гибридов кукурузы, и не приводит к выводу о повышенной засоренности сорняками. В заключение, агрономические данные свидетельствуют о том, что кукуруза 40278 биологически эквивалентна общепринятой кукурузе.
Анализ агрономических характеристик
Агрономические характеристики гибридной кукурузы, содержащей объект DAS-40278-9, по сравнению с почти изогенной линией кукурузы собирали из множества мест полевых испытаний в различных географических областях в течение периода вегетации. Результаты для гибридных линий кукурузы, содержащих объект DAS-40278-9, по сравнению с растениями без вставки представлены в таблице 33.
"Name" - "Название", "Hybrid corn containing event DAS-40278-9" - "Гибридная кукуруза, содержащая объект DAS-40278-9", "Control hybrid corn" - "Контрольная гибридная кукуруза, "Yield" - "Урожайность", "Grain moisture" - "Влажность зерна", "Final population (plants/acre in 1000s)" - "Конечная популяция (растения/акр в тыс.)"]
Данные об урожайности, проценте влажности зерна и конечной популяции для гибридной кукурузы, содержащей объект DAS-40278-9, по сравнению с почти изогенной контрольной линией
Агрономические характеристики гибридных линий кукурузы, содержащих объект DAS-40278-9, и растений без вставки, опрысканных гербицидами квизалофопом (280 г кэ/га) в фазе роста V3 и 2,4-D (2,240 г кэ/га) в фазе роста V6, представлены в таблице 34.
Агрономические данные для объекта DAS 40278-9 по сравнению с почти изогенной контрольной линией
ПРИМЕР 21
2,4-D повышает рост резистентной к 2,4-D сои
Трансгенная соя с трансгеном AAD-12 обеспечивает защиту растению сои, в то время как сорняки уничтожают нанесением 2,4-D. Неожиданно наблюдали, что 2,4-D также повышает рост устойчивой к 2,4-D сои. Этот повышенный рост приводит к повышению высоты растений и/или урожайности опрысканных опытных участков по сравнению с неопрысканными опытными участками.
Повышение роста растений и/или урожайности, являющееся результатом обработки 2,4-D, описывают для растений сои, генетически сконструированных устойчивыми к 2,4-D. Испытания проводили во множестве местностей, покрывающих область выращивания сои в Северной Америке. Входные элементы данных включали элитные линии, в которые интрогрессировали объект DAS-68416-4 (обусловливающий устойчивость к 2,4-D). Варианты экспериментов состояли из неопрысканных растений и растений, опрысканных 2,4-D в фазах роста V3 и R2. Опытные участки исследовали на различные агрономические характеристики на всем протяжении сезона, включая высоту растения и урожай зерна. Сорняки контролировали на всем протяжении сезона на опрысканных и неопрысканных опытных участках для устранения любого конкурентного эффекта. В заключение испытания при анализе данных наблюдали значимое повышение высоты растений и урожайности для тех входных элементов данных, которые опрыскивали 2,4-D, по сравнению с неопрысканными. Повышение урожайности является дополнительным преимуществом контроля сорняков, осуществляемого с помощью 2,4-D в отношении резистентной к 2,4-D сои.
Полевые испытания начинали в 2011 году для сравнения агрономических характеристик объекта сои DAS-68416-4 (международная патентная заявка № 2011/066384), опрысканного 2,4-D, с агрономическими характеристиками неопрысканного объекта сои DAS-68416-4. Полевые испытания включали входные элементы данных 4 элитных линий сои, в которые интрогрессировали объект сои DAS-68416-4, и соответствующие изогенные линии сои без вставок из 4 элитных линий сои, не содержащих объект сои DAS-68416-4. Испытания проводили в различных географических местностях (всего 10 местностей). Эксперимент осуществляли в виде модифицированной расщепленной делянки с двумя повторениями на местность. Целые опытные участки являлись вариантами эксперимента, а подучастки являлись входными элементами данных. Каждый опытный участок состоял из двух рядов длиной 12,5 футов, посаженных с расстоянием 30 дюймов. Опрыскиваемые опытные участки обрабатывали 2,4-D (1120 г кэ/га), наносимым в фазы роста V3 и R2. На всем протяжении сезона полевые опытные участки поддерживали в соответствии с общепринятой агротехникой и очищали от сорняков.
Для определения того, как обработка 2,4-D влияет на агрономические характеристики сои, измеряли различные агрономические характеристики растений сои. Тестируемые агрономические характеристики и фазы роста на момент сбора данных представлены в таблице 35.
Список агрономических характеристик, измеряемых в исследованиях урожая 2011 года для сравнения опрысканного 2,4-D и неопрысканного объекта сои DAS-68416-4
В конце сезона выращивания сои объединяли данные со всех местностей и осуществляли анализ по местностям. Анализ данных осуществляли с использованием JMP® Pro 9.0.3 (SAS, Cary, NC). Средние значения, полученные при анализе методом наименьших квадратов, представлены в таблице 28. Обработка 2,4-D объекта сои DAS-68416-4, содержащего трансген AAD-12, приводила к кондиционному эффекту повышения роста. Повышенный рост приводил к значимо более высокой урожайности и высоте растений, измеряемых на полевых опытных участках, опрысканных 2,4-D, по сравнению с полевыми опытными участками, неопрысканными 2,4-D. Эти повышения являлись очевидными, когда данные анализировали совокупно по всем местностям. В отличие от этого, повышенная урожайность объекта сои DAS-68416-4, опрысканного 2,4-D, снижалась при анализе взаимодействия местность-вариант эксперимента. Средняя высота растений и урожайность повышались приблизительно на 5% при обработке 2,4-D (таблица 36).
Средние значения, полученные методом наименьших квадратов, в анализе по местностям для сравнения объекта сои DAS-68416-4, опрысканного 2,4-D, с неопрысканными растениями. Уровни, не помеченные одной и той же буквой, значимо различаются
Как показано в таблице 37, по меньшей мере в одной из десяти местностей (местность №a3) зарегистрировали значимо более высокую урожайность для неопрысканных растений объекта сои DAS-68416-4 по сравнению с опрысканными 2,4-D растениями объекта сои DAS-68416-4. При суммировании результатов со всех местностей, обработка 2,4-D объекта сои DAS-68416-4, содержащего трансген AAD-12, демонстрировала кондиционный эффект, приводящий к повышенному росту. Например, урожайность растений объекта сои DAS-68416-4, опрысканных 2,4-D, составляла 56,4 бушелей/акр, являясь существенно большей, чем урожайность неопрысканных растений объекта сои DAS-68416-4, составляющая 53,7 бушелей/акр. Аналогично, высота растений объекта сои DAS-68416-4, опрысканных 2,4-D, составляла 81 см, являясь существенно большей, чем высота неопрысканных растений объекта сои DAS-68416-4, составляющая 77 см.
Средние значения урожайности, полученные методом наименьших квадратов, в конкретных местностях для сравнения объекта сои DAS-68416-4, опрысканного 2,4-D, с неопрысканными растениями. Уровни, не помеченные одной и той же буквой, значимо различаются
(бушели/акр)
ПРИМЕР 22
2,4-D повышает рост резистентной к 2,4-D сои в комбинации 2,4-D/глифосат
В 2010 году осуществляли аналогичные полевые испытания, что и в предыдущем примере, но с двумя обработками 2,4-D в комбинации с глифосатом. Результаты показали, что повышенный рост резистентной к 2,4-D сои по высоте растения и/или урожайности опрысканных опытных участков по сравнению с неопрысканными опытными участками является результатом обработки 2,4-D.
Наблюдали значимые эффекты условий для ряда измеряемых параметров. 2,4-D и глифосат наносили в фазы роста V3 и R2. Испытания проводили в различных географических местностях (всего шесть местностей). Тестируемые агрономические характеристики и фазы роста на момент сбора данных представлены в таблице 30. В случае опрысканной сои средняя высота растений повышалась на 6%, и средняя урожайность повышалась на 17% (таблица 38). Кроме того, в случае опрысканной сои средняя масса семян повышалась на 6%.
Средние значения, полученные методом наименьших квадратов, в анализе по местностям для сравнения устойчивой к 2,4-D сои, опрысканной 2,4-D и глифосатом, с неопрысканными растениями. Уровни, не помеченные одной и той же буквой, значимо различаются
Как показано в таблице 39, в этом примере также наблюдали колебания по конкретным географическим местностям. В случае опрысканной сои средняя урожайность повышалась на 21,6% (таблица 39).
Средние значения урожайности, полученные методом наименьших квадратов, в конкретных местностях для сравнения устойчивой к 2,4-D сои, опрысканной 2,4-D и глифосатом, с неопрысканными растениями
ПРИМЕР 23
Результаты испытаний урожайности для сравнения вариантов эксперимента с опрыскиванием и без опрыскивания
Резистентные к 2,4-D трансгенные сельскохозяйственные культуры, трансформированные с использованием арилоксиалканоатдиоксигеназы (AAD), приводили к повышенной урожайности при обработке стимулирующим количеством гербицида, содержащего остаток арилоксиалканоата. Объекты сои, содержащие экспрессирующую кассету гена AAD-12, тестировали в повторных испытаниях урожайности в условиях с опрыскиванием и без него. Проводили одну серию экспериментов, включавших сою с ранним созреванием, адаптированную к северным широтам, и другую серию экспериментов, включавших сою с поздним созреванием, адаптированную к более южным широтам. В предыдущих экспериментах имели место случаи, когда входные элементы данных сои, содержащей экспрессирующую кассету гена AAD-12, обработанные 2,4-D в течение сезона выращивания, проявляли повышенную урожайность относительно неопрысканных контролей.
Для испытаний использовали модифицированный план с расщепленными делянками с 2 повторениями. Каждый опытный участок был шириной в 2 ряда с расстоянием между рядами 30 дюймов и длиной 12,5 футов. Создавали проход шириной от 2,5 до 3 футов между опытными участками, расположенными непрерывной цепью, для обеспечения движения в течение испытания в течение сезона. Опрыскиваемые блоки опрыскивали последовательно (дважды) в течение сезона выращивания 2,4-D-холином + глифосатом (премиксом) при 2185 г кэ/га + AMS при 2% масс.
Список анализируемых местностей для испытаний урожайности со сравнением вариантов экспериментов с опрыскиванием и без него
Первую обработку осуществляли в фазу роста V3, а вторую обработку - в фазу роста R2. Экспериментальные и контрольные опытные участки полевых испытаний очищали от сорняков на всем протяжении сезона с использованием общепринятых гербицидов или ручной прополки. Собирали данные о всходах, мощности проростков, повреждениях сельскохозяйственной культуры, дате цветения, подсчете густоты стояния в R2, частоте заболеваний, повреждении насекомыми, высоте растений, даты созревания, полегании, растрескивании, массе 100 семян и урожайности. Данные анализировали с использованием JMP® Pro 9.0.3. В таблице 40 приведены местности, использованные для конечного анализа. Некоторые местности, в которых осуществляли посадку, не включали в анализ по причине вариабельности по опытному участку.
Анализ по местностям осуществляли для испытаний сои с ранним созреванием и поздним созреванием. В таблицах 41 и 42 представлен дисперсионный анализ урожайности для испытаний сои с ранним созреванием и поздним созреванием, соответственно.
Дисперсионный анализ урожайности по местностям (8 местностей) в испытаниях со сравнением опрысканной и неопрысканной сои с ранним созреванием
В испытаниях сои и с ранним, и с поздним созреванием наблюдали значимый эффект названия (P=0,05). Это являлось ожидаемым, т.к. каждая элитная линия сои, в которую интрогрессировали объект, имела различный генетический фон.
Дисперсионный анализ урожайности по местностям (8 местностей) в испытаниях со сравнением опрысканной и неопрысканной сои с поздним созреванием
данные
Признак
Определяли значимый эффект условий эксперимента в случае испытания сои с ранним созреванием, свидетельствующий о том, что варианты эксперимента с опрыскиванием и без него отличаются по урожайности. В случае испытания сои с поздним созреванием не наблюдали значимого эффекта условий эксперимента, что свидетельствует о том, что опрысканные и неопрысканные опытные участки не отличаются по урожайности.
Таблица средних значений урожайности, полученных методом наименьших квадратов, в испытании урожайности сои с ранним созреванием
В случае испытаний сои и с ранним, и с поздним созреванием эффект взаимодействия название-вариант эксперимента не являлся значимым, что свидетельствует о том, что эффект условий эксперимента (или отсутствие эффекта) являлся одинаковым для каждого входного элемента данных в конкретном испытании.
В таблице 43 представлена средняя урожайность для каждой комбинации входной элемент данных-вариант эксперимента в испытании сои с ранним созреванием, где HOMO означает гомозиготу. Значения, после которых следует одна и та же буква (в пределах указанного сорта), не отличаются по критерию Стьюдента при P=0,05. Наблюдали четыре входных элемента данных, проявляющих более высокую урожайность при последовательном опрыскивании в V3 и R3 с использованием 2,4-D-холина + глифосата (премикса) при 2185 г кэ/га + AMS.
В таблице 44 представлена средняя урожайность для каждой комбинации входной элемент данных-вариант эксперимента. Значения, после которых следует одна и та же буква (в пределах указанного сорта) не отличаются по критерию Стьюдента при P = 0,05. Как указано выше, в случае испытания сои с поздним созреванием не наблюдали значимого эффекта условий эксперимента или эффекта варианта эксперимента по входному элементу данных, поэтому множественные сравнения средних не осуществляли. Буквами в таблице указано, что не наблюдали различий между вариантами эксперимента с опрыскиванием и без него при тестировании сои с поздним созреванием.
Таблица средних значений урожайности, полученных методом наименьших квадратов, в испытании урожайности сои с ранним созреванием 2012 года
Результаты испытаний урожайности, представленные в этом примере, снова свидетельствуют о том, что в некоторых условиях окружающей среды в случае некоторых генотипов сои можно наблюдать повышение урожайности после обработки 2,4-D. В последние два года такое повышение урожайности наблюдали в испытаниях урожайности, проводимых в районе произрастания MG 2.
ПРИМЕР 24
Сравнение сои и кукурузы
Результаты по урожайности в полевых испытаниях сои, содержащей трансген AAD-12, свидетельствуют о том, что обработка 2,4-D может повышать урожайность сои в конкретных условиях окружающей среды для конкретных генотипов сои. Эти результаты являются неожиданными при сравнении с трансгенными объектами кукурузы, содержащими трансген AAD-1. Урожайность AAD-1-трансгенных растений кукурузы не демонстрировала стабильного статистически значимого повышения урожайности после опрыскивания 2,4-D. Эти AAD-1-трансгенные растения кукурузы биологически эквивалентны общепринятой кукурузе.
Дополнительные полевые испытания в различных географических областях проводили с 2010 по 2012 год на гибридных линиях кукурузы. На всем протяжении этих полевых испытаний урожайность линий кукурузы, опрысканных 2,4-D (2,185 г кэ/га и 4,370 г кэ/га), сравнивали с необработанными контрольными линиями кукурузы (например, неопрысканными 2,4-D). Результаты этих экспериментов дополнительно подтверждают то, что растения кукурузы, содержащие трансген AAD-1, не приводят к значимому повышению урожайности в результате опрыскивания 2,4-D. Для сравнения, для некоторых генотипов сои показано повышение урожайности после обработки 2,4-D. Наблюдаемое повышение урожайности у генотипов сои, демонстрируемое после обработки 2,4-D, является неожиданным усовершенствованием, применимым для повышения урожайности сельскохозяйственных культур. Описываемый способ можно распространять на использование обработки 2,4-D для повышения урожайности трансгенных сельскохозяйственных культур, например, экспрессирующих ген AAD-12.
Хотя изложенное выше изобретение для ясности понимания довольно подробно описывали с помощью иллюстраций и примеров, будет очевидно, что конкретные изменения и модификации можно осуществлять на практике в объеме заявленной формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СОБЫТИЕ 416 AAD-12, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ СОИ И ИХ СОБЫТИЕСПЕЦИФИЧНАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2603252C2 |
| УСТОЙЧИВЫЕ К ГЛИФОСАТУ РАСТЕНИЯ И СПОСОБЫ, СВЯЗАННЫЕ С НИМИ | 2013 |
|
RU2627169C2 |
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2721122C2 |
| УСТОЙЧИВЫЕ К ГЛИФОСАТУ РАСТЕНИЯ И СПОСОБЫ, СВЯЗАННЫЕ С НИМИ | 2013 |
|
RU2636037C2 |
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2634411C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| СОБЫТИЕ DAS-40278-9 AAD-1, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ КУКУРУЗЫ И ИХ СОБЫТИЕ-СПЕЦИФИЧЕСКАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2614120C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2632648C2 |







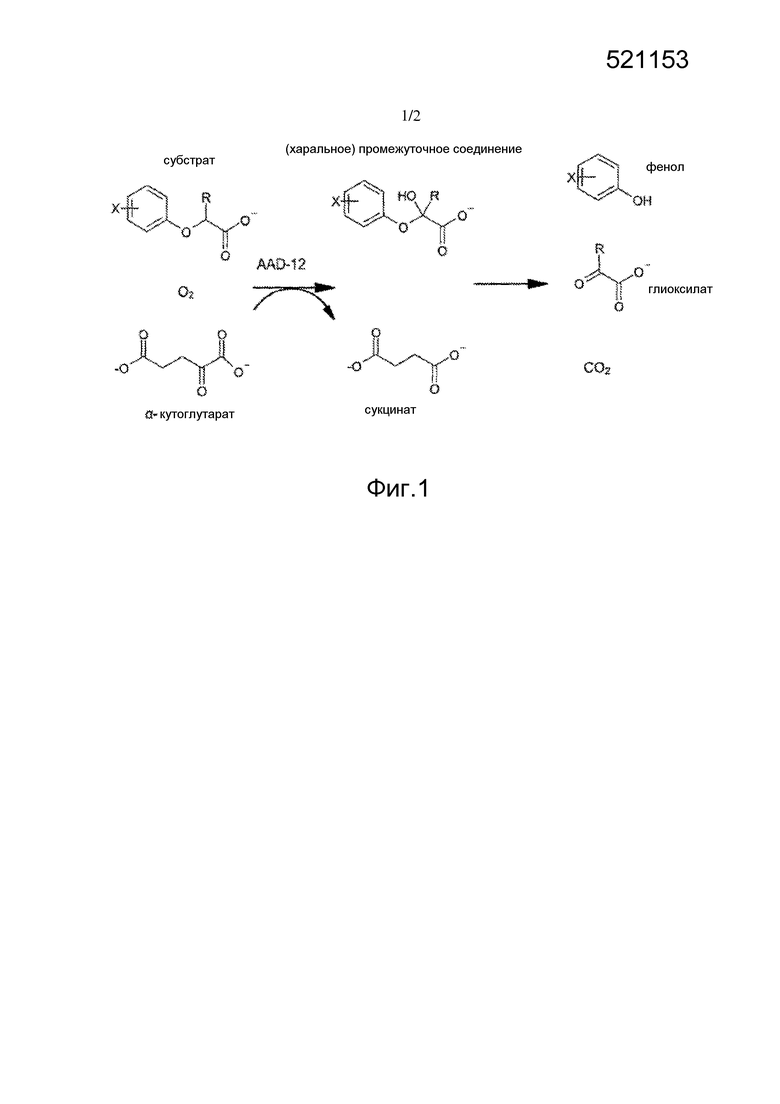

Изобретение относится к сельскому хозяйству. Для повышения урожайности резистентного к 2,4-D растения сои его обрабатывают 2,4-D на стадии роста V3 и R2 при норме внесения от 1000 до 2000 г кэ/га. Для повышения урожайности резистентного к 2,4-D растения сои осуществляют: (а) трансформацию клеток сои молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую арилоксиалканоатдиоксигеназу (AAD); (b) селекцию трансформированных клеток; (c) регенерацию растений сои из трансформированных клеток; и (d) применение 2,4-D на стадии роста V3 и R2 резистентного к 2,4-D растения сои, которое обрабатывают 2,4-D при норме внесения от 1000 до 2000 г кэ/га. Изобретение позволяет повысить урожайность сои. 3 н. и 15 з.п. ф-лы, 3 ил., 44 табл., 24 пр.
1. Способ повышения урожайности резистентного к 2,4-D растения сои, включающий применение 2,4-D на стадии роста V3 и R2 резистентного к 2,4-D растения сои, где резистентное к 2,4-D растение сои обрабатывают 2,4-D при норме внесения от 1000 до 2000 г кэ/га.
2. Способ по п. 1, где обработку осуществляют по меньшей мере три раза.
3. Способ по п. 1, где резистентное к 2,4-D растение сои находится в состоянии стресса.
4. Способ по п. 1, где резистентное к 2,4-D растение сои для борьбы с сорняками также обрабатывают гербицидом, иным, чем 2,4-D.
5. Способ по п. 4, где гербицид, иной, чем 2,4-D, является фосфорным гербицидом или арилоксифеноксипропионовым гербицидом.
6. Способ по п. 5, где фосфорный гербицид содержит глифосат, глуфосинат, их производные или их комбинации.
7. Способ по п. 5, где фосфорный гербицид находится в форме соли аммония, соли изопропиламмония, изопропиламиновой соли или соли калия.
8. Способ по п. 5, где арилоксифеноксипропионовый гербицид содержит хлоразифоп, феноксапроп, флуазифоп, галоксифоп, квизалофоп, их производные или их комбинации.
9. Способ по п. 1, где 2,4-D попадает в резистентное к 2,4-D растение сои посредством поглощения корнями.
10. Способ по п. 5, где фосфорный гербицид попадает в резистентное к 2,4-D растение сои посредством поглощения корнями.
11. Способ по п. 5, где арилоксифеноксипропионовый гербицид попадает в резистентное к 2,4-D растение сои посредством поглощения корнями.
12. Способ повышения урожайности резистентного к 2,4-D растения сои, включающий
(а) трансформацию клеток сои молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую арилоксиалканоатдиоксигеназу (AAD);
(b) селекцию трансформированных клеток;
(c) регенерацию растений сои из трансформированных клеток; и
(d) применение 2,4-D на стадии роста V3 и R2 резистентного к 2,4-D растения сои, где резистентное к 2,4-D растение сои обрабатывают 2,4-D при норме внесения от 1000 до 2000 г кэ/га.
13. Способ по п. 12, где арилоксиалканоатдиоксигеназа (AAD) является AAD-12.
14. Способ по п. 12, где молекула нуклеиновой кислоты содержит селективный маркер, не являющийся арилоксиалканоатдиоксигеназой (AAD).
15. Способ по п. 14, где селективный маркер является геном фосфинотрицинацетилтрансферазы (pat) или геном резистентности к биалафосу (bar).
16. Способ по п. 12, где молекула нуклеиновой кислоты оптимизирована для растений сои.
17. Применение 2,4-D в производстве трансгенных растений с резистентностью к 2,4-D с повышенной урожайностью по сравнению с их нетрансгенными родительскими растениями,
где трансгенным растением является резистентным к 2,4-D растением сои,
где 2,4-D применяют на стадии роста V3 и R2 резистентного к 2,4-D растения сои и где резистентное к 2,4-D растение сои обрабатывают 2,4-D при норме внесения от 1000 до 2000 г кэ/га.
18. Применение по п. 17, где 2,4-D содержит 2,4-D-холин или 2,4-D-диметиламин (DMA).
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Прибор для наведения канцелярских и архивных справок | 1929 |
|
SU14775A1 |

