Группа изобретений относится к биотехнологии и медицине, а именно к новому полипептидному ингибитору α-амилаз млекопитающих из. морской анемоны Heteractis magnifica (Mgf-II), рекомбинантному способу его получения и применению в качестве средства для лечения и/или профилактики постпрандиальной гипергликемии, коррекции лишнего веса и ожирения.
По данным Всемирной организации здравоохранения на сегодняшний день от диабета страдают 422 миллиона жителей планеты, 90% которых больны инсулиннезависимым СД2Т. Данный тип заболевания почти всегда развивается у людей с излишним весом и ожирением. Распространенность СД возрастает быстрее в странах со средним и низким уровнем дохода, а в последние годы стремительно растет количество случаев заболевания среди детей и подростков. Полные пациенты, в т.ч. страдающие диабетом, имеют патологические изменения микро- и макроциркуляторного русла, обусловливающие такие патологии как ретинопатия, нейропатия, нефропатия, атеросклероз сосудов сердца и конечностей. Глобальные изменения наблюдаются и на различных уровнях врожденного и адаптивного иммунного ответа в связи с хроническим фоновым воспалением, что делает данную группу людей более уязвимыми к действию патогенов и повышает риск тяжелого протекания инфекционных заболеваний.
Традиционно, в терапии сахарного диабета 1 и 2 типов основное внимание уделялось контролю показателей уровня глюкозы натощак, но в настоящее время также признана важность снижения показателей постпрандиальной (после еды) глюкозы [Hanefeld М et al. Acarbose reduces the risk for myocardial infarction in type 2 diabetic patients: Meta-analysis of seven long-term studies. Eur. Heart J. 2004, 25, 10-16; Sorkin J.D., et al. The relation of fasting and 2-h postchallenge plasma glucose concentrations to mortality: data from the Baltimore Longitudinal Study of Aging with a critical review of the literature. Diabetes Care 2005, 28, 2626-32]. Для терапии СД2Т на ранних стадиях применяют диетотерапию и пероральные сахароснижающие препараты. Производные сульфонилмочевины (например, Глибенкламид) и глиниды (например, Старликс) стимулируют секрецию инсулина β-клетками островков Лангерганса. Однако, через несколько лет (в среднем около 5) такие препараты приводят к гибели β-клеток, что делает необходимым регулярные инъекции инсулина. Помимо этого, применение производных сульфонилмочевины способствует увеличению массы тела из-за переедания в результате гиперинсулинемии. К побочным эффектам этих препаратов относят вероятность наступления гипогликемии [Inzucchi S., et al. Management of hyperglycaemia in type 2 diabetes, 2015: a patient-centred approach. Update to a position statement of the American Diabetes Association and the European Association for the Study of Diabetes. Diabetologia 2015, 58, 429-42].
Бигуаниды (метформин) оказывают свое влияние на углеводный обмен путем воздействия на гепатоциты (торможение глюконеогенеза), повышения чувствительности тканей к инсулину (преимущественно поперечно-полосатой мускулатуры и жировой ткани), а также замедления всасывания глюкозы в кишечнике. К редким (1:100000) побочным эффектам метформина относится вероятность наступления смертельно опасного лактат-ацидоза. К плюсам можно отнести отсутствие риска гипогликемии, а также способность угнетать аппетит, что приводит к потере веса. Производные сульфонилмочевины, глиниды и бигуаниды можно применять только для пациентов с СД2Т, пациентам с СД1Т они противопоказаны [Inzucchi S., et al. Management of hyperglycaemia in type 2 diabetes, 2015: a patient-centred approach. Update to a position statement of the American Diabetes Association and the European Association for the Study of Diabetes. Diabetologia 2015, 58, 429-42].
В качестве пероральных сахароснижающих средств для всех диабетических пациентов могут быть использованы ингибиторы α-глюкозидаз, глюкозоснижающее действие которых, в отличие от других антидиабетических препаратов, находится вне спектра гормональной регуляции углеводного обмена. Ингибиторы α-глюкозидаз препятствуют гидролизу сложных углеводов до моносахаров, нарушая всасывание глюкозы из кишечника и развитие постпрандиальной гипергликемии и, как следствие, гиперинсулинемии. Ингибирование α-глюкозидаз, в частности α-амилаз, позволяет эффективно контролировать уровень постпрандиальной глюкозы в крови, препятствуя возникновению побочных эффектов, ассоциированных с глюкозотоксичностью.
Дополнительное преимущество таких препаратов заключается в достоверном снижении риска развития сердечно-сосудистых осложнений у пациентов с СД2Т, по сравнению со стимуляторами секреции инсулина [Hanefeld M et al. Acarbose reduces the risk for myocardial infarction in type 2 diabetic patients: Meta-analysis of seven long-term studies. Eur. Heart J. 2004, 25, 10-16]. Применение ингибиторов α-глюкозидаз приводит к нормализации нарушений углеводного и липидного обмена и может быть рекомендовано не только при лечении СД, но и для ранней коррекции нарушений обмена, в том числе для борьбы с излишним весом и ожирением.
Известны несколько олигосахаридных ингибиторов α-глюкозидаз, которые применяются в качестве фармакологических препаратов: миглитол (Glyset™), воглибоза (Voglib™) и акарбоза (Precose™ или Glucobay™). Первые два соединения ингибируют α-глюкозидазы и не оказывают существенного влияния на α-амилазу, в то время как акарбоза ингибирует оба типа ферментов. Таким образом, все три препарата препятствуют полному гидролизу сложных углеводов, однако образующиеся в процессе гидролиза олигосахариды и сами ингибиторы могут ферментироваться бактериями кишечника, что приводит к возникновению диареи и метеоризма. Кроме того, эти ингибиторы хорошо абсорбируются тканями организма, что является нежелательным побочным эффектом, так как действие данных препаратов должно ограничиваться кишечником [Scheen A. J. Drug treatment of non-insulin-dependent diabetes mellitus in the 1990s. Achievements and future developments. Drugs 1997, 54, 355-68; Scott L. J. & Spencer C. M. Miglitol: a review of its therapeutic potential in type 2 diabetes mellitus. Drugs 2000, 59, 521-49; Scheen A. J. Is there a role for alpha-glucosidase inhibitors in the prevention of type 2 diabetes mellitus? Drugs 2003, 63, 933-51]. Все перечисленные соединения обладают средней ингибиторной активностью и для достижения эффекта требуется довольно большая дозировка, сопровождающаяся умеренными побочными эффектами, преимущественно со стороны желудочно-кишечного тракта. Предполагается, что селективные ингибиторы α-амилаз будут обладать меньшими побочными эффектами и большей эффективностью.
Поиск новых ингибиторов α-амилаз активно проводится в природных источниках, таких как лизаты бактерий, экстракты лекарственных растений и животных [RU 2355755, CN 101804045 A, CN 103520374 A, CN 105943625 A, CN 107312106 A, CN 107827992 A, CN 108434177 A, WO 2021248601 A1, US 5726291 A, US 2003124208 A1, JP 2017008096 A, JP 2016121110 A, CN 110279730 A, CN 112971154 A, СТ 113197281 А, CN 113274403 A, CN 113318143 A]. Так, наиболее изученный ингибитор α-амилаз акарбоза был выделен из бактерии Streptomyces dimorphogenes [Yokose K., et al. New alpha-amylase inhibitor, trestatins. I. Isolation, characterization and biological activities of trestatins A, B and C. J. Antibiot. (Tokyo). 1983, 36, 1157-65]. Акарбоза является слабым и неспецифичным ингибитором α-амилаз, константа ингибирования (Ki) свиной панкреатической α-амилазы (PPA) составляет 0,797 мкМ [Yoon S. H. & Robyt J. F. Study of the inhibition of four alpha amylases by acarbose and its 4IV-α-maltohexaosyl and 4IV-α-maltododecaosyl analogues. Carbohydr. Research 2003, 338, 1969-1980]. Более эффективным ингибитором является гликозилированный флавоноид монтбредин А, выделенный из растения семейства ирисовых Crocosmia crocosmiiflora, который ингибирует α-амилазу в наномолярных концентрациях (Ki=8 нМ) [Williams L., et al. The amylase inhibitor montbretin A reveals a new glycosidase inhibition motif. Nature Chem. Biol. 2015, 11, 691-696]. Исследование активности монтбредина А показало, что он эффективно уменьшает уровень глюкозы в крови мышей в модели СД2Т, снижает уровень холестерина, нормализует функции печени. Однако, эффективная доза монтбредина А в исследованиях in vivo составляет 7,5 мг/кг, что практически не отличается от эффективной дозы для акарбозы (10 мг/кг), несмотря на стократную разницу в значении Ki [Yuen V. et al. Glucose lowering effect of montbretin A in Zucker Diabetic Fatty rats. Molecular Cell. Biochem. 2016, 411, 373-381].
Хорошо известно, что пептиды обладают высокой биологической активностью в низких дозах. Так, из морской анемоны Stichodactyla helianthus был выделен эффективный и селективный ингибитор α-амилаз млекопитающих - хелиантамид, полипептид, состоящий из 44 аминокислотных остатков. Он ингибирует панкреатическую α-амилазу человека и свиньи с Ki 0,01 и 0,1 нМ соответственно [Tysoe C., et al. Potent human α-amylase inhibition by the β-defensin-like protein helianthamide. ACS Cent. Science 2016, 2, 154-161; WO 2017/000060 A1].
Еще одним представителем пептидных ингибиторов α-амилаз является магнификамид, выделенный из H. magnifica [Sintsova O., et al. Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a new β-defensin α-amylase inhibitor. J. Proteomics 2018, 173, 12-21]. Он является ближайшим аналогом заявляемого полипептида (идентичность аминокислотных последовательностей составляет 95%) и гомологом хелиантамида (84% идентичности). Константа ингибирования свиной панкреатической α-амилазы магнификамидом составляет 0,17 нМ. Показано, что он также ингибирует человеческую слюнную α-амилазу (HSA) (Ki = 7,7 нМ) [Sintsova O., et al. Magnificamide, a β-Defensin-Like Peptide from the Mucus of the Sea Anemone Heteractis magnifica, Is a Strong Inhibitor of Mammalian α-Amylases. Mar. Drugs 2019, 17, 1-15]. Активность магнификамида не уступает активности хелиантамида и значительно превосходит активность акарбозы, константы ингибирования которой составляют 0,797 и 1,265 мкМ для PPA и HSA соответственно.
Так как хелиантамид и магнификамид принадлежат к семейству β-дефензинов, широко распространенных среди животных и человека, и обладают компактным фолдом, предполагается, что их иммуногенность должна быть минимальной. Данные полипептиды устойчивы к протеолизу, длительному воздействию низких значений pH и высокой температуры, что демонстрирует возможность их применения в качестве оральных сахароснижающих препаратов для контроля постпрандиальной гипергликемии [Tysoe C., et al. Potent human α-amylase inhibition by the β-defensin-like protein helianthamide. ACS Cent. Science 2016, 2, 154-161; WO 2017/000060 A1]. Более того, электрофизиологическое исследование на 12 подтипах потенциал-зависимых калиевых и шести подтипах потенциал-зависимых натриевых каналов, экспрессированных в ооцитах лягушки Xenopus laevis, включая каналы сердца (hERG), показало, что магнификамид не модулирует ионные каналы, следовательно, велика вероятность безопасности этих соединений для экспериментальных животных и человека.
Задача изобретения - расширение арсенала глюкозопонижающих лекарственных средств нового поколения, имеющих направленное действие на молекулярную мишень, а именно ингибирование α-амилаз млекопитающих, которые могут найти применение в медицине в качестве средств для лечения и/или профилактики постпрандиальной гипергликемии, коррекции лишнего веса и ожирения.
Поставленная задача решена получением нового полипептида из морской анемоны Heteractis magnifica, магнификамида-II (Mgf-II), ингибирующего α-амилазы млекопитающих, и имеющего следующую аминокислотную последовательность (SEQ ID NO: 2):
SEGNSCYIYHGVYGICKAKCAEDEKAMAGMGVCEGDLCCYKTPW
Mgf-II является родственным полипептидом хелиантамиду и магнификамиду. Наиболее близкой к Mgf-II структурой обладает магнификамид, выделенный из морской анемоны того же вида [Sintsova O., et al. Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a new β-defensin α-amylase inhibitor. J. Proteomics 2018, 173, 12-21]. Заявляемый полипептид, состоящий из 44 аминокислотных остатков, эффективно ингибирует α-амилазы млекопитающих и может быть выделен как из морской анемоны H. magnifica, так и получен рекомбинантным способом. Рекомбинантный Mgf-II полностью соответствует нативному полипептиду по структуре и биологической активности и может быть использован в качестве средства для лечения и/или профилактики постпрандиальной гипергликемии, коррекции лишнего веса и ожирения у млекопитающих, в частности человека.
Технический результат изобретения заключается в получении нового эффективного ингибитора α-амилаз млекопитающих, Mgf-II, при пероральном введении которого у животных с острым метаболическим синдромом по типу СД1Т наблюдается полная резистентность к крахмалу и уровень глюкозы крови остается неизменным; а также пригодного для коррекции веса и ожирения у животных и человека с клиническими признаками СД2Т.
Mgf-II эффективно снижает уровень постпрандиальной глюкозы крови, вес, а также массу и массовый индекс жировой ткани, как показано в тестах на диком типе мышей с СД1Т и СД2Т соответственно.
В тестах Mgf-II превосходит аналог магнификамид по следующим параметрам:
• более эффективное ингибирование свиной панкреатической и слюной человеческой амилаз;
• достоверное снижение уровня постпрандиальной глюкозы.
Mgf-II может быть использован как терапевтическое средство для ингибирования α-амилаз млекопитающих, эффективного лечения и/или профилактики постпрандиальной гиперкликемии, коррекции лишнего веса и ожирения.
Заявляемый полипептид Mgf-II получают путем 4-ех этапной хроматографической очистки из водно-этанольного экстракта морской анемоны Н. magnifica.
Заявляемый полипептид Mgf-II получают также в виде рекомбинантного белка генно-инженерными методами на основании нуклеотидной последовательности кодирующего его гена.
Для этого получают экспрессионную конструкцию на основе вектора рЕТ-32b(+), содержащую ген слитного белка TrxA-6His-magnificamid-II, который состоит из тиоредоксина, сайга для протеолиза энтерокиназой, шести остатков гистидина и Mgf-II, позволяющую производить контролируемую экспрессию слитного гена.
Способ получения Mgf-II й бактериальной системе Е. соli включает проведение контролируемой экспрессии гена слитного белка TrxA-6His-magnificamid-II в составе экспресеионной конструкции с использованием в качестве индуктора изопропил-β-D-1-тиогалактопиранозида, лизис клеток, отделение осадка, очистку слитного белка ТrхА-6His-magnificamid-II с помощью аффинной хроматографии на сорбенте Ni2+-NTA-агазоре, расщепление слитного белка TrxA-6His-magnificamid-II энтерокиназой, очистку полипептида Mgf-II с помощью обращенно-фазовой хроматографии.
Структура белка-предшественника заявляемого полипептида может быть выведена на основании полученной нуклеотидной последовательности кДНК, транскрибированной с матричной РНК, имеющей нижеследующую структуру (SEQ ID NO: 1):
gggcaagcagtggtatcaacgcagagtacgggggaataaaccatcagtggcgaaattctactactcactgggagctttacttatcagcaaaGGTGTGTACGGGATATGCAAGGCTAAGTGCGCGGAAGACGAAAAAGCCATGGCGGGAATGGGTGTGTGTGAAGGCGATCTCTGCTGTTATААААСACCATGGTAAtcatcggtggattttcgaccaggataatgaaatacaaaacaaataatt
Полученный ген кодирует в открытой рамке считывания 72 аминокислотных остатка и стоп кодон, где сигнальный пептид из 18 аминокислотных остатков предшествует структуре пробелка, содержащего прочасть из 10 аминокислотных остатков и зрелый пептид, который является заявляемым Mgf-II и состоит из 44 аминокислотных остатков. Mgf-II синтезируется в одном экземпляре (в расчете на один пробелок), и никаких посттрансляционных модификаций в его составе не обнаруживается.
Полная аминокислотная последовательность Mgf-II может быть установлена комбинацией методов секвенирования по Эдману и установления нуклеотидной последовательности кодирующего его гена. Последовательность, состоящая из 38 аминокислотных остатков, определяется автоматическим секвенированием алкилированного пептида по методу Эдмана, последовательность оставшихся шести С-концевых аминокислот выводится на основании последовательности кодирующего Mgf-II гена. Средняя измеренная молекулярная масса выделенного Mgf-II составляет 4779 Да. Расчетная масса отличается от измеренной на 0,41 Да. С учетом точности метода определения масс можно утверждать, что природный Mgf-II не имеет в своем составе модифицированных аминокислотных остатков.
Mgf-II облагает пространственной структурой, характерной для β-дефензинов, что предполагает наличие компактного фолда с тремя дисульфидными связями, который придает ему стабильность при введении в организм, обусловливает низкую иммуногенность, и, как следствие этого, улучшает потребительские свойства фармакологических препаратов на его основе, такие как длительность воздействия и простота использования.
Гомология первичной структуры заявляемого полипептида с магнификамидом, являющимся ингибитором α-амилаз млекопитающих, позволяет утверждать, что Mgf-II обеспечивает снижение уровня поспрадиальной глюкозы за счет ингибирующей активности в отношении α-амилаз млекопитающих. Высокий процент идентичности последовательностей магнификамидов позволяет предполагать одинаковую доменную организацию, но не идентичность биологической активности. Различия были показаны при установлении констант ингибирования α-амилаз и доказаны при проведении экспериментов на животных моделях. Так, Mgf-II превосходит магнификамид по эффективности ингибирования свиной панкреатической амилазы (константа ингибирования составляет 0,03 нМ) и слюной человеческой амилазы (константа ингибирования составляет 0,08 нМ), а также в крахмальном тесте. При пероральном введении Mgf-II в дозе 0,01 мг/кг у животных с острым метаболическим синдромом по типу СД1Т наблюдалось достоверное снижение уровня постпрандиальной глюкозы в течение 120 минут, по сравнению с магнификамидом в той же дозе и акарбозой в дозе 3 мг/кг, эффект которых не был достоверным относительно отрицательного контроля (физраствор).
Mgf-II может быть использован как терапевтическое средство для эффективного лечения постпрандиальной гипергликемии, а также как препарат для коррекции лишнего веса и ожирения за счет его способности полностью блокировать расщепление крахмала до моносахаридов, их всасывания в кровь и запасания в форме жировых отложений.
Изобретение иллюстрируют фигуры.
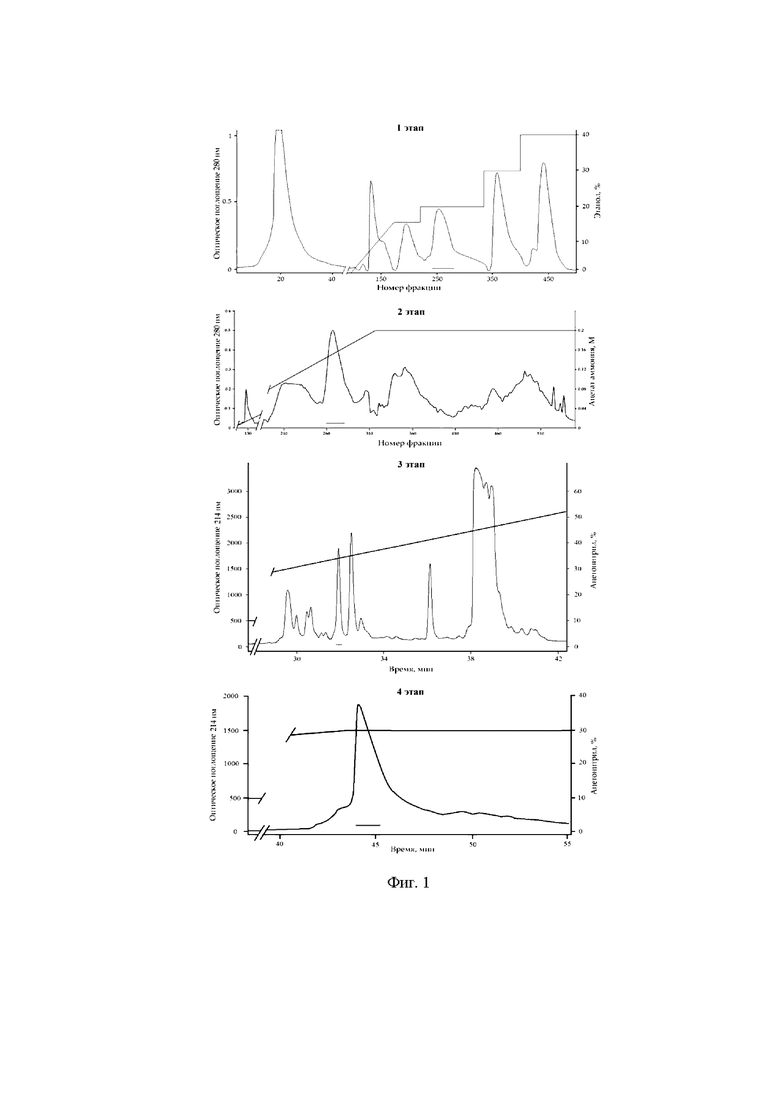
Фиг. 1. Основные стадии хроматографического выделения ингибитора α-амилазы, полипептида Mgf-II, из водно-зткнольного экстракта морской анемоны Н. magnifica. На 1 этапе используют колонку с гидрофобным носителем Полихромом-1 (48×950 мм), этанол и скорость элюирования 120 мл/час, на 2 этапе используют ионообменную колонку Bio-Rex 70 (25×600 мм), 0,2 М аммонийно-ацетатный буфер, pH 4,5, скорость элюции - 22 мл/час, на 3 этапе используют обращено-фазовую колонку Luna С18 (10×250 мм), ацетонитрил, содержащий 0,1% трифторуксусной кислоты и скорость элюции 3 мл/мин, на последнем, 4 этапе используют обращеннофазную колонку Nucleosil С18 (4,6×250 мм), ацетонитрил, содержащий 0,1% трифторуксусной кислоты, скорость элюции - 0,5 мл/мин. На фигуре показан градиент концентрации растворителя и отмечены границы объединения фракций, содержащих Mgf-II.
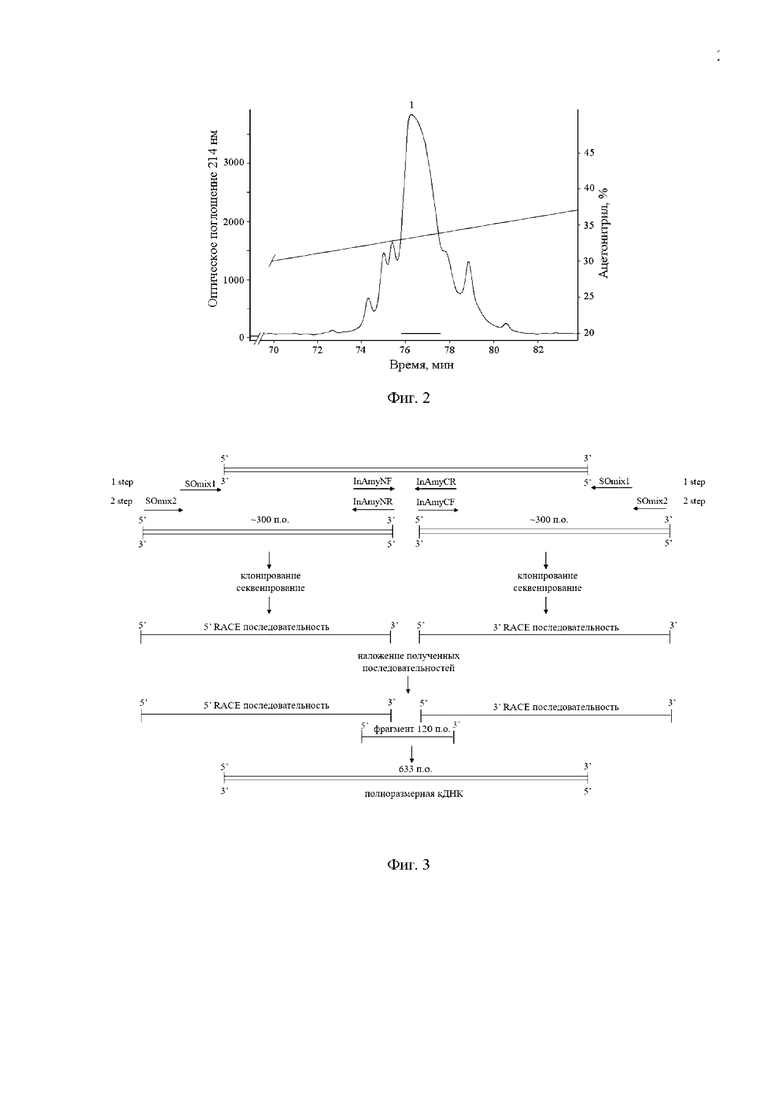
Фиг. 2. ВЭЖХ продуктов алкилирования Mgf-II на колонке с обращенной фазой (Nucleosil C18) 4,6×250 мм, уравновешенной 10% ацетонитрилом в 0,1% растворе трифторуксусной кислоты. Разделение проводят со скоростью 0,5 мл/мин в 10% ацетонитриле в изократическом режиме 30 мин, затем в линейном градиенте концентрации ацетонитрила 10-70% в течение 120 мин. Отмечены фракции, содержащие алкилированный Mgf-II.
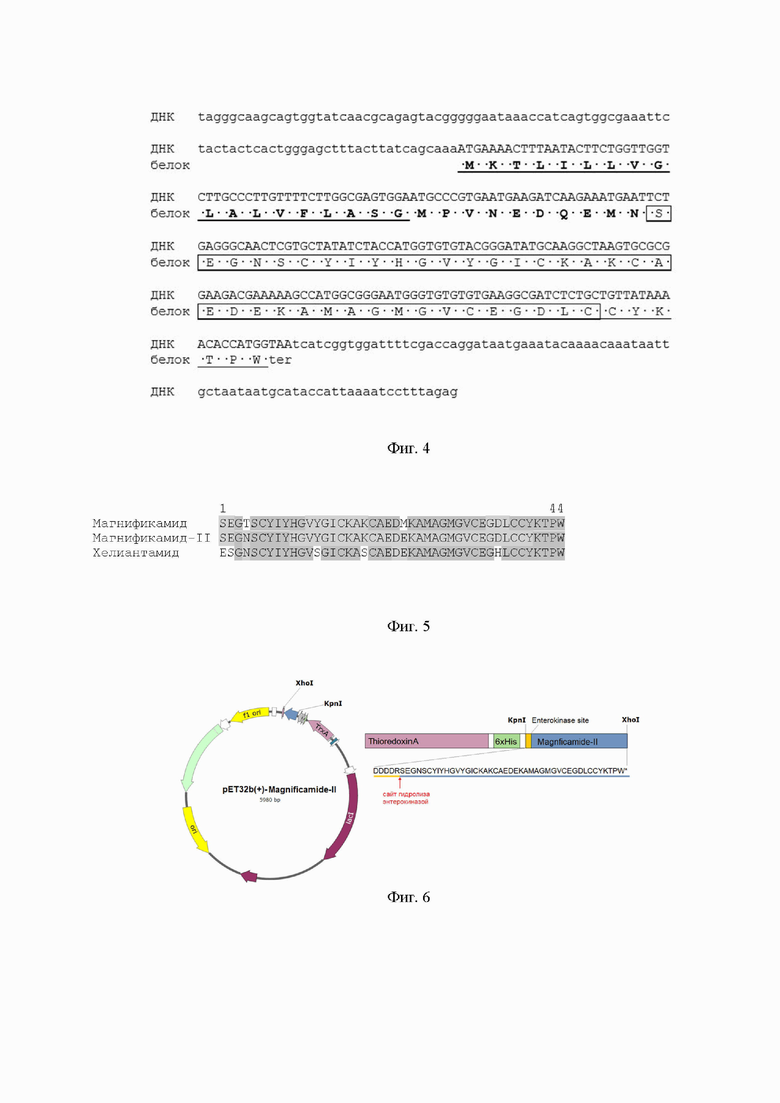
Фиг. 3. Схема проведения быстрой амплификации кДНК концов (Step-Out RACE). Стрелками показано направление стандартных (SOmix1 и SOmix2) и генспецифичных (InAmyNF, InAmyCF, InAmyNR, InAmyCR) праймеров. ПЦР проводят с указанными праймерами в два этапа. В результате получают фрагменты ДНК, которые при наложении и множественном выравнивании дают полноразмерную кДНК. Указан фрагмент, полученный с праймерами 1В и 2В (120 п.о.).
Фиг. 4. Структура нуклеотидной последовательности гена, кодирующего Mgf-II и аминокислотная последовательность его белка-предшественника. Последовательность выщепляемого сигнального пептида и пропептида выделена жирным шрифтом, зрелый полипептид, получаемый при созревании белка-предшественника подчеркнут, последовательность полипептида, соответствующая ранее полученной автоматическим методом секвенирования аминокислотной последовательности, выделена рамкой.
Фиг. 5. Сравнение аминокислотной последовательности Mgf-II и близких структурных гомологов, ингибиторов α-амилаз, относящихся по типу пространственной укладки полипептидной цепи к β-дефензинам. Идентичные аминокислотные остатки выделены серым цветом, различия показаны на белом фоне.
Фиг. 6. Схема экспрессионной конструкции pET32b(+)-Mgf-II и гибридного белка ТrxA-6His-magnificamid-II. Последовательность, содержащая ген Mgf-II, и сайт гидролиза энтерокиназой, клонирован в вектор рЕТ32b(+) по сайтам рестрикции KpnI и XhoI. Аминокислотная последовательность Mgf-II показана.
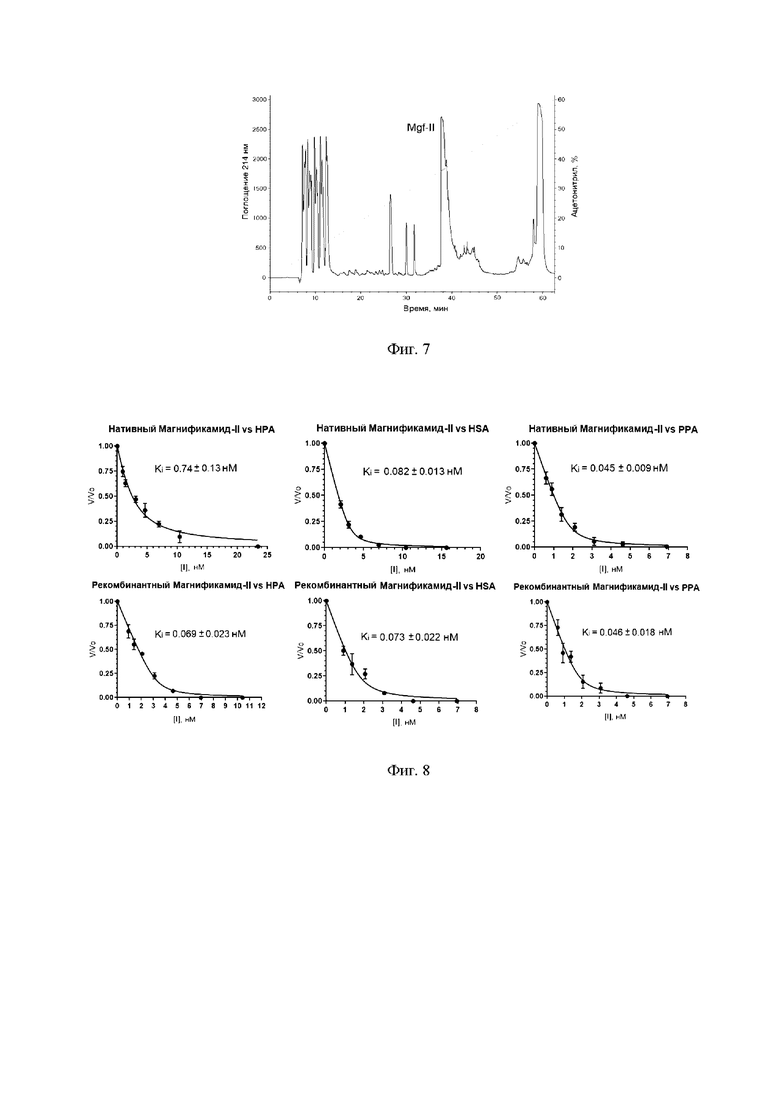
Фиг. 7. ВЭЖХ гидролизата 5 мг слитного белка на обращено-фазовой колонке Jupiter С4 (Phenomenex, США) 10×250 мм. Разделение со скоростью 3 мл/мин проводят в линейном градиенте концентрации ацетонитрила (0-70%) в присутствии 0,1% трифторуксусной кислоты. Детекция при длине волны 214 нм. Фракция, содержащая Mgf-II, обозначена серый цветом.
Фиг. 8. Кривые ингибирования α-амилаз нативным и рекомбинантным Mgf-II. Ферменты; свиная панкреатическая (РРА, 20 нМ), человеческая слюнная (HSA, 100 нМ) и человеческая панкреатическая (HPА, 20 нМ) α-амилазы, инкубируют с Mgf-II в концентрациях от 1 до 10 нМ 10 мин при 37°С. Субстрат 2-хлор-4-нитрофенил-α-D-мальтотриозид в концентрации 1 мМ для РРА, 3,3 мМ для HSA и 1,7 мМ для HPА добавляют в реакционную смесь и выдерживают 10 мин при 37°С, pH буферного раствора 7,0. Продукт реакции детектируют спектрофотометрически при длине волны 405 нм. Представленные данные являются результатом трех независимых экспериментов.
Фиг. 9. Эффект снижения уровня глюкозы полипептида Mgf-II и прототипа - полипептида магнификамида, в дозах 0,005 и 0,01 мг/кг при перроральном введении в крахмальном тесте. Контрольные животные получают 3 мг/кг акарбозы (положительный контроль) или аналогичный объем физраствора (здоровые и отрицательный контроль). Достоверность различий вычисляют t-критерием Стьюдента, достоверным считают значение * - р<0,05, ** - р<0,01, по сравнению с группой отрицательного контроля.
Изобретение иллюстрируется примерами.
Пример 1
Выделение полипептида Mgf-II
Морские анемоны Heteractis magnifica гомогенизируют в холодной дистиллированной воде и прибавляют этиловый спирт до конечной концентрации 70%. Экстракцию проводят 24 час, после чего фильтруют через несколько слоев ткани. Тонкий осадок удаляют центрифугированием при 3500 g, а получаемый супернатант повторно центрифугируют при 8000 g. Супернатант лиофильно высушивают и хранят при температуре (-20)°С до последующего применения.
Выделение и очистку Mgf-II из этанольного экстракта проводят по схеме, включающей гидрофобную хроматографию на полихроме-1 (политетрафторэтилене) (этап 1); ионообменную хроматографию на носителе Bio-Rex 70 (этап 2) и ОФ-ВЭЖХ на колонках Luna 08 (3 этап) и Nucleosil С18 (4 этап) (фиг. 1). Детектирование проводят спектрофотометрически при длине волны 280 нм (этапы 1 и 2) и 214 нм (этапы 3 и 4). Активность промежуточных фракций подтверждают в тестах in vitro по способности полипептидов фракции ингибировать вызванный α-амилазой гидролиз субстрата, 2-хлор-4-нитрофенил-α-D-мальтотриозида (CNPG3), с высвобождением 2-хлор-4-нитрофенола, образование которого измеряется спектрофотометрически.
На первом этапе хроматографию экстракта проводят на колонке (48×950 мм) с гидрофобным носителем полихром-1 (ХромЛаб, Россия), который сорбирует биологически активные полипептиды морских анемон и не задерживает находящиеся в экстракте в значительных количествах гидрофильные примеси и морскую соль. Элюцию пептидов проводят водой, затем ступенчатым градиентом этанола 15, 20, 30 и 40% со скоростью 120 мл/час, объем фракций - 20 мл. Активную фракцию элюируют 20%-ным этанолом.
Ионообменную хроматографию проводят на колонке с носителем Bio-Rex 70 (25×600 мм), уравновешенной в 0,01 М аммонийно-ацетатном буферном растворе (pH 4,5). Активную фракцию элюируют в линейном градиенте аммонийно-ацетатного буферного раствора (0,01-0,2 М). Скорость элюции - 22 мл/час, объем фракций - 5,8 мл.
ОФ-ВЭЖХ осуществляют на колонке Luna С18 (10×250 мм, размер пор 100  , диаметр частиц 5, мкм, Phenomenex), уравновешенной 10% ацетонитрилом в буферном растворе, содержащем 0,1% трифторуксусной кислоты (ТФУ). Пептиды элюируют 10 мин 10% ацетонитрилом в 0,1% ТФУ, затем используют линейный градиент концентрации ацетонитрила 10-70% за 60 мин. Скорость элюции - 3 мл/мин. Финальную очистку Mgf-II осуществляют ОФ-ВЭЖХ на колонке Nucleosil С18 (4,6×250 мм, размер пор 100 , диаметр частиц 5 мкм, Supelco), уравновешенной 10% ацетонитрилом в буферном растворе, содержащем 0,1% ТФУ. Пептиды элюируют 3 мин 10% ацетонитрилом, затем используют линейный градиент концентрации ацетонитрила 10-30% за 40 мин, затем 30% ацетонитрил. Скорость элюции - 0,5 мл/мин. В результате получают гомогенный Mgf-II.
, диаметр частиц 5, мкм, Phenomenex), уравновешенной 10% ацетонитрилом в буферном растворе, содержащем 0,1% трифторуксусной кислоты (ТФУ). Пептиды элюируют 10 мин 10% ацетонитрилом в 0,1% ТФУ, затем используют линейный градиент концентрации ацетонитрила 10-70% за 60 мин. Скорость элюции - 3 мл/мин. Финальную очистку Mgf-II осуществляют ОФ-ВЭЖХ на колонке Nucleosil С18 (4,6×250 мм, размер пор 100 , диаметр частиц 5 мкм, Supelco), уравновешенной 10% ацетонитрилом в буферном растворе, содержащем 0,1% ТФУ. Пептиды элюируют 3 мин 10% ацетонитрилом, затем используют линейный градиент концентрации ацетонитрила 10-30% за 40 мин, затем 30% ацетонитрил. Скорость элюции - 0,5 мл/мин. В результате получают гомогенный Mgf-II.
Пример 2
Установление частичной аминокислотной последовательности Mgf-II
Для определения N-концевой аминокислотной последовательности Mgf-II тиольные группы остатков цистеина восстанавливают дитиотреитолом и алкилируют 4-винилпиридином. Лиофилизированный полипептид растворяют в 100 мкл буферного раствора 0,5 М Трис-НСI (pH 8,5), содержащего 2 мМ ЭДТА и 6 М гуанидингидрохлорид. Добавляют 4 мкл дитиотреитола и инкубируют 4 ч при температуре 40°С. Тиольные группы остатков цистеина алкилируют добавлением 4 мкл смеси изопропанола и 4-винилпиридина в ацетонитриле в соотношении 1:1 (v:v). Инкубируют в течение 20 мин при комнатной температуре без доступа света. К реакционной смеси добавляют 150 мкл 0,1% ТФУ и центрифугируют 10 мин при 10 000 об/мин. Смесь разделяют ВЭЖХ на колонке с обращенной фазой сорбента Nucleosil С18 (4,6×250 мм, размер пор 100 , диаметр частиц 5 мкм, Supelco), уравновешенной 10% ацетонитрилом в буферном растворе, содержащем 0,1% ТФУ (фиг. 2). Алкилированные полипептиды элюируют в градиенте концентрации ацетонитрила (в 0,1% ТФУ): 10% ацетонитрил в изократическом режиме 30 мин, затем в линейном градиенте ацетонитрила 10-70% в течение 120 мин. Скорость элюции - 0,5 мл/мин. Детектирование проводят спектрофотометрически при длине волны 214 нм.
Определение N-концевой аминокислотной последовательности алкилированого Mgf-II проводят методом ступенчатой деградации по Эдману на автоматическом секвенаторе Precise 492 cLC (Applied Biosystems, США) по программе производителя, При деградации полипептидной цепи устанавливают аминокислотную последовательность первых 38 аминокислотных остатков:
SEGNSCYIYHGVYGICKAKC AEDEKAMAGMGVCEGDLC
Пример 3
Установление структуры гена и полной аминокислотной последовательности Mgf-II
Суммарную мРНК получают из свежезамороженного (-70°С) или свежепрепарированного образца морской анемоны (50-100 мг) с использованием коммерческого набора для получения РНК ExtractRNA (ЗАО Евроген, Россия) согласно инструкции производителя. Образец гомогенизируют в 1 мл лизирующего буферного раствора для выделения РНК, инкубируют при комнатной температуре 10 мин. Полученную смесь центрифугируют при комнатной температуре (10 мин, 14000 g), к супернатанту добавляют 0,2 мл хлороформа, встряхивают и инкубируют 5 мин. Полученную смесь центрифугируют при температуре 4°С (15 мин, 12000 g) и отбирают верхнюю водную фазу. Последовательно добавляют 0,5 мл изопропанола и инкубируют при комнатной температуре 10 мин, центрифугируют при комнатной температуре (10 мин, 12000 g). К осадку добавляют 1 мл 75% этилового спирта и осаждают центрифугированием при комнатной температуре (5 мин, 12000 g). Осадок РНК подсушивают на воздухе и растворяют в 50 мкл дистиллированной воды, не содержащей РНКаз. Концентрацию РНК определяют епектрофотомегрически.
Синтез кДНК проводят с помощью коммерческого набора Mint (ЗАО Евроген, Россия) по протоколу производителя. Синтез первой цепи кДНК осуществляют с 10 мкМ 3’-праймером (5’-AAGCAGTGGTATCAACGCAGAGTAC(T)30VN -3’) и 15 мкМ PlugOligo адаптером (5’-AAGCAGTGGTATCAACGCAGAGTACGGGGG-P-3’) и 2 мкг суммарной РНК. Компоненты смешивают, доводят дистиллированной водой объем смеси до 5 мкл, инкубируют 2 мин при температуре 70°С и охлаждают до 42°С. Добавляют 2 мкл 5-кратного реакционного буфера (250 мМ Трис-НСl (pH 8,3 при 25°С), 250 мМ KСl, 20 мМ MgCl2), 1 мкл 20 мМ дитиотреитола, 5 мкл ингибитора рибонуклеаз (IP solusion), 1 мкл 10 мМ dNTP, 1 мкл обратной транскриптазы (Mint Reverse Transcriptase). Реакцию проводят 1,5 ч при температуре 42°С. Для синтеза двухцепочечной кДНК используют 40 мкл воды для ПНР, 5 мкл 10-кратного реакционного буфера, 1 мкл 10 мМ dNTP, 2 мкл 10 мМ праймера M1 (5’-AAGCAGTGGTATCAACGCAGAGT-3’), 1 мкл 50-кратной Encyclo полимеразы и 1 Мкл одноцепочечной кДНК. Амплификацию проводят при следующих условиях: денатурация 95°С 1 мин., элонгация 15 циклов 95°С - 15 с, 66°С - 20 с, 72°С 3 мин, финальная элонгация 66°С - 20 с, 72°С - 3 мин.
Для определения полной нуклеотидной последовательности гена, кодирующего белок-предшественник полипептида Mgf-II, синтезируют вырожденные праймеры, сконструированные на основании аминокислотных последовательностей хелиантамида, магнификамида и N-концевой: последовательности Mgf-II: 1B (ATG YTA YAT НТА YCA YGG YGT) и 2В (ACC ANG GNG TYT TRT ARC AGC А). Полимеразную цепную реакцию проводят при следующих условиях: денатурация 94°С 3 мин., элонгация 35 циклов 94°С - 10 с, 55°С - 15 с, 72°С 10 с, финальная элонгация 72°С - 15 мин. В качестве матрицы используют кДНК Н. magnifica. Получают фрагмент около 120 п. о., который клонируют в плазмиду pTZ57R/T (ЗАО Евроген). Нуклеотидную последовательность нескольких клонов определяют на генетическом анализаторе 3130xl Genetic Analyzer (Applied Biosystems). На основании полученной нуклеотидной последовательности синтезируют генспецифические праймеры InAmyNF (CGG GAT ATG САА GGC ТАА GT), InAmyCF (GGT GTG TGT GAA GGC GAT), InAmyNR (ACT TAG CCT TGC АТА TCC CG) и InAmyCR (АТС GCC TTC АСА CAC ACC) и проводят быструю амплификацию кДНК концов (Step-Out RACE) по схеме (фиг. 3).
Для этого получают продукты ПЦР с праймерами SOmix1 (ЗАО Евроген) и InAmyNF (в эксперименте по определению 3’-концевой последовательности) и InAmyCR (в эксперименте по определению 5’-концевой последовательности), которые используют в качестве матрицы при 100-кратном разведении для 2 раунда ПЦР с праймерами SOmix2 (ЗАО Евроген) и InAmyCF (в эксперименте по определению 3’-концевой последовательности) и InAmyNR (в эксперименте по определению 5’-концевой последовательности). Оба раунда ПЦР проводят при сходных условиях 94°С - 15 с, 55°С - 30 с, 72°С 3 мин, 25 циклов используя Encyclo полимеразу (ЗАО Евроген, Россия). Амплифицированные фрагменты около 350 п.о. клонируют в плазмиду pTZ57R/T (ЗАО Евроген). Нуклеотидные последовательности нескольких клонов определяют на генетическом анализаторе 3130xl Genetic Analyzer (Applied Biosystems). Полноразмерную структуру гена, кодирующего белок-предшественник, определяют после множественного выравнивания всех полученных последовательностей (фиг. 4).
Правильность конечной аминокислотной, последовательности подтверждают сравнением расчетных и измеренных молекулярных масс Mgf-II. Уникальность последовательности Mgf-II подтверждают сравнением с базой данных известных аминокислотных последовательностей UniProt КВ (фиг. 5).
Пример 4
Создание генно-инженерной конструкции для получения рекомбинантного Mgf-II
Для получения генно-инженерной конструкции, способной экспрессировать Mgf-II в клетках Escherichia coli, на основе нуклеотидной последовательности гена, кодирующего зрелый Mgf-II, синтезируют генспецифичные прямой F (5’-AAGGTACCGATGATGATGATCGTTCTGAGGGCAACTCGTGC-3’) и обратный R (5’-СAATATTTTGTGGTACCATTGAGCTСАС-3’)праймеры,содержащие последовательность, кодирующую сайт для энтерокиназы, стоп кодон и сайты для рестриктаз KpnI и XhoI. ПЦР с праймерами F и R, Encyclo полимеразой (ЗАО Евроген, Россия) и кДНК Н. magnifica проводят при следующих условиях: 94°С - 20 сек, 55°С - 20 сек, 72°С 20 сек, 25 циклов. Последовательность Mgf-II вставляют в гибридный белок после фрагмента тиоредоксина (ТrхА) и последовательности 6 остатков гистидина (6xHis) для прокариотической экспрессии в виде водорастворимого слитного белка (фиг. 6). Для этого ПЦР продукт обрабатывают рестриктазами KpnI и XhoI и лигируют с экспрессирующим вектором pET32b+ (Novagen). В результате получают плазмиду, кодирующую слитный белок под контролем Т7 промотера. Полученную плазмиду трансформируют в клетки XL1 и проверяют правильность сборки, лигирования и корректности последовательности Mgf-II секвенированием.
Пример 5
Получение рекомбинантного Mgf-II в бактериальной системе Е. coli
Клетки BL21 (DE3) трансформируют экспрессионной плазмидой pET-32b(+)-Mgf-II. После селекции на чашках с антибиотиком карбенициллином (100 мкг/мл), клетки пересеивают в 600 мл среды LB (карбенициллин, 100 мкг/мл) и выращивают при перемешивании и аэрации до оптической плотности ~0,6 при длине волны 600 нм.
Индукцию экспрессии проводят добавлением изопропил-β-D-1-тиогалактопиранозида (ИПТГ) до концентрации 0,2 мМ, после чего инкубируют клетки еще 18 ч при 18°С. Клетки, осаждают при 8000 об/мин и ресуспендируют в буферном растворе для дезинтеграции (25 мМ Tris-HCl pH 8,0, 500 мМ NaCl, 1,25% глицерин). Клетки дезинтегрируют ультразвуком, осаждают клеточный дебрис и тельца включения центрифугированием при 10000 об/мин. Целевой слитный белок содержится в растворимой фракции. В результате экспрессии созданной конструкции наработка рекомбинантного Mgf-II происходит в составе слитного белка с тиоредоксином, где последовательность тиоредоксина совместно с полигистидиновым фрагментом предшествует активному пептиду. Металлоаффинную хроматографию на носителе Ni2+-NTA-агароза проводят в буферном растворе (25 мМ ТрисHCl pH 8,0, 500 мМ NaCl), для элюирования слитного белка используют буферный раствор (250 мМ имидазола, 500 мМ⋅NaCl, 25 мМ Tris, pH 8,0). Собранный белок обессоливают на центрифужных картриджах Amicon Ultra-10 (Millipbre), используя дистиллированную воду как элюент.
Сконцентрированный слитный белок доводят до концентрации 1 мг/мл буферным раствором (50 мМ Tris-HCl pH 8,0, 1 мМ СаСl2) и подвергают процедуре рефолдинга в течение 5 суток при 14°С добавлением 4 мМ восстановленного (GSH) и 1 мМ окисленного (GSSG) глутатиона, после чего расщепляют энтерокиназой с расчетом 1,5 ед. акт. на 1 мг белка в течение 24 ч в шейкере при 80 об/мин. Смесь продуктов гидролиза фильтруют через насадку на шприц Syringe filter (ТРР).
Далее фильтрат наносят на колонку с обращенной фазой Jupier С4 (Phenomenex) 10×250 мм для выделения целевого полипептида (фиг. 7). Фракционирование проводят в линейном градиенте концентрации ацетонитрила от 0 до 70% (v/v) в 0,1%-ной (v/v) ТФУ в течение 70 мин со скоростью элюции 3 мл/мин. Детекцию осуществляют спектрофотометрически при длине волны 214 нм. Контроль чистоты и правильности синтеза рекомбинантного Mgf-II проверяют при помощи масс-спектрометрического анализа, сравнением хроматографических подвижностей природного и рекомбинантного полипептида на аналитической колонке Luna С18 (Phenomenex) 10×250 мм, а также сравнением ингибиторной активности in vitro.
Пример 6
Определение относительной молекулярной массы
Индивидуальность очищенного полипептида подтверждают масс-спектрометрическим анализом. Масс-спектры получают на MALDI-времяпролетном масс-спектрометре ULTRAFLEX III MALDI-TOF/TOF (Bruker Daltonik, Германия) с идентификацией положительных ионов в линейном режиме. В качестве матрицы используют синапиновую кислоту в концентрации 10 мг/мл. Кислоту растворяют в 50% ацетонитриле, содержащем 0,1% ТФУ. На подложку наносят 2 мкл матрицы, после высыхания наносят на нее 2 мкл образца пептида и сушат потоком воздуха, после чего подложку вносят в масс-спектрометр. Для калибровки прибора используют стандартную смесь пептидов с диапазоном молекулярных масс 700-3500 Да (Sigma, США).
Измеренная средняя молекулярная масса Mgf-II составляет 4779 Да. Расчетная средняя молекулярная масса отличается от измеренной на 0,41 Да. С учетом точности метода определения масс выносится заключение об отсутствии любой посттрансляционной модификации у данного пептида.
Пример 7
Определение констант ингибирования α-амилаз
Перед расчетом реальных констант ингибирования свиной панкреатической (РРА), человеческой слюнной (HSA) и панкреатической (HPА) α-амилаз Mgf-II, определяют константы Михаэлиса (KM) ферментов и подбирают их оптимальные концентрации. Стационарную кинетику ферментов изучают путем наблюдения за протеканием реакции гидролиза хромогенного субстрата 2-хлор-4-нитрофенил-α-D-мальтотриозида (CNPG3), происходящей под действием α-амилазы. Реакции проводят в 96-луночном кварцевом планшете в конечном объеме 100 мкл в буферном растворе, содержащем 50 мМ Nа3РО4, 100 мМ NaCl (pH 7,0), при температуре 37°С. Регистрацию накопления окрашенного продукта 2-хлор-4-нитрофенола проводят в микропланшетном спектрофотометре xMark при длине волны 405 нм в кинетическом режиме на протяжении 30 мин. Результаты измерений в 3 повторах преобразуют в кривые зависимости оптической плотности от времени.
Для определения константы ингибирования (Ki), ферменты РРА (20 нМ), HSA (100 нМ) и HP А (20 нМ) инкубируют с Mgf-II в разных концентрациях (от 1 до 10 нМ) в течение 10 мин при 37°С. Субстрат CNP-G3 в концентрации 1 мМ для РРА, 3,3 мМ для HSA и 1,7 мМ для HSA добавляют в реакционную смесь и выдерживают 10 мин при 37°С. Значения Ki определяют по уравнению Моррисона [Morrison J.F. Kinetics of the reversible inhibition of: enzyme-catalysed reactions by tight-binding inhibitors. Biochim. Biophys. Acta Enzymol. 1969, 185, 269-286] и получают следующие значения; 0,740 и 0,069 нМ в отношений НРА, 0,082 и 0,073 нМ - HSA и 0,045 и 0,046 нМ - РРА для природного и рекомбинантного Mgf-II соответственно (фиг. 8). При сравнении значений Ki с таковыми для магнификамида и хелиантамида делают вывод, что Mgf-II является эффективным ингибитором α-амилаз млекопитающих и обладает большим сродством к панкреатической α-амилазе, чем к слюнной.
Примере 8
Тестирование способности Mgf-II подавлять постпрандиальную гипергликемию Тестирование способности Mgf-II подавлять Постпрандиальную гипергликемию проводят на биомодели острого метаболического синдрома по типу сахарного диабета 1 типа у животных, вызванного стрептозоцином. В эксперименте используют самцов мышей линии ICR в возрасте 10 недель. Мышей делят на 7 групп (3 контрольные и 4 экспериментальные) по 9 особей в каждой, СД1Т индуцируют у 6-ти групп животных путем однократной внутрибрюшинной инъекции стрептозоцина, растворенного в 0,1 М цитратном буферном растворе, pH 4,5, в дозе 150 мг/кг (10 мл/кг). Перед введением стрептозоцина и измерением концентрации глюкозы, животных лишают корма на 4 часа. Концентрацию глюкозы определяют путем забора капли крови небольшим надрезом кончика хвоста на глюкометре Сателлит®Экспресс (СОП Fm/73). Развитие гипергликемии в группе отслеживают на 7 и 14 день эксперимента, У групп животных, которым вводят стрептозоцин, наблюдают значительное повышение концентрации глюкозы в крови на 7 день исследования (16,6 ммоль/л в среднем), относительно начального уровня глюкозы на 1 день исследования (6,9 ммоль/л в среднем) по сравнению с контрольной группой, получившей однократную дозу 10 мл/кг 0,1 М цитратного буферного раствора, pH 4,5, внутрибрюшинно. Крахмальный тест проводят на 14 день эксперимента и наблюдают выраженный ответ (19,2 ммоль/л в среднем) на введение крахмала в дозе 3 г/кг в течение 120 мин у животных с СД1Т по сравнению с контрольной группой (фиг. 9).
Mgf-II растворяют в стерильном физиологическом растворе и вводят в дозах 0,005 и 0,01 мг/кг перорально на 14 день исследования. Акарбозу в дозе 3 мг/кг (перорально) используют в качестве положительного контроля. Ближайший аналог Mgf-II, магнификамид, в дозах 0,005 и 0,01 мг/кг (перорально) применяют для сравнения эффективности. Животные лишаются корма в течение 4 ч перед началом тестирования. Крахмальный тест проводится через 45 мин после введения тестируемых веществ. Группы животных, которым не вводили стрептозоцин (здоровые), и одна группа с СД1 (отрицательный контроль) получают равный объем носителя (физраствор). Измеряют абсолютные значения концентрации глюкозы в крови через 0, 30, 60, 90 и 120 мин после введения крахмала в дозе 3 г/кг. Получают, что Mgf-II в дозе 0,01 мг/кг не приводит к повышению уровня глюкозы после введения крахмала в течение 120 мин и, в отличие от магнификамида и акарбозы, достоверно снижает абсолютные значения концентрации глюкозы по сравнений) с группой отрицательного контроля (фиг. 9). Результаты обрабатывают статистически, достоверность отличий результатов контрольной и экспериментальной группы определяют с помощью теста Стьюдента.
Примеры доказывают, что заявляемое соединение полипептид Mgf-II, состоящий из 44 аминокислотных остатков, эффективно ингибирует α-амилазы млекопитающих и может быть выделен как из морской анемоны Н. magnifica, так и получен в бактериальной системе рекомбинантным способом. Рекомбинантный Mgf-II полностью соответствует нативному полипептиду по структуре и биологической активности и может быть использован самостоятельно или в составе лекарственного средства для лечения и/или профилактики постпрандиальной гипергликемии, коррекции лишнего веса и ожирения у млекопитающих, в частности, человека.
--->
Перечень последовательностей
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3"
fileName="Magnificamide-II.xml" softwareName="WIPO Sequence"
softwareVersion="2.0.0" productionDate="2022-06-06">
<ApplicantFileReference>Patent Mg-II</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение науки Тихоокеанский институт биоорганической
химии им. Г.Б. Елякова Дальневосточного отделения Российской академии
наук </ApplicantName>
<ApplicantNameLatin>Pacific Institute of Bioorganic
Chemistry</ApplicantNameLatin>
<InventorName languageCode="ru">Синцова Оксана
Владимировна</InventorName>
<InventorNameLatin>Sintsova Oksana Vladimirovna</InventorNameLatin>
<InventionTitle languageCode="ru">Новый полипептид из морской
анемоны Heteractis magnifica, ингибирующий α-амилазы млекопитающих,
Mgf-II, экспрессионная конструкция, позволяющая производить
контролируемую экспрессию гена слитного белка для получения Mgf-II
рекомбинантным путем, способ получения Mgf-II в бактериальной системе
и его применение в качестве средства для лечения и/или профилактики
постпрандиальной гипергликемии</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>358</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..358</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q1">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Heteractis
magnifica</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gggcaagcagtggtatcaacgcagagtacgggggaataaaccatcagtg
gcgaaattctactactcactgggagctttacttatcagcaaaatgaaaactttaatacttctggttggtc
ttgcccttgttttcttggcgagtggaatgcccgtgaatgaagatcaagaaatgaattctgagggcaactc
gtgctatatctaccatggtgtgtacgggatatgcaaggctaagtgcgcggaagacgaaaaagccatggcg
ggaatgggtgtgtgtgaaggcgatctctgctgttataaaacaccatggtaatcatcggtggattttcgac
caggataatgaaatacaaaacaaataatt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>44</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..44</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Heteractis
magnifica</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>SEGNSCYIYHGVYGICKAKCAEDEKAMAGMGVCEGDLCCYKTPW</INS
DSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИД ИЗ МОРСКОЙ АНЕМОНЫ HETERACtis crispa, ОБЛАДАЮЩИЙ АНАЛЬГЕТИЧЕСКИМ ДЕЙСТВИЕМ | 2012 |
|
RU2475497C1 |
| ВАРИАНТ МАЛЬТОГЕННОЙ α-АМИЛАЗЫ | 1999 |
|
RU2258739C2 |
| АНАЛЬГЕТИЧЕСКИЙ ПЕПТИД ИЗ МОРСКОЙ АНЕМОНЫ URTICINA GREBELNYI | 2013 |
|
RU2521657C1 |
| ПОЛИПЕПТИД АКТИНИИ, ОБЛАДАЮЩИЙ АНАЛЬГЕТИЧЕСКИМ ДЕЙСТВИЕМ | 2009 |
|
RU2404245C1 |
| АНАЛЬГЕТИЧЕСКИЙ ПЕПТИД ИЗ МОРСКОЙ АНЕМОНЫ | 2016 |
|
RU2614759C1 |
| ПОЛИПЕПТИД АКТИНИИ, ОБЛАДАЮЩИЙ АНАЛЬГЕТИЧЕСКИМ ДЕЙСТВИЕМ | 2008 |
|
RU2368621C1 |
| КОМПОЗИЦИИ НА ОСНОВЕ ПОЛИПЕПТИДА α-АМИЛАЗЫ ИЗ BACILLUS, ВИД 195, И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2459867C2 |
| N,N-КОМПЛЕКС ДИХЛОРОДИ-[3,5-ДИМЕТИЛ-4-((БЕНЗИЛСУЛЬФАНИЛ)МЕТИЛ)-1H-ПИРАЗОЛ]ДИГИДРАТ МЕДИ(II) - СЕЛЕКТИВНЫЙ ИНГИБИТОР ФЕРМЕНТА α-АМИЛАЗА | 2019 |
|
RU2716138C1 |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ TS-23 С ИЗМЕНЕННЫМИ СВОЙСТВАМИ | 2009 |
|
RU2526516C2 |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ С ИЗМЕНЕННЫМИ СВОЙСТВАМИ | 2008 |
|
RU2499044C2 |

Изобретение относится к области биотехнологии, конкретно к новому полипептидному ингибитору альфа-амилаз млекопитающих, и может быть использовано в медицине для лечения или профилактики постпрандиальной гипергликемии, коррекции лишнего веса и ожирения. Предложен полипептид Mgf-II из морской анемоны Heteractis magnifica, обладающий способностью снижать уровень постпрандиальной глюкозы в крови млекопитающих за счет эффективного ингибирования альфа-амилаз. Изобретение позволяет получить эффективный глюкозоснижающий препарат, имеющий направленное действие на молекулярную мишень, который может быть доставлен в организм пероральным путем. 5 н.п. ф-лы, 9 ил., 8 пр.
1. Полипептид из морской анемоны Heteractis magnifica, ингибирующий α-амилазы млекопитающих и имеющий аминокислотную последовательность SEQ ID NO: 2.
2. Применение полипептида по п.1 в качестве ингибитора α-амилаз млекопитающих.
3. Применение полипептида по п.1 в качестве средства для лечения и/или профилактики постпрандиальной гипергликемии.
4. Применение полипептида по п.1 в качестве средства для коррекции лишнего веса.
5. Применение полипептида по п.1 в качестве средства для коррекции ожирения.
| SINTSOVA O | |||
| et al., Magnificamide, a β-Defensin-Like peptide from the mucus of the sea anemone Heteractis magnifica, is a strong inhibitor of mammalian α-Amylases, Mar | |||
| Drugs, 2019, v | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| SINTSOVA O | |||
| et al., Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a | |||

