Изобретение относится к новому штамму бактерий, способному ингибировать рост патогенов растений почвенного происхождения и усиливать рост растений.
Грибковые фитопатогены являются причиной больших экономических потерь в сельском хозяйстве и садоводстве. Было описано множество различных типов грибковых фитопатогенов: эти патогены вызывают болезни растений, такие как вызревание, белая гниль, бурая гниль и корневая гниль. Такие болезни могут погубить появляющиеся проростки, снизить мощность растений и неблагоприятно повлиять на урожай.
Для того чтобы свести к минимуму грибковые инфекции, можно выращивать рассаду на почве, простерилизованной паром или обработанной химическим способом. Однако подобные обработки уничтожают в почве и полезные микроорганизмы, которые в нормальных условиях конкурируют с почвенными грибами. В этих условиях, если в почву будет случайно занесен грибковый патоген, он может быстро распространиться и вызвать широкое распространение болезни.
В сельскохозяйственном производстве почвы, зараженные фитопатогенными грибами, являются неподходящими для выращивания определенных культур. Например, урожаи соевых бобов в Мичигане и в других штатах, выращивающих сою, часто сильно уменьшаются вследствие корневой гнили, вызванной грибом Phytophera megasperma (Filinow and Lockwood, 1985). Виды гриба Pythium широко распространены на части земель в Калифорнии, штате Вашингтон и Айдахо. Pythium ultimum является наиболее часто встречающимся патогенным видом, ответственным за выпревание проростков до и после прорастания. Этот вид является серьезным патогеном пшеницы, гороха и турецкого гороха и других культур, которые выращиваются на этих почвах и на почвах других штатов и других стран (Trapero-Casas и др. , 1990; Stanghellini и Hancock, 1970; Kraft и Burke, 1971; Westerlund и др. , 1988). Применение химических агентов для контроля грибковых фитопатогенов часто не является практичным, так как они дорого стоят, малоэффективны и вызывают появление резистентных штаммов грибов. Помимо этого применение химических фунгицидов нежелательно с точки зрения экологии.
В патенте США N 4595589 описываются штаммы актиномицетов Streptomyces для защиты растений от грибковой инфекции. Кроме того, раскрывается композиция для защиты растений от грибковых инфекций, содержащая актиномицеты Streptomyces и доставочную среду в виде торфа.
Целью настоящего изобретения явилось создание средств биологического контроля для снижения инфекций растений, вызванных грибковыми патогенами.
Поставленная цель была достигнута путем выделения ряда актиномицетов, активных в отношении ингибирования роста грибковых фитопатогенов. В частности, было показано, что один из выделенных актиномицетов, который в настоящем документе именуется Streptomyces WYEC 108 (также упоминается здесь как WYEC 108), демонстрирует сильный антагонизм в отношении целого ряда грибковых патогенов растений, включая патогены, вызывающие болезни растений, известные как выпревание, корневая гниль, белая гниль и бурая гниль. Так одной особенностью настоящего изобретения является биологически чистая культура Streptomyces WYEC 108.
Настоящее изобретение также предлагает различные композиции, удобные для обработки семян или корней растений штаммов Streptomyces WYEC 108. Такие композиции полезны для снижения чувствительности растений к грибковой инфекции и для усиления роста обработанных растений. В предпочтительном варианте осуществления изобретения такие композиции включает биологически чистую культуру Streptomyces WYEC 108 и доставочную среду. В некоторых вариантах осуществления изобретения доставочная среда может включать альгинатный гель, песок или кукурузную муку. В одном варианте осуществления настоящего изобретения поставочная среда, которая включает торфяной мох, песок и кукурузную муку, объединена со Streptomyces WYEC 108. В другом варианте осуществления изобретения доставочная среда включает по меньшей мере 105 колониеобразующих единиц на 1 г доставочной среды.
В еще одном варианте осуществления изобретение включает в себя гранулы альгинатного геля, содержащие Streptomyces WYEC 108. Такие гранулы можно непосредственно добавлять к корням растущих растений или вносить в почвы для земледелия или садоводства для уменьшения повреждения растений, вызванного фитопатогенными грибами.
Настоящее изобретение включает также способы снижения чувствительности растений к грибковой инфекции. В одном варианте осуществления изобретения этот способ включает в себя доставку Streptomyces WYEC 108 к корням растения. В другом варианте осуществления изобретения этот способ включает в себя погружение семян в композицию, содержащую Streptomyces WYEC 108, а затем высаживание обработанных семян в подходящую ростовую среду. В этом способе подходящая композиция включает в себя альгинатный гель, содержащий Streptomyces WYEC 108.
Настоящее изобретение описывает выделение ряда штаммов актиномицетов из почвы. Ряд этих штаммов продемонстрировал свою эффективность в снижении эффектов грибковых патогенов на растения, включая латук, турецкий горох и перец. В частности, настоящее изобретение предлагает выделение штамма, называемого в настоящем документе Streptomyces WYEC 108. Показано, что штамм YEC 108 демонстрирует сильный антагонизм в отношении широкого ряда грибковых фитопатогенов, включая патогены, вызывающие выпревание проростков до и после прорастания, корневую гниль, бурую гниль и белую гниль. В качестве такого антагониста штамм WYEC 108 особенно удобен в качестве биоконтролирующего агента, который можно использовать для защиты растений от инфицирования этими фитопатогенами. Так, Streptomyces WYEC 108 является полезным для использования в способах снижения чувствительности растений к грибковой инфекции. Благодаря этому, растения, обработанные Streptomyces WYEC 108, менее подвержены воздействию грибковой инфекции. Грибковая инфекция чувствительных необработанных растений влияет на некоторые ростовые характеристики этих растений. Например, необработанные растения, зараженные грибковыми патогенами, могут демонстрировать значительное снижение высоты, биомассы и урожайности по сравнению с растениями, не зараженными грибковым патогеном. В предпочтительных вариантах осуществления настоящего изобретения растения, обработанные Streptomyces WYEC 108 и затем зараженные грибковым патогеном, демонстрируют менее выраженные снижения высоты, биомассы и урожайности, чем необработанные растения, зараженные грибковым патогеном. В более предпочтительных вариантах осуществления настоящего изобретения растения, обработанные Streptomyces WYEC 108 и зараженные грибковым патогеном, демонстрируют те же ростовые характеристики, что и необработанные и незараженные растения. В наиболее предпочтительных вариантах осуществления настоящего изобретения растения, обработанные Streptomyces WYEC 108 и зараженные грибковым патогеном, демонстрируют ростовые характеристики, превосходящие таковые необработанных незараженных растений.
Штамм WYEC 108 колонизирует корни растений в присутствии конкуренции микрофлоры ризосферы. Показано, что штамм WYEC 108 усиливает рост растений латука, выращиваемых на почве, стерилизованной паром, и растений перца, выращиваемых на открытом грунте.
Настоящее изобретение также заключает в себе средства производства вегетативных клеток или спор штамма WYEC 108, подходящих для помещения их в доставочную среду. Показано, что композиция, включающая вегетативные клетки и споры WYEC 108 и доставочную среду, имеет длительный срок хранения и удобна для доставки штамма WYEC 108 к растениям.
Материалы и методы.
Среды для роста бактерий.
Все среды для роста бактерий приготавливали, используя дистиллированную воду, и стерилизовали автоклавированием перед использованием. Все бактериальные образцы обрабатывались в соответствии со стандартными асептическими лабораторными методиками для поддержания чистоты.
Среда YGM (дрожжевой экстракт/глюкоза/минеральные соли) включала 0,6% (вес/объем) дрожжевого экстракта (лаборатории Difco, Детройт, Мичиган), 1,0% (вес/объем) глюкозы и раствор фосфатных неорганических солей (5,3 г Na2HPO4, 1,98 г KH2PO4, 0,2 г MgSO4 • 7H2O, 0,2 г NaCl, 0,05 г CaCl2 • 2H2O плюс 1,0 мл микроэлементов (Pridham и Gottlieb, 1948) на 1 л деионизированной воды: pH от 7,1 до 7,2). Раствор микроэлементов состоял из 0,64 г CuSO4 • 5H2O, 0,11 г FeSO4 • 7H2O, 0,79 г MnCl2 • 4H2O, 0,15 г ZnSO4 • 7H2O в 100 мл дистиллированной воды.
Среда WYE (вода/дрожжевой экстракт/агар), модифицированная Reddi и Rao (1971), содержала дрожжевой экстракт (Oxoid, 0,25 г/л) в качестве единственного источника углерода и азота и агар (Oxoid, 18,0 г/л). pH среды доводили до 7,2 - 7,4 буфером K2HPO4 (0,5 г/л). WYEC (вода/дрожжевой экстракт/целлюлоза/агар) представляла агар WYE, к которому добавили тонкий покрывающий слой агара. Покрывающий агар содержал 0,25 г/л целлюлозы (Solka Floc, химическая компания Sigma) и 18,0 г/л агара в дистиллированной воде.
Среда CYD (казаминокислоты/дрожжевой экстракт/декстрозный агар) содержала казаминокислоты (Difco 0,5 г/л), дрожжевой экстракт (Oxoid или Difco : 0,8 г/л), D-глюкозу (0,4 г/л), K2HPO4 (2,0 г/л; pH 7,2-7,4) и 18,0 г/л агара в дистиллированной воде.
Среда УCED (казаминокислоты/дрожжевой экстракт/декстроза/агар), модифицированная среда Reddi; и Rao (1971), содержала дрожжевой экстракт (Oxoid, 0,3 г/л), казаминокислоты (Difro 0,3 г/л), D - глюкозу (0,3 г/л) и агар (Oxoid, 18,0 г/л). pH среды доводили буфером K2HPO4 (2,0 г/л).
Среда CYPC (целлюлоза/дрожжевой экстракт/пептон/экстракт компоста/агар) содержала целлюлозу (Solka Flock, химическая компания Sigma, 5,0 г/л), дрожжевой экстракт (1,0 г/л), пептон (Oxoid, 1,0 г/л), фосфатный буфер (K2HPO4, 0,75 г/л), агар (18,0 г/л) и экстракт компоста (100 мл/л), замещающий 100 мл дистиллированной воды в среде. Эта серия наливается непосредственно и не используется в качестве покрывающего агара.
Среда MSSC (неорганические соли/крахмал/казеин/агар; Turhan, 1981) содержала раствор неорганических солей, состоящий из NaCl (2,0 г/л), MgSO4 • 17H2O (0,05 г/л), CaCO3 (0,02 г/л), FeSO4 • 18H2O (0,01 г/л) и KNO3 (2,0 г/л) плюс органические составляющие, включающие растворимый крахмал (10,0 г/л) и казеин (0,3 г/л) плюс агар (18,0 г/л). К среде прибавляли буфер K2HPO4 (2,0 г/л).
Среда для споруляции (АТСС Medium # 5) содержала дрожжевой экстракт (1,0 г/л), мясной экстракт (1,0 г/л), триптозу (2,0 г/л), FeSO4 (0,01 г/л), глюкозу (10,0 г/л) и агар (15,0 г/л). Перед автоклавированием pH среды доводили до 7,2 (17-е издание каталога ATCC бактерий и бактериофагов).
CYG - среда содержала казаминокислоты (кислотный гидролизат, 5,0 г/л), дрожжевой экстракт (5,0 г/л) и глюкозу (10,0 г/л) в дистиллированной воде, pH 7,1-7,2.
Доставочная среда (включающая песок/кукурузную муку/воду или торфяной мох/песок/кукурузную муку в соотношениях, приведенных ниже) стерилизовалась паром перед использованием. Стерилизацию выполняли обычно путем трехкратного автоклавирования, каждый раз по 90 мин.
Сбор бактериальной культуры.
Для мицелиального роста Streptomyces WYEC 108 в 1-литровые колбы Эрлейнмейера, содержащие 500 мл среды YGM (pH 7,1-7,2) инокулировали 20 мл культуры штамма, приготовленной как описано в примере II, и инкубировали при встряхивании на 250 об/мин при 30oC в течение трех дней. Затем собирали мицелий центрифугированием на 5000 об/мин. в течение 10 мин. Иначе мицелий собирали, оставляя культуры стоять до оседания на дно колб Эрленмейера и спор. Надосадок затем декантировали, а концентрированную суспензию мицелия и спор использовали непосредственно для инокуляции в доставочную среду.
Клетки и споры получали также путем роста на твердой среде (например, на агаре для споруляции). Мицелий и споры собирали с агара, соскребая поверхность агара в дистиллированную воду. Эту суспензию спор и мицелия затем непосредственно смешивали с доставочной средой.
Для получения спор Streptomyces WYEC 108 в 2-литровые колбы Эрленмейера, содержащие 1200 мл среды YGM, инокулировали 50 мл культуры штамма и инкубировали при встряхивании на 250 об/мин. при 30oC в течение 12-18 дней. Споры собирали центрифугированием на 9000 об/мин. в течение 10 мин.
Грибковые патогены.
Pythium ultimum pUMXL получали из коллекции культур в департаменте микробиологии и защиты растений в Horticulture Research International, Worthing Road, Литтлгемптон, Западный Суссекс BN17 6ZP, Великобритания). Грибы белой гнили Phanerochaete chysosporium и Coriolus versicolor;
грибы бурой гнили Postia placenta, Caldariomyces fumago и Gloeophyllum trabeum;
почвенные грибковые патогены Rhijoctonia solani, Fusarium sambucinctum, Geotrichum candidum и Verticillum dahlliae получили из коллекции культур профессора Don Z. Crawford факультет бактериологии университета Айдахо, Москва, Айдахо. Pythium irregulare, Phytophthora capsici, Phytophthora cinnamomi, Phytophthora parasitica, Selerotinia cepivorum и Sclerotinia selerotiorum получили из коллекции культур д-ра Wersley chun, Department of Plant soil Entomology science, университет Айдахо, Москва, Айдахо, Fusarium oxysporum получили из коллекции культур д-ра Arthur D.Partridge, Department of Forest Resources, университет Айдахо, Москва, Айдахо. Все культуры выращивали на картофельном декстрозном агаре или кукурузном агаре при 25oC. Эти штаммы при получении были идентифицированы как "патогены", но повторно на патогенность не проверялись.
Исследование почвы.
Для использования в биологическом исследовании собирали почву, естественно инфицированную Pythium ultimum из нескольких мест района Palouse неподалеку от Москвы, Айдахо. Эту почву собирали из 15-сантиметрового слоя на полях, засеянных и горохом в предыдущие два сезона. Почвенную популяцию видов Pythium определяли следующим образом: взвесь из 1,0 г высушенной воздухом почвы в 50 мл стерилизованной дистиллированной воды тщательно перемешивали с помощью миксера Vortex, 0,1 мл образца тщательно перемешанной извести помещали в виде маленьких капель на трехдневный 2%-ный водный агар на чашках Петри (Stanghellini и Hancock, 1970). Чашки инкубировали при 25oC и просматривали периодически под препаровальной лупой (х 10) с флюоресцентной подсветкой для определения идентичности и количества присутствующих видов Pythium. Колонии на каждой чашке проверяли спустя 12, 48 и 72 ч после инкубации до установления окончательной популяции. Идентификация основывалась на морфологических характеристиках мицелия гриба Pythium под микроскопом и характере роста на 2%-ном (вес/объем) водном агаре. Колонии гриба чистой культуры, выращенные на 2%-ном (вес/объем) агаре, служили контролем для целей визуальной идентификации (Stanghellini и Hancock, 1970; Stasz и др., 1980).
Исследование показало, что популяционная плотность P. ultimum и P. irregulare составляла 354 ± 15 и 194 ± 11 кое/г высушенной воздухом почвы во время посевной (весна 1992) соответственно. Популяционная плотность других видов Pythium составляла 57 ± 9 кое/г высушенной воздухом почвы. P. ultimum и P. irregulare были превалирующими видами в собранной почве.
Пример 1
Выделение штаммов актиномицетов, демонстрирующих антагонизм в отношении грибковых фитопатогенов.
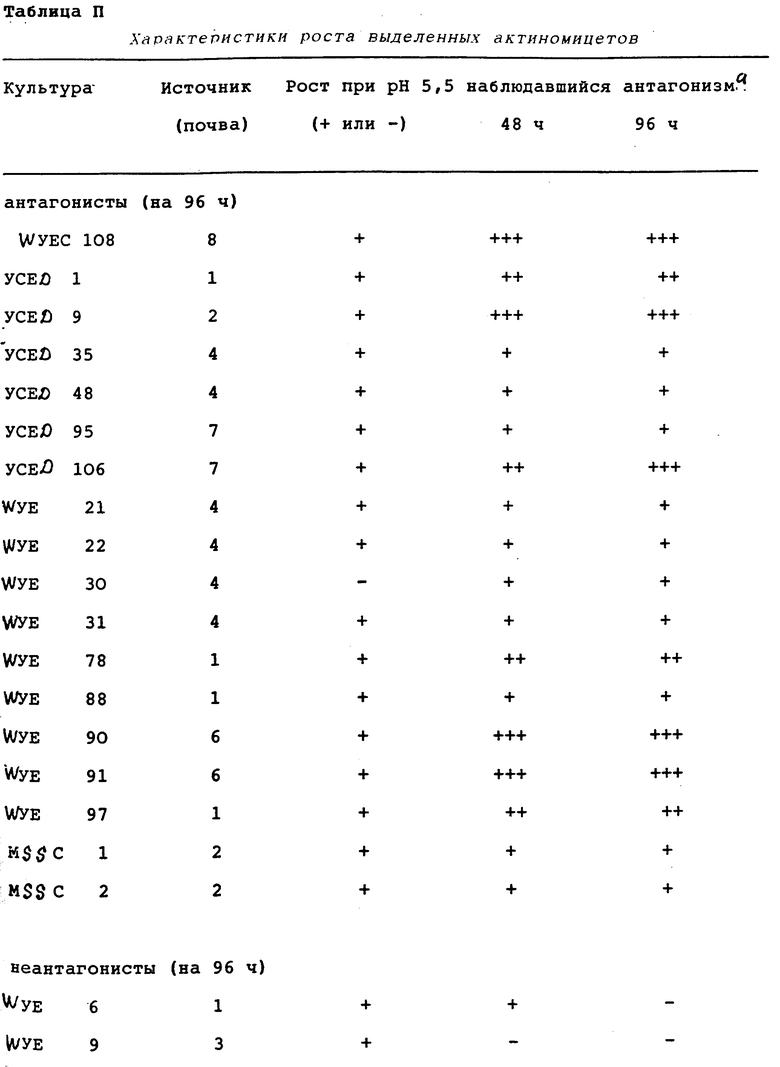
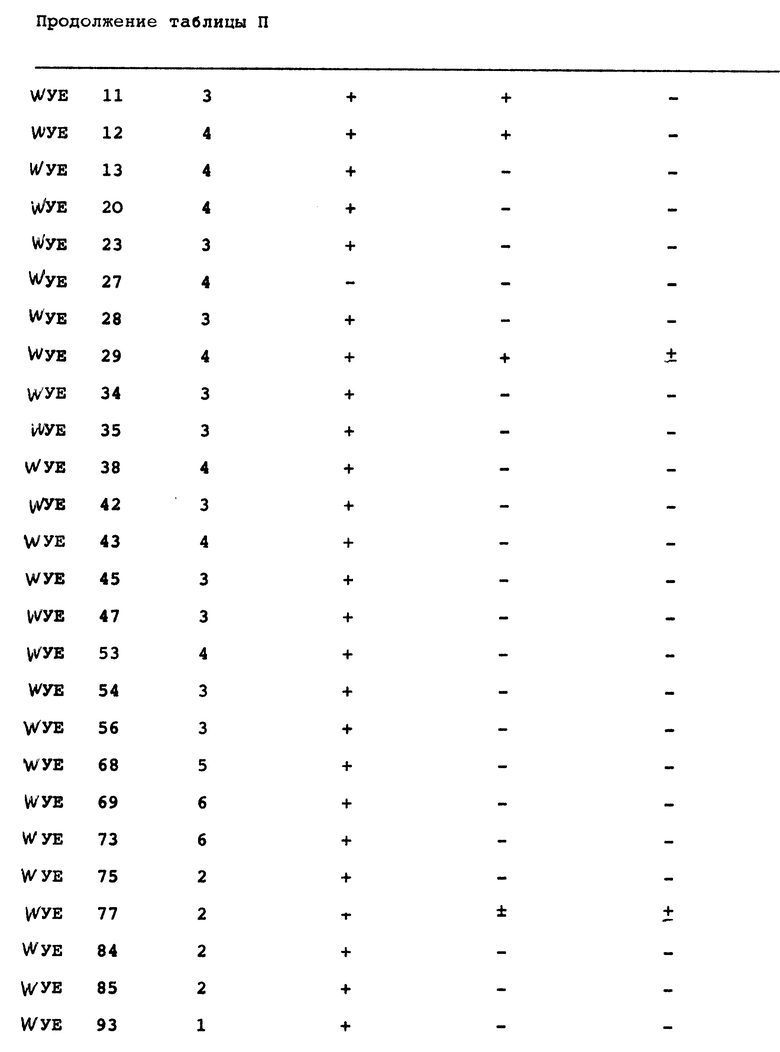
Актиномицетные штаммы выделяли из четырех связанных с ризосферой и четырех несвязанных с ризосферой почвенных образцов. Эти штаммы затем тестировали на пригодность в качестве ингибиторов грибковых фитопатогенов.
Выделение актиномицетов.
Актиномицеты выделяли из 8 различных почв с помощью методики серийного разведения и посева на чашки. Разведения (10-5 - 10-7) засевали на различные селективные агаровые среды. Состав этих сред приводился в разделе "Материалы и методы" выше. Изоляты актиномицетов обозначали согласно среде, на которой они были выделены. Например, WYEC 108 был выделен на среде WYEC, а YCED 9 был выделен на среде YCED. В целом эти среды бедны органическим углеродом, который эффективно контролирует эубактериальный и грибковый рост и помогает выделить медленно растущие актиномицеты. Поскольку WYE и YCED агары были особенно эффективными изоляционными средами, их использовали прежде всего. Разведения на агаре инкубировали при 25oC от 4 до 10 дней, чтобы позволить актиномицетам образовать споры, а затем колонии собирали и помещали на чашки с агаром WYE или YCED для очистки. Чистые колонии переносили с этих чашек на агар YCED или агар CYD (скошенные), инкубировали при 25oC или 37oC до достижения спорообразования и хранили при 5oC вплоть до использования. Маточные культуры пересевали каждые 3 - 4 недели.
Почвы.
(1) Не связанные с ризосферой почвы
Образцы (100 - 200 г) почвы отбирали из поверхностного 7,5-10 см слоя почвы в 4 районах Великобритании, включая культивированный розовый сад (почва S1) в Растингтоне, Западный Суссекс; в междурядье на пшеничном поле (почва S2) на ферме Horticullure Research International (H.R.1.) в Литтлгемптоне, Западный Суссекс; лесную почву (почва S6) из заповедника деревьев с твердой древесиной в Винд Клифф, Южный Уэльс и почву лугопастбищного угодья (почва S7), на которой периодически выпасают овец в Гастингс Хилл, Саут Даунс, Западный Суссекс. Эти почвы считали не связанными с ризосферой, хотя они и содержали корни растений в разных количествах.
(2) Связанные с ризосферой почвы.
Образцы связанной с ризосферой почвы из 4 мест были приготовлены в основном согласно способу Miller и др. (1990). Почва 3 (S3) была связана с корнями растения одуванчика (Taracum officinale) в розовом саду HR1 в Западном Суссексе, Англия. Почва 5 (S5) была связана с с корнями пшеницы и была взята с того же поля, что и S2, пшеничного поля на ферме HR1 в Литттлгемптоне, Западный Суссекс. Почва 4 (S4) была также связана с корнями пшеницы, но почва была взята с поля в Bignor Hill неподалеку от South Downs Way, Западный Суссекс. Почва 8 (S8) была связана с корнями растений льна и была взята с поля, прилегающего к участку, на котором был взят образец S7, на лугопастбищном угодье в Гастингс Хилл, Саут Даунс, Западный Суссекс.
Актиномицеты, выделенные из почв, могут быть разделены на изолированные из почв, связанных с ризосферой, и из почв, не связанных с ризосферой (S3, S4, S5 и S8) и (S1, S2, S6 и S7) соответственно. Во всех образцах оценивали влажность почвы путем высушивания 3 г (сырой вес) образцов (три повторности) при 100oC в течение 58 часов и затем повторного взвешивания. pH почв определяли путем тщательного перемешивания взвеси почвы в воде (1:1), оседания твердых частиц в течение 2 ч, и определения pH надосадка. После отбора почвы хранили при 4oC вплоть до использования (от 24 до 48 ч). То, что изоляты являлись штаммами актиномицетов, подтверждалось при визуальном исследовании, показавшем, что колонии, образованные этими штаммами, являются типичными колониями актиномицетов (твердые и кожистые, с воздушным мицелием, содержавшим споры). Дальнейшее подтверждение идентичности этих штаммов актиномицетам было получено при микроскопии.
Определение пределов pH для роста.
Каждый изолят актиномицетов испытывали на способность расти при pH от 5,5 до 8,0. Культуры засевали на чашки с агаром CYD, pH доводили до 5,5, 6,0, 6,5, 7,0 и 8,0 с помощью комбинаций буфера K2HPO4 и KH2PO4 с концентрацией 100 мМ. Конечное значение pH в каждой среде доводили до ее конечного значения непосредственно перед автоклавированием. Рост культур проверяли спустя 5 - 7 дней инкубации при 25oC или 37oC. Чашки оценивали визуально: малый рост или отсутствие роста (±), небольшой рост (+ или ++) или отличный рост (+++).
Как видно из таблицы 1, почвы, связанные с ризосферой, дали почти в два раза больше изолятов, чем почвы, не связанные с ризосферой. Каждый изолят испытывали на рост на агаре CYD при pH от 5,5 до 8,0. Все изоляты росли в пределах pH от 6,5 до 8,0. Только 9 изолятов не росли при pH 6,0, в то время как 57 (21%) не росли при pH 5,5. Среди изолятов, росших при pH 5,5, рост варьировал от слабого до отличного, в зависимости от изолята. Способность изолятов образовывать споры на агаре CYD определяли также визуально и под микроскопом спустя 5 - 10 дней инкубации.
Исследование антагонизма in vitro.
Отобрали 82 изолята по их способности хорошо расти и образовывать много спор на агаре CYD.
Для испытания способности изолятов ингибировать рост P. ultimum использовали исследование на чашках in vitro. Каждый актиномицет сеяли штрихом на кукурузный агар (КА), на одной половине. Культуру инкубировали при 25oC в течение приблизительно 8 дней или до образования спор. Агаровый блок из КА (0,5 см2), содержавший активно растущий мицелий P. ultimum переносили в асептических условиях в центр чашки. Инкубацию продолжали еще 96 ч. Спустя 48 и 96 ч чашки изучали на предмет ингибирования роста P. ultimum. На ингибирование указывало то, что рост мицелия P. ultimum по направлению к колонии актиномицетов задерживался или предотвращался. Результаты этого исследования представлены в таблице II.
Спустя 96 ч пять изолятов (WYEC 108, YCED 9, WYE 91, WYE 90 и YCED 106) демонстрировали очень сильный антагонизм по отношению к P. ultimum, четыре (YCED 1, YCED 106, WYE 97 и WYE 98) демонстрировали сильный антагонизм, а десять других демонстрировали слабый антагонизм. Остальные изоляты демонстрировали слабый антагонизм. Остальные изоляты демонстрировали или очень слабый антагонизм, или вовсе его не демонстрировали. Культуры, которые четко ингибировали рост P. ultimum, примерно пополам распределялись между связанными и не связанными с ризосферой почвами.
Шестьдесят изолятов, которые росли при pH 5,5, также испытывали на антагонизм in vitro по отношению к грибу белой гнили Phanerochaete chrysosporium на кукурузном агаре (КА). Тринадцать изолятов демонстрировали определенную степень антагонизма по отношению к грибу белой гнили, как показано в таблице III. Степень антагонизма варьировала от очень высокой (+++) до относительно низкой (+), что определяли при размеру зоны ингибирования. Пять из этих культур, которые демонстрировали антагонизм по отношению к P. Chrysosporium (WYEC 108, WYE 78, WYE 90, YCED 9 и MSSC 2) испытывали далее на КА против еще одного гриба белой гнили (Coriolus versicolor) и против двух типов грибов бурой гнили (Postia placenta и Gloephyllum trabeum). Четыре изолята (MSSC 2, YCED 9, WYE 90, WYEC 108) демонстрировали очень сильный антагонизм по отношению к вышеупомянутым грибам белой и бурой гнили. Один изолят, WYEC 78, демонстрировал сильный антагонизм только по отношению к двум грибам белой гнили.
Для исследования 12 изолятов в отношении их воздействия на всхожесть семян и рост латука (Latuca Sativa) применяли методику Zynch и др. (1991, 1992).
Для наблюдения за растениями пластиковые горшки диаметром 9 см наполняли смесью для выращивания растений латука и утрамбовывали чашкой Петри. Десять семян латука помещали на поверхность, слегка вдавливали в смесь и слегка покрывали лишней смесью. Затем горшки помещали на лотки в воду и инкубировали в темноте при 20 - 22oC, пока прорастание не становилось очевидным (приблизительно 3 дня). Лотки затем переносили в теплицу на капиллярные циновки и содержали при 15 - 25oC и поливали по потребности. Количество проросших семян в каждом горшке периодически подсчитывали вплоть до 18-го дня.
Для растений, обработанных актиномицетами, горшки наполняли смесью, инокулированной спорами определенного актиномицета. Смесь инокулировали спорами из штаммов с косячков CYD в количестве в среднем 108 - 109 кое/г (сухой вес) смеси для выращивания. Кое/г в смеси для выращивания определяли подсчетом жизнеспособных колоний на чашках с агаром CYD на время инокуляции. Затем семена латука высаживали, как описано выше, покрывали небольшим количеством инокулированной смеси для выращивания, а затем обрабатывали так же, как контроль.
Для растений, обработанных актиномицетами и грибом выпревания Pythium ultimum (штамм PUMXZ, Lynch и др., 1991), смесь также инокулировали грибковым патогеном в количестве приблизительно 200 спорангиев на 1 г (сухой вес) смеси. Для получения спорангиев патогена и инокуляции смеси использовали методику Lynch и др. (1991, 1992). Подсчет спорангиев производили на гемоцитометре, а патогенность штамма P. ultimum подтверждали перед применением путем пассажа на латуке.
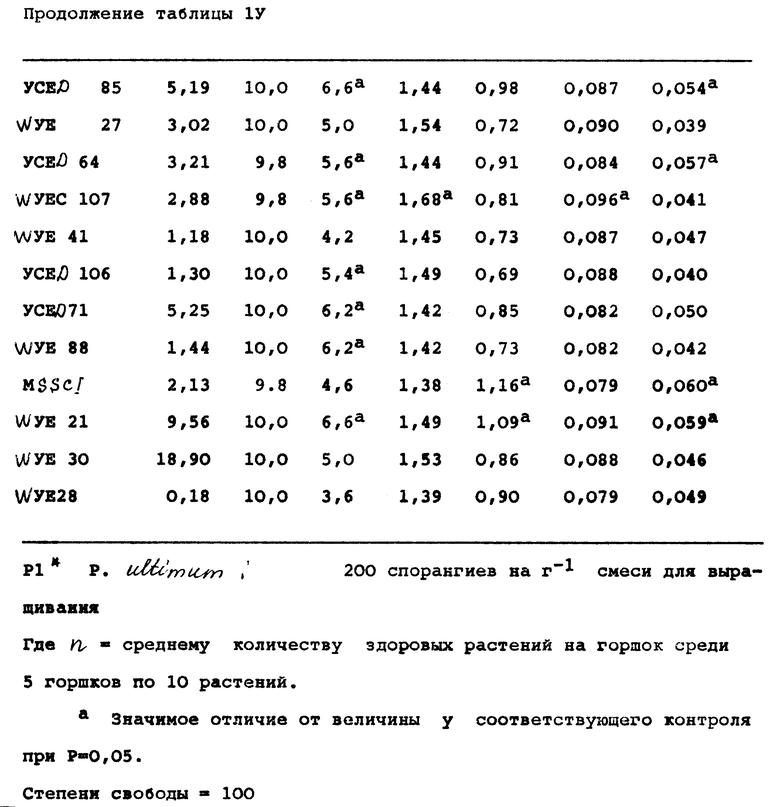
Для всех видов обработки брали по пять горшков. В теплице горшки помещали произвольными блоками, окруженными "сторожевыми растениями"; "сторожевые растения" служили для обеспечения одинаковых условий и действия в качестве буфера. Восемнадцать дней спустя после посадки растения собирали, определяли их количество, а также сухой и сырой вес (надземные листья и стебли). Сырой и сухой вес растений фиксировали как общую биомассу горшка в мг и как среднюю биомассу растений в мг. Величины представляли как средние пяти повторностей ± стандартное отклонение. Так, каждая величина основывалась на 50 семенах, высаженных для дальнейшей обработки (пять горшков по десять семян в каждом). Процент всхожести семян и количество выросших растений оценивали сходным образом (таблица IV).
В отсутствие патогенов (обозначены как -P1 в таблице IV) не было значимой разницы между инокулированными актиномицетами и контрольными горшками (без актиномицетов) в проценте семян, которые взошли и дали здоровые растения. Всхожесть семян и их прорастание усредняли в ≥ 98% всех случаев. Однако присутствие актиномицетов иногда задерживало прорастание семян на 1 - 3 дня (данные не показаны). Подобно этому в отсутствие патогенов обычно не было значимой разницы между инокулированными актиномицетами и контрольными горшками по весу побегов растений, измеренному как сырой и сухой вес. Исключение составляли горшки с WYEC 107, в которых присутствие актиномицетов значимо повышало выход биомассы растений.
В присутствии патогена (+P1) количество здоровых растений на горшок спустя 20 дней после посева в среднем составляло 3,6 из 10,0 в контрольных горшках по сравнению с 9,8 из 10,0 в контрольных горшках без патогена. В присутствии специфических актиномицетов, однако, наблюдались некоторые значимые улучшения в количестве здоровых растений и инокулированных патогенами горшках. Семь из 12 актиномицетов (YCED 85, YCED 64, WYEC 107, YCED 106, YCED 71, WYE 88 и WYE 21) значимо улучшали выход здоровых растений. Из них YCED 85, YCED 64 и WYE 21 также значимо улучшали показатели выхода сухого веса растений по сравнению с контролями, содержавшими только патоген. WYE 21 также значимо улучшал выходы сырого веса растений по сравнению с контролями, содержавшими только патоген. Один штамм, который не улучшал значимо выход здоровых растений (MSSC 1), улучшал показатели сырого и сухого веса побегов растений (таблица IV, колонки 5 и 7). Таким образом, MSSC 1 может оказаться полезным для защиты растений от более низких доз P. ultimum, чем те, которые использовались в настоящем эксперименте.
Пример II.
Выделение Srteptomyces WYEC 108.
Штамм WYEC 108 был идентифицирован как вид Srteptomyces на основании морфологических характеристик рода Srteptomyces, которые определены в руководстве по систематической бактериологии Bergey (1986). WYEC 108 является нитевидной бактерией, которая образует цепочки спор на воздушном мицелии. Как описано выше, Srteptomyces WYEC 108 был выделен в числе других штаммов актиномицетов, выделенных из почвы, взятой из восьми различных мест в Великобритании. Srteptomyces WYEC 108 был выделен среди других актиномицетов путем серийного разведения и посева ризосферной почвы, связанной с корнями растений льна на поле в Гастингс Хилс, Саут Даунс, Западный Суссекс, Англия. Разведения (10-5 - 10-7) этой почвы высевали на селективный агар WYE. Чашки с разведениями инкубировали при 25oC в течение 4 - 10 дней, чтобы дать возможность колониям актиномицетов вырасти и образовать споры. Затем колонии собирали и сеяли штрихом на агар WYEC для очистки. Чистые колонии WYEC 108 переносили с этих чашек на скошенный агар CYD, инкубировали при 25oC до образования спор и хранили при 4oC вплоть до использования. Исходные культуры пересевали каждые 3 - 4 недели.
Идентификация Srteptomyces WYEC 108.
Как описано выше, выделенные штаммы актиномицетов изучали на предмет способности хорошо расти и образовывать много спор на агаре CYD. После этого ряд изолятов испытывали на способность ингибирования рост in vivo фитопатогена Pythium ultimum. Эти изоляты испытывали также на предмет антагонизма in vitro в отношении грибов белой гнили Phanerochaete chrysosporium и Coryolus versicolor, а также грибов бурой гнили Postia placenta и Gloeophyllum trabeum. В результате этих испытаний был выбран один их этих штаммов, в настоящем документе называемый Streptomyces WYEC 108, благодаря его благоприятным характеристикам.
Колонии Streptomyces WYEC 108, выращенные на чашках с агаром казиминокислота /дрожжевой экстракт/декстроза (CYD) исследовали под сканирующим электронным микроскопом. Образцы приготавливали следующим образом:
(1) колонии Streptomyces WYEC 108 на чашках с CYD покрывали 1,5%-ным раствором глутарового альдегида в 0,2 M буфере натрия какодилата и фиксировали в течение по меньшей мере двух часов;
(2) колонии полностью отмывали (2 x) 0,2 M буфером какодилата натрия путем аспирации предыдущей жидкости пипеткой и замены ее раствором буфера. Следили за тем, чтобы образец не высох;
(3) колонии затем снимали путем забора "пробок" агара, содержащих колонии;
(4) "пробки" помещали в отдельные контейнеры с отверстиями и подвергали дегидратации 100%-ным этанолом (2x) (J.T.Baker Inc., Phillipsberg, NJ);
(5) образцы затем высушивали в сушильной "бомбе" критической точки высушивали и помещали на отдельные штыри для образцов, используя окраску коллоидным серебром. Затем их покрывали 60/40 золотом/палладием и просматривали на сканирующем электронном микроскопе.
Определяли также различные физиологические характеристики штамма WYEC 108.
Штамм WYEC 108 не вырабатывает меланин или H2S на пептонном агаре с дрожжевым экстрактом и железом и пептонном агаре с железом (лаборатории Difco, Детройт, Мичиган) соответственно. Споровая масса, образуемая Streptomyces WYEC 108 на CYD была серого цвета. Этот штамм не рос при 45oC. Streptomyces WYEC 108 может принадлежать к виду Streptomyces ludicus, как определено в руководстве по определительной бактериологии Bergey (1986). Соответственно этот организм может быть отнесен к Streptomyces WYEC 108. Для краткости в настоящем документе этот организм упоминается как Streptomyces WYEC 108 или просто WYEC 108.
Присвоение номера АТСС.
Депозит Streptomyces WYEC 108 был осуществлен на условиях Будапештского договора с Американской коллекцией типированных культур (АТСС), Rockvill, MD, 29 июня 1993. Этот штамм зарегистрирован под номером АТСС 55445.
Приготовление исходных культур Streptomyces WYEC 108.
Для быстрого использования Streptomyces WYEC 108 инкубировали на агаре CYD или агаре для спорообразования (косячках) при 25oC до спорообразования и хранили при 4oC вплоть до использования. Для долговременного хранения культур 10 мл споровых суспензий приготавливали путем суспендирования спор с одного скошенного агара или чашки в 10 мл среды YGM. Эту споровую суспензию затем использовали для инокуляции в 250-мл колбы Эрленмейера, содержащие 100 мл среды YGM (дрожжевой экстракт/глюкоза/неорганические соли). Колбы затем инкубировали при встряхивании на 250 об/мин в течение 32 - 36 ч при 30oC для получения стандартного инокулюма.
Образцы из выращенного на YGM стандартного инокулюма также использовали для приготовления глицериновых культур, удобных для долговременного хранения при -70oC и для лиофилизации.
Пример III
Streptomyces WYEC 108 по отношению к грибковым патогенам in vitro
Способность Streptomyces WYEC 108 ингибировать рост ряда выбранных грибковых фитопатогенов измеряли степенью ингибирования роста колоний Streptomyces WYEC 108 сеяли штрихом на одну половину в центре чашек с кукурузным агаром (лаборатории Difco, Детройт, Мичиган). Инокулированные чашки инкубировали при 25oC в течение приблизительно 8 - 12 дней до образования культурами спор. Споруляцию определяли невооруженным глазом как массу серого воздушного мицелия и спор. Споруляцию определяли также под фазово-контрастным микроскопом (x 1000), 5-мм агаровый диск КА, содержавший активно растущий мицелий специфического грибкового фитопатогена, брали с ведущего края грибковой культуры и асептически переносили в центр чашки с агаром. Эти чашки инкубировали при 25oC до тех пор, пока тест-гриб не достигал края контрольной чашки, не содержавшего Streptomyces WYEC 108. Ингибирование грибкого роста оценивали количественно путем определения отношения радиального роста грибкового патогена под влиянием Streptomyces WYEC 108 к отдельному его росту на контрольных чашках. Процент ингибирования грибкового роста регистрировали спустя 48, 96 и 192 ч инкубации, в зависимости от выбранного патогенного гриба. Это исследование повторяли на пяти чашках, ингибирование измеряли по отдельности и выражали как среднее ± стандартное отклонение.
Результаты этих исследований in vitro приведены в таблице V. Эти данные свидетельствуют о том, что Streptomyces WYEC 108 демонстрировал очень сильный антагонизм в отношении широкого спектра грибковых патогенов растений, включая грибы-возбудители выпревания (Pythium Ultimum), корневой гнили (Pythium ultimum, Rhigoetonia solani, Fusarium solani и Phytophthora cinnamomi), белой гнили (Phanerochaete chrysosporium и Coriolus versicolor) бурой гнили (Postia placenta и Gloephillum trabeum) и листовой и стеблевой гнили (Sclerotinia sp).
Пример IV
Использование Streptomyces WYEC 108 для обработки семян
Эффективность Streptomyces WYEC 108 в отношении защиты растений от фитопатогенов определяли обработкой штаммов Streptomyces WYEC 108 непроросших семян турецкого гороха и последующим высевом этих семян на почву, зараженную грибковыми фитопатогенами P.ultimum и P.irregulare.
Внеклеточные метаболиты, вырабатываемые штаммом WYEC 108, получали из культур путем экстракции. Эффект этих метаболитов на грибковую инфекцию прорастающих семян турецкого гороха также определялся.
Рост Streptomyces WYEC 108.
Для роста клеток штамма WYEC 108 1-литровые колбы Эрленмейера, содержавшие 500 мл YGM (pH 7,1-7,2) инокулировали 20 мл исходной культуры и инкубировали при встряхивании на 250 об/мин при 30oC в течение 3 дней для получения клеточной массы. Для получения противогрибковых метаболитов 1-литровые колбы Эрленмейера, содержавшие 500 мл CYD (pH 7,1-7,2), инокулировали 20 мл исходной культуры и инкубировали при встряхивании на 250 об/мин при 30oC в течение 7 дней.
Обработка семян Streptomyces WYEC 108 и противогрибковыми метаболитами.
Мицелиальную суспензию Streptomyces WYEC 108 собирали центрифугировали на 5000 об/мин в течение 10 мин 500 мл трехдневной жидкой YGM культуры. Собранный мицелий ресуспендировали в 200 - 300 мл стерильного 3%-ного (вес/объем) раствора альгинатта натрия до плотности культуры 1,0-1,2•104 кое/мл. Затем к хорошо перемешанной клеточно-альгинатной суспензии добавляли семена турецкого гороха, которые затем переносили по одному в стерильный 0,25 М раствор CaCl2 в дистиллированной воде. Эти семена использовали для эксперимента по биологическому контролю, описанного выше.
Противогрибковые метаболиты, вырабатываемые Steptomyces WYEC 108, получали в чистом виде следующим образом. Семидневные культуры (500 мл) фильтровали для удаления клеток и последовательно экстрагировали 150 мл эфира, используя экстракционную воронку. Затем эфир удаляли путем выпаривания в вакууме, а полученные экстракты вновь растворяли в 1,5 мл дистиллированной воды. Этот раствор затем стерилизовали фильтрованием через стерильный 0,45 мкм фильтр и добавляли в 10 мл 3% (вес/объем) раствора альгината натрия. Очищенные противогрибковые метаболиты можно использовать для защиты растений от грибковой инфекции. Предпочтительно противогрибковые метаболиты нужно очищать так, чтобы они практически не содержали клеток WYEC 108. Однако можно ожидать, что препараты клеток WYEC 108 и/или спор в сочетании с противогрибковыми метаболитами будут также эффективным средством против грибковых фитопатогенов. Суспензию противогрибковый метаболит-альгинат применяли, как описано выше, для обработки семян турецкого гороха в эксперименте по биологическому контролю.
Исследование по биологическому контролю in vivo.
Почва, естественно зараженная P. ultimum и P. irregulare, описана выше в разделе "Материалы и методы". Эта почва из сельскохозяйственных угодий использовалась в эксперименте по биологическому контролю in vivo. pH почвы определяли путем тщательного перемешивания взвеси почва:вода (1:1), оседания твердых частиц в течение 2 ч и определения pH надосадка, который был равен 5,6. Почву крошили, тщательно перемешивали и наполняли ею горшки для рассады (10 см глубиной и 10 см в диаметре).
Исследование по биологическому контролю in vivo выполняли путем посева непроросших семян турецкого гороха, обработанного Streptomyces WYEC 108 или противогрибковыми метаболитами, в зараженную почву. Необработанные семена, посеянные в ту же почву, служили контролем. Эта процедура включала следующие этапы:
1) 1 см торфяного мха укладывали на дно каждого горшка для предотвращения потери почвы и одновременного обеспечения аэрации и дренажа.
2) Горшки наполняли зараженной почвой.
3) Почву затем поливали водой до полного насыщения со стороны дна.
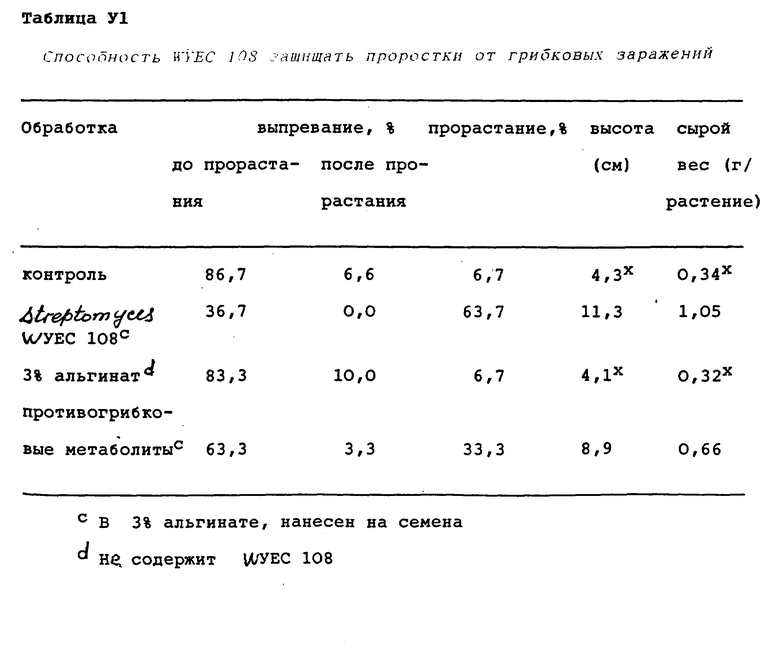
После насыщения поверхности почвы на нее помещали необработанные и обработанные семена турецкого гороха, углубляли на 1,5-2,0 см и покрывали той же почвой. Покрытие становилось влажным в результате капиллярности почвы, поднимающей воду из столба влажной земли под покрытием. В каждый из трех горшков (повторности) засевали по десять семян. К почве не прибавляли удобрения. Для минимизации высыхания и предотвращения образования корки горшки покрывали чистой пластиковой пленкой до появления проростков. Начиная со времени появления проростков, на поверхность горшков распыляли добавочную воду, когда это требовалось. Эксперименты выполняли в теплице при 15 - 30oC с фотопериодом в 12 ч света и 12 ч темноты (16000 лк). Появление проростков подсчитывали периодически, а конечные их подсчеты производились по истечении 20 дней. Эти данные были представлены как средние величины для каждой обработки. Способность Streptomyces WYEC 108 действовать как биоконтролирующий агент основывалась на общем количестве проростков, высоте растений и сыром весе растений по сравнению с контрольными растениями, выросшими из необработанных биоконтролирующим агентом семян. Результаты этого эксперимента представлены в таблице VI.
Как Streptomyces WYEC 108, так и противогрибковые метаболиты, вырабатываемые этими клетками, снижали выпревание турецкого гороха, вызванное Pythium.
Растения демонстрировали мощный рост, если семена покрывались клетками Streptomyces WYEC 108. Наблюдалось значимое снижение высоты и сырого веса растений, появившихся из контрольных семян, по сравнению с теми же показателями растений, выросших из семян, покрытых клетками Streptomyces WYEC 108. Прорастание необработанных семян турецкого гороха было снижено в крайней степени (прорастание 6,7%) из-за гнили семян и выпревания до прорастания, вызванного P. ultimum, когда семена были высажены в почву, естественно зараженную P. ultimum и P. irregulare. В противоположность этому прорастание семян, обработанных клетками Streptomyces WYEC 108 перед посевом, составляло 63,3%. Семена, обработанные только альгинатом, не показали увеличения прорастания. Симптомы, типичные для корневой гнили, вызванной Pythium, включая потерю корневых волосков и обесцвечивание корней, были очевидными в собранных корнях турецкого гороха, выросших из контрольных семян; в то же время эти симптомы отсутствовали у растений, выросших из семян, обработанных клетками Streptomyces WYEC 108. В контроле повреждение турецкого гороха наблюдалось в основном в форме гниения семян и выпревания до прорастания. Проростки турецкого гороха, которые все же взошли и выросли, были низкорослыми, а их корни были сильно заражены P. ultimum.
Прорастание семян турецкого гороха, обработанных противогрибковыми метаболитами в форме растворенного в эфире метаболита, была выше (33,3%), чем прорастание контрольных семян (6,7%), но ниже, чем прорастание семян, покрытых клетками Streptomyces WYEC 108. Растения, выросшие из семян, обработанных противогрибковыми метаболитами, демонстрировали мощный рост, более длинные корни и более высокую плотность корневых волосков по сравнению с контрольными растениями.
Пример V.
Влияние Streptomyces WYEC 108 на корневую инфекцию и гниль семян, вызванные Pythium ultimum.
Проростки турецкого гороха, выросшие, как описано в примере IV, на почве, естественно зараженной Pythium ultimum, с предварительной обработкой WYEC 108 или без нее, изучались с целью определения влияния штамма WYEC 108 на P. ultimum инфекцию.
Корневую инфекцию P. ultimum изучали с контрольными проростками турецкого гороха, собранными на 20-й день роста. P. ultimum, вызывающий корневую гниль, также был выделен из пораженных корней этих контрольных растений. Выделение P. ultimum было выполнено путем отмывания почвы от корней турецкого гороха (корешки и корневые волоски) водопроводной водой и ополаскивания их дважды стерильной дистиллированной водой. Обесцвеченные и тронутые гнилью корни измельчали в асептических условиях с помощью лезвия безопасной бритвы и помещали их на чашки с 3-дневным 2%-ным агаром. Чашки инкубировали 24-48 ч при комнатной температуре и просматривали под фазово-контрастным микроскопом (х 40). P. ultimum, растущий из зараженных корней турецкого гороха, рекультивировали на свежем 2%-ном агаре, а затем идентифицировали, как описано ранее (Ingram и Cook, 1990).
Гниль семян, вызванную P. ultimum, изучали на тронутых гнилью контрольных семенах, собранных на 20-й день после посева. Часть гнилых семян асептически помещали на 3-дневный 2%-ный агар стерильной зубочисткой и инкубировали 24-48 ч при комнатной температуре. Чашки изучали, как описано выше.
Pythium заражал корни необработанных растений турецкого гороха, росших на естественно зараженной почве. P. ultimum был превалирующим видом из тех, что выделяли из сгнивших семян и корней. P. irregulare встречался реже.
Колонизацию корней Streptomyces WYEC 108 изучали на корнях 20-дневных растений турецкого гороха, которые выросли из семян, обработанных суспензией клеток Streptomyces WYEC 108 и альгината, описанной в примере II. Эти растения росли на почве, естественно зараженной P. ultimum и P. irregulare, также описанной в примере II. растения удаляли из горшков и осторожно отмывали водопроводной водой для удаления прилипшей ризосферной почвой. Затем их прополаскивали стерильной дистиллированной водой. Образцы корней подготавливали для микроскопии путем помещения кусочков корней на предметные стекла, добавления капли метиленового синего и покрывания покровным стеклом. Приготовленные образцы затем изучали под фазово-контрастным микроскопом (х 1000).
Streptomyces WYEC 108, присутствующий на семенах, приходит в контакт с появляющимися корнями. По мере удлинения корней Streptomyces WYEC 108 продвигается вместе с ними на корневые волоски и верхушку корня. Наблюдалась обширная колонизация Streptomyces WYEC 108 главного корня, вторичных корней, корневых волосков и верхушки. Растения, которые появлялись из семян, покрытых Streptomyces WYEC 108, были более здоровыми, имели более длинные корни и большую плотность корневых волосков по сравнению с контрольными растениями, которые появлялись из семян, не покрытых WYEC 108. Эта разница была отчетливо связана с колонизацией корней биоконтролирующим агентом. Корни, колонизированные биоконтролирующим агентом, не имели каких-либо симптомов поражения корней. Streptomyces WYEC 108 показали отличную колонизацию корней в присутствии конкуренции со стороны природной микрофлоры ризосферы.
Противогрибковая активность.
В добавление к способности колонизировать корни растений и вырабатывать противогрибковые метаболиты наблюдалась также способность Streptomyces WYEC 108 лизировать клеточную стенку и ооспоры грибов. С помощью сканирующего электронного микроскопа было показано, что гифы Streptomyces WYEC 108 колонизировали поверхность гиф и ооспор гриба, включая гифы и ооспоры Pythium ultimum. Колонизированные гифы и ооспоры разрушились Streptomyces WYEC 108, наиболее вероятно, в результате экстрекции WYEC 108, экстрацеллюлярных ферментов, таких как хитиназы и целлюлазы. Показано, что Streptomyces WYEC 108 вырабатывает обе группы этих ферментов. Вероятно, Streptomyces WYES 108 вырабатывает также ряд других экстрацеллюлярных разрушающих ферментов.
Пример VI.
Включение Streptomyces WYEC 108 в доставочную среду.
Композиция, удобная для длительного хранения жизнеспособных спор Streptomyces WYEC 108 и использования в сельскохозяйственной практике, была составлена следующим образом. 1-литровые колбы Эрленмейера, содержащие 500 мл среды YGM (pH 7,0-7,1) инокулировали 20 мл исходной культуры и инкубировали при встряхивании на 250 об/мин при 30oC в течение трех дней. После инкубации культуру собирали центрифугировали на 5000 об/мин. в течение 10 мин. Собранный материал ресуспендировали в 1600 мл 10% YGM и смешивали со стерильным раствором 8 г NH4Cl в 400 мл дистиллированной воды. Два литра смеси клеток и NH4Cl затем инкубировали в пластиковый контейнер, содержащий 4 кг стерильной доставочной среды, состоявшей из смеси песка, воды и кукурузной муки в соотношении 9:2:1 (вес/вес). Доставочную среду дважды стерилизовали (3 ч при 121oC) перед инокуляцией культуры. Эту смесь затем инкубировали в течение 10 - 14 дней при 25oC, чтобы довести до максимума содержание спор в смеси. Streptomyces WYEC 108 вырабатывали споры в течение 10 - 14 дней инкубации, в результате чего повышалось кое/г доставочной среды (до среднего уровня 108 - 109 кое/г доставочной среды (сухой вес)). Эту смесь затем хранили при 4oC вплоть до использования.
Клетки и споры можно получать и на среде CYG вместо среды YGM. Как альтернативный способ собиранию клеток центрифугированием культуральные колбы (флаконы) можно также оставлять стоять до оседания бактериального мицелия и спор. Затем прозрачный надосадок сливают, а концентрированную суспензию мицелия и спор инокулируют непосредственно в доставочную среду. Когда применяется эта процедура собирания, нет необходимости добавлять к среде NH4Cl, поскольку бактериальная ростовая среда (YGM или CYG) является подходящим источником азота.
Пример VII.
Влияние Streptomyces WYEC 108 на прорастание и сырой вес продуктов лука/
Для определения влияния Streptomyces WYEC 108 на рост латука семена выращивали как на доставочной среде, содержащей Streptomyces WYEC 108, как описано в примере VI выше, так и на почве, простерилизованной паром. На каждую ростовую среду высевали по 30 семян (одно семя на 4 х 13,5- см горшок) и прорастание регистрировали спустя 21 день роста. Сырой вес определяли путем сбора надземных частей растений спустя 35 дней роста. Сырой вес выражали средними величинами. Результаты представлены в таблице VII.
Данные табл. VII свидетельствуют о том, что, обработка семян латука Streptomyces WYEC 108 увеличивала как всхожесть семян, так и сырой вес проростков.
Пример VIII.
Применение Streptomyces WYEC 108 в доставочной среде на поле.
Исследования по биологическому контролю in vivo выполнялись для определения эффективности Streptomyces WYEC 108 как биоконтролирующего агента при инкорпорации в доставочную среду, описанную выше. Streptomyces WYEC 108 был получен и инкорпорирован в доставочную среду, как описано выше в примере VII, при использовании Streptomyces WYEC 108, выращенного в течение трех дней на среде YGM. Исходная популяция Streptomyces WYEC 108 в доставочной среде была определена приблизительно в 1,0-1,2 • 105 кое/г почвы путем подсчета на чашках с агаром CYD непосредственно перед высаживанием ростков в горшки. Эта обработанная почва использовалась затем для наполнения горшков (4 х 13,5 см).
Контрольные растения выращивали в горшках с почвой, простерилизованной паром (100oC в течение 60 мин).
Ростки перца (зеленого, жгучего) затем высаживали в горшки, содержавшие как почву, простерилизованную паром, так и почву, состоящую из смеси почвы, простерилизованной паром, и доставочной среды, содержавшей Streotomyces WYEC 108. Спустя шесть недель роста в теплице эти растения пересаживали на сельскохозяйственное поле. В некоторых случаях 100 г доставочной среды, содержавшей 450 ± 17 кое/г Phytophthora parasitica, были инокулированы в посадочные лунки перед пересаживанием растений.
Высоту растений определяли спустя 55 дней после пересаживания и выражали как среднюю величину. Биомассу растений определили путем сбора и определения сырого веса растений спустя 110 дней культивации после пересаживания. К этому времени на растениях образовывались плоды перца, и вес перцев на каждом растении определяли как среднюю величину. Результаты полевых испытаний представлены в таблице VIII.
Как видно из таблицы VII, обработка проростков перца WYEC 108 в отсутствие P. parasitica обеспечивала статистически значимое увеличение высоты растений, биомассы растений, количества перцев и урожая перцев по сравнению с контрольными растениями, не получившими WYEC 108. Сравнение горизонтальных рядов 3 и 4 в таблице VIII показывает, что штамм WYEC 108 защищал растения перца от вредного воздействия P.parasitica. Помимо этого наблюдалось значимое усиление роста растений, обработанных штаммом WYEC 108 в отсутствие P. parasitica по сравнению с необработанными растениями без P. parasitica.
Помимо экспериментов, описанных в этом примере, было показано, что WYEC 108 был эффективен для защиты ряда растений от грибковой инфекции. Растения, участвовавшие в испытаниях, включают приведенные в таблице IX.
Испытания сравнивали растения, обработанные WYEC 108, с необработанными контрольными растениями и/или с растениями, обработанными WYEC 108 и специфическим грибковым патогеном (Pythium, Aphanomyces, Rhiroctonia, Fusarium, Phytopthora или Phytomatotrichum).
В каждом из этих случаев WYEC 108 защищал растения от грибковых заболеваний.
Пример IX
Образование спор Streptomyces WYEC 108 в жидкой среде.
Биоконтролирующие агенты должны сохранять жизнеспособность в течение длительных периодов времени, чтобы соответствовать нуждам перевозки и временному характеру сельскохозяйственных работ. Применение спор штамма WYEC 108 в большей степени, чем вегетативных клеток в композициях для биоконтроля, увеличивает срок хранения биоконтролирующей композиции, поскольку споры сохраняют жизнеспособность при неблагоприятных условиях и в течение длительных периодов времени.
Обычно споры видов Streptomyces образуются только на твердых средах. Однако, как объясняется ниже, был обнаружен следующий способ получения этих спор в жидкой культуре.
В двухлитровые колбы Эрленмейера, содержавшие по 200 мл среды YGM (pH 6,5), инокулировали 50 мл исходной культуры (полученной, как описано в примере II) и инкубировали при встряхивании на 250 об/мин при 30oC в течение 12-18 дней. За образованием спор следили путем просмотра культур под фазово-контрастным микроскопом (x 1000, окрашивание метиленовым синим). Споры собирали центрифугированием на 9000 об/мин в течение 10 мин.
После этого споры ресуспендировали в 600 мл стерильной 10%-ной жидкой среды YGM и добавляли 400 мл стерильного раствора, содержавшего 8 г NH4Cl в дистиллированной воде (для получения конечной плотности спор 1,0 - 1,2 • 107 кое/мл). Эту среду затем непосредственно инокулировали в 4 кг стерильной доставочной среды, состоящей из песка, воды и кукурузной муки в соотношении 9 : 2 : 1. Доставочную среду стерилизовали дважды в автоклаве (3 ч при 121oC) до инокуляции спор.
Получение спор непосредственно в жидкой культуре указанным способом избавляет от необходимости дальнейшей инкубации смеси. Доставочную среду, содержавшую споры, затем хранили при 4oC вплоть до использования.
Споры, полученные описанным способом в жидкой культуре, испытывали на жизнеспособность спустя четыре месяца хранения при 4oC. 1 мл суспензии спор инокулировали во флаконы, содержавшие 100 мл стерильной 10%-ной жидкой среды YGM (pH 6,5) и инкубировали при встряхивании на 250 об/мин при 30oС. Прорастание спор наблюдали с помощью фазово-контрастного микроскопа (х 1000, окрашивание метиленовым синим). Споры полностью прорастали в течение приблизительно 8 дней. Это простое испытание показало отсутствие потери жизнеспособности после указанного периода хранения. Streptomyces WYEC 108, инкорпорированный в доставочную среду (песок, вода, кукурузная мука; 9 : 2 : 1), испытывали на жизнеспособность следующим способом. 1 г образца доставочной среды, содержавшей Streptomyces WYEC 108, серийно разводили и высевали на чашки с агаром CYD). Чашки инкубировали при 25oC до образования колоний. В образцах, которые хранили 30 дней, зафиксировали средний уровень в 108 - 109 кое/г доставочной среды (сухой вес).
Иначе споры с чашки с агаром для спорообразования ресуспендировали в 10 - 20 мл стерильной дистиллированной воды или бульона YGM и примешивали к 10 - 100 г доставочной среды, для получения 1012 - 1014 кое/г. Эту смесь затем высушивали на воздухе, тщательно перемешивали и хранили при 4oC вплоть до использования. Эта рецептура является концентрированным продуктом, который можно разводить дополнительным количеством доставочной среды до любого желаемого более низкого содержания кое/г.
Пример X
Стабильность композиции с альгинатным гелем.
Мицелий Streptomyces WYEC 108 собирали центрифугированием на 5000 об/мин в течение 10 мин из 500 мл 3-дневной жидкой культуры YGM. Собранный мицелий ресуспендировали в 125 мл 10% YGM и добавляли 125 мл стерильного 5%-ного раствора альгината натрия до плотности культуры 1,0 - 1,2 • 104 кое/мл. Получали альгинатные гранулы, содержавшие мицелий Streptomyces WYEC 108, путем добавления клеточно-альгинатной суспензии по каплям в стерильный раствор 0,25 М CaCl2 в дистиллированной воде.
Для определения жизнеспособности альгинатных гранул, полученных этим способом, альгинатные гранулы, содержавшие культуру, равномерно распределяли на стерильной пластиковой чашке Петри (10 • 10 см) и высушивали в течение одного часа в вытяжном шкафу с ламинарным потоком стерильного воздуха. Гранулированный Streptomyces WYEC 108 хорошо образовывал споры после хранения в течение 6 - 8 месяцев при 25oC (до среднего уровня 108 - 109 кое/г высушенных альгинатных гранул). Эти споры хорошо прорастали, будучи помещенными в стерильную воду при 25oC. Прорастание спор наблюдали с помощью фазово-контрастного микроскопа (x 1000, окрашивание метиленовым синим).
Пример XI
Предпочтительные композиции доставочной среды, включающей Streptomyces WYEC 108.
После изложения способов выделения штаммов актиномицетов, тестирования этих штаммов на их пригодность в качестве биоконтролирующих агентов, способов получения этих биоконтролирующих агентов в форме мицелия и спор, подходящей доставочной среды и в предпочтительном варианте Streptomyces WYEC 108 специалисту будет ясно, что настоящее изобретение можно модифицировать целым рядом способов, не отступая от идеи настоящего изобретения.
Ниже приводятся примеры альтернативных вариантов осуществления настоящего изобретения вместе с описанием особенно предпочтительных вариантов осуществления.
Оптимальные условия культивирования
Оптимальные условия для роста штамма WYEC 108 включают температуры между 20 и 30oC, значения pH между 5,5 и 7,5 и скорости перемешивания в ферментере между 200 и 300 об/мин. Streptomyces WYEC 108 обычно достигает максимальных выходов клеточной массы около 5,3 г сухого веса биомассы на литр в жидкой среде YGM при условиях культивирования 30oC, pH 6,5 и встряхивании на 250 об/мин, в течение 72 ч (до окончания log фазы). Время удвоения во время логарифмической фазы роста составляет приблизительно 10 ч. 72-часовое время инкубации может быть значительно сокращено при использовании более высоких уровней инокуляции клеток в log фазе.
Иначе споры можно получать на твердой агаровой среде, такой как агар для спорообразования. Эти споры можно собирать непосредственным соскабливанием в подходящую жидкую среду, такую как 10% YGM, и затем непосредственно вводить в доставочную среду.
Этот подход избавляет от необходимости выращивания культуры в жидкой среде и, таким образом, сокращает процесс получения продукта.
Предпочтительная и альтернативная доставочные среды
Streptomyces WYEC 108 можно инкорпорировать в доставочную среду для использования в сельском хозяйстве и садоводстве. Пример VI описывает одну рецептуру доставочной среды, которая включает песок/воду/кукурузную муку в соотношении 9 : 2 : 1 (в/в). Специалист должен понять, что рецептура доставочной среды может диктоваться конкретным вариантом применения, для которого предназначен биоконтролирующий агент. Например, к доставочной среде можно прибавлять различные органические и неорганические наполнители, такие как глина, вермикулит, пшеничные отруби, кукурузные кочерыжки или хитин. Соотношение компонентов в доставочной среде определяется на основании требуемых текстуры и физических свойств. Например, могут быть важными такие свойства, как способность удерживать влагу, легкий вес для удобства применения и транспортировки, полезность для обеспечения пространства для роста и распространения мицелия и корней растений. Альтернативно можно добавлять вегетативный мицелий или споры Streptomyces WYEC 108 к альгинатной суспензии для получения гранул этого микроорганизма, заключенного в альгинат. Способы получения альгинатных гранул известны специалистам и описываются в патенте США N 4668512 и др. В эти гранулы также можно инкорпорировать другие ингредиенты, такие как удобрения.
В предпочтительном варианте осуществления изобретения, авторы определили, что особенно удобна доставочная среда, включающая торфяной мох/песок/кукурузную муку в соотношении 1 : 3,5 : 1 (вес). Это соотношение создает подходящую плотность и влагоудерживающую способность для использования этого продукта в сельском хозяйстве и садоводстве. Однако, как было отмечено выше, другие соотношения этих компонентов и других компонентов также являются приемлемыми для доставочной среды. Например, эффективная альтернативная доставочная среда включает торфяной мох (620 г)/песок (3380 г)/кукурузную муку (270 г)/хитин (10 г).
В одном варианте осуществления изобретения приблизительно 1,6 л собранного культурального бульона (клетки в log-фазе, например приблизительно 72 ч культура), содержавшего мицелий Streptomyces WYEC 108, выращенный на бульоне YGM, как описано выше, смешивали с 400 мл стерильного раствора NH4Cl (содержащего 8 г NH4Cl в 400 мл дистиллированной воды) и инокулировали в пластиковые контейнеры, содержавшие 4 кг стерильной доставочной среды, состоявшей из торфяного мха, песка и кукурузной муки. Доставочную среду дважды стерилизовали (3 ч при 121oC) перед инокуляцией Streptomyces WYEC 108. Инокулированные контейнеры инкубировали при 30oC в течение 10 - 14 дней для максимального спорообразования. Контейнеры затем хранили при 4oC вплоть до использования.
Использование NH4Cl в доставочной среде создает источник азота для прорастающих спор Streptomyces WYEC 108. Специалисту будет ясно, что для этой цели можно использовать и другие источники азота помимо NH4Cl. Например, и как описано в настоящем документе, когда споры ресуспендируют в ростовой среде для бактерий (такой как 10% YGM) перед инкорпорацией в доставочную среду, добавление этого источника азота уже не является необходимым. В предпочтительных вариантах осуществления настоящего изобретения доставочная среда включает доставочное количество источника азота. Специалисту будет ясно, что "достаточное количество" источника азота можно определить по эффекту на прорастание повышенных или пониженных количеств определенного источника азота или по эффекту смены источника азота. Достаточное количество источника азота представляет такое количество определенного источника азота, которое облегчает прорастание спор Streptomyces WYEC 108.
В альтернативном варианте осуществления изобретения, как описано в примере XI, споры Streptomyces WYEC 108 получали в жидкой среде и инкорпорируют непосредственно в предпочтительную доставочную среду, которую затем хранят при 4oC.
В предпочтительном варианте осуществления настоящего изобретения Streptomyces WYEC 108 добавляют к доставочной среде до конечной концентрации по меньшей мере 1 • 105 кое/г. В более предпочтительных вариантах осуществления настоящего изобретения конечная концентрация Streptomyces WYEC 108 в доставочной среде находится в пределах между 1 • 105 кое/г и 1 • 108 кое/г.
Пример XII
Подробное описание рецептуры доставочной среды, включающей Streptomyces WYEC 108.
Процедура приготовления предпочтительной композиции доставочной среды, включающей Streptomyces WYEC 108, была в основном следующей. Все процедуры, описанные ниже, производились с помощью стандартной асептической методики (например, в камере с ламинарным потоком воздуха, простерилизованной УФ-лучами), чтобы гарантировать асептические условия вплоть до вскрытия упакованных пакетов потребителями.
Получение клеток.
1) Суспендировали споры со скошенного агара CYD со Streptomyces WYEC 108 в 10 мл стерильного бульона YGM или CYD (pH 6,5). Эту суспензию использовали для инокуляции в культуральные флаконы.
2) Инокулировали шесть 250-мл флаконов, содержавших 100 мл YGM (pH 6,5). Использовали 10 мл суспензии спор на флакон в качестве инокулюма. После инокуляции флаконы инкубировали при встряхивании на 200 об/мин и 30oC в течение приблизительно 36 ч.
3) Инокулировали шесть 2-литровых флаконов, содержавших 1,1 л бульона YGM (pH 6,5) мицелиальным инокулюмом, приготовленным на предыдущем этапе (100 мл инокулюма на флакон). После инокуляции флакона инкубировали при встряхивании на 200 об/мин и 30oC в течение приблизительно 24 - 48 ч или больше (до 4 дней). Эта становилось инокулюмом для ферментера.
Ферментация.
Приблизительно 7,2 л исходной культуры, приготовленной, как описано выше, инокулировали в ферментер, содержавший 40 л стерильного бульона YGM (pH 6,5) (= 15% инокулюма по объему; подход состоял в инокуляции настолько высокой плотности клеточной суспензии, насколько это оправдано практикой). В ферментере производилось перемешивание (200 об/мин) при 30oC в течение приблизительно 72 ч (почти до окончания log фазы).
Сбор культуры из ферментера.
1) Культуральный бульон из ферментера, содержавший клетки WYEC 108, (приблизительно после 72 ч инкубации) собирали в асептических условиях в стерильные 20-литровые пластиковые бутыли.
2) К собранному культуральному бульону, содержавшему клетки WYEC 108, добавляли стерильный раствор H4Cl (использовали 16 г NH4Cl, растворенных в 800 мл дистиллированой воды, на 3,2 л собранного культурального бульона; предварительно раствор стерилизовался автоклавированием). Получившийся 1,2 л объем содержавшей NH4Cl клеточной суспензии затем тщательно перемешивали путем встряхивания бутыли до ее инокуляции в предварительно приготовленную доставочную среду.
Приготовление доставочной среды.
1) Каждый компонент доставочной среды по отдельности тщательно отмеряли и добавляли в кювету большого размера, которую можно было подвергать стерилизации, или в другой подходящий контейнер. Комбинированную смесь определяли как доставочную среду. Последняя состояла из торфяного мха, песка и кукурузной муки (540 г : 2700 г : 540 г; 1:3,5:1 (вес./вес.)).
2) Доставочную среду тщательно перемешивали и покрывали прочной алюминиевой фольгой или хлопковой ватой, а затем стерилизовали дважды (90 мин за один раз при 121oC с 12-часовым интервалом).
3) Доставочную среду охлаждали до комнатной температуры после второй стерилизации и перед инокуляцией собранного культурального бульона, содержавшего штамм WYEC 108 и раствор NH4Cl (приготовленный, как описано выше). Инкорпорация Streptomyces WYEC 108 в доставочную среду для создания композиции из торфяного мха, песка, воды, кукурузной муки и NH4Cl.
1) Приблизительно 0,5 л собранного культурального бульона, содержавшего штамм WYEC 108 и раствор NH4Cl (приготовленные, как описано выше) аккуратно инокулировали в каждый из необходимого количества предварительно простерилизованных пластиковых контейнеров, содержавших 3,78 кг доставочной среды.
2) Инокулированные контейнеры затем инкубировали при 30oC в течение 10 - 14 дней (инкубация может быть оптимальной до 20 дней), после чего их можно было хранить при 4oC вплоть до использования (композиция стабильна в течение месяцев).
Упаковка и транспортировка.
1) Приготовленную композицию, содержащую Streptomyces WYEC 108, асептически переносили в стерильные трехслойные пластиковые пакеты, используя маленькую простерилизованную лопатку или эквивалентный инструмент, предпочтительно в камере с ламинарным потоком воздуха со стерилизацией УФ-лучами.
2) Упакованные пакеты затем завязывали и помещали в 1,5 фт3 ящики для транспортировки. Каждый ящик затем запечатывали прочной лентой.
Пример XII
Включение композиции, содержавшей Streptomyces WYEC 108 в рассадочные грядки.
Композицию, содержавшую биоконтролирующий агент Streptomyces WYEC 108 и доставочную среду, описанную в примере XII, смешивали с почвой для рассадочных грядок или смесь для рассадочных горшков до конечной концентрации Streptomyces ≥ 1,0 - 1,2 • 105 или более кое/г почвы. Процедура высаживания была следующей.
1) На дно каждого горшка (или грядки) помещали около 1,0 см торфяного мха для предотвращения потерь почвы (или рассадочной смеси) и одновременного обеспечения аэрации и дренажа.
2) Рассадочные горшки затем наполняли почвой (грядочной или смесью для рассады) приблизительно 3 см, не доходя до края горшков (или грядки). Горшки (грядки) затем поливали до насыщения.
3) Затем горшки (грядки) заполняли сверху приблизительно 1,5 см композиции, содержавшей Streptomyces WYEC 108 и доставочную среду.
Если это желательно, композицию можно также смешивать предварительно с почвой под рассаду или смесью для горшков для увеличения объема и доведения содержания кое/г. Однако для оптимальной эффективности содержание кое/г в конечной смеси должно поддерживаться на уровне по меньшей мере 105 кое/г.
4) Семена помещали на поверхность приготовленных рассадочных горшков или грядок, а затем покрывали дополнительно 1,5 см (приблизительно) почвы или смеси для горшков.
5) Затем добавляли небольшое количество воды для увлажнения почвы и семян.
6) Для минимизации высыхания и предотвращения образования корки горшки обычно закрывали чистым черным пластиком до появления проростков (это можно не делать, если увлажнение контролируется).
7) На поверхность горшков (или грядок) распыляли дополнительно воду, как это необходимо после появления проростков.
После приведения примеров осуществления настоящего изобретения и предпочтительных вариантов осуществления специалисту будет ясно, что могут быть произведены изменения и модификации без отступления от идеи настоящего изобретения и его более широких аспектов. Прилагаемые пункты формулы изобретения даны с тем, чтобы охватывать все подобные изменения и модификации, находящиеся в пределах объема настоящего изобретения и его главной идеи.
Список литературы
1. ATCC Catalogue of Bacteria and Bacteriophages, 17th Edition, 1989. Amterican Type Culture Collection, Rockville, MD.
2. Locci, R. 1989. "Streptomyces and Related Genera", in Bergey's Manual of Systematic Bacteriology, Williams and Wilkens, Baltimore, MD. 4:2451-2492.
3. Filnow, A. B. and J.L Lockwood. 1985. Evaluation of several actinomycetes and the fungus Hypochytrium catenoides as biocontrol agents of Phylophthora root rot of soybean. Plant Disease 69:1033-1036.
4. Ingram, D. M. and R. J. Cook. 1990. Pathogenicity of four Pythium species to wheat, barley, peas, and lentils. Plant Pathology 39:110-117.
5. Kraft, J. M. and D.W. Burke. 1971. Pythium ultimum as a pathogen of beans and peas in Washington. Plant Dis. Rep. 55:1056-1060.
6. Lynch, J. M. , R. D. Lumsden, P.T. Atkey, and M. A. Ousley. 1992. Prospects for control of Pythium damping-off of lettuce with Trichoderma, Gliocladium, and Enterobacier 15 spp. Biol Fertil. Soils 12:95-99.
7. Lynch, J. M. , K. L. Wilson, M. A. Ousley, and J. M. Whipps. 1991. Response of lettuce to Trichoderma treatment. Lett. Appl. Microbiol. 12: 59-61.
8. Miller, J. J. E. Liljeroth, G. Henken, and J. A. van Veen. 1990. Fluctuations in the fluorescent pseudomonad and actinomycete populations of rhizosphere and rhizoplane during the growth of spring wheat. Can. J. Microbiol. 36:254-258.
9. Pridham, T.G. and D. Gottlieb. 1948. The utilization of carbon compounds by some actinomycetales as an aid for species determination. J. Bacteriol. 56:107-114.
10. Reddi, G. S., and A. S. Rao. 1971. Antagonism of soil actinomycetes to some soil borne plant pathogenic fungi. Indian Phytopathol. 24: 649-657.
11. Stanghellini, M.E. and J.G. Hancock. 1970. A Quantitative Method for the Isolation of Pythium ultimum from Soil. Phytopathology. 60:551- 552.
12. Stasz, Т.Е., G.E. Harman and G.A. Marx. 1980. Time and site of infection of resistant and susceptible germinating pea seeds by Pythium ultimum. Phytopathology. 70:730-733.
13. Trapero-Casas, A. , W.J. Kaiser and D.M. Ingram. 1990. Control of Pythium seed rot and preemergence damping-off of chickpea in the U.S. pacific northwest and Spain. Plant Dis. 74:563-569.
14. Westerlund. F. V..Jr., R.N. Campbell and K.A. Kimble. 1974. Fungal root rots and wilt of chickpea in California. Phytopathology 64:432-436.













Изобретение относится к биотехнологии, в частности к новому штамму актиномицетов, способному ингибировать рост фитопатогенов и усиливать рост растений, к средствам защиты растений от грибковой инфекции и снижению чувствительности растений к грибковой инфекции. Штамм актиномицета Streptomyces lydicus WYEC 108 выделен из почвы, депонирован в Американской коллекции типовых культур (АТСС) под 55445. Ингибирует рост таких фитопатогенов, как Pythium ultimum, Phanerochaete chrysosporium, Coryolus versicolor, Rhizoctonia solani, Fusarium sambucinetum. На основе вышеуказанного штамма создана композиция, содержащая штамм и доставочную среду, включающую по крайней мере один из компонентов: альгинатный гель, торфяной мох, песок, кукурузную муку. Дополнительно может включаться и источник азота в виде хлорида аммония. Способ снижения чувствительности растения к грибковой инфекции предусматривает обработку корней или семян вышеуказанным штаммом или композицией, содержащей штамм. Данные изобретения позволяют защитить растения от фитопатогенных грибов и стимулировать рост растений. 5 с. и 6 з.п. ф-лы, 9 табл.
| US 4595589 A, 17.06.86 | |||
| Способ защиты растений от увядания, вызванного фитопатогенными грибами | 1983 |
|
SU1477231A3 |
| Штамм актиномицета SтRертомYсеS VIoLaceoNIGeR для получения нигроцида активного против фитопатогенной микрофлоры | 1989 |
|
SU1792614A1 |

