Изобретение относится к рекомбинантной нейраминидазе гриппа, вектору экспрессии, с помощью которого рекомбинантная нейраминидаза может экспрессироваться в клетках-хозяевах методом продуцирования и очистки рекомбинантной нейраминидазы, вакцинам против гриппа и использованию рекомбинантной нейраминидазы в соответствии с настоящим изобретением.
Эпидемии гриппа, вызываемые вирусами А и Б, приводят к значительному дискомфорту пораженных ими людей и оказывают очень большое воздействие на социальную и экономическую жизнь. Грипп является причиной высокого уровня смертности у пожилых людей и пациентов, страдающих хроническими заболеваниями. С момента применения в 40-х годах нашего столетия инактивированные вакцины на основе вирусного материала, выращенного в куриных эмбрионах, показали свою очевидную эффективность против гриппа и привели к значительному сокращению смертности людей, входящих в группу риска.
Вирусы гриппа уникальны среди других вирусов, развивающихся в бронхиолах, поскольку они претерпевают значительные антигенные изменения (так называемый "дрейф") в двух их поверхностных антигенах, а именно гемагглютинине (HА) и нейраминидазе (NА).
Кроме того, грипп А благодаря феномену "сдвига" (shift) способен обходить распространенный иммунитет. Возникающий у людей вирус представляет собой ген нейраминидазы, который появляется из животных источников генов гриппа. В 1957 году наиболее распространенный в то время вирус типа NA1 сменился новым вирусом типа NА2. С 1977 года вирусы типа NА1 вновь вернулись в популяцию человека. Применяемые в настоящее время вакцины должны поэтому предпочтительно быть направлены против обоих типов вирусов NA1 и NА2.
Нейраминидаза катализирует удаление концевых остатков сиаловой кислоты гликозильных групп, при этом разрушаются потенциальные рецепторы для гемагглютинина (Gottschalk 1957; Burnet and Stone, 1947). Считается, что нейраминидаза играет важную роль, препятствуя агрегации вируса и подавляя его способность распространяться из клетки в клетку (Colman and Ward, 1985).
Каждая молекула нейраминидазы (Мr = 240000) имеет структуру, которая напоминает поганку и состоит из четырех идентичных полипептидных цепочек, образованных двумя димерами, которые связаны дисульфидными мостиками и далее скрепляются друг с другом нековалентными связями (Bucher and Kilbourne, 1972; Laver and Valentine, 1969; Vargese et al., 1983; Ward et al., 1983). В отличие от гемагглютинина нейраминидаза закрепляется в липидной мембране с помощью неподвергнутой сплайсингу NА-концевой липофильной последовательности (Fields et al. , 1983; Вlоск et al., 1982), так называемого мембранного якоря. Наибольшая часть всей структуры выступает за мембрану и образует там периферическую "головную" часть, имеющей форму коробочки, которая размещается поверх удлиненной области "стебля" (Wrigley et al., 1973). Внутри головки содержится мономер, который имеет свой собственный каталитический сайт и содержит по крайней мере четыре NА-связанные гликозильные группы (Colman et al. , 1983; Ward et al., 1982). Наличие O-гликозилирования было установлено совсем недавно.
Вследствие их внешнего расположения антигены гемагглютинин и нейраминидаза являются наиболее важными мишенями в структуре вируса для иммунной системы хозяина. Что касается антигенов, которые специфично связываются с гемагглютинином, то считается, что они нейтрализуют инвазивную способность вируса, вероятно путем блокирования ранних стадий инфекции (Hirst, 1942; Kida et al., 1983). Нейраминидаза-специфичные антитела обычно не препятствуют первичной инфекции клетки-мишени (Jahiel and Kilbourne, 1966; Kilbourne et al. , 1968; Johanssen et al., 1988), а блокируют именно распространение вируса. Далее, вследствие протекания конкурентных процессов иммунный ответ на нейраминидазу, видимо, частично подавляется в пользу более часто встречающегося антигена гемагглютинина (Johanssen et al., 1987; Kilbourne, 1976). В итоге воздействие иммунитета нейраминидазы обычно затмевается антителами, нейтрализующими гемагглютинин. По этой причине внимание разработчиков вакцины в течение длительного времени было почти исключительно сфокусировано на гемагглютинине.
Однако ряд экспериментальных наблюдений указывает на то, что нейраминидаза на самом деле способна играть важную роль в создании защитного иммунитета против гриппа (Schulman et al., 1968; Johansen and Kilbourne, 1990; Johansen et al. , 1993). Фундаментальные исследования иммуногенного потенциала нейраминидазы требуют очень чистых антигенов в достаточном количестве и с правильной трехмерной структурой. До настоящего времени нейраминидазу получают обработкой оболочек вируса детергентами (Gallgher et al., 1984; Kilbourne еt al., 1968) или протеолитическим расщеплением головки белка, часто с помощью проназы (Seto et al., 1966; Rott et al., 1974), с последующей очисткой нейраминидазы. Хотя эти методы в какой-то степени применимы, они имеют значительные ограничения с точки зрения выхода и чистоты продукта.
В связи с этим целью настоящего изобретения является рекомбинантная нейраминидаза гриппа, которая обладает антигенными свойствами, соответствующими свойствам природной нейраминидазы, и корректно свернута.
В соответствии с настоящим изобретением указанная рекомбинантная нейраминидаза в практически очищенной форме может быть получена:
а) культивированием в культуральной среде клеток-хозяев, трансформированных с помощью вектора экспрессии нейраминидазы или инфицированных вирусом, который трансформирован с помощью вектора экспрессии нейраминидазы, при этом вектор экспрессии включает по крайней мере часть кодирующей области гена нейраминидазы вируса гриппа минус область, кодирующая мембранный якорь, или ее модифицированный вариант, которой предшествует присоединенная в фазе сигнальная последовательность; и
б) выделением продуктов экспрессии нейраминидазы из культуральной среды.
Рекомбинантная нейраминидаза по настоящему изобретению, секретируемая в культуральную среду, может, например, использоваться в фундаментальных исследованиях, в которых проводится отдельная вакцинация нейраминидазой, с целью определения роли нейраминидазы в вакцине. На практике рекомбинантная нейраминидаза вероятно все еще будет использоваться в сочетании с гемагглютинином, чтобы увеличить степень защиты (процента получившего прививки населения, которое эффективно защищено от инфекции) и устойчивость защиты (защиты от более поздних эпидермических штаммов).
В настоящем изобретении, в частности, заявляется рекомбинантная нейраминидаза гриппа NА2, которую можно получить выращиванием клеток-хозяев в подходящей для выращивания питательной среде и выделением продукта экспрессии нейраминидазы из питательной среды. На практике эта процедура заключается в переносе рекомбинантного модуля экспрессии из рАс21VNАS в бакуловирус дикого типа или его производное. Клетки-хозяева затем инфицируют с помощью этого рекомбинантного бакуловируса.
Клетки-хозяева, которые используются для получения рекомбинантной нейраминидазы гриппа, преимущественно получают из низших эукариотных организмов, таких как насекомые, преимущественно из клеточной линии sf9 насекомых, однако могут также использоваться дрожжевые клетки, такие как Saccharomyces или Pichia.
Настоящее изобретение относится также к двух векторам для экспрессии способной секретироваться нейраминидазы гриппа, содержащим сайт инициации репликации, по крайней мере часть кодирующей области гена нейраминидазы гриппа минус область, которая кодирует мембранный якорь, или ее модифицированный вариант, сигнальную последовательность, расположенную в 5'-положении от кодирующего района и связанную с ним в фазе, промотор, расположенный в 5'-положении от сигнальной последовательности, и терминатор транскрипции, расположенный в 3'-положении от кодирующей области. В изобретении, в частности, заявляется вектор для экспрессии способной секретироваться нейраминидазы гриппа NA2, который содержит сайт инициации репликации, кодирующую область гена штамма гриппа A/Victoria/3/75 минус область, кодирующая мембранный якорь, или ее модифицированный вариант.
Для экспрессии в клетках насекомых указанный вектор помещают в клетку вместе с бакуловирусом дикого типа или его производным. Вследствие двойной гомологичной рекомбинации получают рекомбинантный бакуловирус, при этом модуль экспрессии из вектора вводится в геном вируса. После очистки бляшек получают запас рекомбинантных бакуловирусов, который затем можно использовать, например, для инфицирования клеток sf9.
Предпочтительно используют сигнальную последовательность гена гемагглютинина вируса A/Victoria/3/75 (Н3N2) гриппа NA2. Изобретение преимущественно включает вектор рАс21VNАS, зарегистрированный 3 января 1994 года в Коллекции плазмид Лаборатории молекулярной биологии (Laboratorium voor Moleculaire Biologie - Plasmidencollectie (LMBP)) по адресу: K.L. Ladeganckstraat 35, B-9000 Ghent, Belgium, и получивший при депонировании регистрационный номер LMBP 2976, который используют для трансформации вируса, такого как, например, бакуловирус, с помощью двойной гомологичной рекомбинации. В соответствии с настоящим изобретением модуль экспрессии вектора, включающий сигналы регуляции транскрипции, сигнальную последовательность и кодирующую область, помещают в геном вируса.
В еще одном варианте осуществления настоящего изобретения используют второй вектор по настоящему изобретению. Указанный вектор рассчитан на использование в дрожжах и включает, например, сайт инициации репликации, кодирующая область гена нейраминидазы штамма A/Victoria/3/75 гриппа NА2 минус области, которые кодируют соответственно мембранный якорь и стеблевую часть нейраминидазы, или ее модифицированные варианты, сигнальную последовательность, расположенную в 5'-положении от кодирующей области и связанную с ней в фазе, промотор, расположенный в 5'-положении от сигнальной последовательности, и терминатор транскрипции, расположенный в 3'-положении от кодирующей области.
Последовательности промотора и терминатора предпочтительно являются гомологичными и выделяются из метилотрофных дрожжей Pichia pastoris таких как последовательности гена алкогольоксидазы 1. В сигнальной последовательности, например, располагается сигнал секреции препро-фактора, спаривания α Saccharomyces cerevisiae.
Указанный вектор pPP1IVNAf1s зарегистрирован 3 января 1995 года в Laboratorium voor Moleculaire Biologie - Plasmidencollectie (LMBP), K.L. Ledeganckstraat 35, B-9000 Ghent, Belgium, и получил при депонировании регистрационный номер LMBP 3223.
Рекомбинантная нейраминидаза по настоящему изобретению оказалось способна генерировать защитный иммунитет против вирусов гриппа, в частности вирусов гриппа типа NA2. Изобретение относится также к вакцине против гриппа, которая содержит рекомбинантную нейраминидазу.
Кроме того, изобретение относится к способу получения рекомбинантной нейраминидазы и к способу ее очистки.
В описании настоящего изобретения и в формуле термин "NAS" обозначает секретируемую (рекомбинантную) нейраминидазу; "рNА" относится к природной нейраминидазе, обработанной проназой; "NA" обозначает нейраминидазу.
Настоящее изобретение далее поясняется приведенными ниже примерами, которые приводятся лишь для иллюстрации, а не для ограничения объема притязаний по настоящему изобретению.
Пример 1. Экспрессия, очистка и изучение свойств рекомбинантной нейраминидазы гриппа NА2
Материалы и методики
1. Конструирование гена, кодирующего секретируемую нейраминидазу, и его интегрирование в систему экспрессии бакуловируса
а. Плазмиды
Плазмида рV6/21 является производной pBR322, содержащей копию гена нейраминидазы вируса A/Victoria/3/75 гриппа (Van Rompuy et al., 1982). pSV 51 и как pSV 23m, так и рS 24m представляют собой соответственно поздний и ранний SV40-замещенные секторы и описываются в других местах (Huylenbroeck et al., 1988). pSRS-8 представляют собой плазмиду на основе pPLa2311, которая содержит стартовую последовательность гена гемагглютинина (NА) из A/Victoria/3/75 (Н3N2) (Huylenbroeck et al., 1988). Бакуловирусный трансферный вектор pVL941 разработали Luckow and Summers (1989).
b. Субклонирование сигнальной пептидной последовательности гемагглютинина (pIV-рrеНА)
Bst NI фрагмент pSR S-8 размером 1830 пар оснований вставляют с помощью фермента Кленова. Далее располагают Pvu II-линкеры (GCAGCTGC). Полученный фрагмент и pBR322 расщепляют Pvu I и Pvu II и выделяют фрагменты размером соответственно 731 пара оснований и 1699 пар оснований. Их лигированием получают pIv рrеНА, которая содержит 5'-нетранслируемую область гена гемагглютинина, начиная с G16 (АТG = +1, +2, +3), за которой следует интактная сигнальная последовательность гемагглютинина и первые несколько кодонов зрелого гемагглютинина.
с. Конструирование химерной последовательности, которая кодирует способную секретироваться нейраминидазу: pATIVNAS
pV 6/21 раскрывают с помощью Pvu I и обрабатывают экзонуклеазой Ва131. К этой смеси лигируют Hind III-линкеры, а затем расщепляют Hind III. Фрагмент нейраминидазы размером приблизительно 1500 пар оснований отделяют и клонируют в уникальный Hind III - сайт pSV 23m (рSV 23mIVNA). Плазмиды со вставкой в противоположной ориентации затем расщепляют Fnud и Sal I и выделяют фрагмент размером 1291 пара оснований, содержащий ген нейраминидазы минус последовательность мембранного якоря. Затем pIVpreNA инкубируют с Рst I и Pvu II и получают В61br - фрагмент с сигнальной последовательностью гемагглютинина. Оба фрагмента в конце концов сливают методом лигирования по тупым концам Pvu II - Fnud II и встраивают во фрагмент Sal I/Pst I размером 2253 пары оснований из рАТ153, получая pATIVNAs. Эта плазмида содержит последовательность, которая кодирует сигнальную последовательность гемагглютинина, включая первые несколько аминокислот зрелого гемагглютинина, за которыми непосредственно следует последовательность нейраминидазы, в которой отсутствует сигнальный пептид/мембранный якорь и часть области, кодирующей "стебель". Лигирование фрагментов гемагглютинина и нейраминидазы приводит к замене лишь одной аминокислоты (Gly на Аlа), которая соответствует позиции 5 в зрелом гемагглютинине. На основании информации, опубликованной Minjou et al. (1988) и Van Rompuy et al. (1982), предсказанные последовательности ДНК и аминокислотных остатков, фланкирующие места лигирования в нейраминидазе, показаны на фиг. 2.
d. Интеграция нейраминидазы в бакуловирусный вектор переноса
XbaI/SalI фрагмент из pATIV NAS размером 1368 пар оснований лигируют с фрагментом SаlI/ЕсоRI из рSV 51 размером 5562 пары оснований и фрагментом EcoRI/XbaI из рSV24m размером 624 пары оснований. Копию фрагмента BamHI размером 1647 пар оснований, содержащего ген нейраминидазы и поли(А)-сайт SV40, затем встраивают в единственный сайт рестрикции pVL941, учитывая правильную ориентацию по отношению к промотору полиэдрина, и получают pAc21VNAS. Эта конструкция обеспечивает гомологичную рекомбинантную с ДНК AcNPV дикого типа после котрансфекции клеток sf9. Потомство рекомбинантного вируса выделяют последующей очисткой бляшек, как описано Summers and Smith (1987).
2. Культура клеток насекомого - продукция нейраминидазы
Обычную культуру sf9 клеток насекомого поддерживают в виде конфлюентных клеточных монослоев в среде ТС100, содержащей 10% сыворотки эмбриона теленка и 50 мкг/мл гентамицина. Для проведения инфекции рекомбинантным бакуловирусом культуры переносят в суспензии объемом 200 мл, которые выращивают в роллер-флаконах емкостью 850 мл (25 об/мин). Затем суспензии клеток по окончании их лог-фазы (2•106 клеток на миллилитр) инфицируют рекомбинантным бакуловирусом при moi ("множественности заражения", т.е. количестве инфекционных вирусных частиц на клетку), равной 1,0. Через два часа инфицированные клетки переносят в свежую порцию среды ТС100, не содержащую сыворотку, и инкубируют в суспензию в течение 48 часов. Нейраминидазу очищают из культуральной жидкости, как описано ниже.
3. Выращивание вируса Х-47 гриппа
Штамм Х-47 гриппа используют в качестве источника для получения природной нейраминидазы A/Victoria/3/75 после обработки проназой. Вирус Х-47 выращивают в полости желточного мешка 11-дневного куриного эмбриона. После двух дней выдерживания в термостате при температуре 25,5oС яйца оставляют на ночь охлаждаться до 4oС и собирают жидкость желточного мешка для дальнейшей переработки.
4. Буферные системы
Обычно применяют следующие буферные растворы:
буфер А: 20 мМ диэтаноламин/HCl с pH 8,5;
буфер В: 50 мМ NaAc с рН 5,5;
буфер С: 10 мМ NaP с рН 7,4, 150 мМ NaCl;
буферы А и С дополнительно содержат 4% бутанола (если специально не оговаривается) и 2 мМ хлорида кальция.
5. Очистка рекомбинантной нейраминидазы
а. Фракционирование с помощью сульфата аммония
Суспензии культуры sf9 (обычно около 1 литра) собирают после заражения (см. выше) и остатки клеток осаждают центрифугированием при 4000•g в течение 15 минут. Все остальные операции проводят при температуре 4oС. На первой стадии очистки к растворам добавляют 5 мМ раствор азида натрия. Осветленную сырую среду подвергают фракционированию с помощью сульфата аммония при рН 7,5. Вещество, которое осаждается в интервале концентраций от 20% до 60% сульфата аммония, отделят центрифугированием (10000•g, 60 минут) и растворяют в буфере А (без бутанола) + 20 мМ хлорида натрия в количестве, равном одной десятой от первоначального объема. Вновь растворенный осадок подвергают в течение 24 часов диализу (с молекулярным весом отсечки, равным 25 килодальтон) против 50 объемов того же самого буфера в течение 24 ч, при этом буфер трижды меняют. Нерастворимые компоненты удаляют центрифугированием при ускорении 20000•g в течение 15 минут.
b. Анионообменная хроматография на сефарозе Q
К раствору после диализа вначале добавляют бутанол до 4% и помещают в колонку с сефарозой Q (2,5 см•10 см), которую уравновешивают буфером А + 20 мМ хлорида натрия со скоростью потока 25 мл/ч. После промывки колонки тем же самым буфером проводят элюирование в линейном градиенте концентраций хлорида натрия в промывочном буфере до достижения концентраций 250 мМ (250 мл; 25 мл/ч). Фракции объемом 2,5 мл, содержащие рекомбинантную нейраминидазу, идентифицируют, измеряя активность фермента и его содержание по методу твердофазного иммуноферментного анализа (ELISA). Активность нейраминидазы элюируется из колонки в виде одного пика.
с. Афинная хроматогарфия на агарозе с N-(п-аминофенил)оксамовой кислотой
Использование указанной афинной матрицы описано для очистки нейраминидазы (нерекомбинантной) вируса гриппа и бактериальных NА-ферментов (Cuatracassas and Illiano, 1971; Bucher, 1977). Корректное функционирование афинной матрицы достигается лишь после изменения первоначально рекомендованных буферных условий. Активные фракции после разделения на сефарозе Q собирают и добавляют к ним равный объем 200 мМ раствора ацетата натрия с рН 5,5. Затем активные фракции помещают в колонку на основе агарозы с N-(п-аминофенил)оксамовой кислотой (1,5 • 5 см), уравновешенную буфером В, 100 мМ NaCl. Затем колонку промывают уравновешивающим буфером и обессоливают буфером В. Вторую стадию промывки проводят, используя буфер А. Рекомбинантную нейраминидазу в конце концов элюируют буфером А, содержащим 1 М хлорид натрия со скоростью потока 10 мл/ч (собирают фракции объемом 2 мл).
d. Гель-хроматография на Superdex 200
Элюат, полученный после афинной колонки, концентрируют до объема 2,0 мл, используя концентраторы Centriprep ТМ (от компании "Amicon", молекулярная масса отсечки 30 килодальтон). Концентрат хроматографируют во фракциях объемом 1,0 мл на колонке для гель-хроматографии с Superdex 200 (1,5•60 см), которую уравновешивают буфером С, содержащим 4% бутанола. Колонку элюируют уравновешивающим буфером при скорости потока 10 мл/ч и отбирают фракции объемом 1,0 мл. Для хранения в течение длительного времени при температуре минус 20oС отбирают соответствующие фракции, концентрируют как описано ранее и добавляют к ним глицерин до конечной концентрации 50%.
Для оценки молекулярного веса очищенных белков гель-хроматографическую колонку калибруют апоферритином из селезенки лошади (443 килодальтон), бета-амилазой из сладкого картофеля (200 килодальтон), алкогольдегидрогеназой из дрожжей (150 килодальтон), бычьим сывороточным альбумином (67 килодальтон) и карбонангидразой (29 килодальтон) (все от компании "Sigma Chemical Со.").
6. Получение и очистка природной нейраминидазы, обработанной проназой (pNA)
а. Обработка проназой
Желточный мешок куриных эмбрионов, инфицированных с помощью Х-47, вначале осветляют центрифугированием на малой скорости (1000•g, 10 минут), а затем подвергают центрифугированию при 13000•g в течение 16 часов, чтобы осадить вирус. Осадок вируса вновь суспендируют в объеме 10 мл буфера С на эквивалент 100 инфицированных яиц и, не проводя дальнейшую очистку вируса, добавляют проназу в количестве от 2 мг/мл. Смесь инкубируют в течение 16 часов при температуре 20oС, слегка перемешивая. Оставшиеся части вируса и нерастворенные проназой компоненты затем удаляют на ультрацентрифуге (100000•g, 1 час) при температуре 4oС. Супернатант, содержащий освобожденные головки нейраминидазы, очищают колоночной хроматографией.
b. Катионообменная хроматография на сефарозе S
Хроматографию проводят при температуре 4oС. Сырой образец рNА разбавляют в пять раз и добавляют к 50 мМ раствора ацетата натрия с рН 5,5, содержащего 2 мМ хлорида кальция и 1% бутанола. Затем раствор помещают на колонку с сефарозой S (1,5 см • 10 см), которую уравновешивают буфером В + 1% бутанола и 50 мМ хлорида натрия. Связанное вещество элюируют в линейном градиенте до концентрации 500 мМ хлорида натрия в том же самом буфере. Собирают фракции, показывающие наибольшую активность фермента, и концентрируют до объема 2,0 мл с помощью концентраторных трубок Centriprep ТМ ("Amicon"; молекулярная масса отсечки: 30 килодальтон).
d. Гель-фильтрация на Superdex 200
Гель-фильтрацию на Superdex 200 проводят аналогично тому, как проводят очистку нейраминидазы (за исключением того, что концентрация бутанола составляет 1%). Очищенную pNA хранят при температуре минус 20oС в 50%-ном глицерине.
7. Ферментативный анализ нейраминидазы
Анализ каталитической активности нейраминидазы основан на методе Potier et al. (1979). Если коротко, то испытания фермента проводят в реакционном объеме 100 мкл в растворе 20 мМ ацетата натрия с рН 6,5, 2 мМ хлорида кальция и 1% бутанола в присутствии 1 мМ 2'-(4-метилумбеллиферил)-альфа-D-N-ацетилнейраминовой кислоты в качестве субстрата. После инкубации при температуре 37oС в течение от 30 до 60 минут реакцию прекращают, добавляя 0,5 мл 133 мМ глицина, 83 мМ бикарбоната натрия, 60 мМ хлорида натрия, рН 10,7. Определяют содержание 4-метилумбеллиферона, измеряя поглощение на длине волны 365 нм. Одну единицу определяют как количество фермента, которое освобождает один нмоль 4-метилумбеллиферона в минуту.
8. Иммунологические методы
а. Получение поликлонального анти-рNА IgG
Поликлональную антисыворотку против очищенной pNA, обработанной проназой, получают в трехмесячных кроликах линии New Zealand. Первичную иммунизацию проводят, вводя в каждую лапу внутримышечно четыре дозы объемом 500 мкл, содержащей 50 мкг рNА/доза и 75%-ный полный адъювант Фрейнда. Через шесть недель животные получают две соответствующие бустерные инъекции в обе задние лапы. Для получения фракций IgG собранную сыворотку очищают адсорбцией на протеин А-сефарозе ("Pharmacia LKB").
b. ELISA
Ячейки планшета для микротитрования покрывают анти-рNА IgG кролика. Испытуемые образцы разбавляют забуференным фосфатом солевым физиологическим раствором, содержащим 0,1% бычьего сывороточного альбумина. Связанный антиген определяют с помощью обработанного биотином анти-рNА IgG кролика с последующим добавлением конъюгата стрептавидин-щелочная фосфатаза ("Boehrinqer"). Ферментативная реакция развивается при инкубировании планшетов с п-нитрофенилфосфатом ("Sigma Chemical Co."). Значения абсорбции определяют на длине волны 405 нм с помощью ридера микропланшетов.
9. Аналитические методы
Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия проводят по методу Laemmli (1970) на 10%-ном разделяющем геле (если специально не указано). Все образцы, если специально не оговаривается, денатурируют в присутствии бета-меркаптоэтанола. В качестве белков-маркеров в 10%-ном геле используют фосфорилазу b (94 килодальтон), бычий сывороточный альбумин (67 килодальтон), овальбумин (43 килодальтон), карбонангидразу (29 килодальтон) и ингибитор трипсина (20,1 килодальтон, не всегда виден) ("Pharmacia LKB"). Градиентные гели калибруют следующими стандартами массы: миозин (22 килодальтон), бета-галактозидаза (116 килодальтон), фосфорилаза b, бычий сывороточный альбумин и овальбумин ("Biorad"). Окрашивание серебром в гелях проводят по модифицированному методу, приведенному Morrisey (1981). Концентрацию белка определяют по методу Bradford (1976), используя в качестве стандарта овальбумин.
10. Анализ методом перекрестного связывания
Молекулу ВS3 для перекрестного связывания используют в виде свежеприготовленного 1,0 М раствора в 10 мМ буфере Hepes с рН 7,4. Белки перекрестно связывают добавлением BS3 до концентрации 0,5 мМ в реакционном объеме 30 мкл. Инкубацию проводят в течение 1 ч при комнатной температуре. Затем реакцию прерывают, добавляя 5 мкл 1,0 М буфера Тris с рН 8,0. Полипептидные образцы анализируют с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия.
11. Углеводный анализ
Образцы белков (от 0,1 мкг до 1 мкг) денатурируют кипячением в 500 мМ Тris/НСl с рН 8,0, 0,5% додецилсульфата натрия, 50 мМ бета-меркаптоэтанола. После добавления нейраминидаза-октилглюкозида до концентрации 2,5%, что приводит к по крайней мере семикратному превышению над конечной концентрацией додецилсульфата натрия, добавляют нейраминидаза-глюканазу (около 0,5 единиц; количество единиц указано в соответствии с данными производителя) и реакционную смесь инкубируют при температуре 37oС в течение 16 часов. Картины расщепления анализируют электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия.
Результаты
1. Очистка pNА
При проведении описываемого здесь типичного эксперимента перерабатывают 186 инфицированных яиц. Различные стадии очистки суммированы в табл. 1. После сбора жидкости из желточных мешочков и осаждения вируса добавляют проназу до концентрации 2 мг/мл и смесь инкубируют при температуре 20oС в течение 16 часов. После ультрацентрифугирования в жидкости над осадком определяют приблизительно 60% активности нейраминидазы. Было показано, что в указанных условиях потеря активности в основном связана с неполным извлечением головок нейраминидазы вирусных частиц. Более высокие концентрации проназы, более длительные периоды инкубации или более высокая температура не приводят к увеличению выхода, поскольку нейраминидаза постепенно деградирует (данные не показаны). Сырой материал рNА затем растворяют, доводят рН до 5,5 и помещают на катионообменную колонку с сефарозой S. Для максимального выхода рNА, все последующие растворы содержат 1% бутанола. Большая часть белка не удерживается в колонке с сефарозой S и после градиентного элюирования при значении концентрации хлорида натрия приблизительно 400 мМ собирают единственный пик (не показано). Это вещество содержит в значительной степени очищенную рNА: поскольку никаких загрязняющих полос после электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия не обнаружено (фиг. 3А, дорожка 3). Далее, окрашивание серебром не показывает никаких отличий между пулом после сефарозы S и после дополнительной гель-фильтрации на Superdex 200 (фиг. 3А, дорожки 3 и 4). После последней колонки собирают единственный колоколообразный пик во фракции 60 (не показано), который соответствует молекулярному весу приблизительно 210 килодальтон. Последовательные стадии очистки поясняются на фиг. 3А.
Во время электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия мономер рNА на самом деле виден как дублет двух полос, соответствующих соответственно приблизительно 54 килодальтон и 52 килодальтон, причем последняя встречается наиболее часто, о чем свидетельствует относительно интенсивности при окрашивании серебром. Наиболее вероятно, что эта амбивалентность появляется вследствие предпочтительного расщепления проназой в двух различных местах стеблевой области. Перекрестное связывание химическим агентом BS3 подтверждает, что pNA выделяется в виде аутентичного тетрамерного белка (фиг. 3В).
2. Конструирование и экспрессия рекомбинантной нейраминидазы
Ген нейраминидазы штамма A/Victoria/3/75 гриппа NA2 отделяют от его концевого мембранного якоря нейраминидазы и присоединяют вместо этого к 5'-последовательности гена гемаглютинина A/Victoria/3/75, который содержит сайт сплайсинга сигнального пептида. Становится возможным синтез секретируемого растворимого продукта. Полученный химерный ген содержит сигнальную последовательность гемагглютинина, включающую кодоны для первых четырех концевых аминокислот зрелого гемагглютинина, непосредственно за которыми следуют последовательность нейраминидазы, в которой отсутствует трансмембранная часть (якорь) и часть стеблевой области (аминокислоты с 1 по 45). Обе ДНК-последовательности располагаются в одной рамке считывания информации, при этом не вводится дополнительных аминокислот. Лигирование приводит лишь к одной замене аминокислоты, соответствующей позиции 5 зрелого белка гемагглютинина (фиг. 2). Копию этой химерной последовательности которая теперь в основном кодирует способный секретироваться белок, интегрируют позади промотора полиэдрина бакуловируса AcNPV, используя pVL941 в качестве вектора переноса. После заражения клеток sf9 насекомого быстро определяют активность нейраминидазы в этой среде и показывают, что действительно получает растворимый белок.
Из фиг. 4 видно, что активность нейраминидазы в среде достигает уровня плато приблизительно через 48 часов после инфицирования. Дальнейшая инкубация не приносит результатов, поскольку общая концентрация белка начинает резко уменьшаться, вероятно вследствие избыточного лизиса клеток. Было показано, что экспрессия наиболее интенсивно проходит в том случае, когда промежуточный перенос между исходным монослоем sf9 и суспензионной культурой ограничен к минимуму (данные не приведены). На основании разнообразных экспериментов по очистке было установлено, что уровень экспрессии нейраминидазы варьирует от 6 до 8 мг/л, что соответствует довольно низкой производительности, однако она сравнима с выходами, которые сообщаются для других секретируемых комплексных гликопротеинов, продуцируемых в данной системе (Jarvis et al., 1990).
3. Очистка рекомбинантной нейраминидазы
Среду ТС100 собирают через 48 часов после инфицирования в тот момент, когда удельная ферментативная активность растворимого белка достигает пикового значения (фиг. 4). Различные стадии очистки рекомбинантной нейраминидазы суммированы в табл. 1. Осаждение сырой среды сульфатом аммония в интервале насыщения от 20 до 60% дает среднее двоекратное обогащение и позволяет осуществить концентрирование продукта. После интенсивного диализа и удаления нерастворимых продуктов добавляют бутанол до концентрации 4%. Было показано, что добавление бутанола оказывает значительное благоприятное воздействие на общий выход рекомбинантной нейраминидазы, особенно при низких концентрациях белка. Возможно, для предотвращения образования нерастворимых агрегатов необходима определенная гидрофобность среды. Затем раствор фракционируют анионообменной хроматографией на сефарозе Q (фиг. 5). Активность нейраминидазы элюируется в начале градиента соли в виде практически симметричного пика. В соответствии с твердофазным иммуносорбентным анализом оставшиеся фракции не содержат относящегося к нейраминидазе вещества. На этой стадии выделяют приблизительно 97,5% исходного количества белка, что приводит к увеличению удельной активности приблизительно в 20 раз. Затем рН раствора перед загрузкой в колонку с N-(п-аминофенил)оксамовой кислотой - агарозой понижают до величины 5.5. Из ранних исследований известно, что нейраминидаза-замещенные оксамовые кислоты являются сильными обратимыми ингибиторами нейраминидазы гриппа (Edmond et al., 1966). Использование N-(п-аминофенил)оксамовой кислоты - агарозы в качестве селективного абсорбента нейраминидазы гриппа или бактерий впервые описали Cuatrecasas and Illiano (1971) и позднее Bucher (1977). В соответствии с оригинальной методикой нейраминидазу элюируют буфером с высоким рН (100 мМ бикарбоната натрия, рН 9,1). Однако в наших экспериментах эти условия позволяют осуществить лишь частичное и медленное извлечение рекомбинантной нейраминидазы. Однако эффективной десорбции можно достичь, сочетая увеличение значения рН с высокой концентрацией соли. Перед элюированием значительное количество неспецифично связанного белка удаляют из колонки, проведя дополнительную стадию промывки при рН 8,5 при низкой концентрации соли. Если отдать предпочтение диэтаноламину как буферному агенту перед бикарбонатом натрия, то можно добиться абсорбции 2 мМ хлорида кальция без осаждения. Неоднократно указывалось, что сохранение активности нейраминидазы несколько зависит от концентрации ионов кальция (Chond et al., 1966; Dimmock, 1971). Так ли это, в настоящем исследовании детально не анализируется.
Чтобы удалить следы остаточных загрязняющих веществ элюат концентрируют методом ультрафильтрации и подвергают гель-фильтрации на Superdex 200 (фиг. 6). Проверка с использованием А280 показывает три пика с неравными поглощениями, которые элюируются, соответственно, приблизительно как 220 килодальтон, приблизительно 130 килодальтон и приблизительно 54 килодальтон. Было показано, что картины иммунореактивности элюата, измеренные методом иммуноферментного твердофазного анализа, являются надежным представлением профилей A280 для каждого из зарегистрированных трех пиков, что свидетельствует о том, что все соединения являются NАs-специфичными. Анализ фракций пиков с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия показывает интенсивную полосу в ожидаемой области размером приблизительно 55 килодальтон, хотя с увеличением номера фракции наблюдается небольшое понижение молекулярного веса (фиг. 7А). Пик размером 220 килодальтон анализом по методу перекрестного связывания с использованием ВS3 идентифицируют как тетрамерную рекомбинантную нейраминидазу, а два других пика с меньшей молекулярной массой являются, как было обнаружено, димерной и мономерной нейраминидазой, соответственно, при этом последняя форма имеет незначительную количественную значимость (фиг. 7В). Полагают, что благодаря своей палочкообразной структуре димерная рекомбинантная нейраминидаза элюируется несколько выше значения своего молекулярного веса по сравнению с тетрамерной и мономерной рекомбинантной нейраминидазой, которые, как полагают, имеют круглую форму. Особо следует отметить, что каталитическая активность требует полностью собранной тетрамерной структуры рекомбинантной нейраминидазы. Возможно, формирование тетрамера вызывает несколько локальных конформационных изменений, которые имеют важное значение для ферментной активности.
Последовательность проведения операций очистки приведена в табл. 2.
4. Свойства рекомбинантной нейраминидазы
Денатурация кипящим раствором додецилсульфата натрия в присутствии бета-меркаптоэтанола вызывает полную диссоциацию рекомбинантной нейраминидазы на мономерные цепочки с молекулярным весом близким к 55 килодальтон (фиг. 7А). Тетрамерная и димерная рекомбинантная нейраминидаза гомогенно очищены, если судить по окрашиванию серебром гелей при электрофорезе в полиакриламидном геле в присутствии додецилсульфата натрия. Мономерная рекомбинантная нейраминидаза имеет несколько худшее качество, поскольку наблюдается несколько следов загрязняющих соединений. Когда их денатурируют в отсутствие восстановителя, тетрамерная и димерная рекомбинантная нейраминидаза мигрируют в виде димерных цепочек размером приблизительно 110 килодальтон (не показано). Полученные результаты свидетельствуют о том, что димеры рекомбинантной нейраминидазы действительно связываются дисульфидными мостиками и могут далее ассоциироваться посредством нековалентных взаимодействий, образуя тетрамер, который соответствует структурной организации природной нейраминидазы.
Неоднократно сообщалось, что клетки насекомого дают картину гликозилирования нейраминидазы, отличающуюся в некоторой степени от картины, которую дают клетки млекопитающих и другие клетки высших животных (Hsieh and Robbins, 1984; Butter and Hughes 1981; Butters et al., 1981; Kiroda et al., 1990). В связи с этим проводят изучение количества связанного нейраминидазой углевода рекомбинантной нейраминидазы в сравнении с природной нейраминидазой, обработанной проназой. Отдельные образцы белка обрабатывают ферментом NА-глюканазой, а затем анализируют электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия (фиг. 9). На основании относительного смещения полос можно сделать вывод, что общее количество NА-связанного углевода рекомбинантной нейраминидазой несколько меньше по сравнению с природной молекулой (сравните фиг. 9А и 9В), и это наблюдение согласуется с наблюдением для других гликопротеинов, экспрессирующихся в этой системе (Kiroda et al., 1986; Domingo and Trowbridge, 1988; van Drunen Littel et al., 1991). Было также установлено, что денатурированная ферментативно дегликозилированная рекомбинантная нейраминидаза образует миграты с той же самой электрофоретической мобильностью независимо от их первоначальной олигомерной структуры, а это подтверждает тот факт, что первичная рекомбинантная нейраминидаза синтезируется в виде полипептида с однородной длиной цепи (фиг. 9В, сравните дорожки 3, 5 и 9). Молекулярной вес полипептидной цепи после обработки NА-глюканазой, составляет по оценке 47,5 килодальтон, что соответствует теоретическому значению 47717 дальтон на основании предсказанной последовательности аминокислотных остатков. Достаточно интересно отметить, что степень гликозилирования нейраминидазы вероятно связана со способностью образовывать тетрамеры, поскольку полосы, соответствующие гликозилированной димерной и мономерной нейраминидазе, движутся несколько быстрее в геле, чем полосы гликозилированной тетрамерной нейраминидазы (фиг. 9В, дорожки 4 и 6 по сравнению с дорожкой 2; см. также фиг. 7А). Действительно, было высказано предположение, что связанный с нейраминидазой углеводород, а именно олигосахаридная цепочка, присоединенная к Аsn200, может играть роль в установлении тетрамерной структуры за счет взаимодействия с прилегающими субъединицами (Varghese et al., 1983; Varghese et Colman, 1991).
Вклад в каталитические свойства рекомбинантной нейраминидазы вносит лишь тетрамерный белок. Выделенная тетрамерная рекомбинантная нейраминидаза обладает удельной активностью, практически идентичной удельной активности очищенной природной нейраминидазы, обработанной проназой (табл. 1 и 2). То наблюдение, что форма рекомбинантной нейраминидазы низшего структурного порядка ферментативно неактивны, даже несмотря на то, что каждый мономер имеет каталитическую полость, вероятно, отражает роль структуры в четвертичных взаимодействиях при функционировании нейраминидазы гриппа.
С целью проверки антигенных свойств рекомбинантной нейраминидазы, образцы белка равной концентрации дважды последовательно разбавляют и испытывают в сэндвич-иммуноферментном твердофазном анализе с использованием поликлонального анти-рNА IgG (фиг. 10). Кривая титрования тетрамерной рекомбинантной нейраминидазы идентична кривой сравнения природной нейраминидазы, обработанной проназой, указывая на то, что обе они имеют идентичные или очень похожие антигенные свойства. Несмотря на отсутствие заметной ферментативной активности, антигенная активность димерной и мономерной рекомбинантной нейраминидазы остается практически неизменной, хотя может наблюдаться небольшой сдвиг антигенности. Это небольшое отличие в антигенности очевидно и из профиля гель-фильтрации (фиг. 6), где отношение антигенная активность/A280 показывает некоторое превосходство пика тетрамера. Возможно, некоторые молекулы антигена, генерированные против природной тетрамерной структуры, не способны эффективно связываться с неполностью собранной рекомбинантной нейраминидазой, например, такие молекулы, которые распознают контактные области между соседними субъединицами. Локальные конформационные изменения, вызванные формированием тетрамера, также могут приводить к незначительным антигенным различиям.
Обсуждение результатов
Основной целью настоящего изобретения является синтез антигена нейраминидазы гриппа в виде секретируемого и правильным образом свернутого белка вместе с выбором процедуры очистки с целью получения гомогенного продукта, который можно использовать в качестве средства для вакцинации. Из гена нейраминидазы штамма A/Victoria/3/75 гриппа NA2 конструируют химерный ген, в котором первоначальный NA-концевой район, который обладает комбинированной функцией сигнальной последовательности -
мембранного якоря замещена частью 5'-последовательности гена нейраминидазы гриппа. Полученная генно-инженерная конструкция благодаря способному расщепляться сигнальному пептиду, полученному из гемагглютинина, и кодирующая в основном секретируемую нейраминидазу, затем включается в вектор экспрессии бакуловируса при транскрипционном регулировании со стороны мощного промотора полиэдрина. После инфекции клетки-хозяина sf9 в культуральную среду действительно секретируется рекомбинантная нейраминидаза. На основании результатов очистки уровень экспрессии оценивают величиной 6-8 мг/л. Было показано, что в процессе инфекции бакуловируса способность клетки-хозяина производить белки путем секреции значительно понижается (Jarvis and Summers, 1989). Тем не менее описанная система продуцирования применима для изучения вакцинаций в условиях лаборатории и подходит для использования в больших масштабах.
Очистка рекомбинантной нейраминидазы в основном заключается в проведении четырехстадийного процесса, первой стадией которого является фракционирование с помощью сульфата аммония, за которой следуют три последовательные стадии хроматографии. На основании выхода ферментативной активности количество рекомбинантной нейраминидазы, выделенной в виде белка, оценивают приблизительно в 25%. С помощью хроматографии на гель-хроматографической колонке рекомбинантную нейраминидазу субфракционируют на три популяции с различным размером молекул, которые идентифицируют методом перекрестного связывания, соответственно, как тетрамерную, димерную и мономерную рекомбинантную нейраминидазу, причем последняя форма присутствует лишь в очень малых количествах. Две основные формы, тетрамерную и димерную рекомбинантную нейраминидазу, получают приблизительно в равных количествах, как показывает электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия с последующим окрашиванием серебром.
Чтобы оценить ферментативные и иммунологические свойства рекомбинантной нейраминидазы, необходимо выделить природную нейраминидазу, обработанную проназой, в качестве белка сравнения. Головка нейраминидазы A/Victoria/3-75 вируса Х-47 расщепляют обработкой проназой, а затем очищают катионообменной и гель-хроматографией. Проведение перекрестного связывания подтверждает, что рекомбинантная нейраминидаза сохраняет тетрамерную структуру интактной нейраминидазы, присоединенной к мембране.
Каталитические свойства рекомбинантной нейраминидазы весьма примечательны, поскольку ферментативную активность проявляет лишь тетрамерный белок. Удельная активность тетрамерной рекомбинантной нейраминидазы практически равна удельной активности природной нейраминидазы, обработанной проназой. Вероятно, димерная и мономерная рекомбинантная нейраминидаза неактивны по той причине, что представляют собой денатурированные белки, поскольку в процессе очистки указанные формы прочно удерживались при аффинной хроматографии в местах связывания сусбтрата, указывая на то, что ферментативная полость функционально не затрагивается, однако последующий каталитический сдвиг может не наблюдаться.
Обработка NА-гликоназой показывает, что в целом содержание углеводов в рекомбинантной нейраминидазе несколько ниже, чем в природной нейраминидазе, обработанной проназой, что также наблюдается для других гликопротеинов, экспрессируемых в данной системе (Kuroda et al., 1986; Domingo and Trowbridge, 1986; van Drunen Littel et al., 1991). Гипогликозилирование, видимо, более выражено для димерной и мономерной рекомбинантной нейраминидазы.
Структурные исследования с помощью рентгеновского дифракционного анализа показывают, что углеводная цепочка, присоединенная к А200, тесно контактирует с соседними субъединицами, заставляя предположить: что это может способствовать возникновению дополнительных взаимодействий, укрепляющих четвертичную структуру (Vaqhese et al., 1983; Vaghese and Colman, 1991).
Реакционноспособность тетрамерной рекомбинантной нейраминидазы по отношению к поликлональным иммуноглобулинам, генерированным против очищенной природной нейраминидазы, обработанной проназой, является практически полной, указывая на то, что оба белка обладают очень схожими антигенными свойствами. Можно наблюдать небольшой сдвиг антигенности в случае димерной и мономерной рекомбинантной нейраминидазы. Отсюда следует, что, вероятно, можно выделить моноклональные антитела, которые связываются лишь с тетрамерной структурой нейраминидазы гриппа. Подобные антитела, вероятно, вступают во взаимодействие с поверхностными детерминантами, образованными соседними субъединицами, или же они способны распознавать эпитопы, возникающие после конформационных перегруппировок в процессе формирования тетрамера. Кроме того, различия в составе углевода также могут модулировать антигенные свойства.
Пример 2. Секреция рекомбинантной нейраминидазы Pichia pastoris
Введение
С целью выяснить, можно ли использовать дрожжи помимо клеток насекомых для продуцирования рекомбинантной нейраминидазы гриппа, конструируют вектор экспрессии, содержащий обладающую ферментативной активностью область "шляпки" нейраминидазы.
Материалы и методики
1. Вектор и хозяин
Для конструирования полигенного экспрессирующего кластера используют плазмиду рРIС9 из Pichia pastoris (Invitrogen). Эта плазмида содержит сайт инициации репликации, ген устойчивости к ампициллину, области промотора и терминатора вводимого гена алкогольоксидазы 1 (АОХ1) из Р.pastoris, препро-сигнал секреции альфа-фактора Saccharomyces cerevisiae и HIS4 маркер из Р. pastoris.
В качестве хозяина используют метилотрофные дрожжи Pichia pastoris ("Invitrogen").
2. Конструирование полигенного экспрессирующего кластера
Методом сайт-ориентированного мутагенеза сайт рестрикции Stu I вводят в последовательность кДНК гена нейраминидазы А/Victoria/3/75. Сайт рестрикции расположен в положении Рro79. С помощью этого сайта рестрикции можно выделить иммуногенную "последовательность шляпки" гена нейраминидазы, содержащего активный центр в виде фрагмента Stu I/Hind III, и клонировать ее в сайте рестрикции Sna BI плазмиды рРIС9 из Р.pastoris. На фиг. 15 приведена диаграмма плазмиды рРIС9. На фиг. 16 показан вид области слияния между препро-сигнальной последовательностью и рекомбинантной нейраминидазой. Пропептид расщепляют по методу Гольджи посредством эндогенной протеазы КЕХ2. Дипептид (Glu-Ala)2 выделяют, используя дипептидиламинопептидазу STЕ13-типа. Лишний остаток тирозина не расщепляется и сохраняется в рекомбинантной нейраминидазе в виде N-концевого остатка, однако он не нужен.
Полученную плазмиду линеаризуют по положению НIS 4 маркера селекции расщеплением с помощью SаlI, а затем трансформируют в протопласты GTS115 (his 4) Р. pastoris в присутствии полиэтиленгликоля. ДНК, выделенную из трансформированных клеток, анализируют по методу саузерн-блоттинга. Проведенный анализ показывает, что вектор экспрессии интегрируется посредством гомологичной рекомбинации в положение внутреннего (однако неполного) локуса his 4. Большинство трансформированных клеток содержит от 1 до 2 копий плазмиды, однако было обнаружено, что трансформированные клетки с большей секретирующей способностью содержат несколько копий, которые интегрируются по типу голова к хвосту в геноме хозяина в тандемной структуре. Количество копий может достигать до 25 на одну трансформированную клетку.
3. Экспрессия нейраминидазы
Трансформированные клетки предварительно выращивают в минимальном количестве глицерина, содержащего буфер (рН 6,0) и через 48 часов переносят в минимальное количество буферного раствора, содержащего 0,5% метанола. Вводят промотор алкогольоксидазы 1 и экспрессируют "шляпку" нейраминидазы. Используя известный метод анализа по нозерн-блоттингу, проводят оценку количества мРНК нейраминидазы в клетке. Анализ показывает, что произошла весьма эффективная индукция.
Анализ по методу вестерн-блоттинга жидкости над осадком показывает, что экспрессируется рекомбинантная нейраминидаза с молекулярным весом приблизительно 70 кДа (см. фиг. 17).
Секретируемый продукт подвергают дегликозилированию с помощью РNGазы F. Получают продукт "сердцевины" с ожидаемым размером, составляющим 43 кДа. Было обнаружено, что в зависимости от количества копий выход рекомбинантной нейраминидазы в среде меняется от 1 до 1,5 мг/л.
Пример 3. Иммунизация
Материалы и методики
1. Животные
Возраст женских инбредных особей мышей Balb/c (от компании "SСК Моl", Бельгия) в начале проведения иммунизации составлял 8 недель. При проведении пассивной иммунизации возраст мышей-реципиентов составлял 12 недель. Мышей разбивают на группы по три животных на клетку (410 куб.см) и вдоволь дают пищу и воду.
2. Вирусы
Штаммы вируса получены от доктора A. Douglas и доктора J. Skehel ("MCR Laboratories", Милл-Хилл, Лондон). Лабораторные вирусы Х-31 и Х-47 имеют H3N2-антигенный состав и их получают путем генетической реаранжировки из A/PR/8//34 (Н1N1) соответственно с А/Аiсhi/2/68 (H3N2) и A/Victoria/3-75 (H3N2). Оба источника вирусов адаптируют несколькими перепрививками в легкие, так чтобы они вызывали смерть у мышей.
3. Секретируемая рекомбинантная нейраминидаза
Нейраминидазу гриппа A/Victoria/3/75 вводят в виде очищенного рекомбинантного белка, продуцируемого с помощью системы экспрессии бакуловирус-клетки насекомого, как это описано в Примере 1. Очищенные препараты рекомбинантной нейраминидазы, которые используют для описываемых экспериментов по иммунизации, содержат смесь тетрамерных и димерных молекул в забуференном фосфатном солевом физиологическом растворе.
4. Адъюванты
Подходящие адъюванты подбирают на основе исследований иммунизации с использованием рекомбинантного гемагглютинина гриппа, которые проводят в нашей лаборатории. Адъювант компании "Ribi" (с монофосфориллипидом A (MPLA), трегалоза-6,6-димиколятом (ТМD), скваленом и Tween 80) и Sаlmоnеlla typhimurium MPLA заполняют в бутылочки в соответствии с рекомендациями изготовителя ("Ribi Immunochem Rеsеаrсh"). Мурамилдипептид (МДР) покупают у компании "Sigma Chemical Со."
5. Методика иммунизации
Мышам вводят подкожно с интервалом в три недели три дозы объемом 200 мкл, каждая из которых содержит 1 мкг рекомбинантной нейраминидазы. Для первой иммунизации рекомбинантную нейраминидазу эмульгируют в половине обычной дозы Ribi для мыши (соответствует 25 мкг MPLA, 25 мкг TMD, 2 мкл сквалена и 0,1% Tween 80). Активные иммунизации проводят, добавляя 25 мкг MPLA и 25 мкг MDP к рекомбинантной нейраминидазе. Контрольные животные получают адъювант, растворенный в забуференном фосфатом солевом физиологическом растворе.
6. Пассивная иммунизация
Через три недели после проведения третьей иммунизации берут анализы крови у мышей-доноров методом пункции из сердца и препараты сыворотки от соответствующих подвергнутых вакцинации мышей объединяют. Мыши-реципиенты получают внутрибрюшинно одну инъекцию иммунной или контрольной сыворотки объемом 400 мкл.
7. Заражение гриппом
Под легким эфирным наркозом мышей заражают дозой 20 LD50 конкретного вируса через три недели после проведения последней активной иммунизации или через один день после пассивной иммунизации. За развитием инфекции следят в течение 10 дней, измеряя ректальную температуру и вес тела в период после прививки.
8. Серологический метод
За один день до начала проведения методики вакцинации (предыммунная сыворотка) и через две недели после каждой иммунизации берут пробы крови из хвостовой артерии. Индивидуальные образцы сыворотки исследуют на NА-специфичные антитела с помощью иммуноферментного твердофазного анализа. Планшеты для титрования ("Nunc Maxisorp") покрывают очищенной рекомбинантной нейраминидазой (50 нг на ячейку) и сыворотку разбавляют в соотношении 1:5. Связывание специфичного антитела количественно оценивают, добавляя кроличьи антитела к анти-(IgG мыши), конъюгированные с щелочной фосфатазой ("Sigma Chemical Co."), а затем планшеты выдерживают в инкубаторе вместе с раствором п-нитрофенола (от компании "Sigma Chemical Co."), который является субстратом. Величину оптической плотности определяют на длине волны 405 нм с помощью ридера планшетов для микротитрования. Титр антигена нейраминидазы выражают в виде обратной величины logs-разбавления сыворотки и получают величину поглощения, которая на 0,05 выше, чем в контрольных ячейках (обработанных предыммунной сывороткой).
Результаты
1. Порядок проведения исследований
Три группы по 12 мышей вакцинируют рекомбинантной нейраминидазой в соответствии с вышеприведенной методикой иммунизации. Равное количество контрольных мышей параллельно обрабатывают забуференным фосфатом солевым физиологическим раствором. Парные группы вакцинированных и контрольных мышей затем заражают адаптированными для мышей Х-47 и Х-31. В альтернативном методе они служат в качестве доноров сыворотки для проведения экспериментов по пассивной иммунизации.
2. Серологический ответ
За ответными реакциями антигенов против рекомбинантной нейраминидазы следят методом иммуноферментного твердофазного анализа в сыворотке случайно выбранных 12 вакцинированных и 12 контрольных животных (по одному животному из каждой клетки) (фиг. 11). Применение рекомбинантной нейраминидазы для проведения иммунизации у мышей вызывает устойчивый рост содержания антител к рекомбинантной нейраминидазе в сыворотке. Первая активная иммунизация вызывает увеличение количества антител приблизительно на величину в три lоgs, в то время как реиммунизация приводит к еще одному приблизительно пятикратному увеличению титра антител к рекомбинантной нейраминидазе. У контрольных мышей инъекция одного лишь адъюванта не приводит к значительному продуцированию аспецифичных антител, которые взаимодействуют с рекомбинантной нейраминидазой.
3. Гомовариантная защита с помощью рекомбинантной нейраминидазы
Через три недели после вакцинации мышей, подвергнутых иммунизации, и контрольных мышей изучают на иммунную активность, назначая дозу 20 LD50 гомо-NА-вариантного вируса Х-47 (фиг. 12). Все контрольные животные серьезно заболели, что определяют измерением возрастающего падения температуры тела и по потере веса тела. На четвертый день после инфекции обнаруживаются первые жертвы, а по прошествии девяти дней после заражения все контрольные животные умерли. Напротив, у животных, которым вводили вакцины рекомбинантной нейраминидазы, наблюдается лишь временное и весьма умеренное изменение клинических параметров. Все иммунизированные животные перенесли инфекцию.
Исследуют также, может ли быть получен тот же уровень защитного иммунитета без третьей иммунизации просто за счет применения компенсации в виде больших доз рекомбинантной нейраминидазы и/или адъюванта. Эти испытания проводят по той же самой схеме. Хотя мыши, вакцинированные указанным образом, обычно показывают хорошую резистентность, несколько отдельных особей серьезно заболевают, а ряд подвергнутых вакцинации животных в конце концов умирают, хотя процент выживших животных редко составляет меньше 80%. Однако было установлено, что уровень защитного иммунитета, достигаемый путем трех вакцинаций, превосходит во всех отношениях уровень защиты после двух вакцинаций.
4. Гетеровариантная защита с помощью рекомбинантной нейраминидазы
Параллельные группы вакцинированных и контрольных мышей исследуют на иммунитет с помощью дозы 20 LD50 гетеро-NА-вариантного вируса Х-31, который содержитнейраминидазу, выделенную из рекомбинантной нейраминидазы из A/Victoria/3/75 после 7 лет антигенного дрейфа (фиг. 13). Мы также наблюдали при проведении иммунизации с использованием X-47, что клинические последствия инфекции в контрольной группе были драматическими. Контрольные животные начали умирать уже через пять дней после инфекции. Смертность достигла максимума на восьмой день, когда осталось лишь одно животное. Мыши, которым вводили вакцины рекомбинантной нейраминидазы, показывают 100%-ную выживаемость после обычно летальных гетеровариантных инфекций. Точно так же, как и в исследовании гомовариантной иммунизации, подвергнутые вакцинации животные оказались также способны поддерживать свою температуру тела на разумно нормальном уровне. Потеря веса тела выражена несколько ярче, однако все мыши начинают восстанавливаться на шестой день.
5. Протективный иммунитет может быть достигнут пассивным переносом NАS-иммунной сыворотки.
Для определения, имеют ли значение гуморальные защитные механизмы, которые главным образом ответственны за индукцию протективного иммунитета, тестируют защиту животных с помощью пассивной иммунизации. Для этого мышам-донорам делают прививки по стандартной методике. Берут кровь и получают от каждого животного приблизительно 400 мкл сыворотки. Объединяют, с одной стороны, образцы контрольной сыворотки, а с другой стороны, образцы иммунной сыворотки и мышам-реципиентам внутрибрюшинно вводят одну дозу сыворотки в количестве 400 мкл. Перед заражением дозой адаптированного вируса Х-47 в количестве 20 LD50 выжидают в течение 24 часов с тем, чтобы молекула антитела системно распространились в организме мыши. В то время как у мышей, которые получали контрольные дозы сыворотки, позднее развивается острая гипотермия и наблюдается быстрая потеря веса, что в конце концов приводит к смерти, введение иммунной сыворотки, содержащей рекомбнантную нейраминидазу, защищает мышей практически так же, как и в случае активной вакцинации (фиг. 14). Отсюда можно сделать вывод, что присутствие уже циркулирующих NА-антител достаточно для обеспечения полной защиты.
Обсуждение результатов
Иммунитет по отношению к гриппу в течение длительного времени изучали практически исключительно как функцию антитела гемагглютинина, в то время как важный вклад в иммунитет нейраминидазы в значительной степени игнорировался. Это частично объясняется тем наблюдением, что лишь антитела, способные связывать гемагглютинин, обладали способностью непосредственно нейтрализовать частицы вируса (Нirst, 1942; Davanport et al., 1964; Kida et al., 1983), в то время как антитела против нейраминидазы в широком диапазоне концентраций очевидно не могли предотвращать первичную инфекцию (Jahiel and Kilbourne, 1966; Kilbourne et al., 1968; Johanson, 1989). Эта толерантность, вероятно, отражает ту недавно выявленную роль, которую играет рекомбинантная нейраминидаза в жизненном цикле вируса гриппа, препятствуя вновь сформировавшемуся вирусу агрегироваться на поверхности инфицированной клетки (Colman and Ward, 1985; Brown and Laver, 1968). Более того, было обнаружено, что в отличие от гемагглютинина нейраминидаза составляет меньшую часть оболочки вируса гриппа и этот факт также может вносить вклад в неспособность антител нейраминидазы оказывать нейтрализующее воздействие (Schulman et al., 1968). Это различие молярном присутствии также оказывает влияние на относительные антительные ответы к индивидуальным антигенам. Повторяющееся доминирование гемагглютинина по отношению к нейраминидазе благодаря успешной конфронтации со всем вирусом гриппа могло привести к подавлению продуцирования антител нейраминидазы, вероятно, вследствие ослабленной помощи, которую оказывают нейраминидаза-специфичные Т-клетки (Kilbourne, 1976; Johanson et al., 1987; Kilbourne et al., 1987; Johanson et al., 1987). С целью изучения защитного иммунитета нейраминидазы, необходимо разработать систему, в которой устраняется мешающее воздействие антител, нейтрализующих гемагглютинин, и удается избежать подавления иммунного ответа нейраминидазы вследствие конкуренции антигенов гемагглютинина и нейраминидазы. Классические подходы базировались либо на выделении компонента природной нейраминидазы (Schulman et al., 1968; Johanson and Kilbourne, 1990; Gallagher et al., 1984), либо же на комбинированном назначении определенных серий штамма гриппа с селологически различающимися антигенами гемагглютинина и нейраминидазы (Rott et al., 1974; Kilbourne, 1976). Полученные же в настоящей работе результаты непосредственно демонстрируют защитную иммунизацию с помощью очищенного рекомбинантного белка нейраминидазы. Ген нейраминидазы вируса A/Victoria/3/75 (Н3N2) трансформируется в ген, который кодирует способный секретироваться белок (рекомбинантную нейраминидазу), путем замещения области, кодирующей мембранный якорь, сигнальной последовательностью гена гемагглютинина гриппа (см. Пример 1).
Методами in vitro уже установлено, что антитела нейраминидазы могут эффективно подавлять рост выхода вируса путем ингибирования выхода и распространения частиц вируса (Jahiel and Kilbourne, 1968; Kilbourne et al., 1968). Аналогичные заключения были сделаны для животных, иммунизированных с помощью нейраминидазы, при измерении уменьшенных титров вируса в легких и снижении вероятности развития легочных поражений (11, 12, 13).
Хотя значительное внимание уделено воздействию NА-иммунитета на репликацию вируса в легких, оставалось под вопросом, может ли иммунизация чистым белком нейраминидазой предотвратить клинические симптомы заболевания или может увеличить шанс выжить после потенциально летальной инфекции гриппа. На этот вопрос пока не было получено удовлетворительного ответа. Приведенные в настоящем описании результаты ясно показывают, что путем иммунизации с помощью чистой рекомбинантной нейраминидазы может быть достигнута полная защита против обычно летальной дозы инфекции, при этом исключается любой возможный вклад со стороны анамнестического антигемагглютининового иммунного механизма или эффектов клеточно-опосредованной иммунной памяти против антигенов консервативных внутренних вирусных белков.
Во время проведения описываемых здесь экспериментов мышей иммунизируют тремя дозами 1 мкг рекомбинантной нейраминидазы, которые назначают с интервалами в три недели. Вакцинированные животные способны полностью пережить летальную инфекцию вируса гриппа, при этом вирус экспрессирует гомо- и гетеровариантную рекомбинантную нейраминидазу. Ввиду высоких доз инфекции вируса было очень удивительно, что у животных после хорошо проведенной иммунизации не наблюдается симптомов заболевания, которые определяют по изменению температуры и веса тела. Важно отметить, что адъюванты, которые назначают вместе с рекомбинантной нейраминидазой, все имеют низкие реактогенные свойства, так что методика иммунизации, приведенная в настоящем описании, непосредственно может быть использована для вакцинации людей. Вакцины по настоящему изобретению могут также использоваться для других млекопитающих и птиц.
Пассивное перенесение сыворотки мышей, иммунизированных рекомбинантной нейраминидазой, мышам-реципиентам приводит к тому же самому уровню защиты, указывая на то, что защитное воздействие иммунизаций с помощью рекомбинантной нейраминидазы может быть объяснено циркулирующими антигенами нейраминидазы.
Что касается рассмотренной в настоящем описании гетеровариантной защиты, то важно рассмотреть структурные связи между антигеном нейраминидазы вакцины А/Victoria/3/75 и нейраминидазы из A/Aichi/2/68, которая присутствует в варианте инфекционного вируса Х-31. К сожалению, не получены данные о последовательности аминокислотных остатков для нейраминидазы из А/Aichi/2/68 (H3N2), однако можно провести сравнение с последовательностью аминокислотных остатков нейраминидазы из штамма А/NТ/60/68 (Н3N2) (Bentley and Brownley, 1982), выделенного в том же году, что и штамм Aichi. При внимательном изучении области головки обоих вариантов нейраминидазы, можно обнаружить замещения аминокислот в положениях 28, при этом большинство их находится на поверхности молекулы.
Вероятно, вакцина по настоящему изобретению также может обеспечить защиту от еще более удаленных за счет дрейфа вариантов. Возможно также, что с помощью генных модификаций гена нейраминидазы варианты могут быть аранжированы в ее антигенную структуру. При этом становится возможным, например, получить "коктейль" различных версий нейраминидазы и добиться интенсивной защиты от различных штаммов гриппа.
Фигуры
На фиг. 1 приведена стратегия конструирования гена секретируемой нейраминидазы и его интеграции в вектор переноса бакуловируса. Указаны только необходимые сайты рестрикции. Тонкими линиями показаны последовательности бактериальной плазмиды, в то время как жирные линии показывают гемагглютинин-специфичные (закрашенные) и нейраминидаза-специфичные (составлены точками) последовательности. Сигнальная последовательность гемагглютинина обозначена одной штриховкой. Сигнальная последовательность нейраминидазы/последовательность мембранного якоря обозначены двойной штриховкой.
На фиг. 2 показана нуклеотидная последовательность позитивной ветви кДНК и последовательность аминокислотных остатков фланкирующих областей сайта лигирования между сигнальным пептидом гемагглютинина и нейраминидазы, при этом мембранный якорь нейраминидазы удален.
На фиг. 2А приведен непроцессированный гемагглютинин и детально сайт рестрикции сигнальной пептидазы между Аlа16 и Gln17 (вертикальная пунктирная линия). Терминальный сегмент гемагглютинина, который используют для секреции рекомбинантной нейраминидазы, обозначен стрелкой.
На фиг. 2В детально показана "стеблевая" область нейраминидазы. Усеченная последовательность, участвующая в конструировании рекомбинантной нейраминидазы, обозначена стрелкой.
На фиг. 2С показано, как последовательность рекомбинантной нейрамидиназы конструируется из А и В. Детально показана область слияния между гемагглютинин-специфичной и нейраминидаза-специфичной последовательностями. Рекомбинантная нейраминидаза, видимо, начинается с четырех терминальных аминокислот нейраминидазы, полученных из зрелого гемагглютинина, за которыми следует подвергнутый мутации кодон (подчеркнут пунктирной линией).
На фиг. 3 приведен результат анализа очищенной природной нейраминидазы, обработанной проназой, методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Фиг. 3А относится к анализу образцов белка, взятых на различных стадиях в процессе очистки природной нейраминидазы, обработанной проназой. Дорожка 1 показывает белки-маркеры; дорожка 2 соответствует сырому материалу природной нейраминидазы, обработанной проназой (1 мкг; полосы природной нейраминидазы, обработанной проназой, ниже предела обнаружения); дорожка 3 показывает объединенный элюат после сефарозы S (1 мкг); дорожка показывает объединенный элюат после Superdex 200 (1 мкг). На фиг. 3В приведен градиентный от 5,0% до 7,5% гель 1 мкг природной нейраминидазы, обработанной проназой, которая перекрестно связана с помощью BS3. Дорожка 1 соответствует белкам-маркерам; дорожка 2 соответствует природной нейраминидазе, обработанной проназой, после перекрестного связывания. Дополнительные полосы появились около 105 кДа (димер), около 160 кДа (тример) и около 210 кДа (тетрамер).
На фиг. 4 показано изменение с течением времени удельной активности  обнаруженной в среде для выращивания культуры, которую оценивают из уровней ферментной активности
обнаруженной в среде для выращивания культуры, которую оценивают из уровней ферментной активности  и общей концентрации белка
и общей концентрации белка  после заражения клеток sf9 рекомбинантным бакуловирусом.
после заражения клеток sf9 рекомбинантным бакуловирусом.
На фиг. 5 показан результат анионообменной хроматографии в колонке с сефарозой Q. После растворения и диализа осадка, полученного с помощью (20-60)% сульфата натрия, раствор (97,5 мг белка, 117000 единиц) помещают в колонку с сефарозой Q. Несвязанные вещества вымываются перед элюированием, которое проводят в градиенте хлорида натрия (---) до концентрации 250 мМ, добавляемого к исходному буферному раствору. После концентрации белка в элюате проводят измерения А280(_____). Собирают фракции по 2,5 мл и изучают ферментативную активность  и антигенность иммуноферментным твердофазным анализом
и антигенность иммуноферментным твердофазным анализом 
На фиг. 6 приведен результат гель-фильтрации рекомбинантной нейраминидазы на Superdex 200. Элюат после разделения на N-(п-аминофенил)оксамовая кислота - агароза (2,63 мг белка, 49100 единиц) концентрируют до объема 2,0 мл и затем хроматографируют, собирая фракции по 1,0 мл, на колонке с Supеrdеx 200 со скоростью подачи 10 мл/ч. Непрерывно проводят контроль значения A280(_ _ _ __). Индивидуальные фракции (1,0 мл) испытывают на ферментативную  а также антигенную активность
а также антигенную активность  Стрелки показывают объем элюции калибровочных белков (см. текст): 443 кДа (1), 200 кДа (2), 150 кДа (3), 67 кДа (4) и 29 кДа (5).
Стрелки показывают объем элюции калибровочных белков (см. текст): 443 кДа (1), 200 кДа (2), 150 кДа (3), 67 кДа (4) и 29 кДа (5).
На фиг. 7 приведен результат анализа методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия очищенной природной нейраминидазы, обработанной проназой. Каждая дорожка соответствует номеру специфической фракции из колонки с Superdex 200 после гель-фильтрации. На фиг. 7А приведена картина электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия после денатурации 10 мкл образца (добавляют бета-меркаптоэтанол). Дорожки А и В соответствуют белкам-маркерам. На фиг. 7В образцы белка перекрестно сшивают с помощью ВS3 и затем разделяют электрофорезом в градиентном геле от 5,0 до 7,5% в присутствии додецилсульфата натрия, но в невосстановительных условиях. Фракции от 57 до 68 показывают объем образца 10 мкл; фракции от 70 до 77 показывают объем образца 25 мкл. Тетрамерная рекобинантная нейраминидаза дает полосы около 220 кДа (тетрамер) и около 110 кДа (димер). Димерная и мономерная рекомбинантная нейраминидаза остаются видимыми в виде полос соответственно около 110 кДа и около 5 кДа.
На фиг. 8 приведен результат анализа методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия образцов белка, взятых на различных стадиях процесса очистки рекомбинантной нейраминидазы. Дорожка 1 соответствует белкам-маркерам; дорожка 2 соответствует сырой среде (5 мкг); дорожка 3 осадку в (20-60)% сульфата натрия (5 мкг); дорожка 4 соответствует объединенному элюату после хроматографии на сефарозе Q (2,5 мкг); дорожка 5 соответствует объединенному элюату после разделения на N-(п-аминофенил)оксамовой кислоте - агарозе (1 мкг); дорожка 6 соответствует объединенному элюату фракций тетрамерной и димерной рекомбинантной нейраминидазы после гель-фильтрации на Superdex 200 (1 мкг).
На фиг. 9 приведен сравнительный анализ содержания углевода, связанного с природной нейраминидазой, обработанной проназой, и углевода, связанного с рекомбинантной нейраминидазой, который проводят на основании ферментации нейраминидаза-глюканазой и анализа электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия. Фермент нейраминидаза-глюканаза виден в виде полосы около 35 кДа. На фиг. 9А дорожка 1 соответствует белкам-маркерам; дорожка 2 - не подвергнутой ферментации природной нейраминидазе (1 мкг); дорожка 3 - природной нейраминидазе, обработанной нейраминидаза-глюканазой (1 мкг). На фиг. 9В дорожки 1 и 8 соответствуют белкам-маркерам; дорожка 2 - не подвергнутой ферментации тетрамерной рекомбинантной нейраминидазе; дорожка 3 - тетрамерной рекомбинантной нейраминидазе, обработанной нейраминидаза-глюканазой; дорожка 4 - не подвергнутой ферментации димерной рекомбинантной нейраминидазе; дорожка 5 - димерной рекомбинантной нейраминидазе, обработанной нейраминидаза-глюканазой; дорожка 6 - не подвергнутой ферментации мономерной рекомбинантной нейраминидазе; дорожка 7 - мономерной рекомбинантной нейраминидазе, обработанной нейраминидаза-глюканазой.
Фиг. 10 показывает близкую к идентичности антигенность рекомбинантной нейраминидазы и природной нейраминидазы, обработанной проназой. Создают равные концентрации белков рекомбинантной нейраминидазы и природной нейраминидазы, а затем последовательно разбавляют в соотношении 1:2 и проводят иммуноферментный твердофазный анализ. На чертеже представлена синусообразная кривая антигенности, полученная для специальных антигенов. Значения для природной нейраминидазы, обработанной проназой, обозначены  ; значения для тетрамерной рекомбинантной нейраминидазы обозначены
; значения для тетрамерной рекомбинантной нейраминидазы обозначены  ; значения для димерной рекомбинантной нейраминидазы обозначены
; значения для димерной рекомбинантной нейраминидазы обозначены  ; и значения для мономерной рекомбинантной нейраминидазы обозначены
; и значения для мономерной рекомбинантной нейраминидазы обозначены 
Антигенный отклик на рекомбинантную нейраминидазу приведен на фиг. 11. Через четырнадцать дней после каждой иммунизации (показаны стрелками) у мышей берут образцы крови и присутствие антител рекомбинантной нейраминидазы определяют методом иммуноферментного твердофазного анализа (детали эксперимента приведены в тексте). Полностью закрашенные прямоугольники и заштрихованные прямоугольники соответствуют средним титрам (± стандартное отклонение) соответственно вакцинированных и контрольных животных. На фиг. 12 показана гомовариантная защита. Вакцинированным [--- в (А);  в (В) и (C)] и контрольным [_ __ в (А); • в (В) и (С)] животным назначают дозу 20 LD50 гомовариантного адаптированного для мышей вируса Х-47. За развитием инфекции следят, регистрируя уровень смертности (А) и измеряя ректальную температуру (В) и вес тела (С) мышей (см. детали эксперимента в тексте). Цифровые значения являются средними (± стандратное отклонение).
в (В) и (C)] и контрольным [_ __ в (А); • в (В) и (С)] животным назначают дозу 20 LD50 гомовариантного адаптированного для мышей вируса Х-47. За развитием инфекции следят, регистрируя уровень смертности (А) и измеряя ректальную температуру (В) и вес тела (С) мышей (см. детали эксперимента в тексте). Цифровые значения являются средними (± стандратное отклонение).
Гетеровариантная защита показана на фиг. 13. Вакцинированным [--- в (А);  в (В) и (С)] и контрольным [___ в (А); • в (В) и (С)] животным назначают дозу 20 LD50 гетеровариантного адаптированного для мышей вируса Х-31. За развитием инфекции следят, регистрируя уровень смертности (А) и измеряя ректальную температуру (В) и вес тела (С) мышей (см. детали эксперимента в тексте). Цифровые значения являются средними (± стандартное отклонение).
в (В) и (С)] и контрольным [___ в (А); • в (В) и (С)] животным назначают дозу 20 LD50 гетеровариантного адаптированного для мышей вируса Х-31. За развитием инфекции следят, регистрируя уровень смертности (А) и измеряя ректальную температуру (В) и вес тела (С) мышей (см. детали эксперимента в тексте). Цифровые значения являются средними (± стандартное отклонение).
Фиг. 14 иллюстрирует пассивную иммунизацию. Группы мышей пассивно иммунизируют с помощью внутрибрюшинной инъекции иммунной сыворотки рекомбинантной нейраминидазы [--- в (А);  в (В) и (C)] и контрольной сыворотки [__ _ в (А); • в (В) и (С)]. Через двадцать четыре часа мышам вводят дозу 20 LD50 адаптированного для мышей вируса Х-47 (см. детали эксперимента в тексте). Показаны соответственно уровень смертности (А), ректальная температура (В) и вес тела (С) мышей. Цифровые значения являются средними (± стандартное отклонение).
в (В) и (C)] и контрольной сыворотки [__ _ в (А); • в (В) и (С)]. Через двадцать четыре часа мышам вводят дозу 20 LD50 адаптированного для мышей вируса Х-47 (см. детали эксперимента в тексте). Показаны соответственно уровень смертности (А), ректальная температура (В) и вес тела (С) мышей. Цифровые значения являются средними (± стандартное отклонение).
На фиг. 15 приведена диаграмма плазмиды рР1С9, которая помимо последовательности промотора АОХ1 и терминатора содержит маркер H1S4 из Р.pastoris и сигнал секреции препро-гена альфа-фактора из Saccharomyces cerevisiae. Позади сигнала секреции расположен составной сайт клонирования.
На фиг. 16 дан обзор режимов слияния между препро-сигналом секреции и рекомбинантной частью "шляпки" нейраминидазы. "КЕХ2" указывает место, где пропептид расщепляется по Гольджи с помощью эндогенной КЕХ-2-протеазы. Дипептид (Glu-Ala)2 удаляется с помощью дипептидиламинопептидазы типа STЕ13. Остаток тирозина не привносится из нейраминидазы, однако не удаляется. Следующий пролин соответствует позиции 79 нейраминидазы из Х-47.
На фиг. 17 приведен анализ по методу вестерн-блоттинга на 12,5%-ном полиакриламидном геле пяти образцов среды индивидуальных трансформированных клеток. Дорожка 1 содержит образец среды нетрансформированного штамма Р.pastoris. На каждую дорожку помещают вещество, выделенное из 1 мл среды для выращивания культуры путем осаждения с помощью трихлоруксусной кислоты.
Табл. 1 относится к одному типичному эксперименту по очистке (детали см. в тексте). Объем после Superdex 200 соответствует объединенному количеству после двух операций хроматографии.
Табл. 2 относится к одному типичному эксперименту по очистке (детали см. в тесте). Указанные объемы после гель-фильтрации на Superdex 200 соответствует объединенным фракциям рекомбинантной нейраминидазы после двух операций хроматографии.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПСЕВДОАДЕНОВИРУСНАЯ ЧАСТИЦА НА ОСНОВЕ ГЕНОМА АДЕНОВИРУСА ЧЕЛОВЕКА 5 СЕРОТИПА, ПРОДУЦИРУЮЩАЯ ГЕМАГГЛЮТИНИН ВИРУСА ГРИППА ШТАММА A/Brisbane/59/2007(H1N1) И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ | 2012 |
|
RU2523599C1 |
| MDCK клетка-продуцент белков вируса гриппа (варианты) | 2018 |
|
RU2681482C1 |
| Экспрессионный вектор на основе аденовируса человека 5 серотипа, индуцирующий кросс-протективный иммунитет к вирусам гриппа А субтипа Н1, и фармацевтическая композиция на его основе | 2023 |
|
RU2802753C1 |
| ЭФФЕКТИВНЫЕ ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ (VLPS) ГРИППА | 2004 |
|
RU2369405C2 |
| Вирусоподобная частица вируса гриппа и способ ее получения | 2018 |
|
RU2681439C1 |
| КОНСТРУКЦИЯ ДНК (ВАРИАНТЫ), ДНК-ВЕКТОР, ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГРИППА, СПОСОБ ИНДУКЦИИ ИММУННОГО ОТВЕТА, ВАКЦИНА И СПОСОБ ВАКЦИНАЦИИ | 1994 |
|
RU2193065C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИВАЛЕНТНОЙ ВАКЦИНЫ ОТ ГРИППА | 2018 |
|
RU2701953C1 |
| КОНЦЕНТРАТ ВАКЦИНЫ ОТ ГРИППА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2018 |
|
RU2706544C1 |
| ПОЛИВАЛЕНТНАЯ ВАКЦИНА ПРОТИВ ГРИППА | 2018 |
|
RU2706191C1 |
| Кассета, предназначенная для получения плазмидных векторов, используемых для создания клеток-продуцентов вирусоподобных частиц (ВПЧ) вируса гриппа | 2018 |
|
RU2680703C1 |











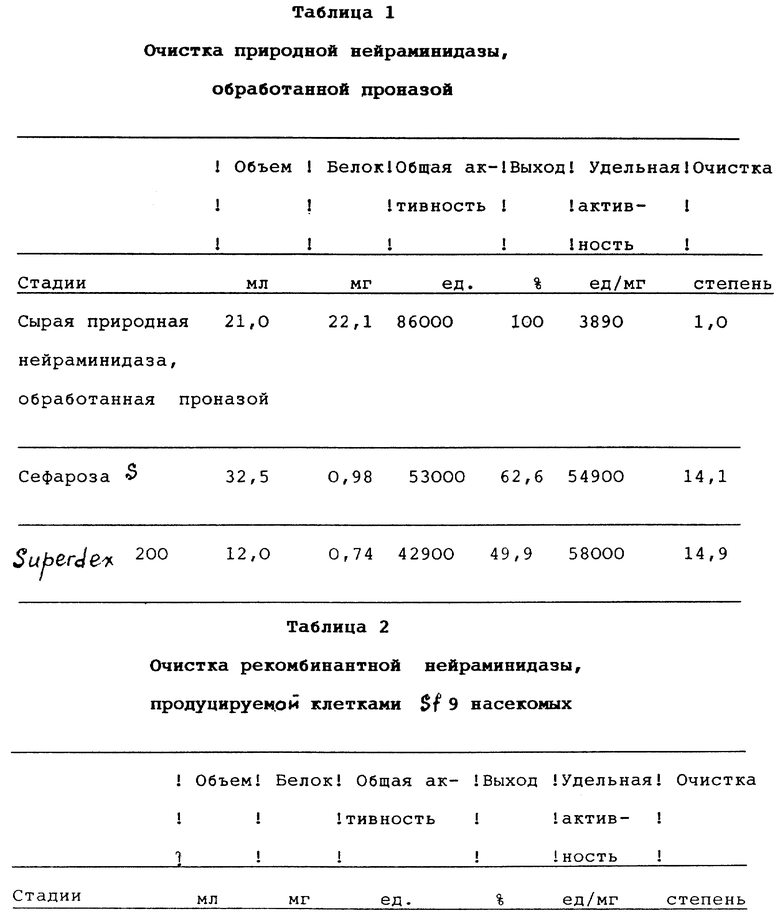
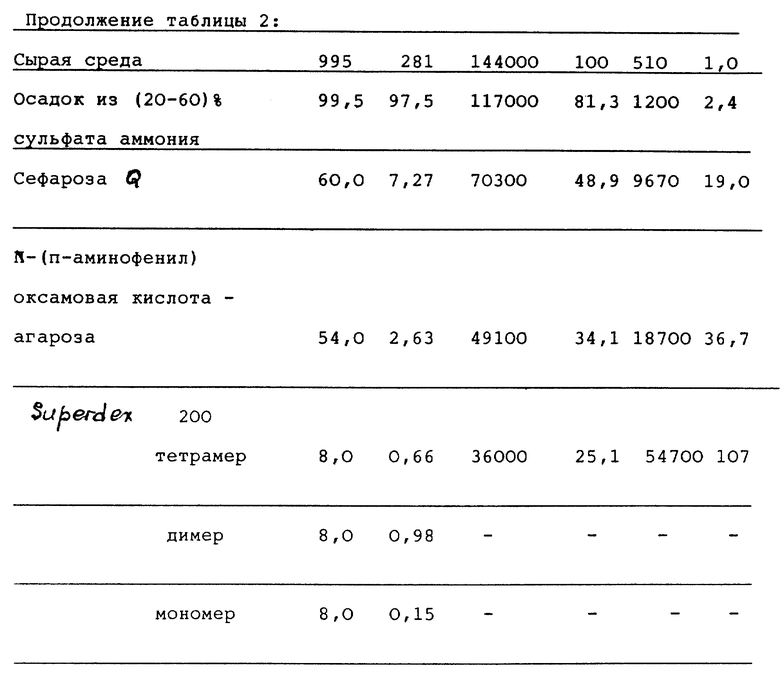
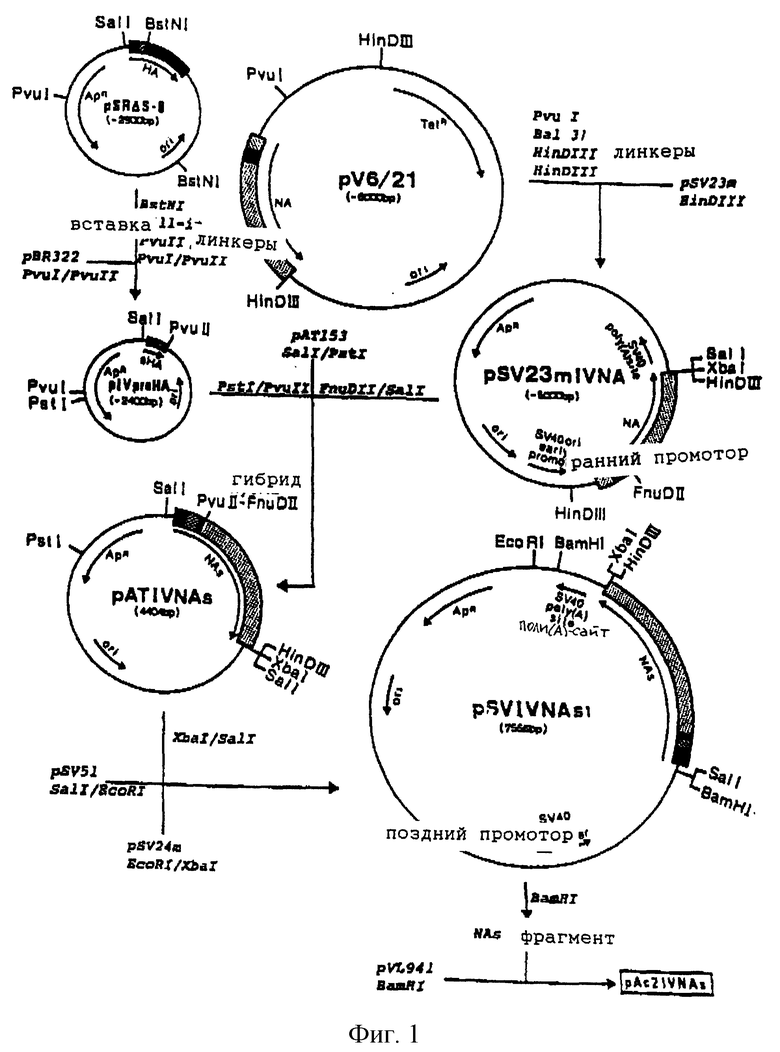
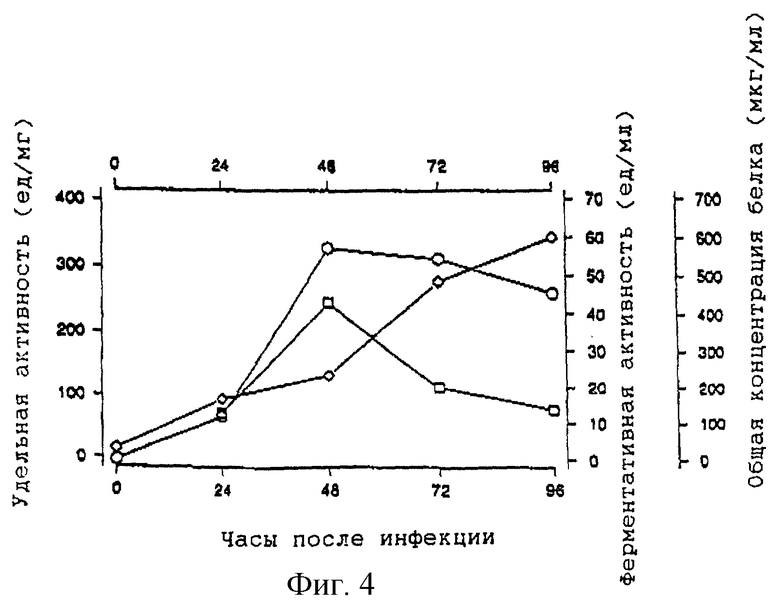
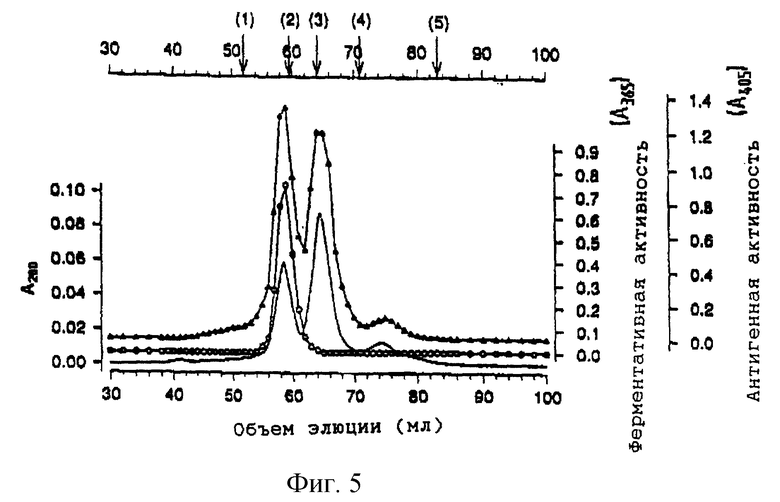
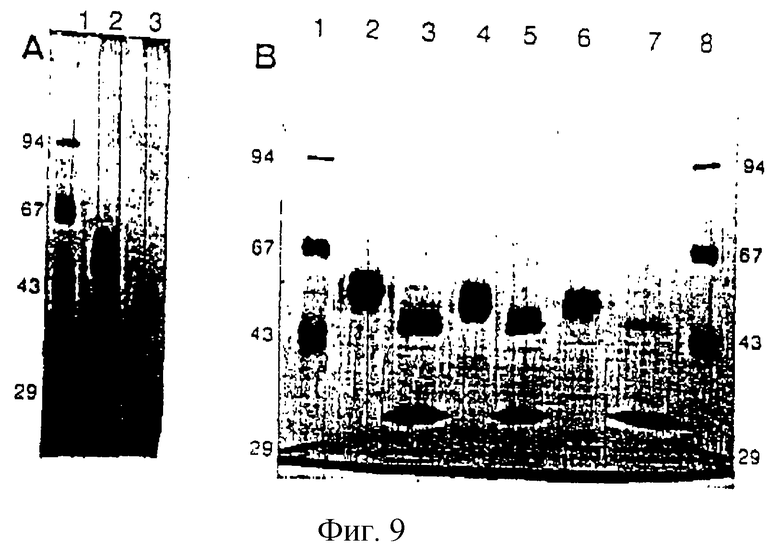
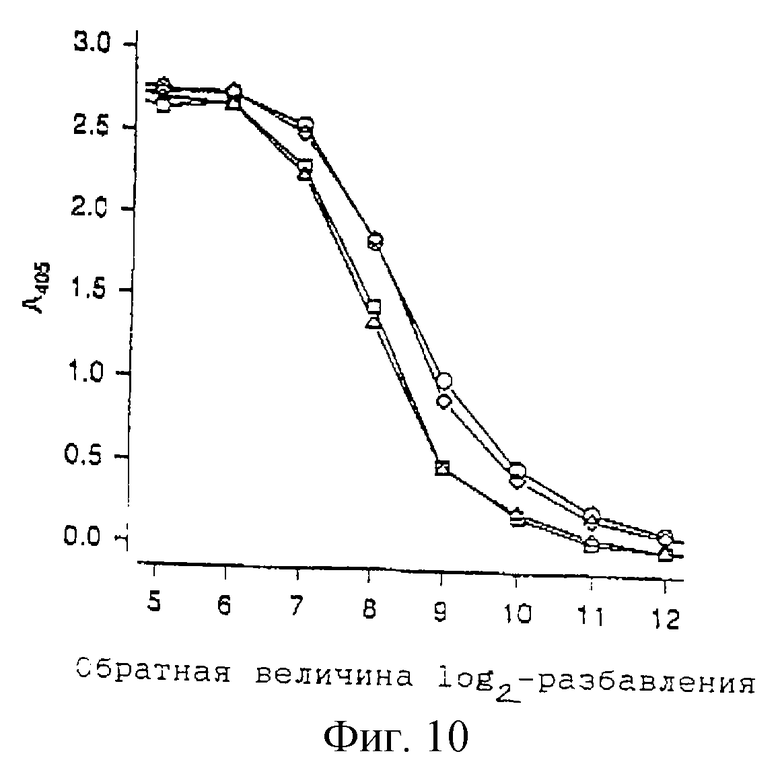
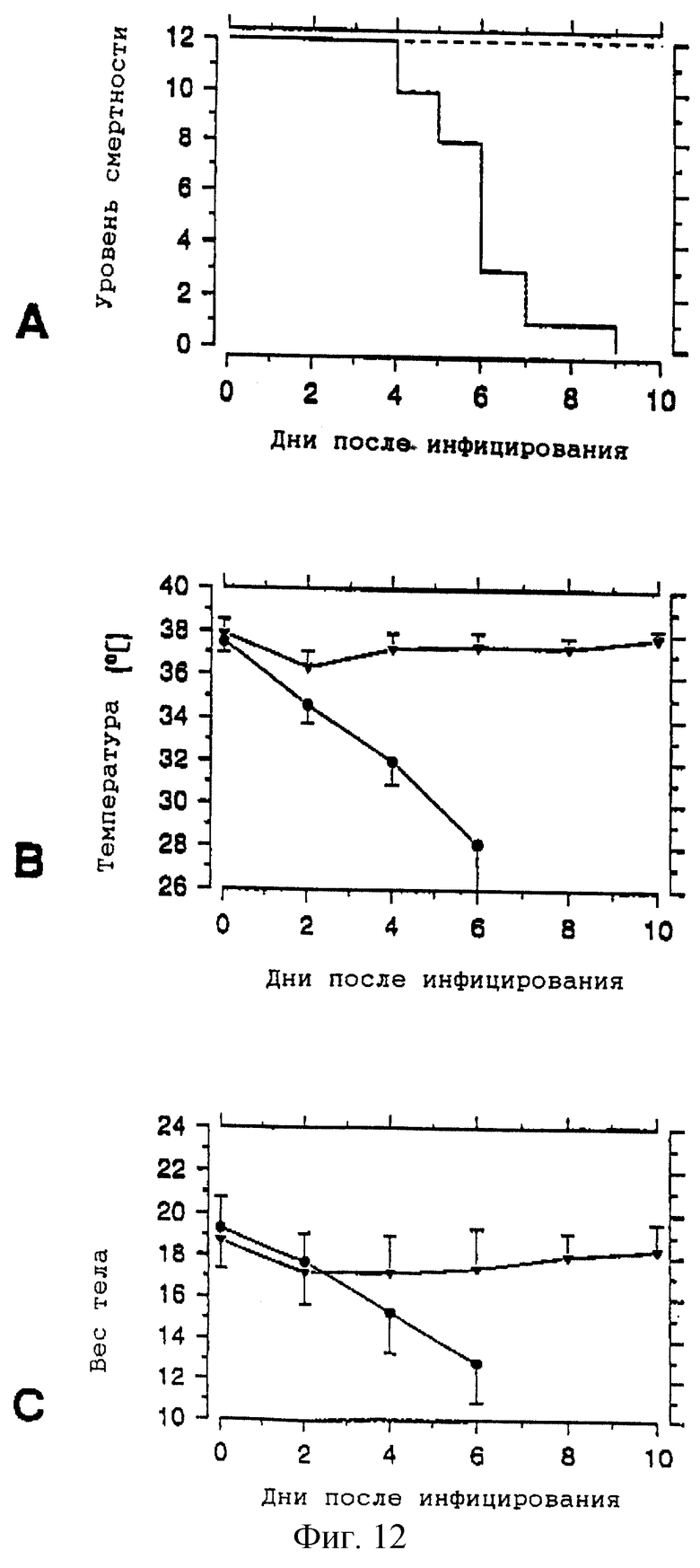
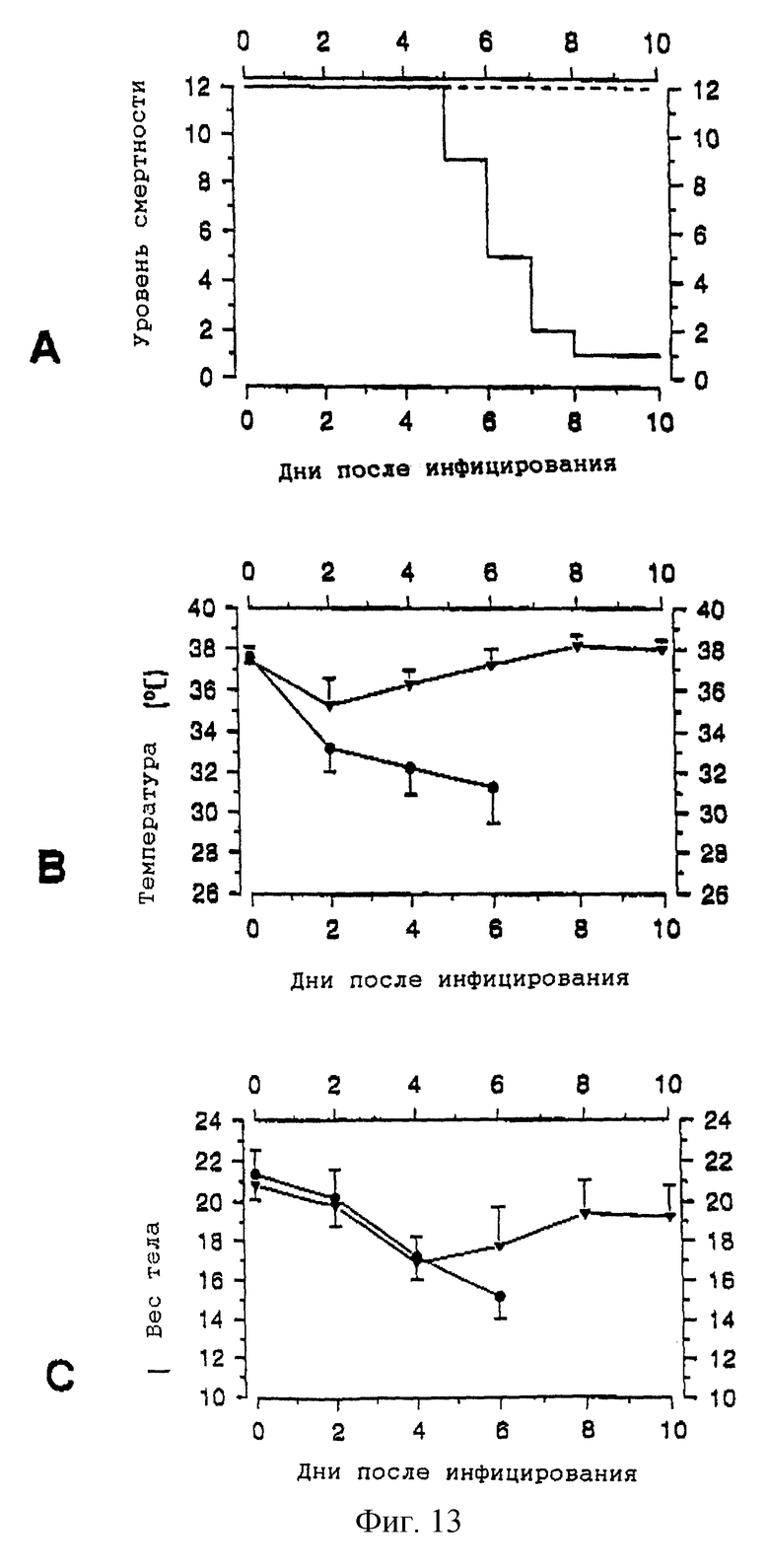
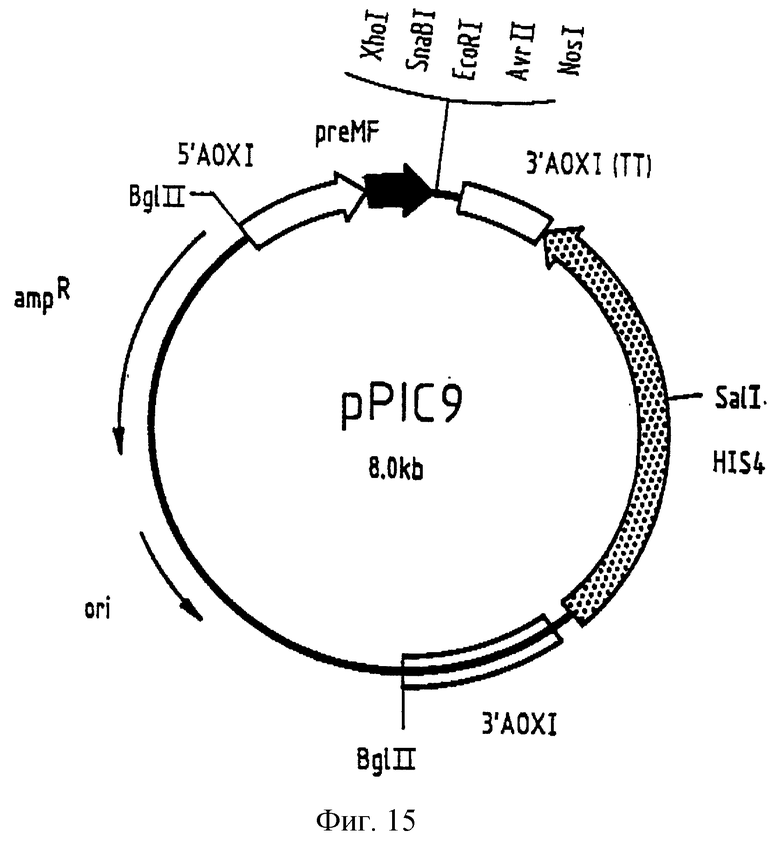
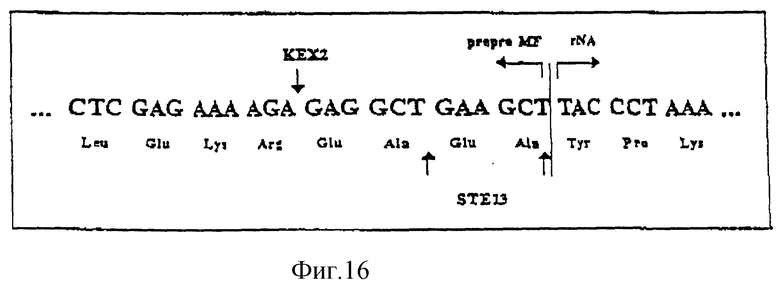
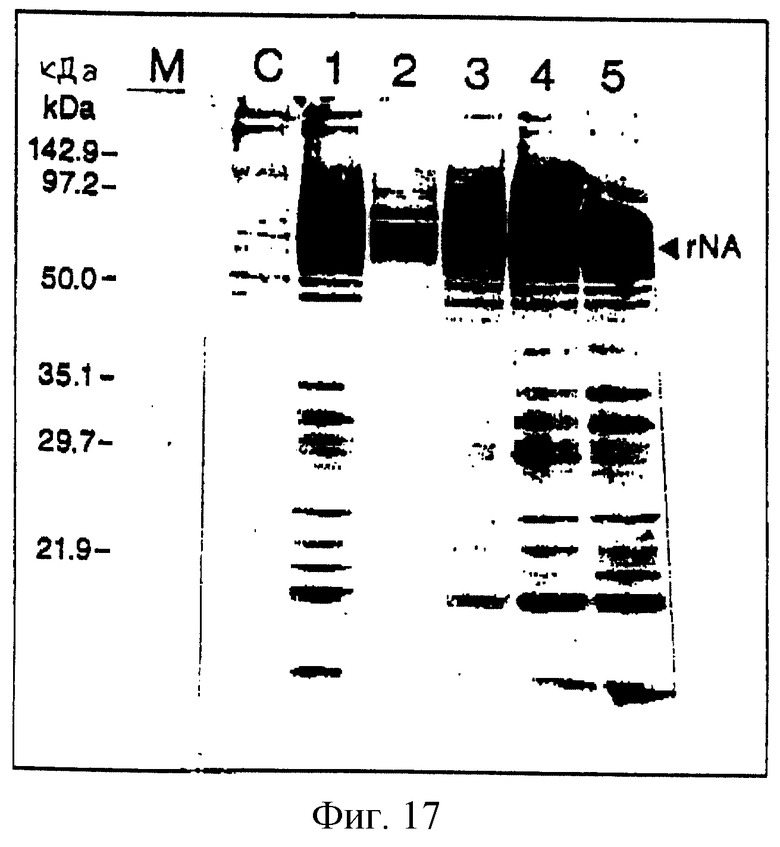
Изобретение относится к генной инженерии. Рекомбинантную нейраминидазу гриппа типа NA2 получают культивированием в подходящей среде инфицированных клеток насекомого или дрожжевых клеток. Клетки насекомого инфицируют вирусом, трансформированным посредством гомологичной рекомбинации его генома с помощью вектора экспрессии рAc21VNAS. Дрожжевые клетки трансформируют вектором экспрессии рРР1IVNAf1S. Рекомбинантную нейраминидазу используют в вакцине против гриппа. Изобретение позволяет получать вакцину, эффективную против вируса гриппа типа NA2. 2 с. и 3 з.п. ф-лы, 17 ил., 2 табл.
| WO 8607383 А1, 15.07.1986 | |||
| CRISO A.D | |||
| et al | |||
| Modification of infections bursal disease virus antigen VP for cell location fails to enhance immunogenicity | |||
| Virus-Res | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| МАНИАТИС Г | |||
| и др | |||
| Методы генетической инженерии | |||
| - М.: Мир, 1984, с.9. | |||
