Изобретение относится к медицинской технике и практике, в частности, к оториноларингологии, конкретно к моделированию процессов, происходящих в периферическом отделе слухового анализатора.
Известно, что улитка внутреннего уха человека действительно напоминает по форме улитку - конусообразную спираль с 21/2 - 23/4 завитками. Ее высота приблизительно равна 5 мм, диаметр у основания около 9 мм и длина ее средней части около 35 мм. Заключенный в улитку перепончатый лабиринт совпадает с ее формой и имеет длину порядка 32 мм, переменную ширину и толщину. При этом апикальный участок ее ответвлен за восприятие низких звуковых частот, а противоположный (базальный) - за восприятие высоких частот.
Прежние представления о структурах внутреннего уха и их функциях, на которых строились биофизические модели слуха, обладают серьезными недостатками. Эти представления являются настолько упрощенными, что рассматривают улитку в виде всего двух однотипных камер (пространств, полостей, каналов), заполненных оба перилимфой и разделенных так называемой эквивалентной мембраной. Модели улитки не учитывают ни анатомических особенностей структур внутреннего уха, ни физиологических (функциональных) свойств как жидкостей, заполняющих соответствующие пространства внутреннего уха, так и мембран, их разделяющих, ни физических свойств структур внутреннего уха, - и поэтому они не раскрывают реальные биофизические процессы, устанавливающие природу слуха.
Однако на их основе создано две принципиально различных модели слуховой рецепции и достаточно большое число их разновидностей и комбинаций.
Одной из первой явилась модель слуха, которую предложил H.L.F.Helmholtz [1] (излагается по [2, 3]. По H.Helmholtz внутреннее ухо рассматривается состоящим из двух камер, заполненных перилимфой и разделенных вдоль улитки пополам базилярной пластинкой. Процессы, определяющие природу слуха по H. Helmholtz, устанавливают, что механические колебания окна преддверия передаются перилимфе, которая, в свою очередь, вызывает колебания базилярной пластинки. Последняя же, имея частотно-зависимые сегменты, резонирует в ответ на звуковые сигналы только тех частот, которые совпадали с собственной частотой конкретных сегментов. Современной и достаточно полно описывающей многие явления эта модель стала на основена экспериментов G.Bekesy [4] и дальнейших биофизических разработок, полный обзор которых изложен В.С.Шупляковым [5]. Эта модель H.Helmholtz и в соответствии ей построенный механизм слуха до сих пор излагаются практически без вариаций во всех существующих учебниках по физиологии [6 - 11], анатомии [12] и гистологии [13].
Описание механизма слуха по H.Helmholtz испытывает до сих пор несколько затруднений [2, 3]. Одно из них заключается в том, что базилярная пластика должна находиться постоянно в состоянии напряжения, которое в эксперименте [4] не наблюдается. Другая проблема теории слуха по H.Helmholtz заключается в том, что она не может объяснить появление "отсутствующей основной" - восприятия ухом частоты в 100 Гц, отсутствующей физически, в присутствии только тонов в 1100, 1200 и 1300 Гц.
Наиболее известной альтернативной моделью слуха является так называемая телефонная теория, которую предложил W.Rutherford [14] (излагается по [3]). Согласно W. Rutherford улитка не является частотно-зависимой вдоль своей длины: все ее участки реагируют на все частоты сразу и улитковый механизм передает все волновые параметры звукового сигнала в ЦНС, где и совершается анализ звука. Эта теория также испытывает затруднения в объяснении механизма периферической слуховой рецепции. Первая проблема - невозможность передачи сигнала частотой свыше 1 кГц из-за наличия абсолютного рефрактерного периода нервного волокна (длительностью около 1 мс); положительному разрешению этой проблемы способствовало создание принципа залпов, который предложил E.G. Wever [15]. Другая трудность модели состоит в том, что она не способна объяснить "апикально-базаальный эффект", заключающийся в потере уха воспринимать высокочастотную составляющую волнового сигнала при нарушениях апикальной части базилярной пластинки.
Все остальные модели уха и механизмы, объясняющие природу слуховой рецепции, являются комбинированными представленных.
В литературе представлено несколько математических моделей, определяющих частоты звуковых волн, воспринимаемых соответствующими участками (сегментами) базилярной пластинки [9 - 10, 18, 19, 20], но не раскрывающие механизма слуха.
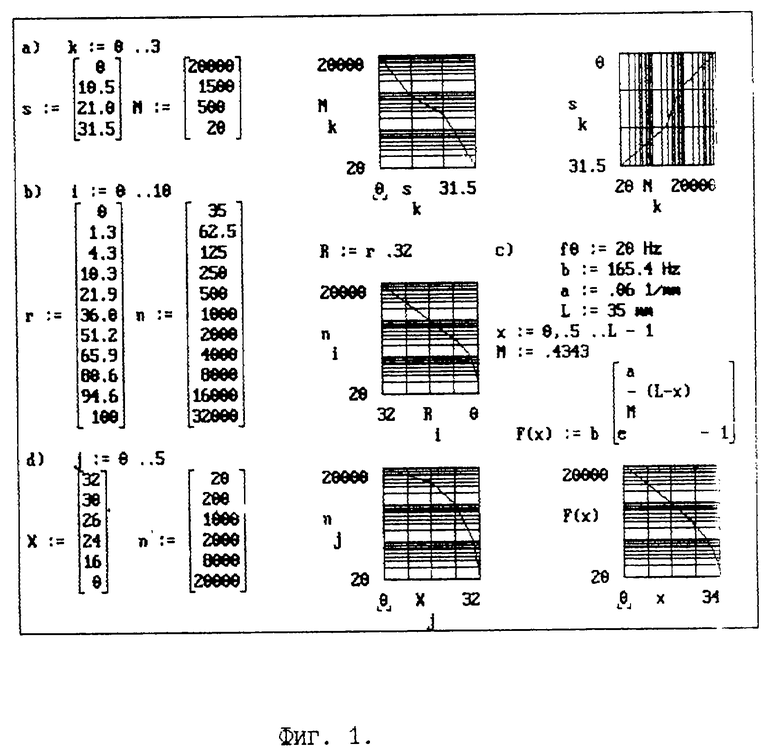
На представленной фиг. 1 дана графическая интерпретация некоторых моделей с использованием компьютерной системы MathCAD [24]. В блоке a) задан цикл выполнения расчета первой модели [9 -10], приведены данные и представлены их графическая интерпретация в виде графика Nk(Sk) - видна достаточно сложная нелинейная зависимость между координатой базилярной пластинки и логарифмом частоты звука.
Если представить данные [9 - 10] в виде зависимости Sk(Nk), то можно заметить, что она напоминает диаграмму соответствия частоты звука (выраженную в герцах) высоте тона (выраженную в мелах) [21]. Очевидно, что данные [9 - 10] устанавливают значения высоты тонов звука, которые были ошибочно отождествлены с частотой звука. Поэтому эта математическая модель, являясь первой, недостаточно проработана биофизически.
Существуют данные, собранные Г.И.Ратниковой по многочисленным источникам [18]. Их математическая обработка приводит к подобным же результатам и поэтому здесь они не приводятся.
В блоке b) приводятся данные так называемой "new frequency scale", или Koenig's scale [19], (излагается по [20], и их графическая интерпретация как ni(Ri). Здесь уже заметна некоторая функциональная зависимость, математическая трактовка которой, проанализированная по [22], представлена в блоке c). Эта модель по сравнению с первой явилась уже хорошим приближением к экспериментам. Однако, кроме аппроксимационного соотношения между частотой звука F(x) и координатой базилярной пластинки x, воспринимающей указанную частоту F, она не позволяет исследовать никакие другие процессы во внутреннем ухе, что заметно умаляет ее информативную значимость. Следует также отметить, что длина базилярной пластинки в этой модели L взята заведомо больше наблюдаемой, равной 32 мм, а экспериментальные данные [19], на основании которых устанавливалось приведенное в блоке c) соотношение [22], даны при усреднении длин базилярных пластинок, что является не совсем корректной процедурой.
В блоке d) приведены экспериментальные данные так называемой Schuknecht's scale [20]. Ее результаты дают зависимость nj(xj), подобную приведенной в блоках b) и c) и отличающуюся от предыдущей и характером, и числовыми значениями, но совпадающими с данными Bekesy [4].
Целью данного изобретения является выявление (на базе анатомо-гистологических представлений о структурах внутреннего уха и их физиологических функциях) биофизических процессов для установления механизма слуха (с использованием явлений дисперсии звуковых волн от поступающего во внутреннее ухо волнового пакета в перилимфатической камере и интерференции звуковых волн в спиральном органе) с последующей их интерпретацией в качестве биофизической (волновой) модели слуха.
В качестве аналогов предлагаемого способа выявления биофизических процессов, устанавливающих механизм слуха человека, взяты модели, предложенные как H.L.F.Helmholtz [1] (с экспериментальными данными H.G.Schuknecht [20] и G.Bekesy [4]), так и W.Rutherford [14].
Сущность изобретения заключается в следующем:
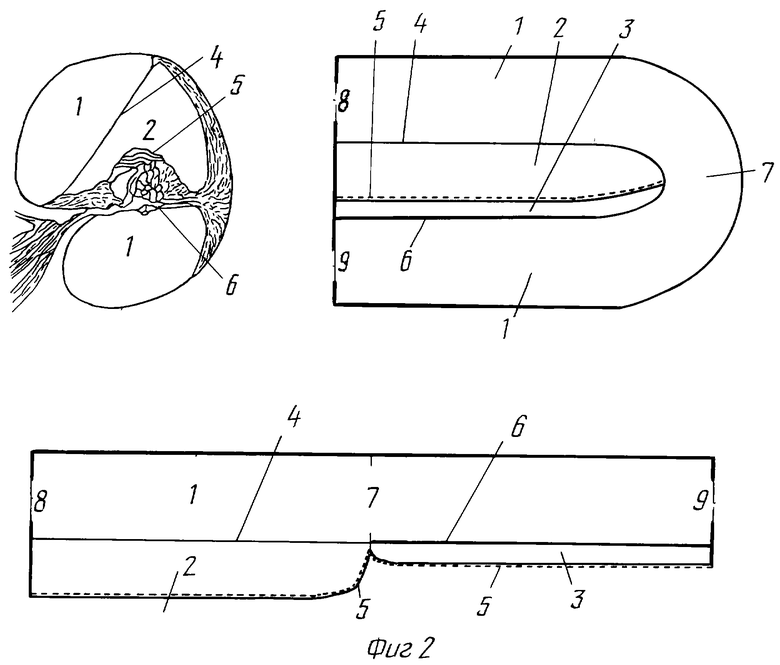
Современные данные об анатомо-гистологических структурах внутреннего уха и их физиологических функций (фиг. 2) свидетельствуют, что внутреннее ухо состоит из трех отдельных камер, заполненных пери-, эндо- и кортилимфой, имеющих различный химический состав и физиологические свойства [16].
Перилимфатическая камера (1), заполненная перилимфой, ограничена каменистой частью костного лабиринта самой улитки и преддверной мембраной (4) со стороны окна преддверия (8) и каменистой частью улитки и базилярной пластинкой (6) со стороны мембраны окна улитки (9) и простирается, соединяясь в апикальной части улитки посредством геликотремы (7) в единый канал, от окна преддверия до мембраны окна улитки.
Эндолимфатическая камера (2), заполненная эндолимфой, ограничена преддверной мембраной (4), той частью костного лабиринта улитки, которая покрыта сосудистой полоской, и отделена от других отделов внутреннего уха системой двух мембран - покровной и сетчатой (5) и зубчатой частью базилярной пластинки (6), покрытой клетками Гензена и Клаудиса.
Кортилимфатическая камера (3), заполненная кортилимфой, ограничена базилярной пластинкой (6) и системой двух мембран (5) - покровной и сетчатой, которую следует рассматривать [17] как достаточно прочное и слитное соединение, надежно отделяющее содержимое этой камеры от других, но также и достаточно мягкое, что позволяет покровной мембране совершать колебательные движения относительно волосковых клеток.
Анатомо-гистологические и физиологические данные о структурах внутреннего уха и их функциях предполагают, что внутрилабиринтные жидкости и разделяющие их мембраны обладают теми физиологическими (функциональными) свойствами, которые обуславливают биофизические процессы, раскрывающие механизм слуха человека, реализуются на использовании эффектов, характеризующих звук как волновое явление.
Первичным эффектом, реализующимся как биофизический процесс во внутреннем ухе человека, является дисперсия звуковых волн в перилимфатической камере, так как при распространении в реальной среде звук имеет тенденцию к диспергированию [4]. При этом в качестве предположения допустимо принять, что промежуток времени, в течение которого звуковые волны различных частот достигают соответствующих координат базилярной пластинки, должен быть одинаковым для волн любой частоты (во избежание ненаблюдаемого в жизни несовпадения одновременно посылаемых в ухо сигналов разных частот).
Это возможно, поскольку существуют такие среды, степень дисперсии в которых достигает 50% [23], каковая, возможно, наблюдается и для перилимфатической камеры, в которой должно выполняться условие lf20/lf20000 = Vf20/Vf20000 = 0,5. В соответствии с экспериментальными данными H.G.Schuknecht [20] скорость распространения звуковых волн V(f) в перилимфатической камере и координата базилярной пластинки l(f) как функции воспринимаемой ухом частоты звука f могут быть представлены следующим распределением по частотам, полученным на основании исследования рекуррентной зависимости между дискретными значениями экспериментальных данных [20]: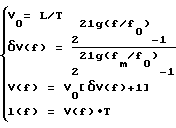 (1)
(1)
где T - время прохождения волнами волнового пакета по перилимфатической камере от окна преддверия до соответствующей координаты l(f), L - длина базилярной пластинки, V0 = L/T - скорость звука для минимальной частоты f0, δV(f) = ΔV(f)/V0 - относительное приращение скорости от V0 до V(f) при изменении частоты от f0 до f, fm - максимальная воспринимаемая ухом частота звук f.
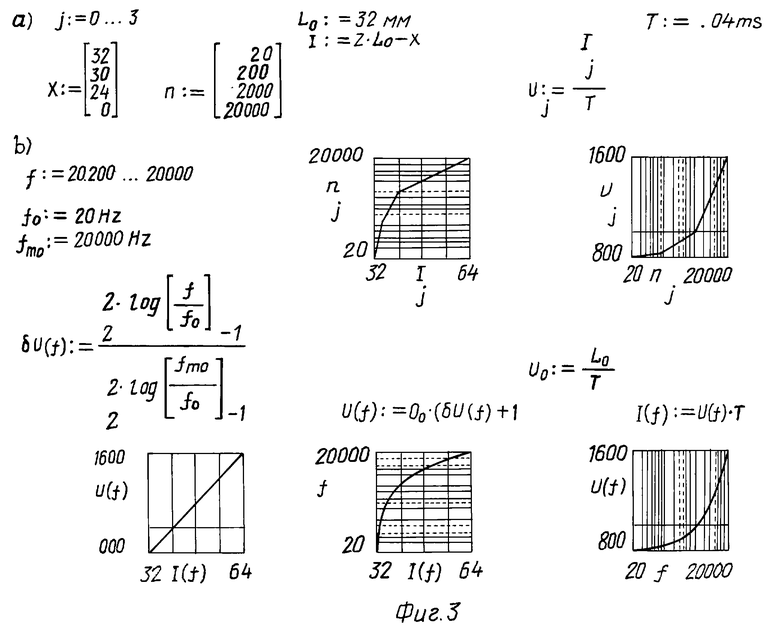
На фиг. 3 представлены результаты обработки для некоторых фиксированных (20, 200, 2000, 20000 Гц) значений экспериментальных данных [20] в соответствии с соотношениями (1), выполненные в системе MathCAD [24] с учетом общей длины X0 = 2L0 перилимфатической камеры (1).
В блоке a) приведены начальные условия: L0 = 32 мм и T = 0,04 мс - время, рассчитанное по формуле для скорости звука V0 = L0/T = 800 м/с для частоты f0 = 20 Гц, и цикл расчета для конкретных частот nj (20, 200, 2000, 20000 Гц), для которых повторены [4, 20] значения координат базилярной пластинки xj, воспринимающие указанные частоты. Приведены преобразование координат x(l) в соответствии с анатомо-гистологическими и физиологическими представлениями о структурах внутреннего уха и их функциях для продольной координаты l(f) в виде соотношения
l(f) = 2L0 - x(f), (2)
формула расчета скорости звуковых волн конкретных частот nj звука, диспергированного в перилимфатической камере
Vj = lj/T. (3)
Этот блок дает графическую интерпретацию расчета экспериментальным данным [4, 20] по формулам (2) и (3). При этом представлены графики тех величин, которые могут быть определены этой моделью: соотношениями между скоростью распространения звуковых волн произвольной частоты, частотой звука и координатой базилярной пластинки, воспринимающей звук этой частоты.
В блоке b) приведены соотношения (1), определяющие математическое представление биофизических процессов, устанавливающих механизм слуха: формула для расчета относительного приращения скорости δV(f) от V0 до V(f) при изменении частоты от f0 до f и координат базилярной пластинки l(f), воспринимающих частоты f с учетом формулы (2). Далее представлена графическая интерпретация расчета модели (1) по указанным формулам.
Сравнение результатов расчета, выполненного по экспериментальным данным [20] и по формулам (1), показывает, что математическое соотношение (1) распределения V(f) и l(f) по частотам для выявленных биофизических процессов, раскрывающих механизм слуха, адекватно экспериментам отражает эффект восприятия ухом соответствующих частот.
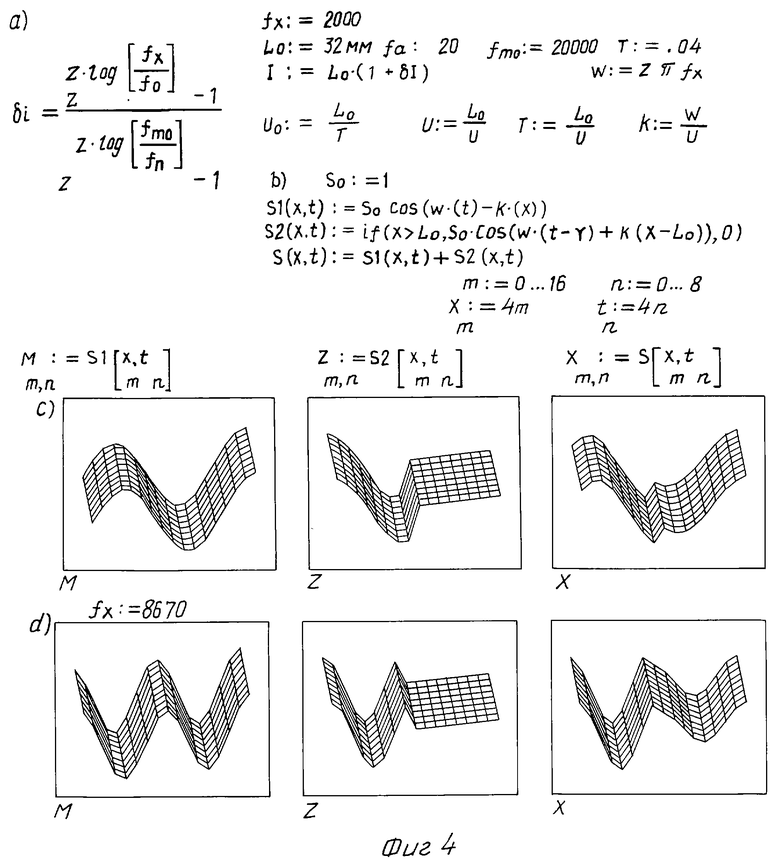
Второй биофизический процесс, определяющий механизм слуха, реализуется образованием на преддверной мембране стоячей волны, как результат интерференции бегущих вдоль нее и отраженных от ее незакрепленного в апикальной части края волн. Расчет показывает, что координаты x(f) интерференционных максимумов для волн с частотой f на преддверной мембране находятся в точках, расположенных симметрично l(f) относительно точки L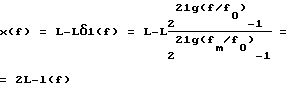 (4)
(4)
На фиг. 4 в блоке a) представлены начальные условия для расчета максимума интерференции бегущей вдоль преддверной мембраны волны s1(x,t), волны s2(x, t), отраженной от ее свободного конца, и результирующей s(x,t) = s1(x, t) + s2(x,t), уравнения которых определены в блоке b). Расчет, выполненный в режиме трехмерной графики системы MathCAD для частот fx = 2000 в блоке c) и 8670 Гц в блоке d), показывает, что действительно отмечается симметрия координат x(f) интерференционных максимумов от преддверной мембраны для волн с частотой fx на покровной и сетчатой мембранах координатам l(f) относительно точки L, причем с ростом частоты f координаты x(f) и l(f) смещаются от апикальной части улитки к окну преддверия и к круглому окну соответственно.
Важно отметить, что роль преддверной мембраны усложняется. Разделяя пери- от эндолимфы и деформируясь при распространении в перилимфатической камере звукового сигнала, она становится способной к созданию волн в эндолимфатической камере. Из-за узости эндолимфатической камеры распределение ее звукового поля можно считать аналогичным результату интерференции для преддверной мембраны. Звуковое поле эндолимфы вызывает колебания системы из покровной и сетчатой мембран, частота которых будет равна частоте исходного звукового сигнала.
Базилярная пластинка, разделяющая пери- от эндо- и кортилимфы, так же как и преддверная мембрана, деформируется при дальнейшем распространении звуковой волны в перилимфатической камере после геликотремы и вызывает колебания самих волосковых клеток.
Третий тип биофизических процессов, протекающих во внутреннем ухе и приводящих к генерации нервного импульса, - суперпозиция однонаправленных колебаний в кортилимфатической камере, создаваемых базилярной пластинкой и системой из покровной и сетчатой мембран, приводящая в соответствующей координате, определяемой соотношениями (1, 4), к максимуму механического воздействия на нервные окончания волосковых клеток (максимуму деформации их сдвига) и, как следствие, - к генерации нервного импульса, вызывающего слуховые ощущения.
Описанные дисперсионные и интерференционные процессы, происходящие в камерах внутреннего уха, а также возможность замены спиралевидной формы улитки на линейно-протяженную [21] и представление перилимфатического канала как единого волновода, - составляют совокупность биофизических процессов, устанавливающих механизм слуха человека, а соотношения (1, 4) дают их математическую интерпретацию.
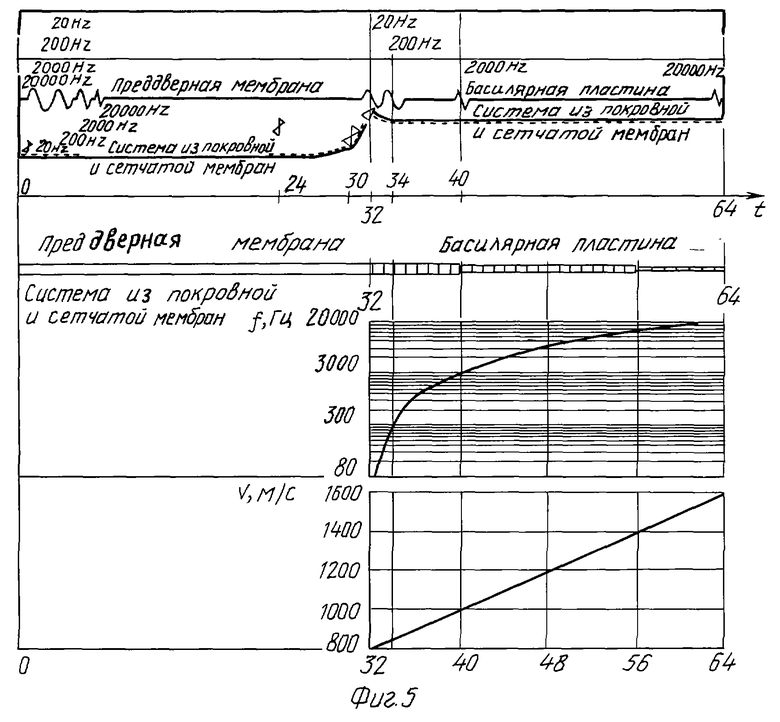
На фиг. 5 дано иллюстративное представление этих процессов как биофизической модели, определяющей механизм слуха человека: приведены схема строения внутреннего уха с отображением анатомо-физиологических представлений о его строении и функциях его структур (с указанием положения в пространстве преддверной мембраны, базилярной пластинки и системы из покровной и сетчатой мембран) и ее биофизические характеристики (зависимости от координаты базилярной пластинки частоты воспринимаемого ею звукового сигнала и скорости распространения звуковых волн этих частот).
Установленные биофизические процессы, определяющие механизм слуха человека, выявленные на основе анатомо-гистологических и физиологических представлений о структурах внутреннего уха и их функциях, и математическая их интерпретация, как биофизическая модель механизма слуха, объясняют феномены моделей, которые предложили H.Helmholtz [1] и W.Rutherford [4].
Они отражают принцип сегментации базилярной пластинки, ответственной за восприятие звуковых сигналов конкретных частот, который предложил H.Helmholtz [1], но совершенно иначе объясняют механизм слуха.
Здесь прослеживаются также и идеи телефонной теории, которую высказал W. Rutherford [14] и по которой звуковые волны любой частоты можно обнаружить в любой точке перилимфатического пространства. Это действительно возможно, но нервные окончания волосковых клеток, в результате суперпозиции колебаний системы покровной и сетчатой мембран и базилярной пластики в кортилимфатической камере, реагируют с генерацией нервного импульса только на волны тех частот, которые удовлетворяют соотношение (1).
В отличие от модели H.Helmholtz [1] предлагаемый механизм слуха не требует постоянного напряжения базилярной пластинки (в соответствии с воззрениями G.Bekesy [4]), так как базилярная пластинка испытывает напряжение только в случае максимума интерференции именно для координат l(f) тех своих участков, которым соответствуют частоты, присутствующие в данном волновом пакете.
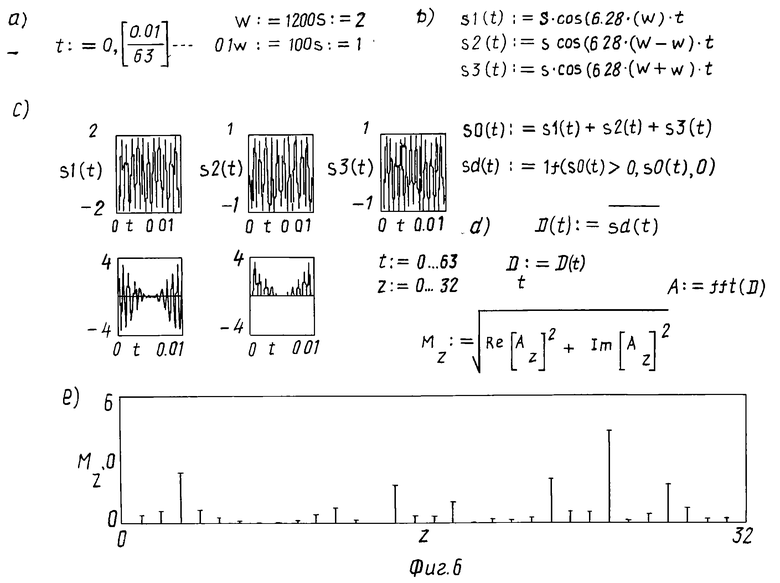
Для объяснения появления "отсутствующей основной" в данной модели рассмотрим результат сложения трех гармоник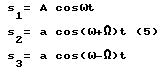 (5)
(5)
Психоакустический и физиологический подход [3] восприятия ухом тона, соответствующего частоте 100 Гц и отсутствующего в исходном сигнале, объясняет этот эффект восприятием двух равных комбинационных (первичных разностных) тонов с равными частотами Ω
[ω+Ω-ω] = [ω-(ω-Ω)] = Ω = 100 Гц,
которые, вследствие их суперпозиции, дают хорошо воспринимаемый низкий тон, соответствующий указанной частоте Ω.
Биофизический анализ интерференции трех данных гармоник с Ω = 100 Гц и ω = 1200 Гц, распространяющихся в одном направлении, показывает, что результирующее колебание s0 имеет вид
s0= s1+s2+s3= A(1+2a/AcosΩt)cosωt (6)
При Ω ≪ ω (что здесь и наблюдается) результирующее колебание s0 представляет собой модулированное колебание с условным периодом
T0= 2π/ω, (7)
переменной амплитуды
S0,max= A(1+2a/AcosΩt),
период изменения которого
T = 2π/Ω, (8)
Огибающая этого колебания воспроизводится преддверной мембраной и создает в эндолимфатической камере дополнительную волну с частотой Ω, воздействующую на систему покровной и сетчатой мембран и вызывающую ее колебания. Волна той же частоты создается и базилярной пластинкой в перилимфатической камере после прохождения ею геликотремы.
В результате суперпозиции этих колебаний от системы покровной и сетчатой мембран и от базилярной пластинки в кортилимфатической камере производится максимум воздействии на нервные окончания волосковых клеток с генерацией нервного импульса и возможностью уха воспринимать звук, соответствующий не только наличествующим физически частотам в 1100, 1200 и 1300 Гц, но и "отсутствующей" в 100 Гц.
На фиг. 6 в блоках a), b) и c) представлены формулы и вид всех трех гармоник s1, s2, s3 и расчет результирующего колебания s0, а также в блоках d) и e) результат спектрального Фурье-разложения, выполненное в системе MathCAD [24] . При этом оказывается, что результирующее колебание имеет не только составляющие в 1100, 1200 и 1300 Гц, но несколько других составляющих, с меньшими частотами, в том числе и отсутствующую частоту в 100 Гц достаточно большой амплитуды и поэтому воспринимаемой ухом значительнее остальных.
В этом случае следует признать, что ухо способно воспринимать как несущую частоту ω, так и частоту модуляции Ω, соответствующую комбинационному (первичному разностному) тону.
Суперпозиця волн в кортилимфатической камере способна объяснить механизм самозащиты внутреннего уха от чрезмерных механических нагрузок при воздействии звуков большой интенсивности, Интерференционный аспект на преддверной мембране способен объяснить.
Важно отметить, что выявленные биофизические процессы, устанавливающие механизм слуха, и их математическая интерпретация, как биофизическая (волновая) модель слуха, могут объяснить природу апикально-базального эффекта, возможность самозащиты уха при звуках большой интенсивности, эффекты маскировки звука иметь другие практические приложения. Этим существенно повышается информативность как самой биофизической модели, так и основанных на ней аудиометрических методов исследования внутреннего уха.
Литература
1. Helmholz H.L.F. Die Lehre von Tonempfindungen. Draunschweig. - 1863. - (Рус. пер. Петрова. - СПб. - 1875).
2. Руководство по оториноларингологии. /Под ред. И.Б. Солдатова. - М.: Медицина. - 1994.
3. Гельфанд С. А. Слух: введение в психологическую физиологическую акустику. /Пер. с англ. - М.: Медицина. - 1984.
4. Bekesy G. Experiments in Hearing. - N.J.: McGraw - Hill. 1960.
5. Шупляков В.С. //Сб.: Сенсорные системы. Слух. - Л.: Наука. - 1982. - С. 3-17
6. Физиология человека /Под ред. Г.И. Косицкого. //Изд. - 3. М.: Медицина. - 1985.
7. Основы физиологии. /Под ред. П. Стерки. - М.: Мир. - 1984.
8. Физиология человека: Compendium. /Под ред. Б.И. Ткаченко и В.Ф. Пятина. - Спб. - 1996.
9. Human Anatomy and Physiology. /3 Ed. E.N. Marieb. - The Benjamin Commings Publishing Company, Inc. - California. - 1995.
10. The World's Best Anatomical Charts. /Anatomical Chart Co. - Skokle, IL - 1993.
11. Тамар Г. Основы сенсорной физиологии. /Пер. с англ. - М.: Мир. - 1976
12. Привес М. Г. , Лысенко Н.К., Бушкович В.И. Анатомия человека. /Под ред. М.Г. Привеса. - М.: Медицина. - 1985.
13. Гистология. /Под ред. В.Г. Елисеева, Ю.И. Афанасьева, Е.А. Юриной. - М.: Медицина. - 1983.
14. Rutherford W. //J. Anat. Physiol. 1986. - V. 21. - P. 166-168.
15. Wever E.G. Theory of Hearing. - N.J.: Dover. - 1949.
16. Rauch C., Kostlin A. //Pract. Otol. - 1958. - V. 20. - P. 287-291.
17. Laurence M. //Laryngoscope, - 1966. - V. 76. - P. 1318-1337.
18. Гершуни Г.В. //Механизмы слуха. М.: Наука. - 1967. - С. 3 - 32.
19. Koenig W. A new frequency scale for acoustic measurements. Bell Laboratory Record. - 1949.
20. Schuknecht H. F. Pathology of the Ear. A Commonwealth Fund Book. / Harward Univ. Press. - Cambridgem Massachusets. - 1974.
21. Stevens S.S., Volkman J. // Amer. J. Psych. -1940. -V.53. - P. 329 - 353.
22. Физиология человека . Т.2: Органы чувств / Пер. с анг. под ред. П.Г. Костюка. - М.: Мир. - 1985.
23.Полякова А.А. // БСЭ. - Т.8. - 1972. - С. 306 - 307.
24. Дьяконов В.П. Система MathCAD. - М: Радио и связь. - 1993.
Способ может быть использован в медицине, а именно в оториноларингологии. Выявляют дисперсию звука в прелимфатической камере. Выявление проводят на базе анатомо-гистологических структур внутреннего уха и их физиологических функций по соотношению распределения скорости звука по частотам. Выявляют образование стоячих волн на пред-дверной мембране в результате интерференции бегущих вдоль нее и отраженных от ее свободного у геликотремы края волн на основании рекуррентной зависимости между дискретными значениями экспериментальных данных. Выявляют соотношения распределения скорости звука и координат базилярной пластинки по частотам при максимуме механического воздействия на нервные окончания волосковых клеток, генерирующих нервные импульсы. Выявленные соотношения соответствуют частоте исходного звукового сигнала. Способ обеспечивает точность выявления. 6 ил.
Способ моделирования биофизических процессов, реализующих механизм и биофизическую (волновую) модель слуха человека, включающий выявление анатомо-гистологических структур внутреннего уха и их физиологических функций, отличающийся тем, что выявляют дисперсию звука в перилимфатической камере на базе анатомо-гистологических структур внутреннего уха и их физиологических функций по соотношению распределения скорости звука по частотам, образование стоячих волн на преддверной мембране в результате интерференции бегущих вдоль нее и отраженных от ее свободного у геликотремы края волн на основании рекуррентной зависимости между дискретными значениями экспериментальных данных, соотношения распределения скорости звука и координат базилярной пластинки по частотам при максимуме механического воздействия на нервные окончания волосковых клеток, генерирующих нервные импульсы, соответствующие частоте исходного звукового сигнала.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Руководство по отоларингологии | |||
| /Под ред | |||
| СОЛДАТОВА И.Б | |||
| - М.: Медицина, 1994, с | |||
| Мяльно-трепальный станок для обработки тресты лубовых растений | 1922 |
|
SU200A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| ГЕЛЬФАНД С.А | |||
| Слух: введение в психологическую и физиологическую акустику | |||
| - М.: Медицина, 1984, с | |||
| Приспособление для уменьшения дымовой тяги паровоза | 1920 |
|
SU270A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| ГЕРШУНИ Г.В | |||
| Механизмы слуха | |||
| - М.: Наука, 1967, с | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

