1
Изобретение относится к сельскому хозяйству, в частности к способам выявления холодоустойчивости растений.
Цель изобретения - ускорение процесса определения устойчивости отдельно взятого растения.
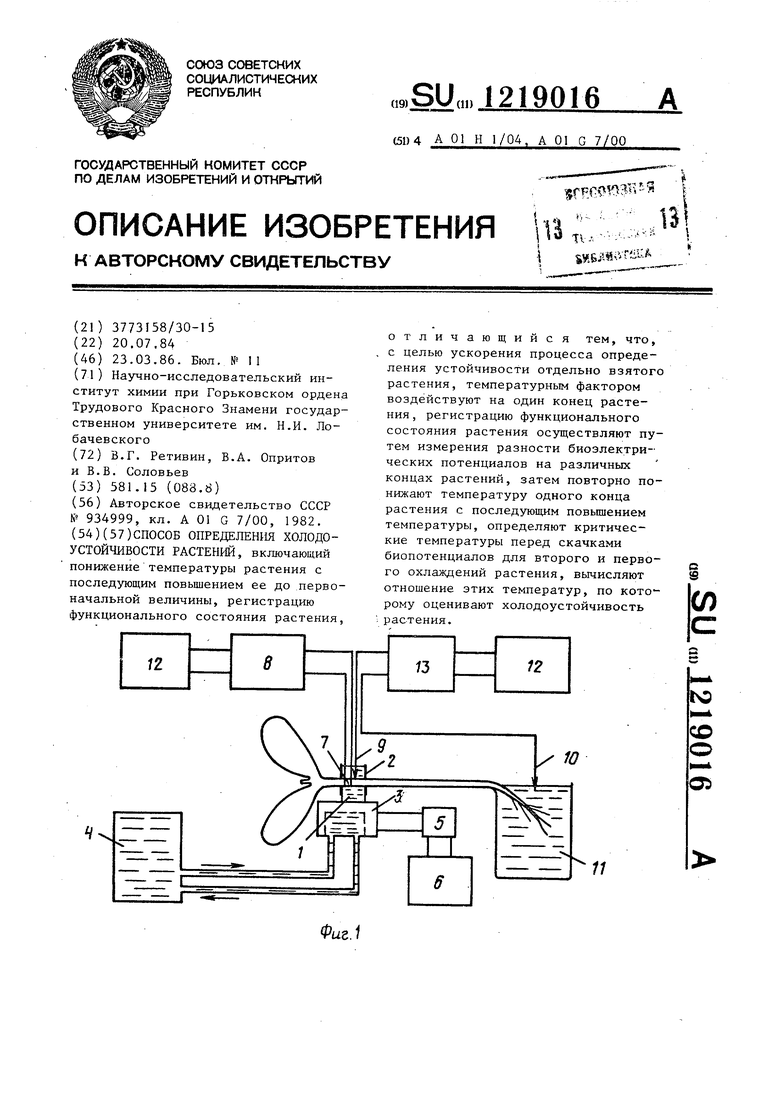
На фиг. 1 представлено устройство для реализации способа определения холодоустойчивости растений; на фиг. 2 - графики зависимостей биопотенциалов на концах растений во времени при термических воздействиях.
Устройство реализации способа содержит кювету 1 с боковьми прорезями 2, заполненную водой с температурой 21-23 С. Дно кюветы 1 контактирует с поверхностью полупроводникового охлаждающего элемента 3. Жидкость, охлаждающая элемент, подают из термостата 4, Охлаждающий элемент 3 связан через переключатель 5 с выпрямителем 6. В кювете 1 установлен термистор 7, расположенный на уровне нижней поверхности укрепленного стебля и электрически связанный с уравновешенным мостом В. Измерительный хлорсеребряный электрод 9 контактирует с водой в кювете а электрод 10 для сравнения с омывающим корни раствором 11. Электроды 9 и 10 связаны с самопшпущим потенциометром 12 и рИ - метром 3.
Способ осуществляют следующим образом.
Участок стебля или листового черешка растения постепенно, с градиентом.О ,5-3 C/f iHH, охлаждают в зоне жидкостного контакта с измерительным электродом 9. Электрод для сравнения располагают наозначительном удалении от первого, напримерS он контактирует, с корнями или с листовой пластинкой. Ведут синхронную запись изменения разности потенциалов между электродами 9 и 10 и температуры. Последнюю снртаают от исходной до любой низкой полохсительной температуры, но всегда не менее, чем до критичр-ской температуры возникновения фазы быстрой негативации участка стебля растения в зоне измерительного электрода, что соответствует быстрой деполяризации охлаждаемых клеток. Затем с тем же или близким градиентом восстанавливают исходную температуру. Фиксируют температур190162
ный режим как на верхнем, так и на нижнем уровне.
Последующий циклический термоимпульс подают нерез временной интер- 5 вал, подбираемый для данного вида растений таким образом, что не происходит полного восстановления исходной разности потенциалов между электродами 9 и 10 после первого воздей- 0 ствия. В этом случае происходит смещение критической температуры возникновения резкой деполяризации охлаждаемых клеток в сторону более низких температур. Смещение температур- 5 ного порога характеризует адаптивные способности исследуемого растения к холодовому воздействию, т.е. холодоустойчивость. Определение холодоустойчивости можно проводить как на 20 целых растениях в любой фазе онтоге- неза, так и на заранее отделенных органах (побегах или отдельных че-. решках).
Пример. Выбирают огурцы сред- 25 неспелого сорта Перосимый 40, которые прора.шд1вают на фильтровальной бумаге при 22-23 С. Половина растений подвергается процедуре закаливания. Для этого их выдерживают при 30 1 и°С с 3 по 7 день (считая с момента замачивания) и затем возвращают в исходные условия. Контрольные растения все время находятся в исходных условиях. Известно, что после 35 процедуры закаливания растения становятся более холодоустойчивыми. Сравнение проводят между контрольными и закаленными растениями на 9-iO день. Поскольку сравнивают 40 растения не только одного вида, но и одного сорта, т.е. одного и того же генотипа, то все об.наруженные изменения в характере ответа на термоимпульс должны быть отнесены 45 к различию устойчивости растений к холодовому воздействию. .
Характер восстановительных процессов, т.е. медленной реполяри- зации, протекающей при возвращении 50 температуры на исходный уровень, также различен у закаленных и незакаленных растений. Но наиболее яркие различия наблюдаются в ходе ответной реакции на последующее 55 воздействие, которое наносили через 15 мин после окончания первого термоимпульса. За этот промежуток времени закаленные холодоустойчивые растения восстанавливают свои свойства в большей степени и поэтому обнаруживают меньший сдвиг критических температур быстрой деполяризации в сторону охлаждения, чем незакаленные. По сравнению с последними они начинают отвечать резкой деполяризацией раньше по времени, т.е. при более высоких температурах, чем незакаленные.
На графиках приведены типичные результаты опыта на закаленных растениях (изображено сплошной линией: .1 - первое воздействие 1.2 - второе воздействие) и контрольного опыта (изображено пунктиром: 2.1 и 2.2 - первое и второе воздействие соответственно) с проростками огурцов сорта Неросимьш 40. Точками обозначено изменение температуры. Как видно из совмещенной записи эксперимента, первоначально наблюдается медленное, изменение регистрируемого параметра разности потенциалов между электродами. Очевидно сначала протекает пассивный процесс уменьшения мембранного потенциала охлаждаемых клеток в соответствии с линейной зависимостью электродиффузионных процессов на плазматических мембранах от температуры. Затем при некой критической температуре Тц,р , характерной для каждого тдельного растения, наблюдается рез. 90 64
Кое увеличение скорости деполяризации постепенно oxлaлцae Iыx клеток. Оно уже не объясняется простыми физико-химическими закономерностями и
5 является активной реакцией клеток растения. Характерно, что деполяризация клеток у закаленных растений сменяется интенсивной реакцией репо- ляризации, т.е. восстановления мем10 бранного потенциала. Продолжающееся действие охлаждения снова вызывает деполяризацию, затем реполяризацию и так с затуханием несколько раз вплоть до выхода на новый устойчи15 вый уровень. У незакаленных растений кинетика перехода обычно имеет более простой характер.
В качестве индивидуального показателя холодоустойчивости каждого
20 исследованного растения относительный коэффициент холодовой адаптации К, который равен отношению критических температур возникновения реакций быстрой деполяризации при пов25 торном (в данном примере через
5 мин) и исходном термовоздействиях
К - /Т ,р г (к,-Т
,/т,
/т.
кр 1.7 itPl. l
0,528).
- 0,869; TI-TI P,/А кр г.1 Чем меньше коэффициент, тем хуже адаптационные способности, тем хуже устойчивость к холодовому воздействию.
Таким образом ускорягот процесс определения устойчивости растения.
.2
вништи
Тирахс 679
Заказ 1171/4 Подписное
Филиал Ш1П Патент, г.Ужгород, ул.Проектная,

| название | год | авторы | номер документа |
|---|---|---|---|
| Способ оценки холодоустойчивости растений огурца | 1990 |
|
SU1701175A1 |
| СРЕДСТВО ДЛЯ ПОВЫШЕНИЯ ХОЛОДОУСТОЙЧИВОСТИ РАСТЕНИЙ | 2000 |
|
RU2195809C2 |
| Способ оценки холодоустойчивости растений | 1990 |
|
SU1732857A1 |
| Способ определения устойчивости растения к изменению фактора внешней среды | 1983 |
|
SU1123586A1 |
| Средство для повышения холодоустойчивости и продуктивности растений | 2019 |
|
RU2732627C1 |
| Способ получения холодоустойчивого посадочного материала батата | 2022 |
|
RU2787700C1 |
| Способ диагностики морозоустойчивости растений винограда | 1986 |
|
SU1450787A1 |
| СПОСОБ ЭЛЕКТРОСТИМУЛЯЦИИ ЖИЗНЕДЕЯТЕЛЬНОСТИ РАСТЕНИЙ | 2002 |
|
RU2261588C2 |
| СПОСОБ ОЦЕНКИ СТРЕССОУСТОЙЧИВОСТИ РАСТЕНИЙ | 2009 |
|
RU2412585C1 |
| Способ определения устойчивости растений к стрессовым воздействиям | 1990 |
|
SU1761047A1 |
| Способ определения низких и высоких температурных границ зон закаливания растений | 1980 |
|
SU934999A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
