Настоящее изобретение касается LACTOBACILLUS BULGARICUS, обладающих уменьшенным продуцированием кислоты и/или продуцированием улучшенного аромата и вкуса.
Настоящее изобретение также относится к пищевому составу, в особенности к нежному йогурту или продуктам, похожим на нежный йогурт, содержащим Lactobacillus Bulgaricus согласно изобретению.
Предпосылки изобретения и уровень развития техники в данной области
Йогурт образуется в результате роста ассоциаций двух видов кисломолочных бактерий, Streptococcus thermophilus и Lactobacillus bulgaricus. Они развиваются вместе в молоке, где они ферментируют лактозу, которая присутствует в количестве 40-50 г на 1 л лактата. В общем ферментация обычно происходит в течение времени от 3-х до 5-ти часов при 40oC, при этом pH понижается от нейтрального значения до pH 4,2. При хранении в течение нескольких дней при 4-12oC pH снижается далее до значений ниже 4,0. Параллельно возросшей кислотности развивается появление горькости, которая снижает органолептическое качество продукта.
Именно L. bulgaricus вызывают типичный приятный вкус йогурта во время ферментации. Следовательно, отсутствие в йогурте или наличие в нем пониженного титра метаболических активных клеток L.bulgaricus приводит к отсутствию в обычном йогурте ароматических компонентов. Таким образом, такой йогурт /или продукты, похожие на йогурт/ могут быть очень нейтральными, т.е. безвкусными. Эта проблема является в особенности значительной в производстве нежного или очень нежного йогурта, который в результате ограниченной скорости роста L.bulgaricus имеет склонность к большой потере аромата.
Количество потребителей, предпочитающих более нежный, но все таки ароматный, с приятным вкусом йогурт, в последние годы возросло. Следовательно, способ регулирования продуцирования кислоты в процессе ферментации молока и предотвращения последующего подкисления за счет снижения pH во время хранения является поистине желательным для производителей йогурта.
До настоящего времени было предложено несколько способов или усовершенствований исходного штамма для регулирования подкисления и послеподкисления йогурта. В большинстве из них уменьшают количество активных клеток в исходной культуре или уничтожают живые клетки в конечном продукте путем пастеризации. Альтернативно можно использовать слабоподкисляющие штаммы.
В патенте США N A-4734361 (Murao et. al.) описан такой штамм Lactobacillus bulgaricus (OLL 1074), который был осажден в Японии под номером FERM BP 1041.
В этом варианте показана слабая тенденция в направлении образования молочной кислоты при пониженной температуре.
При использовании этого варианта можно производить сброженное молоко или молочнокислый напиток, в котором послеподкисление при пониженной температуре уменьшается.
В патенте США N 5.071.763 (Somkutu et. al.) описано, что мутантные штаммы Streptococcus thermophilus, имеющие недостаточные системы транспорта лактозы и имеющие gluS, lacS-, SucS+ и βgal+ фенотип, являются эффективными для использования в способах, где важен гидролиз лактозы. Термоустойчивость этих штаммов, а также продуцирование β- галактозидазы обеспечивают гидролиз лактозы до и во время пастеризации. Эти организмы обеспечивают пищевую промышленность усовершенствованными способами приготовления молочных продуктов с уменьшенным содержанием лактозы.
В L.bulgaricus лактоза поглощается лактозной проницаемой системой и расщепляется посредством β- галактозидазы на две части: глюкозу и галактозу. Глюкоза затем метаболизируется в пируват, большая часть которого превращается в лактат посредством D-лактатдегидрогеназы (D-LDH).
Однако часть пирувата декарбоксилируется до ацетальдегида, важного компонента приятного запаха йогурта, или расходуется на другие пути обмена, в результате чего получают приятный запах или предшествующие элементы приятного запаха.
Thomas and al. (J. of Bacteriology, May 1974, с. 329-333) и Smart and al (Applied and Environmental Microbiology Mar, 1987, с. 533-541) описывают, что для Lactic streptococci гомоферментативное брожение можно изменить в гетероферментативное брожение посредством уменьшения активности LDH.
Payton and al. (F. ERM Mictobiology letters, 26, 1985, с. 333-336) описывают, что вычитание LDH из мутанта Bacillus Stearo thermophilus приводит к увеличению количества полученного этанола (при осуществлении ферментации глюкозы), поскольку они теряют способность вырабатывать лактат.
В документе Jornal of Bacteriology (т. 144; п. 1, 1980, Baltimore, США, сс. 217-221) показано, что когда штамм Lactobacillus bulgaricus NLS-4 развивается анаэробно в целостной культуре при ограничении глюкозы, сдвиг pH среды от кислой до щелочной области заставляет обычно гомоферментативную бактерию катаболизировать глюкозу гетероферментативным образом. Изменения при ферментации сопровождаются уменьшением биосинтеза лактатдегидрогеназы (LDH) в щелочных условиях.
Йогурт находится в кислой среде, однако в этом документе не говорится о том, что можно получить уменьшение биосинтеза лактатдегидрогеназы в кислых условиях.
Цели изобретения
Целями настоящего изобретения являются обеспечение Lactobacillus bulgaricus, обладающих продуцированием улучшенного аромата и вкуса.
Другой целью настоящего изобретения является обеспечение Lactobacillus bulgaricus, также обладающих уменьшенным подкислением и послеподкислением.
Другой целью настоящего изобретения является получение пищевого состава, предпочтительно нежного йогурта или продуктов, похожих на нежный йогурт, содержащего (их) упомянутые Lactobacillus bulgaricus, имеющие повышенное продуцирование аромата и вкуса и/или уменьшенное подкисление предпочтительно при низкой и высокой температуре.
Описание изобретения
Настоящее изобретение касается Lactobacillus bulgaricus, имеющих более низкую активность лактатдегидрогеназы (LDH), чем у штамма дикого типа Lfi5 (осажденного под наименованием CNCM 1-800 в институте Пастера, 26 rue du Docteur ROux, 75024 Paris Cedex 15, FRANCE).
Преимущественно упомянутая активность лактатдегидрогеназы составляет менее чем 50%, предпочтительно менее чем 10% активности лактатдегидрогеназы этого штамма дикого типа.
Преимущественно активность лактатдегидрогеназы составляет менее 250 единиц, предпочтительно менее чем 150 единиц активности фермента / 10 мл культуры.
Активность лактатдегидрогеназы измеряли посредством последующего уменьшения поглощения при 340 нм из-за превращения NADH в NAD+ в 1 мл реакционной смеси, 8 мМ пирувата натрия, 0,15 мМ NADH 50 мМ трис-хлорида pH 7,5. Одну единицу активности фермента определяли как количество, окисляющее 1 μ моль NADH в минуту при 25oC. L.bulgaricus в соответствии с изобретением также отличаются более высокой активностью - галактозидазы (β-gal), чем у штамма дикого типа Lfi5.
Преимущественно активность упомянутой β- галактозидазы составляет более, чем 200%, предпочтительно более чем 500% активности β- галактозидазы штамма дикого типа Lfi5.
Преимущественно активность β- галактозидазы составляет более чем 300 единиц, предпочтительно более чем 500 единиц активности фермента/10 мл культуры.
Активность β-галактозидазы определяют способом Miller J.H. (Experiments in molecular genetics, Cold Spring Habor Laboratory N.V, 1972).
50 мл бесклеточного экстракта добавили в 1 мл буфера Z, 0,1 М фосфата натрия pH 7,0, 10 мМ KCl, 1 мМ MgSO4, 50 мМ 2-меркаптоэтанола и приводили в равновесное состояние при 28oC. Добавили предварительно нагретые 200 μл ONPG и инкубировали при 28oC в течение 20 минут. Реакцию завершили путем добавления 0,5 мл 1 М Na2CO3.
Измеряли поглощение OD420. Одну единицу активности фермента определяли как количество, которое гидролизовало 1 μмоль ONPG в минуту при 28oC.
Предпочтительно Lactobacillus bulgaricus в соответствии с изобретением характеризуются активностью LDH, которая составляет менее чем 0,8, предпочтительно менее чем 0,3 активности β-галактозидазы.
Lactobacillus bulgaricus в соответствии с изобретением (выделенные мутанты) представляют пути метаболизма, которые более направлены к пирувату, чем к продуцированию молочной кислоты. Предпочтительно Lactobacillus bulgaricus в соответствии с изобретением выбирают из группы, составленной штаммами Lactobacillus bulgaricus CNCM 1-1348, 1-1349, 1-1350.
Штаммы характеризуются следующими свойствами:
- происхождение: мутант, выделенный из коммерческого штамма L.bulgaricus коллекции Nestle;
- морфология: прямые нежгутиковые Bacilli, без споруляции, Грам+ микроорганизмы, отрицательная каталаза и возможно анаэробные;
- сахаридная ферментация: продуцирование молочной кислоты из:
- D-глюкозы;
- D-фруктозы;
- D-маннозы
- лактозы;
- смешанные;
- уменьшенная активность фермента LDH (лактатдегидрогеназы)
- микроструктурные свойства (продуцирование экзополисахарида).
Мутанты Lactobacillus bulgaricus, имеющие уменьшенную активность LDH, увеличивают продуцирование соединений с приятным запахом и вкусом. Анализ выделенных мутантов путем сравнения активности LDH с активностью β- галактозидазы (отношение LDH/ β -гал) показывает, какой из мутантов является не только с низкой активностью LDH (высокое продуцирование соединений с приятным запахом и низкое продуцирование молочной кислоты), но в действительности показывает, что большая часть углеродного потока (активность β- галактозидазы) переходит в продуцирование соединений с приятным запахом и вкусом.
Настоящее изобретение касается также пищевого состава, содержащего упомянутые Lactobacillus bulgaricus.
Преимущественно упомянутым пищевым составом является нежный йогурт или продукты, похожие на нежный йогурт, содержащий (ие) в соответствии с изобретением Lactobacillus bulgaricus и Streptococcus thermophilus.
Йогурт или продукты, похожие на йогурт, в соответствии с изобретением отличаются увеличенным ароматом и приятным вкусом и уменьшением подкисления и послеподкисления.
Краткое описание фигур
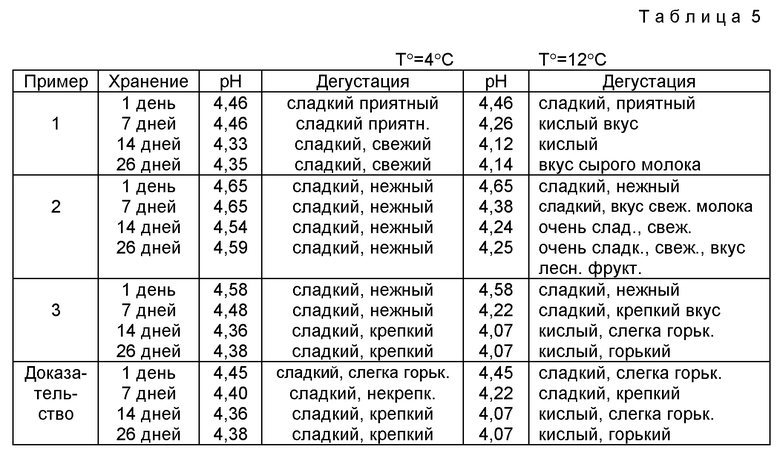
Фиг. 1 представляет поведение роста (• ОД 600, ▿LDH /белок • 100,  L. bulgaricus Lfi 5.
L. bulgaricus Lfi 5.
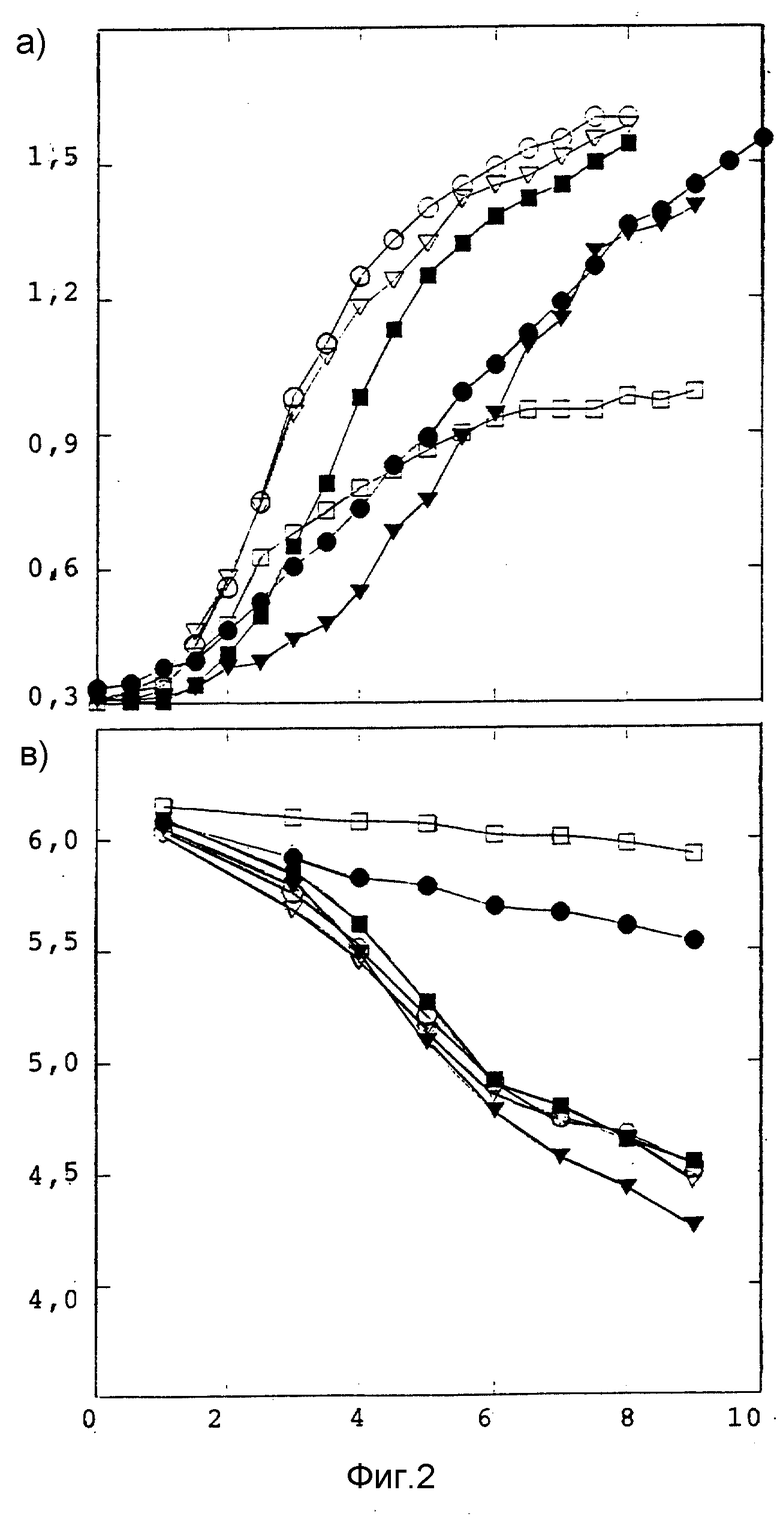
Фиг. 2 (a, b) представляет поведение роста мутантов с низкой активностью LDH на Lfi 5 (oLfi 5,  ).
).
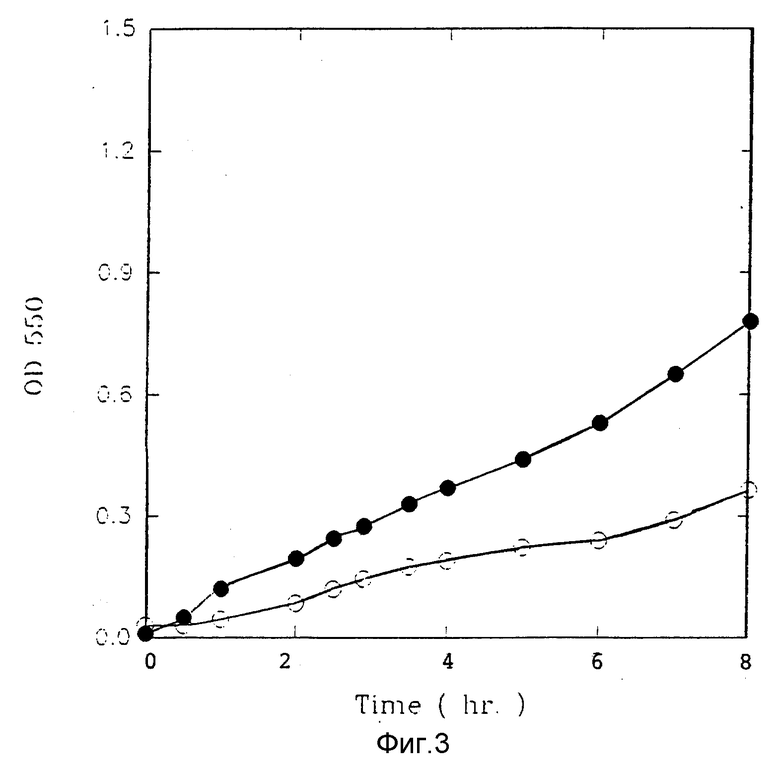
Фиг. 3 представляет воздействие LDH в среде на KTL 50/OKTL 50 без LDH, KTL 50 с LDH).
Описание предпочтительного варианта изобретения
A. Оптимизация мутагенеза на средах Lactobacillus bulgaricus и бактериях
Lactobacillus bulgaricus выращивали в 10 мл Lactobacillus MRS бульона/0,5% дрожжевой экстракт, 1% мясной экстракт, 1% пептон, 0,1% Tween 80, 0,2% цитрат аммония, 0,5% ацетат натрия, 0,2% K2HPO4, 0,01% MgSO4, 0,005% MnSO4/ (x) при 42oC. Чашка с агаровой средой содержала MRS с 15 г агара на 1 л MRS X-Jal чашки с агаровой средой содержали X-Jal (5-бром-4-хлор-3-индолил -β-D- галактопиранозид) при концентрации 25 мг/мл. Мутагенез штамма L. bulgaricus Lfi5 получали с помощью ультрафиолетового света или посредством N-метил-N'-нитро-N-нитрозогуанидина (MNNG) в соответствии со способом Silhavy (T. J. Silhavy, M.L. Berman и W. Enguist (1984), Experiments with genefusions: Cold Spring Harbor Laboratory).
Частота мутации мутагенеза MNNG была в 7 раз выше таковой мутагенеза с помощью ультрафиолетового света.
B. Отсев мутантов с низкой активностью лактатдегидрогеназы
Переночевавшие культуры мутагенезных клеток разбавили, поместили на MRG чашку с агаровой средой и инкубировали при 42oC всю ночь. Отобрали одиночные колонии, которые инокулировали в 250 μл MRS бульона на чашках (FALCON 3072) и выращивали при 42oC всю ночь. 50 μл каждой клеточной суспензии поместили на мембранный фильтр (Dupont Gene Sireen). Фильтр разместили на 3 М фильтре Whatman, насыщенном раствором клеточного лизиса, 1 мг/мл лизицима и 50 μг/мл мутанолизина в воде и инкубировали при 37oC в течение 30 минут. Затем фильтр подвергали действию паров хлороформа в течение 30 минут, воздушной сушке и замораживали при 80oC в течение 30 минут. Затем его промыли 50 мМ трис-хлоридного буфера pH 8,0 для удаления остатков клеток. Промытый фильтр пропитывали в течение 1-4 минут красящим раствором, содержащим 134 мМ натриевой соли D-молочной кислоты, 2 мМ хлорида иодонитротетразолия, 1,4 мМ NAD+, 0,5 мМ N-метилфеназонийметилсульфата, 50 мМ трис-хлорида pH 8,0.
Для прекращения реакции фильтр промыли 0,1 N HCl. Активность лактатдегидрогеназы соответствовала интенсивности (глубине) красного цвета отдельных колоний на чашках. Клетки, показывающие слабые сигналы, переносили в 10 мл свежего MRS бульона и инкубировали при 42oC всю ночь. Клетки собрали центрифугированием и еще раз суспендировали в 1 мл MRS бульона, содержащего 15% глицерина. Суспензии клеток хранили при -80oC.
Приготовление бесклеточных экстрактов
Предварительно нагретые 10 мл свежего MRS бульона, содержащего 2% лактозы, инокулировали 2% переночевавшей культуры и инкубировали при 42oC в течение 7-и часов. Клетки собрали центрифугированием и промыли дважды буфером 50 мМ трис-хлорида pH 8,0, 100 мМ NaCl, 2 мМ EDTA (этилендиаминтетрауксусной кислоты), 1 мМ PMSF, 1 мМ MDTT и еще раз суспендировали в 1 мл буфера. Добавили 100 мл раствора лизоцима концентрацией 10 мг/мл и 50 μл раствора мутанолизина концентрацией 1 мг/мл и инкубировали при 37oC в течение 10 минут. Остатки клеток удаляли центрифугированием при скорости 12000 оборотов в минуту в течение 30 минут.
Испытание лактатдегидрогеназы
Активность лактатдегидрогеназы измеряли посредством последующего уменьшения поглощения при 340 мм из-за превращения NADH в NAD+, в 50 мМ трис-хлорида, pH 7,5. Одну единицу активности фермента определяли как количество, которое окисляло 1 μмоль NADH в минуту при 25oC.
Испытание β-галактозидазы
Активность β -галактозидазы определяли методом Miller (Experiments in Molecular genetics, Cold Spring Habor Laboratory, N.V, 1972). 50 л бесклеточного экстракта добавили в 1 мл Z буфера, 0,1 М фосфата натрия, pH 7,0 10 мМ KCl, 1 мМ MgSO4, 50 мМ 2-меркаптоэтанола и установили равновесие при 28oC. Добавили предварительно нагретые 200 μл ONPG и инкубировали при 28oC в течение 20 минут. Реакцию завершили путем добавления 0,5 мл 1 М Na2CO3. Измерили поглощение OD420. Одну единицу активности фермента определили как количество, которое гидролизовало 1 μ моль ONPG в минуту при 28oC.
Анализ белка
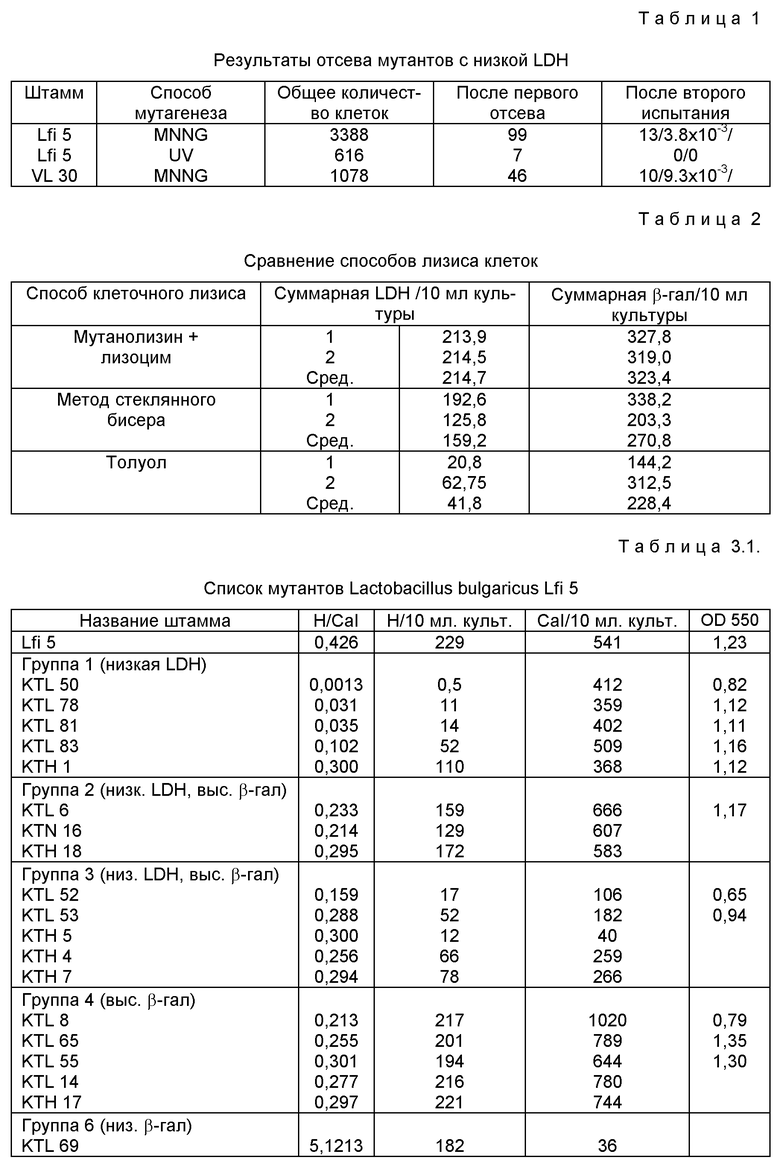
Общую концентрацию белка определяли путем анализа связывания красителя по Bradford (M.M. Bradford (1976), Anal. Biochem. 72: 248 - 254) с использованием комплекта биорадиологии (Bio-Rad). Lactobacillus bulgaricus, Lfi5 и VL30 мутогенизировали как описано ранее (таблица 1). 3388 колоний мутагенизированных MNNG клеток Lfi5, 616 колоний мутагенизированных ультрафиолетовым излучением клеток Lfi5 и 1078 колоний клеток VL30, мутагенизированных MNNG, отобрали и подвергли испытаниям. 152 колонии идентифицировали как мутантов LDH при первом отсеве мембранным испытанием. Фиг. 1 представляет пример такого результата вычисления.
Кандидатов, идентифицированных при этом отсеве, использовали для измерения активности LDH. Каждого кандидата выращивали в среде MRS с 2% лактозы и собирали на ранней стационарной фазе логарифмического роста, что обеспечивало самую высокую активность. Клетки подвергали лизису путем инкубации с муатанолизином и лизоцимом. Другие методы проверяли также. Они суммированы в таблице 2. Метод стеклянного бисера описан Abbondi (M.E. Abbondi S. Pandian, J. Food Sci, 56: 948-953).
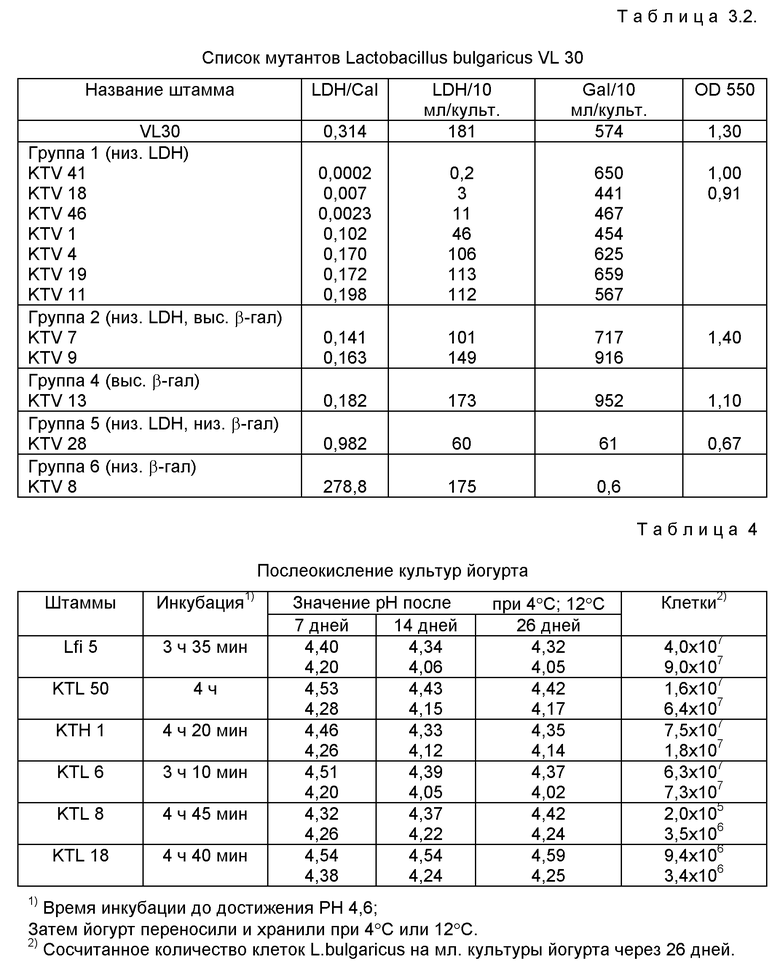
Активность LDH определяли с помощью удельной активности суммарных единиц LDH на суммарный клеточный белок бесклеточного экстракта. Удельная активность LDH непрерывно увеличивалась с ростом клетки (фиг. 2). Поскольку β- галактозидаза является также важным ферментом гликолизного пути обмена, отношение активности LDH к активности β- галактозидазы будет постоянным. В действительности отношение является почти стабильным в период от средней фазы логарифмического роста до ранней стационарной фазы. Мутантов идентифицировали посредством оценки этого отношения, определяли суммарную активность LDH в сравнении с активностью β- галактозидазы. Результаты показаны в таблице 3.
Мутантов классифицировали на 6 групп. Мутанты группы 1 показывали низкую суммарную активность LDH, которая составляла ниже средней на 20%, но при этом сохранялся обычный уровень активности β- галактозидазы. Низкое отношение LDH/β- галактозидаза существовало благодаря только пониженным мутациям LDH.
Группа 2 также имела низкую активность LDH, но активность ее β- галактозидазы была больше средней мутантов дикого типа.
Мутанты групп 1, 2, 3, 4 являются мутантами с низкой LDH. Они имеют низкое отношение LDH/β- галактозидаза и низкие суммарные активности LDH. 13 мутантов Lfi5 и 10 мутантов VL30 распределили по категориям на эти группы. Частота мутации для каждого штамма составила соответственно 3,8 • 10-3 и 8,3 • 10-3.
KTL 50, 78, 81, KTV 41, 18 и 46 имели пониженные отношения LDH/β- галактозидаза (от 1:100 до 1:200) по сравнению со штаммами дикого типа. Кроме того, суммарная активность их β- галактозидазы сохранялась.
Мутанты группы 5 показывали обычную активность. Однако активность их β- галактозидазы увеличивалась.
KTL 50 и KTH 1 группы KTL6 группы 2, KTL 52 и группы 3 и KTL 8 группы 5 исследовали на предмет поведения роста и послеокисления.
C. Характеристики мутантов с низкой LDH
Подкисление мутантов с низкой активностью LDH
Кривая роста:
Мутантов инокулировали на 2% в пробирках с культурой, изготовленных из пирекса (16 х 150 мм) в содержащих 17 мл предварительно нагретого MRS с 2% лактозы, и инкубировали при 42oC. Оптическую плотность OD550 контролировали каждые 30 минут.
Кривая pH:
Мутантов инокулировали на 2% в 10 мл предварительно нагретого MRS бульона с 2% лактозы и инкубировали при 42oC. pH контролировали каждые 30 минут.
Послеокисление мутантов с низкой активностью LDH в смешанных испытаниях культуры йогурта
Исходный материал для пересева состоял из стандартных коммерческих штаммов S. thermophillus, включающих 1,5% Sfi 16, 1,5% Sfi 21 и 0.8% кандидата L. bulgaricus. Йогурт приготавливали из 150 мл пастеризованного молока, 3% снятого молока с 1,5% молочного жира, инокулированного вышеприведенной смешанной культурой. Культуры выращивали при 40oC до достижения pH около 4,6. Затем их охлаждали до 4oC всю ночь и хранили при 4oC или 12oC в течение 26-ти дней. Значение pH, органического свойства и количество клеток контролировали через 7 дней, 14 дней и 26 дней инкубации.
Результаты показаны в таблице 4. Йогурт, хранившийся при 4oC, показал слабое послеподкисление только через 26 дней. Мутанты и штаммы дикого типа сохраняли значение pH выше 4,3. Существовала небольшая разница в значениях pH (около 0,1) у дикого типа и у мутантов с низкой LDH. Культура, хранившаяся при 12oC, показала послеподкисление. В то время как штамм дикого типа Lfi 5 подкислял значение pH до 4,0 через 26 дней значение pH мутантов с низкой LDH, KTL 50 и KTH 1 оставалось все еще равным 4,2.
Штаммы Lactobacillus bulgaricus KTL 1, 18 и 42 зарегистрированы в соответствии с Будапештским договором в Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 28, rue du Docteur Roux, 75024 Paris ledex 15, France под N CNCM 1-1348, CNCM 1-1349 и CNCM 1-1350.
Изобретение будет проиллюстрировано следующими примерами.
Пример 1.
К 500 мл снятого молока в виде порошка добавили 0,1% дрожжевого экстракта и стерилизовали в автоклаве 15 минут при 121oC, затем добавили 10% об. активной культуры штамма L.bulgaricus CNCM 1-1348 (KTL 1), содержащего около 5•108 микроорганизмов/см3.
Путем инкубирования этой смеси в течение 4-х часов при 40oC получили закваску, содержащую около 2,5•108 микроорганизмов/см3.
В соответствии с тем же самым способом получили из активной культуры коммерческого штамма закваску, содержащую около 5•108 растущих S.thermophilus/см3.
К стандартной загрузке молока с содержанием жиров 1,5% добавили 3% снятого молока в виде порошка и пастеризовали в течение 30 минут при 90oC, добавили 1% об. закваски штамма L.bulgaricus CNCM 1-1348 (KTL 1) и 3% об. коммерческого штамма S.thermaphilus
После слабого перемешивания приготовление осуществляли в бидонах и инкубировали в течение 4 часов 20 минут при 40oC до достижении pH 4,62.
Полученный йогурт имел хорошую структуру и очень интересный вкус для этого вида продукта. Было насчитано около 1•109 S.themophilus/см3 и около 1•108 L.Bulgaricus/см3.
Пример 2.
Как и в пример 1 после 8-ми часов инкубации получали закваску L.bulgaricus CNCM 1-1349 (KTL 18), содержащую около 3•108 микроорганизмов/см3.
При добавлении 1% закваски L.bulgaricus CNCM 1-1349 (KTL 18) и 3% закваски S. thermophilus к стандартной загрузке молока (как описано в примере 1) получили йогурт с pH 4,61.
После 4-х часов 40 мин инкубации при 40oC насчитали около 3•108 S.thermophilus и около 1•107 L.bulgaricus/см3.
Этот йогурт с очень маслянистой структурой имел очень интересный вкус свежего молока.
Пример 3.
Штамм L. bulgaricus CNCM 1-1350 (KTL 42) получили в соответствии со способом, описанным в вышеприведенных примерах.
Через 5 часов получили закваску, содержащую около 5•108 микроорганизмов/см3.
В соответствии с примером 1 после 3-х часов 45 мин инкубации получили йогурт с pH 4,62. Насчитали около 1•109 S.thermophilus/см3 и около 2•108 L. bulgaricus/см3.
В дополнение к тому, что йогурт был очень сладким, он имел очень крепкий типичный вкус йогурта. Эти продукты испытывали на сохранение вкуса при 4oC и 12oC, упомянутые испытания включали проверку pH и проверку вкуса соответственно после 1, 7, 14 и 26 дней хранения. Эти результаты представлены в таблице 5.
Сравнительный опыт
В качестве доказательства для получения йогурта в соответствии со способом, описанным в примере 1, использовали первоначальный штамм трех мутантов CNCM 1-1348, CNCM 1-1349 и CNCM 1-1350 (KTL 1, 18, 42).
После 3 часов 45 мин инкубации получили йогурт с pH 4,54, содержащий около 1•109 S.thermophilus/см3 и около 2•108 L.bulgaricus/см3.
Вышеприведенные примеры показывают, что мутанты CNCM 1-1348, CNCM 1-1349 и CNCM 1-1350 (KTL 1, 18 и 42), имеющие уменьшенную активность LDH, могут вырабатывать йогурт с очень интересными структурой, приятным вкусом и последовательными свойствами. Кроме того могут быть получены очень различные вкусы при очень высоких pH.
Изобретение может быть применено в производстве пищевых молочных продуктов, в частности нежного йогурта. Штамм бактерии Lactobacillus bulgaricus CNCM 1-1348 характеризуется отношением лактатдегидрогеназной активности к β-галактозидазной активности менее 0,3 и лактатдегидрогеназную активность менее 150 ед. ферментной активности на 10 мл культуры. Мутированная культура позволяет получать молочные продукты с улучшенными органолептическими свойствами. 3 з.п. ф-лы, 3 ил., 6 табл.
| EP 0518096 A1, 16.12.92 | |||
| Приспособление к химическим пипеткам | 1949 |
|
SU81705A1 |
| Способ регулирования процесса турбинного бурения | 1958 |
|
SU122104A1 |
| Устройство для цементирования обсадных колонн | 1986 |
|
SU1425303A1 |
