ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится, inter alia, к новым вариантам исходных Термамил-подобных альфа-амилаз, в особенности к вариантам, проявляющим измененный характер расщепления (относительно исходной альфа-амилазы), которые являются выгодными в отношении применений этих вариантов, в частности в промышленной переработке крахмала (например, разжижении или осахаривании крахмала).
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Альфа-амилазы (альфа-1,4-глюкан-4-глюканогидролазы, ЕС 3.2.1.1) составляют группу ферментов, которые катализируют гидролиз крахмала и других неразветвленных или разветвленных 1,4-глюкозидных олиго- и полисахаридов.
Имеется большая масса патентной и научной литературы, относящейся к этому очень важному в промышленном отношении классу ферментов. Ряд альфа-амилаз, таких как варианты Термамил-подобных альфа-амилаз, известны, например, из WO 90/11352, WO 95/10603, WO 95/26397, WO 96/23873, WO 96/23874 и WO 97/41213.
Среди недавних описаний, относящихся к альфа-амилазам, в WO 96/23874 приведены данные по трехмерной, рентгеновской кристаллографии для Термамил-подобной альфа-амилазы, называемой ВА2, которая состоит из 300 N-концевых аминокислотных остатков альфа-амилазы В. amyloliquefaciens, содержащей аминокислотную последовательность, показанную здесь в SEQ ID NO:6, и С-концевых аминокислот 301-483 альфа-амилазы В. licheniformis, содержащей аминокислотную последовательность, показанную здесь в SEQ ID NO:4 (последняя доступна коммерчески под товарным названием Термамил™ ), и которая, следовательно, является близкородственной промышленно важным альфа-амилазам Bacillus (которые в данном контексте охватываются понятием "Термамил-подобные альфа-амилазы" и которые включают в себя, inter alia, альфа-амилазы В. licheniformis, В. amyloliquefaciens и В. stearothermophilus). В WO 96/23874 описана, кроме того, методология для конструирования на основе анализа структуры исходной Термамил-подобной альфа-амилазы вариантов исходной Термамил-подобной альфа-амилазы, которые проявляют измененные свойства относительно исходного фермента.
В WO 96/23874 и WO 97/41213 (Novo Nordisk) описаны варианты Термамил-подобной альфа-амилазы с измененным характером расщепления, содержащие мутации в аминокислотных остатках V54, D53, Y56, Q333, G57 и А52 последовательности, показанной здесь в SEQ ID NO:4.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к новым альфа-амилолитическим вариантам (мутантам) Термамил-подобной альфа-амилазы, в частности вариантам, проявляющим измененный характер расщепления (относительно исходного фермента), которые являются выгодными в связи с технологией производства крахмала (разжижения крахмала, осахаривания крахмала и т.п.).
Авторы изобретения неожиданно обнаружили варианты с измененными свойствами, в частности измененным характером расщепления, которые имеют пониженную способность расщепления субстрата вблизи точки ветвления и, кроме того, имеют повышенную субстратную специфичность и/или повышенную удельную активность, в сравнении с описанными в WO 96/23874 вариантами Термамил-подобной альфа-амилазы с измененным характером расщепления, содержащими мутации в аминокислотных остатках V54, D53, Y56, Q333, G57 и А52 последовательности, показанной здесь в SEQ ID NO:4.
Далее, данное изобретение относится к конструкциям ДНК, кодирующим варианты данного изобретения, композициям, содержащим варианты данного изобретения, способам получения вариантов данного изобретения и применению вариантов и композиций данного изобретения, отдельно или в комбинации с другими альфа-амилолитическими ферментами, в различных промышленных процессах, например в разжижении крахмала, и в детергентных композициях, таких как композиции для стирки белья, мытья посуды и чистки твердых поверхностей; в производстве этанола, например в производстве топливного, питьевого и промышленного этанола; в расшлихтовке текстильных изделий, тканей или одежды и т.д.
Номенклатура
В данном описании и формуле изобретения используются общепринятые однобуквенные и трехбуквенные коды для аминокислотных остатков. Для легкости ссылки варианты альфа-амилазы данного изобретения описаны с использованием следующей номенклатуры:
Исходная аминокислота (аминокислоты):положение (положения): замененная аминокислота (аминокислоты)
Согласно этой номенклатуре, например, замена аланина на аспарагин в положении 30 представляется как:
Ala30Asn или A30N
делеция аланина в том же самом положении представляется как:
Аlа30* или А30*
и инсерция дополнительного аминокислотного остатка, такого как лизин, представляется как:
*30aLys или *30аК
Делеция последовательного отрезка аминокислотных остатков, таких как аминокислотные остатки 30-33, указывается как (30-33)* или Δ (А30-N33) или дельта(A30-N33).
Если конкретная альфа-амилаза содержит "делецию" в сравнении с другими альфа-амилазами и произведена инсерция в таком положении, это указывается как:
*36aAsp или *36aD
для инсерции аспарагиновой кислоты в положении 36.
Множественные мутации разделяются знаками плюс, т.е.:
Ala30Asp+Glu34Ser или A30N+E34S
что обозначает мутации в положениях 30 и 34, заменяющие аланин и глутаминовую кислоту на аспарагин и серин соответственно. Множественные мутации могут также разделяться следующим образом, т.е. обозначая то же самое, что и знак плюс:
Ala30Asp/Glu34Ser или A30N/E34S
При встраивании одного или нескольких альтернативных аминокислотных остатков в конкретное положение это указывается как
A30N,E или
A30N или А30Е
Кроме того, когда положение, пригодное для модификации, идентифицируется здесь без предложения какой-либо конкретной модификации, или А30Х, следует понимать, что любой аминокислотный остаток может быть заместителем для аминокислотного остатка, присутствующего в этом положении. Так, например, при упоминании модификации аланина в положении 30, но без уточнения или указания в виде X, должно быть понятно, что этот аланин может быть делегирован или заменен любой другой аминокислотой, т.е. любой из R, N, D, С, Q, Е, G, H, I, L, К, М, F, Р, S, Т, W, Y, V.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Термамил-подобная альфа-амилаза
Хорошо известно, что ряд альфа-амилаз, продуцируемых Bacillus spp., являются высокогомологичными на аминокислотном уровне. Например, было обнаружено, что альфа-амилаза В. licheniformis, содержащая аминокислотную последовательность, показанную в SEQ ID NO:4 (коммерчески доступная как Термамил™ ), является гомологичной приблизительно на 89% альфа-амилазе В. amyloliquefaciens, содержащей аминокислотную последовательность, показанную в SEQ ID NO:6, и гомологичной приблизительно на 79% альфа-амилазе В. stearothermophilus, содержащей аминокислотную последовательность, показанную в SEQ ID NO:8. Дополнительные гомологичные альфа-амилазы включают в себя альфа-амилазу, полученную из штамма Bacillus sp. NCIB 12289, NCIB 12512, NCIB 12513 или DSM 9375, все они описаны подробно в WO 95/26397, и альфа-амилазу № 707, описанную Tsukamoto et al., Biochemical and Biophysical Research Communications, 151 (1988), pp. 25-31.
Другие гомологичные альфа-амилазы включают в себя альфа-амилазу, продуцируемую штаммом В. licheniformis, описанным в ЕР 0252666 (АТСС 27811), и альфа-амилазы, идентифицированные в WO 91/00353 и WO 94/18314. Другими коммерческими Термамил-подобными альфа-амилазами В. licheniformis являются Optitherm™ и Takatherm™ (доступные из Solvay), Maxamyl™ (доступная из Gist-Brocades/Genencor), Spezym АА™ и Spezyme Delta AA™ (доступные из Genencor), и Keistase™ (доступная из Daiwa).
Вследствие существенной гомологии, обнаруженной между этими альфа-амилазами, считают, что они принадлежат к одному и тому же классу альфа-амилаз, а именно классу "Термамил-подобных альфа-амилаз".
Таким образом, в данном контексте, термин "Термамил-подобная альфа-амилаза" обозначает альфа-амилазу, которая на аминокислотном уровне проявляет существенную гомологию относительно Термамил™ , т.е. альфа-амилазы В. licheniformis, имеющей аминокислотную последовательность, показанную здесь в SEQ ID NO:4. Другими словами, Термамил-подобная альфа-амилаза представляет собой альфа-амилазу, которая имеет последовательность, показанную здесь в SEQ ID NO:2, 4, 6 или 8, и аминокислотную последовательность, показанную в SEQ ID NO:1 или 2 WO 95/26397 или в Tsukamoto et al., 1988, или i) которая обнаруживает по меньшей мере 60%-ную, по меньшей мере 70%-ную, предпочтительно по меньшей мере 75%-ную, более предпочтительно по меньшей мере 80%-ную, более предпочтительно по меньшей мере 85%-ную, особенно предпочтительно по меньшей мере 90%-ную, еще более предпочтительно по меньшей мере 95%-ную гомологию, более предпочтительно по меньшей мере 97%-ную, более предпочтительно по меньшей мере 99%-ную гомологию по меньшей мере с одной из указанных аминокислотных последовательностей, и/или ii) проявляет иммунологическую перекрестную реактивность с антителом, индуцированным по меньшей мере против одной из указанных альфа-амилаз, и/или iii) кодируется последовательностью ДНК, которая гибридизуется с последовательностями ДНК, кодирующими указанные выше альфа-амилазы, которые представлены в SEQ ID NO:1, 3, 5 и 7 данной заявки и SEQ ID NO: 4 и 5 WO 95/26397 соответственно.
В связи со свойством i) "гомология" может быть определена с использованием любого общепринятого алгоритма, предпочтительно с использованием программы GAP из версии 7.3 пакета GCG (июнь 1993) с использованием величин штрафа за вставку по умолчанию, которые равны величинам штрафа за открытие вставки 3,0 и штрафа за удлинение вставки 0,1 (Genetic Computer Group (1991) Programme Manual for the GCG Package, version 7, 575 Science Drive, Madison, Wisconsin, USA 53711).
Структурное сопоставление между альфа-амилазой Термамил и Термамил-подобной альфа-амилазой может быть использовано для идентификации эквивалентных/соответствующих положений в других Термамил-подобных альфа-амилазах. Одним из способов получения указанного структурного сопоставления является применение программы Pile up из пакета GCG с использованием величин штрафа за вставку по умолчанию, т.е. штрафа за открытие вставки 3,0 и штрафа за удлинение вставки 0,1. Другие способы структурного выравнивания включают в себя гидрофобный кластерный анализ (Gaboriaud et al., (1987, FEBS LETTERS 224, pp. 149-155) и обратный фолдинг (Huber, Т; Torda, AE, PROTEIN SCIENCE Vol. 7, No. 1, pp. 142-149 (1998). Свойство ii) альфа-амилазы, т.е. иммунологическая перекрестная реактивность, может анализироваться с использованием антитела, индуцированного по меньшей мере против одного эпитопа или реактивного по меньшей мере с одним эпитопом релевантной Термамил-подобной альфа-амилазы. Это антитело, которое может быть моноклональным или поликлональным, может быть получено способами, известными в данной области, например, как описано Hudson et al., Practical Immunology, Third edition (1989), Blackwell Scientific Publications. Иммунологическая перекрестная реактивность может быть определена анализами, известными в данной области, примерами которых являются Вестерн-блоттинг или анализ радиальной иммунодиффузии, например, описанный Hudson et al., 1989. В этом отношении была обнаружена иммунологическая перекрестная реактивность между альфа-амилазами, имеющими последовательности SEQ ID NO:2, 4, 6 или 8 соответственно.
Олигонуклеотидный зонд, используемый в характеристике Термамил-подобной альфа-амилазы в соответствии со свойством iii), указанным выше, может быть легко получен на основе полной или частичной нуклеотидной или аминокислотной последовательности рассматриваемой альфа-амилазы.
Подходящие условия для тестирования гибридизации включают в себя предварительное замачивание в 5× SSC и предгибридизацию в течение 1 часа при -40° С в растворе 20% формамида, 5х растворе Денхардта, 50 мМ фосфате натрия, рН 6,8, и 50 мг денатурированной обработанной ультразвуком ДНК тимуса теленка, с последующей гибридизацией в том же самом растворе, дополненном 100 мМ АТФ, в течение 18 час при -40° С, с последующими тремя промывками фильтра 2× SSC, 0,2% ДСН при 40° С в течение 30 мин (низкая жесткость), предпочтительно при 50° С (умеренная жесткость), более предпочтительно при 65° С (высокая жесткость), даже более предпочтительно при ~75° С (очень высокая жесткость). Более подробные детали относительно способа гибридизации могут быть найдены в Sambrook et al., Molecular Cloning. A Laboratory Manual, 2nd Ed., Cold Spring Harbor, 1989.
В данном контексте "происходящая из" обозначает не только альфа-амилазу, продуцированную или продуцируемую рассматриваемым штаммом организма, но также альфа-амилазу, кодируемую последовательностью ДНК, выделенной из такого штамма, и продуцируемую в организме-хозяине, трансформированном указанной последовательностью ДНК. Наконец, этот термин обозначает альфа-амилазу, которая кодируется последовательностью ДНК синтетического происхождения или кДНК-происхождения и которая имеет идентифицирующие характеристики рассматриваемой альфа-амилазы.
Этот термин указывает также, что исходная альфа-амилаза может быть вариантом природной альфа-амилазы, т.е. вариантом, который является результатом модификации (инсерции, замены, делении) одного или нескольких аминокислотных остатков природной альфа-амилазы.
Исходные гибридные альфа-амилазы.
Исходная альфа-амилаза может представлять собой гибридную альфа-амилазу, т.е. альфа-амилазу, которая содержит комбинацию частичных аминокислотных последовательностей, происходящих по меньшей мере из двух альфа-амилаз.
Исходная гибридная альфа-амилаза может быть альфа-амилазой, которая может быть определена на основе аминокислотной гомологии и/или иммунологической перекрестной реактивности и/или гибридизации ДНК (как определено выше) как принадлежащая к семейству Термамил-подобных альфа-амилаз. В этом случае гибридная альфа-амилаза обычно состоит по меньшей мере из одной части Термамил-подобной альфа-амилазы и части (частей) одной или нескольких других альфа-амилаз, выбранных из Термамил-подобных альфа-амилаз или не-Термамил-подобных альфа-амилаз микробного (бактериального или грибкового) происхождения и/или происходящих из млекопитающих.
Таким образом, исходная гибридная альфа-амилаза может содержать комбинацию частичных последовательностей, происходящих по меньшей мере из двух Термамил-подобных альфа-амилаз или по меньшей мере из одной Термамил-подобной альфа-амилазы и по меньшей мере одной грибковой альфа-амилазы. Термамил-подобная альфа-амилаза, из которой происходит частичная аминокислотная последовательность, может быть, например, любой из указанных здесь Термамил-подобных альфа-амилаз.
Например, исходная альфа-амилаза может содержать С-концевую часть альфа-амилазы, происходящей из штамма В. licheniformis, и N-концевую часть альфа-амилазы, происходящую из штамма В. amyloliquefaciens или из штамма В. stearothermophilus. Например, исходная альфа-амилаза может содержать по меньшей мере 430 аминокислотных остатков С-концевой части альфа-амилазы В. licheniformis и может содержать, например, а) аминокислотный сегмент, соответствующий 37 N-концевым аминокислотным остаткам альфа-амилазы В. amyloliquefaciens, имеющей аминокислотную последовательность, показанную в SEQ ID NO:6, и аминокислотный сегмент, соответствующий 445 С-концевым остаткам альфа-амилазы В. licheniformis, имеющей аминокислотную последовательность, показанную в SEQ ID NO:4, или b) аминокислотный сегмент, соответствующий 68 N-концевым аминокислотным остаткам альфа-амилазы В. stearothermophilus, имеющей аминокислотную последовательность, показанную в SEQ ID NO:8, и аминокислотный сегмент, соответствующий 415 С-концевым аминокислотным остаткам альфа-амилазы В. licheniformis, имеющей аминокислотную последовательность, показанную в SEQ ID NO:4.
В предпочтительном варианте исходная Термамил-подобная альфа-амилаза является гибридной Термамил-подобной альфа-амилазой, идентичной альфа-амилазе В. licheniformis, показанной в SEQ ID NO:4, за исключением того, что 35 N-концевых аминокислотных остатков (зрелый белок) заменены 33 N-концевыми аминокислотными остатками зрелого белка альфа-амилазы В. amyloliquefaciens (BAN), показанной в SEQ ID NO:6. Указанный гибрид может дополнительно иметь следующие мутации: H156Y+A181T+N190F+A209V+Q264S (с использованием нумерации в SEQ ID NO:4), и этот гидрид назван LE174.
Другой предпочтительной исходной альфа-амилазой является LE429, показанная в SEQ ID NO:2.
Не-Термамил-подобной альфа-амилазой может быть, например, грибковая альфа-амилаза, альфа-амилаза млекопитающего или растения или бактериальная альфа-амилаза (отличающаяся от Термамил-подобной альфа-амилазы). Конкретные примеры таких альфа-амилаз включают в себя альфа-амилазу ТАКА Aspergillus oryzae, альфа-амилазу A. niger, альфа-амилазу Bacillus subtilis, свиную панкреатическую альфа-амилазу и альфа-амилазу ячменя. Все эти альфа-амилазы имеют выясненные структуры, которые заметно отличаются от структуры типичной Термамил-подобной альфа-амилазы, на которую здесь ссылаются.
Грибковые альфа-амилазы, указанные выше, т.е. происходящие из А. niger и A. oryzae, являются высокогомологичными на аминокислотном уровне и обычно считаются принадлежащими к тому же самому семейству альфа-амилаз. Грибковая альфа-амилаза, происходящая из Aspergillus oryzae, является коммерчески доступной под товарным названием Фунгамил™ .
Кроме того, при упоминании конкретного варианта Термамил-подобной альфа-амилазы (варианта данного изобретения) - общепринятым образом - ссылкой на модификацию (например, делецию или замену) конкретных аминокислотных остатков в аминокислотной последовательности конкретной Термамил-подобной альфа-амилазы, должно быть понятно, что сюда включены и варианты другой Термамил-подобной альфа-амилазы, модифицированной в эквивалентном положении (положениях) (как определено из сопоставления наиболее возможных аминокислотных последовательностей между соответствующими аминокислотными последовательностями).
Предпочтительный вариант данного изобретения является вариантом, происходящим из альфа-амилазы В. licheniformis (в качестве исходной Термамил-подобной альфа-амилазы), например одним из указанных выше, таким как альфа-амилаза В. licheniformis, имеющая аминокислотную последовательность, показанную в SEQ ID NO:4.
Конструирование вариантов изобретения
Конструирование представляющего интерес варианта может быть выполнено культивированием микроорганизма, содержащего последовательность ДНК, кодирующую этот вариант, в условиях, которые являются благоприятными для получения данного варианта. Затем этот вариант может быть извлечен из полученной культуральной среды. Это описано подробно дополнительно ниже.
Измененные свойства
Далее обсуждается взаимосвязь между мутациями, которые могут присутствовать в вариантах данного изобретения, и желательными изменениями в свойствах (относительно свойств исходной Термамил-подобной альфа-амилазы), которые могут возникать из них.
В первом аспекте данное изобретение относится к варианту исходной Термамил-подобной альфа-амилазы, содержащему изменение в одном или нескольких положениях, выбранных из группы:
W13, G48, Т49, S50, Q51, А52, D53, V54, G57, G107, G108, A111, S168, M197, где (а) это изменение (эти изменения) представляют собой независимо:
(i) инсерцию аминокислоты справа от аминокислоты, которая занимает это положение;
(ii) делецию аминокислоты, которая занимает это положение, или
(iii) замену аминокислоты, которая занимает это положение, отличающейся аминокислотой,
(b) этот вариант имеет альфа-амилазную активность и (с) каждое положение соответствует положению аминокислотной последовательности исходной Термамил-подобной альфа-амилазы, имеющей аминокислотную последовательность SEQ ID NO:4.
В предпочтительном воплощении вышеуказанные варианты данного изобретения содержат мутацию в положении, соответствующем по меньшей мере одной из следующих мутаций в аминокислотной последовательности, показанной в SEQ ID NO:4:
V54N, A52S, A52S+V54N, T49L, T49+G107A, A52S+V54N+T49L+G107A, A52S+V54N+T49L, G107A, Q51R, Q51R+A52S, A52N; или
T49F+G107A, T49V+G107A, T49D+G107A, T49Y+G107A, T49S+G107A, T49N+G107A, T49I+G107A, T49L+A52S+G107A, T49L+A52T+G107A, T49L+A52F+G107A, T49L+A52L+G107A, T49L+A52I+G107A, T49L+A52V+G107A; или
T49V, T49I, T49D, T49N, T49S, T49Y, T49F, T49W, Т49М, Т49Е, T49Q, Т49К, T49R, А52Т, A52L, A52I, A52V, А52М, A52F, A52Y, A52W, V54M, G107V, G107I, G107L, G107C.
В предпочтительном воплощении вариант данного изобретения содержит по меньшей мере одну мутацию в положении, соответствующем следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4:
W13F,L,I,V,Y,A;
G48A,V,S,T,I,L;
*48aD или *48aY (т.е. инсерция D или Y);
T49X;
*49aX (т.е. инсерция любого возможного аминокислотного остатка)
S50X, в частности D,Y,L,T,V,I;
Q51R,K;
А52Х, в частности A52S,N,T,F,L,I,V;
D53E, Q,Y,I,N,S,T,V,L;
V54X, в частности V54I,N,W,Y,F,L;
G57S,A,V,L,I,F,Y,T;
G107X, в частности G107A,V,S,T,I,L,C;
G108X, в частности G108A,V,S,T,I,L;
A111V,I,L;
S168Y;
M197X, в частности Y,F,L,I,T,A,G.
В предпочтительном воплощении вариант данного изобретения содержит следующие мутации, соответствующие следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4: T49X+A52X+V54N/I/L/Y/F/W+G107A, и может дополнительно включать в себя G108A.
В предпочтительном воплощении вариант данного изобретения содержит по меньшей мере одну мутацию, соответствующую следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4:
T49L+G107A;
T49I+G107A;
T49L+G107A+V54I;
T49I+G107A+V54I;
A52S+V54N+T49L+G107A;
A52S+V54I+T49L+G107A;
A52S+T49L+G107A;
A52T+T49L+G107A;
A52S+V54N+T49I+G107A;
A52S+V54I+T49I+G107A;
A52S+T49I+G107A;
T49L+G108A;
T49I+G108A;
T49L+G108A+V54I;
T49I+G108A+V54I.
Все из вышеуказанных вариантов данного изобретения имеют измененные свойства (т.е. усиленные или ослабленные свойства), в частности, по меньшей мере одно из следующих свойств, относительно исходной альфа-амилазы; пониженную способность расщеплять субстрат вблизи точки ветвления, повышенную субстратную специфичность и/или повышенную удельную активность, измененный профиль рН-активности, измененный профиль рН-стабильности, измененную устойчивость к окислению, измененную Са2+-зависимость.
Стабильность
В контексте данного изобретения мутации (в том числе аминокислотные замены и/или делеции), важные в отношении достижения измененной стабильности, в частности улучшенной стабильности (т.е. более высокой или более низкой), особенно при низком рН (т.е. рН 4-6), включают в себя все мутации, перечисленные в разделе "Измененные свойства" выше, и варианты, указанные непосредственно ниже.
Следующие варианты: Q360A,K; N102A, N326A,L; N190G, N190K; Y262A,K,E (с использованием нумерации BAN, т.е. SEQ ID NO:6) испытывали на их рН-стабильность. Предпочтительной исходной альфа-амилазой может быть ВА2, описанная выше. рН-стабильность определяли, как описано в разделе "Материалы и способы".
Устойчивость к Ca2+
Измененная устойчивость к Ca2+ обозначает улучшенную стабильность фермента при низком содержании Ca2+, т.е. более высокую или более низкую стабильность. В контексте данного изобретения мутации (в том числе аминокислотные замены), важные в отношении достижения измененной устойчивости к Ca2+, в частности улучшенной устойчивости к Ca2+, т.е. более высокой или более низкой устойчивости, особенно при низком рН (т.е. рН 4-6), включают в себя любые из мутаций, перечисленных в разделе "Измененные свойства" выше.
Удельная активность
В дополнительном аспекте данного изобретения важные мутации в отношении получения вариантов, проявляющих измененную удельную активность, в частности увеличенную или уменьшенную активность, особенно при температурах 60-100° С, предпочтительно 70-95° С, особенно 80-90° С, включают в себя любые из мутаций, перечисленных в разделе "Измененные свойства" выше.
Удельную активность LE174 и LE429 определяли до 16000 NU/мг с использованием анализа Phadebas® , описанного в разделе "Материалы и способы".
Измененный характер расщепления
В процессе разжижения крахмала желательным является использование альфа-амилазы, которая способна разрушать молекулы крахмала на длинные, разветвленные олигосахариды, а не альфа-амилазы, которая образует более короткие разветвленные олигосахариды (подобно общепринятым Термамил-подобным альфа-амилазам). Короткие разветвленные олигосахариды (предшественники панозы) не гидролизуются удовлетворительно пуллуланазами, которые используют после обработки альфа-амилазами в процессе разжижения или одновременно с осахаривающей амилоглюкозидазой (глюкоамилазой) или перед добавлением осахаривающей амилоглюкозидазы (глюкоамилазы). Таким образом, в присутствии предшественников панозы смесь продуктов, присутствующая после глюкоамилазной обработки, содержит значительную долю короткого, разветвленного, так называемого предельного (остаточного) декстрина, а именно трисахарида панозы. Присутствие панозы значительно снижает выход осахаривания и, следовательно, является нежелательным.
Ранее сообщалось (патент США 5234823), что при осахаривании глюкоамилазой и пуллуланазой присутствие остаточной альфа-амилазной активности, возникающей из процесса разжижения, может приводить к более низким выходам глюкозы, если альфа-амилаза не инактивируется перед стадией осахаривания. Эту инактивацию можно обычно проводить доведением рН до менее 4,7 при 95° С перед снижением температуры до 60° С для осахаривания.
Причина этого отрицательного действия на выход глюкозы полностью не выяснена, но предполагается, что разжижающая альфа-амилаза (например, Термамил 120 L из В. licheniformis) генерирует "предельные (остаточные) декстрины" (которые являются плохими субстратами для пуллуланазы) гидролизом 1,4-альфа-глюкозидных связей вблизи от точек ветвления или по обе стороны от точек ветвления в амилопектине. Гидролиз этих предельных декстринов глюкоамилазой приводит к накоплению трисахарида панозы, которая лишь слабо гидролиэуется глюкоамилазой.
Получение термостабильной альфа-амилазы, которая не страдает этим недостатком, было бы значительным усовершенствованием, так как не требовалась бы отдельная стадия инактивации.
Таким образом, целью данного изобретения является получение мутантной альфа-амилазы, имеющей подходящим образом модифицированные характеристики деградации крахмала, но сохраняющей термостабильность исходной Термамил-подобной альфа-амилазы.
Таким образом, данное изобретение относится к варианту Термамил-подобной альфа-амилазы, который имеет улучшенную пониженную способность расщеплять субстрат вблизи точки ветвления и дополнительно имеет улучшенную субстратную специфичность и/или улучшенную удельную активность.
Особый интерес представляет вариант, который расщепляет субстрат-амилопектин, от восстанавливающего конца, на расстоянии более одного глюкозного звена от точки ветвления, предпочтительно на расстоянии более двух или трех глюкозных звеньев от точки ветвления, т.е. при большем расстоянии от точки ветвления, чем это расстояние при применении альфа-амилазы В. licheniformis дикого типа.
Здесь можно упомянуть, что согласно WO 96/23874 ожидается, что варианты, содержащие по меньшей мере одну из следующих мутаций, предотвращают расщепление вблизи от точки ветвления:
V54L,I,F,Y,W,R,K,H,E,Q;
D53L,I,F,Y,W;
Y56W;
Q333W;
G57, все возможные аминокислотные остатки;
А52, аминокислотные остатки, большие чем А, например A52W,Y, L,F,I.
Мутации, представляющие особенный интерес в связи с получением вариантов в соответствии с данным изобретением, имеющие улучшенную уменьшенную способность расщеплять субстрат вблизи точки ветвления и, кроме того, имеющие улучшенную субстратную специфичность и/или улучшенную удельную активность, включают в себя мутации в следующих положениях в альфа-амилазе В. licheniformis, SEQ ID NO:4:
Н156, А181, N190, А209, Q264 и 1201.
Следует подчеркнуть, что могут быть использованы не только специально указанные ниже Термамил-подобные альфа-амилазы.
Могут быть также использованы другие коммерческие Термамил-подобные альфа-амилазы. Неограничивающий перечень таких альфа-амилаз включает: альфа-амилазы, продуцируемые штаммом В. licheniformis, описанные в ЕР 0252666 (АТСС 27811), и альфа-амилазы, идентифицированные в WO 91/00353 и WO 94/18314. Другими коммерческими Термамил-подобными альфа-амилазами В. licheniformis являются Optitherm™ и Takatherm™ (доступные из Solvay), Maxamyl™ (доступная из Gist-Brocades/Genencor), Spezym АА™ и Spezyme Delta AA™ (доступные из Genencor), и Keistase™ (доступная из Daiwa).
Все Термамил-подобные альфа-амилазы могут быть выгодным образом использованы в качестве скелета макромолекулы для получения вариантов данного изобретения.
В предпочтительном варианте данного изобретения исходная Термамил-подобная альфа-амилаза является гибридной альфа-амилазой SEQ ID NO:4 и SEQ ID NO:6. Конкретно, исходная гибридная Термамил-подобная альфа-амилаза может быть гибридной альфа-амилазой, содержащей 445 С-концевых аминокислотных остатков альфа-амилазы В. licheniformis, показанной в SEQ ID NO: 4, и 37 N-концевых аминокислотных остатков зрелой альфа-амилазы, происходящей из В. amyloliquefaciens, показанной в SEQ ID NO:6, которая может выгодно иметь дополнительно следующие мутации: H156Y+A181T+N190F+A209V+Q264S (с использованием нумерации SEQ ID NO:4). Этот гибрид называют LE174. Гибрид LE174 может быть объединен с дополнительной мутацией I201F с образованием исходной гибридной Термамил-подобной альфа-амилазы, имеющей следующие мутации H156Y+A181T+N190F+A209V+ Q264S+I201F (с использованием SEQ ID NO:4 для нумерации). Этот гибридный вариант показан в SEQ ID NO:2 и использован в примерах, приведенных ниже, и назван LE429.
Также LE174 или LE429 (SEQ ID NO: 2) или альфа-амилаза В. licheniformis, показанная в SEQ ID NO:4, содержащая одну или несколько следующих мутаций, могут быть использованы в качестве скелета макромолекулы (с использованием SEQ ID NO:4 для нумерации мутаций):
Е119С;
S130C;
D124C;
R127C;
А52 все возможные аминокислотные остатки;
S85 вce возможные аминокислотные остатки;
N96 вce возможные аминокислотные остатки;
V129 вce возможные аминокислотные остатки;
А269 все возможные аминокислотные остатки;
А378 все возможные аминокислотные остатки;
S148 вce возможные аминокислотные остатки, в частности S148N;
Е211 все возможные аминокислотные остатки, в частности E211Q;
N188 вce возможные аминокислотные остатки, в частности N188S, N188P;
М197 все возможные аминокислотные остатки, в частности М197Т, М197А, M197G, M197I, M197L, M197Y, M197F, M197I;
М138 все возможные аминокислотные остатки, в частности W138Y;
D207 вce возможные аминокислотные остатки, в частности D207Y;
Н133 все возможные аминокислотные остатки, в частности H133Y;
Н205 все возможные аминокислотные остатки, в частности Н205Н, Н205С, H205R;
S187 все возможные аминокислотные остатки, в частности S187D;
А210 все возможные аминокислотные остатки, в частности A210S, А210Т;
Н405 все возможные аминокислотные остатки, в частности H405D;
К176 все возможные аминокислотные остатки, в частности K176R;
F279 все возможные аминокислотные остатки, в частности F279Y;
Q298 вce возможные аминокислотные остатки, в частности Q298H;
G299 все возможные аминокислотные остатки, в частности G299R;
L308 все возможные аминокислотные остатки, в частности L308F;
Т412 все возможные аминокислотные остатки, в частности Т412А;
Кроме того, альфа-амилаза В. licheniformis, показанная в SEQ ID NO:4, содержащая по меньшей мере одну из следующих мутаций, может быть использована в качестве скелета макромолекулы:
М15все возможные аминокислотные остатки;
А33все возможные аминокислотные остатки.
При использовании LE429 (показанной в SEQ ID NO: 2) в качестве скелета макромолекулы. (т.е. в качестве исходной Термамил-подобной альфа-амилазы) объединением LE174 с мутацией I201F (нумерация SEQ ID NO:4) могут быть произведены мутации/изменения, в частности замены, делеции и инсерции в соответствии с данным изобретением в одном или нескольких положениях для улучшения уменьшенной способности расщеплять субстрат вблизи точки ветвления и для улучшения субстратной специфичности и/или улучшенной удельной активности: W13, G48, Т49, S50, Q51, А52, D53, V54, G57, G107, G108, A111, S168, М197 (с использованием нумерации SEQ ID NO:4), где (а) это изменение (эти изменения) представляют собой:
(i) инсерцию аминокислоты справа от аминокислоты, которая занимает это положение;
(ii) делецию аминокислоты, которая занимает это положение, или
(iii) замену аминокислоты, которая занимает это положение, отличающейся аминокислотой,
(b) этот вариант имеет альфа-амилазную активность и (с) каждое положение соответствует положению аминокислотной последовательности исходной Термамил-подобной альфа-амилазы, имеющей аминокислотную последовательность SEQ ID NO:4.
В предпочтительном воплощении вариант данного изобретения содержит по меньшей мере одну мутацию в положении, соответствующем следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4:
V54N, A52S, A52S+V54N, T49L, T49+G107A, A52S+V54N+T49L+G107A, A52S+V54N+T49L, G107A, Q51R, Q51R+A52S, A52N; или
T49F+G107A, T49V+G107A, T49D+G107A, T49Y+G107A, T49S+G107A, T49N+G107A, T49I+G107A, T49L+A52S+G107A, T49L+A52T+G107A, T49L+A52F+G107A, T49L+A52L+G107A, T49L+A52I+G107A, T49L+A52V+G107A; или
T49V, T49I, T49D, T49N, T49S, T49Y, T49F, T49W, Т49М, Т49Е, T49Q, Т49К, T49R, А52Т, A52L, A52I, A52V, А52М, A52F, A52Y, A52W, V54M, G107V, G107I, G107L, G107C.
В предпочтительном воплощении вариант данного изобретения содержит по меньшей мере одну мутацию в положении, соответствующем следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4:
W13F,L,I,V,Y,A;
G48A,V,S,T,I,L;
*48aD или *48aY (т.е. инсерция D или Y);
T49X;
*49aX (т.е. инсерция любого возможного аминокислотного остатка)
S50X, в частности D,Y,L,T,V,I;
Q51R,K;
А52Х, в частности A52S,N,T,F,L,I,V;
D53E,Q,I,N,S,T,V,L;
V54X, в частности V54I,N,W,Y,F,L;
G57S,A,V,L,I,F,Y,T;
G107X, в частности G107A,V,S,T,I,L,C;
G108X, в частности G108A,V,S,T,I,L;
A111V,I,L;
S168Y;
M197X, в частности Y,F,L,I,T,A,G.
В предпочтительном воплощении вариант данного изобретения содержит по меньшей мере одну мутацию в положении, соответствующем следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4:
T49X+A52X+V54N/I/L/Y/F/W+G107A, и может дополнительно включать в себя G108A.
В предпочтительном воплощении вариант данного изобретения содержит по меньшей мере одну мутацию в положении, соответствующем следующим мутациям в аминокислотной последовательности, показанной в SEQ ID NO:4:
T49L+G107A;
T49I+G107A;
T49L+G107A+V54I;
T49I+G107A+V54I;
A52S+V54N+T49L+G107A;
A52S+V54I+T49L+G107A;
A52S+T49L+G107A;
A52T+T49L+G107A;
A52S+V54N+T49I+G107A;
A52S+V54I+T49I+G107A;
A52S+T49I+G107A;
T49L+G108A;
T49I+G108A;
T49L+G108A+V54I;
T49I+G108A+V54I.
Общие мутации в вариантах данного изобретения
Может быть предпочтительным, чтобы вариант данного изобретения содержал одну или несколько модификаций, кроме описанных выше. Так, может быть выгодным, чтобы один или несколько остатков пролина, присутствующих в части варианта альфа-амилазы, которая является модифицированной, был/были заменены другим остатком (не остатком пролина), который может быть любым из возможных природных остатков, отличных от пролина, и который предпочтительно является аланином, глицином, серином, треонином, валином или лейцином.
Аналогично, может быть предпочтительным, чтобы один или несколько остатков цистеина среди аминокислотных остатков, которыми модифицирована исходная альфа-амилаза, был/были заменены другим остатком (не остатком цистеина), таким как серин, аланин, треонин, глицин, валин или лейцин.
Кроме того, вариант данного изобретения может быть - либо в виде единственной модификации, либо в комбинации с любой из вышеописанных модификаций - модифицирован таким образом, что один или несколько Asp и/или Glu, присутствующих в аминокислотном фрагменте, соответствующем аминокислотному фрагменту 185-209 SEQ ID NO:4, является замененным Asn и/или Gln соответственно. Также представляет интерес замена, в Термамил-подобной альфа-амилазе, одного или нескольких остатков Lys, присутствующих в аминокислотном фрагменте, соответствующем аминокислотному фрагменту 185-209 SEQ ID NO:4, посредством Arg.
Должно быть понятно, что данное изобретение включает в себя варианты, содержащие две или несколько вышеописанных модификаций.
Кроме того, может быть выгодным введение точковых мутаций в любом из описанных здесь вариантов.
Способы получения вариантов альфа-амилазы
Несколько способов введения мутаций в гены известны в данной области. После краткого обсуждения клонирования кодирующих альфа-амилазу последовательностей ДНК будут обсуждены способы генерирования мутаций в специфических сайтах в кодирующей альфа-амилазу последовательности.
Клонирование последовательности ДНК, кодирующей альфа-амилазу
Последовательность ДНК, кодирующая исходную альфа-амилазу, может быть выделена из любой клетки или микроорганизма, продуцирующих рассматриваемую альфа-амилазу, с использованием различных способов, хорошо известных в данной области. Во-первых, должна быть сконструирована библиотека геномной ДНК и/или библиотека кДНК с использованием хромосомной ДНК или матричной РНК из организма, который продуцирует исследуемую альфа-амилазу. Затем, если аминокислотная последовательность этой альфа-амилазы известна, могут быть синтезированы гомологичные меченые олигонуклеотидные зонды и использованы для идентификации кодирующих альфа-амилазу клонов из геномной библиотеки, полученной из рассматриваемого организма. Альтернативно, меченый олигонуклеотидный зонд, содержащий последовательность, гомологичную известному гену альфа-амилазы, мог бы быть использован в качестве зонда для идентификации кодирующих альфа-амилазу клонов с использованием условий гибридизации и промывки низкой жесткости.
Еще один способ для идентификации кодирующих альфа-амилазу клонов включает в себя встраивание фрагментов геномной ДНК в экспрессирующий вектор, такой как плазмида, трансформацию альфа-амилаза-отрицательных бактерий полученной библиотекой геномной ДНК и затем посев трансформированных бактерий на агар, содержащий субстрат для альфа-амилазы, что позволяет в результате идентифицировать клоны, экспрессирующие альфа-амилазу.
Альтернативно, последовательность ДНК, кодирующая этот фермент, может быть получена синтетически при помощи установленных стандартных способов, например фосфороамидитного способа, описанного S.L.Beaucage and M.H. Caruthers (1981), или способа, описанного Matthes et al., (1984). В фосфороамидитном способе олигонуклеотиды синтезируют, например, в автоматическом ДНК-синтезаторе, очищают, лигируют и клонируют в подходящих векторах.
Наконец, последовательность ДНК может быть смешанного геномного и синтетического происхождения, смешанного синтетического и кДНК-происхождения или смешанного геномного и кДНК-происхождения, полученная лигированием фрагментов синтетического, геномного или кДНК-происхождения (если требуется, фрагментов, соответствующих различным частям полной последовательности ДНК), в соответствии со стандартными способами. Последовательность ДНК может быть также получена полимеразной цепной реакцией (ПЦР) с использованием специфических праймеров, например, как описано в патенте США 4683202 или R.K. Saiki et al. (1988).
Сайт-направленный мутагенез
После выделения кодирующей альфа-амилазу последовательности ДНК и идентификации желательных сайтов для мутации мутации могут быть введены с использованием синтетических олигонуклеотидов. Эти олигонуклеотиды содержат нуклеотидные последовательности, фланкирующие желательные сайты мутации; мутантные нуклеотиды встраивают во время синтеза олигонуклеотидов. В конкретном способе одноцепочечный разрыв ДНК, соединяющей кодирующую альфа-амилазу последовательность, создают в векторе, несущем ген альфа-амилазы. Затем синтетический нуклеотид, несущий желаемую мутацию, отжигают с гомологичной частью этой одноцепочечной ДНК. Затем оставшийся пропуск заполняют с использованием ДНК-полимеразы I (фрагмента Кленова) и эту конструкцию лигируют при помощи лигазы Т4. Характерный пример этого способа описан в Morinaga et al., (1984). Патент США 4760025 описывает введение олигонуклеотидов, кодирующих множественные мутации, выполнением минорных изменений этой кассеты. Однако еще большее разнообразие мутаций может быть введено одномоментно по способу Morinaga, так как может быть введено множество олигонуклеотидов различной длины.
Другой способ введения мутаций в кодирующую альфа-амилазу ДНК описан в Nelson and Long (1989). Он включает в себя 3-стадийное генерирование ПЦР-фрагмента, содержащего желательную мутацию, введенную с использованием химически синтезированной цепи ДНК в качестве одного из праймеров в реакциях ПЦР. Из ПЦР-генерированного продукта фрагмент ДНК, несущий мутацию, может быть выделен расщеплением рестрикционными эндонуклеазами (рестриктазами) и повторно встроен в экспрессионную кассету.
Случайный мутагенез
Случайный мутагенез удобно проводить либо в виде локализованного, либо в виде область-специфического случайного мутагенеза по меньшей мере в трех частях гена, транслируемого в аминокислотную последовательность, или в целом гене.
Случайный мутагенез последовательности ДНК, кодирующей исходную альфа-амилазу, может быть удобным образом выполнен с использованием любого известного в данной области способа.
В связи с вышесказанным, следующий аспект данного изобретения относится к способу генерирования варианта исходной альфа-амилазы, в котором, например, этот вариант проявляет пониженную способность расщепления олигосахаридного субстрата вблизи точки ветвления и дополнительно проявляет повышенную субстратную специфичность и/или повышенную удельную активность относительно исходного фермента, причем этот способ предусматривает:
(a) подвергание последовательности ДНК, кодирующей исходную альфа-амилазу, случайному мутагенезу,
(b) экспрессию мутированной последовательности ДНК, полученной в стадии (а), в клетке-хозяине и
(c) скрининг на клетки-хозяева, экспрессирующие вариант альфа-амилазы, который имеет измененное свойство (т.е. термостабильность) относительно исходной альфа-амилазы.
Стадию (а) указанного выше способа данного изобретения предпочтительно выполняют с использованием легированных (doped) праймеров. Например, случайный мутагенез может быть выполнен с использованием подходящего физического или химического мутагенного агента, с использованием подходящего олигонуклеотида, или подверганием последовательности ДНК генерируемому ПЦР мутагенезу. Кроме того, случайный мутагенез может быть выполнен с использованием любой комбинации этих мутагенных агентов. Мутагенный агент может, например, быть агентом, который индуцирует транзиции, трансверсии, инверсии, перетасовку, делеции и/или инсерции (вставки).
Примеры физического или химического мутагенного агента, пригодного для этой цели, включают в себя облучение ультрафиолетом (УФ), гидроксиламин, N-метил-N'-нитро-N-нитрозогуанидин (MNNG), О-метилгидроксиламин, азотистую кислоту, этилметансульфонат (EMS), бисульфит натрия, муравьиную кислоту и аналоги нуклеотидов. При использовании таких агентов мутагенез обычно проводят инкубированием последовательности ДНК, кодирующей исходный фермент, который должен быть мутагенизирован, в присутствии предпочтительного мутагенного агента при подходящих условиях для осуществления мутагенеза и отбора на мутированную ДНК, имеющую желательные свойства. При проведении мутагенеза с использованием олигонуклеотида этот олигонуклеотид может быть легирован (doped) или снабжен (spiked) тремя вставленными, не содержащимися в исходном ферменте нуклеотидами во время синтеза этих олигонуклеотидов в положениях, которые должны быть изменены. Это "легирование" или "вставка" может быть выполнено таким образом, что кодоны для нежелательных аминокислот избегаются. "Легированный" или "снабженный вставленными нуклеотидами" олигонуклеотид может быть включен в ДНК, кодирующую альфа-амияазный фермент, любым опубликованным способом, с использованием, например, ПЦР, ЛЦР или любой ДНК-полимеразы и лигазы, которые считаются подходящими. Предпочтительно, "легирование" проводят с использованием "постоянного случайного легирования", в котором процент дикого типа и мутации в каждом положении является предварительно определенным. Кроме того, "легирование" может проводиться в направлении предпочтения для введения определенных нуклеотидов и, тем самым, предпочтения для введения одного или нескольких специфических аминокислотных остатков. "Легирование" может быть выполнено, например, таким образом, чтобы сделать возможным введение 90% дикого типа и 10% мутаций в каждом положении. Дополнительное соображение в выборе схемы "легирования" основано на генетических, а также белковых структурных сдерживаниях. Схема "легирования" может быть построена с использованием программы DOPE, которая, inter alia, гарантирует, что введение стоп-кодонов избегается. При использовании ПЦР-генерируемого мутагенеза химически обработанный или не обработанный ген, кодирующий исходную альфа-амилазу, подвергают ПЦР в условиях, которые увеличивают ошибочное включение нуклеотидов (Deshler 1992; Leung et al., Technique, Vol. 1, 1989, pp. 11-15). Мутирующий штамм Е. coli (Fowler et al., Molec. Gen. Genet., 133, 1974, pp. 179-191), S. cerevisiae или любой другой микробный организм может быть использован для случайного мутагенеза ДНК, кодирующей альфа-амилазу, например, трансформацией плазмиды, содержащей исходную гликозилазу, в мутирующем штамме, выращиванием мутирующего штамма с этой плазмидой и выделением мутированной плазмиды из мутирующего штамма. Затем мутированной плазмидой может быть трансформирован экспрессирующий организм. Подлежащая мутагенезу последовательность ДНК может для удобства присутствовать в библиотеке геномной ДНК или кДНК, полученной из организма, экспрессирующего исходную альфа-амилазу. Альтернативно, эта последовательность ДНК может присутствовать на подходящем векторе, таком как плазмида или бактериофаг, который в таком виде может быть инкубирован с мутагенным агентом или иным образом экспонирован мутагенному агенту. ДНК, которая должна быть мутагенизирована, может также присутствовать в клетке-хозяине либо будучи интегрированной в геном этой клетки, либо присутствуя на векторе, уловленном в этой клетке. Наконец, ДНК, которая должна быть мутагенизирована, может быть в выделенной форме. Должно быть понятно, что последовательность ДНК, подлежащая случайному мутагенезу, является предпочтительно кДНК-последовательностью или последовательностью геномной ДНК. В некоторых случаях может быть удобной амплификация мутированной последовательности ДНК перед выполнением стадии экспрессии b) или стадии скрининга с). Такая амплификация может проводиться в соответствии со способами, известными в данной области, причем предпочтительным в настоящее время способом является ПЦР-генерируемая амплификация с использованием олигонуклеотидных праймеров, полученных на основе последовательности ДНК или аминокислотной последовательности исходного фермента. После инкубации с мутагенным агентом или экспонирования мутагенному агенту мутированную ДНК экспрессируют культивированием подходящей клетки-хозяина, несущей эту последовательность ДНК, в условиях, обеспечивающих протекание экспрессии. Клеткой-хозяином, используемой для этой цели, может быть любая клетка, которая была трансформирована мутированной последовательностью ДНК, необязательно присутствующей на векторе, или клетка, которая несла последовательность ДНК, кодирующую исходный фермент, во время мутагенной обработки. Примерами подходящих клеток-хозяев являются следующие: грамположительные бактерии, такие как Bacillus subtilis. Bacillus licheniformis, Bacillus lentus, Bacillus brevis, Bacillus stearothermophilus, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus coagulans, Bacillus circulans, Bacillus lautus, Bacillus megaterium, Bacillus thuringiensis, Streptomyces lividans или Streptomyces murinus; и грамотрицательные бактерии, такие как Е. coli. Мутированная последовательность ДНК может дополнительно содержать последовательность ДНК, кодирующую функции, делающие возможной экспрессию этой мутированной последовательности ДНК.
Локализованный случайный мутагенез
Случайный (спонтанный) мутагенез может быть выгодно локализован в части рассматриваемой исходной альфа-амилазы. Это может быть, например, выгодным, когда определенные области этого фермента были идентифицированы как области, особенно важные для конкретного свойства этого фермента, и ожидается, что при модификации они приводят к варианту, имеющему улучшенные свойства. Такие области могут быть обычно идентифицированы, когда третичная структура исходного фермента была выяснена и соотнесена с функцией этого фермента.
Локализованный или область-специфический спонтанный мутагенез удобно проводить с использованием способов ПЦР-генерируемого мутагенеза, как описано выше, или любым другим подходящим способом, известным в данной области. Альтернативно, последовательность ДНК, кодирующая часть последовательности ДНК, которая должна быть модифицирована, может быть выделена, например, встраиванием в подходящий вектор, и указанная часть может быть подвергнута мутагенезу при помощи любого из обсуждаемых выше способов мутагенеза.
Альтернативные способы обеспечения вариантов альфа-амилазы
Альтернативные способы для обеспечения вариантов данного изобретения включают в себя способ перетасовки генов, известный в данной области, в том числе способы, описанные в WO 95/22625 (из Affymax Technologies N.V.) и WO 96/00343 (из Novo Nordisk A/S).
Экспрессия вариантов альфа-амилазы
Согласно данному изобретению последовательность ДНК, кодирующая вариант, полученная описанными выше способами или любыми альтернативными способами, известными в данной области, может быть экспрессирована, в форме фермента, с использованием экспрессирующего вектора, который обычно включает в себя регуляторные последовательности, кодирующие промотор, оператор, сайт связывания рибосом, сигнал инициации трансляции и, необязательно, репрессорный ген или различные гены-активаторы.
Рекомбинантный экспрессирующий вектор, несущий последовательность ДНК, кодирующую вариант альфа-амилазы данного изобретения, может быть любым вектором, который может быть легко подвергнут процедурам рекомбинантных ДНК, и выбор вектора будет часто зависеть от клетки-хозяина, в которую он должен вводиться. Таким образом, этот вектор может быть автономно реплицирующимся вектором, т.е. вектором, который существует в виде внехромосомной сущности, репликация которой является независимой от хромосомной репликации, например плазмидой, бактериофагом или внехромосомным элементом, минихромосомой или искусственной хромосомой. Альтернативно, этот вектор может быть вектором, который при введении в клетку-хозяина интегрируется в геном клетки-хозяина и реплицируется вместе с хромосомой (хромосомами), в которую (в которые) он был интегрирован.
В этом векторе последовательность ДНК должна быть функционально соединена с подходящей промоторной последовательностью. Промотор может быть любой последовательностью ДНК, которая обнаруживает транскрипционную активность в выбранной клетке-хозяине и может быть получена из генов, кодирующих белки, либо гомологичные, либо гетерологичные относительно клетки-хозяина. Примерами подходящих промоторов для направления транскрипции последовательности ДНК, кодирующей вариант альфа-амилазы данного изобретения, в частности в бактериальном хозяине, являются промотор lac-оперона Е. coli, промоторы гена агаразы dagA Streptomyces coelicolor, промоторы гена альфа-амилазы (amyL) В. licheniformis, промоторы гена мальтогенной амилазы (amyМ) В. stearothermophilus, промоторы гена альфа-амилазы (amyQ) В. amyloliquefaciens, промоторы генов xylА и xylB Bacillus subtilis. Для транскрипции в грибковом хозяине примерами применимых промоторов являются промоторы, происходящие из гена, кодирующего ТАКА-амилазу A. oryzae, протеиназу аспарагиновой кислоты Rhizomucor miehei, нейтральную альфа-амилазу A. niger, кислотоустойчивую альфа-амилазу A. niger, липазу Rhizomucor miehei, щелочную протеазу A. oryzae, триозо-фосфатизомеразу A. oryzae или ацетамидазу A. nidulans.
Экспрессирующий вектор данного изобретения может также содержать подходящий терминатор транскрипции и, в эукариотах, последовательности полиаденилирования, функционально связанные с последовательностью ДНК, кодирующей вариант альфа-амилазы данного изобретения. Последовательности терминации и полиаденилирования могут быть подходящим образом получены из тех же самых источников, что и промотор.
Вектор может дополнительно содержать последовательность ДНК, позволяющую вектору реплицироваться в рассматриваемой клетке-хозяине. Примерами такой последовательности являются точки начала репликации плазмид pUC19, pACYC177, pUB110, рЕ194, pAMB1 и pIJ702.
Вектор может также содержать селектируемый маркер, например ген, продукт которого восполняет дефект в клетке-хозяине, такой как гены dal из Bacillus subtilis или В. licheniformis, или ген, который придает устойчивость к антибиотику, например устойчивость к ампициллину, канамицину, хлорамфениколу или тетрациклину. Кроме того, вектор может содержать селектируемые маркеры Aspergillus, такие как amdS, argB, niaD и sC, маркер, сообщающий устойчивость к гигромицину, или отбор может быть выполнен с использованием котрансформации, например, как описано в WO 91/17243.
Хотя внутриклеточная экспрессия может быть выгодной в некоторых отношениях, например при использовании некоторых бактерий в качестве клеток-хозяев, обычно предпочтительно, чтобы эта экспрессия была внеклеточной. Обычно альфа-амилазы Bacillus, указанные здесь, содержат пре-район, делающий возможной секрецию экспрессируемой протеазы в культуральную среду. Если желательно, этот пре-район может быть заменен отличающимся пре-районом или сигнальной последовательностью, что удобно выполняется заменой последовательностей ДНК, кодирующих соответствующие пре-районы.
Процедуры, используемые для лигирования конструкции ДНК данного изобретения, кодирующей вариант альфа-амилазы, промотор, терминатор и другие элементы соответственно, и для инсертирования их в подходящие векторы, содержащие информацию, необходимую для репликации, хорошо известны специалистам в данной области (см., например, Sambrook et al., (Molecular Cloning. A Laboratory Manual, 2nd Ed., Cold Spring Harbor, 1989).
Клетку данного изобретения, содержащую либо конструкцию ДНК, либо экспрессирующий вектор данного изобретения, определенные здесь, выгодно используют в качестве клетки-хозяина в рекомбинантном получении варианта альфа-амилазы данного изобретения. Эта клетка может быть легко трансформирована конструкцией ДНК данного изобретения, кодирующей этот вариант, путем интеграции этой конструкции ДНК (в виде одной или более копий) в хромосому хозяина. Обычно считается, что эта интеграция является выгодной, так как эта последовательность ДНК с большей вероятностью стабильно сохраняется в клетке. Интеграция конструкций ДНК в хромосому хозяина может выполняться в соответствии с общепринятыми способами, например гомологичной или гетерологичной рекомбинацией. Альтернативно, клетка может быть трансформирована экспрессирующим вектором, как описано выше, в связи с различными типами клеток-хозяев.
Клетка данного изобретения может быть клеткой высшего организма, такого как млекопитающее или насекомое, но предпочтительно она является микробной клеткой, например бактериальной или грибковой (в том числе дрожжевой) клеткой.
Примерами подходящих бактерий являются грамположительные бактерии, такие как Bacillus subtilis, Bacillus licheniformis, Bacillus lentus, Bacillus brevis, Bacillus stearothermophilus, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus coagulans, Bacillus circulans, Bacillus lautus, Bacillus megaterium, Bacillus thuringiensis, Streptomyces lividans или Streptomyces murinus, и грамотрицательные бактерии, такие как Е. coli. Трансформация бактерий может выполняться, например, трансформацией протопластов или с использованием компетентных клеток известным per se способом.
Дрожжевой организм может быть предпочтительно выбран из видов Saccharomyces или Schizosaccharomyces, например Saccharomyces cerevisiae. Нитевидный гриб может предпочтительно принадлежать к видам Aspergillus, например Aspergillus oryzae или Aspergillus niger. Грибковые клетки могут быть трансформированы способом, включающим в себя образование протопластов и трансформацию этих протопластов с последующей регенерацией клеточной стенки известным per se образом. Подходящая процедура для трансформации клеток-хозяев Aspergillus описана в ЕР 238023.
Еще в одном аспекте данное изобретение относится к способу получения варианта альфа-амилазы данного изобретения, предусматривающему культивирование клетки-хозяина, описанной выше, в условиях, способствующих образованию этого варианта, и извлечение этого варианта из клеток и/или культуральной среды.
Среда, используемая для культивирования клеток, может быть любой общепринятой средой, пригодной для выращивания рассматриваемой клетки-хозяина и получения экспрессии варианта альфа-амилазы данного изобретения. Подходящие среды доступны от коммерческих поставщиков или могут быть приготовлены в соответствии с опубликованными руководствами (например, как описано в каталогах Американской Коллекции Типовых Культур).
Вариант альфа-амилазы, секретируемый из клеток-хозяев, может быть извлечен из культуральной среды хорошо известными процедурами, в том числе отделением клеток от среды центрифугированием или фильтрованием и осаждением белковых компонентов среды высаливанием, например, при помощи сульфата аммония, с последующим применением хроматографических процедур, таких как ионообменная хроматография, аффинная хроматография или т.п.
Промышленная применимость
Варианты альфа-амилазы данного изобретения обладают ценными свойствами, позволяющими разнообразные промышленные применения. В частности, варианты фермента данного изобретения применимы в качестве компонента в моющих композициях для стирки, мытья посуды и чистки твердых поверхностей. Многочисленные варианты применимы, в частности, в производстве подсластителей и этанола, например топливного, питьевого или промышленного этанола, из крахмала, и для расшлихтовки текстильных изделий. Условия для общепринятых процессов превращения крахмала, в том числе процессов разжижения крахмала и/или осахаривания крахмала, описаны, например, в патенте США 3912590 и в патентных публикациях ЕР с номерами 252730 и 63909.
Получение подсластителей из крахмала:
Традиционный способ превращения крахмала во фруктозные сиропы обычно состоит из трех последовательных ферментативных процессов, а именно процесса разжижения с последующими процессом осахаривания и процессом изомеризации. Во время процесса разжижения крахмал деградируется до декстринов альфа-амилазой (например, Термамилом™ ) при величинах рН между 5,5 и 6,2 и при температурах 95-160° С в течение периода приблизительно 2 часов. Для обеспечения оптимальной стабильности фермента при этих условиях добавляют 1 мМ кальций (40 м.д. свободных ионов кальция).
После процесса разжижения декстрины превращают в декстрозу добавлением глюкоамилазы (например, AMG™ ) и деветвящего фермента, такого как изоамилаза или пуллуланаза (например, Промозим™ ). Перед этой стадией рН снижают до величины менее 4,5, поддерживая высокую температуру (выше 95° С), и разжижающая альфа-амилазная активность денатурируется. Температуру понижают до 60° С и добавляют глюкоамилазу и деветвящий фермент. Процесс осахаривания протекает в течение 24-72 час.
После процесса осахаривания рН увеличивают до величины в диапазоне 6-8, предпочтительно 7,5, и кальций удаляют посредством ионообмена. Затем декстрозный сироп превращают во фруктозный сироп с использованием, например, иммобилизованной глюкозоизомеразы (такой как Sweetzyme™ ).
Можно представить себе по меньшей мере одно ферментативное усовершенствование этого процесса: уменьшение зависимости от кальция разжижающей альфа-амилазы. Добавление свободного кальция требуется для гарантии адекватной высокой стабильности альфа-амилазы, но свободный кальций сильно ингибирует активность глюкоизомеразы и требует удаления с использованием операции дорогостоящего узла до такой степени, которая снижает свободный кальций до менее 3-5 м.д. Экономия могла бы быть получена, если можно было бы избежать такой операции и процесс разжижения мог бы выполняться без добавления свободных ионов кальция.
Для достижения этого требуется менее кальций-зависимая Термамил-подобная альфа-амилаза, которая является стабильной и высокоактивной при низких концентрациях свободного кальция (<40 м.д.). Такая Термамил-подобная альфа-амилаза должна иметь оптимум рН при рН в диапазоне 4,5-6,5, предпочтительно в диапазоне 4,5-5,5.
Данное изобретение относится также к композиции, содержащей смесь одного или нескольких вариантов данного изобретения, происходящих из (в качестве исходной Термамил-подобной альфа-амилазы) альфа-амилазы В. stearothermophilus, имеющей последовательность, показанную в SEQ ID NO:8, и Термамил-подобной альфа-амилазы, происходящей из альфа-амилазы В. licheniformis, имеющей последовательность, показанную в SEQ ID NO:4.
Далее, данное изобретение относится также к композиции, содержащей смесь одного или нескольких вариантов данного изобретения, происходящих из (в качестве исходной Термамил-подобной альфа-амилазы) альфа-амилазы В. stearothermophilus, имеющей последовательность, показанную в SEQ ID NO:8, и гибридной альфа-амилазы, включающей в себя часть альфа-амилазы В. amyloliquefaciens, показанной в SEQ ID NO:6, и часть альфа-амилазы В. licheniformis, показанной в SEQ ID NO:4. Последняя указанная гибридная Термамил-подобная альфа-амилаза содержит 445 С-концевых аминокислотных остатков альфа-амилазы В. licheniformis, показанной В SEQ ID NO: 4, и 37 N-концевых аминокислотных остатков альфа-амилазы, происходящей из В. amyloliquefaciens, показанной в SEQ ID NO:6. Указанная последняя гибридная альфа-амилаза может предпочтительно содержать следующие мутации: H156Y+A181T+N190F+A209V+Q264S (с использованием нумерации SEQ ID NO:4). Предпочтительно, указанная последняя гибридная альфа-амилаза может предпочтительно содержать следующие мутации: H156Y+A181T+ N190F+A209V+Q264S+I201F (с использованием нумерации SEQ ID NO:4). В примерах ниже указанную последнюю гибридную Термамил-подобную альфа-амилазу, называемую LE429 (показанную в SEQ ID NO:2), используют для получения вариантов данного изобретения, которые могут быть использованы в композициях данного изобретения.
Вариант альфа-амилазы данного изобретения или композиция данного изобретения могут быть в аспекте данного изобретения использованы для разжижения крахмала, в детергентных композициях, таких как композиции для стирки, мытья посуды и чистки твердых поверхностей, для производства этанола, такого как топливный, питьевой и промышленный этанол, расшлихтовки текстильных изделий, ткани и одежды.
МАТЕРИАЛЫ И СПОСОБЫ
Ферменты:
LE174: гибридный вариант альфа-амилазы:
LE174 представляет собой гибридную Термамил-подобную альфа-амилазу, идентичную последовательности Термамила, т.е. альфа-амилазы В. licheniformis, показанной в SEQ ID NO:4, за исключением того, что 35 N-концевых аминокислотных остатков (зрелого белка) были заменены 33 N-концевыми остатками BAN (зрелого белка), т.е. альфа-амилазы В. amyloliquefaciens, показанной в SEQ ID NO:6, которая дополнительно имеет следующие мутации: H156Y+A181T+N190F+A209V+Q264S (SEQ ID NO:4). LE429: гибридный вариант альфа-амилазы:
LE429 представляет собой гибридную Термамил-подобную альфа-амилазу, идентичную последовательности Термамила, т.е. альфа-амилазы В. licheniformis, показанной в SEQ ID NO:4, за исключением того, что 35 N-концевых аминокислотных остатков (зрелого белка) были заменены 33 N-концевыми остатками BAN (зрелого белка), т.е. альфа-амилазы В. amyloliquefaciens, показанной в SEQ ID NO:6, которая дополнительно имеет следующие мутации: H156Y+A181T+N190F+A209V+Q264S+I201F (SEQ ID NO:4). LE429 показана как SEQ ID NO:2 и была сконструирована с использованием SOE-PCR (Higuchi et al. 1988, Nucleic Acids Research 16:7351).
Декстрозим™ Е: сбалансированная смесь глюкоамилазы (AMG) и пуллуланазы, полученная из выбранных штаммов Aspergillus niger и Bacillus deramificans (доступных из Novo Nordisk A/S).
Ферментация и очистка вариантов альфа-амилазы
Штамм В. subtilis, несущий релевантную экспрессирующую плазмиду, высевают штрихом на чашку с LB-агаром с 10 мкг/мл канамицина из исходного раствора, хранящегося при -80° С, и выращивают в течение ночи при 37° С.
Колонии переносят в 100 мл среды ВРХ, дополненной 10 мкг/мл канамицина, в колбе для встряхивания на 500 мл. Состав среды ВРХ, г/л:
Картофельный крахмал 100
Ячменная мука 50
BAN 5000 SKB 0,1
Казеинат натрия 10
Соевая мука 20
NaH2PO4· 12 Н2O 9
Плуроник™ 0,1
Культуру встряхивают при 37° С при 270 об/мин в течение 5 дней.
Клетки и клеточный дебрис удаляют из ферментационного бульона центрифугированием при 4500 об/мин в течение 20-25 мин. После этого супернатант фильтруют с получением полностью прозрачного раствора. Фильтрат концентрируют и промывают на УФ-фильтре (мембране с отсечением 10000) и буфер заменяют на 20 мМ ацетат рН 5,5. УФ-фильтрат наносят на S-сефарозу F.F. и элюцию проводят ступенчатой элюцией 0,2 М NaCl в том же самом буфере. Элюат диализуют против 10 мМ Трис, рН 9,0 и наносят на Q-сефарозу F.F. и элюируют линейным градиентом 0-0,3 М NaCl на протяжении протекания 6 колоночных объемов. Фракции, которые содержат активность (измеренную при помощи анализа Phadebas), объединяют, рН доводят до рН 7,5 и остаточную окраску удаляют обработкой 0,5% мас./об. активированным углем в течение 5 минут.
Определение активности - (KNU)
Одной альфа-амилазной килоединицей (1 KNU) является количество фермента, которое разрушает 5,26 г крахмала (Merck, Amylum Solubile, Erg. В 6, Batch 9947275) в час в стандартном способе Novo Nordisk для определения альфа-амилазы, основанном на следующих условиях:
Субстрат растворимый крахмал
Содержание кальция в растворителе, М 0,0043
Время реакции, мин 7-20
Температура, ° C 37
рН 5,6
Подробное описание аналитического способа Novo Nordisk (AF 9) доступно по запросу.
Анализ на альфа-амилазную активность
Альфа-амилазную активность определяют по способу с использованием таблеток Phadebas® в качестве субстрата. Таблетки Phadebas (Амилазный тест Phadebas® , поставляемый Pharmacia Diagnostic) содержат сшитый нерастворимый полимер крахмала синего цвета, который был смешан с бычьим сывороточным альбумином и буферным веществом и таблетирован.
Для каждого отдельного измерения одну таблетку суспендируют в пробирке, содержащей 5 мл 50 мМ буфера Бриттона-Робинсона (50 мМ уксусная кислота, 50 мМ фосфорная кислота, 50 мМ борная кислота, 0,1 мМ CaCl2, pH, доведенный до нужной величины при помощи NaOH). Тест проводят в водяной бане при представляющей интерес температуре. Тестируемую альфа-амилазу разбавляют в × мл 50 мМ буфера Бриттона-Робинсона. 1 мл этого раствора альфа-амилазы добавляют в 5 мл 50 мМ буфера Бриттона-Робинсона. Крахмал гидролизуется альфа-амилазой с образованием растворимых синих фрагментов. Поглощение полученного синего раствора, измеренное спектрофотометрически при 620 нм, является функцией альфа-амилазной активности.
Важно, чтобы измеренное поглощение при 620 нм после 10 или 15 минут инкубации (времени тестирования) находилось в диапазоне 0,2-2,0 единиц поглощения при 620 нм. В этом диапазоне абсорбции существует линейность между активностью и абсорбцией (закон Ламберта-Беера). Таким образом, разбавление фермента должно быть скорректировано для удовлетворения этого критерия. При указанном наборе условий (температуре, pH, времени реакции, буферных условиях) 1 мг конкретной альфа-амилазы будет гидролизовать определенное количество субстрата и будет образовываться синяя окраска. Интенсивность окраски измеряют при 620 нм. Измеренная абсорбция прямо пропорциональна удельной активности (активность/мг белка чистой альфа-амилазы) рассматриваемой альфа-амилазы при данном конкретном наборе условий.
Определение удельной активности
Удельную активность определяют с использованием теста Phadebas (Pharmacia) в виде активность/мг фермента.
Измерение рН-профиля активности (рН-стабильности)
Вариант альфа-амилазы хранят в 20 мМ Трисе рН 7,5, 0,1 М СаСl2 и тестируют при 30° С, 50 мМ буфера Бриттона-Робинсона, 0,1 мМ CaCl2. Активность измеряют при рН 4,0, 4,5, 5,0, 5,5, 6,0, 7,0, 8,0, 9,0, 9,5, 10 и 10,5 с использованием анализа Phadebas, описанного выше.
Определение активности AGU и AGU/мг
Одна амилоглюкозидазная единица Novo (AGU) определяется как количество фермента, которое гидролизует 1 микромоль мальтозы в минуту при 37° С и рН 4,3. Подробное описание этого аналитического способа можно получить по запросу из Novo Nordisk.
Активность определяют в виде AGU/мл по способу, модифицированному по (AEL-SM-0131), с использованием набора Glucose GOD-Perid Kit из Boehringer Mannheim, 124036. Стандарт: AMG-стандарт, партия 7-1195, 195 AGU/мл.
375 мкл субстрата (1% мальтозы в 50 мМ ацетате натрия, рН 4,3) инкубируют 5 минут при 37° С. Добавляют 25 мкл фермента, разбавленного в ацетате натрия. Реакцию останавливают после 10 минут добавлением 100 мкл 0,25 М NaOH. 20 мкл переносят в 96-луночный микротитрационный планшет и добавляют 200 мкл раствора GOD-Perid. После 30 минут при комнатной температуре поглощение измеряют при 650 нм и активность рассчитывают в AGU/мл на основании AMG-стандарта.
Затем рассчитывают удельную активность в AGU/мг из активности (AGU/мл), деленной на концентрацию белка (мг/мл).
ПРИМЕРЫ
ПРИМЕР 1
Конструирование вариантов Термамила в соответствии с данным изобретением
Термамил (альфа-амилазу В. licheniformis SEQ ID NO:4) экспрессируют в В. subtilis из плазмиды, названной pDN1528. Эта плазмида содержит полный ген, кодирующий Термамил, amyL, экспрессия которого направляется его собственным промотором. Далее, эта плазмида содержит точку начала репликации, ori, из плазмиды pUB110 и ген cat из плазмиды рС194, придающий устойчивость к хлорамфениколу. pDN1528 показана на фиг.9 WO 96/23874. Получали вектор специфического мутагенеза, содержащий основную часть кодирующей области SEQ ID NO:3. Важные признаки этого вектора, названного pJeEN1, включают в себя начало репликации, происходящее из плазмид pUC, ген cat, придающий устойчивость к хлорамфениколу, и содержащую сдвиг рамки версию гена blа, дикий тип которого обычно придает устойчивость к ампициллину (фенотип аmрR). Эта мутированная версия приводит к фенотипу аmрS. Плазмида pJeEN1 показана на фиг.10 WO 96/23874, и на этой плазмиде показаны начало репликации Е. coli, ori, bla, cat, 5'-укороченная версия гена Термамил-амилазы и сайты рестрикции.
Мутации вводят в amyL по способу, описанному Deng and Nickoloff (1992, Anal. Biochem. 200, pp. 81-88), за исключением того, что плазмиды с включенным "праймером отбора" (праймером № 6616; см. ниже) выбраны на основе фенотипа аmрR трансформированных клеток Е. coli, несущих плазмиду с репарированным геном bla, вместо применения отбора расщеплением рестриктазами, описанного Deng and Nickoloff. Химикалии и ферменты, используемые для мутагенеза, получали из набора для мутагенеза  из Stratagene (номер по каталогу 200509).
из Stratagene (номер по каталогу 200509).
После подтверждения последовательности ДНК в плазмидах с вариантом укороченный ген, содержащий желательное изменение, субклонировали в pDN1528 в виде PstI-EcoRI-фрагмента и им трансформировали дефицитный по протеазе и амилазе штамм Bacillus subtilis SHA273 (описанный в WO 92/11357 и WO 95/10603) для экспрессии вариантного фермента.
Вариант V54W Термамила конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PG GTC GTA GGC ACC GTA GCC ССА АТС CGC TTG (SEQ ID NO:9)
Вариант Термамила A52W+V54W конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PG GTC GTA GGC ACC GTA GCC ССА АТС ССА TTG GCT CG (SEQ ID NO:10)
Прaймер № 6616 (написанный 5'-3', слева направо); Р обозначает 5'-фосфат):
Р CTG TGA CTG GTG AGT ACT CAA ССА AGT С (SEQ ID NO:11)
Вариант Термамила V54E конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGG TCG TAG GCA CCG TAG CCC ТСА ТСС GCT TG (SEQ ID NO:12)
Вариант Термамила V54M конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGG TCG TAG GCA CCG TAG CCC ATA TCC GCT TG (SEQ ID NO:13)
Вариант Термамила V54I конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGG TCG TAG GCA CCG TAG CCA ATA TCC GCT TG (SEQ ID NO:14)
Варианты Термамила Y290E и Y290K конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGC AGC ATG GAA CTG CTY ATG AAG AGG САС GTC ААА С (SEQ ID NO:15)
Y обозначает равную смесь С и Т. Присутствие кодона, кодирующего глутамат или лизин в положении 290, подтверждали секвенированием ДНК.
Вариант Термамила N190F конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
РСА TAG TTG CCG ААТ ТСА TTG GAA ACT TCC С (SEQ ID NO:16)
Вариант Термамила N188P+N190F конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
РСА TAG TTG CCG ААТ ТСА GGG GAA ACT TCC CAA TC (SEQ ID NO:17)
Вариант Термамила H140K+H142D конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
РСС GCG CCC CGG GAA АТС ААА ТТТ TGT CCA CGC ТТТ ААТ TAG (SEQ ID NO:18)
Вариант Термамила H156Y конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
РСА ААА TGG ТАС САА ТАС САС ТТА ААА TCG CTG (SEQ ID NO:19)
Вариант Термамила А181Т конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
РСТ ТСС САА ТСС САА GTC ТТС ССТ TGA AAC (SEQ ID NO:20)
Вариант Термамила A209V конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
РСТТ ААТ ТТС TGC ТАС GAC GTC AGG ATG GTC АТА АТС (SEQ ID NO:21);
Вариант Термамила Q264S конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PCG CCC AAG ТСА ТТС GAC CAG ТАС ТСА GCT ACC GTA AAC (SEQ ID NO:22)
Вариант Термамила S187D конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGC CGT ТТТ CAT TGT CGA СТТ CCC ААТ CCC (SEQ ID NO:23)
Вариант Термамила DELTA(K370-G371-D372) (т.е. делетированные аминокислотные остатки с номерами 370, 371 и 372) конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGG ААТ ТТС GCG CTG ACT AGT CCC GTA CAT АТС CCC (SEQ ID NO:24)
Вариант Термамила DELTA(D372-S373-Q374) конструировали с использованием следующего мутагенного праймера (написанного 5'-3', слева направо):
PGG CAG GAA TTT CGC GAC CTT TCG TCC CGT АСА ТАТ С (SEQ ID NO:25)
Варианты термамила А181Т и A209V объединяли с A181T+A209V расщеплением А181T-содержащей рDN1528-подобной плазмиды (т.е. pDN1528, содержащей в гене amyL мутацию, приводящую к изменению А181Т) и А209V-содержащей рDN1528-подобной плазмиды (т.е. pDN1528, содержащей в гене amyL мутацию, приводящую к изменению A209V) рестриктазой ClaI, которая разрезает pDN1528-подобные плазмиды дважды, приводя к образованию фрагмента длиной 1116 п.н. и векторной части (т.е. содержит точку начала репликации плазмиды) длиной 3850 п.н. Фрагмент, содержащий мутацию A209V, и векторную часть, содержащую мутацию А181Т, очищали с использованием набора для экстракции гелей QIAquick (купленного в QIAGEN) после разделения на агарозном геле. Этот фрагмент и вектор лигировали и им трансформировали дефицитный по протеазе и амилазе штамм Bacillus subtilis, указанный выше. Плазмиду из аmу+ (просветленные зоны на содержащих крахмал чашках с агаром) и устойчивые к хлорамфениколу трансформанты анализировали на присутствие обеих мутаций на плазмиде.
Подобно описанному выше, объединяли H156Y и A209V с использованием рестриктаз Асс651 и EcoRI с получением H156Y+A209V.
H156Y+A209V и A181T+A209V объединяли в H156Y+A181T+A209V с использованием рестриктаз Асс651 и HindIII.
35 N-концевых остатков зрелой части варианта Термамила H156Y+A181T+A209V заменяли 33 N-концевыми остатками альфа-амилазы В. amyloliquefaciens (SEQ ID NO:4) (которую в данном контексте называют BAN) с использованием методики SOE-PCR (Higuchi et al. 1988, Nucleic Acids Research 16:7351) следующим образом:
Праймер 19364 (последовательность 5'-3'): ССТ CAT TCT GCA GCA GCA GCC GTA AAT GGC ACG CTG (SEQ ID NO:26)
Праймер 19362: ССА GAC GGC AGT AAT ACC GAT ATC CGA TAA ATG TTC CG (SEQ ID NO:27)
Праймер 19363: CGG ATA TCG GTA TTA CTG CCG TCT GGA TTC (SEQ ID NO:28)
Праймер 1С: CTC GTC CCA ATC GGT TCC GTC (SEQ ID NO:29)
Стандартную ПЦР, полимеразную цепную реакцию, проводили с использованием термостабильной полимеразы Pwo из Boehringer Mannheim в соответствии с инструкциями изготовителя и с температурным циклом: 5 мин при 94° С, 25 циклов (94° С в течение 30 сек, 50° С в течение 45 сек, 72° С в течение 1 мин), 72° С в течение 10 мин.
Фрагмент длиной приблизительно 130 п.н. амплифицировали в первой ПЦР, обозначаемой ПЦР1, с праймерами 19364 и 19362 на ДНК-фрагменте, содержащем ген, кодирующий альфа-амилазу В. amyloliquefaciens.
Фрагмент длиной приблизительно 400 п.н. амплифицировали в другой ПЦР, обозначаемой ПЦР2, с праймерами 19363 и 1С на матрице pDN1528.
Продукты ПЦР1 и ПЦР2 выделяли из агарозного геля и использовали в качестве матриц в ПЦРЗ с праймерами 19364 и 1С, что приводило к фрагменту длиной приблизительно 520 п.н. Таким образом, этот фрагмент содержал одну часть ДНК, кодирующей N-конец из BAN, слитый с частью ДНК, кодирующей Термамил, от 35-ой аминокислоты.
Фрагмент длиной 520 п.н. субклонировали в pDN1528-подобную плазмиду (содержащую ген, кодирующий вариант Термамила H156Y+A181T+A209V) расщеплением рестриктазами PstI и SacII, лигированием и трансформацией штамма В. subtilis, как описано ранее. Последовательность ДНК между сайтами рестрикции PstI и SacII проверяли ДНК-секвенированием в экстрагированных плазмидах из аmу+ и устойчивых к хлорамфениколу трансформантов.
Конечная конструкция, содержащая правильный N-конец из BAN и H156Y+A181T+A209V, была названа BAN(1-35)+H156Y+A181T+A209V.
N190F объединяли с BAN(1-35)+H156Y+A181T+A209V с получением BAN(1-35)+H156Y+A181T+N190F+A209V проведением мутагенеза, как описано выше, за исключением того, что последовательность amyL в pJeENI заменяли последовательностью ДНК, кодирующей вариант Термамила BAN(1-35)+H156Y+A181T+A209V.
Q264S объединяли с BAN(1-35)+H156Y+A181T+A209V с получением BAN(1-35)+H156Y+A181T+A209V+Q264S проведением мутагенеза, как описано выше, за исключением того, что последовательность amyL в pJeEN1 заменяли последовательностью ДНК, кодирующей вариант Термамила BAN(1-35)+H156Y+A181T+A209V.
BAN(1-35)+H156Y+A181T+A209V+Q264S и BAN(1-35)+H156Y+A181T+N190F+A209V объединяли в BAN(1-35)+H156Y+A181T+N190F+A209V+Q264S с использованием рестриктаз BsaHI (BsaHI-сайт вводили вблизи от мутации A209V) и PstI.
I201F объединяли с BAN(1-35)+H156Y+A181T+N190F+A209V+Q264S с получением BAN(1-35)+H156Y+A181T+N190F+A209V+Q264S+I201F (SEQ ID NO:2) проведением мутагенеза, как описано выше. Использовали праймер мутагенеза AM100, вводили замену I201F и удаляли одновременно сайт рестрикции ClaI, что облегчает легкое получение точковых мутантов.
Праймер AM100:
5' GATGTATGCCGACTTCGATTATGACC 3' (SEQ ID NO:30)
ПРИМЕР 2
Конструирование вариантов Термамил-подобной альфа-амилазы с измененным характером расщепления в соответствии с данным изобретением
Вариант термостабильной альфа-амилазы В. licheniformis, содержащий 445 С-концевых аминокислотных остатков альфа-амилазы В. licheniformis, показанной в SEQ ID NO:4, и 37 N-концевых аминокислотных остатков альфа-амилазы, происходящей из В. amyloliquefaciens, показанной в SEQ ID NO:6, и дополнительно содержащий следующие мутации: H156Y+A181T+N190F+A209V+ Q264S+I201F (конструирование этого варианта описано в примере 1, а аминокислотная последовательность показана в SEQ ID NO:2), имел пониженную способность расщепления субстрата вблизи точки ветвления.
В попытке дальнейшего улучшения пониженной способности расщепления субстрата вблизи точки ветвления указанного варианта альфа-амилазы проводили сайт-направленный мутагенез с использованием Мега-праймерного способа, описанного Sarkar and Sommer, 1990 (BioTechniques 8:404-407):
Конструирование LE313: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+V54N:
Ген-специфический праймер 27274 и мутагенный праймер AM115 используют для амплификации при помощи ПЦР ДНК-фрагмента длиной приблизительно 440 п.н. из рDN1528-подобной плазмиды (несущей мутации BAN(1-35)+H156Y+A181T+N190F+I120F+A209V+ Q264S в гене, кодирующем амилазу, из SEQ ID NO:4).
Фрагмент длиной 440 п.н. выделяют из агарозного геля и используют в качестве Мега-праймера вместе с праймером 113711 во второй ПЦР, проводимой на той же самой матрице.
Полученный фрагмент длиной приблизительно 630 п.н. расщепляют рестриктазами EcoRI и Асс651 и полученный ДНК-фрагмент длиной приблизительно 370 п.н. очищают и лигируют с pDN1528-подобной плазмидой, расщепленной теми же самыми ферментами. Компетентные клетки Bacillus subtilis SHA273 (с низким содержанием амилазы и протеазы) трансформируют с лигированием и устойчивые к хлорамфениколу трансформанты проверяют ДНК-секвенированием для подтверждения присутствия правильных мутаций на плазмиде.
Праймер 27274:
5' CATAGTTGCCGAATTCATTGGAAACTTCCC 3' (SEQ ID NO:31)
Праймер 1В:
5' CCGATTGCTGACGCTGTTATTTGC 3' (SEQ ID NO:32)
Праймер AM115:
5' GCCAAGCGGATAACGGCTACGGTGC 3' (SEQ ID NO:33)
Конструирование LE314: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+A52S проводят сходным образом, за исключением того, что используют мутагенный праймер AMI 16.
АМ116:
5' GAACGAGCCAATCGGACGTGGGCTACGG 3' (SEQ ID NO:34)
Конструирование LE315: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+A52S+V54N проводят сходным образом, за исключением того, что используют мутагенный праймер АМ117.
АМ117:
5' GGAACGAGCCAATCGGATAACGGCTACGGTGC 3' (SEQ ID NO:35)
Конструирование LE316; BAN/Термамил гибрид+H156Y+A181T+ N190F+A209V+Q264S+T49L проводят сходным образом, за исключением того, что используют мутагенный праймер АМ118.
АМ118:
5' GCATATAAGGGACTGAGCCAAGCGG 3' (SEQ ID NO:36)
Конструирование LE317: BAN/Термамил гибрид+ H156Y+A181T+N190F+A209V+Q264S+T49L+G107A проводят сходным образом, за исключением того, что используют одновременно мутагенный праймер AM118 и мутагенный праймер AM119.
АМ119:
5' CAACCACAAAGCCGGCGCTGATGCG 3' (SEQ ID NO:37)
Конструирование LE318: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+A52S+V54N+T49L+G107A проводят сходным образом, за исключением того, что используют одновременно мутагенный праймер АМ120 и мутагенный праймер АМ119.
АМ120:
5' GCATATAAGGGACTGAGCCAATCGGATAACGGCTACGGTGC 3' (SEQ ID NO:38)
Конструирование LE319: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+A52S+V54N+T49L проводят сходным образом, за исключением того, что используют мутагенный праймер АМ120.
Конструирование LE320: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+G107A проводят сходным образом, за исключением того, что используют мутагенный праймер АМ119.
Конструирование LE322: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+Q51R+A52S проводят сходным образом, за исключением того, что используют мутагенный праймер АМ121.
АМ121:
5' GAACGAGCCGATCGGACGTGGGCTACGG 3' (SEQ ID NO:39)
Конструирование LE323: BAN/Термамил гибрид+H156Y+A181T+N190F+A209V+Q264S+A52N проводят сходным образом, за исключением того, что используют мутагенный праймер АМ122.
АМ122:
5' GAACGAGCCAAAACGACGTGGGCTACGG 3' (SEQ ID NO:40)
ПРИМЕР 3
Испытание вариантов LE429 (осахаривание)
Использовали следующие стандартные условия реакции:
Концентрация субстрата 30% мас./мас.
Температура 60° С
Исходный рН (при 60° С) 5,5
Доза фермента
Глюкоамилаза 0,18 AGU/г DS
Пуллуланаза 0,06 PUN/г DS
Альфа-амилаза 10 мкг фермента/г DS
Декстрозим™ использовали для обеспечения глюкоамилазной и пуллуланазной активностей.
Субстраты для осахаривания готовили растворением обычного кукурузного крахмала в деионизованной воде и доведением сухого вещества до приблизительно 30% мас./мас. рН доводили до 5,5 (измеряли при 60° С) и аликвоты субстрата, соответствующие 10 г сухого веса, переносили в стеклянные колбы с синими крышками.
Затем эти колбы помещали во встряхивающую водяную баню, уравновешенную при 60° С, и добавляли ферменты. Если необходимо, рН повторно доводили до 5,5. Пробы брали после 48 часов осахаривания; рН доводили до приблизительно 3,0 и затем нагревали в кипящей водяной бане в течение 15 минут для инактивации ферментов. После охлаждения пробы обрабатывали приблизительно 0,1 г смешанного слоя ионообменной смолы (BIO-RAD 501 Х8 (D)) в течение 30 минут на ротационной мешалке для удаления солей и растворимого N. После фильтрования углеводный состав определяли при помощи ВЭЖХ. Получали следующие результаты:
Исходной альфа-амилазой для этих вариантов является LE429 (табл. 1).

В сравнении с контролем присутствие активного варианта альфа-амилазы данного изобретения во время разжижения приводит к уменьшенному содержанию панозы (DP3).
Особенно вариант T49L+G107A LE429 и вариант A52S+V54N+T49L LE429 соответственно приводят к очень сильно уменьшенному содержанию панозы (DР3). Если эти варианты альфа-амилазы используются для разжижения крахмала, исчезнет необходимость в инактивации этого фермента перед началом осахаривания.
ПРИМЕР 4
Разжижение и осахаривание с использованием вариантов LE429
Эксперимент примера 3 повторяли для ряда других вариантов LE429 при тех же самых условиях.
Результат показан в табл.2.

ПРИМЕР 5
Эксперимент в примере 3 повторяли для ряда вариантов LE429, за исключением того, что разжижение проводили при 95° С, рН 6,0, а осахаривание при 60° С, рН 4,5, 40 м.д. CaCl2, с последующей инактивацией. Этот вариант называют ниже вариантом LE429. Были получены следующие результаты (табл.3).
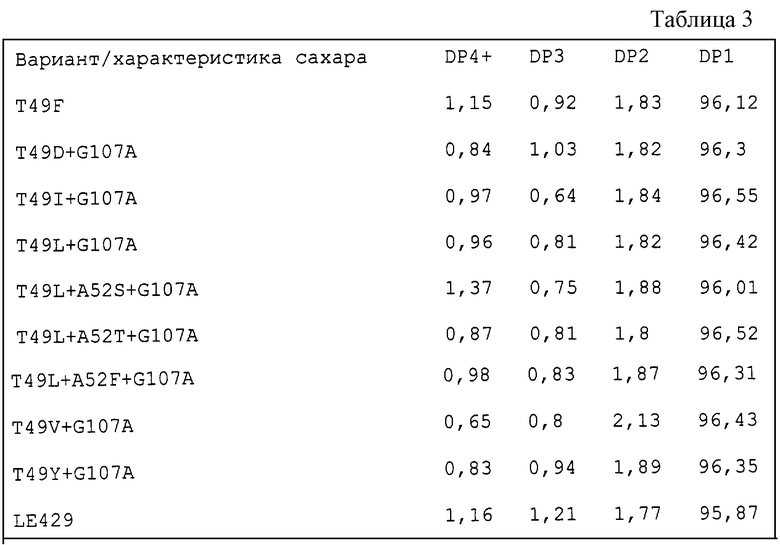
Список последовательностей приведен в конце описания.
Источники информации
Klein, С., et al., Biochemistry 1992, 31, 8740-8746.
Mizuno, H., et al.. J. Mol. Biol. (1993) 234, 1282-1283.
Chang, С., et al, J. Mol. Biol. (1993) 229, 235-238.
Larson, S.B., J. Mol. Biol. (1994) 235, 1560-1584.
Lawson, C.L., J. Mol. Biol. (1994) 236, 590-600.
Qian, M., et al., J. Mol. Biol. (1993) 231, 785-799.
Brady, R.L., et al., Acta Crystallogr. sect. B, 47, 527-535.
Swift, H.J.. et al., Acta Crystallogr. sect. B, 47, 535-544.
A. Kadziola, Ph.D. Thesis: "An alpha-amylase from Barley and its Complex with a Substrate Analogue Inhibitor Studied by X-ray Crystallography", Department of Chemistry University of Copenhagen 1993.
MacGregor, E.A., Food Hydrocolloids, 1987, Vol.1, No. 5-6. p. B.Diderichsen and L.Christiansen, Cloning of a maltogenic amylase from Bacillus stearothemophilus, FEMS Microbiol. letters: 56: pp.53-60 (1988).
Hudson et al., Practical Immunology, Third edition (1989), Blackwell Scientific Publications.
Sambrook et al., Molecular Cloning: A Laboratory Manual. 2nd Ed., Cold Spring Harbor, 1989.
S.L.Beaucage and M.H. Caruthers, Tetrahedron Letters 22, 1981, pp. 1859-1869.
Matthes et al., The EMBO J. 3, 1984, pp. 801-805.
R.K.Saiki et al., Science 239, 1988, pp.487-491.
Morinaga et al., (1984, Biotechnology 2:646-639).
Nelson and Long, Analytical Biochemistry 180, 1989, pp.147-151.
Hunkapiller et al., 1984, Nature 310:105-111.
R.Higuchi, B.Krummel, and R.K.Saiki (1988). A general method of in vitro preparation and specific mutagenesis of DNA fragments: study of protein and DNA interactions. Nucl. Acids Res. 16:7351-7367.
Dubnau et al., 1971, J. Mol. Biol. 56, pp. 209-221.
Gryczan et al., 1978, J.Bacteriol. 134, pp. 318-329.
S.D.Erlich, 1977, Proc. Natl. Acad. Sci. 74, pp.1680-1682.
Boel et al., 1990, Biochemistry 29, pp. 6244-6249.
Sarkar and Sommer, 1990, BioTechniques 8, pp. 404-407.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ САХАРОВ И СИРОПОВ | 2014 |
|
RU2705243C2 |
| Трансформант Ogataea haglerorum - продуцент термостабильной α-амилазы | 2022 |
|
RU2795707C1 |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ С ИЗМЕНЕННЫМИ СВОЙСТВАМИ | 2008 |
|
RU2499044C2 |
| ВАРИАНТЫ ЛИПАЗЫ И КОМПОЗИЦИИ В ВИДЕ МИКРОКАПСУЛ, СОДЕРЖАЩИЕ ТАКИЕ ВАРИАНТЫ ЛИПАЗЫ | 2018 |
|
RU2779301C2 |
| МОЮЩИЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ЛИПАЗЫ | 2018 |
|
RU2775700C2 |
| ВАРИАНТЫ ПРОТЕАЗЫ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2014 |
|
RU2670946C9 |
| СПОСОБ ЭКСПРЕССИИ | 2012 |
|
RU2642324C2 |
| ВАРИАНТЫ ГЛЮКОАМИЛАЗЫ | 2006 |
|
RU2439153C2 |
| ВАРИАНТЫ СУБТИЛАЗЫ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2015 |
|
RU2710720C2 |
| БЕТА-ЛАКТАМАЗА, СПОСОБ ЕЕ МОДИФИКАЦИИ, ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2011 |
|
RU2570551C2 |






















Изобретение относится к биотехнологии. Вариант исходной Термамил-подобной альфа-амилазы содержит изменение в одном или нескольких положениях исходной альфа-амилазы. Такие изменения представляют собой инсерции, делеции и замены аминокислот в аминокислотной последовательности. При этом указанный вариант сохраняет альфа-амилазную активность и проявляет измененные свойства, такие как пониженная способность расщепления субстрата вблизи точки ветвления, улучшенная субстратная специфичность. Данное изобретение повышает эффективность использования альфа-амилазы в моющих композициях, при производстве этанола, для обработки тканей или одежды. 12 з.п. ф-лы, 3 табл.
G48A, V, S, Т, I, L;
*48aD или *48aY (т.е. инсерция D или Y);
T49X;
*49aX (т.е. инсерция любого аминокислотного остатка);
S50X, в частности, D, Y, L, Т, V, I;
Q51R, K;
A52S, N, Т, V;
D53Q, N, S, T, V;
G107X, в частности, G107A, V, S, Т, I, L, С;
G108X, в частности, G108A, V, S, Т, I, L;
A111V, I, L;
S168Y;
M197Y, F, G.
T49L+G107A;
T49I+G107A;
T49L+G107A+V54I;
T49I+G107A+V54I;
A52S+V54N+T49L+G107A;
A52S+V54I+T49L+G 107А;
A52S+T49L+G107A;
A52T+T49L+G107A;
A52S+V54N+T49I+G107A;
A52S+V54I+T49I+G107A;
A52S+T49I+G107A;
T49L+G108A;
T49I+G108A;
T49L+G108A+V54I;
T49I+G108A+V54I.
| WO 9741213, 30.04.1997 | |||
| US 5989169, 23.11.1999 | |||
| WO 9418314, 18.08.1994 | |||
| MACGREGOR, J | |||
| PROTEIN CHEMISTRY, № 7, 1988, р.399-415. |
