Заявляемое изобретение относится к области биодозимерии помещений и может быть применено для биодозиметрии радона, а также выявления создаваемых им с другими поллютантами жилых помещений синергических биологических эффектов.
Основным методом, используемым повсеместно для измерения радона, является определение его интегральной объемной активности (заявка РФ №95111197, G 01 T 1/167, опубл. 20.06.1997; патент РФ №2123192, G 01 T 1/167, опубл. 10.12.1998).
Все ныне существующие приборы для дозиметрии радона измеряют концентрацию радона или единомоментно (однако этот способ, несмотря на быстроту, не может дать точную оценку активности радона в помещении, т.к. его содержание может значительно изменяться в течение суток), или же угольные детекторы измеряют концентрацию радона в воздухе в течение нескольких суток. Общим недостатком всех этих детекторов является невозможность отслеживания совокупных эффектов радона и других поллютатнтов (их синергические эффекты) (заявка РФ №95111197, G 01 T 1/167, опубл. 20.06.1997; патент РФ №2123192, G 01 T 1/167, опубл. 10.12.1998) и прямой оценки вызываемых эффектов непосредственно у живых организмов. Для применяемых в настоящий момент биологических тест-систем и тест-объектов для оценки генотоксичности окружающей среды (они реагируют на возможные синергические эффекты поллютантов) отсутствуют данные по эффектам воздействия радона на них (возможно, они не чувствительны к радоновой радиации) (патент РФ №2145711, G 01 T 1/167; патент РФ №2092870, G 01 T 1/167) и не построена шкала для его количественной оценки. Обычно указанные тест-объекты дают ответ по принципу “да-нет” (есть ли эффект загрязнения или его нет). Каждый из существующих ныне тест-обектов использует, как правило, один критерий, каждый из которых обладает различной чувствительностью к стрессовым воздействиям. В настоящее время нет тест-систем, использующих несколько критериев на одном тест-объекте.
Для выяснения генотоксичности окружающей среды широкое распространение получили методы биоиндикации с использованием различных видов традесканции (Grant W.F., Lee H.G., Logan D.M. et al. The use of Tradescantia and Vicia faba bioassays for the in situ detection of mutagens in an aquatic environment // Mutat. Res. - 1992. - V. 270. - Р. 53-64). Эти исследования проводят, как правило, на волосках тычинок, которые в случае присутствия в среде мутагенов в результате генных мутаций изменяют цвет тычиночной нити (Sparrow A.H., Schairer L.A., Villalobos-Piltrini R. Comparison of somatic mutation rates induced in Tradescantia by chemical and physical mutagens. // Mutat. Res. - 1974. - V.26, №4. - Р.265-276; Nauman C.H., Sparrow A.H., Schairer L.A. Comparative effects of ionizing radiation and two gaseosus chemical mutagens on somatic mutation induced in one mutable and two non-mutable clones of Tradescantia. // Mutat. Res. - 1976. - V.38, №1. - Р.53-70.; Ichikava S., Shima N., Ishii C. et al. Variation of spontaneous somatic mutation frequency in the stamen hairs of Tradescantia clone BNL 02. // Genes & Genetic systems. - 1996. - V.71, №3. - P.159-165 и многие др.), для выявления генотоксичности некоторых соединений применяют тест на хромосомные аберрации в апикальной меристеме корней (Инге-Вечтомов С.Г. Генетика с основами селекции. - М., 1989. - С.592-594). Единственная известная нам попытка оценить полученную от радона дозу с использованием биообъектов была выполнена на фибробластах легкого (анализировали частоту сестринских хромосомных обменов) (Bauchinger M., Braselmann H., Kulka U. et al. Quantification of FISH-painted chromosome aberrations after domestic radon exposure // Int. J. Radiat. Biol. - 1996. - V.70, №6. - Р.657-663), однако метод не дал положительных результатов из-за слабой чувствительности. Работ по исследованию влияния радоновой радиации на растительные объекты в изученной нами литературе не обнаружено.
Заявляемое изобретение предназначено для оценки загрязнения радоном жилых помещений и возможных биологических эффектов от синергического взаимодействия радона и других поллютантов жилых помещений.
При его осуществлении можно провести оценку радонового загрязнения жилых помещений и его биологических эффектов от синергических взаимодействий этого благородного газа и других поллютантов, а также установить пригодность помещений для проживания людей по показателю “радоновое загрязнение”.
Вышеуказанная задача решается тем, что проводят анализ нескольких цитогенетических показателей в апикальной меристеме корня на одном биоиндикаторе, сравнивают их с контролем и созданной шкалой, отражающей изменчивость цитогенетических показателей биоиндикатора при облучении различными эквивалентными равновесными объемными активностями (ЭРОА) радона, затем определяют интегральный биологический эффект радонового загрязнения.
В качестве биоиндикатора используется Zebrina pendula Schnizl., а в качестве цитогенетических показателей - ядрышковые характеристики, митотическая активность, уровень и спектр нарушений митоза, микроядерный тест.
При их анализе ведут подсчет общего количества просмотренных клеток (не менее 400), количества делящихся клеток и из них числа клеток, имеющих патологии митоза, количества метафаз, анафаз, телофаз с остаточным ядрышком, учитывают количество клеток с 1, 2, 3... n ядрышками. У традесканции при облучении подсчитывают количество клеток с микроядрами и с остаточными ядрышками в цитоплазме интерфазных клеток. На основании полученных данных определяют: 1) митотическую активность, показателем которой является митотический индекс (MI) (%-ное отношение числа делящихся клеток к общему числу клеток), 2) количество клеток с патологиями митоза от общего числа делящихся клеток (%), 3) ядрышковую активность, под которой понимают %-ное соотношение клеток с 1, 2, 3... n ядрышками в изучаемой выборке. % клеток с n ядрышками считают как отношение клеток с n ядрышками к общему числу интерфазных клеток. Под термином “увеличение ядрышковой активности” подразумевают возрастание доли клеток с большим количеством ядрышек, а под термином “уменьшение ядрышковой активности” -возрастание количества клеток с меньшим числом ядрышек по сравнению с контролем, 4) число ядрышек на клетку, 5) % клеток с микроядрами (отношение числа клеток с микроядрами к общему числу интерфазных клеток), 6) % клеток с остаточным ядрышком в цитоплазме (отношение числа клеток с остаточным ядрышком в цитоплазме к общему числу интерфазных клеток).
Данные, полученные в опыте, сравнивают с результатами контрольного эксперимента и приведенной ниже шкалы. Сравнение результатов опыта и контроля проводят по митотической активности, ядрышковой активности по t-критерию Стьюдента; по остаточным ядрышкам, нарушениям митоза, микроядерному тесту - по Х-критерию Ван-дер-Вардена и U-критерию Вилкоксона.
Свежесрезанные побеги Zebrina pendula Schnizl. помещаются в стандартные 0,7 л банки с отфильтрованной водопроводной водой. Воду отделяют от окружающего воздуха двумя плотно подогнанными слоями полиэтиленовой пленки во избежание его попадания в воду. Банку с побегами помещают в исследуемое помещение на солнечную сторону (параллельно ставится контроль в помещении с контролируемыми условиями), температура в исследованных помещениях должна быть не ниже +20°С. По истечение 14 дней придаточные корни, которые выросли за время эксперимента, в 15 часов дня фиксируют в растворе ацетоалкоголя (3 части 96% этилового спирта:1 часть ледяной уксусной кислоты). Время нахождения материала в фиксаторе не менее 1 суток, в фиксаторе материал может храниться продолжительное время при температуре +4°С. После фиксации материал извлекается из ацетоалкоголя и из апикальной меристемы корня изготавливаются давленые микропрепараты по следующей методике:
1) для мацерации тканей кончики корешков традесканции помещают в 18%-ную соляную кислоту и подогревают (несколько раз) до температуры 60°С;
2) материал ополаскивают 45%-ной уксусной кислотой;
3) заливают красителем ацетогематоксилином корни традесканции на 25 минут при комнатной температуре;
4) отдифференцировают от избыточного красителя дистиллированной водой;
5) отрезают кончик корешка и помещают его на предметное стекло в каплю жидкости Гойера. Слегка раздавливают корешок препаравальной иглой и накрывают покровным стеклом. Препарат подогревают (но не сильно!) над пламенем горелки и раздавливают спичкой поверх покровного стекла, добиваясь равномерного распределения клеток на микропрепарате (Буторина А.К., Калаев В.Н. Анализ чувствительности различных критериев цитогенетического мониторинга. // Экология. - 2000. - №3. - С.206-210).
Изготовленный таким образом давленый микропрепарат может храниться в закрытой коробке в течение 2-3 лет.
Анализ микропрепаратов проводится на микроскопе при увеличении 10×1,5×40 и 10×1,5×100. Всего анализируется в каждом опыте не менее 7 микропрепаратов.
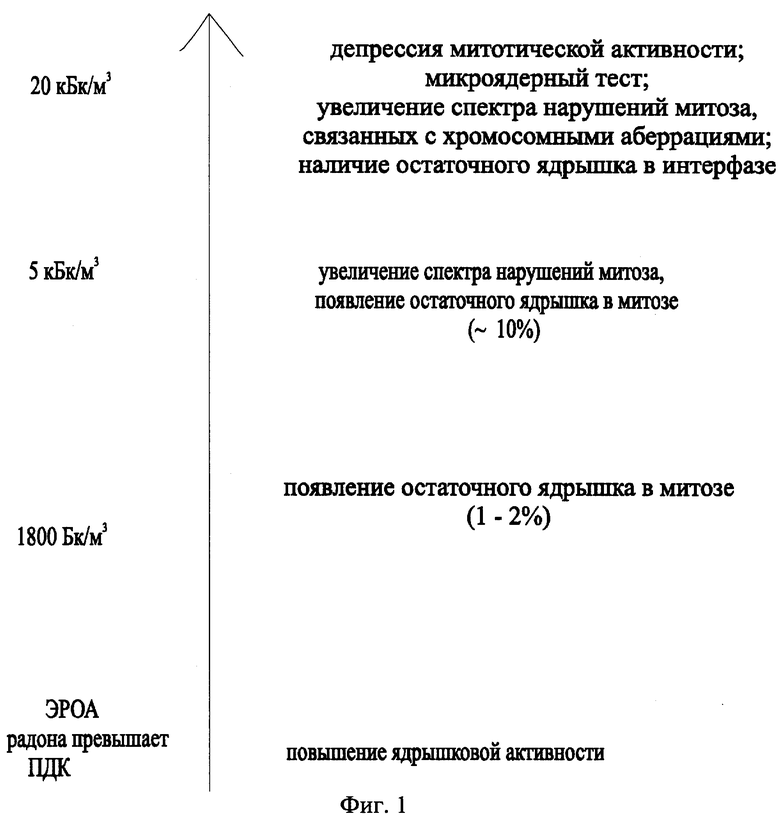
На фиг.1 представлена шкала для определения ЭРОА радона в воздухе по цитологическим аномалиям в меристеме корней Zebrina pendula.
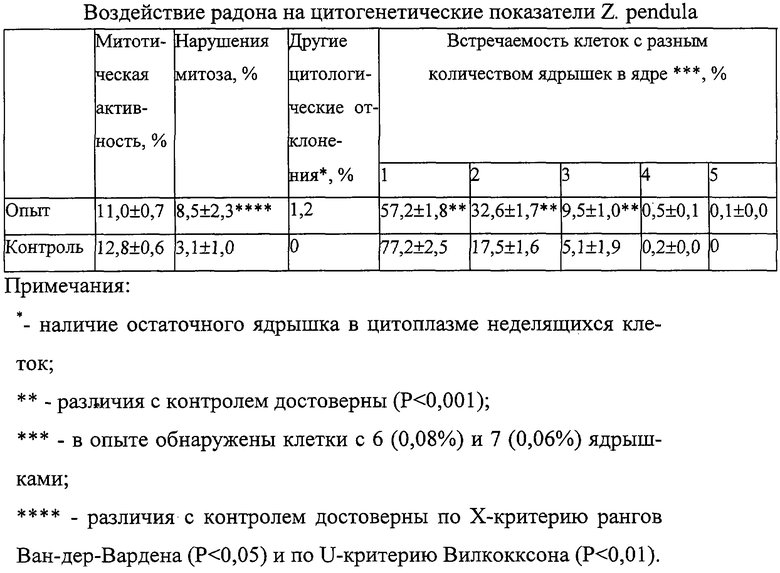
На фиг.2 представлены значения митотического индекса (а) и уровня патологий митоза (б) в корнях Zebrina pendula, облученной ЭРОА радона 100, 1800, 6100 Бк/м3 в течение 28 дней.
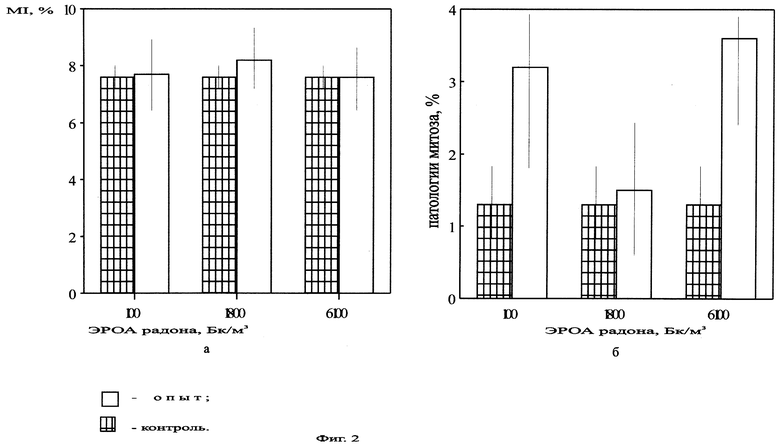
На фиг.3 представлена ядрышковая активность в корнях Zebrina pendula, облученной ЭРОА радона 100, 1800, 6100 Бк/м3 в течение 28 дней.
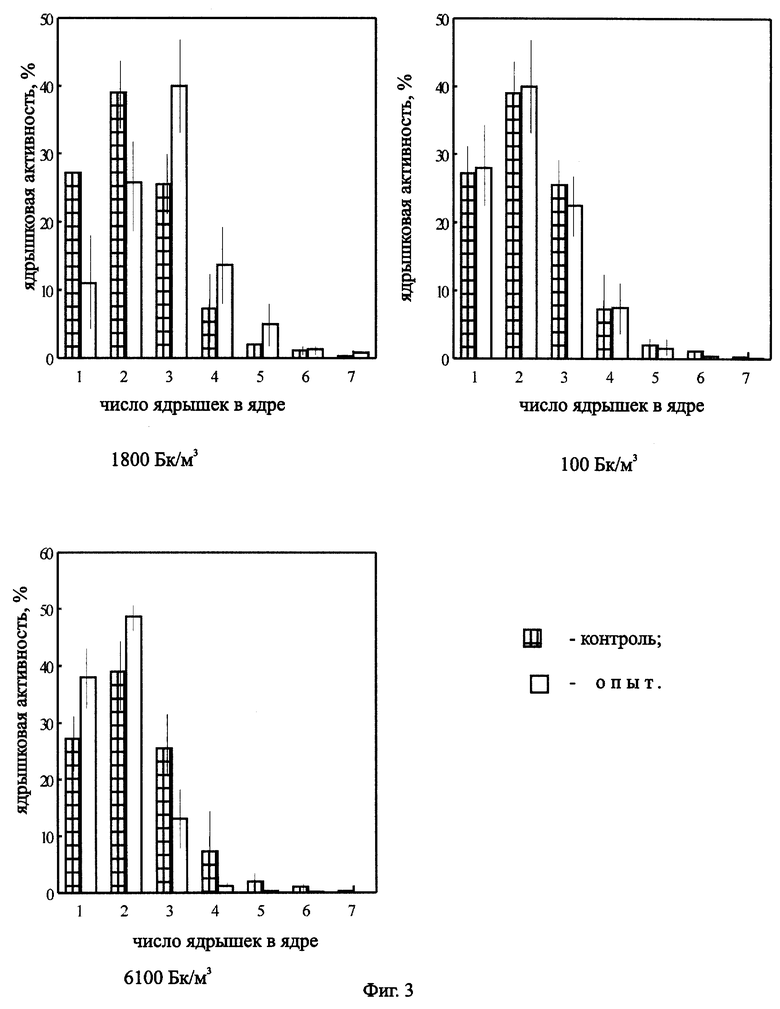
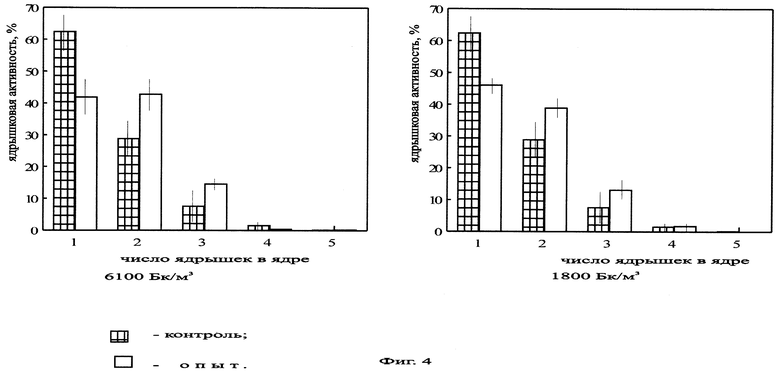
На фиг.4 представлена ядрышковая активность в корнях Zebrina pendula, облученной ЭРОА радона 1800 и 6100 Бк/м3 в течение 14 дней.
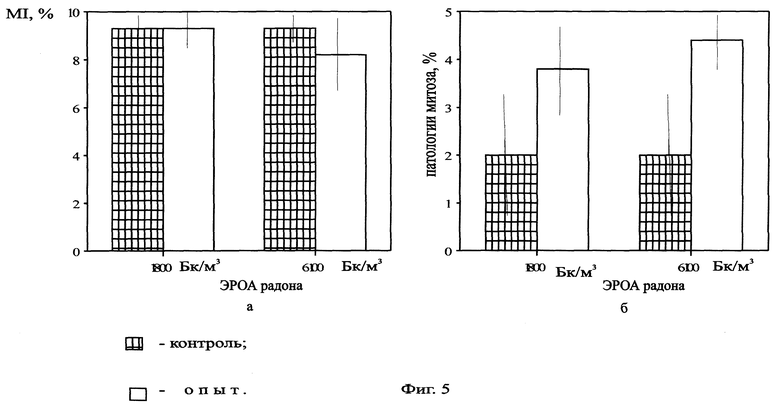
На фиг.5 представлена митотическая активность (а) и уровень патологий митоза (б) в корнях Zebrina pendula, облученной ЭРОА радона 1800 и 6100 Бк/м3 в течение 14 дней.
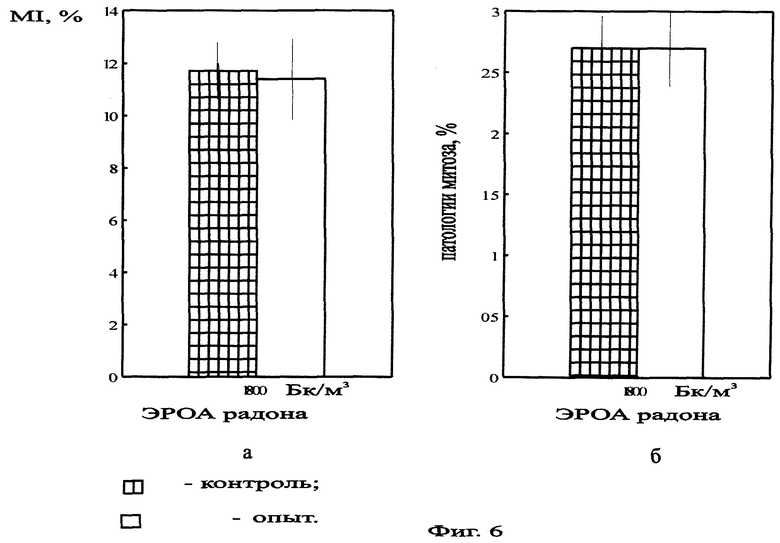
На фиг.6 представлена митотическая активность (а) и уровень патологий митоза (б) в корнях Zebrina pendula, облученной ЭРОА радона 1800 Бк/м3 в течение 3 месяцев.
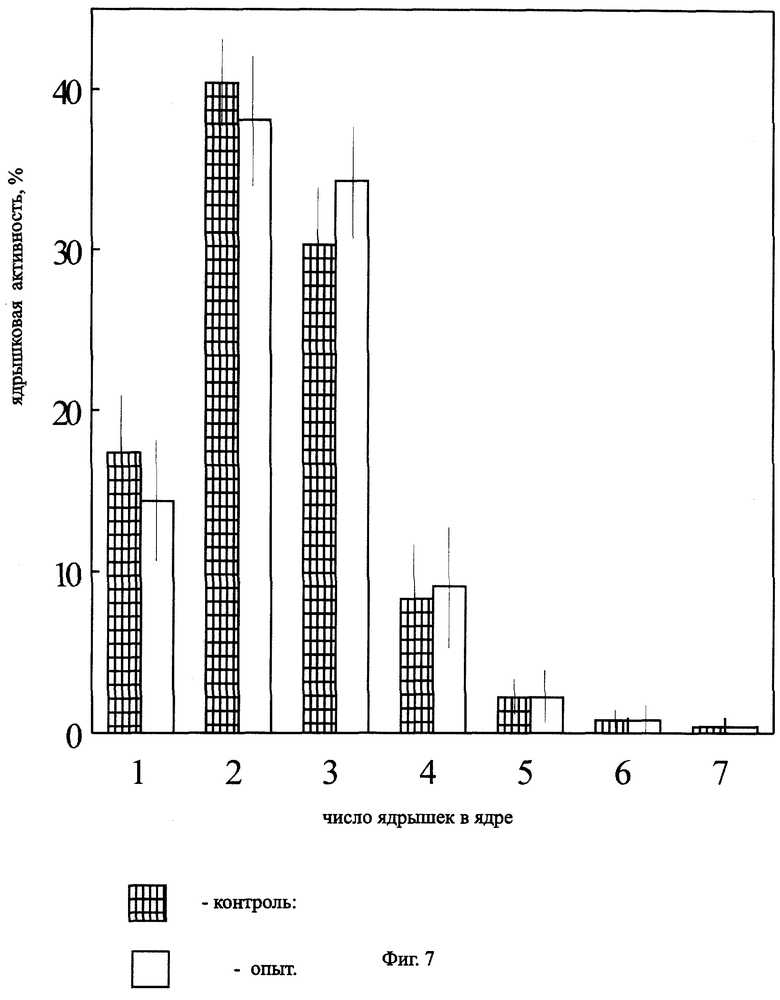
На фиг.7 представлена ядрышковая активность в корнях Zebrina pendula, облученной ЭРОА радона 1800 Бк/м3 в течение 3 месяцев.
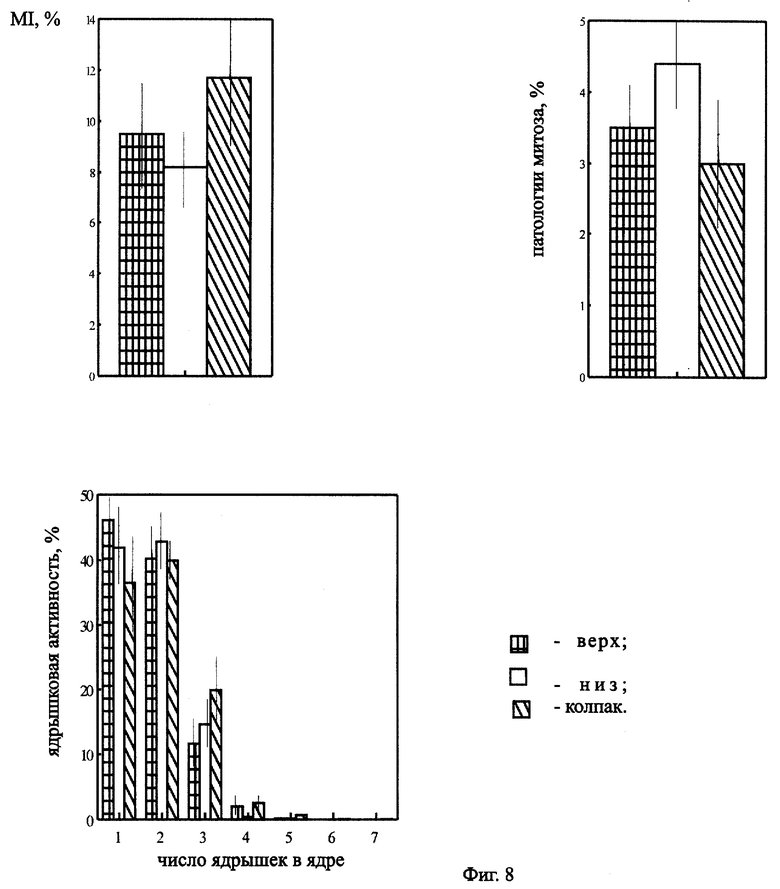
На фиг.8 представлено влияние эффекта положения на цитогенетические показатели Zebrina pendula, облученной ЭРОА радона 6100 Бк/м3 в течение 14 дней.
Опираясь на концепцию беспорогового воздействия радиации (Дубинин, 1961, 1990; Ли, 1963; Шевченко, Померанцева, 1985), мы можем сказать, что любое статистически достоверное повышение ядрышковой активности свидетельствует о превышении предельно допустимых концентраций (200 и 400 Бк/м3).
Появление остаточного ядрышка в метафазе, анафазе и телофазе митоза (~1-2%) говорит о том, что концентрация радона превышает уровень 1800 Бк/м3.
Увеличение частоты встречаемости остаточного ядрышка в митозе (~до 10%), а также расширение спектра аномалий митоза (появление слипания хромосом) показывают, что объемная концентрация радона превышает уровень 5500 Бк/м3.
Появление остаточного ядрышка в цитоплазме неделящихся клеток, статистически достоверное увеличение патологий митоза указывают на повышение концентраций радона до уровня 27500 Бк/м3.
Статистически достоверное уменьшение митотической активности связано со значением ЭРОА более 27500 Бк/м3.
Таким образом, используя цитогенетические критерии, можно эффективно проводить дозиметрию объемных концентраций радиоактивного газа радона в жилых помещениях, используя созданную шкалу для оценки загрязнения жилых помещений радоном (фиг.1).
Пример 1.
Нами были проведены работы по изучению цитогенетических характеристик зебрины повислой при облучении радоном в течение 14 дней (ЭРОА 1800 Бк/м3, 6100 Бк/м3), в течение 28 дней (ЭРОА 100 Бк/м3, 1800 Бк/м3) и 3 месяцев (объемная концентрация 1800 Бк/м3). Источником радона служил сернокислый уран. Активности создавались в прозрачных колпаках объемом 0,5 м3. Подготовку материала к облучению, его фиксацию и изготовление микропрепаратов проводили по описанной выше методике. Результаты эксперимента представлены на фиг.2, 3, 4, 5, 6, 7, 8. При облучении растений радоном объемной активностью 100 Бк/м3 отклонений цитогенетических показателей от контрольных не обнаружено (фиг.2, 3). Это можно объяснить исходя из концепции беспорогового действия радиации (Дубинин, 1961, 1990; Ли, 1963; Шевченко, Померанцева, 1985), возникающими скрытыми повреждениями, которые данным методом не могут выявляться, или недостаточной концентрацией мигрирующих из листьев в корни радиотоксинов, способных вызвать биологический эффект. Увеличение ЭРОА радона до 1800 Бк/м3 (при времени облучения 14 дней) приводило к возрастанию ядрышковой активности (фиг.4, 5), появлению остаточного ядрышка в метафазе-телофазе митоза (до 2,7%), в контроле такое нарушение отсутствовало. Наблюдаемое усиление ядрышковой активности, а также пуфинг хромосом в метафазе митоза в период облучения могут быть обусловлены ослаблением под действием радиации степени конденсации хроматина ядрышкообразующих районов за счет нарушения взаимодействия ДНК-белок. Следствием этого будет недостаточная инактивация рибосомных генов в хромосомах. Усиление ядрышковой активности можно рассматривать как компенсаторный механизм клеток, обеспечивающий синтез необходимого количества белков в условиях подавления функции многих уникальных генов радиацией. Характер пуфинга метафазных хромосом, завершающегося выбрасыванием ядрышкоподобного тельца в цитоплазму, свидетельствует о возможности увеличения числа рибосомных генов путем их амплификации. Появление остаточного ядрышка в метафазе, анафазе и телофазе митоза наблюдалось Буториной у проростков дуба черешчатого из района Чернобыльской АЭС (Буторина, Исаков, 1989; Butorma et al., 1997).
При более длительном времени облучения (28 дней) при той же ЭРОА (1800 Бк/м3) происходило возрастание ядрышковой активности (фиг.3). Однако клетки с остаточным ядрышком в митозе исчезали. Митотическая активность различалась с контролем по F-критерию Фишера (Р<0,05). Это, скорее всего, связано с тем, что при увеличении времени воздействия радиации начинается процесс элиминации наиболее радиочувствительных клеток, а хронический приток радиотоксинов из облучаемых листьев создает стрессовые условия радиационной интоксикации корней.
При облучении в течение 3 месяцев ЭРОА 1800 Бк/м3 наблюдались различия только между значениями митотической активности по F-критерию Фишера (Р<0,05) (фиг.6, 7). Это свидетельствует о завершении процесса элиминации радиочувствительных клеток и возможном неравномерном распределении радиотоксинов между корнями растения.
При увеличении объемной активности радона до 6100 Бк/м3 (14 дней) происходило возрастание ядрышковой активности (фиг.4) и увеличение количества метафаз-телофаз с остаточным ядрышком (до 9,6%) по сравнению с контрольными данными. Однако при сопоставлении с данными по облучению объемной активностью 1800 Бк/м3 в течение 14 дней статистически достоверных различий не наблюдалось. Предположительно, это, скорее всего, связано с выходом системы на новый триггерный уровень. Это согласуется с концепцией Гераськина (Гераськин, 1995) о возможности существования в области малых доз интервалов, на которых клетка способна справиться с увеличивающимся стрессовым воздействием без изменения функциональных показателей.
Дополнительно исследовали зависимость эффекта от положения облучаемого объекта (вверху (активность радона 5500 Бк/м3) и внизу (активность радона 6100 Бк/м3) колпака, а также под полиэтиленовым навесом). Наличие эффекта могло быть обусловлено дополнительным облучением растений оседающими продуктами распада радона. Данные представлены на фиг.8. Статистически достоверных различий в зависимости от положения не обнаружено. На основании этого можно говорить о незначительности влияния эффекта положения традесканции в опыте на цитогенетические показатели. Следовательно, возможно предположить, что основное воздействие происходит за счет внешнего и внутреннего облучения радоном и осевших на листья аэрозолей (продуктов распада) и инкорпорированного облучения этими же частицами, а не за счет облучения частицами, находящимися вблизи (т.е. на поверхности колпака) растения.
Пример 2.
Проводили облучение зебрины повислой ЭРОА радона 27500 Бк/м3. Источником радона служил сернокислый уран. Активности создавались в прозрачных колпаках объемом 0,5 м3. Подготовки материала к облучению, его фиксации и изготовление микропрепаратов проводили по описанной выше методике.
Данные по митотической активности, количеству нарушений митоза, ядрышковой активности у биоиндикатора представлены в таблице.
Под действием радона у исследуемого вида наблюдали широкий спектр аномалий митозов. Спектр патологий митозов у Z. pendula в контроле был представлен (классификация по Алову (1965)) отставанием хромосом в метакинезе. Типы отклонений в протекании митоза у облученных растений -слипание хромосом, отставание хромосом в метакинезе и анафазе, мосты.
Следует особо отметить, что при облучении у 0,3% клеток встречались микроядра (4 на 1596 просмотренных интерфазных клеток), которые являются прямым тестом на действие радиации. У Z. pendula происходило статистически достоверное увеличение числа патологий митоза (для Х-критерия рангов Вандер-Вардена Р<0,05, для U-критерия Вилкокксона Р<0,01). Отмечена также тенденция к снижению митотической активности.
Обнаружено статистически достоверное увеличение (Р<0,001) ядрышковой активности по сравнению с контролем, что можно трактовать как реакцию на стрессовое воздействие облучения и рассматривать изменение ядрышковых характеристик как наиболее чувствительный критерий при воздействии радона.
У зебрины повислой при воздействии радоном наблюдалось явление присутствия оптически плотного, отличающегося по окраске от ядра и сходного по окраске с ядрышком ядрышкоподобного образования в цитоплазме интерфазных клеток, которое можно квалифицировать как остаточное ядрышко.
В связи с тем, что происходило облучение верхней части растения, а радиационные эффекты наблюдались в необлученных корешках, можно говорить о опосредованном воздействии радиации. Сходные опыты были проведены Кузиным (1986), зарегистрировано ингибирование корнеобразования у спиреи при облучении верхушечных почек веток (Krjukova, 1979). А.М.Кузин с соавторами полагают, что ответственны за опосредованное действие облучения радиотоксины - вещества, возникающие в облученных клетках и выполняющие очень важную роль в индукции лучевого патогенеза (Копылов и др., 1966; Кузин, Копылов, 1983; Кузин, 1984; Кузин, 1986).
Заявляемое изобретение относится к области биодозимерии помещений и может быть применено для биодозимерии радона, а также выявления создаваемых им с другими поллютантами жилых помещений синергических биологических эффектов. Способ заключается в проведении анализа по нескольким цитогенетическим показателям в апикальной меристеме корня на одном растительном биоиндикаторе, в качестве которого используется Zebrina pendula Schnizl. Анализируют митотическую активность, ядрышковые характеристики, уровень и спектр патологии митоза. Технический результат: повышение достоверности оценки радонового загрязнения помещений. 2 с. и 8 з.п. ф-лы, 1 табл., 8 ил.
| СПОСОБ ВЫЯВЛЕНИЯ ВОЗДЕЙСТВИЯ ИЗЛУЧЕНИЯ НА БИОЛОГИЧЕСКИЙ ОБЪЕКТ | 1999 |
|
RU2145711C1 |
| ДЕССЛЕР Х.Г | |||
| Влияние загрязнения воздуха на растительность | |||
| - М.: Лесная промышленность, 1981, с.52-54 | |||
| РАДИОМЕТР ДЛЯ ОПЕРАТИВНОГО ИЗМЕРЕНИЯ ОБЪЕМНОЙ АКТИВНОСТИ РАДОНА, ТОРОНА И ДОЧЕРНИХ ПРОДУКТОВ ИХ РАСПАДА В ВОЗДУХЕ | 1996 |
|
RU2123192C1 |
| US 5235190 А, 10.08.1993. | |||

