Изобретение относится к области морской аквакультуры и предназначено для получения массы высококачественных живых кормовых организмов для использования в качестве живых кормов при выращивании в аквакультуре ранних стадий развития морских ракообразных и рыб, в том числе личинок ценных морских рыб, в морской аквариумистике и для проведения экспериментальных работ в области научных исследований по биологии, эмбриологии, генетике, физиологии, биохимии и биологического тестирования.
Известно, что характерной чертой личинок многих ценных пород морских рыб является облигатное питание живыми кормами соответствующей пищевой ценности и размеров, особенно, для стадий развития в период раннего метаморфоза. Обычно, в интенсивной марикультуре для кормления личинок рыб используют в качестве живых кормов последовательно солоноватоводных коловраток Brachionus plicatilis и науплиев ракообразных гиперсоленых озер Artemia spp., а в естественных условиях личинки питаются смесью планктонных каланодных копепод и кладоцер, обеспечивающих их нормальное развитие. Использование морских копепод в массовых количествах в условиях аквакультуры разработано, но в накопительной культуре их популяции имеют смешанную структуру, состоящую из 6 науплиальных, 5 копеподитных и взрослых жизненных стадий, для которых необходимо функционирование многочисленных водных бассейнов больших объемов, поэтому был предложен «Способ длительного хранения покоящихся яиц копепод акарций для получения массовой синхронной культуры одновозрастных науплиев» (Ханайченко, 2017), как дополнение к применению «Способа интенсивного когортного культивирования акарций (морских каланоидных копепод)» (Ханайченко, 2018). Каланоидные копеподы являются оптимальным естественным кормом личинок морских рыб, обеспечивающим их необходимым количеством и соотношением незаменимых эссенциальных компонентов пищи, таких как фосфолипиды, полиненасыщенные жирные кислоты, свободные аминокислоты и высокоэффективные антиоксиданты (каротиноиды) - астаксантин. Однако индивидуальная биомасса копепод невелика, а личинкам в процессе метаморфоза необходимо получать наиболее энергетически-емкие и малоподвижные живые корма, при поимке которых личинки могут получать значительное количество энергии при незначительных энергетических расходах.
Известен способ накопительного культивирования моин («Seawater domestication and indoor high-density culture method of Moina mongolica Daday»), по которому моин при начальной плотности 100/л помещают в бассейн, и постепенно повышают соленость на 1-2 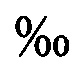 до достижения солености 35 , поддерживают освещенность 5000-7000 лк при фотопериоде L:D=12 ч: 12 ч, температуре 25-28°С, производя ежедневное кормление хлореллой (2 млн. кл/мл), а при достижении плотности моин 2000-5000/л, их переводят в проточный бассейн при непрерывном добавлении в него смеси хлореллы и дрожжей, и при поддержании уровня растворенного кислорода в воде 6-8 мг/л, рН воды 7-8 доращивают моин до плотности 15000-20000/л, производят сбор биомассы и повторяют цикл. Недостатками вышеприведенного метода накопительного культивирования является невозможность контроля перехода от партеногенетического (бесполого, или однополого) способа размножения к половому, неконтролируемый биохимический состав и возможность бактериального загрязнения кормов. При этом известно, что при неконтролируемом внезапном переходе к половому типу размножения самки моин производят массу покоящихся яиц в эфиппиумах, что приводит к снижению количества реальной продукции самок моин, т.е. продукции однородных живых организмов, пригодных для кормления личинок рыб. Количество молоди, производимой самками моин при этих способах культивирования, непредсказуемо. При использовании этого метода в бассейны для выращивания личинок могут быть перенесены патогенные микроорганизмы, влияющие на выживаемость личинок рыб. При питании хлореллой и дрожжами содержание ненасыщенных жирных кислот в составе моин не может удовлетворять потребности личинок.
до достижения солености 35 , поддерживают освещенность 5000-7000 лк при фотопериоде L:D=12 ч: 12 ч, температуре 25-28°С, производя ежедневное кормление хлореллой (2 млн. кл/мл), а при достижении плотности моин 2000-5000/л, их переводят в проточный бассейн при непрерывном добавлении в него смеси хлореллы и дрожжей, и при поддержании уровня растворенного кислорода в воде 6-8 мг/л, рН воды 7-8 доращивают моин до плотности 15000-20000/л, производят сбор биомассы и повторяют цикл. Недостатками вышеприведенного метода накопительного культивирования является невозможность контроля перехода от партеногенетического (бесполого, или однополого) способа размножения к половому, неконтролируемый биохимический состав и возможность бактериального загрязнения кормов. При этом известно, что при неконтролируемом внезапном переходе к половому типу размножения самки моин производят массу покоящихся яиц в эфиппиумах, что приводит к снижению количества реальной продукции самок моин, т.е. продукции однородных живых организмов, пригодных для кормления личинок рыб. Количество молоди, производимой самками моин при этих способах культивирования, непредсказуемо. При использовании этого метода в бассейны для выращивания личинок могут быть перенесены патогенные микроорганизмы, влияющие на выживаемость личинок рыб. При питании хлореллой и дрожжами содержание ненасыщенных жирных кислот в составе моин не может удовлетворять потребности личинок.
Задачей Способа культивирования моин является разработка технологии культивирования партеногенетических самок моин; стимуляции полового размножения, получения, сбора и хранения эфиппиумов для получения в искусственных условиях массовой моновидовой популяции (культуры) моины.
Технический результат от решения поставленной задачи заключается в том, что сочетание технических приемов позволяет манипулировать составом искусственной популяции моин: поддерживать ее постоянно в состоянии партеногенеза для получения массы самок, или создавать специфические условия для получения покоящихся стадий. Технический результат заключается также в пролонгированном хранении диапаузиирующих яиц моин в эфиппиумах в условиях пониженного метаболизма, применении разработанной системы активации метаболизма и запрограммированном по времени единовременном выклеве большой массы партеногенетических самок моин.
Заявленный технический результат достигается тем, что Способ культивирования моин для использования в качестве живых кормов в аквакультуре предусматривает, что изъятых из естественной зоопланктонной пробы самок моин выдерживают в дезинфекционном растворе с добавлением микроводорослей в течение 6-8 часов, помещают в контейнеры со стерильной морской водой при минимальном объеме 30 мл на самку, и постоянной освещенностью 5000 лк, осуществляют кормление микроводорослями. Затем полученное потомство используют для накопительного культивирования при партеногенетической стадии размножения, которое проводят при температуре 24±2°С, освещенности на поверхности культиватора 5000 лк, фотопериоде не менее 16 часов освещения и 8 часов темноты, при последовательном наращивании плотности самок до 300 экз./л, в несменяемой среде, с ежедневным внесением микроводорослей. После чего при поддержании постоянной плотности самок около 300 экз/л, проводят наращивание объема культуры, при ежедневном добавлении стерильной морской воды с микроводорослями, после чего проводят культивирование моин с ежедневным изъятием около 0,25 объема культуры, заменой данного объема стерильной морской водой и внесением микроводорослей, в качестве которых используют Isochrysis galbana, Tetraselmis suecica и Rhodomonas sp., поддерживая их концентрацию во время партеногенетической стадии размножения от 3 до 10 мкг/мл. По завершении наращивания биомассы, партеногенетических самок моин переводят в стадию полового размножения, увеличивая плотность культуры до 3000-5000 экз/л и выдерживая при этой плотности 48 часов, снижая за это время температуру воды с 25°С до 15°С, при освещенности на поверхности культиватора 500 лк, фотопериоде 5-6 часов освещения и 18-19 часов темноты, и концентрации микроводорослей 1-2 мкг/л. Полученные в результате полового размножения эфиппиумы высушивают и хранят в течение 6-24 месяцев. Для перевода покоящихся эмбрионов моин в активное состояние и получения ювенильных партеногенетических самок моин, эфиппиумы помещают при плотности около 300 экз/л на 24-48 часов в стерильную холодную морскую воду соленостью 18 , промывают несколько раз, и постепенно в течение 12 час поднимают температуру воды до 25°С, одновременно повышая уровень кислорода до 85-100% насыщения, при постоянной освещенности на поверхности инкубатора не менее 1000 лк.
Отличительные признаки изобретения, совокупное осуществление которых позволяет достигнуть заявленный технический результат, включают:
- создание комплексных условий для накопительного культивирования моин с использованием партеногенетической фазы цикла;
- создание комплексных условий стимуляции продуцирования самками моин массового количества эфиппиумов с диапаузиирующими яйцами;
- одновременный сбор массы эфиппиумов в течение не позднее 12 час после их откладки;
- перевод массы диапаузиирующих яиц в эфиппиумах в состояние длительного покоя в одинаковых условиях для обеспечения хранения яиц и синхронизации выклева партеногенетических самок моин после активации;
- соблюдение стабильных стандартизованных условий длительного хранения диапаузиирующих яиц моин в эфиппиумах для поддержания их жизнеспособности;
- создание стандартных условий активации диапаузиирующих яиц для синхронного вылупления из эфиппиумов партеногенетических самок моин.
Заявляемый способ соответствует критериям «новизна» и «изобретательский уровень» т.к. впервые предложены оптимальные температурные, трофические и плотностные условия для синхронизации и стандартизации процессов продуцирования постоянного однополого размножения и воспроизводства исключительно партеногенетических самок солоноватоводных моин. Заявленный способ может быть осуществлен промышленным способом с использованием известного стандартного оборудования.
Способ поясняется графиками, представленными на Фиг. 1-Фиг. 4:
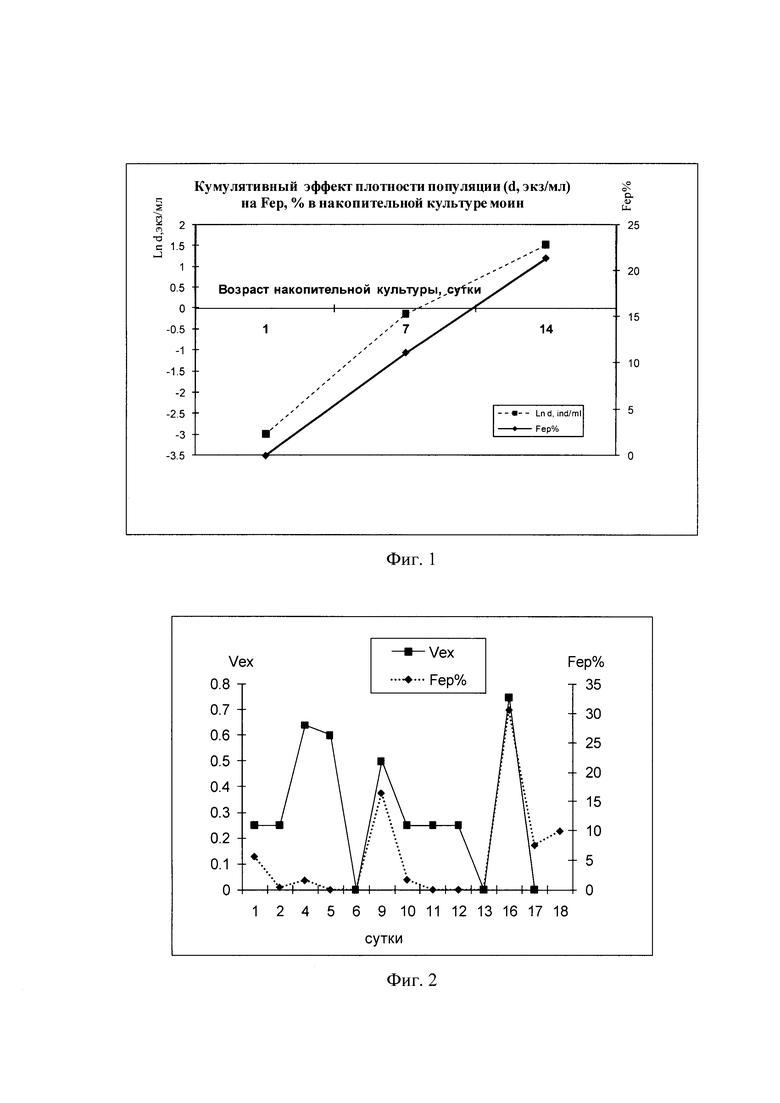
Фиг. 1 - Зависимость доли самок с эфиппиумами от возраста накопительной культуры и плотности моин;
Фиг. 2 - Влияние объема сменяемой среды (Vex, доля объема) в полупроточной культуре моин на долю самок с эфиппиумами (Fep%);
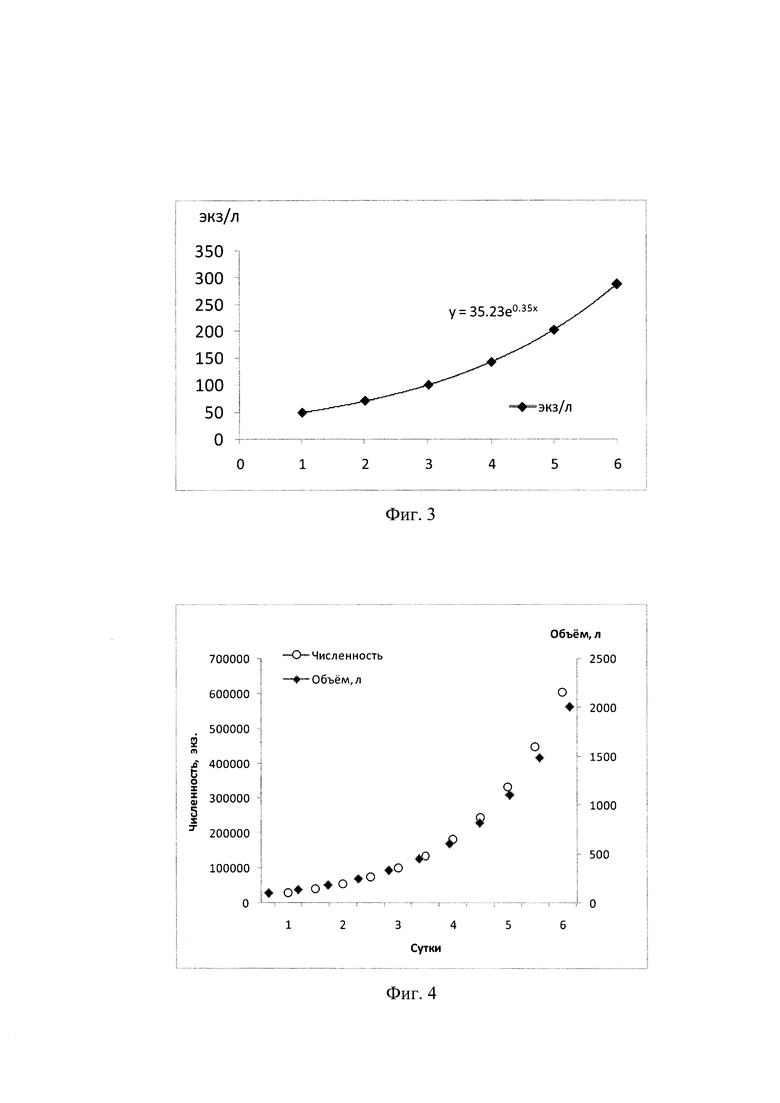
Фиг. 3 - Наращивание плотности моин (от 50 до 300 экз/л) в 100-л объеме при 24±2°С с ежедневным добавлением пищевых микроводорослей (около 5 мкг/мл);
Фиг. 4 - Наращивание объема культуры моин при поддержании постоянной плотности (300 экз/л) от начального 100 л до объема массовой эксплуатации - 2 м3 при ежедневном добавлении дезинфицированной морской воды с микроводорослями при 24±2°С. По оси ординат - рост численности популяции моин (экз.) соответственно увеличению объема (л).
В связи с недостаточной изученностью биологии питания морских кладоцер (Penilia spp., Podon spp., Evadne spp., Pleopis spp.) (Egloff et al., 1997) методы их культивирования до настоящего времени не разработаны. Однако в ларвикультуре морских рыб можно успешно использовать галофильных кладоцер, Moina salina Daday, 1888 (синоним Moina mongolica Daday, 1901 no He, 1987) (Crustacea: Branchiopoda: Cladocera: Moinidae), для которых характерен циклический партеногенез, характеризующийся разделением бесполой (иначе, однополой, а именно, партеногенетических самок) и половой (миктических самок и самцов) фаз размножения (Hebert, 1978).
В течение бесполой (однополой) фазы цикла воспроизводства популяции моин партеногенетическая самка в течение жизненного цикла проходит серию личиночных стадий (от 1-й до 3-й) и половозрелых стадий самки (от 4-й до 6-7-й). Переход от одной стадии к другой осуществляется в результате морфологических изменений, нарастания массы и размеров тела и смены покровов (линьки). После перехода (линьки) в половозрелую стадию самки, моина не достигает сразу терминальных размеров тела, а продолжает расти. Единовременно с созреванием каждой очередной порции яиц (прохождении зародышами моин полного эмбрионального развития в выводковой камере), происходит рост карапакса самки. Вылупление молоди моин из эмбриональной оболочки происходит одновременно с разрывом общей оболочки кладки в выводковой камере, и линькой самки с образованием нового карапакса. Таким образом, происходит дальнейший прирост биомассы половозрелой самки моины за счет отложения запасных веществ и формирования генеративной продукции (яиц). Прохождение очередной линьки самок моин обусловлено запрограммированными генетически механизмами и зависит от комплекса модифицирующих их температурных и трофических условий содержания.
Половая фаза цикла. Комплекс неблагоприятных факторов (повышение плотности организмов в среде, резкое изменение обеспеченности пищей, накопление метаболитов в среде), воздействующие на ювенильных самок в процессе развития стимулируют к изменению условий развития яйцеклеток, и приводят к тому, что яйцеклетки проходят мейоз, в результате которого они становятся гаплоидными (1n). В отсутствии оплодотворения развивающийся из гаплоидной яйцеклетки организм становится гаплоидным - самцом, производящим гаметы (сперматозоиды). При условии оплодотворения самцами ювенильных самок с яйцеклетками, прошедшими мейоз, происходит слияние гамет, и у самок развиваются оплодотворенные диплоидные яйца (1-2), которые покрываются хитиновой оболочкой, называемой эфиппиумом, и, эмбрионы в них находятся в состоянии диапаузы, и могут выдерживать длительный период покоя до наступления благоприятных условий. При наступлении сочетания благоприятных условий диапауза эмбриона моин прерывается, и через 1-2 суток происходит вылупление ювенильных партеногенетических самок, которые дают начало новой популяции партеногенетических самок
Экспериментальное культивирование моин. Для разработки данного метода в экспериментальных условиях были исследованы условия поддержания партеногенетической фазы цикла в культуре моин, а также влияние некоторых факторов на переход от бесполого типа размножения (партеногенеза), к половому (продукции самками самцов, и последующему производству оплодотворенными самками покоящегося яйца в цисте - эфиппиума).
Оценку различных факторов среды на параметры роста популяции моин проводили в условиях накопительного культивирования (постепенно возрастающего по мере роста численности популяции и объема культиватора) и полупроточного культивирования (в условиях одного и того же объема культиватора при периодическом изъятии части объема культуры вместе с пропорциональной частью популяции и замены его новой средой того же объема). Исследования показали, что любое изменение (1-3) в питании, а именно: смена вида кормовой микроводоросли (1); биохимический состав микроводорослей, изменяющегося от экспоненциальной к стационарной фазе культивирования) (2); количества корма (3) может вызывать изменение скорости воспроизводства и изменить выживаемость потомства моин, но не влияет на смену типа размножения моин с однополого (партеногенеза) на половое размножение.
На смену типа размножения и долю самок с эфиппиями в массовой культуре влияет нарастание плотности животных (соответственно, снижение обеспеченности пищей и возрастание концентрации метаболитов их жизнедеятельности) в несменяемой среде при накопительном культивировании (Фиг. 1), или снижение скорости разбавления среды с извлечением части объема культивационной жидекости с частью популяции моин и замена ее свежей средой с кормом с целью снижения плотности животных (Фиг. 2).
При накопительном культивировании при плотности посадки партеногенетических самок моин 50 экз/л в популяции отсутствуют самцы и самки, несущие диапаузиирующие яйца в эфиппиях. В течение первой недели выращивания моин нарастание численности популяции в накопительной культуре происходит со скоростью около 0,42 сут-1 до плотности животных 990 экз/л. К окончанию первой недели процент самок, несущих диапаузиирующие яйца в эфиппиях вырастает до 10% от общей численности популяции. В течение второй недели выращивания скорость роста численности моин в накопительной культуре замедляется вдвое, составляя около 0,21 сут-1, в результате перехода части самок на половое размножение, и процент самок, несущих диапаузиирующие яйца в эфиппиях вырастает до 22% от общей численности популяции (Фиг. 1).
При выращивании моин в режиме полу-проточного культивирования доля самок с эфиппиумами звисит от скорости разбавления среды (подмены культивационной жидкости чистой водой с добавлением пропорционально количества пищи) (Фиг. 2). Так, в несменяемой в течение 3-4 суток (с 5 по 9 сут и с 12 по 16 сут, при нарастании плотности от 100 до 500 экз/л) среде доля самок с эфиппиумами вырастает до около 20% и около 30%, соответственно, а при ежедневной подмене -30% культуры свежей средой с добавлением пищи их доля снижается до 0, моины размножаются исключительно партеногенезом, продуцируя идентичных диплоидных самок.
Пример реализации способа
Выделение моим в культуру. Инокулят для получения искусственной популяции моин - интактные партеногенетические самки - были выделены из солоноватоводных прибрежных водоемов Керченского полуострова Крыма (характеризующихся большими сезонными колебаниями температуры (0-30°С) и солености (4-25%), в планктоне которых сезонно доминируют моины (Семик, Ушакова, 2017). Сразу же после получения живой пробы, в лаборатории, здоровых половозрелых самок моины отделяли и перемещали в дезинфекционный раствор. Дезинфекционный раствор готовили на основе стерилизованной морской воды стандартной черноморской солености 17,5-18,5%. Изолированных интактных самок при плотности 0,05 экз/мл, освещенности ~1000 люкс и температуре 22±2°С выдерживали в дезинфекционном растворе с добавлением микроводорослей (концентрация ~5⋅104 клеток на мл) в течение 6-8 часов. После процедуры дезинфекции самок моин промывали стерильной морской водой и помещали в контейнеры со стерильной морской водой (при минимальном объеме 30 мл на самку) Кормление моин производили монокультурами или смесью зеленых морских микроводорослей (при концентрации, варьирующей в процессе культивирования от 3 до 10 мкг/мл), выращенными на среде Тамия при постоянном освещении 5000 люкс. Полученное потомство от «диких» самок моин было использовано для начала накопительного культивирования моин с использованием партеногенетической фазы цикла. В благоприятных температурных, трофических и световых условиях происходило однополое размножение (партеногенетическая фаза), во время которой популяция моин состоит из амиктических самок, у которых в яйцеклетках не проходит мейоз, и они остаются диплоидными (2n); партеногенетические самки производят из партеногенетических диплоидных яиц генетически идентичное потомство самок (клоны). При оптимальных температурных (24±2°С) и трофических условиях (около 106 кл/сут/экз) самки моины размножались партеногенетически, т.е. производили идентичных себе партеногенетических самок. Длина карапакса ювенильных самок моин после вылупления из паргеногенетическош яйца составляла 0,45±0,02 мм. До достижения половозрелости партеногенетические самки проходят три последовательные линьки в течение 4-5 суток, и при длине карапакса 0,63±0,03 мм начинают размножаться партеногенетически. Созревание очередной порции диплоидных партеногенетических яиц в выводковой сумке и вылупление очередного потомства сопровождается очередной линькой самки (от 3 до 5 в течение жизненного цикла), что приводит к увеличению размера ее карапакса. Терминальный размер карапакса самки моины к концу жизненного цикла в условиях культивирования составляет 1,23(0,03) мм. Средняя численность молоди моин, полученной из одной кладки партеногенетичской самки, может варьировать от 2 до 17 в зависимости от возраста самки и изменения концентрации пищи от 1 до 10 мкг/мл. Созревание порции яиц (кладки) в выводковой сумке самки происходит в среднем через 2-3 сут. Длительность эмбрионального развития (от 3 до 1 сут), времени генерации (от 11 до 4 сут) и минимальный временной интервал между кладками (от 6 до 1 сут) обратно пропорциональны возрастанию температуры от 15±1 до 26±2°С при поддержании концентрации пищи в диапазоне 3-10 мкг/мл и плотности моин менее 300 экз/л. Партеногенетическая фаза длится, пока не нарушаются благоприятные условия (сочетание оптимальных температур, обилия пищи, содержания кислорода и других факторов окружающей среды). Длительность жизненного цикла партеногенетических самок обратно пропорциональна температуре культивирования.
Накопительное культивирование моин для получения массовой культуры для кормления личинок камбалы калкан проводили при последовательном наращивании плотности самок от 0,05 до 0,3 экз/мл. Культивирование (воспроизводство искусственной моновидовой популяции) моин осуществляли на стерилизованной (путем ультрафильтрации и УФ облучения черноморской воде около 18%). Моин кормили смесью культивируемых морских микроводорослей Haptophyceae (Isochrysis galbana), Prasinophyceae Tetraselmis suecica and Cryptophyceae (Rhodomonas sp.) выращенными на среде Тамия. Смесь водорослей вносили ежедневно, концентрация их варьировала в процессе питания моин в пределах 3-10 мкг/мл. Освещенность на поверхности культиваторов поддерживали около 5000 люкс на поверхности воды, при фотопериоде >16 ч: 8 ч и 24±2°С.
Для получения массовой партеногенетической культуры моин производили накопительное культивирование моин (наращивание от начальной до максимальной плотности 300 экз/л в несменяемой среде с ежедневным внесением корма) проводили сначала в 20 л, а затем в 100 л объемах (Фиг. З). После достижения оптимальной плотности партеногенетической популяции моин в массовой культуре производили наращивание объема культуры моин при поддержании постоянной плотности (300 экз/л) от начального 100 л до объема массовой эксплуатации - 2 м3 при ежедневном добавлении дезинфицированной морской воды с микроводорослями (Фиг. 4).
После достижения культуры моин в объеме массовой эксплуатации - 2 м3 производили полу-проточное культивирование моин при ежедневном сборе части популяции (около 0,25 объема) для кормления личинок (>150000 экз/сут) с ежедневной подменой данного объема свежей средой и внесением корма.
Перевод культуры моин в фазу полового размножения для получения эфиппиев. Наиболее эффективно индукция полового размножения производится при сочетании негативных стимулов, действующих синергетически на метаболизм моин. По завершении цикла наращивания биомассы партеногенетических самок моин для кормления личинок рыб производили индуцирование продуцирование эфиппиумов (путем сгущения моин до плотности 3-5 тыс./л, выдерживания при этой плотности около 2 суток при поддержании количества кормовых микроводорослей на уровне 1-2 мкг/л при снижении освещенности до 500 люкс и сокращении светового дня до 5-6 час (соотношение света/темноты 5-6/19-18 час и снижение в течение 2 суток температуры воды от 25 до 15°С). При переходе большинства особей моин от однополого к половому размножению производили ежедневный сбор продуцируемых ими эфиппиумов со дна эксплуатируемого бассейна, и затем производили полный слив бассейна, собирая эфиппиумы моин на газ, на котором их отмывали от детритных частиц и промывали дезинфицирующим раствором, постепенно понижая температуру до 10°С. При правильной манипуляции с культурой моин, можно получить биомассу эфиппиумов около 0,5 г/л. Полученные порции яиц эфиппиумов подготавливали к состоянию покоя - длительного хранения, концентрировали, высушивали и помещали в герметичные условия без доступа кислорода и света на холод при +3(±1)°С, где их сохраняли в консервированном виде в темноте в течение 6-24 месяцев.
Содержание более 12 часов (после рождения) новорожденных самок в несменяемой культуральной среде с накопленными метаболитами от взрослых самок действовало как стимул к смене типа размножения, то есть к прохождению мейоза в яйцеклетках. Миктические самки могут производить смешанное потомство в единовременной кладке (6-12 яиц) с варьируемым (от 0,3 до 4) соотношением самцов и самок. Самцы, помимо ряда морфологических особенностей, отличаются от самок меньшими терминальными размерами, после нескольких линек достигая длины карапакса 0,73±0,02 мм. Оплодотворение самцами самок происходит только в течение первых 12 часов после их вылупления. В результате оплодотворения самок происходит слияние гаплоидной яйцеклетки и сперматозоида, и восстанавливается диплоидность. Утолщенные хитиновые структуры эфиппиумов образуются из стенок раковины самок в области выводковой камеры, обволакивают оплодотворенные диапаузиирующие яйца, которые сбрасываются вместе с раковиной при линьке самки. Оплодотворенные самки (0,63 - 1 мм в длину) производят в одной кладке непарные (в одной из выводковых сумок) или парные эфиппии (в двух выводковых сумках) размером 0,35±0,03×0,48±0,05 мм, с 1-2 покоящимися (диапаузиирующими) яйцами (около 0,20±0,02 мм × 0,30±0,05 мм). За жизненный цикл миктическая самка может произвести до 6 кладок эфиппиев. Для производства последовательных кладок оплодотворенных покоящихся диплоидных яиц, заключенных в эфиппии, самкам необходимо повторно спариваться с самцами, так как у них отсутствуют приспособления для хранения спермы.
При условии перемещения свежеполученных (в течение первых суток после откладки) эфиппиев в оптимальные условия (чистую среду с добавлением микроводорослей при постоянном освещении, аэрации и температуре 24±2°С), через 19±7 часов из 90% оплодотворенных диплоидных яиц вылуплялись самки, которые давали начало новой партеногенетической популяции.
Сбор эфиппиумов производили сифоном со дна, их концентрировали, промывали и сохраняли в сухом, прохладном, темном месте в высушенном состоянии или в рапе до следующего цикла выращивания. При условии содержания эфиппиумов в несменяемой культуральной среде более суток, при повышении солености выше оптимальной, снижении температуры, высушивании и помещении в темноту для перехода в состояние длительной диапаузы с целью их длительного хранения (от нескольких месяцев до нескольких лет) в сухом темном месте (например, в эксикаторе с силикагелем) или в холодильнике при температуре ниже 4°С. Длительность воздействия факторов активации диапаузиирующих яиц увеличивается, а процент вылупления жизнеспособных особей снижается с увеличением срока хранения эфиппиев. После 4 лет хранения диапаузирующих яиц моин, защищенных неповрежденными структурами эфиппиумов, единовременный выклев жизнеспособных моин может снизиться до 10%.
Для перевода покоящихся эмбрионов моим в активное состояние и получения ювенильных партеногенетических самок моин за 24 - 48 ч до вылупления эфиппиумы помещали при плотности 300-500 эфиппиумов в 1 л в стерильную морскую воду соленостью 18%, промывали несколько раз, и постепенно (в течение 12 час) поднимали температуру воды до 25°С, одновременно повышая уровень кислорода до 85-100% насыщения и при постоянной освещенности на поверхности инкубатора не менее 1000 люкс. Диплоидные самки, вылупившиеся из эфиппиумов, при перемещении их в благоприятные условия среды, свежую воду с высоким содержанием кислорода, оптимальную температуру и освещенность, при питании смесью микроводорослей ad libitum, могут продуцировать в первой кладке 17±1 яиц со 100% выживаемостью ювенильных партеногенетических самок.
Культивируемые автором изобретения моины были впервые применены в качестве живых кормов при искусственном выращивании личинок черноморского калкана (Khanaychenko, 1990; Bityukova et al., 1990; Khanaychenko et al., 1993; Khanaychenko et al., 1994; Ханайченко, 1996), и в дальнейшем использованы неоднократно. Питание моинами личинок черноморского калкана в возрасте от 20 до 45 сут способствует более быстрому достижению ими большей массы тела к началу перехода на питание инертными кормами, и, соответственно, повышает скорость их роста и улучшает их выживаемость. Размеры моин, как то длина карапакса от 400 мкм у ювенильных особей и до 1250 мкм у половозрелых особей, их масса около 0,25 мг, и при оптимальном культивировании их химический состав с содержанием белка не менее 50% и незаменимой, эссенциальной для роста аминокислоты метионина около 1,5% (Qian et al., 2016) полностью удовлетворяют потребностям личинок в течение периода метаморфоза.
Согласно исследованиям автора изобретения, максимальный часовой рацион во время пика активности калкана ~ 1 см составил 100 экз. моин (25 мг по сырой массе) по сравнению с 205 экз. акарций (9,9 мг) и 1200 экз (15,6 мг) метанауплиев артемий. Личинки калкана отлавливали быстрее всего (или в первую очередь) самый крупный и наименее подвижный пищевой объект из смеси кормов - моин. Визуальный контроль за перевариванинм метанауплиев артемий, акарций и моин личинками калкана показал различия не только в скорости потребления разных организмов, но и в скорости прохождения через пищеварительный тракт и, соответственно, степени переваривания. Моины были полностью переварены, и после них обнаруживались только пустые хитиновые оболочки. Напротив, артемий плохо переваривались личинками калкана. Было установлено, что съеденные моины задерживались в отдельных участках пищеварительного тракта более длительный период времени, в результате чего и происходило их более полноценное переваривание, в то время как артемий быстро проходили через пищеварительный тракт личинок недостаточно переваренными, что свидетельствует о том, что ферментативная активность личинок не соответствует жировому составу артемий.
Источники информации, принятые во внимание
1. Семик A.M., Ушакова А.О. 2017. Особенности развития зоопланктонного сообщества соленого Акташского озера. Труды Южного научно-исследовательского института рыбного хозяйства и океанографии. 2017; 54(1):123-30.
2. Пат. 2614644 Российская Федерация. МПК A01K 61/00 Способ интенсивного когортного культивирования акарций (морских каланоидных копепод). / Ханайченко А.Н.; заявитель и патентообладатель Федеральное государственное бюджетное учреждение науки «Институт морских биологических исследований имени А.О. Ковалевского РАН» - №2015147611; заявл. 05.11.2015; опубл. 28.03.2017, Бюл. №10.
3. Пат. 2670159 Российская Федерация. МПК A01K 61/00. Способ длительного хранения яиц каланоидных копепод акарций для получения синхронной культуры одновозрастных науплиев [Текст] / Ханайченко А.Н. (RU); заявитель и патентообладатель Федеральное государственное бюджетное учреждение науки «Институт морских биологических исследований имени А.О. Ковалевского РАН» - №2017142059; заявл. 01.12.2017; опубл. 18.10.2018 Бюл. №29.
4. Egloff D. Α., Fofonoff P. W., & Onbe, T. 1997. Reproductive biology of marine cladocerans. Advances in marine biology, 31, 79-167; Katechakis, Α., & Stibor, H. (2004). Feeding selectivities of the marine cladocerans Penilia avirostris, Podon intermedius and Evadne nordmanni. Marine Biology, 145(3), 529-539
5. He ZH, Qin JG, Wang Y, Jiang H, Wen Z. Biology of Moina mongolica (Moinidae, Cladocera) and perspective as live food for marine fish larvae. Hydrobiologia. 2001 Aug;457(1):25-37.
6. Hebert, P. D. N., 1978. The population biology of Daphnia (Crustacea, Daphnidae). Biol. Rev. 53: 387-426.
7. Qian J, Li H, Yang Z, Li Y, Wang X. Effects of different algae on survival and reproduction of Moina mongolica. Jiangsu Agricultural Sciences. 2016;44(7):477-80.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ интенсивного когортного культивирования акарций (морских каланоидных копепод) | 2015 |
|
RU2614644C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ МОРСКИХ ЦИКЛОПОИДНЫХ КОПЕПОД OITHONA DAVISAE | 2022 |
|
RU2788532C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ КАЛАНОИДНЫХ КОПЕПОД CALANUS EUXINUS (ЧЕРНОМОРСКОГО КАЛЯНУСА) | 2014 |
|
RU2541458C1 |
| СПОСОБ ДЛИТЕЛЬНОГО ХРАНЕНИЯ ЯИЦ КАЛАНОИДНЫХ КОПЕПОД АКАРЦИЙ ДЛЯ ПОЛУЧЕНИЯ СИНХРОННОЙ КУЛЬТУРЫ ОДНОВОЗРАСТНЫХ НАУПЛИЕВ | 2017 |
|
RU2670159C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ МОРСКИХ ГЕТЕРОТРОФНЫХ ДИНОФЛАГЕЛЛЯТ OXYRRHIS MARINA | 2023 |
|
RU2810308C1 |
| Способ получения живых кормов для личинок морских рыб | 2019 |
|
RU2717990C1 |
| СПОСОБ ИНТЕНСИВНОГО ВЫРАЩИВАНИЯ МАЛЬКОВ КАМБАЛЫ КАЛКАН | 2014 |
|
RU2548106C1 |
| СПОСОБ ИСКУССТВЕННОГО ПОЛУЧЕНИЯ МОЛОДИ КАМБАЛЫ КАЛКАН | 1992 |
|
RU2017413C1 |
| Способ снижения численности бактерий-оппортунистов в средах выращивания личинок морских рыб и их кормов | 2015 |
|
RU2614604C1 |
| СПОСОБ ВЫРАЩИВАНИЯ ГИГАНТСКОЙ УСТРИЦЫ CRASSOSTREA GIGAS В ЧЕРНОМ МОРЕ | 2014 |
|
RU2541459C1 |
Способ относится к области морской аквакультуры и предназначен для получения массы высококачественных живых кормовых организмов для использования в качестве живых кормов при выращивании личинок ценных морских рыб и для проведения экспериментальных работ в области научных исследований по биологии, эмбриологии, генетике, физиологии, биохимии и биологического тестирования. Изъятых из естественной зоопланктонной пробы самок моин выдерживают в дезинфекционном растворе с добавлением микроводорослей в течение 6-8 часов, помещают в контейнеры со стерильной морской водой при минимальном объеме 30 мл на самку, и постоянной освещенностью 5000 лк, осуществляют кормление микроводорослями. Затем полученное потомство используют для накопительного культивирования при партеногенетической стадии размножения, которое проводят при температуре 24±2°С, освещенности на поверхности культиватора 5000 лк, фотопериоде не менее 16 часов освещения и 8 часов темноты, при последовательном наращивании плотности самок до 300 экз./л, в несменяемой среде, с ежедневным внесением микроводорослей. После чего при поддержании постоянной плотности самок около 300 экз/л проводят наращивание объема культуры, при ежедневном добавлении стерильной морской воды с микроводорослями, после чего проводят культивирование моин с ежедневным изъятием около 0,25 объема культуры, заменой данного объема стерильной морской водой и внесением микроводорослей, в качестве которых используют Isochrysis galbana, Tetraselmis suecica и Rhodomonas sp., поддерживая их концентрацию во время партеногенетической стадии размножения от 3 до 10 мкг/мл. По завершении наращивания биомассы партеногенетических самок моин переводят в стадию полового размножения, увеличивая плотность культуры до 3000-5000 экз/л и выдерживая при этой плотности 48 часов, снижая за это время температуру воды с 25°С до 15°С, при освещенности на поверхности культиватора 500 лк, фотопериоде 5-6 часов освещения и 18-19 часов темноты и концентрации микроводорослей 1-2 мкг/л. Полученные в результате полового размножения эфиппиумы высушивают и хранят в течение 6-24 месяцев. Для перевода покоящихся эмбрионов моин в активное состояние и получения ювенильных партеногенетических самок моин эфиппиумы помещают при плотности около 300 экз/л на 24-48 часов в стерильную холодную морскую воду соленостью 18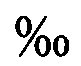 , промывают несколько раз и постепенно в течение 12 часов поднимают температуру воды до 25°С, одновременно повышая уровень кислорода до 85-100% насыщения, при постоянной освещенности на поверхности инкубатора не менее 1000 лк. Изобретение обеспечивает получение массовых высококачественных живых кормов однородного состава с высоким содержанием белка и незаменимых аминокислот. 4 ил., 1 пр.
, промывают несколько раз и постепенно в течение 12 часов поднимают температуру воды до 25°С, одновременно повышая уровень кислорода до 85-100% насыщения, при постоянной освещенности на поверхности инкубатора не менее 1000 лк. Изобретение обеспечивает получение массовых высококачественных живых кормов однородного состава с высоким содержанием белка и незаменимых аминокислот. 4 ил., 1 пр.
Способ культивирования моин для использования в качестве живых кормов в аквакультуре, характеризующийся тем, что изъятых из естественной зоопланктонной пробы самок моин выдерживают в дезинфекционном растворе с добавлением микроводорослей в течение 6-8 часов, помещают в контейнеры со стерильной морской водой при минимальном объеме 30 мл на самку, и постоянной освещенностью 5000 лк, осуществляют кормление микроводорослями, полученное потомство используют для накопительного культивирования при партеногенетической стадии размножения, которое проводят при температуре воды 24±2°С, освещенности на поверхности культиватора 5000 лк, фотопериоде не менее 16 часов освещения и 8 часов темноты, при последовательном наращивании плотности самок до 300 экз./л, в несменяемой среде, с ежедневным внесением микроводорослей, затем, при поддержании постоянной плотности самок около 300 экз/л, проводят наращивание объема культуры, при ежедневном добавлении стерильной морской воды с микроводорослями, после чего проводят культивирование моин с ежедневным изъятием около 0,25 объема культуры, заменой данного объема стерильной морской водой и внесением микроводорослей, в качестве которых используют Isochrysis galbana, Tetraselmis suecica и Rhodomonas sp., поддерживая их концентрацию во время партеногенетической стадии размножения от 3 до 10 мкг/мл, по завершении наращивания биомассы партеногенетических самок моин переводят в стадию полового размножения, увеличивая плотность культуры до 3000-5000 экз/л и выдерживая при этой плотности 48 часов, снижая за это время температуру воды с 25°С до 15°С, при освещенности на поверхности культиватора 500 лк, фотопериоде 5-6 часов освещения и 18-19 часов темноты и концентрации микроводорослей 1-2 мкг/л, полученные в результате полового размножения эфиппиумы высушивают и хранят в течение 6-24 месяцев, для перевода покоящихся эмбрионов моин в активное состояние и получения ювенильных партеногенетических самок моин, эфиппиумы помещают при плотности около 300 экз/л на 24-48 часов в стерильную холодную морскую воду соленостью 18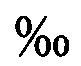 , промывают несколько раз и постепенно в течение 12 часов поднимают температуру воды до 25°С, одновременно повышая уровень кислорода до 85-100% насыщения, при постоянной освещенности на поверхности инкубатора не менее 1000 лк.
, промывают несколько раз и постепенно в течение 12 часов поднимают температуру воды до 25°С, одновременно повышая уровень кислорода до 85-100% насыщения, при постоянной освещенности на поверхности инкубатора не менее 1000 лк.
| Способ интенсивного когортного культивирования акарций (морских каланоидных копепод) | 2015 |
|
RU2614644C1 |
| Способ получения живых кормов для личинок морских рыб | 2019 |
|
RU2717990C1 |
| Рентгеновский измеритель толщины проката | 1988 |
|
SU1534307A1 |

