Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности, к микробиологическому синтезу L-треонина с использованием бактерии вида Escherichia coli.
Уровень техники
Традиционно L-аминокислоты в промышленном масштабе могут быть получены методом ферментации с использованием штаммов микроорганизмов, выделенных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (US 4278765). Указанные методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот и/или уменьшении чувствительности целевого фермента к обратному ингибированию продуцируемой L-аминокислотой (WO 9516042, US 5661012, US 6040160).
Известны различные штаммы, использующиеся для производства L-треонина методом ферментации. Это штаммы с увеличенными активностями ферментов, вовлеченных в биосинтез L-треонина (US 5175107; US 5661012; US 5705371; US 5939307; ЕР 219027), штаммы, устойчивые к некоторым химических реагентам, таким как L-треонин и его аналоги (WO 0114525, ЕР 301572, US 5376538), штаммы с инактивированными ферментами системы деградации треонина (US 5939307 и US 6297031), штаммы, в которых устранена чувствительность целевого фермента к ингибированию продуцируемой аминокислотой или ее побочными продуктами по типу обратной связи (US 5175107 и US 5661012).
Известен штамм-продуцент L-треонина Е. coli ВКПМ В-3996 (SU 1694643, US 5175107 и US 5705371), полученный путем введения в штамм Е. coli ВКПМ В-7 следующих мутаций и плазмиды:
- мутантный ген thrA (мутация thrA442) кодирует белок аспартокиназа-гомосериндегидрогеназу I, который устойчив к ингибированию треонином по типу обратной связи;
- мутантный ген ilvA (мутация ilvA442) кодирует белок треониндеаминазу, обладающую пониженной активностью, которая выражается в пониженном уровне биосинтеза изолейцина и в фенотипе с недостатком по изолейцину типа "leaky". В бактерии с мутацией ilvA442 транскрипция оперона thrABC не репрессируется изолейцином, что дает положительный эффект на продукцию L-треонина;
- инактивация гена tdh приводит к предотвращению деградации L-треонина, в указанный штамм была введена генетическая детерминанта ассимиляции сахарозы (гены scrKYABR);
- увеличение экспрессии генов, контролирующих биосинтез L-треонина, достигнута путем введения в штамм плазмиды pVIC40, содержащей мутантный треониновый оперон thrA442BC.
Штамм Е. coli ВКПМ В-3996 в ходе ферментации пробирках продуцирует 13 г/л L-треонина (RU 2182173), при культивировании в ферментере - 85 г/л L-треонина (US 5175107).
Технической проблемой, на решение которой направлено заявляемое изобретение является поиск новых генов-мишеней, модификация которых приводит к увеличению продукции L-треонина.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является расширение арсенала штаммов Escherichia coli, способных к продукции L-треонина.
Он достигается тем, что получен штамм бактерии Escherichia coli ВКПМ В-14269 с инактивированным геном yqeG, обладающий способностью продуцировать L-треонин.
Краткое описание чертежей
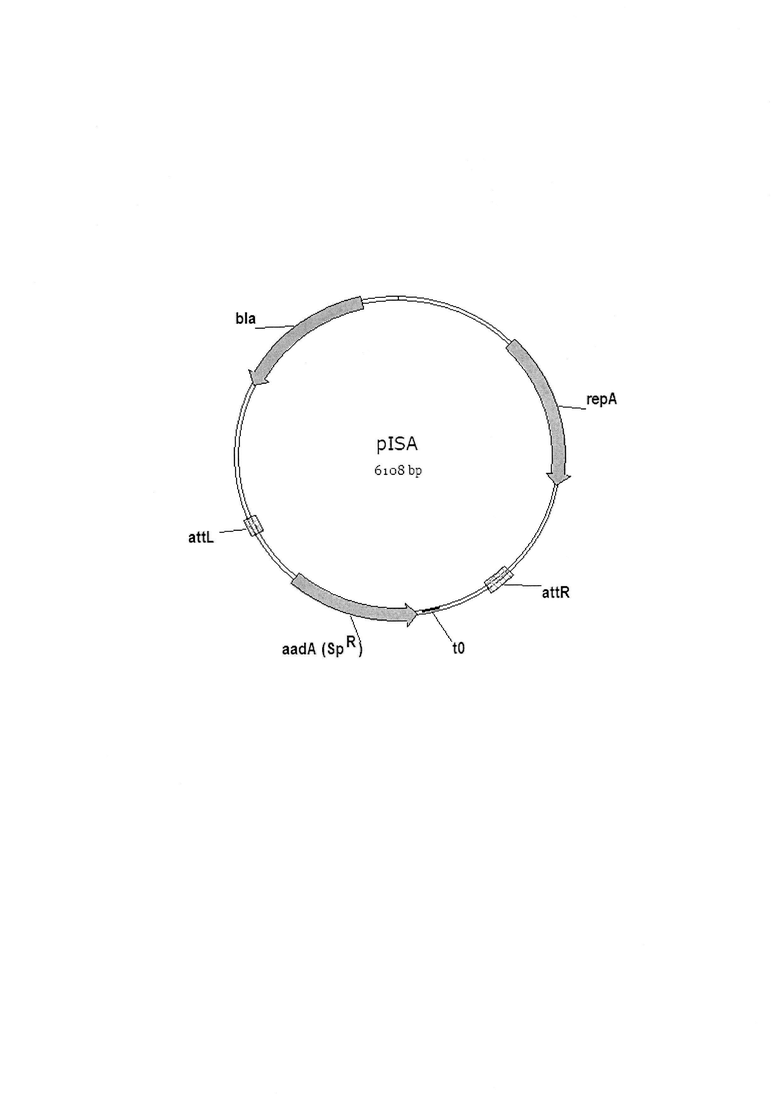
На фигуре показана схема плазмиды pISA.
Осуществление изобретения
Термин "ген yqeG инактивирован" означает, что целевой ген модифицирован таким образом, что такой модифицированный ген кодирует мутантный белок со сниженной активностью, или полностью неактивный белок. Также возможно, что естественная экспрессия модифицированного участка ДНК невозможна из-за делеции целевого гена или его части, сдвига рамки считывания данного гена или введения missense/nonsense мутации или модификации прилегающей к гену областей, которые включают последовательности, контролирующие экспрессию гена, такие как промотор(ы), энхансер(ы), аттенуатор(ы), сайт(ы) связывания рибосомы, и т.д.
Инактивация гена может быть осуществлена любым возможным методом, например, такими как мутагенез с использованием УФ излучения или обработка нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, инактивации гена с помощью гомологичной рекомбинации, или/и инсерционно-делеционного мутагенеза (Yu, D. et al., Proc. Natl. Acad. Sci. USA, 2000, 97: 12: 5978-83); (Datsenko K.A. and Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97: 12: 6640-45), также называемого "Red-зависимая интеграция".
В данной работе инактивацию гена yqeG проводят путем замены открытой рамки считывания гена на селективный маркер aadA, ген устойчивости к спекциномицину. Полученный таким образом штамм не способен синтезировать белок YqeG, кодируемый геном yqeG.
Пример 1. Конструирование штамма с инактивированным геном yqeG
Инактивацию гена осуществляют в штамме ВКПМ В-13207, который получен на основе дикого штамма Escherichia coli K-12 MG1655 (АТСС 47076) в результате проведения нескольких раундов мутагеза различными мутирующими агентами с целью отбора мутантов наиболее устойчивых к L-треонину, и последующего введения направленных генетических модификаций: замена промотора генов треонинового оперона, введение десенсибилизирующей мутации в ген thrA, оверэспрессия rhtA, инактивация гена sstT, инактивация гена рохВ, кодирующего пируватоксидазу.
Делецию гена yqeG осуществляют путем замены его ORF (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 2,985,847 -> 2,987,076), на селективный маркер устойчивости к спектиномицину, методом ПЦР с использованием следующих олигонуклеотидов:
yqeGCinsF 5'-atgtctaatatttggtctaaagaagaaactctgtggagtttcgcgctctagccaagcttgcatgcctgc
yqeGinsR 5'-ttagctaaagaacattaccgatacgcacaggatacccacgatcagcgtaagggatcctctagagtcgag
В качестве матрицы для синтеза используют плазмиду pISA (Биотехнология 2019 Т. 35 №4 с. 42-54). В своем составе эта плазмида (фиг. 1) несет следующие генетические элементы: ген aadA, обуславливающий устойчивость клеток к спектиномицину для прямого отбора трансформантов; последовательности фага λ attL и attR - сайты узнавания системы рекомбинации Int/Xis; repA - репликон; ген bla, кодирующий бета-лактамазу, обуславливающий устойчивость к ампициллину.
Амплификацию фрагмента проводят с использованием полимеразы KAPA (KAPAbiosystems) для того, чтобы избежать ошибок в последовательности. Используют следующий температурный профиль для ПНР: денатурация при 95°С в течение 3 мин; 25 циклов: 20 сек при 98°С, 20 сек при 60°С, 2 мин при 72°С; и заключительная полимеризация: 2 мин при 72°С. ПЦР продукт размером 2037 п.о. очищают методом экстракции из агарозного геля и далее используют для трансформации штамма ВКПМ В-13207, который предварительно трансформируют плазмидой pKD46, которая необходима для интеграции продукта ПЦР в хромосому штамма. Плазмида pKD46 (Datsenko, K.А. and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 2000, 97: 12: 6640-45) содержит фрагмент ДНК фага λ (инвентарный номер последовательности J02459 в базе данных GenBank) длиной 2,154 нуклеотида (31088-33241), а также содержит гены Red гомологичной системы рекомбинации (β, γ, ехо гены) под контролем промотора ParaB; индуцируемого арабинозой.
Электрокомпетентные клетки получают следующим образом: ночную культуру штамма Е. coli ВКПМ В-13207 выращивают при 30°С в жидкой среде LB (мас. %): триптон - 1, NaCl - 1, дрожжевой экстракт - 0,5, вода -остальное, рН 7,0 с добавкой ампициллина (125 мг/л), разводят в 100 раз при помощи 5 мл среды SOB (мас. %): триптон 2, дрожжевой экстракт - 0,5, MgCl2 - 0,0956, MgSO4⋅7H2O - 0,252, NaCl - 0,0058, KCl - 0,0185, вода остальное с добавлением ампициллина (100 мг/л) и L-арабинозы (1 мМ). Полученную культуру растят с перемешиванием при 30°С до достижения оптической плотности ОП660нм 0,6 ед., после чего делают клетки электрокомпетентными путем концентрации в 100 раз и трехкратного отмывания ледяной деионизированной Н2О. Электропорацию проводят с использованием 40 мкл клеток и 1 мкг продукта ПЦР. После электропорации клетки инкубируют в 1 мл среды SOC (мас. %): триптон 2, дрожжевой экстракт - 0,5, глюкоза - 0,36, MgCl2 - 0,0956, MgSO4⋅7H2O - 0,252, NaCl - 0,0058, KCl - 0,0185, вода - остальное при 37°С в течение 2,5 часов, после чего высевают на чашки со средой LA (мас. %): агар-агар - 2, триптон - 1, NaCl - 1, дрожжевой экстракт - 0,5, вода - остальное, рН 7,0 с добавлением спектиномицина (75 мкг/мл).
Отбор трансформантов, несущих делецию в гене yqeG, проводят на чашках со средой LA с добавлением спектиномицина. Наличие инсерции в локусе yqeG у отобранных клонов подтверждают методом ПЦР. Для амплификации фрагмента используют полимеразу Taq (Thermo Fisher Scientific) и следующие олигонуклеотиды:
yqeGoutF 5'-cctggtttatggtaaattgcc
yqeGoutR 5'-acgagagaagtgatagggtg
Для проведения ПЦР используют следующий температурный профиль: денатурация при 94°С в течение 4 мин; 25 циклов: 20 сек при 94°С, 20 сек при 55°С, 2 мин 0 сек при 72°С; и заключительная полимеризация: 5 мин при 72°С.
Наличие продукта размером 2192 п.о. интерпретируют как интеграцию кассеты в целевой локус. Для удаления вспомогательной плазмиды pKD46, проводят 2 пассажа на среде LA со спектиномицином при 42°С и полученные колонии тестируют на чувствительность к ампициллину.
В результате отобрано 5 клонов, несущих делецию в гене yqeG.
Пример 2. Отбор наиболее продуктивного клона
Для проверки влияния делеции гена yqeG на продукцию L-треонина проводят пробирочную ферментацию пяти отобранных клонов, в качестве контрольного штамма используют родительский штамм ВКПМ В-13207.
Штаммы выращивают на чашках со средой LA в течение 24 часов при 37°С. Для приготовления инокулята используют посевную среду следующего состава (мас. %)
В пробирки объемом 50 мл с рабочим объемом 5 мл посевной среды вносят биомассу клеток до стартового значения оптической плотности равной ОП660нм 0,1 ед. Пробирки инкубируют на роторной качалке в течение 5 часов при 37°С и скорости перемешивания 220 об/мин. Для основного процесса ферментации используют среду следующего состава (мас. %):
Растворы глюкозы, сульфата марганца и сульфата магния стерилизуют отдельно автоклавированием при 121°С в течение 40 мин. Навески СаСО3 по 40 мг стерилизуют в стеклянных пробирках автоклавированием при 121°С в течение 20 мин. рН доводят до значения 7,0. Раствор сульфата железа стерилизуют фильтрованием через мембрану диаметром пор 22 мкм.
Полученный инокулят вносят в пробирки объемом 50 мл с рабочим объемом 2 мл ферментационной среды до стартовой оптической плотности ОП660 нм 0,1 ед. Клетки культивируют в течение 24 часов при 37°С на роторной качалке - 220 об/мин.
После выращивания количество накопленного в среде L-треонина определяют с методом ВЭЖХ (Waters 2695, Alliance). Результаты ферментации пяти клонов приведены в табл. 1.
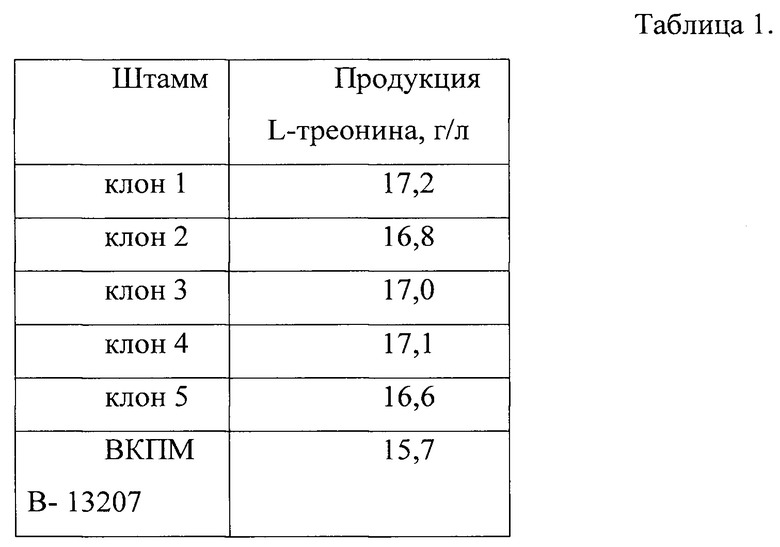
Как видно из табл. 1, клон 1, содержащий делецию гена yqeG, накапливает большее количество L-треонина по сравнению с контрольным штаммом ВКПМ В-13207. Отобранный клон 1 депонирован в Биоресурсном Центре Всероссийская Коллекция Промышленных Микроорганизмов (БРЦ ВКПМ) НИЦ «Курчатовский институт» (117545 Москва, 1-ый Дорожный пр-д, д. 1) как штамм Escherichia coli ВКПМ В-14269.
Заявляемый штамм Escherichia coli ВКПМ В-14269 характеризуется следующими признаками:
Культурально-морфологические характеристики
Грамм-отрицательная бактерия. Суточная культура в жидкой LB представлена слабо подвижными клетками округлой формы 1 мкм в диаметре. При культивировании на L-агаре в течение 18-24 часов при 37°С образует круглые, беловатый, полупрозрачные на свет колонии колонии 1-2 мм, поверхность колонии гладкая, края ровные или слегка волнистые, центр колоний приподнят, структура однородная, консистенция пастообразная, легко эмульгируется. При культивировании на агаризованной среде Эндо (мас. %: пептон - 1, лактоза - 1, Na2SO3 - 0,33, K2HPO4 - 0,25, фуксин основной - 0,03, агар-агар -2%, вода - остальное) при 37°С образуются колонии, круглой формы с ровным четко очерченным краем малиново-красного цвета с металлическим блеском. Диаметр колоний 0,5 - 1,5 мм.
Физиолого-биохимические характеристики.
Факультативный анаэроб. Сахара не сбраживает. Ассимилирует: D-глюкозу, L-арабинозу, D-маннитол, D-ксилозу, в меньшей мере: D-галактозу, D-лактозу. Отсутствует способность к гидролизу крахмала. Отсутствует потребность в факторах роста. Штамм устойчив к хлорамфениколу. Оптимальное значение рН для роста 7,2-7,4. Не растет при температурах свыше 45°С. Оптимальная температура роста 37°С. Штамм не патогенен.
Таким образом, получен штамм Escherichia coli ВКПМ В-14269 с инактивированным геном yqeG, способный синтезировать до 17,2 г/л L-треонина при культивировании в пробирках.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм Escherichia coli с инактивированным геном lysP - продуцент L-треонина | 2020 |
|
RU2758269C1 |
| Штамм Escherichia coli с инактивированным геном sdaC - продуцент L-треонина | 2021 |
|
RU2774071C1 |
| Штамм Escherichia coli с инактивированным геном ydgI - продуцент L-треонина | 2020 |
|
RU2748676C1 |
| Штамм Escherichia coli с инактивированным геном yjeM - продуцент L-треонина | 2021 |
|
RU2775206C1 |
| Штамм Escherichia coli с инактивированным геном ttdT - продуцент L-треонина | 2019 |
|
RU2731282C1 |
| Штамм Escherichia coli с инактивированным геном ychE - продуцент L-треонина | 2019 |
|
RU2728251C1 |
| Штамм Escherichia coli с инактивированным геном yhjE - продуцент L-треонина | 2022 |
|
RU2787585C1 |
| Бактерия вида Escherichia coli - продуцент L-треонина, способ микробиологического синтеза L-треонина с ее использованием. | 2018 |
|
RU2697499C1 |
| Штамм Escherichia coli - продуцент L-треонина | 2019 |
|
RU2728242C1 |
| БАКТЕРИЯ РОДА ESCHERICHIA - ПРОДУЦЕНТ L-ТРЕОНИНА И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА L-ТРЕОНИНА С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2515095C1 |
Настоящее изобретение относится к микробиологии. Описан штамм бактерии Escherichia coli ВКПМ В-14269 с инактивированным геном yqeG, обладающий способностью продуцировать L-треонин. Техническим результатом является расширение арсенала штаммов Escherichia coli, способных к продукции L-треонина. 1 ил., 1 табл.
Штамм Escherichia coli ВКПМ В-14269 с инактивированным геном yqeG - продуцент L-треонина.
| Штамм Escherichia coli с инактивированным геном ttdT - продуцент L-треонина | 2019 |
|
RU2731282C1 |
| Штамм Escherichia coli с инактивированным геном ychE - продуцент L-треонина | 2019 |
|
RU2728251C1 |
| Штамм Escherichia coli с инактивированным геном ydgI - продуцент L-треонина | 2020 |
|
RU2748676C1 |

