Рассматриваемая заявка заявляет приоритет относительно U.S. предварительной заявки номер 61/378831, поданной 31 августа 2010, полное содержание которой включено сюда по ссылке во всех отношениях.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ НАСТОЯЩЕЕ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области невирусной доставки РНК для иммунизации.
ИЗВЕСТНЫЙ УРОВЕНЬ ТЕХНИКИ
Доставка нуклеиновых кислот для иммунизации животных являлась задачей в течение нескольких лет. Были тестированы различные подходы, включая использование ДНК или РНК, вирусных или невирусных носителей доставки (или даже без носителей доставки, в "оголенной" вакцине), реплицирующихся или нереплицирующихся векторов, или вирусных или невирусных векторов.
Остается необходимость в дальнейшем усовершенствовании вакцин из нуклеиновых кислот и, в частности, разработке усовершенствованных способов доставки вакцин из нуклеиновых кислот.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением иммунизацию нуклеиновыми кислотами обеспечивают путем введения РНК, инкапсулированной в липосомы. РНК кодирует представляющий интерес иммуноген, и диаметр липосом находится в интервале 60-180 нм, и идеально в интервале 80-160 нм. Такой размер сравним, например, с диаметром ~40 нм для лишенного оболочки изометрического белкового капсида альфавируса. Комбинация эффективного инкапсулирования РНК (особенно самореплицирующейся РНК) внутри маленькой липосомы обеспечивает эффективную доставку для выработки сильной иммунной реакции.
Таким образом, в настоящем изобретении предложена липосома, внутри которой инкапсулирована РНК, кодирующая представляющий интерес иммуноген, где диаметр липосомы находится в интервале 60-180 нм. Такие липосомы можно использовать для in vivo доставки РНК в клетки позвоночных, и таким образом они оказываются полезными в качестве компонентов фармацевтических композиций для иммунизации субъектов против различных заболеваний.
В настоящем изобретении также предложен способ получения содержащих РНК липосом, включающий стадию смешивания РНК с одним или более из липидов, в таких условиях, в которых липиды образуют липосомы с диаметром в интервале 60-180 нм и в которых РНК инкапсулирована.
Липосомы
В настоящем изобретении используют липосомы, внутри которых инкапсулированы кодирующие иммуноген РНК. Таким образом, РНК (как и в природном вирусе) оказывается изолированной от любой внешней среды. Обнаружено, что инкапсулирование в липосомы защищает РНК от расщепления РНКазой. Такие липосомы могут включать некоторые внешние РНК (например, на их поверхности), но по меньшей мере, половина РНК (а в идеале все они) инкапсулированы в ядре липосом. Инкапсулирование внутри липосом отличается от, например, комплексов липид/РНК, раскрытых в ссылке 1, где РНК смешивают с предварительно сформированными липосомами. Различные амфифильные липиды могут образовывать двойные слои в водном окружении, чтобы инкапсулировать РНК-содержащее водное ядро, по типу липосомы. Указанные липиды могут иметь анионные, катионные или цвиттерионные концевые группы. Образование липосом из анионных фосфолипидов относится к 1960-м, и липиды, образующие катионные липосомы, исследовали, начиная с 1990-х. Некоторые фосфолипиды являются анионными, тогда как другие являются цвиттерионными а другие являются катионными. Подходящие классы фосфолипидов включают, но ими не ограничиваются, фосфатидилэтаноламины, фосфатидилхолины, фосфатидилсерины и фосфатидилглицерины и некоторые полезные фосфолипиды перечислены в таблице 1. Подходящие катионные липиды включают, но ими не ограничиваются, диолеоилтриметиламмонийпропан (DOTAP), 1,2-дистеарилокси-N,N-диметил-3-аминопропан (DSDMA), 1,2-диолеилокси-N,N-диметил-3-аминопропан (DODMA), 1,2-ди-О-октадеценил-3-триметиламмонийпропан (DOTMA), 1,2-дилинолеилокси-N,N-диметил-3-аминопропан (DLinDMA), 1,2-дилиноленилокси-N,N-диметил-3-аминопропан (DLenDMA). Цвиттерионные липиды включают, но ими не ограничиваются, ацил-цвиттерионные липиды и эфир-цвиттерионные липиды. Примерами подходящих цвиттерионных липидов являются DPPC, DOPC, DSPC, додецилфосфохолин, 1,2-диолеоил-sn-глицеро-3-фосфатидилэтаноламин (DOPE), и 1,2-дифитаноил-sn-глицеро-3-фосфоэтаноламин (DPyPE). Указанные липиды могут быть насыщенными или ненасыщенными. Для получения липосом, предпочтительно использовать по меньшей мере один ненасыщенный липид. Если ненасыщенный липид имеет две концевые группы, обе концевые группы могут быть ненасыщенными, или у него может быть одна насыщенная концевая группа и одна ненасыщенная концевая группа. Липиды могут включать стероидную группу на одном конце, например, как в RV05 (см. также ФИГ.16A и C-K).
Так, в некоторых вариантах, в настоящем изобретении предложены липосомы, содержащие липидный бислой, инкапсулирующий водное ядро, где: (i) липосомы имеют диаметр в интервале 60-180 нм; и (ii) водное ядро включает РНК, которая кодирует иммуноген.
Липосомы настоящего изобретения можно сформировать из одного липида или из смеси липидов. Такая смесь может включать (i) смесь анионных липидов, (ii) смесь катионных липидов, (iii) смесь цвиттерионных липидов, (iv) смесь анионных липидов и катионных липидов, (v) смесь анионных липидов и цвиттерионных липидов, (vi) смесь цвиттерионных липидов и катионных липидов, или (vii) смесь анионных липидов, катионных липидов и цвиттерионных липидов. Аналогично, смесь может включать как насыщенные, так и ненасыщенные липиды. Например, смесь может включать DSPC (цвиттерионный, насыщенный), DlinDMA (катионный, ненасыщенный) и/или DMG (анионный, насыщенный). Если используют смесь липидов, вовсе не все компоненты липидов в смеси должны быть амфифильными, например, один или более из амфифильных липидов может быть смешан с холестерином.
Если липосомы настоящего изобретения образуются из смеси липидов, предпочтительно, чтобы доля указанных катионных липидов составляла от 20 до 80% от полного количества липидов, например, между 30-70%, или между 40-60%. Остальное может составлять, например, холестерин (например, 35-50% холестерина) и/или DMG (необязательно пегилированный) и/или DSPC. Использование таких смесей представлено далее. Приводимые процентные значения являются мольными процентами.
Липосомы могут включать амфифильные липиды, гидрофильная часть которых пегилирована (т.е. модифицирована за счет ковалентного присоединения полиэтиленгликоля). Такая модификация может повысить стабильность и предотвратить неспецифическую адсорбцию указанных липосом. Например, липиды могут быть конъюгированы с ПЭГ с использованием способов, таких как те, что раскрыты в ссылках 2 и 3. ПЭГ снабжает липосомы оболочкой, которая может придавать подходящие фармакокинетические характеристики. Можно использовать ПЭГ различной длины, например, между 0,5 и 8 кДа.
Итак, липосомы можно сформировать из катионных липидов (например, DlinDMA, RV05), цвиттерионных липидов (например, DSPC, DPyPE), холестерина и пегилированных липидов. В представленных примерах используют смесь DSPC, DlinDMA, ПЭГ-DMG и холестерина, также как некоторые другие смеси.
Липосомы обычно подразделяют на три группы: многослойные пузырьки (MLV); маленькие однослойные пузырьки (SUV); и крупные однослойные пузырьки (LUV). MLV имеют множество бислоев в каждом пузырьке, образующих несколько отдельных водных отделений. SUV и LUV имеют один бислой, инкапсулирующий водное ядро; SUV обычно имеют диаметр ≤50 нм, и LUV имеют диаметр >50 нм. Липосомы настоящего изобретения представляют собой идеальные LUV с диаметром в интервале 60-180 нм, и предпочтительно в интервале 80-160 нм. Указанные липосомы предпочтительно являются практически сферическими. Если они не являются сферическими, указанный термин "диаметр " относится к наибольшему диаметру поперечного сечения липосомы.
Липосомы настоящего изобретения могут составлять часть композиции, включающей множество липосом, и липосомы, составляющие указанное множество, могут иметь различные диаметры. Для композиции, включающей популяцию липосом с различными диаметрами: (i) по меньшей мере 80% от числа липосом должны иметь диаметры в интервале 60-180 нм, и предпочтительно, в интервале 80-160 нм, и/или (ii) средний диаметр (определенный по интенсивности, например, Z-среднее) популяции идеально находится в интервале 60-180 нм, и предпочтительно, в интервале 80-160 нм.
Идеально, если распределение липосом по размерам (по интенсивности) имеет только один максимум, т.е. существует одна популяция липосом с распределением вокруг среднего значения (вид), скорее не имеющая двух максимумов. Диаметры внутри популяции липосом в идеале должны иметь показатель полидисперсности <0,2, и иногда <0,1. Ожидают, что комплексы липосома/РНК, раскрытые в ссылке 1, должны иметь диаметры в интервале 600-800 нм и должны отличаться высокой степенью полидисперсности.
Аппаратура для определения среднего диаметра частиц в суспензии липосом и распределения по размерам, коммерчески доступна. Указанные типично используемые методы динамического светорассеяния и/или оптического определение одиночных частиц, представляет собой например, серии приборов Accusizer™ и Nicomp™, доступные от Particle Sizing Systems (Santa Barbara, USA), или приборы Zetasizer™ от Malvern Instruments (UK), или анализатор Particle Size Distribution Analyzer от Horiba (Kyoto, Japan). Метод динамического светорассеяния является предпочтительным методом, с помощью которого определяют размеры липосом. Для популяции липосом, предпочтительным способом определения среднего диаметра липосом в композициях настоящего изобретения является определение величины Z-среднего, т.е. средневзвешенной интенсивности гидродинамического размера общей совокупности липосом, измеренного методом динамического светорассеяния (DLS). Z-среднее определяют с помощью кумулятивного анализа измеренной корреляционной кривой, где принимают размер отдельной частицы (диаметр липосомы) и применяют простое экспоненциальное приближение к автокорреляционной функции. Алгоритм кумулятивного анализа не дает распределения, но, в дополнении к определенному по средневзвешенной интенсивности Z-среднему, определяет коэффициент полидисперсности.
Способы получения подходящих липосомы хорошо известны специалистам, например, смотри ссылки 4-6. Один подходящий способ раскрыт в ссылке 7 и включает смешивание (i) этанольного раствора липидов, (ii) водного раствора нуклеиновой кислоты и (iii) буфера, с последующим перемешиванием, уравновешиванием, разбавлением и очисткой. Предпочтительные липосомы настоящего изобретения можно получить таким способом смешивания. Для получения липосом с нужным диаметром(ами), смешивание можно осуществить, используя способ, в котором два сырьевых потока водного РНК раствора объединяют в единой зоне смешивания с одним потоком этанольного липидного раствора, причем все потоки поступают с одинаковой скоростью например, в микрофлюидизированный канал, как раскрыто далее.
РНК
Липосомы настоящего изобретения включают РНК молекулы, которые (в отличие от siРНК (короткая интерферирующая РНК)) кодируют иммуноген. После in vivo введения частиц, РНК выделяются из частиц и транслируются внутрь клетки для создания иммуногена in situ.
РНК является +-цепочечной, и поэтому она может транслироваться клетками без необходимости в каких-либо промежуточных стадиях репликации, таких как обратная транскрипция. РНК может также связаться с TLR7 рецепторами, экспрессируемыми иммунными клетками, тем самым инициируя эффект адъюванта.
Предпочтительные +-цепочечные РНК являются самореплицирующимися. Самореплицирующиеся молекулы РНК (репликоны) могут, если они введены в клетки позвоночных животных, даже без каких-либо белков, привести к продуцированию множества дочерних РНК за счет собственной транскрипции (через антисмысловые копии, которые они создают сами из себя). Самореплицирующиеся молекулы РНК являются типичными +-цепочечными молекулами, которые могут непосредственно транслироваться после доставки в клетку, и в результате такой трансляции получают РНК-зависимую РНК-полимеразу, которая затем продуцирует как антисмысловые, так и смысловые транскрипты из введенной РНК. Таким образом, введенная РНК приводит к продуцированию множества дочерних РНК. Такие дочерние РНК, также как коллинеарные субгеномные транскрипты, могут быть транслированы ими самими, что приводит к in situ экспрессии кодированного иммуногена, или могут быть транскрибированы, обеспечивая дальнейшие транскрипты с тем же смыслом, что и введенные РНК, которые транслируются для обеспечения in situ экспрессии иммуногена. Итоговым результатом указанной последовательности транскрипций является колоссальное увеличение количества введенных репликоновых РНК, и таким образом кодированный иммуноген становится основным полипептидным продуктом клеток.
Одной из подходящих систем для достижения саморепликации является использование РНК репликона на основе альфавируса. Такие +-цепочечные репликоны транслируются после доставки в клетку, приводя к образованию репликазы (или репликаза-транскриптазы). Такая репликаза транслируется как полипротеин, который авто-расщепляется, обеспечивая репликацию комплекса, который создает копии геномной --цепочки из +-цепочки, поставляемой РНК. Указанные --цепочечные транскрипты сами могут быть транскрибированы с созданием дальнейших копий +-цепочечных исходных РНК и также обеспечивают получение субгеномного транскрипта, который кодирует иммуноген. Таким образом, трансляция субгеномного транскрипта приводит к in situ экспрессии указанного иммуногена инфицированной клеткой. Подходящие альфавирусные репликоны могут использовать репликазу из Sindbis вируса, вируса леса Семлики, вируса восточного энцефалита лошадей и вируса венесуэльского энцефалита лошадей и т.д. Можно использовать последовательности мутантных или дикого типа вирусов, например, ослабленный TC83 мутант VEEV был использован в репликонах [8].
Предпочтительные самореплицирующиеся молекулы РНК кодируют (i) РНК-зависимую РНК полимеразу, которая может транскрибировать РНК из самореплицирующейся РНК молекулы, и (ii) иммуноген. Такая полимераза может быть альфавирусной репликазой, например, включающей один или более из альфавирусных белков nsP1, nsP2, nsP3 и nsP4.
Несмотря на то что природные альфавирусные геномы кодируют структурные белки вирионов в дополнение к неструктурным полипротеинам репликазы, предпочтительно, чтобы самореплицирующиеся молекулы РНК настоящего изобретения не кодировали структурные альфавирусные белки. Таким образом, предпочтительные самореплицирующиеся РНК могут привести к продуцированию своих копий геномных РНК в клетке, но не к продуцированию содержащих РНК вирионов. Неспособность продуцировать указанные вирионы означает, что в отличие от альфавируса дикого типа самореплицирующаяся молекула РНК не может сохраняться в инфекционной форме. Структурные белки альфавируса, которые необходимы для сохранения в вирусах дикого типа, отсутствуют в самореплицирующихся РНК настоящего изобретения и их место занимает ген(ы), кодирующий представляющий интерес иммуноген, так что субгеномный транскрипт кодирует скорее иммуноген, нежели структурные белки альфавирусного вириона.
Так, самореплицирующиеся РНК молекулы, которые можно использовать в настоящем изобретении, могут иметь две открытые считывающие рамки. Первая (5’) открытая считывающая рамка кодирует репликазу; вторая (3’) открытая считывающая рамка кодирует иммуноген. В некоторых вариантах РНК могут иметь (например, в прямом направлении) открытые считывающие рамки, например, для кодирования дополнительных иммуногенов (см. далее) или для кодирования вспомогательных полипептидов.
Самореплицирующаяся РНК молекула может иметь 5’ последовательность, которая совместима с закодированной репликазой.
Самореплицирующиеся молекулы РНК могут быть различной длины, но обычно они состоят из 5000-25000 нуклеотидов в длину, например, 8000-15000 нуклеотидов или 9000-12000 нуклеотидов. Таким образом, РНК оказывается длиннее, чем видно в siРНК доставке.
РНК молекула, которую можно использовать в настоящем изобретении, может иметь 5’-кэп (например, 7-метилгуанозин). Такой кэп может усилить in vivo трансляцию РНК.
5’-нуклеотид молекулы РНК, который можно использовать в настоящем изобретении, может содержать 5’-трифосфатную группу. В кэпированной РНК указанная группа может быть связана с 7-метилгуанозином посредством 5’-до-5’ мостика. 5’ трифосфат может усиливать RIG-I связывание, и таким образом промотировать эффекты адъюванта.
Молекула РНК может содержать 3’ поли-A концевую группу. Она также может включать последовательность, распознающую поли-А полимеразу (например, AAUAAA) вблизи ее 3’ конца.
Молекула РНК, которую можно использовать в настоящем изобретении, обычно является одноцепочечной. Одноцепочечные РНК обычно могут инициировать эффект адъюванта, путем связывания с TLR7, TLR8, РНК-геликазами и/или PKR. РНК, введенная в двухцепочечной форме (дцДНК), может связываться с TLR3, и указанный рецептор также может быть стимулирован за счет дцДНК, которая образуется или во время репликации одноцепочечной РНК или внутри вторичной структуры одноцепочечной РНК.
Молекулы РНК, которые можно использовать в настоящем изобретении, обычно удобно получать путем in vitro транскрипции (IVT). IVT может использовать (кДНК) матрицу, созданную и размноженную в плазмидной форме в бактериях, или созданную синтетически (например в результате использования способов генного синтеза и/или полимеразной цепной реакции). Например, ДНК-зависимая РНК полимераза (такая как бактериофаг T7, T3 или SP6 РНК полимеразы) может быть использована для транскрибирования РНК из ДНК матрицы. Подходящие реакции кэппинга и поли-А присоединения можно использовать при необходимости (хотя поли-А репликона обычно кодируется внутри ДНК матрицы). Такие РНК полимеразы могут выдвигать жесткие требования в отношении транскрибированного 5’ нуклеотида (нуклеотидов) и в некоторых вариантах указанные требования должны удовлетворять требованиям кодированной репликазы, чтобы обеспечить способность IVT-транскрибированной РНК функционировать эффективно в качестве субстрата для ее самокодирующейся репликазы.
Как обсуждалось в ссылке 9, самореплицирующиеся РНК могут включать (в дополнении к любой 5’-кэп структуре) один или более из нуклеотидов, содержащих модифицированное нуклеооснование. Так, РНК может включать m5C (5-метилцитидин), m5U (5-метилуридин), m6A (N6-метиладенозин), s2U (2-тиоуридин), Um (2’-O-метилуридин), m1A (1-метиладенозин); m2A (2-метиладенозин); Am (2’-О-метиладенозин); ms2m6A (2-метилтио-N6-метиладенозин); i6A (N6-изопентинаденозин); ms2i6A (2-метилтио-N6-изопентинаденозин); io6A (N6-(цис-гидроксиизопентин)аденозин); ms2io6A (2-метилтио-N6-(цис-гидроксиизопентин)аденозин); g6A (N6-глицинилкарбамоиладенозин); t6A (N6-треонилкарбамоиладенозин); ms2t6A (2-метилтио-N6 треонилкарбамоиладенозин); m6t6A (N6-метил-N6-треонилкарбамоиладенозин); hn6A(N6.-гидроксинорвалилкарбамоиладенозин); ms2hn6A (2-метилтио-N6-гидроксинорвалилкарбамоиладенозин); Ar(p) (2’-О-рибозиладенозин (фосфат)); I (инозин); ml1 (1-метилинозин); m’Im (1,2’-О-диметилинозин); m3C (3-метилцитидин); Cm (2T-О-метилцитидин); s2C (2-тиоцитидин); ac4C (N4-ацетилцитидин); f5C (5-формилцитидин); m5Cm (5,2-О-диметилцитидин); ac4Cm (N4ацетил2TOметилцитидин); k2C (лизидин); m1G (1-метилгуанозин); m2G (N2-метилгуанозин); m7G (7-метилгуанозин); Gm (2’-О-метилгуанозин); m22G (N2,N2-диметилгуанозин); m2Gm (N2,2’-О-диметилгуанозин); m22Gm (N2,N2,2’-О-триметилгуанозин); Gr(p) (2’-О-рибозилгуанозин (фосфат)); yW (вибутозин); o2yW (пероксивибутозин); OHyW (гидроксивибутозин); OHyW* (недомодифицированный гидроксивибутозин); imG (виозин); mimG (метилгуанозин); Q (куэозин); oQ (эпоксикуэозин); galQ (галтаклозил-куэозин); manQ (маннозил-куэозин); preQo (7-циано-7-деазагуанозин); preQi (7-аминометил-7-деазагуанозин); G* (архаэозин); D (дигидроуридин); m5Um (5,2’-О-диметилуридин); s4U (4-тиоуридин); m5s2U (5-метил-2-тиоуридин); s2Um (2-тио-2’-О-метилуридин); acp3U (3-(3-амино-3-карбоксипропил)уридин); ho5U (5-гидроксиуридин); mo5U (5-метоксиуридин); cmo5U (уридин 5-оксиуксусная кислота); mcmo5U (уридин 5-оксиуксусной кислоты метиловый эфир); chm5U (5-(карбоксигидроксиметил)уридин)); mchm5U (5-(карбоксигидроксиметил)уридина метиловый эфир); mcm5U (5-метоксикарбонилметилуридин); mcm5Um (S-метоксикарбонилметил-2-O-метилуридин); mcm5s2U (5-метоксикарбонилметил-2-тиоуридин); nm5s2U (5-аминометил-2-тиоуридин); mnm5U (5-метиламинометилуридин); mnm5s2U (5-метиламинометил-2- тиоуридин); mnm5se2U (5-метиламинометил-2-селеноуридин); ncm5U (5-карбамоилметилуридин); ncm5Um (5-карбамоилметил-2’-О-метилуридин); cmnm5U (5-карбоксиметиламинометилуридин); cnmm5Um (5-карбоксиметиламинометил-2-L-O-метилуридин); cmnm5s2U (5-карбоксиметиламинометил-2-тиоуридин); m62A (N6,N6- диметиладенозин); Tm (2’-О-метилинозин); m4C (N4-метилцитидин); m4Cm (N4,2-О-диметилцитидин); hm5C (5-гидроксиметилцитидин); m3U (3-метилуридин); cm5U (5-карбоксиметилуридин); m6Am (N6,T-О-диметиладенозин); rn62Am (N6,N6,-О-2-триметиладенозин); m2’7G (N2,7-диметилгуанозин); m2’2’7G (N2,N2,7-триметилгуанозин); m3Um (3,2T-О-диметилуридин); m5D (5-метилдигидроуридин) f5Cm (5-формил-2’-О-метилцитидин); m1Gm (1,2’-О-диметилгуанозин); m’Am (1,2-O-диметиладенозин)иринометилуридин); tm5s2U (S-тауринометил-2-тиоуридин)); imG-14 (4-деметилгуанозин); imG2 (изогуанозин); или ac6A (N6-ацетиладенозин), гипоксантин, инозин, 8-оксо-аденин, 7-замещенные их производные, дигидроурацил, псевдоурацил, 2-тиоурацил, 4-тиоурацил, 5-аминоурацил, 5-(C1-C6)-алкилурацил, 5-метилурацил, 5-(C2-C6)-алкенилурацил, 5-(C2-C6)-алкинилурацил, 5-(гидроксиметил)урацил, 5-хлорурацил, 5-фторурацил, 5-бромурацил, 5-гидроксицитозин, 5-(C1-C6)-алкилцитозин, 5-метилцитозин, 5-(C2-C6)-алкенилцитозин, 5-(C2-C6)-алкинилцитозин, 5-хлорцитозин, 5-фторцитозин, 5-бромцитозин, N2-диметилгуанин, 7-деазагуанин, 8-азагуанин, 7-деаза-7-замещенный гуанин, 7-деаза-7-(C2-C6)алкинилгуанин, 7-деаза-8-замещенный гуанин, 8-гидроксигуанин, 6-тиогуанин, 8-оксогуанин, 2-аминопурин, 2-амино-6-хлорпурин, 2,4-диаминопурин, 2,6-диаминопурин, 8-азапурин, замещенный 7-деазапурин, 7-деаза-7-замещенный пурин, 7-деаза-8-замещенный пурин, или абазический нуклеотид. Например, самореплицирующиеся РНК могут включать одно или более из модифицированных пиримидиновых нуклеооснований, таких как остатки псевдоуридина и/или 5-метилцитозина. В некоторых вариантах, однако, РНК включают немодифицированные нуклеооснования, и могут включать немодифицированные нуклеотиды т.е. все нуклеотиды в РНК представляют собой стандартные A, C, G и U рибонуклеотиды (за исключением любых 5’-кэп структур, которые могут включать 7’-метилгуанозин). В других вариантах, РНК могут включать 5’-кэп, включающий 7’-метилгуанозин, и первые 1, 2 или 3 5’-рибонуклеотида могут быть метилированы по 2’-положению рибозы.
РНК, которые используют в настоящем изобретении, в идеале включают только фосфодиэфирные связи между нуклеозидами, но в некоторых вариантах они могут содержать фосфорамидатные, фосфоротиоатные и/или метилфосфонатные связи.
Идеально, если липосомы настоящего изобретения включают меньше, чем 10 различных видов РНК, например, 5, 4, 3 или 2 различных вида; наиболее предпочтительно, если липосомы включают один вид РНК, т.е. все молекулы РНК в липосомах содержат одинаковые последовательности, причем одинаковой длины.
Количество РНК в липосоме может варьироваться. Число индивидуальных самореплицирующихся молекул РНК в липосоме обычно составляет ≤50, например, <20, <10, <5 или 1-4 на липосому.
Иммуноген
Молекулы РНК, используемые в настоящем изобретении, кодируют полипептидный иммуноген. После введения липосом РНК транслируется in vivo, и указанный иммуноген может вызвать иммунную реакцию у реципиента. Указанный иммуноген может вызвать иммунную реакцию против бактерии, вируса, грибков или паразита (или, в некоторых вариантах, против аллергена; и в других вариантах, против опухолевого антигена). Иммунная реакция может включать реакцию антител (обычно включающую IgG) и/или опосредованную клеткой иммунную реакцию. Полипептидный иммуноген обычно вызывает иммунную реакцию, которая распознает соответствующий бактериальный, вирусный, грибковый или паразитарный (или аллергена или опухоли) полипептид, но в некоторых вариантах такой полипептид может действовать как мимеотоп, вызывая иммунную реакцию, которая распознает бактериальный, вирусный, грибковый или паразитарный сахарид. Указанный иммуноген обычно представляет собой поверхностный полипептид, например, адгезин, гемагглютинин, оболочковый гликопротеин, спайковый гликопротеин и т.д.
Молекула РНК может кодировать один полипептидный иммуноген или множественные полипептиды. Множественные иммуногены могут присутствовать в виде одного полипептидного иммуногена (фьюжн полипептид) или в виде отельных полипептидов. Если иммуногены экспрессируются в виде отдельных полипептидов репликона, тогда один или более из них может быть снабжен в обратном направлении IRES или дополнительным вирусным промоторным элементом. Альтернативно, множественные иммуногены могут экспрессироваться из полипротеина, который кодирует отдельные иммуногены, слитые с короткой автокаталитической протеазой (например, 2A белком вируса ящура), или в виде интеинов.
В отличие от ссылок 1 и 10, РНК кодирует иммуноген. Во избежание сомнений, настоящее изобретение не включает РНК, которые кодируют люциферазу светлячков или кодируют рекомбинантный белок E.coli β-галактозидазы или кодируют зеленый флуоресцентный белок (GFP). Такие полипептиды могут быть полезны в качестве маркеров, или даже в контексте генной терапии, но настоящее изобретение касается доставки РНК стимуляции системы иммунологической реакции. Оптимальный диаметр липосом для генной терапии может отличаться от диаметра липосом для целей иммунизации, так как мишеневые клетки и ткани различны для указанных двух подходов. Так иммуноген также не является самостоятельным белком, который вводят для дополнения или замены дефективного белка хозяина (что происходит в генной терапии). Также указанная РНК не является полной РНК мышиного тимуса.
В некоторых вариантах иммуноген вызывает иммунную реакцию против одной из указанных бактерий:
Neisseria meningitidis: подходящие иммуногены включают, но ими не ограничиваются, мембранные белки, такие как адгезины, автотранспортеры, токсины, накапливающие железо белки и белок, связывающий фактор H. Комбинация их трех подходящих полипептидов раскрыта в ссылке 11.
Streptococcus pneumoniae: подходящие полипептидные иммуногены раскрыты в ссылке 12. Они включают, но ими не ограничиваются, RrgB плюс субъединицу, предшественник бета-N-ацетилгексозаминидазы (spr0057), spr0096, белок общего стресса GSP-781 (spr2021, SP2216), серин/треонинкиназу StkP (SP1732) и пневмококковый поверхностный адгезин PsaA.
Streptococcus pyogenes: подходящие иммуногены включают, но ими не ограничиваются, полипептиды, раскрытые в ссылках 13 и 14.
Moraxella catarrhalis
Bordetella pertussis: Подходящие коклюшевые иммуногены включают, но ими не ограничиваются, токсин коклюша или коклюшевый токсоид (PT), филаментный гемагглютинин (FHA), пертактин, и агглютиногены 2 и 3.
Staphylococcus aureus: Подходящие иммуногены включают, но ими не ограничиваются, полипептиды, раскрытые в ссылке 15, такие как гемолизин, esxA, esxB, белок, связывающий феррихром (sta006) и/или sta011 липопротеин.
Clostridium tetani: типичный иммуноген представляет собой столбнячный токсоид.
Corynebacterium diphtheriae: типичный иммуноген представляет собой дифтерийный токсоид.
Haemophilus influenzae: Подходящие иммуногены включают, но ими не ограничиваются, полипептиды, раскрытые в ссылках 16 и 17.
Pseudomonas aeruginosa
Streptococcus agalactiae: подходящие иммуногены включают, но ими не ограничиваются, полипептиды, раскрытые в ссылке 13.
Chlamydia trachomatis: Подходящие иммуногены включают, но ими не ограничиваются, PepA, LcrE, ArtJ, DnaK, CT398, OmpH-подобный, L7/L12, OmcA, AtoS, CT547, Eno, HtrA и MurG (например, как раскрыто в ссылке 18. LcrE [19] и HtrA [20] представляют собой два предпочтительных иммуногена).
Chlamydia pneumoniae: Подходящие иммуногены включают, но ими не ограничиваются, полипептиды, раскрытые в ссылке 21.
Helicobacter pylori: Подходящие иммуногены включают, но ими не ограничиваются, CagA, VacA, NAP и/или уреазу [22].
Escherichia coli: Подходящие иммуногены включают, но ими не ограничиваются, иммуногены, полученные из энтеротоксигенной E. coli (ETEC), энтероаггреативной E. coli (EAggEC), диффузно приклеивающейся E. coli (DAEC), энтеропатогенной E. coli (EPEC), экстраинтестинальной патогенной E. coli (ExPEC) и/или энтерогеморрагической E. coli (EHEC). ExPEC штаммы включают уропатогенную E.coli (UPEC) и связанную с менингитом/сепсисом E.coli (MNEC). Подходящие UPEC полипептидные иммуногены раскрыты в ссылках 23 и 24. Подходящие MNEC иммуногены раскрыты в ссылке 25. Подходящим иммуногеном для некоторых типов E.coli является AcfD [26].
Bacillus anthracis
Yersinia pestis: Подходящие иммуногены включают, но ими не ограничиваются, те, что раскрыты в ссылках 27 и 28.
Staphylococcus epidermis
Clostridium perfringens или Clostridium botulinums
Legionella pneumophila
Coxiella burnetii
Brucella, такие как B. abortus, B.canis, B.melitensis, B.neotomae, B.ovis, B.suis, B.pinnipediae.
Francisella, такие как F.novicida, F.philomiragia, F.tularensis.
Neisseria gonorrhoeae
Treponema pallidum
Haemophilus ducreyi
Enterococcus faecalis или Enterococcus faecium
Staphylococcus saprophyticus
Yersinia enterocolitica
Mycobacterium tuberculosis
Rickettsia
Listeria monocytogenes
Vibrio cholerae
Salmonella typhi
Borrelia burgdorferi
Porphyromonas gingivalis
Klebsiella
В некоторых вариантах иммуноген вызывает иммунную реакцию против одного из следующих вирусов:
Orthomyxovirus: Подходящие иммуногены могут быть из вирусов гриппа A, B или C, таких как гемагглютинин, нейраминидаза или матричные M2 белки. Если иммуноген представляет собой гемагглютинин вируса гриппа A, он может быть из любого субтипа, например, H1, H2, H3, H4, H5, H6, H7, H8, H9, H10, H11, H12, H13, H14, H15 или H16.
Paramyxoviridae viruses: Вирусные иммуногены включают, но ими не ограничиваются, иммуногены, полученные из пневмовирусов (например, респираторного синцитиального вируса, RSV), Rubulaviruses (например, вируса свинки), Paramyxoviruses (например, вируса парагриппа), Metapneumoviruses и Morbilliviruses (например, вируса кори).
Poxviridaevirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Orthopoxvirus, такие как Variola vera, включая, но ими не ограничиваясь, Variola major и Variola minor.
Picornavirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые полученные из Picornaviruses, такие как энтеровирусы, риновирусы, гепаРНКвирусы, кардиовирусы и Aphthoviruses. В одном варианте, энтеровирус представляет собой полиовирус, например, типа 1, типа 2 и/или типа 3 полиовирус. В другом варианте, энтеровирус представляет собой EV71 энтеровирус. В другом варианте, энтеровирус представляет собой вирус Коксэки A или B.
Bunyavirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Orthobunyavirus, такие как калифорнийский энцефалитный вирус, Phlebovirus, такие как вирус лихорадки Рифт-Вэлли, или Nairovirus, такие как вирус конго-крымской геморрагической лихорадки.
Heparnavirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Heparnavirus, такие как вирус гепатита A (HAV).
Filovirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из филовируса, такие как вирусы Эбола (включая эболавирус Заира, берега слоновой кости, Рестона или Судана) или Марбург-вирус.
Togavirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Togavirus, такие как Rubivirus, Alphavirus или an Arterivirus. Перечисление включает рубеллавирус.
Flavivirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Flavivirus, такие как вирус клещевого энцефалита (TBE), Денге (типы 1, 2, 3 или 4) вирус, вирус желтой лихорадки, вирус японского энцефалита, вирус болезни Кьясанурского леса, вирус лихорадки Западного Нила, вирус лихорадки Сент-Луиса, вирус российского весенне-летнего энцефалита, вирус энцефалита Повассан.
Pestivirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Pestivirus, такие как вирус диареи крупного рогатого скота (BVDV), вирус классической чумы свиней (CSFV) или вирус пограничной болезни овец (BDV).
Hepadnavirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Hepadnavirus, такие как вирус гепатита В. Композиции могут включать поверхностный антиген вируса гепатита В (HBsAg).
Другие гепатитные вирусы: Композиции могут включать иммуноген из вируса гепатита С, вируса гепатита дельта, вируса гепатита E или вируса гепатита G.
Rhabdovirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Rhabdovirus, такие как лиссавирус (например, вирус бешенства) и везикуловирусы (VSV).
Caliciviridae: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из Caliciviridae, такие как Норволквирус (Норовирус), и Норволк-подобные вирусы, такие как вирус Гавайских островов и вирус снежных гор.
Coronavirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из SARS коронавируса, инфекционного вируса птичьего гриппа (IBV), вируса мышиного гепатита (MHV) и передаваемого вируса гастроэнтерита свиней (TGEV). Иммуноген коронавируса может быть спайк-полипептидом.
Retrovirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из онковирусов, лентивирусов (например, HIV-1 или HIV-2) или спумавирусов.
Reovirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из ортореовирусов, ротавирусов, орбивирусов или колтивирусов.
Parvovirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из парвавируса B19.
Herpesvirus: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из герпесвируса человека, такие как (только в качестве примера), простой герпесвирус (HSV) (например, HSV типов 1 и 2), вирус ветряной оспы (VZV), вирус Эпштейна-Барра (EBV), цитомегаловирус (ЦМВ), человеческий герпесвирус 6 (HHV6), человеческий герпесвирус 7 (HHV7), и человеческий герпесвирус 8 (HHV8).
Papovaviruses: Вирусные иммуногены включают, но ими не ограничиваются, те, которые получены из папилломавирусов и полиомавирусов. Указанные (человеческие) папилломавирусы могут принадлежать к серотипам 1, 2, 4, 5, 6, 8, 11, 13, 16, 18, 31, 33, 35, 39, 41, 42, 47, 51, 57, 58, 63 или 65, например, из одного или более из серотипов 6, 11, 16 и/или 18.
Adenovirus: Вирусные иммуногены включают те, которые получены из аденовируса серотипа 36 (Ad-36).
В некоторых вариантах, иммуноген вызывает иммунную реакцию против вирусов, которые инфицируют рыбу, таких как: инфекционный вирус анемии лосося (ISAV), вирус панкреатита лосося (SPDV), инфекционный вирус некроза поджелудочной железы (IPNV), вирус болезни американского проточного сома (CCV), вирус лимфотического заболевания рыб (FLDV), инфекционный вирус гематопоэтического некроза (IHNV), вирус герпеса карпов кои, пикорна-подобный вирус лосося (также известный как пикорна-подобный вирус атлантического лосося), вирус садкового лосося (LSV), ротавирус атлантического лосося (ASR), вирус язвенной болезни форели (TSD), вирус опухоли кижуча (CSTV) или вирус вирусной геморрагической септицемии (VHSV).
Грибковые иммуногены можно получить из Dermatophytres, включая: Epidermophyton floccusum, Microsporum audouini, Microsporum canis, Microsporum distortum, Microsporum equinum, Microsporum gypsum, Microsporum nanum, Trichophyton concentricum, Trichophyton equinum, Trichophyton gallinae, Trichophyton gypseum, Trichophyton megnini, Trichophyton mentagrophytes, Trichophyton quinckeanum, Trichophyton rubrum, Trichophyton schoenleini, Trichophyton tonsurans, Trichophyton verrucosum, T. verrucosum var. album, var. discoides, var. ochraceum, Trichophyton violaceum и/или Trichophyton faviforme; или из Aspergillus fumigatus, Aspergillus flavus, Aspergillus niger, Aspergillus nidulans, Aspergillus terreus, Aspergillus sydowi, Aspergillus flavatus, Aspergillus glaucus, Blastoschizomyces capitatus, Candida albicans, Candida enolase, Candida tropicalis, Candida glabrata, Candida krusei, Candida parapsilosis, Candida stellatoidea, Candida kusei, Candida parakwsei, Candida lusitaniae, Candida pseudo-tropicalis, Candida guilliermondi, Cladosporium carrionii, Coccidioides immitis, Blastomyces dermatidis, Cryptococcus neoformans, Geotrichum clavatum, Histoplasma capsulatum, Klebsiella pneumoniae, Microsporidia, Encephalitozoon spp., Septata intestinalis и Enterocytozoon bieneusi; реже встречаются Brachiola spp, Microsporidium spp., Nosema spp., Pleistophora spp., Trachipleistophora spp., Vittaforma spp Paracoccidioides brasiliensis, Pneumocystis carinii, Pythiumn insidiosum, Pityrosporum ovale, Sacharomyces cerevisae, Saccharomyces boulardii, Saccharomyces pombe, Scedosporium apiosperum, Sporothrix schenckii, Trichosporon beigelii, Toxoplasma gondii, Penicillium marneffei, Malassezia spp., Fonsecaea spp., Wangiella spp., Sporothrix spp., Basidiobolus spp., Conidiobolus spp., Rhizopus spp, Mucor spp, Absidia spp, Mortierella spp, Cunninghamella spp, Saksenaea spp., Alternaria spp, Curvularia spp, Helminthosporium spp, Fusarium spp, Aspergillus spp, Penicillium spp, Monolinia spp, Rhizoctonia spp, Paecilomyces spp, Pithomyces spp и Cladosporium spp.
В некоторых вариантах иммуноген вызывает иммунную реакцию против паразитов рода Plasmodium, таких как P. falciparum, P.vivax, P.malariae или P.ovale. Таким образом, настоящее изобретение можно использовать для иммунизации против малярии. В некоторых вариантах указанный иммуноген вызывает иммунную реакцию против паразитов семейства Caligidae, особенно принадлежащих к родам Lepeophtheirus и Caligus, например, морских вшей, таких как Lepeophtheirus salmonis или Caligus rogercresseyi.
В некоторых вариантах указанный иммуноген вызывает иммунную реакцию против: пыльцевых аллергенов (аллергенов пыльцы деревьев, лекарственных растений, сорняков и трав); аллергенов насекомых или клещей (аллергенов летучих веществ, слюны и яда, например, аллергенов клещей, аллергенов тараканов и комаров, аллергенов яда перепончатокрылых насекомых); аллергенов шерсти животных и аллергенов перхоти животных (например, собак, кошек, лошадей, крыс, мышей и т.д.); и пищевых аллергенов (например, глиадина). Важными пыльцевыми аллергенами деревьев, трав и лекарственных растений являются аллергены таксономических отрядов Fagales, Oleales, Pinales и Platanaceae, включая, но ими не ограничиваясь, березу (Betula), ольху (Alnus), орех (Corylus), граб (Carpinus) и оливу (Olea), кедр (Cryptomeria и Juniperus), платан (Platanus), отряд Poales, включая травы родов Lolium, Phleum, Poa, Cynodon, Dactylis, Holcus, Phalaris, Secale, и Sorghum, отряды Asterales и Urticales, включая травы родов Ambrosia, Artemisia и Parietaria. Другими важными аллергенами летучих веществ являются аллергены клещей домашней пыли родов Dermatophagoides и Euroglyphus, амбарных клещей, например, Lepidoglyphys, Glycyphagus и Tyrophagus, аллергены тараканов, москитов и мух, например, Blatella, Periplaneta, Chironomus и Ctenocepphalides, и аллергены млекопитающих, таких как кошки, собаки и лошади, аллергены ядов, включая яды жалящих и кусающих насекомых, таких или такие как насекомые отряда жесткокрылых, включая пчел (Apidae), ос (Vespidea) и комаров (Formicoidae).
В некоторых вариантах иммуноген представляет собой опухолевый антиген, выбранный из: (a) антигенов рака яичек, таких как NY-ESO-1, SSX2, SCP1, также как RAGE, BAGE, GAGE и MAGE семейств полипептидов, например, GAGE-1, GAGE-2, MAGE-1, MAGE-2, MAGE-3, MAGE-4, MAGE-5, MAGE-6 и MAGE-12 (которые можно использовать, например, при лечении меланомы, опухолей легких, головы и шеи, NSCLC, груди, желудочно-кишечного тракта и мочевого пузыря; (b) мутантных антигенов, например, p53 (связанных с различными твердыми опухолями, например, раком прямой кишки, легких, головы и шеи), p21/Ras (связанных, например, с меланомой, раком поджелудочной железы и раком прямой кишки), CDK4 (связанных, например, с меланомой), MUM1 (связанных, например, с меланомой), капсазы-8 (связанной, например, с раком головы и шеи), CIA 0205 (связанных, например, с раком мочевого пузыря), HLA-A2-R1701, бета-катехина (связанным, например, с меланомой), TCR (связанных, например, с T-клеточной лимфомой не-Ходжкина), BCR-abl (связанных, например, с хронической миелогенной лейкемией), триосефосфатизомеразы, KIA 0205, CDC-27 и LDLR-FUT; (c) сверхэкспрессированных антигенов, например, галектина 4 (связанного, например, с раком прямой кишки), галектина 9 (связанного, например, с болезнью Ходжкина), протеиназы 3 (связанной, например, с хронической миелогенной лейкемией), WT 1 (связанных, например, с различными лейкемиями), карбоангидразы (связанной, например, с раком почек), альдолазы A (связанной, например, с раком легких), PRAME (связанным, например, с меланомой), HER-2/neu (связанным, например, с раком груди, прямой кишки, легких и яичников), маммаглобина, альфа-фетопротеина (связанного, например, с гепатомой), KSA (связанной например, с раком прямой кишки), гастрина (связанного, например, с раком поджелудочной железы и желудка), каталитического белка теломеразы, MUC-1 (связанного, например, с раком молочной железы и яичников), G-250 (связанных, например, с клеточной карциномой почек), p53 (связанных, например, с раком груди и прямой кишки) и карциноэмбрионного антигена (связанного, например, с раком груди, раком легких, раковыми заболеваниями желудочно-кишечного тракта, такими как рак прямой кишки); (d) перекрестно-реагирующих антигенов, например, меланома-меланоцитных дифференцирующих антигенов, такие как MART-1/Melan A, gp100, MC1R, рецепторов меланоцит-стимулирующего гормона, тирозиназы, тирозиназо-родственного белка-1/TRP1 и тирозиназо-родственного белка-2/TRP2 (связанных, например, с меланомой); (e) связанных с простатой антигенов, таких как PAP, PSA, PSMA, PSH-P1, PSM-P1, PSM-P2, связанные с раком простаты, например; (f) идиотипов иммуноглобулина (связанных, например, с миеломой и B клеточными лимфомами). В некоторых вариантах опухолевые иммуногены включают, но ими не ограничиваются, p15, Hom/Mel-40, H-Ras, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, антигены вируса Эпштейн-Барра, EBNA, антигены человеческого папилломавируса (HPV), включая E6 и E7, антигены вирусов гепатита В и С, антигены человеческого Т-клеточного лимфотропического вируса, TSP-180, p185erbB2, p180erbB-3, c-met, mn-23H1, TAG-72-4, CA 19-9, CA 72-4, CAM 17.1, NuMa, K-ras, p16, TAGE, PSCA, CT7, 43-9F, 5T4, 791 Tgp72, бета-HCG, BCA225, BTAA, CA 125, CA 15-3 (CA 27.29\BCAA), CA 195, CA 242, CA-50, CAM43, CD68\KP1, CO-029, FGF-5, Ga733 (EpCAM), HTgp-175, M344, MA-50, MG7-Ag, MOV18, NB/70K, NY-CO-1, RCAS1, SDCCAG16, TA-90 (Mac-2-связывающий белок/циклофилин C-связанный белок), TAAL6, TAG72, TLP, TPS и т.п.
Фармацевтические композиции
Липосомы настоящего изобретения можно использовать в качестве компонентов фармацевтических композиций для иммунизации субъектов против различных заболеваний. Такие композиции обычно включают в дополнение к липосомам фармацевтически приемлемый носитель. Исчерпывающее обсуждение фармацевтически приемлемых носителей можно найти в ссылке 29.
Фармацевтические композиции настоящего изобретения могут включать одну или больше из маленьких молекул иммуностимуляторов. Например, такие композиции могут включать агонисты TLR2 (например, Pam3CSK4), агонисты TLR4 (например, аминоалкилглюкозаминидфосфат, такие как E6020), агонисты TLR7 (например, имиквимод), агонисты TLR8 (например, резиквимод) и/или агонисты TLR9 (например, IC31). В идеале любой такой агонист имеет молекулярный вес <2000 Да. В некоторых вариантах такой агонист(ы) также инкапсулированы с РНК внутри липосом, но в других вариантах они не являются инкапсулированными.
Фармацевтические композиции настоящего изобретения могут включать липосомы в чистой воде (например, в воде для инъекций) или в буфере, например, в фосфатном буфере, в Трис-буфере, в боратном буфере, в сукцинатном буфере, в гистидиновом буфере или в цитратном буфере. Содержание солей в буфере обычно находится в интервале концентраций 5-20 мМ.
Фармацевтические композиции настоящего изобретения могут иметь pH между 5,0 и 9,5, например, между 6,0 и 8,0.
Композиции настоящего изобретения могут включать соли натрия (например, хлорид натрия) для обеспечения тоничности. Типичной является концентрация 10±2 мг/мл NaCl и обычно, например, около 9 мг/мл.
Композиции настоящего изобретения могут включать ионы металлов в качестве хелатирующих агентов. Они могут пролонгировать стабильность РНК, удаляя ионы, которые могут ускорять фосфодиэфирный гидролиз. Так, композиции могут включать один или более из EDTA, EGTA, BAPTA, триамина пентауксусной кислоты и т.д. Такие хелатирующие агенты обычно присутствуют в концентрациях между 10-500 мкМ, например, 0,1 мМ. Цитратная соль, такая как цитрат натрия, также может действовать как хелатирующий агент, при этом с выгодой обеспечивая буферирующую активность.
Фармацевтические композиции настоящего изобретения могут иметь значение осмоляльности между 200 мОсм/кг и 400 мОсм/кг, например, между 240-360 мОсм/кг, или между 290-310 мОсм/кг.
Фармацевтические композиции настоящего изобретения могут включать один или более из консервантов, таких как тиомерсал или 2-феноксиэтанол. Предпочтительны композиции, которые не содержат ртуть, и можно приготовить вакцины, которые не содержат консервантов.
Фармацевтические композиции настоящего изобретения предпочтительно являются стерильными.
Фармацевтические композиции настоящего изобретения предпочтительно являются непирогенными, например, содержащими <1 EU (единиц эндотоксина, стандартное измерение) на дозу, и предпочтительно <0,1 EU на дозу.
Фармацевтические композиции настоящего изобретения предпочтительно не содержат глютена.
Фармацевтические композиции настоящего изобретения можно приготовить в единичной дозовой форме. В некоторых вариантах единичная доза может иметь объем между 0,1-1,0 мл, например, около 0,5 мл.
Композиции настоящего изобретения можно приготовить в виде, пригодном для инъекций, или в виде растворов или в виде суспензий. Такие композиции можно приготовить для пульмонарного введения, например, с помощью ингаляции, используя мелкокапельный спрей. Указанную композицию можно приготовить для назального, ушного или окулярного введения, например, в виде спрея или капель. Типичными являются препараты для инъекций для внутримышечного введения.
Композиции включают иммунологически эффективное количество липосом, также как любые другие необходимые компоненты. Под выражением "иммунологически эффективное количество" подразумевают, что введение указанного количества индивидууму, или в виде одной дозы, или как часть серии, является эффективным для лечения или профилактики. Указанное количество меняется в зависимости от состояния здоровья и физического состояния подлежащего лечению индивидуума, возраста, таксономической группы подлежащего лечению индивидуума (например, нечеловеческого примата, примата, и т.д.), способности иммунной системы индивидуума синтезировать антитела, от степени необходимой защиты, от лекарственной формы вакцины, от оценки лечащим врачом медицинской ситуации и других важных факторов. Ожидают, что такое количество попадает в относительно широкий интервал значений, который можно определить, используя рутинные испытания. Содержание липосом и РНК в композициях настоящего изобретения обычно выражают в терминах количества РНК на дозу. Предпочтительная доза содержит ≤100 мкг РНК (например, от 10 до 100 мкг, например, около 10 мкг, 25 мкг, 50 мкг, 75 мкг или 100 мкг), величина дозы может быть выражена в гораздо более низких уровнях, например, ≤1 мкг/дозу, ≤100 нг/дозу, ≤10 нг/дозу, ≤1 нг/дозу и т.д.
В настоящем изобретении также предложены устройства для введения (например, шприц, небулайзер, распылитель, ингалятор, накожный пластырь и т.д.), содержащие фармацевтическую композицию настоящего изобретения. Указанные устройства можно использовать для введения композиций настоящего изобретения позвоночным.
Липосомы настоящего изобретения не должны содержать рибосом.
Способы лечения и использование в медицине
В противоположность частицам, раскрытым в ссылке 10, липосомы и фармацевтические композиции настоящего изобретения предназначены для in vivo использования для выработки иммунной реакции на представляющий интерес иммуноген.
В настоящем изобретении предложен способ усиления иммунной реакции у позвоночных, включающий стадию введения эффективного количества липосом или фармацевтической композиции настоящего изобретения. Иммунная реакция является предпочтительно защитной и, предпочтительно, включает иммунитет, опосредованный антителами и/или клетками. Указанный способ может вызвать бустерную реакцию.
В настоящем изобретении также предложены липосома или фармацевтическая композиция настоящего изобретения для использования в способе для усиления иммунной реакции у позвоночных.
В настоящем изобретении также предложено использование липосом настоящего изобретения при изготовлении лекарственных средств для усиления иммунной реакции у позвоночных.
За счет усиления иммунной реакции у позвоночных с помощью указанных лекарственных средств и способов, позвоночные могут быть защищены от различных заболеваний и/или инфекций, например, от бактериальных и/или вирусных заболеваний, как здесь раскрыто. Указанные липосомы и композиции являются иммуногенными, и более предпочтительно, являются композициями вакцин. Вакцины в соответствии с настоящим изобретением могут быть или профилактическими (т.е. предназначенными для предотвращения инфекции), или терапевтическими (т.е. предназначенными для лечения инфекции), но обычно они являются профилактическими.
Позвоночное является, предпочтительно, млекопитающим, таким как человек или крупное ветеринарное млекопитающее (например, лошади, крупный рогатый скот, олени, козы, свиньи). Если вакцина предназначена для профилактического использования, человеком, предпочтительно, является ребенок (например, начинающий ходить ребенок или младенец) или тинэйджер; если вакцина предназначена для терапевтического использования, человеком, предпочтительно, является тинэйджер или взрослый. Вакцину, предназначенную для детей, можно также вводить взрослым, например, для оценки безопасности, дозировки, иммуногенности и т.д.
Вакцины, полученные в соответствии с настоящим изобретением, можно использовать для лечения как детей, так и взрослых. Так возраст пациентов-людей может быть меньше 1 года, меньше 5 лет, в интервале 1-5 лет, 5-15 лет, 15-55 лет, или по меньшей мере 55 лет. Предпочтительными пациентами, которым вводят вакцину, являются старшие (например, ≥50 лет, ≥60 лет, и предпочтительно ≥65 лет), молодые (например, ≤5 лет), госпитализированные пациенты, медицинские работники, военнослужащие и военный персонал, беременные женщины, хронические больные или пациенты с иммунодефицитом. Однако указанные вакцины не предназначены исключительно для указанных групп и их можно более широко использовать для населения.
Композиции настоящего изобретения обычно вводят непосредственно пациенту. Прямое введение можно осуществить, используя парентеральные инъекции (например, подкожные, внутрибрюшинные, внутривенные, внутримышечные, внутрикожные или инъекции в интерстициальное пространство тканей; в отличие от ссылки 1, инъекции в язык в настоящем изобретении обычно не используют). Альтернативные способы введения включают ректальный, пероральный (например, в виде таблеток, спреев), буккальный, сублингвальный, вагинальный, наружный, трансдермальный или транскутанный, интраназальный, окулярный, ушной, пульмонарный или другие способы введения через слизистую. Чрескожное и внутримышечное введение являются двумя предпочтительными способами. Инъекции можно осуществлять через иглы (например, используя шприцы для инъекций), но альтернативно можно использовать шприцы без иголки. Типичная доза для внутримышечного введения составляет 0,5 мл.
Настоящее изобретение можно использовать для выработки системного иммунитета и/или иммунитета слизистых оболочек, предпочтительно для выработки повышенного системного иммунитета и/или иммунитета слизистых оболочек.
Дозы можно вводить или по схеме разовой дозы, или по схеме введения нескольких доз. Множество доз можно использовать в схеме первичной иммунизации и/или в схеме бустерной иммунизации. В схеме множества доз различные дозы можно вводить одним и тем же или различными способами, например, как парентеральную первичную и через слизистую бустерную, через слизистую первичную и парентеральную бустерную и т.д. Множественные дозы обычно вводят с промежутком по меньшей мере в 1 неделю (например, с интервалом около 2 недель, около 3 недель, около 4 недель, около 6 недель, около 8 недель, около 10 недель, около 12 недель, около 16 недель и т.д.). В одном варианте множественные дозы можно вводить через примерно 6 недель, 10 недель и 14 недель после рождения, например, в возрасте 6 недель, 10 недель и 14 недель, как часто используют по расширенной программе иммунизации Всемирной Организации Здравоохранения ("EPI"). В альтернативном варианте, две первичные дозы вводят с промежутком около двух месяцев, например, с промежутком около 7, 8 или 9 недель, после чего вводят одну или более из бустерных доз через от около 6 месяцев до 1 года после введения второй первичной дозы, например, спустя около 6, 8, 10 или 12 месяцев после второй первичной дозы. В следующем варианте три первичные дозы вводят с промежутком около двух месяцев, например, с промежутком около 7, 8 или 9 недель, после чего вводят одну или более из бустерных доз через примерно 6-12 месяцев после третьей первичной дозы, например, около 6, 8, 10 или 12 месяцев после третьей первичной дозы.
Общая часть
В практике настоящего изобретения используют, если не указано иначе, обычные способы химии, биохимии, молекулярной биологии, иммунологии и фармакологии, которые известны специалистам в указанных областях. Такие способы полностью раскрыты в литературе. См., например, ссылки 30-36 и т.д.
Термин "включающий" охватывает "включая", также как "состоящий" например, композиция "включающая" X может состоять исключительно из X или может включать что-либо дополнительно, например, X+Y.
Термин "около" в отношении численного значения x является необязательным и означает, например, x±10%.
Выражение "практически" не исключает "полностью", например, композиция, которая "практически не содержит" Y может полностью не содержать Y. При необходимости, выражение "практически" можно опустить из определения настоящего изобретения.
Ссылки на заряд, катионы, анионы, цвиттерионы и т.д. получены при pH 7.
TLR3 представляет собой Toll-подобный рецептор 3. Он представляет собой простой одиночный трансмембранный рецептор, который играет ключевую роль во врожденной иммунной системе. Известные агонисты TLR3 включают поли(I:C). "TLR3" представляет собой принятое название HGNC для гена, кодирующего указанный рецептор, и его уникальное HGNC ID является HGNC: 11849. Ссылочная последовательность для человеческого TLR3 представляет собой GI:2459625.
TLR7 представляет собой Toll-подобный рецептор 7. Он представляет собой одиночный трансмембранный рецептор, который играет ключевую роль во врожденной иммунной системе. Известные агонисты TLR7 включают, например, имиквимод. "TLR7" представляет собой принятое HGNC название для гена, кодирующего указанный рецептор, и его уникальная HGNC ID представляет собой HGNC: 15631. Ссылочная последовательность для человеческого TLR7 гена представляет собой GI:67944638.
TLR8 представляет собой Toll-подобный рецептор 8. Он представляет собой одиночный трансмембранный рецептор, который играет ключевую роль во врожденной иммунной системе. Известные агонисты TLR8 включают, например, резиквимод. "TLR8" представляет собой принятое HGNC название для гена, кодирующего указанный рецептор, и его уникальный HGNC ID представляет собой HGNC: 15632. Ссылочная последовательность для человеческого TLR8 гена представляет собой GI:20302165.
Семейство RIG-I-подобных рецепторов ("RLR") включает различные РНК геликазы, которые играют ключевую роль во врожденной иммунной системе [37]. RLR-1 (также известный как RIG-I или ген I, индуцируемый ретиновой кислотой) имеет два домена рекрутирования каспазы вблизи его N-конца. Принятое HGNC название для гена, кодирующего геликазу RLR-1, представляет собой "DDX58" (для DEAD (Asp-Glu-Ala-Asp) бокс-полипептид 58) и уникальный HGNC ID представляет собой HGNC: 19102. Сравнительная последовательность для человеческого RLR-1 гена представляет собой GI:77732514. RLR-2 (известный также как MDA5 или ген 5, ассоциированный с дифференцированием меланомы) также содержит два домена рекрутирования каспазы вблизи своего N-конца. Принятое название HGNC для гена, кодирующего геликазу RLR-2, представляет собой "IFIH1" (для интерферона, индуцируемого доменом 1 геликазы С) и уникальный HGNC ID представляет собой HGNC: 18873. Сравнительная последовательность для человеческого гена RLR-2 представляет собой GI: 27886567. RLR-3 (также известный как LGP2 или лаборатория генетики и физиологии 2) не содержит доменов рекрутирования каспазы. Принятое наименование HGNC для гена, кодирующего геликазу RLR-3, представляет собой "DHX58" (для DEXH (Asp-Glu-X-His) бокс-полипептида 58) и уникальный HGNC ID представляет собой HGNC:29517. Сравнительная последовательность для человеческого гена RLR-3 представляет собой GI: 149408121.
PKR представляет собой двухцепочечную РНК-зависимую протеинкиназу. Она играет ключевую роль во врожденной иммунной системе. "EIF2AK2" (для эукариотного фактора инициации трансляции 2-альфакиназы 2) представляет собой принятое название HGNC для гена, кодирующего указанный фермент, и его уникальный HGNC ID представляет собой HGNC:9437. Сравнительная последовательность для человеческого гена PKR представляет собой GI:208431825.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГ.1 демонстрирует гель с окрашенной РНК. Полоски демонстрируют (1) маркеры, (2) оголенный репликон, (3) репликон после обработки РНКазой, (4) репликон, инкапсулированный в липосоме, (5) липосома после обработки РНКазой, (6) липосома, обработанная РНКазой, и затем подвергнутая экстрагированию фенолом/хлороформом.
ФИГ.2 представляет собой электронную микрофотографию липосомы.
ФИГ.3 демонстрирует экспрессию белка (в виде относительных световых единиц, RLU) в дни 1, 3 и 6 после введения РНК в крупные (нижняя линия) или мелкие (верхняя линия) липосомы.
ФИГ.4 демонстрирует гель с окрашенной РНК. Полоски демонстрируют (1) маркеры, (2) оголенный репликон, (3) репликон, инкапсулированный в липосому, (4) липосома, обработанная РНКазой и затем подвергнутая экстрагированию фенолом/хлороформом.
ФИГ.5 демонстрирует экспрессию белка в дни 1, 3 и 6 после введения РНК в виде репликона, упакованного в вирион (квадраты), в виде оголенной РНК (звездочки), или в липосомы (+=0,1 мкг, x=1 мкг).
ФИГ.6 демонстрирует экспрессию в дни 1, 3 и 6 после введения четырех различных инкапсулированных в липосомы РНК.
ФИГ.7 демонстрирует титры анти-F IgG у животных, которым вводили упакованный в вирион репликон (VRP или VSRP), 1 мкг оголенной РНК и 1 мкг инкапсулированной в липосомы РНК.
ФИГ.8 демонстрирует титры анти-F IgG у животных, которым вводили VRP, 1 мкг оголенной РНК и 0,1 г или 1 мкг инкапсулированной в липосомы РНК.
ФИГ.9 демонстрирует титры нейтрализующих антител у животных, которым вводили VRP, или 0,1 г или 1 мкг инкапсулированных в липосомах РНК.
ФИГ.10 демонстрирует уровни экспрессии после введения репликона в виде оголенной РНК (окружности), инкапсулированной в липосомах РНК (треугольники и квадраты), или в виде липоплекса (перевернутые треугольники).
ФИГ.11 демонстрирует титры F-специфических IgG (2 недели после второй дозы) после введения репликона в виде оголенной РНК (0,01-1 мкг), инкапсулированной в липосомы РНК (0,01-10 мкг), или упакованной в виде вириона (VRP, 106 инфекционная доза или IU).
ФИГ.12 демонстрирует титры F-специфических IgG (окружности) и титры PRNT (квадраты) после введения репликона в виде оголенной РНК (1 мкг), инкапсулированной в липосомы РНК (0,1 или 1 мкг), или упакованной в виде вириона (VRP, 106 IU). Также показаны титры у интактных мышей. Жирные линии демонстрируют геометрию.
ФИГ.13 демонстрирует продуцирование внутриклеточного цитокина после повторной стимуляции синтетическими пептидами, представляющими основные эпитопы F белка, через 4 недели после второй дозы. По оси y отложен % цитокинов+ из CD8+CD4-.
ФИГ.14 демонстрирует титры F-специфических IgG (средние log10 титры ± ср.откл.) через 210 дней после иммунизации телят. Три линии четко различимы к 63 дню и представляют собой, снизу вверх: PBS негативный контроль; доставленные липосомами РНК; и "Треугольники 4" продукт.
ФИГ.15 демонстрирует увеличение (относительное) титров анти-F через две недели после первой дозы репликона, кодирующего F белок. Значения титров отложены против среднего диаметра липосом Z (нм).
ФИГ.16A-16M демонстрируют структуру альтернативных катионных липидов: (A) RV05; (B) RV02; (C) RV04; (D) RV07; (E) RV03; (F) RV08; (G) RV09; (H) RV14; (I) RV10; (J) RV11; (K) RV15; (L) RV16; (M) RV17.
ФИГ.17 демонстрирует структуру полезных "сплит" ПЭГ-конъюгированных липидов. Полный молекулярный вес ПЭГ внутри бокса составляет 2000 для тестированных липосом.
ФИГ.18A-18E демонстрируют структуры различных ПЭГ-конъюгированных липидов, где R представляет собой ПЭГ необходимой длины.
ПРЕДПОЧТИТЕЛЬНЫЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
РНК репликоны
Далее использованы различные репликоны. Вообще они основаны на гибридном геноме альфавируса с неструктурными белками вируса Венесуэльского лошадиного энцефалита (VEEV), сигнала упаковки из VEEV, и 3’ UTR из Синдбис вируса или VEEV мутанта. Указанный репликон имеет около 10 кб в длину и содержит поли-A концевую группу.
Плазмидная ДНК, кодирующая репликоны альфавируса (называемые: pT7-mVEEV-FL.RSVF или A317; pT7-mVEEV-SEAP или A306; pSP6-VCR-GFP или A50), служит в качестве матрицы для синтеза РНК in vitro. Такие репликоны содержат генетические элементы альфавируса, необходимые для репликации РНК, но не содержат тех кодирующих генных продуктов, которые необходимы для сборки частиц; структурные белки вместо этого заменены представляющим интерес белком (или репортером, таким как SEAP или GFP, или иммуногеном, таким как полной длины RSV F белок) и поэтому указанные репликоны не способны индуцировать образование инфекционных частиц. Промотор бактериофага (T7 или SP6) в обратном направлении от альфавирусной кДНК облегчает синтез РНК репликона in vitro, и рибозим вируса дельта гепатита (HDV) непосредственно в прямом направлении от поли(A)-концевой группы образует правильный 3’-конец за счет активности саморасщепления.
После линеаризации плазмидной ДНК, расположенной в прямом направлении от рибозима HDV, подходящей рестрикционной эндонуклеазой, ран-офф транскрипты синтезируют in vitro, используя ДНК-зависимые РНК полимеразы бактериофагов T7 или SP6. Ионы транскриптов выдерживают в течение 2 часов при 37°C в присутствии 7,5 мМ (T7 РНК полимеразы) или 5 мМ (SP6 РНК полимеразы) каждого из нуклеозидтрифосфатов (ATP, CTP, GTP и UTP) в соответствии с инструкциями, предоставленными изготовителем (Ambion). После транскрипции матричную ДНК переваривают, используя TURBO ДНКазу (Ambion). РНК репликон осаждают, используя LiCl и восстанавливают в не содержащей нуклеазы воде. Для некэпированной РНК создают кэп-группировку посттранскриптционно, используя кэпирующий вакцину фермент (Vaccinia Capping Enzyme (VCE)), используя систему кэпинга ScriptCap m7G (Epicentre Biotechnologies) в соответствии с рекомендациями для пользователя; репликонам, снабженным такой кэп-группой присваивают приставку "v", например, vA317 представляет собой A317 репликон, кэпированный VCE. Посттранскрипционно кэпированные РНК осаждают, используя LiCl, и восстанавливают в не содержащей нуклеазы воде. Концентрацию образцов РНК определяют, измеряя OD260нм. Целостность in vitro транскриптов подтверждают, используя электрофорез в агарозном геле в условиях денатурации.
Липосомальное инкапсулирование
РНК инкапсулируют в липосомы практически по способу, представленному в ссылках 7 и 38. Липосомы получают из 10% DSPC (цвиттерионный), 40% DlinDMA (катионный), 48% холестерина и 2% ПЭГ-конъюгированного DMG (2 кДа ПЭГ). Указанные пропорции относятся к молярному % в полном количестве липосом.
DlinDMA (1,2-дилинолеилокси-N,N-диметил-3-аминопропан) синтезируют, используя способ, раскрытый в ссылке 2. DSPC (1,2-диастеароил-sn-глицеро-3-фосфохолин) получают от Genzyme. Холестерин получают от Sigma-Aldrich. ПЭГ-конъюгированный DMG (1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин-N-[метокси(полиэтиленгликоль), аммониевая соль), DOTAP (1,2-диолеоил-3-триметиламмонийпропан, хлоридная соль) и DC-chol (3P-[N-(N’,N’-диметиламиноэтан)карбамоил]холестерин гидрохлорид) получают от Avanti Polar Lipids.
Короче, липиды растворяют в этаноле (2 мл), РНК репликон растворяют в буфере (2 мл, 100 мМ цитрата натрия, pH 6) и все это смешивают с 2 мл буфера, затем уравновешивают в течение 1 часа. Полученную смесь разбавляют 6 мл буфера, затем фильтруют. Полученный продукт содержит липосомы с эффективностью инкапсулирования ~95%.
Например, в одном конкретном способе свежие липидные исходные растворы приготавливают в этаноле. 37 мг DlinDMA, 11,8 мг DSPC, 27,8 мг холестерина и 8,07 мг ПЭГ-DMG взвешивают и растворяют в 7,55 мл этанола. Свежеприготовленный липидный исходный раствор осторожно встряхивают при 37°C в течение около 15 мин, получая гомогенную смесь. Затем 755 мкл исходного раствора добавляют к 1,245 мл этанола, получая рабочий липидный исходный раствор в количестве 2 мл. Указанное количество липидов используют для создания липосом с 250 мкг РНК. 2 мл рабочего раствора РНК также приготавливают из исходного раствора ~1 мкг/мкл в 100 мМ цитратном буфере (pH 6). Три стеклянные ампулы объемом 20 мл (с якорями мешалки) промывают раствором RNase Away (Molecular BioProducts) и промывают большим количеством MilliQ воды перед тем, как используют для очищения ампул от РНКаз. Одну из ампул используют для РНК рабочего раствора и другие для сбора смесей липида и РНК (как раскрыто далее). Рабочие липидный и РНК растворы нагревают при 37°C в течение 10 мин перед тем, как помещают в 3 см3 шприцы с люэровским клапаном. 2 мл цитратного буфера (pH 6) помещают в другой 3 см3 шприц. Шприцы, содержащие РНК и липиды, соединяют с T миксером (PEEK™ 500 мкм ID соединение, Idex Health Science), используя FEP трубку (фторированный этилен-пропилен; все используемые FEP трубки имеют внутренний диаметр 2 мм и внешний диаметр 3 мм; получены от Idex Health Science). На выходе из T миксера находится также FEP трубка. Третий шприц, содержащий цитратный буфер, соединен с отдельным отрезком трубки. Затем все шприцы прокачивают со скоростью 7 мл/мин, используя шприцевый инфузионный насос. Выходы трубок расположены таким образом, чтобы собирать смеси в 20 мл стеклянные ампулы (при перемешивании). Якоря мешалок удаляют и дают возможность этанольному/водному раствору уравновеситься при комнатной температуре в течение 1 часа. 4 мл смеси помещают в 5 см3 шприц, который соединен с отрезком FEP трубки, и в другой 5 см3 шприц, соединенный с равной длины отрезком FEP трубки, помещают одинаковое количество 100 мМ цитратного буфера (pH 6). Два шприца прокачивают со скоростью потока 7 мл/мин, используя шприцевый инфузионный насос, и конечную смесь собирают в 20 мл стеклянную ампулу (при перемешивании). Затем смесь, собранную на второй стадии смешивания (липосомы), пропускают через мембрану Mustang Q (анионообменная подложка, которая связывает и удаляет анионные молекулы, получена от Pall Corporation). Перед использованием указанной мембраны для липосом, через нее последовательно пропускают 4 мл 1 M NaOH, 4 мл 1 M NaCl и 10 мл 100 мМ цитратного буфера (pH 6). Липосомы нагревают в течение 10 мин при 37°C перед тем, как их пропускают через мембрану. Затем липосомы концентрируют до 2 мл и осуществляют диализ против 10-15 объемов 1X PBS, используя фильтрование тангенциальным потоком перед тем, как выделяют конечный продукт. Систему TFF и половолоконные фильтрационные мембраны закупают у Spectrum Labs (Rancho Dominguez) и используют в соответствии с рекомендациями изготовителей. Используют полисульфоновые половолоконные фильтрационные мембраны с отсечением по размеру пор 100 кД и площадью поверхности 8 см2. Для экспериментов in vitro и in vivo полученные композиции разбавляют 1X PBS до необходимой концентрации РНК.
ФИГ.2 представляет собой пример электронной микрофотографии липосом, полученных указанными способами. Эти липосомы содержат инкапсулированные РНК, кодирующие полной длины антиген RSV F. По данным динамического светорассеяния одна партия имеет средний диаметр 141 нм (по интенсивности) или 78 нм (по числу).
Процент инкапсулированных РНК и концентрацию РНК определяют, используя реагентный набор Quant-iT RiboGreen RNA (Invitrogen), в соответствии с инструкциями изготовителей. Рибосомальный РНК стандарт, предоставленный в наборе, используют для построения стандартной кривой. Липосомы разбавляют 10× или 100× в 1X TE буфере (из набора) перед добавлением красителя. Отдельно липосомы разбавляют 10× или 100× в 1X TE буфере, содержащем 0,5% Triton X перед добавлением красителя (для разрушения липосом и определения таким образом полного содержания РНК). После этого равное количество красителя добавляют в каждый раствор и затем ~180 мкл каждого раствора после добавления красителя помещают в двойном экземпляре в 96-луночные планшеты для культуры тканей. Интенсивность флуоресценции (возбуждение на 485 нм, эмиссия на 528 нм) считывают, используя считывающее устройство для микропланшетов. Все композиции липосом дозируют in vivo на основании количества инкапсулированных РНК.
Было показано, что инкапсулирование в липосомы защищает РНК от расщепления РНКазой. В экспериментах используют 3,8 мАЕд (mAU - единицы оптической плотности) РНКазы A на микрограмм РНК, инкубированные в течение 30 минут при комнатной температуре. РНКазу инактивируют, используя протеиназу K при 55°C в течение 10 минут. Затем добавляют смесь A 1:1 об/об образца к смеси 25:24:1 об/об/об фенол:хлороформ:изоамиловый спирт для экстрагирования РНК из липидов в водную фазу. Образцы перемешивают на вортексе в течение нескольких секунд и затем помещают в центрифугу на 15 минут при скорости 1200 об/мин. Водную фазу (содержащую РНК) удаляют и используют для анализа РНК. Перед загрузкой (400 нг РНК на ячейку) все образцы инкубируют с формальдегидным красителем, денатурируют в течение 10 минут при 65°C и охлаждают до комнатной температуры. Маркеры Ambion Millennium используют для аппроксимации молекулярного веса конструкции РНК. Гель обрабатывают при 90 Вольт. Гель окрашивают, используя 0,1% SYBR золото в соответствии с рекомендациями изготовителей в воде на качалке при комнатной температуре в течение 1 часа. ФИГ.1 демонстрирует, что РНКаза полностью расщепила РНК в отсутствии инкапсуляции (полоска 3). РНК не детектируется после инкапсуляции (полоска 4), и никаких изменений не наблюдают, для липосом, не обработанных РНКазой (полоска 4). После того, как обработанные РНКазой липосомы экстрагируют фенолом, видны нерасщепленные РНК (полоска 6). Даже после 1 недели при 4°C можно наблюдать РНК без какой-либо фрагментации (ФИГ.4, стрелки). Экспрессия белка in vivo остается неизменной после 6 недель при 4°C и одного цикла замораживание-оттаивание. То есть, инкапсулированные в липосомы РНК стабильны.
Для оценки экспрессии РНК in vivo фермент-репортер (SEAP; секретируемая щелочная фосфатаза) кодируют скорее в репликон, чем в иммуноген. Уровни экспрессии определяют в сыворотке, разбавленной 1:4 в 1X Phospha-Light буфере для разбавления, используя хемилюминесцентный щелочной фосфатный субстрат. 8-10 недельным мышам BALB/c (5/группу) внутримышечно вводят инъекции в день 0,50 мкл в лапу дозу РНК 0,1 мкг или 1 мкг. Тот же самый вектор вводят без липосом (в не содержащем РНКазы 1X PBS) в дозе 1 мкг. Тестируют также упакованные в вирион репликоны. Используемые здесь упакованные в вирион репликоны (называемые "VRP") получают по способу ссылки 39, где альфавирусный репликон получают из мутантного VEEV или химеры, полученной из генома VEEV, сконструированного так, чтобы содержать 3’ UTR Синдбис вируса и сигнал упаковки Синдбис вируса (PS), упакованные путем совместной их электропорации в BHK клетки с дефективными хелперными РНК, кодирующими гены капсида Синдбис вируса и гликопротеин.
Как представлено на ФИГ.5, инкапсулирование повышает SEAP уровни на около ½log при дозе 1 мкг, и в день 6 экспрессия за счет 0,1 мкг инкапсулированной дозы соответствует значениям для 1 мкг неинкапсулированной дозы. К 3 дню уровни экспрессии превышают уровни, достигнутые с VRP (квадраты). Таким образом, экспрессия возрастает, если РНК инкапсулированы в липосомы при сравнении с контрольными оголенными РНК, даже при дозе, которая в 10 раз ниже. Экспрессия также оказывается выше по сравнению с VRP контролем, но кинетики экспрессии значительно отличаются (см. ФИГ.5). Введение РНК путем электропорации приводит к возрастанию экспрессии по сравнению с контрольными оголенными РНК, но эти уровни оказываются ниже, чем с липосомами.
Для выяснения того, связан ли наблюдаемый эффект в группе липосом просто с компонентами липосом, или он связан с инкапсулированием, репликон вводят в инкапсулированной форме (для двух различных протоколов очистки, 0,1 мкг РНК), или в смеси с липосомами после их образования (неинкапсулированный "липоплекс", 0,1 мкг РНК), или в виде оголенной РНК (1 мкг). ФИГ.10 демонстрирует, что использование липоплекса приводит к самым низким уровням экспрессии, свидетельствуя о том, что инкапсулирование существенно для эффективной экспрессии.
Дальнейшие SEAP эксперименты демонстрируют отчетливую зависимость реакции от дозы in vivo, когда экспрессия заметна уже после введения столь малого количества, как 1 нг РНК (ФИГ.6). Дальнейшие эксперименты по сравнению экспрессии за счет инкапсулированных и оголенных репликонов показывают, что эффект от 0,01 мкг инкапсулированных РНК эквивалентен 1 мкг оголенных РНК. В дозе 0,5 мкг РНК инкапсулированный материал обеспечивает в 12 раз более высокую экспрессию в день 6; в дозе 0,1 мкг уровни экспрессии оказываются в 24 раза выше в день 6.
Были исследованы не только средние значения в группе животных, но исследовали также отдельных животных. Хотя несколько животных не реагировали на оголенные репликоны, инкапсулирование исключает неотвечающие особи.
В дальнейших экспериментах DlinDMA заменяют на DOTAP. Хотя DOTAP липосомы приводят к более высокой экспрессии, чем оголенные репликоны, их результаты оказались хуже, чем для DlinDMA липосом (различие в 2-3 раза в день 1).
Для оценки in vivo иммуногенности конструируют репликон для экспрессии полной длины F белка из респираторно-синтициального вируса (RSV). Его вводят оголенным (1 мкг), инкапсулированным в липосомы (0,1 или 1 мкг), или упакованным в вирионы (106 IU; "VRP") в дни 0 и 21. ФИГ.7 демонстрирует титры анти-F IgG через 2 недели после введения второй дозы, и отчетливо видно, что липосомы повышают иммуногенность. ФИГ.8 демонстрирует титры спустя 2 недели, причем к этому моменту не наблюдается статистических различий между инкапсулированными РНК в дозе 0,1 мкг, инкапсулированными РНК в дозе 1 мкг, или VRP группой. Титры нейтрализации (измеренные как 60% уменьшения бляшек, "PRNT60") заметно не отличались от указанных трех групп через 2 недели после введения второй дозы (ФИГ.9). ФИГ.12 демонстрирует титры как IgG, так и PRNT, через 4 недели после введения второй дозы.
ФИГ.13 подтверждает тот факт, что РНК вызывает значительную реакцию CD8 T клеток.
В дальнейших экспериментах проводят сравнение титров F-специфичных IgG у мышей, которым вводили VRP, 0,1 мкг инкапсулированных в липосомы РНК, или 1 мкг инкапсулированных в липосомы РНК. Отношения титров (VRP: липосомы) в различные моменты времени после введения второй дозы оказались следующими:
Таким образом, инкапсулированные в липосомы РНК индуцируют иммунную реакцию практически той же величины, что наблюдается при введении вирионов.
Дальнейшие эксперименты демонстрируют превосходящие F-специфические IgG реакции в дозе 10 мкг, эквивалентные реакции для доз 1 мкг и 0,1 мкг и более слабые реакции в дозе 0,01 мкг. ФИГ.11 демонстрирует титры антител IgG для мышей, которым вводили репликон в оголенной форме в 3 различных дозах, в липосомах в 4 различных дозах, или в виде VRP (106 IU). Реакция, которую наблюдали при введении 1 мкг инкапсулированных в липосомы РНК, оказалась статистически незначительной (ANOVA) при сравнении с VRP, но более значительная реакция, которую наблюдали при введении 10 мкг инкапсулированных в липосомы РНК, оказалась статистически значимой (p<0,05) по сравнению с обеими указанными группами.
Дальнейшие исследования подтверждают, что 0,1 мкг инкапсулированных в липосомы РНК вызывают гораздо более сильные анти-F IgG реакции (через 15 дней после введения второй дозы), чем 0,1 мкг введенной ДНК, и оказываются даже более иммуногенными чем 20 мкг плазмидной ДНК, кодирующей F антиген, введенный с помощью электропорации (Elgen™ ДНК Delivery System, Inovio).
Хлопковые хомяки
В исследованиях используют хлопковых хомяков (Sigmodon hispidis) вместо мышей. В дозе 2 мкг инкапсулирование липосом увеличивает титры F-специфических IgG в 8,3 раза по сравнению с оголенными РНК и повышает титры PRNT в 9,5 раза. Интенсивность реакции антител оказалась эквивалентной интенсивности реакции, вызванной 5×106 ИмЕд VRP. Как оголенные, так и инкапсулированные в липосомы РНК оказались способны защитить хлопковых хомяков от заражения RSV (1×105 бляшкообразующих единиц), уменьшая концентрацию вируса в легких по меньшей мере в 3,5 log. Инкапсулирование усиливает уменьшение примерно в 2 раза.
В дальнейшей работе с хлопковыми хомяками используют четыре различных репликона: vA317 экспрессирует полной длины RSV-F; vA318 экспрессирует усеченный (с удаленными трансмембранным и цитоплазмическим концевыми сегментами) RSV-F; vA142 экспрессирует RSV-F с исключенным фьюжн пептидом; vA140 экспрессирует усеченный RSV-F также без своего пептида. Хлопковым хомякам, от 4 до 8 животных в группе, проводят внутримышечную вакцинацию (100 мл в одну лапу) в дни 0 и 21 четырьмя различными репликонами, введенными в липосомы в двух дозах (1,0 и 0,1 мкг), используя 2 кДа ПЭГ-конъюгированный DMG по способу (D), но с порцией 150 мкг РНК. Контрольным группам вводят вакцину белка RSV-F-субъединицы (5 мкг) с адъювантом - квасцами (8 животных/группу), VRP экспрессирующие полной длины RSV-F (1×106 IU, 8 животных/группу), или интактный контроль (4 животных/группу). Сыворотку собирают для анализа антител в дни 0, 21 и 34.
Титры F-специфических сывороточных IgG и титры сывороточных антител, нейтрализующих RSV, в дни 21 и 34 были:
Все четыре репликона, которые оценивают в данном исследовании (vA317, vA318, vA142, vA140), являются иммуногенными для хлопковых хомяков, если их вводят в липосомах, хотя титры нейтрализации сыворотки были по меньшей мере в десять раз ниже, чем в случае индуцирования белковыми вакцинами с адъювантами, или в случае VRP. Вакцины липосомы/РНК вызывают выработку сывороточных F-специфических IgG и нейтрализующих RSV антител после первой вакцинации, и вторая вакцинация эффективно усиливает реакцию. Титры F-специфических IgG после второй вакцинации 1 мкг репликонов оказываются в 2-3 раза выше, чем после второй вакцинации 0,1 мкг репликонов. Указанные четыре репликона приводят к сопоставимым титрам антител, позволяя предположить, что полной длины и усеченный RSV-F, каждый с фьюжн пептидами или без них, оказываются одинаково иммуногенными для хлопковых хомяков.
В следующем эксперименте с хлопковыми хомяками снова используют vA317, vA318 и vA142 репликоны. Хлопковым хомякам, по 2-8 животных в группе, проводят внутримышечные вакцинации (100 мкл в одну лапу) в дни 0 и 21, используя репликоны (0,1 или 1 мкг), инкапсулированные в RV01 липосомы (с ПЭГ-2000), полученные по способу (D), но с порцией 150 мкг РНК. Контрольным группам вводят RSV-F субъединичную белковую вакцину (5 мкг), дополненную квасцами или VRP эксперессирующими полной длины RSV-F (1×106 IU, 8 животных/группу). Всем животным проводят третью вакцинацию (день 56), используя RSV-F субъединичную белковую вакцину (5 мкг) с адъювантом - квасцами. Кроме того, был интактный контроль (4 животных/группу). Кроме того, дополнительной группе проводят внутримышечно бактериальные вакцинации (50 мкл в лапу) в дни 0 и 56, используя 1 мкг vA317 РНК в липосомах, но не проводят третью вакцинацию субъединичной белковой вакциной.
Сыворотку собирают для анализа антител в дни 0, 21, 35, 56, 70, плюс дни 14, 28 и 42 для дополнительной группы. Титры сывороточного F-специфического IgG (GMT) оказались следующими:
Титры нейтрализации сыворотки были следующими (60% RSV титр нейтрализации для 2 групп по 3-4 животных в группе, GMT для указанных 2 групп были на группу):
Сывороточные титры и титры нейтрализующих антител для дополнительной группы были следующими:
Подтверждено, что указанные репликоны являются иммуногенными для хлопковых хомяков, вырабатывая сывороточные F-специфические IgG и нейтрализующие RSV антитела после первой вакцинации. Вторая вакцинация повышает эффективность реакций. Титры F-специфических IgG после второй вакцинация 1,0 мкг репликона оказались в 1,5-4 раза выше, чем после второй вакцинации 0,1 мкг репликона.
Третья вакцинация (белок в 56 день) не повышает титры у хлопковых хомяков, которые ранее были вакцинированы F тримерной субъединицей + квасцы, но она обеспечила значительное увеличение титров у хлопковых хомяков, которые предварительно были вакцинированы репликонами. В большинстве случаев сывороточные титры нейтрализующих RSV антител после двух вакцинаций репликонами с последующим введением белка оказались равными или больше, чем титры, стимулированные двумя или тремя последовательными вакцинациями белком.
В рассматриваемом исследовании также оценивают кинетические особенности реакции антител на 1,0 мкг vA317. Титры сывороточных F-специфических IgG и титры нейтрализующих RSV антител, стимулированные одной вакцинацией, достигают своих пиковых значений примерно на 21 день, и сохраняются по меньшей мере до 56 дня (50-70% капель в титр F-специфических IgG, мало изменений в титре нейтрализующих RSV антител). Вторую гомологичную вакцинацию проводят для указанных животных в день 56, и повышают титры антител до уровня по меньшей мере равного тому, которого достигают, если вторую вакцинацию проводят в день 21.
Дальнейшие эксперименты включают заражение вирусами. vA368 репликон кодирует полной длины дикого типа поверхностный фьюжн гликопротеин RSV, из которого удален фьюжн пептид, причем экспрессию стимулируют EV71 IRES. Хлопковым хомякам, по 7 в группе, проводят внутримышечную вакцинацию (100 мкл в лапу) в дни 0 и 21, используя vA368 в липосомах, полученный по способу (H), размер порции 175 мкг РНК, или VRP с тем же самым репликоном. Указанные липосомы включают 2 кДа ПЭГ, конъюгированного с DMG. Контрольной группе вводят 5 мкг белка, дополненного квасцами, и включают также интактную контрольную группу.
Для всех групп осуществляют интраназальное заражение (i.n.), используя 1×106 БОЕ RSV, через четыре недели после окончательной иммунизации. Сыворотку собирают для анализа антител в дни 0, 21, 35. Титры вируса в легких измеряют на 5 день после заражения. Получены следующие результаты:
Таким образом, РНК вакцина снижает содержание вирусов в более чем 1000 раз, с приблизительно 106 БОЕ/г у невакцинированных контрольных хлопковых хомяков до менее чем 103 БОЕ/г у вакцинированных контрольных хлопковых хомяков
Исследования, проводимые на крупных млекопитающих
Исследования осуществляют на крупном рогатом скоте. Проводят иммунизацию телят (4-6 недельные, ~60-80 кг, 5 в группе), используя 66 мкг репликона vA317, кодирующего полной длины F белок RSV, в дни 0, 21, 86 и 146. Указанные репликоны включены в липосомы по способу (E), но с добавлением 1,5 мг РНК; они содержат 40% DlinDMA, 10% DSPC, 48% холестерина и 2% ПЭГ-2000, конъюгированного с DMG. Один только PBS используют в качестве негативного контроля, и в качестве позитивного контроля используют лицензионную вакцину ("Triangle 4" от Fort Dodge, содержащую убитый вирус). Всем телятам вводят 15 мкг F белка, дополненного MF59 эмульсией в день 146.
РНК вакцины кодируют человеческий RSV F, тогда как вакцины "Triangle 4" содержат коровий RSV F, но RSV F белок является высоко консервативным между BRSV и HRSV.
Телятам вводят по 2 мл каждой экспериментальной вакцины, внутримышечно как инъекции 2×1 мл в обе стороны шеи. Напротив, вакцину "Triangle 4" вводят как инъекцию одной 2 мл дозы в шею.
Сыворотку собирают для анализа антител в дни 0, 14, 21, 35, 42, 56, 63, 86, 100, 107, 114, 121, 128, 135, 146, 160, 167, 174, 181, 188, 195 и 202. Если у отдельного животного значение титра находится ниже предела определения, этот титр оценивают как 5.
ФИГ.14 демонстрирует титры F-специфических IgG за 210 дней. В течение первых 63 дней РНК репликон является иммуногенным для телят за счет липосом, хотя он дает более низкие титры, чем лицензионная вакцина. У всех вакцинированных телят наблюдают F-специфические антитела после второй дозы, и их титры оказываются очень стабильными в течение 2-6 недель после введения второй дозы (и оказываются особенно стабильными для РНК вакцины). Титры вплоть до дня 202 были следующими:
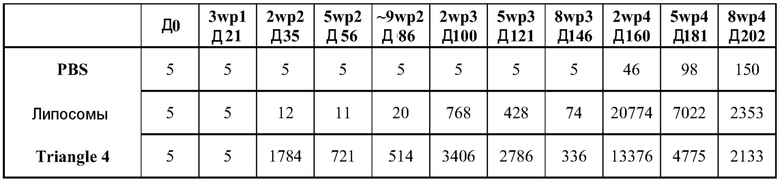
Титры сывороточных антител, нейтрализующих RSV, были следующими:
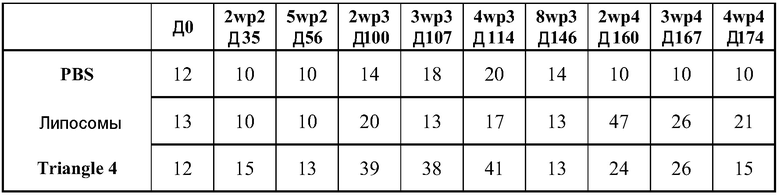
Материал, который был использован для второй липосомальной дозы, не был свежеприготовленным, и та же партия РНК продемонстрировала снижение эффективности в исследованиях иммуногенности на мышах. Поэтому существует вероятность того, что указанная вакцина была бы более иммуногенной, если бы для всех вакцинаций были использованы свежие материалы.
При проведении анализов с комплементом, нейтрализующие антитела детектировались у всех вакцинированных телят. В указанном анализе у всех вакцинированных телят были хорошие титры нейтрализующих антител после второй РНК вакцинации. Кроме того, РНК вакцина приводит к появлению титров сывороточных F-специфических IgG, которые наблюдались у нескольких телят после второй вакцинация и у всех телят после третьей вакцинации.
RSV-F с добавленным в качестве адъюванта MF59 оказался способным бустировать реакцию IgG у всех ранее вакцинированных телят, и бустировать титры комплемент-независимых нейтрализующих антител у телят, предварительно вакцинированных РНК.
Доказательство концепции РНК вакцины для крупных животных является особенно важным в свете потерь эффективности, наблюдавшейся ранее для вакцин на базе ДНК, при переходе от моделей с мелкими животными к крупным животным и людям. Типичная доза ДНК вакцины для коров должна составлять 0,5-1 мг [40, 41], и поэтому весьма обнадеживает тот факт, что иммунные реакции можно вызывать, используя всего лишь 66 мкг РНК.
Влияние диаметра липосом
Для получения более мелких липосом способ со шприцом/трубкой заменяют способом, в котором растворы липидов и РНК смешивают в каналах микрожидкостного чипа.
Приготавливают свежие исходные растворы липидов в этаноле. Взвешивают 37 мг DlinDMA, 11,8 мг DSPC, 27,8 мг холестерина и 8,07 мг ПЭГ-DMG и растворяют в 7,55 мл этанола. Свежеприготовленный липидный исходный раствор осторожно встряхивают при 37°C в течение около 15 мин для получения гомогенной смеси. Затем 226,7 мкл исходного раствора добавляют к 1,773 мл этанола для получения 2 мл исходного рабочего раствора липидов. 4 мл рабочего раствора РНК также приготавливают из исходного раствора ~1 мкг/мкл в 100 мМ цитратном буфере (pH 6). Четыре 20 мл стеклянные ампулы (с магнитными якорями) промывают раствором РНКазы Away и обильно промывают водой MilliQ перед тем, как используют для очищения ампул от РНКаз. Две из ампул используют для рабочего раствора РНК (по 2 мл в каждой ампуле) и остальные используют для сбора смесей липидов и РНК. Рабочие липидный и РНК растворы нагревают при 37°C в течение 10 мин перед тем, как помещают в 3 см3 шприцы с люэровским клапаном. Шприцы, содержащие РНК и липиды, соединяют с Mitos Droplet junction Chip (стеклянное микрожидкостное устройство от Syrris, Part № 3000158), используя PTFE трубки с внутренним диаметром 0,076 см и внешним диаметром 0,16 см, (Syrris) используя 4-сторонний краевой разъем. Два потока РНК и один липидный поток направляют с помощью шприцевых насосов, и смешение этанольной и водной фаз происходит в X соединении чипа (100 мкм × 105 мкм). Скорости всех трех потоков поддерживают при 1,5 мл/мин, следовательно, отношение полного количества водной фазы к этанольной составляет 2:1. Выходную трубку располагают таким образом, чтобы собирать смеси в 20 мл стеклянные ампулы (при перемешивании). Магнитные якоря убирают и этанольный/водный раствор оставляют уравновешиваться до комнатной температуры в течение 1 часа. Затем полученную смесь помещают в 5 см3 шприц, снабженный кусочком PTFE трубки с внутренним диаметром 0,076 см и внешним диаметром 0,16 см, и в другой 5 см3 шприц с равной длины PTFE трубкой, и вводят равные объемы 100 мМ цитратного буфера (pH 6). Скорости в шприцах поддерживают 3 мл/мин, используя шприцевые насосы, и конечную смесь собирают в 20 мл стеклянную ампулу (при перемешивании). Затем липосомы концентрируют до 2 мл и осуществляют диализ против 10-15 объемов 1X PBS, используя систему TFF, перед тем, как выделяют конечный продукт. Используют половолоконные фильтрационные мембраны с отсечением по размеру пор 100 кДа и площадью поверхности 20 см2. Для in vitro и in vivo экспериментов состав разбавляют до нужной концентрации РНК, используя 1X PBS.
Если липосомы, полученные с использованием способа шприц/трубка для 75 мкг РНК имеют средний диаметр Z 148 нм и показатель полидисперсности 0,122, смешивание с использованием микрожидкостного чипа обеспечивает получение среднего диаметра Z 97 нм и с показателем полидисперсности 0,086. Доля инкапсулированных РНК несколько снижается с 90% до 87%. Указанные диаметры и показатели полидисперсности измеряют, используя зетасайзер (прибор для измерения размеров наночастиц) Zetasizer Nano ZS (Malvern Instruments, Worcestershire, UK) в соответствии с инструкциями изготовителя. Перед измерением липосомы разбавляют в 1X PBS.
Полученные липосомы вводят 8-10 недельным BALB/c мышам путем внутримышечной инъекции в день 0, 50 мкл в лапу. Образцы синус-орбитального кровотечения отбирают в дни 1 и 3, и окончательно в день 6. Измеряют сывороточные уровни SEAP, используя хемилюминесцентный анализ. Как видно на ФИГ.3, более мелкие липосомы повышают уровни SEAP примерно в 2 раза в день 1 и примерно в 5 раз в день 6.
Липосомы, полученные двумя различными способами, оценивают также в отношении доставки репликонов, кодирующих полной длины RSV-F белок. Титры F-специфического сывороточного IgG у мышей, по 8 животных в группе, определяют после внутримышечных вакцинаций в дни 0 и 21. Сыворотку собирают для анализа антител в дни 14 (2wp1) и 35 (2wp2). Если титры у отдельного животного оказываются <25 (предел детектирования), их оценивают как титры 5. Далее представлены результаты в виде среднего геометрического значения для титров каждой группы:

Таким образом, более мелкие липосомы, полученные способом смешивания в чипе, обеспечивают примерно двукратное увеличение GMT для 2wp1 и 2wp2. Вариации различных липосом с различными диаметрами также используют для доставки репликонов, кодирующих полной длины RSV F белок. На ФИГ.15 представлен график зависимости полных титров IgG против F белка через две недели после первой дозы от диаметров липосом.
Способы получения липосом
Вообще для получения липосом в соответствии с настоящим изобретением были использованы восемь различных способов. В тексте их обозначают как способы (A) до (H) и они отличаются, главным образом, стадиями фильтрации и TFF стадиями. Далее приводятся подробности способов:
(A) Приготавливают свежие исходные липидные растворы в этаноле. Взвешивают 37 мг DlinDMA, 11,8 мг DSPC, 27,8 мг холестерина и 8,07 мг ПЭГ DMG 2000 и растворяют в 7,55 мл этанола. Свежеприготовленный липидный исходный раствор осторожно встряхивают при 37°C в течение около 15 мин для получения гомогенной смеси. Затем 755 мкл исходного раствора добавляют к 1,245 мл этанола для получения 2 мл исходного рабочего липидного раствора. Указанное количество липидов используют для создания липосом с 250 мкг РНК. Также приготавливают 2 мл рабочего раствора РНК из исходного раствора ~1 мкг/мкл в 100 мМ цитратном буфере (pH 6). Три стеклянные 20 мл ампулы (с магнитными якорями) промывают раствором РНКазы Away (Molecular BioProducts, San Diego, CA) и обильно промывают водой MilliQ перед тем, как используют для очистки ампул от РНКаз. Одну из ампул используют для рабочего раствора РНК и остальные для сбора смесей липидов и РНК (как раскрыто далее). Рабочие липидный и РНК растворы нагревают при 37°C в течение 10 мин перед тем, как помещают в 3 см3 шприцы с люэровским клапаном. 2 мл цитратного буфера (pH 6) помещают в другой 3 см3 шприц. Шприцы, содержащие РНК и липиды, соединяют в T смесителе (PEEK™ с 500 мкм внутренним диаметром соединения, Idex Health Science, Oak Harbor, WA), используя FEP трубки (фторированный этилен-пропилен; причем размер всех FEP трубок: 2 мм внутренний диаметр × 3 мм внешний диаметр, поставка от Idex Health Science). На выходе T смесителя также расположена FEP трубка. Третий шприц, содержащий цитратный буфер, соединен с отдельным отрезком FEP трубки. Скорость потока во всех шприцах поддерживается шприцевыми насосами на уровне 7 мл/мин. Выходы трубок располагают таким образом, чтобы собирать смеси в 20 мл стеклянную ампулу (при перемешивании). Магнитный якорь удаляют, и этанольно/водный раствор оставляют для уравновешивания до комнатной температуры в течение 1 часа. 4 мл полученной смеси помещают в 5 см3 шприц, который соединен с отрезком FEP трубки, и в другой 5 см3 шприц, соединенный с равной длины FEP трубкой, и загружают равные количества 100 мМ цитратного буфера (pH 6). Скорость потока через оба шприца, которую обеспечивают шприцевые насосы, составляет 7 мл/мин, и конечную смесь собирают в 20 мл стеклянную ампулу (при перемешивании). Затем смесь, собранную на второй стадии смешивания (липосомы), пропускают через мембрану Mustang Q (анионообменная подложка, которая связывает и удаляет анионные молекулы, от Pall Corporation, AnnArbor, MI, USA). Перед тем, как через Mustang мембрану пропускают липосомы, через нее последовательно пропускают 4 мл 1 M NaOH, 4 мл 1 M NaCl и 10 мл 100 мМ цитратного буфера (pH 6). Липосомы нагревают в течение 10 мин при 37°C перед тем, как их пропускают через мембрану. Затем липосомы концентрируют до объема 2 мл и осуществляют диализ против 10-15 объемов 1X PBS, используя TFF, перед выделением конечного продукта. Систему TFF и половолоконные фильтрационные мембраны закупают у Spectrum Labs и используют в соответствии с рекомендациями изготовителя. Используют полисульфонные половолоконные фильтрационные мембраны (№ по каталогу P/N: X1AB-100-20P) с отсечением по размеру пор 100 кДа и площадью поверхности 8 см2. Для in vitro и in vivo экспериментов композиции разбавляют до нужной концентрации РНК, используя 1X PBS.
(B) По способу (A) за исключением того, что после встряхивания 226,7 мкл исходного раствора добавляют к 1,773 мл этанола для получения 2 мл исходного липидного раствора, тем самым изменяя отношение липиды:РНК.
(C) По способу (B) за исключением того, что фильтрацию с использованием системы Mustang опускают, при этом липосомы поступают из 20 мл стеклянной ампулы в систему TFF для диализа.
(D) По способу (C) за исключением того, что в TFF используют полиэфирсульфонные (PES) половолоконные мембраны (номер по каталогу P-C1-100E-100-01N) с отсечением по размеру пор 100 кДа и площадью поверхности 20 см2.
(E) По способу (D) за исключением того, что используют мембрану Mustang, как в способе (A).
(F) По способу (A) за исключением того, что фильтрацию на системе Mustang опускают, при этом липосомы поступают из 20 мл стеклянной ампулы в систему TFF диализа.
(G) По способу (D) за исключением того, что 4 мл рабочего раствора РНК приготавливают из исходного раствора ~1 мкг/мкл в 100 мМ цитратном буфере (pH 6). Затем четыре 20 мл стеклянные ампулы подготавливают аналогичным образом. Две из них используют для рабочего раствора РНК (2 мл в каждой ампуле) и остальные используют для сбора смесей липидов и РНК, по способу (C). Вместо использования T миксера, шприцы, содержащие РНК и липиды, соединяют с чипом Mitos Droplet junction Chip (стеклянное микрожидкостное устройство, полученное от Syrris, номер по каталогу 3000158), используя PTFE трубки (внутренний диаметр 0,076 см × 0,16 см внешний диаметр), используя 4-сторонний краевой разъем (Syrris). Два потока РНК и один липидный поток перемещаются с помощью шприцевых насосов, и смешивание этанольной и водной фаз происходит в X соединении (100 мкм × 105 мкм) чипа. Скорости всех трех потоков поддерживают на 1,5 мл/мин, поэтому отношение количества полной водной фазы к этанольной фазе составляет 2:1. Выход трубки расположен таким образом, чтобы собирать смеси в 20 мл стеклянную ампулу (при перемешивании). Магнитный якорь удаляют и этанольно/водный раствор оставляют для уравновешивания до комнатной температуры в течение 1 часа. Затем смесь помещают в 5 см3 шприц, который соединен с другим отрезком FEP трубки; и в другой 5 см3 шприц, соединенный с равной длины FEP трубкой, загружают равное количество 100 мМ цитратного буфера (pH 6). Скорость потока через оба шприца, которую обеспечивают шприцевые насосы, составляет 3 мл/мин и конечную смесь собирают в 20 мл стеклянную ампулу (при перемешивании). Затем липосомы концентрируют до 2 мл и осуществляют диализ против 10-15 объемов 1X PBS, используя TFF, по способу (D).
(H) По способу (A), за исключением того, что 2 мл рабочего исходного липидного раствора приготавливают, перемешивая 120,9 мкл липидного исходного раствора с 1,879 мл этанола. Также после смешивания в T миксере липосомы из 20 мл ампулы помещают в кассету для диализа Pierce Slide-A-Lyzer Dialysis Cassette (Thermo Scientific, extra strength, 0,5-3 мл емкости) и осуществляют диализ против 400-500 мл 1X PBS в течение ночи при 4°C в подвергнутом обработке в автоклаве пластиковом контейнере перед выделением конечного продукта.
RSV иммуногенность
vA317 самореплицирующийся репликон, кодирующий белок RSV F, вводят мышам штамма BALB/c, по 4 или 8 животных в группе, путем бактериальных внутримышечных вакцинаций (50 мкл в лапу) в дни 0 и 21 только с репликоном (1 мкг), или в виде липосом с DlinDMA ("RV01"), или DOTAP ("RV13"), или с липидами, как представлено на ФИГ.16A-16M ("RV05"). RV01 липосомы содержат 40% DlinDMA, 10% DSPC, 48% холестерина и 2% ПЭГ-DMG, но различные количества РНК. RV05 липосомы содержат или 40% RV05, 10% DSPC, 48% холестерина и 2% ПЭГ-DMG или 60% RV05, 38% холестерина и 2% ПЭГ-DMG. RV13 липосомы содержат 40% DOTAP, 10% DOPE, 48% холестерина и 2% ПЭГ-DMG. Для сравнения оголенные плазмидные ДНК (20 мкг), экспрессирующие тот же самый RSV-F антиген, вводят или используя электропорацию, или с RV01(10) липосомами (0,1 мкг ДНК). Четырех мышей используют в качестве интактной контрольной группы.
Липосомы приготавливают по способу (A) или по способу (B). Для некоторых липосом, полученных по способу (A), используют двойное или половинное количество РНК. Средний диаметр частиц Z и коэффициент полидисперсности составляют:
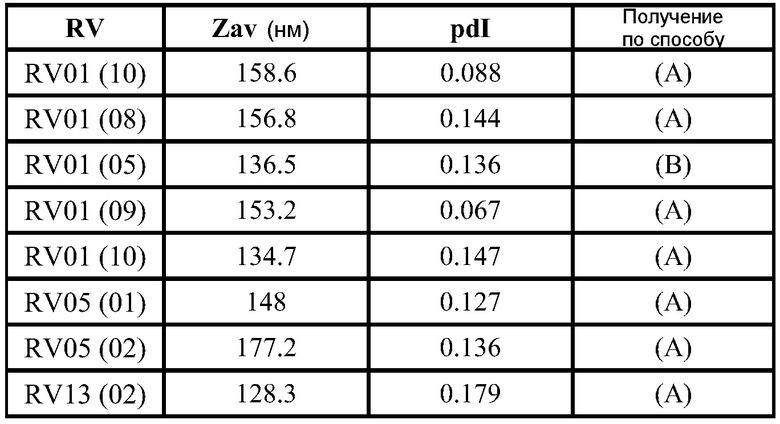
Сыворотку собирают для анализа антител в дни 14, 36 и 49. Селезенки извлекают в день 49 для анализа T клеток.
Титры сывороточных F-специфических IgG (GMT) следующие:
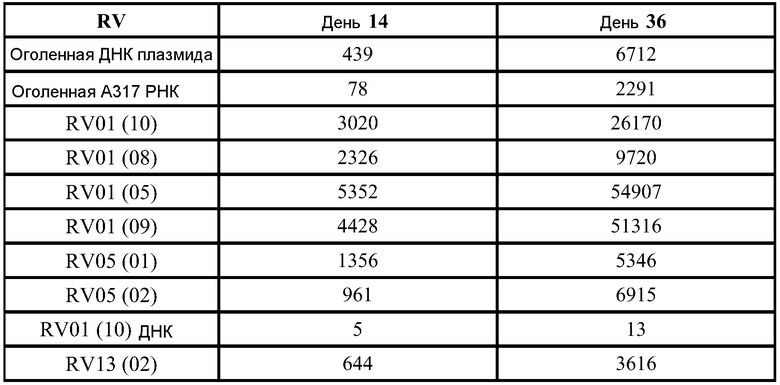
Количество цитокин-положительных Т клеток и специфических в отношении пептида RSV F51-66 оказались следующими, приведены только те значения, которые статистически значимы выше 0:
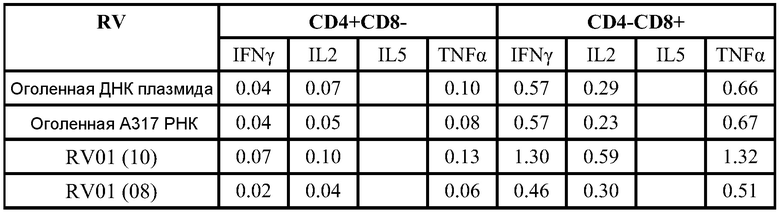
Таким образом, липосомальные композиции значительно повышают иммуногенность в отношении оголенных РНК контролей, что определено по увеличению титров F-специфических IgG и количеству T клеток. Композиции плазмидных ДНК с липосомами, или оголенные, введенные с использованием электропорации, оказываются значительно менее иммуногенными, чем самореплицирующиеся РНК в композиции с липосомами.
Далее RV01 липосомы приготавливают по способу (H), используя или короткий (2 кДа) или длинный (5 кДа) ПЭГ, конъюгированный с DMG, и инкапсулируя 150 мкг РНК (vA375 репликон, кодирующий поверхностный фьюжн гликопротеин RSV), или инкапсулируя только буфер. Указанные липосомы содержат 40% DlinDMA, 10% DSPC, 48% Chol, и 2% ПЭГ-DMG. Размеры и инкапсулирование были следующими:
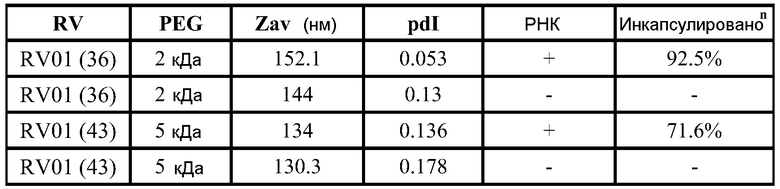
Указанные липосомы вводят BALB/c мышам (по 10 в группе) путем бактериальных внутримышечных инъекций (50 мкл в лапу) в дни 0 и 21. Дозы составляют 0,01, 0,03, 0,1, 0,3 или 1 мкг. Титры сывороточных F-специфических IgG и титры PRNT60 (GMT) через 2 недели после первой или второй инъекции были следующими:
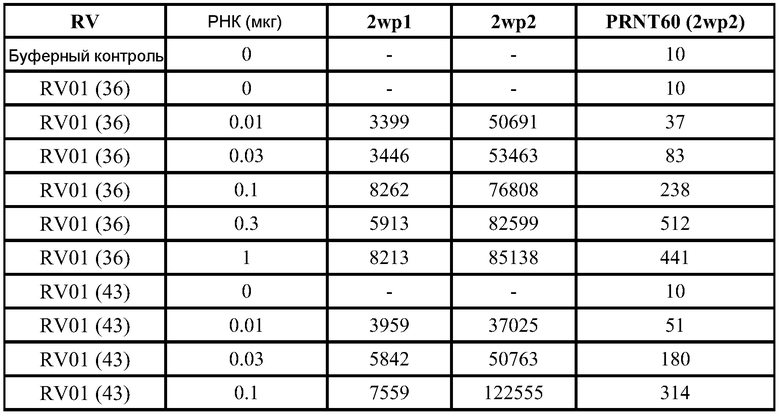

Липосомы - требования для инкапсулирования
Как было указано выше со ссылкой на ФИГ.10, способ инкапсулирования является существенным для эффективной экспрессии. В дальнейших экспериментах используют три различные РНК: (i) ‘vΑ317’ репликон, который экспрессирует RSV-F т.е. поверхностный фьюжн гикопротеин RSV; (ii) ‘vΑ17’ репликон, который экспрессирует GFP; и (iii) ‘vΑ336’, который является репликационно-дефектным и кодирует GFP. РНК вводят или оголенными, или в липосомах, полученных по способу (D). Пустые липосомы получают по способу (D), но совсем без каких-либо РНК. Липосомные композиции имеют следующие характеристики:
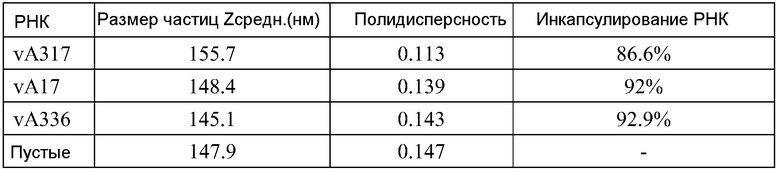
BALB/c мышам, по 5 животных в группе, проводят бактериальную внутримышечную вакцинацию (50 мкл в лапу) в дни 0 и 21, используя:
Группа 1 оголенные самореплицирующиеся RSV-F РНК (vA317, 0,1 мкг)
Группа 2 самореплицирующиеся RSV-F РНК (vA317, 0,1 мкг), инкапсулированные в липосомы
Группа 3 самореплицирующиеся RSV-F РНК (vA317, 0,1 мкг), добавленные к пустым липосомам
Группа 4 F субъединичный белок (5 мкг)
Сыворотку собирают для анализа антител в дни 14, 35 и 51. Измеряют титры F-специфических сывороточных IgG (GMT); если у отдельных животных титр оказывается меньше <25 (предел детектирования), его оценивают как титр 5. Кроме того в день 51 извлекают селезенки для анализа Т клеток, для определения клеток, которые оказываются цитокин-позитивными и специфическими в отношении RSV F51-66 пептида (CD4+) или в отношении RSV пептидов F85-93 и F249-258 (CD8+).
Титры IgG для 10 групп и для неиммунизированных контрольных мышей следующие:
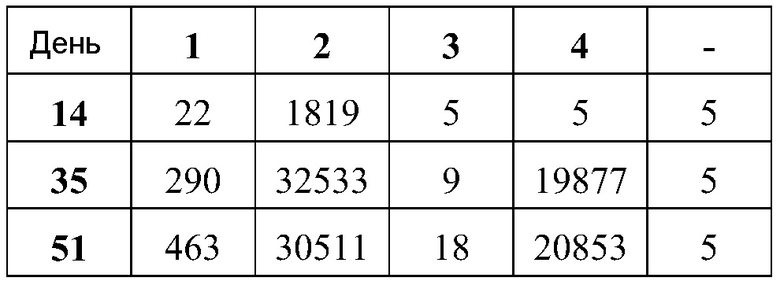
Титры сывороточных нейтрализующих RSV антител в день 51 были следующими:
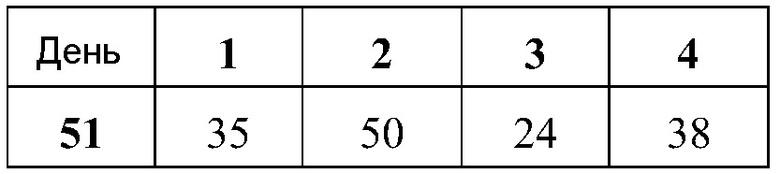
Для животных, демонстрирующих RSV F-специфические CD4+ селезеночные T клетки в день 51, были следующими, где число (% позитивных клеток) приводится, только если статистическая значимость стимулированной реакции оказывается больше нуля:
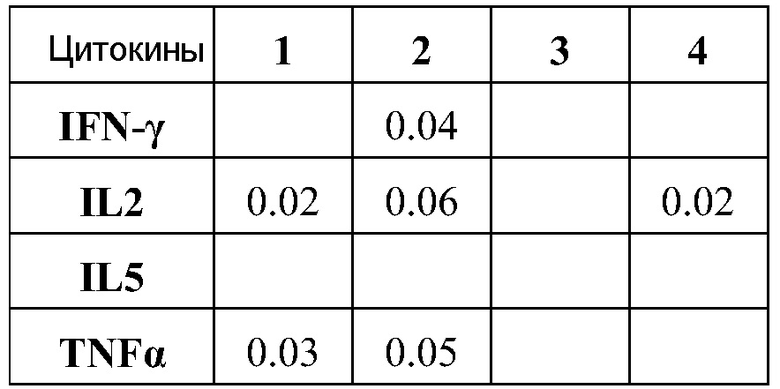
Для животных, демонстрирующих RSV F-специфические CD8+ селезеночные T клетки в день 51, были следующими, где число (% позитивных клеток) приводится, только если статистическая значимость стимулированной реакции оказывается больше нуля:
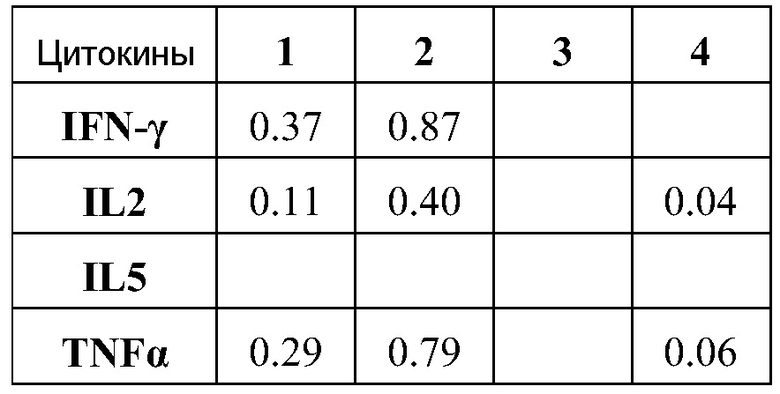
Таким образом, инкапсулирование РНК внутри липосом является необходимым условием для высокой степени иммуногенности, в то время как простая смесь РНК и указанных липосом (группа 3) оказалась неиммуногенной (в действительности, менее иммуногенной, чем оголенные РНК).
Различные катионные липиды с vA317 RSV репликоном
В дальнейших экспериментах проводят сравнение четырех различных катионных липидов (DlinDMA, RV02, RV04 и RV07). Все липосомы содержат 2% ПЭГ-DMG 2000, но остальная часть липидной композиции варьируется. Состав и физические характеристики приводятся далее:
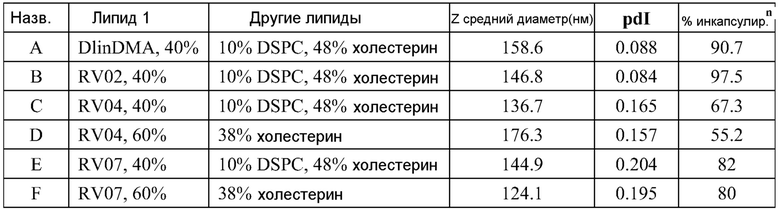
BALB/c мышам, по 8 в группе, проводят бактериальную внутримышечную вакцинацию (50 мкл в лапу) в дни 0 и 21, используя оголенный репликон (1 мкг) или 0,1 мкг инкапсулированной РНК. Титры сывороточных F-специфических IgG (GMT) через 2 недели после двух указанных инъекций оказались следующими:
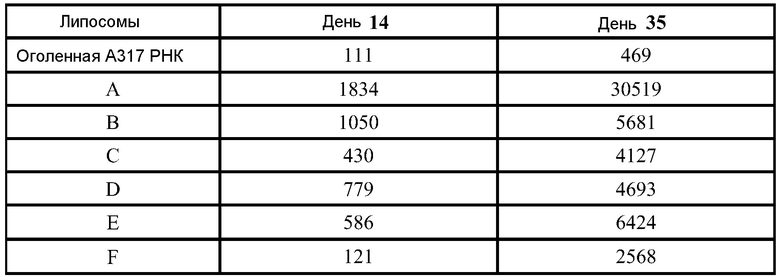
Для RV07 отсутствие DSPC вызывает значительное снижение иммуногенности.
Аналогичным образом тестируют другие липиды (RV03, RV08, RV09, RV14 [42]):
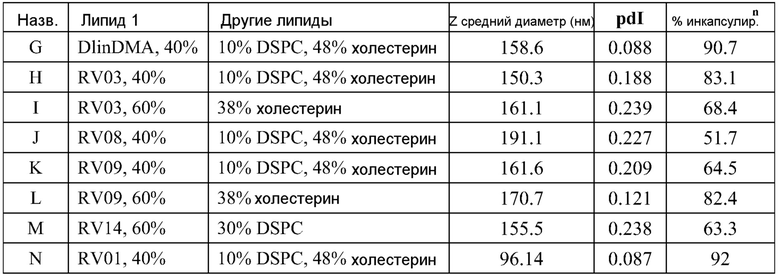
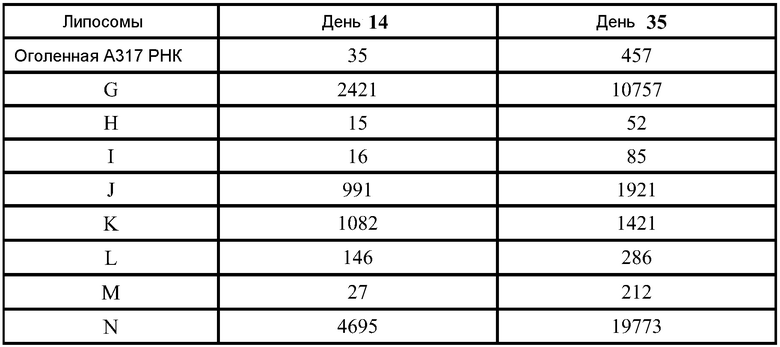
Результаты для липосомы M (с DC-холестерином) оказались неудовлетворительными, даже хуже, чем для контрольной оголенной РНК. Напротив, для остальных катионных липидов результаты оказались удовлетворительными. Липосомы способа N были получены способом (способ (G) с микрожидкостным чипом), который отличался от способа получения липосом G (способ (D)) и такие более маленькие липосомы обеспечили лучшие результаты с примерно таким же инкапсулированием.
Другие липиды (RV01, RV10, RV11, RV15) тестируют аналогичным образом:
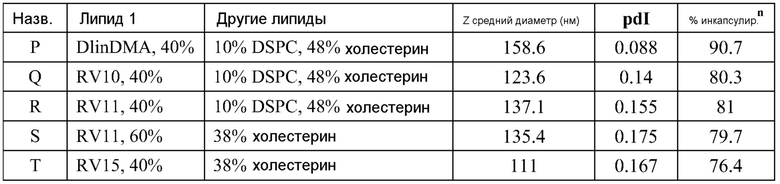
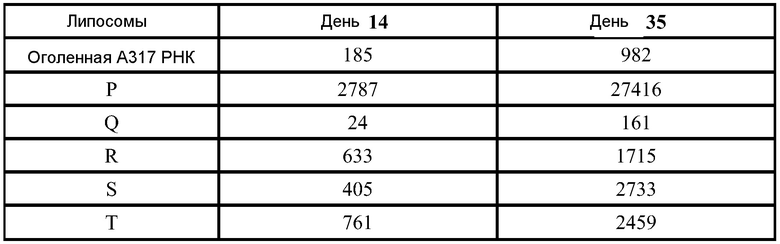
За исключением липосом способа Q, результаты для каждой из указанных липосом были лучше, чем для контроля. RV10 липид в липосомах Q имеет значение pKa 7,86, что по-видимому слишком велико, чтобы их можно было использовать in vivo. Однако даже внутри подходящего интервала значений pKa 5,0-7,6, хотя результаты были неплохими, ни один из липидов с одним алкильным концевым сегментом и одним стероид-содержащим концевым сегментом не обеспечил столь же хорошие результаты как RV01.
Другие липосомы получают с RV05. Все указанные липосомы содержат 40% RV05 и 2% пегилированного липида, но остальные компоненты варьируют (хотя во всех случаях холестерин включают). Далее приводятся физические характеристики:
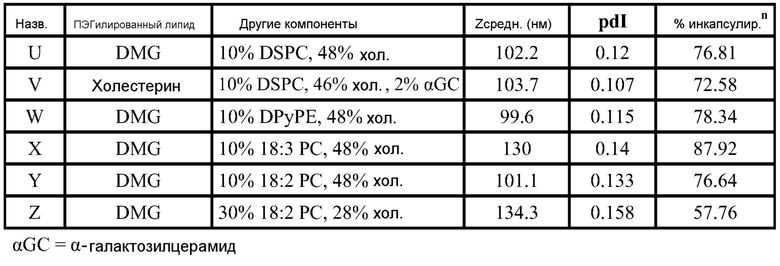
BALB/c мышей тестируют как и раньше:
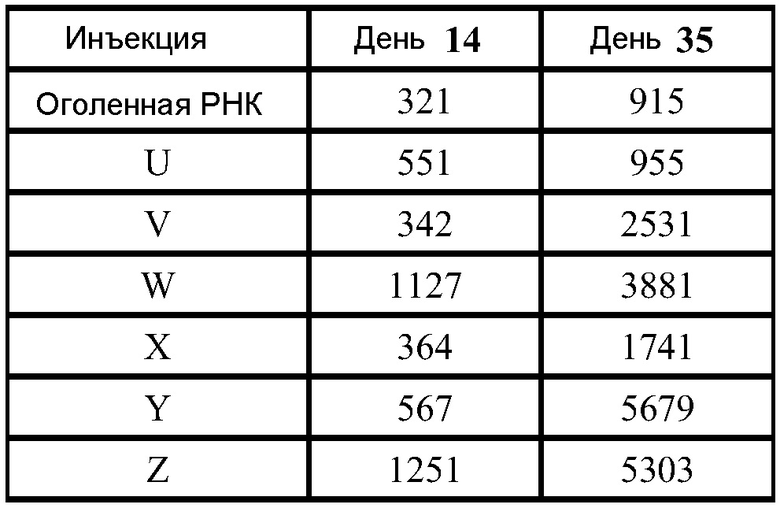
Для катионных липидов с асимметричными концевыми сегментами (алкил + холестерин), замена нейтрального липида из DSPC (насыщенный C18 липидный сегмент) на 18:2 или 18:3 PC (с 2 и 3 ненасыщенными двойными связями в концевом сегменте) увеличивает титры полного IgG. Сравнимые результаты получают при замене DSPC на DPyPE.
Другие различные катионные липиды с vA317 RSV репликоном
Катионные липиды, раскрытые в ссылке 43, также используют для получения липосом для vA317 репликона. Указанные катионные липиды имеют значение pKa между 5,8 и 6,1. Для сравнения также тестируют DODMA, DlinDMA и DOTMA. Катионный липид всегда присутствует в количестве 40%. Все липосомы содержат холестерин и 2% пегилированный DMG (ПЭГ2000, за исключением липосом E, которые содержат ПЭГ5000), и их получают по способу (H). Физические характеристики следующие:
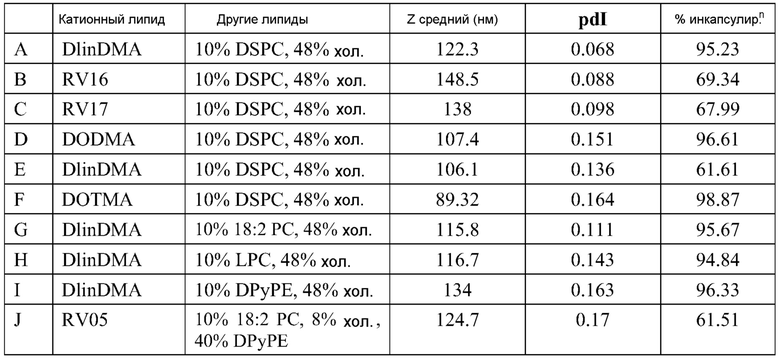
Указанные липосомы используют для вакцинации BALB/c мышей, также как раньше. Титры сывороточных F-специфических IgG (GMT) были следующими:
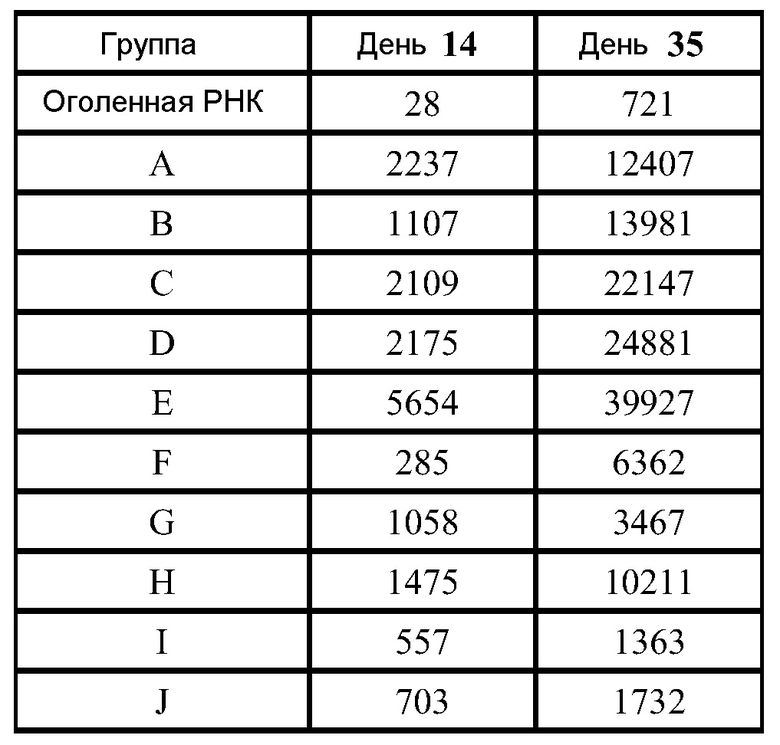
Указанные RV05 липосомы оказались более иммуногенными, чем оголенные РНК, но менее иммуногенными, чем RV01 липосомы. Селезенку вырезают на 49 день для анализа Т клеток. Все липосомы дают F-специфические цитокин-позитивные T клеточные частоты (CD4+ и CD8+), статистическая значимость которых оказывается выше нуля.
Различные липиды и различные длины ПЭГ
vA317 репликоны вводят в липосомы с различными липидами с различными длинами ПЭГ. Все указанные липосомы содержат 40% DlinDMA, 10% DSPC и 48% холестерина, но остальные 2% варьируют, используя различные пегилированные липиды (например, ФИГ.18A-18E) и различной длины ПЭГ.
Физические характеристики липосом, полученных по способу (H), следующие:
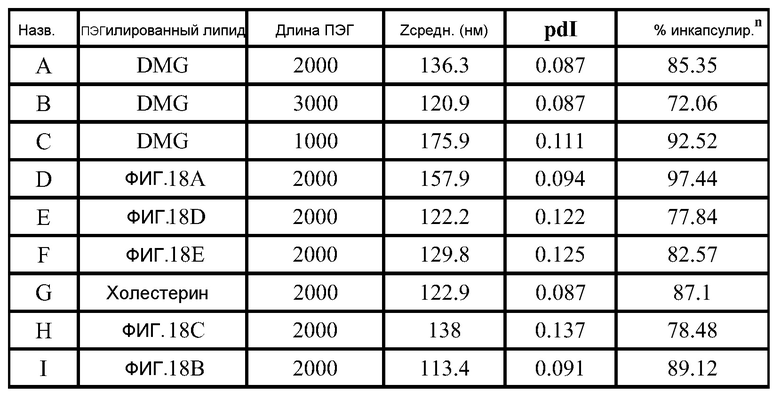
BALB/c мышам, по 8 в группе, проводят внутримышечные вакцинации (50 мкл в лапу) в дни 0 и 21 репликонами, или оголенными (1 мкг) или инкапсулированными (0,1 мкг). Сыворотку собирают для анализа антител в дни 14 и 35.
Титры сывороточных F-специфических IgG (GMT) через 2 недели после двух инъекций (2wp1) были следующими:
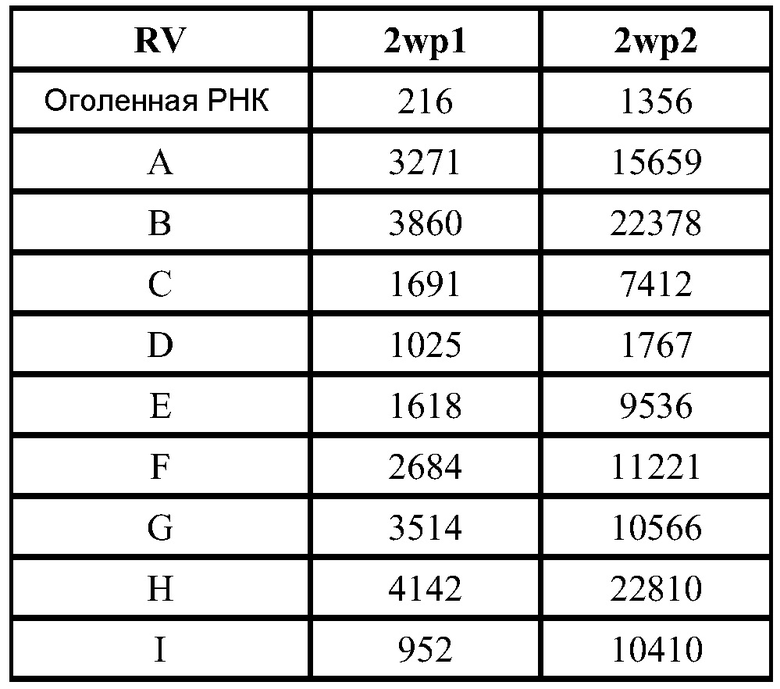
Указанные результаты демонстрируют тенденцию, свидетельствующую о том, что ПЭГ концевые группы с более высоким молекулярным весом более иммуногены. По мере увеличения длины DMG-конъюгированного ПЭГ (увеличение со 1000 Да до 3000 Да) титры 2wp2 F-специфических IgG увеличиваются с 7412 до 15659 до 22378.
Изменение линкерного участка со сложноэфирного до простого эфира практически не влияют на титры. Также, притом же самом молекулярном весе концевой группы (2000) наблюдается тенденция того, что увеличение длины липидных концов снижает титры (H с C14 диалкилом против I с C18 диалкилом). Замена ПЭГ диалкильного липидного концевого сегмента холестерином оказывает мало влияния на иммуногенность (A с DMG против G с холестерином).
Аналогичные эксперименты осуществляют с различными липидами, в которых 2 кДа ПЭГ расщеплен на 2× 1 кДа группы (ФИГ.17). Снова используют vA317 репликон, BALB/c мышами, по 8 в группе, проводят бактериальную вакцинацию (50 мкл в лапу) в дни 0 и 21, используя 1 мкг оголенных РНК или 0,1 мкг инкапсулированных в липосомы РНК. Все липосомы содержат 40% катионный липид (DlinDMA), 10% DSPC и 48% холестерина, но остальные 2% варьируют, используя различные пегилированные липиды (но все содержат 2 кДа ПЭГ). Их получают по способу (H).
Физические характеристики липосом оказываются:
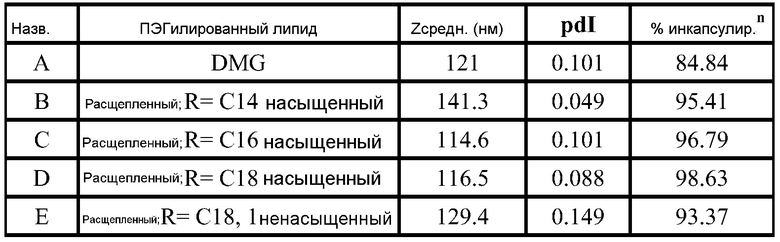
Далее липосомы получают, используя RV05. Все липосомы содержат 40% катионного липида (RV05) и 2% пегилированного липида (2 кДа ПЭГ), но остальные компоненты варьируют (хотя холестерин включают всегда). Указанные липосомы получают по способу (H), но при pH 5. Физические характеристики следующие:
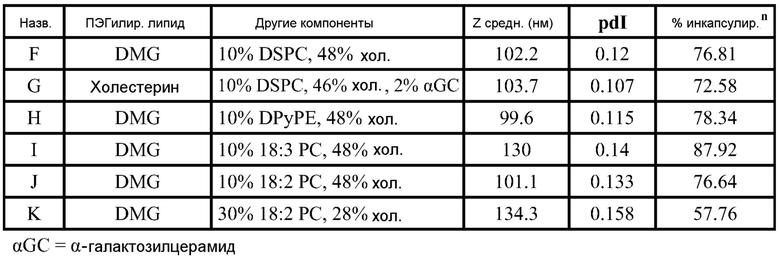
BALB/c мышам, по 8 в группе, проводят бактериальную внутримышечную вакцинацию (50 мкл в лапу) в дни 0 и 21, используя репликон, или оголенный (1 мкг) или инкапсулированный (0,1 мкг). Сыворотку собирают для анализа антител в дни 14 и 35. Титры сывороточных F-специфических IgG (GMT) через 2 недели после двух инъекций оказались следующими (2wp1):
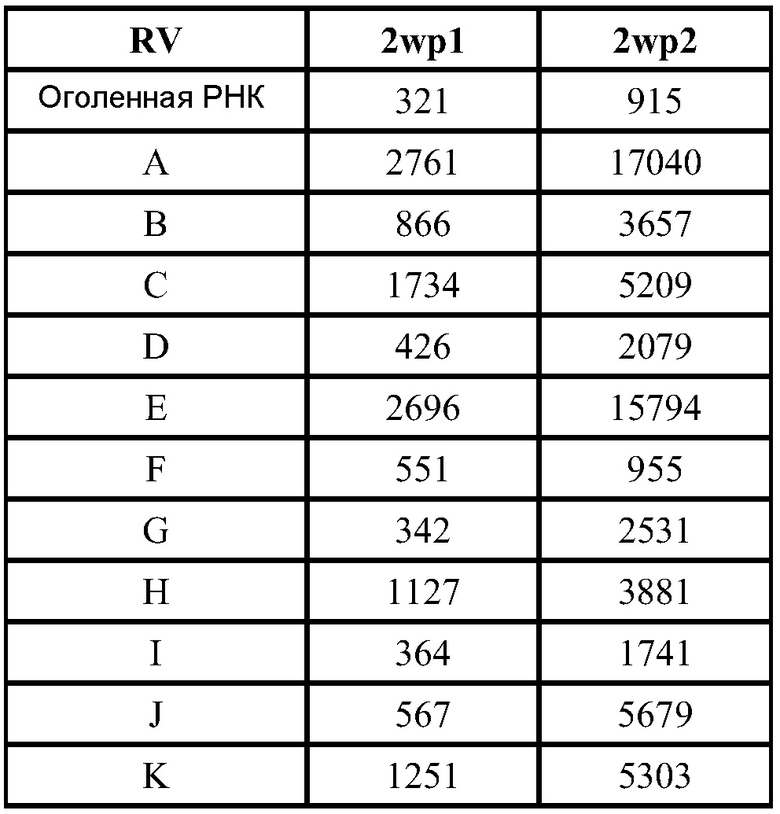
Таким образом, расщепление концевых сегментов ПЭГ понижает титры in vivo. Включение двойной связи (1 степень ненасыщенности на алкильный концевой сегмент) в ПЭГ липидные концевые сегменты повышает титры IgG в 6 раз в день 14 и в 7 раз в день 35. Для катионных липидов с асимметрическими липидными концевыми сегментами (алкил + холестерин), изменение нейтрального липида с DSPC (насыщенный, C18 липидная концевая группа) на 18:2 или 18:3 PC (с 2 и 3 ненасыщенными двойными связями на концевой сегмент) приводит к повышению титров полного IgG. Сопоставимые результаты наблюдают при замене DSPC на DPyPE.
ЦМВ иммуногенность
RV01 липосомы с DLinDMA в качестве катионного липида используют для доставки РНК репликонов, кодирующих цитомегаловирусные (ЦМВ) гликопротеины. Репликон "vA160" кодирует полной длины гликопротеины H и L (gH/gL), тогда как репликон "vA322" кодирует растворимую форму (gHраств./gL). Указанные два белка находятся под контролем отдельных субгеномных промоторов в одном репликоне; совместное введение двух отдельных векторов, один из которых кодирует gH и один кодирует gL, не дает хороших результатов.
BALB/c мышам, по 10 в группе, проводят бактериальную внутримышечную вакцинацию (50 мкл в лапу) в дни 0, 21 и 42, используя VRP, экспрессирующие gH/gL (1×106 ИмЕд (IU иммунизирующие единицы)), VRP, экспрессирующие gHраств./gL (1×106 ИмЕд) и PBS в качестве контролей. Двум тестируемым группам вводят 1 мкг vA160 или vA322 репликонов, заключенных в липосомы (40% DlinDMA, 10% DSPC, 48% холестерина, 2% ПЭГ-DMG; полученные по способу (D), но содержащие порции 150 мкг РНК).
Zсредн. диаметр vA160 липосом составляет 168,8 нм, pdI равно 0,144, и процент инкапсулирования 87,4%. Zсредн. диаметр vA322 липосомы составляет 162 нм, pdI равно 0,131, и процент инкапсулирования 90%.
Указанные репликоны способны экспрессировать два белка из одного вектора.
Сыворотку для иммунологических анализов собирают в день 63 (3wp3). Титры антител, нейтрализующих ЦМВ (обратная величина от разбавлений сыворотки, обеспечивающих 50% снижение количества позитивных вирусных очагов на лунку, относительно контроля), имеют следующие значения:

РНК, экспрессирующие или полной длины или растворимую форму комплекса ЦМВ gH/gL, вызывают высокие титры нейтрализующих антител, по оценкам, проведенным для эпителиальных клеток. Средние титры, вызванные инкапсулированными в липосомах РНК, оказываются по меньшей мере столь же высокими, что и соответствующие значения для VRP.
Повторные эксперименты подтверждают тот факт, что репликон способен экспрессировать два белка из одного вектора. РНК репликон приводит к значению титра 3wp3 11457, по сравнению с 5516 для VRP.
Следует принять во внимание, что настоящее изобретение было раскрыто с помощью только одного примера, и модификации можно осуществить, при этом оставаясь в рамках объема и сути настоящего изобретения.
подходящие фосфолипиды
Ссылки


| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕГИЛИРОВАННЫЕ ЛИПОСОМЫ ДЛЯ ДОСТАВКИ КОДИРУЮЩЕЙ ИММУНОГЕН РНК | 2012 |
|
RU2628705C2 |
| ЛИПОСОМЫ С ЛИПИДАМИ, ИМЕЮЩИМИ ПРЕИМУЩЕСТВЕННОЕ ЗНАЧЕНИЕ РКА, ДЛЯ ДОСТАВКИ РНК | 2011 |
|
RU2589503C2 |
| ЛИПИДЫ, ПОДХОДЯЩИЕ ДЛЯ ЛИПОСОМНОЙ ДОСТАВКИ КОДИРУЮЩЕЙ БЕЛОК РНК | 2011 |
|
RU2577983C2 |
| КАТИОННЫЕ ЭМУЛЬСИИ "МАСЛО-В-ВОДЕ" | 2011 |
|
RU2625546C2 |
| ПЛАТФОРМЫ ДОСТАВКИ АНТИГЕНОВ | 2011 |
|
RU2597974C2 |
| КОМПОЗИЦИИ БЕЛКА F RSV И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2010 |
|
RU2585227C2 |
| КАТИОННЫЕ ЭМУЛЬСИИ МАСЛО-В-ВОДЕ | 2012 |
|
RU2649133C2 |
| ЭМУЛЬСИИ ТИПА "МАСЛО В ВОДЕ", КОТОРЫЕ СОДЕРЖАТ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2012 |
|
RU2606846C2 |
| ПЕГИЛИРОВАННЫЕ ЛИПОСОМЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2796539C2 |
| РНК-ЧАСТИЦЫ, ВКЛЮЧАЮЩИЕ ПОЛИСАРКОЗИН | 2019 |
|
RU2792644C2 |
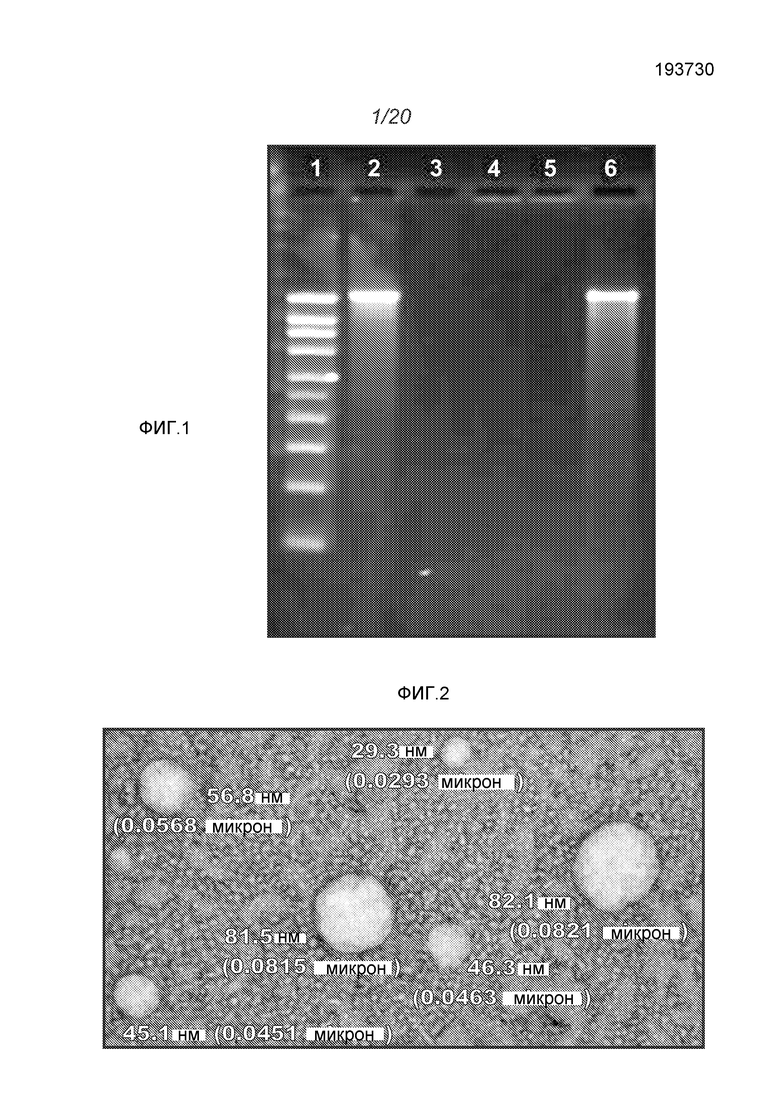
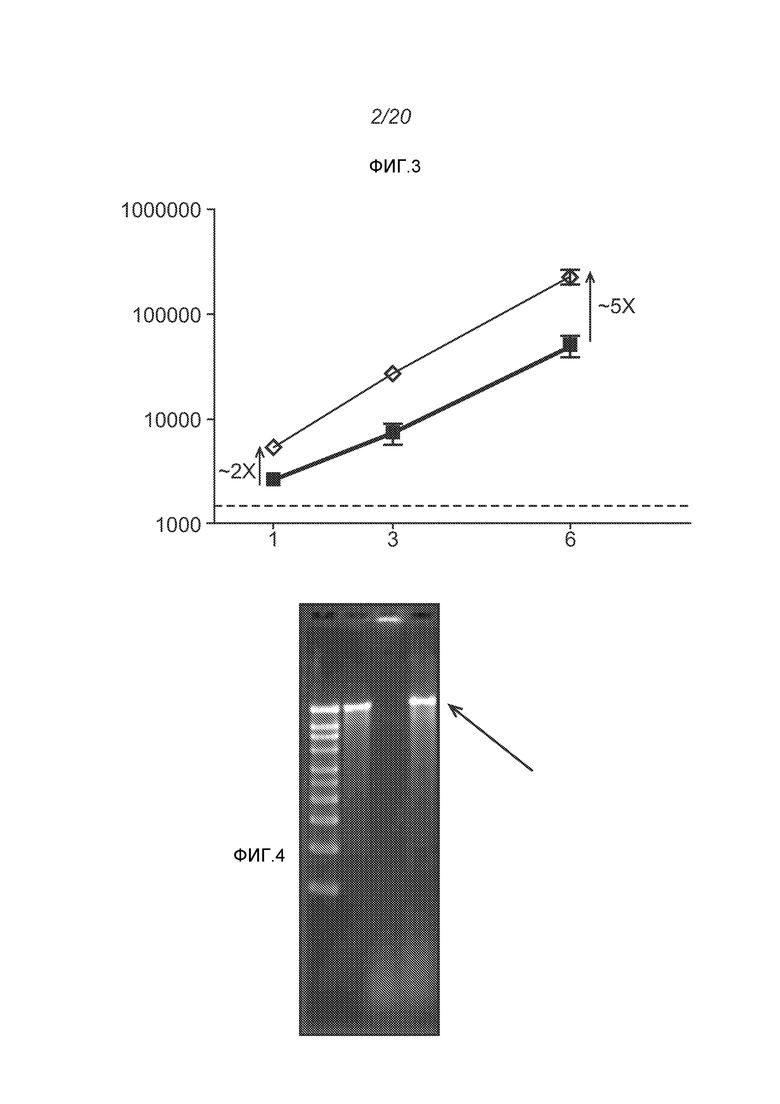
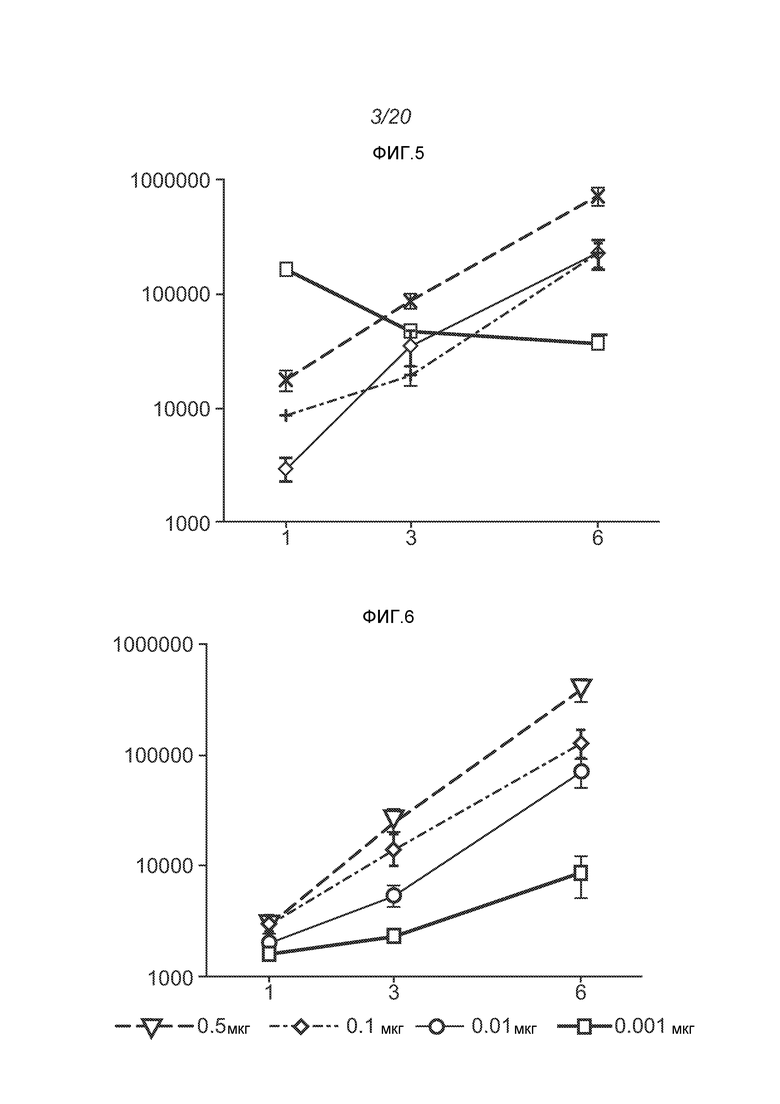
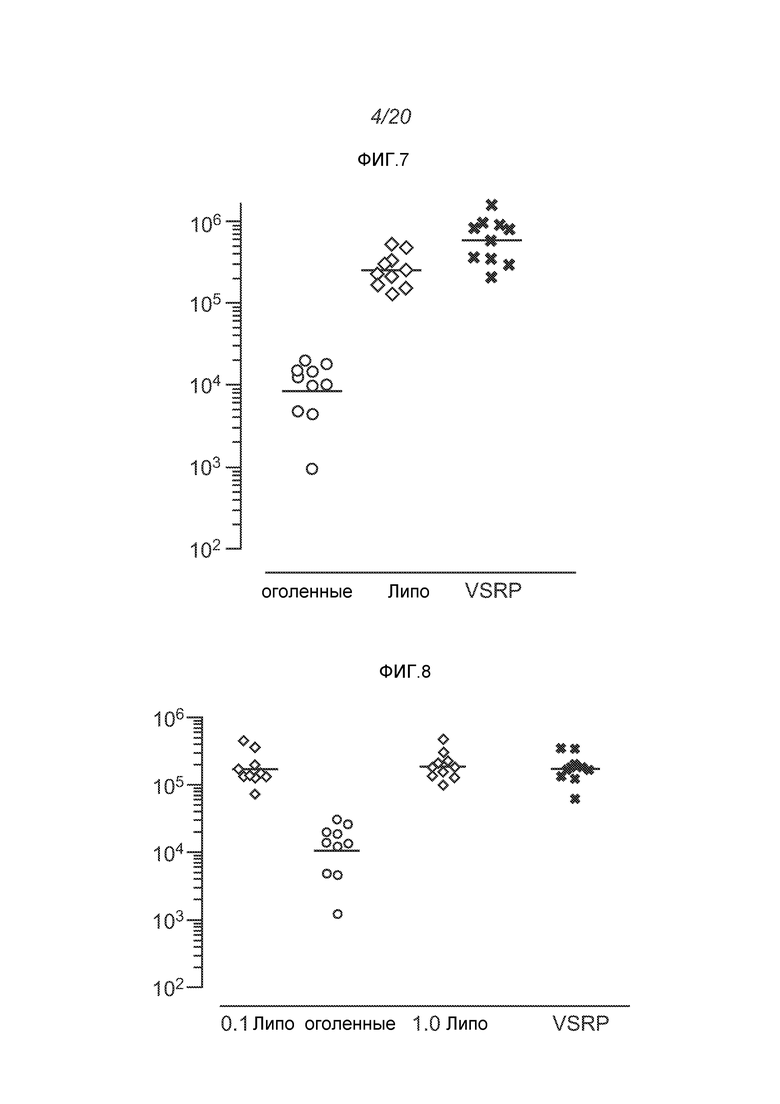
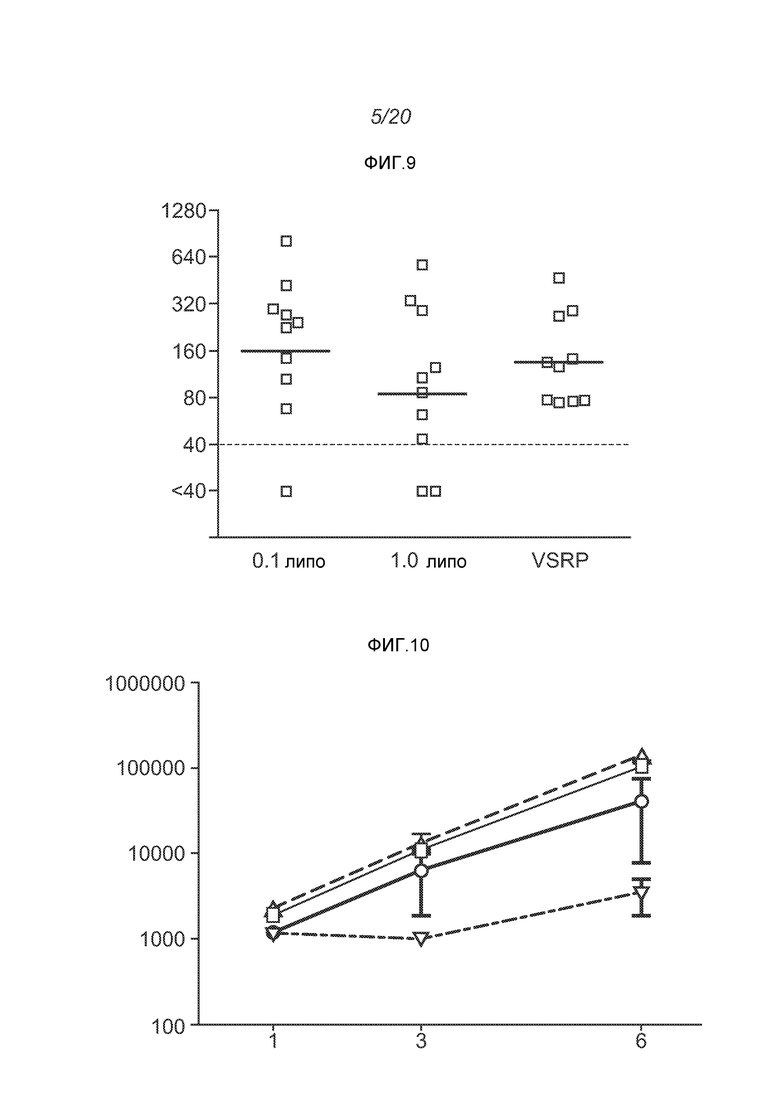
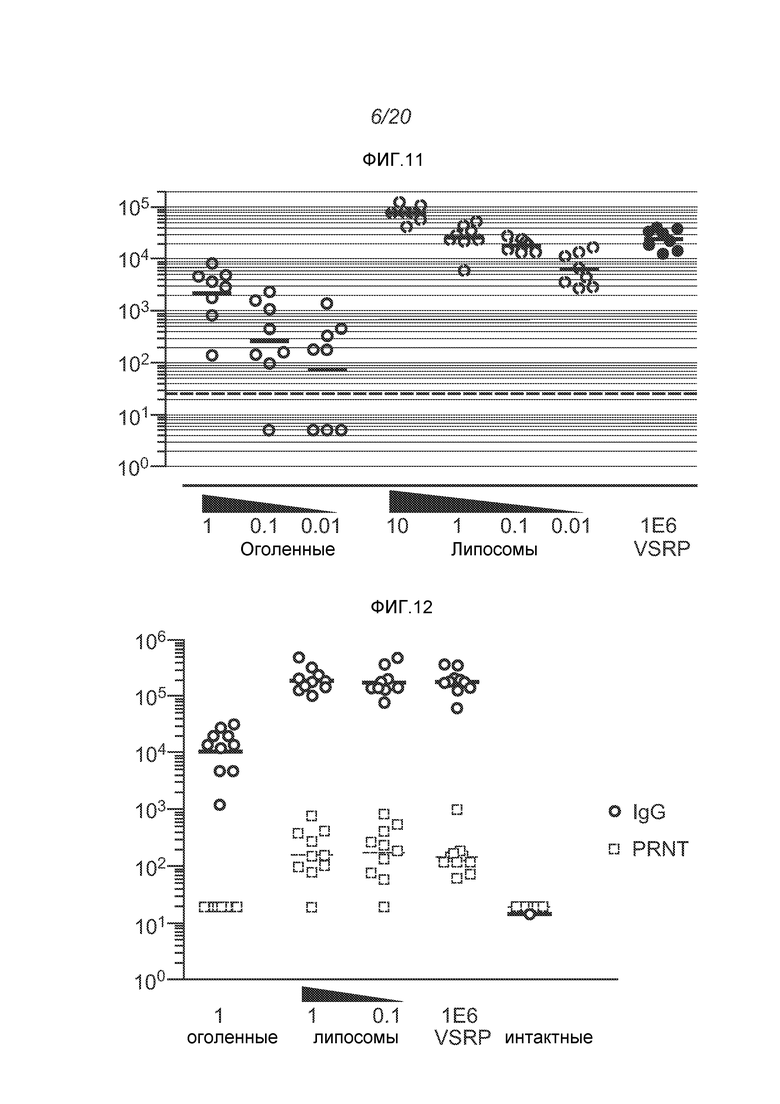
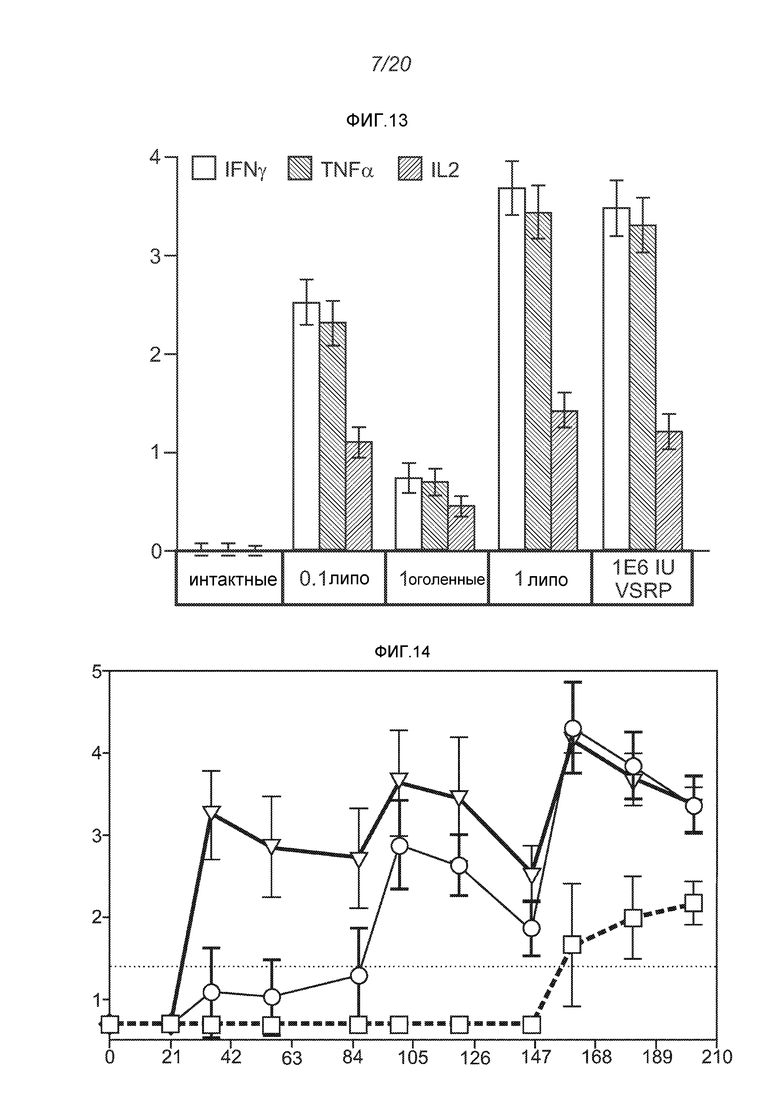
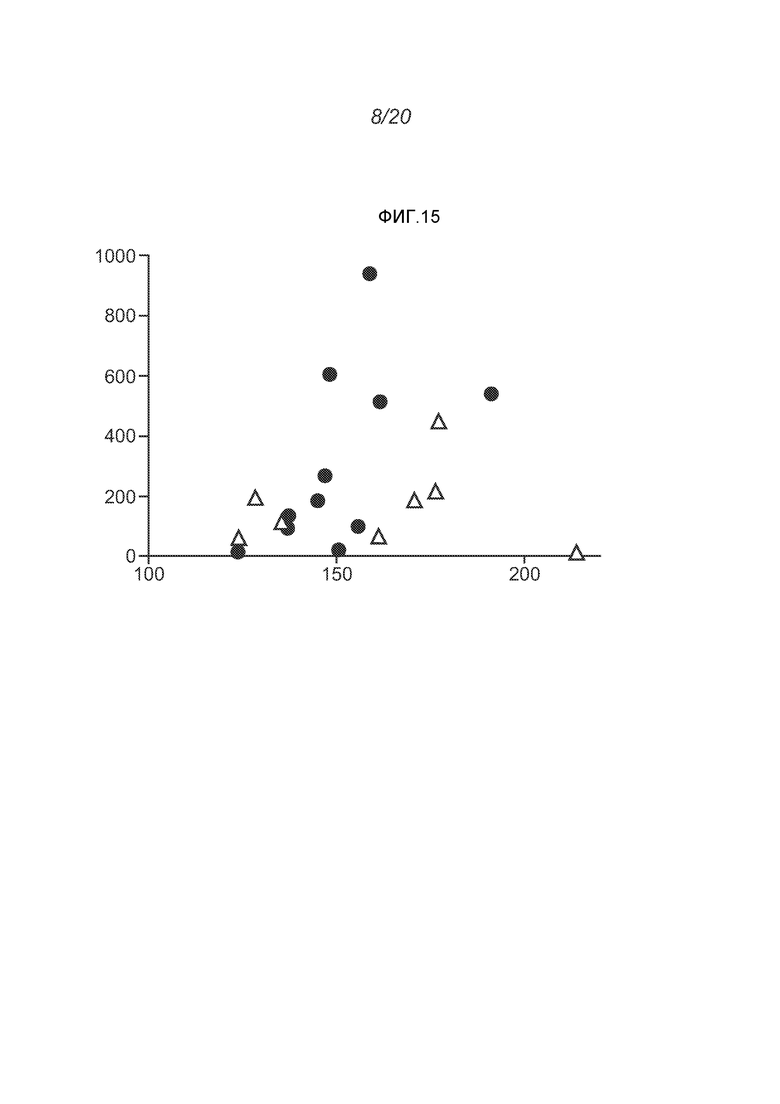
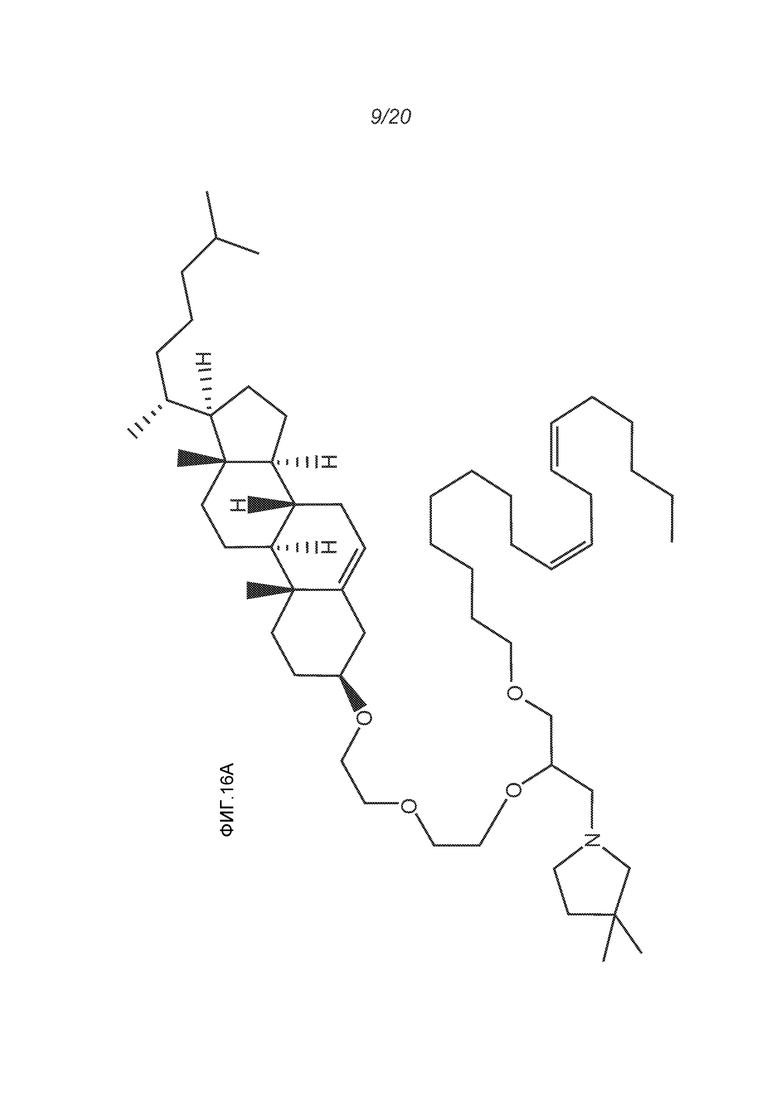
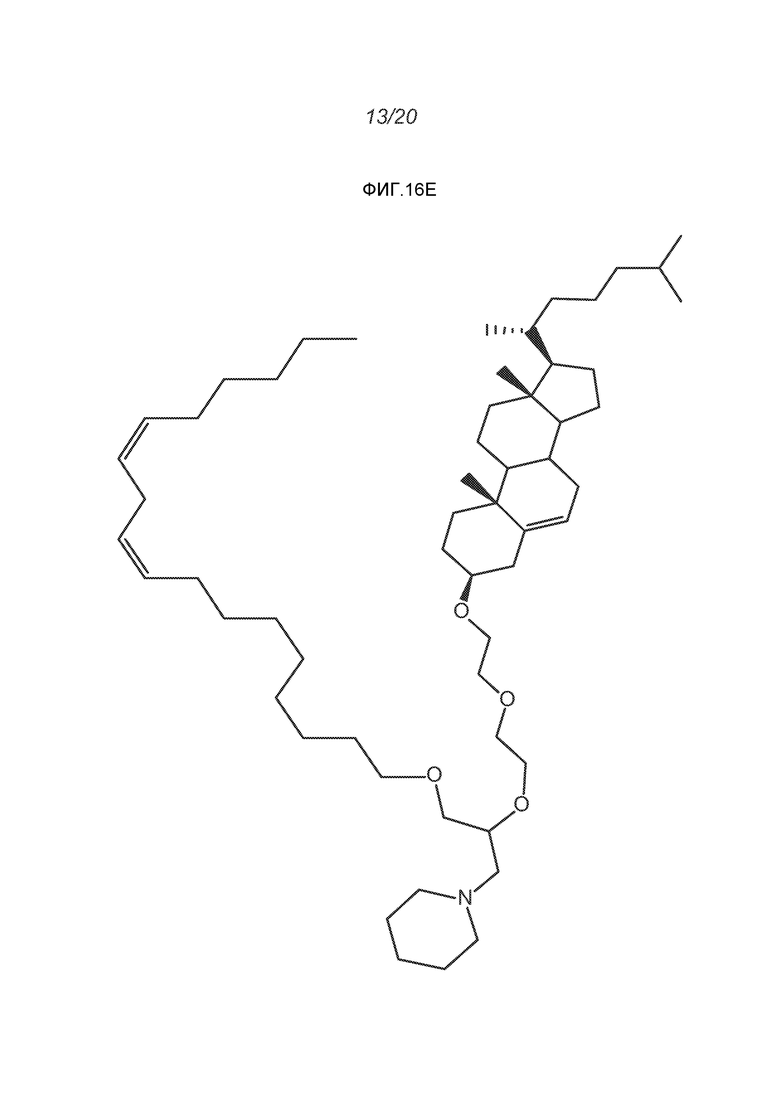
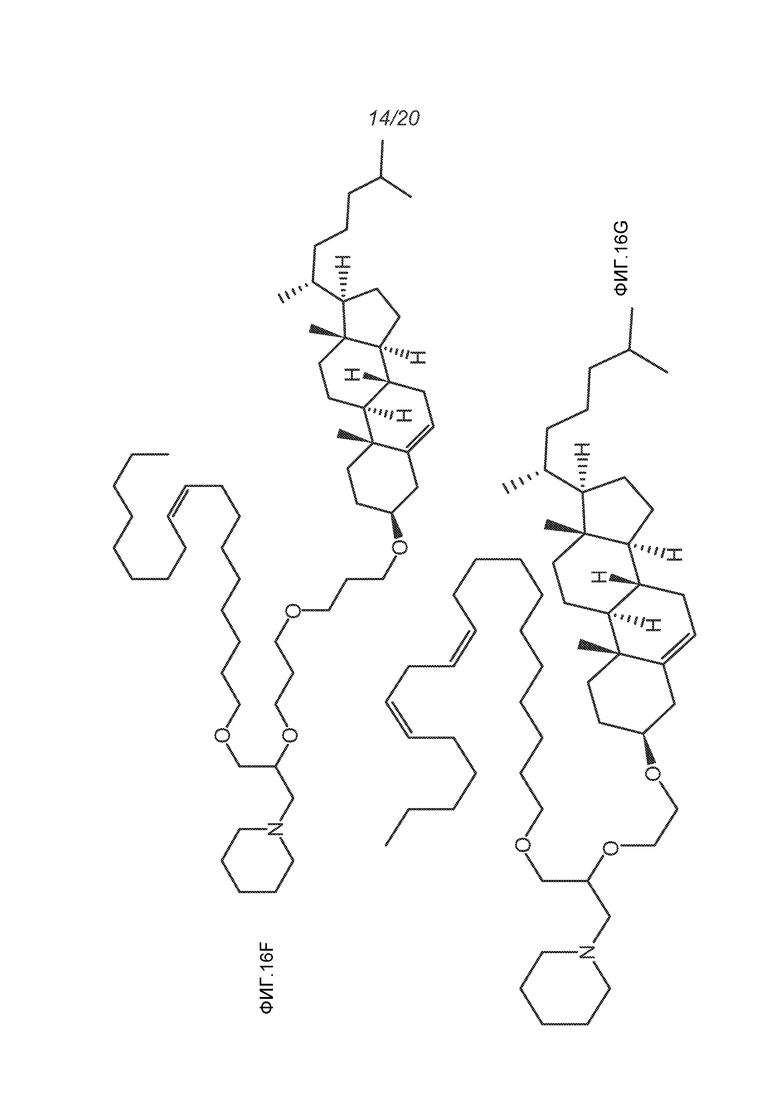
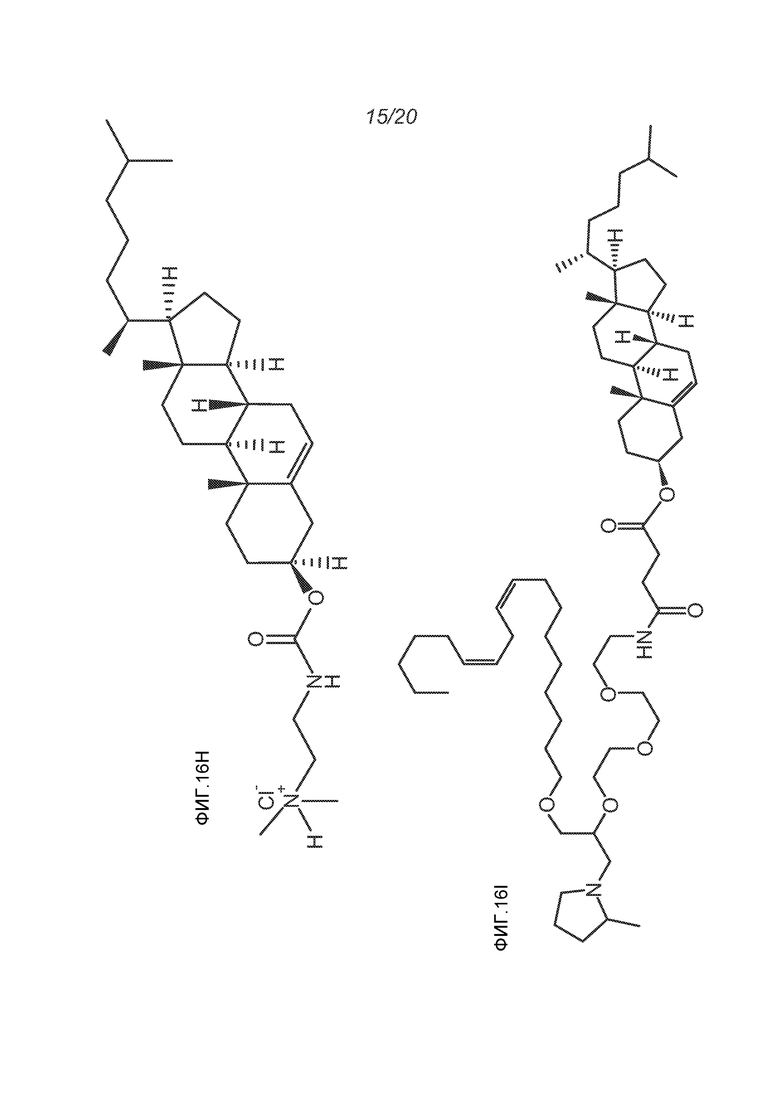
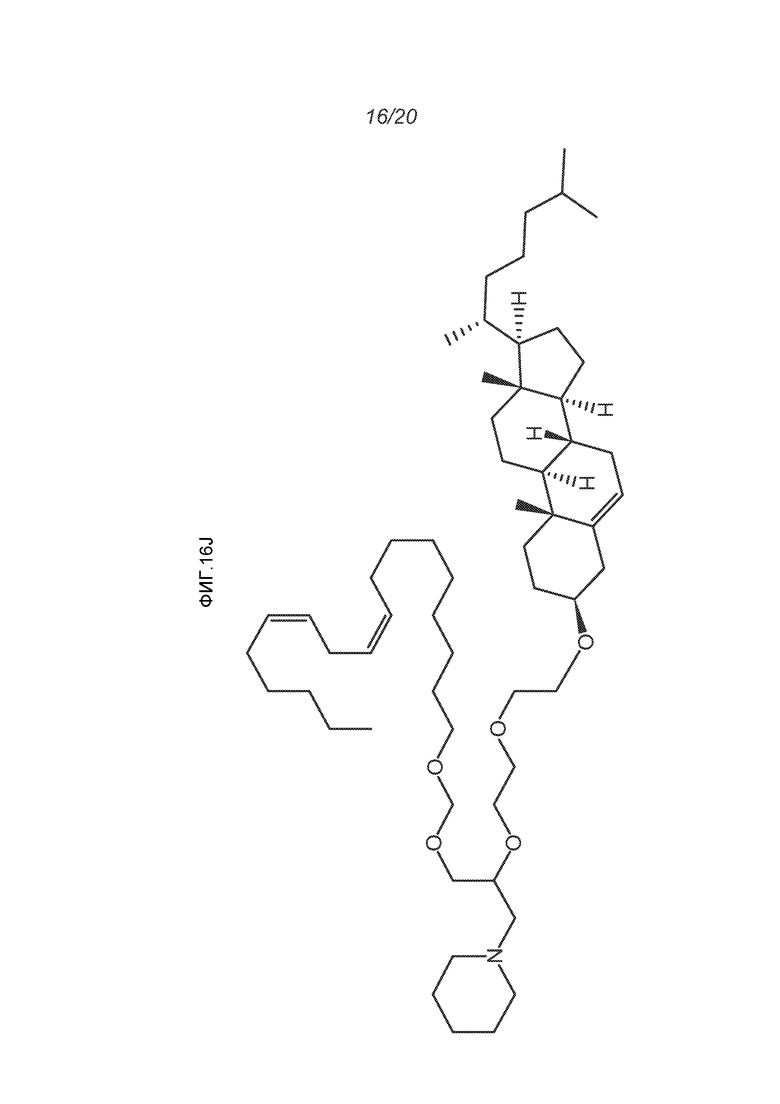
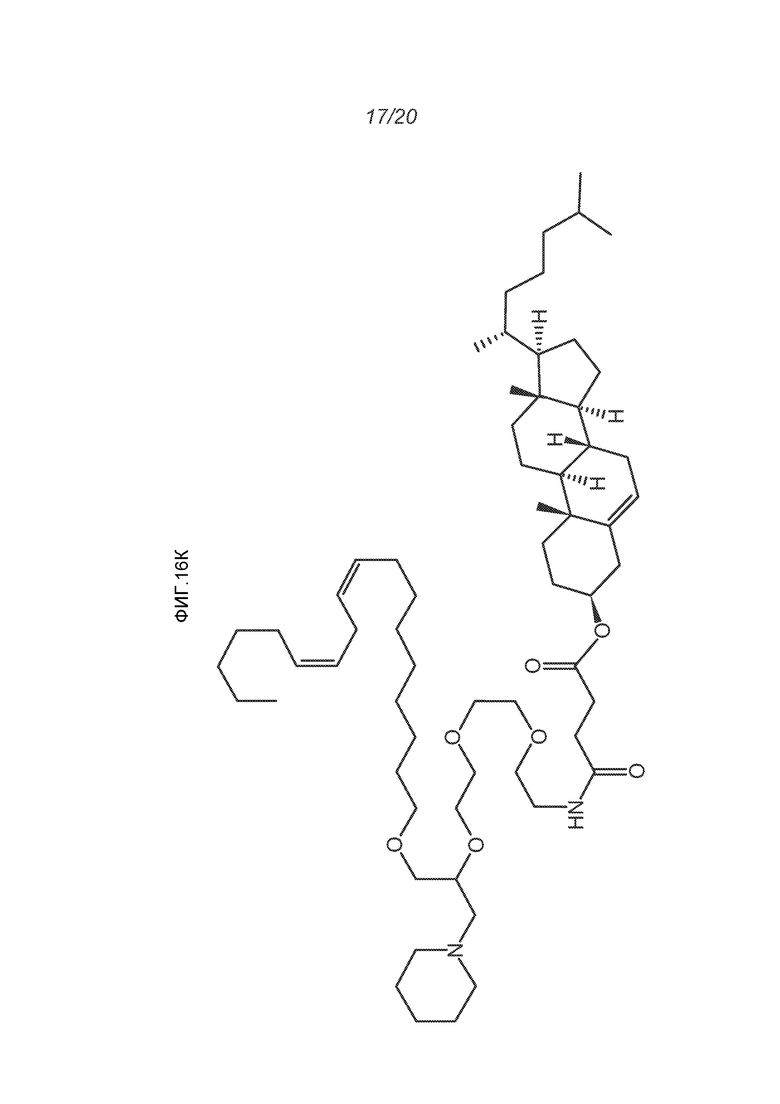
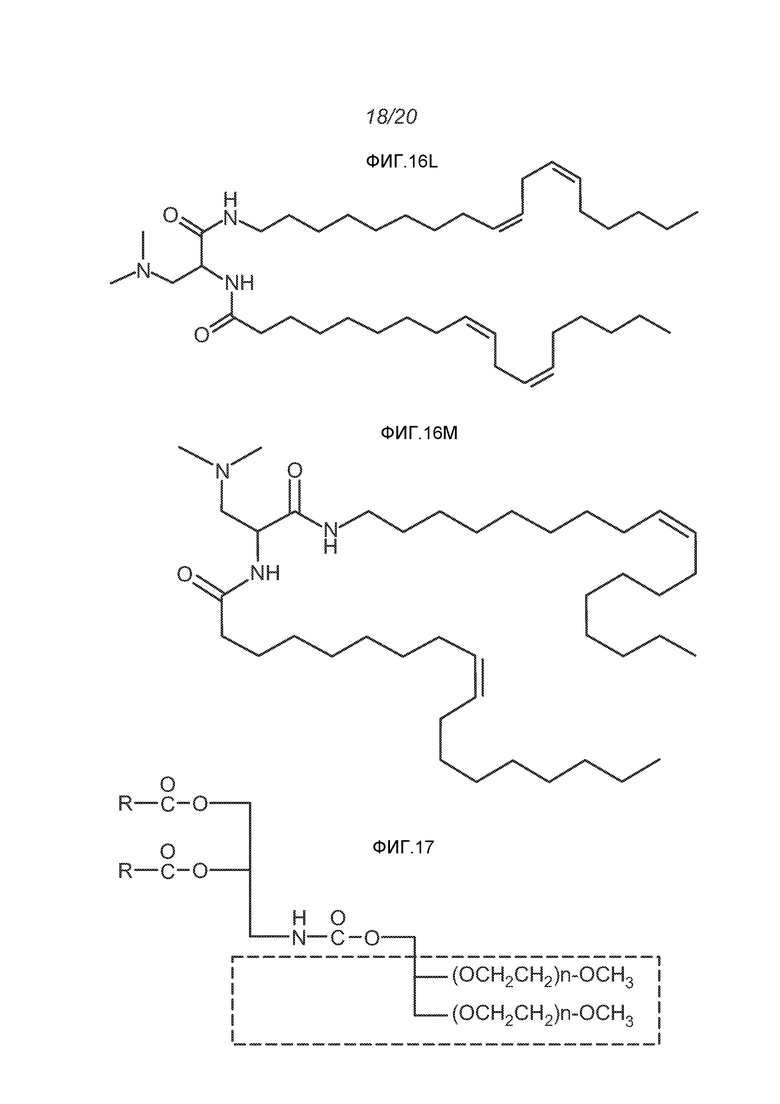
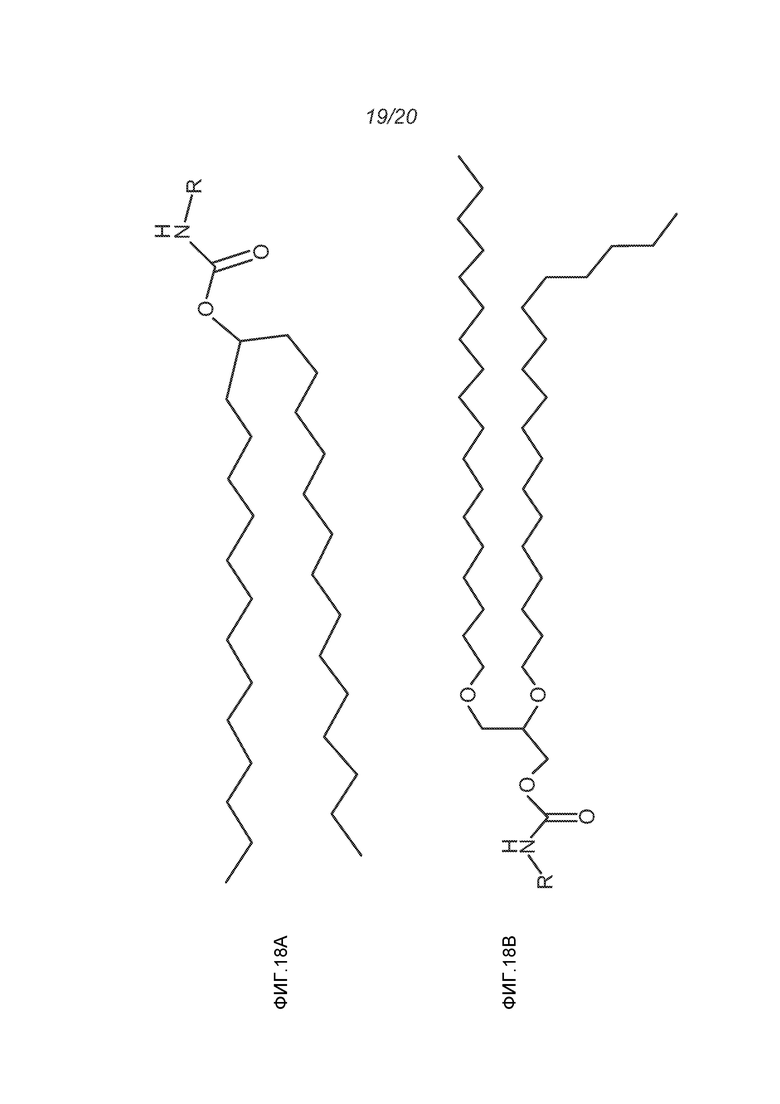
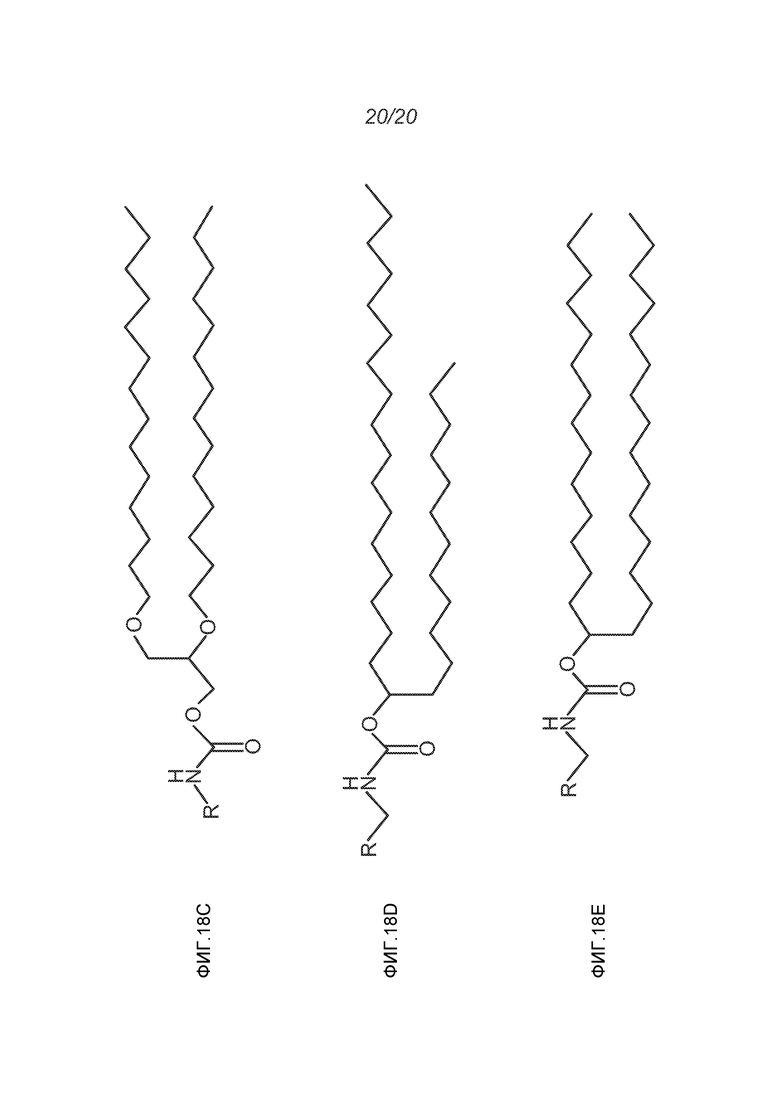
Настоящее изобретение относится к биотехнологии, в частности к липосоме для in vivo доставки РНК в клетку позвоночного. Указанная липосома содержит инкапсулированную самореплицирующуюся РНК, кодирующей представляющий интерес иммуноген, в молярных процентах от 20% до 80% катионного липида, от 35% до 50% холестерина и цвиттерионный липид. Липосома имеет диаметр в интервале значений 60-180 нм. Указанный иммуноген, который инкапсулирован в липосому, вызывает иммунный ответ против бактерии, вируса, грибка или паразита. Настоящее изобретение раскрывает фармацевтическую композицию для иммунизации субъектов, содержащую эффективное количество указанной липосомы, и способ выработки защитной иммунной реакции у позвоночных с её использованием. Настоящее изобретение позволяет улучшить доставку вакцин из нуклеиновых кислот в организм субъекта. 3 н. и 7 з.п. ф-лы, 18 ил., 1 табл., 1 пр.
1. Липосома для in vivo доставки РНК в клетку позвоночного, в которой инкапсулирована самореплицирующаяся РНК, кодирующая представляющий интерес иммуноген, и где липосома имеет диаметр в интервале значений 60-180 нм, где липосома содержит (i) от 20% до 80% (молярный процент) катионного липида, (ii) цвиттерионный липид и (iii) от 35% до 50% (молярный процент) холестерина, где иммуноген вызывает иммунный ответ против бактерии, вируса, грибка или паразита.
2. Липосома по п.1, где липосома имеет диаметр в интервале 80-160 нм.
3. Липосома по п.1, отличающаяся тем, что липосома имеет диаметр около 97 нм.
4. Липосома по п. 1, где указанная липосома содержит DlinDMA (1,2-дилинолеилокси-N,N-диметил-3-аминопропан), DSPC (1,2-диастеароил-sn-глицеро-3-фосфохолин), DPyPE (1,2-дифитаноил-sn-глицеро-3-фосфоэтаноламин), холестерин, ПЭГилированный липид или комбинацию перечисленного.
5. Липосома по п.1, где самореплицирующаяся РНК кодирует (i) РНК-зависимую РНК полимеразу, которая может транскрибировать РНК из самореплицирующейся РНК, и (ii) иммуноген.
6. Липосома по п.1, где самореплицирующаяся РНК содержит две открытые считывающие рамки, причем первая из них кодирует альфавирусную репликазу и вторая из них кодирует иммуноген.
7. Липосома по п.1, где длина самореплицирующейся РНК составляет 9000-12000 нуклеотидов.
8. Липосома по п.1, где длина самореплицирующейся РНК составляет 5000-25000 нуклеотидов.
9. Фармацевтическая композиция для иммунизации субъектов, содержащая эффективное количество липосомы по любому из пп.1-8.
10. Способ выработки защитной иммунной реакции у позвоночных, включающий стадию введения позвоночному эффективного количества липосомы по любому из пп. 1-8.
| US 2006083780 A1, 20.04.2006 | |||
| F | |||
| MARTINON, S | |||
| KRISHNAN et al., Induction of virus-specific cytotoxic T lymphocytes in vivo by liposome-entrapped mRNA, Eur | |||
| J | |||
| Immunol., 1993, vol.23, pp.1719-1722 | |||
| S | |||
| SAXENA et al., Induction of immune responses and protection in mice against rabies using a self-replicating RNA vaccine encoding rabies virus glycoprotein, Veterinary Microbiology, 2009, vol.136, pp.36-44 | |||
| СИСТЕМА ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ | 2002 |
|
RU2294192C2 |

