Настоящее изобретение относится к области биоорганической химии и, в частности, касается новых дипептидных и псевдодипептидных соединений, имеющих в своем составе имидазольную или индольную группу, способа получения этих и известных соединений подобного строения, а также их применения в медицине в качестве потенциальных лекарственных средств.
Объектом изобретения являются производные пептидов общей формулы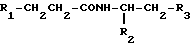 (I)
(I)
или их фармацевтически приемлемые соли, где R1 представляет собой COOH, NH2 или углеводородный радикал с C1-C3, замещенный функциональной группой, например амино, амидной с C1-C3, уретановой с C1-C7 или карбоксильной группой; или углеводородный радикал с C1-C3, одновременно замещенный амино- и карбоксильной группой, причем карбоксильная группа может быть этерифицирована, а аминогруппа может быть замещена ацильным заместителем или эфиром угольной кислоты; R2 - атом водорода или функциональная группа, например карбоксил, который может быть этерифицирован; R3 - индол или его метильные и/или гидроксильные производные, причем гидроксильная группа может быть ацилирована, алкилирована или аралкилирована; 5-6-членные насыщенные и ненасыщенные циклические и гетероциклические заместители, содержащие кислород, серу и/или 1-3 атома азота, или их метильные производные, причем при R1=NH2, NH2-CH2- R2=H, R3 не означает -4-Im, -3-(5-OMe-lnd), -3-(5-OH-Ind); при R1= NH2, NH2-CH2- R2=COOH, R3 не означает -4-Im; а при R1= HOOC-CH(NH2)- R2=H, R3 не означает -3-Ind, -3-(5-OH-Ind).
Предпочтительными являются производные пептидов общей формулы I,
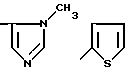
где R1=HOOC-(CH2)n-, n=0-2, C6H5CH2-OCO-NH-CH2-, NH2-CH2-, 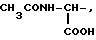
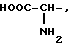
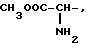
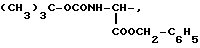 R2=H, -COOH, COOCH3
R2=H, -COOH, COOCH3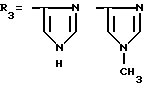
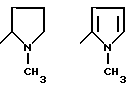
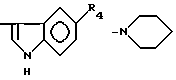
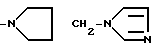
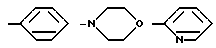
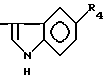
причем R4= -H, -OH, -OCH3, -OCH2C6H5.
Наиболее показательные примеры новых дипептидов и псевдодипептидов, соответствующих общей формуле (I), представлены ниже:
Сукцинилгистамин (II)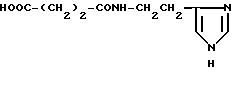
Глутарилгистамин (III)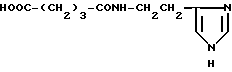
Адипинилгистамин (IV)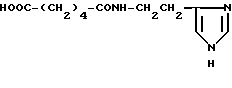
Nα-ацетил-L-γ- глутамилгистамин (V)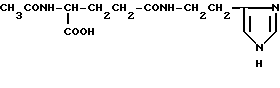
Метиловый эфир γ-L-глутамил-L-гистидина (VI)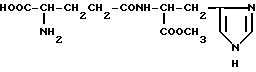
Метиловый эфир глутарил-L-гистидина (VII)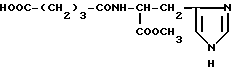
Метиловый эфир γ-аминобутирил-L-гистидина (VIII)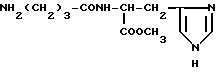
α-Метиловый эфир-γ-L-глутамилгистидина (IX)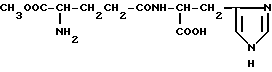
γ- Аминобутирилтриптамин (X)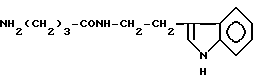
Глутарил-O-алкилсеротонин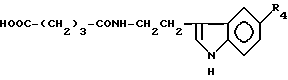
где R4= -OH (XI), -OCH3 (XII), -OCH2-C6H5 (XIII).
Глутарил-4-(2-аминоэтил)морфолин (XIV)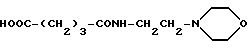
Глутарил-2-(2-аминоэтил)пиридин (XV)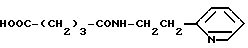
Глутарилфенилэтиламин (XVI)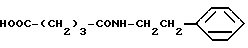
Известные псевдодипептиды, соответствующие общей формуле (I), где
при R1=NH2-, NH2-CH2-, NH2-CH(COOH)- R2=H, R3=-4-Im,
при R1=NH2-CH2- (COOH)- R2=H, R3=-3-Ind или -3(5-OH)Ind.
Примеры известных псевдодипептидов, соответствующих общей формуле (I), представлены ниже:
γ-D-глутамилгистамин (XVII)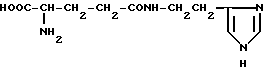
β-Аланилгистамин (XVIII)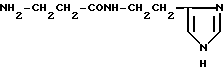
γ-Аминобутирилгистамин (XIX)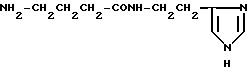
γ- L-глутамилтриптамин (XX)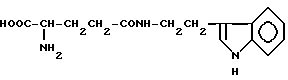
γ-L-глутамилсеротонин (XXI)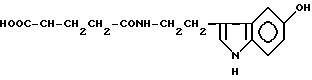
Примеры соединений общей формулы (I) приведены в табл. 1 (см. в конце описания).
Соединения общей формулы (I) получают путем ацилирования аминогруппы амина или аминокислоты активированным по карбоксильной группе производным дикарбоновой или N-защищенной аминокислоты.
Синтез дипептидов и псевдодипептидов, содержащих N-аминоацильный заместитель, осуществляют классическими методами пептидной химии с применением предпочтительно активированных, например, N- оксисукцинимидных эфиров. Наилучший вариант заключается в применении пентафторфениловых эфиров, как наиболее активных из известных. В качестве активированных производных дикарбоновых кислот применяют, как правило, их циклические внутренние ангидриды.
α-Аминогруппу карбоксильного компонента замещают различными обычно применяемыми группами, предпочтительно трет.- бутилоксикарбонильной (Boc-) или бензилоксикарбонильной (Z-) защитами.
α-Карбоксильную функцию глутаминовой кислоты защищают предпочтительно бензильной (Bzl-) группой.
Карбоксильная группа аминокомпонента - гистидина в соединениях общей формулы (I) находится в виде метилового эфира или остается незамещенной.
Синтез соединений, содержащих N-аминоацильный заместитель, представлен на следующей схеме 1:
Схема 1.
Стадия 1. R1-NH-A-COO-X+NH2-B--->R1-NH-A-CONH-B
Стадия 2. R1-NH-A-CONH-B--->NH2-A-CONH-B
Стадия 3.
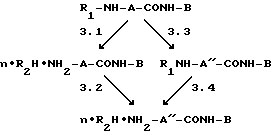
Стадия 4. n-R2H•NH2-A''-CONH-B--->NH2-A''-CO-NH-B,
где R1=Boc, Z; NH2-A-CO- = GABA-, H-L-Glu-OBzl; NH2-B=HA, TrpA, H-His-OH, H-His-OMe; NH2-A''-CO=H-L-Glu-, H-L-Glu-OMe, n•R2H=HHal(HCI); CF3COOH n= 1, 2.
Стадию 1 проводят, как правило, в среде безводного апротонного растворителя, предпочтительно диметилформамида (DMFA), 18-48 час при комнатной температуре, за исключением дипептида Boc-L-Glu(L-His)-OBzl. Последний получают действием 3-кратного избытка Boc-L-Glu(ONSu)-OBzl на незащищенный L-гистидин в водно-диоксановой среде (1:1). Преимуществом данного способа является упрощение процесса вследствие уменьшения числа стадий (отсутствия необходимости вводить и удалять C-защиту гистидина) и возможности получения дипептида, селективно защищенного по одной из 2-х карбоксильных групп.
Если это необходимо, проводят отщепление защитных групп промежуточного соединения R1-NH-A-CONH-B в соответствии со стадиями 2 и 3.
Стадия 2 проводится лишь в случае R1=Z, NH2-A-CO-= GABA- путем каталитического гидрогенолиза.
Когда это необходимо, осуществляется стадия 3 в 2-х различных модификациях при наличии в промежуточном соединении R1-A-CONH-B Nα-Boc- и Bzl-групп, а именно для производных глутаминовой кислоты. Способ 3.1 заключается в ацидолитическом отщеплении Nα-Boc-защиты, например, действием хлористого водорода в органическом растворителе, преимущественно диоксане, метаноле или их смеси; или трифторуксусной кислотой с последующим удалением Bzl-группы каталитическим гидрогенолизом (3.2). По способу 3.3 сначала проводится гидрогенолиз, а затем ацидолитическое отщепление Nα-защиты (3.4). В результате осуществления стадии 3 получают продукты в виде соответствующих солей.
При необходимости получения целевых соединений в виде свободных оснований проводят стадию 4.
В случае, если соединение не содержит незащищенных карбоксильных групп, оно может быть получено в виде свободного основания путем добавления органического (Et3N) или неорганического (NaOH) основания с последующим отделением соли этого основания от целевого продукта. Кроме того, для этой цели можно использовать ионообменную хроматографию в соответствии с методологией, описанной ранее [Евстигнеева Р.П., Желтухина Г.А., Огрель С.А., Небольсин В.Е. / Синтез псевдопептидов на основе биогенных аминов.// Докл. АН СССР. -1995. -т. 345. -N 4. -с. 493- 495].
Кроме того, когда это необходимо, соединение в форме основания может быть представлено в виде соли переходного металла с образованием хелата.
Известное соединение γ-L-Glu-HA [Koniski Н., Kakimoto Y./ Formation of γ-glutamylhistamine from histamine in rat brain.// J. Neurochem. -1976. -vol. 27. -pp. 1461-1463], являющееся исходным для получения нового производного (V), может быть получено описанным в литературе способом [Mc Caman М. W. , Stetzler J., Clark В. / Synthesis of γ-Glutamyldopamine and Other Peptidoamines in the Nervous System of Aplysia californica.// J. Neurochem. -1985. -vol. 45. -N 6. -pp. 1828-1835]. N- ацетильное производное γ-L-Glu-HA-Ac-γ-L-Glu-HA (V) может быть получено в соответствии с предложенной нами схемой 2.
Схема 2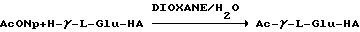
Схема 2 введения ацетильной группы в дипептид имеет преимущества по сравнению с использованием N-ацетильного производного глутаминовой кислоты в качестве исходного продукта для создания пептидной связи, т.к. уменьшает риск рацемизации. Применение n-нитрофенилацетата предпочтительно по сравнению с уксусным ангидридом, т.к. его использование не сопровождается побочными реакциями по имидазольной группе, которые заключаются в образовании ацетильного производного по имидазолу и соответствующей соли последнего с молекулой выделяющейся уксусной кислоты.
Синтез соединений общей формулы (I), содержащих остаток дикарбоновой кислоты, может быть осуществлен различными способами, предпочтительно в соответствии со схемой 3, где в качестве С-активированного карбоксильного компонента применяют его внутренний циклический ангидрид.
Схема 3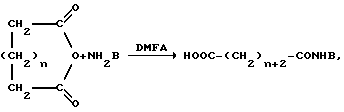
где n=0-1, NH2B=HA, H-His-OMe,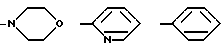
где R4=-H, -OH, -OCH3, -OCH2C6H5.
В случае, если подобный ангидрид не является доступным, например, для адипиновой кислоты, синтез псевдопептида может быть осуществлен DCC-методом. При этом сначала проводится реакция дикарбоновой кислоты с DCC при соотношении 2:1 моль/моль, а затем добавляют аминокомпонент (амин или производное аминокислоты).
Известные соединения, соответствующие общей формуле (I), а именно β-аланилгистамин (XIII) и γ-аминобутирилгистамин (XIV), были получены ранее методами классической пептидной химии [Евстигнеева Р.П., Желтухина Г.А., Огрель С.А., Небольсин В.Е. /Синтез псевдопептидов на основе биогенных аминов. // Докл. АН СССР. -1995. -т. 345. -N 4. -с. 493-495; Mc Caman M. W., Stetzler J. , Clark B. / Synthesis of γ- Glutamyldopamine and Other Peptidoamines in the Nervous System of Aplysia californica. // J. Neurochem. -1985. -vol. 45. -N 6. -pp. 1828-1835; Евстигнеева Р.П., Желтухина Г.А., Агеева Е.А., Бабижаев М.А. /Липопепоксидазная активность карнозина и карцинина. // Докл. АН СССР. -1993. -т. 333. -N 1. -с. 104-106], а также ферментативным способом - путем сочетания аминокислоты и гистамина в присутствии фермента типа гидролазы [Seguin М. C., Babizhayev M. / Product de couplage de l'un acide amine, procede de preparation, et applications therapeutigues et cosmetologigues. // Patent Fr 2701947. 1994. C1 C 07 C 237/04. A 61 K 31/195. 7/48].
Наиболее близкие по структуре к новым соединениям дипептидной природы эфиры N-ацильных производных γ- глутамилдипептидов [Floyd D. /N-Acyl-γ-glutamylimino and amino acids and esters.// Patent US 4568489. 1986. C1 C 07 C 103/52. C 07 D 217/00. C 07 D 548/470] были получены ранее DCC-методом, который, как известно, может приводить к побочным реакциям по имидазольной и индольной группе [Шредер Э., Любке К. Пептиды. - M.- Мир.- 1967.- с. 249]. В [Seguin М. C., Babizhayev M. /Product de couplage de l'un acide amine, procede de preparation, et applications therapeutigues et cosmetologigues.// Patent Fr 2701947. 1994. C1 C 07 C 237/04. A 61 K 31/195. 7/48; Floyd D. / N-Acyl-γ-glutamylimino and amino acids and esters.// Patent US 4568489. 1986. C1 C 07 C 103/52. C 07 D 217/00. C 07 D 548/470; Babizhayev M., Seguin М.C. /Pseudodipeptide product having on imidazole grouping and applications. // WO 95/12581. C1 C 07 D 233/64. C 12 P 17/10. A 61 K 31/415. 7/42] не приведены формулы предлагаемых нами гистидинсодержащих дипептидов, методики их синтеза и константы.
Ниже представлены примеры химического синтеза соединений общей формулы (I).
Производные глутаминовой кислоты, используемые в синтезах, L-ряда, D-производные указаны особо. Индивидуальность полученных соединений проверяют методом ТСХ на пластинках "Silufol" фирмы Kavalier, UV-254 (Чехословакия) в системах растворителей: хлороформ-метанол 9: 1 (1), хлороформ-метанол 8:2 (2), н-бутанол-уксусная кислота-вода 4:1:5 (3); на пластинках "Kieselgel" фирмы "Merck" в системе растворителей хлороформ- метанол-25% водный аммиак 5: 3: 1 (4); на пластинках "Silufol": хлороформ-метанол 9,3:0,7 (5); хлороформ-метанол-25% водный амииак 5:3:0,5 (6), хлороформ-метанол 8,5:1,5 (7); изопропанол-вода - 25% водный аммиак 6:3:1 (8).
Хроматограммы проявляют хлор-толидиновым раствором, реактивами Паули и Эрлиха, нингидрином и по свечению в УФ-свете.
Температуры плавления веществ определяют на приборе "Boetius" (Германия).
1H-ЯМР спектры снимают на приборе Brucker WM-250" (Германия) и "Varian XL-400" (Япония) с ТМС в качестве внутреннего стандарта.
Масс-спектрометрию осуществляют на приборе МСБХ (Украина, г. Сумы) методом плазменно-десорбционной ионизации осколками ядер калифорния 252.
Аналитическую обращенно-фазовую ВЭЖХ проводят в условиях (1): колонка МПС-270 C-18 (4,0 х 250 мм), 10 мкм, элюция 4% ацетонитрилом в воде, содержащей 0.1% TFA; в условиях (2): колонка та же, элюция градиентом от 10% до 50% фазы B в фазе A за 20 мин; фаза A - 0.1% TFA в воде, фаза B - 0.09% TFA в смеси ацетонитрила и воды 60:40; в условиях (3): колонка МПС 300 C18T (4,0 х 250 мм), 10 мкм, элюция градиентом от 0% до 40% фазы В за 20 мин, фаза A - 0.1% TFA в воде, фаза В - 0.09% TFA в смеси ацетонитрила и воды (60:40); в условиях (4): колонка МПС 270 C18 (4,0 х 250 мм) 10 мкм, элюция 0.1 M Na2HPO4, pH 2.3; в условиях (5): колонка та же, но элюция 0.1 M Na2HPO4, pH 2.7; в условиях (6): колонка Диасорб 130C18T (4.0 х 150 мм), 7 мкм, элюция градиентом от 0% до 42% ацетонитрила в 0.1% TFA; в условиях (7): колонка МПС 300 C18T (4.0 х 250 мм), 10 мкм, элюция градиентом от 0% до 18% ацетонитрила в 0.1% TFA за 20 мин; в условиях (8): колонка Lichrosorb RP-18 (4.6 х 250 мм), 5 мкм, элюция градиентом от 6% до 24% ацетонитрила в 0.1% TFA за 20 мин; в условиях (9): колонка Диасорб 130 C16T (4.0 х 150 мм), элюция 0,1% TFA; в условиях (10); колонка Диасорб 130 C16T (4.0 х 150 мм), 7 мкм, элюция градиентом от 0% до 24% ацетонитрила в 0.1% TFA; в условиях (11): колонка та же, элюция градиентом от 3% до 54% ацетонитрилом в 0.1% TFA за 30 мин; в условиях (12): колонка та же, элюция градиентом от 0% до 30% ацетонитрила в 0.1% TFA за 30 мин; в условиях (13): колонка МПС C18T (4.0 х 250 мм), элюция градиентом от 12% до 60% ацетонитрила в 0.1% TFA за 20 мин, в условиях (14): колонка МПС-270 (4.0 х 250 мм), 10 μ , элюция градиентом от 0% до 60% ацетонитрила в 0.1% TFA за 20 мин.
Во всех случаях ВЭЖХ проводилась при скорости элюции 1 мл/мин с детекцией при 214 нм.
Пример 1.
Сукцинилгистамин (II).
К раствору 0.080 г (0.72 ммоль) гистамина в 4 мл DMF при перемешивании добавляют 0.072 г (0.72 ммоль) янтарного ангидрида. Реакционную смесь перемешивают 1 ч и оставляют на 20 ч при 20oC. Растворитель удаляют в вакууме. Маслообразный остаток растворяют в 1.5 мл этанола, прибавляют 4 мл сухого эфира, растирают и выдерживают 30 мин при 4oC. Осадок отделяют и трижды перекристаллизовывают из метанола. Выход 0.075 г (49.2 %). Rf 0.41(4). Т.пл. 153-155oC. ВЭЖХ в условиях (6): один пик, время выхода 7.77 мин. 1H-ЯМР спектр (CD3OD), δм.д. : 2.42 (т, 2H, -CH2CONH), 2.55 (т, 2H, HOOC-CH2), 2.8 (т, 2H, β- CH2-HA), 3.4 (т, 2H, α- CH2-HA), 7.0 (с, 1H, CH-4-Im), 8,0 (с, 1H, CH-2-Im). Масс-спектр; m/z: [М+1H]+ 212.2.
Пример 2.
Глутарилгистамин (III).
К раствору 0.366 г (3.3 ммоль) гистамина в 5 мл DMF прибавляют 0.376 г (3.3 ммоль) глутарового ангидрида. Реакционную смесь перемешивают 3 ч и оставляют на 20 ч при 20oC. Белый осадок отделяют, сушат в вакууме, перекристаллизовывают. Выход 0.510 г (70,0%). Rf 0.36 (6), 0.34 (4). Т.пл. 187-189oC. ВЭЖХ в условиях (7): один пик, время выхода 14.36 мин. 1H-ЯМР спектр (D2O), δ, м. д. : (м, 2H, β- CH2), 2.18 (м, 4H, α,γ- CH2), 2.85 (т, 2H, β- CH2-HA), 3.5 (т, 2H, α- CH2-HA), 7.25 (с, 1H, CH-4-Im), 8.5 (с, 1H, CH-2-Im). Масс-спектр, m/z: [М+1H]+ 226.1.
Пример 3.
Хлоргидрат адипинилгистамина (IV).
К раствору 0.197 г (1.35 ммоль) адипиновой кислоты в 2.5 мл DMF при 0oC прибавляют 0.278 г (1.35 ммоль) DCC. Реакционную смесь перемешивают при 0oC 30 мин и прибавляют раствор 0.150 г (1.35 ммоль) гистамина в 1 мл DMF и оставляют при 20oC на 20 часов. Осадок DCU отделяют фильтрованием. К реакционной смеси прибавляют 10 мл сухого эфира и оставляют на 1 час при 0oC. Маслообразный остаток растворяют в этаноле и прибавляют 0.2 мл 4 н. HCl в диоксане. Растворитель удаляют в вакууме. Остаток промывают эфиром, растворяют в смеси хлороформ-этанол 1.5:1 и очищают на колонке с силикагелем 40/100 (22 х 175 мм). Элюируют смесью хлороформа и этанола от 7:3 до 2:8, этанолом, метанолом и смесью метанола-AcOH- H2O 8:1:0.5. Фракции, содержащие целевое вещество с Rf 0.25 (6), объединяют, растворитель удаляют в вакууме, сушат над P2O5. Выход 0.129 г (40%). Rf 0.25 (6). ВЭЖХ в условиях (11): один пик, время выхода 3.6 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 1.6 (м, 4H, β,γ- CH2), 2.3 (м, 4H, α,δ- CH2), 3.0 (т, 2H, β- CH2-HA), 3.55 (т, 2H, α- CH2-HA), 7.25 (с, 1H, CH-4-Im), 8.65 (с, 1H, CH-2-Im).
Пример 4.
Nα-АЦЕТИЛ-γ-L- глутамилгистамин (V).
К 0.10 (0.405 ммоль) γ-L- глутамилгистамина прибавляют 5 мл воды и перемешивают до растворения основной массы вещества. К реакционной смеси прибавляют 2.5 мл диоксана и 0.073 г (0.405 ммоль) n-нитрофенилацетата, перемешивают 2 ч и оставляют на 18 ч при 20oC. Растворитель удаляют в вакууме при 40oC. Остаток растворяют в минимальном количестве метанола и очищают колоночной хроматографией на Kieselgel 60, элюируют метанолом. Фракции, содержащие целевое вещество с Rf 0.3 (4), объединяют, растворитель удаляют в вакууме. Получают бесцветное стеклообразное вещество. Выход 0.046 г (40.0%). Rf 0.3 (4). ВЭЖХ в условиях (3): один пик, время выхода 10.77 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 2.0-2.3 (м, 2H, β-CH2), 2.19 (с, 3H, CH3CO), 2.45 (т, 2H, γ- CH2), 3.07 (т, 2H, β- CH2-HA), 3.64 (т, 1H, α- CHAHB-HA), 3.65 (т, 1H, α- CHAHB-HA), 4.42 (т, 1H, α- CH), 7.42 (д, 1H, CH-4-Im), 8.69 (д, 1H, CH-2-Im).
Пример 5.
Метиловый эфир Nα-ТРЕТ.-бУТИЛОКСИКАРбОНИЛ-α- бензил-L-глутамил-гистидина (VIa).
К раствору 0.30 г (1.16 ммоль) дигидрохлорида метилового эфира L-гистидина, полученному нагреванием до 40oC с 4 мл безводного метанола с последующим охлаждением до 0oC, прибавляют холодный раствор метилата натрия, полученный из 0.053 г металлического натрия (2.32 ммоль) и 1 мл метанола. Оставляют на 20 мин при 0oC, затем прибавляют равный объем сухого эфира и оставляют на 20 мин при 0oC. Осадок хлористого натрия отделяют, растворитель из фильтрата удаляют в вакууме. Остаток растворяют в 3.5 мл DMF и прибавляют 0.604 г (1.16 ммоль) Boc-L-Glu(OPfp)-OBzl. Реакционную смесь перемешивают 2 ч и оставляют на 20 ч. DMF удаляют в вакууме. Маслообразный остаток очищают на колонке 30 х 1.6 см с силикагелем 100/160, элюируют смесью хлороформ: метанол (9: 1). Фракции, содержащие целевой продукт, объединяют, растворитель удаляют в вакууме. Целевое вещество сушат над P2O5. Выход 0.334 г (55.0%). Rf 0.35 (1). 1H-ЯМР спектр (CD3OD): 1.45 (c, 9H, (CH3)3O, 2.0 (м, 2H, β-CH2), 2.3 (т, 2H, γ-CH2), 3.0 (д, 2H, β-CH2- His), 3.7 (с, 3H, CH3O), 4.1 (т, 1H, α-CH), 4.55 (т, 1H, α-CH-His), 5.15 (м, 2H, CH2-Bzl), 6.85 (с, 1H, CH-4-Im), 7.35 (м, 5H, C6H5-Bzl), 7.6 (с, 1H, CH-2-Im).
Дигидрохлорид метилового эфира γ-L-глутамил-L-гистидина (VI).
К раствору 0.30 г Boc-L-Glu(HisOMe)-OBzl (Vla) в 1 мл MeOH прибавляют 3 мл 4 н. раствора хлористого водорода в диоксане. Через 30 мин прибавляют 5 мл сухого эфира. Растворители удаляют в вакууме, добавляют сухой эфир и также удаляют в вакууме. Остаток растирают с сухим эфиром. Эфир отделяют декантацией. Белое твердое вещество сушат над P2O5 в вакууме. Получают 0.25 г 2HCI•L-Glu(HisOMe)OBzl.
К раствору 0.140 г полученного вещества в 4.5 мл безводного метанола прибавляют 0.10 г 10%-го палладия на активированном угле и перемешивают 2.5 ч, периодически пропуская ток водорода. Катализатор отделяют, промывают на фильтре MeOH. Растворитель из объединенного фильтрата удаляют в вакууме. К остатку добавляют сухой эфир и также удаляют в вакууме. Вещество сушат в вакууме над P2O5. Выход 0.103 г (90.3%). Rf 0.35 (6). ВЭЖХ в условиях (9): один пик, время выхода 6.16 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 2.15 (м, 2H, β-CH2), 2.55 (т, 2H, γ-CH2), 3.15 (т, 1H, α-CH), 3.75 (с, 3H, CH3O), 4.0 (т, 1H, α-CH-His), 4.8 (д, 2H, β- CH2-His), 7.4 (с, 1H, CH-4-Im), 8.81 (с, 1H, CH-2-Im). Масс-спектр, m/z: 299.1.
Пример 6.
Метиловый эфир глутарил-L-гистидина (VII).
К раствору 0.30 г (1.16 ммоль) дигидрохлорида метилового эфира L-гистидина, полученному нагреванием до 40oC в 4 мл безводного метанола с последующим охлаждением до 0oC, прибавляют холодный раствор метилата натрия, полученный из 0.053 г (2.32 ммоль) металлического натрия и 1 мл метанола. Реакционную смесь оставляют на 20 мин при 0oC, затем прибавляют равный объем сухого эфира и оставляют на 20 мин при 20oC. Осадок хлористого натрия отделяют, растворитель удаляют в вакууме. Остаток растворяют в 3.5 мл DMF и прибавляют 0.132 г (1.16 ммоль) глутарового ангидрида. Реакционную смесь перемешивают 2 ч и оставляют на 20 ч при 20oC. Растворитель удаляют в вакууме. Маслообразный остаток очищают на колонке (30 х 1.6 см) с силикагелем 100/160, элюируют метанолом. Фракции, содержащие целевой продукт, объединяли. Растворитель удаляют в вакууме, сушат над P2O5. Выход 0.095 г (28%). Rf 0.43 (10). ВЭЖХ в условиях (8): один пик, время выхода 9.95 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 1.85 (м, 2H, β-CH2), 2.25 (т, 4H, α,γ- CH2), 3.05 (д, 2H, β- CH2-His), 3.7 (с, 3H, CH3O), 4.67 (т, 1H, α- CH-His), 6.92 (с, 1H, CH-4-Im), 7.72 (с, 1H, CH-2-Im). Масс-спектр, m/z: 284.4.
Пример 7.
Пентафторфениловый эфир N-бензилоксикарбонил - γ - аминомасляной кислоты (VIIIa).
К 0.60 г (2.53 ммоль) N- бензилоксикарбонил- γ -аминомасляной кислоты прибавляют 9 мл безводного этилацетата, перемешивают и охлаждают до 0oC. Прибавляют 0.52 г (2.53 ммоль) DCC и 0.465 (2.53 ммоль) пентафторфенола. Реакционную смесь перемешивают при охлаждении 2 ч, после чего оставляют на 20 ч при 20oC. Осадок DCU отделяют, промывают безводным этилацетатом. Фильтраты объединяют, растворитель удаляют в вакууме. Получают белые, слегка желтоватые кристаллы Z-GABA-OPfp, которые сушат в вакууме. Выход 1.05 г (98.0%). Rf 0.85 (1).
Метиловый эфир N-бензилоксикарбонил -γ- аминобутирил-L-гистидина (Vlllб).
К 0.30 г (1.16 ммоль) дихлоргидрата метилового эфира L-гистидина прибавляют 4 мл безводного метанола и нагревают до растворения. Раствор охлаждают до 0oC и прибавляют охлажденный раствор метилата натрия, приготовленный из 0.053 г (2.32 ммоль) металлического натрия и 1 мл безводного метанола. Реакционную смесь оставляют при 0oC на 20 мин, прибавляют равный объем сухого эфира и оставляют при 20oC на 20 мин. Осадок хлористого натрия отделяют. Растворитель из фильтрата удаляют в вакууме. К маслообразному остатку прибавляют 5 мл безводного DMF, 0.50 г (1.24 ммоль) Z-GABA-OPfp и оставляют на 20 ч при 20oC. Растворитель удаляют в вакууме. Остаток очищают колоночной хроматографией на силикагеле L 40/100, элюируют хлороформом и градиентом метанола в хлороформе до соотношения 2:8. Фракции, содержащие целевое вещество с Rf 0.46 (2), объединяют, растворитель удаляют в вакууме. К полученному остатку прибавляют избыток сухого эфира с 1 каплей триэтиламина, растирают. Белое твердое вещество отделяют, промывают сухим эфиром, сушат в вакууме. Выход 0.243 г (65.0%), Rf 0.46(2). Масс-спектр, m/z: 375.
Метиловый эфир γ- аминобутирил-L-гистидина (VIII).
К 0.04 г (0.124 ммоль) Z-GABA-L-His-OMe прибавляют 7 мл безводного метанола, 0.04 г 10%-го палладия на активированном угле и гидрируют 1 час при перемешивании. После полного превращения исходного вещества с Rf 0.46 (2) в целевой продукт с Rf 0 (2) катализатор отделяют, промывают метанолом. Фильтраты объединяют, растворитель удаляют в вакууме. Получают бесцветное вязкое масло. Выход 0.030 г (90.0%). Rf 0.05 (3), Rf 0.1 (4). ВЭЖХ в условиях (1): один пик, время выхода 4.8 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 1.85 (м, 2H, β- CH2), 2.23 (т, γ- CH2), 2.95 (д, 2H, β- CH2-His), 3.1 (т, α- CH2), 3.7 (с, 3H, CH3O), 4.65 (т, 1H, α- CH-His), 6.85 (с, 1H, CH-4-Im), 7.6 (с, 1H, CH-2-Im).
Пример 8.
Nα-трет.-бутилоксикарбонил-α- бензил-L-глутамилгистидин (IXа).
К раствору 0.100 г (0.645 ммоль) L-гистидина в 3 мл воды при перемешивании прибавляют 0.75 мл диоксана и затем порциями в течение 2 ч 0.560 г (1.29 ммоль) Boc-L-Glu(ONSu)OBzl и 2.25 мл диоксана (до соотношения диоксан: вода 1:1). Образовавшуюся суспензию перемешивают 2 ч и оставляют на 20 ч при 20oC, после чего суспензия превращается в раствор. Растворитель удаляют в вакууме. Маслообразный остаток очищают на колонке 30 х 1.6 см с силикагелем 100/160, элюируют смесью хлороформ-метанол 5:1, постепенно увеличивая содержание MeOH до 50%. Фракции, содержащие целевой продукт, объединяют, растворитель удаляют в вакууме. Получают белое твердое вещество.
Выход 0.150 г (45.7%). Rf 0.48 (3). 1H-ЯМР спектр (CD3OD), δ, м.д.: 1.4 (с, 9H, (CH3)3C), 1.98 (м, 2H, β- CH2), 2.3 (т, 2H, γ- CH2-Glu), 3.1 (м, 2H, β- CH2-His), 4.07 (т, 1H, α- CH), 4.5 (т, 1H, α- CH-His), 5.18 (м, 2H, CH2-Bzl), 7.17 (с, 1H, CH-4-Im), 7.35 (м, 5H, C6H5), 8,4 (с, 1H, CH-2-Im).
Дихлоргидрат α- метилового эфира -γ- L-глутамилгистидина (IX).
К раствору 0.140 г Boc-L-Glu(His)-OBzl (IXа) в 1 мл безводного метанола прибавляют 3 мл 4 н. раствора хлористого водорода в диоксане и выдерживают 40 мин при 20oC. К реакционной смеси прибавляют эфир, и растворители удаляют в вакууме. К остатку дважды прибавляют порциями эфир и удаляют в вакууме. Осадок растирают с эфиром до образования белого твердого вещества. Эфир декантируют, остаток сушат в вакууме и очищают препаративной бумажной хроматографией в системе бутанол-уксусная кислота-вода 4:1:5, верхняя фаза. Получают 0.12 г (95.8%) дихлоргидрата дипептида, Rf 0.26 (3). К раствору 0.117 г полученного продукта в 4.5 мл безводного метанола прибавляют 0.096 г 10%-го палладия на активированном угле. Реакционную смесь перемешивают 2.5 ч, периодически пропуская ток водорода. Катализатор отделяют, промывают на фильтре 5 мл MeOH. Фильтраты объединяют, растворитель удаляют в вакууме. Остаток сушат в вакууме над P2O5. Выход 0.091 г (96.1%). Rf 0.3(6). ВЭЖХ в условиях (10): один пик, время выхода 5.40 мин. Масс-спектр, m/z: 299.2. 1H-ЯМР спектр (CD3OD), δ, м.д.: 2.15 (м, 2H, β- CH2), 2.55 (т, 2H, γ- CH2), 3.16 (т, 1H, α- CH), 3.75 (с, 3H, CH3O), 3.99 (т, 1H, α- CH-His), 4.8 (дд, 2H, β- CH2-His), 7.38 (с, H, CH-4-Im), 8.80 (с, H, CH-2-Im).
Пример 9.
N-бензилоксикарбонил -γ- аминобутирилтриптамин (Xа).
К раствору 0,252 г (0,625 ммоль) Z-GABA-OPfp в 5 мл DMF прибавляют 0.10 г (0.625 ммоль) триптамина и перемешивают при 25oC 2 ч. Реакционную смесь оставляют на 16 ч при той же температуре, после чего прибавляют 7-кратный избыток воды (по объему). Белый творожистый осадок отфильтровывают, промывают на фильтре водой, сушат. Выход 0.22 г (93.0%). Rf 0.6 (5), Rf 0.54 (3). Масс-спектр, m/z: [М+1H]+ 380.6.
γ- Аминобутирилтриптамин (X).
К 0.133 г (0.35 ммоль) Z-GABA-TrpA прибавляют 7 мл безводного метанола, перемешивают до растворения основной массы вещества, прибавляют 0.133 г 10%-го палладия на активированном угле и гидрируют 1 ч. После полного превращения исходного вещества с Rf 0.6 (5) в целевой продукт с Rf 0 (5) катализатор отделяют, промывают метанолом. Фильтраты объединяют, растворитель удаляют в вакууме. Получают бесцветное вязкое масло. Выход 0.086 г (99.0%). Rf 0.43 (3). ВЭЖХ в условиях (2): один пик, время выхода 18.5 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 1.65 (м, 2H, β- CH2), 2.05 (т, 2H, γ- CH2), 2.85 (т, 2H, β- CH2-TrpA), 3.0 (т, 2H, α- CH2), 3.45 (м, 2H, α- CH2-TrpA), 6.95 (с, 1H, CH-2-Ind), 6.99 (м, 1H, CH-5-Ind), 7.05 (м, 1H, CH-6-Ind), 7.28 (д, 1H, CH-7-Ind), 7.48 (д, 1H, CH-4-Ind).
Пример 10.
Глутарил-O-бензилсеротонин (XI).
К суспензии 0.20 г (0.66 ммоль) хлоргидрата O-бензилсеротонина в 3 мл DMF прибавляют при перемешивании 0.09 мл (0.66 ммоль) триэтиламина, а затем 0.075 г (0.66 ммоль) глутарового ангидрида. Реакционную массу перемешивают 1 ч и оставляют на 20 ч при 20oC. Осадок хлоргидрата триэтиламина отделяют, растворитель удаляют в вакууме. Маслообразный остаток очищают на колонке 31 х 1.5 см с силикагелем Silica gel 60, 0.063- 0.2 мм (Merck). Элюируют смесью хлороформ - метанол 9:1. Фракции, содержащие целевой продукт, объединяют, растворитель удаляют в вакууме. Получают бесцветный стеклообразный продукт. Выход 50 мг (20 %). Rf 0.48 (1). ВЭЖХ в условиях (14): один пик, время выхода 23.8 мин. 1H- ЯМР спектр (CD3OD), δ, м.д.: 2.05 (м, 2H, β- CH2-GA), 2.45 (м, 4H, α,γ- CH2-GA), 3.1 (т, 2H, β- CH2), 3.67 (т, 2H, α- CH2), 5.3 (с, 2H, CH2-Bzl), 7.0 (дд, 1H, CH-6-Ind), 7.1- 7.7 (м, 8H, CH-7-Ind, CH-2-Ind, CH-4-Ind, C6H5).
Пример 11.
N-глутарилсеротонин (XII).
К 25 мг (0.065 ммоль) глутарил-O- бензилсеротонина прибавляют 4 мл безводного метанола. К раствору добавляют 30 мг катализатора - палладия на активированном угле и гидрируют 1 ч при перемешивании. Катализатор отфильтровывают. Растворитель из фильтрата удаляют в вакууме. Выход 17 мг (90 %). Rf 0.31 (1). ВЭЖХ в условиях (14): один пик, время выхода 16.27 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 2.0 (м, 2H, β- CH2-GA), 2.5 (м, 4H, α,γ- CH2-GA), 3.0 (т, 2H, β- CH2), 3.65 (т, 2H, α- CH2), 6.85 (дд, 1H, 6-CH-Ind), 7.15 (д, 1H, CH-4-Ind), 7.25 (с, 1H, CH-2-Ind), 7.4 (д, 1H, CH-7-Ind).
Пример 12.
Глутарил-5-O-метилсеротонин (XIII).
К раствору 0.20 г (1.05 ммоль) O-метилсеротонина в 3 мл DMF прибавляют при перемешивании 0.12 г (1.05 ммоль) глутарового ангидрида. Реакционную массу перемешивают 1 ч и оставляют на 20 ч при 20oC. Растворитель удаляют в вакууме. Маслообразный остаток очищают на колонке 31 х 1.5 см с силикагелем Silica gel 60, 0.063-0.2 мм (Merck). Элюируют смесью хлороформа и метанола 9:1. Фракции, содержащие целевой продукт, объединяют, растворитель удаляют в вакууме. Получают бесцветное стеклообразное вещество. Выход 0.095 г (29.8%). Rf 0.51 (1). ВЭЖХ в условиях (14): один пик, время выхода 19.0 мин. 1H-ЯМР спектр (CDCl3+CD3OD), δ, м.д.: 1.95 (м, 2H, β- CH2-GA), 2.37 (м, 4H, α,γ- CH2-GA), 3.0 (т, 2H, β- CH2), 3.56 (т, 2H, α- CH2), 3.92 (с, 3H, CH3O), 6.83 (дд, 1H, CH-6-Ind), 7.1 (д, 2H, CH-2,4-Ind), 7.31 (д, 1H, CH-7-Ind).
Пример 13.
Глутарил-4-(2-аминоэтил)морфолин (XIV).
К раствору 0.5 г (4.39 ммоль) глутарового ангидрида в 1 мл DMF при перемешивании и охлаждении водой прибавляют 0.57 мл (4.39 ммоль) 4-(2-аминоэтил) морфолина. Перемешивают 30 мин, оставляют на 20 ч при 20oC. Растворитель удаляют в вакууме. Маслообразный остаток промывают эфиром, растворитель удаляют в вакууме, сушат и оставляют при +4oC на 20 ч. Образующееся кристаллическое вещество промывают и растирают с эфиром (3 х 1.5 мл), а затем с ацетоном (4 х 1.5 мл), отделяют от растворителей, сушат в вакууме. Получают 0.265 г (24.8%), Rf 0.41 (6). ВЭЖХ в условиях (11): один пик, время выхода 3.93 мин. 1H-ЯМР спектр (CD3OD), δ, м.д.: 0,5 (м, 2H, β- CH2-GA), 0.9 (м, 4H, α,γ- CH2-GA), 1.3 (м, 8H, CH2-2,3,5,6-Morph), 2.0 (т, 2H, β- CH2), 2.38 (т, 2H, α- CH2).
Пример 14.
Глутарил-2-(2-аминоэтил)-пиридин (XV). К раствору 0.3 г (2.63 ммоль) глутарового ангидрида в 1 мл DMF прибавляют при перемешивании и охлаждении водой 0.315 мл (2.63 ммоль) 2-(2- аминоэтил)-пиридина. Перемешивают 1 ч и оставляют на 20 ч при 20oC. Выпавший белый осадок отфильтровывают, многократно промывают эфиром и ацетоном, сушат. Выход 0.29 г (46.7 %). Rf 0.27 (1). ВЭЖХ в условиях (12): один пик, время выхода 14.33 мин. 1H-ЯМР спектр (CD3OD), δ, м. д.: 0.41 (м, 2H, β- CH2-GA), 0.81 (м, 4H, α,γ- CH2-GA), 1.6 (т, 2H, β- CH2), 2.15 (т, 2H, α- CH2), 5.9 (м, 2H, CH-4,5-Py), 6.4 (м, 1H, CH-3-Py), 7.05 (м, 1H, CH-6-Py).
Пример 15.
Глутарилфенилэтиламин (XVI).
К раствору 0.5 г (4.39 ммоль) глутарового ангидрида в 1 мл DMF прибавляют при перемешивании и охлаждении водой 0.55 мл (4.39 ммоль) фенилэтиламина. Перемешивают 30 мин, оставляют на 20 ч при 20oC. Растворитель удаляют в вакууме, остаток - густое масло - очищают на колонке 25 х 230 мм с Kieselgel 40 для колоночной хроматографии ("Fluka"). Элюируют градиентом хлороформ - MeOH (9:1). Фракции, содержащие целевой продукт с Rf 0.57(1) объединяют, растворитель удаляют в вакууме, сушат. Выход 1.02 г (99.03%). ВЭЖХ в условиях (13): один пик, время выхода 15.12 мин. 1H-ЯМР спектр (CDCl3), δ, м.д.: 1.9 (м, 2H, β- CH2-GA), 2.15 (т, 4H, α,γ- CH2-GA), 2.8 (т, 4H, α,β- CH2), 7.2 (м, 5H, C6H5).
По аналогичным типовым методикам получают также новые соединения общей формулы (I), приведенные в таблице 1а (см. в конце описания)
Фармакологические свойства.
Ранее, исходя из антиокислительных свойств известных соединений общей формулы (I), а именно β- аланилгистамина (XIII), γ- аминобутирилгистамина (XIV), а также α-L-Glu-HA, было предложено использовать их для лечения ряда заболеваний, в частности атеросклероза, аллергии [Seguin М.C., Babizhayev М. / Product de couplage de l'un acide amine, procede de preparation, et applications therapeutigues et cosmetologigues.// Patent Fr 2701947. 1994. C1 C 07 C 237/04. A 61 K 31/195. 7/48; Babizhayev M., Seguin M.C. / Pseudodipeptide product having on imidazole grouping and applications. // WO 95/12581. C1 C 07 D 233/64. C 12 P 17/10. A 61 K 31/415. 7/42]. Однако эти показания к применению не были подтверждены экспериментально in vivo, а был продемонстрирован только один из многих возможных механизмов действия веществ в опытах in vitro. Как было показано нами ранее [Рожкова Е.А., Огрель C. А. , Григорьев Д.Н., Небольсин В.Е., Желтухина Г.А., Евстигнеева Р.П. / Взаимосвязь между конформацией и антиоксидантными свойствами в ряду топохимических аналогов карнозина и карцинина с различными N-ацильными заместителями.// Журнал биоорганической химии. -1996, -т. 22, -N 10-11, -с.838-45], γ- аминобутирилгистамин обладает лишь слабой антиоксидантной активностью, a α-L-Glu-HA ингибирует образование супероксиданиона лишь в небольшой степени при высоких концентрациях, не влияя на образование гидроксильного радикала [Babizhayev M. , Seguin M.C. / Pseudodipeptide product having on imidazole grouping and applications.// WO 95/12581. -C1 C 07 D 233/64. -C 12 P 17/10. -A 61 K 31/415. -7/42]. Кроме того, можно прогнозировать большую неустойчивость α-L-Glu-HA к действию ферментов in vivo, чем у соответствующего γ- аналога.
Для наиболее близких по структуре к соединениям общей формулы (I) аналогов γ- глутамилдипептидов, а именно N-ацильных производных γ- глутамиламинокислот и/или их эфиров, была показана их способность ингибировать ангиотензинпревращающий фермент и энкефалиназу и предложено применять их для понижения кровяного давления и анальгезии /6/.
Исходя из этих данных, нами предпринято расширенное исследование биологической активности соединений общей формулы (I). В результате для них были выявлены неизвестные ранее виды фармакологической активности, экспериментально подтверждены в опытах in vivo известные и предложены механизмы их действия на основании экспериментов in vitro. Так, было показано, что соединения общей формулы (I) обладают антиоксидантной и антирадикальной активностями, а также могут воздействовать на систему цитохрома P-450, модулировать метаболизм арахидоновой кислоты, гормонов коры надпочечников.
Совокупность вышеотмеченных свойств определяет ценность соединений общей формулы (I) в качестве фармакологических средств для лечения ряда заболеваний.
Примеры разнообразных видов биологической активности, проявляемых соединениями общей формулы (I) in vitro и in vivo, приведены ниже.
Пример 16.
Влияние соединений, отвечающих общей формуле (I), на продолжительность гексеналового сна у животных.
Изменения длительности гексеналового сна (ГС) - показатель состояния системы цитохрома Р-450 печени, измеряемый in vivo.
Исследования проводили на мышах-самцах линии BALB/c•h75, C 57 В 1/6, CBA, DBA/2, гибридах первого поколения CBF1 и BDF1 с исходной массой 18-20 г, в каждой группе использовали по 10 мышей, и на беспородных морских свинках-самцах с исходной массой 250-300 г, группы содержали по 24 животное. Все соединения исследуемого ряда вводили перорально в питьевой воде и в течение 3-х суток в дозах 50 и 500 мкг/кг. Гексенал-гидрохлорид в дозе 36 мг/кг массы животного вводили через 24 часа после последнего введения препарата, за исключением мышей линии BALB/c•h75, которым вводили гексенал в дозе 60 мг/кг в те же сроки, и морских свинок, для которых доза гексенала составляла 30 мк/кг. Длительность ГС определяли в минутах.
Результаты обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -M. -Наука. -1978. -365 с.].
В таблице 2 представлены данные изменения длительности гексеналового сна (ГС) у мышей различных линий, гибридов BDF1 и морских свинок под влиянием одного из соединений общей формулы (I). Показано, что препарат снижал длительность ГС у всех подопытных животных как в дозе 50, так и 500 мкг/кг.
При изучении изменения длительности ГС под влиянием ряда веществ общей формулы (I) на мышах CBF показано, что препараты снижали длительность ГС от 13% до 54% по сравнению с контролем.
Таким образом, все исследованные соединения уменьшают длительность гексеналового сна, что свидетельствует об индукции ими системы цитохрома P-450 печени.
Пример 17.
Изменение состояния системы цитохрома P-450 печени под влиянием одного из соединений общей формулы (I).
Изменение состояния монооксигеназной системы печени изучали на беспородных морских свинках-самцах с исходной массой 250-300 г, в каждой группе содержалось по 12 животных. Одно из исследованных соединений и фенобарбитал (ФБ) вводили трехкратно за 72, 48 и 24 часа до проведения исследования монооксигеназной системы. Содержание цитохромов Р-450 и в5 измеряли по методу [Omura Т. , Sato R./The monooxide binding pigment of liver microsomes. 11. Solubillization, purification and properties,// J.Chem. -1964. -vol. 239. -pp. 2379-2385.] в микросомальной фракции печени, выделенной методом дифференциального центрифугирования [Ahokas J., Pelkonen O., Karkin N. /Characterisation of BP-hydroxylase of Trout-liver.// Cancer res. -1977. -vol. 37. -pp. 3737-3743]. Для определения групп цитохромов Р-450В и Р-450Л использовали приоритетный метод, разработанный М. И. Изотовым [Изотов М.В., Щербаков В.М., Девиченский В.М. и др. / Способ определения содержания изоферментов цитохрома Р-450 в микросомах печени.// А.С. N 1488738. -Б.И. -1989. -N3. -6.06.09, МКИ, 01. -N 33/15, N 33/48.]. Содержание микросомального белка определяли модифицированным методом Лоури [Hartree E. /Determination of protein: a modification of the Lowry method, that gives a linear photometric respones.//Ann. Biochem. -1972. -vol. 48. -pp. 422-427].
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты, представленные в таблице 3, свидетельствуют, что у морских свинок, предобработанных исследованным соединением и ФБ, достоверно повышается общее содержание цитохромов Р-450 и в5 печени. При этом у животных групп 2 и 3 отмечалось достоверное увеличение количества цитохромов Р-450В, а у животных, получавших изучаемое соединение общей формулы (I), - также соотношения цитохромов Р-450В к Р-450Л и соотношения цитохромов в5 и Р- 450.
Выявлено, что исследованное соединение изменяет систему цитохрома Р-450 печени подобно ФБ, однако имеет свои особенности, такие как повышение соотношения цитохромов Р-450В/Р-450Л и цитохрома в5 к Р-450.
Пример 18.
Изменение гормонального статуса животных под влиянием одного из соединений общей формулы (I).
Исследования проводили на беспородных морских свинках-самцах с исходной массой 250-300 г, в каждой группе использовали по 12 животных. Сенсибилизацию животных осуществляли как описано в примере 25. Кровь для определения гормонов у животных отбирали до начала эксперимента и через 18 часов после последнего введения препарата с 10 до 11 часов. Вещество вводили сенсибилизированным и интактным животным в дозе 50 мкг/кг трехкратно перорально за 72, 48 и 18 часов до повторного забора крови. Сравнительный анализ изменения гормонального статуса проводили по индивидуальным изменениям у каждого животного в группе. В таблице 4 приведены средние значения показателей по группам, выраженные в нмоль/л.
Гормоны - прогестерон, 17-оксипрогестерон, 11- дезоксикортизол и кортизол определяли радиоиммунологическим методом в сыворотке крови с использованием наборов фирмы "Белорис". Концентрация гормона в пробах определялась из графика зависимости активности осажденного связанного меченного гормона от концентрации гормона в калибровочных пробах.
Полученные данные обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты эксперимента представлены в таблице 4. Введение одного из исследуемых соединений интактным животным приводило к достоверному повышению количества свободного кортизола (F), его предшественников оксипрогестерона (17-OН-Р) и дезоксикортизола, и отмечалась тенденция к увеличению содержания прогестерона (Р). У сенсибилизированных морских свинок выявлено достоверное уменьшение количества F и 17-OН-Р и тенденция к снижению P в сыворотке крови. У сенсибилизированных животных, которым вводили соединение, отмечалась нормализация содержания F и дезоксикортизола в сыворотке крови и достоверное увеличение количества P и 17-OН-Р.
При введении экспериментальным животным, как интактным, так и сенсибилизированным, одного из известных соединений отмечались аналогичные изменения гормонального статутса.
Пример 19.
Влияние одного из соединений общей формулы (I) на метаболизм [C14]-арахидононовой кислоты в гомогенате легочной ткани.
Исследования проводили на мышах-самках линии CBA, находившихся на стандартном рационе вивария. Животным вводили соединение в дозе 50 мкг/кг и фенобарбитал в дозе 80 мг/кг в течение трех дней. Затем животных забивали, извлекали легкие, замораживали их в жидком азоте, гомогенизировали в стеклянном гомогенизаторе фирмы "Wheaton" (США) при +4oC в 10 объемах 0,05 М трис-HCl буфера. Аликвоты (0,5 мл) супернатанта инкубировали с 0,5 мкКю [1- C14] -арахидоновой кислоты ([C14]-AA), "Amersham", Англия; удельная активность 50-60 мКю/ммоль при +37oC в течение 30 мин. Экстракцию неметаболизированной [C14] -AA и продуктов ее метаболизма осуществляли в 20 объемах смеси хлороформа и метанола (1:1) при эффективности экстракции не менее 90%, оцененной с помощью [C14]- ПГF2α. Разделение и идентификацию [C14]-AA и ее метаболитов осуществляли при помощи тонкослойной хроматографии (пластины Kieselgel 60 фирмы "Merck", Германия) с использованием органической фазы системы растворителей (этилацетат, изооктан, уксусная кислота, вода - 110:50:20:100) и меченных стандартов. Авторадиохроматограммы, полученные на рентгеновской пленке X-Omat AR ("Kodak", США) и HS 11 ("ORWO", Германия), денситометрировали на денсискане KS 3 ("Kipp and Zonnen", Голландия). Количественный анализ отдельных эйкозаноидов проведен с помощью радиометрии фракций, полученных высокоэффективной жидкостной хроматографией (HPLC-система фирмы "Gilson", Франция; колонка ZORBAX C8 фирмы "Du Pont", США) и элюированием пятен на ТСХ-пластинках.
Результаты эксперимента представлены в таблице 5. Выявлено, что одно из исследованных соединений стимулирует образование циклооксигеназных метаболитов АА, а именно увеличивает синтез простагландина E2 (ПГЕ2), 6-кето-ПГF1α,ПГF2α, и липоксигеназный путь метаболизма AA повышает образование 5-НЕТЕ и ННТ. При этом наблюдается снижение метаболизма AA в системе цитохрома Р-450 - 12-НЕТЕ и 15-НЕТЕ. Следует отметить, что изменение профиля метаболитов AA у животных, предобработанных одним из соединений общей формулы (I), сходно с тем, которое выявлено у животных, получавших фенобарбитал - известный индуктор системы цитохрома Р-450.
Пример 20.
Изменение антиген-зависимой секреции гистамина перитонеальными тучными клетками сенсибилизированных крыс под влиянием одного из соединений общей формулы (I).
Сенсибилизацию крыс-самцов Вистар с исходной массой 200-250 г проводили по методу [Guschin I.S., Voitenko V.G., Sviridov B.D. et al./A polyfunctional molecule produce by the conjugation of synthetic polyionimmunostimulant with specific antigen and an inhibitor of mast cell activation. Effects on histamine release. // Agents and Actions. -1989. -vol. 27. -S. -pp. 75-78]. На 14-й день сенсибилизации у животных по стандартной методике [Fredholm В. В. , Gyschin I.S., Flwin К. et al./ Cyclic AMP-independ inhibition by papaverine of histamine release induced by compound 48/80.//Biochem. Pharmacol. -1976. -vol. 25. -pp. 1583-1588.] выделяли клеточную взвесь из брюшной полости, определяли спонтанную и стимулированную куриным овальбумином (ОВА) секрецию гистамина тучными клетками (ТК), а также содержание гистамина в ТК контрольных и опытных животных. Клеточную взвесь в количестве 2 мл, содержащую 0,1-0,2•106 ТК/мл инкубировали в присутствии 200 мкг/мл OBA. Секрецию гистамина выражали в процентах к его общему содержанию. Количество гистамина в образцах определяли спектрофлуориметрически [Short P.A., Burkhalter A., Cohn V.N./ A method for fluometric assay of histamine in tissues.// J.Pharmacol., exp. Ther. -1959. -voL 127. -pp. 182-186.]
Вещества вводили животным внутрибрюшинно по следующей схеме - в течение 3-х дней перед исследованием ТК: исследуемое соединение - в дозе 50 мкг/кг (2 группа), супрастин - в дозе 1000 мкг/кг (3 группа). Животные контрольной группы получали внутрибрюшинно физиологический раствор (1 группа). В каждой группе использовали по 10 животных.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов.- М.- Наука.- 1978.- 365 с.].
Результаты исследования представлены в таблице 6. Показано, что введение одного из соединений общей формулы (1) достоверно снижает антиген-зависимую секрецию гистамина ТК. При этом отмечено достоверное повышение спонтанной секреции гистамина ТК. Данные изменения происходят на фоне снижения количества гистамина в ТК сенсибилизированных животных, получавших одно из исследованных соединений.
Пример 21.
Влияние соединений общей формулы (I) на образование активных форм кислорода в модельных системах.
Изучалось влияние соединений общей формулы (I) на изменение хемилюминесценции (ХЛ), обусловленной образованием гидроксильного радикала (.ОН) и супероксидного анион-радикала (O2 -), в модельных химических и ферментативных системах.
Активные формы кислорода (АФК) различной природы генерировали в следующих системах:
А. Гидроксильный радикал - в смеси FeSO4 с H2O2 (реактив Фентона) [Halliwell В. / Superoxide-dependent formation of hydroxyl radicals in the presence of iron salts.// FEBS Lett. -1978. -vol. 96. -pp. 238-241].
Инкубационная среда состояла из 5 мМ KH2PO4 (pH 7,4); 5 мМ FeSO4; 2 мМ люминола. H2O2 в конечной концентрации 5 мМ вводили в кювету через диспендер после того, как было зарегистрировано фоновое свечение смеси реагентов. Исследуемые соединения, растворенные в воде, в необходимой концентрации вносили в кювету в объеме 5-10 мкл. Общий объем пробы составлял 0,5 мл.
В. Супероксидный анион - радикал - в смеси ксантина и ксантиноксидазы [Afanasev I. , Suslova Т. , Cheremisina Z. et al. / Study of antioxidant properties of metal aspartates.// Analyst. -1995. -vol. 120. -pp. 859-862].
Инкубационная среда содержала следующие инградиенты: 5 мМ KH2PO4; 0,2 МЕ/мл ксантиноксидазы. Ксантин в концентрации 1 мМ вводили в пробу через диспендер. В качестве сенсибилизатора свечения в данной системе использовали люцигенин (0,2 мМ). Исследуемые вещества вводили аналогично тому как при исследовании гидроксильного радикала.
В предварительных исследованиях было показано, что соединения общей формулы (I) не влияли на активность ксантиноксидазы. Измерение ХЛ описанных систем проводили при 25oC в режиме пульсового (в момент введения реагентов через диспендер) перемешивания. Индикация сигнала ХЛ осуществлялась путем его интегрирования каждые 10 сек - 5 мин. Длительность регистрации вспышки ХЛ после смешивания инградиентов зависела от кинетики конкретного процесса. Для каждой системы определяли светосумму ХЛ (мВ) в контрольных пробах без препаратов (I-) и в пробах в присутствии соответствующих концентраций препаратов (I+). Для оценки степени ингибирования (активации) ХЛ в изученных системах находили отношение I+/I- (относительные единицы).
Образование АФК и влияние на этот процесс исследуемых соединений регистрировали на приборе Luminometer-1251 (LKB, Швеция).
А. Влияние соединений общей формулы (1) на образование гидроксильного радикала в реактиве Фентона.
При введении H2O2 в фосфатный буфер, содержащий сульфат железа, возникала вспышка ХЛ, которая была обусловлена образованием в реакционной смеси преимущественно гидроксильного радикала. Вспышка носила кратковременный характер, максимум интенсивности достигался на 10-й секунде и в течение следующих 10-20 секунд свечение гасло и показатели ХЛ уменьшались до фоновых значений.
В таблице 7 представлены данные исследования действия соединений на генерацию гидроксильного радикала в реактиве Фентона. Результаты показывают, что исследованные соединения ингибировали образование гидроксильного радикала в изученной системе.
В. Влияние соединений общей формулы (I) на образование супероксидного анион-радикала в системе ксантин-ксантиноксидаза.
Введение ксантина в среду, содержащую ксантиноксидазу и люцигенин, приводило к возникновению вспышки ХЛ, которая достигала максимума за 3-5 минут, после чего в изучаемой системе начиналось очень медленное уменьшение интенсивности хемилюминесценции. Добавление исследуемых соединений в смесь ксантин-ксантиноксидазы принципиально не меняло форму кривой ХЛ-ответа, варьировались только значения максимальной интенсивности ХЛ. Вследствие такой "растянутости" кинетической кривой регистрировалась светосумма хемилюминесценции за первые 3-5 минут, которая отражала суммарное количество квантов света, вырабатываемых в системе до достижения максимальных значений интенсивности ХЛ.
Как видно из представленных данных (таблица 8) исследованные соединения обладают способностью достоверно ингибировать образование супероксидного анион-радикала.
Пример 22.
Изучение антигипоксической активности соединений общей формулы (I).
Для изучения антигипоксической активности соединений общей формулы (1) был использован тест определения средней продолжительности жизни животных в замкнутом объеме [Методические рекомендации по экспериментальному изучению препаратов, предлагаемых для клинического изучения в качестве антигипоксических средств. // Фармакологический комитет Минздрава СССР. -1990. -с. 10]. Согласно данному тесту мышей сажают по одной в стеклянные банки, которые закрывают герметично и регистрируют продолжительность жизни.
В эксперименте использовано 90 беспородных мышей-самцов с исходной массой 20±0.5 г, содержавшихся по 10 особей в клетке на стандартном рационе вивария. Предварительно была определена средняя продолжительность жизни мышей в замкнутых сосудах объемом 250 мл, которая составляла 22-25 минут. Во всех группах было по 10 животных.
Схема эксперимента:
а) - предварительная гипоксия в течение 12 минут;
б) - первое введение соединения сразу после предварительной гипоксии;
в) - второе введение соединения через 24 часа после первого, затем ежедневно в течение трех дней;
г) - регистрация времени жизни животных в условиях гипоксии в герметическом пространстве через 2 часа после последнего введения соединения.
Исследуемые соединения растворяли в дистиллированной воде и вводили животным перорально в течение 5 дней.
Экспериментальные группы:
1 группа - контрольная - без предварительной гипоксии с пероральным введением в течение 5 дней дистиллированной воды;
2 и 3 группы - введение 1-го соединения по п.п. б), в) и г) с использованием двух доз - 50 и 500 мкг/кг соответственно;
4 группа - введение оксибутирата натрия перорально в дозе 300 мкг/кг за 2 часа до гипоксии. Оксибутират натрия рекомендован в качестве препарата сравнения в Методических рекомендациях /20/;
5 группа - контрольная - с предварительной гипоксией в течение 12 минут с последующим введением в течение 5 дней дистиллированной воды;
6 и 7 группы - введение 1-го соединения по п.п. а), б), в) и г) с использованием двух доз - 50 и 500 мкг/кг соответственно;
8 и 9 группы - введение 2-го соединения по п.п а), б), в) и г) с использованием двух доз - 50 и 500 мкг/кг соответственно.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Данные, представленные в таблице 9, свидетельствуют о том, что соединения общей формулы (I) обладают выраженной антигипоксической активностью, достоверно увеличивая продолжительность жизни экспериментальных животных на 20-50 процентов. Следует отметить, что исследованные соединения повышали продолжительность жизни мышей сопоставимо с действием препарата сравнения - оксибутирата натрия при использовании их в дозах на три-четыре порядка ниже.
Пример 23.
Изменение тяжести пищевой анафилаксии под влиянием одного из соединений общей формулы (I).
Исследования проводили на беспородных морских свинках-самцах с исходной массой 250-300 г, содержавшихся на общевиварном рационе. Сенсибилизацию животных проводили по методу [Шатерников В.А., Марокко И.Н., Пятницкий Н.Н., Ширина Л.И., Горгошидзе Л.Ш., Касьяненко В.В., Сугоняева Н.П., Жминченко В. М. , Внокурова Н.М. / Экспериментальное воспроизведение пищевой анафилаксии. // Вопр. питания. -1982. - N 2. -с. 27-31] куриным овальбумином (OBA), однократно перекристаллизованным, производства Олайненского НПО "БИОХИМРЕАКТИВ" в дозе 50 мг на животное в сутки в течение 3 дней.
Через 14 дней после окончания сенсибилизации у животных вызывали активный анафилактический шок (ААШ) внутривенным введением разрешающей дозы гомологичного белка в дозе 5 мг в 0,5 мл физиологического раствора. Тяжесть ААШ оценивали по уровню летальности, количеству судорожных проявлений и по величине анафилактического индекса [Weigle W. , Cochrane C., Dixon F. / Anaphylactogenic hroherties of soluble antigen-antibody complexes in the guinea pigs and rabbits.// J. Immunology. -1960. -vol. 85. -pp. 469-477].
Одно из исследуемых соединений и кларитин (группа 2 - 30 морских свинок и группа 3 - 24 животных соответственно) вводили животным перорально в течение 3-х дней перед разрешением в дозе 50 мкг/кг и 1000 мкг/кг соответственно. Животные контрольной группы получали в соответствующие сроки физиологический раствор (группа 1 - 30 морских свинок).
Определение достоверности различий между группами проводили с использованием метода углового преобразования Фишера [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Показано (табл. 10) достоверное снижение тяжести проявлений анафилактического шока у морских свинок, получавших одно из исследуемых соединений как по сравнению с контрольной группой, так с группой животных, получавших Кларитин.
Пример 24.
Исследование влияния соединений общей формулы (I) на выраженность бронхоспазма.
Антиастматическое действие исследуемых веществ изучалось на модели антиген-индуцированного бронхоспазма у активно сенсибилизированных морских свинок по методу Andersson [Andersson P. /Antigen induced bronchial anaphylaxis in actively sensitized guinea pig.// Allergy. -1980. -vol. 35. -pp. 65-71] . Данная модель наиболее адекватна атонической бронхиальной астме, так как спазм бронхов у морской свинки развивается в результате аллергической реакции между антигеном и гомоцитотропными антителами lgE класса.
Морские свинки-самцы сенсибилизировались внутримышечной инъекцией 0,5 мл суспензии, содержавшей 20 мкг овальбумина (ОВА, производства фирмы Sigma /grade 111/) и 100 мг Al(ОН)3 на животное. Разрешающую дозу 150-200 мкг/кг ОВА вводили внутривенно (v.jugularis) в 0,1 мл физиологического раствора на 26 день после сенсибилизации. Индукция бронхоспазма и измерение параметров внешнего дыхания проводились по методу, описанному в работе [Yu- Hong L., Barnes P. , Rogers D. / Inhibition of neurogenic plasma exudation and bronchoconstriction by a K+ chennel activator, BRL 38227, in guinea pig airways in vivo.// Europ. J. Pharmacol. -vol. 239. -pp. 257- 259]. Животное наркотизировали внутрибрюшинным введением этаминала натрия (70 мг/кг массы), обнажали трахею, проводили трахеотомию и вставляли в трахею канюлю. Канюля присоединялась через специальный тройник к респиратору (Ugo Basel, модель 7025), который работал в течение эксперимента в постоянном режиме: объем вентилируемого воздуха 8 мл, частота дыхания 70 в минуту. Измерение параметров дыхания осуществлялось с помощью трансдуцера (Ugo Basel, модель 7020), соединенного с канюлей, и самописца (Миллихром), регистрировавшего амплитуду дыхания. Величина амплитуды отражала степень сопротивления гладкой мускулатуры бронхов воздушному потоку. После установления у морской свинки нормального ритма дыхания животному вводили в v. jugularis разрешающую дозу антигена. Через 1-2 минуты развивался бронхоспазм, который выражался в резком увеличении сопротивления бронхов (вследствие их сужения) и амплитуды дыхания в 8-10 раз по сравнению с исходным значением. Динамику бронхоспазма наблюдали в течение 30-60 минут. Эффективность соединений определяли по изменению величины бронхоспазма.
Исследовали ряд соединений общей формулы (I), которые растворяли в физиологическом растворе и вводили животным трехкратно внутрижелудочно за 72, 48 и 18 часов до индукции бронхоспазма в дозе 50 мкг/кг. В качестве препарата сравнения использовали Интал - препарат, получивший широкое распространения при лечении бронхиальной астмы. Интал вводили животным в дозе 5 мг/кг по той же схеме, что и исследуемые вещества. Контрольная группа животных получала эквивалентное количество физиологического раствора.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Все тестированные соединения снижали величину бронхоспазма на 20-70% по сравнению с контрольными значениями. Интал уменьшал величину бронхоспазма почти на 50%.
В таблице 11 представлены результаты, свидетельствующие о том, что два из соединений общей формулы (I) снижали величину бронхоспазма более чем на 50% по сравнению с контрольными значениями.
Таким образом, исследованные соединения проявляли активность в отношении снижения величины бронхоспазма, сравнимую с действием препарата сравнения - Интал. Однако действующая доза исследованных веществ была на два порядка ниже, чем Интала.
Пример 25.
Изучение изменения реакции пассивной кожной анафилаксии под влиянием одного из соединений общей формулы (I).
Эксперименты проводили на нелинейных мышах-самцах с исходной массой 22-24 г. Каждая группа содержала по 10 животных. Сенсибилизацию животных осуществляли внутрикожным введением 20-30 мкл сыворотки мышей, полученной от предварительно иммунизированных животных, содержавшей реагиновые антитела против куриного овальбумина (OBA). Одно из исследуемых веществ вводили животным перорально в течение 3-х дней, начиная со дня сенсибилицации кожи, в дозах 50, 150 и 500 мкг/кг. Аналогично вводили препараты сравнения - Супрастин в дозе 10 мг/кг и Кларитин в дозе 2 мг/кг. Через 48 часов после сенсибилизации мышам внутривенно вводили разрешающую дозу OBA (1 мг) и 1 мг синьки Эванса в 0,2 мл физиологического раствора. Интенсивность реакции пассиной кожной анафилаксии (ПКА) определяли через 30 минут по величине (площади) синего пятна на внутренней поверхности кожи в месте введения реагиновых антител. Величину пятна измеряли в двух взаимно перпендикулярных направлениях и результат выражали в мм2.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.]
Результаты, представленные в таблице 12, свидетельствуют, что введение исследуемого соединения в дозе 50 мкг/кг мышам достоверно снижало выраженность реакции ПКА на 42%, а в дозах 150 и 500 мкг/кг - на 34,3% и 30,0% соответственно. Введение Супрастина экспериментальным животным не изменяло проявлений ПКА. При применении Кларитина наблюдалось выраженное снижение реакции ПКА.
Таким образом, исследуемое соединение обладало способностью снижать проявления ПКА в большей степени, чем Супрастин. Выраженность реакции ПКА при введении животным Кларитина была подавлена в большей степени, чем при введении исследуемого соединения, однако его действующая доза была на порядок выше, чем у изучаемого соединения.
Пример 26.
Изучение влияния одного из соединений общей формулы (I) на аллергическую реакцию замедленного типа.
Исследования проводили на нелинейных белых мышах-самцах с исходной массой 22-24 г. Каждая группа содержала по 10 животных. Мышей сенсибилизировали внутривенной инъекцией взвеси 2•105 эритроцитов барана в 0,05 мл физиологического раствора. На 5 сутки в подушечку задней лапы вводили взвесь 108 эритроцитов барана в 0,05 мл физиологического раствора. В качестве контроля вводили растворитель - физиологический раствор в эквивалентном количестве. Интенсивность реакции оценивали через 24 часа по разности масс лапок животных.
Одно из исследуемых соединений вводили перорально в течение трех дней в дозах 50 и 500 мкг/кг по схеме - 3, 4 и 5 дни, 5-й день - введение разрешающей дозы.
Результаты обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Как видно из данных, представленных в таблице 13, введение животным соединения в дозах 50 и 500 мкг/кг достоверно снижает интенсивность реакции гиперчувствительности замедленного типа.
Пример 27.
Изучение антиоксидантной активности одного из соединений общей формулы (I) в опыте in vivo.
Антиоксидантная активность тестировалась в опыте in vivo на модели острого токсического поражения печени четыреххлористым углеродом (CCl4).
Эксперимент проведен на 40 беспородных крысах-самцах с исходной массой 190-200 г, содержавшихся на стандартном рационе вивария. В каждой группе было по 10 животных. Поражение печени (экспериментальный гепатит) у животных вызывали введением CCl4 внутрижелудочно в виде 50% раствора в вазелиновом масле в объеме 0,25 мл на 100 г массы тела в течение 3 дней [Венгеровский А. И., Чучалин B.C., Паульс О.В., Саратиков А.С. / Влияние гепатопротекторов на метаболизм липидов при CCl4 - гепатите.// Бюлл. экспер. биол. -1987. -N 4. -с. 430-432] . Одно из исследуемых соединений вводили животным внутрижелудочно в дозах 50 и 500 мкг/кг в течение трех суток в дни введения CCl4 (3 и 4 группы). Животным контрольной группы вводили CCl4 как описано выше (2 группа). Интактные животные получали перорально физиологический раствор в эквивалентном количестве (1 группа).
Образцы крови и печени брали на анализ через 18 часов после последнего введения CCl4.
Содержание первичных продуктов перекисного окисления липидов (ПОЛ) - диеновые конъюгаты, диеновые кетоны и триены определяли методом [Волчегорский И. А., Налимов А. Г., Яровинский Б.Г., Лифшиц Р. И. /Сопоставление различных подходов к определению продуктов перекисного окисления липидов в гептан-изопропанольных экстрактах крови.// Вопр. мед. химии. -1989. - N1. -с. 127-131] . Расчет содержания продуктов ПОЛ проводили, соотнося величины соответствующих экстинкций к 1 мл исследуемой пробы.
Количество конечных продуктов ПОЛ - малонового диальдегида (МДА) - определяли по тесту с тиобарбитуровой кислотой (ТБК) [Коробейникова Э.Н. / Модификация определения продуктов перекисного окисления липидов в реакции с тиобарбитуровой кислотой. // Лаб. дело. -1989. -N7. -с. 8-10]. Концентрацию ТБК-активных продуктов рассчитывали с помощью уравнения регрессии. Содержание МДА в печени экспериментальных животных оценивали модифицированным методом [Стальная И.Д., Гаришвили Т.Г. / Метод определения малонового диальдегида с помощью тиобарбитуровой кислоты.// В кн: Современные методы в биохимии. -М. -Медицина. -1977. -с. 66-69], проводя предварительную экстракцию липидов по Фолчу [Кейтс М. Техника липидологии.- М.- Мир.- 1975.- с. 74-76].
Результаты эксперимента обрабатывались статистически с применением t-критерия Стьюдента [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты таблицы 14 свидетельствуют, что введение крысам CCl4 приводило к накоплению продуктов ПОЛ в сыворотке крови и печени. У животных экспериментальных групп, которым вводили одно из исследуемых соединений, отмечалось достоверное снижение содержания первичных продуктов ПОЛ в печени и МДА в сыворотке крови и в печени.
Таким образом, одно из соединений общей формулы (1) обладает выраженным антиоксидантным действием в модели острого токсического поражения печени.
Пример 28.
Изучение гепатопротекторной активности соединений общей формулы (I).
Исследование гепатопротекторных свойств двух из исследованных соединений проводили на модели субхронического поражения печени (гепатита) четыреххлористым углеродом.
В эксперименте использовано 70 беспородных крыс-самок с исходной массой 190-200 г, содержавшихся на стандартном рационе вивария. Животные были разделены на 7 групп (по 10 в каждой):
1 группа - контрольная, животным в течение первых четырех дней эксперимента вводили 0,2 мл вазелинового масла подкожно;
2 группа - животным в течение первых четырех дней вводили подкожно 50%-ный раствор CCl4 в вазелиновом масле из расчета 0,1 мл на 100 г массы тела;
3 группа - на фоне введения CCl4 вводили Легалон в крахмальном геле перорально в течение 8 дней в дозе 30 мг/кг;
4 и 5 группы - аналогично 3 группе применяли 1-е исследуемое соединение в дозах 50 и 500 мкг/кг соответственно;
6 и 7 группы - аналогично 3 группе использовали 2-е исследуемое соединение в дозах 50 и 500 мкг/кг соответственно.
Исследуемые соединения и Легалон животным вводили за 1 час до введения CCl4 Через 24 часа после последнего введения препаратов животных декапитировали.
Гепатопротекторную активность исследованных соединений оценивали по следующим показателям:
1) в сыворотке крови:
- активность аланинаминотрансферазы (АЛТ) и аспартат амино-трансферазы (АСТ);
- холестерин общий (ХС-общий);
- холестерин в составе липопротеинов высокой плотности (ХС-ЛПВП);
- холестерин в составе липопротеинов низкой и очень низкой плотности (ХС-ЛПНП и ХС-ЛПОНП соответственно)
- триглицериды (ТГ);
- малоновый диальдегид (МДА);
2) в печени:
- липидный состав: фосфолипиды (ФЛ), свободный холестерин (СХС), триглицериды (ТГ);
- МДА.
Активность трансаминаз - АЛТ и АСТ в сыворотке крови оценивали общепринятым методом Френкеля-Райтмана [Лабораторные методы исследования в клинике. -Под ред. Меньшикова В.В. -М. -Медицина. -1969. -302 с].
Содержание ХС-общего в сыворотке крови определяли по методу Илька [Биохимические исследования в клинике.- Под. ред. А.А. Покровского. -М. -Медицина. -1969. -с. 300-302]. Количество ХС-ЛПВП измеряли в супернатанте после гепарин-марганцевой преципитации [Титов В.Н., Бренер Е.Д., Халтаев Н.Г., Задоя А.А., Творогова М.Г. / Метод и диагностическая значимость исследования содержания холестерина в α- липопротеидах. // Лаб. дело. -1979. -N1. -с. 36-41] методом Илька [Биохимические исследования в клинике. -Под. ред. А.А. Покровского. -М. -Медицина. -1969. -с. 300-302]. Содержание ХС-ЛПНП и ХС-ЛПОНП рассчитывали по формуле [Friedwald W.T., Levy R.I., Fredrickson D.S. /Estimation of concentration of low-density lipoprotein cholesterol in plasma, without use of preparative ultracentrifuge.// Clin. Chem. -1972. -vol. 18. -pp. 499-502]:
ХС-ЛПНП=ХС-общий-(ХС-ЛПВП+ТГ/5),
где ТГ/5 соответствует содержанию в сыворотке крови ХС-ЛПОНП.
Для оценки количества ТГ в сыворотке крови использовали метод [Родионова Л. П. /Модификация метода определения содержания триглицеридов в сыворотке крови. // Лаб. дело. -1980. -N 5. -с. 297-299].
Содержание конечных продуктов ПОЛ - МДА в сыворотке крови определяли методом [Коробейникова Э. Н. / Модификация определения продуктов перекисного окисления липидов в реакции с тиобарбитуровой кислотой.// Лаб. дело. -1989. -N7. -с. 8-10] . Концентрацию ТБК-активных продуктов рассчитывали с помощью уравнения регресси:
C=0,21+0,26 Д,
где C- концентрация ТБК-активных продуктов (в нмоль МДА на 1 мл сыворотки), Д - показатель Д535-Д580 (в ед. опт. плотности).
Суммарные липиды в печени экстрагировали модифицированным методом Фолча [Кейтс М. Техника липидологии. -М. -Мир. -1975. -с. 74-76]. Количественное содержание липидных фракций оценивали методом тонкослойной хроматографии (ТСХ), используя систему растворителей гексан:диэтиловый эфир:уксусная кислота в соотношении 80:20:2. Зоны индивидуальных липидных фракций определяли с помощью 10%-го спиртового раствора фосфорномолибденовой кислоты, которые после элюирования анализировали спектрофотометрически при 600 нм.
Содержание МДА в печени определяли методом [Стальная И.Д., Гаришвили Т. Г. / Метод определения малонового диальдегида с помощью тиобарбитуровой кислоты. // В кн: Современные методы в биохимии. -М. -Медицина. -1977. -с. 66-69] . Количество МДА рассчитывали, используя величину молярного коэффициента экстинции окрашенного триметинового комплекса, образованного МДА с двумя молекулами ТБК:
E=1,56•105 см-1•М-1.
Результаты эксперимента обрабатывались статитически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты эксперимента свидетельствуют (таблица 15), что у животных, обработанных CCI4, отмечалась выраженная гиперферментемия. Введение соединений общей формулы (I) сопровождалось нормализацией активности АЛТ и ACT в сыворотке крови, причем наибольшая выраженность гепатозащитного эффекта отмечалась при применении второго вещества.
Анализ полученных данных липидного состава сыворотки крови экспериментальных животных показал (таблица 16), что токсическое поражение печени сопровождается гипертриглицеридемией. В распределении холестерина во фракциях липопротеинов при воздействии CCl4 отмечались некоторые особенности - наряду с повышением содержания общего холестерина и ХС-ЛПОНП, возросло количество ХС-ЛПВП и снизилось содержание ХС-ЛПНП. На фоне введения Легалона и исследуемых соединений наблюдались тенденция к снижению содержания общего холестерина, нормализация количества ХС-ЛПНП, ХС-ЛПОНП и ХС-ЛПВП.
Как следует из результатов, представленных в таблице 17, введение животным CCl4 приводило к увеличению содержания в печени ТГ и снижению количества холестерина. Исследуемые соединения нормализовали содержание СХС и снижали количество ТГ. При этом наблюдалось повышение содержания фосфолипидной фракции при применении исследуемых веществ.
В таблице 18 приведены данные изменения конечного продукта ПОЛ - МДА в сыворотке крови и в печени при поражении CCl4 и введении исследуемых соединений. Токсическое поражение печени сопровождалось повышением содержания МДА как в сыворотке, так и в печени. Обработка животных Легалоном и соединениями общей формулы (I) приводила к нормализации количества МДА в исследованных тканях.
Таким образом, исследуемые соединения общей формулы (I) обладали выраженным антиоксидантным и липидрегулирующим действием, сравнимым с эффектом препарата сравнения Легалоном, а по ряду показателей превосходя его. При этом следует отметить, что исследованные соединения применялись в дозах на 2-3 порядка ниже, чем Легалон.
Пример 29.
Гиполипидемическая активность соединений общей формулы (I).
Гиполипидемическую активность соединений общей формулы (I) изучали на модели экспериментальной гиперлипидемии [Arichi Н., Kumura Н.О. / Effects of stibens compounds of roots of polygonum cuspidatium on the lipid metabolism. // Chem. Pharm. Bull. -1982. -vol. 30. -N 5. -pp. 1766-1767] у беспородных крыс-самцов с исходной массой 220-250 г, получавших в течение 10 дней внутрижелудочно на фоне стандартного рациона масляную суспензию, содержавшую 10% холестерина и 1% холевой кислоты (из расчета 1 мл суспензии на 100 г массы тела). Каждая группа животных содержала по 10 крыс. Восемь из исследуемых соединений вводили животным перорально в дозах 50 и 500 мкг/кг в течение последних четырех дней эксперимента. В качестве препарата сравнения использовали никотиновую кислоту, которую вводили животным в течение 10 дней на фоне атерогенной нагрузки в дозе 10 мг/кг. Образцы крови брали на анализ через 18 часов после последнего введения изучаемых веществ, в течение которых у крыс отнимали пищу.
Определяли следующие показатели: холестерин общий (ХС-общий), холестерин липопротеидов высокой плотности (ХС-ЛПВП), холестерин липопротеидов низкой плотности и липопротеидов очень низкой плотности (ХС-ЛПНП и ХС-ЛПОНП), триглицериды (ТГ). Содержание ХС в сыворотке крови определяли методом Илька [Биохимические исследования в клинике. -Под. ред. А.А. Покровского. -М. -Медицина. -1969. -с. 300-302], ХС-ЛПВП оценивали в супернатанте после гепарин-марганцевой преципитации ЛПНП+ЛПОНП [Титов В.Н., Бренер Е.Д., Халтаев Н. Г. , Задоя А.А., Творогова М.Г. / Метод и диагностическая значимость исследования содержания холестерина в α- липопротеидах.// Лаб. дело. -1979. -N 1. -с. 36-41]. Определение ХС-ЛПНП проводили путем расчета по формуле, представленной в работе Friedewald W.T. et al [Friedewald W.T., Levy K.J., Leus R. //Fat transhort in lipoproteins an integrated approach to mechanism and disoders.// New Eugl.J.Med. -1967. -vol. 276. -p. 32].
Для оценки влияния исследуемых соединений на соотношение атерогенных и антиатерогенных липопротеидов плазмы крови вычисляли холестериновый индекс (Kxc) по формуле, приведенной в работе Климова А.Н. с соавторами [Климов А. Н. , Никульчева Н.Г. Липопротеиды, дислипопротеидемии и атеросклероз. -М. -Медицина. -1984. -165 с.].
Содержание ТГ в сыворотке крови определяли общепринятым методом [Friedwald W.T., Levy R.I., Fredrickson D.S. /Estimation of concentration of low-density lipoprotein cholesterol in plasma, without use of preparative ultracentrifuge.// Clin. Chem. -1972. -vol. 18. -pp. 499-502].
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Введение животным жировой суспензии сопровождалось достоверным повышением содержания ХС в сыворотке крови за счет атерогенных фракций липопротеидов (ЛПНП и ЛПОНП) на фоне снижения количества антиатерогенной фракции - ЛПВП. Индекс атерогенности возрос в 3 раза. Отмечалось также значительное повышение ТГ в сыворотке крови (табл. 19). При введении животным изучаемых соединений в обоих дозах отмечалось снижение общего холестерина по сравнению с животными с атерогенной нагрузкой на 10-26% за счет снижения ХС-ЛПНП и ЛПОНП при достоверном повышении ХС-ЛПВП на 20-100%. Выявлено снижение холестеринового индекса атерогенности Kxc при введении всех соединений практически до контрольных значений. Количество ТГ в крови животных уменьшалось до контрольных значений при введении препаратов. В качестве примера действия исследуемых соединений на изменение липидного состава крови в таблице 20 представлены результаты влияния одного из соединений общей формулы (I) на липидный состав крови у животных, получавших атерогенную нагрузку. Показано, что при введении этих соединений наблюдалась нормализация всех исследованных показателей практически до контрольных значений.
Таким образом, изменения липидного состава сыворотки крови при введении экспериментальным животным исследованных веществ сопоставимы с гиполипидемическим эффектом препарата сравнения - никотиновой кислотой (табл. 19).
Пример 30.
Экспериментальное изучение гипогликемической активности соединений общей формулы (I).
Исследования проведены на крысах-самцах Вистар массой 250-300 г. Экспериментальный диабет вызывали однократным внутривенным введением стрептозотоцина (кооп. "Синтез" при ИМБГ АН Украины) в дозе 42 мг/кг крысам, предварительно голодавшим в течение 24 часов с допуском к пище сразу после инъекции. Крыс отбирали в опыт через 2 недели после индукции диабета с уровнем гликемии 120-180 мг%. В каждой группе в опыте использовано по 12 животных.
Вещества вводили интактным животным и крысам со стрептозотоциновым диабетом внутрижелудочно в течение 4 дней в суточных дозах 50 мкг/кг и 500 мкг/кг в водном растворе из расчета 1 мл на 200 г массы тела. Контрольное определение содержания глюкозы в крови проводили в день эксперимента, затем животным вводили исследуемые соединения и лишали пищи. Контрольные животные получали соответствующий объем воды. Эффект оценивали по изменению уровня глюкозы в крови через 2 и 5 часов. Далее животные получали корм и через 24 часа после последнего введения препарата вновь брали кровь для исследования. Кровь брали из хвостовой вены в объеме 0,1 мл. Содержание глюкозы определяли о-толуидиновым методом.
Содержание глюкозы рассчитывали в мг% с помощью стандартных растворов глюкозы. Далее определяли степень изменения содержания глюкозы по отношению к исходному количеству для каждого животного, выражая ее в процентах от исходного содержания. Окончательный расчет проводили для каждой группы животных. Количество животных в каждой группе составляло 10 штук.
Результаты опытов обработаны статистически с использованием t-критерия Стьюдента [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Как следует из результатов таблицы 21, одно из исследованных соединений не влияет на кривую изменений содержания глюкозы в крови интактных животных после кратковременного голодания, тогда как другое исследованное соединение, введенное в дозе 500 мкг/кг, предотвращало снижение количества глюкозы в крови интактных крыс через 2 часа после отмены пищи.
Данные, приведенные в таблице 22, свидетельствуют, что 4-дневное внутрижелудочное введение одного из соединений общей формулы (I) в суточных дозах 50 и 500 мкг/кг приводило к достоверному снижению глюкозы в крови через 2 часа после последнего введения препарата животным. Гипогликемический антидиабетический эффект сохранялся через 5 часов после введения, однако был менее выражен.
Таким образом, соединения, соответствующие общей формуле (I), либо не влияли, либо стабилизировали содержание глюкозы в крови интактных животных и обладали глюкозопонижающей активностью у крыс со стрептозотоциновым диабетом, выраженой в равной степени при применении как в дозе 50 мкг/кг, так и 500 мкг/кг.
Пример 31.
Влияние соединений, соответствующих общей формуле (I), на активность перитонеальных макрофагов.
Изучали изменение активности перитонеальных макрофагов (ПМ) под влиянием трех из исследуемых соединений.
Эксперименты проводили на беспородных белых мышах-самцах с исходной массой 20-22 г, в каждой группе было по 10 животных. Соединения растворяли в физиологическом растворе (ФР). Исследуемые вещества вводили животным перорально в течение 3 дней в дозах 50 и 500 мкг/кг. Животные контрольной группы получали равный объем ФР. Мышей забивали через 20 часов после последнего введения препарата, брюшную полость промывали 1,5 мл раствора Хенкса, содержавшего 5 ед./мл гепарина. 0,1 мл смыва инкубировали в течение 30 мин при 37oC с 0,1 мл 0,2% раствора нитросинего тетразолия (НТС), затем делали мазки. Ядра докрашивали 0,1% раствором нейтральным красным. Результаты выражали в процентах активных клеток [Климов В.В., Коловкина Т.В. /Тест восстановления нитросинего тетразолия, стимулированный пирогеналом.// Лаб. дело. -1982. -N 10. -с. 624-625]. Степень активности ПМ оценивали по количеству включений восстановленного НТС (формазана) в баллах:
0 - нет включений формазана, неактивные ПМ;
1 - единичные включения формазана;
2 - включения заполняют до 1/3 цитоплазмы ПМ;
3 - включения заполняют 1/2 цитоплазмы ПМ;
4 - включения заполняют всю цитоплазму.
Результаты обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты таблицы 23 свидетельствуют, что исследованные соединения увеличивали долю активных макрофагов за счет снижения количества неактивных форм. Причем у одного из исследуемых соединений это действие выражено при исследовании обоих доз вещества, тогда как у другого соединения эффект более выражен при использовании дозы 500 мкг/кг, а у третьего соединения - 50 мкг/кг.
Таким образом, исследованные соединения обладали способностью увеличивать долю активных форм макрофагов, но в разных дозах.
Пример 32.
Изучение влияния соединений, соответствующих общей формуле (I), на развитие пострессорных состояний у мышей.
Исследования проводили на инбредных мышах-самцах линии Balb с исходной массой 16-18 г, в каждой группе использовано по 12 животных. Для развития эмонационально-двигательного стресса использовали модель плавания животных в теплой воде в течение 20 минут. Развитие стресса оценивали по активности естественных киллерных лимфоцитов (НК-клетки) и по функционированию системы интерферона [Сухих Т.Г., Носик Н.Н., Паршина О.В., Ванько Л.В., Меерсон Ф.З. , Ершов Ф. И. / Взаимоотношение системы естественной клеточной цитотоксичности и системы интерферона при иммобилизационном стрессе.// Бюлл. экспер. биол. -1984. -с. 593-595].
Активность НК-клеток определяли в тесте освобождения Н3-уридина из меченых клеток-мишеней YAC-1 (клеточная лимфома мыши, поддерживаемая пассажами in vitro во взвеси в среде RPMI-1640 с 10% фетальной сыворотки). В клеточную взвесь в концентрации 1 млн. клеток/мл вносили H3-уридин (3-5 мкюри/кл./мл) на 60 мин при встряхивании с интервалом в 15 мин с последующим отмыванием средой.
Источником НК-клеток служили спленоциты мышей, которые получали методом [Nossik N. N. , Bopegamage S.A., Yershov F.I. / Properties of interferons produced by different cell populations.// Acta microbiologica Hungarica. -1988. -vol. 35. -Iss 4. -pp. 397-403].
При исследовании интерфероновой реакции спленоцитов мышей клетки селезенки помещали в инкубационную среду (среда RPMI 1640+10% фетальной сыворотки коров), доводили до концентрации 1-3•108 клеток/мл и инкубировали при 37oC в течение 24 часов. В качестве индукторов интерферона был использован митоген ФГА для индукции γ- интерферона [Nossik N.N., Bopegamage S.A., Yershov F.I. /Properties of interferons produced by different cell populations. // Acta microbiologica Hungarica. -1988. -vol. 35. -Iss 4. -pp. 397-403].
Животные были разделены на 4 группы по 36 мышей в каждой.
1 группа - интактные животные, не подвергавшиеся стрессу и получавшие физиологический раствор (ФР).
2 группа - животные, которые были подвергнуты стрессовому воздействию и получавшие ФР.
3 группа - животные, подвергнутые стрессовому воздействию и получавшие соединение 1.
4 группа - животные, подвергнутые стрессу и получавшие соединение 2.
Исследуемые соединения в дозе 50 мкг/кг и ФР вводили животным перорально четырехкратно по следующей схеме: в течение 2-х дней до стрессового воздействия (первое и второе введение), за два часа до стресса (третье введение) и через 24 часа после третьего введения (четвертое введение).
Активность НК-клеток и способность спленоцитов к продукции γ-интерферона определяли через 4 и 24 часов после стресса.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты, представленные в таблице 24, свидетельствуют, что введение исследуемых соединений предотвращало снижение активности НК-клеток, вызванное стрессом. Кроме этого, введение животным исследуемых соединений повышало способность спленоцитов к продукции γ- интерферона (табл. 25).
Таким образом, у животных, которым вводили исследуемые соединения, последствия стрессовой реакции были менее выражены, чем у мышей, их не получавших.
Пример 33.
Исследование влияния соединений, соответствующих общей формуле (I), на резистентность животных к микробным инфекциям.
Изучали влияние двух соединений общей формулы (I) на изменение средней продолжительности жизни экспериментальных животных, инфицированных Salmonella sp.
В эксперименте использованы нелинейные белые мыши обоих полов массой 18-22 г. Соединения общей формулы (I) и препарат сравнения нуклеинат натрия вводили перорально в течение 3 суток (всего 6 раз) до и после заражения (схема -3, -2, -1, 0, 1, 2, 3, где 0 - день заражения). Для инфицирования использовалась суточная культура Salmonella sp., выращенная на агаре Хоттингера. Для приготовления суспензии клеток использовали физиологический раствор. Условия заражения отработаны в предварительных экспериментах. Суспензию клеток Salmonella sp. вводили подкожно в дозе 5•108 кл/мышь в 0,5 мл.
Испытуемые соединения вводили в дозе 500 мкг/кг, а препарат сравнения - нуклеинат натрия - в дозе 50 мг/кг в 0,5мл физиологического раствора.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Оба исследуемых соединения в дозе 500 мкг/кг достоверно увеличивали среднюю продолжительность жизни мышей, инфицированных Salmonella sp. (табл. 26), оказывая защитное действие в жестких условиях эксперимента (100% летальность в контрольной группе). Также обращает внимание тот факт, что наблюдалось достоверное повышение выживаемости мышей, получавших 1-е соединение, на 2 сутки эксперимента по сравнению с контрольной группой, а животных, которым вводили 2-е соединение, на первые сутки. При этом установлено, что характер распределения смертельных исходов животных контрольной группы не является нормальным, тогда как под влиянием 1-го и 2-го соединений оно становится таковым, что подтверждает эффективность исследованных соединений.
Защитный эффект двух исследованных соединений в отношении бактериальной инфекции был сравним с действием нуклеината натрия, известного стимулятора иммунологических реакций, в том числе фагоцитоза [Лазарева Д.Л., Алехин Е.К. Стимуляторы иммунитета. -1985. -286 с.]. При этом эффективная доза исследованных соединений была на два порядка ниже, чем препарата сравнения.
Таким образом, соединения, соответствующие общей формуле (I), обладали выраженным защитным эффектом против микробной инфекции.
Пример 34.
Противовирусная активность соединений, соответствующих общей формуле (I).
А. Защитный эффект одного из соединений общей формулы (I) при инфекции, вызванной вирусом энцефаломиокардита.
Исследования проводили на беспородных мышах обоего пола с исходной массой 10-11 г. В контрольной и опытных группах использовали по 30 животных. Исследуемое соединение и препарат сравнения ридостин - индуктор интерферона вводили однократно внутрибрюшинно через 24 часа после заражения мышей вирусом энцефаломиокардита в дозах 30 мкг/кг и 5,0 мг/кг соответственно. Вирус энцефаломиокардита вводили в дозе 100 ЛД50. Противовирусную активность определяли по изменению средней продолжительности жизни (СПЖ) мышей и степени защиты от смертельной вирусной инфекции.
Результаты эксперимента обрабатывались статистически [Гублер Е.В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Полученные результаты представлены в таблице 27. Показано, что исследованное соединение обладало выраженной противовирусной активностью в дозе на два порядка ниже по сравнению с дозой препарата сравнения.
Б. Влияние соединений общей формулы (I) на выраженность экспериментальной гриппозной инфекции у мышей.
Исследование проводили на белых беспородных мышах обоего пола массой 8-10 г. Вирус гриппа человека, тип A, штамм Aichi (аллантоисная жидкость), вводили мышам интраназально в дозе 100 ЛД50. Было изучено
действие 5 исследуемых соединений, которые вводили животным перорально в течение 3-х дней до заражения и 10 дней после заражения. В качестве препаратов сравнения использовали Арбидол (специфический препарат) в дозе 100 мг/кг, который вводили за 24 и 2 часа до и 3 дня после заражения, и Тимоген (неспецифический препарат) в дозе 10 мкг/кг, вводившийся животным по той же схеме, что и исследуемые соединения. Противовирусную активность определяли по изменению СПЖ мышей и степени защиты от смертельной дозы вирусной инфекции.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты исследования представлены в таблице 28. Показано, что увеличение СПЖ, выживаемости и степень защиты наиболее выражены при введении 1-го соединения в дозе 500 мкг/кг. Применение других исследованных соединений в дозе 50 и 500 мкг/кг и Арбидола сопровождалось изменением показателей в равной степени. Действие Тимогена в качестве противовирусного препарата было выражено более слабо.
Таким образом, представленные данные свидетельствуют о выраженном противовирусном действии соединений общей формулы (I) при экспериментальной вирусной инфекции у мышей, вызванной вирусом гриппа человека, тип A.
В. Влияние соединений общей формулы (I) на репродукцию вируса иммунодефицита человека при острой инфекции лимфобластоидных клеток.
Исследование проводили на культуре лимфобластоидных клеток человека МТ-4. В эксперименте использовали вирус иммунодефицита человека, тип 1, изолят ВИЧ-1/ИВ17 из коллекции Института вирусологии им. Д.И. Ивановского. Изучали действие трех соединений общей формулы (I) на репродукцию вируса иммунодефицита человека в концентрациях 1,0; 0,1 и 0,01 мкг/мл. Препарат сравнения - азидотимидин в дозе 0,05 мкМ/мл. Соединения вносили в культуру клеток МТ-4 за 1 час до внесения вируса в дозе 1000 ТЦИД50. Оценку жизнеспособности клеток МТ-4 проводили на 7 день эксперимента.
Основным параметром эффективности действия исследуемых соединений являлась жизнеспособность клеток.
В предварительных экспериментах было показано, что исследуемые соединения в использованных концентрациях нетоксичны для лимфобластоидных клеток в концентрациях от 1,0 мкг/мл и менее.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты, представленные в таблице 29, показывают, что исследуемые соединения обладали защитным эффектом, выраженным в разной степени, от цитодеструктивного действия ВИЧ-1 в концентрациях 1,0 и 0,1 мкг/мл. Причем влияние 1-го соединения на сохранение жизнеспособности клеток было сопоставимо с действием препарата сравнения - азидотимидина.
Пример 35.
Изучение влияния одного из соединений, соответствующих общей формуле (I), на гемокоагуляцию.
Эксперимент проводили на 30 беспородных белых крысах с исходной массой 200-230 г. Животные были разделены на 3 группы по 10 особей в каждой: 1 группа - контроль - интактный; 2 группа - животные получали перорально в течение 3-х дней соединение в дозе 50 мкг/кг; 3 группа - животным вводили перорально в течение 3-х дней соединение в дозе 500 мкг/кг. Животные контрольной группы получали физиологический раствор в том же объеме. Изучение свертываемости крови проводили через 18-20 часов после последнего введения исследуемого соединения.
Для изучения гемокоагуляции был использован метод электрокоагулографии, разработанный У. А. Ватмахером с соавторами [Колобов Л.Ф. Методы и приборы для клинических лабораторных исследований. -М. -Медицина. -1979. -с. 51-54]. Кровь из подязычной вены крыс помещали в ячейку коагулографа Н-334. Ячейка совершала колебательные движения со скоростью 6 колебаний в минуту. Регистрировали электрическое сопротивление между двумя электродами, расположенными на дне ячейки. Свертывание крови приводило к увеличению сопротивления электрическому току. На основе полученной коагулограммы определяли:
- время T1 (сек) - начало свертывания - 1 и 2 стадия свертывания крови;
- время Т2 (сек) - время образования сгустка - 3 стадия свертывания крови;
- время Т3 (сек) - время свертывания крови - общая длительность свертывания крови.
Результаты эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Данные, представленные в таблице 30, свидетельствуют, что одно из исследованных соединений, вводимое в дозе 50 мкг/кг, не оказывало влияние на показатели свертываемости крови. Введение соединения в 500 мкг/кг приводило к достоверному увеличению времени 1 и 2 фазы свертывания крови и общего времени свертывания крови.
Пример 37.
Влияние соединений, соответствующих общей формуле (I), на метастазирование карциномы легкого Льюиса.
Изучали влияние одиннадцати соединений общей формулы (1) на процесс метастазирования карциномы легких Льюиса (LLC).
Исследование проводили на мышах-самках BDF1 с исходной массой 25-30 г. Каждая группа животных содержала 15 особей. Взвесь опухолевой ткани LLC вводили внутримышечно в объеме 0,3 мл. В качестве растворителя использовали раствор Хенкса. Исследуемые соединения растворяли в воде и вводили животным перорально в течение 24 суток в дозе 50 и 500 мкг/кг в 0,2 мл растворителя, начиная со вторых суток после перевивки опухоли. На 25 сутки эксперимента животных забивали, извлекали легкие и фиксировали их в растворе Буэна с применением микроскопа МБС-9, увеличение 8х2. Через 24 часа после фиксации производили подсчет метастатических колоний в легких. Расчет индекса метастазирования проводили в соответствии с методическими рекомендациями [Методические рекомендации по доклиническому изучению средств, обладающих способностью ингибировать процесс метастазирования и повышать эффективность цитостатической терапии злокачественных опухолей. -М. -1992. -13 с.].
Результаты эксперимента обрабатывались статистически [Гублер Е.В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Выявлено, что протестированные соединения ингибировали процесс спонтанного метастазирования перевиваемой опухоли LLC в присутствии первичного опухолевого узла в разной степени (от 20 до 70%). В таблице 31 представлены результаты, демонстрирующие высокую антиметастатическую активность исследуемых веществ.
Критерием активности для антиметастатических препаратов является их способность ингибировать метастазирование на 35%.
Пример 38.
Влияние одного из соединений, соответствующих общей формуле (I), на выносливость (работоспособность) мышей.
Для изучения влияния соединения общей формулы (I) на выносливость мышей использована модель, применяемая в токсикологии. Мерой выносливости являлось время плавания мышей до их погружения на дно.
Работа выполнена на беспородных белых мышах-самцах массой 20-22 г. Животные получали вещество в течение 3-х суток в дозах 50 и 500 мкг/кг. Длительность плавания регистрировали через 18 часов после последнего введения исследуемого вещества.
Данные эксперимента обрабатывались статистически [Гублер Е. В. Вычислительные методы анализа и распознавание патологических процессов. -М. -Наука. -1978. -365 с.].
Результаты эксперимента приведены в таблице 32. Выявлено, что животные, получавшие соединение в течение 3-х дней, плавали достоверно дольше, чем контрольные животные.
Результаты проведенных экспериментов показывают высокую биологическую активность ряда соединений, соответствующих общей формуле (I).
Как видно из приведенных результатов (пример 16), исследованные соединения индуцируют систему цитохрома Р-450 печени, снижая длительность гексеналового сна. Причем по ряду параметров (пример 17) изменения монооксигеназной системы печени сравнимы с изменениями при введении фенобарбитала: снижение длительности гексеналового сна, увеличение общего содержания цитохрома Р-450 и группы цитохромов Р-450В. Однако изменения системы цитохрома Р-450 печени под влиянием одного из соединений имеет свои особенности - повышается соотношение цитохромов Р-450В/Р-450Л и соотношение цитохрома в5 к цитохрому Р-450. Известно, что увеличение относительного содержания цитохрома в5 связано с повышением использования оксикомплекса цитохрома Р-450 печени, что ускоряет протекание реакций монооксигеназного цикла [Вознесенский А.И., Канаева И. Н. , Корнева Е. Н. / Реконструкция микросомальной монооксигеназной системы печени в гигантских полисомах. 11. Роль цитохрома в5.// Биол. мембраны. -1990. -N 3. -с. 130-134]. Одним из объяснений стимулирующего действия цитохрома в5 на цитохром Р-450 является увеличение доли цитохрома Р-450, содержащего железо в высокоспиновом состоянии [Кобляков В.А. /Цитохромы семейства Р-450 и их роль в активации проканцерогенеза.// ВИНИТИ. Итоги науки и техники. Сер. биол. химия. -М. -1990. -т.35. -192 с.].
Наблюдаемые изменения (пример 18) содержания глюкокортикоидов в крови экспериментальных животных являются, по- видимому, прямым следствием индукции монооксигеназной системы надпочечников, так как при введении исследуемых соединений отмечается не только увеличение содержания конечного продукта цикла синтеза гормона - кортизола, но и его предшественников. Выявлено, что в крови интактных животных при введении им соединений общей формулы (I) увеличивается содержание кортизола в 1,3 раза, оксипрогестерона в 1,4 раза и дезоксикортизола в 1,5 раза.
Эндогенное повышение содержания предшественников и глюкокортикоидных гормонов под влиянием исследуемых соединений может быть использовано при лечении заболеваний, связанных со снижением синтеза этих гормонов в организме. Известно, что экзогенное введение гормональных препаратов приводит, как правило, к системным осложнениям и является фактором риска развития онкологических заболеваний [Parker K.L., Schimmer В.Р. /The Role of Nuclear Receptors in Steroid-Hormones Production. //Sevinars in Cancer Biology. -1994. -Vol. 5. -Iss 5. -pp. 317-325], а также угнетает синтез соответствующих гормонов организмом. В частности, можно прогнозировать положительный эффект использования соединений общей формулы (I) при патологиях, сопровождающихся снижением содержания кортикостероидов: аллергические заболевания, поражения печени различной этиологии, гинекологические заболевания. Выявленное повышение количества оксипрогестерона в крови (пример 18) может иметь важное значение при лечении ряда гинекологических заболеваний, таких как бесплодие, угроза выкидыша, аменорея, дисфункциональные маточные кровотечения и др., так как известно, что при лечении перечисленных патологий используется оксипрогестерон [Машковский М.Д. Лекарственные средства. -М. -Медицина. -1993. -т. 1. -с. 357-361. -с. 691-692].
Известно, что изменение активности системы цитохрома Р-450 сопровождается изменением метаболизма незаменимых полиненасыщенных жирных кислот, в частности арахидоновой кислоты (AK) [Oyekan A.O. / The Suppression by Lipopolysaccharide of Cytochrome P450 - Dependent Renal Vasodilation in the Rat Is Mediated by Nitric- Oxide.// Europ. J. Pharmacol. -1995. -vol. 277. -Iss 2-3. -pp. 123-132], которая является предшественником таких биологически активных соединений, как простагландины (ПГ) и лейкотриены (ЛТ). В наших экспериментах показано, что одно из исследуемых соединений, например, модулирует метаболизм AK подобно фенобарбиталу - известному индуктору монооксигеназной системы. При этом повышается образование ПГ и снижается синтез 12- и 15-гидрокси-6,8,11,14- эйкозатетраеновой кислоты (12- и 15-НЕТЕ). Уменьшение синтеза 12-НЕТЕ, предшественника гепоксилинов, обладающих прометастатическим действием, указывает на перспективность применения соединений общей формулы (I) в качестве противоопухолевых средств [Honn K.V., Tang D.G., Gao X., Butovich I.A., Liu B., Timar J., Hagmann W./12- lipoxygenases and 12(S)-HETE: role in cancer metastasis.// Cancer and Metastasis Reviews. -1994. -vol. 13. -Iss 3-4. -pp. 365-96. -Ref: 195].
Тяжесть клинических проявлений аллергических реакций, как известно, во многом зависит от физиологического состояния клеток-мишеней - базофилов и тучных клеток. Полученные результаты свидетельствуют, что в тучных клетках животных, получавших одно из исследованных соединений, достоверно снижаются содержание гистамина и его антиген-стимулированная секреция и повышается спонтанная секреция гистамина клетками (пример 20), что также может влиять на тяжесть проявлений аллергических реакций.
Общим звеном в патогенезе большого круга патологических состояний (стресс, физическая нагрузка, радиация, аллергические заболевания, атеросклероз и сопутствующие заболевания, поражение печени различной этиологии, сахарный диабет и другие) является гипоксия. Известно, что при гипоксии повышается перекисное окисление липидов (ПОЛ), снижается содержание цитохрома Р-450 и активность Р-450-зависимых ферментов [Proulx М., Dusouich P. /Acute Moderate Hypoxia in Conscious Rabbits - Effect on Hepatic Cytochrome-P450 and on Reactive Oxygen Species.// J. Pharm. Pharmacol. -1995. -vol. 47. -Iss 5. -pp. 392-397; Barakat M., du Souich P./ Effect of nifedipine on the elimination of theophylline in the rabbit subjected to hypoxia or to an inflammatory reaction. // J. Pharm. Pharmacol. -1996. -vol. 48. -pp. 906-910]. При этом повышается активность супероксид-дисмутазы и снижается содержание в печени глутатиона и активность глутатионпероксидазы. Эти изменения приводят к повышению количества реактивных радикалов кислорода, которые могут повреждать мембраносвязанные ферменты, в частности ферменты системы цитохрома Р-450 [Proulx М., Dusouich P. /Acute Moderate Hypoxia in Conscious Rabbits - Effect on Hepatic Cytochrome-P450 and on Reactive Oxygen Species.// J. Pharm. Pharmacol. -1995. -vol. 47. -Iss 5. -pp. 392-397], одной из центральных ферментных систем, поддерживающей гомеостаз внутренней среды организма [Ковалев И. Е., Полевая О.Ю. Биохимические основы иммунитета к низкомолекулярным соединениям. -М. -Медицина. -1985. -303 с.]. ПОЛ - важный физиологический процесс, постоянно протекающий в клеточной мембране и в норме играющий важную роль в жизнедеятельности клеток, например, при биосинтезе простагландинов, лейкотриенов, а также при фагоцитозе [Соколов Е. И. Сахарный диабет и атеросклероз. -М. -Наука. -1996. -405 с.]. Однако на фоне развившейся тканевой гипоксии этот процесс становится плохо управляемым и образуется большое количество свободно-радикальных соединений, которые не успевают нейтрализоваться. Нормализация ПОЛ может служить важным критерием при лечении названных заболеваний.
Другим возможным механизмом подавления активности монооксигеназ при гипоксии может служить преобладание анаэробного пути окислительно-восстановительных процессов в печени, описанных при сенсибилизации различными антигенами [Плешкова С.М. Окислительно-восстановительные процессы при аллергических реакциях немедленного и замедленного типов. -Автореф. докт. дисс. -Киев. -1978. -40 с.; Федоров И. И. О механизмах аллергических реакций.- В кн.: Аллергия в клинике и эксперименте. -Киев. -1968. -с. 5-13]. При использовании ингибиторов клеточного дыхания в экспериментах с изолированными гепатоцитами: цианида для митохондрий и метирапона для цитохрома Р-450 показано, что доля каждой из этих двух основных электрон-транспортных редокс-цепей составляет около 40% [Лукьянова Л.Д., Гулак П.В., Дудченко А.М., Зайцев В.В. , Новиков К.Н., Орлов С.Н., Покудин Н.И., Попова О.А., Уголев А.Т. Гепатоцит: функционально-метаболические свойства. -М. -Наука. -1985. -271 с.]. Сродство цитохрома Р-450 к молекулярному кислороду очень высоко, величина кажущейся KM (O2) составляет 10-6-10-5 М, что лишь незначительно ниже, чем у митохондриальной цитохромоксидазы. При окислении различных ксенобиотиков изолированной печенью и изолированными гепатоцитами величина кажущейся KM(O2) практически не меняется [Iones D., Mason Н./ Gradientes of O2 concentration in hepatocetes. // J. Biol. Chem. -1978. -v. 253. -pp. 4874-4880], что свидетельствует об отсутствии существенного градиента кислорода между внеклеточным пространством и эндоплазматическим ретикулумом. Следовательно, в нормальных условиях кислород, видимо, не лимитирует скорости монооксигеназных реакций. Однако в гипоксических условиях, возникающих при развитии ряда патологических состояний, в частности при клинических и экспериментальных формах сенсибилизации, концентрация кислорода снижается, уменьшается активность цитохром-с(Р-450)-оксидазы [Плешкова С.М. Окислительно-восстановительные процессы при аллергических реакциях немедленного и замедлнного типов. -Автореф. докт. дисс. -Киев. -1978. -40 с.; Федоров И.И. О механизмах аллергических реакций. -В кн.: Аллергия в клинике и эксперименте. -Киев. -1968. -с. 5-13] , превалируют анаэробные процессы метаболизма, развивается ацидоз [Федоров И. И. О механизмах аллергических реакций. -В кн.: Аллергия в клинике и эксперименте. -Киев. -1968. -с. 5-13], что приводит к разбалансировке системы цитохрома Р-450. Развитие ацидоза (накопление в организме ацетона и других кетоновых тел) сопровождается индукцией цитохрома Р-450 2E1 [Freeman J.E., Stirling D., Russell A.L., Wolf C.R. /CDNA Sequence, Deduced Amino-Acid-Sequence, Predicted Gene Structure and Chemical-Regulation of Mouse CYP2E1. // Biochem. J. -1992. -Vol. 281. -Iss FEB. -pp. 689-695; Bestervelt L. L. , Vaz A. D.N., Coon M.J. /Inactivation of Ethanol-Inducible Cytochrome-P450 and Other Microsomal P450 Isozymes by Trans-4-Hydroxy-2-Nonenal, a Major Product of Membrane Lipid-Peroxidation.// Proc. Nat. Acad. Sci. USA. -1995. -Vol. 92. -Iss 9. -pp. 3764-3768], который проявляет выраженную активность в образовании пероксидов липидов [Bestervelt L.L., Vaz A.D.N., Coon M.J. /Inactivation of Ethanol-Inducible Cytochrome-P450 and Other Microsomal P450 Isozymes by Trans-4- Hydroxy-2-Nonenal, a Major Product of Membrane Lipid- Peroxidation.// Proc. Nat. Acad. Sci. USA. -1995. -Vol. 92. -Iss 9. -pp. 3764-3768], что в еще большей степени способствует повышению ПОЛ.
С другой стороны, показано, что при гипоксии, развивающейся при сенсибилизации различными антигенами, происходит снижение активности изоцитратдегидрогеназы и α- кетоглутаратдегидрогеназы [Плешкова С.М. Окислительно-восстановительные процессы при аллергических реакциях немедленного и замедленного типов. - Автореф. докт. дисс. -Киев. -1978. -40 с], которые осуществляют связь обмена цикла Кребса - изоцитрата и α- кетоглутарата через НАДФН-зависимую изоцитратдегидрогеназную реакцию с микросомальной цитохром Р-450-зависимой системой [Weigle К., Sies Н. /Drug oxidation dependent on cytochrome P-450 in isolated hepatocytes.// Europ. J. Biochem. -1977. -vol. 77. -pp. 401-408]. В реакциях, катализируемых изоцитратдегидрогеназой, образуется 65-75% эндогенного НАДФН [Kauffman F., Evans R., Thurman R. / Alterations in nicotinamide and adenine nucleotide systems during mixedfunction oxidation livers from normal and phenobarbital-treated rats.// Biochem. J. -1977. -vol. 166. -pp. 583-592], являющегося донором электронов для системы микросомального окисления [Арчаков А.И. Микросомальное окисление. М. -Наука. -1975. -328 с] и, следовательно, одним из главных лимитирующих факторов монооксигеназной реакции. Наряду с этим, при гипоксии, характеризующейся накоплением НАДН [Плешкова С.М. Окислительно-восстановительные процессы при аллергических реакциях немедленного и замедленного типов. -Автореф. докт. дисс. -Киев. -1978. -40 с], выявлено торможение реакции окисления цитохрома в5, поскольку установлено, что в анаэробных условиях происходит восстановление этого гемопротеина до дитионитного уровня [Карякин А.В., Арчаков А.И. /Межмембранный перенос электронов.// Успехи совр. биологии. -1981. -т. 91. -вып. 1. -с. 74-89]. Как отмечалось выше, повышение относительного содержания цитохрома в5 приводит к улучшению функционирования всего монооксигеназного цикла.
Результаты, полученные в экспериментах, описанных в примере 21, свидетельствуют, что исследуемые соединения обладают выраженным антирадикальным действием в опытах in vitro: ингибируют образование активных форм кислорода - гидроксильного радикала и супероксидного анион-радикала (O2 -). Особо следует подчеркнуть факт снижения образования гидроксильного радикала (.ОН), так как в живом организме нет ферментов, способных обезвреживать эти радикалы. В норме существуют механизмы эффективного удаления перекиси водорода и супероксидного анион-радикала, что делает образование .ОН невозможным [Хочачка П., Сомеро Дж. Биохимическая адаптация. -М. -Мир. -1988. -568 с.]. Однако в стрессовых, патологических ситуациях избыток образования или неполное обезвреживание свободных радикалов приводит к образованию гидроксильного радикала, что в свою очередь увеличивает пероксидацию липидов.
Описанные биохимические изменения под влиянием исследуемых соединений - изменение состояния системы цитохрома Р-450, изменение гормонального фона, антирадикальная активность служат объяснением повышения устойчивости животных к экспериментальной гипоксии, увеличивая среднюю продолжительность жизни животных на 20-50% (пример 22).
Полученные результаты изменения биохимических показателей под влиянием соединений общей формулы (I) достаточно полно объясняют выраженное антиаллергическое действие исследуемых соединений. Так, введение экспериментальным животным одного из представленных соединений достоверно уменьшает интенсивность проявлений пищевой анафилаксии у морских свинок, достоверно снижая процент летальности в 2,9 раза, судорожный синдром - в 3 раза, анафилактический индекс - в 2,57 раза по сравнению с контрольными значениями (пример 23). Тестирование влияния ряда исследуемых соединений на выраженность бронхоспазма выявляет достоверное снижение тяжести бронхоспазма у животных, получавших эти соединения (пример 24). Одним из объяснений уменьшения выраженности проявлений аллергических реакций немедленного типа (ГНТ; пищевая анафилаксия, бронхоспазм) является снижение интенсивности реакции пассивной кожной анафилаксии (ПКА), так как известно, что выраженность ПКА определяется количеством анафилактических антител, которые у морской свинки относят к иммуноглобулинам G (lgG), а у людей - к lgE. В работе [Кржечковская В.В. Состояние системы цитохрома Р-450 печени при развитии экспериментальной пищевой анафилаксии и под влиянием гипоаллергенных алиментарных воздействий. -Автореф. канд. дисс. -М. -1986. -24с.] была выявлена прямая корреляционная зависимость между тяжестью проявлений пищевой анафилаксии и интенсивностью протекания ПКА. Результаты экперимента свидетельствуют (пример 25), что введение экспериментальным животным одного из соединений общей формулы (I) приводит к достоверному снижению выраженности проявлений ПКА. Наряду с уменьшением тяжести проявлений гиперчувствительности немедленного типа (пищевая анафилаксия, бронхиальная астма), показано, что одно из исследованных соединений достоверно снижает интенсивность аллергической реакции замедленного типа (ГЗТ; пример 26).
Описанные изменения тяжести проявлений экспериментальных аллергических реакций хорошо согласуются с литературными данными и могут быть объяснены результатами собственных исследований (примеры 16, 17, 18, 19, 20, 21 и 27).
Как было показано выше, сенсибилизация животных и аллергические заболевания у людей приводят к развитию тканевой гипоксии и соответственно каскаду биохимических изменений, связанных с этим состоянием.
Известно, что у людей, страдающих аллергическими заболеваниями, а также при сенсибилизации и иммунизации животных в эксперименте отмечается снижение функционального состояния монооксигеназной системы печени и снижение содержания цитохрома Р- 450 [Ковалев И.Е., Кошелева Н.А., Хлопушина Т.Г., Лысенкова Е.М., Паутова И.Д./Угнетение цитохром-Р-450-зависимой монооксигеназной системы печени при адъювантном артрите у крыс.// Патол. физиология и эксперим. терапия. -1983. -N 4. -с. 29-31.; Ковалев И.Е., Пирузян Л.А., Шатерников В. А. , Марокко И.Н., Хлопушина Т.Г., Азизов Р.Г. /Участие системы цитохрома Р-450 в аллергических реакциях немедленного типа.// Докл. АН СССР. - 1982. - т. 266.- с. 247-249; Лебедев Д.А., Тихомиров А.В. / Проблемы патогенеза системной красной волчанки и склеродермии.- В кн.: X Европейский конгресс ревматологов. -М. -1983. -тез. N 1000; Ковалев И.Е., Хлопушина Т.Г. , Марокко И.Н., Лысенкова Е.М. / Роль системы цитохрома Р-450 в развитии анафилаксии.// Патол. физиология и эксперим. терапия. -1983. -N 4. -с. 22-25.] . Показано, что развитие экспериментальных аллергических реакций формируется на фоне снижения содержания терминальной оксигеназы и дестабилизированного состояния компонентов монооксигеназной системы печени, оцененного по обновляемости цитохромов в5 и Р-450 [Кржечковская В.В., Мальцев Г.Ю., Марокко И. Н. / Обновление цитохромов Р-450 и в5 в печени при развитии пищевой сенсибилизации.// 4-й Всесоюз. симп. по мед. энзимологии. -1983. -с. 137]. Активация ферментов системы цитохрома Р-450 индукторами типа фенобарбитала приводит к ослаблению тяжести проявлений экспериментальных аллергических реакций [Ковалев И.Е., Полевая О.Ю. Биохимические основы иммунитета к низкомолекулярным соединениям. -М. -Медицина. -1985. -303 с.; Марокко И.Н., Кржечковская В.В., Маликова Н.А., Изотов М.В., Бенедиктова С.А., Спиридонова С. М. / Взаимосвязь тяжести проявлений экспериментальной пищевой анафилаксии и соотношения цитохромов P- 450W и P-450L.// Бюлл. эксп. биол. мед. -1991. -N 8. -с. 200-202.]. В то же время при введении сенсибилизированным животным метирапона, ингибитора системы цитохрома Р-450 и синтеза кортизола, отмечается усиление бронхоспазма [Fornhem С., Kumlin М., Lundberg J.M., Alving К. /Allergen-Induced Late-Phase Airways Obstruction in the Pig - Mediator Release and Eosinophil Recruiment.// Eur. Resp. J. -1995. -vol. 8. -Iss 7. -pp. 1100- 1109; Fornhem C., Lundberg J.M., Alving K./ Allergen-Induced Late-Phase Airways Obstruction in the Pig - The Role of Endogenous Cortisol.// Eur. Resp. J. -1995. -vol. 8. -Iss 6. -pp. 928-937]. Известно, что состояние сенсибилизации у людей и у экспериментальных животных сопровождается снижением содержания глюкокортикоидов в крови и в моче. Нормализация количества гормонов коры надпочечников приводит к улучшению клинической картины заболевания у людей и снижению тяжести проявлений аллергической реакции у животных [Марокко И. Н., Кржечковская В.В., Маликова Н.А. /Взаимосвязь организации микросомальной монооксигеназной системы печени, метаболизм 17-оксикортикостероидов и выраженность экспериментальных аллергических реакций немедленного типа. //Матер. Всес. конф. "Цитохром Р-450 и модификация макромолекул". -Ялта. -1989. -с. 339; Мачарадзе Д.Ш., Марокко И.Н., Балаболкин И.И. , Юхтина Н.В., Маликова Н.А. /Особенности метаболизма глюкокортикоидных гормонов при бронхиальной астме у детей.// Педиатрия. -1994. -N3. - с. 9- 12.], а снижение их количества - к их усилению [Fornhem С., Kumlin М., Lundberg J. M. , Alving К. /Allergen-Induced Late-Phase Airways Obstruction in the Pig - Mediator Release and Eosinophil Recruiment.// Eur. Resp. J. -1995. -vol. 8. -Iss 7. -pp. 1100-1109; Fornhem С., Lundberg J.M., Alving K./ Allergen-Induced Late-Phase Airways Obstruction in the Pig - The Role of Endogenous Cortisol. // Eur. Resp. J. -1995. -vol. 8. -Iss 6. -pp. 928-937.]. В наших исследованиях показано, что введение животным соединений общей формулы (I) приводит к индукции системы цитохрома Р-450 печени (пример 17), повышению содержания кортизола и его предшественников (пример 18) и снижению тяжести проявлений аллергических реакций (примеры 23, 24, 25, 26). Ранее было показано, что снижение длительности гексеналового сна у морских свинок сопровождается повышением количества группы цитохромов Р-450В и относительного содержания неполярных форм 17-оксикортикостероидов. Выявлена обратная корреляционная зависимость между проявлениями экспериментальной анафилаксии и количеством группы цитохромов Р-450В печени [Марокко И.Н., Кржечковская В.В. , Маликова Н. А. /Взаимосвязь организации микросомальной монооксигеназной системы печени, метаболизм 17-оксикортикостероидов и выраженность экспериментальных аллергических реакций немедленного типа. //Матер. Всес. конф. "Цитохром Р-450 и модификация макромолекул. -Ялта. -1989. -с. 339]. Наиболее мощным из известных индукторов системы цитохрома Р-450 печени, который увеличивает содержание группы цитохромов Р-450В и снижает тяжесть проявлений пищевой анафилаксии, является фенобарбитал [Марокко И.Н., Кржечковская В.В., Маликова Н.А., Изотов М.В., Бенедиктова С.А., Спиридонова С.М. / Взаимосвязь тяжести проявлений экспериментальной пищевой анафилаксии и соотношения цитохромов P-450W и P-450L.// Бюлл. эксп. биол. мед. -1991. -N 8. -с. 200-202. ] . Недостатком таких индукторов при лечении является наличие выраженных побочных эффектов (например, сонливость, привыкание) и использование сравнительно высоких доз препаратов. Преимуществом исследованных соединений является их высокая эффективность, достигаемая при существенно более низких эффективных дозах, и отсутствие отрицательных побочных эффектов.
Как показано в примере 19, соединения общей формулы (I) усиливают циклооксигеназный путь метаболизма арахидоновой кислоты (AK), а именно увеличивают синтез ПГЕ2. Известно, что последний обладает бронхолитическим эффектом [Knickle L.C., Bend J.R./ Bioactivation of Arachidonic-Acid by the Guinea-Pig Lung - The Orthologue of Cytochrome-P450-2B4 Is Solely Responsible for Formation of Epoxyeicosatrienoic Acids.// Mol. Pharmacol. -1994. -vol. 45. -Iss 6. -pp. 1273-1280]. При этом такие метаболиты AK, образующиеся в системе цитохрома Р-450, как 5,6-эпоксиэйкозотриеновая кислота (5,6-EET) потенциируют синтез и секрецию ПГЕ2 [Carroll M.A., Balazy М., Margiotta P., Falck J. R. , Mcgiff J.C. / Renal Vasodilator Activity of 5,6-Epoxyeicosatrienoic Acid Depends upon Conversion by Cecljjxygenase and Release of Prostaglandins.// J. Biol. Chem. -1993. -vol. 268. -Iss 17. -pp. 12260-12266; Sakairi Y., Jacobson H.R., Noland T.D., Capdevila J.H., Falck J.R., Breyer M. D. / 5,6-Eet Inhibits Ion-Transport in Collecting Duct by Stimulating Endogenous Prostaglandin Synthesis. // Amer. J. Physiol-Renal.FI. Elect. -1995. -Vol. 37. -Iss 5. -pp. F931-F939.]. По мнению авторов [Федосеева Г.Б. Механизмы обструкции легких. -Санкт-Петербург. -Медицинское информационное агенство. -1995. -334 с. -с. 129-132.] ПГЕ2 ингибирует анафилактогенный выброс гистамина и других медиаторов аллергии и воспаления из тучных клеток, с одной стороны. С другой, гистамин потенциирует выделение ПГЕ2, что рассматривается как один из механизмов обратной связи функционирования тучных клеток. Наблюдаемое в наших исследованиях повышение спонтанной секреции гистамина тучными клетками под влиянием одного из соединений общей формулы (I) (пример 20) может оказывать такое стимулирующее действие на этот процесс, что может являться одной из причин снижения тяжести проявлений экспериментальных аллергических состояний.
Увеличение спонтанной секреции гистамина и снижение антиген-стимулированной секреции и содержания гистамина в тучных клетках (ТК) животных, получавших исследуемые соединения, вероятно, связано с тем, что в ТК содержится фермент трансглутаминаза (ТГлу) [Fesus L,. Szucs Е., Barrett К. et al. / Activation of Transglutaminase and Production of Protein-bound-Glutamylhistamine in Stimulated Mouse Mast Cells.// J. Biol. C. Chem. -1985. -N 25. -pp. 13771-13778. ] . Данный фермент синтезирует белок-связанный гамма-глутамилгистамин, являющийся структурным аналогом соединений общей формулы (I), которые, в свою очередь, могут выступать в роли ингибиторов ТГлу. Показано [Fesus L, . Szucs Е., Barrett К. et al./ Activation of Transglutaminase and Production of Protein- bound-Glutamylhistamine in Stimulated Mouse Mast Cells. / J. Biol. C. Chem. -1985. -N 25. -pp. 13771-13778.], что активность ТГлу повышается как при антигенной, так и антиген-независимой стимуляции тучных клеток. При этом достоверно повышается содержание белок-связанного гамма-глутамилгистамина, что может служить защитным механизмом неконтролируемого выделения гистамина из тучных клеток при введении антигена в сенсибилизированный организм. Введение исследуемых соединений до повторного антигенного воздействия, по-видимому, снижает активность данного фермента, в связи с чем повышается спонтанная секреция гистамина и снижается его количество в клетке. Как говорилось выше, гистамин стимулирует синтез ПГЕ2, а спонтанное выделение гистамина может активировать этот процесс еще в большей степени, с одной стороны. С другой стороны, происходит снижение содержания гистамина в тучных клетках к моменту введения в сенсибилизированный организм антигена. Этот процесс, вероятно, может являться одним из звеньев в цепи взаимодействий биологически активных веществ в организме, приводящей к снижению тяжести проявлений аллергических реакций.
Кроме этого, соединения общей формулы (I) обладают выраженным антиоксидантным действием (примеры 21, 27). Снижение тяжести проявлений анафилаксии, бронхоспазма и ГЗТ под влиянием исследуемых веществ патогенетически может быть связано и с данным их действием, так как известно, что у больных бронхиальной астмой при контакте с аллергеном происходит накопление свободных радикалов и активируется перекисное окисление липидов (ПОЛ). В период ремиссии выявлено значительное снижение ПОЛ [Федосеева Г. Б. Механизмы обструкции легких. -Санкт-Петербург. -Медицинское информационное агентство. -1995. -334 с. -с. 129-132.].
Таким образом, результаты проведенных исследований позволяют рекомендовать соединения общей формулы (I) в качестве средств, обладающих антиаллергической и антиастаматической активностью. Так как известно, что глюкокортикоиды и сосудорасширяющие препараты применяются при лечении кожных проявлений аллергических заболеваний, а также ряда кожных заболеваний, в том числе псориаза, то исследованные соединения могут быть использованы при этих заболеваниях.
Показано, что введение четыреххлористого углерода (CCl4) приводит к характерным изменениям печени с признаками портобилиарного цирроза, повышением перекисного окисления липидов и является общепризнанной моделью поражения печени [Kapil A., Koul I.В., Suri О.Р./ Antihepatotoxic Effects of Chlorogenic Acid from Anthocephalus- Cadamba.// Phytother. Res. -1995. -vol. 9. -lss 3. -pp. 189-193. ] . Метаболизм CCl4 происходит в основном в эндоплазматической сети гепатоцитов: в микросомальной фракции он осуществляется в наибольшей степени [Арчаков А.И., Карузина И.И. / Молекулярные механизмы взаимодействия четыреххлористого углерода с мембранами эндоплазматического ретикулума. // Успехи гепатологии. -Рига. -1973. -вып. 4. -с. 39-59; Poli G., Chiarpeno В., Albano E. /Biochemical evidence for chemical and/or topographic differences in the lipoperoxidative processes induced by CCl4 and iron. // Chem. Biol. Interact. -1983. -vol. 43. -N 3. -pp. 253- 261]. Обладая высокой липофильностью, CCl4 связывается с гидрофобным участком апофермента или с гемовой группой цитохрома Р-450. Показано, что в наибольшей степени активация CCl4 происходит цитохромом Р-448 [Губский Ю. И., Прадий Т.П. /Влияние тетрахлорметана на α-кетоглутаратдегидрогеназную и сукцинатдегидрогеназную активность в митохондриях печени крыс в условиях предварительного введения полициклических углеводородов.// Укр. биохим. журн. -1977. -т. 49. -N 4. -С. 26- 30], изоформой, индуцируемой 3-метилхолантреном (3-МХ), что соответствует в современной номенклатуре цитохромам Р-4501A1 и Р- 4501A2. В работе [Марокко И.Н., Кржечковская В.В., Маликова Н.А., Изотов М.В., Бенедиктова С. А. , Спиридонова С.М. / Взаимосвязь тяжести проявлений экспериментальной пищевой анафилаксии и соотношения цитохромов P-450W и P-450L.// Бюлл. эксп. биол. мед. -1991. -N 8. -с. 200-202.] показано, что введение 3-МХ приводит к повышению содержания группы цитохромов Р-450Л, активный центр которых имеет липофильное микроокружение [Изотов М.В., Щербаков В.М., Девиченский В.М. и др. / Способ определения содержания изоферментов цитохрома Р-450 в микросомах печени.// А.С. N 1488738. -Б.И. -1989. -N 3. -6.06.09, МКИ 01. -N 33/15, N 33/48.]. Полагают, что гем является как местом связывания и активации, так и мишенью при метаболизме CCl4 цитохромом Р-450 [Арчаков А.И. Микросомальное окисление. М. -Наука. -1975. -328 с.]. Биоактивация CCl4 происходит не только в микросомах печени, но также и цитохромом Р-450 микросомальной фракции коры надпочечников [Colby H.D., Purcell Н., Kominami S. , Takemori S., Kossor D.C. /Adrenal Activation of Carbon-Tetrachloride - Role of Microsomal P450.// Toxicology. -1994. -vol. 94. -Iss 1-3. -pp. 31- 40]. При этом наблюдается прямая взаимосвязь между активацией ПОЛ и деградацией цитохрома Р-450 [Colby H.D., Purcell Н., Kominami S., Takemori S., Kossor D.C. /Adrenal Activation of Carbon- Tetrachloride - Role of Microsomal P450. // Toxicology. -1994. -vol. 94. -Iss 1-3. -pp. 31-40; Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в биологических мембранах. - М. - Наука. -1972. -259 с.; Jahn F., Reuter A., Karge E., Danz M., Klinger W. / Age- Dependent Different Influence of Carbon-Tetrachloride on Biotransformation of Xenobiotics, Glutathione Content, Lipid- Peroxidation and Histopathology of Rat-Liver.// Experimental and Toxicologic Pathology. -1993. -vol 45. -Iss 2-3, -pp. 101-107.]. При поражении печени CCl4 подавляются механизмы утилизации супероксидных радикалов и гидроперекисей, что способствует накоплению активных форм кислорода (в том числе, гидроксильных радикалов) и проявлению их токсического действия на мембранные структуры гепатоцитов [Матюшин Б. Н. , Логтнов А. С. , Шустова С.Г., Рослова Е.М. /Энзимная утилизация активных форм кислорода и липидный состав печени при ее токсических поражениях.// VI Симпозиум по биохимии липидов. -Тез. докл. -М. -1995. -с. 64. ] . В результате поражения печени CCl4 происходит изменение фосфолипидного состава микросомальной мембраны, увеличение содержания холестерина и уменьшение соотношения содержания фосфолипидов к холестерину (Ф/ХС). Такие изменении липидного состава микросомальной мембраны приводят к снижению ее текучести, что снижает функциональную активность микросомальных ферментов, в частности системы цитохрома Р-450. Это подтверждается тем, что изменение соотношения Ф/ХС в мембранных структурах клетки достоверно коррелирует с изменением аминопирин-N-деметилазной активности системы цитохрома Р-450 печени [Buters J.T.M., Zysset Т., Reichen J. / Metabolism of Antipyrine in-Vivo in 2 Rat Models of Liver-Cirrhosis - Its Relationship to Intrinsic Clearance in-Vitro and Microsomal Membrane Lipid Composition.// Biochem. Pharmacol. -1993. -vol. 46. -Iss 6. -pp. 983-991; Reichen J., Buters J.T.M., Sojcic Z., Roos F.J./ Abnormal Lipid-Composition of Microsomes from Cirrhotic Rat-Liver-Does It Contribute to Decreased Microsomal Function.// Experientia. -1992. -Vol. 48. -Iss 5. -pp. 482-486]. Действие CCl4 обусловлено прежде всего повреждением эндоплазматического ретикулума, лизосом и других мембранных структур клетки вследствие активации ПОЛ, индуцированного образованием свободнорадикальных метаболитов CCl4 [Венгеровский А.И., Чучалин B. C. , Паульс О.В., Саратиков А.С. / Влияние гепатопротекторов на метаболизм липидов при CCl4 - гепатите.// Бюлл. экспер. биол. -1987. -N 4. -с. 430-432; Долгов А.В., Душкин М.И., Морозов А.В., Морозова И.Ю. /Изменение содержания липидов печени, плазмы крови, аорты и активности холестеролэстераз печени крысы при воздействии тетрахлорметана./ Вопр. мед.химии. -1986. -N 1. -с. 55-58.]
Антиоксидантное и гепатопротекторное действие исследуемых соединений в опыте in vivo изучалось на модели поражения печени при введении животным CCl4 в токсических и субтоксических дозах (примеры 27 и 28). Выявлено (пример 27), что при остром токсическом поражении печени CCl4 введение животным одного из соединений общей формулы (I) приводит к достоверному снижению содержания первичных продуктов перекисного окисления липидов - диеновых конъюгатов, диеновых кетонов и триенов в печени и конечного продукта ПОЛ - МДА в печени и сыворотке крови. На модели субхронического поражения печени CCl4 (пример 28) показано, что введение животным соединений общей формулы (I) предотвращает некроз печеночной ткани, выражающийся в нормализации содержания маркерных ферментов - АЛТ и ACT, количества ХС-ЛПНП, ХС-ЛПОНП и ХС-ЛПВП в сыворотке крови, а также СХС и ТГ в печени. Кроме того, отмечается нормализация содержания конечного продукта ПОЛ - МДА как в сыворотке крови, так и в печени. Причем выраженность действия исследованных соединений сравнима с эффективностью препарата сравнения - Легалона, однако его действующая доза на 2-3 порядка выше, чем у исследованных соединений.
Полученные данные по снижению ПОЛ под влиянием соединений общей формулы (I) хорошо согласуются с увеличением содержания в печени группы цитохромов Р-450В и снижением содержания группы цитохромов Р-450Л (пример 17). По-видимому, активация CCl4 осуществляется в системе цитохрома Р-450 изоформами, активный центр которых имеет липофильное микроокружение [Арчаков А.И., Карузина И.И. / Молекулярные механизмы взаимодействия четыреххлористого углерода с мембранами эндоплазматического ретикулума.// Успехи гепатологии. -Рига. -1973. -вып. 4. -с. 39-59; Губский Ю.И., Прадий Т.П. /Влияние тетрахлорметана на α- кетоглутаратдегидрогеназную и сукцинатдегидрогеназную активность в митохондриях печени крыс в условиях предварительного введения полициклических углеводородов.// Укр. биохим. журн. -1977. -т. 49. -N 4. -с. 26- 30. ]. Исследование системы цитохрома Р-450, использованное в настоящей работе, позволяет определить количественно содержание в микросомальной фракции печени изоформ с активным центром, ориентированным в цитозоль и имеющим гидрофильное окружение (Р-450В), и с активным центром, расположенным внутри микросомальной мембраны с липофильным микроокружением (Р-450Л) [Изотов М.В., Щербаков В. М., Девиченский В.М. и др. / Способ определения содержания изоферментов цитохрома Р-450 в микросомах печени.// А.С. N 1488738. -Б.И. -1989. -N 3. -6.06.09, МКИ 01. -N 33/15, N 33/48.]. Снижение группы цитохромов Р-450Л, с которой, вероятно, связана биоактивация CCl4, сопровождается уменьшением образования биоактивных свободных радикалов CCl4 и соответственно понижением ПОЛ и нормализацией обмена липидов. Другим объяснением нормализации ПОЛ может служить повышение содержания кортизола (пример 18), так как известно, что стероидные гормоны ингибируют этот процесс [Teare J.P. , Greenfield S. M. , Marway J.S., Preedy V.R., Punchard N.A., Peters T.J., Thompson R. P. H. /Effect of Thyroidectomy and Adrenalectomy on Changes in Liver Glutathione and Malonaldehyde Levels After Acute Ethanol Injection.// Free Radical Biology and Medicine. -1993. -vol. 14. -Iss 6. -pp. 655-660.].
Кроме того, результаты экспериментов (пример 21) свидетельствуют, что исследованные соединения обладают антирадикальной активностью в опытах in vitro.
Вероятно, введение исследуемых соединений в организм приводит к функциональной перестройке многих систем, что в итоге и определяет их высокую биологическую активность в качестве антиоксидантов и гепатопротекторов.
Полученные результаты по влиянию соединений, соответствующих общей формуле (I), позволяют рекомендовать их при воздействии на организм таких факторов, как стресс, физическая нагрузка, радиация, профилактика побочных явлений при радиотерапии, для активации процессов репарации; при лечении заболеваний печени, например циррозов и гепатитов различной этиологии; состояний, связанных с интоксикацией веществами, активирующими перекисное окисление липидов, например при алкоголизме; в качестве профилактических препаратов на производствах, связанных с получением химических препаратов, и при контактах с токсическими веществами.
Известно, что процессы старения также связаны с окислительным стрессом, сопровождающимся повышением ПОЛ и, как следствие этого, образованием МДА, который, взаимодействуя с лизином белков, образует липофусциноподобный пигмент. Это, вероятно, ускоряет липофусциноз, что вызывает необратимые морфологические сдвиги. Снижение содержания МДА (примеры 27, 28) уменьшает вероятность образования данного соединения, что может приводить к понижению вероятности развития возрастных заболеваний. Кроме того, подтверждением возможности применения исследуемых соединений при возрастных заболеваниях могут служить результаты, полученные в работе Эмануэля с соавторами [Эмануэль Н.М. , Обухова Л. К., Найдич В.И., Мурза Л.И., Бунто Т.В. / Замедление старения посредством активации системы микросомальных оксидаз фенобарбиталом.// Докл. АН СССР. -1977. -т. 235. -N 4. -с. 957- 960], по увеличению средней продолжительности жизни экспериментальных животных при использовании малых доз фенобарбитала. В примерах 16 и 17 показано, что соединения общей формулы (I) вызывают схожие с ФБ изменения системы цитохрома Р-450 печени, и результатом их влияния на организм может быть снижение скорости старения. Комплекс геронтологических заболеваний включает такие патологические состояния, как старческие психозы, катаракта, старческие изменения кожных покровов. Окислительный стресс в нервной ткани (увеличение ПОЛ) является одним из звеньев патогенеза ряда заболеваний нервной системы, в частности болезни Паркинсона. На основании полученных данных можно предложить применение исследованных веществ при лечении болезни Паркинсона, катаракты, старческих изменений кожных покровов.
Нарушения липидного обмена, характеризующиеся повышением содержания триглицеридов, общего холестерина (ХС), холестерина в составе липопротеинов низкой и очень низкой плотности (ЛПНП и ЛПОНП) и снижением содержания холестерина в составе липопротеинов высокой плотности, приводят к развитию таких широко распространенных заболеваний, как атеросклероз и ожирение, и как следствие - ишемической болезни сердца и головного мозга, инфаркта миокарда, инсульта, а также служат фактором риска манифестации сахарного диабета. Следует также отметить, что причиной нарушения обмена холестерина может быть изменение структуры ЛПНП и ЛПОНП, а также активности клеточных рецепторов. Выявлено, что у больных атеросклерозом и сахарным диабетом в ЛПНП повышено содержание триглицеридов, в ЛПОНП - холестерина. С другой стороны, показано, что у лиц с повышенным содержанием глюкозы в крови активно происходит процесс гликозилирования ЛПНП, а при повышении продуктов ПОЛ, в частности МДА, образуются перекисно- модифицированные липопротеины, в первую очередь - ЛПНП. Последние в силу своей цитотоксичности могут повреждать эндотелиальный покров сосудов. Выявлена положительная корреляция между содержанием продуктов ПОЛ в ЛПНП и площадью поражения коронарных сосудов. Авторы считают, что именно модифицированные ЛПНП обладают высокой атерогенностью [Климов А.Н., Никульчева Н. Г. Липопротеиды, дислипопротеидемии и атеросклероз. -М. -Медицина. -1984. -165 с.].
Показано, что изменение содержания и соотношения липидов в плазме отражает их изменения в мембранных структурах паренхиматозных органов. Состав мембран клетки, например микросомальных, прямо зависит от состава рациона экспериментальных животных [Wade A., Harred W. / Effect of dietary lipid on drug-metabolising. // Feder. Proct. -1976. -vol. 55. -pp. 2475-2479]. Введение животным холестерина вызывает накопление его в мембранах клеток, уменьшая ее текучесть, что, в свою очередь, приводит к изменению функционального состояния ферментов [Buters J.T.M., Zysset Т., Reichen J. / Metabolism of Antipyrine in-Vivo in 2 Rat Models of Liver-Cirrhosis - Its Relationship to Intrinsic Clearance in-Vitro and Microsomal Membrane Lipid Composition.// Biochem.Pharmacol. -1993. -vol. 46. -Iss 6. -pp. 983-991; Reichen J., Buters J.T.M., Sojcic Z., Roos F.J./ Abnormal Lipid- Composition of Microsomes from Cirrhotic Rat-Liver - Does It Contribute to Decreased Microsomal Function.// Experientia. -1992. -Vol. 48. -Iss 5. -pp. 482-486; Татонь Я.Н. Ожирение патофизиология, диагностика, лечение. - Варшава.- Польское Медицинское издательство. -1981. -364 с.]
Моделирование нарушений обмена липидов, как правило, связано с добавлением в рацион животных тех или иных жировых композиций или повышенного количества холестерина. Использованная в наших исследованиях модель гиперхолестеринемии вызывает значительные изменения количества и соотношения липидов в крови: повышение триглицеридов (ТГ), общего холестерина (ХС), холестерина липопротеинов низкой и очень низкой плотности (ЛПНП и ЛПОНП - "атерогенные"липопротеины), холестеринового индекса (КXC) и снижение количества липопротеинов высокой плотности (ЛПВП - "антиатерогенные"липопротеины). Эти изменения липидного состава крови сходны с изменениями у больных атеросклерозом, ожирением и сахарным диабетом [Wade A., Harred W. / Effect of dietary lipid on drug-metabolising.// Feder. Proct. -1976. -vol. 55. -pp. 2475- 2479; Татонь Я.Н.
Ожирение патофизиология, диагностика, лечение. - Варшава.- Польское Медицинское издательство. -1981. -364 с.; Балаболкин М.И. Сахарный диабет.- М.- Медицина. -1994. -384 с.].
При изучении эффективности действия соединений общей формулы (I) на показатели липидного обмена отмечается (пример 29) достоверное снижение содержания ТГ, общего XC, ЛПНП, ЛПОНП и KXC и увеличение ЛПВП. Особо следует подчеркнуть важность того факта, что под влиянием исследуемых веществ происходит достоверное увеличение ЛПВП, так как известно, что данная фракция липопротеидов оказывает защитное действие на вероятность развития атеросклероза и ишемической болезни сердца. Протективное действие ЛПВП связывают с тем, они участвуют в транспорте XC из тканей в печень, снижая его накопление в стенке сосудов [Соколов Е. И. Сахарный диабет и атеросклероз. -М. -Наука. -1996. -405 с.], а также задерживают образование перекисно-модифицированных ЛПНП [Климов А.Н., Никульчева Н.Г. Липопротеиды, дислипопротеидемии и атеросклероз. -М. -Медицина. -1984. -165 с.]. Кроме того, ЛПВП предотвращают образование тромбов на поверхности атеросклеротических бляшек. ЛПВП могут также блокировать рецепторы ЛПНП, после чего снижается накопление XC в клетках. В эксперименте показано, что даже при повышенном содержании XC в сыворотке крови наличие высокого количества ЛПВП предотвращает образование атеросклеротических бляшек в сосудах [Соколов Е. И. Сахарный диабет и атеросклероз. -М. -Наука. -1996. -405 с.].
Следует особо выделить снижение содержания ТГ в сыворотке крови при введении изучаемых соединений, так как известно, что ТГ могут являться независимым фактором риска развития ишемической болезни, оказывая токсическое действие на эндотелий сосудов, приводящее к повышенной агрегации тромбоцитов и снижению фибринолитической активности крови [Соколов Е. И. Сахарный диабет и атеросклероз. -М. -Наука. -1996. -405 с.].
Липидрегулирующее действие соединений общей формулы (I) имеет неоднозначное объяснение. Как было показано ранее (примеры 16 и 17), изучаемые соединения обладают выраженным потенциирующим действием на систему цитохрома Р-450 печени, а в примере 18 представлены результаты влияния исследованных соединений на гормональный статус, где было выявлено достоверное повышение содержания кортизола в крови экспериментальных животных. Известно, что система цитохрома Р-450 печени принимает активное участие в метаболизме липидов, а модуляция ее функционального состояния может приводить к качественному изменению обмена липидов. Повышение количества кортизола может, вероятно, оказывать определенное положительное влияние на содержание и соотношение липопротеинов, так как у людей с повышенной массой тела (ожирением) выявлено снижение содержания данного гормона в крови. Кроме того, чувствительность тканей к действию кортизола у этих людей понижена [Соколов Е. И. Сахарный диабет и атеросклероз. -М. -Наука. -1996. -405 с.]. Эти изменения биохимических показателей могут прямо влиять на наблюдаемый в эксперименте (пример 29) эффект нормализации липидного обмена при атерогенной нагрузке. Также большое значение имеет тот факт, что исследуемые соединения обладают антиоксидантным действием (примеры 21, 27, 28), так как нарушение липидного обмена, как правило, сопровождается повышением ПОЛ.
Полученные результаты свидетельствуют, что соединения общей формулы (I) могут быть использованы при лечении атеросклероза и сопутствующих заболеваний (ишемическая болезнь сердца и головного мозга, инфаркт миокарда, инсульт), ожирения и сахарного диабета, при профилактике тромбообразования.
В ходе дальнейших исследований было показано, что исследованные соединения достоверно снижают содержание глюкозы в крови у животных с экспериментальным стрептозотоциновым диабетом, не снижая или стабилизируя данный показатель у интактных животных (пример 30). По-видимому, уменьшение количества глюкозы в крови животных с экспериментальным стрептозотоциновым диабетом связано с нормализацией липидного обмена и снижением перекисного окисления липидов у этих животных, в частности снижением образования гидроксильного радикала, играющего важную роль в формировании патологического процесса [Ohkuwa Т. , Sato Y., Naoi-M. /Hydroxyl Radical Formation in Diabetic Rats Induced by Streptozotocin// Life Sci. -1995. -Vol 56. -Iss 21. -pp. 1789-1798] . Результатом данного действия соединений, вероятно, является разрыв порочного круга, наблюдаемого при сахарном диабете, когда изменение метаболизма углеводов влечет нарушение обмена липидов и наоборот, так как известна тесная взаимосвязь этих двух сторон обмена веществ в организме [Соколов Е. И. Сахарный диабет и атеросклероз. -М. -Наука. -1996. -405 с.]. Кроме этого, соединения общей формулы (I) изменяют функциональное состояние системы цитохрома Р-450, разбалансировка которой наблюдается при сахарном диабете [Shimojo N., Ishizaki Т., Imaoka S., Funae Y., Fujii S., Okuda K. / Changes in Amounts of Cytochrome-P450 Isozymes and Levels of Catalytic Activities in Hepatic and Renal Microsomes of Rats with Streptozocin-Indiced Diabets.// Biochem.Pharmacol. -1993. -vol. 46. -Iss 4. -pp. 621-627], что в свою очередь приводит к нарушению гормонального статуса у этих больных. Нормализация гормонального статуса, наблюдаемая у сенсибилизированных животных при введении им исследуемых соединений (пример 18), приводит к достоверному снижению тяжести бронхоспазма у животных (пример 24). По аналогии с этими результатами можно предположить, что у животных с экспериментальным сахарным диабетом также происходит гармонизация гормонального статуса, что может являться одним из факторов, приводящим к нормализации содержания глюкозы в крови.
Таким образом, данные результаты подтверждают, что соединения, соответствующие общей формуле (I), стабилизируют гомеостаз организма, что дает основание рекомендовать их для лечения сахарного диабета.
Важным звеном в поддержании гомеостаза внутренней среды организма являются фагоциты. Фагоцитирующие клетки представлены в организме в большом количестве. К ним, в частности, относятся нейтрофилы периферической крови и макрофаги (МФ). Активация МФ является одним из адаптивных механизмов гомеостаза, способствующих элиминации патогенов, и противоопухолевой резистентности [Иммунология. -Под ред. Пола У. -М. -Мир. -1989. -т. 3. -с. 202- 228; Пальцев М.А., Иванов А.А. Межклеточные взаимодействия. -М. -Медицина. -1995. -224 с.]. Известно, что эти клетки несут на внешней мембране глюкокортикоидные рецепторы [Маянский А.Н., Пикуза О.И. Клинические аспекты фагоцитоза. Казань. -"МАГАРИФ". -1993. -192 с.]. Одним из механизмов активации нейтрофилов и МФ является повышение содержания глюкокортикоидных гормонов.
Полученные результаты по изучению изменения количества активных перитонеальных МФ свидетельствуют об увеличении количества активных клеток (пример 31). Также выявлена нормализация активности НК-клеток, выделенных из селезенки, и содержания γ-интерферона при введении животным исследованных соединений (пример 32), которые были снижены стрессовым воздействием.
Объяснением повышения резистентности животных к микробным и вирусным инфекциям (примеры 33, 34) служит повышение количества активных перитонеальных макрофагов у животных, получавших соединения общей формулы (I), и активных НК-клеток при стрессовом воздействии, принимающих активное участие в поддержании иммунного статуса организма [Маянский А.Н., Пикуза О.И. Клинические аспекты фагоцитоза. Казань. -"МАГАРИФ". -1993. -192 с.], а также повышение содержания γ- интерферона, который ингибирует in vitro пролиферацию клеток, трансформированных вирусом [Кетлинский С.А., Симбирцев А.С., Воробьев А.А. Эндогенные иммуномоделяторы. -1992. -Санкт-Петербург. -Гиппократ. -256 с]. Особо следует подчеркнуть защитный эффект веществ от цитодеструктивного действия вируса иммунодефицита человека. Противовирусное действие соединений также может быть связано с нормализацией перекисного окисления липидов, так как известно, что повышение содержания активных форм кислорода вследствие индукции цитохрома Р-4501A1, который участвует в их образовании, приводит к повышению титра ВИЧ-вируса в экспериментальной системе [Yao Y., Hoffer A., Chang C.Y., Puga A. /Dioxin Activates HIV-1 Gene-Expression by an Oxidative Stress Pathway Requiring a Functional Cytochrome-P450 Cyp1A1 Enzyme. // Environ. Health Perspect. -1995. -Vol. 103. -Iss 4. -pp. 366- 371.].
Таким образом, исследованные соединения можно рекомендовать при развитии бактериальных и вирусных инфекций и, что особо следует подчеркнуть, при лечении ВИЧ-инфекции.
Известно, что активные формы ФГ способны продуцировать большое количество биологически активных факторов (монокинов), в частности фактора некроза опухоли, с которым связано повышение противоопухолевой резистентности организма [Маянский А.Н., Пикуза О.И. Клинические аспекты фагоцитоза. Казань. -"МАГАРИФ". -1993. -192 с.].
Известно, что процесс образования и развития метастазов во многом определяется состоянием внутрисосудистой свертываемости крови, а в результате развития опухолевого заболевания и ряда сопутствующих факторов активируется свертывающая система крови. Показано, что применение антикоагулянтов способно восстановить разбалансированность свертывающей и антисвертыващей систем крови [Буров Ю.В., Сыркин А.Б., Кинзирский А.С., Королева А.М /Фармакологические аспекты профилактики и лечения метастазов злокачественных опухолей. // Химико-фармацевтическое производство. -Обзорная информация. -вып. 11. -42 с].
Результаты собственных экспериментов показывают, что соединения общей формулы (I) увеличивают время свертывания крови, то есть обладают определенной антикоагулянтной активностью (пример 35).
Известно, что фактор свертывания крови XIIIa является ферментом трансглутаминазой [Yee V.C., Pedersen L.C., Letrong I., Bishop P.D., Stenkamp R.E. , Teller D.C. / 3-Dimensional Structure of a Transglutaminase - Human Blood-Coagulation Factor-XIII.// Proceedings of the National Academy of Sciences of the United States of America. -1994. -vol. 91. -Iss 15. -pp. 7296-7300] и принимает участие в стабилизации фибрина [Greenberg C.S., Birckbichler P.J., Rice R.H. / Transglutaminases - Multifunctional Cross-Linking Enzymes That Stabilize Tissues. // Faseb J. -1991. -vol. 5. -Iss 15. -pp. 3071-3077.], одного из основных составляющих свертывающей системы, играющего большую роль в тромбообразовании. Соединения общей формулы (I) по химической структуре аналогичны метаболитам трансглутаминазы и вероятно способны оказывать ингибирующее действие на этот фермент, что может служить одним из объяснений их антикоагулянтной активности.
Данные, представленные в примере 36, свидетельствуют, что исследованные соединения достоверно ингибируют процесс метастазирования. Этот эффект может быть связан с рядом биологических действий соединений общей формулы (I): с изменением состояния системы цитохрома Р-450, изменением гормонального статуса, с их антиоксидантными и антирадикальными свойствами, с увеличением времени свертываемости крови, так как известно, что все перечисленные системы патологически изменены при развитии онкологических заболеваний. Следует особо подчеркнуть способность исследованных соединений повышать количество перитонеальных активных макрофагов, НК-клеток и γ-интерферона, принимающих активное участие в противоопухолевом иммунитете [Маянский А.Н., Пикуза О.И. Клинические аспекты фагоцитоза. Казань. -"МАГАРИФ". -1993. -192 с.; Кетлинский С.А., Симбирцев А.С., Воробьев А.А. Эндогенные иммуномоделяторы. -1992. -Санкт-Петербург. -Гиппократ. -256 с]. Еще одним объяснением, отмеченным выше, антиметастатического действия исследованных соединений может также быть снижение образования 12-НЕТЕ, так как известно, что этот метаболит арахидоновой кислоты потенциирует процесс метастазирования [Honn K.V., Tang D. G. , Gao X., Butovich I.A., Liu B., Timar J., Hagmann W./12-lipoxygenases and 12(S)-HETE: role in cancer metastasis.// Cancer and Metastasis Reviews. -1994. -vol. 13. -Iss 3-4. -pp. 365-96. -Ref: 195]. Кроме этого, вероятно, определенную роль в ингибировании метастазирования опухолей играет также снижение активности трансглутаминазы под влиянием исследуемых соединений, так как известно, что ее активность частично коррелирует с развитом процесса метастазирования [Elsasser Н.Р., Macdonald R., Dienst M., Kern H.F. / Characterization of a Transglutaminase Expressed in Human Pancreatic Adenocarcinoma Cells./ Europ. J. Cell Biol. -1993. -vol. 61. -Iss 2. -pp. 321- 328] , что может быть связано с тем, что соединения общей формулы (I) являются структурными аналогами метаболитов данного фермента. Факт снижения количества метастазов имеет большое практическое значение, поскольку скорость течения этого процесса исключительно важна и ее торможение приводит к более благоприятному прогнозу при канцерогенезе.
На основании полученных данных можно рекомендовать исследованные соединения при лечении опухолевых заболеваний.
Повышение выносливости (работоспособности) животных (пример 37) - увеличение длительности плавания мышей после применения одного из соединений общей формулы (I) в течение 3-х дней может быть связано со снижением активности перекисного окисления липидов. Это может служить показанием для приема исследованных соединений людям, связанным с большими физическими нагрузками, например на производствах и спортсменам. Наличие этого феномена также очень важно и для больных сахарным диабетом, атеросклерозом, так как у этих людей снижена адаптация к физической нагрузке.
Широкий спектр действия соединений, соответствующих общей формуле (I), может быть отчасти объяснен тем, что эти вещества действуют на центральные системы организма, стабилизирующие гомеостаз, например систему цитохрома Р-450, ПОЛ и систему фагоцитоза. Известно, что в живом организме изменение активности одной системы влечет за собой изменения в других и эти изменения принимают каскадный характер. При введении веществ общей формулы (I) в организм наступает нормализация многих его функций, что позволяет отнести их к адаптогенам.
В заключение следует отметить, что исследованные соединения проявляют биологическую активность в дозах на два-три порядка ниже по сравнению с препаратами сравнения при практически одинаковой эффективности. При этом при исследовании токсичности не найдена среднесмертельная доза, так как даже при введении животным, например, одного из исследуемых соединений в дозе 5000 мкг/кг перорально и 10000 мкг/кг парентерально не вызывает смертельного исхода у экспериментальных животных.
Ряд соединений, соответствующих общей формуле (I), можно вводить внутримышечно, внутривенно, интраназально, перорально, сублингвально, интраректально и применять в виде ингаляций. Соответственно они могут применяться в различных лекарственных формах: в виде инъекционных растворов, таблеток, свечей, мазей. При использовании исследованных соединений в виде инъекционных растворов содержание действующего начала в них составляет 0,01-0,03% вес. (0,03-0,50 мг/кг массы тела). В качестве разбавителя вещества могут содержать 0,9% раствор натрия хлорида, дистиллированную воду, раствор новокаина для инъекций, раствор Рингера, раствор глюкозы. Соединения общей формулы (I) в виде таблеток и свечей содержат действующее вещество в количестве 3,0-30,0 мг на одну таблетку или свечу. Для таблеток и свечей в качестве фармацевтического наполнителя используют любую фармацевтически пригодную основу. Соединения в любой форме применяют в течение 5-10 дней по 1 разу в день. Для ряда соединений, соответствующих общей формуле (I), не выявлено отрицательных побочных действий и не обнаружено противопоказаний к применению. Готовят лекарственные формы ряда соединений общей формулы (I) общеизвестным методом.
На основании результатов проведенных исследований предлагается использование соединений общей формулы (I) для лечения и профилактики следующих состояний:
1. стрессовые состояния;
2. аллергические заболевания - бронхиальная астма, пищевая аллергия;
3. атеросклероз, ишемическая болезнь (сердца, головного мозга), ожирение;
4. сахарный диабет 1 и 2 типа;
5. алкоголизм, алкогольная интоксикация;
6. профилактика тромбообразования;
7. гепатит, цирроз печени;
8. токсические поражения печени;
9. заболевания нервной системы, болезнь Паркинсона;
10. геронтологические заболевания: катаракта, старческое изменение кожных покровов;
11. радиационное поражение, профилактика радиотерапии;
12. вирусные и бактериальные инфекции, в частности ВИЧ-инфекция;
13. онкологические заболевания;
14. тяжелая физическая нагрузка.
Список сокращений.
ААШ - активный анафилактический шок
АК - арахидоновая кислота
АЛТ - аланинаминотрансфераза
АН - активированные нейтрофилы
АсК - аскорбиновая кислота
ACT - аспартатаминотрансфераза
АФК - активные формы кислорода
ВЭЖХ - высокоэффективная жидкостная хроматография
ГЗТ - гиперчувствительность замедленного типа
ГНТ - гиперчувствительность немедленного типа
ГС - гексеналовый сон
ЛД - летальная доза
ЛК - линолевая кислота
ЛО - липоксигеназный путь
ЛПВП - липопротеины высокой плотности
ЛПНП - липопротеины низкой плотности
ЛПОНП - липопротеины очень низкой плотности
ЛТ - лейкотриены
МДА - малоновый диальдегид
МФ - макрофаги
НК - нейтральный красный
НК-клетки - натуральные киллеры
НТС - нитросиний тетразолий (формазан)
ОВА - куриный овальбумин
ПГ - простагландины
ПКА - пассивная кожная анафилаксия
ПМ - перитонеальные макрофаги
ПОЛ - перекисное окисление липидов
СПЖ - средняя продолжительность жизни
СХС - свободный холестерин
ТБК - тиобарбитуровая кислота
ТГ - триглицериды
ТГлу - трансглутаминаза
ТК - тучные клетки
ТМС - тетраметилсилан
ТХУ - трихлоруксусная кислота
ТСХ - тонкослойная хроматография
ТЦИД - тканевая цитопатическая доза
Ф - фосфолипиды
ФБ - фенобарбитал
ФГ - фагоциты
ФР - физиологический раствор
ХЛ - хемилюминесценция
ХС - холестерин
ЦНС - центральная нервная система
ЦО - циклооксигеназный путь
ЭДТА - этилендиамид-тетраацетат
ЯМР - ядерный магнитный резонанс
ButOH - трет-бутанол
But - трет-бутил
Bzl - бензил
Boc - трет-бутилоксикарбонил
DCC - N,N'-дициклогексилкарбодиимид
DCU - N,N'-дициклогексилмочевина
DMF - N,N'-диметилформамид
Et3N - триэтиламин
EtOAc - этилацетат
EtOH - этанол
EET - эпоксиэйкозотриеновая кислота
GA - глутаровая кислота
Glu - глутаминовая кислота
F - кортизол
HA - гистамин
HETE - (S)-гидрокси-6,8,11,14-эйкозатетраеновая кислота
5-HT - 5-гидрокситриптамин (серотонин)
Ind - индол
Im - имидазол
MeCN - ацетонитрил
MeOH - метанол
OBut - трет-бутиловый эфир
ONsu - N-оксисукцинил
OPfp - пентафторфенил
Py - пиридин
Р-450В - группа цитохромов Р-450 с активным центром, ориентированным в цитозоль
Р-450Л - группа цитохромов Р-450 с активным центром, ориентированным в фосфолипидную мембрану
TrpA - триптамин
TFA - трифторуксусная кислота
Z - бензилоксикарбонилл
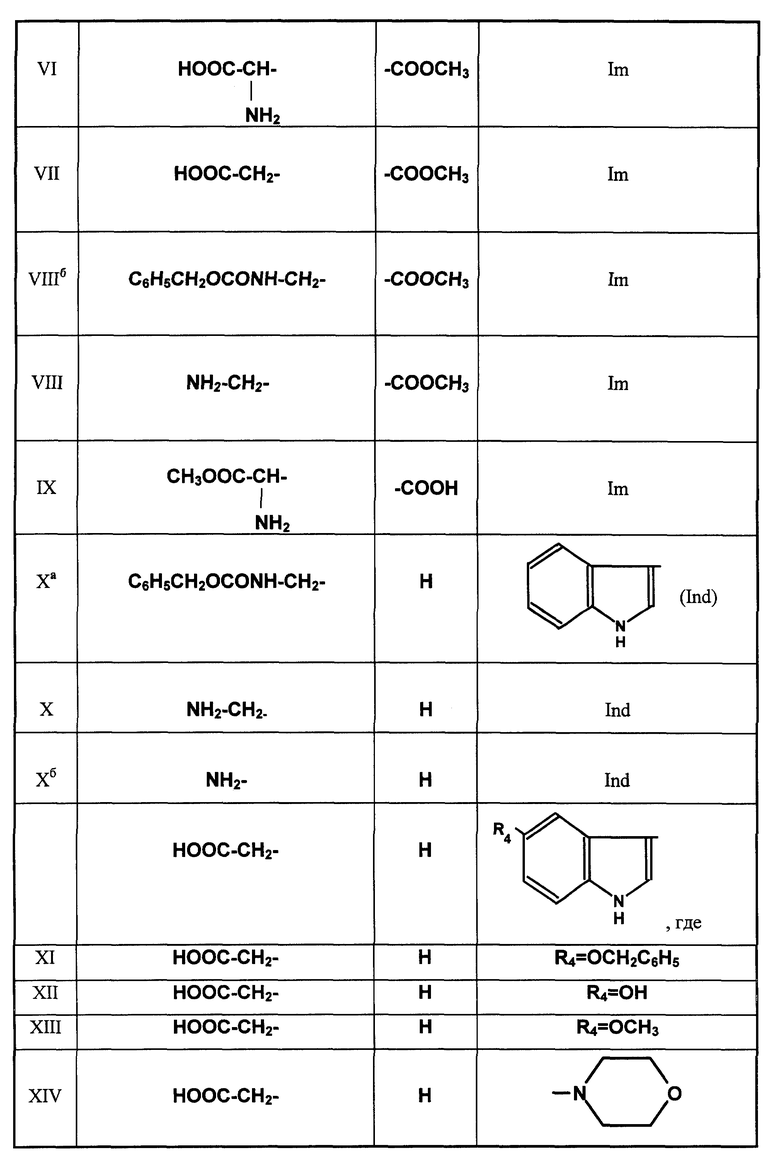
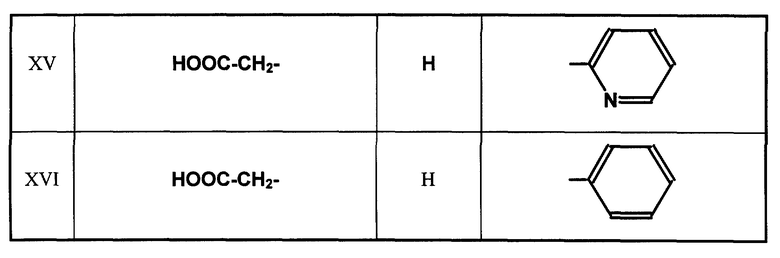
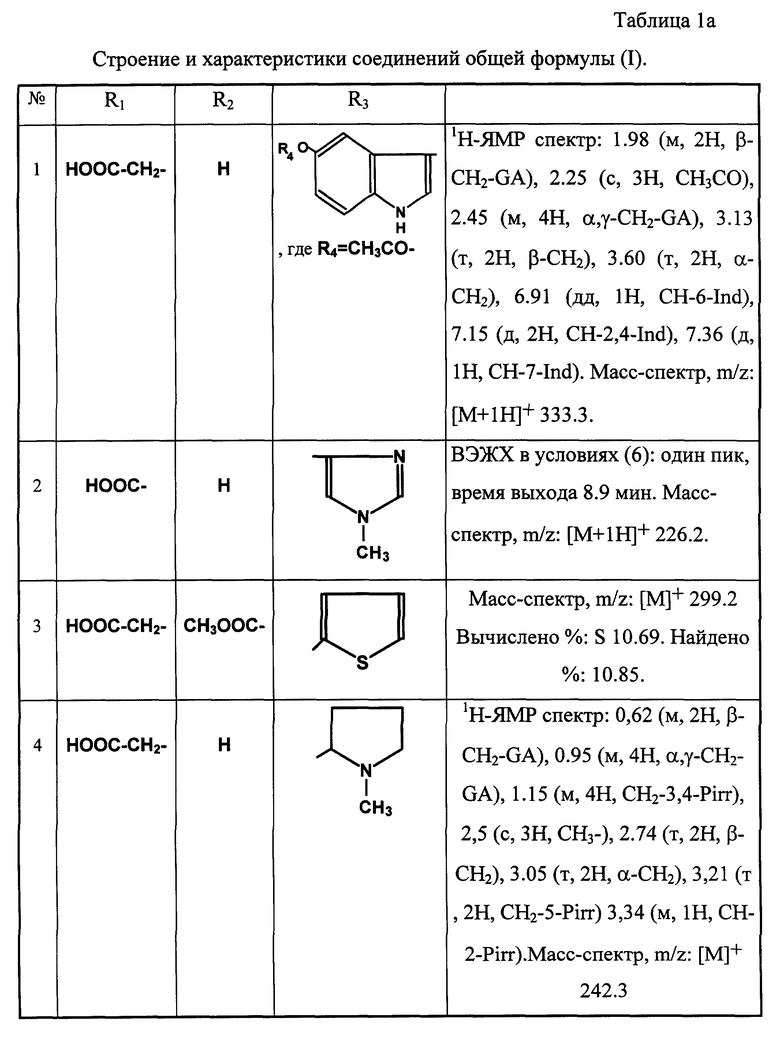
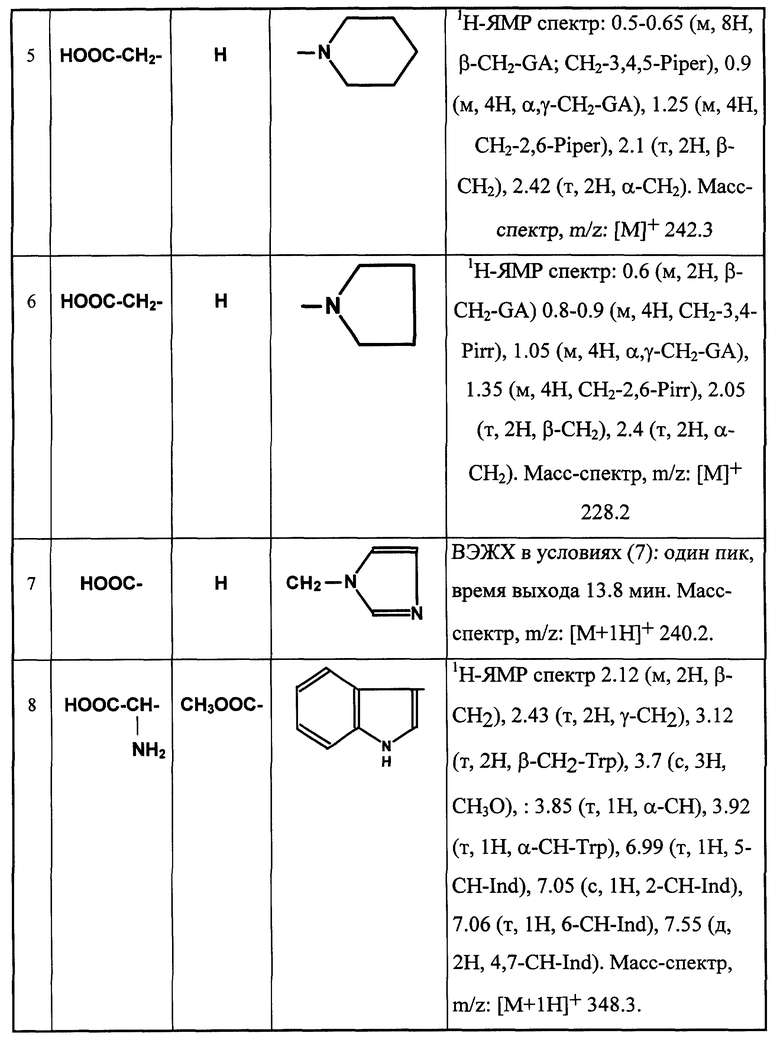
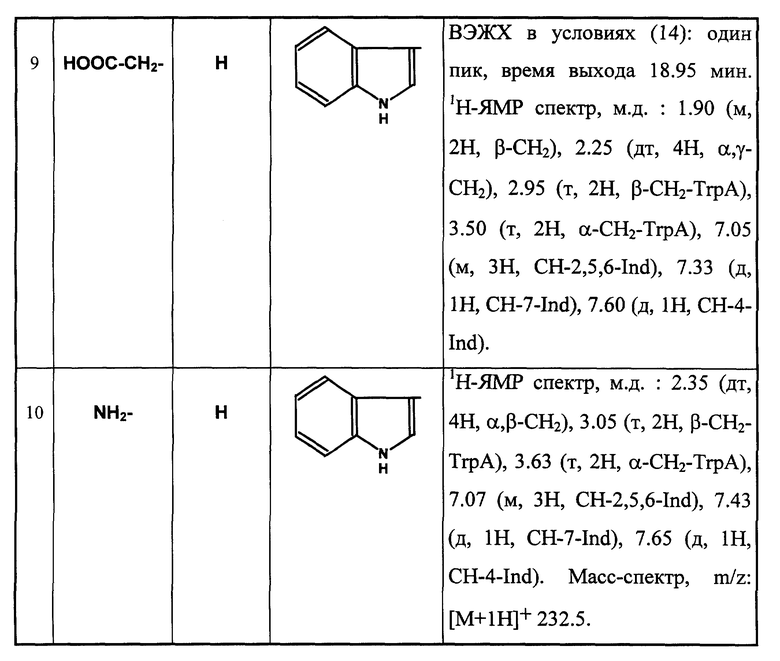
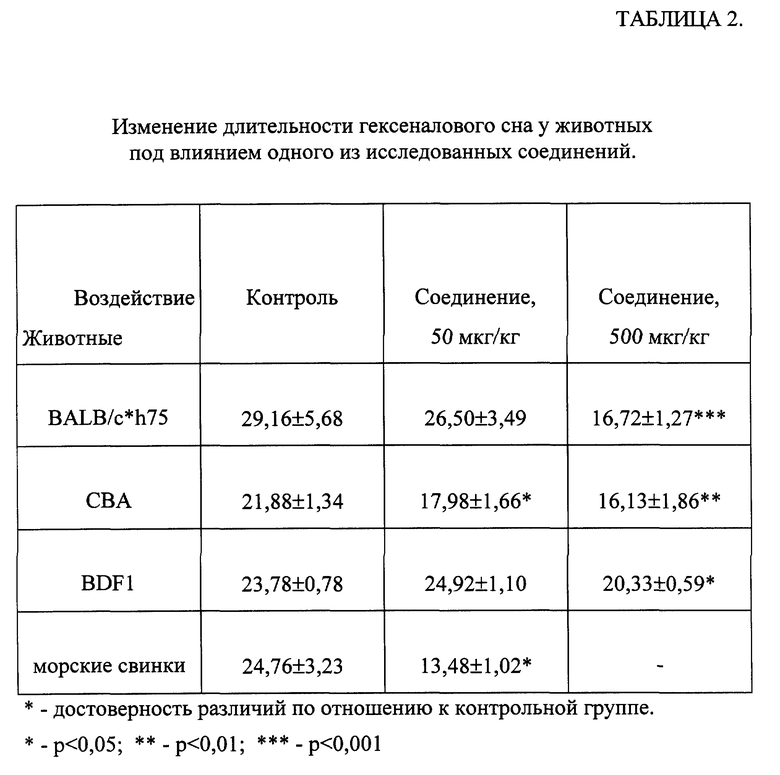
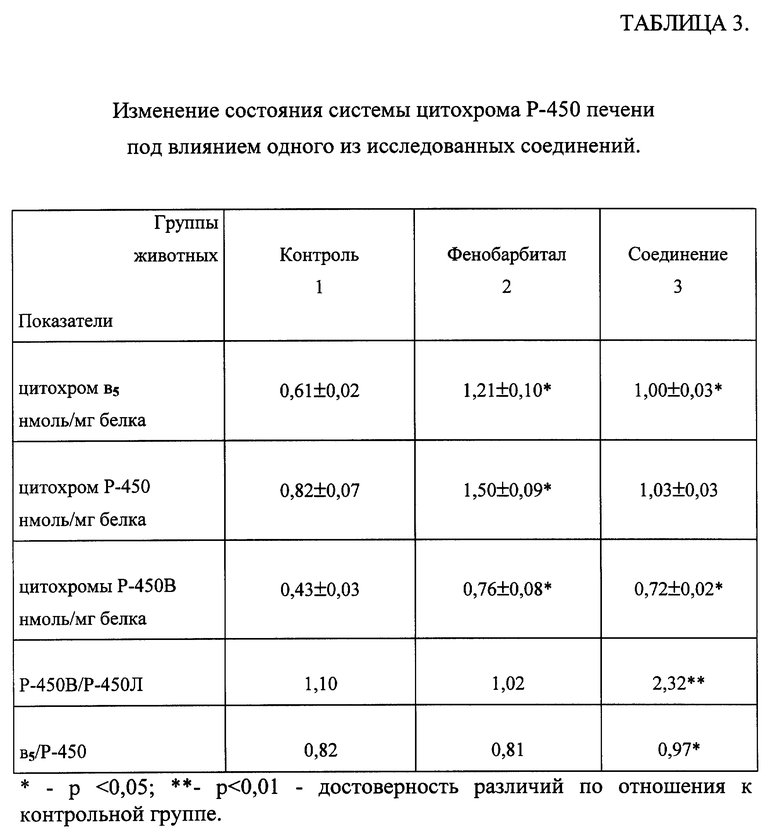
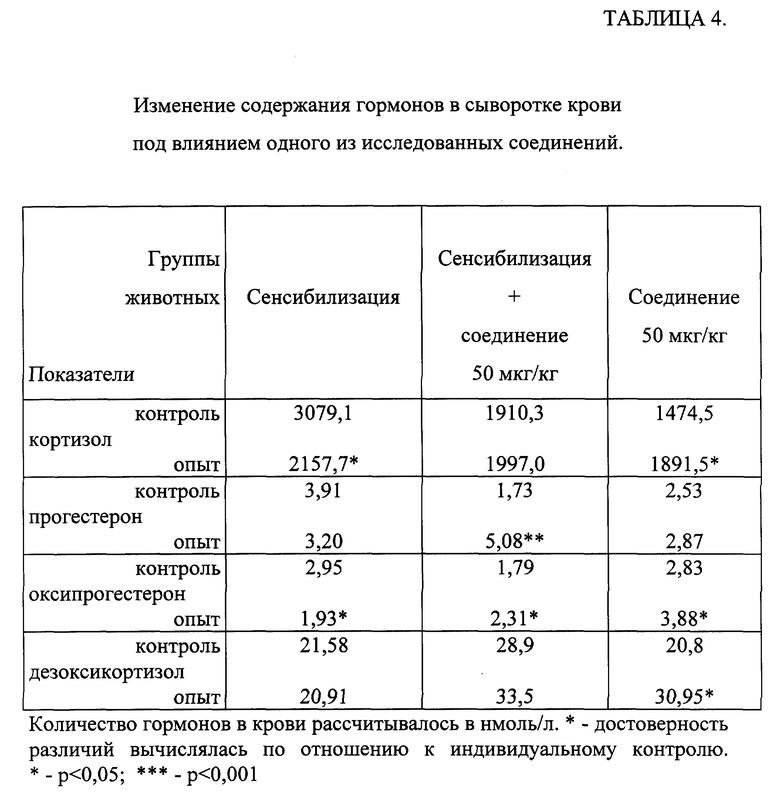
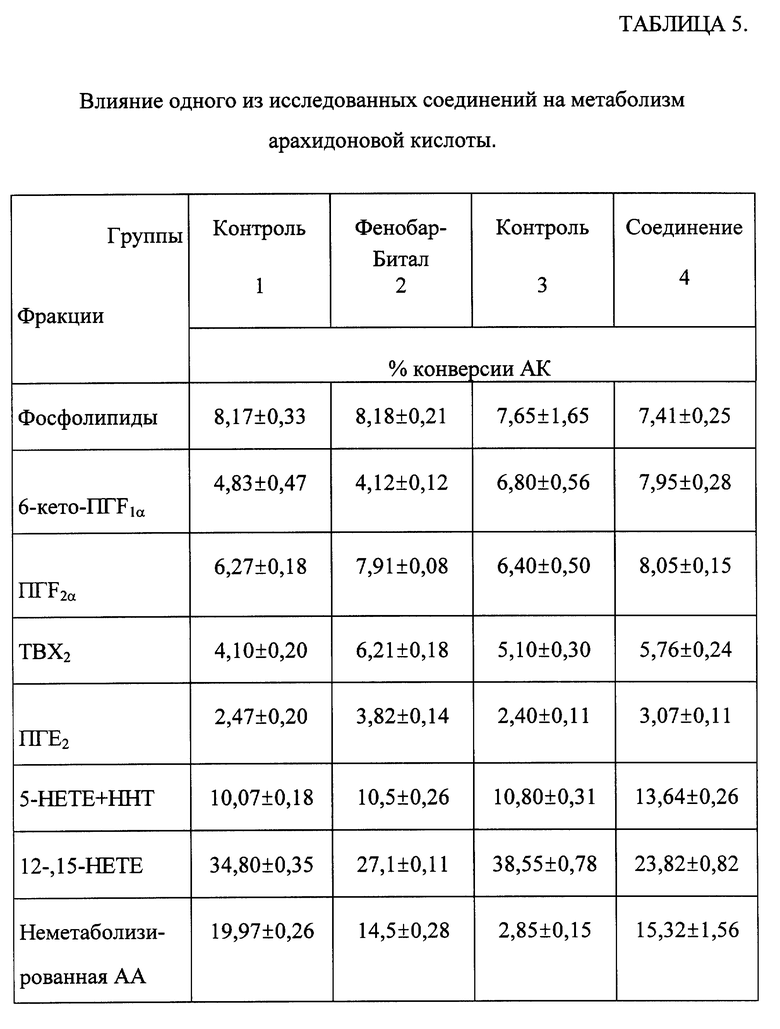
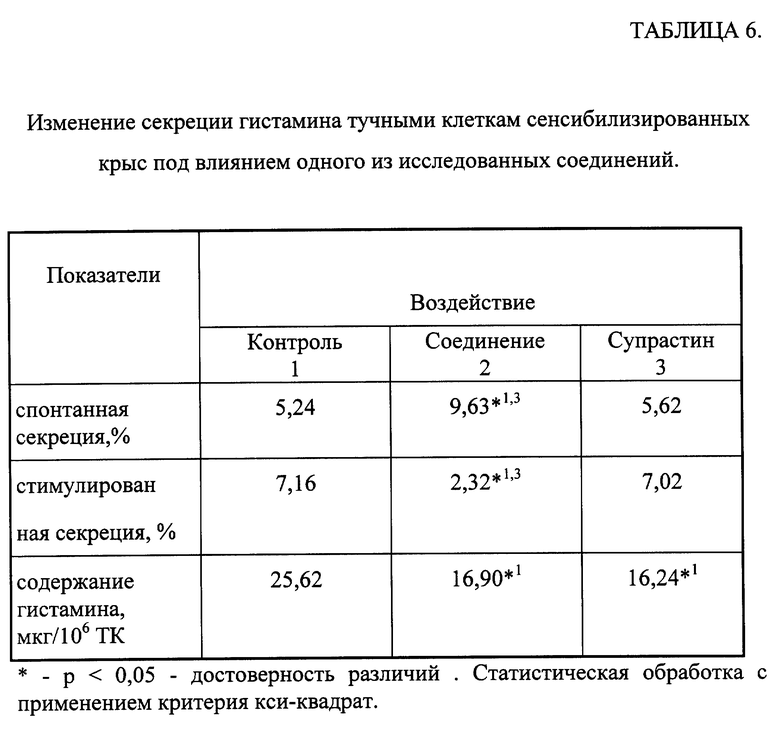
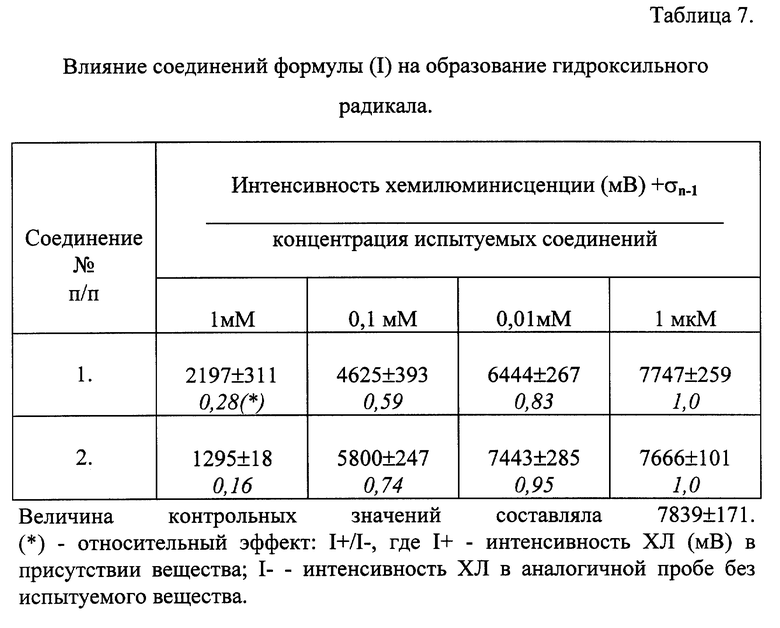
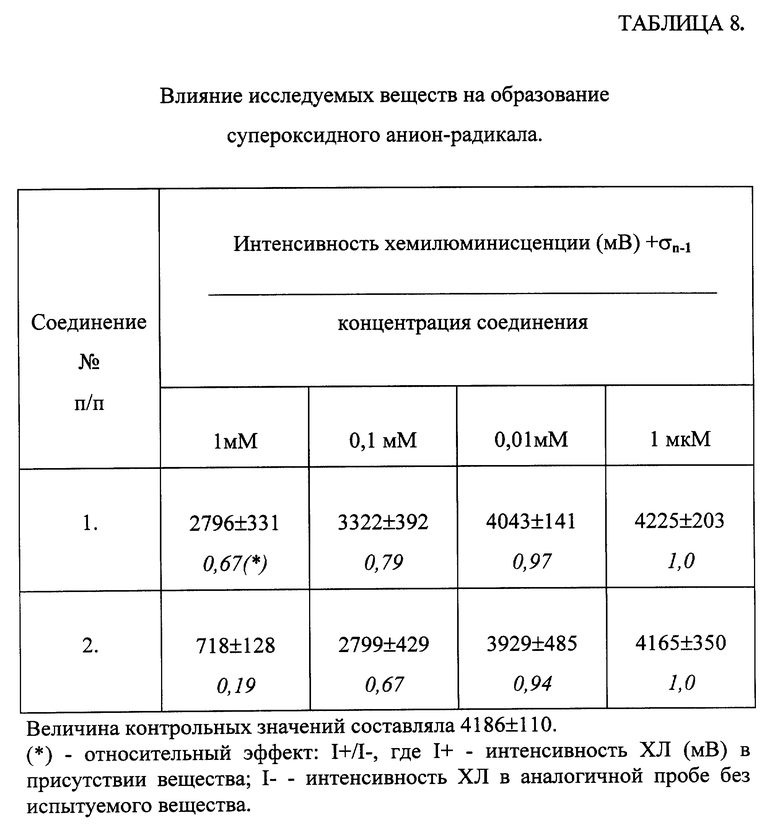
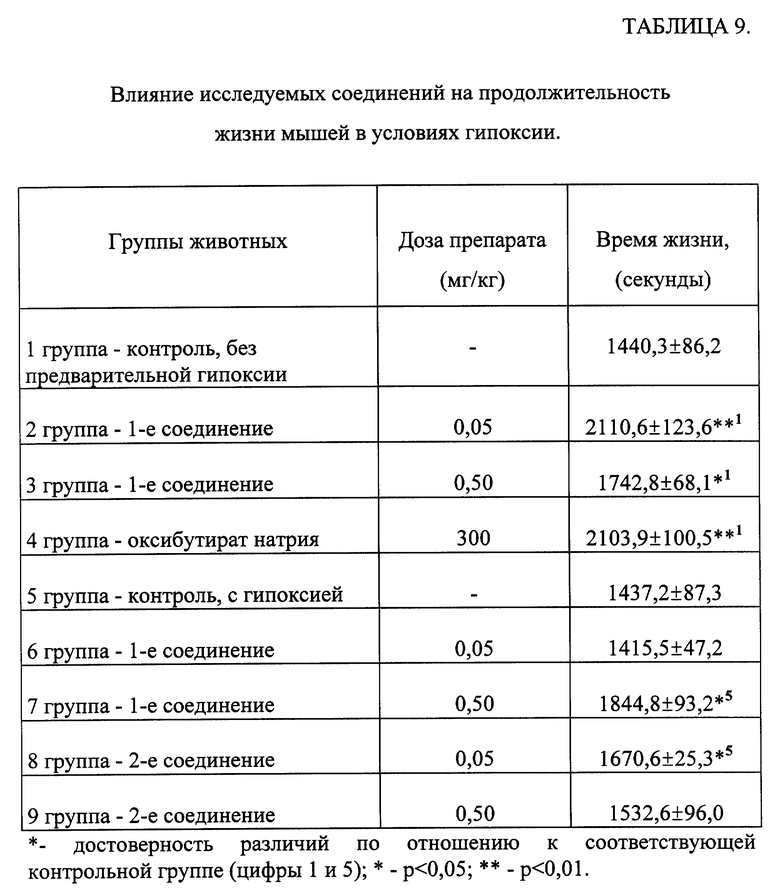
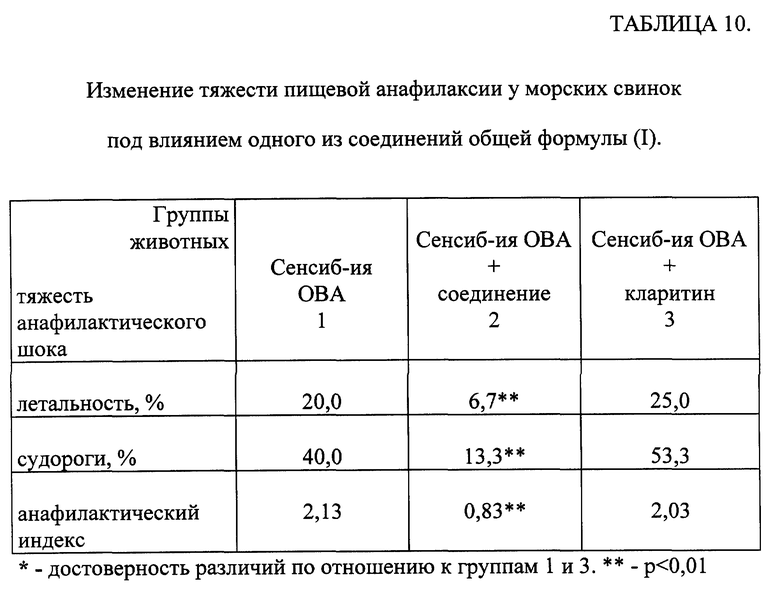

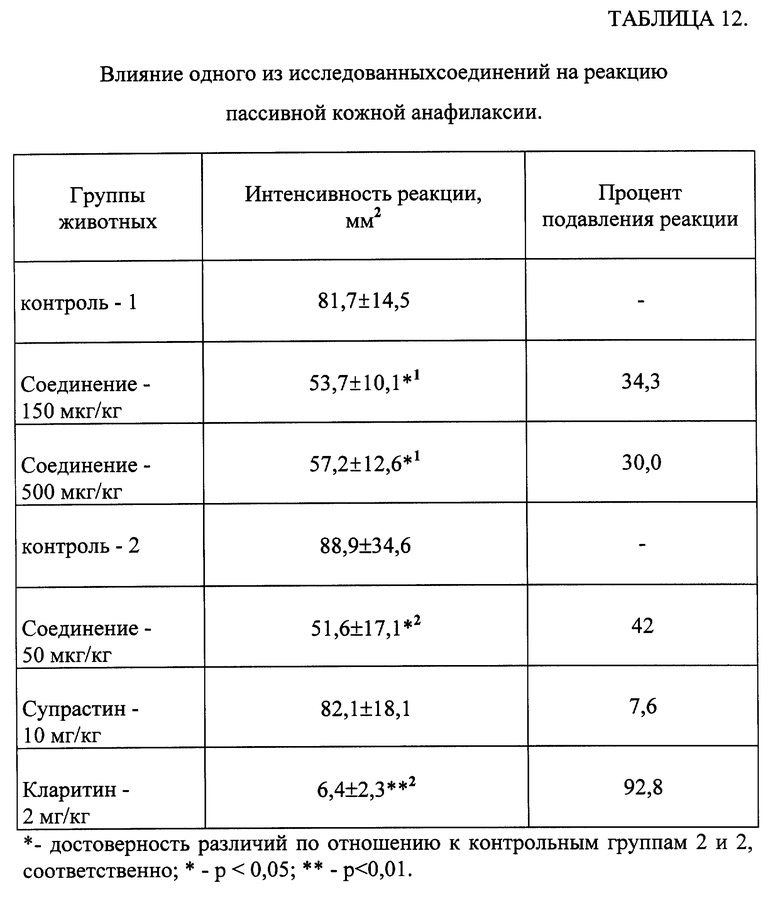
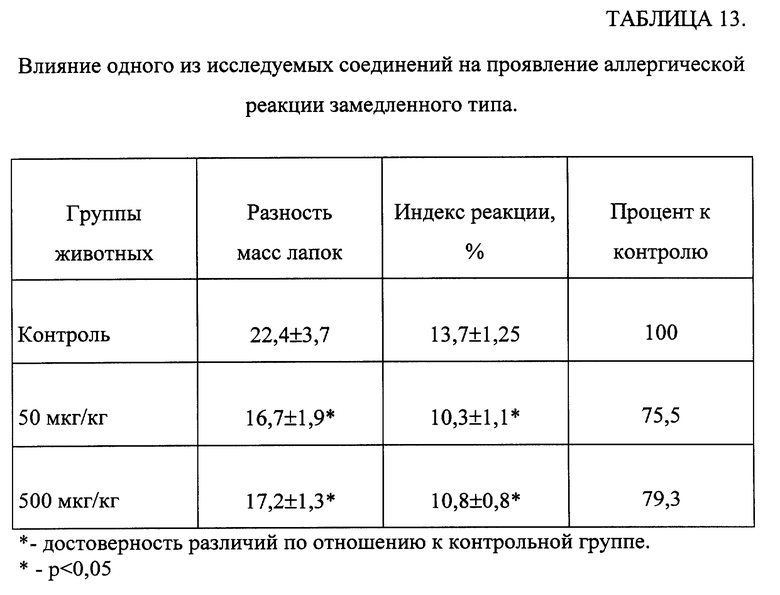
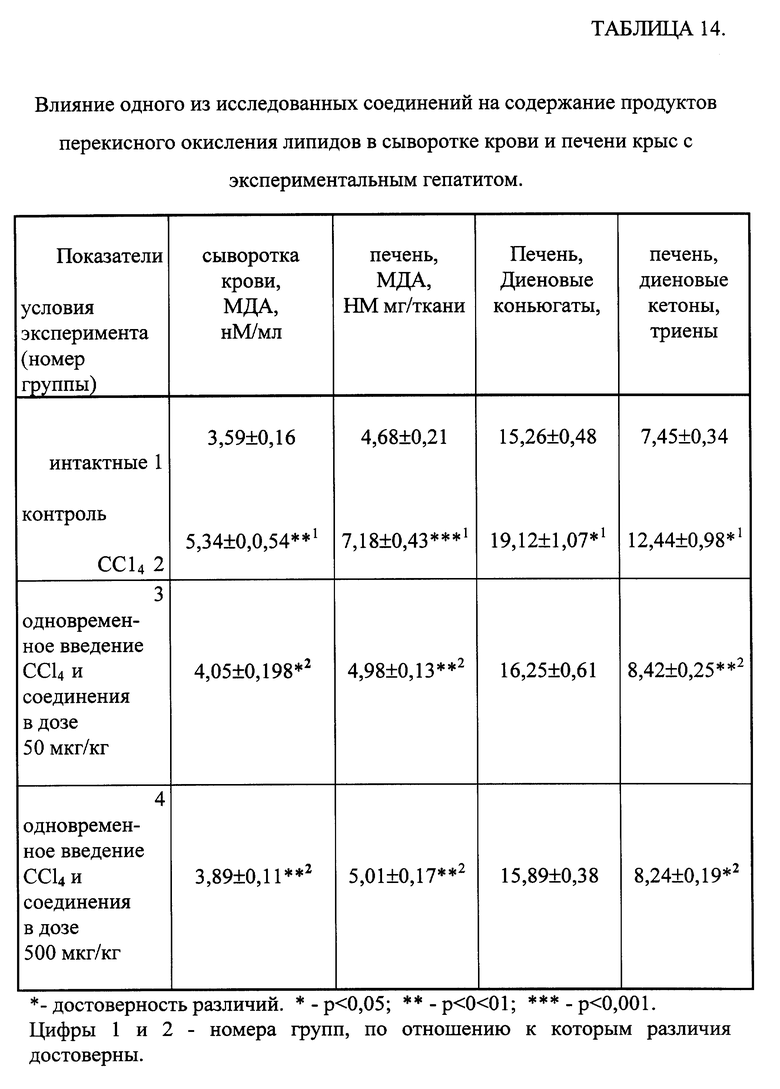
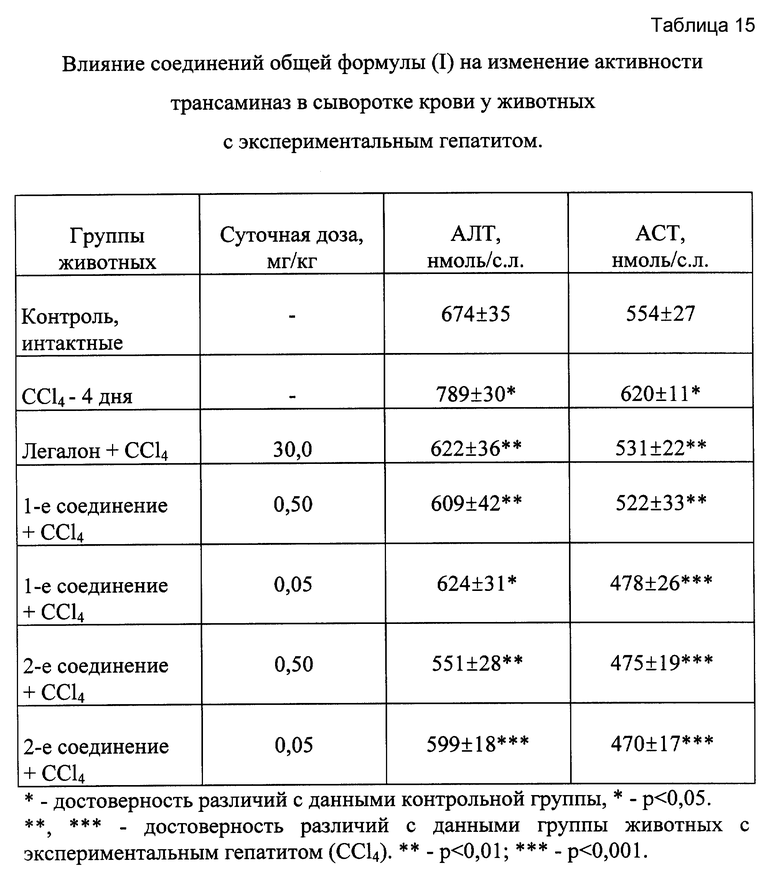
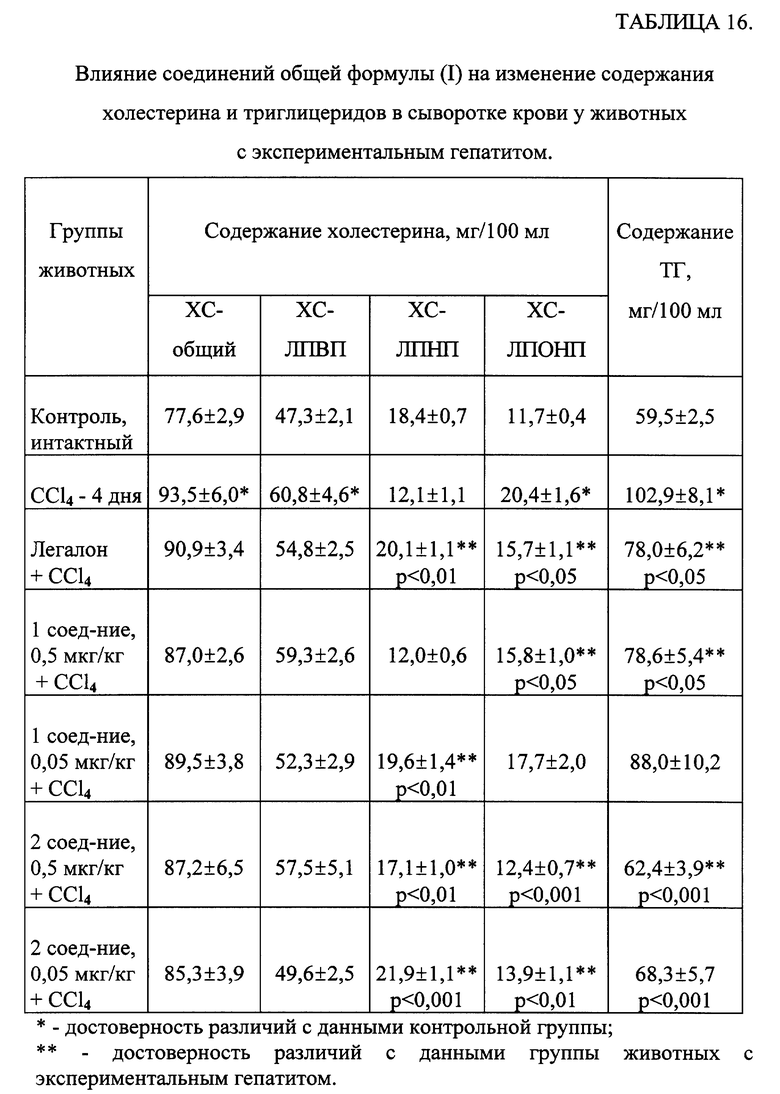
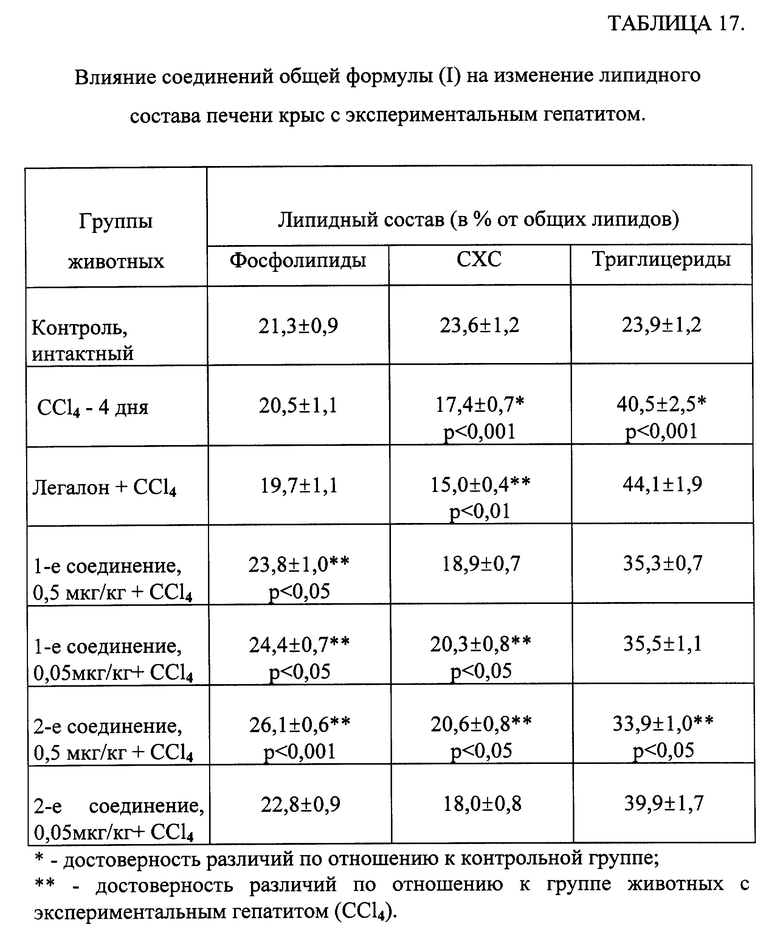
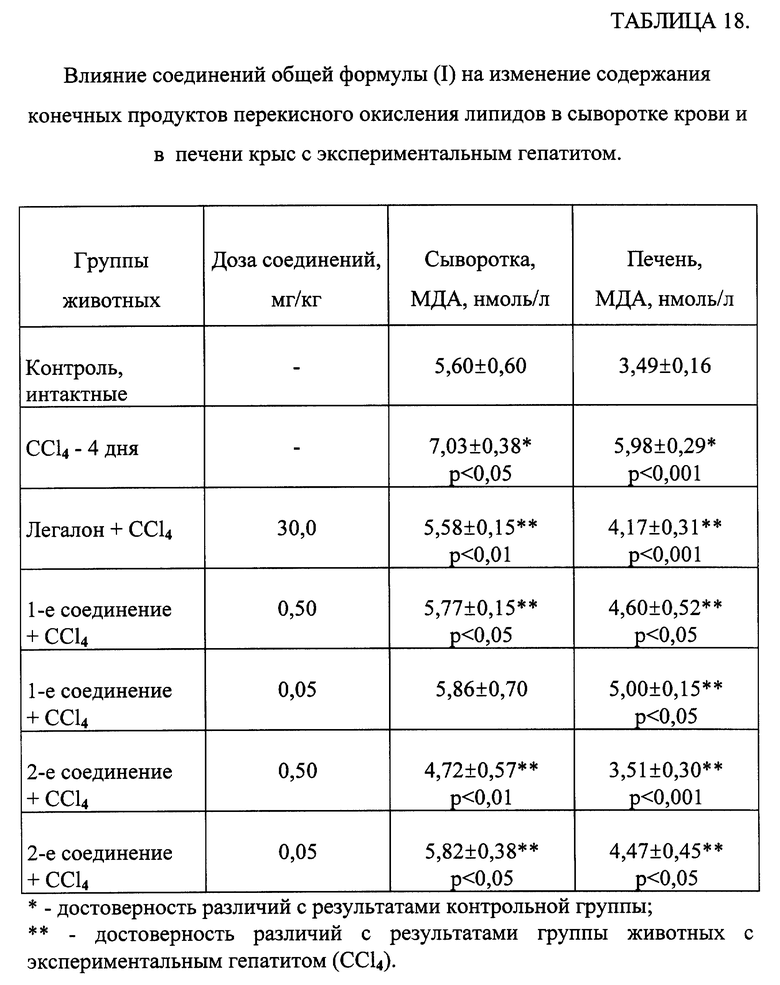
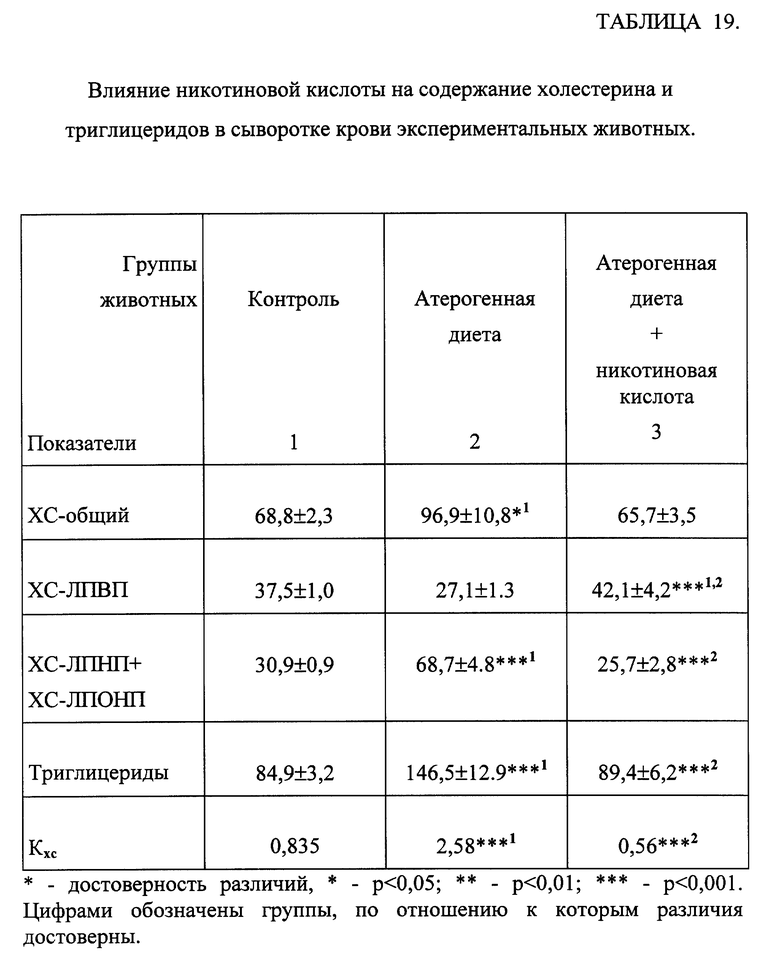
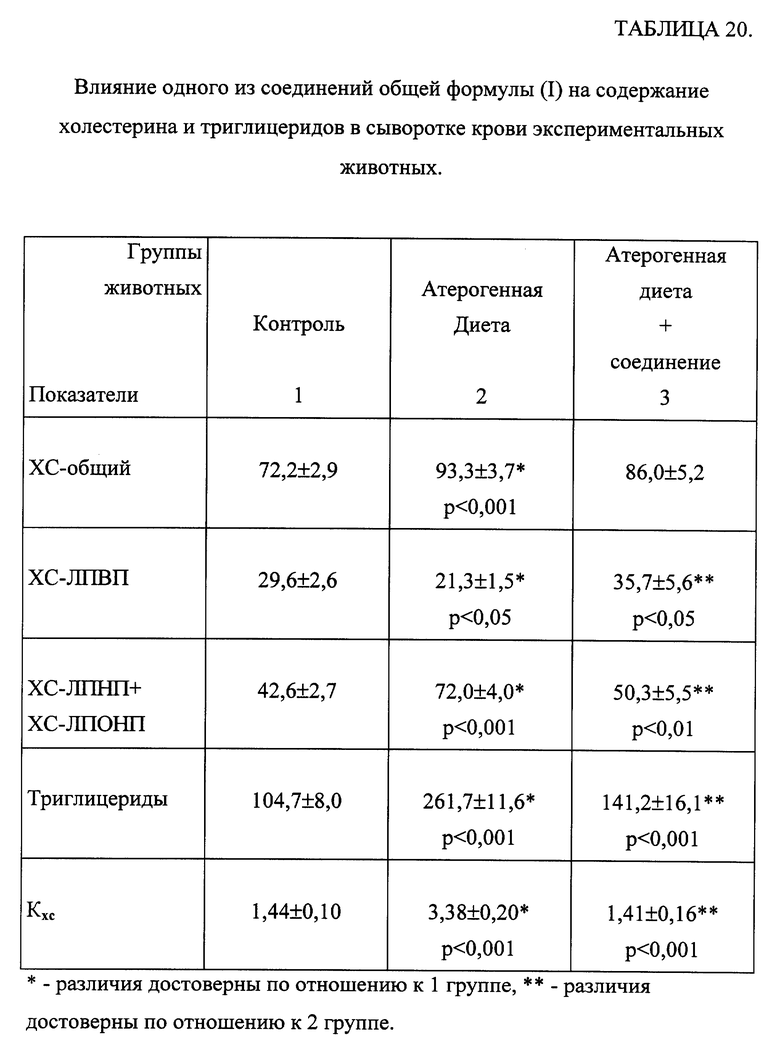
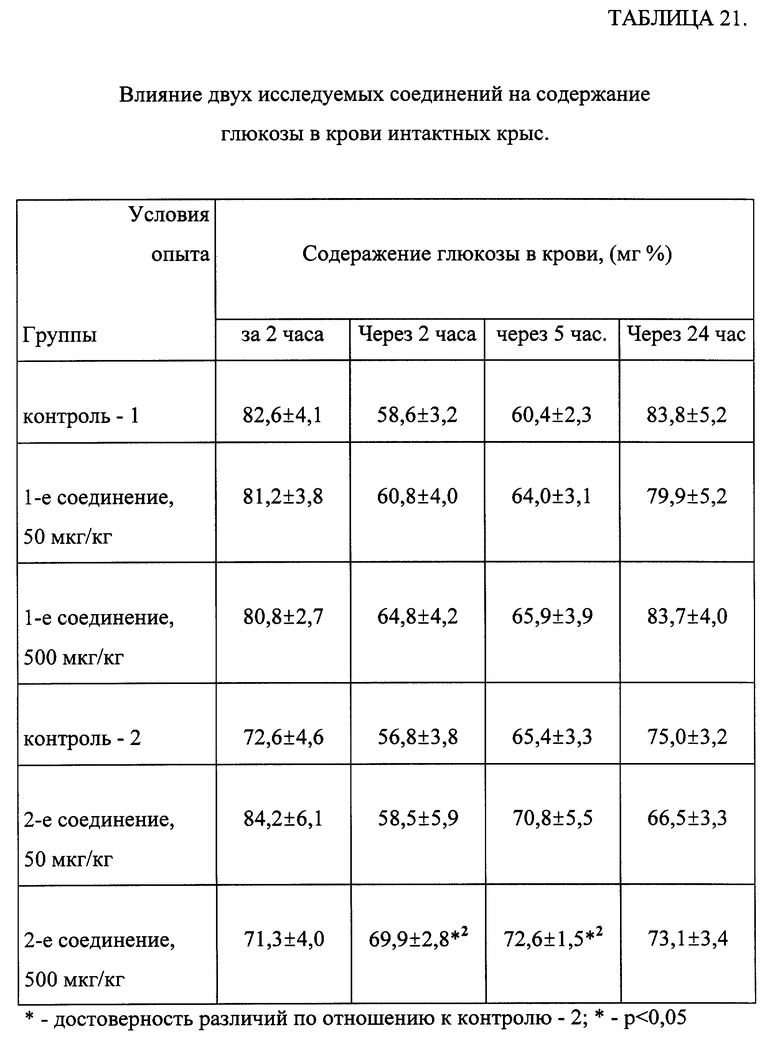

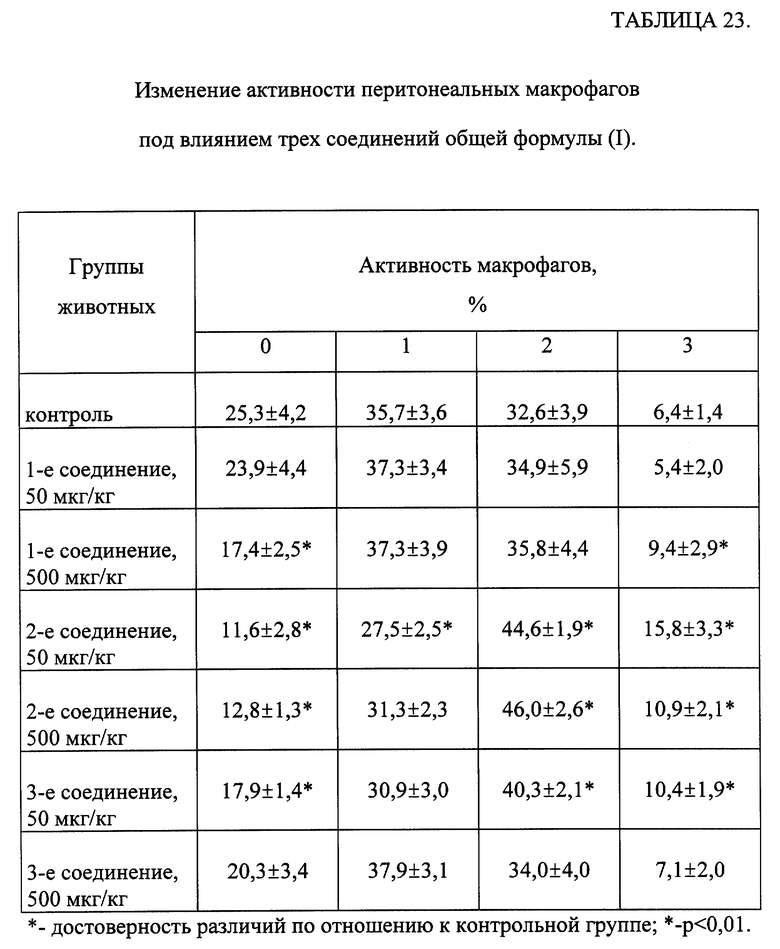
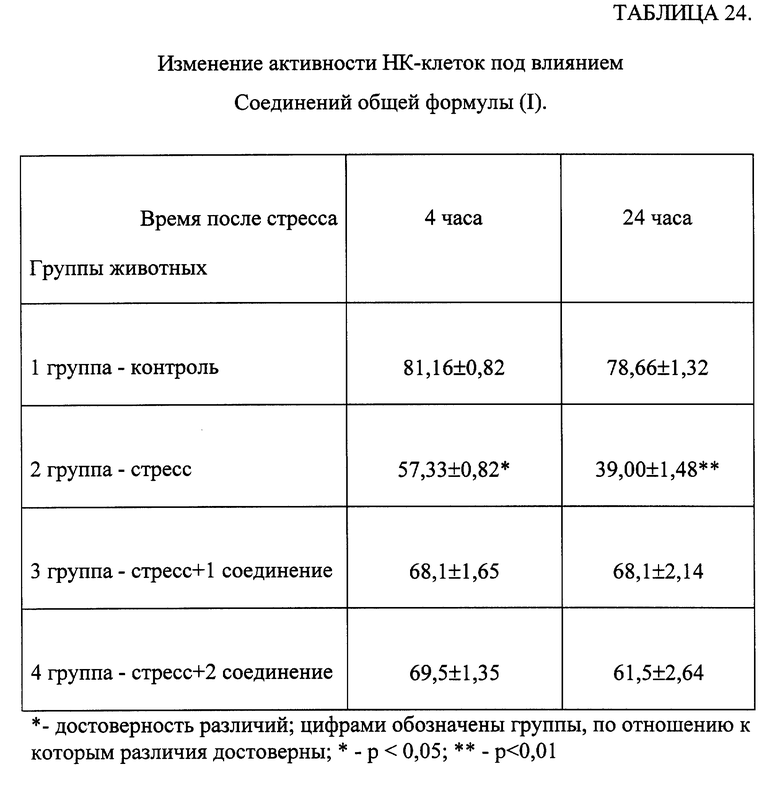
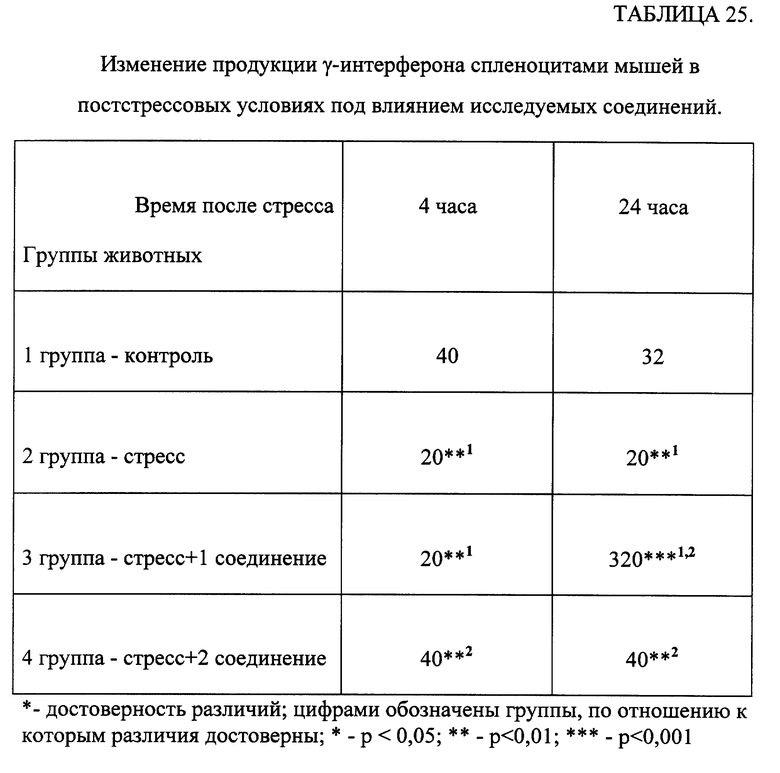
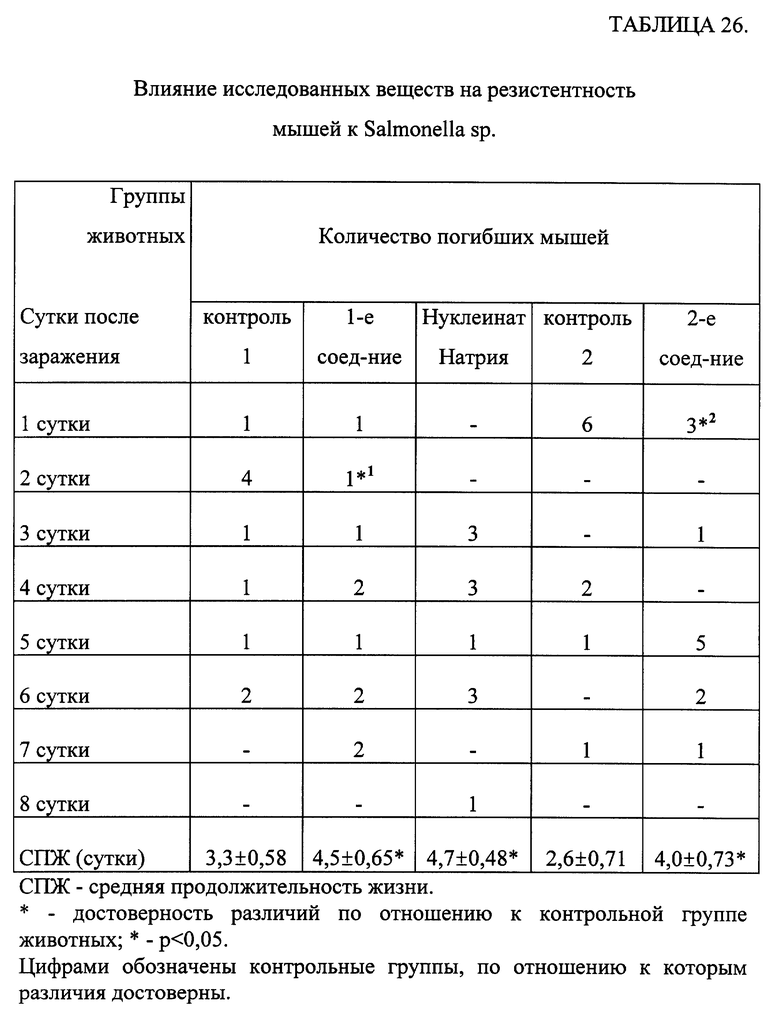
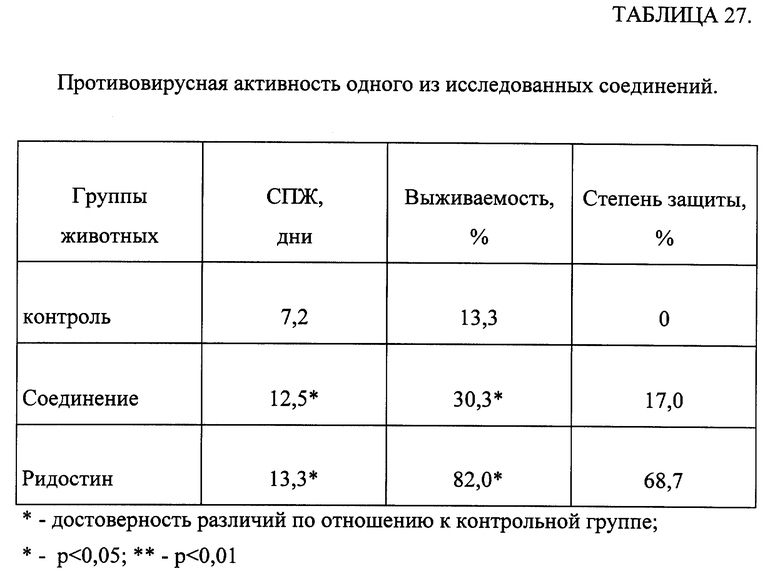
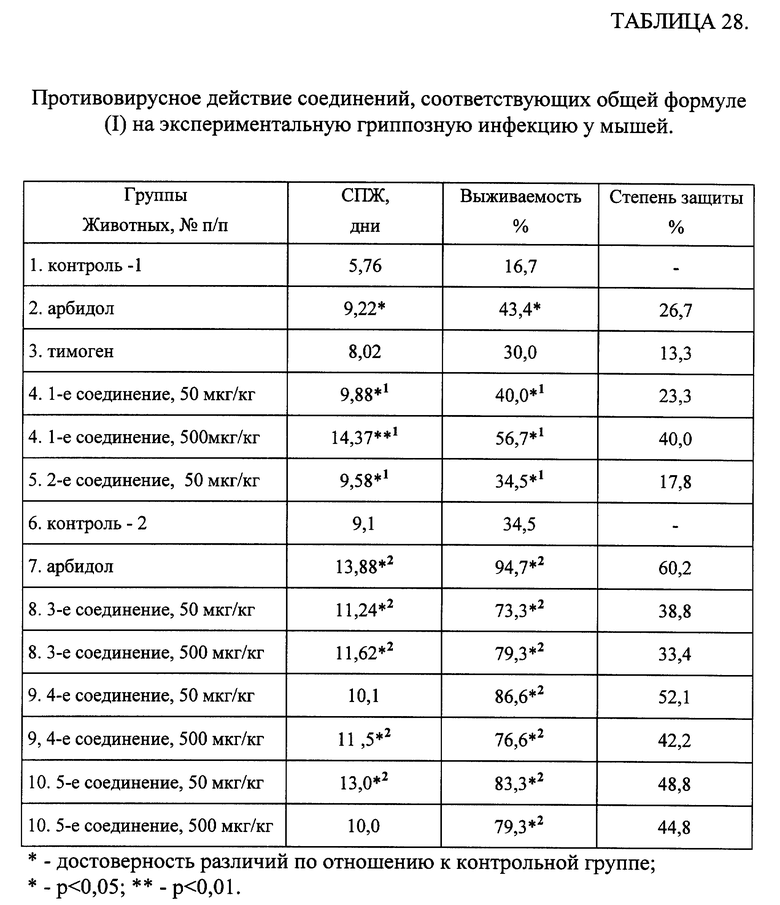
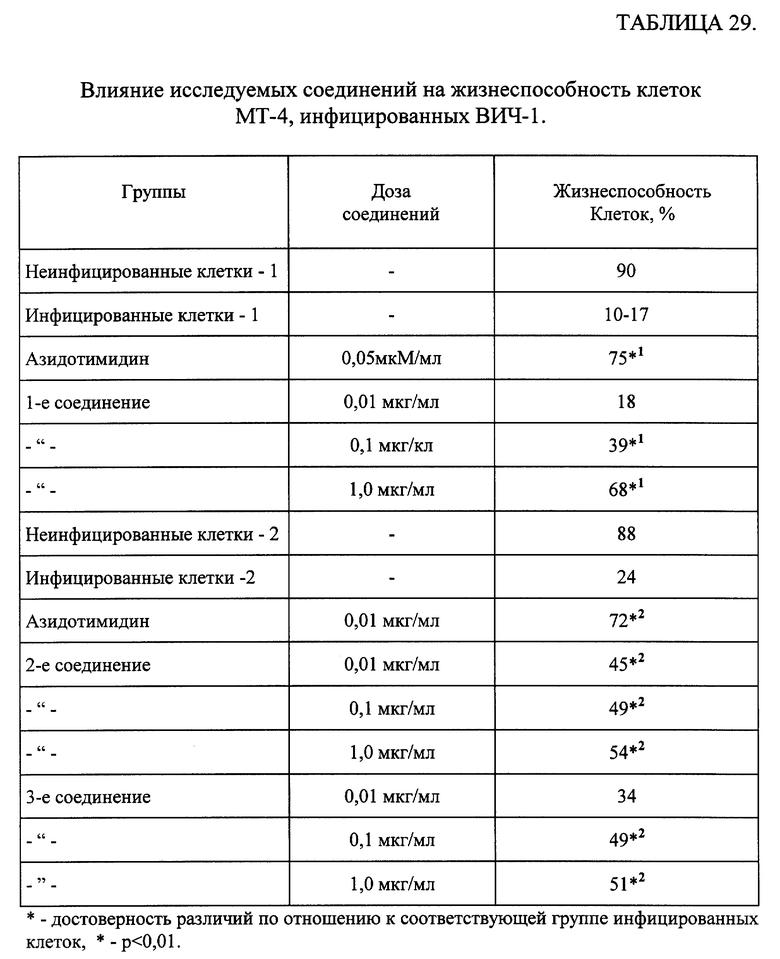
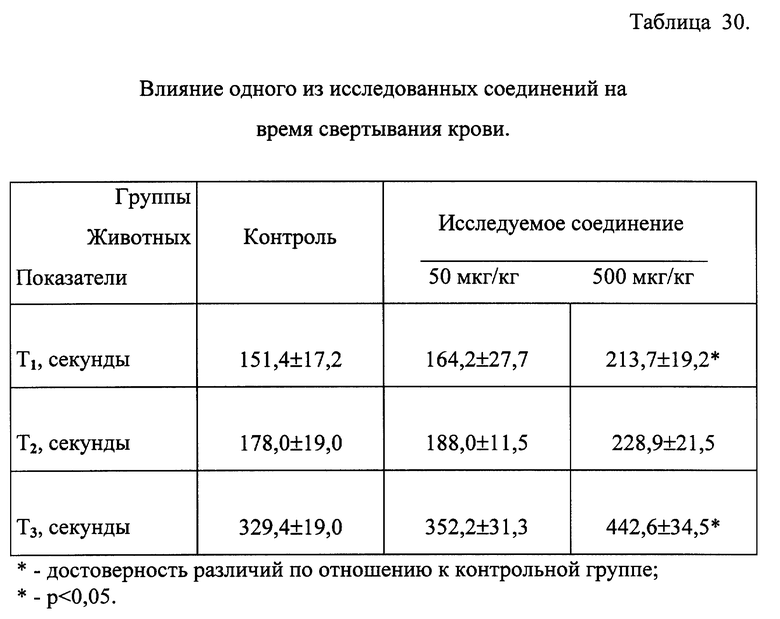
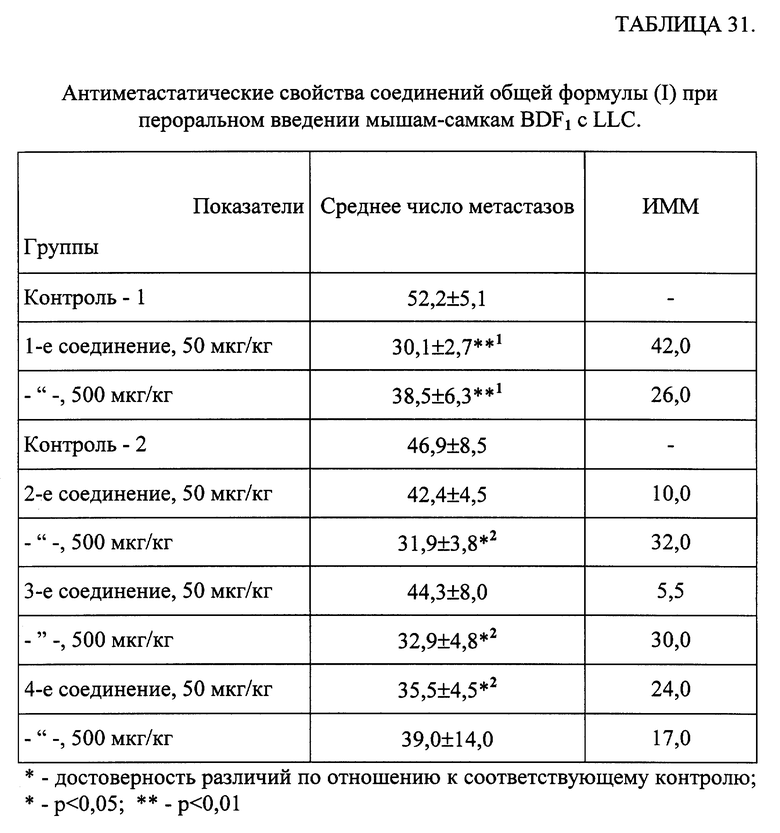
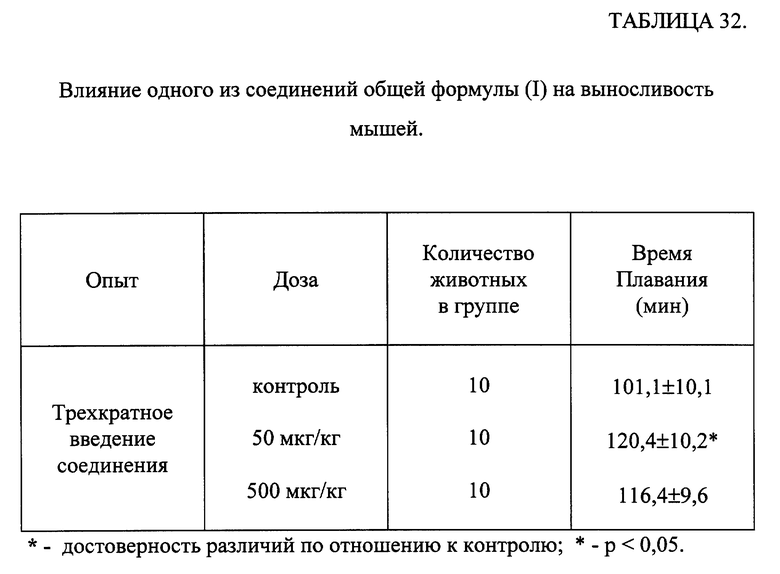
Производные пептидов общей формулы I:
R1 -GH2-СН2-CONH-CH(R2)CH2-R3
или их фармацевтически приемлемые соли,
где R1 представляет собой водород или углеводородный радикал с C1-С3, замещенный функциональной группой, например амино, амидной с C1-С5, уретановой с C1-С7 или карбоксильной группой, или углеводородный радикал с C1-С3, одновременно замещенный амино- и карбоксильной группой, причем карбоксильная группа может быть этерифицирована, а аминогруппа может быть замещена ацильным заместителем или эфиром угольной кислоты, R2 - атом водорода или функциональная группа, например карбоксил, который может быть этерифицирован, R3 - индол или его метильные и/ или гидроксильные производные, причем гидроксильная группа может быть ацилирована, алкилирована или аралкилирована, 5-6-членные насыщенные и ненасыщенные циклические и гетероциклические заместители, содержащие кислород, серу и/или 1-3 атома азота, или их метильные производные, причем при R1=NH2, NH2-CH2- R2=H, R3 не означает -4-Im, -3-(5-OMe-Ind), -3-(5-ОН-Ind), при R1=NH2, NH2CH2 R2=COOH, R3 не означает -4-Im, а при R1=HOOC-CH(NH2)- R2=H, R3 не означает -3-Ind, -3-(5-OH-Ind). Соединения I обладают свойствами модуляторов системы цитохрома Р-420, метаболизма арахидоновой кислоты, гормонов коры надпочечников, активности макрофагов, а также антиоксидантным, противоастматическим, антигипоксическим, противовирусным, липидрегулирующим, антиметастатическим, сахаропонижающим, адаптогенным и другими видами терапевтического действия. Соединения формулы I получают ацилированием аминогруппы в соединении II
NН2-СН-(R2)-СН2-R3
карбоксильным компонентом
R1-СН2-СН2-СОХ,
где Х - активирующая группировка. 4 с. и 19 з.п. ф-лы, 33 табл.
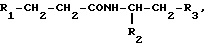
или их фармацевтически приемлемые соли,
где R1 представляет собой COOH, NH2 или углеводородный радикал с С1 - С3, замещенный функциональной группой, например, амино, амидной с С1 - С5, уретановой с С1 - С7 или карбоксильной группой; или углеводородный радикал с С1 - С3, одновременно замещенный амино- и карбоксильной группой, причем карбоксильная группа может быть этерифицирована, а аминогруппа может быть замещена ацильным заместителем или эфиром угольной кислоты;
R2 - атом водорода или функциональная группа, например карбоксил, который может быть этерифицирован;
R3 - индол или его метильные и/или гидроксильные производные, причем гидроксильная группа может быть ацилирована, алкилирована или аралкилирована;
5 - 6 членные насыщенные и ненасыщенные циклические и гетероциклические заместители, содержащие кислород, серу и/или 1 - 3 атома азота, или их метильные производные, причем при R1=NH2, NH2-CH2-, R2=H, R3= не означает -4-Im, -3-(5-Ome-Ind), -3-(5-OH-Ind); при R1-NH2, NH2-CH2-, R2=COOH, R3= не означает -4-Im; а при R1=HOOC-CH(NH2)-, R2=H, R3= не означает -3-Ind, -3-(5-OH-Ind).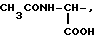
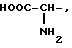
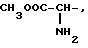
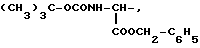
R2=H, -COOH, -COOOCH3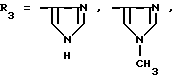
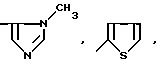
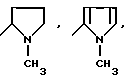
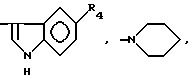
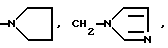
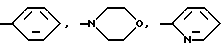
причем R4=-H, -OH, -OCH3, -OCH2C6H5, а при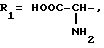
R2=H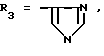
H - остаток глутаминовой кислоты -D-конфигурации.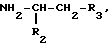
активированным по карбоксильной группе карбоксильным компонентом общей формулы
R1-CH2-CH2-COX,
где Х - активирующая группировка.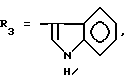
а при R1=NH2-CH2-, или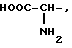
R2= -COOCH3,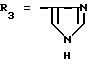
в качестве ацилирующего агента используют пентафторфениловые эфиры N-бензилоксикарбонил-γ-аминомасляной кислоты или α-бензилового эфира N-трет. -бутилоксикарбонил-L-глутаминовой кислоты и процесс ведут в органическом растворителе с последующим отщеплением от защищенных производных дипептидов N-бензилоксикарбонильной защиты и α-бензилового эфира каталитическим гидрогенолизом, с последующим отщеплением N-трет.-бутилоксикарбонильной группы в безводной кислой среде.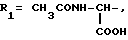
R2=H,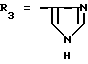
в качестве ацилирующего агента используют п-нитрофенилацетат.
где R1=HOOC-(CH2)n-, n=0-2, C6H5CH2OCO-NH-CH2-, NH2-CH2-, R1=NH2-,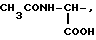
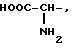
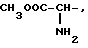
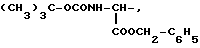
R2=H, -COOH, -COOCH3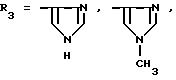
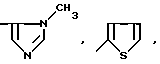
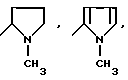
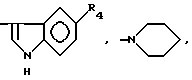
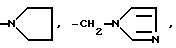
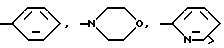
причем R4= -H, -OH, -OCH3, -OCH2C6H5, а при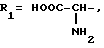
R2=H,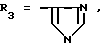
H - остаток глутаминовой кислоты -D-конфигурации,
в эффективном количестве и целевые добавки.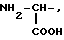
R2=H, COOH,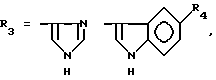
где R4= -H, -OH, -OCH3, -OCH2C6H5,
в количестве средств, обладающих антигипоксическим, антиоксидантным, антирадикальным, липидрегулирующим, гипогликемическим и антикоагулянтным действием, а также способностью индуцировать систему цитохрома Р-450, модулировать метаболизм арахидоновой кислоты, гормонов коры надпочечников и щитовидной железы, снижать содержание и антигензависимую секрецию гистамина перитонеальными тучными клетками, модулировать активность макрофагов.
| SU 1287756 A3, 1987 | |||
| US 5274076 A, 1993 | |||
| Нитепроводник для сновальных рам | 1928 |
|
SU12401A1 |
| EP 0144290 A, 1985 | |||
| RU 94007653 A1, 1995 | |||
| СПОСОБ ПОЛУЧЕНИЯ КАРНОЗИНА | 1990 |
|
RU2030422C1 |

