Область техники
Настоящее изобретение относится к области медицины и к области доставки генов для применений в вакцинации и генной терапии. Более конкретно настоящее изобретение относится к эффективному и сбалансированному двунаправленному промотору для экспрессии двух трансгенов с помощью рекомбинантных векторов, таких как плазмидные векторы, вирусные векторы и рекомбинантные вирусы.
Предпосылки изобретения
Рекомбинантные векторы широко используются при многих путях применения молекулярной биологии для экспрессии гетерологичных белков, в том числе, например, их применяют в генной терапии и вакцинации. Для таких путей применения генной терапии и вакцинации, векторы, в том числе вирусные векторы, применяют в качестве носителей для гена или генов, представляющих интерес для введения в клетки-хозяева. Например, вирусные векторы можно применять для экспрессии гена или его части, кодирующих необходимый антиген для вызова иммунного ответа.
Наиболее ранние вирусные векторы включали, как правило, только один трансген, и опубликовано много стратегий для векторов раннего поколения. Например, в опубликованных стратегиях сообщается о применении многих разных аденовирусных (rAd) векторов, и в них показано, что кассету для экспрессии трансгена можно поместить в разные участки rAd, например, в участок E1, участок E3 или между E4 и правым ITR. Для целей вакцинации, однако, часто требуется более одного антигена или такой же антиген из нескольких разных штаммов для достижения защиты и широкого охвата. Следовательно, в определенных случаях является необходимой экспрессия по меньшей мере двух антигенов из одного вектора. Были описаны разные подходы для кодирования двух антигенов в одном вирусном векторе.
В первом подходе с двумя антигенами с использованием rAd одну кассету для экспрессии антигена помещали в участок E1 и вторую кассету помещали в участок E3 (например (Vogels et al., 2007)). В отличном подходе с двумя антигенами с использованием rAd одну кассету для экспрессии антигена помещали в E1 и вторую кассету помещали между E4 и правым ITR (например (Holman et al., 2007; Pham et al., 2009; Schepp-Berglind et al., 2007)). Еще один подход с двумя антигенами с использованием rAd заключается в применении двух кассет для экспрессии антигена, помещенных в участок E1 в расположении голова-к-хвосту с применением двух разных промоторных последовательностей с целью предотвратить генетическую нестабильность посредством рекомбинации (например (Belousova et al., 2006; C. D. Harro et al., 2009)).
Еще один пример подхода с двумя антигенами заключается в применении внутреннего сайта связывания рибосомы (IRES) РНК-вирусов с положительной цепью, например полученного из вируса энцефаломиокардита (EMCV) для получения одного транскрипта, который транслируется в два белка (например, (Amendola, Venneri, Biffi, Vigna, & Naldini, 2005; Na & Fan, 2010)). Другие примеры включают применение сплайсингового механизма клетки-хозяина или применение "расщепленных" пептидов, полученных из РНК-вирусов с положительной цепью, таких как последовательность 2A эпизоотического стоматита или эквивалентов из других вирусов, для получения полипротеина, который расщеплен на два белка. В соответствии с опубликованными отчетами все из данных стратегий могут быть в равной степени применимы и успешны.
В качестве альтернативы применение двунаправленных промоторов представляет собой еще один подход для экспрессирования двух антигенов с помощью вирусных векторов. Например, разные двунаправленные промоторы были описаны для лентивирусных векторов (Heilbronn & Weger, 2010) и аденовирусных векторов (Na & Fan, 2010; Post & Van Meir, 2001; Robbins & Ghivizzani, 1998; Walther & Stein, 2000).
В целом, известны два разных типа двунаправленных промоторов для применения, встречающиеся в природе последовательности с двунаправленными свойствами и построенные синтетическим путем двунаправленные промоторы. Встречающиеся в природе последовательности с двунаправленными свойствами можно найти в составе геномов вирусов, растений или млекопитающих (Andrianaki, Siapati, Hirata, Russell, & Vassilopoulos, 2010; Barski, Siller-Lopez, Bohren, Gabbay, & Aguilar-Cordova, 2004). Например, сообщалось, что многие промоторы в геноме человека имеют некоторые двунаправленные свойства. Промоторы человека с двунаправленными свойствами характеризуются преобладанием сайтов GABP (Collins, Kobayashi, Nguyen, Trinklein, & Myers, 2007).
В отличие от встречающихся в природе последовательностей, синтетические двунаправленные промоторы могут быть построены для извлечения пользы из необходимых свойств разных однонаправленных промоторов. Например, Amendola et al. создали два разных синтетических двунаправленных промотора для применения в лентивирусных векторах посредством комбинирования минимального промотора, полученного из цитомегаловируса человека (minCMV), с промотором фосфоглицераткиназы (PGK) человека или промотором убиквитина С (UBI C) человека (Amendola et al., 2005). Для конструирования двунаправленных промоторов однонаправленным промоторам придавали конфигурацию в противоположной ориентации (голова к голове) с применением только одного энхансера. В соответствии с Amendola et al., когда сильный минимальный промотор комбинировали с полным промотором млекопитающих в данной конфигурации, в качестве результата получали координированную экспрессию с обоих сторон. Важные признаки для новосозданных поливалентных векторов включают, например, генетическую стабильность во время увеличения масштаба, продуктивность вектора в крупном масштабе, эффективную экспрессию обоих антигенов, сбалансированную экспрессию обоих антигенов и ограничения размеров антигенов, экспрессированных из встроенных кассет экспрессии.
В недавно описанной стратегии, при применении которой показаны особенно хорошие результаты по сравнению с ранее раскрытыми способами, применяли двунаправленный промотор цитомегаловируса мыши (mCMV) для экспрессии двух трансгенов (WO 2016/166088). В приведенном источнике первый трансген был функционально связан с двунаправленным промотором mCMV в одном направлении, и второй трансген был функционально связан с двунаправленным промотором mCMV в другом направлении. Было установлено, что rAd с двунаправленным промотором mCMV является генетически стабильным, обеспечивая генетическую стабильность, которая была сравнима с rAd только с одним трансгеном. Кроме того, основываясь на анализе ELISPOT и ELISA иммуногенности экспрессированных антигенов в отношении T-клеточного и B-клеточного ответов было установлено, что оба трансгена экспрессировались в достаточной мере, чтобы вызывать иммуногенные ответы на оба антигена. Двунаправленный промотор mCMV, таким образом, был описан как превосходящий несколько других ранее описанных стратегий. Однако, было установлено, что сбалансированность уровней экспрессии между двумя сторонами промотора mCMV может быть дополнительно улучшена. Уровень экспрессии антигена, расположенного на правой стороне (3'-конце) двунаправленного промотора mCMV был примерно в 10 раз выше по сравнению с антигеном, расположенным на левой стороне (5'-конце) промотора. Дисбаланс в экспрессии двух закодированных антигенов приводит к более сильному иммунному ответу, направленному против экспрессированного на высоком уровне антигена, по сравнению с антигеном, экспрессированным на более низком уровне. Такой тип дифференциальной экспрессии может быть полезным для определенных применений, но для других применений также является необходимым иметь стратегию, которая объединяет несколько преимуществ промотора mCMV с более сбалансированной экспрессией, т. е. двунаправленный промотор, который является как эффективным, так и более сбалансированным, чем двунаправленный промотор mCMV и другие двунаправленные промоторы, которые были описаны в литературе.
Таким образом, сохраняется потребность в идентификации двунаправленных промоторов, которые являются эффективными, относительно короткими, не имеют или имеют ограниченное количество длинных внутренних отрезков идентичных последовательностей, и имеют улучшенную сбалансированность в отношении экспрессии с обеих сторон по сравнению с двунаправленным промотором mCMV, и в получении рекомбинантных вирусов, которые являются генетически стабильными в отношении эффективной и сбалансированной экспрессии двух трансгенов.
Краткое описание изобретения
Настоящее изобретение предусматривает молекулы рекомбинантной нуклеиновой кислоты, содержащие двунаправленный промотор hCMV-rhCMV и векторы, в том числе, например, плазмидные векторы, вирусные векторы и вирусы, содержащие двунаправленный промотор hCMV-rhCMV. Рекомбинантные векторы по настоящему изобретению содержат два трансгена, где направление транскрипции (от 5' к 3') частей hCMV и rhCMV двунаправленного промотора hCMV-rhCMV расходится друг от друга (конфигурация голова к голове), при этом первый трансген функционально связан в одном направлении с левой стороной, при этом экспрессия регулируется частью hCMV двунаправленного промотора, и второй трансген функционально связан в противоположном направлении с правой стороной, при этом экспрессия регулируется частью rhCMV двунаправленного промотора. Энхансер hCMV помещают посередине между двумя разными промоторами в направлении части промотора hCMV. Поскольку энхансеры могут быть ориентационно-независимыми, энхансер обеспечивает координированную экспрессию обоих трансгенов, функционально связанных с частями hCMV и rhCMV двунаправленного промотора. См., например, на фиг. 1D показаны идентичность и ориентация для разных структурных блоков иллюстративного промотора hCMV-rhCMV. Предпочтительно промотор hCMV-rhCMV в соответствии с настоящим изобретением содержит нуклеотидную последовательность, которая на по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95% и до 100% идентична с SEQ ID NO: 4.
В определенных вариантах осуществления рекомбинантные вирусы и рекомбинантные вирусные векторы представляют собой рекомбинантные аденовирусы (rAd) и векторы на основе rAd. Полученный rAd с двунаправленным промотором hCMV-rhCMV по настоящему изобретению является генетически стабильным, без полос делеций, выявляемых посредством анализа ПЦР, до пассажа 13 (p13), таким образом обеспечивая генетическую стабильность, которая сравнима с вирусами только с одним трансгеном. Кроме того, двунаправленный промотор hCMV-rhCMV является относительно коротким двунаправленным промотором, состоящим всего лишь из 943 нуклеотидов, и он обеспечивает эффективную и очень сбалансированную экспрессию двух трансгенов. Таким образом, двунаправленный промотор hCMV-rhCMV по настоящему изобретению является подходящим для применения в генной терапии и путях применения в вакцинах с рекомбинантными (вирусными) векторами, и, в частности, где является важной сбалансированная и эффективная экспрессия и/или где является целесообразным небольшой размер двунаправленного промотора hCMV-rhCMV.
Общие и предпочтительные варианты осуществления определены, соответственно, с помощью независимых и зависимых пунктов формулы изобретения, прилагаемой к данному документу, которые для краткости включены в данное описание с помощью ссылки. Другие предпочтительные варианты осуществления, признаки и преимущества различных аспектов настоящего изобретения станут очевидными из приведенного ниже подробного описания, рассматриваемого вместе с прилагаемыми графическими материалами.
В одном варианте осуществления настоящее изобретение предусматривает двунаправленный промотор hCMV-rhCMV, содержащий промотор hCMV на левой стороне и промотор rhCMV на правой стороне, где двунаправленный промотор hCMV-rhCMV функционально связан с первым трансгеном в одном направлении на левой стороне и двунаправленный промотор hCMV-rhCMV функционально связан со вторым трансгеном на правой стороне в другом направлении.
В другом варианте осуществления настоящее изобретение также предусматривает способ получения рекомбинантного вируса, содержащего первый и второй трансгены, при этом способ включает: получение конструкции, содержащей двунаправленный промотор hCMV-rhCMV, функционально связанный с первым трансгеном в одном направлении и вторым трансгеном в противоположном направлении, и встраивание указанной конструкции в геном рекомбинантного вируса.
В определенных вариантах осуществления рекомбинантный вирус представляет собой рекомбинантный аденовирус.
В определенных вариантах осуществления рекомбинантный аденовирус предусматривает делецию в участке E1 и в определенных вариантах осуществления содержит двунаправленный промотор hCMV-rhCMV и первый, и второй трансген в данном участке E1. В качестве альтернативы также можно применять другие участки рекомбинантных аденовирусов. Например, кассету экспрессии, содержащую двунаправленный промотор, также можно поместить на правом конце генома, между участком E4 и правым ITR рекомбинантного аденовируса.
В определенных вариантах осуществления первый и второй трансгены являются разными, и по меньшей мере один из них кодирует антиген. В определенных вариантах осуществления оба кодируют разный антиген.
В определенных вариантах осуществления аденовирус представляет собой серотип 26 аденовируса человека или серотип 35 аденовируса человека.
В другом варианте осуществления настоящее изобретение также предусматривает способ экспрессирования по меньшей мере двух трансгенов в клетке, при этом способ предусматривает доставку в клетку рекомбинантного вектора в соответствии с настоящим изобретением.
В другом варианте осуществления настоящее изобретение также предусматривает способ индуцирования иммунного ответа против по меньшей мере двух антигенов, при этом способ предусматривает введение субъекту рекомбинантного вектора в соответствии с настоящим изобретением.
В другом варианте осуществления настоящее изобретение также предусматривает молекулу рекомбинантной ДНК, содержащую геном рекомбинантного аденовируса в соответствии с настоящим изобретением.
В другом варианте осуществления настоящее изобретение также предусматривает фармацевтическую композицию, содержащую рекомбинантный вектор, такой как рекомбинантный аденовирус в соответствии с настоящим изобретением, и фармацевтически приемлемые носитель или вспомогательное вещество. В определенных вариантах осуществления фармацевтическая композиция представляет собой вакцину.
Краткое описание графических материалов
Фиг. 1. Схематические представления исследуемых конструкций двунаправленного промотора, включая аннотации идентичности и ориентации структурных блоков для разных последовательностей двунаправленного промотора. P: промотор, Enh: энхансер, I: интрон.
Фиг. 2. Экспрессию люциферазы и eGFP с разными конструкциями двунаправленного промотора оценивали с помощью временных трансфекций в клетках HEK293. Экспрессию люциферазы измеряют в виде относительных световых единиц (RLU) и экспрессию eGFP измеряют в виде среднего значения интенсивности флуоресценции (MFI) с помощью FACS. Показаны результаты трех разных экспериментов с отбором разных конструкций промотора. Показаны столбчатые диаграммы результатов экспрессии люциферазы из левой стороны и экспрессии eGFP из правой стороны разных двунаправленных промоторов. Положительный контроль: Люцифераза или eGFP под контролем однонаправленного промотора hCMV; в качестве отрицательного контроля применяли нетрансфицированные клетки.
Фиг. 3. (A) Организация кассеты двунаправленной экспрессии для двунаправленного промотора hCMV-rhCMV в pshuttle26, включая идентичность и местоположение сайтов рестрикции, применяемых для вставки трансгенов на обе стороны конструкции двунаправленного промотора. P: промотор, Enh: энхансер, TG: трансген, pA: сигнал полиаденилирования, полученный из SV40 (правая сторона) или бычьего гормона роста (BGH) (левая сторона). Представление в плазмидном векторе pshuttle26. Такую же организацию кассеты двунаправленной экспрессии применяли в pAdapt35. (B) Схематическое представление двунаправленного промотора hCMV-rhCMV, включая положения нуклеотидов структурных блоков. Стрелки представляют направление транскрипции.
Фиг. 4. Экспрессия трансгенов люциферазы и eGFP либо на левой, либо на правой стороне конструкций двунаправленного промотора в векторах на основе rAd Ad26 (A) и векторах на основе rAd Ad35 (B) с инфекциями в клетках A549 при 1000 VP/клетка. Экспрессию люциферазы измеряют в виде относительных световых единиц (RLU) и экспрессию eGFP измеряют в виде среднего значения интенсивности флуоресценции (MFI) с помощью FACS. Результаты для разных конструкций двунаправленного промотора hCMV-rhCMV по сравнению с положительными контролями, составляющие 100 VP/клетка и 1000 VP/клетка, для люциферазы или eGFP под контролем однонаправленного промотора hCMV и клеток, инфицированных с помощью пустого вектора. В случае векторов на основе rAd Ad26, hCMV-rhCMV дополнительно сравнивают с двунаправленным промотором mCMV.
Фиг. 5. Тестирование генетической стабильности посредством серийного размножения с последующей ПЦР на геноме вектора Ad26, содержащем двунаправленный промотор hCMV-rhCMV в участке E1 и кодирующего eGFP и люциферазу либо на правой, либо на левой стороне двунаправленного промотора hCMV-rhCMV. На секциях сверху вниз показаны продукты ПЦР для 5 бляшек на вектор после серийного размножения в клетках PER.C6 на P5, P10 и P13. На линиях 1-5 на каждой секции виден двунаправленный промотор hCMV-rhCMV с люциферазой слева и eGFP справа. На линиях 6-10 на каждой секции виден двунаправленный промотор hCMV-rhCMV с eGFP слева и люциферазой справа. На линии 11 виден маркер размера в тысячах нуклеотидов. На линии 12 виден плазмидный положительный контроль для Ad26.Luc.hCMV-rhCMV.eGFP. На линии 13 виден плазмидный положительный контроль для rAd26.eGFP.hCMV-rhCMV.Luc. На линии 14 виден плазмидный контроль размером с продукт ПЦР кассеты экспрессии без трансгена. На линии 15 виден отрицательный водный контроль ПЦР. Введение меток: P5, P10, P13: номер вирусного пассажа. Дополнительные полосы, присутствующие помимо ожидаемых продуктов ПЦР, представляют собой неспецифические продукты ПЦР. Примечание: отсутствие полос делеции подтверждали на передержанных снимках.
Подробное описание изобретения
Описанное в данном документе представляет собой экспериментальные результаты сравнения новых конструкций двунаправленного промотора в отношении эффективности и сбалансированности. Из результатов видно, что двунаправленный промотор hCMV-rhCMV обеспечивает эффективную и очень сбалансированную экспрессию двух трансгенов, исходя из временной трансфекции клеток HEK293 плазмдными векторами pAdApt и вирусными инфекциями, вызванными rAd26 и rAd35, содержащими двунаправленный промотор hCMV-rhCMV, при этом первый трансген функционально связан с двунаправленным промотором hCMV-rhCMV в одном направлении, и второй трансген функционально связан с двунаправленным промотором hCMV-rhCMV в другом направлении. Двунаправленный промотор hCMV-rhCMV также представляет собой относительно короткий двунаправленный промотор, состоящий всего лишь из 943 нуклеотидов. Кроме того, rAd с двунаправленным промотором hCMV-rhCMV являются генетически стабильными, без полос делеции, определяемых посредством анализа ПЦР, до пассажа 13 (p13), таким образом обеспечивая генетическую стабильность, которая сравнима с вирусами только с одним трансгеном. Таким образом, rAd по настоящему изобретению с двунаправленным промотором hCMV-rhCMV являются подходящими для применения в генной терапии и путях применения в вакцинах, где первостепенное значение имеет очень сбалансированная и эффективная экспрессия и/или где целесообразным является небольшой размер двунаправленного промотора hCMV-rhCMV, например, чтобы оставить больше места для трансгенов в ограниченном размере вектора или вирусного генома, по сравнению с другими, более длинными двунаправленными промоторами.
Настоящее изобретение, следовательно, относится к молекулам рекомбинантной нуклеиновой кислоты, содержащим двунаправленный промотор hCMV-rhCMV, функционально связанный с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении, где транскрипционное направление (от 5' к 3') частей hCMV и rhCMV двунаправленного промотора hCMV-rhCMV расходится друг от друга, и где экспрессия из левой стороны регулируется с помощью части hCMV двунаправленного промотора, и экспрессия из правой стороны регулируется с помощью части rhCMV двунаправленного промотора. В определенных вариантах осуществления настоящее изобретение относится к применению векторов, вирусных векторов и вирусов, содержащих двунаправленный промотор hCMV-rhCMV для экспрессирования двух трансгенов в клетке.
В определенных вариантах осуществления настоящее изобретение относится к применению плазмидных векторов, позволяющих клеткам-хозяевам продуцировать гетерологичные белки. Например, плазмидные векторы, содержащие двунаправленный промотор hCMV-rhCMV, можно применять для экспрессирования двух разных компонентов гетеромерного белкового комплекса из нескольких субъединиц. Такие плазмидные векторы могут представлять собой последовательности ДНК, содержащие, например, (1) двунаправленный промотор hCMV-rhCMV; (2) последовательности, обеспечивающие mRNA сайтами связывания рибосом для каждого трансгена; (3) кодирующий участок для каждого трансгена, т. е. последовательность нуклеотидов, которая кодирует необходимый полипептид; (4) консенсусную последовательность Kozak для каждого трансгена для инициации трансляции; (5) последовательность терминации для каждого трансгена, которая позволяет останавливать трансляцию, когда весь код для каждого трансгена был прочитан; и (6) если вектор не является непосредственно встроенным в геном, точку начала репликации, которая позволяет воспроизвести весь вектор после его попадания в клетку. Затем остается индуцировать включение вектора клеткой-хозяином, например, посредством трансфекции или электропорации и вырастить клетки-хозяева таким образом, чтобы экспрессировать два трансгена в рамках функции клетки-хозяина.
В определенных вариантах осуществления настоящее изобретение относится к rAd и векторам на основе rAd, содержащим двунаправленный промотор hCMV-rhCMV, и к способам получения и применения rAd и векторов на основе rAd, при этом rAd и векторы на основе rAd содержат двунаправленный промотор hCMV-rhCMV и два трансгена, где первый трансген функционально связан с двунаправленным промотором hCMV-rhCMV в одном направлении, и второй трансген функционально связан с двунаправленным промотором hCMV-rhCMV в другом направлении.
rAd по настоящему изобретению может быть получен в больших количествах или партиях. 'Партия' rAd представляет собой композицию, которая была получена за один производственный цикл в одной промышленной емкости, или, в качестве альтернативы, она может относиться ко множеству частиц rAd в композиции, которая присутствует в одном контейнере (например, биореакторе, резервуаре, колбе, флаконе, многодозовом флаконе, однодозовом флаконе, шприце и т. д.). Партия rAd в соответствии с настоящим изобретением или композиция, содержащая rAd в соответствии с настоящим изобретением, предпочтительно содержит по меньшей мере 107 частиц rAd и в определенных вариантах осуществления содержит по меньшей мере 108, 109, 1010, 1011, 1012, 1013, 1014, 1015, 1016, 1017, 1018 или большее количество частиц rAd, до 1020 частиц rAd (например, при получении в биореакторе промышленного масштаба за один производственный цикл). Партия или композиция может содержать или не содержать дополнительные соответствующие компоненты кроме rAd.
Используемый в данном документе термин 'рекомбинантный' в отношении рекомбинантного аденовируса подразумевает, что он, в отличие от аденовирусов дикого типа, был модифицирован человеком, например, он содержит гетерологичные ген, гены или их части и двунаправленный промотор hCMV-rhCMV.
Последовательности в данном документе представлены в направлении от 5' к 3', как принято в данной области.
"Капсидный белок аденовируса" относится к белку на капсиде аденовируса, который вовлечен в процесс установления серотипа и/или тропизма конкретного аденовируса. Капсидные белки аденовирусов, как правило, включают в себя белки файбер, пентон и/или гексон. rAd определенного серотипа (или 'полученный из определенного серотипа') в соответствии с настоящим изобретением, как правило, содержит белки файбер, пентон и/или гексон этого определенного серотипа, и предпочтительно содержит белок файбер, пентон и гексон этого определенного серотипа. Данные белки, как правило, кодируются с помощью генома rAd. rAd определенного серотипа необязательно может содержать и/или кодировать другие белки других серотипов аденовируса.
Используемый в данном документе rAd, 'полученный из' аденовируса, получен путем образования из дикого типа, по меньшей мере по последовательности. Его можно получать молекулярным клонированием с использованием генома дикого типа или его частей в качестве исходного материала. Также возможно применение известной последовательности генома аденовируса дикого типа для получения (частей) генома de novo с помощью синтеза ДНК, который может быть выполнен с использованием стандартных процедур компаниями, предоставляющими услуги в области синтеза ДНК и/или молекулярного клонирования (например, GeneArt, GenScripts, Invitrogen, Eurofins). Таким образом, в качестве неограничивающего примера rAd, который содержит гексон, пентон и файбер из Ad35, считается rAd, полученным из Ad35, и т. д.
Аденовирусные векторы по настоящему изобретению называют векторами на основе rAd. Получение векторов на основе rAd хорошо известно из уровня техники.
В определенных вариантах осуществления вектор на основе rAd в соответствии с настоящим изобретением является дефектным по по меньшей мере одному существенно важному функциональному гену участка E1, например участка E1a и/или участка E1b, аденовирусного генома, который требуется для вирусной репликации. В определенных вариантах осуществления аденовирусный вектор в соответствии с настоящим изобретением является дефектным по меньшей мере в части участка E3, не являющегося существенно важным. В определенных вариантах осуществления вектор является дефектным по по меньшей мере одному существенно важному функциональному гену участка E1 и по меньшей мере в части участка E3, не являющегося существенно важным. Аденовирусный вектор может быть "множественно дефектным", что означает, что данный аденовирусный вектор является дефектным по одному или нескольким существенно важным функциональным генам в каждом из двух или более участков аденовирусного генома. Например, вышеупомянутые аденовирусные векторы с дефектом по E1 или с дефектом по E1, E3 могут дополнительно являться дефектными по по меньшей мере одному существенно важному гену участка E4 и/или по меньшей мере одному существенно важному гену участка E2 (например, участка E2A и/или участка E2B).
Аденовирусные векторы, способы их конструирования и способы их размножения хорошо известны из уровня техники и описаны, например, в патентах США №№ 5559099, 5837511, 5846782, 5851806, 5994106, 5994128, 5965541, 5981225, 6040174, 6020191, 6113913 и 8932607, и Thomas Shenk, "Adenoviridae and their Replication" M. S. Horowitz, "Adenoviruses", главы 67 и 68, соответственно, в Virology, B. N. Fields et al., eds., 3d ed., Raven Press, Ltd., New York (1996), и других источниках, упомянутых в данном документе. Как правило, конструирование аденовирусных векторов включает применение стандартных молекулярно-биологических методик, которые хорошо известны из уровня техники, таких как описанные, например, в Sambrook et al., Molecular Cloning, a Laboratory Manual, 2d ed., Cold Spring Harbor Press, Cold Spring Harbor, N.Y. (1989), Watson et al., Recombinant DNA, 2d ed., Scientific American Books (1992), и Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, NY (1995), и других источниках, упомянутых в данном документе.
Аденовирус в соответствии с настоящим изобретением принадлежит к семейству Adenoviridae и предпочтительно представляет собой аденовирус, принадлежащий к роду Mastadenovirus. Он может представлять собой аденовирус человека, а также аденовирус, который инфицирует другие виды, в том числе без ограничения аденовирус крупного рогатого скота (например, аденовирус 3 крупного рогатого скота, BAdV3), аденовирус собак (например, CAdV2), аденовирус свиней (например, PAdV3 или 5) или аденовирус обезьян (который включает аденовирус нечеловекообразных обезьян и аденовирус человекообразных обезьян, как, например, аденовирус шимпанзе или аденовирус горилл). Предпочтительно аденовирус представляет собой аденовирус человека (HAdV или AdHu; в настоящем изобретении при обозначении Ad без указания видов подразумевают аденовирус человека, например краткое обозначение "Ad5" означает то же самое что и HAdV5, который представляет собой аденовирус человека серотипа 5) или аденовирус обезьян, такой как аденовирус шимпанзе или гориллы (ChAd, AdCh или SAdV).
Наиболее углубленные исследования были проведены с применением аденовирусов человека, при этом аденовирусы человека являются предпочтительными в соответствии с определенными аспектами настоящего изобретения. В определенных предпочтительных вариантах осуществления рекомбинантный аденовирус в соответствии с настоящим изобретением получен из аденовируса человека. В предпочтительных вариантах осуществления рекомбинантный аденовирус получен из аденовируса человека серотипа 5, 11, 26, 34, 35, 48, 49 или 50. В соответствии с конкретным предпочтительным вариантом осуществления настоящего изобретения, аденовирус представляет собой аденовирус человека одного из серотипов 26 и 35. Преимуществом данных серотипов является низкое доминирование серотипа и/или низкие титры ранее существующих нейтрализующих антител в популяции человека. Получение векторов на основе rAd26 описано, например, в WO 2007/104792 и в (Abbink et al., 2007). Иллюстративные последовательности генома Ad26 находятся в GenBank под номером доступа EF 153474 и под SEQ ID NO: 1 в WO 2007/104792. Получение векторов на основе rAd35 описано, например, в U.S. 7270811, в WO 00/70071 и в (Vogels et al., 2003). Иллюстративные последовательности генома Ad35 находятся в GenBank под номером доступа AC_000019 и показаны на фиг. 6 в WO 00/70071.
Аденовирусы обезьян обычно также характеризуются низким доминированием и/или низкими титрами ранее существующих нейтрализующих антител в популяции человека, и было опубликовано значительное количество работ с применением векторов на основе аденовируса шимпанзе (например, U.S. 6083716 и WO 2005/071093; WO 2010/086189 и WO 2010085984; (Bangari & Mittal, 2006; Cohen et al., 2002; Farina et al., 2001; Kobinger et al., 2006; Lasaro & Ertl, 2009; Tatsis et al., 2007). Следовательно, в других предпочтительных вариантах осуществления рекомбинантный аденовирус в соответствии с настоящим изобретением получен из аденовируса обезьян, например аденовируса шимпанзе. В определенных вариантах осуществления данный рекомбинантный аденовирус получен из аденовируса обезьян типа 1, 7, 8, 21, 22, 23, 24, 25, 26, 27.1, 28.1, 29, 30, 31.1, 32, 33, 34, 35.1, 36, 37.2, 39, 40.1, 41.1, 42.1, 43, 44, 45, 46, 48, 49, 50 или SA7P. Также векторы на основе аденовируса макака-резуса были описаны как пригодные кандидатные векторы (например, (Abbink et al., 2015); WO 2014/078688). Таким образом, в других предпочтительных вариантах осуществления рекомбинантный аденовирус по настоящему изобретению получен из аденовируса макака-резуса, к примеру, из одного из неограничивающих примеров RhAd51, RhAd52 или RhAd53 (или sAd4287, sAd4310A или sAd4312; см., например, (Abbink et al., 2015) и WO 2014/078688).
В дополнение к аденовирусам, специалистам в данной области будет понятно, что другие вирусы также являются подходящими для применения в качестве вирусных векторов с применением двунаправленных промоторов по настоящему изобретению. Например, аденоассоциированные вирусы (AAV), вирус простого герпеса (HSV), поксвирус и лентивирус также могут быть сконструированы для включения двунаправленных промоторов по настоящему изобретению. См., например, обзоры разных векторов, которые рассматриваются в (Heilbronn & Weger, 2010; Robbins & Ghivizzani, 1998; Walther & Stein, 2000).
Известны последовательности большинства выше упомянутых аденовирусов человека и аденовирусов, не являющихся аденовирусами человека, а для других они могут быть получены с использованием стандартных процедур.
Рекомбинантные аденовирусы в соответствии с настоящим изобретением могут быть репликативно-компетентными или дефектными по репликации.
В определенных вариантах осуществления аденовирус является дефектным по репликации, например, потому, что он предусматривает делецию в участке E1 генома. "Делеция в участке E1" означает делецию в этом участке по сравнению с аденовирусом дикого типа и означает делецию в по меньшей мере одном из кодирующих участков E1A, E1B 55K или E1B 21K, предпочтительно делецию кодирующих участков E1A, E1B 55K и E1B21K. Как известно специалисту, в случае делеций существенно важных участков из генома аденовируса функциональные элементы, кодируемые этими участками, должны быть обеспечены в транс-положении, предпочтительно клеткой-продуцентом, т. е., если части или целые участки E1, E2 и/или E4 удалены из аденовируса, то они должны присутствовать в клетке-продуценте, к примеру, быть встроенными в ее геном или находиться в форме так называемого вспомогательного аденовируса или вспомогательных плазмид. Аденовирус также может предусматривать делецию в участке E3, который не является существенным для репликации, и, следовательно, такую делецию не следует комплементировать.
Клетка-продуцент (также иногда называемая в данной области техники и в данном документе как "пакующая клетка", или "комплементирующая клетка", или "клетка-хозяин"), которую можно использовать, может представлять собой любую клетку-продуцента, в которой требуемый аденовирус может быть размножен. Например, размножение векторов но основе рекомбинантного аденовируса осуществляют в клетках-продуцентах, которые комплементируют дефекты в аденовирусе. Предпочтительно такие клетки-продуценты содержат в своем геноме по меньшей мере последовательность E1 аденовируса, и таким образом они способны к комплементированию рекомбинантных аденовирусов с делецией в участке E1. Можно применять любую Е1-комплементирующую клетку-продуцента, например клетки сетчатки глаза человека, иммортализированные с помощью E1, например клетки 911 или PER.C6 (см., например, U.S. 5994128), E1-трансформированные амниоциты (см., например, EP 1230354), E1-трансформированные клетки A549 (см., например, WO 98/39411, U.S. 5891690), клетки GH329:HeLa (Gao, Engdahl, & Wilson, 2000), клетки 293 и т. п. В определенных вариантах осуществления клетками-продуцентами являются, например, клетки HEK293, или клетки PER.C6, или клетки 911, или клетки IT293SF и т. п.
Для E1-дефектных аденовирусов, которые не происходят из подгруппы аденовирусов C или E, предпочтительной является замена кодирующей последовательности E4-orf6 аденовируса, не относящегося к подгруппе C или E, на E4-orf6 аденовируса из подгруппы C, например Ad5. Это позволяет осуществлять размножение таких аденовирусов в хорошо известных комплементирующих клеточных линиях, которые экспрессируют гены E1 из Ad5, таких как, например, клетки 293 или клетки PER.C6 (см., например (Havenga et al., 2006); WO 03/104467, включенный в данный документ посредством ссылки в полном объеме).
В альтернативных вариантах осуществления нет необходимости помещать гетерологичный участок E4orf6 (например, из Ad5) в аденовирусный вектор, но вместо этого E1-дефектный вектор на основе аденовируса, не относящегося к подгруппе C или E, размножают в клеточной линии, которая экспрессирует как E1, так и совместимый E4orf6, например, в клеточной линии 293-ORF6, которая экспрессирует как E1, так и E4orf6 из Ad5 (см., например, (Brough, Lizonova, Hsu, Kulesa, & Kovesdi, 1996), описывающие получение клеток 293-ORF6; (Abrahamsen et al., 1997; Nan et al., 2003), в каждой из которых описано получение векторов на основе аденовируса, не относящегося к подгруппе С, с удаленным Е1 с применением такой клеточной линии).
В качестве альтернативы можно применить комплементирующую клетку, которая экспрессирует E1 из серотипа, подлежащего размножению (см., например, WO 00/70071, WO 02/40665).
Для аденовирусов подгруппы B, таких как Ad35, с делецией в участке E1 предпочтительным является сохранение 3'-конца открытой рамки считывания E1B 55K в аденовирусе, к примеру, 166 п. о., расположенных непосредственно выше открытой рамки считывания pIX, или фрагмента, содержащего их, как, например, фрагмента из 243 п. о., расположенного непосредственно выше старт-кодона pIX (ограниченных на 5'-конце сайтом рестрикции Bsu36I в геноме Ad35), поскольку это повышает стабильность аденовируса, потому что промотор гена pIX частично расположен в данной области (см., например, (Havenga et al., 2006); WO 2004/001032, включенные в данный документ посредством ссылки).
"Гетерологичная нуклеиновая кислота" (также называемая в данном документе "трансген") в векторах или (адено)вирусах по настоящему изобретению представляет собой нуклеиновую кислоту, которая в естественных условиях не присутствует в векторе или (адено)вирусе. Ее вводят в вектор или (адено)вирус, к примеру, посредством стандартных методик молекулярной биологии. В определенных вариантах осуществления она может кодировать белок, представляющий интерес, или его часть. Например, ее можно клонировать в удаленный участок E1 или E3 аденовирусного вектора. В предпочтительных вариантах осуществления настоящего изобретения кассету экспрессии с двумя трансгенами под контролем двунаправленного промотора hCMV-rhCMV помещают в участок E1 аденовирусного генома. Трансген обычно функционально связан с последовательностями, контролирующими экспрессию. Это можно осуществить, к примеру, путем помещения нуклеиновой кислоты, кодирующей трансген(-ы), под контроль промотора. Для экспрессии трансгена(-ов) можно использовать множество промоторов, и они известны специалистам в данной области техники.
Известно, что гомологичные отрезки нуклеиновой кислоты могут приводить к нестабильности. Например, в результате более обширных испытаний оказалось, что применение двух идентичных промоторов (hCMV) в одном аденовирусном векторе, о возможности чего сообщалось ранее, приводит к генетической нестабильности аденовируса (WO 2016/166088). Исходя из этого, в настоящем изобретении сделана попытка свести к минимуму применение структурных блоков промотора с обширными отрезками идентичности последовательностей при конструировании двунаправленных промоторов для предотвращения возникновения делеций, происходящих по механизму гомологичной рекомбинации, в аденовирусном векторе. Важно отметить, что в настоящем документе показана генетическая стабильность аденовирусных векторов с трансгенами, регулируемыми с помощью двунаправленного промотора hCMV-rhCMV по настоящему изобретению.
Используемые в данном документе термины "промотор", или "промоторный участок", или "промоторный элемент" применяются взаимозаменяемо и относятся к сегменту последовательности нуклеиновой кислоты, как правило без ограничения к ДНК, которая контролирует транскрипцию последовательности нуклеиновой кислоты, с которой она связана по функции. Промоторный участок включает специфические последовательности, которые являются достаточными для распознавания РНК-полимеразой, связывания и инициации транскрипции. Кроме того, промоторный участок может необязательно включать последовательности, которые модулируют активность такого распознавания, связывания и инициации транскрипции РНК-полимеразой. Данные последовательности могут быть действующими в цис-положении или могут быть чувствительны к факторам, действующим в транс-положении. Кроме того, промоторы могут быть конститутивными или регулируемыми, в зависимости от природы регуляции.
Специалисту в данной области будет известно, что промоторы образованы из отрезков последовательностей нуклеиновой кислоты и часто содержат элементы или функциональные единицы в этих отрезках последовательностей нуклеиновой кислоты, такие как сайт начала транскрипции, сайт связывания для РНК-полимеразы, сайты связывания общего фактора транскрипции, такие как TATA-бокс, сайты связывания специфического фактора транскрипции и т. п. Также могут присутствовать дополнительные регуляторные последовательности, такие как энхансеры и иногда интроны, на конце промоторной последовательности. Такие функциональные единицы ниже в данном документе называются "структурными блоками", и их можно объединять в отрезок нуклеиновой кислоты для построения функциональной промоторной последовательности. Структурные блоки могут быть непосредственно прилегающими один к другому, но также могут быть разделены отрезками нуклеиновой кислоты, которая не играет прямой роли в функции промотора. Специалисту в данной области известно как проверить, являются ли нуклеотиды в составе отрезка нуклеиновой кислоты соответствующими функции промотора, и как удалить или добавить структурные блоки и/или нуклеотиды в данную промоторную последовательность посредством стандартных способов молекулярной биологии, например свести к минимуму его длину, сохраняя при этом активность промотора, или оптимизировать активность.
Используемые в данном документе термины "энхансер" или "структурный блок, представляющий собой энхансер" относятся к регуляторным последовательностям ДНК, например 50-1500 п. о., которые могут быть связаны белками (активирующими белками) для стимуляции или усиления транскрипции гена или нескольких генов. Данные активирующие белки (также известные как факторы транскрипции) взаимодействуют с медиаторным комплексом и способствуют привлечению полимеразы II и общих факторов транскрипции, которые затем начинают транскрипцию генов. Энхансеры обычно являются действующими в цис-положении, но могут быть расположены либо выше, либо ниже сайта начала транскрипции гена или генов, которые они регулируют. Кроме того, энхансер может быть либо в прямом, либо в обратном направлении и не должен быть расположен возле сайта инициации транскрипции, чтобы оказывать влияние на транскрипцию, так как некоторые были обнаружены на несколько сотен тысяч пар оснований выше или ниже сайта начала транскрипции. Энхансеры также можно обнаружить в интронах.
Термин "двунаправленный промотор" относится к непрерывным регуляторным последовательностям гена, которые кроме элементов, представляющих собой промотор, могут содержать элементы, представляющие собой энхансеры, и элементы, представляющие собой интроны, и определены как структурные блоки, которые описаны в данном документе. Данные двунаправленные промоторы управляют экспрессией гена в двунаправленном режиме, контролируя экспрессию трансгенов, помещенных на обе стороны последовательности двунаправленного промотора. Например, двунаправленный промотор по настоящему изобретению управляет транскрипцией двух разных трансгенов в двунаправленном режиме и включает энхансерный структурный блок, фланкированный первым промоторным структурным блоком с одной стороны и вторым промоторным структурным блоком с другой стороны так, что трансгены расположены ниже соответствующих промоторных структурных блоков. Следует отметить, что фланкированные и смежные необязательно означают непосредственно граничащие, так как между структурными блоками могут быть некоторые дополнительные нуклеотиды, но предпочтительно добавляют не слишком длинную дополнительную последовательность, таким образом двунаправленный промотор сохраняет компактный размер. Также следует отметить, что термины "выше" и "ниже" относятся к направлению транскрипции, и их применение является общеизвестным в данной области. Например, в соответствии с соглашением термины выше и ниже относятся к направлению от 5' к 3', в котором происходит транскрипция РНК. Под "выше" подразумевается направление к 5'-концу молекулы РНК, и под "ниже" подразумевается направление к 3'-концу. При рассмотрении двухцепочечной ДНК, под "выше" подразумевается направление к 5'-концу кодирующей цепи для рассматриваемого гена, и под "ниже" подразумевается направление к 3'-концу. В силу антипараллельной природы ДНК это означает, что 3'-конец матричной цепи располагается выше гена, и 5'-конец располагается ниже. См., например, на фиг. 1D, представлен предпочтительный двунаправленный промотор hCMV-rhCMV, содержащий основной немедленно-ранний энхансер цитомегаловируса человека (энхансер hCMV) в качестве энхансерного структурного блока, фланкированный основным немедленно-ранним промотором цитомегаловируса человека (hCMV) в качестве первого промоторного структурного блока на одной стороне и промотором CMV макака-резуса (rhCMV) в качестве второго промоторного структурного блока на другой стороне. Двунаправленный промотор hCMV-rhCMV по настоящему изобретению функционально связан с двумя трансгенами так, что первый трансген функционально связан с промоторным структурным блоком hCMV, и второй трансген функционально связан с промоторным структурным блоком rhCMV так, что как первый, так и второй трансгены расположены ниже от соответствующего промотора, и так, что первый и второй трансгены транскрибируются в наружном направлении от энхансера hCMV.
Предпочтительный двунаправленный промотор по настоящему изобретению представляет собой двунаправленный промотор hCMV-rhCMV, содержащий SEQ ID NO:4, с расположениями последовательностей для разных элементов, как показано на фиг. 3B, но специалисту в данной области будет понятно, что длина или идентичность последовательностей разных структурных блоков и вставочных последовательностей могут варьировать до некоторой степени, так что могут быть получены по существу аналогичные результаты. Например, разные энхансеры могут быть протестированы в отношении замещения, и/или энхансерные последовательности могут быть модифицированы, так что может быть получена по существу аналогичная экспрессия. Аналогично, интрон можно добавлять в смежном положении и ниже одного или обоих промоторных структурных блоков, и ожидается, что двунаправленный промотор hCMV-rhCMV будет сохранять активность. Возможно, это может привести даже к усиленной экспрессии, однако в любом случае это может происходить за счет места для трансгена, поскольку интрон(-ы) будут занимать место в векторе или вирусе. Энхансеры, указанные в данном документе, являются предпочтительными, имея подходящие размеры, обеспечивая сбалансированную экспрессию и устойчивые конструкции в отношении аденовирусного вектора. Структурные блоки двунаправленного промотора по настоящему изобретению как таковые могут быть известны по отдельности, но их никогда не объединяли, и даже не предполагали такого их объединения в конформации в соответствии с настоящим изобретением, что приводит к получению эффективного, очень сбалансированного и относительно короткого двунаправленного промотора. Как показано в данном документе, неожиданно было обнаружено, что, несмотря на наличие относительно короткой последовательности всего лишь из 943 нуклеотидов, эта новая двунаправленная комбинация энхансера hCMV с промоторами hCMV и rhCMV оказалась способной управлять эффективной и сбалансированной транскрипцией двух функционально связанных трансгенов, в то же время сохраняя стабильную конфигурацию двунаправленного промотора со связанными трансгенами в сложном контексте аденовирусного вектора. Из данных, представленных в данном документе, видно, что создание таких двунаправленных промоторов на основе известных аналогичных структурных блоков было непредсказуемым, так как несколько других аналогично построенных двунаправленных промоторов не имели сильной активности промотора и/или приводили к несбалансированной экспрессии, в результате чего экспрессия трансгена, функционально связанного с одной частью двунаправленного промотора, была выражена значительно сильнее (например, по меньшей мере 5-кратное различие) по сравнению с трансгеном, функционально связанным с другой частью такого двунаправленного промотора. Было заведомо невозможно спрогнозировать, будет ли вообще получен какой-либо промотор, который бы отвечал требованиям аналогичных уровней экспрессии из обоих сторон (например, менее чем 2-кратное различие между экспрессией из обоих сторон) и стабильности в контексте аденовирусных векторов. Настоящее изобретение, неожиданно, предусматривает двунаправленные промоторы, которые отвечают этим требованиям и имеют небольшой размер, что может быть очень выгодным в контексте ограничений по размеру векторов, несущих трансгены (т. е. можно расположить большие трансгены, и/или векторы могут оставаться более стабильными).
Дополнительные регуляторные последовательности также могут быть добавлены к конструкциям, содержащим двунаправленные промоторы по настоящему изобретению. В данном документе термин "регуляторная последовательность" используют взаимозаменяемо с термином "регуляторный элемент", и он относится к сегменту нуклеиновой кислоты, как правило без ограничения к ДНК, которая модулирует транскрипцию последовательности нуклеиновой кислоты, с которой она связана по функции, и таким образом действует как транскрипционный модулятор. Регуляторная последовательность часто содержит последовательности нуклеиновой кислоты, которые представляют собой транскрипционные домены связывания, которые распознаются доменами связывания нуклеиновой кислоты транскрипционных белков и/или факторов транскрипции, энхансерами или репрессорами и т. д. Например, регуляторная последовательность может включать одну или несколько последовательностей операторов оперона тетрациклина (tetO), так что экспрессия ингибируется в присутствии белка-репрессора оперона тетрациклина (tetR). В отсутствие тетрациклина белок tetR способен связываться с сайтами tetO и подавлять транскрипцию гена, функционально связанного с сайтами tetO. Однако в присутствии тетрациклина конформационное изменение белка tetR предотвращает его связывание с операторными последовательностями, что обеспечивает транскрипцию функционально связанных генов. В определенных вариантах осуществления rAd по настоящему изобретению может необязательно включать tetO, функционально связанный с двунаправленным промотором hCMV-rhCMV так, что экспрессия одного или нескольких трансгенов ингибируется в векторах, которые продуцируются в линии клеток-продуцентов, в которой экспрессируется белок tetR. Следовательно, экспрессия не будет ингибирована, если вектор введен субъекту или в клетки, которые не экспрессируют белок tetR (см. например, WO 07/073513). В других определенных вариантах осуществления вектор по настоящему изобретению может необязательно включать систему переключения генов с использованием кумата, в которой регуляция экспрессии опосредуется связыванием репрессора (CymR) с сайтом-оператором (CuO), помещенным ниже промотора (см., например, (Mullick et al., 2006)).
Используемый в данном документе термин "репрессор" относится к объектам (например, белкам или другим молекулам), обладающим способностью ингибировать, препятствовать, замедлять и/или подавлять продуцирование гетерологичного белкового продукта рекомбинантного вектора экспрессии. Например, с помощью воздействия на сайт связывания в подходящем месте вдоль вектора экспрессии, как например в кассете экспрессии. Примеры репрессоров включают tetR, CymR, lac-репрессор, trp-репрессор, gal-репрессор, лямбда-репрессор и другие соответствующие репрессоры, известные из уровня техники.
Кроме того, рекомбинантный вектор, вирус или аденовирус по настоящему изобретению содержит двунаправленный промотор hCMV-rhCMV, где транскрипционное направление (от 5' к 3') частей hCMV и rhCMV двунаправленного промотора hCMV-rhCMV расходится друг от друга, и где двунаправленный промотор hCMV-rhCMV функционально связан с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении. Таким образом, двунаправленный промотор будет направлять экспрессию первого трансгена к первому концу вектора или (адено)вирусного генома и второго трансгена к другому концу вектора или (адено)вирусного генома. Специалисту в данной области известно, что мутации могут быть осуществлены в представленных последовательностях и могут быть протестированы в отношении активности промотора с помощью обычных способов. Как правило, последовательность, имеющая по меньшей мере 90% идентичности с указанными промоторными последовательностями (не включая энхансерные последовательности), все еще будет иметь функциональную активность и, таким образом, будет считаться двунаправленным промотором hCMV-rhCMV. Таким образом, двунаправленный промотор hCMV-rhCMV по настоящему изобретению предпочтительно характеризуется по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с указанными промоторными последовательностями (кроме энхансерной последовательности). Предпочтительно двунаправленный промотор hCMV-rhCMV содержит последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 4. В определенных предпочтительных вариантах осуществления двунаправленный промотор hCMV-rhCMV содержит структурные блоки, представленные на фиг. 1D, где двунаправленный промотор hCMV-rhCMV содержит энхансер hCMV в качестве энхансерного структурного блока, фланкированный промотором hCMV в качестве первого промоторного структурного блока на одной стороне и промотором CMV макака-резуса (rhCMV) в качестве второго промоторного структурного блока на другой стороне. В определенном другом предпочтительном варианте осуществления двунаправленный промотор hCMV-rhCMV характеризуется 100% идентичности с SEQ ID NO:4 с расположениями последовательностей для разных элементов, как показано на фиг. 3, но специалисту в данной области будет понятно, что длина последовательностей разных структурных блоков и вставочных последовательностей может до некоторой степени варьировать, и идентичность элементов, представляющих собой энхансеры, также может варьировать, так что могут быть получены по существу аналогичные результаты.
Термины "функционально связан" или "связанный по функции" в данном документе применяются взаимозаменяемо и относятся к функциональной связи последовательностей нуклеиновой кислоты с регуляторными последовательностями нуклеотидов, таких как промоторы, энхансеры, транскрипционные и трансляционные стоп-сайты и другие сигнальные последовательности, и указывают, что два или более сегментов ДНК соединены вместе, так что они функционируют согласно их предполагаемым целям. Например, связь по функции последовательностей нуклеиновой кислоты, как правило ДНК, с регуляторной последовательностью или промоторным участком относится к физической и функциональной связи между ДНК и регуляторной последовательностью или промотором, так что транскрипция такой ДНК инициируется от регуляторной последовательности или промотора с помощью РНК-полимеразы, которая специфически распознает, связывает и транскрибирует ДНК. С целью оптимизировать экспрессию и/или транскрипцию in vitro, может быть необходимо модифицировать регуляторную последовательность для экспрессии нуклеиновой кислоты или ДНК в типе клеток, для которых она экспрессируется. Целесообразность или необходимость такой модификации может быть установлена эмпирическим путем.
При экспрессии, контролируемой любой частью двунаправленного промотора hCMV-rhCMV, трансген является эффективно экспрессированным. Используемые в данном документе термины "эффективно экспрессируемый" или "эффективная экспрессия" означают, что экспрессия из любой части двунаправленного промотора hCMV-rhCMV, измеренная, например, посредством разных методик выявления белка, таких как вестерн-блоттинг, анализ FACS или другие виды анализа с использованием люминесценции или флуоресценции, составляет по меньшей мере 10%, предпочтительно по меньшей мере 20%, более предпочтительно по меньшей мере 30% от экспрессии из моновалентного вектора, экспрессирующего один трансген под контролем однонаправленного промотора hCMV (с SEQ ID NO: 9). Следует отметить, что однонаправленный промотор hCMV намного сильнее по сравнению с другими широко используемыми однонаправленными промоторами, такими как промоторы PGK, UBI C или RSV LTR (Powell, Rivera-Soto, & Gray, 2015). Таким образом, двунаправленный промотор, который является менее сильным, чем промотор hCMV (например, приводящий к уровню экспрессии, который составляет по меньшей мере 10% от такого однонаправленного промотора hCMV) может все еще считаться эффективным. Промоторы hCMV получены из основного немедленно-раннего (mIE) участка цитомегаловируса человека и часто применяются для эффективной однонаправленной экспрессии гена в векторах для вакцины и генной терапии. Например, последовательность промотора hCMV можно получить из локуса mIE штамма AD169 hCMV (X03922), и она включает сайты связывания NF1, энхансерный участок, TATA-бокс и часть первого экзона. Известны другие последовательности промотора hCMV, которые могут быть короче (например, содержащие только энхансерный и промоторный участок и не содержащие сайтов связывания NF1) или длиннее (например включающие дополнительные сайты связывания клеточных факторов и первую интронную последовательность). Как оказалось, данные промоторы hCMV, которые различаются длиной, являются эффективными, повсеместно активными промоторами. Для сравнения уровней экспрессии, описанных в данном документе, промоторная последовательность hCMV представляла собой SEQ ID NO:9. Например, уровень экспрессии из любой части двунаправленного промотора hCMV-rhCMV по настоящему изобретению в rAd составляет предпочтительно по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 95% от уровня экспрессии из rAd, где трансген находится под контролем однонаправленного промотора hCMV с SEQ ID NO:9. В определенных вариантах осуществления уровень экспрессии из любой части двунаправленного промотора hCMV-rhCMV составляет приблизительно 10-60%, например 20-50%, например, приблизительно 30% от уровня экспрессии из rAd, где трансген находится под контролем однонаправленного промотора hCMV с SEQ ID NO:9. Кроме того, известно, что в случае rAd, экспрессирующего один антиген под контролем промотора hCMV, экспрессия является достаточной для вызова значительных иммунных ответов с участием T-клеток и B-клеток. Аналогично, ожидается, что экспрессия двух трансгенов, экспрессируемых с помощью двунаправленного промотора hCMV-rhCMV по настоящему изобретению, будет вызывать значительный иммунный ответ с участием T-клеток и B-клеток на оба трансгена. Например, если два трансгена кодируют антигены, которые вызывают иммунный ответ при введении субъекту, то ожидается, что эффективная экспрессия двух трансгенов будет вызывать измеряемый иммунный ответ на оба антигена.
Экспрессия из обоих сторон двунаправленного промотора hCMV-rhCMV также является очень сбалансированной. Используемые в данном документе термины "сбалансированная экспрессия", "сбалансированность экспрессии", "баланс экспрессии" или "сбалансированная" в отношении экспрессии означают, что экспрессия из одной стороны двунаправленного промотора, измеренная, например, посредством разных методик выявления экспрессии белка, таких как вестерн-блоттинг, анализ FACS или другие виды анализа с использованием люминесценции или флуоресценции, сравнима с экспрессией из другой стороны двунаправленного промотора. Например, уровень экспрессии из одной стороны двунаправленного промотора hCMV-rhCMV по настоящему изобретению предпочтительно составляет по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% уровня экспрессии из другой стороны двунаправленного промотора. В определенных вариантах осуществления уровень экспрессии из одной стороны двунаправленного промотора hCMV-rhCMV составляет приблизительно 70-130%, например 80-120%, например 90-110%, например приблизительно 100% от уровня экспрессии из другой стороны двунаправленного промотора. В другом примере соотношение экспрессии из двух сторон двунаправленного промотора hCMV-rhCMV находится в диапазоне 1/1, 1,1/1, 1,2/1, 1,3/1, 1,4/1, 1,5/1, 1,6/1, 1,7/1, 1,8/1, 1,9/1 и 2/1. Кроме того, известно, что в случае rAd, экспрессирующего один антиген под контролем промотора hCMV, экспрессия является достаточной для вызова значительных иммунных ответов с участием T-клеток и B-клеток. Таким образом, сбалансированная экспрессия двух антигенов, экспрессируемых с помощью двунаправленного промотора hCMV-rhCMV по настоящему изобретению, может, вероятно, вызывать сопоставимый иммунный ответ с участием T-клеток и B-клеток на оба антигена, хотя это также может зависеть от экспрессии антигена с течением времени и свойственной самим антигенам способности вызывать иммунные ответы определенных типов. Таким образом, двунаправленный промотор hCMV-rhCMV по настоящему изобретению является улучшенным в отношении сбалансированности экспрессии по сравнению с двунаправленным промотором mCMV. Для сравнения, уровень экспрессии антигена, расположенного на правой стороне (3'-конец) двунаправленного промотора mCMV, был примерно в 10 раз выше, по сравнению с таковым антигена, расположенного на левой стороне (5'-конец) двунаправленного промотора mCMV (который был описан в WO 2016/166088), который уже считался относительно хорошо сбалансированным, но, очевидно, является гораздо менее сбалансированным, чем двунаправленный промотор по настоящему изобретению.
Важным аспектом векторов, как ДНК-векторов, таких как плазмидные векторы, так и вирусных векторов, таких как аденовирусные векторы, является способность этих векторов размещать необходимые трансгенные последовательности. Такая способность может ограничиваться ограничениями по размеру векторов, которые, например, могут стать нестабильными, или даже их производство может стать невозможным, если превышены определенные пределы размеров. Место, занимаемое промотором, особенно двунаправленным промотором, который может контролировать экспрессию более чем одного трансгена, является, таким образом, важным фактором, который следует учитывать при разработке новых векторов, помимо тех функциональных способностей, которыми должны обладать такие промоторы. Данный двунаправленный промотор имеет преимущество в том, что он является относительно коротким, что означает, что при определенном пределе размера вектора остается больше места для трансгена, например, позволяя включить больше эпитопов, если трансген представляет собой иммуноген, или позволяя осуществлять экспрессию больших белков, по сравнению с другими двунаправленными промоторами большего размера.
Термины "кодирующая последовательность", "последовательность, которая кодирует", или "кодирующая" в данном документе применяются взаимозаменяемо и относятся к последовательности нуклеиновой кислоты, которая транскрибируется (ДНК) и транслируется (mRNA) в полипептид in vitro или in vivo, при функциональной связи с соответствующими регуляторными последовательностями.
Сигнал полиаденилирования, например сигнал полиА гена бычьего гормона роста (U.S. 5122458), может располагаться позади трансгенов. Предпочтительно, каждый трансген имеет сигнал полиА, и предпочтительно сигнал полиА для первого трансгена отличается от сигнала полиА для второго трансгена. В одном варианте осуществления первый сигнал полиА представляет собой сигнал полиА SV40, и второй сигнал полиА представляет собой сигнал полиА гена бычьего гормона роста.
Последовательность, содержащую интрон, можно также размещать на одном или на обоих концах двунаправленного промотора по настоящему изобретению. Например, известно, что интроны могут повышать экспрессию белка, в частности in vivo. Интрон, используемый в данном документе, имеет нормальную функцию и структуру, как известно из уровня техники, и представляет собой полинуклеотидную последовательность в нуклеиновой кислоте, которая не кодирует информацию для синтеза белка и удаляется перед трансляцией информационной РНК посредством процесса, известного как сплайсинг. Интрон содержит сайт донора сплайсинга (5'-конец интрона, обычно последовательность GU) и сайт акцептора сплайсинга (3'-конец интрона, обычно последовательность GA). В соответствии с настоящим изобретением потенциально можно применять множество разных интронов, хотя предпочтительным является применение относительно коротких интронов и интронов, модифицированных в более короткие интроны, чтобы не занимать слишком много места в вирусном векторе, так что для трансгенов в рекомбинантном аденовирусе остается больше места. Предпочтительным является применение первого интрона на одной стороне двунаправленного промотора и второго, отличающегося интрона, на другой стороне двунаправленного промотора, т. е. каждому трансгену предшествует разная интронная последовательность. В определенных вариантах осуществления интрон может быть химерным интроном. Специалисту известно, что много разных интронов являются доступными, и их можно применять. Однако, преимуществом данного промотора является то, что для надлежащей экспрессии ему не требуются такие интроны, и, следовательно, в предпочтительных вариантах осуществления между промоторными структурными блоками двунаправленного промотора и соответствующими трансгенами на обеих сторонах интроны отсутствуют.
Двунаправленный промотор по настоящему изобретению может в определенных вариантах осуществления применяться, например, для направления экспрессии двух антигенов с целью вызвать иммунный ответ на эти антигены в применении в вакцинах. Однако специалисту в данной области сразу станет ясно, что уровни сбалансированной экспрессии трансгена также могут быть существенными для трансгенов, для которых иммунный ответ не является основной целью, например для двух разных трансгенов, которые применяют для целей генной терапии, для экспрессии комплексов гетерологичных белков или для пропорциональной экспрессии двух цепей антитела. Таким образом, настоящее изобретение может быть осуществлено на практике с любой комбинацией трансгенов, для которых является необходимой экспрессия из одного рекомбинантного вектора, например, аденовирусного вектора. Следовательно, идентичность трансгена, подходящего, например, для векторов или аденовирусов, содержащих какой-либо трансген, не является темой настоящего изобретения. Подходящие трансгены хорошо известны специалисту в данной области и, к примеру, могут включать открытые рамки считывания трансгенов, к примеру открытые рамки считывания, кодирующие полипептиды с терапевтическим эффектом, например для целей генной терапии, или полипептиды, против которых необходим иммунный ответ, если вектор на основе rAd применяют для целей вакцинации. Особенно предпочтительными гетерологичными нуклеиновыми кислотами являются представляющие интерес гены, кодирующие антигенные детерминанты, в отношении которых необходимо повышение иммунного ответа. Как правило, такие антигенные детерминанты также называют антигенами. Если рекомбинантный аденовирус вводят субъекту, будет повышаться иммунный ответ против антигена(-ов). Любой необходимый антиген может кодироваться аденовирусным вектором. В типичных вариантах осуществления в соответствии с настоящим изобретением антигены представляют собой пептиды, полипептиды или белки из организмов, которые могут вызывать заболевание или патологическое состояние. Следовательно, в дополнительном предпочтительном варианте осуществления указанная гетерологичная нуклеиновая кислота, представляющая интерес, кодирует иммуногенную (или антигенную) детерминанту. Более предпочтительно указанная иммуногенная детерминанта представляет собой антиген из бактерии, вируса, дрожжей или паразита. Заболевания, вызванные такими организмами, обычно называют "инфекционными заболеваниями" (и, таким образом, не ограничены организмами, которые "инфицируют", но также включают такие, которые попадают в хозяина и вызывают заболевание). Так называемые "аутоантигены", например опухолевые антигены, также образуют часть существующего уровня техники и могут кодироваться гетерологичными нуклеиновыми кислотами в рекомбинантных аденовирусах в соответствии с настоящим изобретением. Неограничивающими примерами, из которых выбраны антигенные детерминанты (или антигены), являются организмы, вызывающие малярию, как, например, Plasmodium falciparum, организмы, вызывающие туберкулез, как, например, Mycobacterium tuberculosis, дрожжи или вирусы. В других предпочтительных вариантах осуществления антигены из таких вирусов, как флавивирусы (например, вирус лихорадки Западного Нила, вирус гепатита C, вирус японского энцефалита, вирус денге), вирус Эбола, вирус иммунодефицита человека (HIV) и вирус Марбург, можно применять в композициях в соответствии с настоящим изобретением. В одном варианте осуществления указанный антиген представляет собой белок CS или его иммуногенную часть из P. falciparum (например, аденовирусные векторы, кодирующие CS, см., например, (Havenga et al., 2006; Ophorst et al., 2006); WO 2004/055187, все из которых включены в данный документ в полном объеме посредством ссылки). В другом варианте осуществления антигенная детерминанта представляет собой белок одного антигена или слитый белок нескольких антигенов из M. tuberculosis, такой как белки Ag85A, Ag85B и/или TB10.4, или их иммуногенную(-ые) часть(части) (о конструировании и получении таких противотуберкулезных вирусных вакцин см., например, WO 2006/053871, включенный в данный документ посредством ссылки). В еще одном варианте осуществления указанная антигенная детерминанта представляет собой вирусный гликопротеин или его иммуногенную часть, такую как GP из филовируса, такого как вирус Эбола или вирус Марбург (например, (Geisbert et al., 2011; Sullivan et al., 2006; Sullivan et al., 2003). В еще одних вариантах осуществления указанная иммуногенная детерминанта получена из белка HIV, такого как gag, pol, env, nef или их вариантов (в отношении примеров вакцин против HIV на основе аденовируса, см., например, в WO 2009/026183, WO 2010/096561, WO 2006/120034, WO 02/22080, WO 01/02607). В других вариантах осуществления указанная антигенная детерминанта представляет собой белок HA, NA, M или NP или иммуногенную часть любого из них из вируса гриппа (например, (Hu et al., 2011; Zhou et al., 2010); обзорная публикация (Vemula & Mittal, 2010)). В других вариантах осуществления антигенная детерминанта представляет собой белок HA или его иммуногенную часть из вируса кори (например, WO 2004/037294). В других вариантах осуществления антигенная детерминанта представляет собой гликопротеин из вируса бешенства (например, (Zhou, Cun, Li, Xiang, & Ertl, 2006)). В дополнительных вариантах осуществления антиген представляет собой антиген из респираторного синцитиального вируса (RSV), например белок RSV F (см., например, WO 2013/139911 и WO 2013/139916), или белок RSV G, или оба, или другие белки RSV. В других вариантах осуществления антиген представляет собой антиген из другого вируса, такого как вирус папилломы человека или другие вирусы и т. д. Рекомбинантный аденовирус может кодировать два разных антигена из такого же организма. Рекомбинантный аденовирус также может кодировать комбинации антигенов из разных организмов, например первый антиген из первого организма и второй антиген из второго организма. Также является возможным кодировать антиген и, к примеру, адъювант в тот же аденовирус, например, антиген и агонист толл-подобного рецептора (TLR), такой как агонист TLR3, как, например, dsRNA или его миметик и подобное (например, WO 2007/100908). В определенных вариантах осуществления рекомбинантный вектор, например рекомбинантный аденовирус, кодирует два разных антигена, при этом каждый находится под контролем двунаправленного промотора hCMV-rhCMV. В других вариантах осуществления вектор или рекомбинантный (адено)вирус кодирует антиген и иммуномодулятор, при этом каждый находится под контролем двунаправленного промотора hCMV-rhCMV. В определенных вариантах осуществления дополнительные гетерологичные последовательности или трансгены могут присутствовать в векторе или рекомбинантном (адено)вирусе, помимо первого и второго трансгенов, которые находятся под контролем двунаправленного промотора hCMV-rhCMV.
Настоящее изобретение также предусматривает способ получения генетически стабильного рекомбинантного аденовируса, содержащего первый и второй трансгены, где каждый эффективно экспрессируется, при инфицировании аденовирусом целевой клетки, при этом способ включает:
получение конструкции, содержащей двунаправленный промотор hCMV-rhCMV, функционально связанный с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении, и встраивание указанной конструкции в геном рекомбинантного аденовируса. Получение конструкции, соответственно, охватывает применение стандартных способов молекулярного клонирования, которые хорошо известны (см., например, (Holterman et al., 2004; Lemckert et al., 2006; Vogels et al., 2003); Sambrook, Fritsch and Maniatis, Molecular Cloning: A Laboratory Manual, 2nd edition, 1989; Current Protocols in Molecular Biology, Ausubel FM, et al, eds, 1987; серия Methods in Enzymology (Academic Press, Inc.); PCR2: A Practical Approach, MacPherson MJ, Hams BD, Taylor GR, eds, 1995), как известно специалисту и обычно выполняются в области технологии рекомбинантных аденовирусов, и приведены в данном документе в качестве примера. Двунаправленный промотор hCMV-rhCMV обладает признаками, которые описаны выше, и может быть получен посредством обычных способов. Для удобства специалист может осуществлять манипуляции с геномом аденовируса посредством клонирования в небольшие фрагменты, такие как первая часть для левой части генома до участка E1 для простоты манипуляции и введения трансгенов в плазмидную форму, и вторая, большая, часть для остального генома, которая может в результате рекомбинации с первой частью привести к образованию полного генома аденовируса (см., например WO 99/55132).
rAd по настоящему изобретению имеет преимущество в том, что он может экспрессировать два трансгена и остается генетически стабильным, в отличие от аденовирусов, полученных посредством различных альтернативных подходов экспрессирования двух трансгенов, которые предусматриваются в уровне техники, при этом также обеспечивая сбалансированную экспрессию двух трансгенов, осуществляемую при участии двунаправленного промотора. Таким образом, применение двунаправленного промотора hCMV-rhCMV позволяет решить проблему генетической нестабильности аденовирусов, которые экспрессируют два трансгена, и несбалансированной экспрессии двух трансгенов, и благодаря его относительно небольшому размеру обеспечивает значительно больше места для трансгенных последовательностей, чем при применении определенных других двунаправленных промоторов, которые имеют больший размер (например, он является приблизительно на 1 т. п. о. короче, чем двунаправленный промотор mCMV, таким образом теоретически в данном векторе с ограничением емкости можно разместить трансгены, которые суммарно приблизительно на 1 т. п. о. длиннее, чем в таком же векторе, где трансгены направляются промотором mCMV).
Для тестирования генетической стабильности rAd "спасают" и размножают в соответствующей клеточной линии, например вспомогательной клеточной линии PER.C6®. Вирусную ДНК выделяют при определенном количестве пассажей и целостность генома rAd можно анализировать посредством одного или нескольких из следующих: анализа ПЦР для определения наличия участка трансгена или отсутствия полос делеции, рестрикционного расщепления генома rAd для определения наличия или отсутствия различий по фрагментам рестрикции и/или секвенирования генома rAd или продуктов ПЦР генома rAd для определения наличия или отсутствия мутаций в последовательностях rAd. В отношении rAd по настоящему изобретению, "генетически стабильный" означает, что нуклеотидная последовательность не изменится в составе плазмид, применяемых для получения rAd, на более поздние стадии продуцирования rAd, так что rAd, экспрессирующий два трансгена, имеет такую же генетическую стабильность, как сравнимый rAd с одним трансгеном (например, позади промотора hCMV), подходящую для получения партий большого масштаба. Например, анализ ПЦР с применением праймеров, фланкирующих кассету экспрессии, который не показывает фрагментов делеции (полос) по сравнению с более ранними номерами пассажей rAd или исходным материалом, и/или секвенированием продукта ПЦР участков E1, E3 и E4, подтверждает, что нуклеотидная последовательность не изменяется. Предпочтительно секвенирование участка, содержащего кассету экспрессии с двунаправленным промотором подтверждает, что нуклеотидная последовательность не изменяется на участке, содержащем кассету экспрессии.
Генетическая стабильность в данном исследовании тщательно оценивается, по сравнению с другими способами тестирования, как, например, тестовое расщепление в одиночной партии вируса. Чувствительность анализа повышают с помощью следующих способов: несколько вирусных популяций (бляшек) выделяют и подвергают длительному пассированию. Длительное пассирование, объединенное с анализом ПЦР с применением праймеров, фланкирующих кассету экспрессии, позволяет выявить малую долю мутантов с делецией в популяции rAd, что может быть упущено при применении других способов. Кроме того, выполняется анализ секвенирования для исключения возникновения точечных мутаций, таких как введение стоп-кодонов в открытую рамку считывания трансгена. Более конкретно, так как вирусные мутации всегда представляют случайное событие, одна бляшка может быть стабильной, тогда как другая может представлять полосу делеции. Следовательно, для правильной оценки генетической стабильности необходимо протестировать несколько вирусных популяций (бляшек). В случае появления мутации, что позволяет вектору осуществлять репликацию эффективнее, чем родительский вектор, это может привести к увеличению количества мутантных версий, что часто наблюдается только после длительного пассирования, как описано в данном исследовании. Предпочтительно, rAd по настоящему изобретению является генетически стабильным в течение по меньшей мере 10 пассажей и даже более предпочтительно в течение по меньшей мере 13 пассажей в применяемой тестовой системе, так что вирус является достаточно стабильным для производственных циклов большого масштаба. Недавно было обнаружено, что рекомбинантный аденовирус, который содержит два трансгена, которые находятся под контролем двунаправленного промотора mCMV, является генетически стабильным, см. например, WO 2016/166088 (в котором также описано, что разные другие решения, которые были описаны в данной области техники для экспрессии двух антигенов из одного вектора, не привели к получению стабильного rAd или эффективной экспрессии, так что промотор mCMV был описан в нем как наиболее предпочтительное решение этой проблемы). В настоящей заявке показано, что рекомбинантный аденовирус, который имеет два трансгена, которые находятся под контролем двунаправленного промотора hCMV-rhCMV по настоящему изобретению, также является генетически стабильным и, более того, характеризуется более сбалансированной экспрессией двух трансгенов по сравнению с ситуацией, когда они находятся под контролем двунаправленного промотора mCMV.
Рекомбинантные аденовирусы, полученные в соответствии со способами по настоящему изобретению, могут быть получены в соответствии с вариантами осуществления, описанными выше для рекомбинантного аденовируса.
Настоящее изобретение также предусматривает способ экспрессирования по меньшей мере двух трансгенов в клетке, при этом способ включает обеспечение клетки вектором или рекомбинантным вирусом, например рекомбинантным аденовирусом в соответствии с настоящим изобретением. Доставку в клетку рекомбинантного аденовируса можно осуществлять посредством введения аденовируса субъекту или посредством введения (например, инфицирования) аденовируса в клетку in vitro или ex vivo. В определенных вариантах осуществления настоящее изобретение предусматривает рекомбинантный аденовирусный вектор для применения в экспрессировании по меньшей мере двух трансгенов в клетке, например посредством введения рекомбинантного аденовируса субъекту.
Настоящее изобретение также предусматривает способ индуцирования иммунного ответа против по меньшей мере двух антигенов, включающий введение субъекту вектора, например рекомбинантного (адено)вируса в соответствии с настоящим изобретением. Настоящее изобретение также предусматривает вектор или рекомбинантный (адено)вирус в соответствии с настоящим изобретением для применения в индуцировании иммунного ответа против по меньшей мере двух антигенов.
Настоящее изобретение также предусматривает молекулу рекомбинантной ДНК, содержащую двунаправленный промотор hCMV-rhCMV по настоящему изобретению или геном рекомбинантного аденовируса по настоящему изобретению. Специалисту в данной области будет известно, что это также может быть комбинация по меньшей мере двух разных молекул рекомбинантной ДНК, которые вместе могут образовать одну молекулу рекомбинантной ДНК по настоящему изобретению. Такие молекулы являются применимыми в осуществлении манипуляций с геномом и создании новых рекомбинантных аденовирусов. Геном кодирует белки, которые требуются для репликации аденовируса и упаковки в подходящих клетках.
Термин "приблизительно" для числовых значений, используемых в данном изобретении, означает значение ± 10%.
Клетки-продуценты культивируют для увеличения количества клеток и вирусов и/или титров вирусов. Культивирование клетки выполняют с обеспечением ей возможности метаболизма, и/или роста, и/или деления, и/или получения вируса, представляющего интерес в соответствии с настоящим изобретением. Этого можно достичь с помощью способов, которые хорошо известны специалистам в данной области и включают без ограничения обеспечение клетки питательными веществами, к примеру в соответствующей питательной среде. Подходящие питательные среды хорошо известны специалистам в данной области и обычно могут быть получены в больших количествах из коммерческих источников или произведены по заказу согласно стандартным протоколам. Культивирование можно проводить, к примеру, в чашках, роллер-флаконах или в биореакторах с применением периодических, подпитываемых, непрерывных систем и т. п. Известны подходящие условия для культивирования клеток (см., например, Tissue Culture, Academic Press, Kruse and Paterson, editors (1973) и R.I. Freshney, Culture of animal cells: A manual of basic technique, fourth edition (Wiley-Liss Inc., 2000, ISBN 0-471-34889-9).
Как правило, аденовирус будут подвергать воздействию соответствующей клетки-продуцента в культуре, обеспечивая возможность поглощения вируса. Обычно оптимальное перемешивание составляет от приблизительно 50 до 300 об./мин., как правило, приблизительно 100-200, например приблизительно 150, как правило, DO составляет 20-60%, например 40%, оптимальный pH составляет от 6,7 до 7,7, оптимальная температура составляет от 30 до 39°C, например 34-37°C, оптимальная MOI составляет от 5 до 1000, например приблизительно 50-300. Как правило, аденовирус инфицирует клетки-продуценты спонтанно, и приведение клеток-продуцентов в контакт с частицами rAd является достаточным для инфицирования клеток. Обычно посевной материал аденовируса добавляют в культуру для инициации инфицирования, и в дальнейшем аденовирус размножается в клетках-продуцентах. Все это является обычным для специалиста в данной области.
После инфицирования аденовирусом вирус реплицируется внутри клетки и таким образом амплифицируется, процесс, называемый в данном документе размножением аденовируса. В итоге аденовирусная инфекция приводит к лизису инфицированной клетки. Следовательно, литические характеристики аденовируса обеспечивают возможность получения вируса двумя различными способами. Первый способ представляет собой сбор вируса до лизиса клетки с применением внешних факторов для лизирования клеток. Второй способ представляет собой сбор вирусной надосадочной жидкости после (почти) полного лизиса клеток полученным вирусом (см., например, U.S. 6485958, описывающий сбор аденовируса без лизиса клеток-хозяев с помощью внешних факторов). Предпочтительным является применение внешних факторов для активного лизирования клеток для сбора аденовируса.
Способы, которые можно использовать для активного лизиса клеток, известны специалисту в данной области, и они рассмотрены, например, в WO 98/22588, p. 28-35. Применимыми в этом отношении способами являются, например, замораживание-размораживание, разрушение клеток абразивными материалами, гипертонический и/или гипотонический лизис, разрушение клеток жидкими материалами, разрушение ультразвуком, экструзия под высоким давлением, лизис детергентом, комбинации вышеперечисленных и подобное. В одном варианте осуществления настоящего изобретения клетки лизируют с использованием по меньшей мере одного детергента. Преимущество использования детергента для лизиса заключается в том, что этот способ простой и его легко масштабировать.
Детергенты, которые можно использовать, и способ их применения обычно известны специалисту в данной области. Например, несколько примеров рассматриваются в WO 98/22588, p. 29-33. Детергенты могут включать анионные, катионные, цвиттерионные и неионные детергенты. Концентрация детергента может варьировать, например, в диапазоне приблизительно 0,1% - 5% (вес/вес). В одном варианте осуществления используемым детергентом является Triton X-100.
Нуклеазы можно использовать для удаления загрязняющих, т. е. главным образом из клетки-продуцента, нуклеиновых кислот. Иллюстративные нуклеазы, подходящие для применения в настоящем изобретении, включают Benzonase®, Pulmozyme® или любую другую ДНКазу и/или РНКазу, широко используемые в данной области. В предпочтительных вариантах осуществления нуклеаза представляет собой Benzonase®, которая быстро гидролизирует нуклеиновые кислоты путем гидролиза внутренних фосфодиэфирных связей между определенными нуклеотидами, за счет чего тем самым уменьшается вязкость клеточного лизата. Benzonase® может быть коммерчески доступна от Merck KGaA (код W214950). Концентрация, при которой используют нуклеазу, предпочтительно находится в диапазоне 1-100 единиц/мл. В качестве альтернативы или в дополнение к обработке нуклеазой также является возможным селективно осаждать ДНК клетки-хозяина из препаратов аденовируса во время очистки аденовируса с применением селективных осаждающих средств, таких как домифена бромид (см., например, U.S. 7326555; (Goerke, To, Lee, Sagar, & Konz, 2005); WO 2011/045378; WO 2011/045381).
Способы сбора аденовируса из культур клеток-продуцентов подробно описаны в WO 2005/080556.
В определенных вариантах осуществления собранный аденовирус дополнительно очищают. Очистку аденовируса можно осуществлять в несколько этапов, предусматривающих осветление, ультрафильтрацию, диафильтрацию или разделение посредством хроматографии, как описано, к примеру, в WO 05/080556, включенном в данный документ посредством ссылки. Осветление можно осуществлять с помощью этапа фильтрации, удаляя клеточный детрит и другие примеси из клеточного лизата. Ультрафильтрацию используют для концентрирования раствора вируса. Диафильтрация или замена буфера с применением ультрафильтров представляет собой способ удаления или замены солей, сахаров и подобного. Специалист в данной области знает, как подобрать оптимальные условия для каждого этапа очистки. Также в WO 98/22588, включенном в данный документ посредством ссылки в полном объеме, описываются способы получения и очистки аденовирусных векторов. Способы включают выращивание клеток-хозяев, инфицирование клеток-хозяев аденовирусом, сбор и лизирование клеток-хозяев, концентрирование неочищенного лизата, замену буфера в неочищенном лизатe, обработку лизата нуклеазой и дополнительную очистку вируса с применением хроматографии.
Предпочтительно при очистке используется по меньшей мере один этап хроматографии, как, например, рассматривается в WO 98/22588, p. 61-70. Описано много способов дополнительной очистки аденовирусов, где в способ включены этапы хроматографии. Специалист в данной области будет осведомлен о данных способах и может вносить изменения в точный способ применения хроматографических стадий для оптимизации способа. К примеру, возможна очистка аденовирусов с помощью этапов анионообменной хроматографии, см., к примеру, WO 2005/080556. Было описано много других способов очистки аденовируса, и они доступны специалисту. Дополнительные способы получения и очистки аденовирусов раскрыты, например, в WO 00/32754, WO 04/020971, WO 2006/108707 и U.S. 5837520 и 6261823, все из которых включены в данный документ посредством ссылки.
Для введения людям в настоящем изобретении могут применяться фармацевтические композиции, содержащие вектор или рекомбинантный вирус, например rAd, и фармацевтически приемлемые носитель или вспомогательное вещество. В настоящем контексте термин "фармацевтически приемлемый" означает, что носитель или вспомогательное вещество в используемых дозах и концентрациях не вызовет каких-либо нежелательных или неблагоприятных эффектов у субъектов, которым их вводят. Такие фармацевтически приемлемые носители и вспомогательные вещества хорошо известны из уровня техники (см. Remington's Pharmaceutical Sciences, 18th edition, A. R. Gennaro, Ed., Mack Publishing Company [1990]; Pharmaceutical Formulation Development of Peptides and Proteins, S. Frokjaer and L. Hovgaard, Eds., Taylor & Francis [2000]; и Handbook of Pharmaceutical Excipients, 3rd edition, A. Kibbe, Ed., Pharmaceutical Press [2000]). Очищенный rAd предпочтительно составляют и вводят в виде стерильного раствора, хотя также возможно применение лиофилизированных препаратов. Стерильные растворы получают с помощью стерилизующей фильтрации или с помощью других способов, известных per se в данной области. Затем растворы лиофилизируют или заполняют ими контейнеры, предназначенные для лекарственных форм. Значение pH раствора обычно находится в диапазоне pH от 3,0 до 9,5, например pH от 5,0 до 7,5. Как правило, rAd находится в растворе с подходящим буфером, при этом раствор rAd может также содержать соль. Необязательно может присутствовать стабилизирующее средство, такое как альбумин. В определенных вариантах осуществления добавляют детергент. В определенных вариантах осуществления rAd можно составлять в виде инъекционного препарата. Данные составы, содержащие эффективные количества rAd, являются либо стерильными жидкими растворами, жидкими суспензиями либо лиофилизированными вариантами и необязательно содержат стабилизаторы или наполнители. Также аденовирусная вакцина может быть в виде аэрозоля для интраназального введения (см., например, WO 2009/117134).
К примеру, аденовирус можно хранить в буфере, который также применяют для Adenovirus World Standard (Hoganson et al., 2002): 20 мМ Tris с pH 8, 25 мМ NaCl, 2,5% глицерина. Другой применимый буфер для составления, подходящий для введения людям, представляет собой 20 мМ Трис, 2 мМ MgCl2, 25 мМ NaCl, 10% вес/об. сахарозы, 0,02% вес/об. полисорбата-80. Другой буфер для составления, который подходит для рекомбинантного аденовируса, содержит 10-25 мМ цитратного буфера с pH 5,9-6,2, 4-6% (вес/вес) гидроксипропил-бета-циклодекстрина (HBCD), 70-100 мМ NaCl, 0,018-0,035% (вес/вес) полисорбата-80 и необязательно 0,3-0,45% (вес/вес) этанола. Безусловно, можно применять много других буферов, и известно несколько примеров подходящих составов для хранения и для фармацевтического введения препаратов очищенного (адено)вируса, в том числе те, которые, к примеру, можно найти в EP0853660, U.S. 6225289 и в WO 99/41416, WO 99/12568, WO 00/29024, WO 01/66137, WO 03/049763, WO 03/078592, WO 03/061708.
В определенных вариантах осуществления композиция, содержащая аденовирус, дополнительно содержит один или несколько адъювантов. Адъюванты, известные в данной области, для дополнительного повышения иммунного ответа на применяемую антигенную детерминанту, и фармацевтические композиции, содержащие аденовирус и подходящие адъюванты, раскрыты, например, в WO 2007/110409, включенной в данный документ посредством ссылки. Термины "адъювант" и "иммуностимулятор" используются в данном документе взаимозаменяемо, и их определяют как одно или несколько веществ, вызывающих стимуляцию иммунной системы. В данном контексте адъювант используют для усиления иммунного ответа на аденовирусные векторы по настоящему изобретению. Примеры подходящих адъювантов включают соли алюминия, такие как гидроксид алюминия и/или фосфат алюминия; композиции на основе масляной эмульсии (композиции масло-в-воде), в том числе эмульсии сквалена в воде, такие как MF59 (см., например, WO 90/14837); составы на основе сапонина, такие как, например QS21 и иммуностимулирующие комплексы (ISCOMS) (см., например, U.S. 5057540 и WO 90/03184, WO 96/11711, WO 2004/004762, WO 2005/002620); производные бактерий или микроорганизмов, примеры которых представляют собой монофосфориллипид A (MPL), 3-O-деацетилированный MPL (3dMPL), олигонуклеотиды, содержащие мотив CpG, ADP рибозилированные бактериальные токсины или их мутанты, такие как термолабильный энтеротоксин LT E. coli, холерный токсин CT и подобное. Также является возможным применение адъюванта, кодируемого вектором, например посредством применения гетерологичной нуклеиновой кислоты, кодирующей слияние домена олигомеризации C4-связывающего белка (C4bp) с антигеном, представляющим интерес (Ogun, Dumon-Seignovert, Marchand, Holder, & Hill, 2008), или гетерологичной нуклеиновой кислоты, кодирующей агонист толл-подобного рецептора (TLR), такой как агонист TLR3, как, например, dsRNA (см., например, WO 2007/100908), или подобное. Такой rAd в соответствии с настоящим изобретением может, к примеру, кодировать антиген, представляющий интерес, на одной стороне двунаправленного промотора и агонист TLR3 на другой стороне двунаправленного промотора. Такой rAd является особенно подходящим для введения через слизистую оболочку, например перорального введения (см., например, WO 2007/100908). В определенных вариантах осуществления композиции по настоящему изобретению содержат алюминий в качестве адъюванта, например в форме гидроксида алюминия, фосфата алюминия, фосфата алюминия-калия или их комбинаций, в концентрациях, составляющих 0,05-5 мг, например 0,075-1,0 мг, алюминия на дозу.
В других вариантах осуществления композиции не содержат адъюванты.
Фармацевтическая композиция в соответствии с настоящим изобретением в определенных вариантах осуществления может представлять собой вакцину.
Композиции на основе аденовируса можно вводить субъекту, например субъекту-человеку. Как известно квалифицированному практику общая доза аденовируса, обеспечиваемая субъекту при однократном введении, может варьировать и обычно составляет от 1×107 вирусных частиц (vp) до 1×1012 vp, предпочтительно от 1×108 vp до 1×1011 vp, например от 3×108 до 5×1010 vp, например от 109 до 3×1010 vp.
Введение композиций на основе аденовируса можно осуществлять с применением стандартных путей введения. Неограничивающие варианты осуществления включают парентеральное введение, такое как инъекция, например внутрикожная, внутримышечная и т. д., или подкожное или чрескожное введение или введение через слизистые оболочки, например интраназальное, пероральное и подобное. В одном варианте осуществления композицию вводят посредством внутримышечной инъекции, например в дельтовидную мышцу руки или латеральную широкую мышцу бедра. Специалисту известны различные варианты введения композиции, например вакцины, для индуцирования иммунного ответа к антигену(-ам), присутствующему(-им) в вакцине.
Субъект, используемый в данном документе, предпочтительно представляет собой млекопитающее, например грызуна, например мышь, или отличного от человека примата, или человека. Предпочтительно, субъект является субъектом-человеком.
Также возможным является обеспечение одного или нескольких бустерных введений одной или нескольких аденовирусных вакцин. При проведении бустерной вакцинации, как правило, такую бустерную вакцину будут вводить одному и тому же субъекту с промежутком от одной недели до одного года, предпочтительно от двух недель до четырех месяцев, после введения композиции субъекту в первый раз (что в данном случае называется "примирующей вакцинацией"). В альтернативных бустерных режимах также возможным является введение различных векторов, например одного или нескольких аденовирусов различных серотипов или других векторов, таких как на основе MVA, или ДНК, или белка, субъекту в качестве примирующей или бустерной вакцинации.
Различные публикации, которые могут включать в себя патенты, опубликованные заявки, технические статьи и научные статьи, процитированы в данном описании в скобках, а полные библиографические данные каждой из них могут быть найдены в конце описания. Каждая из этих процитированных публикаций включена в данный документ посредством ссылки в полном объеме.
Примеры
Без дополнительного описания полагают, что специалист в данной области может с применением предшествующего описания и следующих иллюстративных способов и примеров создать и применять настоящее изобретение и осуществлять на практике заявленные способы. Следующие рабочие примеры, следовательно, конкретно указывают на определенные варианты осуществления, признаки и преимущества настоящего изобретения, и не должны быть истолкованы как ограничивающие каким-либо образом остальную часть настоящего раскрытия. Примеры служат лишь для объяснения настоящего изобретения.
Способы
Клеточная культура
Клетки PER.C6® (Fallaux et al., 1998) поддерживали в среде Игла, модифицированной по способу Дульбекко (DMEM), с 10% фетальной бычьей сыворотки крови (FBS), дополненной 10 мМ MgCl2.
Конструирование аденовирусного вектора в плазмидах pAdApt35 и pshuttle26
Разные конструкции двунаправленного промотора клонировали в плазмиды pAdApt35 (Vogels et al. 2007) или pshuttle26. Pshuttle26 конструировали, основываясь на ранее описанной плазмиде pAdapt26 (Abbink et al., 2007). Фрагмент из 2 т. о., содержащий правую часть генома вектора Ad26, синтезировали и субклонировали в pAdApt26.Luc, в которой сайт SpeI в промоторе CMV изначально устраняли посредством введения замены размером в одну п. о. В результате pshuttle26 можно применять для конструирования аденовирусного вектора посредством гомологичной рекомбинации с космидой Ad26 или посредством гомологичной рекомбинации с плазмидой Ad26 с полноразмерным геномом.
Поскольку плазмиды pAdapt35 и pshuttle26 содержат только стандартную однонаправленную кассету экспрессии с одним промотором и одним сигналом полиА, полученным из SV40, для размещения другого трансгена с сигналом полиА BGH сайты рестрикции добавляли посредством ПЦР с перекрывающимися праймерами. Продукт ПЦР с перекрывающимися праймерами, содержащий SpeI, NotI - BGH полиA - EcoRI- люцифераза - KpnI, SalI, AvrII вставляли в плазмиды в правильной ориентации посредством молекулярного клонирования с помощью сайтов рестрикции SpeI и AvrII. В результате однонаправленный промотор hCMV мог быть заменен на последовательности двунаправленного промотора с применением фланкирующих сайтов рестрикции AvrII и HindIII. Трансгены помещали на обе стороны двунаправленного промотора с применением сайтов рестрикции HindIII и XbaI на одной стороне и AvrII, SalI или KpnI на другой стороне (фиг. 3A). Выбор AvrII, SalI или KpnI зависел от уникальности сайтов рестрикции в плазмидной последовательности. Полные кассеты двунаправленной экспрессии с разными конструкциями двунаправленного промотора клонировали в плазмиды pShuttle26 и переносили в плазмиды pAdapt35 с применением сайтов рестрикции SpeI или NotI и XbaI. Последовательность Kozak (5' GCCACC 3') включали непосредственно перед каждым старт-кодоном ATG и два стоп-кодона (5' TGA TAA 3') добавляли в конце каждой кодирующей последовательности. Как описано в данном документе, рекомбинантные аденовирусы и векторы обычно относятся к rAd или векторам на основе rAd и более конкретно к rAd35 или rAd26 и векторам на их основе.
Получение аденовируса, инфицирование и размножение.
Все аденовирусы выращивали в клетках PER.C6 посредством гомологичной рекомбинации и получали как описано ранее (для rAd35: (Havenga et al., 2006); для rAd26: (Abbink et al., 2007)). Вкратце, клетки PER.C6 трансфицировали с помощью плазмид на основе вектора rAd с применением липофектамина в соответствии с инструкциями, предоставленными производителем (Life Technologies). Для "спасения" векторов rAd35 применяли плазмиды pAdApt35 и космиду pWE/Ad35.pIX-rITR.dE3.5orf6, тогда как для векторов rAd26 применяли плазмиды pShuttle26 и космиду pWE.Ad26.dE3.5orf6. Клетки собирали через один день после полного цитопатического эффекта (CPE), подвергали замораживанию-размораживанию, центрифугировали в течение 5 мин. при 3,000 об./мин. и хранили при  20°C. Далее вирусы очищали с помощью способа бляшкообразования и амплифицировали в клетках PER.C6, культивируемых в одной лунке 24-луночного планшета для культуры тканей. Дополнительную амплификацию проводили в клетках PER.C6, культивируемых с применением колбы T25 для культуры тканей.
20°C. Далее вирусы очищали с помощью способа бляшкообразования и амплифицировали в клетках PER.C6, культивируемых в одной лунке 24-луночного планшета для культуры тканей. Дополнительную амплификацию проводили в клетках PER.C6, культивируемых с применением колбы T25 для культуры тканей.
Анализ экспрессии
Для оценки эффективности экспрессии и сбалансированности экспрессии, вирусные векторы выращивали с помощью репортерных генов, кодирующих усиленный зеленый флуоресцентный белок (номер доступа белка eGFP AAB02572.1) и люциферазу светлячка (номер доступа люциферазы белка ACH53166). Относительное среднее значение интенсивности флуоресценции (MFI) eGFP и относительные световые единицы (RLU) люциферазы записывали для каждого промотора и комбинации репортерных генов с использованием клеток HEK293 (временная трансфекция векторами pAdApt или векторами pshuttle) или клеток A549 (вирусная инфекция). Активность люциферазы измеряли в клеточных лизатах в присутствии 0,1% DTT (1 M), в микропланшетном люминометре Luminoskan™ Ascent. Флуоресценцию eGFP измеряли в проточном цитометре (FACS) путем трипсинизации, центрифугирования и повторного суспендирования клеточных осадков в PBS/1%FBS (невирусный материал) или в CellFix (вирусный материал).
Тестирование генетической стабильности аденовирусных векторов в клетках PER.C6.
Тестирование генетической стабильности векторов для вакцин осуществляли для подтверждения генетической стабильности в способе получения, которое включает несколько пассажей в клетках PER.C6. Выращивания, очистки способом бляшкообразования и приращивания до формата T25 рекомбинантных векторов для вакцин достигали как описано выше. Вкратце, рекомбинантные вирусы выращивали путем плазмидных трансфекций в E1-комплементирующей клеточной линии PER.C6 и очищали способом бляшкообразования. Выбирали 5 бляшек для увеличения масштаба с 24-луночного планшета (MW24) в колбу T25. Следовательно, новые клетки PER.C6 инфицировали в формате T25 до номера 13 вирусного пассажа. Размножение вирусов осуществляли с применением предварительно заданного инфекционного объема, который бы обеспечивал полный цитопатический эффект через 2 дня после инфицирования, и который был ретроспективно определен в диапазоне соотношения вирусной частицы на клетку, составляющего 50 для rAd35 и 900 для rAd26. Вирусную ДНК выделяли из материала p13 и тестировали на наличие полной кассеты для экспрессии трансгена посредством анализа ПЦР. Векторы для вакцин размножали до пассажа под номером 13 в клетки PER.C6. Размножение осуществляли таким образом, чтобы обеспечить полный CPE через два дня после инфицирования. Вирусы rAd35 собирали через 2 дня после полного CPE, тогда как вирусы rAd26 собирали через один день после полного CPE. Вирусную ДНК выделяли при пассаже 2, пассаже 5, пассаже 10 и пассаже 13 и отсутствие делеции тестировали посредством анализа ПЦР с применением праймеров, которые фланкируют кассету для экспрессии трансгена. Отсутствие мутантов с делецией определяли с помощью следующих параметров: Размер полосы продукта ПЦР соответствовал положительному контролю (продукт ПЦР плазмиды, применяемой для "спасения" вируса), никаких дополнительных полос ниже ожидаемого продукта ПЦР (если дополнительные полосы не являются неспецифическими продуктами ПЦР, потому что они также присутствуют в положительном контроле), подтвержденный анализ: без полосы в контроле ПЦР H2O. Чтобы дополнительно подтвердить генетическую стабильность секвенировали продукт ПЦР кассеты экспрессии плюс фланкирующие участки некоторых бляшек.
Пример 1: построение конструкции двунаправленного промотора
Эффективный двунаправленный промотор CMV мыши (mCMV) идентифицировали как применимый промотор для экспрессии двух антигенов из кассеты двунаправленной экспрессии в участке E1 аденовирусных векторов в предыдущей работе, WO 2016/166088. Хотя векторы, содержащие двунаправленный промотор mCMV, экспрессировали антигены, были генетически стабильными и индуцировали иммунный ответ против обоих закодированных антигенов, экспрессия антигенов и индуцированный иммунный ответ не были сбалансированы как объясняется ниже. Экспрессия антигена, помещенного на правую сторону двунаправленного промотора была выше, чем экспрессия антигена, помещенного на левую сторону двунаправленного промотора, что приводило к более сильному иммунному ответу против антигена, помещенного на правую сторону двунаправленного промотора. Различие в уровнях экспрессии для двунаправленного mCMV было примерно 10-кратным. Однако, с целью заменить смесь двух векторов, экспрессирующих только один антиген, для определенных применений является необходимым сбалансированный двунаправленный промотор, который индуцирует сравнимые уровни антигенной экспрессии обоих антигенов. Кроме того, было бы целесообразным, если бы размер двунаправленного промотора был относительно небольшим.
С целью идентификации эффективного и более сбалансированного двунаправленного промотора разрабатывали набор новых двунаправленных промоторов. В силу ограничений по общему размеру аденовирусных векторов, с целью сохранить пространство, достаточное для антигенов, отдавали предпочтение двунаправленным промоторам со структурой меньшего размера с максимальным размером 2 т. н. Дополнительно, предпочтение отдавали структурным блокам без обширных отрезков идентичности последовательностей (<15 нуклеотидов) для предотвращения возникновения делеций, происходящих по механизму гомологичной рекомбинации, в аденовирусном векторе. Двунаправленные промоторы по настоящему изобретению управляют экспрессией гена в двунаправленном режиме, контролируя экспрессию генов, помещенных на обе стороны последовательности двунаправленного промотора. Данные двунаправленные промоторы относятся к непрерывным регуляторным последовательностям гена, которые кроме элементов, представляющих собой промоторы, содержат элементы, представляющие собой энхансеры, и элементы, представляющие собой интроны, и определяются структурными блоками, как описано в данном документе. Структурные блоки, применяемые для построения синтетических двунаправленных промоторов, получали из известных эффективных однонаправленных промоторных, энхансерных и интронных последовательностей. Промоторы направляют экспрессию одного гена, помещенного ниже промоторной последовательности, и, как правило, содержат последовательность TATA-бокса и сайт начала транскрипции (TSS). Энхансерные последовательности могут усиливать экспрессию гена из промотора. Как описывалось, интронные последовательности повышают экспрессию гена in vitro и особенно in vivo.
Разрабатывали и тестировали приведенный ниже набор конструкций двунаправленного промотора
1. rCMV-hEF1α I (фиг. 1A, SEQ ID NO:1)
2. rCMV- hEF1α II (фиг. 1B, SEQ ID NO:2)
3. rhCMV-CAG1 (фиг. 1C, SEQ ID NO:3)
4. hCMV-rhCMV (фиг. 1D, SEQ ID NO:4)
5. rCMV-CAG (фиг. 1E, SEQ ID NO:5)
6. rhCMV-CAG2 (фиг. 1F, SEQ ID NO:6)
7. rCMV bidir 1 (фиг. 1G, SEQ ID NO:10)
8. rCMV bidir 2 (фиг. 1H, SEQ ID NO:8)
9. rCMV bidir 1.1 (фиг. 1I, SEQ ID NO:7)
10. hCMV-CAG4 (фиг. 1J, SEQ ID NO:11)
Другие конструкции двунаправленного промотора также тестировали в предыдущей патентной заявке (PCT/EP2016/057982)
1. mCMV bidir.
2. hCMV-CAG
3. mCMV-CAG
Схематические представления построений двунаправленного промотора и их структурных блоков показаны на фиг. 1 и описаны более подробно ниже.
Промоторные и энхансерные структурные блоки, применяемые в разных построениях, получали из немедленно-ранних (IE) участков цитомегаловируса (как правило, называемых в данном документе промотор hCMV и энхансер hCMV, и иногда также называемых hCMV IE), промоторной последовательности бета-актина курицы/бета-глобина кролика и промоторной последовательности человеческого фактора 1 α элонгации (промотор hEF1α). Интроны получали из химерной последовательности бета-актина курицы/бета-глобина кролика, первого интрона hEF1α и интрона гена аполипопротеина A-1 человека (интрон hApoA1).
Основной немедленно-ранний промотор цитомегаловируса человека известен как эффективный промотор в различных клеточных линиях млекопитающих (Powell et al., 2015). hCMV и другие герпесвирусы экспрессируют гены в три фазы: немедленно-раннюю (IE), раннюю и позднюю фазы. Основные немедленно-ранние промоторы активировали гетерологичные гены на высоких уровнях в различных клеточных линиях млекопитающих.
Хотя основной немедленно-ранний промотор и энхансер цитомегаловируса человека наиболее часто применяются в построении кассет экспрессии трансгена (Addison, Hitt, Kunsken, & Graham, 1997; C. Harro et al., 2009), основные немедленно-ранние промоторы и энхансеры цитомегаловирусов, инфицирующие другие виды, такие как мышь (mCMV) (Addison et al., 1997; Chatellard et al., 2007; C. Harro et al., 2009), крыса (rCMV) (Sandford & Burns, 1996; Voigt, Sandford, Hayward, & Burns, 2005) и макаки-резусы (rhCMV) (Barry, Alcendor, Power, Kerr, & Luciw, 1996; Chan, Chiou, Huang, & Hayward, 1996; Chang et al., 1993; Hansen, Strelow, Franchi, Anders, & Wong, 2003), также являются известными и могут применяться в построении кассет эффективной экспрессии. В частности, последовательность CMV макака-резуса получают из основного немедленно-раннего участка вируса герпеса Мартышковых обезьян 5 типа. Короткую часть промотора CMV макака-резуса идентифицировали с помощью выравнивания с короткими промоторами hCMV и CMV шимпанзе. Промотор hEF1α также описывается как эффективная промоторная последовательность экспрессии гетерологичного гена в клетках млекопитающих (Kim, Uetsuki, Kaziro, Yamaguchi, & Sugano, 1990).
Химерный промотор CAG, состоящий из энхансера hCMV, промотора бета-актина курицы(A) и последовательности гибридного интрона бета-актина курицы/бета-глобина кролика (G), описываемый как эффективный промотор экспрессии гетерологичных генов может быть укорочен и может применяться для экспрессии антигена (Richardson et al., 2009). В исследовании Richardson et al. описывает модификации промотора CAG, которая в итоге приводит к появлению версии промотора Δ829CAG, в которой гибридный интрон значительно укорочен. Данную версию промотора Δ829CAG без энхансера hCMV (Δ829AG) применяли в качестве структурного блока для построения определенных двунаправленных промоторов, обозначаемого в графических материалах как часть AG, но называемого CAG в названиях двунаправленных промоторов и называемого CAG или AG по всему тексту.
Далее описано расположение структурных блоков для построения последовательностей двунаправленного промотора.
Укороченный промотор AG и промотор hEF1α содержат интроны, как важные части описываемых эффективных регуляторных последовательностей. Когда при построении двунаправленного промотора применяли промотор AG или промотор hEF1α, на противоположную сторону построения двунаправленного промотора также помещали гетерологичную интронную последовательность. Основываясь на естественной двунаправлености промотора CMV мыши, были созданы разные построения потенциального двунаправленного промотора немедленно-раннего промотора rCMV.
Ранее описывалось, что синтетическая комбинация интрона, полученного из hEF1α, и промоторной последовательности mCMV дает в результате эффективную регуляторную последовательность. Следовательно, последовательности hEF1α объединяли с элементами промотора rCMV (другого промотора, полученного из муромегаловируса, как mCMV) и энхансера для построения последовательностей двунаправленного промотора rCMV-hEF1α I и rCMV-hEF1α II (см., например, фиг. 1A и 1B). Поскольку согласно описанию интрон hEF1α увеличивает уровни экспрессии, но представляет собой очень длинную последовательность, при построениях пытались значительно укоротить последовательность интрона hEF1α, который описан, с помощью экспериментального подхода для промотора CAG (Richardson et al., 2009). С этой целью часть интронной последовательности удаляли, сохраняя при этом описанный сайт донора сплайсинга, сайт акцептора сплайсинга и предполагаемый сайт точки ветвления плюс описанные сайты связывания клеточных факторов. Строили две разные версии укороченного интрона, в которых комбинация промотор/интрон hEF1α I сохраняет больше сайтов чем комбинация промотор/интрон hEF1α II, давая на выходе двунаправленные промоторы rCMV- hEF1α I и rCMV- hEF1α II.
Дополнительные построения двунаправленного промотора основаны на укороченном промоторе AG в комбинации с энхансерными и промоторными последовательностями основных немедленно-ранних промоторов цитомегаловируса, полученных из разных видов, в том числе CMV макака-резуса (rhCMV), CMV крысы (rCMV) и CMV человека (hCMV). Данные двунаправленные промоторы называются rhCMV-CAG1, rhCMV-CAG2, hCMV-CAG4 и rCMV-CAG. Различие между rhCMV-CAG1 и rhCMV-CAG2 заключается в ориентации энхансерной последовательности rhCMV.
Дополнительное построение двунаправленного промотора основывалось на комбинации энхансера hCMV, промотора CMV человека (hCMV) и промотора CMV макака-резуса (rhCMV). Полученная конструкция двунаправленного промотора по всему тексту называется hCMV-rhCMV.
Поскольку ранее было описано (Amendola et al, 2005), что расположение одного энхансера с двумя фланкирующими его промоторами приводит в результате к координированной экспрессии двух генов, представляющих интерес, применение сильных промоторных и энхансерных структурных блоков теоретически должно приводить к получению двунаправленных промоторов сопоставимой эффективности и сбалансированности.
В качестве основы для последовательностей двунаправленного промотора rCMV bidir 1, и rCMV bidir 2, и rCMV bidir 1.1 кроме синтетических построений двунаправленного промотора применяли потенциально естественный двунаправленный промотор, полученный из участка mIE rCMV. Хотя все три построения двунаправленного промотора содержат предполагаемый минимальный промотор rCMV и предполагаемый минимальный промотор vOX2 rCMV, фланкирующий энхансерную последовательность rCMV, построения различаются длиной энхансерного фрагмента и ориентацией энхансерного фрагмента. Промотор vOX2 управляет транскрипцией клеточного гена CD200 (vOX2) крысы непосредственно справа от участка MIE (Voigt et al., 2005).
Пример 2: скрининг разных конструкций промотора на наличие эффективной и сбалансированной экспрессии репортерных генов
В первом эксперименте с осуществлением скрининга экспрессию из разных конструкций двунаправленного промотора оценивали с помощью временных трансфекций в HEK293 с применением репортерных генов люциферазы и eGFP для количественной оценки эффективности. Для временных трансфекций плазмид pAdapt35 двунаправленные промоторы имели трансген люциферазы на левой стороне и трансген eGFP на правой стороне двунаправленного промотора.
Три независимых эксперимента с осуществлением трансфекции проводили с плазмидами, содержащими разные построения двунаправленного промотора. В эксперименте 1 (фиг. 2A) тестировали промоторы rCMV bidir.2, rCMV- hEF1α II, rhCMV- hEF1α I, rhCMV-CAG1 и hCMV-rhCMV. В то время как rCMV bidir.2, rCMV-EF1 α II и rhCMV-CAG1 неизменно показывают функцию двунаправленного промотора, хоть и мене эффективного, чем промотор hCMV с однонаправленным контролем (SEQ ID NO:9), rCMV- hEF1α I, неожиданно, показывает эффективность промотора не выше отрицательного контроля для экспрессии eGFP и очень слабую эффективность промотора для экспрессии люциферазы. В эксперименте 1 hCMV-rhCMV является наиболее эффективным двунаправленным промотором. Неожиданно, hCMV-rhCMV оказался эффективным двунаправленным промотором с очень сбалансированной экспрессией трансгена, хотя и с несколько более низкими уровнями экспрессии, чем у однонаправленного промотора hCMV. В эксперименте 2 с осуществлением скрининга (фиг. 2B) тестировали промоторы hCMV-CAG4, rCMV bidir.1.1, rCMV-CAG. В этом эксперименте промотор hCMV-CAG4 на обеих сторонах показывал эффективную активность промотора, которая была сопоставимой или даже выше таковой контроля, представляющего собой однонаправленный промотор hCMV. Промотор rCMV bidir.1.1 неожиданно показал низкую эффективность промотора на обеих сторонах. Двунаправленный промотор rCMV-CAG показывает эффективность двунаправленного промотора, которая с обеих сторон является более низкой, по сравнению с эффективностью контроля, представляющего собой однонаправленный hCMV, и также с обеих сторон является более низкой и не такой сбалансированной, по сравнению с двунаправленным промотором hCMV-rhCMV (данные по которому представлены на фиг. 2A). В третьем эксперименте с осуществлением скрининга (фиг. 2С) тестировали два новых двунаправленных промотора, rhCMV-CAG2, rCMV bidir.1, и два уже протестированных двунаправленных промотора, hCMV-rhCMV и hCMV-CAG4. В то время как rhCMV-CAG2 продемонстрировал эффективность двунаправленного промотора, хоть и более слабую по сравнению с контролем, представляющим собой однонаправленный hCMV, rCMV bidir 1 индуцировал экспрессию только eGFP, помещенного на правой стороне промотора, а активность люциферазы находилась в диапазоне контроля, представляющего собой нетрансфицированные клетки. В этом эксперименте 3 снова тестировали двунаправленные промоторы hCMV-rhCMV и hCMV-CAG4, подтверждая их перспективные эффективность и сбалансированность. Как и ожидалось в таком эксперименте по клеточной биологической трансфекции, между экспериментом 1 и экспериментом 3 наблюдается некоторая изменчивость. Оба из hCMV-rhCMV и hCMV-CAG4 демонстрируют более низкую эффективность в эксперименте 3, чем в эксперименте 1, по сравнению с контролем, представляющим собой однонаправленный промотор hCMV. Хотя hCMV-rhCMV является менее эффективным, чем hCMV-CAG4, он является более сбалансированным в отношении экспрессии eGFP и люциферазы. Таким образом, hCMV-rhCMV представляет собой эффективный двунаправленный промотор с очень сбалансированной экспрессией трансгенов, а также имеет преимущество в том, что он является относительно коротким.
Неожиданно, не все комбинации структурных блоков обеспечили получение эффективных и сбалансированных двунаправленных промоторов. Например, hCMV-CAG4 и rhCMV-CAG1, и rhCMV-CAG2 являются аналогичными в отношении их структурных блоков. Во всех трех разных построениях применяются описанные сильные энхансер и промотор, полученные из CMV, хоть и из разных видов, и один и тот же ранее описанный сильный промотор CAG. Однако, неожиданно hCMV-CAG4 оказался более эффективным, чем rhCMV-CAG1 и rhCMV-CAG2. Кроме того, было описано, что часть промотора CAG является более сильной, чем промотор hCMV, однако в двунаправленном режиме функционирования экспрессия трансгена, связанного с промоторным структурным блоком hCMV, превышала экспрессию трансгена, связанного с промоторным структурным блоком CAG. Это наглядно демонстрирует непредсказуемость создания новых двунаправленных промоторов из ранее известных структурных блоков при их применении в составе других комплексов.
Из набора тестируемых конструкций двунаправленный промотор hCMV-rhCMV был идентифицирован как наиболее сбалансированный кандидат конструкций эффективного двунаправленного промотора. Исходя из построения промотора hCMV-rhCMV, из структурных блоков описанных эффективных промотора и энхансеров нельзя было предсказать, что этот промотор будет как эффективным (хотя и немного менее эффективным, чем другой новый двунаправленный промотор hCMV-CAG4), так и очень сбалансированным (намного более сбалансированным, чем двунаправленный промотор mCMV, который был описан в WO 2016/166088, и даже несколько более сбалансированным, чем двунаправленный промотор hCMV-CAG4). Дополнительно, двунаправленный промотор hCMV-rhCMV имеет то преимущество, что он значительно короче (с длиной менее 1 т. п. о.), чем другие построения промотора. Это значит, что при применении этого двунаправленного промотора остается больше места для трансгенов (т. е. это делает возможным применение более длинных трансгенов) в векторах, которые имеют ограничение по месту, таких как rAds, по сравнению с другими двунаправленными промоторами. Остальные протестированные синтетические построения двунаправленного промотора, состоящие из разных структурных блоков однонаправленного промотора, в основном показывали хорошие функции двунаправленного промотора, однако они, как правило, были менее эффективными и менее сбалансированными, чем промотор hCMV-rhCMV.
Сравнительно с двунаправленным промотором CMV мыши, немедленно-ранний промотор rCMV может быть построен как двунаправленный промотор, который, однако, является менее эффективным. Из результатов видно, что невозможно предсказать, какая комбинация структурных блоков будет обеспечивать хорошие функциональные возможности двунаправленного промотора (эффективную экспрессию в обоих направлениях, т. е. по меньшей мере 10%, предпочтительно по меньшей мере 20%, более предпочтительно по меньшей мере 30% экспрессии под контролем однонаправленного промотора hCMV).
Схематическое представление hCMV-rhCMV, включающее аннотации идентичности и ориентации структурных блоков, показано на фиг. 3. Правая сторона двунаправленного промотора hCMV-rhCMV включает промоторный структурный блок CMV макака-резуса (rhCMV), а левая сторона двунаправленного промотора hCMV-rhCMV включает промоторный структурный блок hCMV и энхансерный структурный блок hCMV в инвертированной ориентации, так, что он направлен к левой стороне двунаправленного промотора, в том же направлении, что промотор hCMV. Хотя в данном документе термины "левая" и "правая" применяются для простоты описания, специалисту сразу будет понятно, что конструкцию двунаправленного промотора hCMV-rhCMV также можно поворачивать и применять в противоположной ориентации. Следует также отметить, что промотор hCMV-rhCMV, вследствие его относительно небольшого размера, составляющего менее чем 1 т. п. о., хорошо изучен в отношении его применения в качестве двунаправленного промотора в рекомбинантном аденовирусном векторе.
Пример 3: эффективность и сбалансированность экспрессии трансгена из аденовирусных векторов, содержащих кассету экспрессии hCMV-rhCMV
Для дополнительной оценки эффективности и сбалансированности экспрессии из участка E1 аденовирусных векторов, получали векторы Ad26 и Ad35, содержащие кассету двунаправленной экспрессии hCMV-rhCMV в участке E1. Для оценки эффективности и сбалансированности экспрессии репортерного гена после трансдукции некомплементирующих клеток A549 получали четыре разных вектора, а именно Ad26.eGFP.hCMV-rhCMV.Luc, Ad26.Luc.hCMV-rhCMV.eGFP, Ad35.eGFP.hCMV-rhCMV.Luc и Ad35.Luc.hCMV-rhCMV.eGFP. Трансдукции проводили при 100 VP/клетка и 1000 VP/клетка. Поскольку результаты обоих соотношений VP/клетка были аналогичными, только результаты трансдукций 1000 VP/клетка показаны на фиг. 4. С целью оценить десятикратное различие в экспрессии 100 VP/клетка и 1000 VP/клетка, показаны трансдукции векторов Ad.Luc и Ad.eGFP положительного контроля, экспрессирующих репортерные гены под контролем однонаправленного промотора hCMV. На секции 4A видно, что hCMV-rhCMV индуцирует эффективную экспрессию обоих репортерных генов из кассеты двунаправленной экспрессии Ad26 E1 с уровнями экспрессии немного более низкими, чем таковой из контрольных векторов Ad26.Luc и Ad26.eGFP, составляющий 1000 VP/клетка. Дополнительно Ad26.Luc.hCMV-rhCMV.eGFP непосредственно сравнивали с Ad26.Luc.mCMV bidir.eGFP, и из результатов видно сниженную экспрессию трансгена eGFP по сравнению с mCMV bidir., однако также видно в целом более сбалансированную экспрессию трансгена. На секции 4B видна экспрессия трансгена из кассеты двунаправленной экспрессии hCMV-rhCMV в векторах Ad35. Интересно, что профиль экспрессии в векторах Ad35 немного отличался от профиля экспрессии в векторах Ad26. Следовательно, хотя эффективные двунаправленные промоторы можно применять в rAdV, полученных из разных серотипов, отличный промотор может быть оптимальным для применения в одном rAdV относительно другого, дополнительно показывая в качестве примера, что усложненное построение промоторов и кассет экспрессии является необходимым для оптимальных вирусных векторов.
Пример 4: тестирование генетической стабильности аденовирусных векторов, содержащих кассету двунаправленной экспрессии hCMV-rhCMV в участке E1
Кроме экспрессии трансгена, генетическая стабильность во время получения AdV является важным параметром для применимого AdV, экспрессирующего два антигена. Следовательно, генетическую стабильность тестировали, как описано в предыдущей заявке, WO 2016/166088. Вкратце, векторы Ad26.Luc.hCMV-rhCMV.eGFP и Ad26.eGFP.hCMV-rhCMV.Luc получали посредством плазмидной трансфекции в клетках PER.C6 и вирусные популяции выделяли посредством сбора бляшек. Десять бляшек на вектор размножали до номера вирусного пассажа (vpn) 3. Далее выбирали пять бляшек для длительного пассирования до vpn 13. Генетическую стабильность оценивали посредством идентификации ПЦР на участке E1 кассеты экспрессии (фиг. 5) и участке E3 и E4 (данные ПЦР E3 и E4 не показаны). Отсутствие малого количества делеций или точечных мутаций подтверждали посредством стандартного секвенирования по Сэнгеру E1 продукта ПЦР vpn 13. Пять из пяти бляшек обоих Ad26.Luc.hCMV-rhCMV.eGFP и Ad26.eGFP.hCMV-rhCMV.Luc оставались генетически стабильными до vpn 13.
Вывод
Как описано выше, посредством скрининга набора новых конструкций двунаправленного промотора было установлено, что невозможно предсказать, какие конструкции двунаправленного промотора будут обеспечивать необходимые свойства промотора. В действительности, даже конструкции двунаправленного промотора, которые являются аналогичными, необязательно дают такие же результаты. Например, двунаправленный промотор hCMV-rhCMV с промотором CMV человека (hCMV) на левой стороне и коротким промотором CMV макака-резуса (rhCMV) на правой стороне, продемонстрировал особенно сбалансированную экспрессию двух разных трансгенов из участка E1 векторов rAd26 и rAd35. Неожиданно оказалось, что двунаправленный промотор hCMV-rhCMV сочетает эффективность и сбалансированность в отношении экспрессии трансгена, а также является достаточно небольшим, с размером менее 1 т. п. о. Установлено, что rAd с двунаправленным промотором hCMV-rhCMV являются генетически стабильными даже после серийного пассирования на клетках PER.C6 до P13. Таким образом, непредсказуемо, двунаправленный промотор hCMV-rhCMV по настоящему изобретению представляет собой промотор с неожиданно предпочтительными характеристиками для применения в рекомбинантных вирусных векторах, которые можно применять в генной терапии или вакцинах, где является необходимой сбалансированная и эффективная экспрессия и/или является целесообразным небольшой размер двунаправленного промотора hCMV-rhCMV.
Таблица 1
* описано в WO 2016/166088
Ссылки
Патентные документы США
US5057540A (10/15/1991). "Saponin adjuvant". Kensil, Charlotte A.; Marciani, Dante J.
US5122458A (6/16/1992). "Use of a bGH gDNA polyadenylation signal in expression of non-bGH polypeptides in higher eukaryotic cells". Post, Leonard E.; Palermo, Daniel P.; Thomsen, Darrell R.; Rottman, Fritz M.; Goodwin, Edward C.; Woychik, Richard P.
US5559099A (9/24/1996). "Penton base protein and methods of using same". Wickham, Thomas J.; Kovesdi, Imre; Brough, Douglas E.; McVey, Duncan L.; Brader, Joseph T.
US5837511A (11/17/1998). "Non-group C adenoviral vectors". Falck Pedersen, Erik S.; Crystal, Ronald G.; Mastrangeli, Andrea; Abrahamson, Karil
US5837520A (11/17/1998). "Method of purification of viral vectors". Shabram, Paul W.; Huyghe, Bernard G.; Liu, Xiaodong; Shepard, H. Michael
US5846782A (12/8/1998). "Targeting adenovirus with use of constrained peptide motifs". Wickham, Thomas J.; Roelvink, Petrus W.; Kovesdi, Imre
US5851806A (12/22/1998). "Complementary adenoviral systems and cell lines". Kovesdi, Imre; Brough, Douglas E.; McVey, Duncan L.; Bruder, Joseph T.; Lizonova, Alena
US5891690A (4/6/1999). "Adenovirus E1-complementing cell lines". Massie, Bernard
US5965541A (10/12/1999). "Vectors and methods for gene transfer to cells". Wickham, Thomas J.; Kovesdi, Imre; Brough, Douglas E.
US5981225A (11/9/1999). "Gene transfer vector, recombinant adenovirus particles containing the same, method for producing the same and method of use of the same". Kochanek, Stefan; Schiedner, Gudrun
US5994106A (11/30/1999). "Stocks of recombinant, replication-deficient adenovirus free of replication-competent adenovirus". Kovesdi, Imre; Brough, Douglas E.; McVey, Duncan L.; Bruder, Joseph T.; Lizonova, Alena
US5994128A (11/30/1999). "Packaging systems for human recombinant adenovirus to be used in gene therapy". Fallaux, Frits Jacobus; Hoeben, Robert Cornelis; Van der Eb, Alex Jan; Bout, Abraham; Valerio, Domenico
US6020191A (2/1/2000). "Adenoviral vectors capable of facilitating increased persistence of transgene expression". Scaria, Abraham; Gregory, Richard J.; Wadsworth, Samuel C.
US6040174A (3/21/2000). "Defective adenoviruses and corresponding complementation lines". Imler, Jean Luc; Mehtali, Majid; Pavirani, Andrea
US6083716A (7/4/2000). "Chimpanzee adenovirus vectors". Wilson, James M.; Farina, Steven F.; Fisher, Krishna J.
US6113913A (9/5/2000). "Recombinant adenovirus". Brough, Douglas E.; Kovesdi, Imre
US6225289B1 (5/1/2001). "Methods and compositions for preserving adenoviral vectors". Kovesdi, Imre; Ransom, Stephen C.
US6261823B1 (7/17/2001). "Methods for purifying viruses". Tang, John Chu Tay; Vellekamp, Gary; Bondoc, Jr., Laureano L.
US6485958B2 (11/26/2002). "Method for producing recombinant adenovirus". Blanche, Francis; Guillaume, Jean Marc
US7326555B2 (2/5/2008). "Methods of adenovirus purification". Konz, Jr., John O.; Lee, Ann L.; To, Chi Shung Brian; Goerke, Aaron R
US8932607B2 (1/13/2015). "Batches of recombinant adenovirus with altered terminal ends". Custers, Jerome H. H. V.; Vellinga, Jort
Европейские патентные документы
EP1230354B1 (1/7/2004). "PERMANENT AMNIOCYTE CELL LINE, THE PRODUCTION THEREOF AND ITS USE FOR PRODUCING GENE TRANSFER VECTORS". KOCHANEK, Stefan; SCHIEDNER, Gudrun
EP1601776B1 (7/2/2008). "EXPRESSION VECTORS COMPRISING THE MCMV IE2 PROMOTER". CHATELLARD, Philippe; IMHOF, Markus
EP853660B1 (1/22/2003). "METHOD FOR PRESERVING INFECTIOUS RECOMBINANT VIRUSES, AQUEOUS VIRAL SUSPENSION AND USE AS MEDICINE". SENE, Claude
Публикации международных патентных заявок
WO2003049763A1 (6/19/2003). "COMPOSITION FOR THE PRESERVATION OF VIRUSES". SETIAWAN, Kerrie; CAMERON, Fiona, Helen
WO2003061708A1 (7/31/2003). "STABILIZED FORMULATIONS OF ADENOVIRUS". PUNGOR, Erno
WO2003078592A2 (9/25/2003). "METHOD FOR THE PURIFICATION, PRODUCTION AND FORMULATION OF ONCOLYTIC ADENOVIRUSES". MEMARZADEH, Bahram; PENNATHUR-DAS, Rukmini; WYPYCH, Joseph; YU, De Chao
WO2003104467A1 (12/18/2003). "MEANS AND METHODS FOR THE PRODUCTION OF ADENOVIRUS VECTORS". VOGELS, Ronald; BOUT, Abraham
WO2004001032A2 (12/31/2003). "STABLE ADENOVIRAL VECTORS AND METHODS FOR PROPAGATION THEREOF". VOGELS, Ronald; HAVENGA, Menzo, Jans, Emco; ZUIJDGEEST, David, Adrianus, Theodorus
WO2004004762A1 (1/15/2004). "ISCOM PREPARATION AND USE THEREOF". MOREIN, Bror;  BENGTSSON, Karin
BENGTSSON, Karin
WO2004020971A2 (3/11/2004). "CHROMATOGRAPHIC METHODS FOR ADENOVIRUS PURIFICATION". SENESAC, Joseph
WO2004037294A2 (5/6/2004). "NEW SETTINGS FOR RECOMBINANT ADENOVIRAL-BASED VACCINES". HAVENGA, Menzo, Jans, Emco; HOLTERMAN, Lennart; KOSTENSE, Stefan; PAU, Maria, Grazia; SPRANGERS, Mieke, Caroline; VOGELS, Ronald
WO2004055187A1 (7/1/2004). "RECOMBINANT VIRAL-BASED MALARIA VACCINES". PAU, Maria Grazia; HOLTERMAN, Lennart; KASPERS, Jorn; STEGMANN, Antonius, Johannes, Hendrikus
WO2005002620A1 (1/13/2005). "QUIL A FRACTION WITH LOW TOXICITY AND USE THEREOF". MOREIN, Bror; BENGTSSON, Karin;  , Jill; RANLUND, Katarina
, Jill; RANLUND, Katarina
WO2005071093A2 (8/4/2005). "CHIMPANZEE ADENOVIRUS VACCINE CARRIERS". CIRILLO, Agostino; COLLOCA, Stefano; ERCOLE, Bruno, Bruni; MEOLA, Annalisa; NICOSIA, Alfredo; SPORENO, Elisabetta
WO2005080556A2 (9/1/2005). "VIRUS PURIFICATION METHODS". WEGGEMAN, Miranda; VAN CORVEN, Emile Joannes Josephus Maria
WO2006053871A2 (5/26/2006). "MULTIVALENT VACCINES COMPRISING RECOMBINANT VIRAL VECTORS". HAVENGA, Menzo, Jans, Emco; VOGELS, Ronald; SADOFF, Jerald; HONE, David; SKEIKY, Yasir Abdul Wahid; RADOSEVIC, Katarina
WO2006108707A1 (10/19/2006). "VIRUS PURIFICATION USING ULTRAFILTRATION". WEGGEMAN, Miranda
WO2006120034A1 (11/16/2006). "VACCINE COMPOSITION". ERTL, Peter, Franz; TITE, John, Philip; VAN WELY, Catherine Ann
WO2007073513A2 (6/28/2007). "METHOD FOR PROPAGATING ADENOVIRAL VECTORS ENCODING INHIBITORY GENE PRODUCTS". GALL, Jason, G., D.; BROUGH, Douglas, E.; RICHTER, King, C.
WO2007100908A2 (9/7/2007). "CHIMERIC ADENOVIRAL VECTORS". TUCKER, Sean, N.
WO2007104792A2 (9/20/2007). "RECOMBINANT ADENOVIRUSES BASED ON SEROTYPE 26 AND 48, AND USE THEREOF". BAROUCH, Dan H.; HAVENGA, Menzo Jans Emko
WO2007110409A1 (10/4/2007). "COMPOSITIONS COMPRISING A RECOMBINANT ADENOVIRUS AND AN ADJUVANT". HAVENGA, Menzo Jans Emko; RADOSEVIC, Katarina
WO2009026183A1 (2/26/2009). "USE OF CHIMERIC HIV/SIV GAG PROTEINS TO OPTIMIZE VACCINE-INDUCED T CELL RESPONSES AGAINST HIV GAG". NABEL, Gary, J.; YANG, Zhi-Yong; SHI, Wei; BAROUCH, Dan, H.
WO2009117134A2 (9/24/2009). "AEROSOLIZED GENETIC VACCINES AND METHODS OF USE". ROEDERER, Mario; RAO, Srinivas; NABEL, Gary, J.; ANDREWS, Charla, Anne
WO2010085984A1 (8/5/2010). "SIMIAN ADENOVIRUS NUCLEIC ACID- AND AMINO ACID-SEQUENCES, VECTORS CONTAINING SAME, AND USES THEREOF". COLLOCA, Stefano; NICOSIA, Alfredo; CORTESE, Riccardo; AMMENDOLA, Virginia; AMBROSIO, Maria
WO2010086189A2 (8/5/2010). "SIMIAN ADENOVIRUS NUCLEIC ACID- AND AMINO ACID-SEQUENCES, VECTORS CONTAINING SAME, AND USES THEREOF". COLLOCA, Stefano; NICOSIA, Alfredo; CORTESE, Riccardo; AMMENDOLA, Virginia; AMBROSIO, Maria
WO2010096561A1 (8/26/2010). "SYNTHETIC HIV/SIV GAG PROTEINS AND USES THEREOF". NABEL, Gary J.; YANG, Zhi-yong; SHI, Wei; BAROUCH, Dan H.
WO2011045378A1 (4/21/2011). "METHOD FOR THE PURIFICATION OF ADENOVIRUS PARTICLES". DE VOCHT, Marcel, Leo; VEENSTRA, Marloes
WO2011045381A1 (4/21/2011). "PROCESS FOR ADENOVIRUS PURIFICATION FROM HIGH CELL DENSITY CULTURES". DE VOCHT, Marcel, Leo; VEENSTRA, Marloes
WO2013139911A1 (9/26/2013). "VACCINE AGAINST RSV". RADOSEVIC, Katarina; CUSTERS,  H.H.V.; VELLINGA, Jort; WIDJOJOATMODJO, Myra N.
H.H.V.; VELLINGA, Jort; WIDJOJOATMODJO, Myra N.
WO2013139916A1 (9/26/2013). "VACCINE AGAINST RSV". RADOSEVIC, Katarina; CUSTERS, H.H.V.; VELLINGA, Jort; WIDJOJOATMODJO, Myra, N.
Другие ссылочные документы
Книги
Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, NY (1995)
Ausubel F.M., et al. (editors). Current Protocols in Molecular Biology; the series Methods in Enzymology, Academic Press, Inc. (1987)
Freshney, R.I., Culture of animal cells: A manual of basic technique, fourth edition, Wiley-Liss Inc., ISBN 0-471-34889-9 (2000)
Frokjaer S. and Hovgaard L. (editors), Pharmaceutical Formulation Development of Peptides and Proteins, Taylor & Francis (2000)
Gennaro, A.R. (editor), Remington's Pharmaceutical Sciences, 18th edition,., Mack Publishing Company (1990)
Horowitz, M.S., Adenoviruses, Chapter 68, in Virology, (B. N. Fields et al. (editors), 3rd Ed., Raven Press, Ltd., New York (1996)
Kibbe A. (editor), Handbook of Pharmaceutical Excipients, 3rd edition, Pharmaceutical Press (2000)
Kruse and Paterson (editors), Tissue Culture, Academic Press. (1973)
MacPherson M.J., Hams B.D., Taylor G.R. (editors), PCR2: A Practical Approach (1995)
Sambrook et al., Molecular Cloning, a Laboratory Manual, 2nd Ed., Cold Spring Harbor Press, Cold Spring Harbor, N.Y. (1989)
Sambrook, Fritsch and Maniatis, Molecular Cloning: A Laboratory Manual, 2nd Ed., (1989)
Shenk, Thomas, Adenoviridae and their Replication, Chapter 67, in Virology, B. N. Fields et al. (editors)., 3rd Ed., Raven Press, Ltd., New York (1996)
Watson et al., Recombinant DNA, 2nd ed., Scientific American Books. (1992)
Журналы
Abbink, P., Lemckert, A. A., Ewald, B. A., Lynch, D. M., Denholtz, M., Smits, S.,… Barouch, D. H. (2007). Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J Virol, 81(9), 4654-4663. doi: 10.1128/JVI.02696-06
Abbink, P., Maxfield, L. F., Ng'ang'a, D., Borducchi, E. N., Iampietro, M. J., Bricault, C. A.,… Barouch, D. H. (2015). Construction and evaluation of novel rhesus monkey adenovirus vaccine vectors. J Virol, 89(3), 1512-1522. doi: 10.1128/JVI.02950-14
Abrahamsen, K., Kong, H. L., Mastrangeli, A., Brough, D., Lizonova, A., Crystal, R. G., & Falck-Pedersen, E. (1997). Construction of an adenovirus type 7a E1A- vector. J Virol, 71(11), 8946-8951.
Addison, C. L., Hitt, M., Kunsken, D., & Graham, F. L. (1997). Comparison of the human versus murine cytomegalovirus immediate early gene promoters for transgene expression by adenoviral vectors. J Gen Virol, 78 ( Pt 7), 1653-1661. doi: 10.1099/0022-1317-78-7-1653
Amendola, M., Venneri, M. A., Biffi, A., Vigna, E., & Naldini, L. (2005). Coordinate dual-gene transgenesis by lentiviral vectors carrying synthetic bidirectional promoters. Nat Biotechnol, 23(1), 108-116.
Andrianaki, A., Siapati, E. K., Hirata, R. K., Russell, D. W., & Vassilopoulos, G. (2010). Dual transgene expression by foamy virus vectors carrying an endogenous bidirectional promoter. Gene Ther, 17(3), 380-388. doi: 10.1038/gt.2009.147
Bangari, D. S., & Mittal, S. K. (2006). Development of nonhuman adenoviruses as vaccine vectors. Vaccine, 24(7), 849-862. doi: 10.1016/j.vaccine.2005.08.101
Barry, P. A., Alcendor, D. J., Power, M. D., Kerr, H., & Luciw, P. A. (1996). Nucleotide sequence and molecular analysis of the rhesus cytomegalovirus immediate-early gene and the UL121-117 open reading frames. Virology, 215(1), 61-72. doi: 10.1006/viro.1996.0007
Barski, O. A., Siller-Lopez, F., Bohren, K. M., Gabbay, K. H., & Aguilar-Cordova, E. (2004). Human aldehyde reductase promoter allows simultaneous expression of two genes in opposite directions. Biotechniques, 36(3), 382-384, 386, 388.
Belousova, N., Harris, R., Zinn, K., Rhodes-Selser, M. A., Kotov, A., Kotova, O.,… Alvarez, R. D. (2006). Circumventing recombination events encountered with production of a clinical-grade adenoviral vector with a double-expression cassette. Mol Pharmacol, 70(5), 1488-1493.
Brough, D. E., Lizonova, A., Hsu, C., Kulesa, V. A., & Kovesdi, I. (1996). A gene transfer vector-cell line system for complete functional complementation of adenovirus early regions E1 and E4. J Virol, 70(9), 6497-6501.
Chan, Y. J., Chiou, C. J., Huang, Q., & Hayward, G. S. (1996). Synergistic interactions between overlapping binding sites for the serum response factor and ELK-1 proteins mediate both basal enhancement and phorbol ester responsiveness of primate cytomegalovirus major immediate-early promoters in monocyte and T-lymphocyte cell types. J Virol, 70(12), 8590-8605.
Chang, Y. N., Jeang, K. T., Chiou, C. J., Chan, Y. J., Pizzorno, M., & Hayward, G. S. (1993). Identification of a large bent DNA domain and binding sites for serum response factor adjacent to the NFI repeat cluster and enhancer region in the major IE94 promoter from simian cytomegalovirus. J Virol, 67(1), 516-529.
Chatellard, P., Pankiewicz, R., Meier, E., Durrer, L., Sauvage, C., & Imhof, M. O. (2007). The IE2 promoter/enhancer region from mouse CMV provides high levels of therapeutic protein expression in mammalian cells. Biotechnol Bioeng, 96(1), 106-117. doi: 10.1002/bit.21172
Cohen, C. J., Xiang, Z. Q., Gao, G. P., Ertl, H. C., Wilson, J. M., & Bergelson, J. M. (2002). Chimpanzee adenovirus CV-68 adapted as a gene delivery vector interacts with the coxsackievirus and adenovirus receptor. J Gen Virol, 83(Pt 1), 151-155.
Collins, P. J., Kobayashi, Y., Nguyen, L., Trinklein, N. D., & Myers, R. M. (2007). The ets-related transcription factor GABP directs bidirectional transcription. PLoS Genet, 3(11), e208. doi: 10.1371/journal.pgen.0030208
Fallaux, F. J., Bout, A., van der Velde, I., van den Wollenberg, D. J., Hehir, K. M., Keegan, J.,… Hoeben, R. C. (1998). New helper cells and matched early region 1-deleted adenovirus vectors prevent generation of replication-competent adenoviruses. Hum Gene Ther, 9(13), 1909-1917.
Farina, S. F., Gao, G. P., Xiang, Z. Q., Rux, J. J., Burnett, R. M., Alvira, M. R.,… Wilson, J. M. (2001). Replication-defective vector based on a chimpanzee adenovirus. J Virol, 75(23), 11603-11613. doi: 10.1128/JVI.75.23.11603-11613.2001
Gao, G. P., Engdahl, R. K., & Wilson, J. M. (2000). A cell line for high-yield production of E1-deleted adenovirus vectors without the emergence of replication-competent virus. Hum Gene Ther, 11(1), 213-219. doi: 10.1089/10430340050016283
Geisbert, T. W., Bailey, M., Hensley, L., Asiedu, C., Geisbert, J., Stanley, D.,… Sullivan, N. J. (2011). Recombinant adenovirus serotype 26 (Ad26) and Ad35 vaccine vectors bypass immunity to Ad5 and protect nonhuman primates against ebolavirus challenge. J Virol, 85(9), 4222-4233. doi: 10.1128/JVI.02407-10
Goerke, A. R., To, B. C., Lee, A. L., Sagar, S. L., & Konz, J. O. (2005). Development of a novel adenovirus purification process utilizing selective precipitation of cellular DNA. Biotechnol Bioeng, 91(1), 12-21. doi: 10.1002/bit.20406
Hansen, S. G., Strelow, L. I., Franchi, D. C., Anders, D. G., & Wong, S. W. (2003). Complete sequence and genomic analysis of rhesus cytomegalovirus. J Virol, 77(12), 6620-6636.
Harro, C. D., Robertson, M. N., Lally, M. A., O'Neill, L. D., Edupuganti, S., Goepfert, P. A.,… Mehrotra, D. V. (2009). Safety and immunogenicity of adenovirus-vectored near-consensus HIV type 1 clade B gag vaccines in healthy adults. AIDS Res Hum Retroviruses, 25(1), 103-114.
Harro, C., Sun, X., Stek, J. E., Leavitt, R. Y., Mehrotra, D. V., Wang, F.,… Merck, V. Study Group. (2009). Safety and immunogenicity of the Merck adenovirus serotype 5 (MRKAd5) and MRKAd6 human immunodeficiency virus type 1 trigene vaccines alone and in combination in healthy adults. Clin Vaccine Immunol, 16(9), 1285-1292. doi: 10.1128/CVI.00144-09
Havenga, M., Vogels, R., Zuijdgeest, D., Radosevic, K., Mueller, S., Sieuwerts, M.,… Goudsmit, J. (2006). Novel replication-incompetent adenoviral B-group vectors: high vector stability and yield in PER.C6 cells. J Gen Virol, 87(Pt 8), 2135-2143.
Heilbronn, R., & Weger, S. (2010). Viral vectors for gene transfer: current status of gene therapeutics. Handb Exp Pharmacol(197), 143-170. doi: 10.1007/978-3-642-00477-3_5
Hoganson, D.K., Ma, J.C., Asato, L., Ong, M., Printz, M.A., Huyghe, B.G.,… D'Andrea, M.J. (2002). Development of a Stable Adenoviral Vector Formulation. BioProcessing J., 1(1), 43-48.
Holman, D. H., Wang, D., Raviprakash, K., Raja, N. U., Luo, M., Zhang, J.,… Dong, J. Y. (2007). Two complex, adenovirus-based vaccines that together induce immune responses to all four dengue virus serotypes. Clin Vaccine Immunol, 14(2), 182-189.
Holterman, L., Vogels, R., van der Vlugt, R., Sieuwerts, M., Grimbergen, J., Kaspers, J.,… Havenga, M. (2004). Novel replication-incompetent vector derived from adenovirus type 11 (Ad11) for vaccination and gene therapy: low seroprevalence and non-cross-reactivity with Ad5. J Virol, 78(23), 13207-13215. doi: 10.1128/JVI.78.23.13207-13215.2004
Hu, X., Meng, W., Dong, Z., Pan, W., Sun, C., & Chen, L. (2011). Comparative immunogenicity of recombinant adenovirus-vectored vaccines expressing different forms of hemagglutinin (HA) proteins from the H5 serotype of influenza A viruses in mice. Virus Res, 155(1), 156-162. doi: 10.1016/j.virusres.2010.09.014
Kim, D. W., Uetsuki, T., Kaziro, Y., Yamaguchi, N., & Sugano, S. (1990). Use of the human elongation factor 1 alpha promoter as a versatile and efficient expression system. Gene, 91(2), 217-223.
Kobinger, G. P., Feldmann, H., Zhi, Y., Schumer, G., Gao, G., Feldmann, F.,… Wilson, J. M. (2006). Chimpanzee adenovirus vaccine protects against Zaire Ebola virus. Virology, 346(2), 394-401. doi: 10.1016/j.virol.2005.10.042
Lasaro, M. O., & Ertl, H. C. (2009). New insights on adenovirus as vaccine vectors. Mol Ther, 17(8), 1333-1339. doi: 10.1038/mt.2009.130
Lemckert, A. A., Grimbergen, J., Smits, S., Hartkoorn, E., Holterman, L., Berkhout, B.,… Havenga, M. J. (2006). Generation of a novel replication-incompetent adenoviral vector derived from human adenovirus type 49: manufacture on PER.C6 cells, tropism and immunogenicity. J Gen Virol, 87(Pt 10), 2891-2899. doi: 10.1099/vir.0.82079-0
Mullick, A., Xu, Y., Warren, R., Koutroumanis, M., Guilbault, C., Broussau, S.,… Massie, B. (2006). The cumate gene-switch: a system for regulated expression in mammalian cells. BMC Biotechnol, 6, 43. doi: 10.1186/1472-6750-6-43
Na, M., & Fan, X. (2010). Design of Ad5F35 vectors for coordinated dual gene expression in candidate human hematopoietic stem cells. Exp Hematol, 38(6), 446-452. doi: 10.1016/j.exphem.2010.03.007
Nan, X., Peng, B., Hahn, T. W., Richardson, E., Lizonova, A., Kovesdi, I., & Robert-Guroff, M. (2003). Development of an Ad7 cosmid system and generation of an Ad7deltaE1deltaE3HIV(MN) env/rev recombinant virus. Gene Ther, 10(4), 326-336. doi: 10.1038/sj.gt.3301903
Ogun, S. A., Dumon-Seignovert, L., Marchand, J. B., Holder, A. A., & Hill, F. (2008). The oligomerization domain of C4-binding protein (C4bp) acts as an adjuvant, and the fusion protein comprised of the 19-kilodalton merozoite surface protein 1 fused with the murine C4bp domain protects mice against malaria. Infect Immun, 76(8), 3817-3823. doi: 10.1128/IAI.01369-07
Ophorst, O. J., Radosevic, K., Havenga, M. J., Pau, M. G., Holterman, L., Berkhout, B.,… Tsuji, M. (2006). Immunogenicity and protection of a recombinant human adenovirus serotype 35-based malaria vaccine against Plasmodium yoelii in mice. Infect Immun, 74(1), 313-320.
Pham, L., Nakamura, T., Gabriela Rosales, A., Carlson, S. K., Bailey, K. R., Peng, K. W., & Russell, S. J. (2009). Concordant activity of transgene expression cassettes inserted into E1, E3 and E4 cloning sites in the adenovirus genome. J Gene Med, 11(3), 197-206.
Post, D. E., & Van Meir, E. G. (2001). Generation of bidirectional hypoxia/HIF-responsive expression vectors to target gene expression to hypoxic cells. Gene Ther, 8(23), 1801-1807. doi: 10.1038/sj.gt.3301605
Powell, S. K., Rivera-Soto, R., & Gray, S. J. (2015). Viral expression cassette elements to enhance transgene target specificity and expression in gene therapy. Discov Med, 19(102), 49-57.
Richardson, J. S., Yao, M. K., Tran, K. N., Croyle, M. A., Strong, J. E., Feldmann, H., & Kobinger, G. P. (2009). Enhanced protection against Ebola virus mediated by an improved adenovirus-based vaccine. PLoS One, 4(4), e5308. doi: 10.1371/journal.pone.0005308
Robbins, P. D., & Ghivizzani, S. C. (1998). Viral vectors for gene therapy. Pharmacol Ther, 80(1), 35-47.
Sandford, G. R., & Burns, W. H. (1996). Rat cytomegalovirus has a unique immediate early gene enhancer. Virology, 222(2), 310-317. doi: 10.1006/viro.1996.0428
Schepp-Berglind, J., Luo, M., Wang, D., Wicker, J. A., Raja, N. U., Hoel, B. D.,… Dong, J. Y. (2007). Complex adenovirus-mediated expression of West Nile virus C, PreM, E, and NS1 proteins induces both humoral and cellular immune responses. Clin Vaccine Immunol, 14(9), 1117-1126.
Sullivan, N. J., Geisbert, T. W., Geisbert, J. B., Shedlock, D. J., Xu, L., Lamoreaux, L.,… Nabel, G. J. (2006). Immune protection of nonhuman primates against Ebola virus with single low-dose adenovirus vectors encoding modified GPs. PLoS Med, 3(6), e177. doi: 10.1371/journal.pmed.0030177
Sullivan, N. J., Geisbert, T. W., Geisbert, J. B., Xu, L., Yang, Z. Y., Roederer, M.,… Nabel, G. J. (2003). Accelerated vaccination for Ebola virus haemorrhagic fever in non-human primates. Nature, 424(6949), 681-684. doi: 10.1038/nature01876
Tatsis, N., Blejer, A., Lasaro, M. O., Hensley, S. E., Cun, A., Tesema, L.,… Ertl, H. C. (2007). A CD46-binding chimpanzee adenovirus vector as a vaccine carrier. Mol Ther, 15(3), 608-617. doi: 10.1038/sj.mt.6300078
Vemula, S. V., & Mittal, S. K. (2010). Production of adenovirus vectors and their use as a delivery system for influenza vaccines. Expert Opin Biol Ther, 10(10), 1469-1487. doi: 10.1517/14712598.2010.519332
Vogels, R., Zuijdgeest, D., van Meerendonk, M., Companjen, A., Gillissen, G., Sijtsma, J.,… Havenga, M. J. (2007). High-level expression from two independent expression cassettes in replication-incompetent adenovirus type 35 vector. J Gen Virol, 88(Pt 11), 2915-2924.
Vogels, R., Zuijdgeest, D., van Rijnsoever, R., Hartkoorn, E., Damen, I., de Bethune, M. P.,… Havenga, M. (2003). Replication-deficient human adenovirus type 35 vectors for gene transfer and vaccination: efficient human cell infection and bypass of preexisting adenovirus immunity. J Virol, 77(15), 8263-8271.
Voigt, S., Sandford, G. R., Hayward, G. S., & Burns, W. H. (2005). The English strain of rat cytomegalovirus (CMV) contains a novel captured CD200 (vOX2) gene and a spliced CC chemokine upstream from the major immediate-early region: further evidence for a separate evolutionary lineage from that of rat CMV Maastricht. J Gen Virol, 86(Pt 2), 263-274. doi: 10.1099/vir.0.80539-0
Walther, W., & Stein, U. (2000). Viral vectors for gene transfer: a review of their use in the treatment of human diseases. Drugs, 60(2), 249-271.
Zhou, D., Cun, A., Li, Y., Xiang, Z., & Ertl, H. C. (2006). A chimpanzee-origin adenovirus vector expressing the rabies virus glycoprotein as an oral vaccine against inhalation infection with rabies virus. Mol Ther, 14(5), 662-672. doi: 10.1016/j.ymthe.2006.03.027
Zhou, D., Wu, T. L., Lasaro, M. O., Latimer, B. P., Parzych, E. M., Bian, A.,… Ertl, H. C. (2010). A universal influenza A vaccine based on adenovirus expressing matrix-2 ectodomain and nucleoprotein protects mice from lethal challenge. Mol Ther, 18(12), 2182-2189. doi: 10.1038/mt.2010.202.
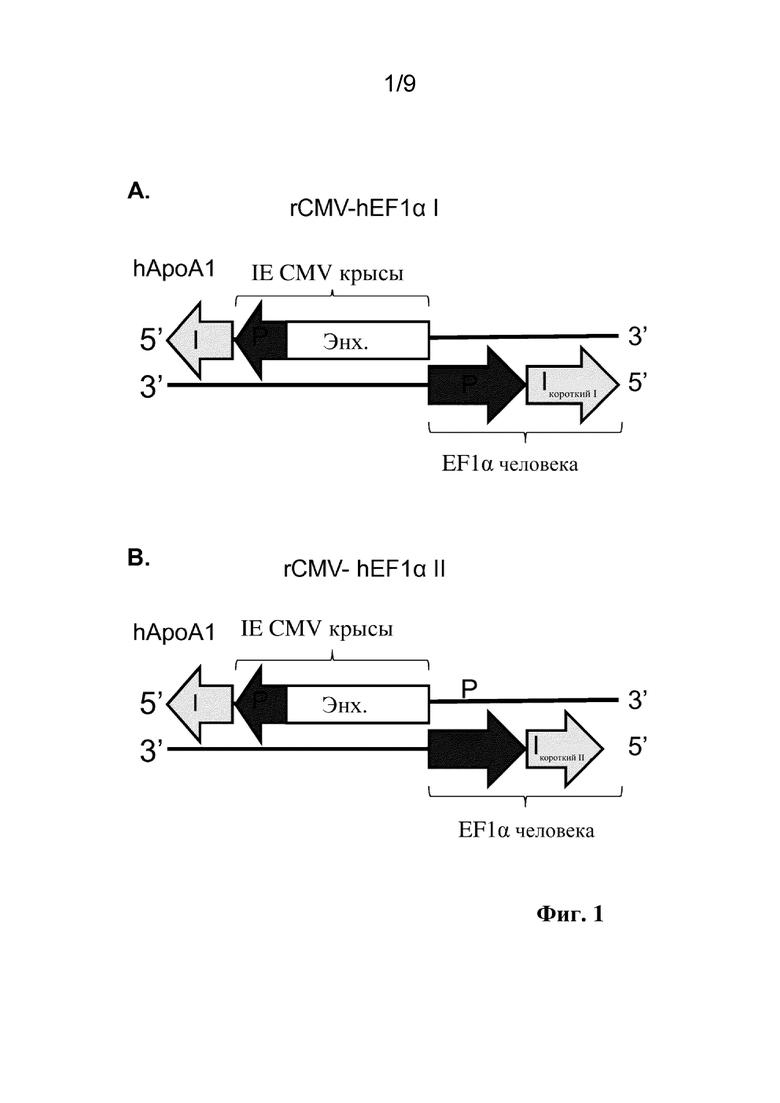
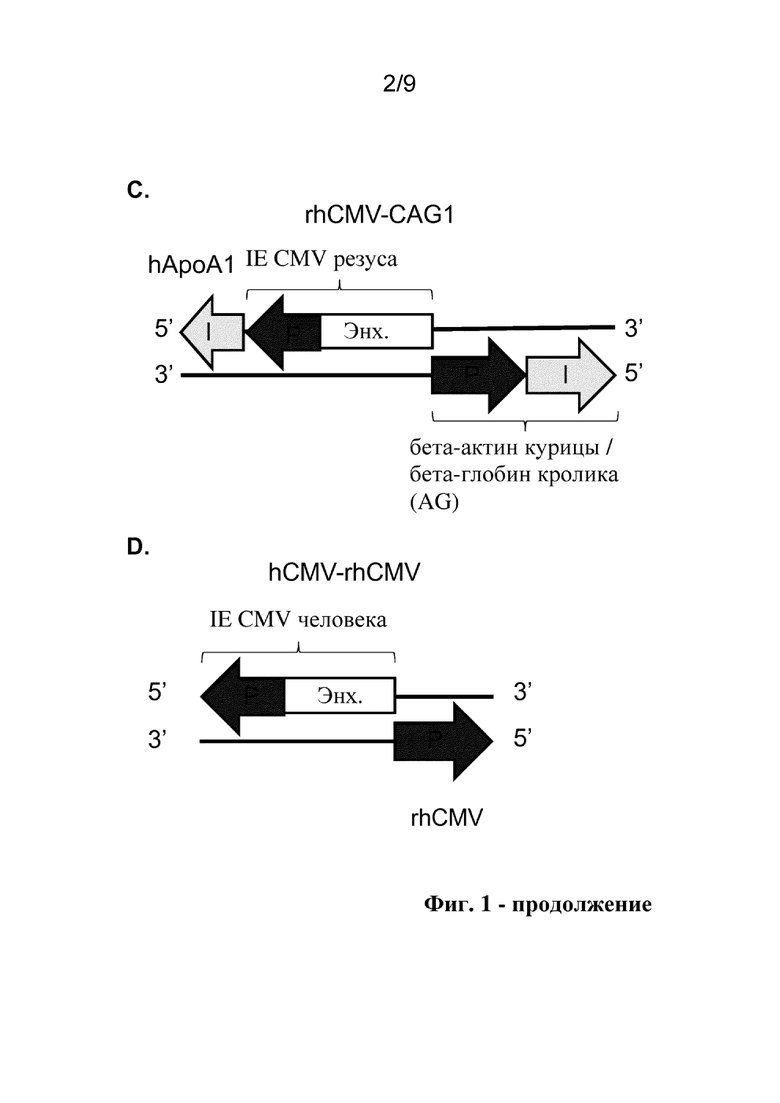
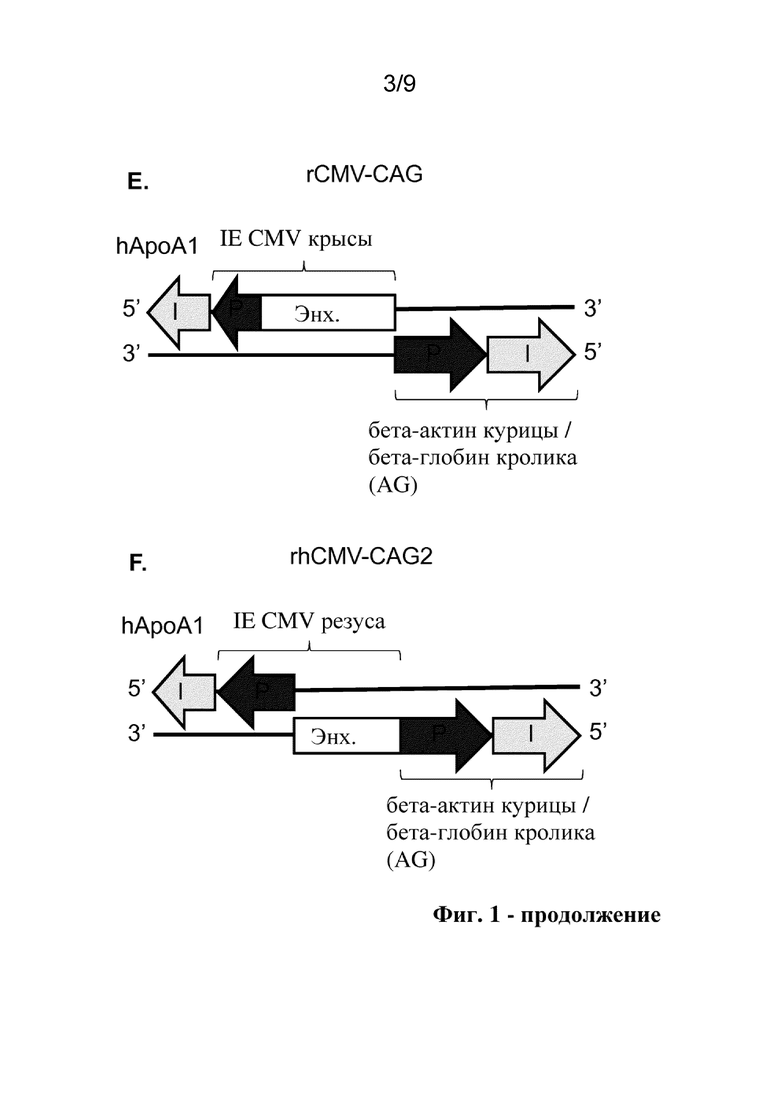
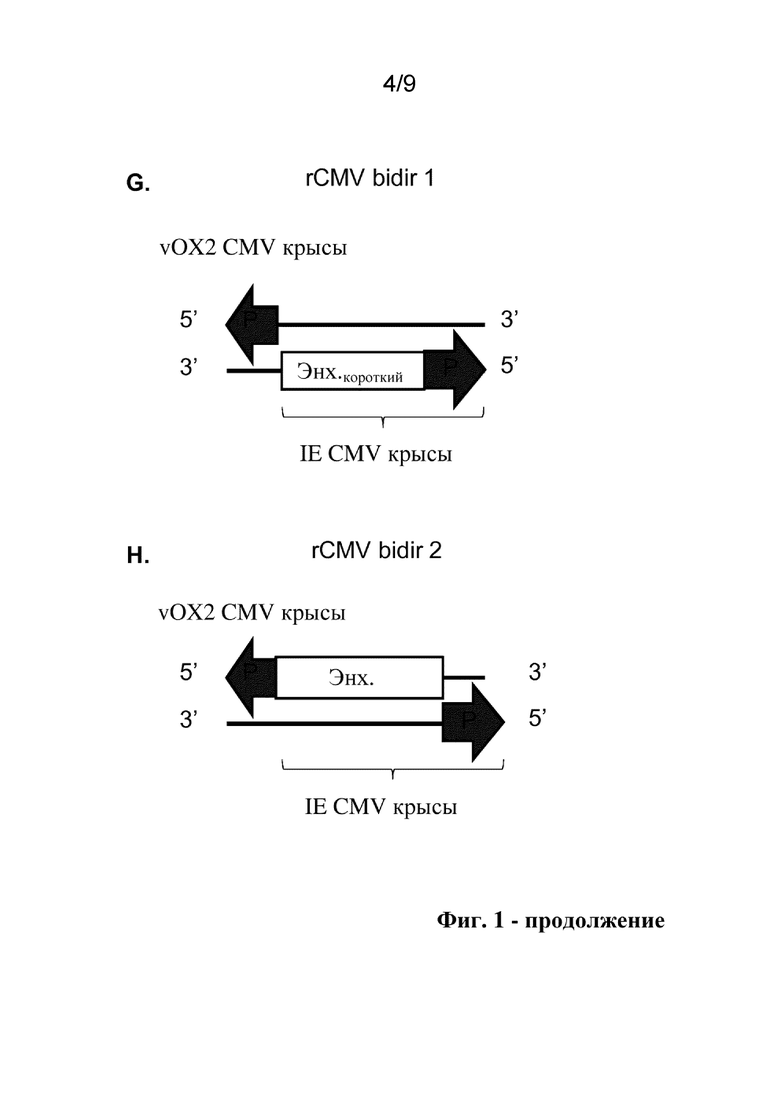
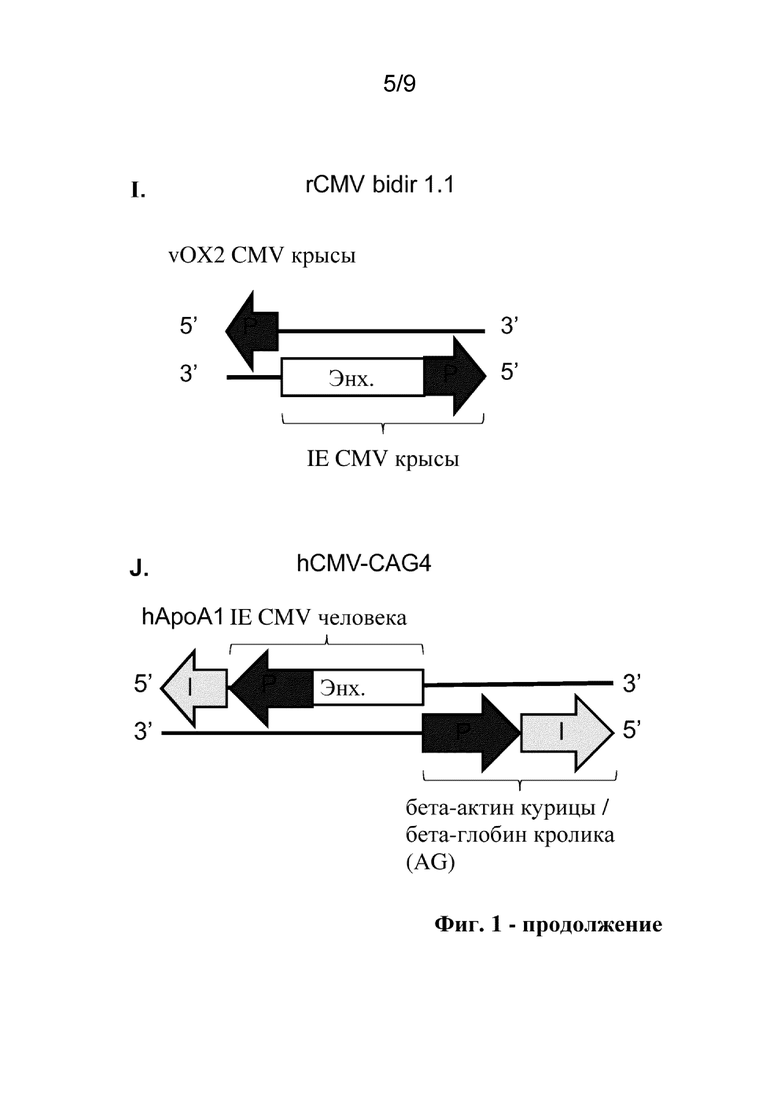
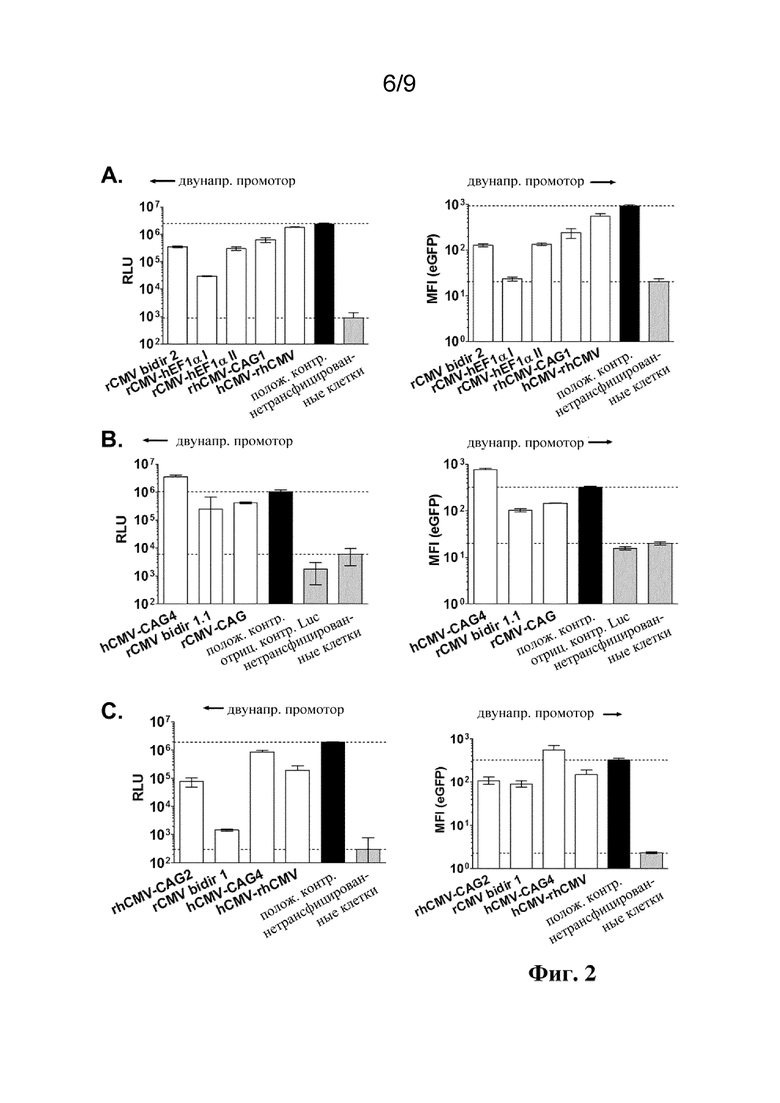
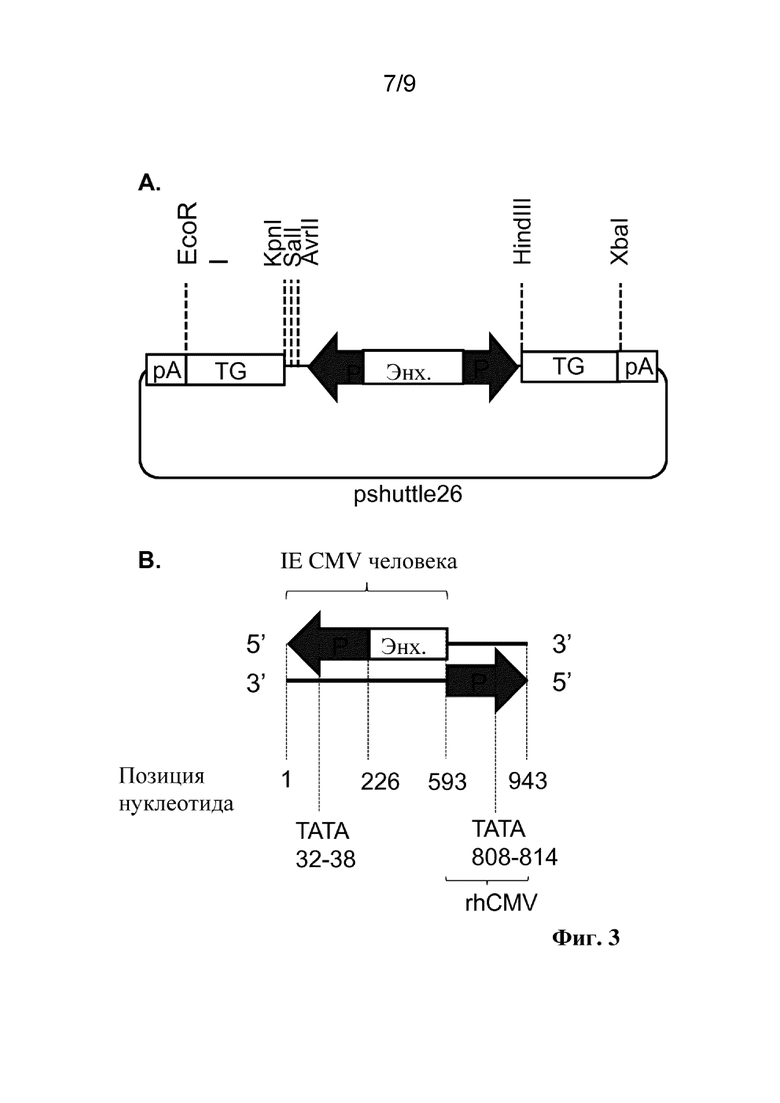
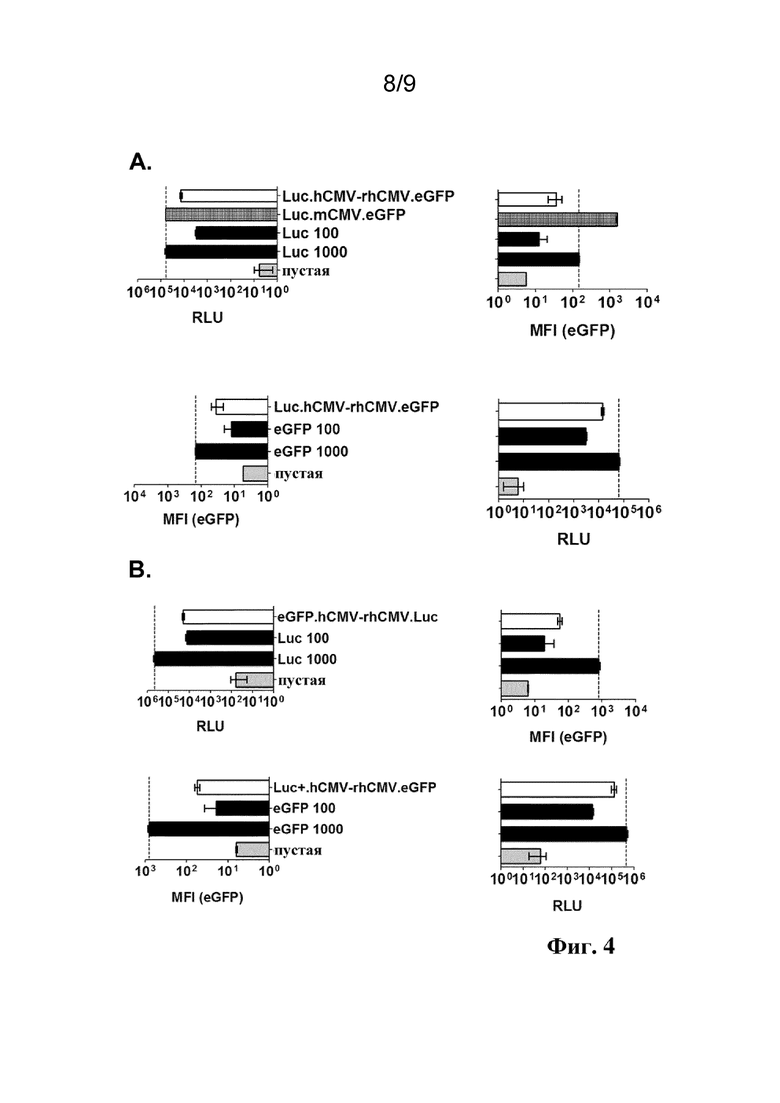
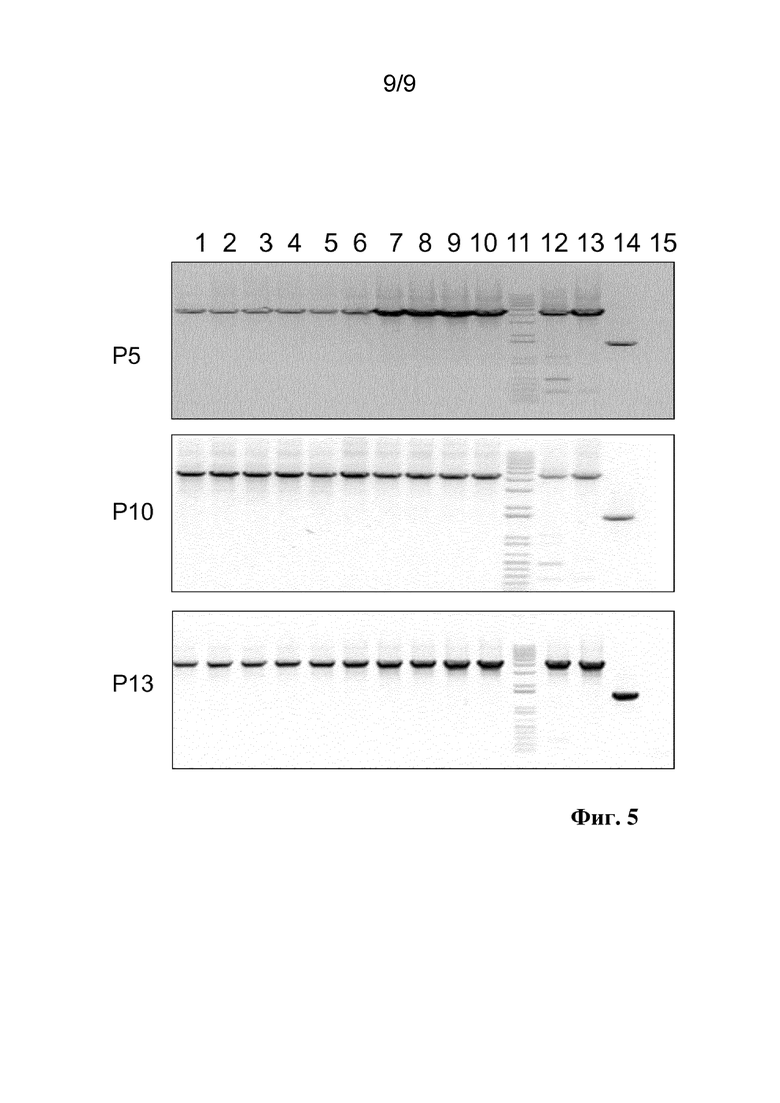
Изобретение относится к биотехнологии. Описана молекула рекомбинантной нуклеиновой кислоты для экспрессии первого и второго трансгена, содержащая двунаправленный промотор (промотор hCMV-rhCMV), функционально связанный с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении, где промотор hCMV-rhCMV содержит: (i) энхансер, фланкированный (ii) основным немедленно-ранним промотором цитомегаловируса человека (промотором hCMV) на одной стороне энхансера и (iii) основным немедленно-ранним промотором цитомегаловируса макака-резуса (промотором rhCMV) на другой стороне энхансера, где (iv) первый трансген расположен ниже промотора hCMV, и (v) второй трансген расположен ниже промотора rhCMV. Представлен рекомбинантный вектор для экспрессии первого и второго трансгена, содержащий молекулу рекомбинантной нуклеиновой кислоты и рекомбинантный вирус для экспрессии первого и второго трансгена, содержащий молекулу рекомбинантной нуклеиновой кислоты. Описан способ получения генетически стабильного рекомбинантного аденовируса, содержащего первый и второй трансгены, где каждый из них эффективно экспрессируется, в случае если аденовирус инфицирует целевую клетку, включающий: a) получение конструкции, содержащей двунаправленный промотор hCMV-rhCMV, функционально связанный с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении; и b) встраивание указанной конструкции в геном рекомбинантного аденовируса. Описан способ экспрессии по меньшей мере двух трансгенов в клетке, включающий доставку в клетку рекомбинантного вектора или рекомбинантного вируса. Описан способ индукции иммунного ответа против по меньшей мере двух антигенов, включающий введение пациенту рекомбинантного вектора или рекомбинантного вируса. Описана молекула рекомбинантной ДНК для экспрессии первого и второго трансгена, содержащая геном рекомбинантного вируса. Описана композиция для экспрессии первого и второго трансгена, содержащая рекомбинантный вектор или рекомбинантный вирус и фармацевтически приемлемые носитель или эксципиент. Изобретение расширяет арсенал средств для экспрессии трансгена. 8 н. и 10 з.п. ф-лы, 1 табл., 4 пр., 5 ил.
1. Молекула рекомбинантной нуклеиновой кислоты для экспрессии первого и второго трансгена, содержащая двунаправленный промотор (промотор hCMV-rhCMV), функционально связанный с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении, где промотор hCMV-rhCMV содержит:
(i) энхансер, фланкированный
(ii) основным немедленно-ранним промотором цитомегаловируса человека (промотором hCMV) на одной стороне энхансера и
(iii) основным немедленно-ранним промотором цитомегаловируса макака-резуса (промотором rhCMV) на другой стороне энхансера, где
(iv) первый трансген расположен ниже промотора hCMV, и
(v) второй трансген расположен ниже промотора rhCMV.
2. Молекула рекомбинантной нуклеиновой кислоты по п.1, где энхансер представляет собой основной немедленно-ранний энхансер цитомегаловируса человека (энхансер hCMV).
3. Молекула рекомбинантной нуклеиновой кислоты по любому из пп.1 или 2, где первый и второй трансгены являются разными, и по меньшей мере один из них кодирует антиген.
4. Рекомбинантный вектор для экспрессии первого и второго трансгена, содержащий молекулу рекомбинантной нуклеиновой кислоты по любому из пп.1-3.
5. Рекомбинантный вектор по п.4, где вектор представляет собой плазмидный вектор.
6. Рекомбинантный вирус для экспрессии первого и второго трансгена, содержащий молекулу рекомбинантной нуклеиновой кислоты по любому из пп.1-3.
7. Рекомбинантный вирус по п.6, где вирус представляет собой аденовирус.
8. Рекомбинантный вирус по п.7, где аденовирус имеет делецию в участке E1.
9. Рекомбинантный вирус по п.7 или 8, где аденовирус представляет собой аденовирус человека серотипа 35 или аденовирус человека серотипа 26.
10. Способ получения генетически стабильного рекомбинантного аденовируса, содержащего первый и второй трансгены, где каждый из них эффективно экспрессируется, в случае если аденовирус инфицирует целевую клетку, включающий:
a) получение конструкции, содержащей двунаправленный промотор hCMV-rhCMV, определенный в п.1, функционально связанный с первым трансгеном в одном направлении и со вторым трансгеном в противоположном направлении; и
b) встраивание указанной конструкции в геном рекомбинантного аденовируса.
11. Способ по п.10, где энхансер представляет собой энхансер hCMV.
12. Способ по любому из пп.10 или 11, где рекомбинантный аденовирус имеет делецию в участке E1 его генома.
13. Способ по любому из пп.10-12, где первый и второй трансгены являются разными, и по меньшей мере один из них кодирует антиген.
14. Способ по любому из пп.10-13, где рекомбинантный аденовирус представляет собой аденовирус человека серотипа 35 или аденовирус человека серотипа 26.
15. Способ экспрессии по меньшей мере двух трансгенов в клетке, включающий доставку в клетку рекомбинантного вектора или рекомбинантного вируса по любому из пп.4-9.
16. Способ индукции иммунного ответа против по меньшей мере двух антигенов, включающий введение пациенту рекомбинантного вектора или рекомбинантного вируса по любому из пп.4-9.
17. Молекула рекомбинантной ДНК для экспрессии первого и второго трансгена, содержащая геном рекомбинантного вируса по любому из пп.7-9.
18. Композиция для экспрессии первого и второго трансгена, содержащая рекомбинантный вектор или рекомбинантный вирус по любому из пп.4-9 и фармацевтически приемлемые носитель или эксципиент.
| Станок для наматывания нитей на шпули | 1929 |
|
SU21009A1 |
| WO 2013184764 A2, 12.12.2013. | |||

