Область техники
Изобретение относится к экспрессии белков в трансформированных дрожжевых клетках, к ДНК-конструкции и векторам для применения в таком способе и дрожжевым клеткам, трансформированным указанными векторами.
Предпосылки изобретения
Трансформированные дрожжевые штаммы широко используются для экспрессии белков. См., например, заявки на европейский патент №№0088632А, 0116201А, 0123294А, 0123544А, 0163529А, 0123289А, 0100561А, 0189998А и 0195986А, РСТ заявки №№ WO 95/01421, 95/02059 и WO 90/10075 и патент США №4546082.
Общей особенностью вышеуказанных методов является то, что дрожжевая плазмида содержит ген-маркер устойчивости к антибиотикам. Такой ген-маркер получают на начальных стадиях клонирования в Е.coli, где он используется для скрининга трансформированных клеток или сохранения плазмид, используемых в качестве векторов. Считается, что гены-маркеры устойчивости к антибиотикам не оказывают вредного влияния на культивирование трансформированной дрожжевой клетки, поэтому такую ДНК, как правило, не удаляют. Кроме того, конструкцию на основе плазмидного вектора обычно исследуют, выделяя плазмиды из трансформированных дрожжевых клеток и трансформируя выделенную плазмиду в Е.coli с последующим отбором антибиотика. Таким образом, в практических целях удобно сохранять ген-маркер устойчивости к антибиотикам.
Хотя условия работы в научно-исследовательских лабораториях и на промышленных предприятиях определяются строгими правилами безопасности, всегда существует небольшая вероятность того, что несколько клеток могут случайно попасть в окружающую среду. Благодаря своей очень сложной природе такие генетически сконструированные микроорганизмы остаются жизнеспособными в течение очень короткого периода времени и вероятность того, что они могут причинить вред окружающей среде, чрезвычайно мала. Именно по этой причине разрешено использовать такие трансформированные микроорганизмы в научно-исследовательских и производственных целях.
Даже если клетки быстро погибают, плазмиды, содержащие ген устойчивости к антибиотикам, могут случайно попасть в окружающую среду, и существует теоретическая вероятность сообщения устойчивости к антибиотикам бактериям в случае случайного поглощения ими такой плазмиды.
Антибиотики имеют важное значение для лечения бактериальных инфекций у людей и животных. Поэтому желательно свести до минимума возможное загрязнение окружающей среды геном, сообщающим устойчивость к антибиотикам.
Необходимы еще более безопасные методы по сравнению с применяемыми в настоящее время, и целью настоящего изобретения является создание таких усовершенствованных методов.
Краткое изложение существа изобретения
Настоящее изобретение относится к способу экспрессии гетерологичных белков или полипептидов в дрожжах, в соответствии с которым штамм дрожжевого трансформанта содержит экспрессирующий вектор, имеющий ген-маркер устойчивости к антибиотикам, используемый на начальных стадиях клонирования, который делают нефункциональным в результате модификации in vitro до трансформации дрожжевой клетки-хозяина. Настоящее изобретение относится также к последовательностям ДНК и экспрессирующим векторам, используемым при осуществлении этого способа, и к трансформированным дрожжевым клеткам.
Одним объектом настоящего изобретения является рекомбинантный дрожжевой экспрессирующий вектор, не способный сообщать устойчивость к антибиотикам бактериальным клеткам и имеющий ген, кодирующий гетерологичный ген и ген-маркер устойчивости к антибиотикам, который делают нефункциональным в результате модификации in vitro.
Другим объектом настоящего изобретения является способ получения целевого полипептида или белка, включающий культивирование дрожжевого штамма, содержащего вектор, не способный сообщать устойчивость к антибиотикам бактериальным клеткам и имеющий ген, кодирующий гетерологичный ген и ген-маркер устойчивости к антибиотикам, который делают нефункциональным в результате модификации in vitro до трансформации дрожжевой клетки-хозяина, и выделение целевого продукта из культуральной среды.
Способ по этому изобретению обычно включает культивирование дрожжевого штамма, содержащего дрожжевую экспрессирующую плазмиду, имеющую функциональный ген-маркер устойчивости к антибиотикам, используемый на начальных стадиях клонирования в бактериях, который делают нефункциональным в результате делеции in vitro части или всего гена-маркера до введения в дрожжевую клетку-хозяина, используемую для экспрессии и секреции целевого полипептида или белка.
Ген-маркер устойчивости к антибиотикам предпочтительно удаляют путем введения приемлемых сайтов рестрикции с обеих сторон гена-маркера устойчивости к антибиотикам, после чего ген-маркер удаляют путем обработки in vitro приемлемыми рестрикционными ферментами.
Настоящее изобретение относится также к трансформированным дрожжевым штаммам, содержащим вектор, не способный сообщать устойчивость к антибиотикам бактериальным клеткам и имеющий ген, кодирующий гетерологичный ген и ген-маркер устойчивости к антибиотикам, который делают нефункциональным в результате модификации in vitro до трансформации дрожжевой клетки-хозяина.
Дрожжевой штамм предпочтительно является штаммом Saccharomyces, в частности штаммом Saccharomyces cerevisiae.
В используемом здесь значении термин "ген-маркер устойчивости к антибиотикам" означает ген, который позволяет производить фенотипический отбор трансформированных бактериальных клеток и амплификацию плазмиды.
Обычно используемыми генами-маркерами устойчивости к антибиотикам в Е.coli являются гены-маркеры, сообщающие устойчивость к ампициллину (ДМР), хлорамфениколу, неомицину, канамицину и тетрацилину.
В используемом здесь значении термин "нефункциональный ген-маркер" означает, что ген-маркер удален или утратил функциональность в результате делеции части гена. Желательно, чтобы ген был удален полностью.
В используемом здесь значении термин "модификация in vitro" означает стадии модификации вектора, выполняемые вне клеточной среды.
В используемом здесь значении выражение "не способен сообщать устойчивость к антибиотикам бактериальным клеткам" означает, что гены-маркеры устойчивости к антибиотикам не могут функционировать ни в одном организме в результате вышеописанной обработки гена.
В используемом здесь значении термин "дрожжевая клетка-хозяин" означает дрожжевой организм, трансформированный или трансфицированный экспрессирующей плазмидой или вектором.
Краткое описание чертежей
Настоящее изобретение далее иллюстрировано прилагаемыми чертежами.
На фиг.1 показана экспрессирующая плазмида рАК729, которая содержит ген, экспрессирующий предшественник инсулина под контролем последовательности промотора TPI и терминатора TPI из S. cerevisiae и сигнальной лидерной последовательности, содержащей сигнальный пептид YAP3 и синтетический лидерный пептид LA19. Конструкция рАК729 описана в заявке на патент WO 97/22706. Эта плазмида содержит также последовательность AMP-R из pBR322/pUC13, включающую ген устойчивости к ампициллину и точку начала репликации ДНК в Е.coli.
На фиг.2 показана плазмидная карта плазмиды рАК729.5, используемой для генерации штамма NN729.5, не имеющего гена АМР, до делеции гена Amp.
На фиг.3 показана плазмидная карта плазмиды рАК729.6, используемой для генерации штамма NN729.6, не имеющего гена АМР, до делеции гена Amp.
На фиг.4 показана плазмидная карта плазмиды рАК729.6-Δаmр, у которой удален ген АМР.
На фиг.5 показана плазмидная карта плазмиды рАК729.7, используемой для генерации штамма NN729.7, не имеющего гена АМР, до делеции гена Аmр.
На фиг.6 показана плазмидная карта плазмиды pKV228, модифицированной заменой кодирующей последовательности EcoRI (940) - Xbal (1403) в рАК729 (фиг.1) кодирующей последовательностью MFalpha*-Arq34GLP-1(7-37).
Подробное описание изобретения
Ген-маркер устойчивости к антибиотикам удаляют in vitro, для чего используют имеющиеся сайты рестрикции или вводят приемлемые сайты рестрикции при помощи PCR, сайтспецифического мутагенеза или других хорошо известных методов манипуляции последовательностями ДНК с последующей обработкой приемлемыми рестрикционными ферментами.
Конструируют четыре модифицированных штамма NN729, чтобы определить, могут ли разные делеции в плазмиде влиять на выход ферментации предшественника инсулина или стабильность штамма во время длительной ферментации (таблица I). Эти штаммы сравнивают с исходным штаммом NN729 в отношении выхода и стабильности ферментации (таблица II). Помимо этого конструируют три модифицированных дрожжевых штамма, продуцирующих вариант GLP-1 Arq34GLP-1(7-37), чтобы определить, могут ли разные делении в плазмиде pKV228, содержащей ген АМР, влиять на выход ферментации Arg37GLP-1(7-37) (таблица III).
Плазмиды и штаммы, в которых удален ген АМР и, возможно, окружающие последовательности, обозначены "ΔАМР".
Модифицированные дрожжевые штаммы получают, трансформируя модифицированные плазмиды рАК729 или pKV228, в которых удален ген-маркер АМР и, возможно, другие последовательности ДНК исходной плазмиды, в штамме S. cerevisiae МТ663 (Е2-7В ХЕ11-36 а/α ΔtpiΔtpi, pep 4-3/pep 4-3) или МЕ1719 (МАТа/αΔуар3::ura3/Δyap3::URA3pep4-3/pep4-3Δtpi::LEU2/-Δtpi::LEU2 leu2/leu2Δurа3/Δurа3).
Модифицированные плазмиды получают, используя приемлемые сайты рестрикции фермента, имеющиеся в плазмиде, или вставляя приемлемые сайты рестрикции фермента так, чтобы можно было удалить ген АМР. Модифицированные плазмиды можно обрабатывать in vitro до трансформации в S. cerevisiae (штамм МТ663) так, чтобы удалить ген АМР или сделать его нефункциональным, в результате чего в полученном дрожжевом штамме отсутствует ген АМР. Таким образом, вероятность загрязнения окружающей среды геном АМР в процессе уничтожения дрожжевых клеток полностью устраняется.
Модифицированные плазмиды рАК729 или pKV228 расщепляют соответствующими рестрикционными ферментами, подвергают электрофорезу в агарозном геле, выделяют, повторно лигируют и трансформируют в компетентных клетках МТ663 и МЕ1719 S. cerevisiae, как это описано в заявке на патент WO 98/01535.
Способом по этому изобретению можно получить любой гетерологичный белок или полипептид, который можно успешно продуцировать в дрожжевой клетке. Примерами таких белков являются апротинин, ингибитор тканевого фактора или ингибиторы других протеаз, инсулин, предшественники или аналоги инсулина, инсулиноподобный фактор роста I или II, гормон роста человека или быка, интерлейкин, активатор плазминогена ткани, трансформирующий фактор роста а или b, глюкагон, глюкагоноподобный пептид 1 (GLP-1), глюкагоноподобный пептид 2 (GLP-2), GRPP, фактор VII, фактор VIII, фактор XIII, тромбоцитарный фактор роста и ферменты, такие как липазы.
Термин "предшественник инсулина" или "предшественник аналога инсулина" означает одноцепочечный полипептид, включая проинсулин, который при помощи одного или нескольких последующих химических и/или ферментативных процессов можно превратить в двухцепочечную молекулу инсулина или аналога инсулина с правильным расположением трех дисульфидных мостиков, имеющихся в натуральном инсулине человека. Предшественники инсулина обычно содержат модифицированный С-пептид, связанный мостиком с А- и В-цепью инсулина. Кроме того, у предпочтительных предшественников инсулина отсутствует аминокислотный остаток В(30). Наиболее предпочтительные предшественники инсулина описаны, например, в европейском патенте №163529 и в заявках на патент РСТ №№95/00550 и 95/07931. В качестве примеров инсулинов можно привести человеческий инсулин, предпочтительно человеческий инсулин des(B30) и свиной инсулин. Предпочтительными аналогами инсулина являются вещества, в которых один или несколько природных аминокислотных остатков, предпочтительно один, два или три, заменены другим кодируемым аминокислотным остатком. Так в положении А21 исходный инсулин может вместо Asn иметь аминокислотный остаток, выбираемый из группы, включающей Ala, Gln, Glu, Gly, His, Ile, Leu, Met, Ser, Thr, Trp, Туr или Val, в частности аминокислотный остаток, выбираемый из группы, включающей Gly, Ala, Ser и Thr. Аналогичным образом в положении В28 исходный инсулин может вместо Pro иметь аминокислотный остаток, выбираемый из группы, включающей Asp, Lys и т.д., и в положении В29 исходный инсулин может вместо Lys иметь аминокислоту Pro.
Термин "кодируемый аминокислотный остаток" в используемом здесь значении означает аминокислотный остаток, который может быть закодирован генетическим кодом, то есть триплетом ("кодоном") нуклеотидов.
Используемые векторные ДНК можно получить синтетическим путем при помощи стандартных методов, например метода на основе фосфоамидита, описанного S.L. Beaucage and М.Н.Caruthers, Tetrahedron Letters 22, 1981, pp.1859-1869, или метода, описанного Matthes et al., EMBO Journal 3, 1984, pp.801-805. В соответствии с методом на основе фосфоамидита синтезируют олигонуклеотиды, например, в автоматическом синтезаторе ДНК, которые затем очищают, дуплецируют и лигируют с получением синтетической векторной ДНК. Предпочтительным методом получения векторной ДНК в настоящее время является полимеразная цепная реакция (PCR), например, описанная в справочнике Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, NY, 1089).
ДНК, кодирующая целевый белок, может быть геномной ДНК или кДНК, например, полученной путем создания библиотеки геномных ДНК или кДНК и скрининга последовательностей ДНК, кодирующих весь полипептид или его часть, либо при помощи гибридизации с использованием синтетических олиго-нуклеотидных зондов в соответствии с известными методами (см. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, 1989).
И наконец, ДНК, кодирующая целевый белок, может представлять собой смесь синтетической и геномной ДНК, смесь синтетической ДНК и кДНК или смесь геномной ДНК и кДНК, которые получают гибридизацией фрагментов синтетической ДНК, геномной ДНК или кДНК (если это необходимо) в соответствии с известными методами, причем эти фрагменты соответствуют разным частям всей векторной ДНК.
Рекомбинантный экспрессирующий вектор может быть автономно реплицирующимся вектором, то есть вектором, который существует в виде внехромосомного элемента, репликация которого не зависит от репликации хромосомы, например, плазмидой, внехромосомным элементом, минихромосомой или искусственной хромосомой. Вектор может иметь любой механизм, обеспечивающий ауторепликацию. Альтернативно можно использовать вектор, который при введении в клетку-хозяина, внедряется в геном и реплицируется вместе с хромосомой (хромосомами), в которую он внедрился. Векторная система может представлять собой один вектор или плазмиду либо два или больше векторов или плазмид, которые вместе содержат полную ДНК, вводимую в геном клетки-хозяина или транспозон.
Рекомбинантный экспрессирующий вектор может содержать последовательность ДНК, кодирующую целевой пептид или полипептид, которая функционально связана с последовательностью приемлемого промотора. Промотор может представлять собой любую последовательность ДНК, которая обладает транскрипционной активностью в дрожжах и может быть получена из генов, кодирующих белки гомологично или гетерологично дрожжам. Промотор предпочтительно получают из гена, кодирующего белок гомологично дрожжам. В качестве примеров приемлемых промоторов можно привести промоторы Sac-charomyces cerevisiae Mα1, TPI, ADH или PGK.
Последовательность ДНК, кодирующая целевой белок или полипептид, может быть также функционально связана с приемлемым терминатором, например, с терминатором TPI (см. Т.Alber and G.Kawasaki, J. Mol. Appl. Genet, 1, 1982, pp.419-434).
Рекомбинантный экспрессирующий вектор по этому изобретению может также содержать последовательность ДНК, позволяющую вектору реплицироваться в дрожжах. Примерами таких последовательностей являются гены репликации REP 1-3 дрожжевой плазмиды 2μ и точка начала репликации. Этот вектор может также включать селектируемый маркер, например ген TPI Schizosaccharomyces pompe, описанный P.R.Russell, Gene 40, 1985, pp.125-130.
И наконец, экспрессирующий вектор предпочтительно содержит сигнальную/лидерную последовательность, обеспечивающую секрецию целевого белка или полипептида в культуральную среду. Сигнальная последовательность представляет собой последовательность ДНК, кодирующую полипептид ("секреторный пептид"), который, являясь компонентом более крупного полипептида, направляет более крупный полипептид по пути секреции клетки, в которой он синтезирован. Более крупный полипептид обычно расщепляют, чтобы удалить секреторный пептид во время передвижения по пути секреции.
Секреторная сигнальная последовательность может кодировать любой сигнальный пептид, который обеспечивает эффективное перемещение экспрессированного полипептида по пути секреции клетки. Сигнальный пептид может быть естественным сигнальным пептидом или его функциональной частью либо он может быть синтетическим пептидом. Полезные сигнальные пептиды для дрожжевых клеток-хозяев получают из генов а-фактора и инвертазы Saccharomyces cerevisiae, сигнального пептида амилазы слюны мыши (см. О.Hagenbuchle et al., Nature 289, 1981, pp.643-646), сигнального пептида модифицированной карбоксипептидазы (см. L.A.Valls et al., Cell 48, 1987, pp.887-897), сигнального пептида дрожжей BAR1 (см. WO 87/02670) или сигнального пептида аспарагиновой протеазы 3 (YAP3) дрожжей (см. М. Egel-Mitani et al., Yeast 6, 1990, pp.127-137).
Для эффективной секреции в дрожжах последовательность, кодирующую лидерный пептид, можно также вставить внизу от сигнальной последовательности и вверху от последовательности ДНК, кодирующей полипептид. Функция лидерного пептида заключается в том, чтобы направлять экспрессированный полипептид из эндоплазматического ретикулюма в аппарат Гольджи и далее в секреторный пузырек для секреции в культуральную среду (то есть осуществлять перенос полипептида через оболочку клетки или по крайней мере через клеточную мембрану в периплазматическое пространство дрожжевой клетки). Лидерный пептид может быть лидерной последовательностью а-фактора дрожжей (использование которой описано, например, в патенте США №4546082, европейских патентах №№16201, 123294, 123544 и 163529). Альтернативно лидерный пептид может быть синтетическим лидерным пептидом, то есть лидерным пептидом, не существующим в природе. Синтетические лидерные пептиды можно сконструировать, как это описано в заявках на патент WO 89/02463 или WO 92/11378, и в научной работе Kjeldsen et al., "Protein Expression and Purification" 9, 331-336 (1997).
Термин "лидерный пептид" означает пептид в виде пропептидной последовательности, функция которой заключается в том, чтобы направлять секрецию гетерологичного белка из эндоплазматического ретикулюма в аппарат Гольджи и далее в секреторный пузырек для выделения в среду (то есть осуществлять перенос экспрессированного белка или полипептида через клеточную мембрану и оболочку клетки, если она имеется, или по крайней мере через клеточную мембрану в периплазматическое пространство клетки, имеющей оболочку).
Методы, используемые для лигирования последовательностей ДНК, кодирующих соответственно целевой белок или полипептид, промотор и терминатор, и их введения в приемлемые дрожжевые векторы, содержащие информацию, необходимую для репликации в дрожжах, хорошо известны специалистам в этой области (см., например, Sambrook et al., в цитируемой работе). Совершенно очевидно, что вектор можно сконструировать, получив сначала векторную ДНК, содержащую всю последовательность ДНК, кодирующую полипептид по этому изобретению, с последующим введением этого фрагмента в приемлемый экспрессирующий вектор или с последующим введением фрагментов ДНК, содержащих генетическую информацию для отдельных элементов (таких как сигнальная последовательность, лидерная последовательность или гетерологичный белок), и лигированием.
При осуществлении способа по этому изобретению используют любой приемлемый дрожжевой организм, который в процессе культивирования продуцирует целевой белок или полипептид в достаточных количествах. Примерами приемлемых дрожжевых организмов могут служить штаммы, выбираемые из видов дрожжей Saccharomyces cerevisiae, Saccharomyces kluyveri, Schizosaccharomyces pombe, Saccharomyces uvarum, Kluyveromyces lactis, Hansenula polymorpha, Pichia pastoris, Pichia methanolica, Pichia kluyveri, Yarrowia lipolytica, Candida sp., Candida utilis, Candida cacaoi, Geotrichum sp. и Geotrichum fermentans, причем предпочтительным видом дрожжей является Saccharomyces cerevisiae.
Дрожжевые клетки можно трансформировать путем получения протопласта с последующей трансформацией известным методом. Для культивирования клеток можно использовать любую известную среду, приемлемую для выращивания дрожжей. Секретированный гетерологичный белок, значительная часть которого присутствует в среде в правильно процессированной форме, можно выделить из среды известными методами, включая отделение дрожжевых клеток от среды центрифугированием или фильтрованием, осаждение белковых компонентов супернатанта или фильтрата при помощи соли, например сульфата аммония, и последующую очистку разными хроматографическими методами, например, при помощи ионообменной хроматографии, аффинной хроматографии и тому подобных. После секреции белка в периплазматическое пространство клетки разрушают ферментативными или механическими методами.
Целевой белок или полипептид можно экспрессировать и секретировать в виде слитого белка с N-концевым удлиняющим сегментом, описанного в заявке на патент WO 97/22706. N-концевой удлиняющий сегмент затем можно удалить из выделенного белка in vitro при помощи химического или ферментативного расщепления, как это хорошо известно в этой области. Расщепление предпочтительно осуществляют при помощи фермента. Примерами таких ферментов являются трипсин или протеаза I Achromobacter lyticus.
Далее дано более подробное описание настоящего изобретения со ссылкой на нижеследующие примеры, которые не ограничивают объем изобретения, представленного в формуле изобретения.
Пример 1
Дрожжевая плазмида рАК729, сконструированная для экспрессии предшественника инсулина (предшественник инсулина с N-концевым удлиняющим сегментом В(1-29)-Ala-Ala-Lys-A(1-21), см. заявку на патент WO 97/22706), содержит два сайта рестрикции фермента АраLI, в частности сайты ApaLI (4477) и ApaLI (5723) (см. фиг.1). Эти сайты рестрикции расположены с обеих сторон гена-маркера АМР. Удаление 1246 нуклеотидов между двумя сайтами ApaLI в плазмиде рАК729 позволяет удалить ген-маркер АМР и дополнительную плазмидную ДНК, выделенную из Е.coli.
Плазмиду рАК729 расщепляют рестрикционным ферментом ApaLI, подвергают электрофорезу в агарозном геле, выделяют, повторно лигируют и затем трансформируют в компетентных клетках S. cerevisiae (MT663, см. европейский патент №В 0163529), что дает трансформированный дрожжевой штамм NN729.1-ΔАМР. Модифицированную экспрессирующую плазмиду повторно выделяют из дрожжевого штамма NN729.1-ΔАМР, последовательности ДНК проверяют после выполнения PCR и субклонируют область ДНК, имеющую делецию. Аналогичным образом последовательности ДНК, кодирующие предшественник инсулина, проверяют с использованием плазмидной ДНК, повторно выделенной из дрожжевого штамма NN729.1-ΔАМР.
Дрожжевой штамм NN729.1-ΔАМР культивируют в среде YPD при 30°С в течение 72 часов. Выход ферментации предшественника инсулина определяют при помощи жидкостной хроматографии высокого давления с обращенной фазой (RP-HPLC).
Пример 2
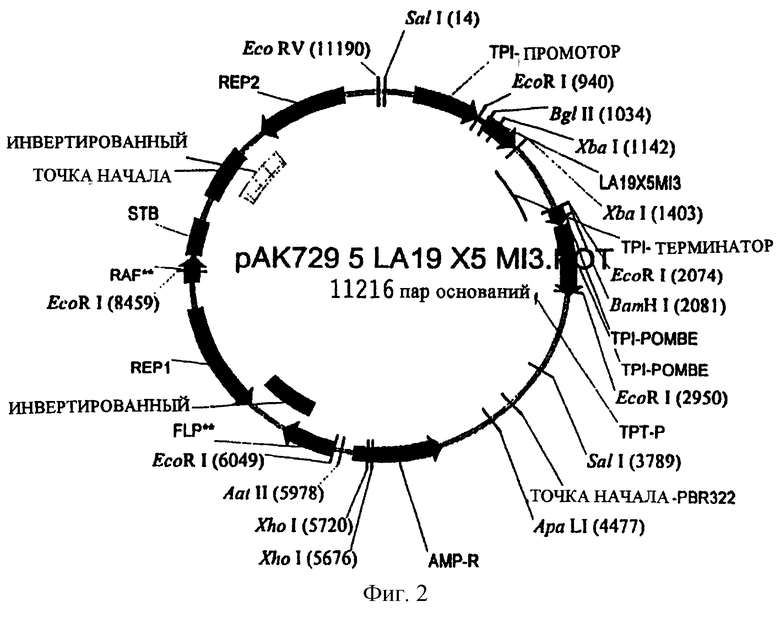
Сайты рестрикции фермента, Xhol (5676) и Xhol (5720), вводят в плазмиду рАК729 при помощи PCR. Затем проверяют отобранные последовательности ДНК полученной плазмиды рАК729.5. Рестриктазная плазмидная карта плазмиды рАК729.5 показана на фиг.2. Фрагмент ДНК, расположенный между сайтами рестрикции фермента Xhol (5676) и Xhol (5720), можно удалить из плазмиды рАК729.5 путем удаления 44 нуклеотидов, расположенных в гене АМР.
Плазмиду рАК729.5 расщепляют рестрикционными ферментами Xhol, подвергают электрофорезу в агарозном геле, выделяют, повторно лигируют и трансформируют в компетентных клетках МТ663 S. cerevisiae, что дает дрожжевой трансформант NN729.5-ΔАМР. Модифицированную экспрессирующую плазмиду повторно выделяют из дрожжевого штамма NN729.5-ΔАМР, последовательности ДНК проверяют после выполнения PCR и субклонируют область ДНК, имеющую делению. Аналогичным бразом последовательности ДНК, кодирующие предшественник инсулина, проверяют с использованием плазмидной ДНК, которую повторно выделяют из дрожжевого штамма NN729.5-ΔАМР. Делеция 44 нуклеотидов в рАК729.5-ΔАМР оказывается такой же эффективной, как и полная делеция гена АМР с точки зрения подавления активности β-лактамазы.
Дрожжевой штамм NN729.5-ΔАМР культивируют в среде YPD при 30°С в течение 72 часов. Выход ферментации предшественника инсулина определяют при помощи RP-HPLC.
Пример 3
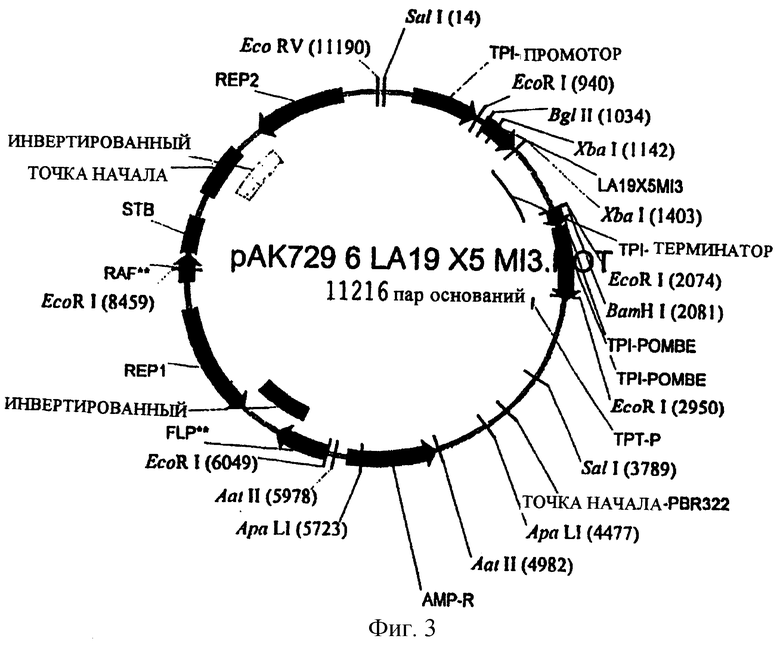
Сайт рестрикции фермента, Aatll (4982), вводят в плазмиду рАК729 при помощи PCR. Затем проверяют отобранные последовательности ДНК полученной плазмиды рАК729.6. Рестриктазная плазмидная карта плазмиды рАК729.6 показана на фиг.3. В плазмиде рАК729.6 можно удалить фрагмент ДНК, расположенный между сайтами рестрикции фермента Aatll (4982) и Aatll (5978), удалив из плазмиды 996 нуклеотидов. Это позволит удалить весь ген АМР и промотор.
Плазмиду рАК729.6 расщепляют рекстрикционным ферментом ДНК Aatll, подвергают электрофорезу в агарозном геле, выделяют, повторно лигируют и трансформируют в компетентных клеткам МТ663 S. cerevisiae. Модифицированную экспрессирующую плазмиду повторно выделяют из дрожжевого штамма NN729.6-ΔАМР, последовательности ДНК проверяют после выполнения PCR и субклонируют область ДНК, имеющую делецию. Аналогичным образом последовательности ДНК, кодирующие предшественник инсулина, проверяют с использованием плазмидной ДНК, повторно выделенной из дрожжевого штамма NN729.6-ΔАМР. Плазмида рАК729.6-ΔАМР, у которой отсутствует ген АМР, показана на фиг.4.
Дрожжевой штамм NN729.6-ΔАМР культивируют в среде YPD при 30°С в течение 72 часов. Выход ферментации предшественника инсулина определяют при помощи RP-HPLC.
Пример 4
Новый сайт рестрикции фермента, Aatll (3801), в плазмиде рАК729.7 вводят в исходную плазмиду рАК729 при помощи PCR. Затем проверяют отобранные последовательности ДНК плазмиды рАК729.7. В плазмиде рАК729.7 можно удалить фрагмент ДНК, расположенный между сайтами рестрикции фермента Aatll (3801) и Aatll (5978), удалив 2177 нуклеотидов из экспрессирующей плазмиды. Плазмиду рАК729.7 конструируют таким образом, чтобы можно было удалить ген АМР и точку начала репликации Е.coli. Рестриктазная плазмидная карта плазмиды рАК729.7 показана на фиг.5.
Плазмиду рАК729.7 расщепляют рестрикционным ферментом ДНК Aatll, подвергают электрофорезу в агарозном геле, выделяют и трансформируют в компетентных клетках МТ663 S. cerevisiae. Модифицированную экспрессирующую плазмиду повторно выделяют из дрожжевого штамма NN729.7-ΔАМР, последовательности ДНК проверяют после выполнения PCR и субклонируют область ДНК, имеющую делению. Аналогичным образом последовательности ДНК, кодирующие предшественник инсулина, проверяют с использованием плазмидной ДНК, повторно выделенной из дрожжевого штамма NN729.7-ΔАМР. Дрожжевой штамм NN729.7-ΔАМР культивируют в среде YPD при 30°С в течение 72 часов. Выход ферментации предшественника инсулина определяют при помощи RP-HPLC.

Новые штаммы NN729-AAMP сравнивают с исходным штаммом NN729 в отношении выхода ферментации предшественника инсулина (таблица II).
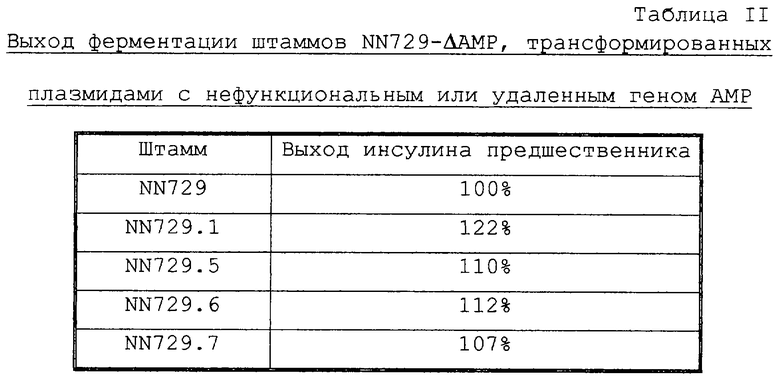
Из вышеизложенного следует, что дрожжевые штаммы, содержащие экспрессирующую плазмиду с частично или полностью удаленным геном-маркером АМР, экспрессируют на 10-20% больше предшественника инсулина по сравнению с исходным дрожжевым штаммом, содержащим экспрессирующую плазмиду с геном АМР.
Пример 5
Экспрессия Arq34GLP-1(7-37) в дрожжах с использованием плазмид с нефункциональным или удаленным геном устойчивости к антибиотикам АМР
Последовательность EcoRI (940) - Xbal (1403) в конструкциях рАК729, кодирующих LA19 Х5 MI3, которые показаны на фигурах 1-5, заменяют кодирующей последовательностью MFalpha*-Arg34GLP-1(7-37) для данного примера (фиг.6). В этой конструкции модифицируют препролидерный пептид MFα1 (Kurjan & Hershowitz, Cell 30, 1982, pp.933), в котором Leu в положении 82 и Asp в положении 83 заменяют соответственно Met и Аlа, вводя в последовательность ДНК расщепляющий сайт Ncol. Лидерную последовательность обозначают MFαl* (Kjeldsen Т. et а1., 1996). Лидерная пептидная последовательность MFαl* сигнальной последовательности MFαl включает двухосновный мотив узнавания Кех2р (Lys-Arg), отделяющий лидерную последовательность от кодирующей последовательности Arg34GLP-1(7-37). Пептид Arg34GLP-1(7-37) является человеческим вариантом GLP-1(7-37) (S.Mojsov, et а1., J. Biol. Chem. 261, 1986, pp.11880-11889), в котором природный аминокислотный остаток в положении 34 заменен остатком Аrg.
Конструируют три экспрессирующие плазмиды Arg34GLP-1(7-37) с удаленным геном устойчивости к антибиотикам АМР, как описано для NN729.1 (пример 1), NN729.5 (пример 2) и NN729.6 (пример 3), и трансформируют их в компетентных клетках МЕ1719 S. cerevisiae (см. заявку на патент WO 98/01535), что дает соответственно дрожжевые трансформанты YES2076, YES2079 и YES2085.
Штамм-хозяин, который используют для экспрессии Arg34GLP-1(7-37), является диплоидным штаммом и имеет фенотипы, у которых отсутствуют аспарагиновые протеазы, то есть (1) дрожжевая аспарагиновая протеаза 3 (YAP3), которая расщепляет С-концевую часть моно- и двухосновных аминокислотных остатков (Egel-Mitani, et al., YEAST 6: 127-137, 1990) и (2) вакуолярная протеаза А, активирующая другие протеазы, такие как протеаза В, карбоксипептидаза Y, аминопептидаза I, РНКаза, щелочная фосфатаза, кислая трегалаза и экзополифосфатаза. Кроме того, удаляют ген триозофосфатизомеразы (TPI), фенотип которого позволяет использовать глюкозу в трансформантах, выращенных на содержащей глюкозу среде. Генетическая среда МЕ1719 представляет собой МАТа/а Dyap3::ura3/Dyap3::URA3 рер4-3/рер4-3 tpi::LEU2/Dtpi::LEU2 leu2/leu2 Dura3/Dura3.
Модифицированные экспрессирующие плазмиды pKV301, pKV307 и pKV304 повторно выделяют из дрожжевых штаммов, последовательности ДНК проверяют после выполнения PCR и субклонируют область ДНК, имеющую делецию. Аналогичным образом последовательности ДНК, кодирующие Arg34GLP-1(7-37), проверяют с использованием плазмидной ДНК, повторно выделенной из дрожжевых штаммов. В таблице III показаны результаты сравнения модифицированных и немодифицированных штаммов.
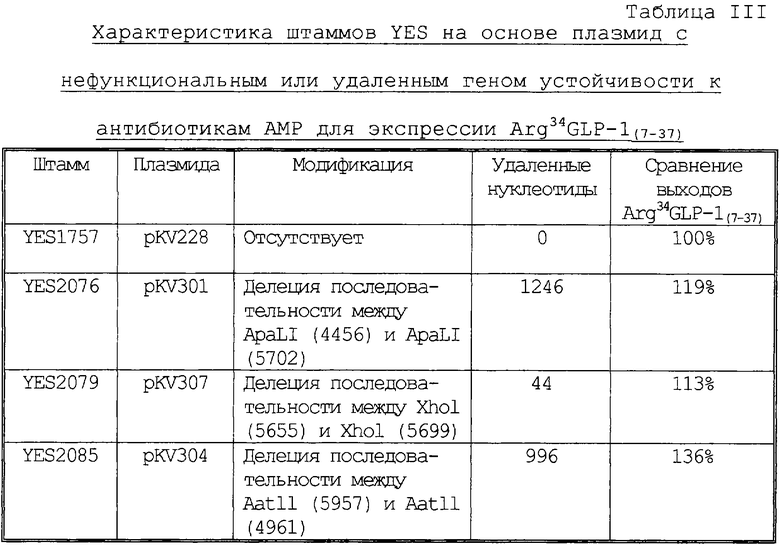
Выходы сравнивают с 5 мл лабораторными ферментациями в среде YPD в течение 72 часов при 30°С. Выходы определяют при помощи HPLC.
Изобретение относится к биотехнологии и представляет собой способ экспрессии полипептидов в дрожжах путем культивирования дрожжевого штамма, который не содержит функционального маркерного гена устойчивости к антибиотику. Трансформированные таким образом дрожжевые клетки продуцируют гетерологичный белок с более высоким выходом. 6 ил., 3 табл.
| EP 0814165 A2, 29.12.1997.EP 0635574 A1, 25.01.1995.WO 9722706, 26.06.1997.RU 2056460 C1, 20.03.1996. |

