ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка истребует приоритет, предоставляемый в связи с подачей Предварительной заявки на патент США № 61/747 672, поданной 31 декабря 2012 года, которая в полном объеме содержится в настоящий документ посредством ссылки.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области дифференцировки клеток. Подробнее, изобретение включает применение специфических гормонов щитовидной железы или их аналогов, и ALK5 ингибиторов в качестве регуляторов HB9 в панкреатических энтодермальных и эндокринных клетках.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Последние достижения в области заместительной клеточной терапии для лечения сахарного диабета 1 типа и нехватка островков Лангерганса для трансплантации заставили обратить внимание на разработку источников инсулин-продуцирующих клеток, или β-клеток, подходящих для приживления трансплантата. Один подход представляет собой получение функциональных β-клеток из плюрипотентных стволовых клеток, таких как, например, эмбриональные стволовые клетки.
При эмбриональном развитии позвоночных плюрипотентные клетки дают начало группе клеток, содержащих три зародышевых листка (эктодерму, мезодерму и энтодерму), в ходе процесса, известного как гаструляция. Ткани, такие как ткань щитовидной железы, вилочковой железы, поджелудочной железы, кишечника и печени, будут развиваться из энтодермы через промежуточную стадию. Промежуточная стадия данного процесса представляет собой образование дефинитивной энтодермы.
К концу гаструляции энтодерма разделяется на передний и задний отделы, которые могут быть распознаны по экспрессии ряда факторов, однозначно выделяющих переднюю, среднюю и заднюю области энтодермы. Например, HHEX и SOX2 идентифицируют переднюю область, в то время как CDX1, 2 и 4 идентифицируют заднюю половину энтодермы.
Миграция ткани энтодермы приближает энтодерму к различным мезодермальным тканям, которые способствуют регионализации кишечной трубки. Это достигается за счет целого ряда секретируемых факторов, таких как FGFs, Wnts, TGF-Bs, ретиноевой кислоты (RA), лигандов BMP и их антагонистов. Например, FGF4 и BMP способствуют экспрессии CDX2 в предполагаемой энтодерме задней кишки и подавляют экспрессию передних генов Hhex и SOX2 (Development 2000 г., 127:1563-1567). Было продемонстрировано, что сигнализация WNT также действует параллельно с сигнализацией FGF, способствуя развитию задней кишки и препятствуя зачаткам передней кишки (Development 2007 г., 134:2207-2217). Наконец, ретиноевая кислота, секретируемая мезенхимой, регулирует границу между передней и задней кишкой (Curr Biol 2002 г., 12:1215-1220).
Уровень экспрессии специфических факторов транскрипции может использоваться для определения типа ткани. Во время преобразования дефинитивной энтодермы в примитивную кишечную трубку кишечная трубка становится разделенной на широкие домены, которые можно наблюдать на молекулярном уровне с помощью ограниченных паттернов экспрессии генов. Например, регионализованный отдел поджелудочной железы в кишечной трубке демонстрирует очень высокую экспрессию PDX1 и очень низкую экспрессию CDX2 и SOX2. PDX1, NKX6.1, PTF1A и NKX2.2 высоко экспрессируются тканью поджелудочной железы; а экспрессия CDX2 высока в кишечной ткани.
Образование поджелудочной железы происходит при дифференцировании дефинитивной энтодермы в панкреатическую энтодерму. Дорсальный и вентральный домены поджелудочной железы формируются из эпителия передней кишки. Передняя кишка также дает начало формирования пищевода, трахеи, легких, щитовидной железы, желудка, печени, поджелудочной железы и системы желчных протоков.
Клетки панкреатической энтодермы экспрессируют панкрео-дуоденальный, содержащий гомеобокс ген PDX1. В отсутствие PDX1 поджелудочная железа не развивается дальше образования вентрального и дорсального зачатков. Следовательно, экспрессия PDX1 является важным этапом органогенеза поджелудочной железы. Зрелая поджелудочная железа содержит как экзокринную, так и эндокринную ткани, которые образуются при дифференцировании панкреатической энтодермы.
D'Amour et al. описывает получение обогащенных культур дефинитивной энтодермы, полученной из эмбриональных стволовых клеток человека в присутствии высокой концентрации активина и низкой концентрации сыворотки (Nature Biotechnology 2005 г., 23:1534-1541; патент США № 7 704 738). Трансплантация этих клеток под капсулу почки у мышей приводила к дифференцированию в более зрелые клетки с характеристиками ткани энтодермы (патент США № 7,704,738). Клетки дефинитивной энтодермы, полученные из эмбриональных стволовых клеток человека, могут быть далее дифференцированы в PDX1-положительные клетки после добавления FGF10 и ретиноевой кислоты (заявка на патент США № 2005/0266554). Последующая трансплантация таких клеток-предшественников панкреатических клеток в жировое тело иммунодефицитных мышей привела к образованию функциональных панкреатических эндокринных клеток с последующей 3-4-месячной стадией созревания (патент США № 7,993,920 и патент США № 7,534,608).
Fisk et al. сообщают о системе получения клеток панкреатических островков из эмбриональных стволовых клеток человека (патент США № 7,033,831). В данном случае процесс дифференцирования был разделен на три стадии. Эмбриональные стволовые клетки человека сначала были дифференцированы в энтодерму при помощи комбинации бутирата натрия и активина A (патент США № 7,326,572). Затем клетки культивировали с антагонистами BMP, такими как Noggin, в комбинации с EGF или β-целлюлином для генерирования PDX1-положительных клеток. Конечное дифференцирование индуцировали никотинамидом.
Низкомолекулярные ингибиторы также применяли для индуцирования клеток-предшественников панкреатических эндокринных клеток. Например, низкомолекулярные ингибиторы рецептора TGF-β и рецепторов ВМР (Development 2011, 138:861-871; Diabetes 2011, 60:239-247) применялись для значительного увеличения количества эндокринных клеток поджелудочной железы. Кроме того, низкомолекулярные активаторы также применяли для генерирования клеток дефинитивной энтодермы или клеток-предшественников панкреатических клеток (Curr Opin Cell Biol 2009 г., 21:727-732; Nature Chem Biol 2009, 5:258-265).
НВ9 (также известный как HIXB9 и MNX1) является транскрипционным активаторным белком bHLH, экпрессируемым на ранней стадии развития поджелудочной железы, начинающейся приблизительно на 8 эмбриональный день. НВ9 также экспрессируется в нотохорде и спинном мозге. Экспрессия HB9 является неравномерна и достигает пика примерно в день 10,5 в эпителии поджелудочной железы, будучи экспрессированной в клетках, экспрессирующих PDX1 и NKX6.1. Примерно в день 12,5 экспрессия HB9 снижается и на поздних стадиях ограничивается только β клетками. В гомозиготах мышей с нулевой мутацией HIXB9, дорсальная доля поджелудочной железы не развивается (Nat Genet 23:67-70, 1999; Nat Genet 23:71-75, 1999). HB9-/- β-клетки экспрессируют низкие уровни глюкозного транспортера, GLUT2 и NKX6.1. Далее, HB9 -/- поджелудочной железы демонстрирует значительное снижение в количестве инсулин-положительных клеток в то время не оказывая значительного влияния на экспрессию прочих гормонов поджелудочной железы. Таким образом, временной контроль HB9 чрезвычайно важен для нормального развития и функционирования β-клетки. В то время как немного известно о факторах, регулирующих экспрессию HB9 в β-клетках, недавнее исследование на данио предполагает, что ретиноевая кислота может положительно регулировать экспрессию HB9 (Development, 138, 4597-4608, 2011).
Гормоны щитовидной железы, тироксин («T4») и трииодотиронин («Т3»), являются гормонами, основанными на тирозине, которые производятся щитовидной железой и первоначально отвечают за регулирование метаболизма. Основной формой гормонов щитовидной железы в крови является Т4, обладающий более длительным периодом полужизни, чем Т3. Соотношение Т4 и Т3, выделяемых в кровь, составляет точно 20 к 1. Т4 конвертируется в более активный Т3 (в три-четыре раза более мощный, чем Т4) внутри клеток с помощью деиодиназы.
Т3 прикрепляется к рецепторам гормонов щитовидной железы, TRα1 и TRβ1 (TR). TR является ядерным рецептором гормонов, который гетеродимеризуется с ретиноидным Х рецептором. Димеры связываются с отвечающими элементами щитовидной железы (TREs) в отсутствии лиганда и действуют как транскрипционные репрессоры. Прикрепление Т3 к TR понижает репрессию TRE-зависимых генов и стимулирует экспрессию различных целевых генов. В то время как многочисленные исследования предполагали, что роль Т3 состоит в увеличении пролиферации β-клеток, снижении апоптоза и улучшении секреции инсулина, его роль в дифференцировке клеток не определена.
Трансформирующий фактор роста бета (TGF-β) относится к большому семейству плеотропных цитокинов, которые задействованы во многих биологических процессах, включая контроль роста, дифференцировку, миграцию, выживание клеток, фиброз и спецификация процесса развития. Представители суперсемейства TGF-β передают сигнал посредством рецепторного комплекса, содержащего рецепторы типа I и типа II. Лиганды TGF-B (такие как активины и GDF (факторы дифференцировки роста)) связывают вместе рецептор типа II с рецептором типа I. Рецептор типа II фосфорилирует и активирует рецептор типа I в комплексе. Существует пять видов рецепторов типа II у млекопитающих: TβR-II, ActR-II, ActR-IIB, BMPR-II, и AMHR-II и семь рецепторов типа I (ALKs 1–7). Активин и родственные ему лиганды сигнализируют через комбинации ActR-II или ActR-IIB и ALK4 или ALK5, и BMP сигнализируют через комбинации ALK2, ALK3, и ALK6 с ActR-II, ActR-IIB, или BMPR-II. AMH сигнализирует через комплекс AMHR-II с ALK6, и лимфоузел, как выяснилось недавно, сигнализирует через комплекс ActR-IIB и ALK7 (Cell. 2003,113(6):685-700). После закрепления лиганда TGF-B на соответствующем рецепторе испускаемые сигналы передаются в ядро, первоначально через активацию комплексов Smad-белков. При активации рецепторы типа I фосфолирируют представителей рецептор-регулируемого подсемейства Smad-белков. Это активирует их и дает возможность формировать комплексы с обычными медиаторами Smad, Smad4. Smad 1, 5, и 8 являются субстратами для ALK 1, 2, 3, и 6, в то время как Smad 2 и 3 являются субстратами для ALK 4, 5, и 7 (FASEB J 13:2105–2124). Активированные комплексы Smad аккумулируются в ядре, где они напрямую задействованы в транскрипции целевых генов, как правило в ассоциации с другими специфическими ДНК-связывающими транскрипционными факторами. Составы, которые избирательно ингибируют рецепторы TGF-β, были разработаны для терапевтического применения и для изменения судьбы клетки в контексте репрограммирования и дифференцировки из различных популяций стволовых клеток. В частности, ингибиторы ALK5 ранее применялись для направления дифференцировки эмбриональных стволовых клеток к эндокринной судьбе (Diabetes, 2011, 60(1):239-47).
В целом, процесс дифференцировки прогениторных клеток в функциональные β-клетки проходит через различные стадии. Известно, что направление стволовых клеток, полученных из эмбрионов человека (hES), в условиях in vitro прогрессивно через стадии коммитирования к клеткам, подобным β-клеткам, является сложной задачей, и получение функциональных β-клеток из клеток hES не является прямолинейным процессом. Каждый шаг в процессе диференцировки прогениторных клеток является уникальной задачей. Несмотря на большие успехи, достигнутые в оптимизации протоколов по получению панкреатических клеток из плюрипотентных стволовых клеток человека, существует необходимость разработки протоколов для получения функциональных эндокринных клеток, и в частности, β-клеток.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фиг. 1A–1С представлены данные анализов ПЦР в реальном времени по экспрессии следующих генов в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных в панкреатические энтодермальные/эндокринные клетки-предшественники, как описано в примере 1: PDX1 (ФИГ. 1A), NKX6.1 (ФИГ. 1B) и HB9 (ФИГ. 1C).
На фиг. 2А-2С представлены результаты анализа FACS клеток линии эмбриональных стволовых клеток человека Н1, дифференцированных в панкреатические энтодермальные/эндокринные клетки-предшественники, как описано в примере 1 для PDX1 (ФИГ. 2A), NKX6.1 (ФИГ. 2B) и HB9 (ФИГ. 2C).
На фиг. 3А и 3В представлены изображения клеток, иммуноокрашенных для NXK6.1 инсулина или НВ9. Клетки были дифференцированы в панкреатические энтодермальные/эндокринные клетки-предшественники, как описано в примере 1. ФИГ. 3А демонстрирует иммунное окрашивание для NKX6.1 (левая часть) и инсулина (правая часть). ФИГ. 3В демонстрирует иммунное окрашивание для HB9 (левая часть) и инсулина (правая часть).
На фиг. 4А, В и С представлены данные FACS для процентной экспрессии PDX1, NKX6.1 и HB9 на стадии 4 день 3 (панель А), стадии 5 день 4 (панель В) и стадии 6 день 3 (панель С) эмбриональный стволовых клеток линии Н1, дифференцированных на стадиях 4-6 как описано в примере 2.
ФИГ. 5А демонстрирует мРНК экспрессию НВ9 в сравнении с человеческими островками на стадиях 2-6 для клеток, как описано в примере 2.
ФИГ. 5В представляет изображения клеток на стадии 4 дня 3, которые были дифференцированы как описано в примере 2, иммунноокрашены для NXK6.1 (левая часть) и НВ9 (правая часть).
На фиг. 6А–6J представлены данные ПЦР в реальном времени при анализе экспрессии следующих генов в эмбриональных стволовых клетках человека линии Н1, дифференцированных до стадии 4, как описано в примере 2, а затем обработаны только на стадии 4, на стадиях 4-5, или на стадиях 4-6. На фиг. 6А-6J представлены данные для следующих: NKX6.1 (ФИГ. 6A); PDX1 (ФИГ. 6B); NKX2.2 (ФИГ. 6C); глюкагон (ФИГ. 6D); инсулин (ФИГ. 6E); соматостатин (ФИГ. 6F); CDX2 (ФИГ. 6G); альбумин (ФИГ. 6H); гастрин (ФИГ. 6I); и SOX2 (ФИГ. 6J).
На фиг. 7А и 7В представлены результаты иммуноокрашивания контроля (ФИГ. 7А) и обработанных культур (ФИГ. 7В) как описано в примере 2. Иммуноокрашивание контроля (ФИГ. 7А) и обработанных культур (ФИГ. 7В) на стадии 6 позволяет обнаружить значительное увеличение количества НВ9 положительных клеток в группе, обработанной Т3 (ФИГ. 7В) по сравнению с контролем (ФИГ. 7А) на стадии 6.
На фиг. 8А и 8В описано иммуноокрашивание для NKX6.1 и НВ9 на стадии 6 день 7 для клеток, дифференцированных на стадии 6 как описано в примере 3. ФИГ 8С описывает данные анализов ПЦР в реальном времени по экспрессии НВ9 в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных на стадии 6 как описано в примере 3.
На фиг. 9А и В описаны данные FACS для стадии 6 день 5 и день 15 соответственно, для НВ9 в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных на стадии 6 как описано в примере 3.
На фиг. 10А-10Е описано иммуноокрашивание для NKX6.1 и НВ9 на стадии 6 день 6 для клеток, дифференцированных в соответствии с протоколом, описанным в примере 4. В зависимости от дозы Т3 в значительной степени увеличил количество НВ9-положительных клеток среди NKX6.1-положительных клеток-предшественников энтодермы поджелудочной железы.
На фиг. 11A–11L представлены данные анализов ПЦР в реальном времени по экспрессии следующих генов в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных на стадии 6 как описано в примере 4: SOX2 (ФИГ. 11A); NKX6.1 (ФИГ. 11B); NKX2.2 (ФИГ. 11C); гастрин (ФИГ. 11D), PDX1 (ФИГ. 11E); NGN3 (ФИГ. 11F); PAX6 (ФИГ. 11G); PAX4 (ФИГ. 11Н); инсулин (ФИГ. 11I); глюкагон (ФИГ. 11J); грелин (ФИГ. 11К) и соматостатин (ФИГ. 11L).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Приведенное ниже подробное описание настоящего изобретения, будет более понятно при изучении вместе с приложенными рисунками. Рисунки предоставлены с целью иллюстрации определенных вариантов осуществления изобретения. Однако, настоящее изобретение не ограничивается только приведенными точными конструкциями, примерами и устройствами. Для четкости описания, а не для ограничения настоящего изобретения, подробное описание настоящего изобретения разделено на подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Настоящее изобретение относится к получению клеток энтодермы поджелудочной железы, которые положительны для NKX6.1, PDX1 и HB9 путем применения определенных гормонов щитовидной железы, или их аналогов, и ALK5 (TGFβ рецептор киназы типа I) ингибиторов в особой культуральной последовательности. Соответственно, настоящее изобретение представляет клеточную культуру в условиях in vitro для дифференцировки клеток, полученных из плюрипотентных стволовых клеток, в клетки, экспрессирующие маркеры, характерные для линии β-клеток, которые экспрессируют NKX6.1, PDX1 и HB9. Изобретение далее представляет способ получения таких клеток путем клеточной культуры в условиях in vitro. В определенных вариантах осуществления, изобретение основано на открытии, согласно которому включения Т3, Т4 или их аналогов, действует как увеличитель экспрессии белка НВ9 в дифференцируемых клетках для упрощения дифференцировки в направлении β-клеток. НВ9 не экспрессируется на белковом уровне на стадии 3 или стадии 4. Соответственно, настоящее изобретение представляет способы дифференцировки стволовых клеток путем регулирования экспрессии белка НВ9. В частности, изобретение представляет получение клеток энтодермы поджелудочной железы, которые положительны для NKX6.1, PDX1 и HB9 путем применения Т3, Т4, или их аналогов, и ALK5 ингибиторов в особой культуральной последовательности.
Определения
Стволовые клетки представляют собой недифференцированные клетки, определяемые как обладающие способностью на одноклеточном уровне к самообновлению и дифференциации. Стволовые клетки могут производить клетки-потомки, включая самообновляющиеся прогениторные клетки, необновляющиеся прогениторные клетки и окончательно дифференцированные клетки. Стволовые клетки также характеризуются своей способностью дифференцироваться in vitro в функциональные клетки различных линий дифференцирования из множества зародышевых листков (энтодермы, мезодермы и эктодермы). Стволовые клетки также дают начало тканям множества зародышевых листков после трансплантации и вносят значительный вклад в образование большинства, если не всех тканей после инъекции в бластоцисты.
Стволовые клетки классифицируются по потенциалу развития. Плюрипотентные клетки способны преобразовываться во все виды эмбриональных клеток.
Дифференцировка представляет собой процесс, посредством которого неспециализированная («некоммитированная») или менее специализированная клетка приобретает свойства специализированной клетки, например нервной клетки или мышечной клетки. Дифференцированная клетка представляет собой клетку, занявшую более специализированное («коммитированное») положение в линии дифференцировки клетки. Термин «коммитированная» применительно к процессу дифференцирования относится к клетке, дошедшей в ходе процесса дифференцирования до стадии, с которой в нормальных условиях она продолжит дифференцироваться до конкретного типа клеток или подмножества типов клеток и не сможет в нормальных условиях дифференцироваться в иной тип клеток или вернуться к менее дифференцированному типу. «Дедифференцировка» обозначает процесс, в ходе которого клетка возвращается к менее специализированной (или коммитированной) позиции в клеточной линии дифференцировки. Применяемый в настоящем документе термин «клеточная линия дифференцировки» означает наследственность клетки, т.е. из каких клеток произошла данная клетка и каким клеткам она может дать начало. В клеточной линии дифференцировки клетка помещается в наследственную схему развития и дифференцировки. Маркер, специфичный для линии дифференцирования, относится к характеристике, специфически ассоциированной с фенотипом клеток интересующей линии дифференцирования, и его можно использовать для оценки дифференцирования некоммитированной клетки в клетки интересующей линии дифференцирования.
В настоящем документе термин «маркеры» означает молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в исследуемых клетках. В данном контексте под дифференциальной экспрессией понимают повышенный уровень положительного маркера и пониженный уровень отрицательного маркера по сравнению с недифференцированной клеткой. Обнаруживаемый уровень маркерной нуклеиновой кислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из множества способов, известных в данной области.
В настоящем документе клетка «положительна по» заданному маркеру или «положительна», если заданный маркер явно обнаруживается в клетке. Аналогично клетка «отрицательна по» заданному маркеру или «отрицательна», если заданный маркер не обнаруживают в клетке. В частности, положительность по FACS как правило выше, чем 2%, в то время как отрицательный предел по FACS как правило менее 1%. Положительность по ПЦР как правило менее 34 циклов (Cts): в время как отрицательность по ПЦР как правило более 34,5 циклов.
В попытках воспроизвести дифференцировку плюрипотентных стволовых клеток в функциональные клетки энтодермы поджелудочной железы в статичных клеточных культурах в условиях in vitro, процесс дифференцировки часто рассматривается как прогрессирование через несколько последовательных стадий. В частности, процесс дифференцировки в целом рассматривается как прогрессирование через шесть стадий. В этой постадийной прогрессии «стадия 1» относится к первому шагу в процессе дифференцировки, дифференцировке плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы (здесь и далее упоминаются как «клетки стадии 1»). «Стадия 2» относится ко второму шагу, дифференцировке клеток, экспрессирующих маркеры, характерные для клеток дефинитивной энтодермы, в клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки (здесь и далее упоминаются как «клетки стадии 2»). «Стадия 3» относится к третьему шагу, дифференцировке клеток, экспрессирующих маркеры, характерные для клеток кишечной трубки, в клетки, экспрессирующие маркеры, характерные для клеток энтодермы передней кишки (здесь и далее упоминаются как «клетки стадии 3»). «Стадия 4» относится к четвертому шагу, дифференцировке клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки кишечной трубки, в клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки (здесь и далее упоминаются как «клетки стадии 4»). «Стадия 5» относится к пятому шагу, дифференцировке клеток, экспрессирующих маркеры, характерные для клеток-предшественников панкреатической передней кишки, в клетки, экспрессирующие маркеры, характерные для панкреатических энтодермальных клеток и/или панкреатиченских эндокринных клеток-предшественников (здесь и далее упоминаются как «панкреатические энтодермальные клетки/эндокринные клетки-предшественники» или альтернативно как «клетки стадии 5»). «Стадия 6» относится к дифференцировке клеток, экспрессирующих маркеры, характерные для панкреатических энтодермальных клеток и / или эндокринных панкреатических клеток-предшественников, в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток (здесь и далее упоминаются как «клетки стадии 6»).
Однако, следует отметить, что не все клетки в отдельно взятой популяции прогрессируют через эти стадии с одинаковой скоростью. Вследствие этого, в клеточных культурах в условиях in vitro нередко обнаруживается присутствие клеток, которые прогрессировали менее или более по пути дифференцировки, чем большинство клеток, присутствующих в популяции, в особенности на поздних стадиях дифференцировки. Например, нередко наблюдается появление маркеров, характерных для панкреатических энтодермальных клеток, во время стадии 5 культивирования клеток. Для иллюстративных нужд настоящего изобретения характеристики различных типов клеток ассоциированы с определенными выше стадиями как описано в настоящем документе.
«Клетки дефинитивной энтодермы», как используется в настоящем документе, относится к клеткам, которые несут в себе характеристики клеток, появившиеся из эпибласта во время гаструляции, и которые формируют желудочно-кишечный тракт и его производные. Клетки дефинитивной энтодермы экспрессируют по меньшей мере один из следующих маркеров: FOXA2 (также известный куак гепатоцитный ядерный фактор 3-β (HNF3β)), GATA4, SOX17, CXCR4, брахиурия, Cerberus, OTX2, goosecoid, C-Kit, CD99, и MIXL1. Маркерные характеристики клеток дефинитивной энтодермы включают CXCR4, FOXA2 и SOX17. Таким образом, клетки дефинитивной энтодермы могут быть охарактеризованы их экспрессией CXCR4, FOXA2 и SOX17. Дополнительно, в зависимости от длительности времени, на протяжении которого клеткам позволяется оставаться на стадии 1, можно наблюдать рост в HNF4α.
«Клетки кишечной трубки», как используется в данном документе, относится к клеткам, произошедшим от дефинитивной энтодермы, которые могут дать начало всем энтодермальным органам, таким как легкие, печень, поджелудочная железа, желудок и кишечник. Клетки кишечной трубки могут быть охарактеризованы их постоянно растущей экспрессией HNF4α, выше, чем экспрессия клеток дефинитивной энтодермы. Например, рост экспрессии HNF4α в мРНК в десять-сорок раз можно наблюдать на стадии 2.
«Энтодерма передней кишки», как используется в данном документе, относится к клеткам энтодермы, которые дают начало пищеводу, легким, желудку, печени, поджелудочной железе, желчному пузырю и части двенадцатиперстной кишки. Клетки энтодермы передней кишки экспрессируют по меньшей мере один из следующих маркеров: PDX1, FOXA2, CDX2, SOX2 и HNF4α. Клетки энтодермы передней кишки могут быть охарактеризованы ростом экспресии PDX1 по сравнению с клетками кишечной трубки. Например, более пятидесяти процентов клеток в культурах стадии 3 типично экспрессируют PDX1.
«Клетка-предшественник панкреатической передней кишки», как используется в данном документе, относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: PDX1, NKX6.1, HNF6, NGN3, SOX9, PAX4, PAX6, ISL1, гастрин, FOXA2, PTF1a, PROX1 и HNF4α. Клетки-предшественники панкреатической передней кишки поджелудочной железы можно охарактеризовать как положительные по отношению к экспрессии PDX1, NKX6.1 и SOX9.
Используемый в настоящей заявке термин, «панкреатическая энтодермальная клетка» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: PDX1, NKX6.1, HNF1 β, PTF1 α, HNF6, HNF4α, SOX9, NGN3; гастрин; HB9, или PROX1. Панкреатические энтодермальные клетки могут характеризоваться отсутствием у них значительной экспрессии CDX2 или SOX2.
«Панкреатическая эндокринная клетка-предшественник», как используется в настоящем документе, относится к клеткам энтодермы поджелудочной железы, обладающим возможностью стать клеткой, экспрессирующей гормон поджелудочной железы. Панкреатические эндокринные клетки-предшественники экспрессируют по меньшей мере один из следующих маркеров: NGN3, NKX2.2, NeuroD1, ISL1, PAX4, PAX6 или ARX. Панкреатические эндокринные клетки-предшественники могут быть охарактеризованы их экспрессией NKX2.2 и NeuroD1.
Используемый в настоящей заявке термин «панкреатическая эндокринная клетка» относится к клеткам, способным к секреции по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин или панкреатический полипептид. В дополнение к этим гормонам, маркерные характеристики панкреатических эндокринных клеток включают один или несколько из NGN3, NeuroD1, ISL1, PDX1, NKX6.1, PAX4, ARX, NKX2.2, и PAX6. Панкреатические эндокринные клетки, экспрессирующие маркеры бета-клеток, могут характеризоваться их экспрессией инсулина и по меньшей мере одного из следующих транскрипционных факторов: PDX1, NKX2.2, NKX6.1, NeuroD1, ISL1, HNF3β, MAFA и PAX6.
В настоящем документе используются попеременно «д1», «1д», и «день1», «д2», «2д», и «день 2» и так далее. Эти комбинации цифр и букв относятся к конкретному дню инкубации на различных стадиях в процессе поэтапного протокола дифференцировки настоящей заявки.
В настоящем документе термин «глюкоза» применяют относительно декстрозы, сахару, обычно встречающемуся в природе.
В настоящем документе термин «NeuroD1» применяют для обозначения белка, экспрессируемого в прогениторных клетках панкреатических эндокринных клеток, и гена, кодирующего его.
«LDN-193189» относится к ((6-(4-(2-(пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил)пиразоло[1,5-a]пиримидин, гидрохлорид; DM-3189)) ингибитор рецептора BMP, доступный под торговой маркой STEMOLECULE™ от Stemgent, Inc., Cambridge, MA, USA.
Характеристики, источник, размножение и культивирование плюрипотентных стволовых клеток
А. Характеристики плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или более стадиеспецифических эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, обнаруживаемые антителами, обозначенными как Tra-1-60 и Tra-1-81 (Thomson et al. 1998, Science 282:1145-1147). Дифференцирование плюрипотентных стволовых клеток in vitro приводит к потере экспрессии Tra-1-60 и Tra-1-81. Недифференцированные плюрипотентные стволовые клетки, как правило, имеют щелочнофосфатазную активность, которую можно обнаружить путем обработки клеток 4%-м раствором параформальдегида, а затем выращивая с Vector Red в качестве субстрата, как описано производителем (Vector Laboratories, штат Калифорния, США). Недифференцированные плюрипотентные стволовые клетки также, как правило, экспрессируют OCT4 и TERT, определяемые с помощью ОТ-ПЦР.
Другим желательным фенотипическим свойством выращенных плюрипотентных клеток является потенциал дифференцировки в клетки всех трех зародышевых листков: в эндодермальные, мезодермальные и эктодермальные ткани. Плюрипотентность стволовых клеток можно подтвердить, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), фиксирования образующихся тератом с помощью 4% параформальдегида и их последующего гистологического исследования на наличие клеточных типов, происходящих от этих трех зародышевых листков. Плюрипотентность можно альтернативно определить по созданию эмбриоидных телец и анализа эмбриоидных телец на наличие маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток можно кариотипировать с использованием стандартного способа G-бэндинга и сравнить с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т. е. эуплоидные клетки, в которых все хромосомы человека присутствуют и не имеют видимых изменений.
В. Источники плюрипотентных стволовых клеток
Примерные типы плюрипотентных стволовых клеток, которые можно применять, включают в себя устойчивые линии плюрипотентных клеток, в том числе преэмбриональной ткани (такой как, например, бластоцист), эмбриональной ткани или ткани плода, взятой в любой момент во время беременности, как правило, но не обязательно перед сроком приблизительно от 10 до 12 недель беременности. Не имеющими ограничительного характера примерами являются устойчивые линии эмбриональных стволовых клеток человека, такие как, например, эмбриональные стволовые клетки человека линий H1, H7 и H9 (WiCell Research Institute, Мэдисон, штат Висконсин, США). Клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивированных в отсутствие питающих клеток, также пригодны для использования. Индуцибельные плюрипотентные клетки (IPS) или перепрограммированные плюрипотентные клетки, полученные из взрослых соматических клеток с помощью принудительной экспрессии ряда факторов, относящихся к плюрипотентным транскрипционным факторам, таким как OCT4, NANOG, SOX2, KLF4 и ZFP42 (Annu Rev Genomics Hum Genet 2011 г., 12:165–185, см. также IPS, Cell, 126(4): 663-676) также могут использоваться. Эмбриональные стволовые клетки человека, применяемые в способах настоящего изобретения, также могут быть подготовлены, как описано Thomson и др. (патент США № 5,843,780; Science, 1998 г.; 282:1145–1147; Curr Top Dev Biol, 1998 г.; 38:133–165; Proc Natl Acad Sci USA 1995, 92:7844-7848). Мутантные линии эмбриональных стволовых клеток человека, такие как BG01v (BresaGen, Athens, Ga.), или клетки, полученные из зрелых соматических клеток человека, такие как клетки, описанные в Takahashi et al., Cell 131: 1-12 (2007) также могут применяться. В определенных вариантах осуществления плюрипотентные стволовые клетки, пригодные для применения в настоящем изобретении, могут быть получены с помощью способов, описанных в: Li et al. (Cell Stem Cell 4: 16-19, 2009); Maherali et al. (Cell Stem Cell 1: 55-70, 2007); Stadtfeld et al. (Cell Stem Cell 2: 230-240); Nakagawa et al. (Nature Biotechnol 26: 101-106, 2008); Takahashi et al. (Cell 131: 861-872 (2007), и в заявке на патент США № 2011/0104805. В определенный вариантах осуществления плюрипотентные клетки могут иметь не-эмбриональное происхождение. Все эти ссылки, патенты и приложения к патентам включены в настоящий документ методом ссылок во всей их полноте, в частности, поскольку они относятся к выделению, культуре, размножению и дифференцировке плюрипотентных клеток.
С. Размножение и культивирование плюрипотентных стволовых клеток
В одном варианте осуществления плюрипотентные стволовые клетки, как правило, культивируют на слое питающих клеток, которые поддерживают плюрипотентные клетки различными путями. Плюрипотентные стволовые клетки альтернативно культивируют в культуральной системе, по существу не содержащей питающих клеток, но тем не менее поддерживающей пролиферацию плюрипотентных стволовых клеток и не допускающей существенного дифференцирования. Рост плюрипотентных стволовых клеток в не содержащей питающих клеток культуральной системе без дифференцирования поддерживают путем использования среды, кондиционированной посредством предварительного культивирования клеток иного типа. Рост плюрипотентных стволовых клеток в не содержащей питающих клеток культуральной системе без дифференцирования альтернативно поддерживают путем использования среды с определенным химическим составом.
Плюрипотентные клетки можно легко размножить в культуре путем использования различных питательных слоев или с помощью сосудов, покрытых матриксными белками. Альтернативно для стандартного размножения клеток можно использовать химически определенные поверхности в комбинации со средами определенного состава, такими как среды mTeSR® 1 (StemCell Technologies, г. Ванкувер, Канада). Плюрипотентные клетки можно легко удалить из планшетов с культурой посредством ферментативной, механической обработки или с применением различных кальцийхелатирующих агентов, таких как EDTA (этилендиаминтетрауксусная кислота). Альтернативно, плюрипотентные клетки можно размножить в суспензии в отсутствие каких-либо матриксных белков или питательного слоя.
Множество различных способов размножения и культивирования плюрипотентных стволовых клеток могут применяться в описываемом изобретении. Например, способы изобретения могут использовать способы Reubinoff et al., Thompson et al., Richard et al. и заявки на патент США № 2002/0072117. Reubinoff et al. (Nature Biotechnology 18: 399-404 (2000)) и Thompson et al. (Science 282: 1145-1147 (1998) описывают культивирование линий плюрипотентных стволовых клеток из человеческих бластоцист с применением слоя питающих клеток из мышиных эмбриональных фибробластов. Richards et al. (Stem Cells 21: 546-556, 2003) оценивает панель из одиннадцати слоев различных человеческих взрослых, эмбриональных и неонатальных питающих клеток по их способности поддерживать культуру плюрипотентных стволовых клеток человека, замечая, что линии человеческих эмбриональных стволовых клеток, культивированных на питательных фибробластах взрослой кожи достигают морфологии эмбриональных стволовых клеток человека и остаются плюрипотентными. В заявке на патент США № 2002/0072117 описаны линии клеток, продуцирующие среду, которая поддерживает рост плюрипотентных стволовых клеток приматов в не содержащей питающих клеток культуре. Использованные клеточные линии представляют собой мезенхимальные и фибробластоподобные клеточные линии, полученные из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. В заявке на патент США № 2002/072117 также описано использование клеточных линий в качестве первичного слоя питающих клеток.
Прочие пригодные способы размножения и культивирования плюрипотентных стволовых клеток описаны, к примеру, в Wang et al., Stojkovic et al., Miyamoto et al. и Amit et al. Wang et al. (Stem Cells 23: 1221–1227, 2005 г. ), описаны способы длительного выращивания плюрипотентных стволовых клеток человека на слоях питающих клеток, полученных из эмбриональных стволовых клеток человека. Stojkovic et al. (Stem Cells 2005 23: 306–314, 2005 г. ), описана система питающих клеток, полученная в результате спонтанного дифференцирования эмбриональных стволовых клеток человека. Miyamoto et al. (Stem Cells 22: 433–440, 2004 г. ), описан источник питающих клеток из плаценты человека. Amit et al. (Biol. Reprod 68: 2150–2156, 2003 г. ) описывает слой питающих клеток, полученных из крайней плоти человека.
В другом варианте осуществления, подходящие способы размножения и культивирования плюрипотентных стволовых клеток описаны, к примеру, в Inzunza et al., патенте США No. 6,642,048, WO 2005/014799, Xu et al. и заявке на патент США № 2007/0010011. Inzunza et al. (Stem Cells 23: 544–549, 2005 г. ), описывает слой питающих клеток, полученных из фибробластов постнатальной крайней плоти человека. В патенте США № 6 642 048 описана среда, поддерживающая рост плюрипотентных стволовых клеток приматов в не содержащей питающих клеток среде, а также клеточные линии, которые можно использовать для получения такой среды. Патент США № 6 642 048 сообщает о мезенхимальных и фибробластоподобных клеточных линиях, полученных из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток; так же как и способах получения таких клеточных линий, обработки среди и выращивания стволовых клеток с применением подобной среды. В международной заявке WO 2005/014799 описана кондиционированная среда для поддержания, пролиферации и дифференцирования клеток млекопитающих. В заявке на патент WO 2005/014799 сообщает, что культуральная среда, изготовленная в соответствии с настоящим изобретением, кондиционируется при помощи секреторной активности клеток мыши, в частности, активности дифференцированных и иммортализованных трансгенных гепатоцитов, именуемых MMH (Met Murine Hepatocyte). Xu et al. (Stem Cells 22: 972–980, 2004 г. ), описана кондиционированная среда, полученная из производных эмбриональных стволовых клеток человека, генетически модифицированных для сверхэкспрессии обратной транскриптазы теломеразы человека. В заявке на патент США № 2007/0010011 описана культуральная среда с определенным химическим составом для поддержания плюрипотентных стволовых клеток.
В альтернативной культуральной системе используют бессывороточную среду, обогащенную факторами роста, способными стимулировать пролиферацию эмбриональных стволовых клеток. Примеры подобных культуральных систем включают, без ограничения, Cheon et al., Levenstein et al. и патентом США № 2005/0148070. Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870, October 19, 2005) описана не содержащая питающих клеток и сыворотки культуральная система, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной, заменяющей сыворотку среде (SR), обогащенной различными факторами роста, способными запустить самообновление эмбриональных стволовых клеток. Levenstein et al. (Stem Cells 24: 568–574, 2006 г. ), описаны способы длительного культивирования эмбриональных стволовых клеток человека в отсутствие фибробластов или кондиционированной среды с применением среды, обогащенной основным фактором роста фибробластов (bFGF). В заявке на патент США № 2005/0148070, описывается способ культивирования человеческих эмбриональных стволовых клеток в среде с определенным составом без сыворотки и без питающих клеток-фибробластов, где данный способ включает: культивирование стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минеральные вещества, по меньшей мере один трансферрин или заменитель трансферрина, по меньшей мере один инсулин или заместитель инсулина, культуральную среду, в основном, не включающую эмбриональную сыворотку млекопитающих и содержащую по меньшей мере приблизительно 100 нг/мл фактора роста фибробластов, способного активировать сигнальный рецептор фактора роста фибробластов, при этом фактор роста фибробластов происходит из источника, иного, чем просто слой питающих клеток-фибробластов, среду, поддерживающую пролиферацию стволовых клеток в недифференцированном состоянии без слоя питающих клеток или кондиционированной среды.
Другие пригодные способы культивирования и размножения плюрипотентных стволовых клеток описаны в заявке на патент США № 2005/0233446, патенте США № 6,800,480, заявке на патент США № 2005/0244962 и WO 2005/065354. В заявке на патент США № 2005/0233446, описана среда с определенным составом, которую можно использовать при культивировании стволовых клеток, включая недифференцированные зародышевые стволовые клетки приматов. Среда в растворе по существу является изотонической по сравнению с культивируемыми стволовыми клетками. В данной культуре конкретная среда содержит основную среду и количество каждого из bFGF, инсулина и аскорбиновой кислоты, необходимое для поддержки по существу недифференцированного роста зародышевых стволовых клеток. В патенте США № 6800480, отмечено, что культуральная среда для выращивания зародышевых стволовых клеток приматов по существу в недифференцированном состоянии, включающая в себя основную среду с низким содержанием эндотоксина и низким осмотическим давлением, которая эффективно поддерживает рост зародышевых стволовых клеток приматов. В описании в патенте 6 800 480 далее сообщается, что основную среду объединяют с питательной сывороткой, которая эффективно поддерживает рост зародышевых стволовых клеток приматов, и субстратом, выбранным из группы, состоящей из питающих клеток и компонента внеклеточного матрикса, полученного из питающих клеток. Эта среда дополнительно включает в себя аминокислоты, не относящиеся к незаменимым, антиоксидант и первый фактор роста, выбранный из группы, состоящей из нуклеозидов и соли пировиноградной кислоты. В заявке на патент США № 2005/0244962 сообщается, что один из аспектов раскрытия представляет способ культивирования эмбриональных стволовых клеток приматов и что стволовые клетки в культивируют в культуре, по существу не содержащей эмбриональную сыворотку млекопитающих (предпочтительно также по существу не содержащей сыворотку любого животного) и при наличии фактора роста фибробластов, полученного из источника, отличного от просто питающего слоя фибробластов.
Международная заявка WO 2005/065354 раскрывает определенную, изотоническую культуральную среду, которая по существу не содержит питающих клеток и сыворотки, и содержит: базальную среду; bFGF; инсулин; и аскорбиновую кислоту. Подробнее, в международной заявке № WO2005/086845, описан способ поддержания недифференцированной стволовой клетки, причем указанный способ включает воздействие на стволовую клетку представителем семейства белков трансформирующего ростового фактора-бета (TGF-β), представителем семейства белков фактора роста фибробластов (FGF) или никотинамидом (NIC) в количестве, достаточном для поддержания клетки в недифференцированном состоянии в течение периода времени, достаточного для получения необходимого результата.
Плюрипотентные стволовые клетки можно высевать на соответствующий культуральный субстрат. В одном варианте осуществления соответствующий культуральный субстрат представляет собой компонент внеклеточного матрикса, такой как, например, полученный из базальной мембраны или компонент, который может участвовать в лиганд-рецепторном взаимодействии с адгезивными молекулами. В одном из вариантов осуществления подходящим культуральным субстратом является МАТРИГЕЛЬ® (Becton Dickenson). МАТРИГЕЛЬ® представляет собой растворимый препарат из клеток опухоли Энгельбрета-Холма-Суорма, который при комнатной температуре превращается в гель и образует восстановленную базальную мембрану.
В качестве альтернативы допустимо использовать другие компоненты внеклеточного матрикса и смеси компонентов. В зависимости от типа пролиферирующих клеток данные компоненты могут включать в себя по отдельности или в различных комбинациях ламинин, фибронектин, протеогликан, энтактин, гепарансульфат и т.п.
Плюрипотентные стволовые клетки можно высевать на субстрат с соответствующим распределением и при наличии среды, поддерживающей выживаемость, размножение и сохранение желательных характеристик клеток. Все данные характеристики улучшаются при тщательном подходе к распределению при посеве и могут быть без труда определены специалистом в данной области. Пригодная культуральная среда может быть изготовлена, например, из следующих компонентов: модифицированная среда Дюльбекко по методу Игла (DMEM), которая продается под торговой маркой Gibco™ (часть № 11965-092) от Life Technologies Corporation, Grand Island, NY; модифицированная среда Дюльбекко по методу Игла нокаут (KO DMEM), которая продается под торговой маркой Gibco™ (часть № 10829-018) от Life Technologies Corporation, Grand Island, NY; базальная следа Хэма F12/50% DMEM; 200 мМ L-глютамин, который продается под торговой маркой Gibco™ (часть № 15039-027) от Life Technologies Corporation, Grand Island, NY; непервостепенный раствор аминокислоты, который продается под торговой маркой Gibco™ (часть № 11140-050) от Life Technologies Corporation, Grand Island, NY; бета-меркаптоэтанол (часть № М7522), которые продает компания Sigma-Aldrich, Company, LLC, Saint Louis, MO; и фактор роста человеческого рекомбинантного базисного фибробласта (bFGF), которые продается под торговой маркой Gibco™ (часть № 13256- 029) компании Life Technologies Corporation, Grand Island, NY.
Дифференцировка плюрипотентных стволовых клеток
По мере того как плюрипотентные клетки дифференцируются в бета-клетки, они дифференцируются через различные стадии, каждая из которых может быть охарактеризована присутствием или отсутствием определенных маркеров. Дифференцировка клеток на этих стадиях достигается путем создания специфических условий культивирования, в том числе присутствие или отсутствие определенных факторов, добавленный в культуральную среду. В целом, эта дифференцировка может задействовать дифференцировку плюрипотентных стволовых клеток в дефинитивные клетки энтодермы. Эти дефинитивные клетки энтодермы могут далее быть дифференцированы в клетки кишечной трубки, которые в свою очередь могут затем быть дифференцированы в клетки энтодермы передней кишки. Клетки энтодермы передней кишки могут быть дифференцированы в клетки-предшественники передней кишки поджелудочной железы, которые, в свою очередь, дифференцируются в клетки энтодермы поджелудочной железы, панкреатические эндокринные клетки-предшественники или и те, и другие. Эти клетки могут быть затем дифференцированы в клетки, продуцирующие гормоны поджелудочной железы (такие как бета-клетки).
Это изобретение представляет пошаговую дифференцировку плюрипотентных стволовых клеток в эндокринные клетки поджелудочной железы с применением гормона щитовидной железы (такого как Т3, аналоги Т3, Т4, аналоги Т4, или их комбинации (совокупно далее именуемых «Т3/Т4»)) и ингибитора ALK5. Это изобретение также представляет пошаговую дифференцировку плюрипотентных стволовых клеток в эндокринные клетки поджелудочной железы с применением гормона щитовидной железы (такого как Т3/Т4) или ингибитора ALK5. Пригодные для применения аналоги гормонов щитовидной железы могут включать: GC-1 (Собертиром), доступный от R & D Systems, Inc. кат. № 4554; DITPA) 3,5-дииодотиропропионовоая кислота); КВ-141, обсуждаемый в J. Steroid Biochem. Мол. Biol. 2008, 111; 262-267 и Proc. Natl. Acad. Sci. US 2003, 100 10067-10072; MB07344, обсужденный в Proc. Natl. Acad. Sci. US 2007, 104 15490-15495; T0681, обсужденный в PLoS One, 2010, 5e8722 и J. Lipid Res. 2009, 50; 938-944; и GC-24, обсужденный в PLoS One, 2010, e8722 и Endocr. Pract. 2012; 18(6): 954-964, содержание которого полностью включено в настоящий документ. Пригодные ALK5 ингибиторы включают: ALK5 ингибитор II (Enzo, Farmingdale, NY); ALK5i (Axxora, San Diego, CA); SD208 (R & D systems (MN)); TGF-B ингибитор SB431542 (Xcess Biosciences (San Diego, CA)); ITD-1 (Xcess Biosciences (San Diego, CA)); LY2109761 (Xcess Biosciences (San Diego, CA)); A83-01 (Xcess Biosciences (San Diego, CA)); LY2157299 (Xcess Biosciences (San Diego, CA)); TGF-β рецептор ингибитор V (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор I (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор IV (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор VII (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор VIII (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор II (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор VI (EMD Chemicals, Gibstown, NJ); TGF-β рецептор ингибитор III (EMD Chemicals, Gibstown, NJ).
Дифференцировка плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для эндокринных клеток поджелудочной железы
Характеристики плюрипотентных стволовых клеток хорошо известны специалистам в данной области, и продолжается выявление дополнительных характеристик плюрипотентных стволовых клеток. К маркерам плюрипотентных стволовых клеток относится, например, экспрессия одного или более из следующих маркеров: ABCG2, cripto, FOXD3, CONNEXIN43, CONNEXIN45, OCT4, SOX2, NANOG, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, TRA-1-60 и TRA-1-81.
Примеры плюрипотентных стволовых клеток включают линию эмбриональных стволовых клеток человека H9 (код NIH: WA09), эмбриональные стволовые клетки человека линии H1 (код NIH: WA01), эмбриональные стволовые клетки человека линии H7 (код NIH: WA07), а также эмбриональные стволовые клетки человека линии SA002 (Cellartis, Швеция). Также подходящими для применения являются клетки, экспрессирующие по меньшей мере один из следующих маркеров, характерных для плюрипотентных клеток: ABCG2, cripto, CD9, FOXD3, CONNEXIN43, CONNEXIN45, OCT4, SOX2, NANOG, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, TRA-1-60 и TRA-1-81.
Также подходящими для применения в рамках настоящего изобретения являются клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцирования дефинитивной энтодермы. В одном из вариантов осуществления настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии дифференцирования дефинитивной энтодермы, представляет собой клетку-предшественника первичной полоски. В альтернативном варианте осуществления клетка, экспрессирующая маркеры, характерные для линии дифференцирования дефинитивной энтодермы, представляет собой мезэнтодермальную клетку. В альтернативном варианте осуществления клетка, экспрессирующая маркеры, характерные для линии дифференцирования дефинитивной энтодермы, представляет собой клетку дефинитивной энтодермы.
Также подходящими для применения в рамках настоящего изобретения являются клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии панкреатической энтодермы. В одном из вариантов осуществления данного изобретения экспрессируемые клеткой маркеры, характерные для линейки панкреатической энтодермы, представляют собой клетки панкреатической энтодермы, в которых экспрессия PDX1 и NKX6.1 значительно превышает экспрессию CDX2 и SOX2. Особенно пригодными являются клетки, в которых экспрессия PDX1 и NKX6.1 по меньшей мере в два раза превышает экспрессию CDX2 или SOX2.
В одном из вариантов осуществления генерируются панкреатические эндокринные клетки, способные экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Подходящей для применения в рамках настоящего изобретения является клетка-предшественник, экспрессирующая по меньшей мере один из маркеров, характерных для линии панкреатических эндокринных клеток. В одном из вариантов осуществления настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. В предпочтительном варианте осуществления панкреатическая эндокринная клетка являются бета-клеткой, производящей инсулин.
В определенных вариантах осуществления для получения клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток, применяется протокол, согласно которому процесс начинается с плюрипотентных стволовых клеток или индуцибельных плюрипотентных клеток, предпочтительно плюрипотентных стволовых клеток. Данный протокол включает следующие стадии.
Стадия 1: Плюрипотентные стволовые клетки, такие как эмбриональные стволовые клетки, полученные для культуры клеточных линий, обрабатываются соответствующими факторами для стимулирования дифференцировки в клетки, экспрессирующие маркеры, характерные для дефинитивных энтодермальных клеток.
Стадия 2: Клетки, полученные на стадии 1, обрабатываются соответствующими факторами для стимулирования дальнейшей дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки.
Стадия 3: Клетки, полученные на стадии 2, обрабатываются соответствующими факторами для стимулирования дальнейшей дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток энтодермы передней кишки.
Стадия 4: Клетки, полученные на стадии 3, обрабатываются соответствующими факторами (в том числе определенными вариантами осуществления Т3/Т4) для стимулирования дальнейшей дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки.
Стадия 5: Клетки, полученные на стадии 4, обрабатываются соответствующими факторами (в том числе определенными вариантами осуществления: (i) T3/T4; (ii) ALK5 ингибитора; или (iii) одновременно Т3/Т4 и ALK5 ингибитора) для стимулирования дальнейшей дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток энтодермы поджелудочной железы/эндокринных клеток-предшественников.
Стадия 6: Клетки, полученные на стадии 5, обрабатываются соответствующими факторами (в том числе определенными вариантами осуществления Т3/Т4, ALK5 ингибитора, или и тем, и другим) для стимулирования дальнейшей дифференцировки в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток.
В том время как настоящее изобретение, в определенных вариантах осуществления, включает дифференцировку плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток, изобретение также включает дифференцировку клеток, являющуюся результатом других промежуточных стадий, ведущих к получению панкреатических эндокринных клеток. В частности, изобретение включает дифференцировку клеток, экспрессирующих маркеры, характерные для клеток-предшественников панкреатической передней кишки, в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток. Более того, хотя процесс описан в отдельных стадиях, обработка, так же как и прогресс клеток через процесс дифференцировки, может быть как последовательным, так и непрерывным.
Способы оценки экспрессии белковых маркеров и маркеров нуклеиновых кислот в культивированных или выделенных клетках являются стандартными для данной области. Эти способы включают в себя количественную полимеразную цепную реакцию с обратной транскриптазой (ОТ-ПЦР), Нозерн-блот, гибридизацию in situ (см., например, Current Protocols in Molecular Biology (Ausubel et al., ред. 2001 г., дополнение)), а также иммунологические анализы (такие как иммуногистохимический анализ среза материала), Вестерн-блоттинг, а для маркеров, доступных в интактных клетках, - способ проточной цитометрии (FACS) (см., например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998 г. )). Далее, эффективность дифференцировки может быть определена путем обработки популяции клеток агентом (например, антителом), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для интересующего типа клеток.
Дифференцированные клетки в дальнейшем также могут быть очищены. Например, после обработки плюрипотентных стволовых клеток с применением способов настоящего изобретения дифференцированные клетки можно очистить путем воздействия на обрабатываемую популяцию клеток веществом (таким как антитело), специфически распознающим белковый маркер, характерно экспрессируемый очищенными дифференцированными клетками.
Стадия 1: Дифференцировка плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы.
Плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы, с использованием любого способа, известного специалистам в данной области, или любого способа, предложенного в настоящем изобретении. Способы, пригодные для дифференцировки плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, обнародованы в: заявке на патент США № 2007/0254359; заявке на патент США № 2009/0170198; заявке на патент США № 2009/0170198; заявке на патент США № 2011/0091971; заявке на патент США № 2010/0015711; заявке на патент США № 2010/0015711; заявке на патент США № 2012/0190111; заявке на патент США № 2012/0190112; заявке на патент США № 2012/0196365; заявке на патент США № 20100015711; заявке на патент США № 2012/0190111; заявке на патент США № 2012/0190112; заявке на патент США № 2012/0196365; заявке на патент США № 20100015711; заявке на патент США № 2012/0190111; заявке на патент США № 2012/0190112; заявке на патент США № 2012/0196365; предварительной заявке на патент США № 61/076900; предварительной заявке на патент США № 61/076908 и предварительной заявке на патент США № 61/076915, которые полностью включены в настоящий документ путем ссылки, поскольку они относятся к плюрипотентным стволовым клеткам и дифференцировке плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы.
В одном из вариантов осуществления плюрипотентные стволовые клетки обрабатываются средой, обогащенной активином А и Wnt3A для получения поколения клеток, экспрессирующизх маркеры, характерные для клеток дефинитивной энтодермы. Обработка может задействовать контакт плюрипотентных стволовых клеток средой, содержащей от приблизительно 50 нг/мл до приблизительно 150 нг/мл, альтернативно от приблизительно 75 нг/мл до приблизительно 125 нг/мл, альтернативно около 100 нг/мл активина А. Обработка также может включать контакт клеток с от приблизительно 10 нг/мл до приблизительно 50 нг/мл, альтернативно от приблизительно 15 нг/мл до приблизительно 30 нг/мл, альтернативно приблизительно 20 нг/мл Wnt3A. Плюрипотентные клетки могут культивировать на протяжении приблизительно двух-пяти дней, предпочтительно приблизительно два-три дня, для обеспечения удобства их дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы. В одном из вариантов осуществления плюрипотентные клетки культивируются в присутствии активина А и Wnt3A на протяжении одного дня, после чего культивирование продолжается в присутствии активина А (без присутствия Wnt3A).
В другом варианте осуществления изобретения плюрипотентные стволовые клетки обрабатывают средой, обогащенной дифференцированным фактором роста 8 («GDF8») и ингибитором гликоген синтазы киназы-3 β («GSK3β») (таким как циклические анилино-пиридинотриазиновые композиции, раскрытые в заявке на патент США № 2010/0015711, полностью включенной в данный документ путем ссылки) для стимулирования дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы. Предпочтительным ингибитором GSK3β является 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазатетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-он, далее именуемый «Композиция MCX». Обработка может включать контакт плюрипотентных стволовых клеток со средой, обогащенной от приблизительно 50 нг/мл до приблизительно 150 нг/мл, альтернативно от приблизительно 75 нг/мл до приблизительно 125 нг/мл, альтернативно приблизительно 100 нг/мл GDF8. Обработка может также включать контакт клеток с от приблизительно 0,1 до 5 мкМ, альтернативно от приблизительно 0,5 до приблизительно 2,5 мкМ, предпочтительно примерно 1 мкМ композиции МСХ. Плюрипотентные клетки могут культивировать на протяжении приблизительно двух-пяти дней, предпочтительно приблизительно три-четыре дня, для обеспечения удобства их дифференцировки в клетки дефинитивной энтодермы. В одном из вариантов осуществления плюрипотентные клетки культивируются в присутствии GDF8 и композиции МСХ на протяжении одного дня, затем культивируются в присутствии GDF8 и более низкой концентрации композиции МСХ на протяжении одного дня, затем культивируются в присутствии GDF8 на протяжении одного дня в отсутствии композиции МСХ. В частности, клетки могут культивироваться в присутствии GDF8 и приблизительно 1мкМ композиции МСХ на протяжении одного дня, затем культивируются в присутствии GDF8 и примерно 0,1 мкМ композиции МСХ на протяжении одного дня, затем культивируются в присутствии GDF8 на протяжении одного дня в отсутствии композиции МСХ. В альтернативном варианте осуществления клетки могут культивироваться в присутствии GDF8 и приблизительно 1мкМ композиции МСХ на протяжении одного дня, затем культивируются в присутствии GDF8 и примерно 0,1 мкМ композиции МСХ на протяжении одного дня.
Образование клеток, экспрессирующих маркеры, характерные для клеток дефинитивной энтодермы, можно определить путем проверки на наличие маркеров до и после выполнения конкретного протокола. Плюрипотентные стволовые клетки, как правило, не экспрессируют данных маркеров. Таким образом, дифференцировка плюрипотентных клеток может быть обнаружена, когда клетки начинают экспрессировать маркеры, характерные для клеток дефинитивной энтодермы.
Стадия 2: Дифференцирование клеток экспрессирующих маркеры, характерные для клеток дефинитивной эндодермы, в клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки.
Клетки, экспрессирующие маркеры, характерные для клеток дефинитивной эндодермы, могут далее дифференцироваться в клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки. В одном из вариантов осуществления формирование клеток, экспрессирующих маркеры, характерные для клеток кишечной трубки, включает культивирование клеток, экспрессирующих маркеры, характерные для клеток дефинитивной энтодермы со средой, содержащей фактор роста фибробластов («FGF»)7 или FGF10 для дифференцировки этих клеток. Например, культуральная среда может включать от приблизительно 25 нг/мл до приблизительно 75 нг/мл, альтернативно от приблизительно 30нг/мл до приблизительно 60 нг/мл, альтернативно приблизительно 50 нг/мл FGF7 или FGF10, предпочтительно FGF7. Клетки можно культивировать при этих условиях на протяжении двух-трех дней, предпочтительно примерно два дня.
В другом варианте осуществления дифференцировка в клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки, включает культивирование клеток, экспрессирующих маркеры, характерные для клеток дефинитивной энтодермы, с FGF7 или FGF10 и аскорбиновой кислотой (витамином С). Культуральная среда может содержать от приблизительно 0,1 мМ до приблизительно 0,5 мМ аскорбиновой кислоты, альтернативно от приблизительно 0,2 мМ до приблизительно 0,4 мМ, альтернативно приблизительно 0,25 мМ аскорбиновой кислоты. Культуральная среда также может включать от приблизительно 10 нг/мл до приблизительно 35 нг/мл, альтернативно от приблизительно 15нг/мл до приблизительно 30 нг/мл, альтернативно приблизительно 25 нг/мл FGF7 или FGF10, предпочтительно FGF7. Например, культуральная среда может содержать приблизительно 0,25 мМ аскорбиновой кислоты и приблизительно 25 нг/мл FGF7. В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы, обрабатываются FGF7 и аскорбиновой кислотой на протяжении 2 дней.
Стадия 3: Дифференцирование клеток, экспрессирующих маркеры, характерные для клеток кишечной трубки, в клетки, экспрессирующие маркеры, характерные для клеток энтодермы передней кишки.
Клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки, могут далее дифференцироваться в клетки, экспрессирующие маркеры, характерные для клеток энтодермы передней кишки. В одном из вариантов осуществления, клетки стадии 2 далее дифференцируются в клетки стадии 3 путем культивирования этих клеток в культуральной среде, обогащенной сглаженным ингибитором рецепторов («SMO») (таким как циклопамин или MRT10 (N-[[[3-бензойламино)фенил]амино]тиоксометил]-3,4,5-триметоксибензамид)) или Sonic Hedgehog («SHH») антагонистом, сигнализирующим о траектории (таким как сглаженный антагонист 1 («SANT-1») ((Е)-4-бензил-N-((3,5-диметил-l-фенил-lH-пиразол-4-ил)метилен-пиперазин-l-амин)), Hedgehog ингибитор траектории 1 («HPI-l) (2-метоксиэтил 1,4,5,6,7,8-гексагидро-4-(3-гидроксифенил)-7-(2-метоксифенил)-2-метил-5-оксо-3-хинолдинкарбоксилат), ретиноевой кислотой и Noggin. Альтернативно, среда может быть обогащена ингибитором SMO, SHH антагонистом, сигнализирующим о траектории, ретиноевой кислотой и Noggin. Клетки можно культивировать на протяжении приблизительно двух-четырех дней, предпочтительно примерно два дня. В одном из вариантов осуществления среда обогащена от приблизительно 0,1 мкМ до приблизительно 0,3 мкМ SANT-1, от приблизительно 0,5 мкМ до приблизительно 3 мкМ ретиноевой кислоты и от приблизительно 75 нг/мл до приблизительно 125 нг/мл Noggin. В другом варианте осуществления среда обогащена приблизительно 0,25 мкМ SANT-1, приблизительно 2 мкМ ретиноевой кислоты и приблизительно 100 нг/мл Noggin.
В альтернативном варианте осуществления, клетки стадии 2 далее дифференцируются в клетки стадии 3 путем обработки клеток стадии 2 средой, обогащенной FGF7 или FGF10, ретиноевой кислотой, ингибитором SMO (таким как MRT10 или циклопамин), или антагонистом, сигнализирующим о траектории, SHH (таким как SANT-1 или HPI-1), активатором белка Киназа С («РКС») (таким как ((2S,5S)-(E,E)-8-(5-(4-(трифлюорометил)фенил)-2,4-пентадиенойламино)бензолактам) («ТРВ»); EMD Chemicals Inc., Gibbstown NJ), форбол-12,13-дибутират («PDBu»), форбол-12-миристат-13-ацетат («PMA») или индолактам V («ILV»), ингибитором морфогенического белка кости («BMP») (таким как LDN-193189, Noggin или Chordin) и аскорбиновой кислотой. В другом варианте осуществления среда может быть обогащена FGF7 или FGF10, ретиноевой кослотой, ингибитором SMO, антагонистом, сигнализирующим о траектории (таким как SANT-1), активатором РКС (таким как ТРВ), ингибитором ВМР (таким как LDN-193189), и аскорбиновой кислотой. Клетки могут культивироваться в присутствии этих факторов роста, низкомолекулярных антагонистов и антагонистов на протяжении приблизительно двух-трех дней.
В одном из вариантов осуществления среда обогащена от приблизительно 15 нг/мл до приблизительно 35 нг/мл FGF7, от приблизительно 0,5 мкМ до приблизительно 2 мкМ ретиноевой кислоты, от приблизительно 0,1 мкМ до приблизительно 0,4 мкМ SANT-1, от приблизительно 100 нМ до приблизительно 300 нМ ТРВ, от приблизительно 50 нМ до приблизительно 200 нМ LDN-193189 и от приблизительно 0,15 мкМ до приблизительно 0,35 мкМ аскорбиновой кислоты. В другом варианте осуществления среда обогащена приблизительно 25 нг/мл FGF7, приблизительно 1 мкМ ретиноевой кислоты, приблизительно 0,25 мкМ SANT-1, приблизительно 200 нМ ТРВ, приблизительно 100 нМ LDN-193189 и приблизительно 0,25 мкМ аскорбиновой кислоты.
Стадии 4-6: Дифференцировка клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки, в клетки, экспрессирующие маркеры, характерные для клеток энтодермы поджелудочной железы, путем обработки культуральной средой, обогащенной гормонами щитовидной железы Т3/Т4 или ингибитором ALK5, или одновременно Т3/Т4 и ингибитором ALK5.
Настоящее изобретение обеспечивает дальнейшую дифференцировку клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки, путем обработки культуральной средой, обогащенной гормонами щитовидной железы Т3/Т4 или ингибитором ALK5, или одновременно Т3/Т4 и ингибитором ALK5. В некоторых вариантах осуществления, изобретение обеспечивает дальнейшую дифференцировку таких клеток на стадиях 4-6 путем обработки культуральной средой, обогащенной (a) Т3, (b) ингибитором ALK5 или (c) Т3 и ингибитором ALK5 на одной ил нескольких из этих стадий.
В одном из вариантов осуществления настоящее изобретение обеспечивает способ получения клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток из плюрипотентных стволовых клеток, включающий следующие стадии:
a. культивирование плюрипотентных стволовых клеток;
b. дифференцировка плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для клеток эндодермы передней кишки; и
c. дифференцировка клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки, в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток, путем обработки культуральной средой, обогащенной (i) Т3/Т4, (ii) ингибитором ALK5, или (iii) одновременно Т3/Т4 и ингибитором ALK5.
В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для панкреатический эндокринных клеток, являются бета-клетками. В другом варианте осуществления, полученные клетки являются положительными для NKX6.1, PDX1 и HB-9. Способ может в значительной степени увеличить количество НВ9-положительных клеток среди NKX6.1-положительных клеток-предшественников энтодермы поджелудочной железы. Способ может также понизить экспрессию NKX2.2 или SOX2 или обоих, так же как и экспрессию альбумина. Способ может дополнительно представить клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток, включая β-клетки, путем культивирования клеток, экспрессирующих маркеры, характерные для энтодермы поджелудочной железы/эндокринных клеток в среде, обогащенной Т3/Т4. Способы получения клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток, из плюрипотентных стволовых клеток, может использовать культуральные условия, показанные в таблицах I-III или описанных в настоящем документе. В одном из вариантов осуществления, ингибитором ALK5 является SD208 (2-(5-хлоро-2-флюорофенил)птеридин-4-ил]пиридин-4-ил-амин). В другом варианте осуществления также может применяться ингибитор ALK5 II ((2-(3-(6-метилпиридин-2-ил)-1-Н-пиразол-4-ил)-1,5-нафтиридин), ALX-270-445, ENZO, Farmingdale, NY).
Обработка клеток на стадиях 4-6 культуральной средой, обогащенной Т3/Т4, ингибитором ALK5 или обоими, дает несколько преимуществ. Например, добавление гормонов щитовидной железы на стадиях 4-6 в значительной степени снижает количество глюкагона, соматостатина и грелина, в то же время умеренно повышая экспрессию инсулина на стадии 5. Добавление Т3/Т4 на стадиях 4-6 также значительно снижает экспрессию NKX2.2, в то же время не оказывая влияния на экспрессию NKX6.1 и PDX1. Далее, добавление Т3/Т4 на стадиях 4-6 подавляет экспрессию SOX2 (маркер желудка) и альбумина (маркер печени), в то же время не влияя на экспрессию CDX2 (маркер кишечника). Более того, по сравнению с необработанным контролем, обработка Т3 на стадии 4 повышает количество НВ9-положительных клеток на стадии 6. Далее, обработка Т3 приводит к увеличению количества NKX6.1-положительных клеток, экспрессирующих НВ9. Продолжительное воздействие ингибитора ALK5 и Т3/Т4 в значительной степени повышает экспрессию НВ9, одновременно поддерживая жесткую экспрессию NKX6.1. Включение Т3/Т4 в культуральную среду, в зависимости от дозы, в значительной степени увеличивает количество НВ9 положительных клеток в NKX6.1-положительных клеток-предшественников энтодермы поджелудочной железы.
Соответственно, в некоторых вариантах осуществления, изобретение представляет способы понижения количества глюкагона, соматостатина и грелина в дифференцированных клетках, полученных на стадиях 4-6, путем обработки средой, обогащенной по меньшей мере гормонами щитовидной железы Т3/Т4. Более того, изобретение представляет методы понижения экспрессии NKX2.2 в дифференцированных клетках, полученных на стадиях 4-6 путем, которые экспрессируют NKX6.1 и PDX1, путем обработки средой, обогащенной по меньшей мере гормонами щитовидной железы Т3/Т4. В дополнение, изобретение представляет методы для увеличения количества NKX6.1-положительных клеток, экспрессирующих HB9, путем культивирования в среде с гормонами щитовидной железы Т3/Т4 и не в обязательном порядке ингибитором ALK5. Для некоторых вариантов осуществления способы применения культуральных условий показаны в таблицах I-III.
Одним из вариантов осуществления изобретения является способ формирования клеток, экспрессирующих маркеры, характерные для β-клеток, включая дифференцировку клеток, экспрессирующих маркеры, характерные для энтодермы передней кишки, в клетки, экспрессирующие маркеры, характерные для бета-клеток, путем обработки средой, обогащенной гормонами щитовидной железы Т3/Т4 и ингибитором ALK5, или обоими одновременно (например, Т3 и ингибитор ALK5). Полученные клетки являются положительными для NKX6.1, PDX1 и HB9. Способ может применяться для увеличения количества НВ9-положительных клеток среди NKX6.1-положительных клеток-предшественников энтодермы поджелудочной железы. Способ может дополнительно применяться для снижения экспрессии NKX2.2. Дополнительно, способ может быть применен для подавления экспрессии SOX2 и альбумина. Гормоном щитовидной железы может быть Т3. Метод также может быть применен для повышения экспресии НВ9, по сравнению с клетками, которые не культивировались в среде, обогащенной Т3 и ингибитором ALK5. Далее, метод включает формирование клеток, экспрессирующих маркеры, характерные для β-клеток путем культивирования клеток, экспрессирующих клеточные маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников в среде, обогащенной Т3/Т4. Способ может использовать культуральные условия, показанные в таблицах I-III или описанные в настоящем документе.
В еще одним вариантом осуществления изобретения является метод понижения количества глюкагона, соматостатина и грелина в клетках, экспрессирующих маркеры, характерные для клеток-предшественников панкреатической передней кишки, клеток, экспрессирующих маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, или клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток, в среде, обогащенной Т3/Т4 и ингибитором ALK5. Среда может быть также обогащена ингибитором SMO, SHH антагонистом, сигнализирующим о траектории (таким как SANT-1), ретиноевой кислотой и аскорбиновой кислотой. Альтернативно, среда может быть обогащена ингибитором SMO или SHH антагонистом, сигнализирующим о траектории, ретиноевой кислотой и аскорбиновой кислотой. Среда, особенно при использовании на стадии 4, может быть предпочтительно обогащена FGF7. Для некоторых вариантов осуществления в способе используются культуральные условия, показаны в таблицах I-III или описанные в настоящем документе.
В частности, в некоторых вариантах осуществления, клетки могут обрабатываться на стадиях 4-6 (т.е. на стадии 4 и стадии 5 и стадии 6, или на стадии 4 и стадии 5, или на стадии 5 и стадии 6, или на стадии 4 и стадии 6), как описано в таблице I ниже, в которой показаны примерные культуральные условия, пригодные для применения в способах настоящего изобретения. В некоторых вариантах осуществления любая обработка на одной стадии (например, стадии 4) может быть скомбинирована с любым видом обработки на других стадиях (например, стадии 5).
В альтернативном варианте осуществления настоящее изобретение представляет клеточную культуру в условиях in vitro для дифференцировки клеток, полученных из плюрипотентных стволовых клеток, в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных β-клеток, так же как и PDX1, NKX6.1 и HB9. Клеточная культура включает культуральный сосуд, среду для дифференцировки и популяцию дифференцированных клеток, полученных из плюрипотентных стволовых клеток. Клеточная культура представляет популяцию дифференцированных клеток, где по меньшей мере десять процентов дифференцированных клеток экспрессируют PDX1, NKX6.1 и НВ9. Виды среды, использованные в клеточной культуре, указаны в таблицах I-III, и предпочтительно содержат Т3/Т4 или ингибитор ALK5, или и то и другое.
Примеры культуральных условий, пригодных для применения в способах настоящего изобретения
SANT-1
Ретиноевая кислота
Аскорбиновая кислота
SANT-1
Ретиноевая кислота
Аскорбиновая кислота
SANT-1
Ретиноевая кислота
Аскорбиновая кислота
ингибитор рецептора ВМР (например, LDN-193189)
PKC активатор (например, TPB)
предпочтительно примерно 3 дня
предпочтительно примерно 3 дня
предпочтительно примерно 3 дня
В то время как Т3 в целом более предпочтителен, вместо него могут применяться другие гормоны щитовидной железы. В частности, Т4 может применяться вместо Т3, так же как и пригодные аналоги Т3 и Т4. Пригодные для применения аналоги гормонов щитовидной железы могут включать: GC-1 (Собертиром), доступный от R & D Systems, Inc. кат. № 4554; DITPA (3,5-дииодотиропропионовоая кислота); КВ-141, обсуждаемый в J. Steroid Biochem. Мол. Biol. 2008, 111; 262-267 и Proc. Natl. Acad. Sci. США 2003, 100 10067-10072; MB07344, обсужденный в Proc. Natl. Acad. Sci. США 2007, 104 15490-15495; T0681, обсужденный в PLoS One, 2010, 5e8722 и J. Lipid Res. 2009, 50; 938-944; и GC-24, обсужденный в PLoS One, 2010, e8722 и Endocr. Pract. 2012; 18(6): 954-964, содержание которого полностью включено в настоящий документ. Количества Т3, ингибитора ALK5, SZANT-1, ретиноевой кислоты, аскорбиновой кислоты, FGF7, LDN-193189 и ТРВ могу варьировать на каждой стадии. Примеры пригодных диапазонов этих компонентов показаны ниже в таблице II.
Примеры объемов культуральных компонентов, пригодных для применения в способах настоящего изобретения
от около 25 нМ до около 150 нМ для стадий 4, 5 и 6
около 25 нМ до около 100 нМ, около 50 нМ до около 150 нМ, около 50 нМ до около 100 нМ, около 25 нМ, около 50 нМ, или около 100 нМ для стадий 4, 5 и 6
около 0,4 мМ
В одном из вариантов осуществления, способы настоящего изобретения включают обработку клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки, средой, обогащенной SANT1, ретиноевой кислотой («RA»), FGF7, LDN-193189, аскорбиновой кислотой и ТРВ на протяжении примерно двух-четырех дней, предпочтительно около трех дней, для их дифференцировки в клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки. В частности, клетки стадии 3 могут быть обработаны средой, обогащенной примерно 0,25 мкМ SANT-1; примерно 100 нМ RA; примерно 2 нг/мл FGF7; примерно 100 нМ LDN-193189; и примерно 0,25 мМ аскорбиновой кислоты; и примерно около 100 нМ ТРВ, на протяжении трех дней. В одном из вариантов осуществления среда обогащена Т3, к примеру, около 1 мкМ Т3. В другом варианте осуществления среда может быть обогащена ингибитором ALK5, к примеру, около 1 мкМ ингибитора ALK5.
В альтернативном варианте осуществления способы настоящего изобретения включают обработку клеток, экспрессирующих маркеры, характерные для клеток-предшественников панкреатической передней кишки, средой, обогащенной SANT1, ретиноевой кислотой («RA»), аскорбиновой кислотой и ингибитором ALK5 на протяжении примерно двух-трех дней для их дифференцировки в клетки, экспрессирующие маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников. В некоторых вариантах осуществления среда может быть также обогащена Т3. В одном из вариантов осуществления клетки стадии 4 дифференцируются в клетки стадии 5 путем обработки клеток средой, обогащенной примерно 0,25 мкМ SANT-1, примерно 50 нМ ретиноевой кислоты, примерно 0,25 мМ аскорбиновой кислоты и примерно 500 нМ ингибитора ALK5. В другом варианте осуществления клетки стадии 4 дифференцируются в клетки стадии 5 путем обработки клеток средой, обогащенной примерно 0,25 мкМ SANT-1, примерно 50 нМ ретиноевой кислоты, примерно 0,25 мМ аскорбиновой кислоты, примерно 1 мкМ ингибитора ALK5 и 0-1000 (например, 100) нМ Т3/T4, на протяжении двух-четырех дней, предпочтительно около трех дней. В одном из вариантов осуществления клетки стадии 4, полученные согласно вариантам осуществления изобретения используются и дифференцируются в клетки стадии 5, в то время как в других вариантах осуществления могут использоваться клетки 4, полученные в соответствии с другими протоколами.
В одном из вариантов осуществления изобретения клетки, экспрессирующие маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, дифференцированные в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток, путем их обработки средой, обогащенной SANT-1, ретиноевой кислотой, аскорбиновой кислотой и либо (1) Т3/Т4 или (2) Т3/Т4 и ингибитором ALK5 на протяжении двух-четырех дней, желательно около трех дней. Например, клетки стадии 5 можно дифференцировать в клетки стадии 6 путем обработки клеток средой, обогащенной примерно 0,25 мкМ SANT-1, примерно 50 нМ ретиноевой кислоты, примерно 0,25 мМ аскорбиновой кислоты и примерно 1 мкМ ингибитора T3/T4, на протяжении трех дней. Альтернативно, клетки стадии 5 можно дифференцировать в клетки стадии 6 путем обработки клеток средой, обогащенной примерно 0,25 мкМ SANT-1, примерно 50 нМ ретиноевой кислоты, примерно 0,25 мМ аскорбиновой кислоты, примерно 500 нМ ингибитора ALK5 и 10 нМ T3/Т4, на протяжении трех дней. Альтернативно, клетки стадии 5 можно дифференцировать в клетки стадии 6 путем обработки клеток средой, обогащенной примерно 0,25 мкМ SANT-1, примерно 50 нМ ретиноевой кислоты, примерно 0,25 мМ аскорбиновой кислоты, примерно 1 мкМ ингибитора ALK5 и 0-1000 нМ T3/Т4, на протяжении трех дней. В дальнейшем клетки могут культивироваться в любой желательной среде, например на протяжении примерно 15 дней.
В одном из вариантов осуществления клетки стадии 5, полученные согласно вариантам осуществления изобретения используются и дифференцируются в клетки стадии 6, в то время как в других вариантах осуществления могут использоваться клетки 5, полученные в соответствии с другими протоколами.
Один из аспектов изобретения представляет способы увеличения экспрессии НВ9 путем обработки клеток стадий 4-6 средой, содержащей Т3/Т4 или ингибитор ALK5 или их комбинацию. Клетки стадии 4, стадии 5 и стадии 6 могут являться клетками-предшественниками панкреатической передней кишки, панкреатическими энтодермальными/эндокринными клетками-предшественниками и панкреатическими эндокринными клетками соответственно. В некоторых вариантах осуществления обработанная популяция клеток экспрессирует по меньшей мере в два раза больше HB9, чем необработанные культуры. В других вариантах осуществления уровень экспрессии инсулина испытывает положительное влияние в обработанных культурах по сравнению с необработанными культурами. Однако, экспрессии соматостатина, грелина и глюкагона понижается в обработанных культурах по сравнению с необработанными. В дополнительных вариантах осуществления клетки на стадии 5 по существу не экспрессируют CDX2 или SOX2.
В других вариантах осуществления настоящее изобретение относится к пошаговому способу дифференцировки плюрипотентных клеток, включающему культивирование клеток стадии 4-6 в среде, содержащей достаточные количества Т3/Т4 или ингибитора ALK5, или их сочетания, для создания популяции клеток панкреатической энтодермальной линии, положительных для белков NKX6.1, PDX1 и НВ9. В других вариантах осуществления по меньшей мере 5% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В других вариантах осуществления по меньшей мере 10% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В альтернативных вариантах осуществления по меньшей мере 20% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В других вариантах осуществления по меньшей мере 30% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В альтернативных вариантах осуществления по меньшей мере 40% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В других вариантах осуществления по меньшей мере 50% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В других вариантах осуществления по меньшей мере 60% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В альтернативных вариантах осуществления по меньшей мере 70% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В других вариантах осуществления по меньшей мере 80% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В других вариантах осуществления по меньшей мере 90% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9. В альтернативных вариантах осуществления по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% PDX1 и NKX6.1 со-положительных клеток экспрессируют белок НВ9.
В некоторых вариантах осуществления популяция панкреатической энтодермальной клеточной линии, состоящая из клеток, положительных относительно PDX1, NKX6.1 и белка НВ9, трансплантируется в животное, больное диабетом, для дальнейшего созревания in vivo до функциональных эндокринных клеток. В другом варианте осуществления настоящее изобретение касается клеток, экспрессирующих инсулин и NKX6.1, полученных способами настоящего изобретения. В еще одном варианте осуществления изобретение включает пошаговый процесс дифференцировки клеток-предшественников, таких как плюрипотентные стволовые клетки, в клетки панкреатической энтодермальной линии, экспрессирующие НВ9. Способы настоящего изобретения содержат один или несколько этих шагов. В частности, способ включает шаг дифференцировки плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для клеток дефинитивной энтодермы. Этот шаг может занять приблизительно три дня. Эти клетки затем дифференцируются в клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки, путем культивирования клеток при соответствующих условиях. В одном из вариантов осуществления клетки можно культивировать приблизительно два дня. Клетки, экспрессирующие маркеры, характерные для клеток кишечной трубки, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для клеток энтодермы передней кишки. Это дифференцировка могут быть осуществлена путем культивирования клеток на протяжении приблизительно двух дней. В некоторых вариантах осуществления плюрипотентные стволовые клетки представляют собой эмбриональные плюрипотентные стволовые клетки человека.
Эти клетки затем дифференцируются в клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки, которые в свою очередь могут затем дифференцироваться в клетки, экспрессирующие маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, которые в свою очередь могу затем дифференцироваться в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток. Для достижения дифференцировки в клетки панкреатической энтодермальной линии, экспрессирующие НВ9, клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки, и панкреатические энтодермальные/эндокринные клетки-предшественники могу быть культивированы с одним или несколькими ингибиторами активина (предпочтительно ингибитор ALK5), и/или Т3/Т4 гормоном щитовидной железы. В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки, и панкреатические энтодермальные/эндокринные клетки-предшественники культивируются с Т3/Т4. В другом варианте осуществления клетки, экспрессирующие маркеры, характерные для клеток-предшественников панкреатической передней кишки, и панкреатические энтодермальные/эндокринные клетки-предшественники культивируются с ингибитором рецептора активина. В альтернативном варианте осуществления клетки культивируются одновременно с ингибитором рецептора активина и Т3/Т4. Способы настоящего изобретения пригодны для многих клеток, которые могут дифференцироваться в клетки линии энтодермы поджелудочной железы, экспрессирующие НВ9. Таблица III иллюстрирует примеры культуральных условий, пригодных для применения в способах настоящего изобретения. Ниже, в таблице III, «MCX» является «композицией МСХ», «АА» - активин, «ALK5 инг.» - ингибитор ALK5, «RA» - ретиноевая кислота, «Вит. С» - аскорбиновая кислота, «инг.» - ингибитор и «акт.» - активатор. В некоторых вариантах осуществления любая обработка на одной стадии (т.е. любой из стадий 1, 2, 3, 4, 5 или 6) может быть скомбинирована с любым видом обработки на других стадиях (т.е. любой из стадий 1, 2, 3, 4, 5 или 6).
ОСТАЛЬНАЯ ЧАСТЬ СТРАНИЦЫ НАМЕРЕННО ОСТАВЛЕНА СВОБОДНОЙ
Примеры культуральных условий, пригодных для применения в вариантах осуществления методов настоящего изобретения
стадии 1
стадии 2
стадии 3
стадии 4
стадии 5
SANT-1
RA
Вит. C
FGF7
BMP рецептор инг. (например LDN-193189)
PKC акт. (например TPB)
SANT-1
RA
Вит. C
SANT-1
RA
Вит. C
В одном из вариантов осуществления настоящее изобретение представляет способ повышения экспрессии HB9 путем культивирования популяции клеток панкреатической энтодермальной линии в среде, содержащей Т3. В некоторых вариантах осуществления популяция клеток панкреатической энтодермальной линии по существу не экспрессирует CDX2 или SOX2. В других вариантах осуществления популяцию клеток панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных клеток. В дополнительных вариантах осуществления плюрипотентные клетки представляют собой эмбриональные плюрипотентные клетки человека.
В одном из вариантов осуществления настоящее изобретение представляет способ повышения экспрессии HB9 путем культивирования популяции клеток панкреатической энтодермальной линии в среде, содержащей ингибитор ALK5. В некоторых вариантах осуществления популяцию клеток панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки представляют собой эмбриональные плюрипотентные клетки человека.
В предпочтительном варианте осуществления настоящее изобретение представляет способ повышения экспрессии HB9 путем культивирования популяции клеток панкреатической энтодермальной линии в среде, содержащей ингибитор ALK5 и Т3. В некоторых вариантах осуществления популяцию клеток панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных клеток. В дополнительных вариантах осуществления плюрипотентные клетки представляют собой эмбриональные плюрипотентные клетки человека.
В другом варианте осуществления изобретение относится к способу повышения экспрессии НВ9 в PDX1 и NKX6.1 со-экспрессирующих клеток путем обработки таких клеток в среде, содержащей достаточный объем Т3, ингибитора ALK5 или их сочетания.
Один из вариантов осуществления настоящего изобретения является способом получения клеток, экспрессирующих маркеры, характерные для β-клеток, из плюрипотентных стволовых клеток, включающий следующие стадии: (а) культивирование плюрипотентных стволовых клеток; (b) дифференцировка плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для клеток энтодермы передней кишки; и (с) дифференцировка клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки, в клетки, экспрессирующие маркеры, характерные для β-клеток, путем обработки средой, обогащенной Т3/Т4, ингибитором ALK5 или обоими. Полученные клетки могут быть положительными для NKX6.1, PDX1 и HB9. Способ может применяться для увеличения количества НВ9-положительных клеток среди NKX6.1-положительных клеток, экспрессирующих маркеры, характерные для клеток-предшественников энтодермы поджелудочной железы. Способ также может применяться для снижения экспрессии NKX2.2. Более того, способ подавляет экспрессию SOX2 и альбумина. Далее, способ может применяться для повышения урожайности клеток, экспрессирующих инсулин.
В одном из вариантов осуществления изобретения применен T3. Способ может включать культивацию клеток в среде, обогащенной Т3 и ингибитором ALK5. Способ также может быть применен для повышения экспрессии НВ9, по сравнению с клетками, которые не культивировались в среде, обогащенной Т3 и ингибитором ALK5. Среда может быть также обогащена одним или несколькими следующими компонентами (т.е. 1, 2, 3 или всеми): ингибитором SMO, SHH антагонистом, сигнализирующим о траектории (таким как SANT-1), ретиноевой кислотой и аскорбиновой кислотой. В одном из вариантов осуществления способ обеспечивает клетки, экспрессирующие маркеры, характерные для β-клеток, путем культивирования клеток, экспрессирующих маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, в среде, обогащенной Т3, которая также может быть позднее обогащена ингибитором ALK5.
Другим вариантом осуществления изобретения является способ формирования клеток, экспрессирующих маркеры, характерные для β-клеток, включая дифференцировку клеток, экспрессирующих маркеры, характерные для энтодермы передней кишки, в клетки, экспрессирующие маркеры, характерные для β-клеток, путем обработки средой, обогащенной Т3/Т4 и ингибитором ALK5, или обоими. В некоторых вариантах осуществления среда далее обогащается ингибитором рецептора ВМР и активатором РКС. Полученные клетки являются предпочтительно положительными для NKX6.1, PDX1 и HB9. Способ может применяться для увеличения количества НВ9-положительных клеток среди NKX6.1-положительных клеток-предшественников энтодермы поджелудочной железы, снижения экспрессии NKX2.2 и/или подавления экспрессии SOX2 и альбумина. В предпочтительных вариантах осуществления изобретения применен T3. Способ может также включать культивацию клеток в среде, обогащенной Т3 и ингибитором ALK5. Способ также может быть применен для повышения экспрессии НВ9, по сравнению с клетками, которые не культивировались в среде, обогащенной Т3 и ингибитором ALK5. Более того, способ включает формирование клеток, экспрессирующих маркеры, характерные для β-клеток путем культивирования клеток, экспрессирующих клеточные маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников в среде, обогащенной Т3 и ингибитором ALK5.
Еще одним вариантом осуществления изобретения является способ повышения экспрессии НВ9 и подавления экспрессии SOX2 и альбумина путем культивирования клеток, экспрессирующих маркеры, характерные для клеток-предшественников панкреатической передней кишки, в среде, обогащенной Т3/Т4 и ингибитором ALK5. Альтернативным вариантом осуществления изобретения является способ понижения количества глюкагона, соматостатина и грелина в клетках, экспрессирующих маркеры, характерные для клеток-предшественников панкреатической передней кишки, клеток, экспрессирующих маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, или клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток, в среде, обогащенной Т3/Т4 и ингибитором ALK5. Среда может быть также обогащена одним или несколькими из следующих компонентов: ингибитором SMO, SHH антагонистом, сигнализирующим о траектории (таким как SANT-1), ретиноевой кислотой и аскорбиновой кислотой. В одном из вариантов осуществления клетки являются клетками стадии 4, а среда обогащена FGF7.
Настоящее изобретение также представляет клетку или популяцию клеток, которые можно получить способами настоящего изобретения. Настоящее изобретение также представляет клетку или популяцию клеток, полученную способами настоящего изобретения.
Изобретение представляет способы обработки. В частности, настоящее изобретение представляет способ лечения пациента, страдающего диабетом или подвергающегося риску развития данного заболевания.
Настоящее изобретение также представляет клетку или популяцию клеток, которые можно получить или полученные способом настоящего изобретения для применения в способе лечения. В частности, настоящее изобретение также представляет клетку или популяцию клеток, которые можно получить или полученные способом настоящего изобретения для применения в способе лечения пациента, страдающего диабетом или подвергающегося риску развития данного заболевания.
Диабет может быть диабетом типа I или типа II.
В одном из вариантов осуществления метод лечения включает имплантацию клеток, которые получены или могут быть получены способом настоящего изобретения, пациенту.
В одном из вариантов осуществления способ лечения включает
дифференцировку плюрипотентных клеток in vitro в клетки стадии 1, стадии 2, стадии 3, стадии 4, стадии 5 или стадии 6, например как описано в настоящем документе,
и имплантации дифференцированных клеток пациенту.
В одном из вариантов осуществления способ также содержит стадию культивирования плюрипотентных стволовых клеток, например как описано в настоящем документе, предшествующую стадии дифференцировки плюрипотентных стволовых клеток.
В одном из вариантов осуществления способ также включает стадию дифференцировки клеток in vivo, после стадии имплантации.
В одном из вариантов осуществления пациент является млекопитающим, предпочтительно человеком.
В одном из вариантов осуществления клетки можно имплантировать в форме дисперсных клеток или клеток, образующих кластеры, которые можно вводить в воротную вену печени способом инфузии. Клетки можно альтернативно вводить в биосовместимые разлагающиеся полимерные опорные материалы, пористые неразлагающиеся устройства или в инкапсулированном виде для защиты от иммунного ответа организма-хозяина. Клетки можно имплантировать в подходящее место в организме реципиента. Места имплантации включают в себя, например, печень, естественную поджелудочную железу, пространство под почечной капсулой, сальник, брюшную полость, субсерозное пространство, кишечник, желудок или подкожный карман.
Для стимуляции дальнейшего дифференцирования, выживаемости или активности имплантированных клеток in vivo, до, одновременно с или после введения клеток можно вводить дополнительные факторы, такие как факторы роста, антиоксиданты или противовоспалительные вещества. Данные факторы могут секретироваться эндогенными клетками и воздействовать на внедренные клетки in situ. Дифференцирование имплантированных клеток можно индуцировать любой комбинацией эндогенных и введенных экзогенно факторов роста, известных в данной области.
Количество клеток, используемых при имплантации, зависит от ряда различных факторов, включая состояние пациента и его реакцию на лечение, и может быть определено специалистом в данной области.
В одном из вариантов осуществления способ лечения также включает встраивание клеток перед имплантацией в трехмерный опорный материал. Клетки можно поддерживать in vitro на данном опорном материале перед имплантацией пациенту. Опорный материал, содержащий клетки, альтернативно можно имплантировать непосредственно в организм пациента без дополнительного культивирования in vitro. В опорный материал может быть необязательно включено по меньшей мере одно фармацевтическое вещество, обеспечивающее выживаемость и функционирование трансплантированных клеток.
Публикации, цитируемые в настоящем документе, полностью включаются в настоящий документ посредством ссылки. Настоящее изобретение дополнительно иллюстрируется, среди прочего, следующими примерами.
ПРИМЕРЫ
Пример 1
Ранее опубликованные протоколы, генерирующие популяции энтодермы поджелудочной железы, полученных из плюрипотентных клеток человека, по существу не экспрессируют белок HB9
Этот пример направлен на идентификацию паттерна экспрессии НВ9 в клетках, полученных из плюрипотентных стволовых клеток, как описано в данном примере. Эмбриональные стволовые клетки человека линии H1 (пассаж 40) рассеивали как одиночные клетки при концентрации 1×105 клеток/см2 на чашки, покрытые MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси), в среде MTESR®1 (StemCell Technologies, г. Ванкувер, Канада) с добавлением 10 мкМ Y27632 (Rock inhibitor, Catalog No. Y0503, Sigma-Aldrich, St. Louis, MO). Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры были затем дифференцированы в панкреатические эндодермальные/эндокринные клетки-предшественники, как описывалось ранее в Diabetes, 61, 2016, 2012. Примененный протокол дифференцировки был следующим:
a. 60-70% конфлюэнтных прикрепленных к субстрату культур недифференцированных клеток Н1, размещенных на поверхностях, покрытых MATRIGEL™, разведенным в соотношении 1:30, были помещены в среду RPMI 1640 (Invitrogen), обогащенную 0,2% эмбриональной бычьей сыворотки (FBS) (Hyclone, Utah), 100 нг/мл активина-A (AA; Pepro-tech; Rocky Hill, NJ), и 20 нг/мл Wnt3A (R&D Systems), только на один день. В течение последующих двух дней клетки культивировались в среде RPMI с 0,5% FBS и 100 нг/мл АА.
b. Клетки, полученные на стадии (а) были помещены в среду DMEM-F12 (Invitrogen), обогащенную 2% FBS и 50 нг/мл FGF7 (Pepro-tech), на три дня.
с. Культуры, полученные на стадии (b) продолжались еще четыре дня в среде DMEM-HG (Invitrogen), обогащенной 0,25 мкМ SANT-1 (Sigma-Aldrich; St. Louis, MO), 2 мкМ ретиноевой кислоты (Sigma-Aldrich), 100 нг/мл Noggin (R&D Systems), и 1% (об/об) добавки, которая продается под торговой маркой B27® (№ кат. 17504044, Life Technologies Corporation, Grand Island, NY).
d. Клетки, полученные на стадии (с) культивировались на протяжении трех дней в среде DMEM-HG, обогащенной 1мкМ ингибитора ALK5 (ALK5i; Farmingdale, NY), 100 нг/мл Noggin, 50 нМ ТРВ ((2S,5S)-(E,E)-8-(5-(4-(трифлюорометил)фенил)-2,4-пентадиенойламино)бензолактам; EMD Chemicals Inc., Gibbstown NJ) и 1% В27 в однослойном формате. Для последнего дня культивирования клетки были обработаны 5 мг-мл диспазы на протяжении 5 минут, после чего осторожно пипетированы для смешивания и разбивки на клеточные кластеры (< 100 микрон). Клеточные кластеры были перенесены в одноразовые полистироловые центрифужные пробирки емкостью 125 мл (Corning) и вращались со скоростью 80-100 об/мин на протяжении ночи в суспензии со средой DMEM-HG, обогащенной 1мкМ ингибитора ALK5, 100 нг/мл Noggin и 1% В27.
В конце стадии (d) мРНК было собрана для ПЦР анализа на релевантные панкреатические энтодермальные / эндокринные гены. Общая РНК был экстрагирована с помощью RNeasy® Mini Kit (Qiagen; Valencia, CA) и обратнотранскрибирована с помощью High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) в соответствии с инструкциями производителя. кДНК была усилена с помощью Taqman® Universal Master Mix and Taqman® Gene Expression Assays, которые были предварительно загружены в Taqman® Arrays (Applied Biosystems). Данные были проанализированы с применением ПО Sequence Detection Software (Applied Biosystems) и нормализованы относительно недифференцированных эмбриональных стволовых клеток человека (hES) с помощью способа ΔΔCt (т.е., результаты ПЦР были откорректированы относительно внутренних контролей (ΔΔCt = ΔCtобразец - ΔCtсправочное)). Все праймеры приобретали в Applied Biosystems. Цитометрия и иммунофлюоресцентный анализ были осуществлены в соответствии с предшествующим описанием (Diabetes, 61, 20126, 2012). Антитело НВ9 было получено от Developmental Studies Hybridoma Bank (University of Iowa, Iowa). Применительно к примерам, Y27632 ((1R,4r)-4-((R)-1-аминоэтил)-N-(пиридин-4-ил)циклогексанкарбоксамид) является клеткопроницающей малой молекулой Rho-ассоциированного ингибитора киназы (ROCK).
На фиг. 1A–1С представлены данные анализов ПЦР в реальном времени по экспрессии следующих генов в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных в панкреатические энтодермальные/эндокринные клетки-предшественники, как описано в примере 1: PDX1 (ФИГ. 1A), NKX6.1 (ФИГ. 1B) и HB9 (ФИГ. 1C). Как показано на ФИГ. 1, жесткая мРНК экспрессия PDX1, NKX6.1 и НВ9 была обнаружена в этих культурах. Кроме того, мРНК-экспрессия НВ9 в этих клетках была эквивалентна или выше, в сравнении с человеческими трупными островками. Однако, как показано на ФИГ. 2, где данные по генной экспрессии PDX1 и NKX6.1 были в соответствии с высокой экспресией корреспондирующих белков, измеренной методом цитометрии, мРНК-экспрессия НВ9 не соответствовало белковой экспрессии НВ9. В день, следующий за завершением стадии (d), т.е. на день 5, приблизительно 1% клеток были положительны для НВ9, в то время как приблизительно 50% клеток были NKX6.1-положительными и приблизительно 90% были PDX1-положительными. Иммуноокрашивание клеточных кластеров также подтвердило данные поточной цитометрии. Как изображено на фиг. 3А-В, значительное количество NRX6.1-положительных клеток и несколько инсулин-положительных клеток присутствовало в кластерах. Однако, иммуноокрашивание не выявило HB9-положительных клеток (ФИГ. 3В).
Пример 2
Добавление Т3 на стадиях 4-6 повышает количество НВ9-положительных клеток
Этот пример направлен на добавление Т3 на стадиях 4-6 для существенного повышения количества НВ9-положительных клеток.
Эмбриональные стволовые клетки человека линии H1 (пассаж 40) рассеивали как одиночные клетки при концентрации 1 X 105 клеток/см2 на чашки, покрытые MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси) в среде mTeSR®1 с добавлением 10 мкМ Y27632. Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры были дифференцированы в клетки, экспрессирующие маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, согласно протоколу, описанному ниже.
а. Стадия 1 (3 дня): Стволовые клетки культивировали в течение одного дня в: среде MCDB-131 (Invitrogen, № по каталогу 10372-019) с добавкой 2%-го БСА без жирных кислот (Proliant, № по каталогу 68700), 0,0012 г/мл бикарбоната натрия (SigmaAldrich, № по каталогу S3187), 1X GlutaMax™ (Invitrogen, № по каталогу 35050-079), 4,5 мМ D-глюкозы (SigmaAldrich, № по каталогу G8769), 100 нг/мл GDF8 (R&D Systems) и композиции 1 мкМ MCX. Затем клетки культивировали на протяжении дополнительного дня в среде MCDB-131 с добавлением 2% не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы, 100 нг/мл GDF8 и 0,1 мкМ композиции MCX. Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 2% не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы и 100 нг/мл GDF8.
b. Стадия 2 (2 дня): Клетки стадии 1 на протяжении двух дней обрабатывались средой MCDB-131, обогащенной 2% BSA без жирных кислот, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы, 0,25 мМ аскорбиновой кислоты (Sigma, MO) и 25 нг/мл FGF7 (R & D Systems, MN).
c. Стадия 3 (2 дня): Клетки стадии 2 были затем обработаны средой MCDB-131, обогащенной 1:200 раствором ITS-X (Gibco® Инсулин,-Трансферрин-Селений-Этаноламин; Invitrogen, Ca); 4.5 мМ глюкозой; 1X GlutaMax™; 0.0017 г/мл бикарбоната натрия; 2% BSA без жирных кислот; 0.25 мкМ SANT-1 (Sigma, MO); 1 мкМ RA (Sigma, MO); 25 нг/мл FGF7; 0.25 мМ аскорбиновой кислоты; 200 нМ TPB (PKC активатор; № по каталогу 565740; EMD Chemicals, Gibbstown, NJ); и 100 нМ LDN (ингибитор рецептора BMP; № по каталогу 04-0019; Stemgent), на протяжении двух дней.
d. Стадия 4 (3 дня): Клетки стадии 3 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0017 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 нМ SANT-1, 100 мкМ ретиноевой кислоты, 2 нг/мл FGF7; 100 нМ LDN-193189; 0,25 мМ аскорбиновой кислоты и 100 нМ TPB в течение трех дней.
е. Стадия 5 (3 дня): Клетки стадии 4 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0015 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 нМ SANT-1, 50 нМ ретиноевой кислоты, 0,25 мМ аскорбиновой кислоты и 500 нМ ингибитора ALK5 SD208 в течение трех дней. SD208 является 2-(5-хлоро-2-флюорофенил)птеридин-4-ил]пиридин-4-ил-амин), обладающим структурой формулы I, и обнародованным в Molecular Pharmacology 2007, 72:152-161. SD208 является 2,4-двухзамещенным птеридином, ATP-конкурентным ингибитором TGF-βR I киназы.
(I)
f. Стадия 6 (3-15 дней): Клетки этапа 5 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0015 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 мкМ SANT-1, 50 нМ RA, 0,25 мМ аскорбиновой кислоты в течение трех дней.
В некоторых культурах на стадиях 4-6 был добавлен 1 мкМ Т3 (T6397, Sigma, MO). По окончании стадий 4-6 контроль и обработанные культуры были проанализированы методами цитометрии и иммуноокращивания. Далее, мРНК была собрана с контролей и обработанных культур на стадиях 2-6.
Фигура 4 описывает данные цитометрии на стадии 4 (ФИГ. 4А), стадии 5 (ФИГ. 4В) и стадии 6 (ФИГ. 4С) для PDX1, NKX6.1 и НВ9. Они соответствуют данным из примера 1; хотя присутствовало значительное количество PDX1 и NKX6.1-положительных клеток на стадиях 4-6, экспрессия НВ9 была значительно ниже. Экспрессия НВ9 достигла пика на стадии 5 и снизилась на стадии 6. В целом, экспрессия НВ9 для клеток, полученных с использованием протокола, описанного в примере 2, была выше по сравнению с клетками, полученными с использованием протокола, описанного в примере 1. ФИГ. 5А демонстрирует мРНК экспрессию НВ9 в сравнении с человеческими островками на стадиях 2-6. Подобно примеру 1, хотя уровень мРНК экспрессии НВ9 на стадиях 3-4 был эквивалентен человеческим островкам, экспрессия белка НВ9 была очень низкой на стадии 4 (ФИГ. 5В).
На фиг. 6А–6J представлены данные ПЦР в реальном времени при анализе экспрессии следующих генов в эмбриональных стволовых клетках человека линии Н1, дифференцированных до стадии 4, как описано в примере 2, а затем обработаны только на стадии 4, на стадии 4 до стадии 5, или на стадии 4 до стадии 6: NKX6.1 (ФИГ. 6A); PDX1 (ФИГ. 6B); NKX2.2 (ФИГ. 6C); глюкагон (ФИГ. 6D); инсулин (ФИГ. 6E); соматостатин (ФИГ. 6F); CDX2 (ФИГ. 6G); альбумин (ФИГ. 6H); гастрин (ФИГ. 6I); и SOX2 (ФИГ. 6J). Добавление Т3 на стадиях 4-6 в значительной степени снижает количество глюкагона, соматостатина и грелина, в то же время умеренно повышая экспрессию инсулина на стадии 5. Добавление Т3 на стадиях 4-6 также значительно снижает экспрессию NKX2.2, в то же время не оказывая влияния на экспрессию NKX6.1 и PDX1. Далее, добавление Т3 подавляет экспрессию SOX2 (маркер желудка) и альбумина (маркер печени), в то же время не влияя на экспрессию CDX2 (маркер кишечника). Иммуноокрашивание контроля и обработанных культур на стадии 6 позволило обнаружить значительное увеличение количества НВ9 положительных клеток в группе, обработанной Т3 (ФИГ. 7В) по сравнению с контролем (ФИГ. 7А) на стадии 6. Более того, возросшее количество NKX6.1-положительных клеток показало экспрессию НВ9 в культурах, обработанных Т3..
Пример 3
Комбинированная обработка Т3 и ингибитором ALK5 на стадии 6 повышает экспрессию НВ9
Этот пример демонстрирует, что комбинация ингибитора ALK5 и Т3 в среде на стадии 6 существенно повышает экспрессию НВ9.
Эмбриональные стволовые клетки человека линии H1 (пассаж 40) рассеивали как одиночные клетки при концентрации 1×105 клеток/см2 на чашки, покрытые MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси) в среде mTeSR®1 с добавлением 10 мкМ Y27632. Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры дифференцировали в линии панкреатических энтодермальных/эндокринных клеток согласно протоколу, описанному ниже:
а. Стадия 1 (3 дня): Клетки культивировали в течение одного дня в: среде MCDB-131 (Invitrogen, № по каталогу 10372-019) с добавкой 2%-го БСА без жирных кислот (Proliant, № по каталогу 68700), 0,0012 г/мл бикарбоната натрия (SigmaAldrich, № по каталогу S3187), 1X GlutaMax™ (Invitrogen, № по каталогу 35050-079), 4,5 мМ D-глюкозы (SigmaAldrich, № по каталогу G8769), 100 нг/мл GDF8 (R&D Systems) и композиции 1 мкМ MCX. Затем клетки культивировали на протяжении дополнительного дня в среде MCDB-131 с добавлением 2% не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы, 100 нг/мл GDF8 и 0,1 мкМ композиции MCX. Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 2% не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы и 100 нг/мл GDF8.
b. Стадия 2 (2 дня): Клетки стадии 1 обрабатывались средой MCDB-131, обогащенной 2% BSA без жирных кислот, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы, 0,25 мМ аскорбиновой кислоты и 25 нг/мл FGF7 в течение двух дней.
c. Стадия 3 (2 дня): Клетки стадии 2 были затем обработаны средой MCDB-131, обогащенной 1:200 раствором ITS-X (Invitrogen, Ca); 4.5 мМ глюкозой; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2% BSA без жирных кислот; 0.25 мкМ SANT-1 (Sigma, MO); 1 мкМ RA (Sigma, MO); 25 нг/мл FGF7; 0.25 мМ аскорбиновой кислоты; 200 нМ TPB (PKC активатор; № по каталогу 565740; EMD Chemicals, Gibbstown, NJ); и 100 нМ LDN (ингибитор рецептора BMP; № по каталогу 04-0019; Stemgent), на протяжении двух дней.
d. Стадия 4 (3 дня): Клетки стадии 3 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0017 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 мкМ SANT-1, 100 нМ ретиноевой кислоты, 2 нг/мл FGF7, 100 нМ LDN-193189, 0.25 мМ аскорбиновой кислоты, 100 нМ TPB и 1 мкМ T3 в течение трех дней.
е. Стадия 5 (3 дня): Клетки стадии 4 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0015 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 мкМ SANT-1, 50 нМ ретиноевой кислоты, 0,25 мМ аскорбиновой кислоты, 1 мкМ ингибитора ALK5 SD208 и 100 нМ T3 в течение трех дней.
f. Стадия 6 (3-15 дней): Клетки стадии 5 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0015 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 нМ SANT-1, 500 нМ ингибитора ALK5, 50 нМ ретиноевой кислоты, 0,25 мМ аскорбиновой кислоты и 10 нМ T3 в течение трех дней.
На фиг. 8А и 8В показано иммуноокрашивание для NKX6.1 и НВ9 на стадии 6, день 7. ФИГ 8С описывает данные анализов ПЦР в реальном времени по экспрессии НВ9 в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных на стадии 6 как описано в примере 3. мРНК экспрессия НВ9 вместе с иллюстрациями иммуноокрашивания показывает, что длительное применение ингибитора ALK5 и Т3 существенно повышает экспрессию НВ9, одновременно поддерживая жесткую экспрессию NKX6.1. На фиг. 9А и 9В описаны данные цитометрического анализа на стадии 6, день 5 и день 15 соответственно. Значительная фракция клеток стадии 6 демонстрирует экспрессию НВ9 в культурах на стадии 6, день 15.
Пример 4
Т3 в зависимости от дозы повышает экспрессию НВ9
Этот пример демонстрирует, что в зависимости от дозы Т3 может применяться для повышения экспрессии НВ9, одновременно поддерживая экспресиию NKX6/1 на стадии 6. Эмбриональные стволовые клетки человека линии H1 (пассаж 40) рассеивали как одиночные клетки при концентрации 1×105 клеток/см2 на чашки, покрытые MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси) в среде mTeSR®1 с добавлением 10 мкМ Y27632. Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры были дифференцированы в клетки, экспрессирующие маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, согласно протоколу, описанному ниже.
а. Стадия 1 (3 дня): Клетки культивировали в течение одного дня в: среде MCDB-131 (Invitrogen, № по каталогу 10372-019) с добавкой 2%-го БСА без жирных кислот (Proliant, № по каталогу 68700), 0,0012 г/мл бикарбоната натрия (SigmaAldrich, № по каталогу S3187), 1X GlutaMax™ (Invitrogen, № по каталогу 35050-079), 4,5 мМ D-глюкозы (SigmaAldrich, № по каталогу G8769), 100 нг/мл GDF8 (R&D Systems) и композиции 1 мкМ MCX. Затем клетки культивировали на протяжении дополнительного дня в среде MCDB-131 с добавлением 2% не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы, 100 нг/мл GDF8 и 0,1 мкМ композиции MCX. Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 2% не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы и 100 нг/мл GDF8.
b. Стадия 2 (2 дня): Клетки стадии 1 на протяжении двух дней обрабатывались средой MCDB-131, обогащенной 2% BSA без жирных кислот, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мМ D-глюкозы, 0,25 мМ аскорбиновой кислоты (Sigma, MO) и 25 нг/мл FGF7 (R & D Systems, MN).
c. Стадия 3 (2 дня): Клетки стадии 2 были затем обработаны средой MCDB-131, обогащенной 1:200 раствором ITS-X (Invitrogen, Ca); 4.5 мМ глюкозой; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2% BSA без жирных кислот; 0.25 мкМ SANT-1 (Sigma, MO); 1 мкМ RA (Sigma, MO); 25 нг/мл FGF7; 0.25 мМ аскорбиновой кислоты; 200 нМ TPB (PKC активатор; № по каталогу 565740; EMD Chemicals, Gibbstown, NJ); и 100 нМ LDN (ингибитор рецептора BMP; № по каталогу 04-0019; Stemgent), на протяжении двух дней.
d. Стадия 4 (3 дня): Клетки этапа 3 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0017 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 мкМ SANT-1, 100 нМ ретиноевой кислоты, 2 нг/мл FGF7, 100 нМ ингибитора LDN-193189, 0,25 мМ аскорбиновой кислоты и 100 нМ TPB, в течение трех дней.
е. Стадия 5 (3 дня): Клетки этапа 4 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0015 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 мкМ SANT-1, 50 нМ ретиноевой кислоты, 0,25 мМ аскорбиновой кислоты, 1 мкМ ингибитора ALK5 SD208 и 0-1000 нМ T3, в течение трех дней.
f. Стадия 6 (6 дней): Клетки этапа 5 обрабатывались средой MCDB-131, обогащенной 1:200 раствором ITS-X, 4,5 мМ глюкозы, 1X GlutaMax™, 0,0015 г/мл бикарбоната натрия, 2% BSA без жирных кислот, 0,25 нМ SANT-1, 500 мкМ ингибитора ALK5, 50 нМ ретиноевой кислоты,0,25 мМ аскорбиновой кислоты и 0-1000 нМ T3, в течение шести дней.
На фиг. 10А-10Е показано иммуноокрашивание для NKX6.1 и НВ9 на стадии 6, день 6. В зависимости от дозы Т3 в значительной степени увеличил количество НВ9-положительных клеток среди NKX6.1-положительных клеток-предшественников энтодермы поджелудочной железы. На фиг. 11A–11L представлены данные анализов ПЦР в реальном времени по экспрессии следующих генов в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных на стадии 6 как описано в примере 4: SOX2 (ФИГ. 11A); NKX6.1 (ФИГ. 11B); NKX2.2 (ФИГ. 11C); гастрин (ФИГ. 11D), PDX1 (ФИГ. 11E); NGN3 (ФИГ. 11F); PAX6 (ФИГ. 11G); PAX4 (ФИГ. 11Н); инсулин (ФИГ. 11I); глюкагон (ФИГ. 11J); грелин (ФИГ. 11К) и соматостатин (ФИГ. 11L).
Хотя изобретение было описано и проиллюстрировано ссылками на различные конкретные материалы, процедуры и примеры, следует понимать, что изобретение не ограничивается конкретными комбинациями материалов и процедур, выбранными для этой цели. Многочисленные изменения, касающиеся таких деталей изобретения, будут очевидными для специалистов в данной области техники. Подразумевается, что все описания и примеры представлены только в качестве иллюстрации и находятся в пределах сущности и объема настоящего изобретения, что отражено в нижеприведенной формуле изобретения. Все ссылки, патенты и заявки на патенты, приведенные в этой заявке, полностью включены в этот документ путем отсылки.
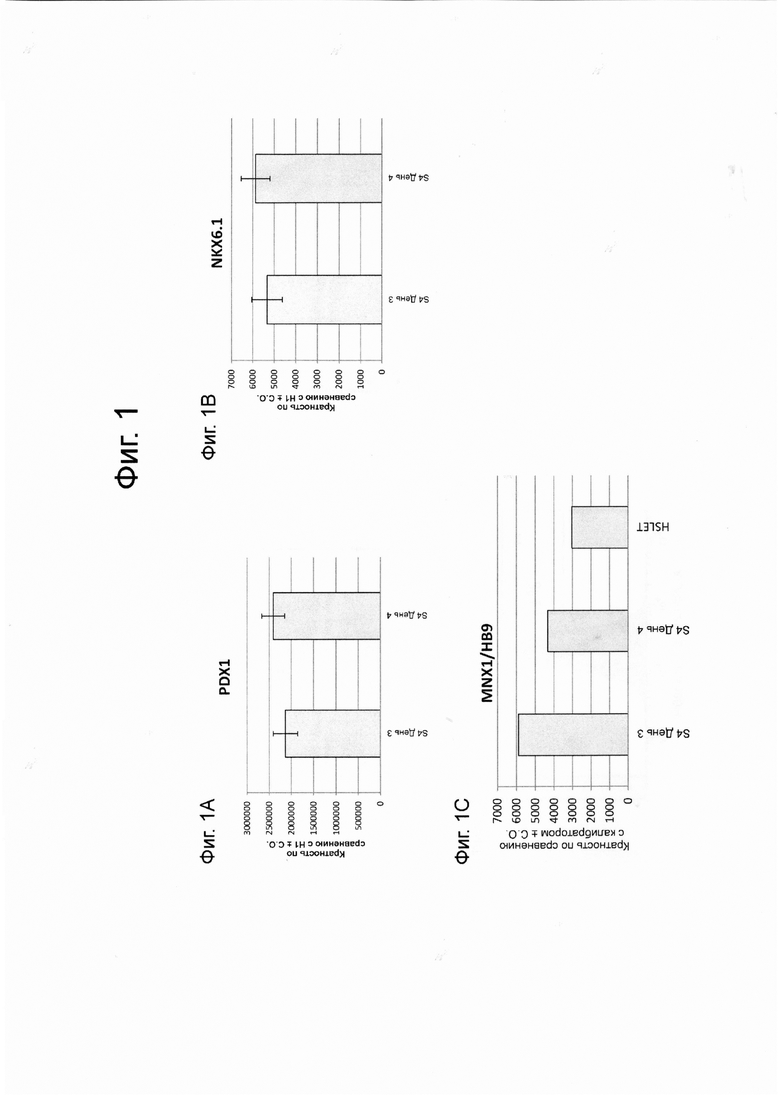
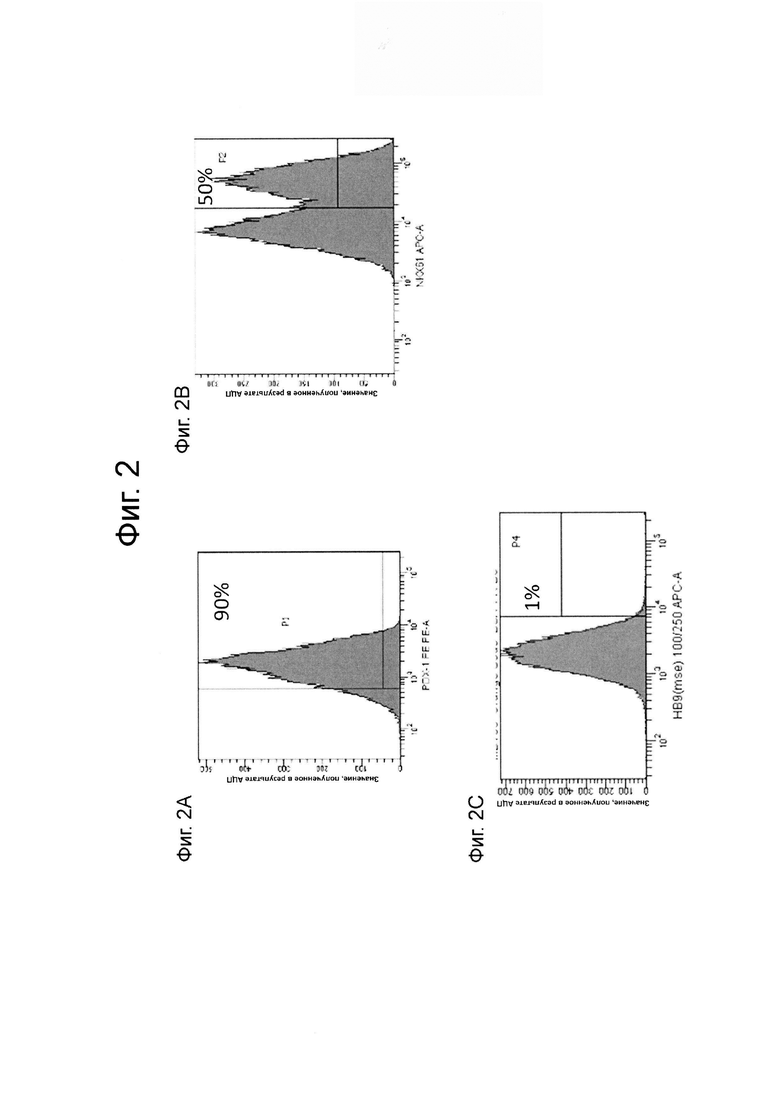
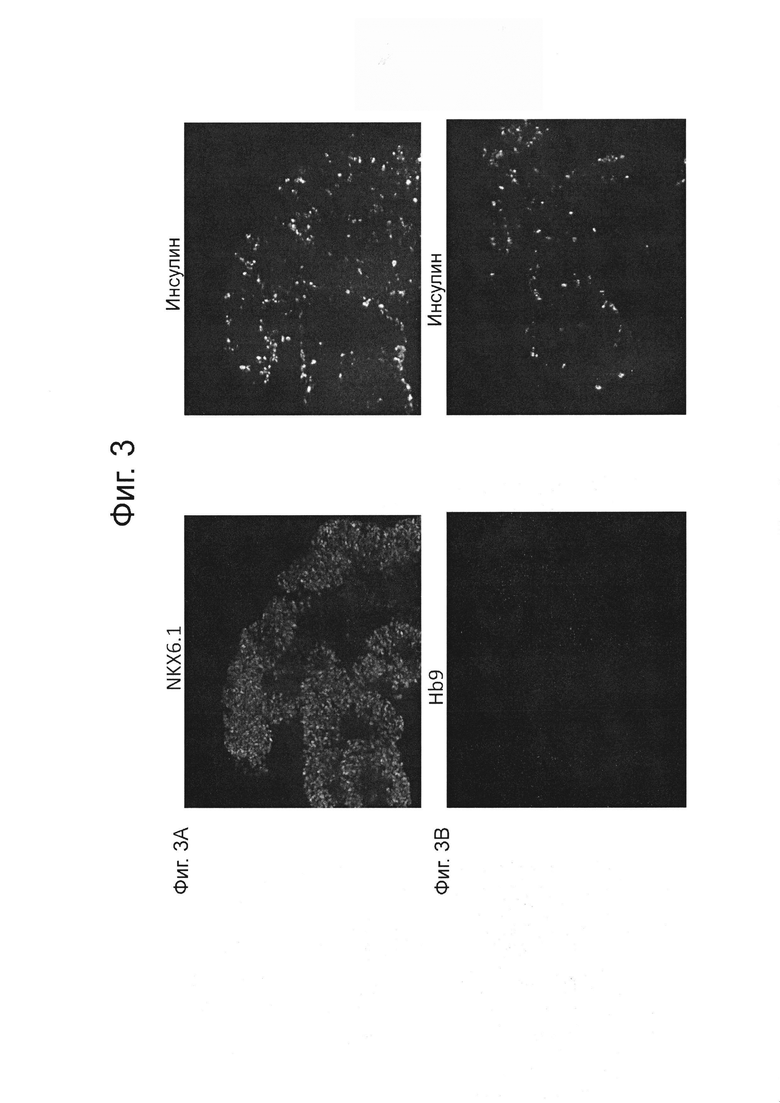
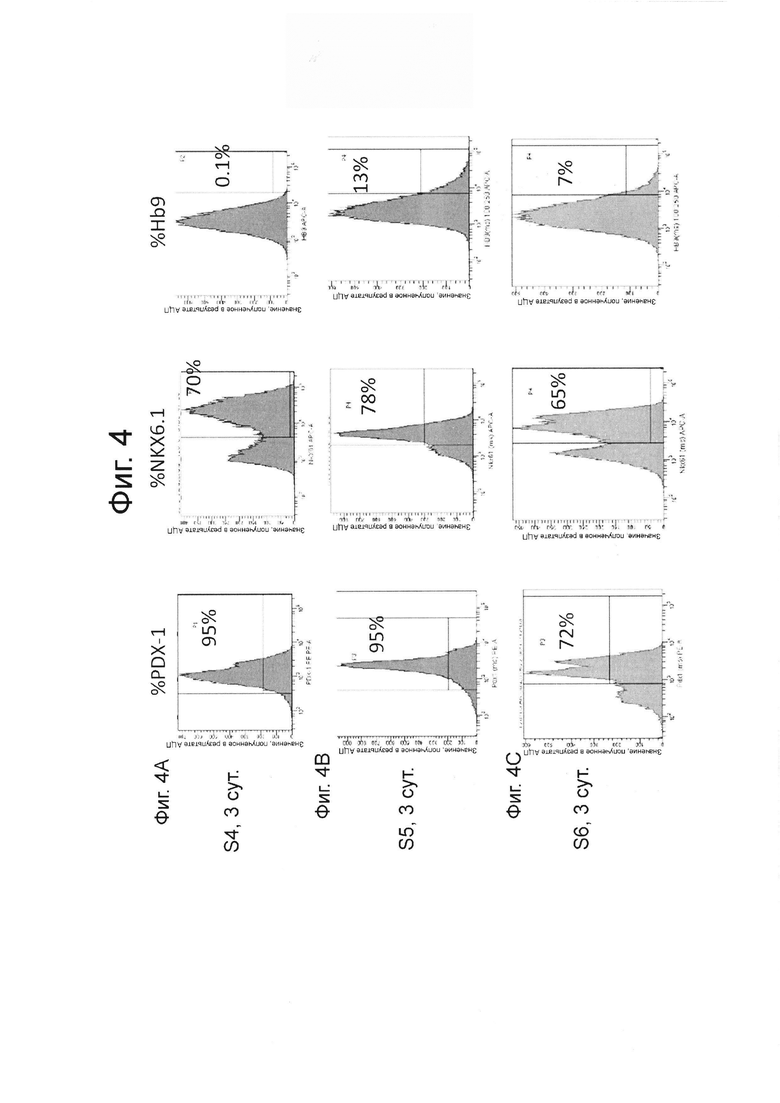
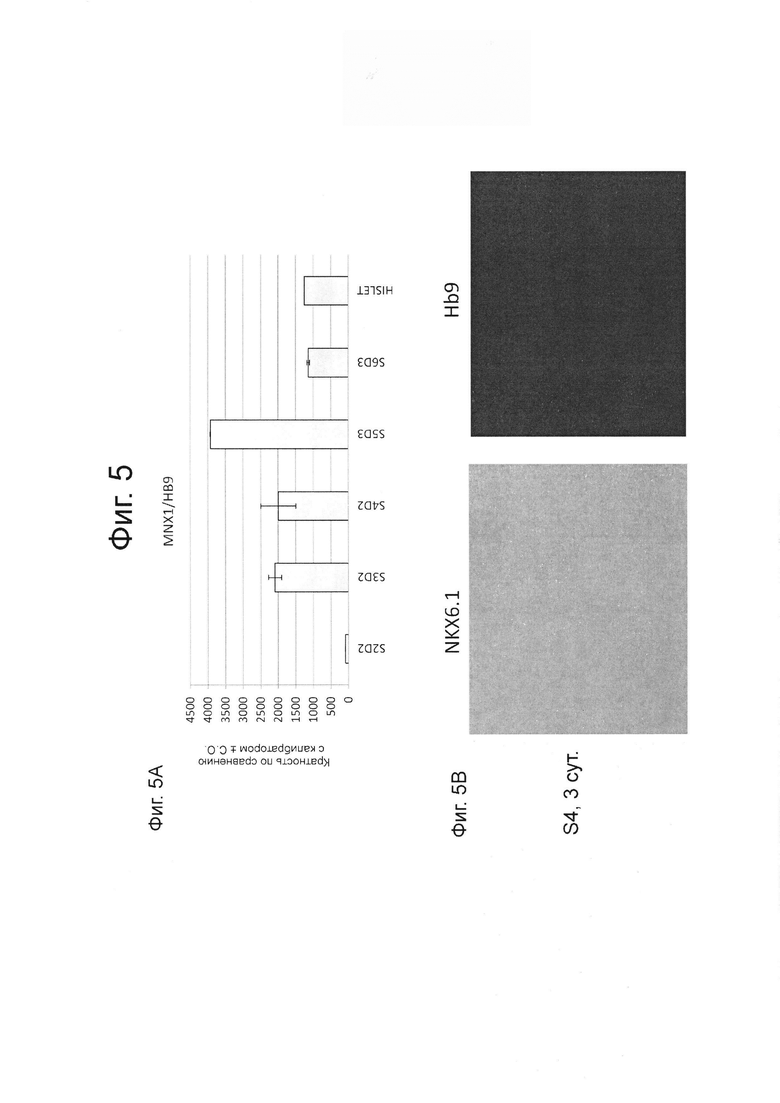
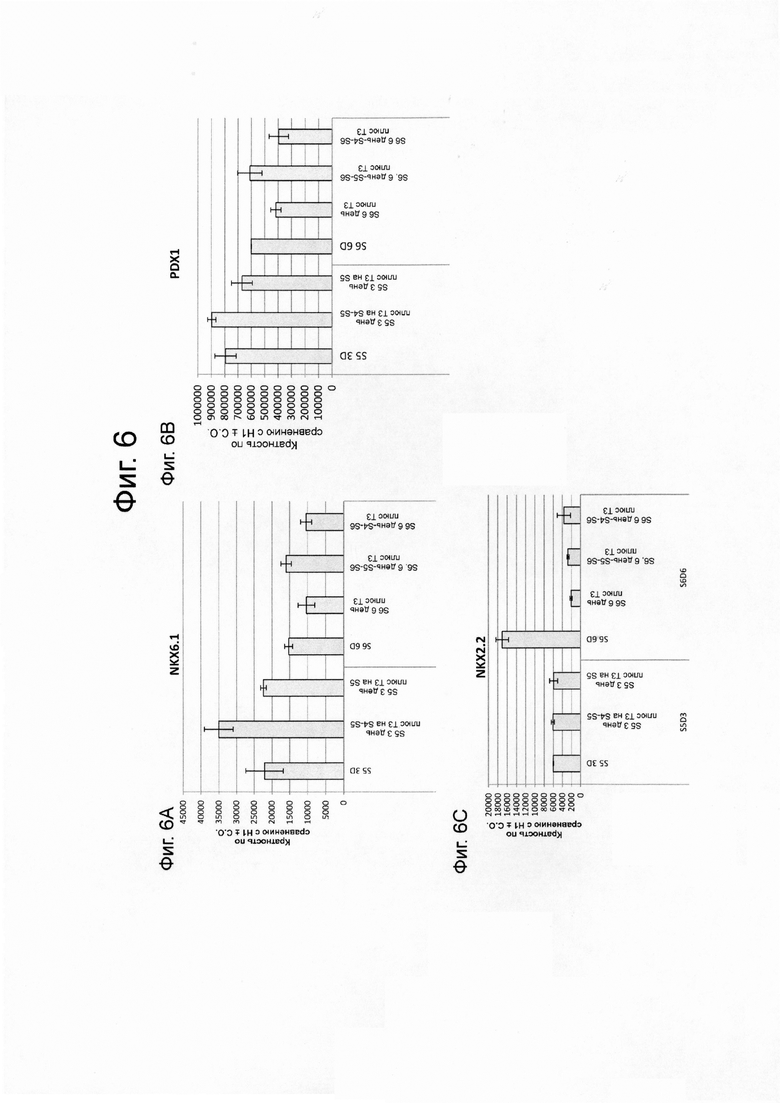
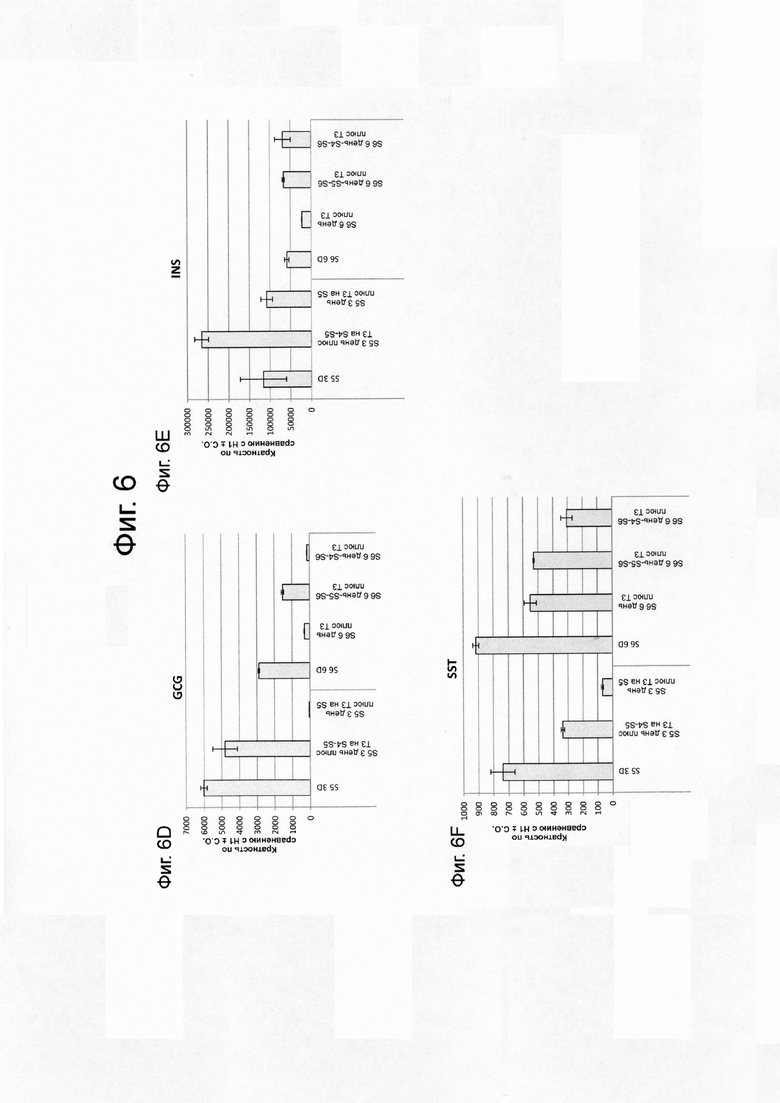
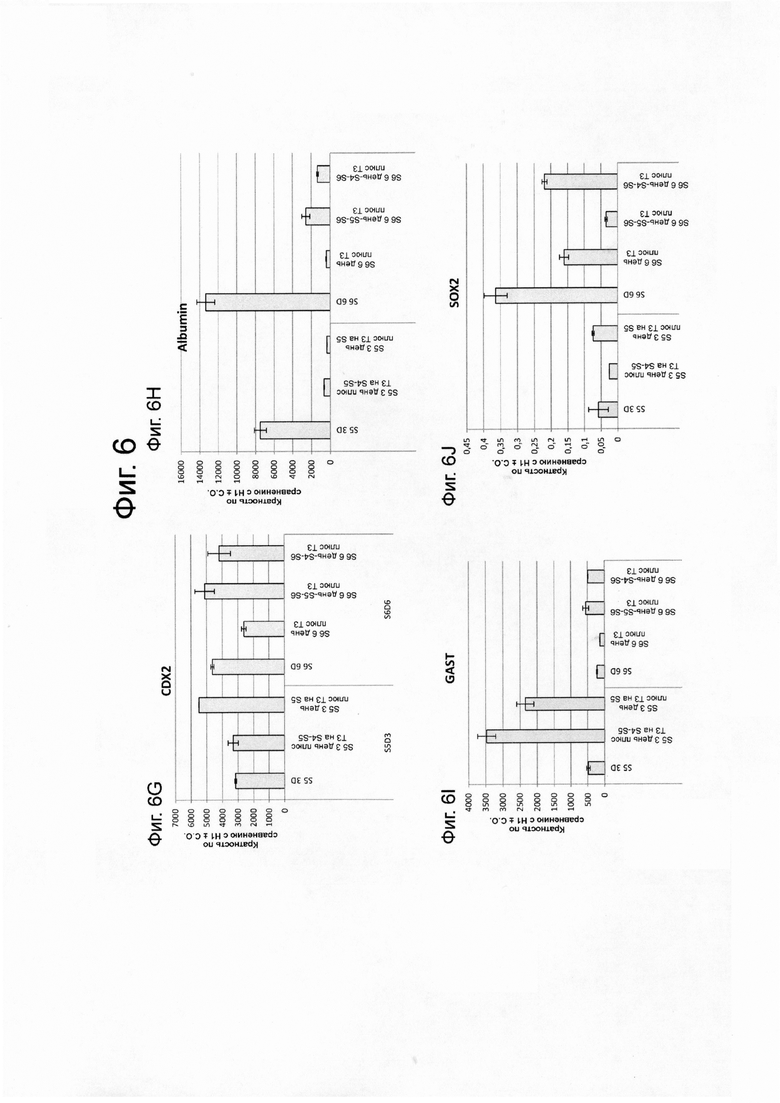
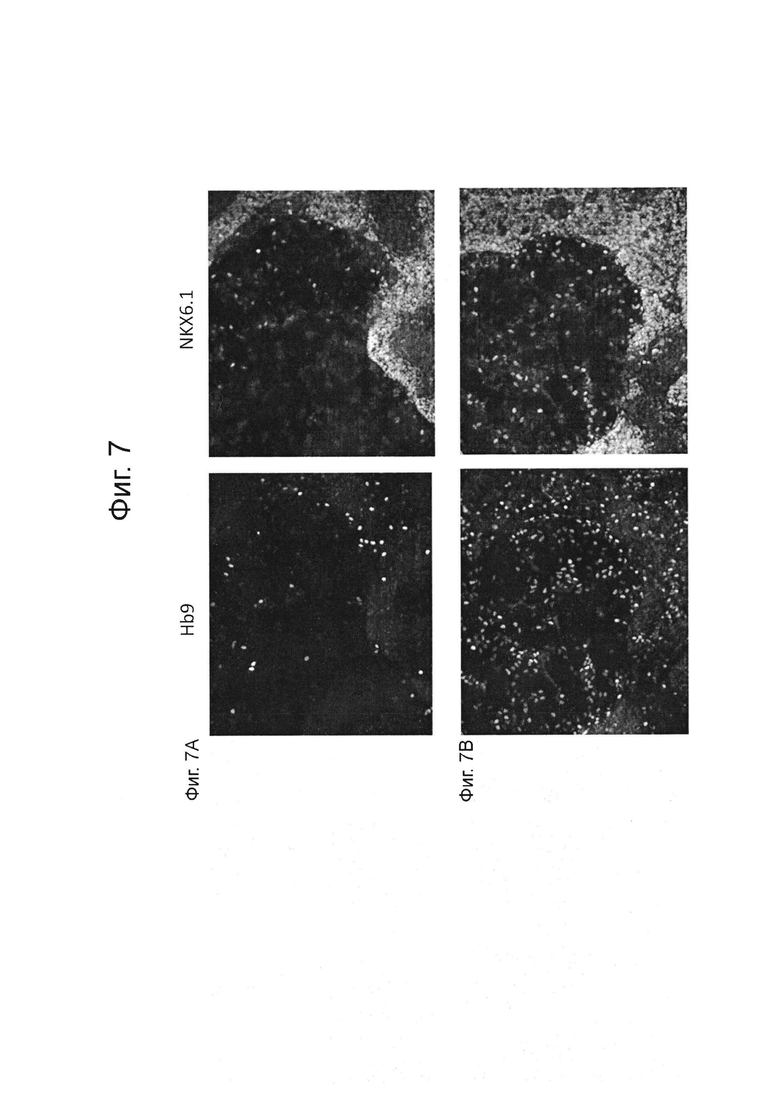

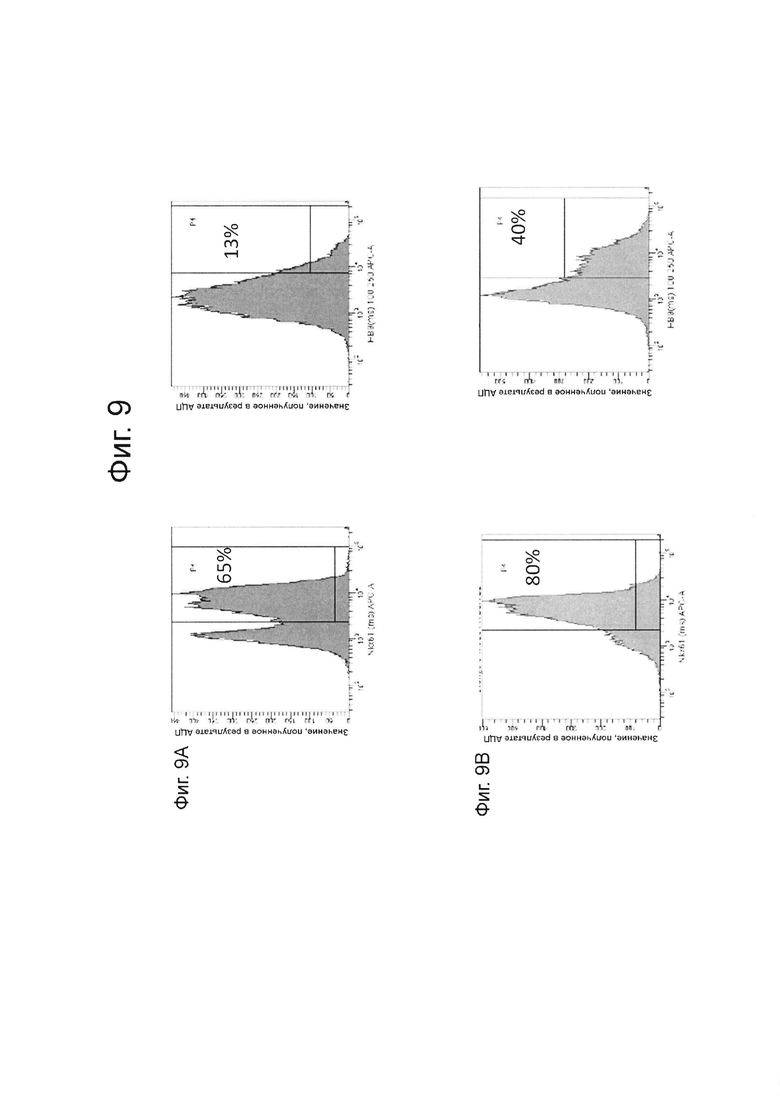
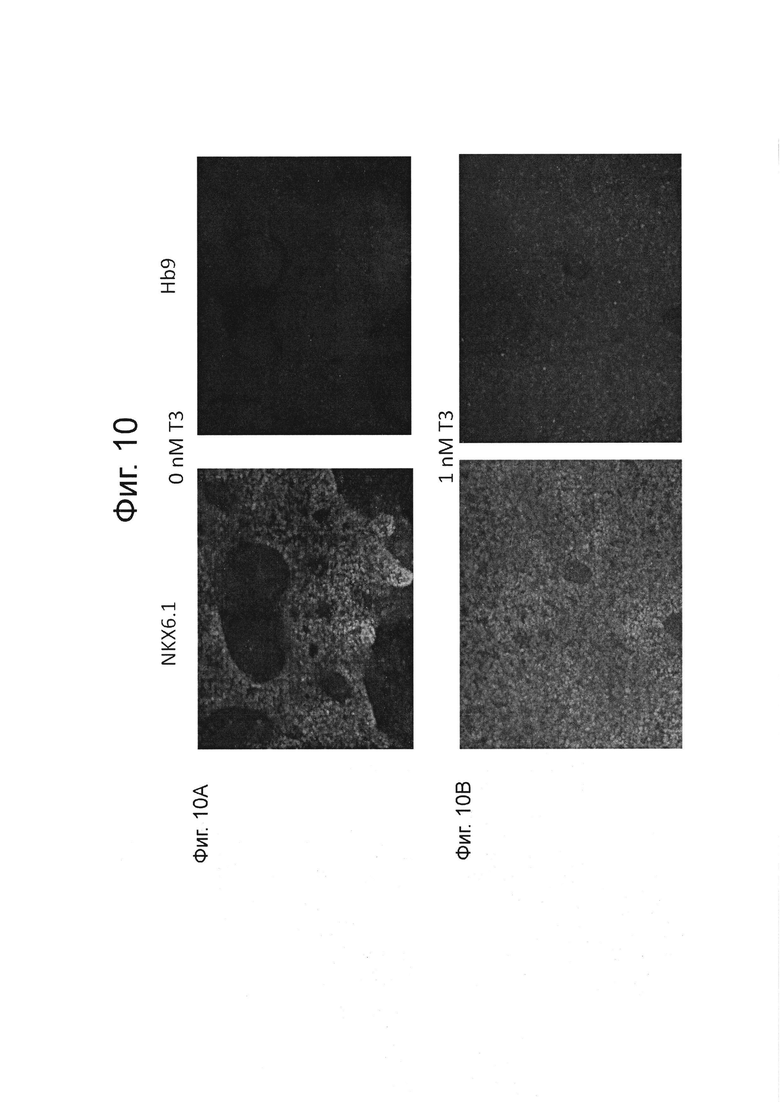


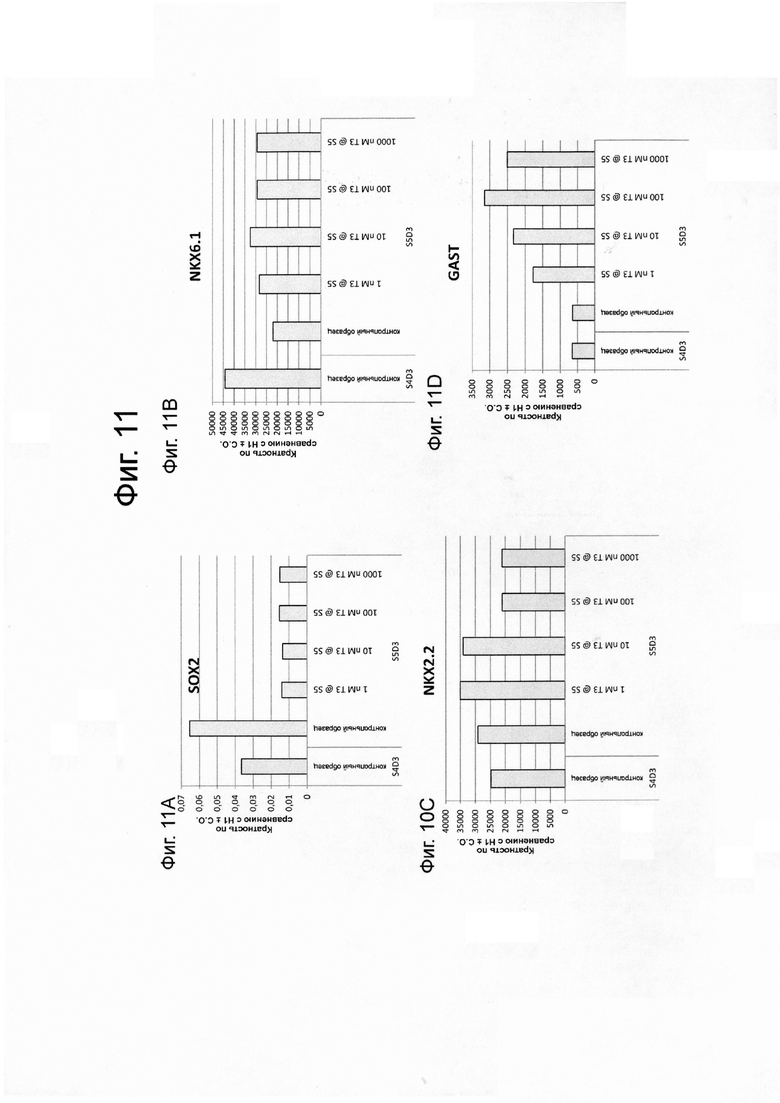


Изобретение относится к области клеточной биологии и биотехнологии, а именно к получению клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток и увеличению выхода β-клеток. Способ включает дифференцировку клеток, экспрессирующих маркеры, характерные для клеток энтодермы передней кишки, или маркеры, характерные для клеток-предшественников панкреатической передней кишки, или маркеры, характерные для панкреатических энтодермальных/эндокринных клеток-предшественников, полученных из неэмбриональных плюрипотентных клеток, линий эмбриональных стволовых клеток линий Н1, Н7, Н9, индуцибельных плюрипотентных клеток или перепрограммированных плюрипотентных клеток, в клетки, экспрессирующие маркеры, характерные для панкреатических эндокринных клеток, путем обработкой гормоном щитовидной железы и, необязательно, ингибитором ALK5. Изобретение позволяет повысить выход указанных клеток и улучшить дифференцировку. 4 н. и 58 з.п. ф-лы, 17 ил., 3 табл., 3 пр.
1. Способ получения панкреатических эндокринных клеток, включающий дифференцировку клеток энтодермы передней кишки в панкреатические эндокринные клетки путем обработки гормоном щитовидной железы, выбранным из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смеси перечисленного, и, необязательно, ингибитором ALK5, где клетки энтодермы передней кишки получены из плюрипотентных клеток неэмбрионального происхождения, линий эмбриональных стволовых клеток человека H1, линий эмбриональных стволовых клеток человека H7, линий эмбриональных стволовых клеток человека H9, индуцибельных плюрипотентных клеток или перепрограммированных плюрипотентных клеток.
2. Способ по п.1, в котором по меньшей мере десять процентов полученных клеток являются положительными для NKX6.1, PDX1 и НВ9.
3. Способ по п.1, в котором увеличивается экспрессия НВ9 в NKX6.1-позитивных клетках-предшественниках панкреатической энтодермы.
4. Способ по п.1, в котором снижается экспрессия NKX2.2.
5. Способ по п.1, в котором подавляется экспрессия SOX2 и альбумина.
6. Способ по п.1, в котором гормоном щитовидной железы является трииодотиронин.
7. Способ по п.1, отличающийся тем, что способ включает обработку гормоном щитовидной и ингибитором ALK5.
8. Способ по п.1, отличающийся тем, что способ включает обработку трииодотиронином и ингибитором ALK5.
9. Способ по п.8, в котором повышается экспрессия НВ9 по сравнению с клетками, не обработанными трииодотиронином и ингибитором ALK5.
10. Способ по п.1, включающий обработку SANT-1, ретиноевой кислотой, аскорбиновой кислотой, гормоном щитовидной железы, выбранным из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смеси перечисленного, и, необязательно, ингибитором ALK5.
11. Способ по п.1, отличающийся тем, что способ также включает формирование панкреатических эндокринных клеток путем обработки панкреатических энтодермальных/эндокринных клеток-предшественников, трииодотиронином.
12. Способ по п.11, включающий обработку трииодотиронином и ингибитором ALK5.
13. Способ по п.1, в котором панкреатические эндокринные клетки производят инсулин.
14. Способ по п. 1 или 7, в котором ингибитор ALK5 выбран из ингибитора ALK5 II, ALK5i, SD208, TGF-B ингибитоора SB431542, ITD-1, LY2109761, A83-01, LY2157299, TGF-β ингбитора рецептора V, TGF-β ингибитора рецептора I, TGF-β ингибитора рецептора IV, TGF-β ингибитора рецептора VII, TGF-β ингибитора рецептора VIII, TGF-β ингибитора рецептора II, TGF-β ингибитора рецептора VI, TGF-β ингибитора рецептора III.
15. Способ по п.14, в котором ингибитором ALK5 является ингибитор ALK5 II.
16. Способ по п. 1 или 7, в котором гормон щитовидной железы, выбран из трииодотиронина, тироксина, GC-1, 3,5-дииодотиропропионовой кислоты, КВ-141, MB07344, T0681 и GC-24.
17. Способ по п.1, в котором гормон щитовидной железы, выбранный из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смеси перечисленного, и, необязательно, ингибитор ALK5 находятся в среде, выбранной из группы, состоящей из: модифицированной среды Дюльбекко по методу Игла (DMEM); MCDB-131; и базальной среды Хэма F12/50% DMEM.
18. Способ получения панкреатических эндокринных клеток, включающий дифференцировку клеток-предшественников панкреатической передней кишки в панкреатические эндокринные клетки путем обработки гормоном щитовидной железы, выбранным из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смесью вышеперечисленного, и, необязательно, ингибитором ALK5, где клетки-предшественники панкреатической передней кишки получены из плюрипотентных клеток неэмбрионального происхождения, линий эмбриональных стволовых клеток человека H1, линий эмбриональных стволовых клеток человека H7, линий эмбриональных стволовых клеток человека H9, индуцибельных плюрипотентных клеток или перепрограммированных плюрипотентных клеток.
19. Способ по п.18, в котором по меньшей мере десять процентов полученных клеток являются положительными для NKX6.1, PDX1 и НВ9.
20. Способ по п.18, в котором увеличивается экспрессия НВ9 в NKX6.1-позитивных клетках-предшественниках панкреатической энтодермы.
21. Способ по п.18, в котором снижается экспрессия NKX2.2.
22. Способ по п.18, в котором подавляется экспрессия SOX2 и альбумина.
23. Способ по п.18, в котором гормоном щитовидной железы является трииодотиронин.
24. Способ по п.18, отличающийся тем, что способ включает обработку гормоном щитовидной железы и ингибитором ALK5.
25. Способ по п.24, включающий обработку трииодотиронином и ингибитором ALK5.
26. Способ по п.25, в котором повышается экспрессия НВ9, по сравнению с клетками, не обработанными трииодотиронином и ингибитором ALK5.
27. Способ по п.18, дополнительно включающий формирование панкреатических эндокринных клеток путем обработки трииодотиронином панкреатических энтодермальных/эндокринных клеток-предшественников.
28. Способ по п.27, включающий обработку трииодотиронином и ингибитором ALK5.
29. Способ по п.18, включающий обработку ингибитором рецептора ВМР, активатором РКС, гормоном щитовидной железы, выбранным из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смесью вышеперечисленного, и, необязательно, ингибитором ALK5.
30. Способ по п.29, в котором ингибитор рецептора ВМР выбран из LDN-193189, Noggin и Chordin, и активатор РКС выбран из TPB, PDBu, PMA и ILV.
31. Способ по п.18, в котором ингибитор ALK5 выбран из ингибитора ALK5 II, ALK5i, SD208, TGF-B ингибитоора SB431542, ITD-1, LY2109761, A83-01, LY2157299, TGF-β ингбитора рецептора V, TGF-β ингибитора рецептора I, TGF-β ингибитора рецептора IV, TGF-β ингибитора рецептора VII, TGF-β ингибитора рецептора VIII, TGF-β ингибитора рецептора II, TGF-β ингибитора рецептора VI, TGF-β ингибитора рецептора III.
32. Способ по п.31, в котором ингибитором ALK5 является ингибитор ALK5 II.
33. Способ по п.18, в котором гормон щитовидной железы, выбран из трииодотиронина, тироксина, GC-1, 3,5-дииодотиропропионовой кислоты, КВ-141, MB07344, T0681 и GC-24.
34. Способ по п.18, в котором панкреатические эндокринные клетки продуцируют инсулин.
35. Способ по п.18, в котором гормон щитовидной железы, выбранный из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смеси перечисленного, и, необязательно, ингибитор ALK5 находятся в среде, выбранной из группы, состоящей из: модифицированной среды Дюльбекко по методу Игла (DMEM); MCDB-131; и базальной среды Хэма F12/50% DMEM.
36. Способ получения панкреатических эндокринных клеток, включающий обработку клеток энтодермы передней кишки, клеток-предшественников панкреатической передней кишки или панкреатических энтодермальных/эндокринных клеток-предшественников, гормоном щитовидной железы, выбранным из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смесью вышеперечисленного, и, необязательно, ингибитором ALK5, где клетки энтодермы передней кишки, клетки-предшественники панкреатической передней кишки или панкреатические энтодермальные/эндокринные клетки-предшественники получены из плюрипотентных клеток неэмбрионального происхождения, линий эмбриональных стволовых клеток человека H1, линий эмбриональных стволовых клеток человека H7, линий эмбриональных стволовых клеток человека H9, индуцибельных плюрипотентных клеток или перепрограммированных плюрипотентных клеток.
37. Способ по п.36, в котором клетки обрабатывают на протяжении периода от примерно трех до примерно девяти дней.
38. Способ по п.36, отличающийся тем, что способ включает обработку гормоном щитовидной железы и ингибитором ALK5.
39. Способ по п.38, в котором клетки обрабатывают гормоном щитовидной железы, но не ингибитором ALK5, на протяжении периода от примерно двух до трех дней, после чего полученную популяцию клеток обрабатывают как гормоном щитовидной железы, так и ингибитором ALK5 на протяжении от примерно трех до шести дней.
40. Способ по п.39, в котором клетки дополнительно обрабатывают на протяжении периода примерно трех дней гормоном щитовидной железы, но не ингибитором ALK5.
41. Способ по п.39, в котором гормон щитовидной железы является трииодотиронином.
42. Способ по п.36, в котором панкреатические эндокринные клетки экспрессируют инсулин.
43. Способ по п.36, в котором ингибитор ALK5 выбран из ингибитора ALK5 II, ALK5i, SD208, TGF-B ингибитоора SB431542, ITD-1, LY2109761, A83-01, LY2157299, TGF-β ингбитора рецептора V, TGF-β ингибитора рецептора I, TGF-β ингибитора рецептора IV, TGF-β ингибитора рецептора VII, TGF-β ингибитора рецептора VIII, TGF-β ингибитора рецептора II, TGF-β ингибитора рецептора VI, TGF-β ингибитора рецептора III.
44. Способ по п.36, отличающийся тем, что способ включает культивирование клеток энтодермы передней кишки.
45. Способ по п.36, отличающийся тем, что способ включает культивирование клеток-предшественников панкреатической передней кишки.
46. Способ по п.36, отличающийся тем, что способ включает культивирование панкреатических энтодермальных/эндокринных клеток-предшественников.
47. Способ по п.36, отличающийся тем, что способ включает культивирование клеток энтодермы передней кишки для дифференциации клеток энтодермы передней кишки в клетки-предшественники панкреатической передней кишки.
48. Способ по п.47, отличающийся тем, что способ включает культивирование клеток-предшественников панкреатической передней кишки для дифференциации клеток-предшественников панкреатической передней кишки в панкреатические энтодермальные/эндокринные клетки-предшественники.
49. Способ по п.48, отличающийся тем, что способ включает культивирование панкреатических энтодермальных/эндокринных клеток-предшественников для дифференциации панкреатических энтодермальных/эндокринных клеток-предшественников в панкреатические эндокринные клетки.
50. Способ по п.36, в котором гормон щитовидной железы, выбранный из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смеси перечисленного, и, необязательно, ингибитор ALK5 находятся в среде, выбранной из группы, состоящей из: модифицированной среды Дюльбекко по методу Игла (DMEM); MCDB-131; и базальной среды Хэма F12/50% DMEM.
51. Способ увеличения выхода β-клеток, включающий обработку клеток гормоном щитовидной железы, выбранным из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смесью вышеперечисленного, и, необязательно, ингибитором ALK5, где клетки представляют собой клетки энтодермы передней кишки, клетки-предшественники панкреатической передней кишки или панкреатические энтодермальные/эндокринные клетки-предшественники, где клетки энтодермы передней кишки, клетки-предшественники панкреатической передней кишки или панкреатические энтодермальные/эндокринные клетки-предшественники получены из плюрипотентных клеток неэмбрионального происхождения, линий эмбриональных стволовых клеток человека H1, линий эмбриональных стволовых клеток человека H7, линий эмбриональных стволовых клеток человека H9, индуцибельных плюрипотентных клеток или перепрограммированных плюрипотентных клеток,.
52. Способ по п.51, в котором гормон щитовидной железы является трииодотиронином.
53. Способ по п.51, в котором клетки, экспрессирующие маркеры, характерные для β-клеток, экспрессируют инсулин.
54. Способ по п.52, отличающийся тем, что способ включает обработку клеток гормоном щитовидной и ингибитором ALK5.
55. Способ по п.51, в котором ингибитор ALK5 выбран из ингибитора ALK5 II, ALK5i, SD208, TGF-B ингибитоора SB431542, ITD-1, LY2109761, A83-01, LY2157299, TGF-β ингбитора рецептора V, TGF-β ингибитора рецептора I, TGF-β ингибитора рецептора IV, TGF-β ингибитора рецептора VII, TGF-β ингибитора рецептора VIII, TGF-β ингибитора рецептора II, TGF-β ингибитора рецептора VI, TGF-β ингибитора рецептора III.
56. Способ по п.51, отличающийся тем, что способ включает культивирование клеток энтодермы передней кишки.
57. Способ по п.51, отличающийся тем, что способ включает культивирование клеток-предшественников панкреатической передней кишки.
58. Способ по п.51, отличающийся тем, что способ включает культивирование панкреатических энтодермальных/эндокринных клеток-предшественников.
59. Способ по п.51, отличающийся тем, что способ включает культивирование клеток энтодермы передней кишки для дифференциации клеток энтодермы передней кишки в клетки-предшественники панкреатической передней кишки.
60. Способ по п.59, отличающийся тем, что способ включает культивирование клеток-предшественников панкреатической передней кишки для дифференциации клеток-предшественников панкреатической передней кишки в панкреатические энтодермальные/эндокринные клетки-предшественники.
61. Способ по п.60, отличающийся тем, что способ включает культивирование панкреатических энтодермальных/эндокринных клеток-предшественников для дифференциации панкреатических энтодермальных/эндокринных клеток-предшественников в панкреатические эндокринные клетки.
62. Способ по п.51, в котором гормон щитовидной железы, выбранный из трииодотиронина, тироксина, аналогов трииодотиронина, аналогов тироксина или смеси перечисленного, и, необязательно, ингибитор ALK5 находятся в среде, выбранной из группы, состоящей из: модифицированной среды Дюльбекко по методу Игла (DMEM); MCDB-131; и базальной среды Хэма F12/50% DMEM.
| US 20110281355 A1, 17.11.2011 | |||
| US 20090170198 A1, 02.07.2009 | |||
| US 20070254359 A1, 01.11.2007 | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| ГИЛБЕРТ С | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| с англ | |||
| - М.: Мир, 1993 | |||
| Приспособление для нагрузки тендеров дровами | 1920 |
|
SU228A1 |

