Способ отбора материнских деревьев Betula pendula, продуцирующих семенное потомство с разной всхожестью и стабильностью генетического материала соматических клеток, по биохимическим параметрам
Изобретение относится к способам выращивания растений, а именно, к способам селекции, и может быть использовано для отбора материнских растений-маточников Betula pendula, продуцирующих семенное потомство с высокой всхожестью и стабильностью генетического материала соматических клеток проростков, по активности пероксидазы в листьях и с проведением проверки ростовой способности потомства по количеству общего белка в семенах и проростках.
В настоящее время существует ряд работ, показывающие, что древесные растения могут продуцировать семенное потомство с разным уровнем цитогенетических нарушений (Артюхов В.Г., Калаев В.Н., Карпова С.С. Цитогенетический полиморфизм семенного потомства деревьев березы повислой (Betula pendula Roth.), произрастающих в различных экологических условиях // Экологическая генетика. - 2009. - Т. 7. - №1. - С. 30-40). Семенное потомство с высоким уровнем цитогенетических нарушений называют мутабильным, с низким - слабомутабильным. Выявлено, что потомство с высоким уровнем цитогенетических нарушений характеризуется худшими ростовыми показателями (по сравнению со слабомутабильным) (Калаев В.Н., Попова А.А. Цитогенетический полиморфизм проростков семян деревьев дуба черешчатого (Quercus robur L.) на территориях с разным уровнем антропогенного загрязнения // Проблемы региональной экологии. 2014. - №2. - С. 176-190). Стоит также отметить, что среди семенного потомства, обладающего низким уровнем цитогенетических нарушений, возможно, будет отмечаться большее фенотипическое разнообразие, что позволяет искать селекционно-ценные формы. Ранее у березы повислой было показано наличие среди семенного потомства форм с высоким и низким уровнем нарушений и дана их цитогенетическая характеристика (Карпова С.С. Влияние антропогенного загрязнения на цитогенетический полиморфизм семенного потомства березы повислой {Betula pendula Roth.) в естественных и искусственных древостоях: дис.... канд. биол. наук / С.С.Карпова. - Воронеж, 2011. - 147 с).
Древесные растения имеют длинный жизненный цикл, долгие сроки наступления генеративного периода, поэтому невозможно проследить в ряду поколений поведение растений. Поскольку селекционный процесс для древесных растений длителен (десятки лет), разрабатываются методики ускоренного получения результатов отбора. Например, ряд исследований для выделения материнских экземпляров использует анализ цитогенетических показателей.
Известен способ отбора материнских растений Rhododendron ledebourii Pojark., продуцирующих семенное потомство с разным уровнем стабильности генетического материала (Патент РФ №2662650, опубл. 2018). Наиболее близким к заявляемому изобретению является способ оценки по цитогенетическим показателям качества семян Rhododendron ledebourii Pojark, где технический результат достигался цитогенетическим анализом (Патент РФ №2654605, опубл. 2018). Недостатком известных способов является трудоемкость при изготовлении и просмотре микропрепаратов, необходимость материальных и временных затрат, наличие высоко квалифицированного персонала, дорогостоящей микроскопической техники. Однако можно отобрать родительские деревья более простым и быстрым способом, чем цитогенетический метод, например, с помощью биохимического анализа.
Некоторые биохимические показатели в частности, активность пероксидазы и других дыхательных ферментов, используются для оценки состояния растений (Воскресенская О.Л. Изменение активности антиоксидантных ферментов у интродуцированных хвойных растений в условиях городской среды / О.Л. Воскресенская, Е.В. Сарбаева, Е.А. Старикова // Вестник Ульяновской государственной сельскохозяйственной академии. - 2015. - №2. - С.56-59), а также качества природной среды (Стаценко А.П. Растительные пероксидазы - маркеры химического загрязнения природных сред / А.П. Стаценко, Л.И. Тужилова, А.А. Вьюговский // Вестник ОГУ. - 2008. - №. 10 (92). - С.188-192).
Известно об изменении показателей состояния защитных систем (пероксидазной активности, содержания общего белка и суммарного количества сульфгидрильных групп защитных пептидов) в листе у березы повислой (Betula pendula Roth) и липы сердцевидной (Tilia cordata Mill.) при действии автотранспортного загрязнения (Ерофеева Е.А. Гормезис и парадоксальные эффекты у растений в условиях автотранспортного загрязнения и при действии поллютантов в эксперименте: дис. … докт. биол. наук / Е.А. Ерофеева. - Нижний Новгород, 2016. - 184 с.). Предлагалось использовать активность пероксидазы Betula pendula как индикатор качества городской среды (Симонова З.А. Активность пероксидазы Betula pendula как индикатор качества городской среды (на примере г. Саратова) / З.А. Симонова, Д.А. Чемаркин // Фундаментальные исследования. - 2013. - №8. - С. 1097-1101). Однако не применялись биохимические параметры (пероксидазная активность, содержание общего белка) в качестве критериев отбора генетически стабильных материнских растений и их семенного потомства.
Изучение цитогенетических характеристик семенного потомства чаще сопряжено с физиологическими параметрами, в частности, всхожестью семян (Филиппова Г.В. Влияние техногенного пылевого загрязнения на физиологические и цитогенетические характеристики семенного потомства лебеды раскидистой (Atriplex patula L.) / Г.В. Филиппова, И.А. Прокопьев, А.А. Шеин // Наука и образование. - 2015. - №3. - С. 89-93) или морфометрическими, например, длиной корней или проростков (Дмитриева С.А. Митотический индекс меристематических клеток и рост корней гороха Pisum sativum при действии модуляторов инозитольного цикла / С.А. Дмитриева, Ф.В. Минибаева, Л.Х. Гордон // Цитология. - 2006. - Т.48, №6. - С. 475-479; Лазарева Е.М. Реорганизация системы микротрубочек клеток корня Medicago sativa L. в условиях акклимации к осмотическому и солевому стрессам / Е.М. Лазарева, Е.Н. Баранова, Е.А. Смирнова // Цитология. - 2017. - Т. 59. - №1. - С. 34-44). Известно о влиянии нитрата свинца на цитогенетические показатели семенного потомства сосны обыкновенной, на активность пероксидазы и других дыхательных ферментов (Белоусов М.В. Влияние нитрата свинца на цитогенетические и некоторые биохимические показатели семенного потомства сосны обыкновенной / М.В. Белоусов, О.А. Землянухина // Вестник Тамбовского университета. Серия: Естественные и технические науки. - Тамбов, 2011. - Т. 16, №5. - С. 1321-1325). Однако никто ранее не использовал биохимические характеристики в селекционных целях для отбора маточников древесных растений, продуцирующих семенное потомство с разной стабильностью генетического материала соматических клеток.
Проведенные авторами исследования выявили наличие прямых корреляционных зависимостей между активностью пероксидазы и цитологическими параметрами: «доля клеток на стадии профазы митоза», «доля клеток на стадии метафазы митоза», «уровень патологий митоза», «уровень клеток с остаточными ядрышками на стадии метафазы-телофазы митоза», а также установлены корреляции между количеством общего белка в семенах и их всхожестью, количеством общего растворимого белка в проростках и цитологическими показателями: «митотический индекс» «уровень патологий митоза», «уровень клеток с остаточными ядрышками», что позволяет производить отбор материнских растений Betula pendula и проверку ростовой способности потомства по биохимическим параметрам.
Задача изобретения - разработка способа отбора по активности пероксидазы в листьях материнских растений Betula pendula, продуцирующих семенное потомство с высокой стабильностью генетического материала соматических клеток и проведение проверки ростовой способности потомства по количеству общего белка в семенах и проростках.
Технический результат - выделение по активности пероксидазы в листьях или количеству общего белка в семенах и проростках материнских растений Betula pendula, продуцирующих семенное потомство с высокой стабильностью генетического материала соматических клеток и проведение проверки ростовой способности потомства по количеству общего белка в семенах и проростках.
Технический результат достигается тем, что в способе отбора по биохимическим параметрам материнских растений Betula pendula, продуцирующих семенное потомство с разной стабильностью генетического материала соматических клеток, включающем выделение не менее пяти учетных фенотипически здоровых деревьев березы повислой, достигших генеративного возраста, на каждой исследуемой территории, срез трех побегов с каждого дерева с распускающимися листьями или почками длиной не менее 30 см, сбор и проращивание семян Betula pendula от каждого материнского растения в отдельности не менее 300 штук, определение активности пероксидазы в распускающихся листьях и количества общего белка в семенах и проростках, согласно изобретению, вычисляют активность пероксидазы материнского растения, полученные значения сравнивают со значением 0,28 мкмоль/мин/мг, выше которого материнский экземпляр продуцирует мутабильное семенное потомство, в противном случае -слабомутабильное; вычисляют количество общего белка в семенах и проростках сравнивают со значениями 0,180 мг/мл и 0,160 мг/мл соответственного, ниже которых материнский экземпляр продуцирует мутабильное семенное потомство, характеризующееся низкой всхожестью и ростовой способностью, в противном случае - слабомутабильное с высокой всхожестью и ростовой способностью.
Способ отбора материнских деревьев Betula pendula, продуцирующих семенное потомство с разной стабильностью генетического материала соматических клеток, по биохимическим параметрам обеспечивает экологическую оценку местопроизрастания и возможность выявления генетически стабильных экземпляров.
Предлагаемый способ отбора материнских растений Betula pendula позволяет относительно быстро оценить стабильность генетического материала семенного потомства материнского растения и определить возможность его использования в качестве маточника по активности пероксидазы в листьях. Кроме того, оценка количества общего белка в семенах и проростках обеспечивают проведение проверки ростовой способности потомства. При использовании предлагаемого способа отбора происходит экономия времени исследования, которое может занимать один вегетационный сезон для сбора и проращивания семян; для получения проростков, анализа пероксидазной активности, оценки количества общего белка в семенах и проростках - 10-20 дней. Всего лишь несколько недель требуется для распускания листьев срезанных побегов в лабораторных условиях (в весеннее время возможно сразу после среза побегов осуществлять анализ активности пероксидазы в распускающихся листьях). Таким образом, значительно сокращается время постановки эксперимента по сравнению с традиционным индивидуальным отбором материнских экземпляров по морфологическим признакам.
Способ реализуется следующим образом.
1. В качестве материнских используют не менее пяти учетных фенотипически здоровых деревьев березы повислой (Betula pendula Roth.), произрастающие в экологически чистой территории, с которых срезают по три побега с распускающимися листьями или почками (длиной не менее 30 см) и собирают семена для проверки ростовой способности потомства (не менее 300 штук с каждого дерева). Если побеги с почками, их ставят в воду до распускания листьев.
Активность пероксидазы определяют в четырех повторностях для каждого дерева по методике, модифицированной О.А. Землянухиной (Землянухина О.А. Сравнительный анализ методов определения активности и изоферментного спектра пероксидаз различного происхождения / О.А. Землянухина, В.Н. Калаев, B.C. Воронина // Успехи современного естествознания 2017. - №9. - С. 13-21). Единицу ферментативной активности вычисляют как количество фермента, превращающего 1 мкм субстрата за 1 мин при 25°С (μmol/min). Измерения оптической плотности проводят на спектрофотометре UNICO 2800 (USA). Удельную активность вычисляют путем отношения общей активности фермента к 1 мг белка в 1 мл ферментативного препарата (мкмоль/мин/мг). Анализ электрофоретических спектров проводят с использованием вертикальной камеры в 7.5% полиакриламидном геле в соответствии со стандартным методом Дэвиса (Davis В.J. Disc Electrophoresis. II. Method and application to human serum proteins / B.J. Davis //Ann. N.Y. Acad. Sci. - 1964. - V. 121. - P. 404-427.). Согласно изобретению, вычисляют среднюю величину активности пероксидазы распускающихся листьев для каждого материнского растения и сравнивают со значением 0,028 мкмоль/мин/мг, выше которого материнский экземпляр продуцирует мутабильное семенное потомство. Если среднюю величину активности пероксидазы 0,028 мкмоль/мин/мг и ниже, то материнский экземпляр продуцирует слабомутабильное семенное потомство.
2. Растительным материалом для исследований потомства служат семена и проростки. Проращивание семян осуществляют по ГОСТу 13056.6-97. Семена проращивают при постоянной температуре +25°С в трех повторностях по 100 шт. Лабораторную всхожесть семян вычисляют как отношение числа проросших семян к общему числу семян и выражают в %, согласно рекомендациям (ГОСТ 13056.6-97). Для получения белковых препаратов используют навеску одного семени или проростка (исследуют по 10 семян и 10 проростков каждого дерева), которую растирают со стеклянным песком в 0.1 М трис-HCl буфере, рН 7.5 и центрифугируют в эппендорфах в течение 10 мин при 20000 g, 4°С, на центрифуге СМ50 ELMI (Латвия). В надосад очной жидкости измеряют содержание общего растворимого белка по методу Брэдфорда, в качестве стандарта используют бычий сывороточный альбумин (Sigma) (Bradford V.V. A rapid and sensitive method for the quantities of protein utilizing the principle of protein-dye binding / V.V. Bradford // Anal. Biochem. - 1976. - V. 72, N4. - P. 417-422). Согласно изобретению, вычисляют среднее количество общего растворимого белка для каждого материнского растения в семенах и сравнивают со значением 0,180 мг/мл и в проростках - 0,160 мг/мл, ниже которых материнский экземпляр продуцирует мутабильное семенное потомство. Если средние величины общего белка в семенах 0,180 мг/мл, а проростках - 0,160 мг/мл и выше, то материнский экземпляр продуцирует слабомутабильное семенное потомство.
Пример 1
Активность пероксидазы определяли по методике, модифицированной О.А. Землянухиной (Землянухина О.А. Сравнительный анализ методов определения активности и изоферментного спектра пероксидаз различного происхождения / О.А. Землянухина, В.Н. Калаев, B.C. Воронина // Успехи современного естествознания 2017. - №9. - С. 13-21).
Для цитогенетического исследования семенного потомства Betula pendula использовали зрелые семена, собранные от каждого в отдельности фенотипически здорового материнского растения (без визуальных повреждений паразитами). Семена проращивали в чашках Петри при температуре +25°С. По достижении корешками длины 0,5-1 см их фиксировали в 9 часов утра в ацетоалкоголе - смеси 96% этилового спирта и ледяной уксусной кислоты (3:1), после чего материал хранили в холодильнике при температуре +4°С. Из корешков проростков готовили постоянно-давленные микропрепараты с использованием жидкости Гойера по ранее описанной методике (Вострикова Т.В. Изучение суточной митотической активности у березы повислой / Т.В. Вострикова, А.К. Буторина // Цитология. - 2004. - Т. 46. - №6. - С. 520-524.)
Анализировали цитогенетические характеристики нечетного числа проростков семян от каждого экземпляра материнского растения (не менее 9). Препараты изучали с помощью микроскопа LABOVAL-4 (Carl Zeiss, Jena) при общем увеличении 40×1,5×10. В каждом микропрепарате (1 препарат соответствует 1 корешку и одному проростку) анализировали около 500 клеток.
На микропрепаратах подсчитывали общее число клеток, число клеток на стадиях митоза: в профазе, метафазе, анафазе-телофазе; число клеток с нарушениями деления (число делящихся клеток с аберрациями); число клеток с остаточными ядрышками на стадии метафазы-телофазы митоза. Остаточные ядрышки выглядят как отдельные круглые или каплеобразные тельца, соединенные с хромосомами на стадии метафазы-анафазы митоза (Вострикова Т.В. Изучение суточной митотической активности у березы повислой / Т.В. Вострикова, А.К. Буторина // Цитология. - 2004. - Т. 46. - №6. - С. 520-524.).
Вычисляли следующие цитогенетические показатели: митотическую активность, показателем которой является митотический индекс - отношение числа делящихся клеток к общему числу подсчитанных клеток (в %), % клеток в профазе (отношение числа клеток в профазе к числу делящихся клеток), % клеток в метафазе (отношение числа клеток в метафазе к числу делящихся клеток), % клеток в анафазе-телофазе (отношение числа клеток в анафазе-телофазе к числу делящихся клеток), уровень патологий митоза (как отношение числа клеток с нарушениями митоза к общему числу делящихся клеток, в %), уровень клеток с остаточными ядрышками на стадии метафазы-телофазы митоза (как отношение числа клеток с остаточными ядрышками от общего числа клеток на указанных стадиях, в %). Классификацию патологических митозов проводили по И.А. Алову (Алов И.А. Цитофизиология и патология митоза. М.: Медицина, 1972. - 264 с.).
Производили компьютерную статистическую обработку данных с помощью пакета программ "Stadia". Процедура группировки данных и их обработка изложены в работе А.П. Кулаичева Методы и средства комплексного анализа данных (А.П. Кулаичев. - М.: ФОРУМ: ИНФА-М, 2006. - 512 с.) Сравнение выборок по уровню патологий митоза и уровню клеток с остаточными ядрышками проводили с использованием Х-критерия рангов Ван-дер-Вардена, так как данные признаки не подчиняются нормальному распределению. Сравнение выборок по остальным цитогенетическим показателям и по флуктуирующей асимметрии производили по t-критерию Стьюдента. Для определения корреляционной зависимости использовали коэффициент корреляции рангов Спирмена (rs).
В качестве критериев стабильности генетического материала соматических клеток семенного потомства использовали совокупность цитогенетических характеристик (митотическая активность клеток (в %), число клеток на стадиях митоза (в %), уровень клеток с патологиями митоза (в %), уровень клеток с остаточными ядрышками на стадии метафазы-телофазы митоза (в %).
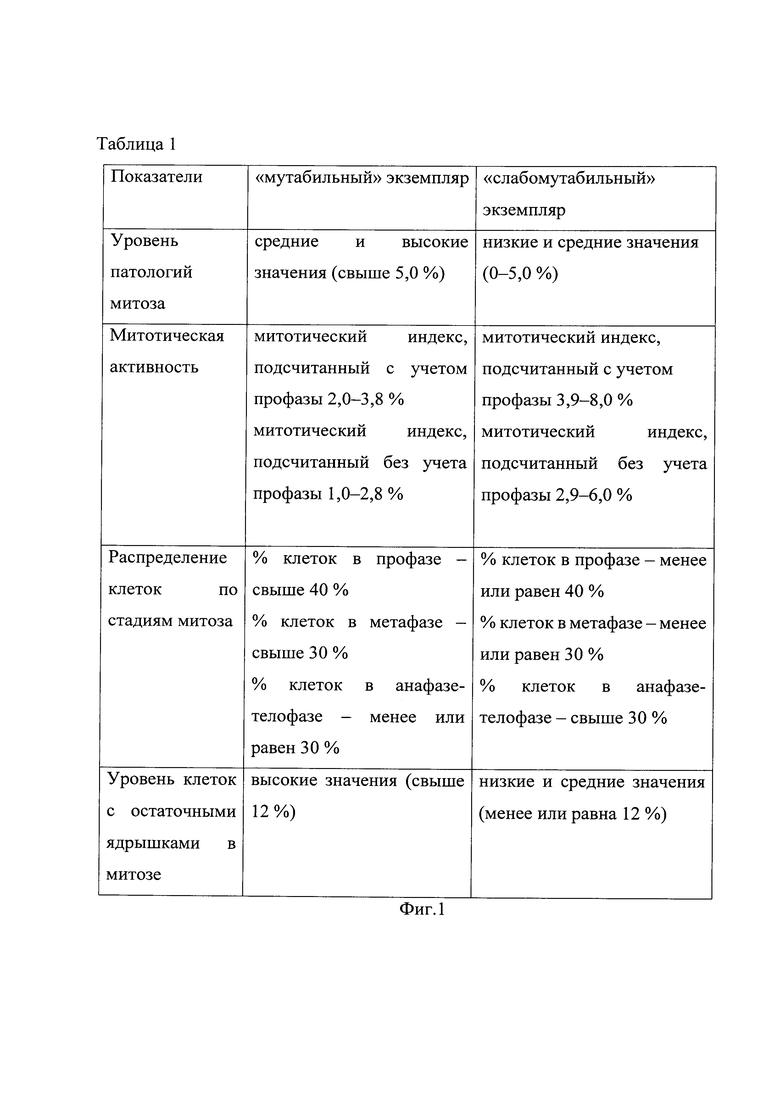
Полученные цитогенетические показатели каждого проростка сравнивали со значениями, представленными в таблице 1, и относили проросток к мутабильной или слабомутабильной группе. Если по некоторым цитогенетическим показателям проросток можно отнести к одной группе, а по другим к другой, то проросток относили к мутабильной или слабомутабильной группе по большинству исследованных цитогенетических характеристик.
На фиг. 1 приведена Таблица 1 критериев отнесения цитогенетических показателей семенного потомства Betula pendula к группам с разной стабильностью генетического материала.
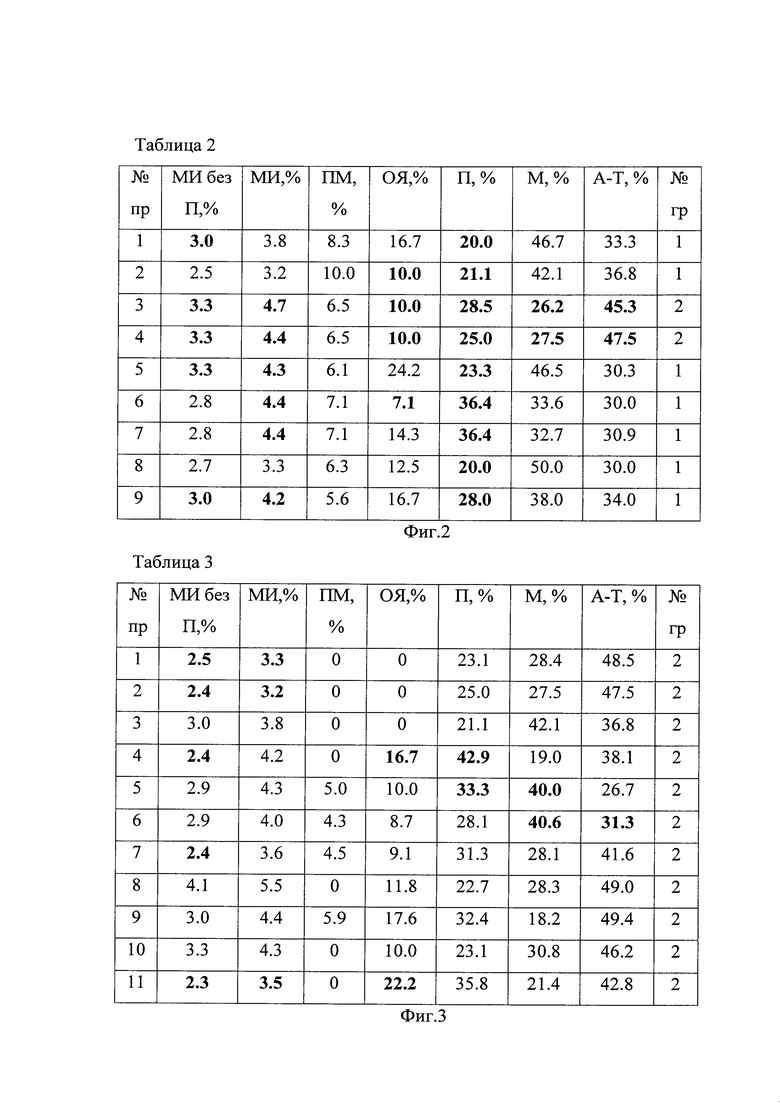
На фиг. 2 и 3 приведены Таблицы 2 и 3 цитогенетических характеристик семенного потомства материнского растения Betula pendula №1 (продуцирующего цитогенетически нестабильное семенное потомство) и материнского растения Betula pendula №2 (продуцирующего цитогенетически стабильное семенное потомство), где № пр. - № препарата; МИ без П - митотический индекс без учета клеток, подсчитанных в стадии профазы, %; МИ, % - митотический индекс, подсчитанный с учетом стадии профазы; ПМ, % - уровень патологий митоза; ОЯ, % - уровень клеток с остаточными ядрышками; П, % - % клеток в стадии профазы; М, % - % клеток в стадии метафазы; А-Т, % - % клеток в стадии анафазы-телофазы; № гр - № группы: 1 - мутабильная, 2 - слабомутабильная.
На фиг. 4 приведена таблица 4 обобщенных цитогенетических характеристик семенного потомства Betula pendula, где * - различия параметров материнского растения 1 и 2 достоверны (Р<0,05).
Обозначения: № др - № дерева; МИ без П - митотический индекс без учета клеток, подсчитанных в стадии профазы, %; МИ, % - митотический индекс, подсчитанный с учетом стадии профазы; ПМ, % - уровень патологий митоза; ОЯ, % - уровень клеток с остаточными ядрышками; П, % - число клеток в стадии профазы; М, % - число клеток в стадии метафазы; А-Т, % - число клеток в стадии анафазы-телофазы; * - различия параметров материнского растения 1 и 2 достоверны (Р<0,05).
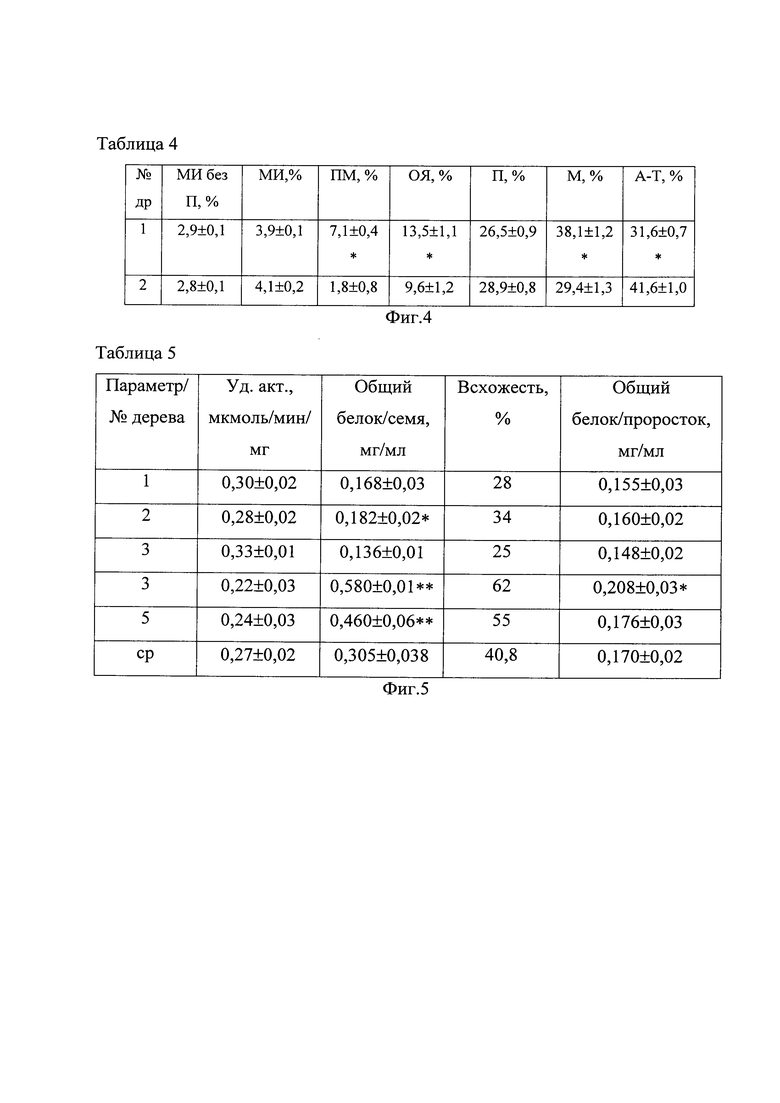
На фиг. 5 приведена Таблица 5 физиолого-биохимических параметров 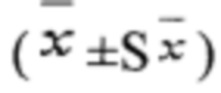 березы повислой (Betula pendula), где № - № дерева; ср - среднее значение параметра; Уд. акт., мкмоль/мин/мг, - удельная активность пероксидазы; * - различия между деревьями выборки достоверны (Р<0,05); ** - различия между деревьями достоверны (Р<0,01).
березы повислой (Betula pendula), где № - № дерева; ср - среднее значение параметра; Уд. акт., мкмоль/мин/мг, - удельная активность пероксидазы; * - различия между деревьями выборки достоверны (Р<0,05); ** - различия между деревьями достоверны (Р<0,01).
Обозначения: № - № дерева; ср - среднее значение параметра; Уд. акт., мкмоль/мин/мг - удельная активность пероксидазы; * - различия между деревьями выборки достоверны (Р<0,05); ** - различия между деревьями достоверны (Р<0,01)
Результаты анализа цитогенетических характеристик семенного потомства материнских растений представили в виде таблицы (фиг. 2-3).
В соответствии со значениями цитогенетических показателей, указанными в таблице 1, семенное потомство тестируемых материнских растений было разделено на две группы: мутабильную и слабомутабильную. Однако не во всех случаях цитогенетические показатели проростков строго соответствовали определенной группе. Например, проростки № 1, 5, 6, 7, 9 материнского экземпляра №1 (табл. 2) имели высокий митотический индекс и низкий % клеток в стадии профазы, как в слабомутабильной группе (табл. 1). Остальные характеристики данных проростков соответствовали мутабильной группе, поэтому по большинству цитогенетических показателей их относили к мутабильной группе. Хотя проростки № 3, 4 (табл. 2) по совокупности цитогенетических характеристик выделили в слабомутабильную группу, большинство проростков материнского растения Betula pendula №1 имели мутабильную группу, поэтому уровень стабильности генетического материала соматических клеток семенного потомства материнского растения № 1 оценили как низкий.
Проростки № 1, 2, 4, 7, 11 материнского экземпляра № 2 имели низкий митотический индекс, проростки № 4, 5 - высокий % клеток в стадии профазы, проростки № 5, 6 - высокий % клеток в стадии метафазы, проросток № 5 - высокий % клеток в стадии анафазы-телофазы, № 4, 11 - высокий уровень клеток с остаточными ядрышками на стадии метафазы-телофазы митоза (табл.3), как в мутабильной группе (табл. 1). Однако по большинству цитогенетических характеристик все проростки материнского растения Betula pendula №2 отнесли к слабомутабильной группе (табл. 3). В соответствии таблицей 1, уровень стабильности генетического материала соматических клеток семенного потомства материнского растения №2 оценивается как высокий. Сравнительный анализ цитогенетических характеристик семенного потомства выявил различия между параметрами материнских деревьев № 1 и № 2 (табл. 4).
Значения физиолого-биохимических параметров приведены в таблице 5 для 5 деревьев Betula pendula. Согласно изобретению, вычисляли среднюю величину активности пероксидазы распускающихся листьев для каждого материнского растения и сравнивали со значением 0,028 мкмоль/мин/мг. У экземпляров № 1 и 3 активность пероксидазы составляет 0,030 мкмоль/мин/мг и 0,033 мкмоль/мин/мг, следовательно, данные деревья продуцируют мутабильное семенное потомство. У экземпляров № 2, 4, 5 активность пероксидазы - 0,028 мкмоль/мин/мг и ниже, что указывает на продуцирование слабомутабильного семенного потомства (табл. 5). Они являются растениями-маточниками Betula pendula, продуцирующими семенное потомство с высокой стабильностью генетического материала соматических клеток проростков.
У материнского растения Betula pendula № 1 (мутабильного), семенное потомство которого характеризуется низким митотическим индексом, большим числом клеток в профазе и метафазе, меньшим числом клеток в анафазе-телофазе, высоким уровнем патологий митоза и уровнем клеток с остаточными ядрышками на стадии метафазы-телофазы митоза, обнаружено увеличение активности пероксидазы распускающихся листьев. Семенное потомство материнского растения Betula pendula №2 (слабомутабильного) показало высокий митотический индекс, меньшее число клеток в профазе и метафазе, больше числом клеток в анафазе-телофазе, низкий уровень патологий митоза и клеток с остаточными ядрышками на стадии метафазы-телофазы митоза. Данное дерево с уровнем активности пероксидазы распускающихся листьев 0,028 мкмоль/мин/мг можно определить как растение-маточник с высокой стабильностью генетического материала соматических клеток проростков.
Высокие положительные корреляции отмечались между активностью пероксидазы и цитогенетическими параметрами: уровнем патологий митоза (rs=0,98 (Р<0,05)), уровнем клеток с остаточными ядрышками (rs=0,98 (Р<0,05)), числом профаз (rs=0,98 (Р<0,05)), числом метафаз (rs=0,88 (Р<0,05)). Это доказывает соответствие показателей и позволяет отбирать материнские растения, продуцирующие стабильное семенное потомство по биохимическим признакам, в частности, по активности пероксидазы.
Пример 2
Проращивание семян и определение всхожести осуществляли по ГОСТу 13056.6-97. Семена проращивали при постоянной температуре +25°С в трех повторностях по 100 шт. Лабораторную всхожесть семян вычисляли как отношение числа проросших семян к общему числу семян и выражали в %, согласно рекомендациям (ГОСТ 13056.6-97). Для получения белковых препаратов использовали навеску одного семени (исследовали по 10 семян каждого дерева), которую растирали со стеклянным песком в 0.1 М трис-НСl буфере, рН 7.5 и центрифугировали в эппендорфах в течение 10 мин при 20000 g, 4°С, на центрифуге СМ50 ELMI (Латвия). В надосадочной жидкости проводили измерение содержания общего растворимого белка по методу Брэдфорда, в качестве стандарта использовали бычий сывороточный альбумин (Sigma) (Bradford V.V. A rapid and sensitive method for the quantities of protein utilizing the principle of protein-dye binding / V.V. Bradford // Anal. Biochem. - 1976. - V.72, № 4. - P. 417-422).
Согласно изобретению, вычисляли среднее количество общего растворимого белка для каждого материнского растения в семенах и сравнивали со значением 0,180 мг/мл и в проростках - 0,160 мг/мл, ниже которых материнский экземпляр продуцирует мутабильное семенное потомство, как у экземпляров № 1 и 3. Если средние величины общего белка в семенах 0,180 мг/мл, а проростках - 0,160 мг/мл и выше, то материнский экземпляр продуцирует слабомутабильное семенное потомство с высокой ростовой способностью, как у экземпляров № 2, 4, 5 (табл. 5).
Нами показано, что количество общего растворимого белка у каждого исследуемого дерева ассоциировано со всхожестью семян: при увеличении первого параметра возрастает второй и наоборот. Это подтверждается высокими положительными корреляциями между данными показателями. Коэффициент корреляции rs=0,9 (Р<0,05). Следовательно, по количеству общего растворимого белка возможно прогнозировать всхожесть семян.
Количество общего белка в проростках характеризует метаболические процессы, происходящие у семенного потомства после прорастания семени. Данный параметр в проростках березы повислой (Betula pendula) был ниже (различия достоверны (Р<0,01)), чем в семенах (табл. 5), но соответствие со всхожестью сохраняется. Растения-маточники Betula pendula № 2, 4, 5 продуцируют семенное потомство с высокой всхожестью и стабильностью генетического материала соматических клеток проростков.
Таким образом, способ отбора материнских растений Betula pendula с разной стабильностью генетического материала соматических клеток проростков по активности пероксидазы распускающихся листьев, обеспечивает возможность выявления экземпляров-маточников, продуцирующих семенное потомство с высокой всхожестью и ростовой способностью (и стабильностью генетического материала). Определение цитогенетических показателей проростков семян и выявление их корреляций с количеством общего растворимого белка в семенах и проростках позволяет провести проверку ростовой способности потомства. Кроме того, по количеству общего растворимого белка возможно прогнозировать всхожесть семян. Исследование занимает 1 вегетационный сезон, т.е. значительно сокращается время постановки эксперимента по сравнению с традиционным индивидуальным отбором по морфологическим признакам.
Изобретение относится к области биотехнологии. Изобретение представляет собой способ отбора материнских деревьев Betula pendula, продуцирующих семенное потомство с разной стабильностью генетического материала соматических клеток, по биохимическим параметрам (количество общего белка в семенах и проростках, активность пероксидазы), включающий выделение не менее пяти учетных фенотипически здоровых деревьев березы повислой, достигших генеративного возраста, на каждой исследуемой территории, срез трех побегов с каждого дерева с распускающимися листьями или почками длиной не менее 30 см, сбор и проращивание семян Betula pendula от каждого материнского растения в отдельности не менее 300 штук, определение активности пероксидазы в распускающихся листьях и количества общего белка в семенах и проростках, при этом вычисляют активность пероксидазы материнского растения, полученное значение сравнивают со значением 0,28 мкмоль/мин/мг, выше которого материнский экземпляр продуцирует мутабильное семенное потомство, в противном случае - слабомутабильное, а вычисленное количество общего белка в семенах и проростках сравнивают со значениями 0,180 мг/мл и 0,160 мг/мл соответственно, ниже которых материнский экземпляр продуцирует мутабильное семенное потомство, характеризующееся низкой всхожестью и ростовой способностью, в противном случае - слабомутабильное с высокой всхожестью и ростовой способностью. 5 ил., 2 пр.
Способ отбора материнских деревьев Betula pendula, продуцирующих семенное потомство с разной стабильностью генетического материала соматических клеток, по биохимическим параметрам (количество общего белка в семенах и проростках, активность пероксидазы), включающий выделение не менее пяти учетных фенотипически здоровых деревьев березы повислой, достигших генеративного возраста, на каждой исследуемой территории, срез трех побегов с каждого дерева с распускающимися листьями или почками длиной не менее 30 см, сбор и проращивание семян Betula pendula от каждого материнского растения в отдельности не менее 300 штук, определение активности пероксидазы в распускающихся листьях и количества общего белка в семенах и проростках, при этом вычисляют активность пероксидазы материнского растения, полученное значение сравнивают со значением 0,28 мкмоль/мин/мг, выше которого материнский экземпляр продуцирует мутабильное семенное потомство, в противном случае - слабомутабильное, а вычисленное количество общего белка в семенах и проростках сравнивают со значениями 0,180 мг/мл и 0,160 мг/мл соответственно, ниже которых материнский экземпляр продуцирует мутабильное семенное потомство, характеризующееся низкой всхожестью и ростовой способностью, в противном случае - слабомутабильное с высокой всхожестью и ростовой способностью.
| АРТЮХОВ В.Г | |||
| и др | |||
| Цитогенетический полиморфизм семенного потомства деревьев Березы повислой (Betula pendula roth), произрастающих в различных экологических условиях, Экологическая генетика, том VII, N1, 2009, с.30-40 | |||
| СПОСОБ ОЦЕНКИ ПО ЦИТОГЕНЕТИЧЕСКИМ ПОКАЗАТЕЛЯМ КАЧЕСТВА СЕМЯН RHODODENDRON LEDEBOURII POJARK | 2016 |
|
RU2654605C2 |
| СТАЦЕНКО А.П | |||
| и др | |||
| Растительные пероксидазы - маркеры химического загрязнения природных сред., | |||

