Изобретение относится к птицеводству, в частности к способам подбора птицы на усиление эффекта гетерозиса по признакам естественной резистентности и адаптационной пластичности потомства.
Известен способ повышения жизнеспособности птицы методом межлинейной и межпородной гибридизации при подборе линий кур для скрещивания в 5-6-месячном возрасте по степени их гетерозиготности на основании полиморфизма овальбумина и овоглобулина белков яиц (а.с. СССР N 655370, A 01 K 67/00, 1975). Недостатком этого способа является то, что требуются значительные средства для комплектования лабораторий и отбора особей уже в период эксплуатации птицы.
Известны способы подбора линий и определения сочетаемости, основанные на экспериментальных данных, полученных при поликроссах и диаллельных скрещиваниях (Сергеев В.А. и др. Оценка комбинационной способности яйценоских линий кур методом диаллельных скрещиваний. Киев: Птахiвництво, 1982, с. 48). Недостатком этих способов является невозможность предсказания сочетаемости линий без экспериментальных скрещиваний, что связано с большими затратами труда и средств.
Наиболее близким к предлагаемому является способ отбора племенной птицы в суточном возрасте, где проводят визуальную оценку молодняка по фенотипическим признакам и формируют однородные группы по продолжительности периода эмбрионального развития (ПЭР) в пределах их физиологической нормы с интервалом 12 ч (патент N 1743519). Недостатком способа является то, что в нем не учитываются показатели естественной резистентности, зависящие от природы генотипа и условий выращивания, поэтому массовая селекция птицы, направленная на повышение продуктивности без учета естественной резистентности, может привести к понижению сохранности поголовья.
Сущность изобретения заключается в том, что в суточном возрасте формируют стада птицы с интервалом 12 ч однородных по продолжительности эмбрионального развития (ПЭР) в пределах физиологической нормы. В период бонитировки (49-56-дневный возраст) или комплектования стада (120-126-дневный возраст) проводят однородный подбор исходных пар по ПЭР (селекционные или прародительские стада) с целью получения потомства с достоверной разницей по показателям и лимитам естественной резистентности или разнородный подбор по ПЭР для получения однородного потомства по этим признакам.
Предлагаемый способ был испытан на исходных линиях уток кросса "Медео" в АО "Инта" Кировского района Ставропольского края. На выводе формирования группы, однородные по продолжительности ПЭР (с коротким от 648 до 653 ч и продолжительным от 653 до 672 ч). Для определения влияния исходных пар с различным ПЭР на естественную резистентность и адаптационную пластичность потомства, проводили однородный подбор (самцов и самок с одинаковым ПЭР) и разнородный: в одной группе самцы с коротким ПЭР, а самки с продолжительным; в другой наоборот, выращивали и содержали их в одинаковых условиях.
От сформированных групп по ПЭР в отцовской и материнской линиях делали закладки яиц (1000 шт.), отвод, мечение и выращивание потомства при одинаковых условиях кормления и содержания.
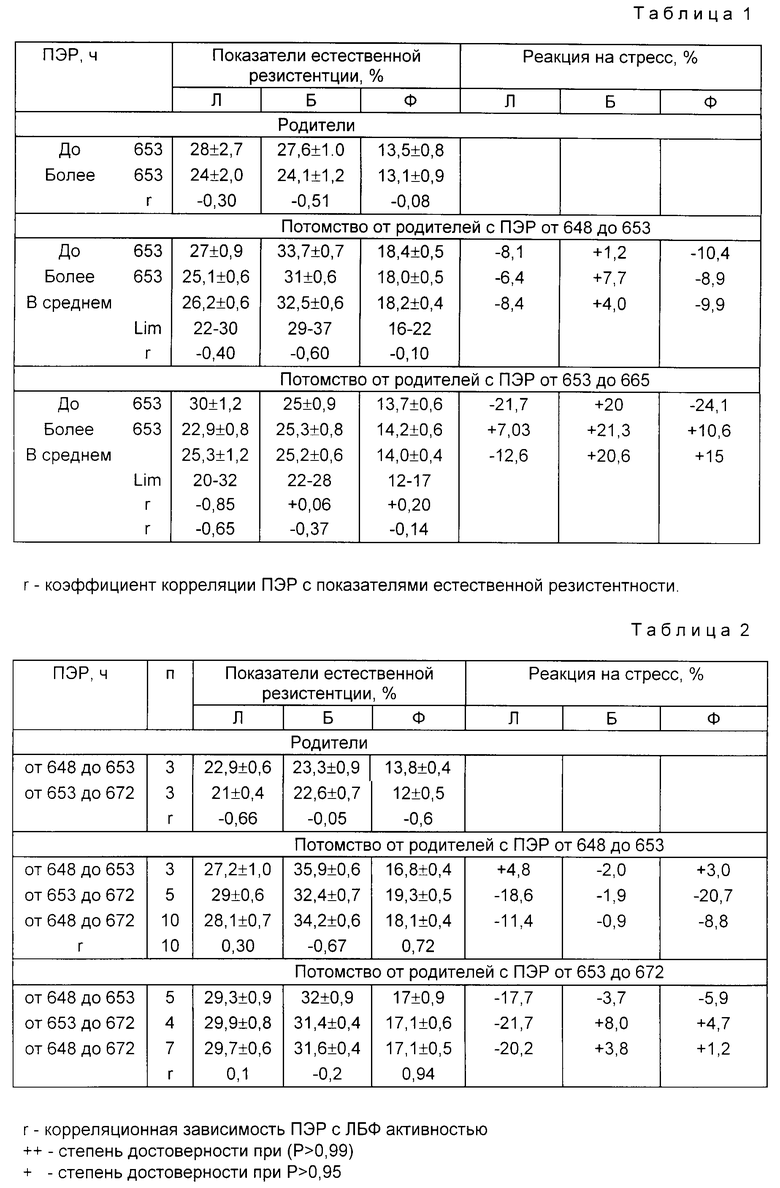
В период бонитировки из каждой группы отбирали методом случайной выборки по 5 голов самцов и самок, брали кровь и определяли показатели естественной резистентности. Данные по наследованию показателей естественной резистентности у потомства по материнской линии кросса "Медео" представлены в табл. 1 и 2, а по отцовской в табл. 3 и 4.
Пример 1. В группу (табл. 1) отбирали потомство (самок) с коротким (от 648 до 653 ч) и продолжительным ПЭР (от 653 до 672 ч) от родителей (самцов и самок) с коротким ПЭР (от 648 до 653 ч.)
Родители самки имели Л. Б. Ф. активность соответственно 28,0±2,7; 27,6±1,0; 13,5±0,8, что достоверно (P>0,95) превосходило самок продолжительного ПЭР соответственно на 3,0; 3,5 и 0,4%
Потомство самки с коротким ПЭР имели Л. Б.Ф. активность 27,0±0,9; 33,7±0,7; 18,0±0,5, что выше на 1,9 (P>0,95); 2,7% (P>0,99) и 0,4% чем у особей с продолжительным ПЭР от этих же родителей. В среднем, все потомство от родителей с коротким ПЭР имело Л.Б.Ф. активность 26,2±0,6; 32,5±0,4% и показатели Lim по этим признакам соответственно; 23,0-30; 29-37; 16-22 и превосходило потомство от родителей с продолжительным ПЭР по активности на 0,9% Б на 7,3% (P>0,999) и Ф на 4,2% (P>0,999), что указывает на наследование этих признаков потомством и сопровождается увеличением корреляционной зависимости ПЭР с показателями Л активности от - 0,30-0,40; Б от 0,5 до 0,60 (P>0,99) и Ф от 0,08 до 0,1.
Пример 2. Во вторую группу (табл. 1) отбирали потомство (самок) с коротким (от 648 до 653 ч) и продолжительным ПЭР (от 653 до 672 ч) от родителей (самцов и самок с продолжительным ПЭР (от 653 до 672 ч). Родители - самки имели показатели Л. Б. Ф активности соответственно: 24,0±2,0; 24,1±1,2; 13,1±0,9.
Потомство самки с короткими ПЭР имело показатели Л.Б.Ф. активности: 30,0±1,2, 25,0±0,9; 13,7±0,6, что достоверно (P>0,999) выше только по лизоцимной активности, чем у потомства с продолжительным ПЭР от этих же родителей. В среднем все потомство от родителей с продолжительным ПЭР, как и родители, имели ниже показатели Л.Б. и Ф. активности и Lim, чем у потомства от родителей с коротким ПЭР.
Пример 3. В группу (табл. 2) отбирали потомство (самцов) с коротким (от 648 до 653 ч) и продолжительным (от 653 до 672 ч) ПЭР от родителей с коротким ПЭР (от 648 до 653 ч).
Родители-самцы с коротким ПЭР имели достоверное (P>0б95) преимущественно перед самцами продолжительного ПЭР только по Л и Ф активности.
Потомство самцы с коротким ПЭР имели Л. Б.Ф. активность 27,2±1,0; 35,9±0,6 и 16,8±0,4, что достоверно (P>0,95) выше только по Б активности, чем у самцов с продолжительным ПЭР от этих же родителей. В среднем показатели Л.Б.Ф. активности составили: 28,1±0,7; 34,2±0,6 и 18,1±0,4, что так же как у самок от этих родителей, достоверно выше (кроме лизоцимной активности), чем у самцов от родителей с продолжительным ПЭР. У них также отмечаются более высокие показатели и генетическое разнообразие по этим признакам и подтверждается усилением корреляционной зависимости ПЭР с Б активностью до 0,67 (P>0,999), с Ф до 0,72 (P>0,999), кроме лизоцимной, что указывает на эффективность подбора родителей с коротким ПЭР и влиянию их на показатели естественной резистентности у потомства.
Пример 4. В 4 группу (табл. 2) отбирали потомство (самцов) с короткими ПЭР и продолжительным от родителей с продолжительным ПЭР (от 653 до 672 ч). Родители-самцы имели показатели Л.Б.Ф. активности соответственно: 21,0±0,4, 22,7±0,7 и 12,0±0,5. Потомство самцы с коротким и продолжительным ПЭР от этих родителей имели Л. Б.Ф. активность соответственно: 29,7±0,6; 31,6±0,4; 17,1±0,5, что достоверно (P>0,95) ниже, чем у потомства самцов от родителей с коротким ПЭР (кроме лизоцимной активности).
Внутри популяции особи (самцы) с разным ПЭР не имели между собой закономерно выраженных различий.
По отцовской линии у потомства отмечали аналогичную закономерность по увеличению показателей естественной резистентности (Л.Б.Ф. активность) от родителей с коротким ПЭР и усиление этих различий у потомства второго поколения табл. 3. Однако определение естественной резистентности у потомства без учета его ПЭР, хотя и отражает общую закономерность, но не дает четких различий между ними, что нужно учитывать при выявлении оптимальных сочетаний.
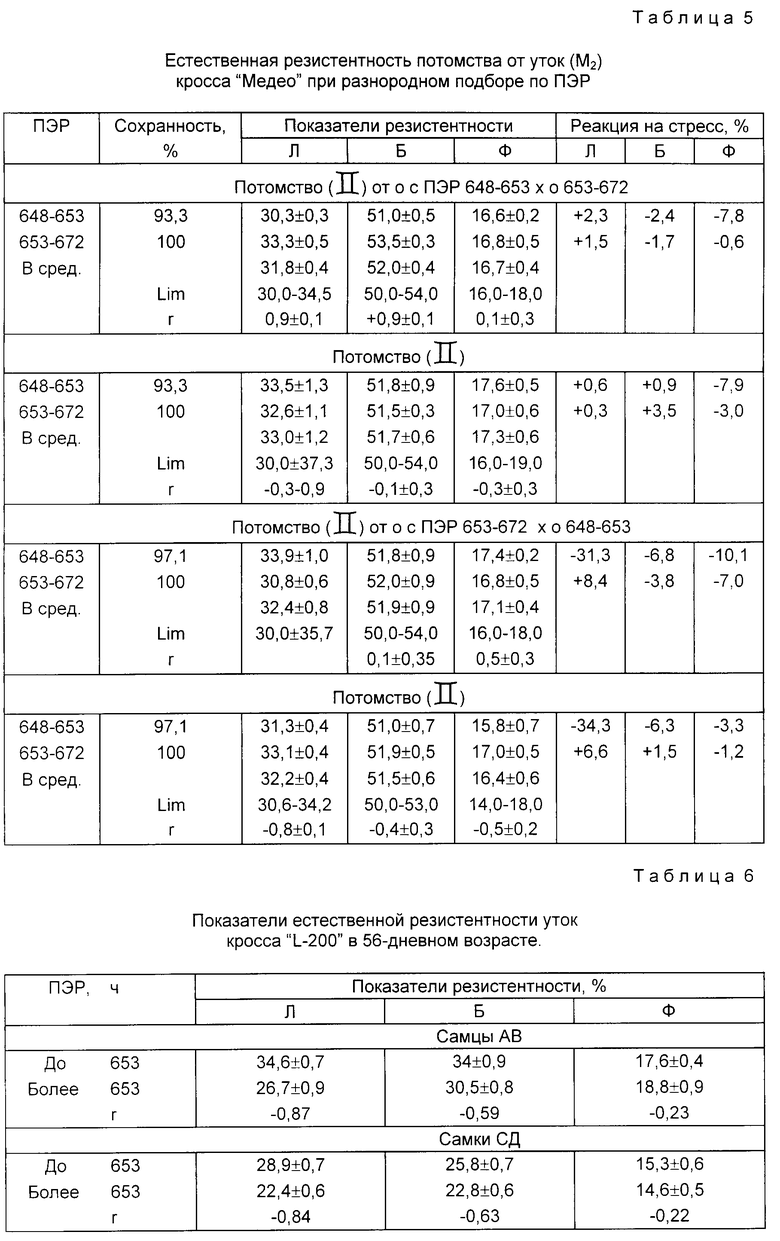
Таким образом при однородном подборе исходных форм уток кросса "Медео" по ПЭР у потомства различия по показателям естественной резистентности возрастают с четким преимуществом по величине лимитов за особями с коротким ПЭР. При разнородном подборе исходных форм по ПЭР однородность потомства по естественной резистентности повышается без существенных различий в лимитах. Эта закономерность проявляется в большей степени у потомства 11 поколения по материнской линии, которому предшествовал однородный подбор по ПЭР (табл. 5). Аналогичную зависимость мы наблюдаем и между потомством от разнородных сочетаний по ПЭР в отцовской линии. От разнородных сочетаний, при более высоких показателях естественной резистентности, у исходных форм был выше вывод утят: по отцовской линии на 5,3% и материнской на 6,8% Как у родителей, так и у потомства выше сохранность на 3,4-1,0% Реакция на стресс более выражена у потомства от тех сочетаний исходных форм, у которых выше уровень естественной резистентности.
Высокие корреляционные связи между ПЭР и показателями естественной резистентности установлены на птице всех изучаемых видов и направлений продуктивности (табл. 6, 7), что дает основание рекомендовать применение способа подбора по продолжительности эмбрионального развития в массовой селекции с целью повышения и контроля за показателями естественной резистентности птицы.


Использование: птицеводство. Сущность изобретения: стада птицы формируют по продолжительности периода эмбрионального развития (ПЭР), а в период бонитировки или комплектования стад для воспроизводства проводят однородный подбор самцов и самок с коротким и продолжительным ПЭР для получения генетически разнообразного по естественной резистентности потомства с достоверной разницей по показателям и лимитах лизоцимной, бактерицидной и фагоцитарной активности - или разнородный подбор по ПЭР родительских пар для получения однородного потомства - без существенных различий и одинаковых лимитах по лизоцимной, бактерицидной и фагоцитарной активности. 6 табл.
Способ подбора исходных пар птицы на повышение естественной резистивности потомства, включающий формирование стад по продолжительности периода эмбрионального развития, отличающийся тем, что в период бонитировки в 49 - 56-дневном возрасте или комплектования стад в 120 126-дневном возрасте используют однородный подбор по продолжительности эмбрионального развития родительских пар для усиления различий у потомства по естественной резистентности или разнородный подбор по продолжительности эмбрионального развития для получения однородного потомства по естественной резистентности.
| SU, авторское свидетельство, 1743519, кл | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
