Область техники
Изобретение относится к медицине, а именно к новым соединениям, представляющим собой модифицированные капсульные полисахариды S. pneumoniae, перспективным для получения вакцинных препаратов для профилактики пневмококковой инфекции.
Уровень техники
Одним из главных возбудителей инфекционных вспышек пневмонии, менингита и безочаговой бактериемии является грамположительный микроорганизм Streptococcus pneumoniae, для которого характерна высокая контагиозность, бессимптомное носительство, причем нескольких серотипов одновременно. В 2016 году из 336 млн. зарегистрированных в мире случаев респираторных заболеваний нижних дыхательных путей 197 млн. пришлось на S. pneumonia (1,2 млн. летальных исходов). В наибольшей степени подвержены этому заболеванию дети до 5 лет и представители возрастной когорты 70+ [1]. Ежегодно в Российской Федерации регистрируется 500000 эпизодов заболевания пневмонией, и у 76% взрослых и 90% детей до 5 лет возбудителем инфекции является S. pneumoniae. При этом смертность от пневмонии в 2013 году составила 26,7 случаев на 100000 человек (51,7% всех летальных исходов от респираторных заболеваний) [2]. Наиболее эффективным средством противодействия распространению S. pneumoniae является вакцинопрофилактика, так как многие циркулирующие и вновь появляющиеся штаммы обладают устойчивостью к антибиотикам.
Современный этап разработки вакцин против S. pneumoniae характеризуется признанием того, что в качестве непременных компонентов протективного препарата должны выступать уникальные по химической структуре капсульные полисахариды (КПС) S. pneumoniae.
Капсульные полисахариды бактерий (КПС), образуя плотную оболочку вокруг клетки, играют, главным образом, защитную роль во взаимодействии микроорганизма с окружающей средой, в частности, с организмом хозяина. В случае S. pneumoniae, КПС не только формируют защитную капсулу вокруг бактериальной клетки, но и являются основным фактором вирулентности, определяют серологическую специфичность каждого из более чем 100 известных к настоящему времени серотипов S. pneumonia и при введении в макроорганизм индуцируют образование протективных антител. Циркуляция в человеческой популяции значительного числа серотипов S. pneumonia заставляет при разработке вакцины не только анализировать и учитывать вклад каждого серотипа в эпидемиологическую ситуацию в географическом регионе в данное время, но принимать во внимание динамику изменений этой ситуации, связанную с проведением массовой вакцинопрофилактики. При этом часто наблюдается заметное снижение в циркулирующей популяции S. pneumonia штаммов, КПС которых, выделяемые из надклеточной жидкости после культивирования и лизиса бактериальных клеток (далее обозначаемые как п-КПС, т.е. природные КПС, выделяемые для получения вакцин), входят в состав применяемых вакцин, при одновременном повышении вклада в эпидемиологическую ситуацию штаммов S. pneumonia не «вакцинных» серотипов. Отмечается также появление новых серотипов, связанное с генетически обусловленной постполимеризационной модификацией полисахаридной цепи КПС (О-ацетилирование, гликозилирование), что помогает пневмококку избегать воздействия защитных механизмов организма хозяина. Таким образом, чем большее число п-КПС эпидемически важных (для данной территории в данное время) серотипов входит в вакцинную композицию, тем эффективнее ее протективное действие. В настоящее время широко используется вакцина PNEUMOVAX 23 (Merck Sharp & Dohme, США), в состав которой входит 23 КПС, выделенных из эпидемически важных для США серотипов S. pneumoniae (эффективность от 56 до 81%).
Чрезвычайно высокомолекулярные КПС всех серотипов S. pneumonia (от 500-900 до 1000 кДа) построены из различающихся по структуре повторяющихся олигосахаридных звеньев, в состав которых входит от 2 до 7 моносахаридных остатков, а также не углеводные фрагменты - О-ацетильные и фосфатные группы, остатки глицерина и пировиноградной кислоты. Моносахарид, расположенный на восстанавливающем конце этого биополимера, присоединен гликозидной связью непосредственно к пептидогликану (ПГ) клеточной стенки [3], в то время как в большинстве изученных грамположительных микроорганизмов связь между полисахаридной цепью и ПГ осуществляется через фосфодиэфирный остаток.
ПГ, в свою очередь, представляет собой прочную замкнутую пространственно «сшитую» структуру в виде так называемого «муреинового мешка», которая обеспечивает механическую и биологическую сохранность внутренних органелл бактериальной клетки. В основе ПГ лежит полисахарид, построенный из дисахаридных повторяющихся звеньев, включающих остаток N-ацетил-D-глюкозамина и N-ацетил-D-мурамовой кислоты (N-ацетил-D-глюкозамина, к которому простой эфирной связью присоединен остаток молочной кислоты). Дисахаридные звенья соединены β-1,4-гликозидными связями, а поперечные связи («сшивки») между полисахаридными цепями осуществляются за счет пептидных мостиков, образующихся между карбоксильными группами остатков N-ацетил-D-мурамовой кислоты соседних полисахаридных цепей. В случае S. pneumonia в состав мостика входят остатки L- и D-аланина, L-изоглутамина и L-лизина. На внешней поверхности «муреинового мешка» расположена липид-содержащая наружная мембрана, в которую «заякорены» соматические антигены белковой и полисахаридной природы, имеющие терминальный, обычно липидный, гидрофобный фрагмент («якорь»).
Наряду с КПС, который присоединяется к ПГ в положение 6 остатков N-ацетил-D-глюкозамина [4], на внешней поверхности бактерий S. pneumonia локализованы другие полимерные молекулы полисахаридной природы, являющиеся одинаковыми (т.е. общими) для S. pneumonia всех серотипов - общие полисахаридные антигены пневмококков (ОПАП). Часть этих биополимеров, имея от 2 до 8 повторяющихся олигосахаридных звеньев и липидный фрагментна восстанавливающем конце, «заякорена» во внешнюю липид-содержащую мембрану бактериальной клетки (F-антиген), тогда как другая часть ОПАП восстанавливающим концом присоединяется ковалентной связью к ПГ клеточной стенки (С-полисахарид, С-ПС) [5]. Строение полисахаридной области обоих антигенов идентично, различен лишь способ их локализации на внешней поверхности бактериальной клетки.
При получении вакцин против S. pneumonia п-КПС обычно выделяют из надклеточной жидкости после культивирования S. pneumonia и удаления клеточной массы центрифугированием. Надклеточную жидкость концентрируют, и низкомолекулярный водорастворимый F-антиген, который наряду с п-КПС содержится в надклеточной жидкости, отделяют физико-химическими методами, а С-ПС при этом теоретически должен оставаться связанным с ПГ клеточной стенки и отделяться при очистке п-КПС от ПГ. Однако при использовании различных методов очистки практически все препараты п-КПС, использующиеся для приготовления вакцин, включая конъюгированные, содержат заметное количество F-антигена и/или С-ПС. Эти полисахаридные молекулы обладают высокой иммуногенностью, однако образующиеся против них антитела не обладают защитным действием против S. pneumonia (Nielsen, Sorensen, & Henrichsen, 1993). Поэтому очистка п-КПС от указанных выше примесей остается важной проблемой, которая до настоящего времени не находила решения. Молекулярная масса п-КПС на порядки выше, чем молекулярная масса ОПАП, однако применение традиционных методов разделения полимерных молекул (различные виды гель-хроматографии, ультрафильтрация с предварительной обработкой ультразвуком, использование детергентов) для удаления примеси ОПАП не приводит к успеху. Количество примесного ОПАП в п-КПС вакцинного качества составляет от 5-7 до 15-18%, и это необходимо учитывать при производстве вакцин против S. pneumoniae, так как доза вакцинного п-КПС в препарате должна быть стандартной вне зависимости от содержания примеси ОПАП. Это существенно усложняет производство вакцин, в связи, с чем разработка методов выделения и очистки п-КПС S. pneumonia от F-антигена является важной задачей не только при создании полисахаридных, но и конъюгированных вакцинных препаратов на их основе. Недавно проведенные исследования показали, что очистка п-КПС от примеси ОПАП возможна с использованием реакции дезаминирования [6], в которую вовлекаются только соединения со свободной аминогруппой [7]. Мишень дезаминирования - остаток 4-амино-2,4,6-тридезоксигалактозы, который входит в состав ОПАП и, как показано авторами исследования, разрушается в результате действия азотистой кислоты (NaNO2 + CH3COOH), что приводит к деполимеризации ОПАП. Низкомолекулярные продукты деградации легко удаляются с помощью ультрафильтрации, и полученные таким способом КПС по данным спектров ЯМР, не претерпевая структурных изменений, освобождаются от примеси ОПАП.
Было установлено, что КПС как грамотрицательных, так и грамположительных микроорганизмов, являясь факторами вирулентности, представляют собой тимус-независимые антигены, которые, при использовании в качестве вакцинных препаратов, индуцируют выработку в основном довольно быстро исчезающих IgM антител, но не способны переключить гуморальный иммунный ответ на выработку долго живущих высоко специфичных антител класса G. Обычно не обнаруживаются при этом и В-клетки памяти. Кроме того, вакцины, содержащие только полисахаридные антигены, не эффективны при проведении профилактики у детей младшего возраста и пожилых людей. В последние годы разработаны конъюгированные полисахарид-белковые вакцины, лишенные этих недостатков. Присоединение к полисахариду белкового носителя (обычно анатоксина) с помощью ковалентной связи придает конъюгату тимус-зависимость и обеспечивает, таким образом, способность активировать иммунную память. В настоящее время для применения в клинической практике разрешены 7-, 10- и 13-валентная конъюгированные вакцины против S. pneumoniae, клинические испытания проходит 20-валентная вакцина [3].
Однако этот подход, несмотря на ряд несомненных достоинств, не лишен некоторых существенных недостатков. Так, в силу невысокой иммуногенности конъюгированных полисахарид-белковых вакцин обычно необходимо присутствие в вакцине адъювантов, чаще всего производных гидроокиси алюминия, которые, по данным последних исследований, обладают рядом нежелательных характеристик.
Высокое содержание в конъюгате белкового компонента (более 50%) приводит к индуцированию мощного экранирующего антибелкового гуморального иммунного ответа. В случае вакцин против S. pneumoniae при проведении конъюгации очень высокомолекулярного КПС (молекулярная масса до 1000 кДа) с различными белковыми матрицами обычно используются методы «рандомного» образования мостиковых связей между полимерными макромолекулами, что приводит к появлению новых структурных эпитопов в полисахаридной (и белковой) части конъюгата и, как следствие, к индукции антител, не характерных для иммунного ответа на нативный КПС.
Конъюгат, в состав которого входит КПС с высокой молекулярной массой и большое количество белка разной степени гидрофобности, зачастую склонен к частичной потере растворимости. В этой связи в случае некоторых КПС приходится понижать его молекулярную массу с помощью различных химических и физико-химических методов.
Помимо этого, обращают на себя внимание сложности, связанные с воспроизводимостью процесса конъюгации, а также анализа полученных конструктов на предмет присутствия в них исходных биополимеров. Перечисленные проблемы подробно обсуждаются в обзорах [4, 5]. Довольно высокая стоимость конъюгированных полисахарид-белковых вакцинных препаратов также существенно ограничивает их применение.
Известно техническое решение US4356170, в котором антигенные менингококковые полисахариды модифицируют путем восстановления моносахарида, расположенного на восстанавливающем конце полисахарида, боргидридом натрия с последующим окислением периодатом натрия для получения терминально расположенной альдегидной группы, которую затем ковалентно связывают со свободной аминогруппой выбранного белка, посредством -CH2-NH-белок связи.
За ближайший аналог можно принять техническое решение WO199640795, в котором описано использование для конъюгирования с белком с помощью восстановительного аминирования свободных альдегидных групп терминальных остатков 2,5-ангидро-D-маннозы, образующихся при деполимеризации капсульных полисахаридов Streptococcus серотипа II или типа III, с помощью мягкого N-дезацетилирования с последующим дезаминированием для конъюгирования с белком с помощью восстановительного аминирования.
Таким образом, несмотря на то, что в распоряжении врачей есть конъюгированные вакцины против S. pneumoniae, все вышеперечисленные проблемы требуют поиска новых подходов к повышению эффективности конъюгированных вакцин на основе КПС S. pneumoniae.
Раскрытие сущности изобретения
Технической задачей настоящего изобретения является повышение эффективности и оптимизации технологии получения конъюгированных вакцин на основе КПС S. pneumoniae.
Технический результат заключается в получении с помощью дезаминирования модифицированных капсульных полисахаридов (м-КПС) S. pneumoniae, не содержащих примеси общего полисахаридного антигена пневмококков (ОПАП), удобных для последующей конъюгации с матрицей белковой или иной природы, включающей активные аминогруппы, обладающие более высокой растворимостью в водных буферах, чем исходные (природные) КПС, что позволяет повысить технологичность процессов конъюгации, и которые могут быть использованы для получения поливалентных пневмококковых полисахаридных вакцин, а также пневмококковых гликоконъюгированных вакцин.
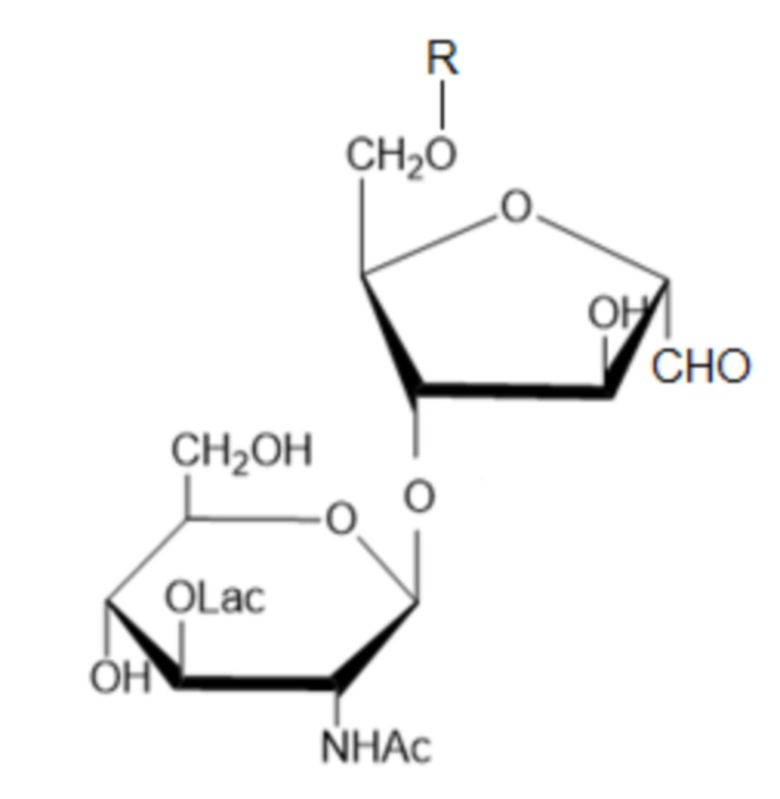
Указанный технический результат достигается тем, что получены соединения общей формулы 4-О-(β-N-ацетил-D-мурамил)-6-О-R-2,5-ангидро-D-манноза:
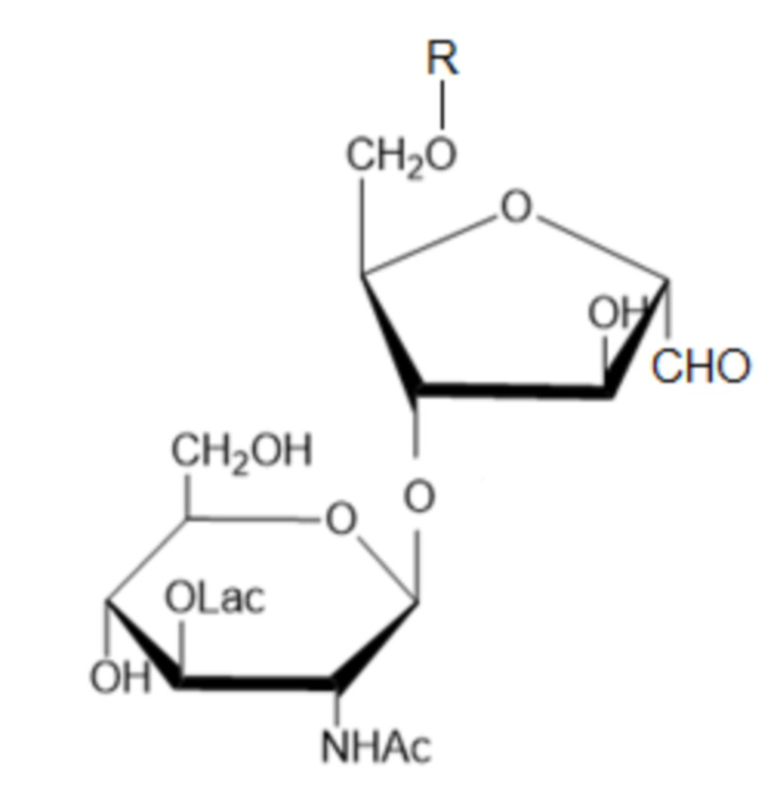 (I),
(I),
где R - остаток капсульного полисахарида S. pneumoniae, любого серотипа, за исключением серотипов 1 и 3, а Lac- обозначает лактил[-CH(CH3)COOH], причем R представляет собой остаток капсульного полисахарида штамма S. pneumoniae серотипа 4, 5, 6А, 6В, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 15F, 18C, 19F, 19A, 22F, 23A, 23F или 33F.
Кроме того, указанный технический результат достигается тем, соединение общей формулы (I) применяется для получения конъюгированных полисахарид-белковых вакцин против S. pneumoniae, при этом полисахарид-белковый конъюгат представляет собой конструкт с мостиковыми связями соединения общей формулы (I) и белковой матрицы (белка-носителя). Кроме того, полисахарид-белковый конъюгат представляет собой конструкт, в котором соединение общей формулы (I) присоединяется к белковой матрице с участием только активной альдегидной группы - СНО остатка 2,5-ангидро-D-маннозы с образованием «звездной» структуры. При этом белковая матрица (белок-носитель) представляет собой бычий сывороточный альбумин, столбнячный анатоксин, или холерный анатоксин, или мутантный дифтерийный анатоксин CRM197, или рекомбинантный детоксифицированный токсин CRM197, или консервативный белковый антиген пневмококка, но не ограничивается ими.
Также технический результат достигается за счет разработки способа получения соединения общей формулы (I), включающий следующие последовательные этапы:
i) культивируют S. pneumoniae с получением культуральной жидкости S. pneumoniae;
ii) проводят центрифугирование или микрофильтрацию полученной культуральной жидкости с разделением надклеточной жидкости и клеточной массы;
iii) выделяют неочищенный исходный капсульный полисахарид (КПС) S. pneumoniae путем ультра- и диафильтрации полученной на этапе ii) надклеточной жидкости с последующей лиофилизацией; получают очищенный исходный КПС S. pneumoniae путем обработки лиофилизата нуклеазами и затем протеиназой К с последующей очисткой диафильтрацией и лиофилизацией;
iv) проводят реакцию дезаминирования исходного КПС S. pneumoniae добавлением 3-8 мас % водного раствора нитрита натрия к раствору очищенного исходного КПС в 10-50 мас % водном растворе уксусной кислоты;
или
iii) промывают и суспендируют клеточную массу, полученную на этапе ii), деионизированной водой с получением клеточной суспензии;
iv) проводят реакцию дезаминирования путем добавления 3-8 мас % водного раствора нитрита натрия к реакционной смеси, представляющей собой смесь полученной клеточной суспензии с 10-50 мас % водным раствором уксусной кислоты;
v) реакционную смесь, полученную в результате реакции дезаминирования на этапе iv), разбавляют деионизированной водой и затем нейтрализуют путем добавления NaOH, KOH, NH4OH или NaHCO3;
vi) полученный раствор подвергают диализации или диафильтрации с последующей лиофилизацией с получением целевого соединения.
Определения и термины
Следующие термины и определения применяются в рамках данного изобретения, если иное не указано явно. Ссылки на методики, используемые при описании данного изобретения, относятся к хорошо известным методам, включая изменения этих методов и замену их эквивалентными методами, известными специалистам.
КПС (капсульный полисахарид) - полисахарид, построенный из повторяющихся олигосахаридных звеньев уникальной структуры, характерной для каждого серотипа S. pneumoniae.
п-КПС (природный капсульный полисахарид) или «исходный КПС» - КПС S. pneumoniae,выделяемый из надклеточной жидкости после культивирования и лизиса бактериальных клеток (в частности, при получении вакцин против S. pneumoniae).
м-КПС (модифицированный капсульный полисахарид) - модифицированный КПС S. pneumoniae, представляющий собой соединение общей формулы (I), где R - остаток капсульного полисахарида S. pneumoniae, причем восстанавливающий конец R связан с остатком 2,5-ангидро-D-маннозы путем образования гликозидной связи в положении 6остатка 2,5-ангидро-D-маннозы; R представляет собой остаток капсульного полисахарида штамма S. pneumoniae любого серотипа (за исключением серотипов 1 и 3) и имеет строение, соответствующее выбранному серотипу.
Восстанавливающий конец полисахарида - терминальный моносахаридный остаток полисахарида со свободной аномерной ОН-группой, не вовлеченный в образование гликозидной связи. При размыкании цикла такого моносахаридного остатка возможно образование свободной альдегидной группы. Линейные полисахариды имеют один невосстанавливающий и один восстанавливающий конец; в разветвленных полисахаридах (в том числе в капсульных полисахаридах) также может быть только один восстанавливающий конец, при этом число невосстанавливающих концевых моносахаридных остатков превышает число разветвлений.
Под серотипом штамма S. pneumoniae подразумевается группа микроорганизмов S. pneumoniae, объединяемых общей антигенной структурой, определяемой первичной химической структурой КПС.
Соединения по изобретению представляют собой любые соединения общей формулы (I), получение которых возможно с точки зрения химии и/или биологии. Это означает, что соединение по изобретению может включать остаток капсульного полисахарида (КПС) любого серотипа S. pneumoniae, строение клеточной стенки которого позволяет получить соединение общей формулы (I). В частности, КПС не должны содержать моносахаридных фрагментов, подверженных деградации в условиях реакции дезаминирования, используемой для получения соединений общей формулы (I) (см. подробнее далее); кроме того, КПС S. pneumoniae должен присоединяться к пептидогликану (ПГ) непосредственно гликозидной связью. Такое строение клеточной стенки присуще подавляющему большинству известных серотипов S. pneumoniae, за исключением серотипов 1 и 3 (см. подробнее далее).
Если не определено отдельно, технические и научные термины в данной заявке имеют стандартные значения, общепринятые в научной и технической литературе.
Краткое описание фигур
Фиг.1 Схема дезаминирования остатка N-дезацетилированного глюкозамина в составе пептидогликана клеточной стенки с получением м-КПС, на восстанавливающем конце которого локализован остаток 2,5-ангидро-D-маннозы (из D-глюкозамина), несущий в положении 4 остаток N-ацетил-D-мурамовой кислоты, а в положении 6 - остаток КПС. Обозначения: Ас - ацетил, MurNAc-остаток N-ацетил-D-мурамовой кислоты, GlcN-остаток D-глюкозамина, 2,5-anhydro-Man - остаток 2,5-ангидро-D-маннозы, Lac - остаток молочной кислоты (лактил, -CH(CH3)COOH), n- целое число, характеризующее количество повторяющихся звеньев в полисахаридной цепи пептидогликана.
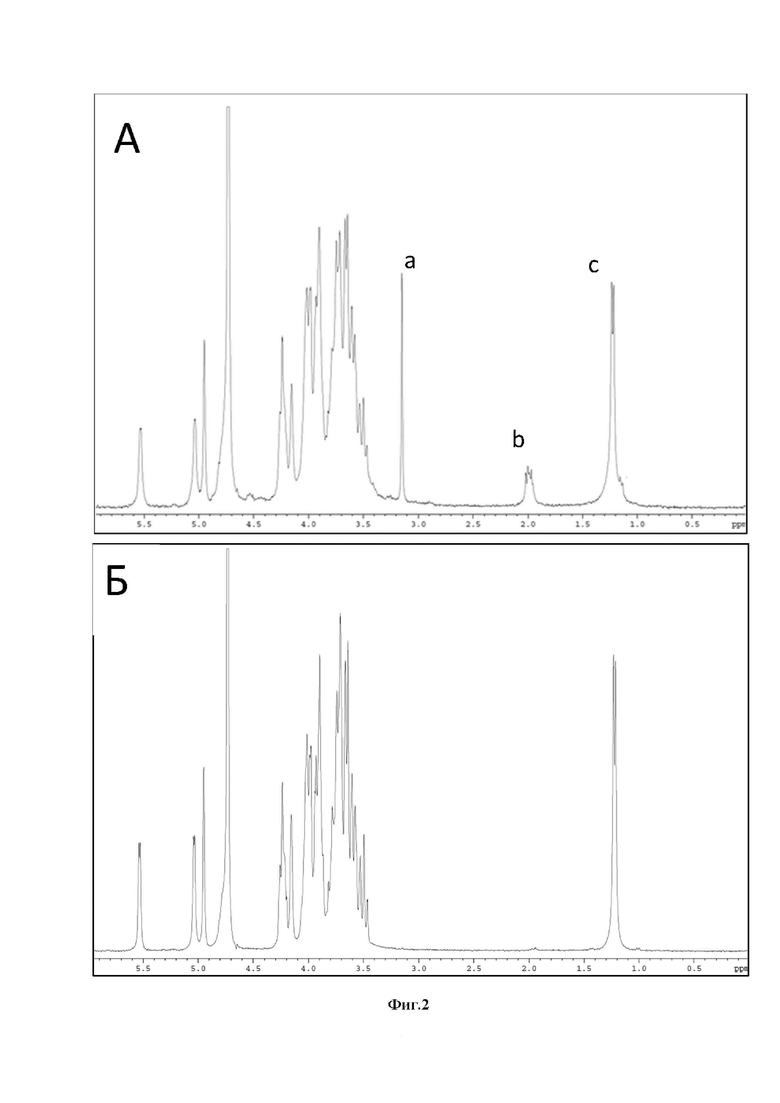
Фиг.2: А - 1H-ЯМР спектр исходного п-КПС бактериального штамма S. pneumonia серотипа 6А, Б - 1H-ЯМР спектр м-КПС бактериального штамма S. pneumonia серотипа 6А.
a- сигнал остатка холина; b - сигналы N-ацетильных групп аминосахаров из ОПАП и ПГ; с - сигнал метильной группы остатка α-L-рамнозы.
Фиг.3: А - Спектр 1Н-ЯМР адипиндигидразидного производного м-КПС 6А; Б - Спектр 1Н-ЯМР стандартного образца адипиндигидразида.
Фиг.4. Динамика веса животных в контрольной и опытных группах в ходе исследования.
Обозначения: PBS - контрольная группа; «КПС 0,5 мкг» - экспериментальная группа животных, иммунизированных м-КПС в дозе 0,5 мкг; «КПС 5 мкг» - экспериментальная группа животных, иммунизированных м-КПС в дозе 5 мкг. И1, И2, И3 - время первой, второй и третьей иммунизации соответственно.
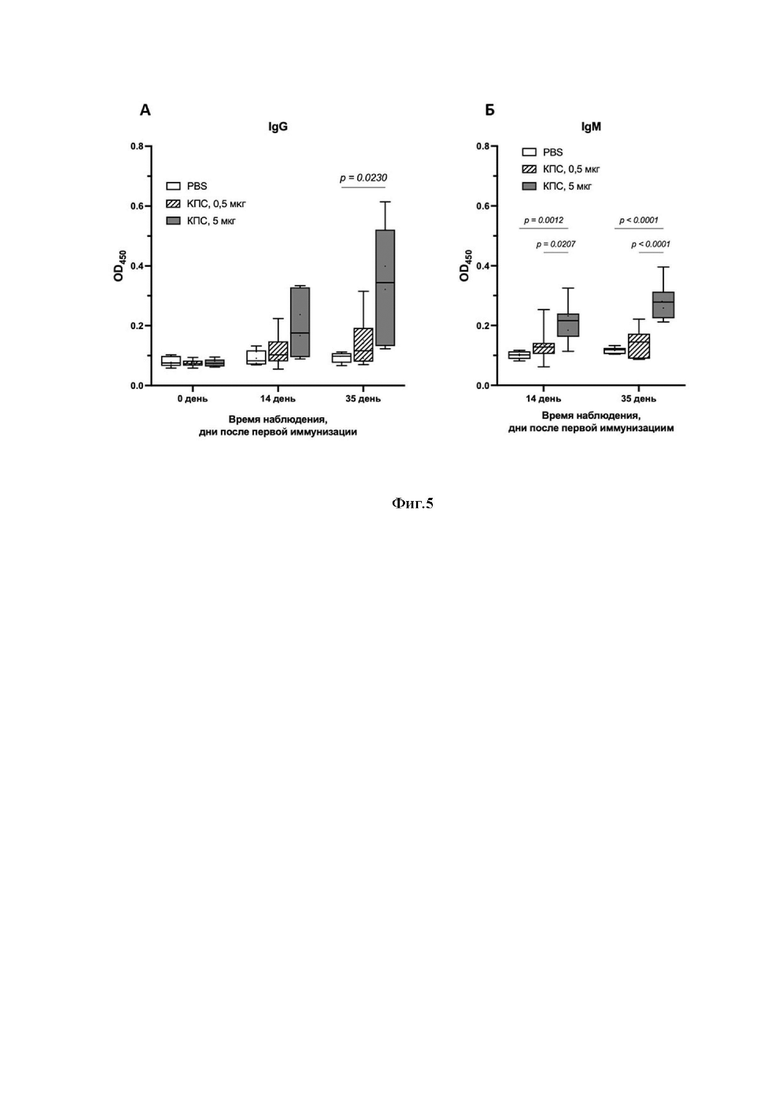
Фиг.5: А- Продукция специфических антител IgG в сыворотке крови иммунизированных животных в ответ на введение м-КПС серотипа 6В; Б - Продукция специфических антител IgМ в сыворотке крови иммунизированных животных в ответ на введение м-КПС серотипа 6В;
Обозначены статистически значимые отличия между группами.
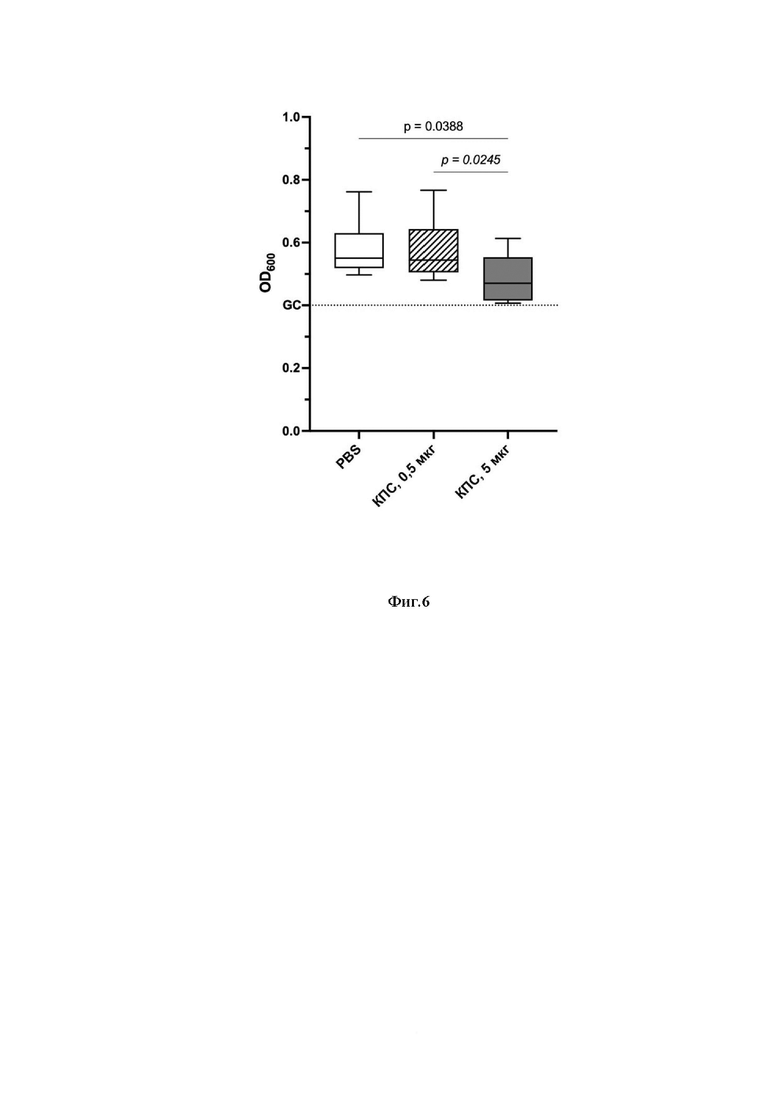
Фиг.6 Бактериостатическая активность сыворотки крови иммунизированных животных после третьей иммунизации.
Обозначения: GC - контроль роста культуры. PBS - контрольная группа; «КПС 0,5 мкг» - экспериментальная группа животных, иммунизированных м-КПС в дозе 0,5 мкг; «КПС 5 мкг» - экспериментальная группа животных, иммунизированных м-КПС в дозе 5 мкг. Обозначены статистически значимые отличия между группами.
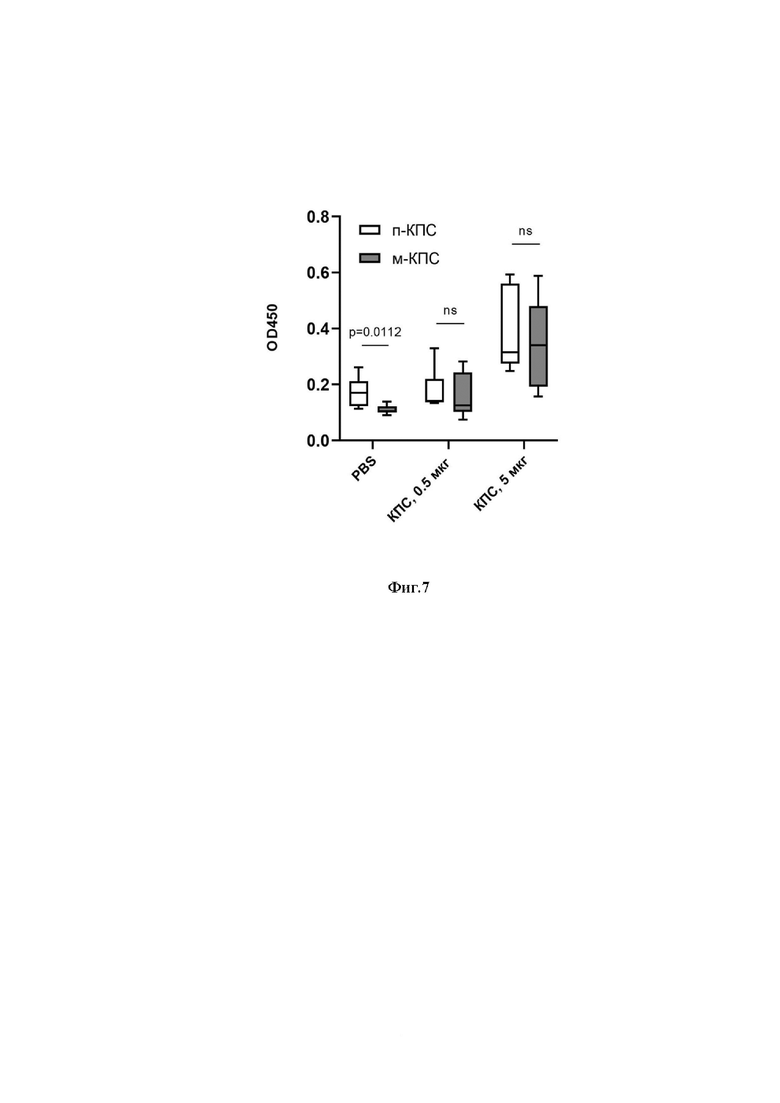
Фиг.7 Уровень IgG антител, связывающихся с п-КПС и м-КПС в сыворотке крови мышей, иммунизированных м-КПС. Сравнение групп с помощью парного двустороннего t-критерия Стьюдента.
Обозначения: GC - контроль роста культуры. PBS - контрольная группа; «КПС 0,5 мкг» - экспериментальная группа животных, иммунизированных м-КПС в дозе 0,5 мкг; «КПС 5 мкг» - экспериментальная группа животных, иммунизированных м-КПС в дозе 5 мкг. Обозначены статистически значимые отличия между группами.
Осуществление изобретения
В рамках проведения исследований, авторами настоящего изобретения, с использованием традиционного поэтапного подхода к выделению и очистке (концентрирование и диализ с помощью тангенциальной ультрафильтрации, обработка нуклеазами и затем протеиназой К, высаживание в спирт) из надклеточной жидкости, полученной в результате культивирования в ферментере, были выделены п-КПС более 20 эпидемически значимых серотипов S. pneumoniae. Данные спектроскопии 1Н-ЯМР полученных п-КПС полностью совпадали с литературными. Каждый и выделенных п-КПС содержал по данным 1Н-ЯМР-спектроскопии от 5 до 17% ОПАП (см. Пример 1).
Для очистки п-КПС от примеси ОПАП был использован способ, описанный в публикации [6]. Однако условия проведения реакции дезаминирования п-КПС были изменены: был использован водный раствор уксусной кислоты в соотношении CH3COOН: Н2О 1:3, вместо 5%-ной уксусной кислоты, которую вводили в реакцию дезаминирования авторы статьи [6]. В результате реакций, проведенных с выделенными п-КПС, были получены модифицированные п-КПС (далее обозначаемые как м-КПС). 1Н-ЯМР-спектры полученных в результате реакции дезаминирования м-КПС, как и следовало ожидать, не отличались от спектров исходных п-КПС, за исключением отсутствия в м-КПС сигнала при 3.2 м.д., характерного для (СН3)3N-группы остатка холина - непременного компонента ОПАП [5].
Однако при анализе молекулярно-массового распределения полученных таким образом м-КПС с помощью ВЭЖХ было обнаружено, что для большинства м-КПС время элюции значительно меньше времени элюции исходных п-КПС, что должно было бы указывать на то, что в результате дезаминирования происходит образование м-КПС с большей молекулярной массой, чем молекулярная масса исходных п-КПС, тогда как реакция дезаминирования должна была привести к дезинтеграции ОПАП без изменения молекулярной массы исходного биополимера. Проведение ВЭЖХ в различных буферах и в присутствии детергентов показало, что порядок элюции исходных п-КПС и м-КПС неизменен. Авторы настоящего изобретения предположили, что с наибольшей вероятностью обнаруженный хроматографический эффект может быть объяснен неспецифическим связыванием с ВЭЖХ-фазой необычных по своей архитектонике исходных молекул п-КПС S. pneumonia и отсутствием этого связывания у молекул м-КПС. Поскольку известно, что КПС и С-ПС S. pneumoniae присоединены ковалентной связью к ПГ[4, 5], авторы настоящего изобретения предположили, что «традиционные» п-КПС S. pneumoniae, используемые для получения вакцин, на самом деле представляют собой образующиеся в результате аутолизиса бактериальных клеток сложные «биоконъюгаты», в которых остатки КПС и С-ПС присоединены посредством гликозидных связей терминальных восстанавливающих моносахаридов к фрагменту ПГ, причем молекула такого трехкомпонентного конструкта может включать не один остаток КПС и С-ПС. Количество «биоконъюгата», образующегося в результате лизиса клеток S. pneumoniae как в процессе глубинного культивирования, так и во время стерилизации клеток после выращивания, зависит от условий проведения этих процедур. Так, в патенте компании VAJET (US) [8] предложен ускоренный метод получения «традиционного» п-КПС с высоким выходом, который включает лизис бактериальных клеток «посредством добавления детергента или другого реагента, который способствует разрушению клеточной стенки и освобождению аутолизинов, которые вызывают ускоренный лизис клеток по достижении стационарной фазы». Таким образом, следует полагать, что все лицензированные вакцинные препараты на основе п-КПС (включая конъюгированные) содержат описанный выше «биоконъюгат», примесь С-ПС из которого (ввиду наличия ковалентной связи С-ПС и КПС с ПГ) удалить обычными методами, естественно, невозможно. Архитектоника трехкомпонентной биомолекулы п-КПС обусловливает технические трудности, связанные с низкой растворимостью вакцинного препарата, и, как следствие, с необходимостью перед конъюгацией с белком снижать его молекулярную массу с помощью химических и физико-химических методов. Описанный выше эффект неспецифического связывания п-КПС в процессе ВЭЖХ обусловлен сложной разветвленной структурой «биоконъюгата» п-КПС. Выделенные из «биоконъюгата» м-КПС таким свойством не обладают, и их молекулярные массы могут быть определены стандартным образом с помощью ВЭЖХ.
В результате дезаминирования происходит деполимеризация С-ПС, что позволяет избавиться от его примеси в п-КПС, и при этом образуется м-КПС [6].
В грамположительном микроорганизме S. pneumonia строение узла связи КПС-ПГ имеет уникальный характер. КПС грамположительных бактерий присоединены в положение С-6 остатка N-ацетил-D-мурамовой кислоты через фосфодиэфирный мостик, в случае же S. pneumoniae местом присоединения КПС является С-6 остатка N-ацетил-D-глюкозамина, причем 80% остатков этого моносахарида и 10% остатков N-ацетил-D-мурамовой кислоты лишены N-ацетильной группы [9]. Наличие в узле связи до 90% N-дезацетилированных остатков обоих аминосахаров очевидно связано с механизмом защиты микроорганизма от действия лизирующих ферментов организма хозяина, которые распознают лишь N-ацетилированные остатки. Ранее [7] было показано, что дезаминирование 2-амино-2-дезоксиглюкозы (глюкозамина) и 2-амино-2-дезоксигалактозы (галактозамина) приводит к перегруппировке шестичленных циклов обоих моносахаридов с образованием 2,5-ангидроманнозы и 2,5-ангидроталозы, соответственно. Если остаток глюкозамина или галактозамина присоединен гликозидной связью, перегруппировка с сужением цикла сопровождается разрывом этой связи, причем в случае локализации остатка глюкозамина или галактозамина внутри полисахаридной цепи (но не в ответвлении) происходит деполимеризация полисахарида с образованием олигосахаридов с остатком 2,5-ангидропроизводного на восстанавливающем конце. При наличии связи остатков КПС с фрагментом ПГ, в котором присутствует до 90% N-дезацетилированных остатков обоих аминосахарных компонентов ПГ, в результате дезаминирования происходит расщепление «биоконъюгата», сопровождающееся освобождением м-КПС, на восстанавливающем конце которого локализован остаток 2,5-ангидро-D-маннозы (образуется из D-глюкозамина), несущий в положении 4 остаток N-ацетил-D-мурамовой кислоты, а в положении 6 - остаток КПС (Фиг.1).
Как показали проведенные в рамках данного изобретения исследования, спектры1H-ЯМР исходных п-КПС и образующихся в результате дезаминирования м-КПС отличаются лишь наличием в спектрах п-КПС минорных сигналов, соответствующих ОПАП и ПГ. В качестве примера на Фиг.2 приведены спектры 1H-ЯМР п-КПС и м-КПС бактериального штамма S. pneumoniae серотипа 6А, повторяющееся звено которого имеет следующую структуру:
[→2)-α-D-GaIp-(1→3)-α-D-Glcp-(1→3)-α-L-Rhap-(1→3)-D-Ribitol-(5→P→]n
Идентифицировать дисахаридный фрагмент с 2,5-ангидро-D-маннозой методом ЯМР-спектроскопии или ГЖХ-масс-спектрометрии не представляется возможным в силу чрезвычайно малого количества этого терминального моносахаридного компонента в м-КПС. Однако, в случае м-КПС бактериальных штаммов S. pneumoniae серотипов 6А и 6В удалось идентифицировать альдегидную группу остатка 2,5-ангидро-D-маннозы и определить ее количественное содержание. п-КПС серотипа 6А резко отличается по физико-химическим и иммунологическим характеристикам от всех остальных КПС. Несмотря на высокую распространенность этого серотипа, он не включен в 23-валентную вакцину из-за своей необъяснимо низкой иммуногенности [10]. Лишь конъюгат с белком на основе п-КПС серотипа 6А приобретает способность к индукции протективных антител (входит в состав всех конъюгированных пневмококковых вакцин). В рамках настоящего исследования установлено, что, в отличие от штаммов S. pneumoniae всех других серотипов (см. выше), время удерживания м-КПС серотипа 6А при ВЭЖХ значительно больше, чем у исходного п-КПС серотипа 6А (м-КПС имеет молекулярную массу, не превышающую 10 кДа), что резко отличает его ото всех остальных м-КПС S. pneumoniae. Это, с одной стороны, позволяет объяснить низкую иммуногенность п-КПС серотипа 6А [11], а, с другой стороны, констатировать, что у «биоконъюгата» с низкой молекулярной массой остатков КПС отсутствует способность к неспецифической адсорбции на ВЭЖХ-фазе.
Прямое доказательство наличия на восстанавливающем конце м-КПС серотипа 6А альдегидной группы было получено с помощью анализа 1Н-ЯМР-спектра гидразидного производного, полученного взаимодействием м-КПС серотипа 6А с избытком адипиндигидразида в условиях реакции восстановительного аминирования [12] (цианборгидрид натрия в натрий-фосфатном буфере, рН 7,2). После очистки с помощью гель-хроматографии на колонке TSK40 (TOYOSODA) в 0,05н АсОН по данным 1Н-ЯМР-спектроскопии было получено производное м-КПС серотипа 6А с H2NNHCO(CH2)4CONHNH-группой на восстанавливающем конце (Фиг.3). Из сравнения величин интегральных интенсивностей сигналов аномерного протона остатка рамнозы (1Н) и сигнала 2,5(4Н)- или 3,4(4Н)- метиленовых групп остатка адипиндигидразида следовало, что на один терминальный гидразидный остаток приходится около 30 повторяющихся олигосахаридных звеньев м-КПС. Таким же способом на примере м-КПС серотипа 6В также было подтверждено наличие в нем терминальной альдегидной группы на восстанавливающем конце. Для этого м-КПС серотипа 6В был подвергнут гель-хроматографии на колонке с TSK55 (TOYOSODA) в 0,2М NaCl, и выделенная при этом самая низкомолекулярная фракция после диализа и лиофилизации была обработана как описано выше для м-КПС серотипа 6А. В 1Н-ЯМР спектре гидразидного производного также присутствовали сигналы H2NNHCO(CH2)4CONHNH-группы, а интегрирование показало, что низкомолекулярный м-КПС серотипа 6В содержит не менее 40 повторяющихся олигосахаридных звеньев.
В рамках проводимого авторами изобретения исследования из всех выделенных эпидемически значимых п-КПС S. pneumonia были дезаминированы п-КПС с получением м-КПС для всех серотипов, кроме серотипов 1 и 3: п-КПС серотипа 1 содержит в своем составе 4-аминосахар, подверженный деградации в условиях этой реакции [6, 13], а п-КПС серотипа 3, в отличие от остальных п-КПС S. pneumoniae, не связан с ПГ [14],
Таким образом, в результате проведенных исследований были получены м-КПС - новые соединения, общая структурная формула которых представлена формулой (I):
,
где R - остаток капсульного полисахарида S. pneumoniae любого серотипа, за исключением серотипов 1 и 3.
Способ получения м-КПС (соединений общей формулы (I)) по изобретению включает следующие последовательные этапы:
I. - культивирование S. pneumoniae с получением культуральной жидкости S. pneumoniae;
II. - центрифугирование или микрофильтрация полученной культуральной жидкости для разделения надклеточной жидкости и клеточной массы;
III. - выделение неочищенного п-КПС S. pneumoniae путем ультра - и диафильтрации полученной надклеточной жидкости с последующей лиофилизацией;
IV. - получение очищенного п-КПС S. pneumoniae путем обработки лиофилизата нуклеазами и затем протеиназой К с последующей очисткой диафильтрацией и лиофилизацией;
V. - дезаминирование п-КПС с последующей очисткой целевого продукта (м-КПС) диафильтрацией и лиофилизацией.
Дезаминирование п-КПС на этапе V. проводят следующим образом: к водному раствору п-КПС добавляют 10-50%-ый водный раствор уксусной кислоты и после этого добавляют 3-8%-ый водный раствор нитрита натрия. Полученную реакционную смесь перемешивают в течение 0,5-2 часов при температуре 4 - 25°С, после чего ее разбавляют водой и нейтрализуют водным раствором неорганического основания, выбранного из NaOH, KOH, NH4OH, или, преимущественно, твердым NaHCO3.Далее полученный раствор подвергают диализации или диафильтрации с последующей лиофилизацией с получением м-КПС (соединения общей формулы (I)).
В некоторых частных вариантах дезаминирование проводят следующим образом: к раствору 20 мг п-КПС в 1-3 мл (в наиболее предпочтительном варианте в 1 мл) деионизованной воды прибавляют при перемешивании 1-2,5 мл (в наиболее предпочтительном варианте 1 мл) раствора уксусной кислоты с концентрацией от 10 до 50 мас % (в наиболее предпочтительном варианте 30 мас %) и затем 1,5-2,5 мл (в наиболее предпочтительном варианте 1 мл) водного раствора нитрита натрия с концентрацией от 3 до 8 мас % 1,5-2,5 мл (в наиболее предпочтительном варианте 5 мас %). Через 0,5 - 2 часа (в наиболее предпочтительном варианте 1 час) перемешивания при температуре 4 - 25°С (в наиболее предпочтительном варианте 20°С) реакционную смесь разбавляют в 3 раза деионизованной водой, нейтрализуют водным раствором неорганического основания (NaOH, KOH, NH4OH), или, преимущественно, твердым NaHCO3. Полученный раствор далее диализуют против деионизованной воды или подвергают диафильтрации с использованием мембраны с пределом отсечения 10 кДа, 20 кДа, 50 кДа или 100 кДа в зависимости от молекулярной массы полученного м-КПС и полученный водный раствор лиофилизуют.
Так как аутолитические процессы, протекающие при культивировании S. pneumoniae и после его окончания, полностью не разрушают ПГ, часть КПС остается с ним связанной, и поэтому дополнительное количество м-КПС может быть получено с помощью реакции дезаминирования из бактериальных клеток S. pneumoniae, выделенных после удаления надклеточной жидкости с помощью центрифугирования (т.е. из клеточной массы, выделенной на этапе II).Для этого проводят дезаминирование бактериальных клеток (т.е. полученной клеточной массы) с последующим удалением центрифугированием нерастворимого компонента и выделением из супернатанта с помощью диафильтрации дополнительного количества целевого продукта (м-КПС). Полученные при этом м-КПС после ультрафильтрации не содержат ОПАП и не требуют дополнительной очистки от белков и нуклеиновых кислот (см. Пример 3).
Как указывалось, выше, при культивировании S. pneumoniae и после его завершения в процессе стерилизации клеток после выращивания происходит лизис микроорганизмов, сопровождающийся разрушением ПГ клеточной стенки с помощью освобождающихся аутолитических ферментов и образованием «биоконъюгата». Таким образом, часть «биоконъюгата» оказывается в надклеточной жидкости, а другая часть - остается в составе не лизированных или частично лизированных бактериальных клеток. Из полученного после центрифугирования осадка таких нативных и полуразрушенных бактериальных клеток с помощью реакции дезаминирования, протекающей в гетерогенной фазе, может быть получено дополнительное количество м-КПС. Для этого полученный клеточный осадок промывают, а затем суспендируют водой. К полученной суспензии добавляют при перемешивании 10-50%-ый водный раствор уксусной кислоты и после этого добавляют 3-8 % водный раствор нитрита натрия. Полученную реакционную смесь перемешивают в течение 0,5-2 часов при температуре 4 - 25°С, после чего ее разбавляют водой и центрифугируют. Полученный супернатант нейтрализуют водным раствором неорганического основания, выбранного из NaOH, KOH, NH4OH, или, преимущественно, твердым NaHCO3. Далее полученный раствор подвергают диализации или диафильтрации с последующей лиофилизацией с получением м-КПС (соединения общей формулы (I)).
В некоторых частных вариантах для получения м-КПС из клеточной массы 20 г влажного клеточного осадка суспендируют в 100-300 мл деионизованной воды (предпочтительно, в 200 мл деионизованной воды), центрифугируют, и полученный осадок обрабатывают еще раз описанным выше способом. Промытый осадок суспендируют в 10 - 30 мл деионизованной воды (предпочтительно, в 20 мл деионизованной воды), к полученной суспензии прибавляют при перемешивании 15 - 45 мл (в наиболее предпочтительном варианте 30 мл) раствора уксусной кислоты с концентрацией от 10 до 50 мас % (в наиболее предпочтительном варианте 30 мас %) и затем 15 - 45 мл (в наиболее предпочтительном варианте 30 мл) водного раствора нитрита натрия с концентрацией от 3 до 8 мас % (в наиболее предпочтительном варианте 5 мас %). Через 0,5 - 2 часа (в наиболее предпочтительном варианте 1 час) перемешивания при температуре 4 - 25°С (в наиболее предпочтительном варианте 20°С) реакционную смесь разбавляют в 3 раза деионизованной водой и центрифугируют. Супернатант нейтрализуют водным раствором неорганического основания (NaOH, KOH, NH4OH), или, преимущественно, твердым NaHCO3. Полученный раствор далее диализуют против деионизованной воды или подвергают диафильтрации с использованием мембраны с пределом отсечения 10 кДа, 20 кДа, 50 кДа или 100 кДа в зависимости от молекулярной массы полученного м-КПС и полученный водный раствор лиофилизуют.
Осуществление изобретения подтверждается следующими примерами.
Пример 1. Культивирование S. pneumoniaе и выделение п-КПС
I. Культивирование S. pneumoniaе
Клинические изоляты бактериальных штаммов S. pneumonia 20-тиразличных эпидемически значимых серотипов (1, 3, 4, 5, 6А, 6В, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 15F, 18C, 19F, 19A, 22F, 23A, 23F, 33F) были получены из коллекции «Национального исследовательского центра эпидемиологии и микробиологии имени почетного академика Н.Ф.Гамалеи» ФГБУ МЗ РФ. Культивирование штаммов проводили методом глубинного культивирования по известной методике [Chase, M.W., Methods of Immunology and Immunochemistry 1, 52 (1967)] в 10-литровом ферментере при температуре 37°С с использованием питательной среды, содержащей дрожжевой экстракт (ультрафильтрат) и декстрозу (25 г/л). Культивирование продолжалось 8-26 часов в зависимости от серотипа S. pneumoniaе при контроле рН 6.0-7.2 добавлением 1н раствора NaOH. Процесс завершался при оптической плотности 2.5-3.5 (измерение при 660 нм) и прекращении поглощения декстрозы, о чем свидетельствовала неизменность величины рН в течение 20-30 минут. Немедленно в ферментер прибавляли 90%-ный водный раствор фенола до концентрации 1% и перемешивание продолжали 12-14 часов при комнатной температуре.
II-III. Отделение надклеточной жидкости и выделение неочищенного п-КПС S. pneumoniaе
Суспензию бактериальных клеток подвергли тангенциальной микрофильтрации с использованием мембраны 0.22 мкм (Владисарт), фильтрат сконцентрировали, для удаления компонентов питательной среды и низкомолекулярных продуктов, образующихся в процессе культивирования, провели диафильтрацию (0.2М хлорид натрия, затем деионизованная вода) с использованием мембраны с пределом отсечения 100 кДа. После концентрирования и лиофилизации получили неочищенный п-КПС с выходом 20-110мг/л в зависимости от серотипа S. pneumoniaе.
IV. Получение очищенного п-КПС S. pneumoniaе
К раствору неочищенного п-КПС в 0.2М хлориде натрия (10 мг/ мл) рН 7.2-7.4, добавляли рибонуклеазу (30-100 мкг/мл) и дезоксирибонуклеазу (10-50 мкг/мл), через 1-2 часа перемешивания при температуре 37°С рН смеси доводили до 8.0, добавляли протеиназу К (10-50 мкг/мл) и перемешивание продолжали 16-18 часов при температуре 56°С. Реакционную смесь подвергли диафильтрации (деионизованная вода) и после концентрирования и лиофилизации получали п-КПС S. pneumoniaе, содержащий не более 1-2% примеси нуклеиновых кислот и белков. Определение проводили в соответствии с рекомендациями Государственной Фармакопеи РФ.
В результате проведенной работы были выделены п-КПС более 20 эпидемически значимых серотипов S. pneumoniae. Подтверждение подлинности полученных п-КПС следовало из сопоставления 1Н-ЯМР-спектров выделенных препаратов и соответствующих литературных данных. Анализ опубликованных ранее результатов исследований показал, что п-КПС каждого серотипа S. pneumoniae обладает уникальным набором сигналов в слабопольной области спектра 1Н-ЯМР (сигналы аномерных протонов) и в области сильного поля (сигналы О- и N-ацетильных групп, 6-дезоксигрупп и остатков пировиноградной кислоты), тогда как сопоставление областей циклических протонов спектров в данной ситуации не информативен. Каждый из выделенных п-КПС содержал по данным 1Н-ЯМР-спектроскопии от 5 до 15% ОПАП. Примеры1Н-ЯМР спектров п-КПС S. pneumoniae приведены на Фиг.2А.
Пример 2. Дезаминирование очищенного п-КПС S. pneumoniaе
На этапе V. к раствору 20 мг п-КПС в 1 мл воды добавляли 1.5 мл 33.3 %-ного водного раствора уксусной кислоты (в соотношении CH3COOН : Н2Оот 1:3 до 1:5) и 1.5 мл 5%-ного водного раствора нитрита натрия, через 60 минут перемешивания при комнатной температуре реакционную смесь разбавляли 10 мл деионизованной воды, нейтрализовали бикарбонатом натрия, диализовали против деионизированной воды, диализат лиофилизовали, раствор сухого остатка в деионизированной воде центрифугировали 60 минут при 20000 об/мин, супернатант лиофилизовали. Выход м-КПС (соединения формулы (I)), составлял 11-17 мг в зависимости от серотипа S. pneumoniaе.
Пример1Н-ЯМР спектра м-КПС бактериального штамма S. pneumoniaе серотипа 6А приведен на Фиг.2Б.
Пример 3. Получение м-КПС путем дезаминирования клеток S. pneumoniaе
Клеточную массу, полученную в результате отделения надклеточной жидкости (см. Пример 1), трижды промывали 0,2 M раствором NaCl и далее деионизованной водой. 10 г влажной бактериальной массы суспензировали в 50 мл деионизированной воды, к перемешиваемой суспензии добавляли 75 мл разбавленной уксусной кислоты (в соотношении CH3COOН: Н2О 1:3) и 75 мл 5%-ного водного раствора нитрита натрия. Через 1 час перемешивания суспензию центрифугировали, супернатант нейтрализовали бикарбонатом натрия и после диафильтрации и лиофилизации получили очищенный м-КПС. Выход м-КПС (соединения формулы (I)), составляет 100 -150 мг в зависимости от серотипа S. pneumoniae.
Пример 4. Получение конъюгата м-КПС S. pneumoniaе, серотип 6B и бычьего сывороточного альбумина (БСА) с использованием многоточечной схемы соединения полисахаридного и белкового антигенов
9 мг м-КПС S. pneumoniaе, серотип 6В растворяли в 1 мл 0,01М раствора периодата натрия, через 30 минут перемешивания при температуре 40°С, что приводило к частичному окислению остатков рибитаи образованию альдегидных групп на полисахаридной цепи (см. строение повторяющегося звена полисахарида на стр. 12), к реакционной смеси прибавляли 50 мкл этиленгликоля, диализовали против воды и лиофилизовали. 7,7 мг полученного лиофилизата растворяли в 1 мл 0,1М натрий-ацетатного буфера, рН 5,8, затем прибавляли последовательно 21,8 БСА и 19,5 мг цианборгидрида, через 16 часов перемешивания при 37°С реакционную смесь нейтрализовали разбавленной соляной кислотой и хроматографировали на колонке (1,5 х 75 см) с сефарозой 6В, уравновешенной 0,05н АсОН. Конъюгат (выход 27,0 мг) элюировался вблизи фронта, не вступившего в реакцию БСА в результате гель-хроматографии не обнаружено. Спектр ЯМР содержал две серии близких по интегральной интенсивности сигналов, принадлежащих исходному полисахариду и БСА.
Пример 5. Получение конъюгата м-КПС S. pneumoniaе, серотип 6B и бычьего сывороточного альбумина (БСА) с использованием активной альдегидной группы остатка 2.5-ангидроманнозы,
К раствору 5,2 мг м-КПС S. pneumoniaе, серотип 6B в 1 мл 0,1М натрий-ацетатного буфера, рН 5,8 прибавляли последовательно 9,8 БСА (избыток) и 9,5 мг цианборгидрида натрия, через 16 часов перемешивания при 37°С реакционную смесь нейтрализовали разбавленной соляной кислотой и хроматографировали на колонке (1,5 х 75 см) с сефарозой 6В, уравновешенной 0,05н АсОН. Конъюгат элюировался вблизи фронта, тогда как БСА (избыток), не вступивший в реакцию, элюировался значительно позже. Выход конъюгата после лиофилизации составил 5,7 мг. Спектр ЯМР конъюгата представлял собой суперпозицию мажорных сигналов спектра исходного м-КПС S. pneumoniaе, серотип 6B и минорных сигналов, характерных для белков.
Пример 6. Исследование антигенной активности м-КПС S. pneumoniaе, серотип 6B
Исследование проводили на мышах линии C57BL/6 (5-6 недель, поставщик ФГБУН «Научный центр биомедицинских технологий федерального медико-биологического агентства» филиал «Столбовая»), методом трехкратной подкожной иммунизации с двухнедельным перерывом между введениями (дни 0, 14, 28). Вес животных к началу введения составлял 18-20 г. Препарат вводили в двух дозах - 0,5 и 5 мкг/животное м-КПС серотипа 6B. В исследование были отобраны животные без признаков отклонений в состоянии здоровья (после клинического осмотра и карантина) с учетом массы тела: разброс массы тела между животными, отобранными в группы, не превышал 10%, среднее значение массы тела статистически не различалось между группами. Животных распределяли по группам случайным образом. Исследуемые группы и дозы приведены в таблице 1.
Перед первой иммунизацией (0 день), перед второй иммунизацией (14 день) и через неделю после третьей иммунизации (35 день) проводили забор образцов крови с последующим получением сывороток. Для всех образцов сывороток проводили анализ продукции иммуноглобулинов классов M и G (IgM и IgG) методом твердофазного иммуноферментного анализа (ИФА); для образцов сывороток, собранных на 35 сутки после первой иммунизации, дополнительно проводили анализ бактериостатической активности с использованием бактериального штамма S. pneumoniae серотипа 6B.
I. Динамика набора массы животными в экспериментальных группах
В результате проведенного исследования было установлено, что динамика набора веса животными контрольной и опытных групп не различалась (Фиг.4 А, Б). Статистически значимые отличия веса мышей не были выявлены вплоть до конца наблюдения.
II. Оценка уровня продукции специфических иммуноглобулинов IgG и IgM методом ИФА
Уровень специфических антител к м-КПС бактериального штамма S. pneumoniae серотипа 6В в сыворотке крови мышей после трехкратной иммунизации оценивали методом непрямого неконкурентного ИФА. Для этого предварительно проводили сорбцию антигена (0,4 мкг/лунку м-КПС серотипа 6B) на плашку (Greiner, Германия) в течение ночи при 4°С. Сыворотку крови мышей разводили ИФА-буфером S011 (Хема, Россия) в соотношении 1:100. Для детекции специфических иммуноглобулинов IgG и IgM использовали, соответственно, конъюгат с пероксидазой хрена поликлональных антител козы к IgG мыши (0,8 мг/мл, HyTest) и конъюгат с пероксидазой хрена поликлональных антител козы к IgM мыши 100X (Alpha Diagnostic H-MsM.211) в соответствии с инструкцией производителя.
Проведенное исследование специфических IgG и IgM к капсульным полисахаридам в сыворотке крови мышей подтвердило формирование иммунного ответа на введение м-КПС серотипа 6B и его достоверное увеличение по сравнению с контрольной группой PBS, начиная с момента введения первой дозы (см. Фиг.5А, Б). При этом уровень продукции антител носит дозозависимый характер и повышается при увеличении дозы иммунизации.
III. Оценка бактериостатической активности сыворотки крови животных после иммунизации
Бактериостатический эффект оценивали по способности сыворотки крови животных ингибировать рост бактериальной культуры исследуемого штамма (S. pneumoniae серотипа 6В) при инкубации в течение 20 часов. Для этого штамм выращивали на питательной среде TSB (триптон-соевый бульон) с добавлением 5% крови КРС (крупного рогатого скота) в течение 18-24 часов при 37°С. Колонии ресуспендировали в стерильном фосфатно-солевом буфере до достижения мутности по стандарту МакФарланда 0,5. Далее культуру разбавляли в 100 раз средой TSB+5% кровь КРС и вносили (50 мкл) в лунки 96-луночного планшета, содержащие 50 мкл исследуемой сыворотки. В качестве контролей использовали смесь бактериальной суспензии и среды (контроль роста) и питательную среду (контроль стерильности). После инкубации полученных смесей при 37°С в течение 20 часов, содержимое лунок хорошо перемешивали и измеряли оптическую плотность при длине волны 600 нм (OD600).
Спустя 20 часов инкубации сывороток с бактериальным штаммом было выявлено снижение роста бактериальной культуры в сыворотке крови животных опытной группы м-КПС 5 мкг, по сравнению с контрольной группой PBS; для сыворотки крови животных опытной группы м-КПС 0,5 мкг статистически значимых эффектов не наблюдалось. Так же, как и в случае исследования иммуногенности, наблюдается дозозависимый эффект введения (см. Фиг.6).
Таким образом, в результате проведенного эксперимента было показано, что введение м-КПС не приводит к серьезным изменениям в общем состоянии животных (не влияет на динамику набора массы в группах), обладает иммуногенностью, вызывая формирование специфических иммуноглобулинов классов M и G в дозозависимой манере, кроме того, приводит к повышению бактериостатической активности сыворотки крови животных в отношении соответствующего штамма S. pneumoniae, что позволяет применять м-КПС в качестве антигена при разработке пневмококковых вакцин на основе полисахаридов.
Пример 7. Анализ различий эпитопного состава м-КПС и п-КПС S. pneumoniaе
Поскольку способ получения КПС может влиять на его эпитопный состав как антигена и, как следствие, на иммуногенные свойства, была проведена оценка возможных различий антигенной структуры м-КПС и п-КПС, полученных из бактериальных штаммов S. pneumoniae серотипа 6В. Для этого методом твердофазного ИФА была исследована способность антител, полученных к м-КПС, связываться с п-КПС.
Образцы сыворотки крови животных, полученные на 35 сутки после первой иммунизации, исследовали методом непрямого неконкурентного ИФА по протоколу ВОЗ (http://www.vaccine.uab.edu/). В лунки ИФА планшетов (Greiner MED BINDING, Ref 655001) вносили по 5 мкг п-КПС или 0,5 мкг м-КПС и сорбировали планшеты 5 часов при 37°С, а затем ночь в холодильнике. Перед использованием планшеты трижды отмывали PBS c 0,1% Tween-20. Сыворотку крови мышей, иммунизированных м-КПС в дозах 0,5 и 5 мкг (см. Пример 4), разводили 1:100 в ИФА-буфере S011 (Хема, Россия) и вносили в лунки по 100 мкл. Планшеты с образцами инкубировали в термошейкере в течение 1 часа при 37°С и перемешивании со скоростью 600 об/мин. После окончания инкубации лунки трижды отмывали от не связавшихся антител, внося в них по 300 мкл PBS с 0,1% Твин-20, после чего инкубировали с поликлональными антивидовыми антителами козы к IgG мыши, конъюгированными с пероксидазой хрена (HyTest, Finland) в разведении 1:25 000. Затем планшеты отмывали 6 раз и визуализировали реакцию, внося в лунки по 100 мкл субстратного буфера с тетраметиленбензидином. Через 15 минут реакцию останавливали 10% раствором HCl и на планшетном спектрофотометре SpectrostarNano (BMG Labtech) измеряли в лунках оптическую плотность при длине волны 450 нм. Результаты измерения уровня антител IgG к п-КПС и м-КПС представлены на Фиг.7.
Как видно на приведенной диаграмме, в условиях сорбции, оптимальных для п-КПС и м-КПС, статистически значимых различий уровней IgG в группах мышей, иммунизированных м-КПС в дозах 0,5 и 5 мкг/животное, не выявлено. Таким образом, антитела, полученные к м-КПС, способны распознавать п-КПС, что может свидетельствовать о наличии у этих препаратов общих эпитопов. Различие уровней IgG, связавшихся с антигеном, в группе контрольных мышей (PBS) может быть обусловлено примесями ОПАП в препарате п-КПС.
Таким образом, технический результат изобретения подтверждается тем, что получены модифицированные капсульные полисахариды (м-КПС), не обладающие недостатками, присущими п-КПС, а именно, не содержащие примеси балластного общего полисахаридного антигена S. pneumoniae. Полученные м-КПС содержат остаток 2,5-ангидро-D-маннозы на восстанавливающем конце, альдегидная группа которой может быть использована для конъюгации с аминосодержащей матрицей белковой или иной природы, без необходимости дополнительной активации м-КПС при проведении конъюгации, при этом полученные м-КПС обладают более высокой растворимостью в водных буферах, чем п-КПС, что позволяет повысить технологичность процессов конъюгации и могут быть использованы для получения поливалентных пневмококковых полисахаридных вакцин, а также пневмококковых гликоконъюгированных вакцин.
Промышленная применимость
Полученные модифицированные капсульные полисахариды (м-КПС), не содержащие примеси балластного общего полисахаридного антигена S. pneumonia и содержащие остаток 2,5-ангидро-D-маннозы на восстанавливающем конце, альдегидная группа которой может быть использована для конъюгации с аминосодержащей матрицей белковой или иной природы, без необходимости дополнительной активации м-КПС при проведении конъюгации могут быть использованы для получения конъюгированных вакцин против S. pneumoniae.
Цитируемые документы
1. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016 / C. Troeger, B. Blacker, I.A. Khalil et al. // The Lancet Infectious Diseases. - 2018. - V.18. - P.1191-210.
2. Immunologic responses of children to serious infections with Streptococcus pneumoniae / C.G. Prober, H. Frayha, M. Klein et al. // J Infect Dis - 1983. - V.148, No.3. - P.427-435.
3. Covalent linkage between the capsular polysaccharide and the cell wall peptidoglycan of Streptococcus pneumoniae revealed by immunochemical methods // Uffe B.SkovSørensen, JørgenHenrichsen, Hao-ChiaChen, Shousun ChenSzu. //Microbial Pathogenesis - 1990. - V.8, Issue 5, - P. 325-334.
4. Streptococcus pneumoniae capsular polysaccharide is linked to peptidoglycan via a direct glycosidic bond to β-D- N-acetylglucosamine.// Thomas R Larson, Janet Yother // Proc Natl Acad Sci U S A. 2017 May 30;114(22):5695-5700. doi: 10.1073/pnas.1620431114.
5. The pneumococcal common antigen C-polysaccharide occurs in different forms. Mono-substituted or di-substituted with phosphocholine.// C Karlsson 1, P E Jansson, U B Skov Sørensen.// Eur J Biochem. 1999 Nov;265(3):1091-7. doi: 10.1046/j.1432-1327.1999.00835.x.
6. Removal of cell wall polysaccharide in pneumococcal capsular polysaccharides by selective degradation via deamination. // Wei Zou, Jianjun Li, Evguenii Vinogradov, Andrew Cox. // Carbohydrate Polymers. — 2019 V. 218, P. 199-207
7. Deamination of carbohydrate amines and related compounds // A Williams // Adv. Carbohydr. Chem. Biochem. - 1975 -31, N1, 9-79
8. Accelerated method of purification for obtaining capsular polysaccharides of streptococcus pneumoniae.// RU2516340C2
9. The pgdA gene encodes for a peptidoglycan N-acetylglucosamine deacetylase in Streptococcus pneumoniae. // W Vollmer , A Tomasz. // J Biol Chem. 2000 Jul 7;275(27):20496-501.
10. Considerations for formulating the second-generation pneumococcal capsular polysaccharide vaccine with emphasis on the cross-reactive types within groups.// J B Robbins, R Austrian, C J Lee, S C Rastogi, G Schiffman, J Henrichsen, P H Mäkelä, C V Broome, R R Facklam, R H Tiesjema, et al.// J Infect Dis. 1983 Dec;148(6):1136-59. doi: 10.1093/infdis/148.6.1136. PMID: 6361173 DOI: 10.1093/infdis/148.6.1136.
11. Studies on immunological paralysis. V. The influence of molecular weight on the immunogenicity, tolerogenicity and antibody-neutralizing activity of the 3 pneumococcal polysaccharide.// J G Howard, H Zola, G H Christie, B M Courtenay.// Immunology. 1971 Sep;21(3):535-46. PMID: 4398137 PMCID: PMC1408154
12. A Strategy for Rational Design of Fully Synthetic Glycopeptide Conjugate Vaccines. // PELE CHONG,* NEVILLE CHAN, ALI KANDIL, BRIAN TRIPET, OLIVE JAMES, YAN-PING YANG, SHAN-PAN SHI, AND MICHEL KLEIN.// INFECTION AND IMMUNITY Vol. 65, No. 12Dec. 1997, p. 4918-4925.
13. Structural analysis and chemical depolymerization of the capsular polysaccharide of Streptococcus pneumoniae type 1. // Stroop, C.é J.M.; Xu, Q.; Retzlaff, M.; Abeygunawardana, C.; Bush, C.A. // Carbohydrate Research 337(4): 335-344. 2002. ISSN/ISBN: 0008-6215 PMID: 11841814 DOI: 10.1016/s0008-6215(01)00318-4.
14. Characterization of the cassette containing genes for type 3 capsular polysaccharide biosynthesis in Streptococcus pneumoniae. // J P Dillard 1, M W Vandersea, J Yother. // J Exp Med. 1995 Mar 1;181(3):973-83. doi: 10.1084/jem.181.3.973. PMID: 7869055 PMCID: PMC2191931 DOI: 10.1084/jem.181.3.973.
15. Type 3-specific synthase of Streptococcus pneumoniae (Cap3B) directs type 3 polysaccharide biosynthesis in Escherichia coli and in pneumococcal strains of different serotypes.//C Arrecubieta 1, R López, E García.// J Exp Med. 1996 Aug 1;184(2):449-55. doi: 10.1084/jem.184.2.449. PMID: 8760798 PMCID: PMC2192726 DOI: 10.1084/jem.184.2.449.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИВАЛЕНТНАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ КОНЪЮГАТЫ ПНЕВМОКОККОВЫЙ ПОЛИСАХАРИД-БЕЛОК | 2013 |
|
RU2605834C2 |
| ПНЕВМОКОККОВЫЕ ПОЛИСАХАРИДЫ И ИХ ПРИМЕНЕНИЕ В ИММУНОГЕННЫХ КОНЪЮГАТАХ ПОЛИСАХАРИД-БЕЛОК-НОСИТЕЛЬ | 2018 |
|
RU2784449C2 |
| ПНЕВМОКОККОВЫЕ ПОЛИСАХАРИДЫ И ИХ ПРИМЕНЕНИЕ В ИММУНОГЕННЫХ КОНЪЮГАТАХ ПОЛИСАХАРИД-БЕЛОК-НОСИТЕЛЬ | 2018 |
|
RU2801304C2 |
| ПНЕВМОКОККОВЫЕ ПОЛИСАХАРИДЫ И ИХ ПРИМЕНЕНИЕ В ИММУНОГЕННЫХ КОНЪЮГАТАХ ПОЛИСАХАРИД-БЕЛОК-НОСИТЕЛЬ | 2018 |
|
RU2785429C2 |
| ПНЕВМОКОККОВЫЕ ПОЛИСАХАРИДЫ И ИХ ПРИМЕНЕНИЕ В ИММУНОГЕННЫХ КОНЪЮГАТАХ ПОЛИСАХАРИД-БЕЛОК-НОСИТЕЛЬ | 2018 |
|
RU2801288C2 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2688831C2 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2743793C1 |
| ИММУНОГЕННЫЙ КОНЪЮГАТ БЕТА-ПРОПИОНАМИД-СВЯЗАННОГО ПОЛИСАХАРИДА С БЕЛКОМ, ИСПОЛЬЗУЮЩИЙСЯ В КАЧЕСТВЕ ВАКЦИНЫ | 1999 |
|
RU2249463C2 |
| МУЛЬТИВАЛЕНТНАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КОНЪЮГАТА ПНЕВМОККОВЫЙ ПОЛИСАХАРИД-БЕЛОК | 2007 |
|
RU2493870C2 |
| МУЛЬТИВАЛЕНТНАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КОНЪЮГАТА ПНЕВМОККОВЫЙ ПОЛИСАХАРИД-БЕЛОК | 2007 |
|
RU2484846C2 |



Изобретение относится к области биотехнологии. Описана группа изобретений, включающая соединение общей формулы 4-О-(β-D-N-ацетилмурамил)-6-О-R-2,5-ангидро-D-манноза, где R - остаток капсульного полисахарида S. Pneumoniae любого серотипа, за исключением серотипов 1 и 3, применение вышеуказанного соединения для получения полисахарид-белковых вакцин против S. Pneumoniae и способ получения вышеуказанного соединения. В одном варианте реализации остаток капсульного полисахарида представляет собой остаток капсульного полисахарида штамма S. pneumoniae серотипа 4, 5, 6А, 6В, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 15F, 18C, 19F, 19A, 22F, 23A, 23F или 33F. Изобретение позволяет получать модифицированные капсульные полисахариды S. pneumoniae, не содержащие примеси общего полисахаридного антигена пневмококков. 3 н. и 4 з.п. ф-лы, 7 ил., 1 табл., 7 пр.
1. Соединение общей формулы 4-О-(β-D-N-ацетилмурамил)-6-О-R-2,5-ангидро-D-манноза
 ,
,
где R - остаток капсульного полисахарида S. pneumoniae любого серотипа, за исключением серотипов 1 и 3, а Lac – обозначает лактил[-CH(CH3)COOH].
2. Соединение по п. 1, в котором остаток капсульного полисахарида представляет собой остаток капсульного полисахарида штамма S. pneumoniae серотипа 4, 5, 6А, 6В, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 15F, 18C, 19F, 19A, 22F, 23A, 23F или 33F.
3. Применение соединения по любому из пп. 1, 2 для получения полисахарид-белковых вакцин против S. pneumoniae.
4. Применение по п. 3, в котором конъюгированная полисахарид-белковая вакцина представляет собой конъюгат с многоточечным соединением по любому из пп. 1, 2 и белковой матрицы.
5. Применение по п. 4, в котором конъюгированная полисахарид-белковая вакцина представляет собой конъюгат, в котором соединение по любому из пп. 1, 2 присоединяется к белковой матрице с участием только активной альдегидной группы -СНО остатка 2,5-ангидро-D-маннозы с образованием звездной структуры конъюгата.
6. Применение по п. 5, в котором белковая матрица представляет собой бычий сывороточный альбумин, столбнячный анатоксин, холерный анатоксин, мутантный дифтерийный анатоксин CRM197, рекомбинантный детоксифицированный токсин CRM197 или консервативный белковый антиген пневмококка.
7. Способ получения соединения по любому из пп. 1, 2, включающий следующие последовательные этапы:
i) культивируют S. pneumoniae с получением культуральной жидкости S. pneumoniae;
ii) проводят центрифугирование или микрофильтрацию полученной культуральной жидкости с разделением надклеточной жидкости и клеточной массы;
iii) выделяют неочищенный исходный капсульный полисахарид (КПС) S. pneumoniae путем ультра- и диафильтрации полученной на этапе ii) надклеточной жидкости с последующей лиофилизацией; получают очищенный исходный КПС S. pneumoniae путем обработки лиофилизата нуклеазами и затем протеиназой К с последующей очисткой диафильтрацией и лиофилизацией;
iv) проводят реакцию дезаминирования исходного КПС S. pneumoniae добавлением 3-8 мас.% водного раствора нитрита натрия к раствору очищенного исходного КПС в 10-50 мас.% водном растворе уксусной кислоты;
или
iii) промывают и суспендируют клеточную массу, полученную на этапе ii), деионизированной водой с получением клеточной суспензии;
iv) проводят реакцию дезаминирования путем добавления 3-8 мас.% водного раствора нитрита натрия к реакционной смеси, представляющей собой смесь полученной клеточной суспензии с 10-50 мас.% водным раствором уксусной кислоты;
v) реакционную смесь, полученную в результате реакции дезаминирования на этапе iv), разбавляют деионизированной водой и затем нейтрализуют путем добавления NaOH, KOH, NH4OH или NaHCO3;
vi) полученный раствор подвергают диализации или диафильтрации с последующей лиофилизацией с получением целевого соединения.
| Приспособление для нивелирования по оборудованной трасе телефонно-телеграфных линий | 1933 |
|
SU34157A1 |
| Дифференциальный манометр | 1933 |
|
SU39427A1 |
| US 11642406 B2, 09.05.2023 | |||
| US 20220105169 A1, 07.04.2022 | |||
| WO 2019139692 A2, 18.07.2019. | |||

