Уровень техники
Ген aad-1 (первоначально выделенный из Sphingobium gerbicidovorans) кодирует белок арилоксиалканоатдиоксигеназы (AAD-1). Данный признак сообщает устойчивость к гербицидам на основе 2,4-дихлорфеноксиуксусной кислоты и арилоксифеноксипропионата (обычно определяемым как ”фоп” гербициды, такие как физалофоп) и может быть использован в качестве селектируемого маркера в процессе трансформации растений и в селекционных питомниках. Ген aad-1 в качестве гена, сообщающего растениям устойчивость к гербицидам, был впервые раскрыт в публикации WO 2005/107437 (см. также US 2009-0093366).
Экспрессия гетерологичных или чужеродных генов в растениях зависит от места инсерции чужеродного гена в хромосому. Данное явление может быть связано, например, со структурой хроматина (например, гетерохроматина) или с близостью расположения элементов, регулирующих транскрипцию (например, энхансеров), от сайта интеграции (Weising et al., Ann. Rev. Genet 22:421-477, 1988). Один и тот же ген в трансгенном растении (или другом организме) одинакового типа может характеризоваться большой изменчивостью уровня экспрессии по сравнению с другими генами. Кроме того, могут иметь место различия в пространственных или временных паттернах экспрессии. Например, различия в относительной экспрессии трансгена в разных тканях растения могут не соответствовать паттернам, ожидаемым от регулирующих транскрипцию элементов во введенной генной конструкции.
Таким образом, часто приходится создавать и исследовать большое число вариантов, чтобы идентифицировать вариант, экспрессирующий введенный для определенной цели ген на удовлетворительном уровне. Для достижения коммерческих целей обычно продуцируют сотни и тысячи разных вариантов, чтобы обнаружить единственный вариант, характеризующийся требуемыми уровнями и паттернами экспрессии трансгена. Вариант, характеризующийся требуемыми уровнями и/или паттернами экспрессии трансгена, имеет важное значение для интрогрессии трансгена в другие генетические среды путем полового ауткроссинга стандартными методами селекции. Потомство таких гибридов сохраняет характеристики экспрессии трансгена первичного трансформанта. Такой метод используют для достижения надежной экспрессии гена в ряде сортов, хорошо адаптированных к локальным условиям выращивания.
Заявки на патент США 20020120964 А1 и 20040009504 А1 относятся к варианту PV-GHGT07(1445) хлопчатника, способам его обнаружения и композициям для осуществления указанных способов. Заявка WO 02/100163 относится к варианту MONI5985 хлопчатника, способам его обнаружения и композициям для осуществления указанных способов. Заявка WO 2004/011601 относится к варианту MON863 кукурузы, способам его обнаружения и композициям для осуществления указанных способов. Заявка WO 2004/072235 относится к варианту MON88913 хлопчатника, способам его обнаружения и композициям для осуществления указанных способов.
Заявка WO 2006/098952 относится к варианту 3272 кукурузы. Заявка WO 2007/142840 относится к варианту MIR162 кукурузы.
Патент США № 7179965 относится к хлопчатнику, включающему вариант cry1F и вариант cry1Ac.
Настоящее изобретение относится к кукурузе AAD-1, включающей специфический вариант, ранее не описанный в научных публикациях.
Сущность изобретения
Настоящее изобретение относится к варианту кукурузы AAD-1, получившему название DAS-40278-9, семена которой депонированы в Американскую коллекцию типовых культур (АТСС) под номером доступа РТА-10244, и к потомству указанной кукурузы. Другими объектами настоящего изобретения являются последующие поколения растений, семена и зерна или регенерируемые части растений и семян и потомство кукурузы, включающей вариант DAS-40278-9, а также изготовленные из них пищевые или кормовые продукты. В объем настоящего изобретения также входят части растений кукурузы, содержащей вариант DAS-40278-9, которые включают, не ограничиваясь ими, пыльцу, семяпочки, цветки, побеги, корни и листья, ядра вегетативных клеток, клетки пыльцы и яйцеклетки. Настоящее изобретение далее относится к растениям кукурузы, обладающим устойчивостью к гербицидам на основе феноксиуксусной кислоты и/или арилоксиалканоата, к новым генетическим композициям варианта DAS-40278-9 кукурузы и некоторым аспектам агрономической продуктивности растений кукурузы, включающих вариант DAS-40278-9.
Настоящее изобретение частично относится к селекции растений и к растениям, устойчивым к гербицидам. Настоящее изобретение относится к новому варианту трансформации гена aad-1 в растениях кукурузы, включающему полинуклеотидную последовательность по настоящему изобретению, которая введена в определенный сайт генома клетки кукурузы.
В некоторых вариантах осуществления изобретения указанный вариант/полинуклеотидная последовательность могут быть объединены с другими признаками, включающими, например, другие гены устойчивости к гербицидам и/или инсектицидные белки. Однако настоящее изобретение относится к растениям, включающим один вариант, рассмотренный в настоящем описании изобретения.
Дополнительные признаки могут быть введены в геном растения в результате селекции растений, повторной трансформации трансгенного растения, содержащего вариант DAS-40278-9 кукурузы, или добавления новых признаков путем направленной интеграции методом гомологичной рекомбинации.
Другие варианты осуществления изобретения относятся к удалению полинуклеотидных последовательностей, включающих вариант DAS-40278-9 кукурузы, в том числе, например, кассету экспрессии гена pat. В результате удаления полинуклеотидной последовательности модифицированный вариант может быть перенацелен в определенный сайт хромосомы, в котором дополнительные полинуклеотидные последовательности объединены с вариантом DAS-40278-9 кукурузы.
Один вариант осуществления настоящего изобретения относится к хромосомному сайту-мишени кукурузы, расположенному на хромосоме 2 на расстоянии примерно 20 сМ между маркером UMC1265 (см. SEQ ID NO:30 и SEQ ID NO:31) и маркером ММС0111 (см. SEQ ID NO:32 и SEQ ID NO:33) простых повторов последовательности (SSR), на расстоянии примерно 20 сМ на карте сцепления генов кукурузы DAS 2008, где сайт-мишень включает гетерологичную нуклеиновую кислоту. Другой вариант осуществления настоящего изобретения относится к сайту-мишени хромосомного набора кукурузы, локализованному в SEQ ID NO:29, и к остаткам указанного сайта, рассмотренным в настоящем описании изобретения и известным специалисту в данной области.
Один вариант осуществления настоящего изобретения относится к способу создания трансгенного растения кукурузы, который включает введение гетерологичной нуклеиновой кислоты в определенное положение хромосомы 2 на расстоянии примерно 20 сМ между маркером UMC1265 (см. SEQ ID NO:30 и SEQ ID NO:31) и маркером ММС0111 (см. SEQ ID NO:32 и SEQ ID NO:33) SSR, на расстоянии примерно 20 сМ на карте сцепления генов кукурузы DAS 2008. В другом варианте осуществления изобретения инсертированная гетерологичная нуклеиновая кислота фланкирована у 5'-конца всей или частью 5'-концевой фланкирующей последовательности, определяемой в настоящем описании изобретения со ссылкой на SEQ ID NO:29, и фланкирована у 3'-конца всей или частью 5'-концевой фланкирующей последовательности, определяемой в настоящем описании изобретения со ссылкой на SEQ ID NO:29.
Кроме того, настоящее изобретение относится к анализам, предназначенным для обнаружения данного варианта в образце (например, в зерне кукурузы). Указанные анализы могут быть выполнены на основе последовательности ДНК рекомбинантной конструкции, инсертированной в геном кукурузы, и на основе геномных последовательностей, фланкирующих сайт инсерции. В объем настоящего изобретения также входят наборы для выполнения указанных анализов и условия их использования.
Таким образом, настоящее изобретение частично относится к клонированию и анализу последовательностей ДНК полноразмерной вставки AAD-1 и ее краевых областей (в линиях трансгенной кукурузы). Указанные последовательности являются уникальными. На основании указанной вставки и краевых последовательностей были созданы вариант-специфические праймеры. Анализ методом ПЦР показал, что указанные варианты могут быть идентифицированы в результате анализа ампликонов ПЦР, созданных с использованием наборов вариант-специфических праймеров. Таким образом, указанные и другие методы могут быть использованы для идентификации линий кукурузы, включающих вариант по настоящему изобретению.
Краткое описание чертежей
На фигуре 1 показана карта плазмиды pDAS1740.
На фигуре 2 показаны компоненты вставки для DAS-40278-9 (pDAS1740).
На фигуре 3 показана рестрикционная карта и компоненты вставки для DAS-40278-9 (pDAS1740).
На фигуре 4 показаны ампликоны, праймеры и метод клонирования ДНК-вставки и краевых последовательностей для DAS-40278-9.
На фигуре 5 показаны местоположения праймеров по отношению к вставке и краевым последовательностям для DAS-40278-9.
На фигуре 6 показаны соединительные области и инсерция для DAS-40278-9.
На фигуре 7 показана схема селекции, описанная в примере 7.
Краткое описание последовательностей
SEQ ID NO:1-28 представляют собой праймеры по настоящему изобретению.
SEQ ID NO:29 представляет собой вставку и фланкирующие последовательности для варианта DAS-40278-9.
SEQ ID NO:30-33 представляют собой праймеры для фланкирующих маркерных последовательностей, описанных в примере 4.
Подробное описание изобретения
Настоящее изобретение частично относится к селекции растений и к растениям, устойчивым к гербицидам. Настоящее изобретение относится к новым вариантам трансформации растений кукурузы (маиса), включающим полинуклеотидные последовательности гена aad-1 по настоящему изобретению, инсертированные в определенный сайт генома клетки кукурузы. В некоторых вариантах осуществления изобретения указанная полинуклеотидная последовательность, например, может быть объединена с другими признаками (такими как другие гены устойчивости к гербицидам и/или гены, кодирующие инсектицидные белки). В некоторых вариантах осуществления изобретения указанные полинуклеотидные последовательности могут быть удалены и затем направленно связаны с дополнительными полинуклеотидными последовательностями. Однако настоящее изобретение относится к растениям, включающим один вариант по настоящему изобретению.
Кроме того, настоящее изобретение относится к анализам, предназначенным для обнаружения в образце варианта по настоящему изобретению. В объем настоящего изобретения входят способы создания и/или продуцирования любых диагностических молекул нуклеиновых кислот, представленных или предложенных в настоящем описании изобретения, в основе которых, в частности, лежит полное или частичное использование фланкирующих последовательностей по настоящему изобретению.
В частности, настоящее изобретение частично относится к варианту DAS-40278-9 трансгенной кукурузы (известному также как pDAS1740-278), линиям растений, включающим указанные варианты, клонированию и анализу последовательностей ДНК указанной вставки и/или ее краевых областей. Линии растений по настоящему изобретению могут быть обнаружены при помощи последовательностей, рассмотренных и предложенных в настоящем описании изобретения.
Некоторые варианты осуществления изобретения относятся к линиям кукурузы, устойчивой к гербицидам, и способам их идентификации. Настоящее изобретение частично относится к обнаружению варианта по настоящему изобретению с целью определения наличия представляющего интерес варианта в потомстве гибрида. Кроме того, в объем настоящего изобретения входит способ обнаружения данного варианта, выполнение которого необходимо, например, для соответствия положениям, требующим предпродажного утверждения и маркировки продуктов, полученных, например, из растений рекомбинантной кукурузы. Вариант по настоящему изобретению можно обнаружить любым хорошо известным методом обнаружения нуклеиновой кислоты, таким как полимеразная цепная реакция (ПЦР) или гибридизация ДНК с помощью зондов нуклеиновой кислоты. Вариант-специфический анализ методом ПЦР описан, например, в публикации Windels et al. (Med. Fac. Landbouww, Univ. Gent 64/5b:459462, 1999). В данной публикации описана идентификация варианта устойчивости к глифосату 40-3-2 сои методом ПЦР с использованием набора праймеров, заполняющих соединение между вставкой и фланкирующей ДНК. В частности, один праймер включал последовательность из вставки и второй праймер включал последовательность из фланкирующей ДНК.
Кукуруза была модифицирована путем введения гена aad-1 из Sphingobium herbicidovorans, кодирующего белок арилоксиалканоатдиоксигеназы (AAD-1). Данный признак сообщает устойчивость к гербицидам на основе 2,4-дихлорфеноксиуксусной кислоты и арилоксифеноксипропионата (обычно определяемых как ”фоп” гербициды, такие как хизалофоп) и может быть использован в качестве селектируемого маркера в процессе трансформации растения и в селекционных питомниках. Трансформация кукурузы при помощи фрагмента ДНК из плазмиды pDAS1740 была выполнена путем селекции с целью создания варианта DAS-40278-9.
Образцы геномной ДНК, выделенные из двадцати растений кукурузы, полученных из пяти поколений и четырех растений в каждом поколении варианта DAS-40278-9, были отобраны для молекулярного исследования варианта DAS-40278-9 кукурузы AAD-1. Экспрессию белка AAD-1 исследовали при помощи специального набора для экспресс-анализа AAD-1. Для последующего молекулярного исследования были отобраны только растения с положительной экспрессией белка AAD-1. Анализ гибридизации методом саузерн-блоттинга подтвердил наличие гена aad-1 в растениях кукурузы с положительной экспрессией белка AAD-1, который был инсертирован в виде одной интактной копии в указанные растения в процессе гибридизации с зондом гена aad-1.
В настоящем описании изобретения также рассмотрено молекулярное исследование ДНК, инсертированной в вариант DAS-40278-9 кукурузы AAD-1. Указанный вариант был получен методом точечной трансформации с использованием фрагмента FspI плазмиды pDAS1740. Анализ методом саузерн-блоттинга был выполнен для определения паттерна интеграции инсертированного фрагмента ДНК и выявления числа вставок/копий гена aad-1 в варианте DAS-40278-9. Были получены данные, свидетельствующие об интеграции и целостности трансгена aad-1, инсертированного в геном кукурузы. Была исследована интеграция некодирующих областей (предназначенных для регуляции кодирующих областей), таких как промоторы и терминаторы, области присоединения матрицы, RB7 Mar v3 и RB7 Mar v4, а также устойчивость вставки трансгена в нескольких поколениях. Устойчивость инсертированной ДНК была продемонстрирована в пяти поколениях растений. Кроме того, с помощью зондов, охватывающих почти всю область остова, фланкирующую сайты рестрикции (FspI) плазмиды pDAS1740, было продемонстрировано отсутствие последовательности области остова трансформирующей плазмиды, включающей ген устойчивости к ампициллину (Ap'). Подробная физическая карта инсерции была создана на основании анализов варианта DAS-40278-9, выполненных методом саузерн-блоттинга.
В тканях кукурузы определяли уровни белка AAD-1. Кроме того, был выполнен анализ состава стеблей и зерен кукурузы для исследования соответствия между линией изогенной нетрансформированной кукурузы и линией трансформированной кукурузы, включающей вариант DAS-40278-9 (неопрысканная кукуруза, кукуруза, опрысканная 2,4-D, опрысканная хизалофопом и опрысканная 2,4-D и хизалофопом). Агрономические характеристики линии изогенной нетрансформированной кукурузы также сравнивали с кукурузой, включающей вариант DAS-40278-9.
Экспрессию, состав питательных веществ и агрономические показатели нетрансгенной контрольной кукурузы и линии гибридной кукурузы, содержащей арилоксиалканоатдиоксигеназу-1 (AAD-1), исследовали в полевых условиях в один и тот же год на шести участках, расположенных в штатах Айова, Иллинойс (2 участка), Индиана, Небраска и в провинции Онтарио, Канада. В настоящем описании изобретения суммированы уровни экспрессии белка AAD-1 в листьях, пыльце, корнях, стеблях, целом растении и зерне и представлены результаты определения агрономических показателей и анализа состава образцов стеблей и зерна контрольной кукурузы и кукурузы AAD-1, включающей вариант DAS-40278-9.
Растворимый экстрагируемый белок AAD-1 измеряли при помощи количественного твердофазного иммуноферментного анализа (ELISA) в листьях, пыльце, корнях, стеблях, целом растении и зерне кукурузы. Хорошие средние значения экспрессии были получены в тканях корней и пыльцы, которые более подробно рассмотрены в настоящем описании изобретения. Значения экспрессии были аналогичны для всех обработок опрыскиванием, а также для участков, которые не опрыскивали и опрыскивали гербицидом 2,4-D и хизалофопом.
Анализы состава, в том числе общих показателей, минеральных веществ, аминокислот, жирных кислот, витаминов, вредных веществ и вторичных метаболитов выполняли для исследования соответствия между кукурузой AAD-1, включающей вариант DAS-40278-9, (обработанной и необработанной гербицидами) и контрольной кукурузой. Результаты, полученные для состава всех образцов кукурузы AAD-1, включающей вариант DAS-40278-9, соответствовали или превосходили (в биологическом и агрономическом отношении) результаты, полученные для контрольных линий и/или обычной кукурузы, при выполнении анализа агрономических данных для участков, на которых произрастала контрольная кукуруза и кукуруза AAD-1, включающая вариант DAS-40278-9.
Как было указано выше в разделе ”Уровень техники”, введение и интеграция трансгена в геном растения вызывает появление некоторых случайных вариантов (отсюда название ”вариант” для данной экспрессируемой инсерции). То есть при использовании многих методов трансформации, таких как трансформация Agrobacterium, ”генное ружье” и точечная трансформация, невозможно предсказать, будет ли введен трансген в геном. Таким образом, идентификация фланкирующей геномной ДНК растения с обеих сторон вставки может иметь важное значение для выявления растения, имеющего данный вариант инсерции. Например, можно создать праймеры для ПЦР, позволяющие получить ампликон ПЦР в области соединения вставки и генома-хозяина. Указанный ампликон ПЦР может быть использован для идентификации уникального или отличающегося типа варианта инсерции.
Так как ”варианты” первоначально являются случайными вариантами, то в качестве части настоящего описания изобретения по меньшей мере 2500 семян линии кукурузы, включающей данный вариант, были депонированы, став достоянием общественности без каких-либо ограничений (но в соответствии с патентными правами), в Американскую коллекцию типовых культур (АТСС) по адресу 10801 University Boulevard, Manassas, VA, 20110. Указанному депозиту был присвоен номер АТСС РТА-10244 (семена гибрида кукурузы Yellow Dent (Zea Mays L.): DAS-40278-9; депонированные компанией Dow AgroScience LLC; дата поступления семян/штамма в АТСС: 10 июля 2009; жизнеспособность семян подтверждена 17 августа 2009). Данный депозит был сделан и будет храниться в соответствии с условиями Будапештского договора о международном признании депонирования семян для целей патентной процедуры. Данный депозит будет храниться без каких-либо ограничений в депозитарии АТСС, который является общедоступным депозитарием, в течение 30 лет или в течение пяти лет после самого последнего запроса, или в течение эффективного времени действия патента в зависимости от того, какой из указанных периодов времени является наиболее продолжительным, и будет заменен, если станет нежизнеспособным в течение указанного срока.
Депонированные семена являются частью настоящего изобретения. Совершенно ясно, что из указанных семян могут быть выращены растения кукурузы, и такие растения также являются частью настоящего изобретения. Настоящее изобретение также относится к последовательностям ДНК в указанных растениях кукурузы, которые пригодны для обнаружения таких растений и их потомства. Методы обнаружения и наборы по настоящему изобретению могут быть использованы для идентификации любого одного, двух или даже всех трех указанных вариантов в зависимости от конечной цели исследования.
Приведенные в настоящем описании изобретения определения терминов и примеры помогают более подробно описать настоящее изобретение и предоставляют специалистам в данной области необходимые указания для осуществления изобретения. За исключением особо оговоренных случаев термины имеют общепринятые значения, известные специалистам в данной области. Номенклатура оснований ДНК использована в соответствии с разделом 37 Свода федеральных правил, §1.822.
В использованном здесь значении термин ”потомство” означает потомство любого поколения родительского растения, включающее вариант DAS-40278-9 кукурузы AAD-1.
Трансгенный ”вариант” получают путем трансформации растительных клеток гетерологичной ДНК, то есть конструкцией нуклеиновой кислоты, включающей представляющий интерес трансген, регенерации популяции растений, полученной в результате инсерции указанного трансгена в геном растения, и отбора конкретного растения, характеризующегося наличием инсерции в определенном положении в геноме. Термин ”вариант” относится к первичному трансформанту и потомству указанного трансформанта, включающему гетерологичную ДНК. Термин ”вариант” также относится к потомству, полученному в результате ауткроссинга трансформанта с другим сортом, включающим геномную/трансгенную ДНК. Даже после повторного обратного скрещивания с родительской формой, с которой гибрид скрещивается вновь, введенная трансгенная ДНК и фланкирующая геномная ДНК (геномная/трансгенная ДНК) из трансформированной родительской формы присутствует в потомстве гибрида в том же положении в хромосоме. Термин ”вариант” также относится к ДНК первичного трансформанта и его потомства, включающего введенную ДНК и фланкирующую геномную последовательность рядом с введенной ДНК, которые должны быть переданы потомству, включая представляющий интерес трансген, в результате скрещивания одной родительской формы, включающей введенную ДНК (например, первичный трансформант и потомство, полученное в результате самоопыления), с родительской формой, не содержащей введенную ДНК.
“Соединительная последовательность” заполняет участок, на котором ДНК, введенная в геном, связывается с ДНК нативного генома кукурузы, фланкирующей участок инсерции, при этом для диагностики варианта достаточно идентифицировать или обнаружить одну или несколько соединительных последовательностей в генетическом материале растения. К таким последовательностям относятся последовательности ДНК, заполняющие места вставки вариантов кукурузы по настоящему изобретению, и фланкирующие ДНК одинаковой длины. В настоящем описании изобретения приведены конкретные примеры таких диагностических последовательностей; однако, другие последовательности, которые перекрывают места соединения инсерций или места соединения инсерций и геномной последовательности, также являются диагностическими и могут быть использованы в соответствии с настоящим изобретением.
Настоящее изобретение относится к идентификации таких фланкирующих, соединительных последовательностей и последовательностей вставок. В объем настоящего изобретения входят соответствующие праймеры и ампликоны ПЦР. В соответствии с настоящим изобретением для обнаружения или идентификации промышленных сортов трансгенной кукурузы или линий, выделенных из линий трансгенной кукурузы, являющихся частной собственностью, могут быть использованы методы ПЦР с применением ампликонов, расположенных рядом с введенной ДНК и ее краевыми последовательностями.
Полные последовательности всех указанных вставок наряду с частями соответствующих фланкирующих последовательностей представлены в настоящем описании изобретения в виде SEQ ID NO:29. Ниже приведены координаты вставки и фланкирующих последовательностей каждого варианта, относящегося в SEQ ID NO:29 (всего 8557 пар оснований). Данный вопрос более подробно рассмотрен в примере 3.8.
Данный инсерционный вариант и его компоненты далее проиллюстрированы на фигурах 1 и 2. Указанные последовательности (в частности, фланкирующие последовательности) являются уникальными. На основании указанной вставки и краевых последовательностей были созданы вариант-специфические праймеры. Анализ методом ПЦР показал, что указанные линии кукурузы могут быть идентифицированы в разных генотипах кукурузы путем анализа ампликонов ПЦР, созданных при помощи наборов вариант-специфических праймеров. Таким образом, для идентификации указанных линий кукурузы могут быть использованы вышеуказанные и другие родственные методы. Последовательности, идентифицированные указанными методами, являются уникальными. Например, поиск с помощью программы BLAST в базах данных GENBANK, не выявил какой-либо значительной гомологии между клонированными краевыми последовательностями и последовательностями в базе данных.
Методы обнаружения по настоящему изобретению особенно полезны в сочетании с селекцией растений для определения потомства растений, включающего данный вариант, который был получен после скрещивания родительского растения, включающий представляющий интерес вариант, с другой линией растений с целью сообщения указанному потомству одного или нескольких дополнительных признаков. Указанные методы ПЦР позволяют эффективно разрабатывать программы селекции кукурузы и контролировать качество, в частности, промышленных семян трансгенной кукурузы. В настоящее время также могут быть созданы и использованы наборы для обнаружения методом ПЦР указанных линий трансгенной кукурузы. Указанные методы также позволяют регистрировать и хранить продукты.
Кроме того, фланкирующие/геномные последовательности кукурузы могут быть использованы для специфической идентификации локализации каждой вставки в геноме. Такая информация может быть использована для создания систем молекулярных маркеров, специфичных к каждому варианту. Указанные последовательности могут быть использованы для ускоренной селекции и получения данных о сцеплении генов.
Информация о фланкирующих последовательностях может быть далее использованы для изучения и исследования процессов интеграции трансгенов, определения сайтов для интеграции трансгенов в геноме, сортировки вариантов, определения устойчивости трансгенов и их фланкирующих последовательностей и экспрессии генов (особенно в отношении сайленсинга генов, паттернов метилирования трансгенов, влияния положения и возможных элементов, определяющих экспрессию, таких как MAR [области присоединения матрицы] и тому подобных).
В свете настоящего описания изобретения должно быть ясно, что в объем настоящего изобретения входят семена, депонированные в АТСС под номером РТА-10244. Настоящее изобретение также относится к растению кукурузы, устойчивому к гербицидам, выращенному из семени, депонированного в АТСС под номером доступа РТА-10244. Настоящее изобретение далее относится к частям указанного растения, таким как листья, образцы тканей, семена, произведенные указанным растением, пыльца и тому подобные.
Настоящее изобретение далее относится к происхождению и/или потомству растений, выращенных из депонированных семян, предпочтительно к растению кукурузы, устойчивому к гербицидам, геном которого включает обнаруживаемую последовательность, соединяющую геномную ДНК дикого типа/вставочную ДНК, по настоящему изобретению. В использованном здесь значении термин ”кукуруза” означает маис (Zea mays), и в определение данного термина входят все сорта, которые могут быть получены в результате селекции кукурузы.
Настоящее изобретение далее относится к способам получения гибридов при использовании растения по настоящему изобретению в качестве по меньшей мере одной родительской формы. Например, настоящее изобретение относится к гибридному растению F1, имеющему в качестве одной или обеих родительских форм любые растения по настоящему изобретению. Кроме того, в объем настоящего изобретения входит семя, произведенное такими гибридами F1 по настоящему изобретению. Настоящее изобретение относится к способу получения семени гибрида F1 путем скрещивания указанного растения с другим (например, инбредной родительской линией) растением и получения гибридного семени. Настоящее изобретение относится к указанному растению, которое является материнской или отцовской родительской формой. Характеристики полученных растений могут быть улучшены в результате тщательного отбора родительских растений.
Растение кукурузы, устойчивой к гербицидам, может быть получено в результате первоначального скрещивания первого родительского растения кукурузы, выращенного из семени любой линии по настоящему изобретению, и второго родительского растения кукурузы, что позволяет получить множество растений первого потомства; и затем отбора растения первого потомства, устойчивого к гербицидам (или содержащего по меньшей мере один из вариантов по настоящему изобретению); и самоопыления растения первого потомства, что позволяет получить множество растений второго потомства; и затем отбора из растений второго потомства одного растения, устойчивого к гербицидам (или содержащего по меньшей мере один из вариантов по настоящему изобретению). Указанные стадии могут далее включать обратное скрещивание растения первого потомства или растения второго потомства со вторым родительским растением кукурузы или третьим родительским растением кукурузы. Затем могут быть посеяны семена кукурузы по настоящему изобретению или их потомство.
Следует также отметить, что могут быть скрещены два разных трансгенных растения с целью получения потомства, содержащего два независимо добавленных экзогенных гена. В результате самоопыления соответствующего потомства могут быть получены растения, которые являются гомозиготными для обоих добавленных экзогенных генов. В объем настоящего изобретения также входит обратное скрещивание родительского растения и ауткроссинг с нетрансгенным растением в качестве вегетативного размножения. В данной области известны другие методы селекции, обычно используемые для сообщения растениям разных признаков и получения сельскохозяйственных культур. Селекцию методом обратного скрещивания используют для переноса генов с целью создания наследственного и хорошо наследуемого признака в требуемом гомозиготном культиваре или инбредной линии, являющейся родительской формой, с которой гибрид скрещивается вновь. Источник передаваемого признака именуется родителем-донором. Предполагается, что полученное растение должно иметь признаки родительской формы, с которой гибрид скрещивается вновь, (то есть культивара) и требуемый признак, перенесенный от родителя-донора. После первичного скрещивания отбирают растения, обладающие фенотипом родителя-донора, и повторно скрещивают (обратное скрещивание) с родительской формой, с которой гибрид скрещивается вновь. Предполагается, что полученная родительская форма должна иметь признаки родительской формы, с которой гибрид скрещивается вновь, (например, культивара) и требуемый признак, перенесенный от родителя-донора.
Молекулы ДНК по настоящему изобретению могут быть использованы в качестве молекулярных маркеров при осуществлении метода селекции с использованием маркеров (МАВ). Молекулы ДНК по настоящему изобретению (такие как маркеры AFLP, маркеры RFLP, маркеры RAPD, SNP и SSR) могут быть использованы в методах, позволяющих идентифицировать генетически связанные агрономически полезные признаки, известные в данной области. Признак устойчивости к гербицидам может быть прослежен в потомстве гибрида растения кукурузы по настоящему изобретению (или в его потомстве и в любом другом культиваре или сорте кукурузы) с помощью методов МАВ. Молекулы ДНК являются маркерами данного признака, и методы МАВ, хорошо известные в данной области, могут быть использованы для отслеживания признака устойчивости к гербицидам в растениях кукурузы, если по меньшей мере одна линия кукурузы по настоящему изобретению или ее потомство было родителем или предком. Способы по настоящему изобретению могут быть использованы для идентификации любого сорта кукурузы, имеющего вариант по настоящему изобретению.
Способы по настоящему изобретению включают получение растения кукурузы, устойчивого к гербицидам, путем селекции растения по настоящему изобретению. В частности, указанные способы могут включать скрещивание двух растений по настоящему изобретению или одного растения по настоящему изобретению и любого другого растения. Предпочтительные способы далее включают отбор потомства указанного гибрида путем исследования потомства в отношении наличия варианта, обнаруживаемого методами по настоящему изобретению. Например, настоящее изобретение может быть использовано для отслеживания варианта по настоящему изобретению на протяжении нескольких циклов селекции растений, включающих другие требуемые признаки, в частности, агрономические признаки, анализируемые в разных примерах, приведенных в настоящем описании изобретения. Растения, включающие вариант по настоящему изобретению и требуемый признак, например, могут быть обнаружены, идентифицированы, отобраны и быстро использованы в последующих циклах селекции. Вариант по настоящему изобретению/признак могут быть также объединены в процессе селекции и отслежены в соответствии с настоящим изобретением наряду с признаком устойчивости к воздействию насекомых и/или другими признаками устойчивости к гербицидам. Один предпочтительный вариант осуществления изобретения относится к растению, выключающему вариант по настоящему изобретению, объединенный с геном, кодирующим устойчивость к гербициду дикамба.
Таким образом, настоящее изобретение может быть, например, объединено с признаками, кодирующими устойчивость к глифосату (например, растения, устойчивые к бактериальным генам EPSPS, GOX, GAT), устойчивость к глюфозинату (например, Pat, bar), устойчивость к гербициду, ингибирующему ацетолактат-синтазу (ALS) (например, имидазолиноны [такие как имазетапир], сульфонилмочевины, триазолопиримидинсульфонанилид, пиримидинилтиобензоат и другие химические вещества [Csr1, SurA и т.д.]), устойчивость к бромоксинилу (например, Bxn), устойчивость к ингибиторам фермента HPPD (4-гидроксифенилпируватдиоксигеназа), устойчивость к ингибиторам фитоендезатуразы (PDS), устойчивость к гербицидам, ингибирующим фотосистему II (например, psbA), устойчивость к гербицидам, ингибирующим фотосистему I, устойчивость к гербицидам, ингибирующим протопорфириногеноксидазу IX (РРО) (например, РРО-1), устойчивость к гербицидам на основе фенилмочевины (например, CYP76B1), ферменты, расщепляющие гербицид дикамба (см., например, заявку на патент США 20030135879), и другими, которые могут быть использованы отдельно или в разных комбинациях, обеспечивая эффективное уничтожение или предотвращение распространения сорняков и/или устойчивость к любому гербициду вышеуказанных классов.
Что касается дополнительных гербицидов, то в качестве некоторых дополнительных предпочтительных ингибиторов ALS (известных также как AHAS) можно указать триазолопиримидинсульфонанилиды (такие как клоранзулам-метил, диклозулам, флоразулам, флуметзулам, метозулам и пеноксзулам), пиримидинилтиобензоаты (такие как биспирибак и пиритиобак) и флукарбазон. Некоторые предпочтительные ингибиторы HPPD включают мезотрион, изоксафлутол и сулкотрион. Некоторые предпочтительные ингибиторы РРО включают флумиклорак, флумиоксазин, флуфенпир, пирафлуфен, флутиацет, бутафенацил, карфентразон, сулфентразон и простые дифениловые эфиры (такие как ацифлуорфен, фомесафен, лактофен и оксифлуорфен).
Кроме того, AAD-1 отдельно или в сочетании с одним или несколькими дополнительными признаками НТС может быть объединен с одним или несколькими дополнительными первичными признаками (такими как, например, устойчивость к воздействию насекомых, устойчивость к грибам или устойчивость к стрессу и т.д.) или вторичными признаками (такими как, например, более высокая урожайность, лучший профиль выхода масла, лучшее качество клетчатки и т.д.). Таким образом, настоящее изобретение может быть использовано для создания полного агрономического набора улучшенных качеств культуры с возможностью гибкой и экономически эффективной борьбы с целым рядом сельскохозяйственных вредителей.
В данной области описаны методы введения полинуклеотидной последовательности в конкретный участок хромосомы растительной клетки путем гомологичной рекомбинации. Например, метод сайт-специфической интеграции, описанный в публикации заявки на патент США № 2009/0111188 А1, предполагает использование рекомбиназ или интеграз, опосредующих введение полинуклеотидной последовательности донора в хромосому-мишень. Кроме того, в международной заявке на патент № WO 2008/021207 описан метод гомологичной рекомбинации, опосредуемой цинкосодержащей пальцеобразной областью, для введения одной или нескольких полинуклеотидных последовательностей донора в определенные положения генома. Для введения полинуклеотидной последовательности в определенный участок хромосомы могут быть использованы рекомбиназы, такие как FLP/FRT, описанные в патенте США № 6720475, или CRE/LOX, описанные в патенте США № 5658772. И наконец, мегануклеазы, используемые для направленного введения донорских полинуклеотидов в определенный участок хромосомы, были описаны в публикации Puchta et al., PNAS USA 93 (1996) pp.5055-5060.
Хорошо известны и широко применяются разные другие методы сайт-специфической интеграции в растительные клетки (Kumor et al., Trands in Plant Sci. 6(4) (2001) pp.155-159). Кроме того, для модификации растений могут быть использованы системы сайт-специфической рекомбинации, идентифицированные в нескольких прокариотических и низших эукариотических организмах. Примеры таких систем включают, не ограничиваясь ими, систему рекомбиназ R/RS из плазмиды pSR1 дрожжей Zygosaccharomyces rouxii (Araki et al. (1985) J. Mol. Biol. 182:191-203) и систему Gin/gix фага Mu (Maeser and Kahlmann (1991) Mol. Gen. Genet. 230:170-176).
В некоторых вариантах осуществления настоящего изобретения может быть желательно интегрировать или присоединить новый трансген(ы) рядом с существующим трансгенным вариантом. Трансгенным вариантом может быть предпочтительный геномный локус, выбранный на основании уникальных характеристик, таких как единственный сайт инсерции, нормальное расщепление Менделиана, устойчивая экспрессия и лучшая комбинация эффективности, включающая устойчивость к гербицидам и агрономическую продуктивность в разных окружениях. Вновь интегрированные трансгены должны сохранять характеристики экспрессии трансгенов существующих трансформантов. Кроме того, разработаны анализы, предназначенные для обнаружения и подтверждения наличия вновь интегрированного варианта, которые позволяют идентифицировать фланкирующие последовательности и положение в хромосоме вновь интегрированного варианта. И наконец, введение нового трансгена в определенный участок хромосомы, связанный с существующим трансгеном, позволит ускорить интрогрессию трансгенов в другие генетические среды в результате ауткроссинга при помощи стандартных методов селекции.
В некоторых вариантах осуществления настоящего изобретения может быть желательно удалить полинуклеотидные последовательности из трансгенного варианта. Например, удаление трансгена, описанное в предварительной заявке на патент США № 61/297628, предполагает использование нуклеаз цинкосодержащей пальцеобразной области для удаления полинуклеотидной последовательности, состоящей из полигенной кассеты экспрессии, из введенного в хромосому трансгенного варианта. Удаляемая полинуклеотидная последовательность может быть селектируемым маркером. После эксцизии и удаления полинуклеотидной последовательности модифицированный трансгенный вариант может быть перенацелен путем инсерции полинуклеотидной последовательности. Эксцизия полинуклеотидной последовательности и последующее перенацеливание модифицированного трансгенного варианта создают такие преимущества как возможность повторного использования селектируемого маркера или устранения случайных изменений транскриптомы растения, возникающих в результате экспрессии специфических генов.
Настоящее изобретение относится к определенному сайту в хромосоме 2 генома кукурузы, который великолепно подходит для инсерции гетерологичных нуклеиновых кислот. Кроме того, в объем настоящего изобретения входят 5'-концевой молекулярный маркер, 3'-концевой молекулярный маркер, 5'-концевая фланкирующая последовательность и 3'-концевая фланкирующая последовательность, используемые для идентификации локализации сайта-мишени в хромосоме 2. Таким образом, настоящее изобретение относится к способам введения представляющих интерес гетерологичных нуклеиновых кислот в заранее определенный сайт-мишень или рядом с указанным сайтом-мишенью. Настоящее изобретение также относится к семени и/или растению кукурузы, включающим любую гетерологичную нуклеотидную последовательность, введенную в указанный сайт-мишень или в непосредственной близости от такого сайта. Одним способом осуществления такой целенаправленной интеграции является эксцизия и/или замена кассеты экспрессии гена pat другой вставкой по настоящему изобретению. Таким образом, целенаправленная гомологичная рекомбинация, например и без ограничений, может быть использована в соответствии с настоящим изобретением.
В использованном здесь значении термин ”стекинг” генов, вариантов или признаков означает объединение желаемых признаков в одной трансгенной линии. Селекционеры растений объединяют трансгенные признаки путем скрещивания родительских форм, каждая из которых обладает требуемым признаком и последующей идентификацией потомства, имеющего оба требуемых признака. Другим способом стекинга генов является одновременный перенос двух или более генов в ядро клетки растения в процессе трансформации. Другим способом стекинга генов является повторная трансформация трансгенного растения другим представляющим интерес геном. Например, стекинг генов может быть использован для объединения двух или более разных признаков, включающих, например, два или более разных признаков устойчивости к воздействию насекомых и признаков устойчивости к болезням, два или более признаков устойчивости к гербицидам и/или признаков устойчивости к воздействию насекомых и признаков устойчивости к гербицидам. Использование селектируемого маркера помимо представляющего интерес гена также может считаться стекингом генов.
Термин ”гомологичная рекомбинация” означает взаимодействие любой пары нуклеотидных последовательностей, имеющих соответствующие сайты, содержащие подобную нуклеотидную последовательность, с помощью которой две указанные нуклеотидные последовательности могут взаимодействовать (рекомбинировать) с образованием новой последовательности рекомбинантной ДНК. Сайты подобной нуклеотидной последовательности определяются в настоящем описании изобретения как ”гомологичная последовательность”. Частота гомологичной рекомбинации обычно возрастает с увеличением длины гомологичной последовательности. Таким образом, хотя гомологичная рекомбинация может происходить между двумя нуклеотидными последовательностями, которые являются недостаточно идентичными, частота (или эффективность) рекомбинации уменьшается при увеличении расхождения между двумя последовательностями. Рекомбинация может быть осуществлена при использовании одной гомологичной последовательности в донорской молекуле и молекуле-мишени, в результате чего образуется продукт рекомбинации ”одного кроссинговера”. Альтернативно в нуклеотидную последовательность-мишень и донорскую нуклеотидную последовательность могут быть введены две гомологичные последовательности. В результате рекомбинации двух гомологичных последовательностей донорской молекулы с двумя гомологичными последовательностями мишени образуется продукт рекомбинации ”двух кроссинговеров”. Если гомологичные последовательности в донорской молекуле фланкируют последовательность, предназначенную для манипуляции (например, представляющую интерес последовательность), рекомбинация молекулы-мишени двумя кроссинговерами позволяет получить продукт рекомбинации, в котором представляющая интерес последовательность заменяет последовательность ДНК, которая первоначально находилась между гомологичными последовательностями в молекуле-мишени. Обмен последовательностями ДНК между мишенью и донором в результате рекомбинации ”двумя кроссинговерами” именуется “заменой последовательности”.
Фермент AAD-1 по настоящему изобретению делает возможной экспрессию трансгена, определяющую устойчивость к комбинациям гербицидов, которые уничтожают почти все широколистные и травянистые сорняки. AAD-1 может быть великолепным признаком устойчивости культуры к гербициду (HTC), объединяемым с другими признаками HTC (такими как, например, устойчивость к глифосату, устойчивость к глюфозинату, устойчивость к имидазолинону, устойчивость к бромоксинилу и т.д.) и признаками устойчивости к воздействию насекомых (Cry1F, Cry1Ab, Cry34/45 и т.д.). Кроме того, AAD-1 может быть селектируемым маркером, помогающим отобрать первичные трансформанты растений, генетически созданные с использованием второго гена или группы генов.
Признаки НТС по настоящему изобретению могут быть использованы в новых комбинациях с другими признаками НТС (которые включают, не ограничиваясь ими, устойчивость к глифосату). Указанные комбинации признаков позволяют создать новые методы уничтожения сорняков (и подобных видов) благодаря вновь приобретенной или наследственной устойчивости к гербицидам (например, к глифосату). Таким образом, в объем настоящего изобретения входят, помимо признаков НТС, новые способы уничтожения сорняков при помощи гербицидов, в которых устойчивость к гербицидам была достигнута в результате введения указанного фермента в трансгенные культуры.
Кроме того, во всем мире преимущественно выращивают сельскохозяйственные культуры, устойчивые к глифосату. При многократном чередовании с другими сельскохозяйственными культурами, устойчивыми к глифосату, могут возникнуть трудности с уничтожением растений-самосевов, устойчивых к глифосату, в севооборотных культурах. Таким образом, использование трансгенных признаков по настоящему изобретению, совместно или отдельно введенных в сельскохозяйственные культуры, делает возможным уничтожение других растений-самосевов с признаками НТС.
Предпочтительное растение или семя по настоящему изобретению содержит в своем геноме последовательности вставок наряду по меньшей мере с 20-500 или большим числом смежных фланкирующих нуклеотидов с обеих сторон вставки. За исключением особо оговоренных случаев фланкирующие последовательности являются последовательностями, представленными в SEQ ID NO:29 (см. приведенную выше таблицу). SEQ ID NO:29 включает гетерологичную ДНК, введенную в первичный трансформант, и иллюстративные фланкирующие геномные последовательности, расположенные рядом с введенной ДНК. Указанные фланкирующие последовательности могут быть полностью или частично переданы потомству, получающему введенную ДНК в результате скрещивания родительской линии, включающей данный вариант.
Настоящее изобретение относится к культурам тканей регенерируемых клеток растения по настоящему изобретению. В объем настоящего изобретения также входит растение, регенерированное из такой культуры тканей, в частности, когда указанное растение может экспрессировать все морфологические и физиологические свойства представленного сорта. Предпочтительные растения по настоящему изобретению обладают всеми физиологическими и морфологическими свойствами растения, выращенного из депонированного семени. Настоящее изобретение далее относится к потомству такого семени и семени, обладающему представляющими интерес качественными признаками.
Манипуляции (такие как мутация, последующая трансфекция и селекция) с растениями или семенами, или их частями могут привести к созданию так называемых ”по существу производных” сортов. Международный союз по защите новых сортов растений (UPOV) выработал следующее руководство для определения сорта, который является существу производным защищенного сорта:
[A] сорт следует считать по существу производным другого сорта (“первоначального сорта”), если
(i) данный сорт в основном выделен из первоначального сорта или из сорта, который сам был выделен из первоначального сорта, при сохранении экспрессии основных характеристик, присущих генотипу или комбинации генотипов первоначального сорта;
(ii) данный сорт имеет явные отличия от первоначального сорта; и
(iii) за исключением отличий, возникших в результате деривации, данный сорт соответствует первоначальному сорту в отношении экспрессии основных характеристик, присущих генотипу или комбинации генотипов первоначального сорта.
UPOV, шестой съезд Международных организаций, Женева, 30 октября 1992 г.; документ подготовлен президиумом Союза.
В использованном здесь значении термин ”линия” означает группу растений, характеризующихся незначительной или отсутствием наследственной изменчивости среди особей в отношении по меньшей мере одного признака. Такие линии могут быть созданы несколькими поколениями самоопыления и селекции или вегетативного размножения из одной родительской формы при помощи методов с использованием культур тканей или клеток.
В использованном здесь значении термины ”культивар” и “сорт” являются синонимами и означают линию, используемую для промышленного производства.
Термин ”устойчивость” или “устойчивый” применительно к определенному компоненту означает сохранение данного компонента из поколения в поколение, предпочтительно на протяжении по меньшей мере трех поколений по существу на одном и том же уровне, например, предпочтительно ±15%, более предпочтительно ±10%, наиболее предпочтительно ±5%. На устойчивость может влиять температура, местоположение, стресс и время посева. При сравнении последующих поколений в полевых условиях указанный компонент должен быть выявлен аналогичным образом.
Термин ”коммерческая полезность” означает хорошую мощность растения и высокую фертильность, благодаря которым фермеры могут выращивать данную сельскохозяйственную культуру с использованием обычных сельскохозяйственных машин и могут экстрагировать из семени масло с требуемыми компонентами при помощи обычного измельчающего и экстрагирующего оборудования. Урожайность коммерчески полезной культуры, измеренная с учетом массы семени, содержания масла и общего количества масла, полученного на акр площади, должна составлять 15% от средней урожайности другого сравниваемого коммерческого сорта, не обладающего первосортными признаками, который был выращен в том же регионе.
Термин ”агрономически элитная” означает линию, которая обладает требуемыми агрономическими характеристиками, такими как урожайность, созревание, устойчивость к болезням и тому подобные, помимо устойчивости к воздействию насекомых, благодаря наличию одного или нескольких вариантов по настоящему изобретению. В объем настоящего изобретения входят агрономические признаки, рассматриваемые отдельно или в любой комбинации в приведенных ниже примерах, которыми обладает растение, включающее вариант по настоящему изобретению. Любые и все указанные агрономические характеристики и данные могут быть использованы для идентификации таких растений на основании одного признака, части диапазона или всего диапазона характеристик, используемых для определения таких растений.
Как должно быть понятно специалисту в данной области в свете настоящего описания изобретения, предпочтительные варианты наборов для обнаружения признаков по настоящему изобретению могут включать, например, зонды и/или праймеры, предназначенные для создания и/или включающие ”соединительные последовательности” или “переходные последовательности” (где геномная фланкирующая последовательность кукурузы стыкуется с последовательностью вставки). Например, данный набор включает полинуклеотидные зонды, праймеры и/или ампликоны, созданные для идентификации одной или обеих соединительных последовательностей (где вставка стыкуется с фланкирующей последовательностью), представленные в таблице 1. Одной общей особенностью такого набора является наличие одного праймера, гибридизирующегося во фланкирующей области, и одного праймера, гибридизирующегося во вставке. Такие праймеры часто имеют длину, равную по меньшей мере примерно ~15 остаткам. Указанные праймеры могут быть использованы для создания/амплификации обнаруживаемого ампликона, указывающего на наличие варианта по настоящему изобретению. Праймеры могут быть использованы для создания ампликона, заполняющего (и включающего) вышеуказанную соединительную последовательность.
Праймер(ы), ”обеспечивающий соединение” во фланкирующей последовательности, обычно не гибридизируется на расстоянии более примерно 200 оснований или за пределами соединения. Таким образом, типичные фланкирующие праймеры включают по меньшей мере 15 остатков цепи в пределах 200 оснований фланкирующих последовательностей от начала вставки. То есть в объем настоящего изобретения входят праймеры, включающие последовательность соответствующей длины, состоящую из остатков ~1674-1873 и/или ~6690-6890 SEQ ID NO:29. Праймеры для вставки могут быть созданы аналогичным образом в любом месте вставки, но для создания таких праймеров могут быть также использованы остатки ~1874-2074 и ~6489-6689.
Специалисту в данной области должно быть также известно, что праймеры и зонды могут быть созданы с возможностью гибридизации в стандартных условиях гибридизации и/или ПЦР с сегментом SEQ ID NO:29 (или комплементом) и комплементарными ей последовательностями, когда праймер или зонд не являются полностью комплементарными приведенной последовательности. То есть допустима некоторая степень ошибочного спаривания. Например, в праймере, состоящем примерно из 20 нуклеотидов, обычно один, два или около того нуклеотидов могут не связываться с противоположной цепью, если ошибочно спаренное основание находится внутри или у конца праймера, противоположного ампликону. Ниже приведены разные приемлемые условия гибридизации. В зондах также могут быть использованы синтетические аналоги нуклеотидов, такие как инозин. Также могут быть использованы пептидные зонды нуклеиновой кислоты (PNA), а также ДНК- и РНК-зонды. Важно, чтобы такие зонды и праймеры позволяли диагностировать (однозначно идентифицировать и выявлять) наличие варианта по настоящему изобретению.
Следует отметить, что при амплификации методом ПЦР могут возникать ошибки, вызывающие, например, незначительные ошибки секвенирования. То есть, за исключением особо оговоренных случаев, последовательности, приведенные в настоящем описании изобретения, были определены в результате создания длинных ампликонов из геномных ДНК кукурузы, после чего ампликоны клонировали и секвенировали. В последовательностях, созданных и определенных таким образом, вполне вероятно, могут быть обнаружены небольшие отличия и незначительные расхождения, связанные с многочисленными циклами амплификации, необходимыми для создания ампликона достаточной длины для секвенирования из геномных ДНК. Специалисту в данной области должно быть понятно, что в объем настоящего изобретения входят любые корректировки, необходимые в связи с возникновением обычных ошибок секвенирования или расхождений указанных типов.
Следует также отметить, что возможно удаление некоторой геномной последовательности, например, при введении последовательности в процессе создания варианта. Таким образом, фланкирующие последовательности по настоящему изобретению могут несколько отличаться от геномных последовательностей, представленных, например, в базе данных GENBANK.
Некоторые из указанных отличий рассмотрены ниже в разделе ”Примеры”. Также могут быть произведены корректировки зондов и праймеров.
Компоненты каждой из ”вставок” показаны на фигурах 1 и 2 и более подробно рассмотрены ниже в разделе ”Примеры”. Полинуклеотидные последовательности ДНК указанных компонентов или их фрагменты могут быть использованы в качестве ДНК-праймеров или зондов при осуществлении способов по настоящему изобретению.
Некоторые варианты осуществления изобретения относятся к способам обнаружения трансгена/инсерционной области генома в растениях, семенах и подобных частях растения кукурузы, и композициям для осуществления указанных способов. Созданы последовательности ДНК, включающие соединительную последовательность трансгена/инсерционной области генома по настоящему изобретению (между остатками 1873-1874 и 6689-6690 SEQ ID NO:29), их сегменты, комплементы приведенных последовательностей и любые их сегменты. Соединительная последовательность инсерционной области заполняет соединение между гетерологичной ДНК, введенной в геном, и ДНК из клетки кукурузы, фланкирующей сайт инсерции. Такие последовательности позволяют диагностировать данный вариант.
На основании указанной вставки и краевых последовательностей могут быть созданы вариант-специфические праймеры. Анализ методом ПЦР показал, что линии кукурузы по настоящему изобретению могут быть идентифицированы в разных генотипах кукурузы путем анализа ампликонов ПЦР, созданных с помощью наборов вариант-специфических праймеров. Для идентификации линий кукурузы по настоящему изобретению могут быть использованы вышеуказанные и другие родственные методы. Таким образом, ампликоны ПЦР, полученные из таких пар праймеров, являются уникальными и могут быть использованы для идентификации указанных линий кукурузы.
Некоторые варианты осуществления изобретения относятся к последовательностям ДНК, включающим смежный фрагмент нового трансгена/инсерционной области генома. В объем настоящего изобретения входят последовательности ДНК, включающие полинуклеотиды последовательности вставки трансгена достаточной длины и полинуклеотиды геномной последовательности кукурузы достаточной длины, из одного или более чем из трех вышеуказанных растений кукурузы, и/или последовательности, пригодные для использования в качестве последовательностей праймеров с целью получения ампликона, позволяющего диагностировать одно или несколько указанных растений кукурузы.
Родственные варианты осуществления изобретения относятся к последовательностям ДНК, включающим по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более смежных нуклеотидов части трансгена последовательности ДНК, приведенной в настоящем описании изобретения (такой как SEQ ID NO:29 и ее сегменты), или комплементарных ей последовательностей, и фланкирующей последовательности ДНК кукурузы аналогичной длины из указанных последовательностей или комплементарных им последовательностей. Такие последовательности могут быть использованы в качестве ДНК-праймеров при осуществлении методов амплификации ДНК. Ампликоны, полученные с помощью указанных праймеров, позволяют диагностировать любые варианты кукурузы по настоящему изобретению. Поэтому настоящее изобретение также относится к ампликонам, полученным с помощью таких ДНК-праймеров и гомологичных праймеров.
Настоящее изобретение также относится к способам обнаружения в образце ДНК, соответствующей варианту кукурузы по настоящему изобретению. Такие способы могут включать: (а) контактирование образца, включающего ДНК, с набором праймеров, которые при использовании в реакции амплификации нуклеиновой кислоты с ДНК по меньшей мере одного из указанных вариантов кукурузы, образуют ампликон, позволяющий диагностировать указанный вариант; (b) выполнение реакции амплификации нуклеиновой кислоты с образованием ампликона; и (с) обнаружение полученного ампликона.
Другие способы обнаружения по настоящему изобретению включают способ обнаружения в образце ДНК, соответствующей по меньшей мере одному из указанных вариантов, который включает: (а) контактирование образца, включающего ДНК, с зондом, который гибридизируется в строгих условиях гибридизации с ДНК по меньшей мере одного из указанных вариантов кукурузы и не гибридизируется в строгих условиях гибридизации с ДНК контрольного растения кукурузы (ДНК не представляющего интереса варианта); (b) гибридизацию образца и зонда в строгих условиях гибридизации; и (с) обнаружение гибридизации зонда с указанной ДНК.
Другие варианты осуществления изобретения относятся к способам создания растения кукурузы, включающего вариант aad-1 по настоящему изобретению, которые включают стадии: (а) полового скрещивания первой родительской линии кукурузы (включающей кассеты экспрессии по настоящему изобретению, сообщающие растениям данной линии признак устойчивости к гербицидам) и второй родительской линии кукурузы (в которой отсутствует указанный признак устойчивости к гербицидам) с образованием многочисленного потомства растений; и (b) отбор растения-потомка при помощи молекулярных маркеров. Такие способы могут необязательно включать стадию обратного скрещивания растения-потомка со второй родительской линией кукурузы с целью получения растения кукурузы с разведением гомозигот, включающего указанный признак устойчивости к воздействию насекомых.
Другим объектом настоящего изобретения являются способы определения зиготности потомства гибрида, включающего любой один (или несколько) из трех указанных вариантов. Указанные способы могут включать контактирование образца, содержащего ДНК кукурузы, с набором праймеров по настоящему изобретению. Указанные праймеры, используемые при осуществлении реакции амплификации нуклеиновой кислоты с геномной ДНК по меньшей мере одного из указанных вариантов кукурузы, образуют первый ампликон, позволяющий диагностировать по меньшей мере один из указанных вариантов кукурузы. Такие способы далее включают выполнение реакции амплификации нуклеиновой кислоты с образованием первого ампликона; обнаружение первого ампликона и контактирование образца, содержащего ДНК кукурузы, с указанным набором праймеров (указанный набор праймеров, используемый при осуществлении реакции амплификации нуклеиновой кислоты с геномной ДНК из растений кукурузы, продуцирует второй ампликон, включающий нативную геномную ДНК кукурузы, гомологичную геномной области кукурузы; и выполнение реакции амплификации нуклеиновой кислоты с образованием второго ампликона. Указанные способы далее включают обнаружение второго ампликона и сравнение первого и второго ампликонов в образце, при этом присутствие обоих ампликонов свидетельствует о том, что образец является гетерозиготным для инсерции трансгена.
Наборы для обнаружения ДНК могут быть созданы с использованием композиций по настоящему изобретению и методов, хорошо известных в области обнаружения ДНК. Указанные наборы могут быть использованы для идентификации ДНК варианта кукурузы по настоящему изобретению в образце в процессе селекции растений кукурузы, содержащих указанную ДНК. Наборы включают последовательности ДНК, гомологичные или комплементарные ампликонам, например, рассмотренным в настоящем описании изобретения, или последовательности ДНК, гомологичные или комплементарные ДНК, содержащейся в генетических элементах трансгена вариантов по настоящему изобретению. Указанные последовательности ДНК могут быть использованы при осуществлении реакций амплификации ДНК или в качестве зондов в способе гибридизации ДНК. Наборы могут также содержать реагенты и вещества, необходимые для выполнения данного способа обнаружения.
Термин ”зонд” означает выделенную молекулу нуклеиновой кислоты, к которой присоединена стандартная детектируемая метка или репортерная молекула (такая как радиоактивный изотоп, лиганд, хемилюминесцентное вещество или фермент). Такой зонд является комплементарным цепи нуклеиновой кислоты-мишени в случае настоящего изобретения, цепи геномной ДНК одного из указанных вариантов кукурузы, из растения кукурузы или из образца, включающего ДНК данного варианта. Зонды по настоящему изобретению включают не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, но также полиамиды и другие вещества зонда, которые специфически связываются с последовательностью ДНК-мишени и могут быть использованы для обнаружения указанной последовательности ДНК-мишени.
Термин ”праймеры” означает выделенные/синтезированные нуклеиновые кислоты, которые гибридизированы с цепью комплементарной ДНК-мишени методом гибридизации нуклеиновых кислот с образованием гибрида между праймером и цепью ДНК-мишени и затем удлинены по цепи ДНК-мишени полимеразой, например, ДНК-полимеразой. Пары праймеров по настоящему изобретению используют для амплификации последовательности нуклеиновой кислоты-мишени, например, при помощи полимеразной цепной реакции (ПЦР) или других стандартных методов амплификации нуклеиновых кислот.
Зонды и праймеры обычно состоят из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500 или большего числа полинуклеотидов. Такие зонды и праймеры специфически гибридизируются с последовательностью-мишенью в строгих условиях гибридизации. Зонды и праймеры по настоящему изобретению предпочтительно обладают полным сходством последовательности с последовательностью-мишенью, хотя стандартными методами могут быть созданы зонды, отличающиеся от последовательности-мишени, но сохраняющие способность гибридизироваться с последовательностями-мишенями.
Методы получения и использования зондов и праймеров описаны, например, в публикации Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Lasboratory Press, Cold Spring Harbor, N.Y., 1989. Пары праймеров для ПЦР могут быть выделены из известной последовательности, например, при помощи компьютерных программ, предназначенных для указанной цели.
Праймеры и зонды на основе фланкирующих последовательностей ДНК и последовательностей вставок по настоящему изобретению могут быть использованы для подтверждения (и при необходимости для коррекции) рассмотренных последовательностей стандартными методами, например, методами повторного клонирования и секвенирования таких последовательностей.
Зонды и праймеры нуклеиновых кислот по настоящему изобретению гибридизируются в строгих условиях с последовательностью ДНК-мишени. Для идентификации ДНК из трансгенного варианта в образце могут быть использованы любые стандартные методы гибридизации или амплификации нуклеиновых кислот. Молекулы нуклеиновых кислот или их фрагменты могут специфически гибридизироваться с другими молекулами нуклеиновых кислот в определенных условиях. Как указано в настоящем описании изобретения, две молекулы нуклеиновых кислот могут специфически гибридизироваться друг с другом, если такие молекулы способны образовывать непараллельную двухцепочечную структуру нуклеиновой кислоты. Считается, что молекула нуклеиновой кислоты является ”комплементом” другой молекулы нуклеиновой кислоты, если указанные молекулы являются полностью комплементарными. Как указано в настоящем описании изобретения, молекулы являются ”полностью комплементарными”, если каждый нуклеотид одной молекулы является комплементарным нуклеотиду другой молекулы. Две молекулы считаются ”минимально комплементарными”, если указанные молекулы способны гибридизироваться друг с другом с достаточной степенью устойчивости для сохранения гибрида по меньшей мере в условиях «пониженной строгости». Аналогичным образом молекулы считаются ”комплементарными”, если указанные молекулы способны гибридизироваться друг с другом с достаточной степенью устойчивости для сохранения гибрида в стандартных строгих условиях. Стандартные строгие условия гибридизации описаны в публикации Sambrook et al., 1989. Отклонения от полной комплементарности допустимы, если такие отклонения полностью не аннулируют способность молекул образовывать двухцепочечную структуру. Молекула нуклеиновой кислоты, которая может быть использована в качестве праймера или зонда, должна иметь достаточно комплементарную последовательность, способную образовывать устойчивую двухцепочечную структуру при определенных концентрациях растворителя и соли.
В использованном здесь значении по существу гомологичная последовательность является последовательностью нуклеиновой кислоты, которая специфически гибридизируется с комплементом последовательности нуклеиновой кислоты, сравниваемой в строгих условиях гибридизации. Термин ”строгие условия” применительно к гибридизации зонда нуклеиновой кислоты с нуклеиновой кислотой-мишенью (то есть с конкретной, представляющей интерес последовательностью нуклеиновой кислоты) методом специфической гибридизации описан в публикации Sambrook et al., 1989, стр. 9.52-9.55. См. также публикацию Sambrook et al., 1989, стр. 9.47-9.52 и 9.56-9.58. Таким образом, нуклеотидные последовательности по настоящему изобретению могут быть использованы благодаря их способности избирательно образовывать дуплексные молекулы с комплементарными участками фрагментов ДНК.
В зависимости от предполагаемого применения можно использовать разные условия гибридизации для достижения разных степеней избирательности зонда в отношении последовательности-мишени. Для применений, требующих высокой избирательности, обычно используют относительно строгие условия для образования гибридов, например, могут быть выбраны условия, характеризующиеся низкой концентрацией соли и/или высокой температурой, например, достигаемые при использовании от около 0,02 М до около 0,15 М NaCl при температуре от около 50
С до около 70
С. Строгие условия, например, могут включать по меньшей мере двукратную промывку фильтра для гибридизации строгим промывающим буфером (0,2-кратный объем SSC, 0,1% SDS, 65
С). Специалистам в данной области (6.3.1-6.3.6) известны соответствующие строгие условия, стимулирующие гибридизацию ДНК, например, 6,0-кратный объем хлорида натрия/цитрата натрия (SSC) при температуре около 45
С с последующей промывкой 2,0-кратным объемом SSC при 50
С. Например, концентрация соли на стадии промывки может быть выбрана в диапазоне от условий пониженной строгости, включающих использование примерно 2,0-кратного объема SSC при 50
С, до строгих условий, включающих использование примерно 0,2-кратного объема SSC при 50
С. Кроме того, температура на стадии промывки может быть повышена от комнатной температуры, примерно 22
С, создаваемой в условиях пониженной строгости, до температуры около 65
С, применяемой в строгих условиях. Могут быть изменены как температура, так и концентрация соли, либо один из параметров, температура или концентрация соли, может оставаться постоянным при изменении другого параметра. Такие избирательные условия допускают незначительные, если вообще допускают, ошибочные спаривания между зондом и матрицей или цепью-мишенью. Специалистам в данной области хорошо известны методы обнаружения последовательностей ДНК путем гибридизации, и в качестве примера таких анализов методом гибридизации можно привести патенты США №№ 4965188 и 5176995.
В особенно предпочтительном варианте осуществления изобретения нуклеиновая кислота по настоящему изобретению специфически гибридизируется в строгих условиях с одним или несколькими праймерами (или ампликонами, или другими последовательностями), приведенными или предложенными в настоящем описании изобретения, включая их комплементы и фрагменты. В одном аспекте настоящего изобретения маркерная молекула нуклеиновой кислоты по настоящему изобретению включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO:3-14, ее комплементы и/или фрагменты.
В другом аспекте настоящего изобретения маркерная молекула нуклеиновой кислоты по настоящему изобретению включает последовательность, которая идентична таким последовательностям нуклеиновых кислот на 80%-100% или 90%-100%. В еще одном аспекте настоящего изобретения маркерная молекула нуклеиновой кислоты по настоящему изобретению идентична такой последовательности на 95%-100%. Такие последовательности могут быть использованы в качестве маркеров в процессе селекции растений для идентификации потомства генетических гибридов. Гибридизацию зонда с молекулой ДНК-мишени можно обнаружить разными методами, известными специалистам в данной области, которые включают, не ограничиваясь ими, флуоресцентные метки, радиоактивные метки, метки на основе антител и хемилюминесцентные метки.
В случае амплификации последовательности-мишени нуклеиновой кислоты (например, методом ПЦР) с использованием конкретной пары праймеров для амплификации ”строгие условия” представляют собой условия, в которых указанная пара праймеров гибридизируется только с последовательностью-мишенью нуклеиновой кислоты, с которой праймер, имеющий соответствующую последовательность дикого типа (или ее комплемент), должен связываться и предпочтительно продуцировать уникальный продукт амплификации, ампликон.
Термин ”специфичный к (последовательности-мишени)” означает, что зонд или праймер гибридизируется в строгих условиях гибридизации только с последовательностью-мишенью в образце, включающем такую последовательность-мишень.
В использованном здесь значении термин ”амплифицированная ДНК” или “ампликон” означает продукт амплификации нуклеиновой кислоты последовательности-мишени нуклеиновой кислоты, являющейся частью матрицы нуклеиновой кислоты. Например, для определения наличия в растении кукурузы, полученном в результате полового скрещивания, геномной ДНК трансгенного варианта из растения кукурузы по настоящему изобретению, ДНК, экстрагированная из образца ткани растения кукурузы, может быть подвергнута амплификации нуклеиновой кислоты с использованием пары праймеров, из которых один праймер выделен из фланкирующей последовательности, локализованной в геноме растения рядом с сайтом инсерции вставленной гетерологичной ДНК, и второй праймер выделен из вставленной гетерологичной ДНК, для продуцирования ампликона, позволяющего диагностировать наличие ДНК варианта. Вариант также можно диагностировать на основании длины и последовательности ампликона. Длина ампликона может быть равна общей длине пар праймеров и одной пары нуклеотидных оснований и/или общей длине пар праймеров и примерно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 30 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500, 750, 1000, 1250, 1500, 1750, 2000 или большему числу пар нуклеотидных оснований (плюс или минус любое число вышеуказанных приращений). Альтернативно пара праймеров может быть выделена из фланкирующей последовательности с обеих сторон вставленной ДНК с образованием ампликона, включающего всю нуклеотидную последовательность вставки. Один член пары праймеров, выделенной из геномной последовательности растения, может быть расположен на расстоянии от вставленной последовательности ДНК. Указанное расстояние может составлять от одной пары нуклеотидных оснований до примерно двадцати тысяч пар нуклеотидных оснований. В определение термина ”ампликон” входят димеры праймеров, которые могут быть образованы при выполнении термической реакции амплификации ДНК.
Нуклеиновая кислота может быть амплифицирована разными методами амплификации нуклеиновой кислоты, известными в данной области, включая полимеразную цепную реакцию (ПЦР). В данной области известны разные методы амплификации, которые описаны, наряду с прочими публикациями, в патенте США № 4683195 и патенте США № 4683202. Методом ПЦР можно амплифицировать до 22 т.п.о. геномной ДНК. При осуществлении настоящего изобретения могут быть использованы указанные методы, а также другие методы амплификации ДНК, известные в данной области. Последовательность ДНК-вставки или фланкирующей геномной последовательности гетерологичного трансгена из варианта кукурузы по настоящему изобретению может быть проверена (и при необходимости скорректирована) путем амплификации таких последовательностей варианта с помощью праймеров, выделенных из последовательностей по настоящему изобретению, стандартными методами секвенирования ДНК ампликона ПЦР или клонированной ДНК.
Ампликон, полученный указанными методами, может быть обнаружен разными методами. Электрофорез в агарозном геле и окрашивание бромидом этидия является общепринятым, хорошо известным методом обнаружения ампликонов ДНК. Другим таким методом является генетический двоичный анализ с использованием олигонуклеотида ДНК, перекрывающего смежную фланкирующую геномную последовательность ДНК и вставленную последовательность ДНК. Указанный олигонуклеотид иммобилизуют в лунках микропланшета. После ПЦР представляющей интерес области (с использованием одного праймера во вставленной последовательности и одного праймера в смежной фланкирующей геномной последовательности) одноцепочечный продукт ПЦР может быть гибридизирован с иммобилизованным олигонуклеотидом и использован в качестве матрицы для выполнения реакции удлинения на одно основание с использованием ДНК-полимеразы и меченых ddNTP, специфичных к предполагаемому следующему основанию. Считывание может быть произведено при помощи флуоресцентного анализа или ELISA. Сигнал указывает на наличие вставки/фланкирующей последовательности вследствие успешной амплификации, гибридизации и удлинения на одно основание.
Другим методом является метод пиросеквенирования, описанный в публикации Winge (Innov. Pharma. Tech. 00:18-24, 2000). При выполнении указанного метода создают олигонуклеотид, который перекрывает место соединения смежной геномной ДНК и вставленной ДНК. Олигонуклеотид гибридизируется с одноцепочечным продуктом ПЦР из представляющей интерес области (один праймер во вставленной последовательности и один праймер во фланкирующей геномной последовательности) и инкубируют в присутствии ДНК-полимеразы, АТР, сульфурилазы, люциферазы, апиразы, аденозин-5'-фосфосульфата и люциферина. Отдельно добавляют DNTP, в результате чего возникает световой сигнал, который измеряют. Световой сигнал свидетельствует о наличии вставки/фланкирующей последовательности трансгена вследствие успешной амплификации, гибридизации и удлинения на одно или несколько оснований.
Поляризация флуоресценции является еще одним методом, который может быть использован для обнаружения ампликона по настоящему изобретению. В соответствии с данным методом создают олигонуклеотид, который перекрывает место соединения геномной фланкирующей последовательности и вставленной ДНК. Олигонуклеотид гибридизируется с одноцепочечным продуктом ПЦР из представляющей интерес области (один праймер во вставленной ДНК и один праймер во фланкирующей геномной последовательности ДНК) и инкубируют в присутствии ДНК-полимеразы и меченного флуоресцентным красителем ddNTP. Удлинение на одно основание вызывает включение ddNTP. Такое включение можно измерить в виде изменения поляризации при помощи флуорометра. Изменение поляризации свидетельствует о наличии вставки/фланкирующей последовательности трансгена вследствие успешной амплификации, гибридизации и удлинения на одно основание.
TAQMAN (PE Applied Biosystems, Foster City, Calif.) является методом обнаружения и количественного определения наличия последовательности ДНК. В кратком изложении данный метод включает создание олигонуклеотидного зонда FRET, который перекрывает место соединения геномной фланкирующей последовательности и вставочной ДНК. Зонд FRET и праймеры для ПЦР (один праймер в последовательности вставочной ДНК и один праймер во фланкирующей геномной последовательности) подвергают циклической обработке в присутствии термостойкой полимеразы и dNTP. В процессе специфической амплификации taq полимераза ДНК отщепляется и высвобождает флуоресцентную часть из гасящей части зонда FRET. Флуоресцентный сигнал свидетельствует о наличии фланкирующей последовательности/последовательности вставки трансгена вследствие успешной амплификации и гибридизации.
Для обнаружения последовательности был предложен метод молекулярных маяков. В кратком изложении данный метод включает создание олигонуклеотидного зонда FRET, который перекрывает место соединения фланкирующей геномной последовательности и вставочной ДНК. Уникальная структура зонда FRET позволяет получить вторичную структуру, содержащую флуоресцентную и гасящую части в непосредственной близости друг от друга. Зонд FRET и праймеры для ПЦР (один праймер в последовательности вставочной ДНК и один праймер во фланкирующей геномной последовательности) подвергают циклической обработке в присутствии термостойкой полимеразы и dNTP. После успешной амплификации методом ПЦР гибридизация зонда FRET с последовательностью-мишенью вызывает удаление вторичной структуры зонда и пространственное отделение флуоресцентной и гасящей частей. Возникает флуоресцентный сигнал. Флуоресцентный сигнал свидетельствует о наличии фланкирующей геномной последовательности/последовательности вставки трансгена вследствие успешной амплификации и гибридизации.
Помимо рассмотренного положения в геноме кукурузы, которое является наиболее подходящим для инсерции, настоящее изобретение также относится к семени кукурузы и/или растению кукурузы, включающему по меньшей мере одну вставку, отличную от гена aad-1, в непосредственной близости от данного положения в геноме. Один вариант осуществления изобретения относится к замене вставки aad-1, рассмотренной в настоящем описании изобретения, другой вставкой. Для указанной цели может быть использована, например, направленная гомологичная рекомбинация. Данный метод описан, например, в WO 03/080809 A2 и соответствующей опубликованной заявке на патент США (US 20030232410). Таким образом, настоящее изобретение относится к растениям и растительным клеткам, включающим гетерологичную вставку (вместо гена или с несколькими копиями гена aad-1), фланкированную полными фланкирующими последовательностями или узнаваемыми частями фланкирующих последовательностей по настоящему изобретению (например, остатками 1-1873 и 6690-8557 SEQ ID NO:29).
Все патенты, заявки на патент, предварительные заявки и публикации, приведенные в настоящем описании изобретения, полностью включены в настоящее описание изобретения в качестве ссылки в той мере, в какой они соответствуют положениям настоящего изобретения.
Приведенные ниже примеры иллюстрируют способы осуществления изобретения и демонстрируют определенные предпочтительные варианты осуществления изобретения. Указанные примеры не ограничивают объем изобретения. Специалистам в данной области должно быть понятно, что методы, описанные в нижеследующих примерах, представляют определенные подходы, используемые для иллюстрации предпочтительных вариантов осуществления изобретения. Однако специалистам в данной области должно быть понятно в свете настоящего описания изобретения, что в конкретные варианты осуществления изобретения могут быть внесены многие изменения, позволяющие получить аналогичные или подобные результаты, не выходя за пределы существа и объема изобретения. За исключением особо оговоренных случаев все проценты являются массовыми процентами, и соотношения смесей растворителей являются объемными соотношениями.
За исключением особо оговоренных случаев в настоящем описании изобретения использованы следующие аббревиатуры.
Примеры
Пример 1. Трансформация и отбор варианта pDAS1740-278 AAD-1
Вариант pDAS1740-278 AAD-1 был получен методом точечной трансформации (WHISKER) линии кукурузы Hi-II. Данный метод трансформации описан в заявке на патент США № 20090093366. В указанную линию кукурузы был перенесен фрагмент FspI плазмиды pDAS1740, также определяемой как pDAB3812 (фигура 1). Конструкция плазмиды содержит кассету экспрессии растения, включающую RB7 MARv3:: промотор v2 убиквитин 1 Zea mays // AAD1 v3 // PER5 3'UTR Zea mays:: транскрипционную единицу растения (PTU) RB 7 MARv4.
Было получено много вариантов. Вариантам, сохранившим жизнеспособность и продуцировавшим здоровую ткань каллюса, устойчивую к галоксифопу, были присвоены однозначные идентифицирующие коды, представляющие предполагаемые трансформационные варианты, после чего указанные варианты были перенесены в свежую селективную среду. Растения были регенерированы из ткани, выделенной из каждого уникального варианта, и перенесены в теплицу.
Были получены образцы листьев для молекулярного анализа, выполняемого для проверки наличия трансгена AAD-1 методом саузерн-блоттинга, методом подтверждения краевых последовательностей ДНК и методом подтверждения с использованием геномного маркера. Положительные растения Т0 опыляли инбредными линиями для получения семени Т1. Растения Т1 с вариантом pDAS1470-278-9 (DAS-40278-9) отбирали, подвергали самоопылению и исследовали на протяжении пяти поколений. В то же время растения Т1 подвергали обратному скрещиванию и переносили гены в элитную зародышевую плазму (ХНН13) путем опосредуемой маркером селекции на протяжении нескольких поколений. Данный вариант был получен из независимого трансформированного изолята. Указанный вариант был отобран на основании уникальных характеристик, таких как единственный сайт инсерции, нормальное менделевское расщепление, устойчивая экспрессия, превосходная комбинация эффективности, включающая устойчивость к гербициду и агрономическую продуктивность в широких генотипических средах и многочисленных окружениях. В приведенных ниже примерах представлены данные, использованные для исследования варианта pDAS-1740-278-9.
Пример 2. Исследование варианта pDAS1740-278-9 методом саузерн-блоттинга
Анализ методом саузерн-блоттинга был выполнен для установления паттерна интеграции инсертированного фрагмента ДНК и определения числа вставок/копий гена aad-1 в варианте pDAS-1740-278-9 (DAS-40278-9). Полученные данные свидетельствовали об интеграции и целостности трансгена aad-1, введенного в геном кукурузы.
Данные саузерн-блоттинга позволили предположить, что вставка фрагмента pDAS1740/Fsp I в вариант DAS-40278-9 кукурузы произошла в виде простой интеграции одной интактной копии транскрипционной единицы aad-1 из плазмиды pDAS1740. Был выполнен детальный анализ методом саузерн-блоттинга с использованием зондов, специфичных к гену, промотору, терминатору и другим регуляторным элементам, находящимся в области плазмиды, и дескриптивных рестрикционных ферментов, имеющих сайты расщепления в плазмиде и продуцирующих гибридизирующиеся фрагменты внутри плазмиды или фрагменты, заполняющие место соединения плазмиды с геномной ДНК кукурузы (краевые фрагменты). Молекулярные массы, установленные в результате гибридизации методом саузерн-блоттинга для комбинации рестрикционного фермента и зонда, были характерны для данного варианта и определяли его идентифицирующие паттерны. Указанные анализы также показали, что фрагмент плазмиды был введен в геномную ДНК кукурузы без реаранжировки транскрипционной единицы aad-1. Идентичные гибридизирующиеся фрагменты были обнаружены в пяти разных поколениях варианта DAS-40278-9 трансгенной кукурузы, что свидетельствует об устойчивом наследовании инсерции PTU aad-1 на протяжении нескольких поколений. Гибридизация со смесью трех зондов к области остова, расположенных за пределами сайта рестрикции Fsp I плазмиды pDAS1740, не позволила обнаружить каких-либо специфических фрагментов ДНК/гена, что указывает на отсутствие гена устойчивости к ампициллину и отсутствие других областей остовов вектора непосредственно рядом с сайтами рестрикции Fsp I плазмиды pDAS1740 в варианте DAS-40278-9 трансгенной кукурузы. Иллюстративная карта вставки варианта DAS-40278-9 кукурузы AAD-1 показана на фигурах 2-3.
Пример 2.1. Получение образцов листьев кукурузы и выделение геномной ДНК (гДНК)
гДНК была получена из листьев отдельных растений кукурузы, содержащих вариант DAS-40278-9 кукурузы aad-1. гДНК экстрагировали из ткани листьев отдельных растений кукурузы, содержащих вариант DAS-40278-9 кукурузы aad-1. Были использованы семена трансгенной кукурузы, полученные из пяти разных поколений варианта DAS-40278-9. Для исследования варианта DAS-40278-9 были отобраны двадцать отдельных растений кукурузы из четырех растений в каждом поколении. Кроме того, гДНК была выделена из растения обычной кукурузы, ХНН13, содержащего генетическую среду, характерную для данной линии с отсутствием гена aad-1.
До выделения гДНК листья были взяты из каждого растения для анализа экспрессии белка aad-1 при помощи набора индикаторных полосок для экспресс-анализа (American Bionostica, Swedesboro, NJ) в соответствии с рекомендациями производителя. Каждый образец пунктата листьев был оценен как + или - в зависимости от наличия или отсутствия aad-1. Для дальнейшего исследования были отобраны только положительные растения из пяти поколений варианта DAS-40278-9.
Образцы листьев кукурузы были получены у отдельных растений, содержащих вариант DAS-40278-9, и обычного контрольного растения XHH13. Образцы листьев быстро замораживали в жидком азоте и хранили примерно при -80°С до использования.
Геномную ДНК экстрагировали из ткани замороженных листьев кукурузы стандартным методом СТАВ. При необходимости некоторые образцы геномной ДНК подвергали дальнейшей очистке при помощи Qiagen Genomic-Tip (Quigan, Valencia, CA) методами, рекомендованными производителем. Экстрагированную ДНК подвергали количественному спектрофлуорометрическому анализу с использованием реагента Pico Green (Invitrogen, Carlsbad, CA). Затем ДНК визуализировали на агарозном геле для подтверждения результатов анализа с использованием реагента Pico Green и определения качества ДНК.
Пример 2.2. Расщепление и отделение ДНК
Для молекулярного исследования ДНК девять микрограммов (9 мкг) геномной ДНК из образца ДНК варианта DAS-40278-9 кукурузы и образца ДНК обычной контрольной кукурузы расщепляли, добавляя к каждому образцу ДНК примерно пять-одиннадцать единиц отобранного рестрикционного фермента на мкг ДНК и соответствующий реакционный буфер. Каждый образец инкубировали примерно при 37°С в течение ночи. Для расщепления образцов использовали рестрикционные ферменты EcoR I, Nco I, Sac I, Fse I и Hind III (New England Biolabs, Ipswich, MA). Положительный гибридизированный контрольный образец, полученный в результате объединения плазмидной ДНК, pDAS1740 (pDAB3812) с геномной ДНК контрольного образца в соотношении, примерно эквивалентом 1 копии трансгена на один геном кукурузы, расщепляли такими же методами с использованием таких же рестрикционных ферментов, что и испытуемые образцы. ДНК из контрольного образца обычной кукурузы (ХНН13) расщепляли такими же методами с использованием таких же рестрикционных ферментов, что и испытуемые образцы, и использовали в качестве отрицательного контрольного образца.
Образцы расщепленной ДНК осаждали при помощи Quick-Precip (Edge BioSystems, Gaithersburg, MD) и повторно суспендировали в 1-кратном объеме наполнителя Blue Juice (Invitrogen, Carlsbad, CA) для достижения объема, необходимого для введения в гель. Образцы ДНК и маркеры молекулярной массы подвергали электрофорезу в 0,8% агарозных гелях с 1-кратным буфером ТВЕ (Fisher Scientific, Pittsburg, PA) под напряжением 55-65 вольт в течение примерно 18-22 часов для отделения фрагмента. Гели окрашивали бромидом этидия (Invitrogen, Carlsbad, CA) и визуализировали ДНК в ультрафиолетовом (UV) свете.
Пример 2.3. Перенос методом саузерн-блоттинга и обработка мембраны
Анализ методом саузерн-блоттинга выполняли в основном в соответствии с описанием, приведенным в публикации Memelink, et al., (1994) Southern, Northern, and Western Blot Analysis. Plant Mol. Biol. Manual F1:1-23. После отделения с помощью электрофореза и визуализации фрагментов ДНК гели депуринизировали 0,25 н. раствором HCl (Fisher Scientific, Pittsburgh, PA) в течение примерно 15 минут, подвергали воздействию денатурирующего раствора (AccuGENE, Sigma, St. Louis, MO) в течение примерно 30 минут и затем нейтрализующего раствора (AccuGENE, Sigma, St. Louis, MO) в течение по меньшей мере 30 минут. Полученные фрагменты ДНК в течение ночи переносили методом саузерн-блоттинга на найлоновые мембраны (Roche Diagnostics, Indianapolis, IN) с использованием системы тампонов с 10-кратным объемом SSC (Sigma, St. Louis, MO). После переноса фрагментов ДНК мембраны промывали 2-кратным раствором SSC и связывали ДНК с мембраной путем перекрестного сшивания в ультрафиолетовом свете. В результате методом саузерн-блоттинга были получены мембраны, готовые к гибридизации.
Пример 2.4. Мечение и гибридизация ДНК-зонда
Фрагменты ДНК, связанные с найлоновой мембраной, были обнаружены с помощью меченого зонда. Зонды, использованные для данного исследования, были созданы путем введения методом ПЦР нуклеотида, меченного дигоксигенином (DIG), [DIG-11]-dUTP, из фрагментов, полученных с помощью праймеров, специфичных к элементам гена и другим областям плазмиды pDAS1740. ДНК-зонды были синтезированы методом ПЦР с использованием набора для синтеза ПЦР DIG-меченных зондов (Roche Diagnostics, Indianapolis, IN) в соответствии с рекомендациями производителя. В таблице 1 приведен перечень зондов, использованных в данном исследовании.
Локализация и длина зондов, использованных при выполнении анализа методом саузерн-блоттинга
Меченые зонды анализировали методом электрофореза в агарозном геле для определения их качестве и количества. Требуемое количество меченого зонда затем использовали для гибридизации с ДНК-мишенью на найлоновых мембранах для обнаружения специфических фрагментов методами, описанными для использования раствора DIG Easy Hyb (Roche Diagnostics, Indianapolis, IN). Содержащие ДНК блоты, фиксированные на найлоновых мембранах, промывали 2-кратным объемом SSC и предварительно гибридизировали с 20-25 мл предварительно нагретого раствора DIG Easy Hyb в сосудах для гибридизации примерно при 50°С в течение как минимум 30 минут в шкафу для гибридизации. Раствор для предварительной гибридизации затем декантировали и заменяли 20 мл предварительно нагретого раствора DIG Easy Hyb, содержащего требуемое количество специфических зондов, предварительно денатурированных путем кипячения в воде в течение 5 минут. Стадию гибридизации выполняли примерно при 40-60°С в течение ночи в шкафу для гибридизации.
Пример 2.5. Обнаружение
В конце гибридизации зондов растворы DIG Easy Hyb, содержащие зонды, декантировали в чистые пробирки и хранили при -20°С. Указанные зонды могут быть повторно использованы 2-3 раза в соответствии с рекомендациями производителя. Блоты на мембранах быстро ополаскивали и дважды промывали в чистых пластиковых емкостях промывающим буфером в условиях пониженной строгости (2-кратный объем SSC, 0,1% SDS) в течение примерно 5 минут при комнатной температуре, затем дважды промывали промывающим буфером в строгих условиях (0,1-кратный раствор SSC, 0,1% SDS) в течение 15 минут примерно при 65°С. Блоты на мембранах затем переносили в другие чистые пластиковые емкости и недолго промывали 1-кратным промывающим буфером из набора промывающих и блокирующих буферов DIG (Roche Diagnostics, Indianapolis, IN) в течение примерно 2 минут, блокировали в 1-кратном блокирующем буфере в течение как минимум 30 минут и инкубировали с антителом против DIG-меченной щелочной фосфатазы (АР) (разведение в отношении 1:5000, Roche Diagnostics, Indianapolis, IN) в 1-кратном блокирующем буфере в течение как минимум 30 минут. После 2-3 промывок 1-кратным промывающим буфером специфические ДНК-зонды оставались связанными с блотами на мембранах, и эталонные образцы DIG-меченной ДНК визуализировали в системе обнаружения нуклеиновой кислоты с помощью хемилюминесцентного красителя CDP-Star (Roche Diagnostics, Indianapolis, IN) в соответствии с рекомендациями производителя. Блоты экспонировали на хемилюминесцентной пленке (Roche Diagnostics, Indianapolis, IN) в течение одного или двух периодов времени для обнаружения гибридизирующихся фрагментов и визуализации эталонов молекулярной массы. Затем пленки проявляли проявителем для пленок All-Pro 100 Plus (Konica SRX-101) и сканировали изображения. Для каждого зонда было документировано число и размеры обнаруженных полос. Маркер II молекулярной массы DIG-меченной ДНК (MWM DIG II), ставший видимым после обнаружения DIG, был использован для определения размера гибридизирующегося фрагмента на блотах, полученных методом саузерн-блоттинга.
Пример 2.6. Отделение зондов
ДНК-зонды отделяли от блотов на мембранах после получения данных гибридизации методом саузерн-блоттинга, после чего блоты на мембранах можно было повторно использовать для гибридизации с другим ДНК-зондом в соответствии рекомендациями производителя (DIG Application Manual for Filter Hybridization, (2003), Roche Diagnostics). После обнаружения сигнала и экспонирования на пленке блоты на мембранах тщательно ополаскивали водой Milli-Q и дважды промывали в отделяющем буфере (0,2 н. раствор NaOH, 0,1% SDS) в течение примерно 15 минут при комнатной температуре или при 37°С. Блоты на мембранах затем недолго промывали 2-кратным объемом SSC, после чего они были готовы к предварительной гибридизации и гибридизации с другим ДНК-зондом. Блоты на мембранах экспонировали на новой хемилюминесцентной пленке для гарантии того, что все ДНК-зонды были удалены до перехода к следующей гибридизации. Повторно экспонированные пленки помещали вместе с ранее полученными данными гибридизации в файл исследований для регистрации.
Пример 2.7. Результаты анализа методом саузерн-блоттинга
В таблице 2 указаны размеры предполагаемых и установленных фрагментов при определенном расщеплении и использовании соответствующего зонда с учетом сайтов известных рестрикционных ферментов фрагмента pDAS1740/Fsp I. В результате выполненных расщеплений и гибридизаций были идентифицированы фрагменты двух типов: внутренние фрагменты, в которых сайты известных ферментов фланкируют область зонда и полностью находятся внутри фрагмента pDAS1740/Fsp I, и краевые фрагменты, в которых сайт известного фермента расположен у одного конца области зонда и наличие второго сайта предполагается в геноме кукурузы. Размеры краевых фрагментов изменяются в зависимости от варианта, так как в большинстве случаев сайты интеграции фрагментов ДНК являются уникальными для каждого варианта. Краевые фрагменты позволяют локализовать сайт рестрикционного фермента по отношению к интегрированной ДНК и определить число инсерций ДНК. По результатам анализов методом саузерн-блоттинга, выполненных в данном исследовании, был сделан вывод о том, что в геном варианта DAS-40278-9 кукурузы была введена одна копия интактной PTU гена aad-1 из плазмиды pDAS1740/Fsp I, как детально показано на карте вставок (фигуры 2-3).
Гибридизирующиеся фрагменты, прогнозированные и установленные при выполнении анализа методом саузерн-блоттинга
ZmUbi1
ZmPer5
остова
# Дублеты в контрольном образце обычной кукурузы, BC3S1, и некоторых образцах BC3S2.
1. Размеры прогнозированных фрагментов указаны на основании плазмидной карты pDAS1740 (pDAB3812), показанной на фигуре 1.
2. Размеры фрагментов, установленных при выполнении указанных анализов, считаются приблизительными и указаны на основании размеров фрагментов маркера II молекулярной массы DIG-меченой ДНК. Из-за введения молекул DIG для визуализации фрагменты маркера обычно имеют длину, которая примерно на 5-10% больше их фактической молекулярной массы.
Для исследования вставки гена aad-1 в вариант DAS-40278-9 были отобраны рестрикционные ферменты с уникальным сайтом рестрикции в плазмиде pDAS1740, EcoR I, Nco I, Sac I, Fse I/Hind III. Было предположено, что краевой фрагмент длиной >3382 п.о., >2764 п.о., >4389 п.о. должен гибридизироваться с зондом к гену aad-1 после расщепления соответственно EcoR I, Nco I и Sac I (таблица 2). Была обнаружена одна полоса гибридизации aad-1, соответствующая ~12000 п.о., ~4000 п.о. и ~16000 п.о. при использовании соответственно EcoR I, Nco I и Sac I, что указывает на наличие одного сайта инсерции гена aad-1 в геноме варианта DAS-40278-9 кукурузы. Расщепление двумя ферментами Fse I и Hind III было выбрано для высвобождения фрагмента длиной 3361 п.о., содержащего транскрипционную единицу aad-1 (PTU, промотор/ген/терминатор) (таблица 2). После расщепления Fse I/Hind III был обнаружен прогнозированный фрагмент длиной 3361 п.о. при использовании зонда к гену aad-1. Результаты, полученные для расщепления образца DAS-40278-9 всеми четырьмя ферментами/комбинацией ферментов с последующей гибридизацией с зондом к гену aad-1, показали, что в геном варианта DAS-40278-9 кукурузы была введена одна копия интактной транскрипционной единицы aad-1 из плазмиды pDAS1740.
Для исследования области промотора (ZmUbi1) для aad-1 в варианте DAS-40278-9 были выбраны рестрикционные ферменты Nco I, Sac I и Fse I/Hind III. Расщепление ферментами Nco I и Sac I предположительно должно вызвать образование фрагмента краевой области длиной соответственно >3472 п.о. и >4389 п.о. при гибридизации с ДНК-зондами, специфичными к области промотора ZmUbi1 (таблица 2). Две полосы гибридизации, соответствующие ~6300 п.о. и ~3600 п.о., были обнаружены при использовании зонда к промотору ZmUbi1 после расщепления Nco I. Однако полоса, соответствующая ~3600 п.о., присутствовала во всех образцах, включая контрольные образцы обычной кукурузы, из чего следует, что полоса, соответствующая ~3600 п.о., является полосой неспецифического сигнала, образующегося в результате гомологичного связывания зонда к выделенному из кукурузы промотору убиквитину (ZmUbi1) с эндогенным геном ubi кукурузы. В отличие от этого полоса сигнала, соответствующая ~6300 п.о., была обнаружена в исследованных образцах DAS-40278-9 и не была обнаружена в контрольных образцах обычной кукурузы, из чего следует, что полоса, соответствующая ~6300 п.о., является специфичной для зонда к промотору ZmUbi1 из плазмиды pDAS1740, и поэтому данная полоса является предполагаемой полосой для Nco I/ZmUbi1, указанной в таблице 2. Аналогичным образом две полосы гибридизации, соответствующие ~3800 п.о. и ~16000 п.о., были обнаружены при использовании зонда к промотору ZmUbi1 после расщепления Sac I. Полоса, соответствующая ~3800 п.о., присутствовала во всех образцах, включая контрольные образцы обычной кукурузы, и поэтому может рассматриваться как неспецифическая гибридизация зонда к промотору ZmUbi1 с эндогенным геном ubi кукурузы. Полоса гибридизации, соответствующая ~16000 п.о., присутствующая только в образцах DAS-40278-9, является предполагаемой полосой Sac I/ZmUbi1. Расщепление двумя ферментами Fse I/Hind III предположительно высвобождает фрагмент RTU aad-1 длиной 3361 п.о., который гибридизируется с зондом к промотору ZmUbi1 (таблица 2). Данная полоса, соответствующая 3361 п.о., и полоса неспецифической гибридизации, соответствующая ~6400 п.о., были обнаружены при использовании зонда к промотору ZmUbi1 после расщепления Fse I/Hind III. Полоса, соответствующая ~6400 п.о., является полосой неспецифического связывания зонда к промотору ZmUbi1 с эндогенным геном ubi кукурузы, так как данная полоса присутствует во всех образцах, включая контрольные образцы обычной кукурузы. Кроме того, в контрольном образце обычной кукурузы BC3S1 и в некоторых образцах BC3S2 была обнаружена другая полоса в непосредственной близости от полосы, соответствующей ~6400 п.о. Указанная дополнительная полоса в непосредственной близости от полосы, соответствующей ~6400 п.о., также считается полосой неспецифического связывания, так как она присутствует в контрольных образцах обычной кукурузы ХНН13 и, наиболее вероятно, ассоциирована с генетической средой ХНН13.
Такие же рестрикционные ферменты/комбинация ферментов Nco I, Sac I и Fse I/Hind III были выбраны для исследования области терминатора (ZmPer5) для aad-1 в варианте DAS-40278-9. Расщепление ферментом Nco I предположительно образует фрагмент краевой области длиной >2764 п.о. при гибридизации с ДНК-зондами, специфичными к области терминатора ZmPer5 (таблица 2). После расщепления ферментом Nco I при использовании зонда к терминатору ZmPer5 были обнаружены две полосы гибридизации, соответствующие ~4000 п.о. и ~3900 п.о. Полоса, соответствующая ~3900 п.о., присутствовала во всех образцах, включая контрольные образцы обычной кукурузы, из чего следует, что полоса, соответствующая ~3900 п.о., является полосой неспецифического сигнала, вероятно, вследствие гомологичного связывания зонда к терминатору (ZmPer5) выделенного из кукурузы гена пероксидазы с эндогенным геном per кукурузы. В отличие от этого полоса сигнала, соответствующая ~4000 п.о., была обнаружена только в исследованных образцах DAS-40278-9 и не была обнаружена в контрольных образцах обычной кукурузы, из чего следует, что полоса, соответствующая ~4000 п.о., является специфичной для зонда к терминатору ZmPer5 из плазмиды pDAS1740 и поэтому представляет собой предполагаемую полосу Nco I/ZmPer5, указанную в таблице 2. Краевой фрагмент длиной >1847 п.о. предположительно гибридизируется с зондом к терминатору ZmPer5 после расщепления ферментом Sac I. После расщепления ферментом Sac I при использовании зонда к терминатору ZmPer5 были обнаружены две полосы гибридизации, соответствующие ~1900 п.о. и ~9000 п.о. Полоса, соответствующая ~9000 п.о., присутствовала во всех образцах, включая контрольные образцы обычной кукурузы, и поэтому считается полосой неспецифической гибридизации зонда к терминатору ZmPet5 с эндогенным геном per кукурузы. Полоса гибридизации, соответствующая ~1900 п.о., которая присутствовала только в образцах DAS-40278-9, считается предполагаемой полосой Sac I/ZmPer5. Расщепление двумя ферментами Fse I/Hind III предположительно высвобождает фрагмент PTU aad-1 длиной 3361 п.о., который гибридизуется с зондом к терминатору ZmPer5 (таблица 2). Полоса, соответствующая 3361 п.о., и дополнительная полоса неспецифической гибридизации, соответствующая ~2100 п.о., были обнаружены при использовании зонда к терминатору ZmPer5 после расщепления ферментами Fse I/Hind III. Дополнительная полоса, соответствующая ~2100 п.о., является полосой неспецифического связывания зонда к терминатору ZmPer5 с эндогенным геном кукурузы, так как указанная полоса присутствует во всех образцах, включая отрицательные контрольные образцы. Результаты, полученные при расщеплении образца DAS-40278-9 указанными ферментами с последующей гибридизацией с зондом к промотору ZmUbi1 и терминатору ZmPer5, снова подтвердили, что в геном варианта DAS-40278-9 кукурузы была введена одна копия интактной PTU aad-1 из плазмиды pDAS1740.
Для исследования остальных компонентов фрагмента pDAS1740/Fsp I в варианте DAS-40278-9 кукурузы AAD-1 были выбраны рестрикционные ферменты Nco I и Sac I (таблица 2). Последовательности ДНК компонентов RB7 Mar v3 и RB7 Mar v4 идентичны более чем на 99,7%, поэтому предполагалось, что ДНК-зонды, специфичные к RB7 Mar v3 или RB7 Mar v4, должны гибридизироваться с фрагментами ДНК, содержащими любой вариант RB7 Mar. Два краевых фрагмента длиной >2764 п.о. и >3472 п.о. предположительно гибридизировали с зондами к RB7 Mar v4 и RB7 Mar v3 после расщепления ферментом Nco I (таблица 2). После расщепления ферментом Nco I в образцах DAS-40278-9 были обнаружены две полосы гибридизации, соответствующие ~4000 п.о. и ~6300 п.о., при использовании зонда к RB7 Mar v4 или RB7 Mar v3. Аналогичным образом два краевых фрагмента длиной >1847 п.о. и >4389 п.о. были обнаружены при использовании зондов к RB7 Mar v4 и RB7 Mar v3 после расщепления ферментом Sac I (таблица 2). После расщепления ферментом Sac I при использовании зонда к RB7 Mar v4 или RB7 Mar v3 в образцах DAS-40278-9 были обнаружены полосы гибридизации, соответствующие ~1900 п.о. и ~16000 п.о.
Вместе взятые результаты гибридизации методом саузерн-блоттинга, полученные при использовании зондов к указанным элементам, показывают, что ДНК, введенная в вариант DAS-40278-9 кукурузы, содержит интактную PTU aad-1 наряду с областями присоединения матрицы RB7 Mar v3 и RB7 Mar v4 соответственно у 5'- и 3'-концов вставки.
Пример 2.8. Отсутствие последовательностей области остова
Комбинация с равным молярным соотношением трех фрагментов ДНК (таблица 1), охватывающих почти всю область остова Fsp I (4867-7143 п.о. в плазмиде pDAS1740) плазмиды pDAS1740, была использована в качестве зонда к области остова для исследования варианта DAS-40278-9 кукурузы AAD-1. Для создания варианта DAS-40278-9 был использован фрагмент плазмиды pDAS1740/Fsp I, поэтому не ожидалось возникновения специфического сигнала гибридизации при использовании указанной комбинации зонда к области остова (таблица 2) после расщепления любым рестрикционным ферментом. Было подтверждено отсутствие специфического сигнала гибридизации при использовании зонда к области остова после расщепления ферментом Nco I или Sac I во всех образцах DAS-40278-9. Положительные контрольные образцы содержали предполагаемые полосы гибридизации, показывающие, что указанные зонды были способны гибридизироваться с любыми фрагментами гомологичной ДНК при наличии их в образцах. Полученные данные позволили предположить, что инсерция в вариант DAS-40278-9 кукурузы не включала последовательность области остова вектора за пределами области Fsp I из плазмиды pDAS1740.
Образцы листьев из пяти разных поколений варианта DAS-40278-9 были использованы для исследования молекулярной массы методом саузерн-блоттинга. Производя расщепление выбранными рестрикционными ферментами и используя комбинации зондов, определяли паттерн интеграции для исследования вставленного гена aad-1, а также некодирующих областей, включающих промотор, терминатор экспрессии гена и области присоединения матрицы.
Исследование методом саузерн-блоттинга ДНК, введенной в вариант DAS-40278-9, показывает, что в вариант DAS-40278-9 была введена одна интактная копия PTU aad-1. Молекулярные массы, установленные путем гибридизации методом саузерн-блоттинга для комбинации рестрикционного фермента и зонда, были уникальными для данного варианта и характеризовались собственными идентифицирующими паттернами. Паттерн гибридизации идентичен для всех пяти поколений, из чего следует, что данная вставка является устойчивой в геноме кукурузы. Гибридизация с зондами, охватывающими область остова за пределами трансформирующего фрагмента pDAS1740/Fsp I из плазмиды pDAS1740, подтверждает отсутствие последовательностей области остова вектора в варианте DAS-40278-9.
Пример 3. Клонирование и исследование последовательности ДНК во вставке и фланкирующих краевых областях варианта DAS-40278-9 кукурузы
Для исследования вставленной ДНК и описания сайта инсерции в геноме определяли последовательности ДНК вставки и краевых областей варианта DAS-40278-9. Было подтверждено в общей сложности 8557 п.о. геномной последовательности варианта DAS-40278-9, включающих 1873 п.о. 5'-концевой фланкирующей краевой последовательности, 1868 п.о. 3'-концевой фланкирующей краевой последовательности и 4816 п.о. ДНК-вставки. ДНК-вставка длиной 4816 п.о. содержит интактную кассету экспрессии гена aad-1, 259 п.о. неполной области MAR v3 у 5'-конца и 1096 п.о. неполной области MAR v4 у 3’-конца. Анализ последовательности показал наличие инсерции длиной 21 п.о. в месте 5'-концевого интеграционного соединения и делецию двух пар оснований из инсерционного локуса генома кукурузы. Инсерция, включающая одну пару оснований, была обнаружена в месте 3'-концевого интеграционного соединения между геномом кукурузы и вставкой DAS-40278-9. Кроме того, замена одного основания (Т на С) была обнаружена во вставке в положении 5212 в некодирующей области 3'-концевой UTR. Ни одно из указанных изменений не повлияло на структуру открытой рамки считывания кассеты экспрессии aad-1.
Амплификация методом ПЦР вставки варианта DAS-40278-9 и краевых последовательностей подтвердила, что краевые области были получены из кукурузы и соединительные области могут быть использованы для вариант-специфической идентификации DAS-40278-9. Анализ последовательности, заполняющей соединительные области, показал, что в результате инсерции ДНК в вариант DAS-40278-9 не были образованы новые открытые рамки считывания (OPF>=200 кодонов) и существующие открытые рамки считывания генома не были нарушены введением DAS-40278-9 в нативный геном кукурузы. Таким образом, исследование вставки и краевых последовательностей варианта DAS-40278-9 кукурузы AAD-1 показало, что в нативный геном кукурузы была введена одна интактная копия кассеты экспрессии aad-1.
Пример 3.1. Экстракция и количественное определение геномной ДНК
Геномную ДНК экстрагировали из лиофилизованных или только что измельченных тканей листьев модифицированным методом СТАВ. Образцы ДНК растворяли в 1-кратном буфере ТЕ (10 мМ трис-буфера, рН 8,0, 1 мМ EDTA) (Fluka, Sigma, St. Louis, MO) и производили количественное определение методом с использованием Pico Green в соответствии с инструкциями производителя (Molecular Probes, Eugene, OR). Для анализа методом ПЦР образцы ДНК разводили водой для молекулярной биологии (5 PRIME, Gaithersburg, MD) до концентрации 10-100 нг/мкл.
Пример 3.2. Праймеры для ПЦР
В таблице 3 представлены последовательности праймеров, которые были использованы для клонирования ДНК-вставки и фланкирующих краевых областей варианта DAS-40278-9, с указанием положений и описаниями, показанными на фигуре 4. В таблице 4 представлены последовательности праймеров, которые были использованы для подтверждения вставки и краевых последовательностей. Положения праймеров показаны соответственно на фигурах 4 и 5. Все праймеры были синтезированы в компании Integrated DNA Technologies, Inc. (Coralville, IA). Праймеры растворяли в воде (5 PRIME, Gaithersburg, MD) до концентрации 100 мкМ для получения исходного раствора и разводили водой до концентрации 10 мкМ для получения рабочего раствора.
Список последовательностей праймеров, использованных при клонировании вставки варианта DAS-40278-9 кукурузы и фланкирующей краевой последовательности
(-): комплементарная последовательность
Список последовательностей праймеров, использованных для подтверждения геномной ДНК кукурузы
(-): комплементарная последовательность
Пример 3.3. «Прогулка по геному» (Genome Walking)
Для клонирования 5'- и 3'-концевых фланкирующих краевых последовательностей варианта DAS-40278-9 кукурузы был использован универсальный набор Genome Walker™ (Clontech Laboratories, Inc., Mountain View, CA). В соответствии с инструкциями производителя примерно 2,5 мкг геномной ДНК из варианта DAS-40278-9 кукурузы AAD-1 расщепляли в течение ночи ферментами EcoR V, Stu I (оба фермента входили в комплект набора) или Sca I (New England Biolabs, Ipswich, MA). Расщепленную ДНК очищали в устройстве для очистки и концентрирования ДНК DNA Clean & Concentrator™-25 (ZYMO Research, Orange, CA) и затем лигировали с адапторами из набора Genome Walker™ для создания библиотек Genome Walker™. Каждую библиотеку Genome Walker™ использовали в качестве матричной ДНК для амплификации при помощи первичной ПЦР с использованием праймера для адаптора АР1, входившего в комплект набора, и специфичных к конструкции праймеров 5End3812_A и 3End3812_C. Один микролитр продукта первичной ПЦР, разведенный в отношении 1:25, использовали в качестве матрицы для амплификации при помощи вторичной ПЦР с использованием вложенного праймера для адаптора АР2 и вложенных специфичных к конструкции праймеров 5End3812_В и 3End3812_D. В процессе амплификации методом ПЦР использовали TaKaRa LA Taq™ HS (Takara Bio Inc., Shiga, Japan). Для получения 50 мкл продукта ПЦР использовали 1 мкл матричной ДНК, 8 мкл 2,5 мМ смеси dNTP, 0,2 мкМ каждого праймера, 2,5 единицы ДНК-полимеразы TaKaRa LA Taq™ HS, 5 мкл 10-кратного буфера II LA для ПЦР (вместе с Mg2+) и 1,5 мкл 25 мМ MgCl2. Условия выполнения ПЦР приведены в таблице 5.
Условия «прогулки по геному» варианта DAS-40278-9 кукурузы AAD-1 для амплификации фланкирующих краевых областей
/
AP1
/
AP2
/
AP1
/
AP2
Пример 3.4. Стандартная ПЦР
Для клонирования и подтверждения ДНК-вставки и краевой последовательности в варианте DAS-40278-9 кукурузы использовали стандартную ПЦР. Амплификацию при помощи стандартной ПЦР производили с использованием TaKaRa LA Taq™ (Takara Bio Inc., Shiga, Japan), ДНК-полимеразы HotStarTaq (Quigen, Valencia, CA), системы для удлинения последовательности при помощи высокоточной ПЦР (Roche Diagnostics, Inc., Indianapolis, IN) или смеси ферментов и рабочего раствора для клонирования при помощи высокоточной ПЦР Easy-A® (Stratagene, LaJolla, CA) в соответствии с рекомендациями производителя. Конкретные условия выполнения ПЦР и описания ампликонов приведены в таблице 6.
Условия амплификации при помощи стандартной ПЦР краевых областей в варианте DAS-40278-9 кукурузы
Пример 3.5. Обнаружение, очистка, субклонирование и секвенирование продуктов ПЦР
Продукты ПЦР исследовали при помощи электрофореза, используя 1,2% или 2% Е-гель (Invitrogen, Carlsbad, CA) в соответствии с инструкциями производителя. Размер фрагмента определяли путем сравнения с маркерами ДНК. При необходимости фрагменты, полученные при помощи ПЦР, очищали, для чего указанные фрагменты переносили из 1% агарозного геля в 1-кратный буфер ТВЕ, окрашенный бромидом этидия, используя набор для экстракции геля QIAquick (Quigen, Carlsbad, CA).
Фрагменты, полученные при помощи ПЦР, субклонировали в pCR®4-TOPO®, используя набор для секвенирования TOPO TA Cloning® (Invitrogen, Carlsbad, CA) в соответствии с инструкциями производителя. В частности, два-пять микролитров клонируемого продукта реакции ТОРО® переносили в химически компетентные клетки ТОР10 One Shot в соответствии с инструкциями производителя. Клонированные фрагменты, правильность которых проверяли с помощью минипрепарата плазмидной ДНК (набор QIAprep Spin Miniprep, Qiagen, Carlsbad, CA), расщепляли рестрикционным ферментом EcoR I или при помощи колониеспецифической ПЦР с использованием праймеров Т3 и Т7, входивших в комплект набора. Плазмидную ДНК или исходные растворы отобранных колоний в глицерине отправляли для секвенирования.
После субклонирования предполагаемые продукты направленной ПЦР сначала секвенировали для подтверждения клонирования ожидаемых фрагментов ДНК. Колонии, содержащие соответствующие последовательности ДНК, отбирали для скрининга с помощью праймеров для определения полных последовательностей ДНК. Секвенирование было выполнено в компании Cogenics (Houston, TX).
Конечную сборку вставки и краевых последовательностей выполняли при помощи программного обеспечения Sequencher (версия 4.8 Gene Codes Corporation, Ann Arbor, MI). Описание вставки и краевых последовательностей варианта DAS-40278-9 кукурузы было произведено при помощи программы Vector NTI (версия 10 и 11, Invitrogen, Carlsbad, CA).
Поиск гомологии был выполнен при помощи программы BLAST на основании базы данных GenBank. Открытую рамку считывания (ORF) анализировали при помощи программы Vector NTI (версия 11, Invitrogen) для идентификации ORF (>=200 кодонов) в полной вставке и фланкирующих краевых последовательностях.
Пример 3.6. 5'-Концевая краевая последовательность
Фрагмент ДНК амплифицировали из каждой библиотеки GenomeWalker™ вариантов DAS-40278-9 кукурузы, используя набор специфических вложенных праймеров для 5'-конца трансгена. Продукт ПЦР длиной примерно 800 п.о. был обнаружен в обеих библиотеках GenomeWalker™ EcoR V и Stu I варианта DAS-40278-9. В библиотеке GenomeWalker™ Sca I был создан продукт длиной около 2 т.п.о. Фрагменты клонировали в pCR®4-TOPO® и произвольно отбирали шесть колоний из каждой библиотеки для секвенирования конца с целью подтверждения наличия во вставке предполагаемых последовательностей. Полное секвенирование путем скрининга вставок с помощью праймеров показало, что фрагменты, амплифицированные из библиотек GenomeWalker™ Stu I, EcoR V и Sca I варианта DAS-40278-9 кукурузы, имели длину, соответственно равную 793, 822 и 2132 п.о. Фрагменты ДНК, полученные из библиотек GenomeWalker™ Stu I и EcoR V, на 100% соответствовали фрагменту ДНК, полученному из библиотеки GenomeWalker™ Sca I, из чего следует, что указанные фрагменты ДНК были амплифицированы из 5'-концевой области вставки трансгена. Поиск с помощью программы BLAST полученной геномной последовательности кукурузы длиной 1873 п.о. выявил высокое сходство с последовательностью клона ВАС кукурузы. Кроме того, анализ последовательности в месте соединения сайта инсерции показал, что были усечены 917 п.о. MAR v3 в 5'-концевой области по сравнению с плазмидой pDAS1740/фрагментом Fsp I, при этом в 5'-концевой области кассеты экспрессии aad-1 была сохранена часть MAR v3 длиной 259 п.о.
Пример 3.7. 3'-Концевая краевая последовательность
Фрагмент ДНК длиной примерно 3 т.п.о. амплифицировали из библиотеки GenomeWalker™ Stu I варианта DAS-40278-9 кукурузы, используя набор специфических вложенных праймеров для 3'-конца трансгена. Фрагмент ДНК клонировали в pCR®4-TOPO® и произвольно отбирали десять колоний для секвенирования конца с целью подтверждения инсерции предполагаемых последовательностей. Полностью секвенировали три клона с предполагаемыми вставками, в результате чего был получен фрагмент ДНК длиной 2997 п.о. Анализ последовательности указанного фрагмента ДНК показал наличие неполного элемента MAR v4 (при отсутствии 70 п.о. в 5'-концевой области) и геномной последовательности кукурузы длиной 1867 п.о. Поиск с помощью программы BLAST показал, что последовательность геномной ДНК длиной 1867 п.о. на 100% соответствовала последовательности такого же клона ВАС кукурузы, которая была идентифицирована в 5'-концевой краевой последовательности.
Пример 3.8. ДНК-вставка и соединительная последовательность
ДНК-вставку и соединительные области клонировали из варианта DAS-40278-9 при помощи ПЦР ранее описанными методами. Были созданы пять пар праймеров на основании 5'- и 3'-концевых фланкирующих краевых последовательностей и предполагаемой последовательности трансгена. В общей сложности клонировали и секвенировали пять перекрывающихся фрагментов ДНК (ампликон 1 длиной 1767 п.о., ампликон 2 длиной 1703 п.о., ампликон 3 длиной 1700 п.о., ампликон 4 длиной 1984 п.о. и ампликон 5 длиной 1484 п.о.) (фигура 4). Всю вставку и фланкирующие краевые последовательности собирали на основании последовательности, перекрывающей все пять фрагментов. В конечной последовательности было подтверждено наличие ДНК-вставки длиной 4816 п.о., выделенной из pDAS1740/Fsp I, 5'-концевой фланкирующей краевой последовательности длиной 1873 п.о. и 3'-концевой фланкирующей краевой последовательности длиной 1868 п.о. ДНК-вставка длиной 4816 п.о. содержит интактную кассету экспрессии гена aad-1, неполную область MAR v3 длиной 259 п.о. у 5'-конца и неполную область MAR v4 длиной 1096 п.о. у 3'-конца (SEQ ID NO:29).
Для скрининга праймерами использовали по меньшей мере два клона для каждой пары праймеров, чтобы получить полную информацию о ДНК-вставке и ее краевых последовательностях. Анализ последовательности выявил инсерцию длиной 21 п.о. у 5'-концевого интеграционного соединения между геномной ДНК кукурузы и вставленной неполной областью MAR v3 из pDAS1740/Fsp I. Результаты поиска с помощью программы BLAST и анализа с помощью программы Vector NTI показали, что ДНК-вставка длиной 21 п.р. не обладала гомологией с ДНК любых видов растений или ДНК плазмиды pDAS1740. Инсерция одной пары оснований была обнаружена у 3'-концевого интеграционного соединения между геномной ДНК кукурузы и неполной областью MAR v4 из pDAS1740/Fsp I. Введение ДНК также сопровождалось делецией двух пар оснований в инсерционном локусе генома кукурузы (фигура 6). Кроме того, было обнаружено отличие одного нуклеотида (замена Т на С) в положении 5212 п.о. в нетранслированной 3'-концевой области UTR ДНК-вставки (SEQ ID NO:29). Однако ни одно из указанных изменений, по-видимому, существенно не влияет на экспрессию гена aad-1 или создания новых открытых рамок считывания (>=200 кодонов) в местах соединения вставки DAS-40278-9.
Пример 3.9. Подтверждение геномных последовательностей кукурузы
Для подтверждения наличия сайта инсерции трансгена варианта DAS-40278-9 в геноме кукурузы выполняли амплификацию методом ПЦР, используя разные пары праймеров (фигура 4). В качестве матрицы использовали геномную ДНК из варианта DAS-40278-9 и других линий трансгенной или нетрансгенной кукурузы. Для амплификации фрагментов ДНК, заполняющих промежуток от гена aad-1 до 5’-концевой краевой последовательности, использовали два праймера, специфичные к гену aad-1, AI5End01 и AI5End02, и два праймера, созданные на основании 5'-концевой краевой последовательности, 1F5End01 и 1F5End02. Аналогичным образом для амплификации фрагмента ДНК, заполняющего промежуток от гена aad-1 до 3'-концевой краевой последовательности, использовали праймер 1F3End05, выделенный из 3'-концевой краевой последовательности, и праймер AI3End01, специфичный к гену aad-1. Фрагменты ДНК предполагаемой длины амплифицировали только из геномной ДНК варианта DAS-40278-9 кукурузы AAD-1, при этом каждая пара праймеров состояла из одного праймера, локализованного у фланкирующего края варианта DAS-40278-9 кукурузы AAD-1, и одного праймера, специфичного к гену aad-1. Из образцов контрольной ДНК не были получены продукты ПЦР при использовании таких же пар праймеров, из чего следует, что клонированные 5’- и 3'-концевые краевые последовательности действительно являются соответственно верхними и нижними последовательностями введенной конструкции гена aad-1. Следует отметить, что слабая полоса, соответствующая примерно 8 т.п.о., была обнаружена во всех образцах кукурузы, включающих вариант DAS-40278-9 кукурузы AAD-1, вариант DAS-40474 кукурузы AAD-1 и линию нетрансгенной кукурузы ХНН13, при использовании для амплификации методом ПЦР пары праймеров 1F5End01 и AI5End01. Обнаруженная слабая полоса (на приготовленном геле) может быть результатом неспецифической амплификации в геноме кукурузы при использовании данной пары праймеров.
Для дальнейшего подтверждения инсерции ДНК в геноме кукурузы были использованы два праймера, локализованные у 5'-концевой краевой последовательности, 1F5End03 и 1F5End04, и два праймера, локализованные у 3'-концевой краевой последовательности, 1F3End03 и 1F3End04, для амплификации фрагментов ДНК, заполняющих инсерционный локус. В результате амплификации методом ПЦР с использованием пары праймеров 1F5End03/1F3End03 или пары праймеров 1F5ERnd04/1F3End04 был получен фрагмент предполагаемой длины, равной примерно 8 т.п.о., из геномной ДНК варианта DAS-40278-9 кукурузы AAD-1. В отличие от этого не были получены продукты ПЦР из геномной ДНК варианта DAS-40474-7 кукурузы AAD-1 или линии нетрансгенной кукурузы ХНН13. При условии, что вариант DAS-40278-9 и вариант DAS-40474-7 кукурузы AAD-1 были созданы в результате трансформации HiII с последующим обратным скрещиванием первичных трансгенных вариантов с линией кукурузы ХНН13, то большая часть генома каждого из указанных двух вариантов теоретически была получена из линии кукурузы ХНН13. Весьма вероятно, что только фланкирующие краевые последовательности, расположенные рядом с трансгеном aad-1, были перенесены из первичной геномной ДНК и сохранились в процессе интрогрессии варианта AAD-1, в то время как другие области геномных последовательности могли быть заменены геномными последовательностями ХНН13. Поэтому неудивительно, что не были амплифицированы фрагменты из геномной ДНК варианта DAS-40474-7 кукурузы AAD-1 и ХНН13 при использовании пары праймеров 1F5End03/1F3End03 или пары праймеров 1F5End04/1F3End04. При использовании пар праймеров 1F5End03/1F3End03 и 1F5End04/1F3End04 были соответственно амплифицированы фрагменты длиной около 3,1 и 3,3 т.п.о. в геномной ДНК линий кукурузы HiII и В73 в отличие от линии кукурузы А188. Полученные результаты показывают, что краевые последовательности были получены из генома линии кукурузы В73.
Было произведено дополнительное клонирование геномной ДНК из кукурузы B73/HiII для подтверждения достоверности фланкирующих краевых последовательностей. Фрагменты, амплифицированные методом ПЦР, секвенировали для подтверждения того, что область ДНК-вставки находится в определенном положении геномной ДНК B73/HiII. Праймеры были созданы на основании полученной последовательности. Для амплификации фрагмента длиной 2212 п.о., заполняющего 5'-3' соединения нативного генома B73/HiII без ДНК-вставки, был использован набор праймеров Amp 1F/Amp 5R. Анализ последовательности показал, что из нативного генома В73 была удалена пара оснований в инсерционном локусе трансгена. Анализ последовательностей ДНК из клонированного фрагмента нативного генома В73 позволил выявить одну открытую рамку считывания (>=200 кодонов), расположенную внизу от 3'-концевой области интеграционного соединения. В первоначальном локусе, в который был интегрирован вариант DAS-40278-9 кукурузы AAD-1, не было обнаружено других открытых рамок считывания. Поиск с помощью программы BLAST также подтвердил, что 5'- и 3'-концевые краевые последовательности из варианта DAS-40278-9 расположены рядом друг с другом в таком же клоне кукурузы ВАС.
С учетом уникальности 5'-концевого интеграционного соединения варианта DAS-40278-9 кукурузы AAD-1 были созданы две пары специфических праймеров для ПЦР, 1F5EndT1F/1F5EndT1R и Corn278-F/Corn278-R, для амплификации указанной области соединения вставки с геномом растения. В соответствии с прогнозом требуемый фрагмент ДНК был создан только в геномной ДНК варианта DAS-40278-9 кукурузы AAD-1 и отсутствовал в любых других линиях трансгенной или нетрансгенной кукурузы. Поэтому две указанные пары праймеров могут быть использованы в качестве вариант-специфических идентификаторов варианта DAS-40278-9 кукурузы AAD-1.
Пример 4. Исследование генома при помощи фланкирующих маркерных последовательностей SSR варианта DAS-40278-9
Для исследования и описания сайта инсерции генома были определены маркерные последовательности, локализованные в непосредственной близости от вставки. Для идентификации и картирования положения трансгена была использована панель полиморфных маркеров SSR. Вариант pDAS1740-278 локализован в хромосоме 2 на расстоянии примерно 20 сМ между маркерами SSR UMC1265 и ММС0111 и на расстоянии примерно 20 сМ на карте сцепления генов кукурузы DAS 2008. В таблице 6 суммирована информация о праймерах для указанных двух маркеров, которые были обнаружены в непосредственной близости от трансгена pDAS1740-278.
Названия праймеров, метки красителей, положения локусов, последовательности верхнего и нижнего праймеров и важные примечания, относящиеся к фланкирующим маркерным последовательностям, ассоциированным с вариантом pDAS1740-278
Пример 4.1. Выделение гДНК
гДНК экстрагировали из пунктатов листьев, используя набор для исследования растений DNEasy 96 (Quiagen, Valencia, California). В процедуру экстракции были внесены изменения, необходимые для автоматизации метода. Выделенную гДНК подвергали количественному анализу с использованием красителя PicoGreen® компании Molecular Probes, Inc. (Eugene, OR). гДНК разводили до концентрации 5 нг/мкл, используя стерильную деионизированную воду для всех образцов.
Пример 4.2. Скрининг гДНК с помощью маркеров
Разведенную гДНК генотипировали совокупностью маркеров простых повторов последовательности (SSR). Маркеры SSR были синтезированы в компании Applied Biosystems (Foster City, California), при этом верхние праймеры метили флуоресцентными метками 6-FAM, HEX/VIC или NED (соответственно синяя, зеленая и желтая). Маркеры были разделены на группы или панели на основании их флуоресцентной метки и размера ампликона для облегчения мультиплексирования и анализа после выполнения ПЦР.
ПЦР выполняли на 384-луночных аналитических планшетах при использовании для каждой реакции 5 нг геномной ДНК, 1,25-кратного буфера для ПЦР (Qiagen, Valencia, California), 0,20 мкМ верхнего и нижнего праймеров, 1,25 мМ MgCl2, 0,015 мМ каждой dNTP и 0,3 единицы ДНК-полимеразы HotStart Taq (Qiagen, Valencia, California). Амплификацию выполняли в системе для ПЦР GeneAmp 9700, оснащенной модулем со сдвоенными головками для 384 лунок (Applied Biosystems, Foster City, California). Была использована следующая программа амплификации: (1) начальная активация Taq при 95°С в течение 12 минут; (2) 30 секунд при 94°С; (3) 30 секунд при 55°С; (4) 30 секунд при 72°С; (5) повтор стадий 2-4 на протяжении 40 циклов и (6) конечное удлинение при 72°С в течение 30 минут. Продукты ПЦР для каждой панели маркеров SSR мультиплексировали, добавляя 2 мкл каждого продукта ПЦР из одного растения в стерильную деионизированную воду до общего объема 60 мкл. Из всех мультиплексированных продуктов ПЦР 0,5 мкл наносили на 384-луночные планшеты, содержащие 5 мкл наполняющего буфера, состоящего из пары оснований GeneScan 500 стандартного размера LIZ и формамида ABI HiDi с соотношением 1:100 (Applied Biosystems, Foster City, California). Затем образцы вводили в анализатор ДНК ABI Prism 3730x1 (Applied Biosystems, Foster City, California) для выполнения капиллярного электрофореза в соответствии с рекомендациями производителя в течение общего времени, равного 36 минутам. Данные маркеров собирали при помощи программного обеспечения для сбора данных автоматического секвенатора ABI Prism 3730x1 версии 4.0 и экстрагировали при помощи программного обеспечения GeneMapper 4.0 (Applied Biosystems) для исследования аллелей и мечения фрагментов соответствующей длины.
Пример 4.3. Результаты скрининга с помощью маркеров SSR
Данные праймеров для фланкирующих маркерных последовательностей, обнаруженных в непосредственной близости от трансгена, приведены в таблице 6. Два ближайших маркера, UMC1265 и MMC0111, локализованы на расстоянии примерно 20 сМ от вставки трансгена в хромосоме 2.
Пример 5. Исследование белка aad-1 в варианте DAS-40278-9
Были исследованы биохимические свойства рекомбинантного белка aad-1, выделенного из варианта DAS-40278-9 трансгенной кукурузы. Биохимические свойства указанного белка исследовали при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE, окрашенном кумасси голубым и красителем для обнаружения гликопротеина), вестерн-блоттинга, анализов при помощи иммунодиагностических полосок, масс-спектрометрии с лазерной десорбцией на матрице/ионизационной времяпролетной масс-спектрометрии (MALDI-TOF MS) и анализа секвенирования белка при помощи тандемной масс-спектрометрии.
Пример 5.1. Анализ при помощи иммунодиагностических полосок
Наличие белка aad-1 в ткани листьев DAS-40278-9 было подтверждено при помощи коммерчески доступных иммунодиагностических полосок компании American Bionostica. Указанные полоски позволяют отличить трансгенные и нетрансгенные растения путем тестирования неочищенных экстрактов листьев (данные не показаны). Экстракты нетрансгенных растений (ХНН13) не содержали иммунореактивный белок в детектируемых количествах. Полученный результат также был подтвержден при помощи анализа методом вестерн-блоттинга.
Экспрессию белка aad-1 исследовали при помощи иммунодиагностических полосок. С каждого растения ХНН13 (контрольное растение) и растения с вариантом DAS-40278-9 получали пунктат из четырех листьев, зажимая ткань между защелкивающимися крышками маркированных 1,5 мл центрифужных пробирок. В лаборатории в каждую пробирку добавляли 0,5 мл буфера для экстракции aad-1 (American Bionostica, Swedesboro, NJ), ткань гомогенизировали при помощи одноразового пестика и встряхивали образец в течение ~10 секунд. После гомогенизации ткани в пробирку вводили индикаторную полоску и оставляли для проявления в течение ~5 минут. Наличие или отсутствие белка aad-1 в растительном экстракте подтверждали на основания появления (или отсутствия) индикаторной линии на иммунодиагностической полоске. После подтверждения экспрессии белка aad-1 для трансгенного варианта собирали ткань стеблей кукурузы, которую лиофилизовали и хранили при температуре около -80°С до использования.
Пример 5.2. Очистка белка aad-1 от кукурузы
Получали иммуноочищенный белок aad-1, выделенный из кукурузы, (молекулярная масса: ~33 кДа) или неочищенные водные экстракты из ткани стебля кукурузы. Все ткани листьев и стеблей собирали и направляли в лабораторию следующим образом: листья срезали с растения ножницами, помещали в тканевые мешки и хранили при температуре около -20°С для последующего использования. Отдельно срезали стебли непосредственно над линией почвы, помещали в тканевые мешки и сразу же замораживали при температуре около -80°С в течение ~6 часов. Затем стебли помещали в лиофилизатор на 5 дней для удаления воды. Полностью высушенные ткани измельчали в тонкий порошок сухим льдом и хранили при температуре около -80°С до использования.
Белок aad-1, выделенный из кукурузы, экстрагировали из лиофилизованной ткани стеблей в фосфатном буфере (см. таблицу 7 для ознакомления с компонентами буфера), для чего ~30 граммов лиофилизованной ткани отвешивали в охлажденный 1000 мл стеклянный блендер и добавляли 500 мл экстрагирующего буфера. Ткань перемешивали с высокой скоростью в течение 60 секунд, и растворимые белки собирали, центрифугируя образец в течение 20 минут с ускорением 30000 × g. Дебрис повторно экстрагировали описанным методом, супернатанты объединяли и фильтровали через 0,45 мкм фильтр. Фильтрованные супернатанты вводили при температуре около +4°С в колонку для иммуноаффинной хроматографии, содержащую антитело против aad-1, которое было конъюгировано с моноклональным антителом, полученным в компании Strategic Biosolution Inc. (моноклональное антитело 473F1 85.1; очищенный протеин А; партия № 609.03С-2-4; 6,5 мг/мл (всего ~35,2 мг)) (Windham, ME); конъюгированным с CNBr-активированной сефарозой 4В (GE Healthcare, Piscataway, NJ). Несвязанные белки собирали и колонку медленно промывали 20 мМ предварительно охлажденного аммоний-бикарбонатного буфера, рН 8,0. Связанные белки элюировали 3,5 М NaSCN (Sigma, St. Louis, MO), 50 мМ трис-буфера (Sigma, St. Louis, MO), рН 8,0. Было получено семь 5 мл фракций, после чего фракции под номерами 2→7 диализовали в течение ночи при температуре около +4°С против 10 мМ трис-буфера, рН 8,0. Указанные фракции исследовали методами SDS-PAGE и вестерн-блоттинга и остальные образцы хранили при температуре около +4°С до использования в последующих анализах.
Коммерчески доступные эталонные вещества, использованные в данном исследовании, приведены в нижеследующей таблице:
Белок, связанный с колонкой для иммуноаффинной хроматографии, исследовали методом SDS-PAGE, при этом полученные результаты показали, что элюированные фракции содержат белок aad-1 с примерной молекулярной массой 33 кДа. Кроме того, было выполнено исследование методом вестерн-блоттинга, результаты которого были положительными в отношении белка aad-1. Белок aad-1 из кукурузы выделяли из ~30 г лиофилизованного вещества стеблей.
Пример 5.3. Методы SDS-PAGE и вестерн-блоттинга
Лиофилизованную ткань из стеблей кукурузы с вариантом DAS-40278-9 и ХНН13 (~100 мг) отвешивали в 2 мл микроцентрифужные пробирки и экстрагировали ~1 мл PBST (Sigma, St. Louis, MO), содержащим 10% коктейля ингибитора растительной протеазы (Sigma, St. Louis, MO). Экстракцию облегчали, добавляя 4 маленьких шарикоподшипника, и измельчали образец в течение 1 минуты. Измельченные образцы центрифугировали в течение 5 минут с ускорением 20000 × g, супернатанты смешивали в отношении 4:1 с 5-кратным буфером для образца Laemmli (2% SDS, 50 мМ трис-буфера, рН 6,8, 0,2 мг/мл бромфенолового синего, 50% (мас./мас.) глицерина, содержащего 10% только что добавленного 2-меркаптоэтанола) и нагревали в течение 5 минут при ~100°С. После непродолжительного центрифугирования 45 мкл супернатанта наносили непосредственно на гель для SDS-PAGE BioRad Criterion (Bio-Rad, Hercules, CA), находящийся в модуле для геля Criterion Cell. Положительный эталонный образец aad-1, выделенного из микроорганизма, ресуспендировали в 1 мг/мл PBST, рН 7,4, и снова разводили PBST. Затем образец смешивали с буфером Laemmli Bio-Rad, содержащим 5% 2-меркаптоэтанола, и обрабатывали ранее описанным методом. Электрофорез выполняли с использованием глицинового трис-буфера с SDS (Bio-Rad, Hercules, CA) при напряжении 150-200 В до достижения красителем конца геля. После отделения гель разрезали пополам, одну половину окрашивали синим красителем для белка GelCode Blue компании Pierce и другую половину при помощи электроблоттинга переносили на нитроцеллюлозную мембрану (Bio-Rad, Hercules, CA), используя ячейку для электрофоретического переноса методом трансблоттинга (Bio-Rad, Hercules, CA), в течение 60 минут при постоянном напряжении 10 вольт. Буфер для переноса содержал 20% метанола и глициновый трис-буфер компании Bio-Rad. Для иммунообнаружения мембрану зондировали специфическим поликлональным антителом кролика против aad-1 (Strategic Biosolution Inc., Newark, DE, поликлональное антитело кролика, очищенное протеином А, партия №: DAS F1 197-151, 1,6 мг/мл). Конъюгат антитела козы против IgG кролика (H+L) и щелочной фосфатазы (Pierce Chemical, Rockford, IL) использовали в качестве вторичного антитела. Для проявления и визуализации полос иммунореактивного белка использовали субстрат BCIP/NBT SigmaFast. Мембрану медленно промывали водой до прекращения реакции и регистрировали результаты при помощи цифрового сканирующего устройства (Hewlett Packard, Palo Alto, CA).
Полоса преобладающего белка aad-1, продуцированного P. fluorescens, которая была визуализирована на гелях для SDS-PAGE, окрашенных кумасси, соответствовала примерно 33 кДа. Как и предполагалось, длина соответствующего белка aad-1, выделенного из кукурузы, (вариант DAS-40278-9) была идентична белкам, экспрессируемым микроорганизмами. Очищенные фракции растения, помимо белка aad-1, содержали незначительное количество неиммунореактивных примесей. Совместно очищенные белки, по-видимому, удерживались на колонке благодаря слабому взаимодействию с матрицей колонки или выщелачиванию моноклонального антитела из колонки в строгих условиях элюирования. Другие исследователи также сообщали о неспецифической адсорбции пептидов и аминокислот на иммуноадсорбентах, представляющих собой сефарозу 4В, активированную цианогенбромидом (Kennedy and Barnes, 1983; Holroyde et al., 1976; Podlaski and Stern, 2008).
Белок aad-1, выделенный из Pseudomonas, характеризовался положительным сигналом предполагаемой длины по результатам анализа методом вестерн-блоттинга с использованием поликлонального антитела. Указанный сигнал также был обнаружен в экстракте стеблей трансгенной кукурузы DAS-40278-9. При выполнении анализа aad-1 методом вестерн-блоттинга в экстракте контрольной кукурузы ХНН13 не были обнаружены иммунореактивные белки, так же как в образцах трансгенной кукурузы отсутствовали белки другой длины (агрегаты или продукты разложения).
Пример 5.4. Обнаружение посттрансляционного гликозилирования
Белок aad-1, выделенный из кукурузы и очищенный при помощи иммуноаффинной хроматографии, (фракция № 3) смешивали в отношении 4:1 с 5-кратным буфером Laemmli. Белок aad-1, выделенный из микроорганизма, ингибитор соевого трипсина, бычий сывороточный альбумин и пероксидазу хрена разводили водой Milli-Q примерно до концентрации белка aad-1, выделенного из растения, и смешивали с буфером Laemmli Bio-Rad. Затем белки нагревали при ~95°С в течение 5 минут и центрифугировали с ускорением 20000 × g в течение 2 минут для получения осветленного супернатанта. Полученные супернатанты наносили непосредственно на гель Bio-RadCriterion и подвергали электрофорезу с использованием рабочего буфера ХТ MES (Bio-Rad, Hercules, CA) в основном в соответствии с приведенным выше описанием за исключением того, что электрофорез выполняли при напряжении 170 В в течение ~60 минут. После электрофореза гель разрезали пополам и одну половину окрашивали синим красителем GelCode Blue до проявления всего белка в соответствии с инструкциями производителей. После окончания окрашивания гель сканировали денситометром Molecular Dynamics для визуальной регистрации геля. Другую половину геля окрашивали с помощью набора для окрашивания гликопротеина GelCode (Pierce Chemical, Rockford, IL) в соответствии с инструкциями производителей для визуализации гликопротеинов. Гликопротеины (с нижним пределом обнаружения, равным 0,625 нг на одну полосу) были визуализированы в виде пурпурных полос на светло-розовом фоне. После окончания окрашивания гликопротеинов гель сканировали при помощи цифрового сканирующего устройства Hewlett Packard для визуальной регистрации геля. После получения изображения окрашивания гликозилирования гель окрашивали синим красителем GelCode Blue для проверки наличия негликозилированных белков. Полученные результаты показали, что белки aad-1, выделенные из кукурузы и микроорганизма, не содержали обнаруживаемых ковалентно связанных углеводов. Данный результат был также подтвержден фингерпринтингом массы пептида.
Пример 5.5. Фингерпринтинг массы пептида при помощи масс-спектрометрии и секвенирование белка aad-1, выделенного из кукурузы и Pseudomonas
Белок aad-1, выделенный из Pseudomonas и кукурузы, анализировали при помощи масс-спектрометрии. Белок aad-1, выделенный из стебля трансгенной кукурузы (вариант DAS-40278-9), расщепляли трипсином в растворе и затем анализировали методами MALDI-TOF MS и ESI-LC/MS. Массы обнаруженных пептидов сравнивали с массами, установленными на основании потенциальных сайтов расщепления протеазой в последовательности белка aad-1, выделенного из кукурузы. Теоретическая модель расщепления была создана с использованием рабочей таблицы для анализа белка (PAWS) в программных средствах компании Proteometrics LLC. Денатурированный белок aad-1 легко расщепляется протеазами и образует многочисленные пептидные пики.
В белке aad-1, выделенном из трансгенной кукурузы (вариант DAS-40278-9) и расщепленном трипсином, обнаруженные пептидные фрагменты охватывали почти всю последовательность белка при отсутствии только одного небольшого триптического фрагмента у С-конца белка, от F248 до R253, и одного короткого (2 аминокислоты) пептидного фрагмента. Данный анализ подтвердил, что аминокислотная последовательность белка, выделенного из кукурузы, соответствует последовательности белка aad-1, выделенного из микроорганизма. Результаты указанных анализов показывают, что аминокислотная последовательность белка aad-1, выделенного из кукурузы, эквивалентна белку, экспрессированному P. fluorescens.
Пример 5.5.1. Секвенирование триптического фрагмента пептида
Помимо фингерпринтинга массы пептида, секвенировали аминокислотные остатки у N- и С-концов белка aad-1, выделенного из кукурузы, (выделенного методом иммунноаффинной хроматографии из варианта DAS-40278-9 кукурузы) и сравнивали с последовательностью белка, выделенного из микроорганизма. Последовательности белков были получены при помощи тандемной масс-спектрометрии для первых 11 остатков белков, выделенных из микроорганизма и кукурузы (таблица 8). Аминокислотные последовательности обоих белков представляли собой A’ H A A L S P L S Q R’’ (SEQ ID NO:30), свидетельствуя о том, что N-концевой метионин был удален аминопептидазой (таблица 8). Предполагалось, что N-концевая последовательность белка aad-1 должна представлять собой M’ A H A A L S P L S Q R’2 (SEQ ID NO:31). Полученные результаты позволяют предположить, что во время и после трансляции последовательностей в кукурузе и P. fluorescens N-концевой метионин отщепляется метионин-аминопептидазой (МАР). МАР быстро расщепляют метионильные остатки, если второй остаток белка имеет небольшую длину, такой как Gly, Ala, Ser, Cys, Thr, Pro и Val (Walsh, 2006). Было установлено, что, помимо удаления метионина, небольшая часть N-концевого пептида белка aad-1 была ацетилирована после отщепления N-концевого метионина (таблица 8). Такое явление часто имеет место в белках, экспрессированных эукариотическими (растительными) организмами, так как модифицировано примерно 80-90% N-концевых остатков (Polevoda and Sherman, 2003). Кроме того, было установлено, что белки, содержащие серин и аланин у N-конца, ацетилируются особенно часто (Polevoda and Sherman, 2002). Два указанных противоположных процесса, отщепление N-концевого остатка метионина и N-концевое ацетилирование, являются наиболее обычными модификациями и происходят в большинстве (~85%) эукариотических белков (Polevoda and Sherman, 2002). Однако примеры, демонстрирующие биологическую значимость, обусловленную N-концевым ацетилированием, являются достаточно редкими (Polevoda and Sherman, 2000).
Краткий обзор данных для N-концевых последовательностей белков aad-1, выделенных из кукурузы и микроорганизма
(SEQ ID NO:30)
(SEQ ID NO:32)
(SEQ ID NO:33)
(SEQ ID NO:34)
2Обнаруженные N-концевые последовательности белков aad-1, выделенных из P. fluorescens и кукурузы.
3Данные тандемной масс-спектрометрии для N-концевых пептидов выявили смесь последовательностей AHAALSPLSQR (ацетилированная) и N-ацетил-AHAALSPLSQR (ацетилированная). Также были обнаружены ”усеченные N-концы” (пептиды, соответствующие аминокислотным последовательностям HAALSPLSQR, AALSPLSQR И ALSPLSQR). Относительное содержание, представляющее собой оценку относительного количества пептидных фрагментов, было определено на основании площадей соответствующих пиков LC, измеренных при 214 нм.
Примечания
Надстрочные индексы (Rx) означают число аминокислотных остатков в последовательности.
Аббревиатуры аминокислотных остатков:
Помимо N-ацетилирования, было также обнаружено незначительное усечение N-конца, произошедшее во время очистки белка aad-1, выделенного из кукурузы (таблица 8). В результате усечения концов были утрачены аминокислоты А2, Н3 и А4 (в разных формах и количествах) в белке, выделенном из кукурузы. Такое усечение, предположительно, произошло во время очистки белка aad-1, так как зонд для вестерн-блоттинга к неочищенным экстрактам листьев содержал одну четкую полосу, соответствующую такой же молекулярной массе (MW), что и белок aad-1, выделенный из микроорганизма. Экстракционный буфер для образцов, исследуемых методом вестерн-блоттинга, содержал избыток смеси ингибиторов протеазы с широкой специфичностью для ингибирования серина, цистеина, аспарагиновой кислоты, а также металлопротеаз и аминопептидаз.
С-концевые последовательности белков aad-1, выделенных из кукурузы и микроорганизма, которые были определены в соответствии с приведенным выше описанием, сравнивали с предполагаемыми аминокислотными последовательностями (таблица 9). Полученные результаты показали, что обнаруженные последовательности были идентичны предполагаемым последовательностям и белки aad-1, выделенные из кукурузы и микроорганизма, были идентичны и не изменены у С-конца.
Краткий обзор данных для С-концевых последовательностей белков aad-1, выделенных из кукурузы и микроорганизма
2Обнаруженные С-концевые последовательности белков aad-1, выделенных из P. fluorescens и кукурузы.
Примечания:
Надстрочные индексы (Rx) означают число аминокислотных остатков в последовательности.
Аббревиатуры аминокислотных остатков:
Пример 6. Экспрессия в полевых условиях, анализ состава питательных веществ и агрономические характеристики гибридной линии кукурузы, содержащей вариант DAS-40278-9
Целью данного исследования было определение уровней белка AAD-1, обнаруженного в тканях кукурузы. Кроме того, был произведен анализ состава стеблей и зерна кукурузы для исследования соответствия между линией изогенной нетрансформированной кукурузы и линией трансгенной кукурузы DAS-40278-9 (которую не опрыскивали гербицидом, опрыскивали гербицидом 2,4-D, опрыскивали хизалофопом и опрыскивали 2,4-D и хизалофопом). Агрономические характеристики линии изогенной нетрансформированной кукурузы также сравнивали с кукурузой DAS-40278-9. Экспрессию в полевых условиях, состав и агрономические признаки определяли на шести опытных участках, расположенных в основных регионах выращивания кукурузы в США и Канаде. Указанные участки представляли регионы с разными агрономическими приемами возделывания кукурузы и окружающими условиями. Опытные участки были расположены в штатах Айова, Иллинойс (2 участка), Индиана, Небраска и в провинции Онтарио, Канада.
Средние значения, полученные на всех участках для образцов контрольных растений, неопрысканных растений AAD-1, растений AAD-1 + хизалофоп, растений AAD-1 + 2,4-D и растений AAD-1 + хизалофоп и 2,4-D, находились в пределах значений для кукурузы, приведенных в научной литературе. Было обнаружено ограниченное число значимых отличий между неопрысканной кукурузой AAD-1, кукурузой AAD-1 + хизалофоп, кукурузой AAD-1 + 2,4-D или кукурузой AAD-1 + хизалофоп и 2,4-D, но такие отличия не были признаны биологически значимыми, так как они были незначительными и находились в пределах диапазонов, установленных для промышленной кукурузы. Графики состава не указывают на какие-либо биологически значимые отличия, вызванные обработкой, у неопрысканной кукурузы AAD-1, кукурузы AAD-1 + хизалофоп, кукурузы AAD-1 + 2,4-D и кукурузы AAD-1 + хизалофоп и 2,4-D и линии контрольной кукурузы. В заключение следует отметить, что результаты, полученные для состава неопрысканной кукурузы AAD-1, кукурузы AAD-1 + хизалофоп, кукурузы AAD-1 + 2,4-D и кукурузы AAD-1 + хизалофоп и 2,4-D, подтвердили соответствие кукурузы AAD-1 (вариант DAS-40278-9) линиям обычной кукурузы.
Пример 6.1. Исследованные линии кукурузы
В таблице 10 представлены растения, выращенные из гибридных семян, содержащих вариант DAS-40278-9, и контрольные растения, выращенные из обычных гибридных семян, относящихся к той же генетической среде, что и исследуемая линия, но не содержащих вариант DAS-40278-9.
Вышеописанные растения кукурузы были выращены на участках, расположенных в основных регионах выращивания кукурузы в США и Канаде. Шесть производственных мощностей, предназначенных для выполнения исследований в полевых условиях, в Ричленде, шт. Айова, Карлайле, шт. Иллинойс, Вайоминге, шт. Иллинойс, Роквилле, шт. Индиана, Йорке, шт. Небраска, и Бранчтоне, провинция Онтарио, Канада (определяемые как IA, IL1, IL2, IN, NE и ON), представляют собой регионы с разными агрономическими приемами выращивания кукурузы и окружающими условиями.
Семена исследуемой и контрольной кукурузы сеяли при норме высева, равной 24 семенам/ряд, на расстоянии около 25 см (10 дюймов) в каждой ряду. На каждом участке были выделены 4 одинаковые делянки для каждой обработки, которые состояли из 2 рядов длиной 7,6 м (25 футов). Делянки были расположены по принципу рандомизированного полного повторения (RCB) с уникальной рандомизацией на каждом участке. Каждая делянка кукурузы была огорожена 2 рядами гибрида нетрансгенной кукурузы аналогичной спелости. Весь опытный участок был окружен как минимум 12 рядами (или 9 м (30 футов)) гибрида нетрансгенной кукурузы аналогичной относительной спелости.
Для получения агрономически приемлемой сельскохозяйственной культуры применяли соответствующие методы борьбы с насекомыми, сорняками и болезнями. Максимальные и минимальные месячные температуры с учетом осадков в виде дождей и полива имели средние значения для участка. Указанные диапазоны являются типичными при выращивании кукурузы.
Пример 6.2. Внесение гербицидов
Обработку гербицидами производили путем опрыскивания в объеме, равном примерно 187 л/га (20 галлонов/акр). Внесение гербицидов в таких количествах соответствует максимальной норме, применяемой в агрономической практике. В таблице 11 представлены использованные гербициды.
bai = активный ингредиент
Гербицид 2,4-D (Weedar 64) вносили разбросным методом в 3 приема, распыляя на опытные растения 4 и 5 (за сезон было внесено в общей сложности 3 фунта эквивалента кислоты/акр). Гербицид вносили до появления всходов и примерно на стадиях роста V4 и V8-V8.5. Нормы направленного внесения были равны 1,0 фунту эквивалента кислоты/акр для гербицида Weedar 64 или 1120 г эквивалента кислоты/га. Фактические нормы внесения составляли 1096-1231 г эквивалента кислоты/акр.
Хизалофоп (Assure II) вносили разбросным методом в один прием, распыляя на опытные растения 3 и 5. Гербицид вносили примерно на стадии роста V6. Норма направленного внесения для Assure II была равна 0,0825 фунта активного ингредиента/акр или 92 г активного ингредиента/га. Фактические нормы внесения составляли 90,8-103 г активного ингредиента/га.
Пример 6.3. Сбор агрономических данных и результаты исследования
Агрономические показатели регистрировали для всех опытных растений в блоках 2, 3 и 4 на каждом участке. В таблице 12 представлены измеренные характеристики.
Был произведен анализ данных, полученных для контрольных растений, неопрысканных растений AAD-1, растений AAD-1 + 2,4-D, растений AAD-1 + хизалофоп и растений AAD-1 + хизалофоп и 2,4-D. При анализе данных, полученных на разных участках, не было обнаружено статистически значимых различий в величинах начальной плотности растений (V1 и V4), мощности проростков, конечной плотности растений, повреждения культур, времени до выметывания пестичных столбиков, времени до сбрасывания пыльцы, ломкости стеблей, ломкости корней, заболеваемости, повреждения насекомыми, числа дней до созревания, высоты растений и жизнеспособности пыльцы (форма и цвет) (таблица 13). В отношении показателей сохранения свежести и высоты початка были обнаружены значимые парные t-критерии для контрольных растений и растений aad-1 + хизолофоп, которые не сопровождались значимым общим эффектом обработки или значениями Р, скорректированными при помощи коэффициента ложного обнаружения (FDR) (таблица 13).
Краткий обзор результатов анализа агрономических показателей на разных участках для кукурузы aad-1 с вариантом DAS-40278-9 (опрысканной и неопрысканной) и контрольной кукурузы
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
bСравнение растений, опрысканных и не опрысканных гербицидом, с контрольными растениями при помощи t-критерия.
cЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
dВизуальная оценка по шкале 1-9; 9 = высокие растения с большими устойчивыми листьями.
eШкала 0-100%; 0 = отсутствие повреждений и 100 = мертвое растение.
fКоличество единиц тепла, накопленных со времени посева.
gШкала 0-100%; % пыльцевых зерен с разрушенными стенками.
hШкала 0-100%; % пыльцевых зерен интенсивного желтого цвета.
iВизуальная оценка по шкале 1-9, в соответствии с которой 1 означает отсутствие видимой зеленой ткани.
jВизуальная оценка по шкале 1-9, в соответствии с которой 1 означает плохую устойчивость к болезням.
kВизуальная оценка по шкале 1-9, в соответствии с которой 1 означает плохую устойчивость к поражению насекомыми.
lNA = статистический анализ не был выполнен из-за отсутствия вариабельности в повторности или обработке опытных участков.
mСтатистическое отличие, определяемое значением Р <0,05.
Пример 6.4. Получение образцов
Для анализа экспрессии и состава были получены образцы, представленные в таблице 14.
Лист
Пыльцаb
Кореньb
Листb
Стебель
Все растение
Зерно
V9
R1
R1
R1
R4
R6
R6-созревание
3 листа
1 растение
1 растение
1 лист
2 растенияс
2 растенияс
1 початок
3
3
3
3
3
3
3
3
3
3
3
3
3
3
Зерно
R6-созревание
5 початков
1
1
bОбразцы пыльцы, корня и листьев были получены на стадии роста R1 с одного растения.
cДва растения измельчали, объединяли и отбирали для анализа экспрессии или 3 растения для анализа состава.
Пример 6.5. Определение белка aad-1 в образцах кукурузы
Образцы кукурузы исследовали для определения количества белка aad-1. Растворимый экстрагируемый белок aad-1 подвергали количественному анализу при помощи набора для твердофазного иммуноферментного анализа (ELISA), приобретенного в компании Beacon Analytical System, Inc. (Portland, ME).
Образцы тканей кукурузы выделяли из опытных растений и готовили для анализа экспрессии, для чего образцы крупно измельчали, лиофилизовали и тонко измельчали (при необходимости) в измельчителе Geno/Grinder (Certiprep, Metuchen, New Jersey). Пыльца не требовала дополнительного приготовления. Белок aad-1 экстрагировали из тканей кукурузы при помощи физиологического раствора с фосфатным буфером, содержащего детергент твин-20 (PBST) и 0,5% бычьего сывороточного альбумина (BSA). Указанный белок экстрагировали из пыльцы при помощи буфера, содержащего 0,5% PBST/BSA, 1 мг/мл аскорбата натрия и 2% коктейля ингибиторов протеазы. Экстракты ткани растения и пыльцы центрифугировали, водный супернатант собирали, разбавляли при необходимости соответствующим буфером и анализировали при помощи набора для исследования aad-1 методом ELISA в виде сэндвича. С помощью указанного набора выполняли следующие стадии. Аликвоту разведенного образца и биотинилированное моноклональное антитело против aad-1 инкубировали в лунках титрационного микропланшета, сенсибилизированного иммобилизованным моноклональным антителом против aad-1. Указанные антитела связываются с белком aad-1 в лунках и образуют ”сэндвич” с белком aad-1, связанным между растворимым и иммобилизованным антителом. Несвязанные образцы и конъюгат затем удаляли из планшета, промывая PBST. В лунки для инкубации добавляют избыточное количество конъюгата стрептавидина с ферментом (щелочной фосфатазой). В конце инкубации несвязанные реагенты удаляли из планшета промыванием. В результате последующего добавления ферментного субстрата образовался окрашенный продукт. Так как белок aad-1 был связан в сэндвиче антител, степень окрашивания соответствовала концентрации белка aad-1 в образце (то есть более низкая концентрация остатков вызывает более слабое окрашивание). Оптическую плотность при 405 нм измеряли при помощи спектрометра для прочтения планшетов V-max Molecular Devices или Max 190 Spectra. Строили калибровочную кривую и вычисляли концентрацию белка aad-1 в неизвестных образцах при помощи алгебраического уравнения регрессии с использованием программного обеспечения Soft-MAX Pro™, совместимым со спектрометром для прочтения планшетов. Образцы анализировали в дублированных лунках и регистрировали среднюю концентрацию для таких дублированных лунок.
Краткий обзор концентраций белка aad-1 (средние значения для участков) в разных матрицах кукурузы представлены в таблице 15. Средняя концентрация белка aad-1 составляли от 2,87 нг/мг сухой массы в корне на стадии R1 до 127 нг/мг в пыльце. Были получены одинаковые результаты экспрессии для делянок, которые не опрыскивали и опрыскивали гербицидом. Белок aad-1 не был обнаружен во всех контрольных образцах за исключением одного контрольного образца корня, полученного с участка в шт. Индиана.
Краткий обзор средних уровней концентрации белка aad-1, измеренных в тканях неопрысканной кукурузы aad-1, кукурузы aad-1 + хизалофоп, кукурузы aad-1 + 2,4-D и кукурузы aad-1 + хизалофоп и 2,4-D
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
13,3
14,2
12,3
6,89
7,16
7,09
4,75-24,5
4,98-26,7
4,07-22,5
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
5,38
6,37
6,52
1,84
2,41
2,38
2,52-9,15
3,03-10,9
3,11-11,1
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
5,70
5,99
6,06
1,63
1,90
2,27
2,70-7,78
2,40-9,42
1,55-10,2
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
108
113
112
29,9
30,2
32,6
52,2-146
37,5-137
45,4-162
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
3,09
3,92
2,87
1,80
2,03
1,23
0,56-6,06
0,91-7,62
1,09-5,56
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
7,16
7,32
6,84
2,84
2,46
2,31
3,05-11,6
2,36-10,6
2,25-10,3
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
4,61
5,16
4,55
2,22
2,53
1,77
0,75-8,77
0,83-10,2
1,30-8,21
AAD-1 + хизалофоп
AAD-1 + 2,4-D
AAD-1 + хизалофоп и 2,4-D
4,63
4,98
4,61
1,51
1,78
1,62
1,07-6,84
2,94-9,10
1,81-7,49
Пример 6.6. Анализ состава
Образцы стеблей и зерна кукурузы анализировали в отношении содержания питательных веществ путем выполнения разных исследований. Анализы стеблей включали определение золы, общего жира, влажности, белка, углевода, сырой клетчатки, клетчатки с кислым детергентом, клетчатки с нейтральным детергентом, кальция и фосфора. Анализы зерна включали определение общих показателей (золы, общего жира, влажности, белка, углевода, сырой клетчатки, клетчатки с кислым детергентом), клетчатки с нейтральным детергентом (NDF), минеральных веществ, аминокислот, жирных кислот, витаминов, вторичных метаболитов и вредных веществ. Результаты анализа питательных веществ в стебле и зерне кукурузы сравнивали со значениями, приведенными в научной литературе (см. публикации Watson, 1982 (4); Watson, 1984 (5); ILSI Crop Composition Database, 2006 (6); OECD Consensus Document on Compositional Considerations for maize, 2002 (7); и Codex Alimentarius Commision 2001 (8)).
Пример 6.6.1. Анализ общих показателей, клетчатки и минеральных веществ в стеблях кукурузы
Был выполнен анализ белка, жира, золы, влажности, углевода, ADF, NDF, кальция и фосфора в образцах стеблей кукурузы, полученных из контрольных растений, неопрысканных растений aad-1, растений aad-1 + хизалофоп, растений aad-1 + 2,4-D и растений aad-1 + хизалофоп и 2,4-D. Краткий обзор результатов для всех участков представлен в таблице 16. При выполнении анализа на всех участках и на одном участке средние значения для всех общих показателей, клетчатки и минеральных веществ находились в пределах диапазонов, указанных в научной литературе. При выполнении анализа на всех участках не было обнаружено статистически значимых различий между контрольными и трансгенными растениями по таким показателям как влажность, ADF, NDF, кальций и фосфор. В отношении белка и золы были обнаружены значимые парные t-критерии для неопрысканных растений AAD-1 (белок), растений aad-1 + хизалофоп (белок) и растений aad-1 + хизалофоп и 2,4-D (зола), которые не сопровождались значимыми общими эффектами обработки или значениями Р, скорректированными при помощи коэффициента ложного обнаружения (FDR). В отношении жира был обнаружен значимый парный t-критерий и скорректированное значение Р для растений aad-1 + хизалофоп по сравнению с контрольными растениями, но значимый общий эффект обработки отсутствовал. В отношении углеводов был обнаружен статистически значимый общий эффект обработки, парный t-критерий и значение Р, скорректированное при помощи FDR, у растений aad-1 + хизалофоп и контрольных растений. Кроме того, в отношении углеводов был обнаружен значимый парный t-критерий для неопрысканных растений aad-1, но отсутствовало значение Р, скорректированное при помощи FDR. Указанные различия не являются биологически значимыми, так как результаты анализов на всех участках для указанных анализируемых показателей находились в пределах диапазонов, указанных в научной литературе для кукурузы, и отличия от контрольных растений были незначительными (<23%).
Краткий обзор анализа общих показателей, клетчатки и минеральных веществ в стеблях кукурузы на всех участках
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
eСтатистически значимое отличие, определяемое значением Р <0,05.
Пример 6.6.2. Анализ общих показателей и клетчатки в зерне
Краткий обзор результатов анализа общих показателей (белка, жира, золы, влажности, холестерина и углеводов) и клетчатки (ADF, NDF и общего содержания клетчатки) в зерне кукурузы на всех участках представлен в таблице 17. Все результаты для общих показателей и клетчатки находились в пределах диапазонов, указанных в научной литературе, и при выполнении анализа на всех участках не были обнаружены значимые отличия между контрольными и трансгенными растениями в отношении жира, золы, NDF и общего содержания клетчатки. В отношении влажности был обнаружен значимый общий эффект обработки, который не сопровождался значимыми парными t-критериями или значениями Р, скорректированными при помощи FDR. В отношении ADF был обнаружен значимый парный t-критерий для растений aad-1 + хизалофоп и 2,4-D, но отсутствовал значимый общий эффект обработки или значение Р, скорректированное при помощи FDR. В отношении белка и углеводов были обнаружены значимые парные t-критерии, скорректированные значения Р и общие эффекты обработки для неопрысканных растений aad-1, растений aad-1 + хизалофоп и растений aad-1 + хизалофоп и 2,4-D. Так как указанные отличия были незначительными (<12%) и все значения находились в пределах диапазонов, указанных в научной литературе, то указанные отличия не были признаны биологически значимыми.
Краткий обзор анализа общих показателей и клетчатки в зерне кукурузы на всех участках
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
eСтатистически значимое отличие, определяемое значением Р <0,05.
fNR = данные в научной литературе отсутствуют.
gNA = статистический анализ не был выполнен, так как большинство данных было <LOQ.
hLOQ = предел количественного определения.
Пример 6.6.3. Анализ минеральных веществ в зерне
Образцы зерна кукурузы анализировали в отношении содержания минеральных веществ, включающих кальций, хром, медь, иод, железо, магний, марганец, молибден, фосфор, калий, селен, натрий и цинк. Краткий обзор результатов анализа для всех участков представлен в таблице 18. Все результаты находились в пределах диапазонов, указанных в научной литературе. При выполнении анализа на всех участках не было обнаружено значимых отличий в содержании кальция, меди, железа и калия. Средние результаты для хрома, иода, селена и натрия были ниже предела количественного определения. В отношении магния и фосфора были обнаружены значимые парные t-критерии для неопрысканных растений aad-1 и растений aad-1 + хизалофоп, которые не сопровождались значимыми общими эффектами обработки или значениями Р, скорректированными при помощи FDR. В отношении марганца и молибдена был обнаружен значимый парный t-критерий для неопрысканных растений aad-1, но при этом не были выявлены значимое значение Р, скорректированное при помощи FDR, и общий эффект обработки. Для растений aad-1 + хизалофоп и 2,4-D был обнаружен значимый парный t-критерий в отношении цинка, но отсутствовало значимое значение Р, скорректированное при помощи FDR, или общий эффект обработки. Кроме того, указанные отличия от контрольных растений были незначительными (<13%) и все значения находились в пределах диапазонов, указанных в научной литературе, когда такие данные были приведены.
Краткий обзор анализа минеральных веществ в зерне кукурузы на всех участках
(0,070, 0,177)
(0,067, 0,173)
(0,206, 0,362)
(0,046, 0,132)
(0,055, 0,150)
(0,085, 0,206)
(0,655, 0,782)
(0,722, 0,839)
(0,454, 0,638)
(0,108, 0,238)
(0,627, 0,768)
(0,027f, 0,085)
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
eNA = статистический анализ не был выполнен, так как большинство данных было <LOQ.
fСтатистически значимое отличие, определяемое значением Р <0,05.
gLOQ = предел количественного определения.
Пример 6.6.4. Анализ аминокислот в зерне
Образцы кукурузы анализировали в отношении содержания аминокислот в контрольных растениях кукурузы, неопрысканных растениях aad-1, растениях aad-1 + хизалофоп, растениях aad-1 + 2,4-D и растениях aad-1 + хизалофоп и 2,4-D; краткий анализ результатов, полученных для всех участков и для каждого участка, приведен в таблице 19. Уровни содержания аминокислот находились в пределах диапазонов, указанных в научной литературе, и при выполнении анализа на всех участках не было обнаружено значимых отличий в содержании аргинина, лизина и тирозина. Значимые отличия были обнаружены в отношении нескольких аминокислот при выполнении анализа на всех участках. В указанных случаях содержание аминокислот в контрольных растениях было ниже, чем в линиях трансгенных растений aad-1, что может быть связано с общим более низким содержанием белка в контрольном зерне по сравнению с линиями aad-1. Для неопрысканных растений aad-1 были обнаружены значимые общие эффекты обработки наряду со значимыми парными t-критериями и значениями Р, скорректированными при помощи FDR, в отношении всех аминокислот за исключением аргинина, глицина, лизина, триптофана и тирозина. Для растений aad-1 + хизалофоп были обнаружены значимые общие эффекты обработки наряду со значимыми парными t-критериями и значениями Р, скорректированными при помощи FDR, в отношении всех аминокислот за исключением аргинина, цистеина, глицина, лизина, триптофана и тирозина. Для растений aad-1 + 2,4-D были обнаружены значимые общие эффекты обработки наряду со значимыми парными t-критериями (со значимыми значениями Р, скорректированными при помощи FDR) в отношении всех аминокислот за исключением аргинина, аспарагиновой кислоты, глицина, гистидина, лизина, тирозина и валина. Для растений aad-1 + хизалофоп и 2,4-D были обнаружены значимые общие эффекты обработки наряду со значимыми парными t-критериями и значениями Р, скорректированными при помощи FDR, в отношении всех аминокислот за исключением аргинина, глицина, лизина, серина, триптофана и тирозина. Хотя было обнаружено много отличий в содержании аиминокислот, такие отличия были незначительными (<15%), не были обнаружены на всех участках и все средние значения находились в пределах диапазонов, указанных в научной литературе.
Краткий обзор анализа аминокислот в зерне кукурузы на всех участках
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
еСтатистически значимое отличие, определяемое значением Р <0,05.
Пример 6.6.5. Анализ жирных кислот в зерне
Образцы зерна кукурузы анализировали в отношении содержания жирных кислот. Краткий обзор результатов, полученных для всех участков, представлен в таблице 20. Все результаты, полученные для контрольных растений, неопрысканных растений aad-1, растений aad-1 + хизалофоп, растений aad-1 + 2,4-D и растений aad-1 + хизалофоп и 2,4-D, анализируемых в отношении содержания жирных кислот, находились в пределах диапазонов, указанных в опубликованной научной литературе. Результаты, полученные для каприловой кислоты (8:0), каприновой кислоты (10:0), лауриловой кислоты (12:0), миристиновой кислоты (14:0), миристолеиновой кислоты (14:1), пентадекановой кислоты (15:0), пентадеценовой кислоты (15:1), гептадекановой кислоты (17:0), гептадеценовой кислоты (17:1), гамма-линоленовой кислоты (18:3), эйкозадиеновой кислоты (20:2), эйкозатриеновой кислоты (20:3) и арахидоновой кислоты (20:4), были ниже предела количественного определения (LOQ) используемого метода. При выполнении анализа на всех участках не было обнаружено значимых отличий для пальмитиновой кислоты (16:0), пальмитолеиновой кислоты (16:1), стеариновой кислоты (18:0), линолевой кислоты (18:2), линоленовой кислоты (18:3) и арахидоновой кислоты (20:0). В отношении олеиновой кислоты (18:1) и эйкозеновой кислоты (20:1) были обнаружены значимые парные t-критерии для неопрысканных растений aad-1 (18:1) и растений aad-1 + 2,4-D (18:1 и 20:1), которые не сопровождались значимыми общими эффектами обработки и значениями Р, скорректированными при помощи FDR. В отношении бегеновой кислоты (22:0) был обнаружен значимый общий эффект обработки и значимые парные t-критерии для растений aad-1 + 2,4-D и растений aad-1 + хизалофоп и 2,4-D при отсутствии значимых значений Р, скорректированных при помощи FDR.
Краткий обзор анализа жирных кислот в зерне кукурузы на всех участках
bОбъединенный диапазон.
cОбщий эффект обработки, определенный при помощи F-критерия.
dСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
eЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
gNA = статистический анализ не был выполнен, так как большинство данных было <LOQ.
gСтатистически значимое отличие, определяемое значением Р <0,05.
Пример 6.6.6. Анализ витаминов в зерне
Содержание витаминов А, В1, В2, В5, В6, В12, С, D, Е, ниацина и фолиевой кислоты в образцах зерна кукурузы определяли для контрольных растений, неопрысканных растений aad-1, растений aad-1 + хизалофоп, растений aad-1 + 2,4-D и растений aad-1 + хизалофоп и 2,4-D. Краткий обзор результатов для всех участков представлен в таблице 21. Витамины В12, D и Е не поддавались количественному определению используемыми аналитическими методами. Все средние значения содержания витаминов были аналогичны значениям, указанным в научной литературе, когда такие данные имелись. Результаты для витаминов, для которых в научной литературе не были указаны диапазоны содержания (витамины В5 и С), были аналогичны значениям, полученным для контрольных растений (отличие <22% от контрольных растений). При выполнении анализа на всех участках не были обнаружены статистически значимые отличия за исключением витаминов В1, С и ниацина. Значимые парные t-критерии были обнаружены в отношении витаминов В1 для контрольных растений, неопрысканных растений aad-1, растений aad-1 + хизалофоп и растений aad-1 + хизалофоп и 2,4-D, которые не сопровождались значимыми общими эффектами обработки или значениями Р, скорректированными при помощи FDR. В отношении витамина С был обнаружен значимый общий эффект обработки наряду со значимыми парными t-критериями и значениями Р, скорректированными при помощи FDR, для растений aad-1 + хизалофоп и растений aad-1 + 2,4-D. Аналогичным образом в отношении ниацина был обнаружен значимый общий эффект обработки наряду со значимыми парными t-критериями и значениями Р, скорректированными при помощи FDR, для растений aad-1 + хизалофоп и растений aad-1 + хизалофоп и 2,4-D. Значимый парный t-критерий для растений aad-1 + 2,4-D был также обнаружен в отношении ниацина для растений aad-1 + 2,4-D, который не сопровождался значимым общим эффектом обработки или значением Р, скорректированным при помощи FDR. Так как отличия не были обнаружены на всех участках и значения находились в пределах диапазонов, указанных в научной литературе (когда такие данные имелись), такие отличия не были признаны биологически значимыми.
Краткий обзор анализа витаминов в зерне кукурузы на всех участках
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
eСтатистически значимое отличие, определяемое значением Р <0,05.
fNR = данные в научной литературе отсутствуют.
gNA = статистический анализ не был выполнен, так как большинство данных было <LOQ.
hLOQ = предел количественного определения.
Пример 6.6.7. Анализ вредных веществ и вторичных метаболитов в зерне
Содержание вторичных метаболитов (кумаровая кислота, феруловая кислота, фурфурал и инозит) и вредных веществ (фитиновая кислота, раффиноза и ингибитор трипсина) в образцах зерна кукурузы определяли для контрольных растений, неопрысканных растений aad-1, растений aad-1 + хизалофоп, растений aad-1 + 2,4-D и растений aad-1 + хизалофоп и 2,4-D. Краткий обзор результатов для всех участков представлен в таблицах 22 и 23. При выполнении анализа на всех участках полученные значения находились в пределах диапазонов, указанных в научной литературе. Не было обнаружено значимых отличий между растениями aad-1 и контрольными растениями при выполнении анализа на всех участках в отношении инозита и ингибитора трипсина. Результаты, полученные для фурфурала и раффинозы были ниже предела количественного определения используемого метода. Значимые парные t-критерии были обнаружены для кумаровой кислоты (неопрысканные растения aad-1, растения aad-1 + 2,4-D и растения aad-1 + хизалофоп и 2,4-D) и феруловой кислоты (растения aad-1 + хизалофоп и растения aad-1 + хизалофоп и 2,4-D). Указанные отличия не сопровождались значимыми общими эффектами обработки или значениями Р, скорректированными при помощи FDR, и были аналогичны результатам, полученным для контрольных растений (отличие <10%). Значимый общий эффект обработки, парный t-критерий и значение Р, скорректированного при помощи FDR, были обнаружены для фитиновой кислоты (неопрысканные растения aad-1). Так как все результаты находились в пределах диапазонов, указанных в научной литературе, и были аналогичны результатам, полученным для контрольных растений (отличие <11%), такие отличия не были признаны биологически значимыми.
Краткий обзор анализа вторичных метаболитов в зерне кукурузы на всех участках
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
eСтатистически значимое отличие, определяемое значением Р <0,05.
fNA = статистический анализ не был выполнен, так как большинство данных было <LOQ.
gLOQ = предел количественного определения.
Краткий обзор анализа вредных веществ в зерне кукурузы на всех участках
bОбщий эффект обработки, определенный при помощи F-критерия.
cСравнение обработки трансгенных растений и контрольных растений при помощи t-критериев.
dЗначения Р, скорректированные при помощи коэффициента ложного обнаружения (FDR).
eСтатистически значимое отличие, определяемое значением Р<0,05.
fNA = статистический анализ не был выполнен, так как большинство данных было <LOQ.
gLOQ = предел количественного определения.
Пример 7. Дополнительные агрономические исследования
Агрономические характеристики линии кукурузы 40278 исследовали в сравнении с близкой изолинией кукурузы в разных окружающих условиях. В исследовании были использованы 4 генетически разных гибрида и соответствующие близкие изолинии контрольных гибридов, выращиваемых в общей сложности на 21 участке.
Четыре опытных гибрида представляли собой среднеспелые и позднеспелые гибриды при относительном созревании от 99 до 113 дней. В эксперименте А был исследован вариант DAS-40278-9 в генетической среде, включающей инбредную форму С с конверсией BC3S1. Указанный гибрид с относительным созреванием в течение 109 дней был исследован на 16 участках (таблица 24). В эксперименте В был исследован гибрид в генетической среде, включающей инбредную форму Е с конверсией BC3S1, характеризующийся относительным созреванием в течение 113 дней. Указанный гибрид исследовали на 14 участках, используя несколько другую совокупность участков, чем в эксперименте А (таблица 24). В экспериментах С и D исследовали гибриды в генетических средах, включающих соответственно инбредную форму D с конверсией BC2S1 и инбредную форму F с конверсией BC2S1. Оба указанных гибрида с относительным созреванием в течение 99 дней исследовали на тех же 10 участках.
Участки, на которых были проведены агрономические исследования
Для каждого исследования использовали схему рандомизированных блоков при наличии двух одинаковых делянок на одном участке с двухрядным расположением растений. Длина ряда была равна 6 м (20 футов), при этом в каждом ряду было посеяно 34 семени. Исследования выполняли в соответствии со стандартными агрономическими приемами возделывания кукурузы в указанных регионах.
Полученные данные анализировали по восьми агрономическим характеристикам: высота растения, высота початка, ломкость стеблей, ломкость корней, конечная плотность растений, влажность зерна, натурная масса зерна и урожайность. Такие параметры, как высота растения и высота початка, позволяют получить информацию о внешнем виде гибридов. Агрономические характеристики, включающие процентное значение ломкости стеблей и ломкости корней, позволяют определить возможности уборки гибрида. Конечная плотность растений позволяет определить качество семян и сезонные условия выращивания, влияющие на урожайность. Процентное значение влажности зерна во время уборки урожая определяет созревание гибрида, урожайность (бушель/акр, скорректированные в отношении влажности) и натурная масса зерна (масса в фунтах бушеля кукурузы, скорректированная в отношении 15,5% влажности) позволяют определить воспроизводительную способность гибрида.
Анализ изменчивости выполняли на полевых участках с использованием линейной модели. Гибриды и участки включали в указанную модель в виде фиксированных величин. Были использованы смешанные модели, в которых участки и гибриды представляли собой произвольные величины, но местоположение гибридов представляет собой лишь небольшую часть изменчивости, и данный компонент изменчивости часто существенно не отличался от нуля. При определении ломкости стеблей и корней была использована логарифмическая трансформация для стабилизации изменчивости, однако, средние значения и диапазоны представлены по первоначальной шкале. Значимые отличия были определены на 95% доверительном уровне. Значимость общего эффекта обработки оценивали при помощи t-критерия.
Результаты исследований указанных агрономических характеристик приведены в таблице 25. Не было обнаружено статистически значимых отличий для всех четырех гибридов 40278 по сравнению с контрольными изолиниями (при р<0,05) в отношении параметров высоты початка, ломкости стеблей, ломкости корней, влажности зерна, натурной массы зерна и урожайности. Показатели конечной плотности и высоты растений имели статистически значимые отличия соответственно в экспериментах А и В, но подобные отличия не были обнаружены при сравнении с другими исследованными гибридами 40278. Некоторая обнаруженная изменчивость может быть связана с низкими уровнями генетической изменчивости, сохраняющейся при обратном скрещивании варианта DAS-40278-9 в элитных инбредных линиях. Общий диапазон значений измеренных параметров находится в пределах значений, полученных для традиционных гибридов кукурузы, и не позволяет сделать вывод о повышенной засоренности сорняками. Таким образом, данные агрономического исследования показывают, что кукуруза 40278 биологически соответствует обычной кукурузе.
Анализ агрономических характеристик
Контрольные растения
95,41
95,00
98,00
Контрольные растения
44,42
40,00
47,00
Контрольные растения
2,49
0,00
28,57
Контрольные растения
0,89
0,00
28,33
Контрольные растения
32,17
27,00
36,00
Контрольные растения
21,84
14,52
31,00
Контрольные растения
54,66
51,00
56,80
Контрольные растения
187,05
99,87
256,72
Контрольные растения
100,79
95,00
104,00
Контрольные растения
45,63
38,00
50,00
Контрольные растения
0,72
0,00
22,22
Контрольные растения
0,40
0,00
9,09
Контрольные растения
30,98
25,00
35,00
Контрольные растения
23,72
13,39
31,10
Контрольные растения
56,67
52,00
60,10
Контрольные растения
205,41
95,35
259,03
Контрольные растения
90,92
90,00
90,00
Контрольные растения
43,75
37,00
46,00
Контрольные растения
5,46
0,00
28,12
Контрольные растения
0,3077
0,00
33,33
Контрольные растения
31,68
24,00
35,00
Контрольные растения
19,36
13,66
26,50
Контрольные растения
55,14
52,80
58,40
Контрольные растения
200,35
129,02
285,58
Контрольные растения
4,17
0,00
39,06
Контрольные растения
31,86
29,00
35,00
Контрольные растения
19,32
13,35
25,70
Контрольные растения
55,50
52,70
57,40
Контрольные растения
199,82
118,56
264,11
Пример 8. Использование сайта инсерции варианта DAS-40278-9 кукурузы для направленной интеграции
Сопоставимая агрономическая продуктивность трансгена варианта DAS-40278-9 кукурузы на протяжении нескольких поколений в полевых условиях позволяет предположить, что области, идентифицированные вокруг сайта инсерции варианта DAS-40278-9 обеспечивают хорошую локализацию в геноме для направленной интеграции других представляющих интерес трансгенных генов. Такая направленная интеграция позволяет решить проблемы, связанные с так называемым ”эффектом положения” и риском создания мутации в геноме при введении трансгена в хозяина. Другие преимущества такой направленной интеграции включают, не ограничиваясь ими, уменьшение большого числа трансформационных вариантов, которые необходимо исследовать и испытывать, прежде чем будет получено трансгенное растение, обладающее требуемым уровнем экспрессии трансгена без возникновения каких-либо нарушений вследствие неправильного введения трансгена в важных локус генома хозяина. Кроме того, такая направленная интеграция позволяет объединять трансгены с созданием линий элитных растений, в которых оба гена функционируют более эффективно.
В соответствии с настоящим изобретением квалифицированный специалист может направленно вводить представляющие интерес полинуклеиновые кислоты в тот же сайт инсерции хромосомы 2, что и вариант DAS-40278-9 кукурузы или в сайт, расположенный в непосредственной близости от данного сайта инсерции в варианте DAS-40278-9 кукурузы. Один такой метод описан в международной заявке на патент № WO2008/021207, которая полностью включена в настоящее описание изобретения в качестве ссылки.
Для фланкирования представляющего интерес одного или нескольких генов, вводимых в хромосому 2 генома путем гомологичной рекомбинации, может быть использована геномная последовательность длиной до 20 т.п.о., фланкирующая 5'-конец сайта инсерции, и геномная последовательность длиной до 20 т.п.о., фланкирующая 3'-конец сайта инсерции (части которых определены со ссылкой на SEQ ID NO:29). Один или несколько представляющих интерес генов могут быть введены непосредственно в сайт инсерции варианта DAS-40278-9 кукурузы или могут быть локализованы в любом положении областей, включающих 20 т.п.о., окружающих сайт инсерции варианта DAS-40278-9 кукурузы, для достижения сопоставимого уровня экспрессии трансгена без вредного влияния на растение. ДНК-векторы, содержащие один или несколько представляющих интерес генов и фланкирующие последовательности, могут быть введены в растительные клетки несколькими методами, известными специалистам в данной области, которые включают, не ограничиваясь ими, трансформацию, опосредованную Agrobacterium. Введение донорского ДНК-вектора в сайт-мишень варианта DAS-40278-9 кукурузы может быть далее усилено несколькими методами, которые включают, не ограничиваясь ими, коэкспрессию или увеличение числа генов, усиливающих рекомбинацию, или уменьшение числа генов супрессии эндогенной рекомбинации. Кроме того, в данной области известно, что специфическое расщепление двухцепочечных последовательностей в геноме может быть использовано для увеличения частоты гомологичной рекомбинации, поэтому введение в сайт инсерции варианта DAS-40278-9 кукурузы и его фланкирующих областей может быть усилено экспрессией природных или созданных эндонуклеаз, специфичных к последовательности, предназначенных для расщепления указанных последовательностей. Таким образом, в соответствии с настоящим изобретением любая гетерологичная нуклеиновая кислота может быть введена в сайт-мишень хромосомы 2, расположенный между 5'-концевым молекулярным маркером и 3'-концевым молекулярным маркером, рассмотренными в примере 4, предпочтительно в SEQ ID NO:29, и/или в области, рассмотренные в настоящем описании изобретения.
Пример 9. Удаление кассеты экспрессии гена pat из варианта DAS-40278-9 кукурузы
Удаление кассеты экспрессии селектируемого гена-маркера благоприятно воздействует на направленную инсерцию в геномные локусы варианта DAS-40278-9 кукурузы. Удаление селектируемого маркера pat из варианта DAS-40278-9 кукурузы позволяет повторно использовать селектируемый маркер pat для направленной интеграции полинуклеиновых кислот в хромосому 4 в последующих поколениях кукурузы.
В соответствии с настоящим изобретением квалифицированный специалист может удалить представляющие интерес полинуклеиновые кислоты из варианта DAS-40278-9 кукурузы. Один такой метод описан в предварительной заявке на патент США № 61/297628, которая полностью включена в настоящее описание изобретения в качестве ссылки.
Созданы эндонуклеазы, специфичные к последовательности, такие как нуклеазы цинкосодержащей пальцеобразной области, которые узнают, связывают и расщепляют специфические последовательности ДНК, фланкирующие кассету экспрессии гена. Нуклеазы цинкосодержащей пальцеобразной области вводят в растительную клетку, скрещивая родительское растение, содержащее кассеты экспрессии нуклеазы цинкосодержащей пальцеобразной области трансгена, со вторым родительским растением, содержащим вариант DAS-40278-9 кукурузы. Полученное потомство выращивают до достижения зрелости и анализируют в отношении утраты кассеты экспрессии pat путем окрашивания листа гербицидом, содержащим глюфозинат. Растения-потомки, не устойчивые к указанному гербициду, подтверждают при помощи молекулярных исследований и используют для самоопыления. Эксцизию и удаление кассеты экспрессии pat подтверждают молекулярными методами в потомстве, полученном в результате самоопыления. В соответствии с настоящим изобретением можно удалить любую гетерологичную нуклеиновую кислоту из хромосомы 2 кукурузы на сайте-мишени, расположенном между 5’-концевым молекулярным маркером и 3'-концевым молекулярным маркером, рассмотренными в примере 4, предпочтительно в SEQ ID NO:29, или в указанных областях.
Пример 10. Сопротивление ломкости
Ломкость означает повреждение стеблей кукурузы под воздействием сильных ветров после внесения гербицидов, регулирующих рост растений, обычно во время периодов быстрого роста. Гибридную кукурузу, содержащую вариант DAS-40278-9, и контрольные растения, не содержащие варианта DAS-40278-9, испытывали методом механического ”толкания”, используя брус для физического воздействия на кукурузу, имитирующего повреждение вследствие сильных ветров. Обработку производили в четырех разных географических регионах и повторяли четыре раза (за исключением одного опыта, который повторяли только три раза). Делянки состояли из восьми рядов: четыре ряда занимал каждый из двух гибридов, при этом два ряда занимала кукуруза, содержащая вариант DAS-40278-9, и два ряда занимала кукуруза без указанного варианта. Длина каждого ряда была равна 6,1 м (20 футов). Растения кукурузы выращивали до стадии развития V4 и вносили коммерческий гербицид, содержащий 2,4-D (Weedar 64, Nufarm Inc., Burr Ridge, IL) при нормах внесения 1120 г эквивалента кислоты/га, 2240 г эквивалента кислоты/га и 4480 г эквивалента кислоты/га. Через семь дней после внесения гербицида проводили испытание методом механического толкания. В соответствии с испытанием на ломкость методом механического толкания брус длиной 1,2 м (4 фута) тянули вдоль двух рядов кукурузы, имитируя повреждение ветром. Высота бруса и скорость движения были заданы с возможностью повреждения стеблей на низком уровне (10% или меньше) по сравнению с растениями, не подвергнутыми воздействию, с целью создания достаточно строгих условий испытания, позволяющих продемонстрировать разницу между воздействиями. Направление испытания на ломкость было противоположным наклону кукурузы.
Два опытных участка подверглись воздействию сильного ветра и грозы через 2-3 дня после внесения гербицида 2,4-D. Через два дня гроза началась на участке Хаксли, шт. Айова. Скорость ветра достигала 2-17 м/сек-1, при этом на полевом участке была зарегистрирована скорость ветра, равная 33 м/сек-1. Направление ветра менялось. Через день гроза достигла участка Лейнсборо, шт. Миннесота. На данном полевом участке была зарегистрирована большая скорость ветра. Кроме того, обе грозы сопровождались дождем. Дождь вместе с ветром вызвал зарегистрированные повреждения, обусловленные ломкостью.
Обусловленные ломкостью повреждения, возникшие в результате испытания методом механического толкания (и ненастья), оценивали визуально в виде процентного значения повреждения. До выполнения механического испытания на ломкость с помощью бруса производили подсчет густоты стояния гибридной кукурузы, содержащей вариант DAS-40278-9, и контрольных растений. Через несколько дней после испытания на ломкость снова производили подсчет густоты стояния растений. Определяли процентное значение наклона и процентное значение уменьшения густоты стояния на делянке (таблица 26). Данные, полученные в результате выполнения указанных испытаний, показали, что гибридная кукуруза, содержащая вариант DAS-40278-9, была менее ломкой по сравнению с контрольными растениями после внесения гербицида 2,4-D.
Сопротивление ломкости кукурузы, содержащей вариант DAS-40278-9, на стадии роста V4 после внесения гербицида 2,4-D. Процентное значение ломкости вычисляли для растений гибридной кукурузы, содержащих вариант DAS-40278-9, и полученный данные сравнивали с контрольными растениями, не содержащими указанный вариант.
Среднее % значение наклона через 7-8 дней после внесения гербицида1
Среднее3 % значение наклона через 11-14 дней после внесения гербицида
Среднее3 % значение уменьшения густоты стояния растений через 11-14 дней после внесения гербицида
2Обработку повторяли четыре раза при рандомизированном расположении делянок (только в одном испытании были произведены три повторные обработки).
3Средние значения, скорректированные относительно природных явлений, имевших место на участках произрастания необработанных растений (термин ”необработанный” означает приближение к нулевой обработке).
Пример 11. Анализ белка в зерне
Из гибридной кукурузы, содержащей вариант DAS-40278-9, было получено зерно с более высоким общим содержанием белка по сравнению с контрольными растениями, не содержащими указанное вариант. Были выполнены два последовательных полевых испытания на нескольких участках, которые включали кукурузу, которую не опрыскивали гербицидами, и кукурузу, обработанную тремя разными комбинациями гербицидов. В 7 из 8 статистически значимых сравнений зерно, полученное из кукурузы, содержащей вариант DAS-40278-9, характеризовалось значительно более высоким общим содержанием белка (таблица 27). Полученные данные подтверждены анализами отдельных аминокислот.
Содержание белка в зерне, полученном при выполнении полевых испытаний на нескольких участках
Пример 12. Дополнительные агрономические испытания
Агрономические характеристики гибридной кукурузы, содержащей вариант DAS-40278-9, по сравнению с близкой изолинией кукурузы были получены в результате выполнения нескольких полевых испытаний в разных географических регионах в течение вегетационного периода. Полученные данные анализировали для определения агрономических характеристик в соответствии с описанием, приведенным в примере 7. Результаты сравнения линий гибридной кукурузы, содержащей вариант DAS-40278-9, с контрольными растениями приведены в таблице 28. Кроме того, в таблице 29 представлены агрономические характеристики для линий гибридной кукурузы, содержащей вариант DAS-40278-9, и контрольных растений, которые опрыскивали гербицидом хизалофопом (280 г эквивалента кислоты/га) на стадии развития V3, и гербицидом 2,4-D (2240 г эквивалента кислоты/га) на стадии развития V6.
Урожайность, процентное значение влажности и конечная плотность растений гибридной кукурузы, содержащей вариант DAS-40278-9, по сравнению с близкой изолинией контрольной кукурузы
Урожайность, процентное значение влажности, процентное значение ломкости стеблей, процентное значение ломкости корней и общая плотность растений линий гибридной кукурузы, содержащей вариант DAS-40278-9, по сравнению с близкой изолинией контрольной кукурузы
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ БОРЬБЫ С СОРНЯКАМИ С ИСПОЛЬЗОВАНИЕМ РАСТЕНИЙ AAD-1 И ПРЕДПОСЕВНОГО И/ИЛИ ДОВСХОДОВОГО ВНЕСЕНИЯ ГЕРБИЦИДОВ | 2012 |
|
RU2610664C2 |
| СОБЫТИЕ 416 AAD-12, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ СОИ И ИХ СОБЫТИЕСПЕЦИФИЧНАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2603252C2 |
| ДЕТЕКЦИЯ AAD-1 ОБЪЕКТА DAS-40278-9 | 2010 |
|
RU2577143C2 |
| ДЕТЕКТИРОВАНИЕ AAD-12-СОБЫТИЯ 416 У СОИ | 2010 |
|
RU2573898C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| ОБЪЕКТ СОИ 9582.814.19.1, ПРИДАЮЩИЙ УСТОЙЧИВОСТЬ К НАСЕКОМЫМ И УСТОЙЧИВОСТЬ К ГЕРБИЦИДАМ | 2012 |
|
RU2628099C2 |
| СПОСОБ ОБНАРУЖЕНИЯ ТРАНСФОРМАНТА СОИ рDAB9582.814.19.1 | 2012 |
|
RU2622079C2 |
| УСТОЙЧИВЫЙ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ И ТОЛЕРАНТНЫЙ К ГЕРБИЦИДАМ СЕЛЕКЦИОННЫЙ ГИБРИД ТРАНСФОРМАНТА СОИ pDAB9582.814.19.1 и pDAB4468.04.16.1 | 2012 |
|
RU2627161C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УРОЖАЙНОСТИ РЕЗИСТЕНТНЫХ К 2,4-D СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР | 2013 |
|
RU2628504C2 |


















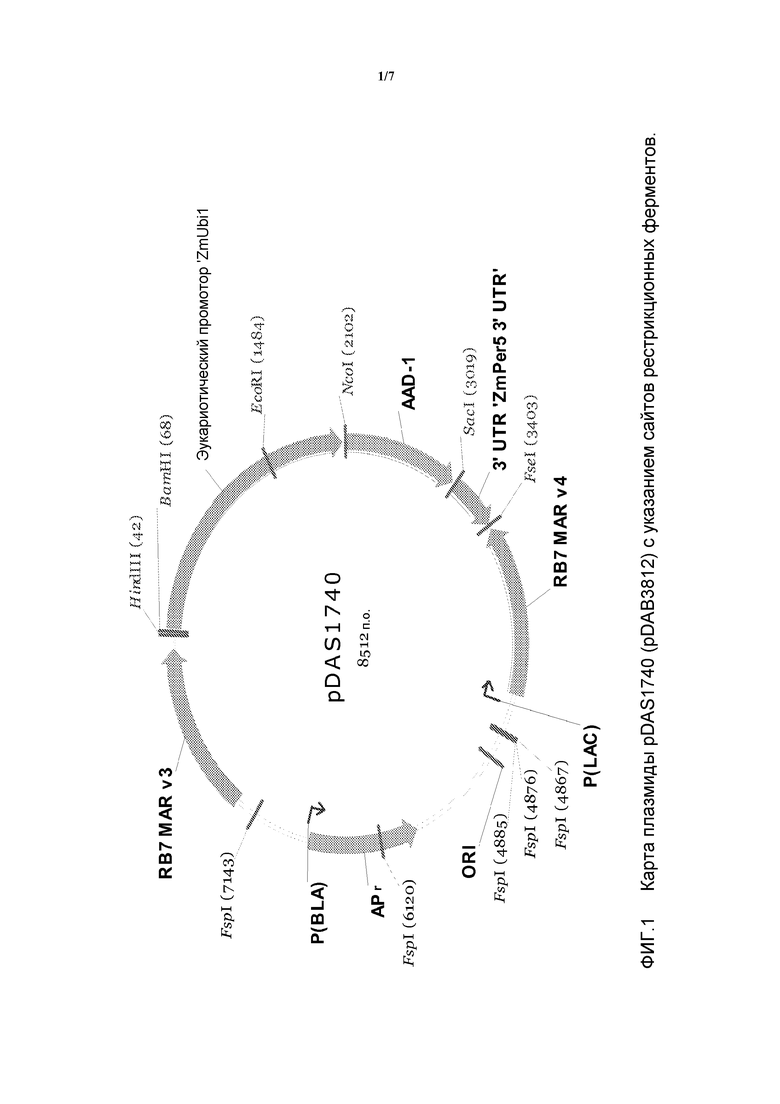
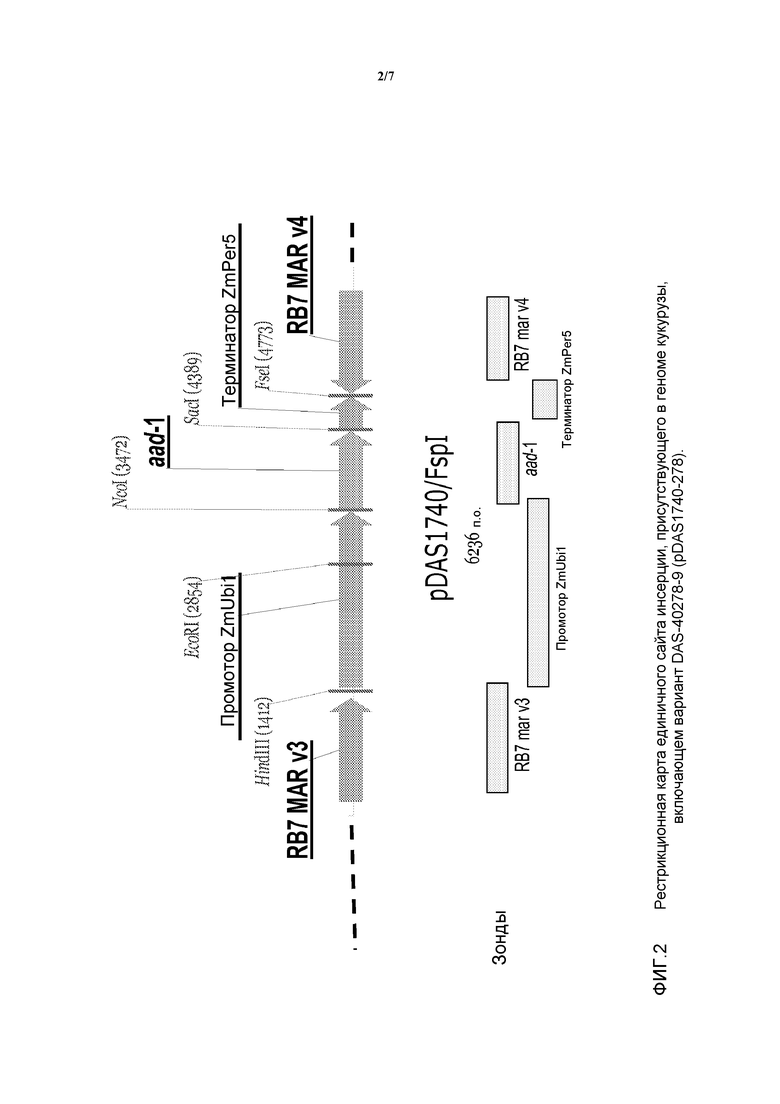
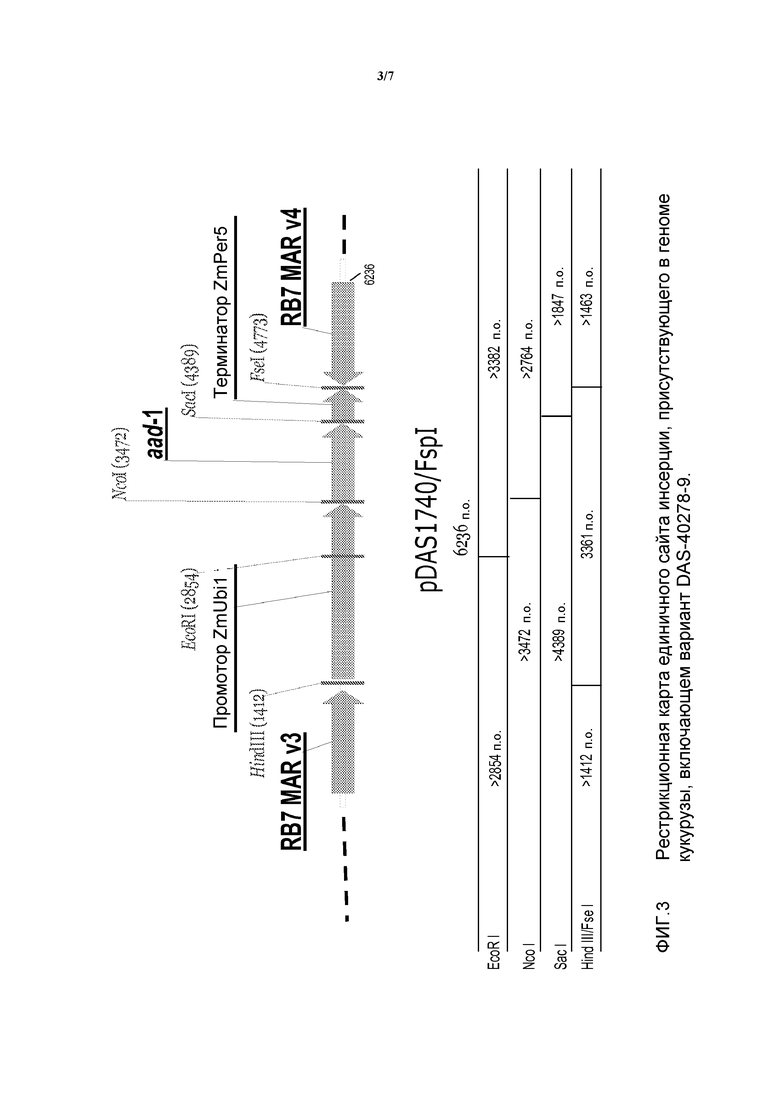
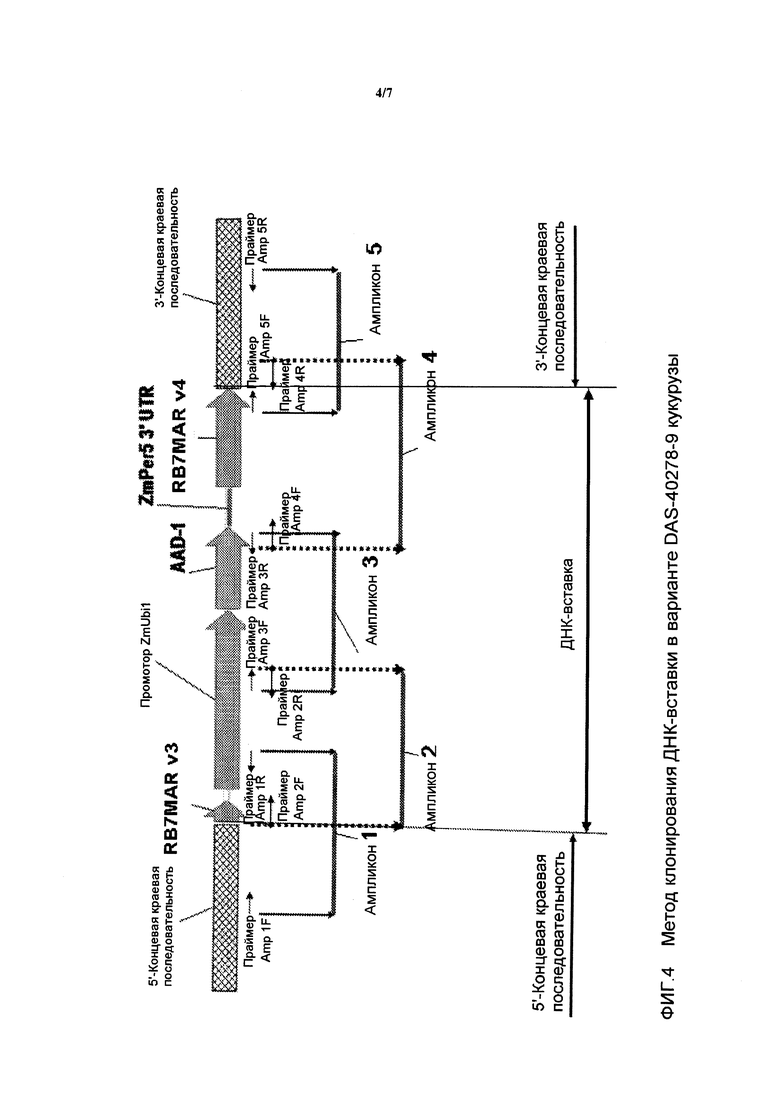
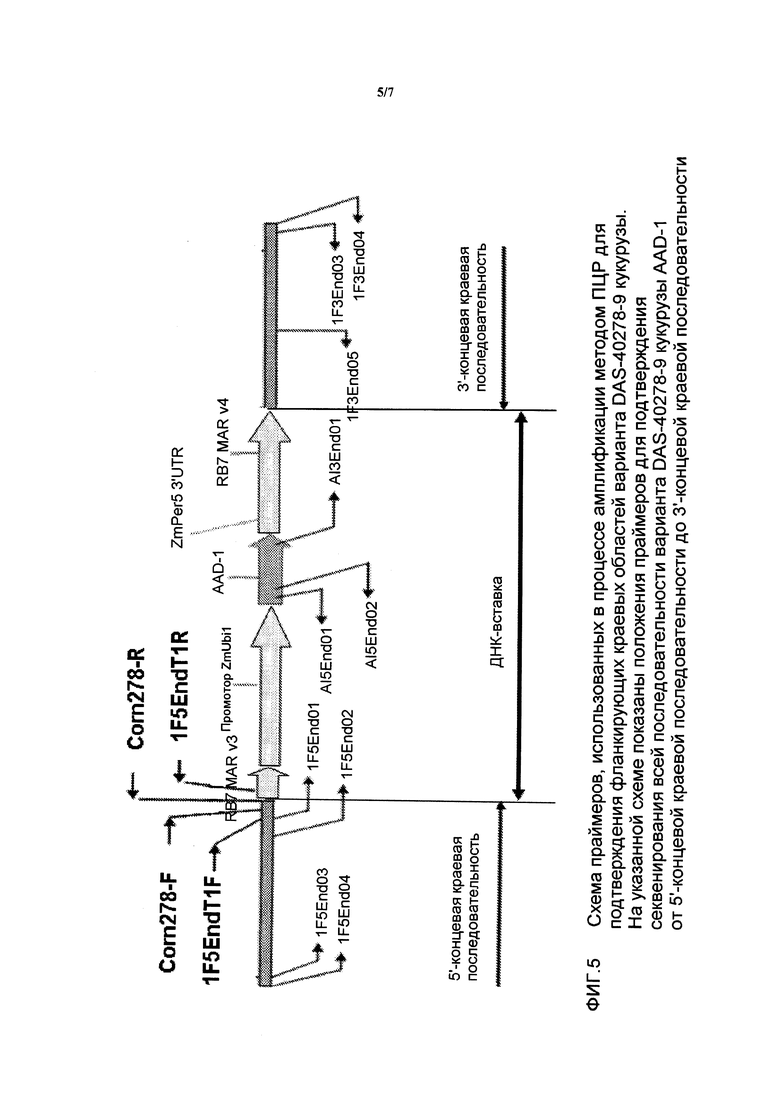

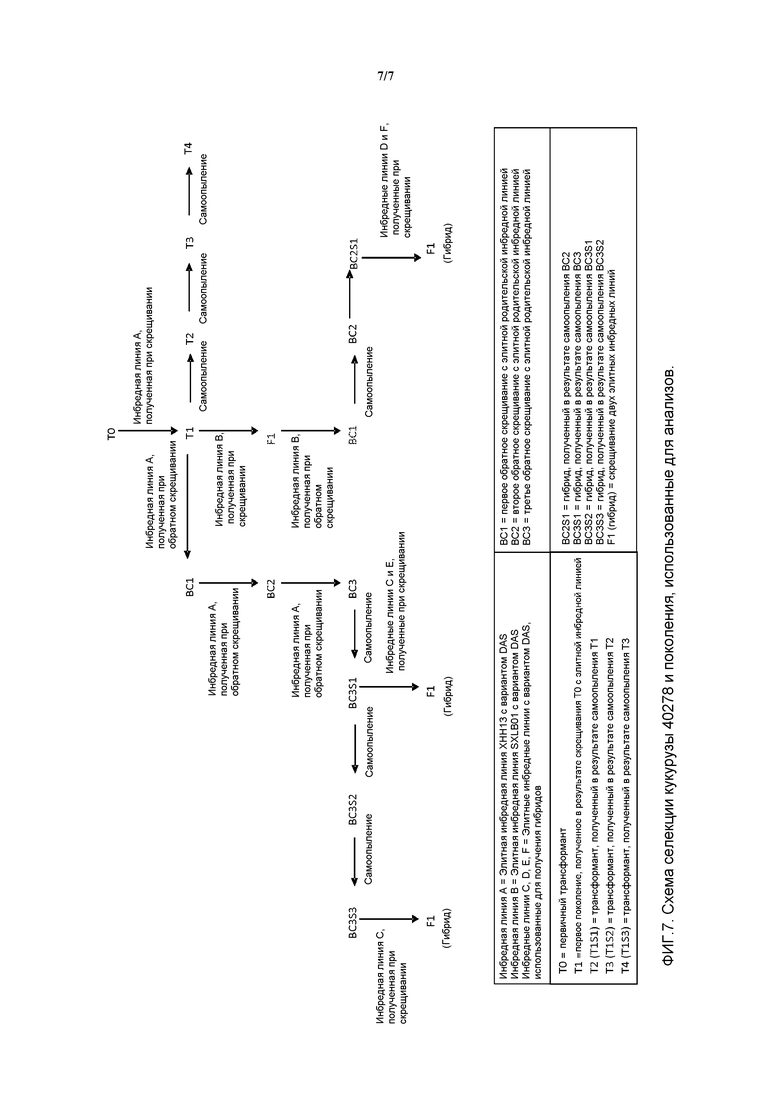
Изобретение относится к области биохимии, в частности к растению трансгенной кукурузы, которое является устойчивым к гербицидам 2,4-D и хизалофопу, его семени и части. Также раскрыт способ обнаружения трансгенного события кукурузы, характеризующегося SEQ ID NO: 29, в образце с использованием полинуклеотида, который характеризуется нуклеотидной последовательностью, выбираемой из группы, состоящей из SEQ ID NOs: 1-28 и 30-33, или с использованием набора, включающего праймеры. Изобретение также относится к полинуклеотиду, который обеспечивает устойчивость к гербицидам 2,4-D и хизалофопу, а также к способу отбора потомства растения кукурузы, содержащего SEQ ID NO: 29, и способу обнаружения потомства растения кукурузы, содержащего признак устойчивости к гербицидам 2,4-D и хизалофопу. Изобретение позволяет эффективно получать растение трансгенной кукурузы, которое является устойчивым к гербицидам 2,4-D и хизалофопу. 12 н. и 3 з.п. ф-лы, 7 ил., 29 табл., 12 пр.
1. Растение трансгенной кукурузы, которое является устойчивым к гербицидам 2,4-D и хизалофопу, где указанное растение содержит геном, включающий последовательность SEQ ID NO: 29.
2. Семя кукурузы для получения растения, являющегося устойчивым к гербицидам 2,4-D и хизалофопу, где указанное семя содержит геном, причем указанный геном включает SEQ ID NO: 29, такую же, которая присутствует в семени, депонированном в Американской коллекции типовых культур (АТСС) под номером доступа РТА-10244, где указанная SEQ ID NO: 29 кодирует арилоксиалканоатдиоксигеназу, которая обеспечивает толерантность к гербицидам 2,4-D и хизалофопу.
3. Трансгенное семя кукурузы для получения растения, являющегося устойчивым к гербицидам 2,4-D и хизалофопу, где указанное семя содержит геном, включающий последовательность SEQ ID NO: 29, где указанная SEQ ID NO: 29 кодирует арилоксиалканоатдиоксигеназу, которая обеспечивает толерантность к гербицидам 2,4-D и хизалофопу.
4. Трансгенное растение кукурузы, которое является устойчивым к гербицидам 2,4-D и хизалофопу, полученное в результате выращивания семени по п. 2, где указанное растение включает нуклеиновую кислоту с последовательностью SEQ ID NO: 29.
5. Часть трансгенного растения по п. 4, которая является устойчивой к гербицидам 2,4-D и хизалофопу, где указанная часть выбрана из группы, состоящей из пыльцы, семяпочки, цветков, побегов, корней и листьев, и указанная часть включает последовательность SEQ ID NO: 29.
6. Выделенный полинуклеотид, который характеризуется нуклеотидной последовательностью, выбираемой из группы, состоящей из SEQ ID NOs: 1-28 и 30-33, где указанный полинуклеотид представляет собой праймер или зонд для обнаружения SEQ ID NO: 29 в трансгенном геноме.
7. Способ обнаружения трансгенного события кукурузы, характеризующегося SEQ ID NO: 29, в образце, содержащем ДНК кукурузы, включающий контактирование указанного образца с меченым зондом или парой праймеров, чтобы обнаружить SEQ ID NO: 29, где указанные зонд и праймеры выбирают из группы, состоящей из SEQ ID NOs: 1-28 и 30-33.
8. Способ по п. 7, который включает контактирование указанного образца с:
a) первым праймером, который связывается с фланкирующей последовательностью, выбираемой из группы, состоящей из остатков 1-1873 SEQ ID NO: 29, остатков 6690-8557 SEQ ID NO: 29 и комплементарных им остатков; и
b) вторым праймером, который связывается с последовательностью вставки, включающей остатки 1874-6689 SEQ ID NO: 29, или комплементарные им остатки;
выполнение полимеразной цепной реакции указанного образца и анализ ампликона, полученного между указанными праймерами.
9. Способ по п. 8, в котором указанные праймеры выбирают из группы, состоящей из SEQ ID NOs: 1-28.
10. Способ по п. 7, в котором указанный полинуклеотид представляет собой меченый зонд, включающий по меньшей мере 30 нуклеотидов и включающий последовательность, выбираемую из группы, состоящей из остатков 1863-1875 SEQ ID NO: 29, остатков 6679-6700 SEQ ID NO: 29 и комплементарных им остатков; где указанный способ дополнительно включает гибридизацию в строгих условиях указанного образца и указанного полинуклеотида и анализ указанного образца на предмет гибридизации указанного полинуклеотида с указанной ДНК.
11. Набор для обнаружения ДНК с SEQ ID NO: 29, который включает первый праймер, выбранный из группы, состоящей из SEQ ID NOs: 5, 7, 12, 14-16, 19-22, 24, 26 и 28, для связывания с остатками 1-1873 или 6690-8557 SEQ ID NO: 29, и второй праймер, выбранный из группы, состоящей из SEQ ID NOs: 1-4, 6, 8-12, 13, 17, 18, 23, 25 и 27, для связывания с остатками 1874-6689 SEQ ID NO: 29, для применения в способе по п. 8.
12. Набор для обнаружения ДНК с SEQ ID NO: 29 для осуществления способа по п. 10, содержащий зонд и гибридизационные реагенты, где указанный зонд содержит последовательность из по меньшей мере 30 нуклеотидов, которая содержит последовательность, выбранную из группы, состоящей из остатков 1863-1875 SEQ ID NO: 29, остатков 6679-6700 SEQ ID NO: 29 и комплементарных им остатков.
13. Полинуклеотид, характеризующийся SEQ ID NO: 29, где указанный полинуклеотид находится в геноме кукурузы и обеспечивает устойчивость к гербицидам 2,4-D и хизалофопу.
14. Способ отбора потомства растения кукурузы, содержащего SEQ ID NO: 29, который включает скрещивание первого растения кукурузы, включающего SEQ ID NO: 29, со вторым растением кукурузы с целью получения третьего растения кукурузы, имеющего геном, и анализ указанного третьего растения кукурузы на наличие SEQ ID NO: 29 в указанном геноме, с использованием способа по п. 7.
15. Способ обнаружения потомства растения кукурузы, содержащего признак устойчивости к гербицидам 2,4-D и хизалофопу, который является таким же, как присутствующий в семени, депонированном в Американской коллекции типовых культур (АТСС) под номером доступа РТА-10244, где указанный способ включает скрещивание первого растения кукурузы, включающего SEQ ID NO: 29, со вторым растением кукурузы с целью получения третьего растения кукурузы, имеющего геном, и анализ указанного третьего растения кукурузы на наличие SEQ ID NO: 29 в указанном геноме, с использованием способа по п. 7.
| WO2008143993 A2, 27.11.2008 | |||
| SUBHASH CHANDER ET AL, Genetic dissection of tocopherol content and composition in maize grain using quantitative trait loci analysis and the candidate gene approach, MOLECULAR BREEDING, 2008, vol | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Замкнутая радиосеть с несколькими контурами и с одной неподвижной точкой опоры | 1918 |
|
SU353A1 |
| ZHANG J ET AL, Mapping quantitative trait loci for oil, starch, and protein concentrations in grain with high-oil maize by SSR markers, EUPHYTICA, 2007, vol | |||
| Деревянное стыковое скрепление | 1920 |
|
SU162A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Способ получения коричневых сернистых красителей | 1922 |
|
SU335A1 |
| WO2007053482 A2, 10.05.2007 | |||
| ТРАНСГЕННЫЕ РАСТЕНИЯ, ЭКСПРЕССИРУЮЩИЕ ГЕН АЦЕТИЛ-СОА-КАРБОКСИЛАЗЫ | 1996 |
|
RU2187555C2 |

