ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к лекарственным средствам и модифицированным бета-лактамазам. Более конкретно настоящее изобретение относится к новым рекомбинантным бета-лактамазам и фармацевтическим композициям, содержащим бета-лактамазы.
Настоящее изобретение также относится к способам модификации бета-лактамазы, способам получения бета-лактамазы, а также к способам лечения или профилактики побочных эффектов, вызванных приемом бета-лактамных антибиотиков. Кроме того, настоящее изобретение относится к бета-лактамазе, которая может использоваться в качестве самостоятельного лекарственного средства или для производства лекарственного средства, предназначенного для лечения и профилактики побочных эффектов, вызванных приемом бета-лактамных антибиотиков.
Настоящее изобретение также относится к полинуклеотиду и клетке-хозяину, содержащей указанный полинуклеотид.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Бета-лактамные антибиотики характеризуются наличием бета-лактамного кольца в своей молекулярной структуре. Целостность бета-лактамного кольца необходима для биологической активности антибиотиков, которая проявляется в инактивации ряда транспептидаз, которые катализируют конечные реакции перекрестного связывания в синтезе пептидогликанов. Членами семейства бета-лактамных антибиотиков являются пенициллины, цефалоспорины, клавамы (или оксапенамы), цефамицины и карбапенемы.
Бета-лактамазы представляют собой защитные ферменты бактерий, которые гидролизуют бета-лактамные антибиотики. Продукция бета-лактамаз является основным механизмом, определяющим устойчивость Грам-отрицательных бактерий к бета-лактамам. Бета-лактамазы катализируют очень эффективный необратимый гидролиз амидной связи бета-лактамного кольца, что приводит к образованию биологически неактивного продукта(ов).
По причине того, что разные типы бета-лактамаз имеют различные ферментные характеристики, было предложено несколько систем для их классификации. Эти классификации базируются на двух основных подходах и представляют собой функциональную и молекулярную классификации.
Схема функциональной классификации бета-лактамаз предложена (Bush et al., 1995, Antimicrob. Agents Chemother. 39: 1211-1233) и включает четыре группы бета-лактамаз, в зависимости от их субстрата и ингибитора. Группа 1 включает цефалоспориназы, которые не очень хорошо ингибируются клавулановой кислотой. Группа 2 включает пенициллиназы, цефалоспориназы и бета-лактамазы широкого спектра, которые, в целом, ингибируются активными сайт-направленными ингибиторами бета-лактамаз. Группа 3 включает металло-бета-лактамазы, которые гидролизуют пенициллины, цефалоспорины и карбапенемы и слабо ингибируются большинством бета-лактам-содержащих молекул. Группа 4 включает пенициллиназы, которые плохо ингибируются клавулановой кислотой. Помимо этого, на основании скорости гидролиза карбенициллина или клоксациллина (оксациллина) среди пенициллиназ из 2 группы выделяют несколько подгрупп.
Наиболее широко используемой является классификация Ambler, которая разделяет бета-лактамазы на четыре класса (A, B, C, D) и базируется на аминокислотной последовательности ферментов (Ambler 1980, Philos Trans R Soc Lond В Biol Sci. 289: 321-331). Классы A, C и D объединяют эволюционно различные группы сериновых бета-лактамазных ферментов, а класс В включает цинк-зависимые ("ЭДТА-ингибируемые") бета-лактамазы (Ambler R.P. et al., 1991, Biochem J. 276:269-270). Классы A, C и D включают сериновые бета-лактамазы, в которых гидролиз бета-лактама опосредуется серином в активном сайте. Сериновые бета-лактамазы относятся к DD пептидазам (D-аланил-D-аланин карбоксипептидаза), мишенями которых являются бета-лактамы. Считается, что механизм, при помощи которого сериновые бета-лактамазы гидролизуют бета-лактамные антибиотики, представляет собой трехэтапный каскад, включающий образование нековалентного комплекса Генри-Михаэлиса (Henri-Michaelis), ковалентного промежуточного ацил-фермента и реакцию деацетилирования (Matagne et al., 1998, Biochem J 330:581-598). Существует мнение, что механизм ацетилирования - это общий механизм для всех групп сериновых бета-лактамаз, между тем как, на основании теоретических расчетов, предполагается, что механизмы деацетилирования субстрата сериновых бета-лактамаз классов A, C и D отличаются. Механизмы деацетилирования имеют как и общие для всех групп черты, так и специфические для каждой группы молекулярные реакции (Hata et al., 2006, Biol Pharm Bull. 29: 2 151- 21 59).
Семейства сериновых бета-лактамаз Bacillus spp и семейства ТЕМ-1, SHV-1 и СТХ-М первоначально классифицировались как бета-лактамазы класса А и как пенициллиназы, обладающие хорошей способностью к гидролизу, например, пенициллина и ампициллина. Бета-лактамазы класса А впервые были идентифицированы у пенициллин-резистентного St. aureus в 1940-х г.г. Ген резистентности к пенициллину ТЕМ-1, связанный с плазмидой, был открыт у E.coli на 20 лет позже. В дальнейшем также было показано, что бета-лактамазы обладают способностью к гидролизу большинства цефалоспоринов и, кроме того, приспособились к гидролизу специфической субпопуляции цефалоспоринов. Большинство указанных бета-лактамаз с расширенным спектром действия (ESBL, от англ. Extended-Spectrum Beta-Lactamases) являются производными ферментов ТЕМ-1, ТЕМ-2 или SHV-1. Последнее время появляется все больше сообщений о выявлении большого числа СТХ-М ферментов, новой группы ESBLs класса А. В настоящее время СТХ-М ферменты являются наиболее часто встречающимися ESBLs и подразделяются на пять основных семейств. Ферменты СТХ-М имеют широкий спектр субстратов, включая пенициллин и первое, второе и третье поколение цефалоспоринов (Bonnet, R. 2004. Antimicrob Agents Chemother. 48:1-14).
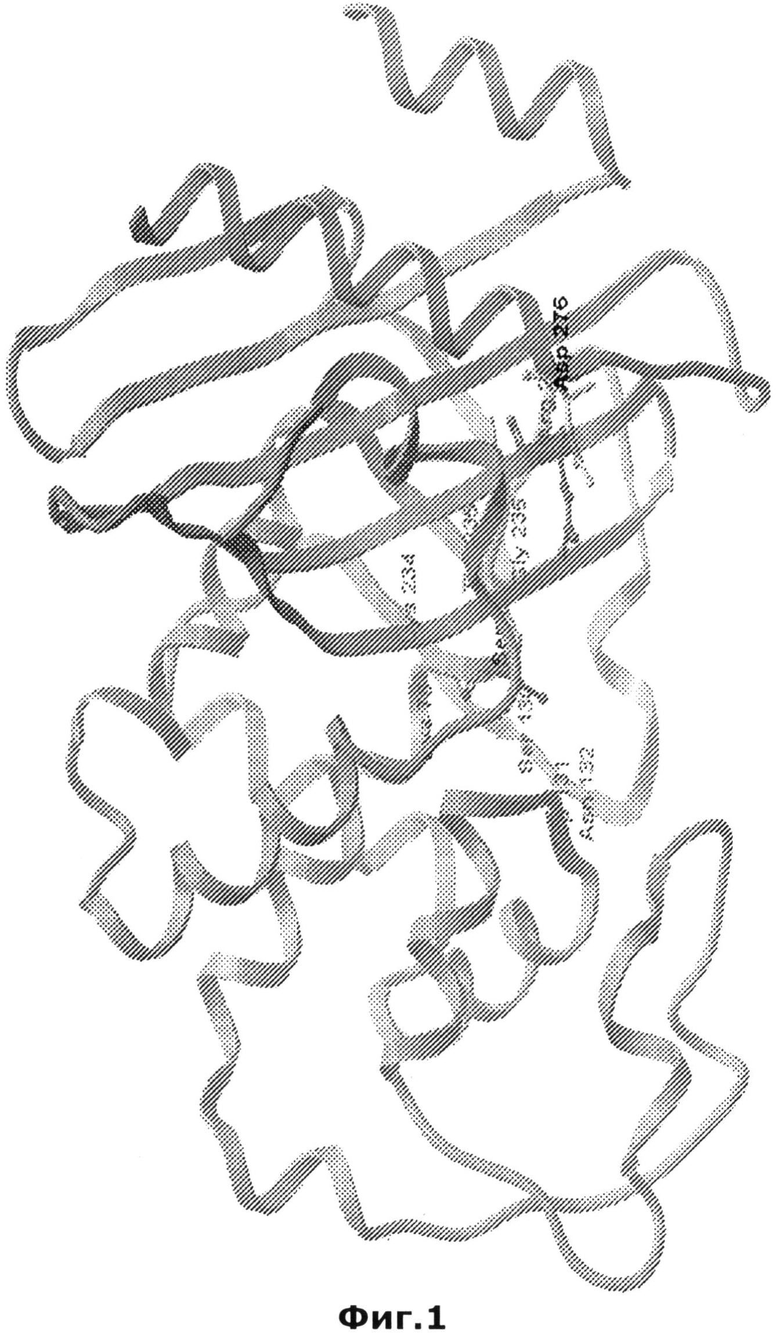
Несмотря на то, что сходство последовательностей бета-лактамаз класса А (ТЕМ, SHV, СТХ-М, бета-лактамазы Bacillus spp.) является незначительным, кристаллические структуры всех сериновых бета-лактамаз имеют высокую степень сходства (Matagne et al., 1998, Biochem J 330:581-598, Tranier S. et al., 2000, J Biol Chem, 275: 28075-28082; Santillana E. et al., 2007, Proc Natl Acad Sci. USA, 104: 5354-5359). Ферменты состоят из двух доменов. Один домен состоит из пятицепочечного бета-складчатого слоя, уложенного вплотную к трем альфа-спиралям, в то время как второй домен, альфа-домен, состоит из восьми альфа спиралей. Карман активного сайта является частью поверхности раздела между указанными двумя доменами и ограничен омега-петлей. Омега-петля представляет собой консервативный структурный элемент всех бета-лактамаз класса А, который главным образом участвует в каталитической реакции (Фиг.1).
У бета-лактамаз класса А были идентифицированы несколько консервативных пептидных последовательностей (элементов), которые имеют отношение к катализу или распознаванию субстрата. Первый консервативный элемент 70-Ser-X-X-Lys-73 (классификация Ambler) включает активный остаток серина в положении 70 в альфа-спиралиг и остаток лизина в положении 73. Второй консервативный элемент представляет собой SNX петлю в альфа домене (в положениях между 130 и 132, согласно классификации Ambler), где она формирует одну сторону каталитического углубления. Третий консервативный элемент (в положениях между 234 и 236, согласно классификации Ambler) находится на самой глубокой цепи бета-складчатой структуры3 и формирует другу сторону каталитического углубления. Третий консервативный элемент обычно представляет собой KTG. Однако, в некоторых исключительных случаях, лизин (К) может быть замещен гистидином (Н) или аргинином (R), а у некоторых бета-лактамаз треонин (Т) может быть замещен серином (S) (Matagne et ai, 1998. Biochem J 330: 581-598).
Устойчивость к бета-лактамам, обусловленная бета-лактамазами, широко распространена среди патогенной и условно-патогенной микрофлоры по причине чрезмерного использования бета-лактамов в последние десятилетия. Более того, устойчивость к антибиотикам представляет собой хорошо изученную клиническую проблему в медицине и ветеринарии; сотни различных бета-лактамаз, выделенных у Грам-положительных и Грам-отрицательных бактерий, были очищены и описаны в научной литературе. Поскольку использование антибиотиков не сокращается и, более того, устойчивость к антибиотикам стала частью повседневной жизни, для решения этой медицинской проблемы постоянно и достаточно срочно требуются новые подходы.
Кишечная микрофлора человека представляет собой сложное бактериальное сообщество, которое играет важную роль в здоровье человека, например, стимулирует систему иммунного ответа, способствует перевариванию пищи и препятствует чрезмерному росту потенциально патогенных бактерий. Известно, что антимикробные агенты, например, бета-лактамы, оказывают воздействие на нормальную микрофлору. Способность антимикробных агентов вызывать изменения нормальной кишечной микрофлоры определяется несколькими факторами, включая дозу препарата, способ его введения и фармакокинетику/фармакодинамику, а также свойства антибиотика (A. et al., 2001, Lancet 1: 101-114). Даже несмотря на то, что кишечная микрофлора обладает способностью восстанавливаться после завершения лечения антибиотиками, сообщается о длительной перстистенции определенных устойчивых условно-патогенных бактерий (Sjolund M. et al., 2003, Ann Intern Med. 139:483-487). Такая персистенция и обмен генами устойчивости к антибиотикам делают условно-патогенные микроорганизмы предполагаемым резервуаром генов устойчивости к антибиотикам.
Определенные бета-лактамы, вводимые парентерально, такие, как ампициллин, цефтриаксон, цефоперазон и пиперациллин частично выводятся с желчью в проксимальные отделы тонкого кишечника (двенадцатиперстной кишки). Оставшиеся неабсорбированные бета-лактамы в кишечнике могут оказывать нежелательное влияние на экологический баланс нормальной кишечной микрофлоры, что проявляется ассоциированной с приемом антибиотиков диареей, избыточным ростом патогенных микроорганизмов, таких, как ванкомицин-резистентные энтерококки (VRE, от англ. Vancomycin-resistant Enterococci), Грам-отрицательные палочки, продуцирующие бета-лактамазы с расширенным спектром действия (ESBL), Clostridium difficWe и грибы, а также отбором устойчивых к антибиотикам штаммов среди нормальной кишечной микрофлоры и потенциально патогенных бактерий.
Терапевтической целью бета-лактамаз является инактивация неабсорбированных антибиотиков в желудочно-кишечном тракте (GIT, от англ. Gastrointestinal Tract), русская аббревиатура ЖКТ, что позволяет поддерживать нормальную кишечную микрофлору и препятствует избыточному росту потенциально патогенных микроорганизмов (WO 93/13795).
Существует, по крайней мере, три основных требования для лекарственных продуктов, содержащих бета-лактамазы и подходящих для направленной терапии ЖКТ. Первое требование заключается в том, что лекарственное средство должно сохранять свою активность в условиях, существующих в ЖКТ. Устойчивость к протеолитическому расщеплению разнообразными протеазами, секретируемыми различными железами в ЖКТ, является важнейшим условием для успешного осуществления лечения бета-лактамазами. Другим важным фактором является разброс величин рН, существующий в различных отделах тонкого кишечника. Указанные величины рН обычно варьируют от 5 (двенадцатиперстная кишка) до 7,5 (подвздошная кишка). Следовательно, для того, чтобы соответствовать требованиям, предъявляемым к препаратам для указанных лечебных целей, бета-лактамазы должны демонстрировать высокую ферментативную активность при величине рН в пределах 5-7,5.
Вторым требованием, предъявляемым к бета-лактамазам или продуктам, их содержащим, является высокая эффективность гидролиза бета-лактамного кольца. Концентрация бета-лактамного антибиотика в химусе тонкого кишечника во время лечения антибиотиками обусловлена, главным образом, элиминацией определенного бета-лактама за счет экскреции с желчью. Подходящая бета-лактамаза должна обладать кинетическими параметрами, которые обеспечивают способность фермента эффективно гидролизировать низкие концентрации бета-лактамов в ЖКТ и снижать их концентрации до уровня, который не приводит к нежелательным нарушениям со стороны кишечной микрофлоры. Идеальный набор кинетических величин включает численно низкую величину константы Михаэлиса КМ в сочетании с численно высокой величиной максимальной скорости реакции Vmax. Высокая Vmax требуется для того, чтобы обеспечить достаточный уровень способности к гидролизу, в то время как низкая Км необходима для реализации активности фермента при низкой концентрации субстрата.
Третье требование, предъявляемое к бета-лактамазам или продуктам, их содержащим, заключается в том, что они должны выдерживать условия производства, такие, как относительно высокие температуры, используемые в процессе получения фармацевтических композиций. Кроме того, в процессе производства смешивание дисперсии водных носителей и лекарственного вещества требует высокого уровня растворимости при соответствующих величинах рН.
Ферментативная терапия, называемая Ipsat P1A, была разработана для профилактики побочных эффектов бета-лактамных антибиотиков в кишечнике. Система доставки Ipsat P1A была разработана для инактивации вводимых парентерально бета-лактамов из группы пенициллинов (например, пенициллина,
амоксициллина, ампициллина и пиперациллина) как вместе с ингибиторами бета-лактамазы, так и без них (например, тазобактам, сульбактам, клавулановая кислота) и экскретируемых с желчью (WO 2008065247; Tarkkanen, A.M. et al., 2009, Antimicrob Agents Chemother. 53:2455-2462). Фермент Р1А представляет собой рекомбинантную форму малой экзо бета-лактамазы Bacillus licheniformis 749/C (WO 2008065247), которая относится к классу А и подгруппе 2а, согласно функциональной классификации. Бета-лактамаза В. licheniformis и ее производное Р1А считаются пенициллиназами, обладающими высоким гидролизующим потенциалом в отношении, например, пенициллина, ампициллина, амоксициллина или пиперациллина (Таблица 1), при этом они, как правило, ингибируются сайт-направленными активными ингибиторами бета-лактамаз, такими, как клавулановая кислота, сульбактам или тазобакам (Bush K. et al., 1995, Antimicrob Agents Chemother 39: 121 1-1233).
Однако фермент Р1А обладает ограниченной способностью к инактивации бета-лактамных антибиотиков, которые относятся к группе цефалоспоринов или карбапенемов. Поскольку используемые бета-лактамазы обладают слабой активностью в отношении цефалоспоринов, они не могут применяться совместно с парентеральной терапией цефалоспоринами для инактивации неабсорбированных бета-лактамов в тонком кишечнике.
Следовательно, совершенно необходимы новые бета-лактамазы или производные Р1А с расширенным субстратным профилем, например, таким, который наблюдается у металло-бета-лактамаз.
Настоящее изобретение предлагает новые генетически созданные производные Р1А бета-лактамазы и более того, новые способы модификации и получения бета-лактамаз.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Новые рекомбинантные производные Р1А бета-лактамазы, заявленные в соответствии с настоящим изобретением, полностью удовлетворяют перечисленным выше трем требованиям, предъявляемым к бета-лактамазам (то есть обладают способностью сохранять ферментативную активность, эффективно гидролизировать бета-лактамы и выдерживать условия производства фармацевтических композиций) и, более того, обладают расширенным субстратным профилем. Бета-лактамазы, заявленные в соответствии с настоящим изобретением, также могут использоваться в сочетании с парентеральной терапией цефалоспоринами для инактивации элиминируемых с желчью бета-лактамов в тонком кишечнике.
Настоящее изобретение раскрывает предварительные и доклинические исследования нового Ipsat P3A фармацевтического белка (D276N замещенного производного Р1А) и предлагает единичную дозированную лекарственную форму на основе данного белка.
Настоящее изобретение относится к быстрым и эффективным способам модификации бета-лактамаз и способам получения модифицированных бета-лактамаз. Кроме того, благодаря настоящему изобретению станут доступны новые более эффективные и специфические способы лечения.
Ферменты, заявленные в соответствии с настоящим изобретением, подходят для широкомасштабного производства лекарственных препаратов, предназначенных для лечения или профилактики побочных эффектов, индуцированных различными группами бета-лактамных антибиотиков.
Задача настоящего изобретения заключается в том, чтобы предложить новые бета-лактамазы, в частности бета-лактамазы В. Licheniformis, а также продукты, способы получения и применения, касающиеся бета-лактамаз. Инструменты для новых разработок в фармацевтической индустрии также представлены в настоящем изобретении.
Настоящее изобретение относится к бета-лактамазе, имеющей аминокислотную последовательность, которая, по крайней мере, на 60% идентична SEQ ID NO:1 и включает гидрофильную аминокислоту в положении SED ID NO:1, соответствующем положению 276, согласно классификации Ambler, а также к вариантам и фрагментам указанной бета-лактамазы.
Изобретение также относится к фармацевтической композиции, содержащей бета-лактамазу, заявленную в соответствии с настоящим изобретением.
Настоящее изобретение также относится к способу модификации бета-лактамазы, имеющей аминокислотную последовательность, которая, по крайней мере, на 60% идентична SEQ ID NO:1, при этом аминокислота бета-лактамазы в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, замещена гидрофильной аминокислотой.
Кроме того, настоящее изобретение относится к способу получения бета-лактамазы, который включает следующие этапы:
а) получение гена, кодирующего бета-лактамазу, заявленную в соответствии с настоящим изобретением;
б) трансформацию клетки-хозяина указанным геном;
в) получение клетки-хозяина, продуцирующей бета-лактамазу;
г) восстановление бета-лактамазы.
Кроме того, настоящее изобретение относится к способу лечения или профилактики побочных эффектов со стороны желудочно-кишечного тракта, вызванных приемом бета-лактамных антибиотиков, который заключается во введении субъекту заявленной бета-лактамазы одновременно с бета-лактамным антибиотиком или после его приема.
Кроме того, настоящее изобретение относится к применению бета-лактамазы в качестве лекарственного средства.
Более того, настоящее изобретение относится к применению бета-лактамазы для производства лекарственного средства, предназначенного для лечения или профилактики побочных эффектов со стороны желудочно-кишечного тракта, вызванных приемом бета-лактамных антибиотиков.
Более того, настоящее изобретение относится к полинуклеотиду, который имеет любую из последовательностей SEQ ID NO:2 или 4, или его варианту, полученному на основе вырожденности генетического кода, или к полинуклеотиду, кодирующему бета-лактамазу, заявленную в соответствии с настоящим изобретением. Изобретение также относится к клетке-хозяину, содержащей указанный полинуклеотид.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На Фиг.1 представлена 3D структура бета-лактамазы Bacillus licheniformis (малая экзо-форма PenP). Обозначены консервативные аминокислотные остатки и остатки боковых цепей R-244 и D-278. Диаграмма была создана при помощи программы MolSof-Browser.
На Фиг.2 показаны нуклеотидная последовательность и выведенная на ее основе аминокислотная последовательность гена бета-лактамазы D276N, Bacillus licheniformis (P1A производное). Аминокислотная последовательность соответствует SEQ ID NO:3, при этом Хаа представляет собой аспарагин (Asn).
Нуклеотидная последовательность соответствует последовательности SEQ ID NO:4, при этом нуклеотидный триплет nnn представляет собой aat. Открытая рамка считывания кодирует полипептид из 299 аминокислот, включающий длинную сигнальную последовательность из 31 аминокислоты (подчеркнута) гена amyQ, происходящего из вектора секреции рКТН141 (WO 2008/065247). Предполагаемый сайт расщепления сигнальной пептидазой находится после аланина (А) в положении -1. Сайт клонирования HindIII, кодирующий NH2-QAS удлинение, выделен жирным шрифтом. Зрелый D276N мутантный фермент начинается с глутамина (Q)b положении +1. Таким образом, зрелая D276N мутантная бета-лактамаза имеет 268 аминокислотных остатка, включая NH2-QAS удлинение, кодируемое HindIII. Единичное аминокислотное замещение аспарагиновой кислоты (D) аспарагином (N) находится в положении 280 (выделено жирным шрифтом), что соответствует положению 276, согласно классификационной системе Ambler, и положению 249 в последовательности SEQ ID NO:3.
NH2-терминальная последовательность очищенного мутантного фермента D276N была определена при помощи автоматической деградации Эдмана в белковом секвенаторе. Анализ показал, что мутантный фермент D276N лишен NH2-QASKT-пентапептида на своем расшифрованном N-конце, как и его родоначальный фермент Р1А (WO 2008/065247). Основная фракция очищенного мутантного фермента D276N, которая использовалась в примерах 4 и 6 данной заявки, начинается с глутаминовой кислоты в положении +6 и состоит из 263 аминокислотных остатков с молекулярной массой 29272.
На Фиг.3 представлены нуклеотидная последовательность и выведенная на ее основе аминокислотная последовательности гена D276R замещенной бета-лактамазы Р1А, выделенной из Bacillus licheniformis. Аминокислотная последовательность соответствует последовательности SEQ ID NO:3, при этом Хаа представляет собой аргинин (Arg). Нуклеотидная последовательность соответствует последовательности SEQ ID NO:4, при этом нуклеотидный триплет nnn представляет собой cgc.
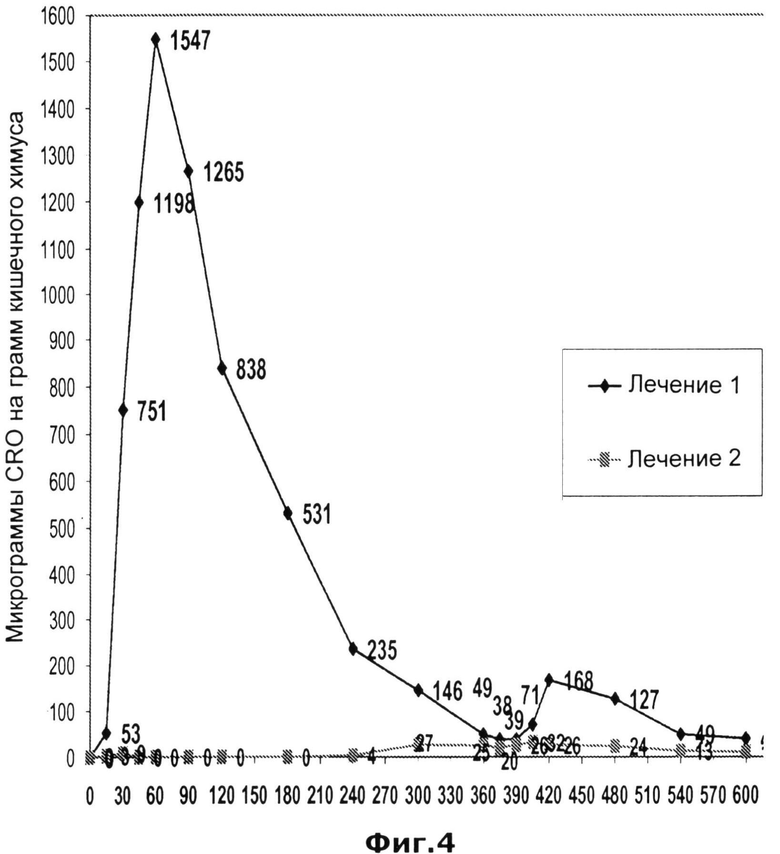
На Фиг.4 представлено влияние перорально вводимых пеллет, покрытых кишечнорастворимой оболочкой и содержащих D276N замещенную бета-лактамазу (Р3А), на концентрацию цефтриаксона в химусе тощей кишки у собак породы бигль (n=5) после внутривенного введения цефтриаксона (30 мг цефтриаксона на кг массы тела) (закрашенные квадраты). Пеллеты бета-лактамазы давали за 10 минут до введения цефтриаксона. Закрашенные ромбики соответствуют концентрации цефтриаксона в тощей кишке после введения единичной дозы цефтриаксона (в/в) без лечения бета-лактамазой.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Бета-лактамазы использовались для инактивации неабсорбированных бета-лактамов в желудочно-кишечном тракте для предотвращения побочных эффектов, вызванных приемом бета-лактамов, включая изменения со стороны нормальной кишечной микрофлоры и избыточный рост устойчивых к бета-лактамам бактерий (WO 199313795, WO 2007065247, WO 2007147945). Настоящее изобретение предлагает новые модифицированные бета-лактамазы Bacillus licheniformis, которые неожиданно демонстрируют расширенный модифицированный субстратный профиль.
Используемый здесь термин бета-лактамаза относится к ферменту, который гидролизует бета-лактамы. Гидролиз амидной связи бета-лактамного кольца приводит к тому, что антимикробные агенты теряют свою биологическую активность. Используемый здесь термин бета-лактамазы класса А (классификация Ambler) относится к сериновым бета- лактамазам, у которых гидролиз бета-лактама опосредуется серином в активном сайте, обычно аминокислотой в положении 70 альфа спирали2. Бета-лактамазы класса А включают, без ограничений указанными, Len-1, SHV-1, ТЕМ-1, PSE-3/PSE-3. ROB1, Bacillus cereus, такие, как 5/В тип 1, 569/Н тип 1 и 569/Н тип 3, Bacillus anthrasis sp, Bacillus licheniformis, такие как PenP, Bacillus weihenstephanensis, Bacillus clausii, Staphylococcus aureus и бета-лактамазы типов РС1, Sme-1, NmcA, IMI-, PER-, VEB-, GES-, KPC-, CME- и СТХ-М.
Идентичность последовательностей пептидов и полинуклеотидов
Аминокислотные последовательности мутантной бета-лактамазы, заявленной в соответствии с настоящим изобретением (D276X, Р1А производное), обозначены в тексте как SEQ ID NO:1 и SEQ ID NO:3. Соответствующие нуклеотидные последовательности приведены в заявке под обозначениями SEQ ID NO:2 и SEQ ID NO:4. SEQ ID NO:1 описывает аминокислотную последовательность, участвующую в формировании вторичной структуры бета-лактамазы. SEQ ID NO:3 описывает полноразмерную аминокислотную последовательность белка, включая длинную сигнальную последовательность из 31 аминокислоты.
Бета-лактамаза, заявленная в соответствии с настоящим изобретением, может иметь аминокислотную последовательность, которая, по крайней мере, на 30, 35, 40, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99,5, 99, 8, 99,9 или 100% идентична последовательности SEQ ID NO:1 или 3.
Согласно специфическому варианту осуществления изобретения, пептид, по крайней мере, на 30, 35, 40, 51, 52, 53, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99,5, 99,8, 99,9 или 100% идентичен последовательности SEQ ID NO:1 или 3.
Согласно одному предпочтительному варианту осуществления изобретения, бета-лактамаза имеет аминокислотную последовательность, которая, по крайней мере, на 60% идентична SEQ ID NO:1. Согласно другому предпочтительному варианту осуществления изобретения, бета-лактамаза имеет аминокислотную последовательность, которая, по крайней мере, на 60% идентична SEQ ID NO:1 или 3.
Согласно одному варианту осуществления изобретения, бета-лактамаза, имеющая аминокислотную последовательность с указанной выше идентичностью с SEQ ID NO:1, содержит гидрофильную аминокислоту, выбранную из группы, включающей аргинин (R), гистидин (Н), лизин (К), аспарагин (N), глутамин (Q), серии (S) и треонин (Т) в положении SEQ ID NO:1, соответствующем положению 276 согласно классификации Ambler.
Согласно предпочтительному варианту осуществления изобретения, пептид имеет последовательность, представленную в SEQ ID NO:1 или 3. Согласно одному варианту осуществления изобретения, бета-лактамаза имеет последовательность, представленную в SEQ ID NO:1 или 3, при этом остаток гидрофильной аминокислоты в положении, соответствующем положению 276, согласно классификации Ambler (обозначенный как Хаа в SEQ ID NO:1 или 3), представлен аргинином (R, Arg). Согласно другому варианту осуществления изобретения, бета-лактамаза имеет последовательность, представленную в SEQ ID NO:1 или 3, при этом остаток гидрофильной аминокислоты в положении, соответствующем положению 276, согласно классификации Ambler (обозначенный какХаа в SEQ ID NO:1 или 3) представлен аспарагином (N, Asn).
Идентичность любой последовательности с последовательностью, представленной в настоящем изобретении, подразумевает идентичность с полной последовательностью согласно настоящему изобретению. Идентичность последовательности может определяться при помощи любого метода биоинформации, например, при помощи BLAST (Basic Alignment Search Tools) или FASTA (FASTA-AII).
Настоящее изобретение также относится к любым вариантам или фрагментам новых бета-лактамаз. Используемые здесь термины "фрагмент" или "вариант" бета-лактамазы относятся к любой части или варианту, который обладает биологической функцией, то есть является ферментативно активным. Вариант относится к пептиду, имеющему незначительные изменения в пептидной последовательности, например, мутации, небольшие делеции или вставки. Фрагменты или варианты должны включать гидрофильную аминокислоту в положении, соответствующем положению 276, согласно классификации Ambler. Гидрофильная аминокислота обычно представляет собой любую кислоту, кроме аспарагиновой кислоты (D).
Существуют различные короткие формы бета-лактамазы, которые можно получить на основе SEQ ID NO:3 и которые секретируются вне клетки. Они называются экзоформами. Экзоформы являются результатом гидролитической активности протеаз в клеточной стенке или культуральной среде.
Термины D276X, D276N, D276R, мутантная форма, R1A производные или РЗА, используемые здесь, относятся к любому активному фрагменту бета-лактамазы и/или варианту SEQ ID NO:3, или варианту, явным образом включающему представленную аминокислотную последовательность (SEQ ID NO:1). В частности, бета-лактамаза, заявленная в соответствии с настоящим изобретением, представляет собой NH2-yce4eHHyra форму, она может иметь одну или более дополнительную аминокислотную делецию, замещение и/или вставку, но при этом обладает бета-лактамазной активностью. Указанные модификации могут представлять собой как существующие в природе варианты, так и мутанты, полученные при помощи искусственной модификации, например, при помощи генных технологий.
Различные усеченные по N-концу экзоформы были выявлены в питательной среде В. licheniformis. Такие формы также включены в настоящее изобретение. Matagne и др. описали различные степени микрогетерогенности внеклеточных форм, продуцируемых естественным хозяином, В. licheniformis 749/С (Matagne A. et al., 1991. Biochem J. 273: 503-510). Были идентифицированы следующие пять различных секретируемых экзоформ с различными аминокислотными остатками на N-конце:
Первичные аминокислотные остатки выделены жирным шрифтом. С-концевые аминокислотные остатки приведены в правой колонке. Экзоформа, начинающаяся с серина (S), обозначается как "большая секретируемая форма" бета-лактамазы В. licheniformis, а форма, начинающаяся с лизина (К), обозначается как "малая секретируемая форма".
Первая альфа-спираль (α1-спираль) начинается с аспарагиновой кислоты (D) (выделено курсивом), а концевая аминокислота последней альфа-спирали (α11-спираль) представлена аспарагином (N) (выделено курсивом). Согласно одному варианту осуществления изобретения, бета-лактамаза включает, по крайней мере, аминокислоты 1-258 SEQ ID NO:1 или аминокислоты 7-264 SEQ ID NO:3, которые принимают участие в формировании вторичной структуры белка (Knox J.R. et al, 1999. J. Mol. Biol. 220:435-455). Согласно другому варианту осуществления изобретения одна или более из указанных аминокислот 1-258 SEQ ID NO:1 или аминокислот 7-264 SEQ ID NO:3 подвергалась замещению или делеции.
Согласно еще одному варианту осуществления изобретения N-конец бета-лактамазы начинается с NH2-KTEMKDD (аминокислоты 4-10 SEQ ID NO:3). Эта, так называемая ES-beta L экзоформа, может быть дополнительно лишена вплоть до 21 последовательных аминокислот, как описано у Gebhard и др. (Gebhard L.G. et al., 2006, J. Mol. Biol. 21:358(1) 280-288). Согласно другому варианту осуществления изобретения, N-конец начинается с глутаминовой кислоты (Е) SEQ ID NO:3, конкретно, он начинается с NH2-EMKDD (аминокислоты 6-10 SEQ ID NO:3), или, альтернативно, он начинается с NH2-MKDD (аминокислоты 7-10 SEQ ID NO:3 или аминокислоты 1-4 SEQ ID NO:1).
Вариабельный участок в N-концевой последовательности бета-лактамазы не обладает ригидной структурой, которая обеспечивает постоянство ферментативных параметров различных форм бета-лактамаз.
Четыре последние аминокислоты карбоксильного конца бета-лактамазы, MNGK-COOH (аминокислоты 265-268 SEQ ID NO:3), не являются частью вторичной структуры, и, следовательно, могут быть удалены без потери ферментативной активности. Согласно другому варианту осуществления изобретения, вплоть до девяти аминокислот С-конца могут быть удалены. С-усеченные формы белка были описаны Santos и соавт.(Santos J. et al., 2004, Biochemistry 43:1715-1723).
Все различные формы бета-лактамазы, приведенные выше, включены в настоящее изобретение, вместе с другими формами белков, обладающих бета-лактамазной активностью.
Полинуклеотид, заявленный в соответствии с настоящим изобретением, может включать или иметь любую из последовательностей SEQ ID NO:2 или SEQ ID NO:4, или представлять собой вариант указанных последовательностей, полученный на основе вырожденности генетического кода. Полинуклеотид, который представляет собой указанный вариант последовательности, представленной как SEQ ID NO:2 или 4, относится к полинуклеотиду, который имеет один или более отличающийся нуклеотид, по сравнению с SEQ ID NO:2 или SEQ ID NO:4, но кодирует ту же аминокислоту. Предпочтительно, нуклеотидный триплет nnn последовательности SEQ ID NO:2 или SEQ ID NO:4 кодирует гидрофильную аминокислоту, наиболее предпочтительно, N или R. Термин "полинуклеотид", используемый здесь, представляет собой последовательность нуклеотидов, такую, как ДНК или РНК последовательность, и может представлять собой как двухцепочечную, так и одноцепочечную нуклеиновую кислоту. Термин полинуклеотид объединяет геномную ДНК, кДНК и мРНК.
Согласно специфическому варианту осуществления изобретения, полинуклеотид, по крайней мере, на 30, 35, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 65, 70, 71, 72, 73, 74, 74, 76, 77, 78, 78, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99,5, 99,8 или 99,9% идентичен любой из нуклеотидных последовательностей SEQ ID NO:2 или SEQ ID NO:4 или их фрагментам.
Согласно одному специфическому варианту осуществления изобретения, полинуклеотид имеет последовательность, представленную в любой из последовательностей SEQ ID NO:2 или SEQ ID NO:4.
Аминокислоты в положении 276 (Ambler) у бета-лактамаз класса А
Аспарагин (Asn, N) в положении 276 присутствует у большого количества разнообразных бета-лактамаз класса А. Функция Asn276 интенсивно изучалась у ТЕМ и SHV бета-лактамаз, у которых Asn276 формирует углеводные связи с гуанидиновой группой аргинина (Arg, R) 244 и, таким образом, ограничивает мобильность Arg244 боковой цепи.
Было обнаружено, что замещение аспарагина (Asn, N) в ТЕМ или SHV ферментах играет ключевую роль в устойчивости к ингибиторам сериновых бета-лактамаз, таких, как клавулановая кислота, сульбактам или тазобактам. N276D (Asp) замещенные варианты ТЕМ-1 бета-лактамазы присутствуют у устойчивых к ингибиторам бета-лактамаз (IRT ферменты, такие, как ТЕМ-35 и ТЕМ-36). Вариант N276D является более устойчивым к клавулановой кислоте и тазобактаму, чем фермент дикого типа ТЕМ-1, однако одновременно каталитическая эффективность (Kcat/Km) варианта N276D по отношению к различным пенициллинам на 50% меньше, чем у фермента дикого типа ТЕМ-1. Каталитическая эффективность варианта N276D по отношению к цефалоспоринам уменьшается по сравнению с эффективностью фермента дикого типа ТЕМ-1 (Saves I et al., 1995, J Biol Chem. 270:18240-18245).
Аналогично ТЕМ-1, замещение N276D в SHV-1 или SHV-5 бета-лактамазах увеличивает устойчивость к ингибиторам сериновых бета-лактамаз, но уменьшает их гидролитическую эффективность по отношению к большинству бета-лактамов (Giakkoupi P. et al., 1999, J Antimicrobiol Chemother, 43: 23-29). Кроме того, замещение N276D в SHV-1 или SHV-5 ферментах умеренно повышает их способность разрушать "четвертое поколение" цефалоспоринов - цефпиром и цефепим.
Для бета-лактамаз типа SHV - ОНЮ-1, было показано, что мутант N276G (Gly) является высокоустойчивым к клавулановой кислоте, при этом мутант N276G на основе ТЕМ-1 бета-лактамаз обладает умеренной устойчивостью к клавулановой кислоте (Bonomo RA et al., 1995, Biochim Biophys Acta. 1247:121-125).
В семействе СТХ-М ферментов аргинин (Arg, R) обычно находится в положении 276 (Bonnet R., 2004, Antimicrob Agents Chemother, 48: 1-14), а мутации Arg276 влияют на продолжительность ферментативной активности. Относительные скорости гидролиза цефотаксима ферментами СТХ-М умеренно снижаются в результате замещения Arg276. Кроме того, мутации Arg276Trp, Arg276Cys, Arg276Ser и Arg276Gly в СТХ-М ферментах не влияют на уровень устойчивости к ингибиторам бета-лактамаз (Bonnet R., 2004, Antimicrob Agents Chemother, 48: 1-14; Perez-Llarena F.J. et al., 2008, J Antimicrobiol Chemother, 61: 792-797).
В настоящем изобретении бета-лактамазы, имеющие аминокислотную последовательность, которая, по крайней мере, на 60% идентична последовательности SEQ ID NO:1 (Bacillus licheniformis PenP производное, то есть Р1А производное), и включающие гидрофильную аминокислоту в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, демонстрируют расширенный бета-лактамный спектр, а также улучшенную каталитическую активность в отношении бета-лактамов.
До настоящего времени влияние аминокислотных замен, таких, как замещение аспарагиновой кислоты (D) в положении 276, на устойчивость к ингибиторам сериновых бета-лактамаз среди бета-лактамаз Bacillus spp., в особенности, бета-лактамазы Б. licheniformis, не изучалась.
Как описано здесь аминокислотный остаток 276, согласно классификации Ambler, соответствует положению 243 в SEQ ID NO:1 и положению 249 в SEQ ID NO:3.
Обычно бета-лактамазы, заявленные в настоящем изобретении, имеют остаток гидрофильной аминокислоты в положении, соответствующем положению 276, согласно классификации Ambler, при этом указанная аминокислота не является аспарагиновой кислотой (D). Аминокислоты классифицируются на основании химических и/или структурных свойств их боковых цепей. Классификационные группы аминокислот включают гидрофильные аминокислоты, которые в свою очередь подразделяются на следующие подгруппы: полярные и положительно заряженные гидрофильные аминокислоты; полярные и нейтрально заряженные гидрофильные аминокислоты; полярные и отрицательно заряженные гидрофильные аминокислоты; ароматические, полярные и положительно заряженные гидрофильные аминокислоты. Используемый здесь термин "гидрофильная аминокислота" включает все описанные выше подгруппы гидрофильных аминокислот, то есть относится к полярным и положительно заряженным гидрофильным аминокислотам, к полярным и нейтрально заряженным гидрофильным аминокислотам, к полярным и отрицательно заряженным гидрофильным аминокислотам и/или к ароматическим, полярным и положительно заряженным гидрофильным аминокислотам (http://www.biomed.curtin.edu.au/biochem/tutorials/AAs/AA.html). "Полярные и положительно заряженные гидрофильные аминокислоты" включают аргинин (R) или лизин (K). "Полярные и нейтрально заряженные гидрофильные аминокислоты" включают аспарагин (N), глутамин (Q), серии (S) или треонин (Т). "Полярные и отрицательно заряженные гидрофильные аминокислоты" включают аспарагиновую кислоту (D) или глутаминовую кислоту (Е). "Ароматические, полярные и положительно заряженные гидрофильные аминокислоты" включают гистидин (Н).
Согласно одному варианту осуществления изобретения, гидрофильная аминокислота представляет собой нейтральную или положительно заряженную гидрофильную аминокислоту, выбранную из группы, включающей аргинин (R), гистидин (Н), лизин (K), аспарагин (N), глутамин (Q), серии (S) и треонин (Т), которая находится в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler.
Согласно предпочтительному варианту осуществления изобретения, гидрофильная аминокислота бета-лактамазы в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, выбрана из группы полярных и положительно заряженных гидрофильных аминокислот, включающей аргинин (R), гистидин (Н) и лизин (K). Более предпочтительно, аминокислота в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, представляет собой аргинин.
Согласно другому предпочтительному варианту осуществления изобретения, гидрофильная аминокислота выбрана из группы полярных и нейтрально заряженных гидрофильных аминокислот, включающей аспарагин (N), глутамин (Q), серии (S) и треонин (Т). Более предпочтительно, когда аминокислотой в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, является аспарагином.
Согласно еще одному предпочтительному варианту осуществления изобретения, гидрофильная аминокислота SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, локализуется в альфа-спирали. Альфа-спираль представляет собой элемент вторичной белковой структуры, напоминающий скрученную конформацию. Альфа-спирали могут иметь особую значимость в формировании элементов связывания ДНК (например, спираль-петля-спираль, лейциновые застежки-молнии и цинковые пальцы). Согласно предпочтительному варианту осуществления изобретения, аминокислотный остаток 276 локализуется в конечной альфа спиралип (Фиг.1). Указанная альфа-спирали11 не является консервативной для бета-лактамаз класса А.
Специфические свойства бета-лактамаз класса А
Одной специфической особенностью бета-лактамаз класса А является гуанидиновая группа Arg276. CTX-M ферменты включают Arg278, Arg244 или Arg220, которые находятся в эквивалентных положениях в трехмерной пространственной структуре. Было показано, что аргинин в положении 220 или 244 является необходимым для каталитической активности ТЕМ-1 (Leu220 и Arg244) ферментов и бета-лактамазы Streptococcus albus G (Arg220 Asn 244). Предполагается, что основная гуанидиновая группа аргинина 244 или аргинина 220 участвует в связывании бета-лактама или в химической инактивации "суицидальных" ингибиторов, таких, как клавулановая кислота (Matagne et al., 1998, Biochem J. 330:582-598; Perez-Llarena et al., 2008, J Antimicrobiol Chemother, 61:792-797). В ферменте PenP Б. licheniformis остаток Arg244 формирует солевой мостик с аспарагиновой кислотой 276 (Herzberg, О. 1991, J Mol Biol. 217:707-719; Knox, J.R., и P.C. Moews, 1991, J Mol Biol. 220:435-555).
Согласно предпочтительному варианту осуществления изобретения, бета-лактамаза дополнительно содержит, по крайней мере, одну аминокислоту, выбранную из группы, включающей Leu220 и Arg244, согласно классификации Ambler, что соответствует Leu189 и Arg212 SEQ ID NO:1.
Бета-лактамаза Bacillus licheniformis (PenP, P1A)
Бета-лактамаза, заявленная в соответствии с настоящим изобретением, секретируется штаммом 749/С Bacillus licheniformis. Бета-лактамаза В. licheniformis 749/С (PenP; пенициллин-амидо-бета-лактамгидролаза, ЕСЗ.5.2.6.) принадлежит к подгруппе 2а класса А бета-лактамаз, согласно функциональной классификации (Bush К. et al., 1995, Antimicrob Agents Chemother 39: 1211-1233). Бета-лактамаза В. licheniformis может рассматриваться как пенициллиназа, которая обладает высокой гидролитической способностью в отношении пенициллина, ампициллина, амоксициллина или пиперациллина, и полностью ингибируется активными сайт-направленными ингибиторами бета-лактамаз, такими, как клавулановая кислота, сульбактам или тазобактам (Bush K et al., 1995, Antimicrob Agents Chemother. 39:1211-1233).
Бета-лактамаза В. licheniformis 749/С экспрессируется как пребелок, включающий 307 аминокислотных остатков. После транслокации и удаления 26-ти аминокислотных остатков длинной сигнальной последовательности он трансформируется в заякоренный в мембране липопротеин, в котором цистеин (С27) на амино-конце формирует тиоэфирную связь с диацилглусеридом. Также были обнаружены секретируемые (внеклеточные) формы бета-лактамазы В. licheniformis, которые представляют собой протеолитические продукты липопротеиновой формы (Azui К. et al., 1980, Bicohemistry 19: 1882-1886; Matagne A et al., 1991, Biochem J, 273: 503-510). Участок гена бета-лактамазы В. licheniformis 749/С, кодирующий небольшую секретируемую форму (малая экзоформа; Р1А) из аминокислотных остатков 43-307, был выбран в качестве ДНК фрагмента для конструирования продукционной системы хозяйского вектора Bacillus subtilis (WO 2008065247).
Функции
Бета-лактамазы гидролизуют бета-лактамные антибиотики, содержащие бета-лактамное кольцо, такие, как пенициллины, цефалоспорины, клавамы (или оксапенемы), цефамицины и карбапенемы. Согласно предпочтительному варианту осуществления изобретения, бета-лактамаза гидролизует пенициллины и/или цефалоспорины. Термин "пенициллины" относится к нескольким естественным или синтетическим вариантам пенициллина, которые обычно получают из организма Penicillinum. К пенициллинами относятся, без ограничений указанными, амоксициллин, ампициллин, азлоциллин, карбенициллин, клоксациллин, диклоксациллин, флуклоксациллин, гетациллин, оксациллин, мезлоциллин, пенициллин G, пенициллин V и пиперациллин.
В случае цефалоспоринов бета-лактамное кольцо соединено с шестичленным дигидротиазиновым кольцом, а также с пятичленным тиазолидиновым кольцом, которое есть и у пенициллинов. На основании их биологической активности цефалоспорины подразделяют на шесть поколений, однако, некоторые цефалоспорины не относятся к определенному поколению. Согласно одному специфическому варианту осуществления изобретения, бета-лактамаза обладает улучшенной каталитической эффективность в отношении цефалоспоринов по сравнению с бета-лактамазами дикого типа. Согласно настоящему изобретению, бета-лактамаза Bacillus licheniformis, в которой аспарагиновая кислота (Asp, D) в положении 276, пронумерованном в соответствии с классификацией Ambler, замещена гидрофильной аминокислотой, такой, как аспарагин (N) или аргинин (R), демонстрирует более продолжительную активность в отношении бета-лактамных антибиотиков, таких, как цефалоспорины.
Согласно одному варианту осуществления изобретения, цефалоспорины выбраны из группы, включающей цефоперазон, цефтриаксон и цефазолин.
Используемый здесь термин "каталитическая эффективность" бета-лактамаз относится к способности гидролизировать бета-лактамные антибиотики. Улучшенная каталитическая эффективность может измеряться при помощи любого традиционного in vitro, ex vivo или in-vivo способа в любом биологическом образце и у любого субъекта.
Способы получения и модификации бета-лактамаз
Бета-лактамаза, заявленная в соответствии с настоящим изобретением, может быть получена путем модификации фермента при помощи любого стандартного способа генной инженерии. Для этого могут применяться такие способы, как рациональное конструирование, выборочный мутагенез, перетасовка ДНК (выборочная рекомбинация), фаговый дисплей, перетасовка целого генома, гетеродуплекс, выборочный химерагенез при транзиентной сборке матриц нужных олигонуклеотидов, мутагенная и непрямая сборка, перестановка экзонов, перестановка блоков, основанная на Y -лигировании, негомологичная рекомбинация, сочетание рационального конструирования и прямой эволюции. Кроме того, мутантные ферменты могут быть получены при помощи сайт-направленного мутагенеза и сплайсинга при использовании технологий перекрывающегося удлинения.
Согласно одному варианту осуществления изобретения, способ модификации бета-лактамазы включает этап модификации бета-лактамазы, имеющей аминокислотную последовательность, которая, по крайней мере, на 60% идентична SEQ ID NO:1, включающий замещение аминокислоты в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler, гидрофильной аминокислотой. Гидрофильной аминокислотой может быть любая гидрофильная аминокислота, например, выбранная из группы, включающей аргинин (R), гистидин (Н), лизин (K)," аспарагин (N), глутамин (Q), серии (S) и треонин (Т).
Согласно одному варианту осуществления изобретения, негидрофильная аминокислота замещается гидрофильной аминокислотой в положении SEQ ID NO:1, соответствующем положению 276, согласно классификации Ambler.
Бета-лактамаза, заявленная в соответствии с настоящим изобретением, также может быть получена, например, при помощи синтетических способов, скажем, пептидного синтеза или рекомбинантной продукции в клетке-хозяине. Согласно предпочтительному варианту осуществления изобретения, фермент является рекомбинантным. Используемый здесь термин "рекомбинантный генетический материал" относится к материалу, который обычно представляет собой комбинацию генетического материала, например, ДНК- цепей различного происхождения, и получен путем объединения или вставки последовательностей. Полинуклеотид, заявленный в соответствии с настоящим изобретением, может быть, например, вставлен под контролем любых эндогенных или экзогенных регуляторов, таких, как промоторы. Рекомбинантный белок получают на основании рекомбинантной ДНК.
По крайней мере, один полинуклеотид или полинуклеотидный фрагмент, представляющий интерес, может быть выделен из клетки или получен синтетическим путем. Указанный полинуклеотид или полинуклеотидный фрагмент может быть трансфомирован в клетку-хозяин. Подходящей клеткой-хозяином для получения любого пептида, заявленного в соответствии с настоящим изобретением, может быть любая прокариотическая или эукариотическая клетка, предпочтительно, бактерия, наиболее предпочтительно, Bacillus spp., например, штамм Bacillus subtilis, Bacillus licheniformis, Bacillus pumilis или Bacillus amyloliquefaciens.
Используемый здесь термин "трансформация" относится к генетическому изменению клетки чужеродным генетическим материалом, предпочтительно, ДНК, что приводит к экспрессии указанного генетического материала. Чужеродный генетический материал может быть введен в клетку сам по себе или быть включенным в другой генетический материал, такой, как вектор, плазмида и т.д. Любой способ генной инженерии или любой способ молекулярного клонирования может использоваться для трансформации клетки-хозяина полинуклеотидом, заявленным в соответствии с настоящим изобретением. Существуют различные способы введения чужеродного материала в эукариотическую клетку. Материалы, такие, как полимеры (например, DEAE-декстран или полиэтиленимин), липосомы и наночастицы (например, золото) использовались в качестве носителей для трансформации. Генетический материал также может вводиться в клетки при помощи, например, вирусов или векторов, которые используют в качестве носителей. Другие способы введения чужеродного материала в клетки включают, без ограничений указанными, нуклеофекцию, электропорацию, конъюгацию, трансфекцию, сонопорацию, тепловой шок и магнетофекцию.
После того, как клетка-хозяин продуцирует пептид, заявленный в соответствии с настоящим изобретением, в подходящих условиях, нужный пептид может быть очищен из клетки, либо секретируемая форма может быть восстановлена, например, из культуральной среды; согласно предпочтительному варианту осуществления изобретения, бета-лактамаза секретируется клеткой.
Фармацевтическая композиция
Фармацевтическая композиция, заявленная в соответствии с настоящим изобретением, включает заявленную бета-лактамазу. Композиция также может содержать одну бета-лактамазу или более, например, по крайней мере, две, три, четыре и т.д. различные бета-лактамазы.
Фармацевтические композиции, заявленные в соответствии с настоящим изобретением, также могут содержать другие активные ингредиенты, помимо бета-лактамазы, заявленной в соответствии с настоящим изобретением.
Фармацевтические композиции могут использоваться, например, в твердой, полутвердой или жидкой форме, а именно, в форме таблеток, пеллет, капсул, растворов, эмульсий или суспензий. Предпочтительно, композиции предназначены для перорального введения или для энтерального введения.
Помимо, по крайней мере, одной бета-лактамазы, заявленной в настоящем изобретении, или полинуклеотида, или клетки-хозяина, включающей заявленные полинуклеотиды, фармацевтическая композиция может включать фармацевтически доступные носители, адъюванты, эксципиенты, вспомогательные эксципиенты, антисептики, стабилизирующие агенты, связывающие агенты, наполняющие агенты, любриканты, суспендирующие агенты, пластификаторы, красители, пленкообразователи, сахар, спирты, глиданты и растворители и/или компоненты, которые обычно можно обнаружить в соответствующих продуктах.
Продукт или фармацевтическая композиция, заявленная в соответствии с настоящим изобретением, содержит бета-лактамазы в количестве, достаточном для достижения желаемого эффекта.
Продукты или фармацевтические композиции могут быть произведены при помощи любого стандартного способа, известного из области техники. Бета-лактамазы могут быть добавлены к любому фармацевтическому продукту или смешаны с любыми агентами на любом этапе производства. Бета-лактамаза, заявленная в соответствии с настоящим изобретением, может быть также получена, например, путем экспрессии гена бета-лактамазы в подходящих условиях в фармацевтическом продукте или в ткани-мишени после разрушения фармацевтического продукта.
Согласно одному предпочтительному варианту осуществления изобретения, бета-лактамаза(ы) и бета-лактамный антибиотик вводятся субъекту одновременно в форме пеллета, покрытого оболочкой, которая растворяется в кишечнике. Покрытия на водной основе считаются наиболее подходящими материалами для создания покрытий для гидрофильного Р1А белка. Водные полимеры, обычно используемые для создания растворимых в кишечнике оболочек и также используемые в соответствии с настоящим изобретением, представляют собой полиметакрилаты, такие, как Eudragit®, полимеры на основе целлюлозы, например, эфиры целлюлозы, например, Duodcell®, или сложные эфиры целлюлозы, например, Aquateric, или сополимеры поливинилацетата, например, Opadry®.
Бета-лактамаза, заявленная в соответствии с настоящим изобретением, или фармацевтическая композиция, заявленная в соответствии с настоящим изобретением, может вводиться субъекту одновременно с бета-лактамным антибиотиком или отдельно от него. Согласно одному варианту осуществления изобретения, бета-лактамаза или фармацевтическая композиция вводится перед введением бета-лактамного антибиотика, например, за 5-30 минут до введения бета-лактамного антибиотика. Бета-лактамаза и бета-лактамный антибиотик/антибиотики могут входить в состав одной лекарственной формы или в состав различных лекарственных форм.
Побочные эффекты бета-лактамных антибиотиков и их лечение
Побочные эффекты, то есть нежелательные лекарственные реакции на введение бета-лактамных антибиотиков, могут включать, без ограничений указанными, диарею, тошноту, сыпь, крапивницу, суперинфекцию, лихорадку, рвоту, эритему, дерматит, ангионевротический отек и псевдомембранозный колит.
Согласно предпочтительному варианту осуществления, настоящее изобретение предназначено для лечения или профилактики побочных эффектов, возникающих в желудочно-кишечном тракте (ЖКТ). Используемый здесь термин "желудочно-кишечный тракт" относится к органам пищеварения, начиная с ротовой полости и заканчивая анальным отверстием. Желудочно-кишечный тракт включает ротовую полость, пищевод, желудок, двенадцатиперстную кишку, тощую кишку, подвздошную кишку, тонкую кишку, толстую кишку, слепую кишку, прямую кишку и анальное отверстие.
Бета-лактамаза, заявленная в соответствии с настоящим изобретением, или фармацевтическая композиция, заявленная в соответствии с настоящим изобретением, могут вводиться субъекту перорально или непосредственно в желудочно-кишечный тракт. Лекарственный продукт, содержащий ферментную комбинацию, предназначен для инактивации неабсорбированного бета-лактамного антибиотика в ЖКТ или в других частях организма, например, в коже или вагине. Фармацевтическая композиция может представлять собой лекарственный продукт для перорального введения или входить в состав дерматологической лекарственной формы или вагинального суппозитория, при этом она может входить в состав жидких лекарственных форм, а также лекарственных форм с немедленным, отсроченным или замедленным высвобождением.
Согласно одному предпочтительному варианту осуществления изобретения, бета-лактамаза(ы) вводятся перорально. Согласно другому предпочтительному варианту осуществления изобретения, бета-лактамаза(ы) вводятся непосредственно в желудочно-кишечный тракт пациента.
Субъектом, который подвергается лечению, может быть человек или животное, такое, как домашний питомец или сельскохозяйственное животное, например, собака, кошка, корова, свинья, курица или лошадь. Согласно предпочтительному варианту осуществления изобретения, субъектом является человек.
Настоящее изобретение иллюстрируется представленными ниже примерами, которые никоим образом не ограничивают его сущности.
Пример 1. Конструирование мутантных ферментов D276N и D276R
Мутанты бета-лактамазы Bacillus licheniformis D276N и D276R конструировали при помощи мутагенеза сплайсингом с перекрывающимся удлинением (SOE, от англ. Splicing-by-Overlap Extension) с использованием плазмиды pRSI-11 0, кодирующей Р1А бета-лактамазу, в качестве матрицы для первичных ПЦР- реакций в соответствии с ранее опубликованными методиками (Horton R.M. et al., 1989, Gene 77:61-68). Праймеры конструировали таким образом, чтобы они обеспечивали получение двух различных ПЦР-продуктов с участком общей последовательности. Затем фрагменты сливали в последовательной ПЦР амплификации за счет добавления перекрывающихся участков. Желаемые мутации получали при помощи мутагенных праймеров в первичной ПЦР.
Для получения мутанта D276N осуществляли мутацию в желаемом положении гена дикого типа, что приводит к замещению кодона GAT кодоном ААТ. Праймеры, которые использовались для первичных ПЦР- амплификации, представлены в Таблице 2. Размер амплифицированных фрагментов в первой ПЦР составлял 800 нуклеотидов и 220 нуклеотидов, которые имели длинный перекрывающийся участок из 21 нуклеотида.
Во второй ПЦР реакции (SOE реакция), два перекрывающихся фрагмента сливали вместе в последовательной реакции удлинения. Включение внешних праймеров (Прямой-1 и Reverse-1) в реакции удлинения амплифицирует составной продукт при помощи ПЦР. Очищенный SOE продукт расщепляли рестрикционным ферментом HindIII и лигировали с HindIII расщепленным рКТН141 вектором секреции, как описано в WO 2008/065247.
Компетентные клетки Bacillus subtilis RS303 трансформировали лигационной смесью. Позитивные клоны на планшетах Лурия-канамицин отбирали путем суспендирования бактериальной массы единичной колонии в растворе нитроцефина. Позитивные клоны эффективно гидролизовали нитроцефин, что проявлялось изменением цвета раствора нитроцефина с желтого на красный. Гибридную плазмиду очищали из клеток единичного клона. Правильную последовательность полученного при помощи ПЦР участка подтверждали при ДНК-секвенированием.
Для получения мутанта D276R, мутацию осуществляли в желаемом положении, что приводило к замещению кодона GAT кодоном CGC. Конструирование мутантного штамма D276R осуществляли аналогично мутанту D276N, с той разницей, что для первичной ПЦР использовали обратный праймер-D276R и прямой праймер-D276K (см. Таблицу 2).
Пример 2. Нуклеотидная последовательность мутантного гена бета-лактамазы D276N (penP)
Экспрессирующий конструкт выделяли из позитивного клона и вставку подвергали ДНК-секвенированию. Анализ полной нуклеотидной последовательности и предсказанной аминокислотной последовательности мутантного гена бета-лактамазы D276N показал, что замещение Asp на Asn произошло корректно в нужном кодоне (Рисунок 2). Кроме того, анализ ДНК последовательности мутантного гена бета-лактамазы D276N выявил наличие в рамке слияния между нуклеотидной последовательностью, кодирующей 31 аминокислоту длинной сигнальной последовательности альфа-амилазы Bacillus amyloliquefaciens, и полноразмерной последовательностью мутантного гена D276N сайта клонирования HindIII. Предполагается, что сигнальная пептидаза будет расщеплять пептидную связь между аланином (А) в положении -1 и глутамином (G) в положении+1. Зрелая бета-лактамаза D276N имеет NH2-концевое удлинение NH2-QAS-трипептида, происходящего из сайта клонирования HindIII в экспрессирующем конструкте. Следовательно, на основании предсказанной аминокислотной последовательности можно установить, что мутантная бета-лактамаза D276N состоит из 268 аминокислотных остатков.
Пример 3. Нуклеотидная последовательность мутантного гена бета-лактамазы D276R (penP)
Для подтверждения желаемого замещения аспарагиновой кислоты в положении 276 (классификация Ambler) в гене бета-лактамазы Bacillus licheniformis, экспрессирующий конструкт выделяли из позитивного клона и нуклеотидную последовательность вставки секвенировали аналогично тому, как описано в Примере 2. Согласно полученной нуклеотидной последовательности, предсказанная аминокислотная последовательность содержит желаемое D276R замещение, а зрелый D276R фермент состоит из 268 аминокислотных остатков (Рисунок 3).
Пример 4. Биохимический анализ D276N мутантной бета-лактамазы (Р3А)
Степень чистоты ферментного препарата составляла более 95% и определялась при помощи анализа SDS-PAGE (данные не представлены).
Кинетические параметры бета-лактамазы дикого типа (Р1А) и мутантной бета-лактамазы D276N (РЗА) В. licheniformis определяли посредством осуществления гидролиза при помощи различных бета-лактамаз; данные приведены в Таблице 3. Ферментативные реакции осуществляли в 20 мМ фосфатном буфере (рН 7) при 30°C с использованием подходящей концентрации фермента и различных концентраций субстратов - пенициллина и цефалоспорина. Величины Kcat и Km получали при помощи способа линеаризации Hanes. Основные результаты приведены ниже.
(i) Пенициллины
Влияние замещения D276N на гидролиз пенициллинов (ампициллина, амоксициллина или пиперациллина) не было существенным и обеспечивало 51-80% ферментативной активности по отношению к ферменту дикого типа. Соответственно, величины Kcat и Km для мутантного фермента D276N при гидролизе пенициллина уменьшались максимально в два раза или менее.
(ii) Цефалоспорины
Как и ожидалось, с учетом результатов, полученных для пенициллинов, бета-лактамаза дикого типа обладала слабой ферментативной эффективностью по отношению к различным цефалоспоринам, включая первое (кафазолин), второе (цефуроксим) и третье(цефтриаксон, цефотаксим, цефтадизим, цефоперазон и цефепим) поколение цефалоспоринов (Таблица 1). Удивительно, что ферментативная эффективность мутантного D276N фермента по отношению к определенным цефалоспоринам, предпочтительно, к цефоперазону и, более предпочтительно, к цефтриаксону, существенно увеличивались по сравнению с ферментативной эффективностью ферментов дикого типа. Константа Km для цефтриаксона и цефоперазона снижалась, а соответствующая обратная величина Kcat для цефтриаксона и цефоперазона увеличивалась по сравнению с величинами для фермента дикого типа (Р1А). Таким образом, замещение аспарагиновой кислоты аспарагином в положении 276 в бета-лактамазе Bacillus licheniformis способствует расширению субстратного профиля бета-лактамазы Bacillus licheniformis.
Пример 5. Биохимическая характеристика D276R мутантного фермента
D276N мутантный фермент конструировали для того, чтобы оценить насколько допустимо замещение Asp276 и насколько замещение D276R способствует увеличению активности бета-лактамазы, характерной для фермента D276N.
Сырые образцы D276R и D276N, выделенные из культурального супернатанта, использовали в качестве тестируемого материала. Степень чистоты и количество ферментных образцов оценивали при помощи анализа SDS-PAGE. Скорость гидролиза мутантных ферментов D276R и D276N для различных бета-лактамов оценивали при помощи величин Vmax. Полученные результаты представлены как относительные величины (%) по сравнению с величинами для фермента D276N и приведены в Таблице 4.
В целом, каталитическая эффективность D276R бета-лактамазы по отношению к пенициллинам и цефалоспоринам сопоставима с эффективностью фермента D276N. По сравнению с ферментом D276N, фермент D276R обладал меньшей активностью по отношению к цефтриаксону, но увеличенной активностью по отношению к цефоперазону. Это исследование показало, что расширение спектра бета-лактамаз может достигаться замещением гидрофильной аминокислотой, такой, как аргинин или аспарагин, аспарагиновой кислоты в положении 276 в бета-лактамазе Bacillus licheniformis. Исследование также показало, что желаемая модификация фермента может достигаться также за счет замещения другого гидрофильного аминокислотного остатка, такого, как глутамин (Q), лизин (K), серии (S) или треонин (Т) аспарагиновой кислотой в положении 276.
Пример 6. In vivo исследование бета-лактамазы D276N
Способность мутантного фермента D276N Bacillus licheniformis инактивировать цефтриаксон (CRO), который экскретировался в желудочно-кишечный тракт во время парентерального лечения, изучали на собаках. Лабораторные собаки породы бигль имели сосочковый клапан, вшитый в тощую кишку приблизительно 170 см дистальнее привратника желудка, через который собирали образцы на анализ. Хирургическое вмешательство не влияет на моторику кишечника. В каждом эксперименте участвовало пять собак породы бигль.
Исследование включало два последовательных этапа лечения: на первом этапе (контрольный эксперимент без терапии бета-лактамазами) животным внутривенно через 20 минут после первого кормления вводили единичную дозу цефтриаксона (30 мг цефтриаксона (CRO) на килограмм массы тела, что соответствует человеческой дозе в 1 грамм). Образцы содержимого тощей кишки собирали в различное время в течение 10 часов. Затем собак через пять часов сорок пять минут после введения цефтриаксона повторно кормили для того, чтобы индуцировать экскрецию цефтриаксона, накопившегося в желчном пузыре, с желчью.
Образцы химуса из тощей кишки немедленно замораживали и хранили при температуре -20°C до начала анализа. Образцы химуса предварительно обрабатывали перхлор-лимонной кислотой для преципитации интерферирующих частиц. Преципитат удаляли при помощи центрифугирования. Для количественного определения цефтриаксона в супернатантах использовали обращено-фазовую хроматографию высокого давления с УФ-регистрацией.
На втором этапе за 10 минут до введения цефтриаксона животным давали мутантную D276N бета-лактамазу в форме пеллет с растворимой в кишечнике оболочкой в составе твердых желатиновых капсул. Лекарственные формы с указанной оболочкой являются распространенной формой для перорального введения в фармацевтической индустрии. Лекарственные формы с кишечнорастворимой оболочкой разработаны таким образом, чтобы они в интактной форме проходили через желудок и высвобождение содержимого происходило только в тонком кишечнике. Твердые лекарственные формы с кишечнорастворимой оболочкой предназначены для защиты лекарственного агента от деструктивного воздействия ферментов и низких величин pH среды желудка, а также для предотвращения обусловленного лекарственным средством раздражения слизистой оболочки желудка, тошноты и кровотечения; кроме того, такие формы позволяют доставить лекарственное средство в нерастворимой форме в нужное место действия в тонком кишечнике. На основании указанных критериев лекарственные формы с кишечнорастворимой оболочкой относятся к лекарственным формам с отсроченным действием. Сополимер полиметакриловой кислоты Eudragit® L 30 D-35 применяли для создания рН-зависимой лекарственной формы с кишечнорастворимой оболочкой. На втором этапе использовали кишечнорастворимые пеллеты с единичной дозой 0,44 мг активной D276N бета-лактамазы на килограмм массы тела.
Полученные результаты после двух этапов лечения представлены на Рисунке 4. Первый этап лечения показал, что цефтриаксон в высоких концентрациях экскретировался в тонкий кишечник во время парентеральной терапии. Наибольшая концентрация цефтриаксона в тощей кишке (приблизительно 1500 микрограмм на грамм кишечного химуса) наблюдалась через 60 минут после введения цефтриаксона. Повышенная концентрация цефтриаксона в химусе тощей кишки наблюдалась и после второго кормления собак (временная точка 340 минут), что указывает на индуцированное приемом пищи накопление цетфтриаксона в желчном пузыре.
Этап 2 показал, что пероральное введение D276N мутантной бета-лактамазы способно снизить концентрацию цефтриаксона в тощей кишке до уровня ниже определяемого (10 микрограмм цефтриаксона на микрограмм кишечного химуса). Полученные результаты показывают, что мутантная D276N бета-лактамаза является потенциальным кандидатом для создания лекарственного средства, позволяющего уменьшить побочные эффекты, связанные с парентеральным введением цефтриаксона. Кроме того, с учетом высокой активности в отношении пенициллинов, такого, как ампициллин, амоксициллин и пиперациллин, мутантные ферменты D276N или D276R могут использоваться в качестве альтернативных лекарственных средств для бета-лактамазаной терапии, описанной в WO 2008065247.


Изобретение относится к биотехнологии и представляет собой бета-лактамазу, включающую аминокислотную последовательность, которая, по крайней мере, на 90% идентична SEQ ID NO: 1. При этом бета-лактамаза обладает бета-лактамазной активностью против пенициллина и цефалоспорина, выбранного из цефтриаксона и цефоперазона, и включает гидрофильный аминокислотный остаток, любой кроме аспарагиновой кислоты (D), в положении, соответствующем положению 276, согласно классификации Ambler, при этом гидрофильный аминокислотный остаток выбран из аргинина (R) и аспарагина (N). Изобретение относится также к способу получения бета-лактамазы, включающему стадии трансформации бактериальной клетки-хозяина геном, кодирующим бета-лактамазу, и выделения бета-лактамазы. Изобретение касается также фармацевтической композиции для применения при лечении или профилактике вызванных бета-лактамными антибиотиками неблагоприятных явлений в желудочно-кишечном тракте, причем фармацевтическая композиция включает бета-лактамазу и фармацевтически приемлемый носитель. Изобретение позволяет получить новую эффективную бета-лактамазу. 7 н. и 10 з.п. ф-лы, 4 ил., 4 табл., 6 пр.
1. Бета-лактамаза, включающая аминокислотную последовательность, которая, по крайней мере, на 90% идентична SEQ ID NO: 1, отличающаяся тем, что бета-лактамаза обладает бета-лактамазной активностью против пенициллина и цефалоспорина, выбранного из цефтриаксона и цефоперазона, и включает гидрофильный аминокислотный остаток, любой кроме аспарагиновой кислоты (D), в положении, соответствующем положению 276, согласно классификации Ambler, при этом гидрофильный аминокислотный остаток выбран из аргинина (R) и аспарагина (N).
2. Бета-лактамаза по п. 1, отличающаяся тем, что аминокислота в положении SEQ ID NO: 1, соответствующем положению 276, представляет собой аспарагин (N).
3. Бета-лактамаза по п. 1, отличающаяся тем, что аминокислота в положении SEQ ID NO: 1, соответствующем положению 276, представляет собой аргинин (R).
4. Бета-лактамаза по п. 1, отличающаяся тем, что гидрофильная аминокислота в положении SEQ ID NO: 1, соответствующем положению 276, локализована в альфа-спирали.
5. Бета-лактамаза по п. 1, отличающаяся тем, что бета-лактамаза дополнительно включает, по крайней мере, одну аминокислоту, выбранную из группы, включающей Leu220 и Arg244, согласно классификации Ambler.
6. Бета-лактамаза по п. 1, отличающаяся тем, что бета-лактамаза обладает улучшенной каталитической эффективностью по отношению к цефалоспоринам по сравнению с бета-лактамазой с последовательностью SEQ ID NO: 1.
7. Бета-лактамаза по п. 6, отличающаяся тем, что цефалоспорины выбраны из группы, включающей цефоперазон и цефтриаксон.
8. Бета-лактамаза по п. 1 в качестве лекарственного средства.
9. Способ получения бета-лактамазы по пп. 1-7, отличающийся тем, что включает следующие стадии:
а) трансформацию бактериальной клетки-хозяина геном, кодирующим бета-лактамазу по любому из пп. 1-7;
б) выделение бета-лактамазы.
10. Способ модификации бета-лактамазы, включающей аминокислотную последовательность, которая по крайней мере на 90% идентична SEQ ID NO: 1, для бета-лактамазной активности против пенициллина и цефалоспорина, выбранного из цефтриаксона и цефоперазона, отличающийся тем, что аминокислота бета-лактамазы в положении SEQ ID NO: 1, соответствующем положению 276, согласно классификации Ambler, замещена гидрофильной аминокислотой, кроме аспарагиновой кислоты (D), при этом указанный гидрофильный аминокислотный остаток выбран из аргинина (R) и аспарагина (N).
11. Полинуклеотид, отличающийся тем, что он кодирует бета-лактамазу по пп. 1-7.
12. Бактериальная клетка-хозяин, содержащая полинуклеотид по п. 10 для получения бета-лактамазы по п. 1.
13. Фармацевтическая композиция для применения при лечении или профилактике вызванных бета-лактамными антибиотиками неблагоприятных явлений в желудочно-кишечном тракте, причем фармацевтическая композиция включает бета-лактамазу по любому из пп. 1-7 и фармацевтически приемлемый носитель.
14. Фармацевтическая композиция по п. 13, приемлемая для введения пациенту последовательно или одновременно с бета-лактамным антибиотиком.
15. Применение бета-лактамазы по п. 1 для производства лекарственного средства, предназначенного для лечения или профилактики побочных эффектов, обусловленных приемом бета-лактамных антибиотиков, в желудочно-кишечном тракте.
16. Применение по п. 15, отличающееся тем, что бета-лактамазу(ы) вводят перорально.
17. Применение по п. 15, отличающийся тем, что бета-лактамазу(ы) вводят непосредственно в желудочно-кишечный тракт пациента.
| Устройство для разработки мерзлых грунтов | 1988 |
|
SU1564286A1 |
| US 20050261270 A1, 24.11.2005 | |||
| WO 2007147945 A1, 27.12.2007 | |||
| MADGWICK PJ et al | |||
| Beta-lactamase I from Bacillus cereus | |||
| Structure and site-directed mutagenesis, Biochem J | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| RU 2007111958 A, 10.10.2008. | |||

