Перекрестная ссылка на родственные изобретения
Данная заявка притязает на приоритет корейской патентной заявки №10-2021-0011010, поданной 26 января 2021 г., полное раскрытие которой включено в настоящий документ как часть настоящего раскрытия.
Настоящее раскрытие относится к новому варианту 5,10-метилентетрагидрофолатредуктазы, штамму Corynebacterium glutamicum, содержащему данный вариант, и к способу продуцирования L-валина с использованием данного штамма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Проводят различные исследования для разработки высокоэффективных микроорганизмов и технологий способов ферментации для производства L-аминокислот и других полезных веществ. Например, главным образом, используют специфичный подход в отношении целевого вещества, при котором увеличивают экспрессию гена, кодирующего фермент, участвующий в биосинтезе L-валина, или при котором удаляют гены, не являющиеся необходимыми для биосинтеза (US 8465962 В2).
Однако все еще необходимо проводить исследования для эффективного увеличения способности к продуцированию L-валина, так как возрастает спрос на L-валин.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Целью настоящего раскрытия является предоставление варианта 5,10-метилентетрагидрофолатредуктазы, состоящего из аминокислотной
последовательности, представленной SEQ ID NO: 1, в которой аргинин (Arg, R), который представляет собой аминокислоту, соответствующую положению 4 аминокислотной последовательности SEQ ID NO: 3, заменен гистидином (His, Н).
Другой целью настоящего раскрытия является предоставление полинуклеотида, кодирующего вариант по настоящему раскрытию.
Еще одной другой целью настоящего раскрытия является предоставление штамма Corynebacterium glutamicum, который содержит вариант по настоящему раскрытию, или полинуклеотид, кодирующий данный вариант, и имеет способность к продуцированию L-валина.
Еще одной другой целью настоящего раскрытия является предоставление способа продуцирования L-валина, который включает культивирование в среде штамма Corynebacterium glutamicum, который содержит вариант по настоящему раскрытию или полинуклеотид, кодирующий данный вариант, и имеет способность к продуцированию L-валина.
Наилучший способ воплощения изобретения
Настоящее раскрытие будет подробно описано следующим образом. Тем временем, каждое из описаний и воплощений, раскрытых в настоящем раскрытии, можно применять к другим описаниям и воплощениям. Другими словами, все комбинации разных элементов, раскрытых в настоящем раскрытии, принадлежат к объему настоящего раскрытия. Кроме того, нельзя считать, что объем настоящего раскрытия ограничивается конкретным описанием, приведенным ниже. Кроме того, во всем настоящем описании изобретения приводятся ссылки целого ряда статей и патентных документов, и указываются их цитирования. Вся полнота содержания, раскрытого в процитированных статьях и патентных документах, включается в настоящее описание изобретения посредством ссылки для того, чтобы более ясно описать уровень технической области, к которой принадлежит настоящее изобретение, и содержание настоящего изобретения.
Согласно одному аспекту настоящего раскрытия предложен вариант 5,10-метилентетрагидрофолатредуктазы, состоящий из аминокислотной
последовательности, представленной SEQ ID NO: 1, в которой аргинин (Arg, R), который представляет собой аминокислоту, соответствующую положению 4 аминокислотной последовательности SEQ ID NO: 3, заменен гистидином (His, Н).
Вариант по настоящему раскрытию может иметь, содержать или по существу состоять из аминокислотной последовательности, представленной SEQ ID NO: 1.
В варианте по настоящему раскрытию аминокислота, соответствующая положению 4 на основе аминокислотной последовательности SEQ ID NO: 3 в аминокислотной последовательности, представленной SEQ ID NO: 1, представляет собой гистидин, и данный вариант может содержать аминокислотную последовательность, имеющую по меньшей мере 70%-ную, 75%-ную, 80%-ную, 85%-ную, 90%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную, 99%-ную, 99,5%-ную, 99,7%-ную или 99,9%-ную или большую гомологию или идентичность с аминокислотной последовательностью, представленной SEQ ID NO: 1. Очевидно то, что варианты, имеющие аминокислотные последовательности, в которых некоторые последовательности удалены, модифицированы, заменены, консервативно заменены или добавлены, также включают в объем настоящего раскрытия, при условии, что аминокислотные последовательности имеют такую гомологию или идентичность и демонстрируют эффективность, соответствующую эффективности варианта по настоящему раскрытию.
Его примеры включают варианты, имеющие присоединение или делецию последовательности, которые не изменяют функцию варианта по настоящему раскрытию, на N-конце, С-конце аминокислотной последовательности и/или внутри данной аминокислотной последовательности, встречающуюся в природе мутацию, молчащую мутацию или консервативную замену.
Термин «консервативная замена» означает замену одной аминокислоты другой аминокислотой, имеющей аналогичные структурные и/или химические свойства. Такая аминокислотная замена обычно может существовать на основе сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природе остатков. Обычно консервативная замена может слегка влиять или не влияет на активность белков или полипептидов.
В настоящем раскрытии термин «вариант» относится к полипептиду, который имеет аминокислотную последовательность, отличную от аминокислотной последовательности данного варианта перед модификацией посредством консервативной замены и/или модификации одной или более чем одной аминокислоты, но сохраняет функции или свойства. Такой вариант обычно может быть идентифицирован посредством модифицирования одной или более чем одной аминокислоты аминокислотной последовательности данного полипептида и осуществления оценки свойств данного модифицированного полипептида. Другими словами, способность данного варианта может быть увеличена, оставлена неизменной или снижена по сравнению со способностью полипептида перед изменением. Некоторые варианты могут включать варианты, в которых одна или более чем одна часть, такая как N-концевая лидерная последовательность или трансмембранный домен, были удалены. Другие варианты могут включать варианты, в которых была удалена часть N- и/или С-конца от зрелого белка. Термин «вариант» можно использовать взаимозаменяемо с такими терминами, как модификация, модифицированный полипептид, модифицированный белок, мутант, мутеин и дивергент, и не ограничивается ими, при условии, что он представляет собой термин, используемый со значением вариации. В целях настоящего раскрытия данный вариант может представлять собой полипептид, содержащий аминокислотную последовательность, представленную SEQ ID NO: 1, в которой аргинин, который представляет собой аминокислоту, соответствующую положению 4 аминокислотной последовательности SEQ ID NO: 3, заменен гистидином.
Данный вариант может содержать делеции или присоединения аминокислот, которые имеют минимальное влияние на свойства и вторичную структуру полипептида. Например, с N-концом данного варианта может быть конъюгирована сигнальная (или лидерная) последовательность, которая котрансляционно или посттрансляционно участвует в транслокации белка. Данный вариант может быть конъюгирован с другими последовательностями или линкерами таким образом, чтобы его идентифицировать, очистить или синтезировать.
В настоящем раскрытии термин «гомология» или «идентичность» означает степень сходства между двумя данными аминокислотными последовательностями или последовательностями оснований и может быть выражена в виде процентной доли. Термины «гомология» и «идентичность» часто можно использовать взаимозаменяемо.
Гомологию или идентичность последовательности консервативного полинуклеотид а или полипептида определяют стандартными алгоритмами выравнивания, и можно совместно использовать штраф за пропуск по умолчанию, установленный применяемой программой. По существу гомологичные или идентичные последовательности обычно способны к гибридизации со всей или с частью последовательности при условиях умеренной или высокой жесткости. Очевидно, что гибридизация также включает гибридизацию полинуклеотид а с полинуклеотидом, содержащим кодон общего типа или вырожденный кодон.
Имеют ли любые две последовательности полинуклеотида или полипептида гомологию, сходство или идентичность, можно определять с использованием известных компьютерных алгоритмов, таких как программа «FASTA», например, с использованием параметров по умолчанию как в Pearson et al., (1988) Proc. Natl. Acad. Sci. USA 85:2444. В качестве альтернативы, гомология, сходство или идентичность могут быть определены с использованием алгоритма Нидлмана Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), как осуществляется в программе Нидлмана пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277) (версия 5.0.0 или более поздняя) (включая программный пакет GCG (Devereux, J., et al., Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S.F., et al., J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994; и CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, для определения гомологии, сходства или идентичности можно использовать BLAST Национального центра биотехнологической информации или ClustalW.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определять посредством сравнения информации по последовательности с использованием, например, компьютерной программы GAP, как, например, Needleman et al. (1970), J Mol Biol. 48:443, как анонсировано, например, в Smith and Waterman, Adv. Appl. Math (1981) 2:482. В заключение, результат программы GAP может быть определен как значение, полученное делением числа аналогичных выровненных символов (а именно: нуклеотидов или аминокислот) на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию программы GAP могут включать (1) матрицу двоичных сравнений (включающую значения 1 для идентичности и 0 для неидентичности) и матрицу взвешенных сравнений Gribskov et al., (1986) Nucl. Acids Res. 14:6745 (или матрицу замен EDNAFULL (EMBOSS версии NCBI NUC4.4)), как раскрыто в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979); (2) штраф 3,0 для каждого пропуска и дополнительный штраф 0,10 для каждого символа в каждом пропуске (или штраф 10 за открытие пропуска, штраф 0,5 за удлинение пропуска); и (3) отсутствие штрафа за концевые пропуски.
В качестве примера по настоящему раскрытию, вариант по настоящему раскрытию может демонстрировать активность 5,10-метилентетрагидрофолатредуктазы. Вариант по настоящему раскрытию может демонстрировать активность таким образом, чтобы иметь повышенную способность к образованию L-валина по сравнению с полипептидом дикого типа.
В настоящем раскрытии термин «5,10-метилентетрагидрофолатредуктаза» представляет собой полипептид, который катализирует превращение 5,10-метилентетрагидрофолата в 5-метилентетрагидрофолат. Термин «5,10-метилентетрагидрофолатредуктаза» по настоящему раскрытию может использоваться взаимозаменяемо с MetF. В настоящем раскрытии последовательность 5,10-метилентетрагидрофолатредуктазы может быть получена из GenBank NCBI - известной базы данных (например, WP_003863792.1, и т.д.). В частности, она может представлять собой полипептид, демонстрирующий активность 5,10-метилентетрагидрофолатредуктазы, кодируемый геном metF, но не ограничивается им.
В настоящем раскрытии термин «соответствующий» относится к аминокислотным остаткам в положениях, перечисленных в полипептиде, или к аминокислотным остаткам, которые являются аналогичными, идентичными или гомологичными остаткам, перечисленным в данном полипептиде. Идентификация аминокислоты в соответствующем положении может определяться специфической аминокислотой в последовательности, которая относится к специфической последовательности. Термин «соответствующая область» в том виде, в котором он здесь используется, обычно относится к аналогичному или соответствующему положению в родственном белке или эталонном белке.
Например, произвольную аминокислотную последовательность выравнивают с SEQ ID NO: 3, и, на основе этого, каждый аминокислотный остаток данной аминокислотной последовательности может быть пронумерован по отношению к аминокислотному остатку SEQ ID NO: 3 и числовому положению соответствующего аминокислотного остатка. Например, алгоритм выравнивания последовательности, как описано в настоящем описании, может определять положение аминокислоты или положение, в котором происходит модификация, такая как замена, вставка или делеция, посредством сравнения с положением в запрашиваемой последовательности (также именуемой «эталонная последовательность»).
Для таких выравниваний, например, можно использовать алгоритм Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), программу Нидлмана пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277) и тому подобные, но программа и алгоритм не ограничиваются ими, и подходящим образом можно использовать программу выравнивания последовательностей, алгоритм попарного сравнения последовательностей и тому подобное, известные в данной области.
Другим аспектом настоящего раскрытия является предоставление полинуклеотид а, кодирующего вариант по настоящему раскрытию.
В настоящем раскрытии термин «полинуклеотид» представляет собой нить ДНК или РНК, имеющую определенную или большую длину, в качестве полимера нуклеотидов, в котором нуклеотидные мономеры соединяются в длинную цепь ковалентными связями, и, более конкретно, означает фрагмент полинуклеотида, кодирующий данный вариант.
Полинуклеотид, кодирующий вариант по настоящему раскрытию, может содержать последовательность оснований, кодирующую аминокислотную последовательность, представленную SEQ ID NO: 1. В качестве примера по настоящему раскрытию полинуклеотид по настоящему раскрытию может иметь или содержать последовательность SEQ ID NO: 2. Полинуклеотид по настоящему раскрытию может состоять или по существу состоит из последовательности SEQ ID NO: 2.
В другом воплощении в полинуклеотиде по настоящему раскрытию основание, соответствующее положению 11 на основе последовательности нуклеиновой кислоты SEQ ID NO: 4 в последовательности нуклеиновой кислоты, представленной SEQ ID NO: 2, представляет собой А, и данный полинуклеотид может содержать последовательность нуклеиновой кислоты, имеющую по меньшей мере 70%-ную, 75%-ную, 80%-ную, 85%-ную, 90%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную, 99%-ную, 99,5%-ную, 99,7%-ную или 99,9%-ную или большую гомологию или идентичность с последовательностью нуклеиновой кислоты, представленной SEQ ID NO: 2. Очевидно, что полинуклеотиды, имеющие аминокислотные последовательности, в которых некоторые последовательности удалены, модифицированы, заменены, консервативно заменены или добавлены, также включены в объем настоящего раскрытия, при условии, что данные последовательности нуклеиновой кислоты имеют такую гомологию или идентичность и кодируют полипептид или белок, демонстрирующий эффективность, соответствующую эффективности варианта по настоящему раскрытию.
В полинуклеотиде по настоящему раскрытию могут быть сделаны разные модификации в кодирующей области при условии, что аминокислотная последовательность варианта по настоящему раскрытию не изменяется при рассмотрении вырожденности предпочтительных кодонов в организмах, которые предназначены для экспрессии варианта по настоящему раскрытию. В частности, полинуклеотид по настоящему раскрытию имеет или содержит последовательность оснований, имеющую 70%-ную или большую, 75%-ную или большую, 80%-ную или большую, 85%-ную или большую, 90%-ную или большую, 95%-ную или большую, 96%-ную или большую, 97%-ную или большую, 98%-ную или большую, но менее, чем 100%-ную гомологию или идентичность с последовательностью SEQ ID NO: 2 или может состоять или по существу состоит из последовательности оснований, имеющей 70%-ную или большую, 75%-ную или большую, 80%-ную или большую, 85%-ную или большую, 90%-ную или большую, 95%-ную или большую, 96%-ную или большую, 97%-ную или большую, 98%-ную или большую, но менее, чем 100%-ную гомологию или идентичность с последовательностью SEQ ID NO: 2, но не ограничиваясь ей. Здесь в последовательности, имеющей гомологию или идентичность, кодон, кодирующий аминокислоту, соответствующую положению 302 в SEQ ID NO: 1, может представлять собой один из кодонов, кодирующих гистидин.
Полинуклеотид по настоящему раскрытию может содержать зонд, который может быть получен из последовательности известного гена, например, последовательности без ограничения при условии, что она представляет собой последовательность, которая может гибридизоваться с комплементарной последовательностью всей или части последовательности полинуклеотида по настоящему раскрытию при жестких условиях. «Жесткие условия» означают условия, которые обеспечивают специфичную гибридизацию между полинуклеотидами. Данные условия конкретно описаны в документах (см. J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, 9.50-9.51, 11.7-11.8). Их примеры включают условия, при которых полинуклеотиды, имеющие более высокую гомологию или идентичность, а именно: полинуклеотиды, имеющие 70%-ную или большую, 75%-ную или большую, 80%-ную или большую, 85%-ную или большую, 90%-ную или большую, 95%-ную или большую, 96%-ную или большую, 97%-ную или большую, 98%-ную или большую, или 99%-ную или большую гомологию или идентичность, гибридизуются друг с другом, тогда как полинуклеотиды, имеющие меньшую гомологию или идентичность, не гибридизуются друг с другом, или условия, при которых промывка осуществляется один раз, в частности, от двух до трех раз при концентрации соли и температуре, эквивалентных 60°С, 1xSSC (раствор цитрата и хлорида натрия), 0,1% SDS (додецилсульфат натрия), в частности, при 60°С, 0,1xSSC, 0,1% SDS, более конкретно, при 68°С, 0,1xSSC, 0,1% SDS, которые представляют собой условия промывки для обычной гибридизации по Саузерну.
Для гибридизации требуется то, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя и допускаются несоответствия между основаниями, в зависимости от жесткости гибридизации. Термин «комплементарный» используется для описания связи между нуклеотидными основаниями, способными к гибридизации друг с другом. Например, в отношении ДНК, аденин является комплементарным тимину, а цитозин является комплементарным гуанину. Следовательно, полинуклеотид по настоящему раскрытию также может содержать по существу аналогичные последовательности нуклеиновой кислоты, а также фрагменты выделенной нуклеиновой кислоты, которые являются комплементарными всей последовательности.
В частности, полинуклеотид, имеющий гомологию или идентичность с полинуклеотидом по настоящему раскрытию, может быть выявлен с использованием условий гибридизации, включая стадию гибридизации при значении Tm 55°С и вышеописанных условиях. Значение Tm может составлять 60°С, 63°С или 65°С, но не ограничивается ими, и его могут подходящим образом корректировать специалисты в данной области согласно цели.
Подходящая жесткость для гибридизации полинуклеотида зависит от длины и степени комплементарно ста данного полинуклеотида, и переменные хорошо известны в данной области (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989).
Другим аспектом настоящего раскрытия является предложение вектора, содержащего полинуклеотид по настоящему раскрытию. Данный вектор может представлять собой экспрессионный вектор для осуществления экспрессии полинуклеотида в клетке-хозяине, но не ограничивается им.
Вектор по настоящему раскрытию может включать ДНК-конструкцию, содержащую последовательность полинуклеотида, кодирующую интересующий полипептид, связанный функциональным образом с подходящей регуляторной областью экспрессии (или регуляторной последовательностью экспрессии) таким образом, что интересующий полипептид может экспрессироваться в подходящем хозяине. Регуляторная область экспрессии может содержать промотор, способный инициировать транскрипцию, любую последовательность оператора для осуществления регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания мРНК с рибосомой, и последовательность, регулирующую терминацию транскрипции и трансляции. Данный вектор можно трансформировать в подходящую клетку-хозяина, и затем он может реплицироваться или функционировать независимо от генома хозяина, или может сам интегрироваться в геном.
Вектор, используемый в настоящем раскрытии, конкретно не ограничивается, но можно использовать любой вектор, известный в данной области. Примеры обычно используемых векторов включают природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и тому подобные, и в качестве плазмидного вектора можно использовать систему pDZ, систему pBR, систему pUC, систему pBluescript II, систему pGEM, систему pTZ, систему pCL, систему рЕТ и тому подобные. В частности, можно использовать векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 и pCC1BAC, и тому подобные.
Например, полинуклеотид, кодирующий интересующий полипептид, можно вставлять в хромосому посредством вектора для внутриклеточной хромосомной вставки. Вставку данного полинуклеотида в хромосому можно осуществлять любым способом, известным в данной области, например, гомологичной рекомбинацией, но не ограничиваясь ей. Данный вектор может дополнительно содержать селективный маркер для подтверждения вставки в хромосому. Данный селективный маркер служит для отбора клеток, трансформированных векторами, то есть, для подтверждения вставки интересующей молекулы нуклеиновой кислоты, и можно использовать маркеры, которые придают селектируемые фенотипы, такие как устойчивость к лекарственным средствам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия поверхностных полипептидов. В среде при обработке селективным агентом выживают или демонстрируют другие фенотипические признаки только клетки, экспрессирующие селективный маркер, и, таким образом, могут быть отобраны трансформированные клетки.
В настоящем раскрытии термин «трансформация» означает то, что вектор, содержащий полинуклеотид, кодирующий целевой полипептид, вводится в клетку-хозяина или микроорганизм таким образом, что полипептид, кодируемый данным полинуклеотидом, может экспрессироваться в клетке-хозяине. Трансформированный полинуклеотид может локализоваться посредством вставки в хромосому клетки-хозяина или локализоваться вне хромосомы, при условии, что он может экспрессироваться в данной клетке-хозяине. Данный полинуклеотид содержит ДНК и/или РНК, кодирующую интересующий полипептид. Данный полинуклеотид может быть вставлен в любой форме, при условии, что он может быть введен в клетку-хозяина и может экспрессироваться. Например, данный полинуклеотид может быть введен в клетку-хозяина в виде экспрессионной кассеты, которая представляет собой генетическую конструкцию, содержащую все элементы, требующиеся для автономной экспрессии. Данная экспрессионная кассета обычно может содержать промотор, связанный функциональным образом с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции. Данная экспрессионная кассета может находиться в виде экспрессионного вектора, способного к автономной репликации. Полинуклеотид может быть введен в клетку-хозяина сам по себе и связан функциональным образом с последовательностью, требующейся для экспрессии в клетке-хозяине, но не ограничиваясь ей.
В приведенном выше термин «связанный функциональным образом» означает то, что последовательность полинуклеотида функционально связана с последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего интересующий вариант по настоящему раскрытию.
Еще одним другим аспектом настоящего раскрытия является предложение штамма Corynebacterium glutamicum, который содержит вариант по настоящему раскрытию или полинуклеотид по настоящему раскрытию.
Штамм по настоящему раскрытию может содержать модифицированный полипептид по настоящему раскрытию, полинуклеотид, кодирующий данный полипептид, или вектор, содержащий полинуклеотид по настоящему раскрытию.
В настоящем раскрытии «штамм (или микроорганизм)» включает все микроорганизмы дикого типа или естественно или искусственно генетически модифицированные микроорганизмы, и он может представлять собой микроорганизм, в котором ослаблен или усилен специфический механизм из-за вставки внешнего гена или усиления активности, или инактивации эндогенного гена, и он может представлять собой микроорганизм, содержащий генетическую модификацию для продуцирования интересующего полипептида, белка или продукта.
Штамм по настоящему раскрытию может представлять собой штамм, содержащий любой один или более чем один вариант по настоящему раскрытию, полинуклеотид по настоящему раскрытию или вектор, содержащий полинуклеотид по настоящему раскрытию; штамм, модифицированный для экспрессии варианта по настоящему раскрытию или полинуклеотида по настоящему раскрытию; штамм (например, рекомбинантный штамм), экспрессирующий вариант по настоящему раскрытию или полинуклеотид по настоящему раскрытию; или штамм (например, рекомбинантный штамм), демонстрирующий активность варианта по настоящему раскрытию, но не ограничивается ими.
Штамм по настоящему раскрытию может представлять собой штамм, имеющий способность к продуцированию L-валина.
Штамм по настоящему раскрытию может представлять собой микроорганизм, имеющий в природе активность 5,10-метилентетрагидрофолатредуктазы и/или способность к продуцированию L-валина, или микроорганизм, в котором вариант по настоящему раскрытию или полинуклеотид, кодирующий данный вариант (или вектор, содержащий данный полинуклеотид), вводят в родительский штамм, который не имеет активности 5,10-метилентетрагидрофолатредуктазы или способности к продуцированию L-валина, и/или в котором способность к продуцированию L-валина придается родительскому штамму, но не ограничивается им.
Например, штамм по настоящему раскрытию представляет собой клетку или микроорганизм, который трансформирован вектором, содержащим полинуклеотид по настоящему раскрытию, или полинуклеотидом, кодирующим вариант по настоящему раскрытию, и экспрессирует вариант по настоящему раскрытию. В целях настоящего раскрытия штамм по настоящему раскрытию может включать все микроорганизмы, которые содержат вариант по настоящему раскрытию и могут продуцировать L-валин. Например, штамм по настоящему раскрытию может представлять собой рекомбинантный штамм, в котором полинуклеотид, кодирующий вариант по настоящему раскрытию, вводят в природный микроорганизм дикого типа или микроорганизм, продуцирующий L-валин, для того, чтобы, таким образом, экспрессировать вариант 5,10-метилентетрагидрофолатредуктазы и иметь повышенную способность к продуцированию L-валина. Рекомбинантный штамм, имеющий повышенную способность к продуцированию L-валина, может представлять собой микроорганизм, имеющий повышенную способность к продуцированию L-валина по сравнению с природным микроорганизмом дикого типа или микроорганизмом, не модифицированным 5,10-метилентетрагидрофолатредуктазой (а именно: микроорганизмом, экспрессирующим 5,10-метилентетрагидрофолатредуктазу дикого типа (SEQ ID NO: 3), или микроорганизмом, который не экспрессирует модифицированный (SEQ ID NO: 1) белок), но не ограничивается им. Например, штамм по настоящему раскрытию, имеющий повышенную способность к продуцированию L-валина, может представлять собой микроорганизм, имеющий повышенную способность к продуцированию L-валина по сравнению с Corynebaterium glutamicum, которая содержит полипептид SEQ ID NO: 3 или полинуклеотид, кодирующий данный полипептид, но не ограничивается им. В качестве примера, микроорганизм, не модифицированный 5,10-метилентетрагидрофолатредуктазой, который представляет собой целевой штамм для сравнения увеличения способности к продуцированию L-валина, может представлять собой штамм АТСС14067 и/или штамм Corynebaterium glutamicum СА08-0072 (KCCM11201P), но не ограничивается им.
Например, рекомбинантный штамм, имеющий повышенную способность к продуцированию, может иметь способность к продуцированию L-валина, повышенную примерно на 1% или более, примерно на 2,5% или более, примерно на 5% или более, примерно на 6% или более, примерно на 7% или более, примерно на 8% или более, примерно на 9% или более, примерно на 10% или более, примерно на 10,5% или более, примерно на 11% или более, примерно на 11,5% или более, примерно на 12% или более, примерно на 12,5% или более, примерно на 13% или более, примерно на 13,5% или более, примерно на 14% или более, примерно на 14,5% или более, примерно на 15% или более, примерно на 15,5% или более, примерно на 16% или более, примерно на 16,5% или более, примерно на 17% или более, примерно на 17,5% или более, примерно на 18% или более, примерно на 18,5% или более, примерно на 19% или более, примерно на 19,5% или более, примерно на 20% или более, примерно на 20,5% или более, примерно на 21% или более, примерно на 21,5% или более, примерно на 22% или более, примерно на 22,5% или более, примерно на 23% или более, примерно на 23,5% или более, примерно на 24% или более, 24,5% или более, примерно 25% или более, примерно 25,5% или более, примерно 25,7% или более, примерно 26% или более, примерно 26,5% или более, примерно на 27% или более, примерно на 27,5% или более, примерно на 28% или более, примерно на 28,5% или более, примерно на 29% или более, примерно на 29,5% или более, примерно на 30% или более, примерно на 30,5% или более, примерно на 31% или более, примерно на 31,5% или более, примерно на 32% или более, примерно на 32,5% или более, примерно на 33% или более, примерно на 33,1% или более, примерно на 33,2% или более, или примерно на 33,3% или более (верхняя граница конкретно не ограничивается и может составлять, например, примерно 200% или менее, примерно 150% или менее, примерно 100% или менее, примерно 50% или менее, примерно 45% или менее, примерно 40% или менее, или примерно 35% или менее) по сравнению со способностью к продуцированию L-валина родительского штамма перед изменением или с немодифицированным микроорганизмом, но повышенное значение не ограничивается им, при условии, что способность к продуцированию имеет повышенное значение плюс значения по сравнению со способностью к продуцированию родительского штамма перед изменением или с немодифицированным микроорганизмом. В другом примере рекомбинантный штамм, имеющий повышенную способность к продуцированию, может иметь способность к продуцированию L-валина, увеличенную примерно в 1,1 раза или более, примерно в 1,12 раза или более, примерно в 1,13 раза или более, примерно в 1,15 раза или более, примерно в 1,16 раза или более, примерно в 1,17 раза или более, примерно в 1,18 раза или более, примерно в 1,19 раза или более, примерно в 1,2 раза или более, примерно в 1,25 раза или более, примерно в 1,3 раза или более, примерно в 1,31 раза или более, примерно в 1,32 раза или более или примерно в 1,33 раза или более (верхняя граница конкретно не ограничивается и может, например, быть примерно в 10 раз или менее, примерно в 5 раз или менее, примерно в 3 раза или менее, или примерно в 2 раза или менее) по сравнению со способностью к продуцированию L-валина родительского штамма перед изменением или с немодифицированным микроорганизмом.
Более конкретно, рекомбинантный штамм, имеющий повышенную способность к продуцированию, может иметь способность к продуцированию L-валина, повышенную примерно на 32,1% (или примерно в 1,32 раза) по сравнению со способностью к продуцированию L-валина родительского штамма перед изменением или с немодифицированным микроорганизмом, но показатель увеличения не ограничивается им. Термин «примерно» представляет собой интервал, включающий все из плюс/минус 0,5; плюс/минус 0,4; плюс/минус 0,3; плюс/минус 0,2; плюс/минус 0,1 и тому подобных, и включает все значения в интервале, равные или аналогичные значению после термина «примерно», но не ограничивается ими.
В настоящем раскрытии «немодифицированный микроорганизм» не исключает штаммы, содержащие мутацию, которая может случаться у микроорганизмов в природе, и может представлять собой штамм дикого типа или сам природный штамм, или может представлять собой штамм перед изменением признака посредством генетической вариации из-за природных или искусственных факторов. Например, данный немодифицированный микроорганизм может представлять собой штамм, в который не вводится или еще не был введен вариант белка, описанный в настоящем описании изобретения. Термин «немодифицированный микроорганизм» можно использовать взаимозаменяемо с фразами «штамм перед модификацией», «микроорганизм перед модификацией», «неизмененный штамм», «немодифицированный штамм», «неизмененный микроорганизм» или «эталонный микроорганизм».
В другом примере настоящего раскрытия микроорганизм по настоящему раскрытию может представлять собой Corynebacterium glutamicum, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium stationis, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium ammoniagenes, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens.
В настоящем раскрытии «ослабление» активности полипептида включает оба случая, где активность снижена по сравнению с эндогенной активностью или отсутствует. Термин «ослабление» можно использовать взаимозаменяемо с такими терминами, как инактивация, недостаточность, понижающая регуляция, снижение, уменьшение и аттенюация.
Ослабление также может включать случай, когда активность самого полипептида снижена или устранена по сравнению с активностью полипептида, которым исходно обладал микроорганизм, посредством изменения полинуклеотида, кодирующего данный полипептид, и тому подобного, случай, где общие уровень активности и/или концентрация (уровень экспрессии) полипептида в клетке ниже по сравнению с уровнем активности или концентрацией природного штамма посредством ингибирования экспрессии гена полинуклеотида, кодирующего полипептид, или ингибирования трансляции в полипептид, случай, где данный полинуклеотид совсем не экспрессируется, и/или случай, где активность полипептида не проявляется даже при экспрессии полинуклеотида. Термин «эндогенная активность» означает активность конкретного полипептида, которой исходно обладал родительский штамм перед изменением признака, или микроорганизм дикого типа, или немодифицированный микроорганизм при изменении признака генетической вариацией из-за природных или искусственных факторов. Фразу «эндогенная активность» можно использовать взаимозаменяемо с фразой «активность перед модификацией». Тот факт, что активность полипептида является «инактивированной, недостаточной, пониженной, подвергнувшейся понижающей регуляции, сниженной или ослабленной» по сравнению с эндогенной активностью означает то, что активность полипептида снижается по сравнению с активностью конкретного полипептида, которой исходно обладал родительский штамм перед изменением признака или микроорганизм дикого типа, или немодифицированный микроорганизм.
Такое ослабление активности полипептида можно осуществлять любым способом, известным в данной области, но данный способ не ограничивается им, и ослабления можно достигнуть применением разных способов, хорошо известных в данной области (например, Nakashima N. et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014; 15(2): 2773-2793, Sambrook et al., Molecular Cloning 2012, и тому подобные).
В частности, ослабление активности полипептида в настоящем раскрытии может представлять собой:
1) делецию всего или части гена, кодирующего полипептид;
2) модификацию регуляторной области экспрессии (или регуляторной последовательности экспрессии) для уменьшения экспрессии гена, кодирующего полипептид;
3) модификацию аминокислотной последовательности, составляющей полипептид, для устранения или ослабления активности полипептида (например, делеция/замена/присоединение одной или более чем одной аминокислоты в аминокислотной последовательности);
4) модификацию последовательности гена, кодирующей полипептид, для устранения или ослабления активности данного полипептида (например, делеция/замена/присоединение одного или более чем одного основания нуклеиновой кислоты в последовательности оснований нуклеиновой кислоты гена полипептида для кодирования полипептида, который был модифицирован для устранения или ослабления активности данного полипептида);
5) модификацию инициирующего кодона транскрипта гена, кодирующего полипептид, или последовательности оснований, кодирующей 5'-UTR (5'-нетранслируемая область) область;
6) введение антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующего полипептид;
7) добавление последовательности, комплементарной последовательности Шайна-Дальгарно, перед последовательностью Шайна-Дальгарно гена, кодирующего полипептид, для того, чтобы образовать вторичную структуру, к которой не может присоединяться рибосома;
8) добавление промотора, подлежащего транскрипции в противоположном направлении относительно 3'-конца открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид (инженерия обратной транскрипции, RTE); или
9) комбинацию двух или более чем двух, выбранных из (1) - (8), но, в частности, не ограничивается ими.
Например:
1) Делецией части или всего гена, кодирующего полипептид, может быть удаление всего полинуклеотида, кодирующего интересующий эндогенный полипептид в хромосоме, или замена полинуклеотидом, в котором некоторые нуклеотиды делетированы, или замена маркерным геном.
2) Модификацией регуляторной области экспрессии (или регуляторной последовательности экспрессии) может быть делеция, вставка, неконсервативная или консервативная замена, или появление изменения в регуляторной области экспрессии (или регуляторной последовательности экспрессии) из-за их комбинации, или замена последовательностью, демонстрирующей более слабую активность. Регуляторная область экспрессии содержит промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, но не ограничивается ими.
3) Модификацией инициирующего кодона транскрипта гена, кодирующего полипептид, или последовательности оснований, кодирующей 5'-UTR область, может быть, например, замена последовательностью оснований, кодирующей другой инициирующий кодон, имеющий меньшую скорость экспрессии полипептида по сравнению с эндогенным инициирующим кодоном, но не ограничивается ей.
4) и 5) Модификацией аминокислотной последовательности или последовательности полинуклеотида может быть делеция, вставка или неконсервативная, или консервативная замена аминокислотной последовательности полипептида или последовательности полинуклеотида, кодирующей данный полипептид, или появление изменения в последовательности из-за их комбинации, или замена аминокислотной последовательностью или последовательностью полинуклеотида, модифицированной так, чтобы демонстрировать более слабую активность, или аминокислотной последовательностью, или последовательностью полинуклеотида, модифицированной так, чтобы быть неактивной, таким образом, что активность данного полипептида ослабевает, но не ограничивается ими. Например, экспрессию гена можно ингибировать или ослаблять посредством введения изменения в последовательность полинуклеотида и образования терминирующего кодона, но модификация не ограничивается ей.
6) Для введения антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующего полипептид, можно сделать ссылку на документы, например, Weintraub, Н. et al., Antisense-RNA as a molecular tool for genetic analysis, Reviews-Trends in Genetics, Vol. 1(1) 1986.
7) Добавление последовательности, комплементарной последовательности Шайна-Дальгарно, перед последовательностью Шайна-Дальгарно, гена, кодирующего полипептид, для того, чтобы образовать вторичную структуру, к которой не может присоединиться рибосома, для того, чтобы сделать невозможной трансляцию мРНК или для замедления скорости трансляции мРНК.
8) Добавление промотора, подлежащего транскрипции в противоположном направлении относительно 3'-конца открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид (инженерия обратной транскрипции, RTE), может служить для ослабления активности посредством получения антисмыслового нуклеотида, комплементарного транскрипту гена, кодирующего полипептид.
В настоящем раскрытии термин «усиление» активности полипептида означает то, что активность полипептида увеличивается по сравнению с эндогенной активностью. Термин «усиление» можно использовать взаимозаменяемо с такими терминами, как активация, повышающая регуляция, сверхэкспрессия и увеличение.
Здесь активация, усиление, повышающая регуляция, сверхэкспрессия и увеличение могут включать как демонстрирование активности, которой исходно не обладали, так и демонстрирование улучшенной активности по сравнению с эндогенной активностью или активностью перед модификацией. Термин «эндогенная активность» означает активность конкретного полипептида, которой исходно обладал родительский штамм перед изменением признака или немодифицированный микроорганизм при изменении признака посредством генетической вариации из-за природных или искусственных факторов. Это можно использовать взаимозаменяемо с «активностью перед модификацией». Тот факт, что активность полипептида «усиливается», «подвергается повышающей регуляции», «сверхэкспрессируется» или «увеличивается» по сравнению с эндогенной активностью означает то, что активность полипептида улучшается по сравнению с активностью и/или концентрацией (уровнем экспрессии) конкретного полипептида, которой исходно обладает родительский штамм перед изменением признака или немодифицированный микроорганизм.
Данное усиление может быть достигнуто посредством введения чужеродного полипептида или увеличения эндогенной активности и/или концентрации (уровня экспрессии) данного полипептида. Усиление активности полипептида может быть подтверждено увеличением степени активности и уровня экспрессии полипептида или количества продукта, продуцируемого из данного полипептида.
Для усиления активности полипептида можно применять разные способы, хорошо известные в данной области, и данный способ не ограничивается, при условии, что активность интересующего полипептида может быть усилена по сравнению с активностью микроорганизма до модификации. В частности, можно использовать генную инженерию и/или белковую инженерию, хорошо известные специалистам в данной области, которые представляют собой традиционные способы молекулярной биологии, но данный способ не ограничивается ими (например, Sitnicka et al., Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol. 2. 1-16; Sambrook et al., Molecular Cloning 2012; и тому подобные).
В частности, усиление активности полипептида по настоящему раскрытию может представлять собой:
1) увеличение числа внутриклеточных копий полинуклеотида, кодирующего полипептид;
2) замену регуляторной области экспрессии гена на хромосоме, кодирующей полипептид, последовательностью, демонстрирующей сильную активность;
3) модификацию инициирующего кодона транскрипта гена, кодирующего полипептид, или последовательности оснований, кодирующей 5'-UTR область;
4) модификацию аминокислотной последовательности полипептида для увеличения активности данного полипептида;
5) модификацию последовательности полинуклеотида, кодирующей полипептид, для усиления активности данного полипептида (например, модификацию последовательности полинуклеотида гена полипептида для кодирования полипептида, который был модифицирован для усиления активности данного полипептида);
6) введение чужеродного полипептида, демонстрирующего активность данного полипептида, или чужеродного полинуклеотида, кодирующего данный полипептид;
7) оптимизацию кодонов полинуклеотида, кодирующего полипептид;
8) анализ третичной структуры полипептида для отбора и модификации или химической модификации экспонированного сайта; или
9) комбинацию двух или более чем двух, выбранных из (1) - (8), но конкретно не ограничиваясь ими.
Более конкретно:
1) Увеличение числа внутриклеточных копий полинуклеотида, кодирующего полипептид, может быть достигнуто посредством введения в клетку-хозяина вектора, который может реплицироваться и функционировать независимо от хозяина, и с которым полинуклеотид, кодирующий данный полипептид, связан функциональным образом. В качестве альтернативы, увеличение может быть достигнуто введением одной копии или двух или более чем двух копий полинуклеотида, кодирующего данный полипептид, в хромосому клетки-хозяина. Введение в хромосому можно осуществлять введением вектора, способного вставлять полинуклеотид в хромосому клетки-хозяина, в клетку-хозяина, но не ограничивается им. Данный вектор является таким, как описано выше.
2) Замена регуляторной области экспрессии гена (или регуляторной последовательности экспрессии) на хромосоме, кодирующей полипептид, последовательностью, демонстрирующей сильную активность, может представлять собой, например, делецию, вставку, неконсервативную или консервативную замену, или появление изменения в последовательности из-за их комбинации, или замену последовательностью, демонстрирующей более сильную активность, таким образом, что активность регуляторной области экспрессии дополнительно усиливается. Регуляторная область экспрессии конкретно не ограничивается ими, но может содержать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, последовательность, регулирующую терминацию транскрипции и трансляции и тому подобные. Например, замена может представлять собой замену исходного промотора сильным промотором, но не ограничивается ей.
Примеры известных сильных промоторов включают промоторы CJ1-CJ7 (US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (US 10584338 В2), промотор O2 (US 10273491 В2), промотор tkt и промотор уссА, но не ограничиваются ими.
3) Модификация инициирующего кодона транскрипта гена, кодирующуего полипептид, или последовательности оснований, кодирующей 5'-UTR область, может представлять собой, например, замену последовательностью оснований, кодирующей другой инициирующий кодон, имеющий более высокую скорость экспрессии полипептида по сравнению с эндогенным инициирующим кодоном, но не ограничивается ей.
4) и 5) Модификация аминокислотной последовательности или последовательности полинуклеотида может представлять собой делецию, вставку, неконсервативную или консервативную замену аминокислотной последовательности полипептида или последовательности полинуклеотида, кодирующей данный полипептид, или появление изменения в данной последовательности из-за их комбинации или замены аминокислотной последовательностью или последовательностью полинуклеотида, модифицированной для демонстрации более сильной активности, или аминокислотной последовательностью или последовательностью полинуклеотида, модифицированной для того, чтобы быть более активной, таким образом, что активность данного полипептида усиливается, но не ограничивается ими. Данную замену можно конкретно осуществлять посредством вставки полинуклеотида в хромосому гомологичной рекомбинацией, но не ограничиваясь ей. Используемый здесь вектор может дополнительно содержать селективный маркер для подтверждения вставки в хромосому. Селективный маркер является таким, как описано выше.
6) Введение чужеродного полипептида, демонстрирующего активность полипептида, может представлять собой введение в клетку-хозяина чужеродного полинуклеотида, кодирующего полипептид, демонстрирующий такую же или аналогичную данному полипептиду активность. Данный чужеродный полинуклеотид не ограничивается по его происхождению или последовательности при условии, что он демонстрирует такую же или аналогичную данному полипептиду активность. Введение можно осуществлять посредством подходящего выбора способа трансформации, известного специалистам в данной области. Поскольку введенный полинуклеотид экспрессируется в клетке-хозяине, может продуцироваться полипептид и может увеличиваться его активность.
7) Оптимизация кодонов полинуклеотида, кодирующего полипептид, может представлять собой оптимизацию кодонов эндогенного полинуклеотида таким образом, чтобы увеличивать транскрипцию или трансляцию в клетке-хозяине, или оптимизацию кодонов чужеродного полинуклеотида таким образом, чтобы осуществлять оптимизированную транскрипцию и трансляцию в клетке-хозяине.
8) Анализ третичной структуры полипептида для выбора и модификации или химическая модификация экспонированного сайта можно осуществлять, например, для определения шаблонного белка-кандидата согласно степени сходства последовательности посредством сравнения информации по последовательности полипептида, подлежащего анализу, с хранилищем информации по последовательностям известных белков базы данных для подтверждения структуры на основе этого и для выбора и модификации или химической модификации экспонированной части, подлежащей модификации или химической модификации.
Такое усиление активности полипептида может представлять собой увеличение активности или концентрации, или уровня экспрессии соответствующего полипептида на основе активности или концентрации полипептида, экспрессируемого в микробном штамме дикого типа или в микробном штамме перед модификацией, или увеличение количества продукта, продуцируемого от полипептида, но не ограничивается ими.
В микроорганизме по настоящему раскрытию частичную или полную модификацию (например, модификация для кодирования варианта белка, как описано выше) полинуклеотида можно индуцировать (а) гомологичной рекомбинацией с использованием вектора для вставки в хромосому в микроорганизме или редактированием генома с использованием генетически модифицированной нуклеазы (например, CRISPR-Cas9), и/или (б) обработкой светом, таким как ультрафиолетовые лучи и излучение, и/или химическими агентами, но не ограничиваясь ими. Способ осуществления модификации части или всего гена может включать способ с использованием технологии рекомбинантной ДНК. Например, посредством введения в микроорганизм нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, гомологичную интересующему гену, для вызова гомологичной рекомбинации можно удалить часть гена или весь данный ген. Введенная нуклеотидная последовательность или вектор может содержать доминантный селективный маркер, но не ограничиваясь им.
В микроорганизме по настоящему раскрытию вариант, полинуклеотид, L-валин и тому подобные являются такими, как описано в других аспектах.
Согласно еще одному другому аспекту настоящего раскрытия предложен способ продуцирования L-аминокислоты, который включает культивирование в среде штамма Corynebacterium glutamicum, содержащего вариант по настоящему раскрытию или полинуклеотид по настоящему раскрытию.
Способ продуцирования L-аминокислоты по настоящему раскрытию может включать культивирование в среде штамма Corynebacterium glutamicum, содержащего вариант по настоящему раскрытию или полинуклеотид по настоящему раскрытию, или вектор по настоящему раскрытию.
Кроме того, L-аминокислота по настоящему раскрытию может представлять собой L-валин.
В настоящем раскрытии термин «культивирование» означает выращивание штамма Corynebacterium glutamicum по настоящему раскрытию при условиях среды, контролируемых подходящим образом. Способ культивирования по настоящему раскрытию можно осуществлять в соответствии с подходящей средой и условиями культивирования, известными в данной области. Такой способ культивирования можно легко корректировать и использовать специалистами в данной области согласно выбранному штамму. В частности, культивирование может быть периодического типа, непрерывного типа и/или типа с подпиткой, но не ограничивается ими.
В настоящем раскрытии термин «среда» означает смешанное вещество, содержащее питательные вещества, требующиеся для культивирования штамма Corynebacterium glutamicum по настоящему раскрытию в качестве главного компонента, и данная среда поставляет питательные вещества, факторы роста и тому подобное, включая воду, которые являются незаменимыми для выживания и развития.
В частности, в качестве среды и других условий культивирования, используемых для культивирования штамма Corynebacterium glutamicum по настоящему раскрытию, можно использовать любые без конкретного ограничения, при условии, что она представляет собой среду, используемую для обычного культивирования микроорганизмов. Штамм Corynebacterium glutamicum по настоящему раскрытию можно культивировать в обычной среде, содержащей правильные источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины, и тому подобное, при одновременном осуществлении контроля температуры, рН и тому подобного при аэробных условиях.
В частности, культуральную среду для штамма рода Corynebacterium можно найти в документе "Manual of Methods for General Bacteriology" by the American Society for Bacteriology (Washington, D.C., USA, 1981).
В настоящем раскрытии источники углерода включают углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, сахароза и мальтоза; сахароспирты, такие как маннит и сорбит; органические кислоты, такие как пировиноградная кислота, молочная кислота и лимонная кислота; аминокислоты, такие как глутаминовая кислота, метионин и лизин; и тому подобное. Можно использовать природные органические питательные вещества, такие как гидролизат крахмала, мелассу, сырую мелассу, рисовые отруби, маниок, остаток сахарного тростника и жидкий кукурузный экстракт. В частности, можно использовать углеводы, такие как глюкоза и стерилизованные пред обработанные мелассы (а именно: мелассы, превращенные в восстанавливающий сахар), и можно использовать подходящие количества других источников углерода разными способами без ограничения. Данные источники углерода можно использовать одиночно или в комбинации с двумя или более чем двумя, но не ограничиваясь ими.
В качестве источников азота можно использовать неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; и органические источники азота, такие как аминокислоты, такие как глутаминовая кислота, метионин и глутамин, пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыба или продукты ее разложения и обезжиренный соевый жмых или продукты его разложения. Данные источники азота можно использовать одиночно или в комбинации двух или более чем двух, но не ограничиваясь ими.
Источники фосфора могут включать монокалия фосфат, дикалия фосфат или соответствующие им натрийсодержащие соли. В качестве неорганических соединений можно использовать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и тому подобное. Помимо них могут содержаться аминокислоты, витамины и/или подходящие предшественники, и тому подобное. Данные компоненты или предшественники можно добавлять в среду порционно или непрерывно, но способ добавления не ограничивается ими.
Во время культивирования штамма Corynebacterium glutamicum по настоящему раскрытию рН среды можно корректировать добавлением в данную среду правильным способом таких соединений, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота. Во время культивирования пенообразование можно подавлять посредством применения пеногасителя, такого как сложный полигликолевый эфир жирной кислоты. В среду можно инъецировать кислород или кислородсодержащий газ для поддержания аэробного состояния данной среды, или газ можно не инъецировать, или можно инъецировать газообразный азот, водород или диоксид углерода для того, чтобы поддерживать анаэробное и микроаэробное состояния, но способ поддержания данного состояния не ограничивается ими.
При культивировании по настоящему раскрытию можно поддерживать температуру культивирования от 20°С до 45°С, в частности, от 25°С до 40°С, и данный штамм можно культивировать в течение примерно от 10 до 160 часов, но условия культивирования не ограничиваются ими.
L-аминокислота, продуцируемая посредством культивирования по настоящему раскрытию, может секретироваться в среду или может оставаться в клетках.
Способ продуцирования L-аминокислоты по настоящему раскрытию может дополнительно включать стадию получения штамма Corynebacterium glutamicum по настоящему раскрытию, стадию приготовления среды для культивирования данного штамма или их комбинацию (в любом порядке), например, перед стадией культивирования.
Способ продуцирования L-аминокислоты по настоящему раскрытию может дополнительно включать стадию выделения L-аминокислоты из среды в соответствии с культивированием (среда, подвергнувшаяся воздействию культуры) или из штамма Corynebacterium glutamicum. После стадии культивирования может быть дополнительно включена стадия выделения.
Выделение может служить для сбора интересующей L-аминокислоты посредством подходящего способа, известного в данной области, согласно способу культивирования микроорганизма по настоящему раскрытию, например, способу периодического, непрерывного культивирования или культивирования с подпиткой. Например, можно использовать центрифугирование, фильтрование, обработку осадителем кристаллизованного белка (высаливание), экстракцию, ультразвуковое разрушение, ультрафильтрацию, диализ, разные виды хроматографии, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, ВЭЖХ (высокоэффективная жидкостная хроматография), или их комбинацию. Интересующую L-аминокислоту можно выделять из среды или микроорганизма посредством подходящего способа, известного в данной области.
Способ продуцирования L-аминокислоты по настоящему раскрытию может дополнительно включать стадию очистки. Очистку можно осуществлять посредством подходящего способа, известного в данной области. В одном примере, когда способ продуцирования L-аминокислоты по настоящему раскрытию включает и стадию выделения, и стадию очистки, данные стадии выделения и очистки можно осуществлять непрерывно или прерывисто, независимо от порядка, или можно осуществлять одновременно, или посредством объединения в одну стадию, но способ осуществления данных стадий не ограничивается ими.
В способе по настоящему раскрытию вариант, полинуклеотид, вектор, штамм и тому подобное являются такими, как описано в других аспектах.
Еще одним другим аспектом настоящего раскрытия является предложение композиции для продуцирования L-аминокислоты, которая содержит штамм Corynebacterium glutamicum, содержащий вариант по настоящему раскрытию, полинуклеотид, кодирующий данный вариант, вектор, содержащий данный полинуклеотид, или полинуклеотид по настоящему раскрытию; среду, в которой культивировали данный штамм Corynebacterium glutamicum; или комбинацию двух или более чем двух из них.
Композиция по настоящему раскрытию может дополнительно содержать произвольные подходящие эксципиенты, которые обычно применяют в композициях для продуцирования аминокислот. Такие эксципиенты могут представлять собой, например, консервант, увлажнитель, диспергирующий агент, суспендирующий агент, буферизующий агент, стабилизатор или изотоничный агент, но не ограничиваются ими.
В композиции по настоящему раскрытию вариант, полинуклеотид, вектор, штамм, среда, L-аминокислота и тому подобные являются такими, как описано в других аспектах.
Полезные эффекты
В случае культивирования штамма Corynebacterium glutamicum, содержащего вариант 5,10-метилентетрагидрофолатредуктазы по настоящему раскрытию, возможна продукция L-валина с более высоким выходом по сравнению со случаем существующих микроорганизмов, имеющих немодифицированные полипептиды.
Краткое описание графических материалов
ФИГ. 1. Схематичная диаграмма плазмиды pDCM2.
Способ осуществления изобретения
Ниже настоящее раскрытие будет более подробно описано посредством Примеров. Однако следующие Примеры являются лишь предпочтительными воплощениями для иллюстрации настоящего раскрытия и, таким образом, не предназначены для ограничения ими объема настоящего раскрытия. Тем не менее, технические вопросы, не описанные в настоящем описании изобретения, могут быть в достаточной степени поняты и легко воплощены специалистами в технической области настоящего раскрытия или в аналогичных технических областях.
Пример 1: конструирование плазмиды
Плазмида (pDCM2, ФИГ. 1, SEQ ID NO: 11) для вставки и замены генов в хромосоме Corynebacterium была сконструирована и синтезирована с использованием службы Gene-synthesis от BIONICS Co., Ltd. Данная плазмида была сконструирована так, чтобы она содержала сайт рестрикционного фермента для легкого применения для клонирования со ссылкой на общеизвестную статью по системе sacB (Gene, 145 (1994) 69-73). Плазмида pDCM2, синтезированная таким образом, имеет следующие свойства:
1) Плазмида pDCM2 имеет репликатор, который работает только в Е. coli, и автономная репликация, таким образом, возможна в Е. coli, но не в Corynebacterium.
2) Плазмида pDCM2 имеет ген устойчивости к канамицину в качестве селективного маркера.
3) Плазмида pDCM2 имеет ген левансахаразы (sacB) в качестве вторичного маркера положительного отбора.
4) В конечном сконструированном штамме не остается информации о гене, происходящем из плазмиды pDCM2.
Пример 2: Конструирование вектора для экспрессии в микроорганизме варианта белка
Для того, чтобы подтвердить влияние варианта (R4H; SEQ ID NO: 1), у которого аргинин (Arg, R) в положении 4 белка, состоящего из аминокислотной последовательности SEQ ID NO: 3, был заменен гистидином (His, Н), на продукцию L-валина, конструировали вектор для конструирования штамма, экспрессирующего данный вариант, следующим образом.
ПЦР (полимеразная цепная реакция) проводили с использованием гДНК (геномная ДНК) Corynebacterium glutamicum дикого типа АТСС14067 в качестве матрицы, пары праймеров, имеющих последовательности SEQ ID NO: 5 и 6, и пары праймеров, имеющих последовательности SEQ ID NO: 7 и 8. Вновь проводили ПЦР с перекрывающимися праймерами с использованием смеси двух фрагментов, полученных выше, в качестве матрицы и пары праймеров, имеющих последовательности SEQ ID NO: 5 и SEQ ID NO: 8, с получением фрагмента. ПЦР проводили следующим образом: денатурация при 94°С в течение 5 минут; 30 циклов при 94°С в течение 30 секунд, 55°С в течение 30 секунд и 72°С в течение 1 минуты 30 секунд; и 72°С в течение 5 минут. Вектор pDCM2 обрабатывали smaI, и ПЦР-продукт, полученный выше, клонировали в него посредством слияния. Клонирование посредством слияния проводили с использованием набора для клонирования In-Fusion® HD (Clontech). Полученную плазмиду называли pDCM2-nadS(P302S). Последовательности праймеров, используемых в данном примере, описаны в следующей Таблице 1:
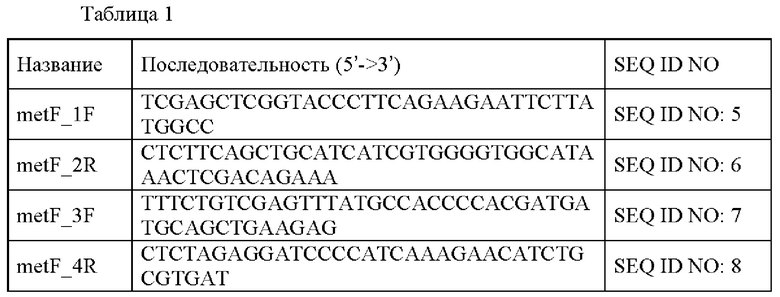
Пример 3. Оценка способности к продуцированию L-валина микроорганизма, экспрессирующего вариант белка
3-1. Конструирование штамма, экспрессирующего вариант белка
Вектор, сконструированный в Примере 2, трансформировали в Corynebacterium glutamicum СА08-0072 (KCCM1120 IP).
Штамм, в который был введен данный вариант, был выбран из штаммов, в которых происходила гомологичная рекомбинация с использованием SEQ ID NO: 9 и 10. Выбранный штамм называли CA08-0072_metF_R4H. Последовательности праймеров, использованных в данном примере, описываются в следующей Таблице 2:
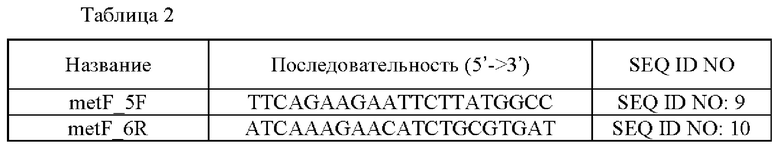
3-2. Сравнение способности к продуцированию L-валина штаммов, экспрессирующих вариант белка
Способность к продуцированию L-валина анализировали посредством оценки титра ферментации в колбе каждого штамма, сконструированного в Примере 3-1, и контрольного родительского штамма.
Сначала каждую колонию субкультивировали в питательной среде, и затем каждый штамм инокулировали в 250 мл колбу с угловыми перегородками, содержащей 25 мл продукционной среды, и подвергали культивированию со встряхиванием при 200 об/мин при 30°С в течение 72 часов. Затем концентрацию L-валина анализировали посредством ВЭЖХ, и проанализированная концентрация L-валина показана в Таблице 3 ниже.
Питательная среда (рН 7,2)
10 г глюкозы, 5 г экстракта говядины, 10 г полипептона, 2,5 г хлорида натрия, 5 г дрожжевого экстракта, 20 г агара, 2 г мочевины (на основе 1 литра дистиллированной воды).
Продукционная среда (рН 7,0)
100 г глюкозы, 40 г сульфата аммония, 2,5 г соевого белка, 5 г твердых веществ кукурузного экстракта, 3 г мочевины, 1 г дикалия фосфата, 0,5 г сульфата магния гептагидрата, 100 мкг биотина, 1 мг тиамина-HCl, 2 мг пантотената кальция, 3 мг никотинамида, 30 г карбоната кальция (на основе 1 литра дистиллированной воды).
Данный эксперимент повторяли 3 раза, и средние значения результатов его анализа представлены в Таблице 3 ниже.
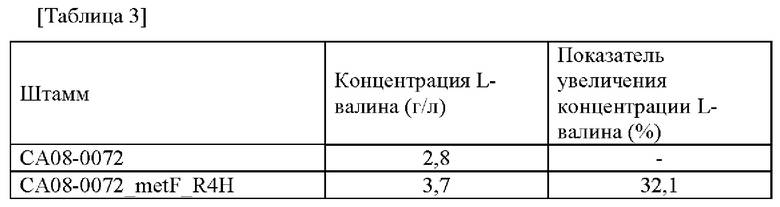
Как представлено в Таблице 3, штамм СА08-0072_metF_R4H демонстрировал повышенную способность к продуцированию L-валина по сравнению со способностью к продуцированию L-валина контрольной группы.
Модифицированный штамм CA08-0072_metF_R4H называли Corynebacterium glutamicum СА08-1746, депонировали в Корейский центр культуры микроорганизмов, учреждение депонирования, работающее согласно Будапештскому соглашению, 2 декабря 2020 г., и ему был присвоен номер доступа KCCM12862P.
Из приведенного выше описания специалистам в технической области, к которой принадлежит настоящее раскрытие, будет понятно то, что настоящее раскрытие можно осуществлять в других конкретных формах без изменения его технической сущности и важных характеристик. В данном отношении следует понимать то, что воплощения, описанные выше, являются во всех отношениях иллюстративными и не ограничивающими. Объем настоящего раскрытия следует истолковывать как включающий все изменения или модифицированные формы, происходящие из значения и объема формулы изобретения, подлежащей описанию ниже, а не из приведенного выше подробного описания и эквивалентных ему идей.
Наименование депозитария: Корейский центр культуры микроорганизмов
Номер доступа: KCCM12862P
Дата депонирования: 02.12.2020
--->
<110> CJ CheilJedang Corporation
<120> Новый вариант 5,10-метилентетрагидрофолатредуктазы и
способ получения L-валина с его применением
<130> OPP20211119KR
<150> KR 10-2021-0011010
<151> 2021-01-26
<160> 11
<170> koPatentIn 3.0
<210> 1
<211> 294
<212> PRT
<213> Artificial Sequence
<220>
<223> вариант 5,10-метилентетрагидрофолатредуктазы
<400> 1
Met Pro Pro His Asp Asp Ala Ala Glu Glu Arg Leu Tyr Arg Ala Ala
1 5 10 15
Glu Val Phe His Asp Leu Gly Ala Ser Phe Val Ser Val Thr Tyr Gly
20 25 30
Ala Gly Gly Ser Thr Arg Glu Arg Thr Ser Arg Ile Ala Arg Arg Leu
35 40 45
Ala Lys Gln Pro Leu Thr Thr Leu Val His Leu Thr Leu Val Asn His
50 55 60
Thr Arg Glu Glu Met Lys Ala Ile Leu Arg Glu Tyr Leu Glu Leu Gly
65 70 75 80
Leu Thr Asn Leu Leu Ala Leu Arg Gly Asp Pro Pro Gly Asp Pro Leu
85 90 95
Gly Asp Trp Val Ser Thr Asp Gly Gly Leu Asn Tyr Ala Ser Glu Leu
100 105 110
Ile Asp Leu Ile Lys Ser Thr Pro Glu Phe Arg Glu Phe Asp Leu Gly
115 120 125
Ile Ala Ser Phe Pro Glu Gly His Phe Arg Ala Lys Thr Leu Glu Glu
130 135 140
Asp Thr Lys Tyr Thr Leu Ala Lys Leu Arg Gly Gly Ala Glu Tyr Ser
145 150 155 160
Ile Thr Gln Met Phe Phe Asp Val Glu Asp Tyr Leu Arg Leu Arg Asp
165 170 175
Arg Leu Val Ala Ala Asp Pro Ile His Gly Ala Lys Pro Ile Ile Pro
180 185 190
Gly Ile Met Pro Ile Thr Ser Leu Arg Ser Val Arg Arg Gln Val Glu
195 200 205
Leu Ser Gly Ala Gln Leu Pro Ser Gln Leu Glu Glu Ser Leu Val Arg
210 215 220
Ala Ala Asn Gly Asn Glu Glu Ala Asn Lys Asp Glu Ile Arg Lys Val
225 230 235 240
Gly Ile Glu Tyr Ser Thr Asn Met Ala Glu Arg Leu Ile Ala Glu Gly
245 250 255
Ala Glu Asp Leu His Phe Met Thr Leu Asn Phe Thr Arg Ala Thr Gln
260 265 270
Glu Val Leu Tyr Asn Leu Gly Met Ala Pro Ala Trp Gly Ala Glu His
275 280 285
Gly Gln Asp Ala Val Arg
290
<210> 2
<211> 885
<212> DNA
<213> Artificial Sequence
<220>
<223> ген, кодирующий вариант 5,10-метилентетрагидрофолатредуктазы
<400> 2
atgccacccc acgatgatgc agctgaagag cgtctttacc gagcagcaga ggtcttccat 60
gacctcggtg catcgtttgt ctccgtgact tatggtgctg gcggatcaac ccgtgagaga 120
acctcacgta ttgctcgacg attagcgaaa caaccgttga ccactctggt gcacctgacc 180
ctggttaacc acactcgcga agagatgaag gcaattcttc gggaatacct agagctggga 240
ttaacaaacc tgttggcgct tcgaggagat ccgcctggag acccattagg cgattgggtg 300
agcaccgatg gaggactgaa ctacgcctct gagctcatcg atcttattaa gtccactcct 360
gagttccggg aattcgacct cggtatcgcc tccttccccg aagggcattt ccgggcgaaa 420
actctagaag aagacaccaa atacactctg gcgaagctgc gtggaggtgc agagtactcc 480
atcacgcaga tgttctttga tgtggaagac tacctgcgac ttcgtgatcg ccttgtcgct 540
gcagacccca ttcatggcgc gaagccaatc attcctggca tcatgcccat tacgagcctg 600
cggtctgtgc gtcgacaggt cgaactctct ggtgctcaat tgccgagcca actagaagaa 660
tcacttgttc gagctgcaaa cggcaatgaa gaagcgaaca aagacgagat ccgcaaggtg 720
ggcattgaat attccaccaa tatggcagag cgactcattg ccgaaggtgc ggaagatctg 780
cacttcatga cgcttaactt cacccgtgca acccaagaag tgttgtacaa ccttggcatg 840
gcgcctgctt ggggagcaga gcacggccaa gacgcggtgc gttaa 885
<210> 3
<211> 294
<212> PRT
<213> Artificial Sequence
<220>
<223> 5,10-метилентетрагидрофолатредуктаза дикого типа
<400> 3
Met Pro Pro Arg Asp Asp Ala Ala Glu Glu Arg Leu Tyr Arg Ala Ala
1 5 10 15
Glu Val Phe His Asp Leu Gly Ala Ser Phe Val Ser Val Thr Tyr Gly
20 25 30
Ala Gly Gly Ser Thr Arg Glu Arg Thr Ser Arg Ile Ala Arg Arg Leu
35 40 45
Ala Lys Gln Pro Leu Thr Thr Leu Val His Leu Thr Leu Val Asn His
50 55 60
Thr Arg Glu Glu Met Lys Ala Ile Leu Arg Glu Tyr Leu Glu Leu Gly
65 70 75 80
Leu Thr Asn Leu Leu Ala Leu Arg Gly Asp Pro Pro Gly Asp Pro Leu
85 90 95
Gly Asp Trp Val Ser Thr Asp Gly Gly Leu Asn Tyr Ala Ser Glu Leu
100 105 110
Ile Asp Leu Ile Lys Ser Thr Pro Glu Phe Arg Glu Phe Asp Leu Gly
115 120 125
Ile Ala Ser Phe Pro Glu Gly His Phe Arg Ala Lys Thr Leu Glu Glu
130 135 140
Asp Thr Lys Tyr Thr Leu Ala Lys Leu Arg Gly Gly Ala Glu Tyr Ser
145 150 155 160
Ile Thr Gln Met Phe Phe Asp Val Glu Asp Tyr Leu Arg Leu Arg Asp
165 170 175
Arg Leu Val Ala Ala Asp Pro Ile His Gly Ala Lys Pro Ile Ile Pro
180 185 190
Gly Ile Met Pro Ile Thr Ser Leu Arg Ser Val Arg Arg Gln Val Glu
195 200 205
Leu Ser Gly Ala Gln Leu Pro Ser Gln Leu Glu Glu Ser Leu Val Arg
210 215 220
Ala Ala Asn Gly Asn Glu Glu Ala Asn Lys Asp Glu Ile Arg Lys Val
225 230 235 240
Gly Ile Glu Tyr Ser Thr Asn Met Ala Glu Arg Leu Ile Ala Glu Gly
245 250 255
Ala Glu Asp Leu His Phe Met Thr Leu Asn Phe Thr Arg Ala Thr Gln
260 265 270
Glu Val Leu Tyr Asn Leu Gly Met Ala Pro Ala Trp Gly Ala Glu His
275 280 285
Gly Gln Asp Ala Val Arg
290
<210> 4
<211> 885
<212> DNA
<213> Artificial Sequence
<220>
<223> ген, кодирующий 5,10-метилентетрагидрофолатредуктазу дикого типа
<400> 4
atgccacccc gcgatgatgc agctgaagag cgtctttacc gagcagcaga ggtcttccat 60
gacctcggtg catcgtttgt ctccgtgact tatggtgctg gcggatcaac ccgtgagaga 120
acctcacgta ttgctcgacg attagcgaaa caaccgttga ccactctggt gcacctgacc 180
ctggttaacc acactcgcga agagatgaag gcaattcttc gggaatacct agagctggga 240
ttaacaaacc tgttggcgct tcgaggagat ccgcctggag acccattagg cgattgggtg 300
agcaccgatg gaggactgaa ctacgcctct gagctcatcg atcttattaa gtccactcct 360
gagttccggg aattcgacct cggtatcgcc tccttccccg aagggcattt ccgggcgaaa 420
actctagaag aagacaccaa atacactctg gcgaagctgc gtggaggtgc agagtactcc 480
atcacgcaga tgttctttga tgtggaagac tacctgcgac ttcgtgatcg ccttgtcgct 540
gcagacccca ttcatggcgc gaagccaatc attcctggca tcatgcccat tacgagcctg 600
cggtctgtgc gtcgacaggt cgaactctct ggtgctcaat tgccgagcca actagaagaa 660
tcacttgttc gagctgcaaa cggcaatgaa gaagcgaaca aagacgagat ccgcaaggtg 720
ggcattgaat attccaccaa tatggcagag cgactcattg ccgaaggtgc ggaagatctg 780
cacttcatga cgcttaactt cacccgtgca acccaagaag tgttgtacaa ccttggcatg 840
gcgcctgctt ggggagcaga gcacggccaa gacgcggtgc gttaa 885
<210> 5
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> metF_1F
<400> 5
tcgagctcgg tacccttcag aagaattctt atggcc 36
<210> 6
<211> 44
<212> DNA
<213> Artificial Sequence
<220>
<223> metF_2R
<400> 6
ctcttcagct gcatcatcgt ggggtggcat aaactcgaca gaaa 44
<210> 7
<211> 44
<212> DNA
<213> Artificial Sequence
<220>
<223> metF_3F
<400> 7
tttctgtcga gtttatgcca ccccacgatg atgcagctga agag 44
<210> 8
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> metF_4R
<400> 8
ctctagagga tccccatcaa agaacatctg cgtgat 36
<210> 9
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> metF_5F
<400> 9
ttcagaagaa ttcttatggc c 21
<210> 10
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> metF_6R
<400> 10
atcaaagaac atctgcgtga t 21
<210> 11
<211> 5803
<212> DNA
<213> Artificial Sequence
<220>
<223> pDCM2
<400> 11
gttcgcttgc tgtccataaa accgcccagt ctagctatcg ccatgtaagc ccactgcaag 60
ctacctgctt tctctttgcg cttgcgtttt cccttgtcca gatagcccag tagctgacat 120
tcatccgggg tcagcaccgt ttctgcggac tggctttcta cgtgttccgc ttcctttagc 180
agcccttgcg ccctgagtgc ttgcggcagc gtgaagctag cttttatcgc cattcgccat 240
tcaggctgcg caactgttgg gaagggcgat cggtgcgggc ctcttcgcta ttacgccagc 300
tggcgaaagg gggatgtgct gcaaggcgat taagttgggt aacgccaggg ttttcccagt 360
cacgacgttg taaaacgacg gccagtgaat tcgagctcgg tacccgggga tcctctagag 420
tcgacctgca ggcatgcaag cttggcgtaa tcatggtcat agctgtttcc tgtgtgaaat 480
tgttatccgc tcacaattcc acacaacata cgagccggaa gcataaagtg taaagcctgg 540
ggtgcctaat gagtgagcta actcacatta attgcgttgc gctcactgcc cgctttccag 600
tcgggaaacc tgtcgtgcca gctgcattaa tgaatcggcc aacgcgcggg gagaggcggt 660
ttgcgtattg ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc ggtcgttcgg 720
ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac agaatcaggg 780
gataacgcag gaaagaacat gtgagcaaaa ggccagcaaa aggccaggaa ccgtaaaaag 840
gccgcgttgc tggcgttttt ccataggctc cgcccccctg acgagcatca caaaaatcga 900
cgctcaagtc agaggtggcg aaacccgaca ggactataaa gataccaggc gtttccccct 960
ggaagctccc tcgtgcgctc tcctgttccg accctgccgc ttaccggata cctgtccgcc 1020
tttctccctt cgggaagcgt ggcgctttct caatgctcac gctgtaggta tctcagttcg 1080
gtgtaggtcg ttcgctccaa gctgggctgt gtgcacgaac cccccgttca gcccgaccgc 1140
tgcgccttat ccggtaacta tcgtcttgag tccaacccgg taagacacga cttatcgcca 1200
ctggcagcag ccactggtaa caggattagc agagcgaggt atgtaggcgg tgctacagag 1260
ttcttgaagt ggtggcctaa ctacggctac actagaagga cagtatttgg tatctgcgct 1320
ctgctgaagc cagttacctt cggaaaaaga gttggtagct cttgatccgg caaacaaacc 1380
accgctggta gcggtggttt ttttgtttgc aagcagcaga ttacgcgcag aaaaaaagga 1440
tctcaagaag atcctttgat cttttctacg gggtctgacg ctcagtggaa cgaaaactca 1500
cgttaaggga ttttggtcat gagattatca aaaaggatct tcacctagat ccttttgggg 1560
tgggcgaaga actccagcat gagatccccg cgctggagga tcatccagcc ctgatagaaa 1620
cagaagccac tggagcacct caaaaacacc atcatacact aaatcagtaa gttggcagca 1680
tcacccgacg cactttgcgc cgaataaata cctgtgacgg aagatcactt cgcagaataa 1740
ataaatcctg gtgtccctgt tgataccggg aagccctggg ccaacttttg gcgaaaatga 1800
gacgttgatc ggcacgtaag aggttccaac tttcaccata atgaaataag atcactaccg 1860
ggcgtatttt ttgagttatc gagattttca ggagctgata gaaacagaag ccactggagc 1920
acctcaaaaa caccatcata cactaaatca gtaagttggc agcatcaccc gacgcacttt 1980
gcgccgaata aatacctgtg acggaagatc acttcgcaga ataaataaat cctggtgtcc 2040
ctgttgatac cgggaagccc tgggccaact tttggcgaaa atgagacgtt gatcggcacg 2100
taagaggttc caactttcac cataatgaaa taagatcact accgggcgta ttttttgagt 2160
tatcgagatt ttcaggagct ctttggcatc gtctctcgcc tgtcccctca gttcagtaat 2220
ttcctgcatt tgcctgtttc cagtcggtag atattccaca aaacagcagg gaagcagcgc 2280
ttttccgctg cataaccctg cttcggggtc attatagcga ttttttcggt atatccatcc 2340
tttttcgcac gatatacagg attttgccaa agggttcgtg tagactttcc ttggtgtatc 2400
caacggcgtc agccgggcag gataggtgaa gtaggcccac ccgcgagcgg gtgttccttc 2460
ttcactgtcc cttattcgca cctggcggtg ctcaacggga atcctgctct gcgaggctgg 2520
ccggctaccg ccggcgtaac agatgagggc aagcggatgg ctgatgaaac caagccaacc 2580
aggaagggca gcccacctat caaggtgtac tgccttccag acgaacgaag agcgattgag 2640
gaaaaggcgg cggcggccgg catgagcctg tcggcctacc tgctggccgt cggccagggc 2700
tacaaaatca cgggcgtcgt ggactatgag cacgtccgcg agggcgtccc ggaaaacgat 2760
tccgaagccc aacctttcat agaaggcggc ggtggaatcg aaatctcgtg atggcaggtt 2820
gggcgtcgct tggtcggtca tttcgaaaaa ggttaggaat acggttagcc atttgcctgc 2880
ttttatatag ttcantatgg gattcacctt tatgttgata agaaataaaa gaaaatgcca 2940
ataggatatc ggcattttct tttgcgtttt tatttgttaa ctgttaattg tccttgttca 3000
aggatgctgt ctttgacaac agatgttttc ttgcctttga tgttcagcag gaagctcggc 3060
gcaaacgttg attgtttgtc tgcgtagaat cctctgtttg tcatatagct tgtaatcacg 3120
acattgtttc ctttcgcttg aggtacagcg aagtgtgagt aagtaaaggt tacatcgtta 3180
ggcggatcaa gatccatttt taacacaagg ccagttttgt tcagcggctt gtatgggcca 3240
gttaaagaat tagaaacata accaagcatg taaatatcgt tagacgtaat gccgtcaatc 3300
gtcatttttg atccgcggga gtcagtgaac aggtaccatt tgccgttcat tttaaagacg 3360
ttcgcgcgtt caatttcatc tgttactgtg ttagatgcaa tcagcggttt catcactttt 3420
ttcagtgtgt aatcatcgtt tagctcaatc ataccgagag cgccgtttgc taactcagcc 3480
gtgcgttttt tatcgctttg cagaagtttt tgactttctt gacggaagaa tgatgtgctt 3540
ttgccatagt atgctttgtt aaataaagat tcttcgcctt ggtagccatc ttcagttcca 3600
gtgtttgctt caaatactaa gtatttgtgg cctttatctt ctacgtagtg aggatctctc 3660
agcgtatggt tgtcgcctga gctgtagttg ccttcatcga tgaactgctg tacattttga 3720
tacgtttttc cgtcaccgtc aaagattgat ttataatcct ctacaccgtt gatgttcaaa 3780
gagctgtctg atgctgatac gttaacttgt gcagttgtca gtgtttgttt gccgtaatgt 3840
ttaccggaga aatcagtgta gaataaacgg atttttccgt cagatgtaaa tgtggctgaa 3900
cctgaccatt cttgtgtttg gtcttttagg atagaatcat ttgcatcgaa tttgtcgctg 3960
tctttaaaga cgcggccagc gtttttccag ctgtcaatag aagtttcgcc gactttttga 4020
tagaacatgt aaatcgatgt gtcatccgca tttttaggat ctccggctaa tgcaaagacg 4080
atgtggtagc cgtgatagtt tgcgacagtg ccgtcagcgt tttgtaatgg ccagctgtcc 4140
caaacgtcca ggccttttgc agaagagata tttttaattg tggacgaatc aaattcagaa 4200
acttgatatt tttcattttt ttgctgttca gggatttgca gcatatcatg gcgtgtaata 4260
tgggaaatgc cgtatgtttc cttatatggc ttttggttcg tttctttcgc aaacgcttga 4320
gttgcgcctc ctgccagcag tgcggtagta aaggttaata ctgttgcttg ttttgcaaac 4380
tttttgatgt tcatcgttca tgtctccttt tttatgtact gtgttagcgg tctgcttctt 4440
ccagccctcc tgtttgaaga tggcaagtta gttacgcaca ataaaaaaag acctaaaata 4500
tgtaaggggt gacgccaaag tatacacttt gccctttaca cattttaggt cttgcctgct 4560
ttatcagtaa caaacccgcg cgatttactt ttcgacctca ttctattaga ctctcgtttg 4620
gattgcaact ggtctatttt cctcttttgt ttgatagaaa atcataaaag gatttgcaga 4680
ctacgggcct aaagaactaa aaaatctatc tgtttctttt cattctctgt attttttata 4740
gtttctgttg catgggcata aagttgcctt tttaatcaca attcagaaaa tatcataata 4800
tctcatttca ctaaataata gtgaacggca ggtatatgtg atgggttaaa aaggatcacc 4860
ccagagtccc gctcagaaga actcgtcaag aaggcgatag aaggcgatgc gctgcgaatc 4920
gggagcggcg ataccgtaaa gcacgaggaa gcggtcagcc cattcgccgc caagctcttc 4980
agcaatatca cgggtagcca acgctatgtc ctgatagcgg tccgccacac ccagccggcc 5040
acagtcgatg aatccagaaa agcggccatt ttccaccatg atattcggca agcaggcatc 5100
gccatgggtc acgacgagat cctcgccgtc gggcatccgc gccttgagcc tggcgaacag 5160
ttcggctggc gcgagcccct gatgctcttc gtccagatca tcctgatcga caagaccggc 5220
ttccatccga gtacgtgctc gctcgatgcg atgtttcgct tggtggtcga atgggcaggt 5280
agccggatca agcgtatgca gccgccgcat tgcatcagcc atgatggata ctttctcggc 5340
aggagcaagg tgagatgaca ggagatcctg ccccggcact tcgcccaata gcagccagtc 5400
ccttcccgct tcagtgacaa cgtcgagaca gctgcgcaag gaacgcccgt cgtggccagc 5460
cacgatagcc gcgctgcctc gtcttggagt tcattcaggg caccggacag gtcggtcttg 5520
acaaaaagaa ccgggcgccc ctgcgctgac agccggaaca cggcggcatc agagcagccg 5580
attgtctgtt gtgcccagtc atagccgaat agcctctcca cccaagcggc cggagaacct 5640
gcgtgcaatc catcttgttc aatcatgcga aacgatcctc atcctgtctc ttgatcagat 5700
cttgatcccc tgcgccatca gatccttggc ggcaagaaag ccatccagtt tactttgcag 5760
ggcttcccaa ccttaccaga gggcgcccca gctggcaatt ccg 5803
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Новый вариант аспарагинсинтазы и способ получения L-валина с его применением | 2021 |
|
RU2793442C1 |
| Новый вариант пролиндегидрогеназы и способ получения L-валина с его применением | 2021 |
|
RU2795067C1 |
| Новый вариант цистеинсульфинат-десульфиназы и способ получения L-валина с его применением | 2021 |
|
RU2792650C1 |
| Новый вариант 2-изопропилмалатсинтазы и способ получения L-валина с его применением | 2021 |
|
RU2794279C1 |
| Новый вариант белка и способ получения L-валина с его применением | 2021 |
|
RU2793440C1 |
| НОВЫЙ ВАРИАНТ ТЕТРАГИДРОДИПИКОЛИНАТ-N-СУКЦИНИЛТРАНСФЕРАЗЫ И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2793468C1 |
| НОВЫЙ ВАРИАНТ АТФ-ФОСФОРИБОЗИЛТРАНСФЕРАЗЫ И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2793399C1 |
| НОВЫЙ ВАРИАНТ БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2793447C1 |
| НОВЫЙ ВАРИАНТ ТРАНСПОРТЕРА ИЗ СЕМЕЙСТВА MFS САХАРНЫХ ПОРТЕРОВ И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2796590C1 |
| НОВЫЙ ВАРИАНТ ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТДЕГИДРОГЕНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2792640C1 |
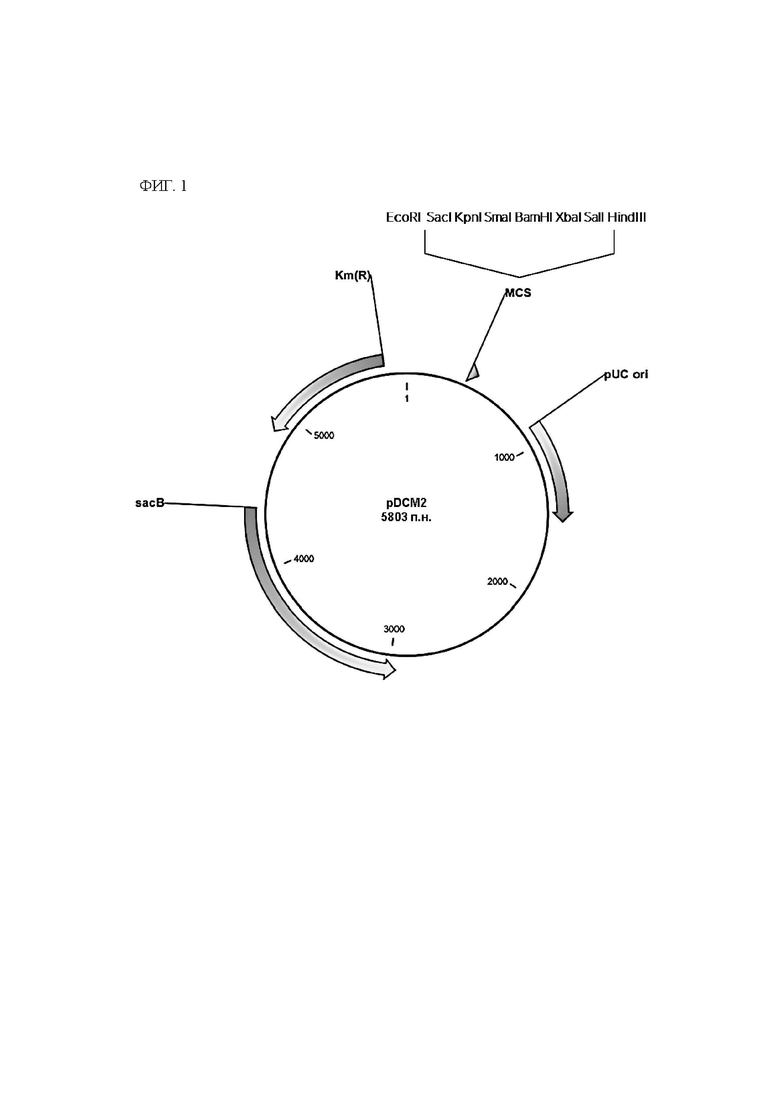
Изобретение относится к биотехнологии. Предложен полипептид, участвующий в продуцировании L-валина, состоящий из аминокислотной последовательности, представленной SEQ ID NO: 1, в которой аргинин, который представляет собой аминокислоту, соответствующую положению 4 SEQ ID NO: 3, заменен гистидином. Также предложены полинуклеотид, кодирующий указанный полипептид, микроорганизм Corynebacterium glutamicum, который продуцирует L-валин и содержит указанный полипептид или полинуклеотид, кодирующий указанный полипептид, и способ продуцирования L-валина. Изобретение позволяет увеличить выход L-валина. 4 н. и 1 з.п. ф-лы, 1 ил., 3 табл., 3 пр.
1. Полипептид, участвующий в продуцировании L-валина, состоящий из аминокислотной последовательности, представленной SEQ ID NO: 1, в которой аргинин, который представляет собой аминокислоту, соответствующую положению 4 SEQ ID NO: 3, заменен гистидином.
2. Полинуклеотид, кодирующий полипептид по п. 1.
3. Микроорганизм Corynebacterium glutamicum для продуцирования L-валина, содержащий полипептид по п. 1 или полинуклеотид, кодирующий данный полипептид.
4. Микроорганизм по п. 3, который имеет повышенную способность к продуцированию L-валина по сравнению с Corynebacterium glutamicum, содержащим полипептид SEQ ID NO: 3 или полинуклеотид, кодирующий данный полипептид.
5. Способ продуцирования L-валина, включающий культивирование микроорганизма Corynebacterium glutamicum, содержащего полипептид по п. 1 или полинуклеотид, кодирующий данный полипептид, в среде.
| База данных NCBI Reference Sequence: WP_011014930.1, 19.06.2019 | |||
| MULTISPECIES: methylenetetrahydrofolate reductase [NAD(P)H] [Corynebacterium] | |||
| Найдено онлайн: https://www.ncbi.nlm.nih.gov/protein/WP_011014930.1?report=genbank&log$=prottop&blast_rank=3&RID=KSK8M16C016 Дата обращения 04.10.2022 | |||
| US 8465962 B2, 18.06.2013 | |||
| US 20170051324 A1, |

